
![Quantification of cerebral cannabinoid receptors subtype 1 (CB1) in healthy subjects and schizophrenia by the novel PET radioligand [ 11C]OMAR](https://static.fdokumen.com/doc/165x107/631341bf3ed465f0570a984c/quantification-of-cerebral-cannabinoid-receptors-subtype-1-cb1-in-healthy-subjects.jpg)






Bahasa
Halaman
Hukum


Enhancement of endocannabinoidneurotransmission through CB1 cannabinoidreceptors counteracts the reinforcingand psychostimulant effects of cocaine
Styliani Vlachou1, Fygaleia Stamatopoulou1, George G. Nomikos2 and George Panagis1
1 Laboratory of Behavioral Neuroscience, Department of Psychology, School of Social Sciences, University of Crete,
Rethymnon, Crete, Greece2 Cannasat Therapeutics, Toronto, Ontario, Canada
Abstract
Cannabinoids, in contrast to typical drugs of abuse, have been shown to exert complex effects on
behavioural reinforcement and psychomotor function. We have shown that cannabinoid agonists lack
reinforcing/rewarding properties in the intracranial self-stimulation (ICSS) paradigm and that the CB1
receptor (CB1R) agonist WIN55,212-2 attenuates the reward-facilitating actions of cocaine. We sought to
determine the effects of the endocannabinoid neurotransmission enhancer AM-404 (1, 3, 10, 30 mg/kg) on
the changes in ICSS threshold and locomotion elicited by cocaine and extend the study of the effects
of WIN55,212-2 (0.3, 1, 3 mg/kg) on cocaine-induced hyperlocomotion. AM-404 did not exhibit reward-
facilitating properties, and actually increased self-stimulation threshold at the highest dose. Cocaine
significantly reduced self-stimulation threshold, without altering maximal rates of responding. AM-404
(10 mg/kg) attenuated this action of cocaine, an effect which was reversed by pretreatment with the
selective CB1R antagonist SR141716A. WIN55,212-2 decreased locomotion at the two highest doses, an
effect that was blocked by SR141716A; AM-404 had no effect on locomotion. Cocaine caused a significant,
dose-dependent increase in locomotion, which was reduced by WIN55,212-2 and AM-404. SR141716A
blocked the effects of WIN55,212-2 and AM-404 on cocaine-induced hyperlocomotion. SR141716A alone
had no effect on ICSS threshold or locomotion. These results indicate that cannabinoids may interfere with
brain reward systems responsible for the expression of acute reinforcing/rewarding properties of cocaine,
and provide further evidence that the cannabinoid system could be explored as a potential drug discovery
target for the treatment of psychostimulant addiction and pathological states associated with psychomotor
overexcitability.
Received 19 December 2007 ; Reviewed 19 January 2008 ; Revised 11 February 2008 ; Accepted 17 February 2008 ;
First published online 1 April 2008
Key words : AM-404, cocaine, locomotion, reinforcement, WIN55,212-2.
Introduction
The endocannabinoid system consists of several
recently discovered lipid mediators that produce
their effects via interaction with specific cannabinoid
binding sites (Rodriguez de Fonseca et al., 2005).
Ample evidence indicates that endocannabinoids play
a significant modulatory role in the control of motor
behaviour, nociception, appetite, cognition, affect
and behavioural reinforcement/reward processes
(Chaperon and Thiebot, 1999). Most of the behavioural
studies have been conducted with direct cannabinoid
agonists, which are known for their undesirable psy-
chotropic effects.
Recently, interest has increased for substances that
enhance endocannabinoid neurotransmission through
inhibition of elimination, and therefore may lack
the adverse psychotropic effects induced by direct
stimulation of the cannabinoid CB1 receptor (CB1R;
Huestis et al., 2001). Although some of these en-
docannabinoid modulators have been examined in
Address for correspondence : Dr G. Panagis, University of Crete,
School of Social Sciences, Department of Psychology, Laboratory
of Behavioral Neuroscience, 74100 Rethymnon, Crete, Greece.
Tel. : (30) (28310) 77544 Fax : (30) (28310) 77548
E-mail : [email protected]
International Journal of Neuropsychopharmacology (2008), 11, 905–923. Copyright f 2008 CINPdoi:10.1017/S1461145708008717
ARTICLE
CINP

various behavioural paradigms (see e.g. Compton and
Martin, 1997 ; de Lago et al., 2004 ; Kathuria et al.,
2003), only a few studies have examined their possible
reinforcing/rewarding properties (Bortolato et al.,
2006 ; Gobbi et al., 2005; Hansson et al., 2007 ; Vlachou
et al., 2006).
Cannabis and compounds that stimulate endo-
cannabinoid neurotransmission may arguably affect
the actions of typical drugs of abuse (del Arco et al.,
2002 ; Gallate et al., 1999 ; Solinas et al., 2005; Vigano
et al., 2004 ; Vlachou et al., 2003). Although interactions
with most drugs of abuse have been extensively
studied, little is known about the interaction between
cannabinoids and psychostimulants, possibly because
most psychostimulants, in contrast to other drugs
of abuse, affect the mesolimbic dopaminergic ter-
minals (increasing extracellular dopamine concen-
trations) directly (Lupica and Riegel, 2005). Human
and animal studies (De Vries et al., 2001 ; Foltin et al.,
1993 ; Lukas et al., 1994; Vinklerova et al., 2002) have
shown that cannabis enhances cocaine’s euphorigenic
and behavioural effects. Inversely, other studies reveal
a counteracting effect of cannabinoids on cocaine-
induced behaviour-stimulating actions (Chaperon
et al., 1998 ; Fattore et al., 1999; Martin et al., 2000;
Parker et al., 2004 ; Schenk and Partridge, 1999;
Vlachou et al., 2003). Inconsistent findings have
also been reported with cannabinoids and other psy-
chostimulants, e.g. amphetamine and MDMA (Braida
et al., 2005; Lamarque et al., 2001 ; Muschamp and
Siviy, 2002). Interestingly, intracerebroventricular co-
administration of the CB1R agonist CP55,940 and
MDMA reduces the mean lever presses for their
combined self-administration, compared to self-
administration for each drug alone (Braida and Sala,
2002). In this direction, we have recently shown that
the CB1R agonist WIN55,212-2, in a dose that does
not affect baseline self-stimulation threshold, reduces
cocaine’s reinforcing/rewarding actions (Vlachou
et al., 2003).
The psychomotor effects of cannabinoids have
attracted interest, since the endocannabinoid system
has been implicated in different psychomotor dis-
orders, such as Parkinson’s and Huntington’s disease
and Tourette’s syndrome (Pacher et al., 2006).
Cannabinoids show a biphasic effect on locomotion,
causing hyperactivity in low doses and hypoactivity
or even catalepsy in high doses (Drews et al., 2005;
Sanudo-Pena et al., 2000 ; Sulcova et al., 1998).
However, our knowledge of the effects of indirect
cannabinoid modulators on either spontaneous loco-
motion or hyperlocomotion induced by psychostimu-
lants remains rather limited.
Accordingly, we investigated the effects of the en-
docannabinoid neurotransmission enhancer AM-404
(Beltramo et al., 1997) on brain stimulation reinforce-
ment and on cocaine’s reward-facilitating action. To
assess the possible involvement of CB1R in these ef-
fects, we also examined whether they could be re-
versed by pretreatment with the selective CB1R
antagonist SR141716A (Rinaldi-Carmona et al., 1994),
since not all of the centrally mediated effects of
endocannabinoids occur through CB1R stimulation.
Additionally, we examined the effects of WIN55,212-2
and AM-404 on spontaneous locomotion and cocaine-
induced hyperlocomotion, and whether SR141716A
could counteract these effects.
Materials and methods
Intracranial self-stimulation (ICSS) studies
Animals and surgery
Male Sprague–Dawley rats weighing 300–350 g at the
time of surgery were used. Before surgery they were
housed in groups of three under a 12-h light–dark
cycle (lights on 08:00 hours) with free access to food
and water. The animals were anaesthetized with in-
tramuscular (i.m.) injection of ketamine hydrochloride
(100 mg/kg) and xylazine (10 mg/kg). Atropine sul-
phate (0.6 mg/kg i.m.) was injected to reduce bron-
chial secretion. Movable monopolar stimulating
electrodes (Model SME-01, Kinetrods, Ottawa, ON,
Canada) were lowered into the medial forebrain
bundle (MFB) at the level of lateral hypothalamus
(coordinates AP x2.5 mm from bregma, L x1.7 mm
from the midline, VD x8.0 from a flat skull), accord-
ing to Paxinos and Watson (1998).
The electrodes consisted of a plastic guiding base
and a 0.25-mm diameter movable stainless-steel wire,
which were insulated with epoxylite except for the
conically shaped tip. The anode was an amphenol pin
connected to five miniature skull screws. Following
implantation and for the entire duration of the ex-
periments, the animals were housed individually.
Apparatus and procedures for self-stimulation
One week after surgery, the animals were tested for
self-stimulation in an operant chamber made of
transparent Plexiglas (25 cm wide, 25 cm deep and
30 cm high). A stainless-steel rodent lever protruded
2 cm from the left wall at a height of 4 cm from the
floor. Each lever press triggered a constant current
generator that delivered a 0.4-s train of rectangular
cathodal pulses of constant duration (0.1 ms) and
906 S. Vlachou et al.

intensity (250 mA) and variable frequency (15–100 Hz,
i.e. 6–40 number of pulses/0.4 s). The pulse frequency,
i.e. the number of pulses within a train, was pro-
gressively increased up to 40 per stimulation train
until the subject showed vigorous self-stimulation.
During the acquisition phase the animals were trained
to self-stimulate for at least three consecutive days
(1 h daily), using stimulation parameters that main-
tained near maximal lever-pressing rates. After self-
stimulation had been acquired and stabilized for a
given pulse frequency, animals were trained to self-
stimulate using four alternating series of ascending
and descending pulse frequencies. The pulse fre-
quency was varied by steps of y0.1 log units. Each
frequency was tested within trials of 60 s duration,
followed by an extinction period of 30 s. At the be-
ginning of each trial, the animals received three
trains of priming stimulation, at the frequency of the
stimulation, which was available for that trial. Thus,
drug-induced changes on the rewarding efficacy of
self-stimulation were inferred using the curve-shift
paradigm. This method, which appears to have the
reward selectivity that is required in psychopharma-
cological research, enabled us to distinguish between
reward and performance, while allowing quantifi-
cation of the drug effect.
Drugs
Cocaine hydrochloride (National Monopoly for
Narcotics, Ministry of Health, Greece) was dissolved
in 0.9% NaCl and injected intraperitoneally (i.p.) at a
volume of 1 ml/kg of body weight. AM-404 (Tocris
Bioscience, Ellisville, MO, USA) and SR141716A
(synthesized by Lilly Research Laboratories, Indian-
apolis, IN, USA) were dissolved into a vehicle solu-
tion that consisted of 5% dimethylsulfoxide, 5%
cremophor EL and 90% of 0.9% NaCl and injected i.p.
at a volume of 3 ml/kg of body weight. The doses
of the cannabinoid compounds tested were within
the range of doses regularly used in a plethora of
functional studies (see e.g. Compton andMartin, 1997 ;
de Lago et al., 2004 ; Kathuria et al., 2003).
Experimental procedure
Drug testing began for each animal when the function
relating lever-pressing rate to pulse frequency (the
rate-frequency function) was stable for at least three
consecutive days. The criterion for stability was met
when the frequency thresholds did not vary by more
than 0.1 log unit. Each drug or vehicle self-stimulation
test consisted of a baseline and a drug rate-frequency
function determination (for 45 min each). Following
the baseline period, each animal was injected with the
drug or its vehicle. The animals were tested 10 min
after the last injection.
In the present study we used a mixed design, i.e.
some animals received only one treatment, whereas
other animals received all doses for only one drug
treatment tested. In this case, the sequence of injec-
tions for the different drug doses was counterbalanced
with respect to order and a 3-d period was allowed
between injections. As we have observed in our pre-
vious studies, this period is considered sufficient
for the behaviour of the animals to return to stable,
pre-treatment levels, and not be affected by prior can-
nabinoid administration.
Experiment 1. Effects of systemically administered AM-
404 on brain stimulation reward. Twenty-seven rats were
used. Seven of them received all doses of AM-404 (1, 3,
10, 30 mg/kg i.p.) or its vehicle in a randomized order,
while 20 received only one drug treatment.
Experiment 2. Effects of the CB1R antagonist SR141716A
on AM-404-induced changes in brain stimulation reward.
Fifteen rats were used. Three of them received i.p.
SR141716A (0.02 mg/kg) or vehicle followed 5 min
later by AM-404 (30 mg/kg) or its vehicle i.p. in a
randomized order, while 12 received only one combi-
nation of SR141716A and AM-404 (or their respective
vehicles).
Experiment 3. Effects of AM-404 on the cocaine-induced
lowering of brain reward thresholds. Sixteen rats were
used. Four of them received AM-404 (0, 10 mg/kg i.p.)
followed 5 min later by cocaine (0, 5 mg/kg i.p.), while
12 received only one combination of AM-404 and
cocaine.
Experiment 4. Reversal of the action of AM-404 on the
cocaine-induced lowering of brain reward thresholds by
pretreatment with SR141716A. Fifteen rats were used.
Three of them received all combinations of SR141716A
(0, 0.02 mg/kg i.p.) and AM-404 (0, 10 mg/kg i.p.),
followed 5 min later by cocaine (0, 5 mg/kg i.p.), while
12 received only one combination of SR141716A, AM-
404 and cocaine.
Data analysis and statistics
Data gathered from pre- and post-injection portions of
each session were curve-fitted and threshold and
asymptote estimates were obtained using the
Gompertz sigmoid model (Coulombe and Miliaressis,
1987) :
f (X)=aexeb(xixx)
:
CB1R stimulation counteracts cocaine’s behavioural actions 907

In this equation, a represents the maximum rate
(asymptote), whereas xi (X at inflection) represents the
threshold frequency. The latter is the pulse number
producing 36.7% of the asymptotic rate, i.e. the rate
lying on the fastest accelerating region of the curve.
Parameter b represents an index of the slope whereas
e is the base of natural logarithms.
The post-treatment threshold and asymptote values
were expressed as percentage of pre-drug values.
The results were statistically evaluated using one-way
(effects of cannabinoid agonists alone) or two-way
(effects of combined administration of cannabinoid
agonists and antagonists) or three-way (effects of com-
bined administration of cocaine, cannabinoid agonists
and antagonists) analyses of variance (ANOVA) fol-
lowed, whenever appropriate, by the LSD test for
multiple contrasts. It should be noted that in our de-
sign, some animals received all treatments and others
received only one (see above). In order to combine
these two datasets, for all rats we expressed drug ef-
fects as a percentage of pre-injection baseline, which
was established independently for each drug or ve-
hicle treatment. Each drug test was thus treated as an
independent measure for statistical analysis.
Histology
At the end of the experiment, the animals were given
a lethal dose of sodium pentothal. The location of the
terminal stimulation site was then marked according
to the protocol described by Vlachou et al. (2006). The
brains were then removed and stored in a solution of
10% formalin until sectioned and stained for verifi-
cation of the electrode tips. Only the rats in which
tracks from the electrode were verified to be located in
the MFB were included in this study.
Spontaneous motor activity
Animals
Male Sprague–Dawley rats (n=576 total) weighing
300–350 g were used for all experiments. Animals
were housed three per cage under a 12-h light–dark
cycle (lights on 08:00 hours) with free access to food
and water.
Assessment of locomotor activity
Spontaneous motor activity was measured using an
activity recording system (Model 7445, Ugo Basile).
Each system consists of an animal cage and an elec-
tronic unit incorporating a counter and a printer. The
rectangular animal cage (56r56r30 cm) has trans-
parent sides and lid to allow observation. The cage
floor has horizontal and vertical infrared sensors. The
counter sums up the photocell disruptions, and a
printer displays the results at preset intervals. In our
studies, a summation of photocell disruptions of
locomotor activity and rearing, for each 5-min interval
period, during the 1 h observation period was
registered. The behavioural testing was performed
between 08:00 and 16:00 hours. All animals were
gently handled for 15 min each for 1 wk before the
beginning of the experimental procedure. Two days
before the drug testing each rat was gently handled for
15 min, injected i.p. with saline and habituated to the
experimental room for 3 h in the home cage. The rats
were also accustomed to the experimental room, for
1 h prior to the experiment. Each animal received only
one drug treatment.
Drugs
Cocaine hydrochloride (National Monopoly for Nar-
cotics, Ministry of Health, Greece) was dissolved in
0.9% NaCl and injected i.p. at a volume of 1 ml/kg
of body weight. WIN55,212-2 (Tocris Bioscience),
AM-404 (Tocris Bioscience) and SR141716A (syn-
thesized by Lilly Research Laboratories) were dis-
solved into a vehicle solution that consisted of 5%
dimethylsulfoxide, 5% cremophor EL and 90% of
0.9% NaCl and injected i.p. at a volume of 3 ml/kg
of body weight.
Study 1
Experiment 1. Effect of WIN55,212-2 on locomotor
activity. Animals (n=44) were injected with
WIN55,212-2 (0.3, 1, 3 mg/kg) or its vehicle and after
10 min placed in the centre of the chamber for 1 h.
Experiment 2. Reversal of WIN55,212-2-induced hypolo-
comotion with the CB1R antagonist SR141716A. Animals
(n=40) were injected with SR141716A (0.02 mg/kg)
or its vehicle, and after 5 min with WIN 55,212-2
(3 mg/kg) or its vehicle, and 10 min later they were
placed in the centre of the chamber for 1 h.
Experiment 3. Effect of WIN55,212-2 on cocaine-induced
hyperlocomotion (5 mg/kg). Animals (n=32) were in-
jected with WIN55,212-2 (1 mg/kg) or its vehicle and
after 5 min with cocaine (5 mg/kg) or saline ; im-
mediately thereafter the animals were placed in the
locomotor activity chambers.
Experiment 4. Reversal of the action of WIN55,212-2
on cocaine-induced hyperlocomotion (5 mg/kg) with
SR141716A. Animals (n=72) were injected with
908 S. Vlachou et al.

SR141716A (0.02 mg/kg) or its vehicle and WIN55,
212-2 (1 mg/kg) or its vehicle, followed after 5 min by
cocaine (5 mg/kg) or saline.
Experiment 5. Effect of WIN55,212-2 on cocaine-induced
hyperlocomotion (10 mg/kg). Two separate experiments
(expts 5 and 6) with a higher dose of cocaine (10 mg/
kg) than the dose used for the ICSS experiments
(5 mg/kg) were added in the study to ascertain the
robustness and the reproducibility of the effects of
WIN55,212-2 on counteracting cocaine-induced hy-
perlocomotion, as cocaine clearly increases locomotion
in a dose-dependent manner. Animals (n=36) were
injected with WIN55,212-2 (1 mg/kg) or its vehicle
and after 5 min with cocaine (10 mg/kg) or saline.
Experiment 6. Reversal of the action of WIN55,212-2
on cocaine-induced hyperlocomotion (10 mg/kg) with
SR141716A. Animals (n=64) were injected with
SR141716A (0.02 mg/kg) or its vehicle and
WIN55,212-2 (1 mg/kg) or its vehicle, followed after
5 min by cocaine (10 mg/kg) or saline.
Study 2
Experiment 1. Effect of AM-404 on locomotor activity.
Animals (n=36) were injected with AM-404 (3, 10,
30 mg/kg) or its vehicle and after 10 min placed in the
centre of the chamber for 1 h.
Experiment 2. Effect of AM-404 (10 mg/kg) on cocaine-
induced hyperlocomotion (5 mg/kg). Animals (n=32)
were injected with AM-404 (10 mg/kg) or its vehicle
and after 5 min with cocaine (5 mg/kg) or saline.
Experiment 3. Reversal of the action of AM-404 (10 mg/kg)
on cocaine-induced hyperlocomotion (5 mg/kg) with
SR141716A. Animals (n=80) were injected with
SR141716A (0.02 mg/kg) or its vehicle and AM-404
(10 mg/kg) or its vehicle, followed after 5 min by co-
caine (5 mg/kg) or saline.
Experiment 4. Effect of AM-404 (10 mg/kg) on cocaine-
induced hyperlocomotion (10 mg/kg). Similarly to the
experiments with WIN55,212-2 (see above), three ad-
ditional experiments (expts 4–6) were run with a
higher dose of AM-404 (30 mg/kg) and a higher dose
of cocaine (10 mg/kg) than the respective doses used
in the ICSS paradigm to confirm that AM-404 had
indeed a robust and reproducible action on cocaine-
induced hyperlocomotion. Animals (n=32) were in-
jected with AM-404 (10 mg/kg) or its vehicle and after
5 min, with cocaine (10 mg/kg) or saline.
Experiment 5. Effect of AM-404 (30 mg/kg) on cocaine-
induced hyperlocomotion (10 mg/kg). Animals (n=36)
were injected with AM-404 (30 mg/kg) or its vehicle
and after 5 min with cocaine (10 mg/kg) or saline.
Experiment 6. Reversal of the action of AM-404 (30 mg/
kg) on cocaine-induced hyperlocomotion (10 mg/kg) by
SR141716A. Animals (n=72) were injected with
SR141716A (0.02 mg/kg) or its vehicle and AM-404
(30 mg/kg) or its vehicle, followed after 5 min by
cocaine (10 mg/kg) or saline.
Statistical analysis
The total locomotor activity and rearing counts (de-
pendent variables) over the 1 h observation period
were statistically evaluated by using one-way (effects
of compounds alone), two-way (effects of combined
administration of two compounds) or three-way (ef-
fects of combined administration of three compounds)
ANOVA, followed by post-hoc analysis using the LSD
multiple comparison test in order to reveal specific
differences between groups.
All animal experiments were carried out in accord-
ancewith theNational Institute of Health Guide for the
Care and Use of Laboratory Animals (NIH publication
no. 86-23, revised 1996). Efforts were made in order to
minimize the number of animals used and reduce
their suffering.
Results
ICSS studies
Experiment 1. Effects of systemically administered
AM-404 on brain stimulation reward. The changes
in self-stimulation threshold and asymptotic rate of
responding after systemic injection of the endo-
cannabinoid neurotransmission enhancer AM-404
are presented in Figure 1(a, c), respectively. AM-404
(1, 3, 10, 30 mg/kg i.p.) significantly increased self-
stimulation thresholds at the highest dose tested
[F(4, 50)=4.239, p=0.005], while it had no effect in
the asymptotic rate of responding [F(4, 50)=1.100,
p=0.367]. Post-hoc analysis with the LSD test showed
that these effects on the self-stimulation thresholds
were significant at the highest dose tested (30 mg/
kg i.p.), compared with the other groups (p=0.011,
p=0.024 and p=0.012, respectively) and with the
vehicle-treated group (p<0.001).
Experiment 2. Effects of SR141716A on AM-404-induced
changes in brain stimulation reward. Figure 1(b, d) pre-
sents the changes in self-stimulation threshold and
asymptotic rate of responding after systemic injection
CB1R stimulation counteracts cocaine’s behavioural actions 909
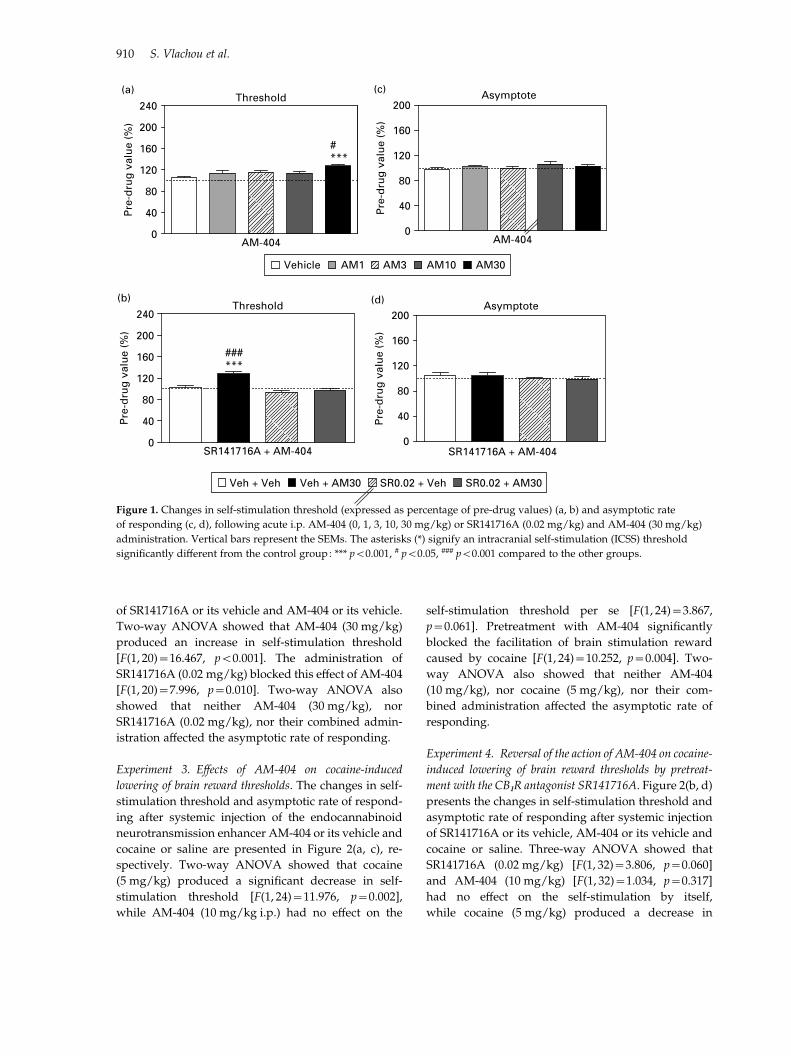
of SR141716A or its vehicle and AM-404 or its vehicle.
Two-way ANOVA showed that AM-404 (30 mg/kg)
produced an increase in self-stimulation threshold
[F(1, 20)=16.467, p<0.001]. The administration of
SR141716A (0.02 mg/kg) blocked this effect of AM-404
[F(1, 20)=7.996, p=0.010]. Two-way ANOVA also
showed that neither AM-404 (30 mg/kg), nor
SR141716A (0.02 mg/kg), nor their combined admin-
istration affected the asymptotic rate of responding.
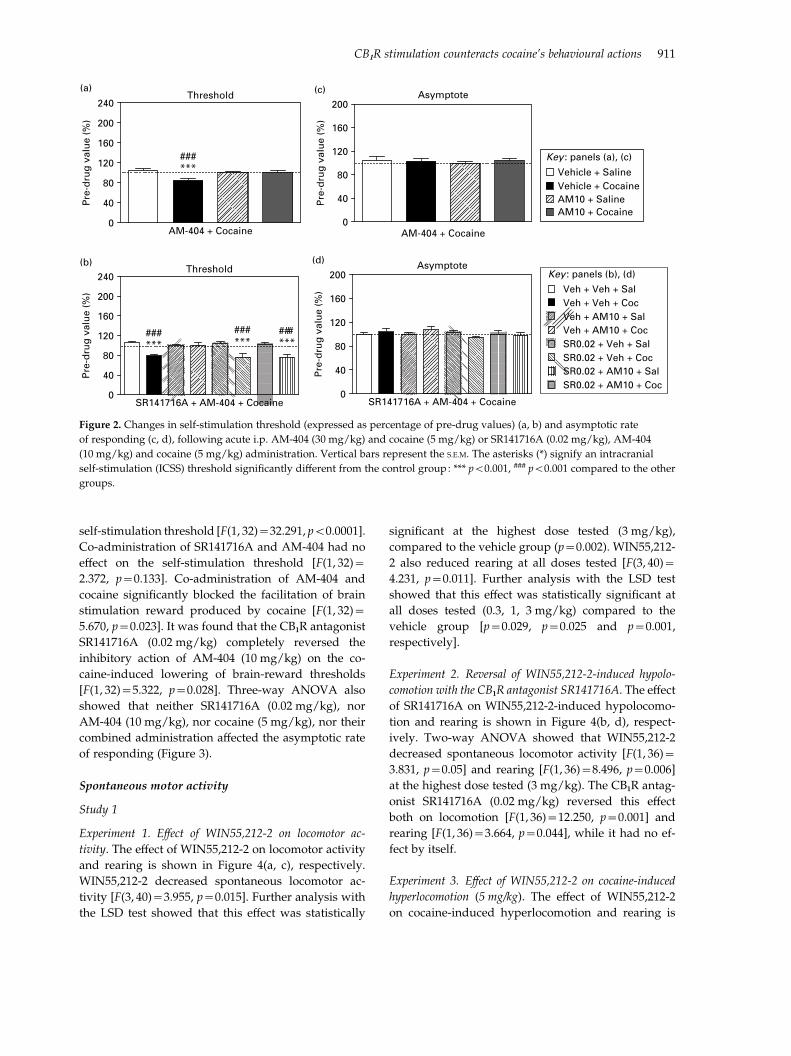
Experiment 3. Effects of AM-404 on cocaine-induced
lowering of brain reward thresholds. The changes in self-
stimulation threshold and asymptotic rate of respond-
ing after systemic injection of the endocannabinoid
neurotransmission enhancer AM-404 or its vehicle and
cocaine or saline are presented in Figure 2(a, c), re-
spectively. Two-way ANOVA showed that cocaine
(5 mg/kg) produced a significant decrease in self-
stimulation threshold [F(1, 24)=11.976, p=0.002],
while AM-404 (10 mg/kg i.p.) had no effect on the
self-stimulation threshold per se [F(1, 24)=3.867,
p=0.061]. Pretreatment with AM-404 significantly
blocked the facilitation of brain stimulation reward
caused by cocaine [F(1, 24)=10.252, p=0.004]. Two-
way ANOVA also showed that neither AM-404
(10 mg/kg), nor cocaine (5 mg/kg), nor their com-
bined administration affected the asymptotic rate of
responding.
Experiment 4. Reversal of the action of AM-404 on cocaine-
induced lowering of brain reward thresholds by pretreat-
ment with the CB1R antagonist SR141716A. Figure 2(b, d)
presents the changes in self-stimulation threshold and
asymptotic rate of responding after systemic injection
of SR141716A or its vehicle, AM-404 or its vehicle and
cocaine or saline. Three-way ANOVA showed that
SR141716A (0.02 mg/kg) [F(1, 32)=3.806, p=0.060]
and AM-404 (10 mg/kg) [F(1, 32)=1.034, p=0.317]
had no effect on the self-stimulation by itself,
while cocaine (5 mg/kg) produced a decrease in
0
40
80
120
160
200
240
(a) (c)
(d)(b)
Threshold
#***
Pre
-dru
g v
alu
e (%
)P
re-d
rug
val
ue
(%)
Pre
-dru
g v
alu
e (%
)P
re-d
rug
val
ue
(%)
0
40
80
120
160
200Asymptote
AM-404AM-404
0
40
80
120
160
200
240
SR141716A + AM-404 SR141716A + AM-404
Threshold
###***
0
40
80
120
160
200Asymptote
Vehicle AM1 AM3 AM10 AM30
Veh + Veh Veh + AM30 SR0.02 + Veh SR0.02 + AM30
Figure 1. Changes in self-stimulation threshold (expressed as percentage of pre-drug values) (a, b) and asymptotic rate
of responding (c, d), following acute i.p. AM-404 (0, 1, 3, 10, 30 mg/kg) or SR141716A (0.02 mg/kg) and AM-404 (30 mg/kg)
administration. Vertical bars represent the SEMs. The asterisks (*) signify an intracranial self-stimulation (ICSS) threshold
significantly different from the control group: *** p<0.001, # p<0.05, ### p<0.001 compared to the other groups.
910 S. Vlachou et al.
self-stimulation threshold [F(1, 32)=32.291, p<0.0001].
Co-administration of SR141716A and AM-404 had no
effect on the self-stimulation threshold [F(1, 32)=2.372, p=0.133]. Co-administration of AM-404 and
cocaine significantly blocked the facilitation of brain
stimulation reward produced by cocaine [F(1, 32)=5.670, p=0.023]. It was found that the CB1R antagonist
SR141716A (0.02 mg/kg) completely reversed the
inhibitory action of AM-404 (10 mg/kg) on the co-
caine-induced lowering of brain-reward thresholds
[F(1, 32)=5.322, p=0.028]. Three-way ANOVA also
showed that neither SR141716A (0.02 mg/kg), nor
AM-404 (10 mg/kg), nor cocaine (5 mg/kg), nor their
combined administration affected the asymptotic rate
of responding (Figure 3).
Spontaneous motor activity
Study 1
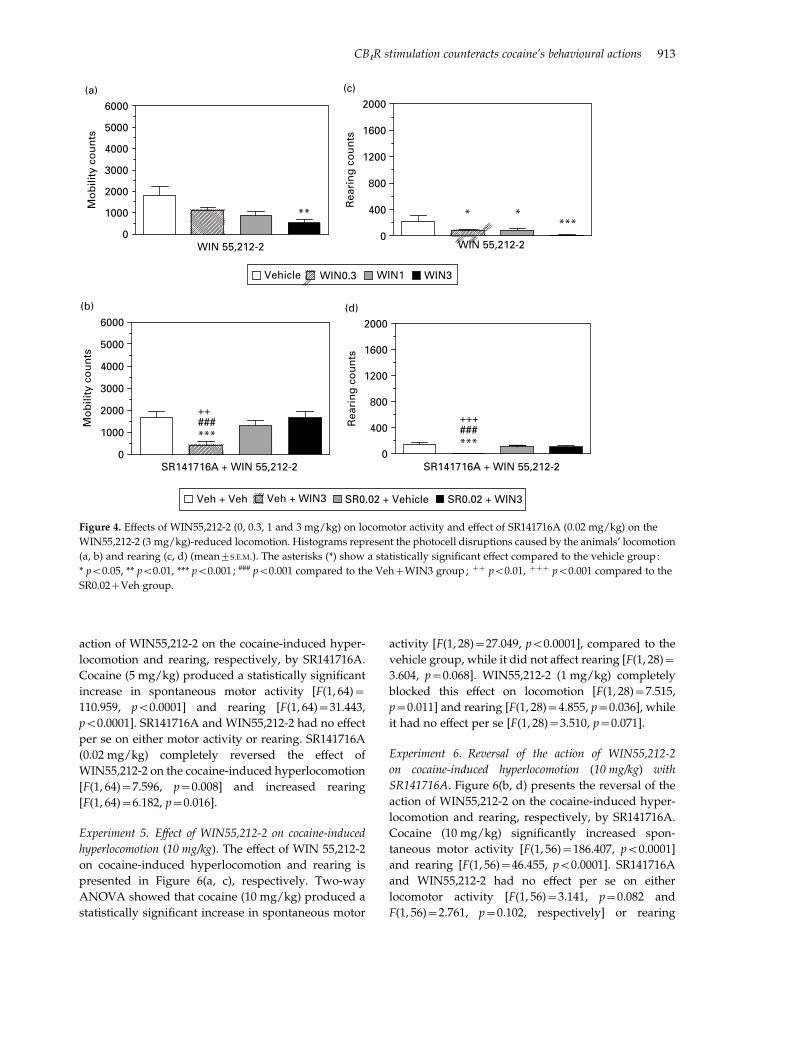
Experiment 1. Effect of WIN55,212-2 on locomotor ac-
tivity. The effect of WIN55,212-2 on locomotor activity
and rearing is shown in Figure 4(a, c), respectively.
WIN55,212-2 decreased spontaneous locomotor ac-
tivity [F(3, 40)=3.955, p=0.015]. Further analysis with
the LSD test showed that this effect was statistically
significant at the highest dose tested (3 mg/kg),
compared to the vehicle group (p=0.002). WIN55,212-
2 also reduced rearing at all doses tested [F(3, 40)=4.231, p=0.011]. Further analysis with the LSD test
showed that this effect was statistically significant at
all doses tested (0.3, 1, 3 mg/kg) compared to the
vehicle group [p=0.029, p=0.025 and p=0.001,
respectively].
Experiment 2. Reversal of WIN55,212-2-induced hypolo-
comotion with the CB1R antagonist SR141716A. The effect
of SR141716A on WIN55,212-2-induced hypolocomo-
tion and rearing is shown in Figure 4(b, d), respect-
ively. Two-way ANOVA showed that WIN55,212-2
decreased spontaneous locomotor activity [F(1, 36)=3.831, p=0.05] and rearing [F(1, 36)=8.496, p=0.006]
at the highest dose tested (3 mg/kg). The CB1R antag-
onist SR141716A (0.02 mg/kg) reversed this effect
both on locomotion [F(1, 36)=12.250, p=0.001] and
rearing [F(1, 36)=3.664, p=0.044], while it had no ef-
fect by itself.
Experiment 3. Effect of WIN55,212-2 on cocaine-induced
hyperlocomotion (5 mg/kg). The effect of WIN55,212-2
on cocaine-induced hyperlocomotion and rearing is
0
40
80
120
160
200
240
(a)
(b)
(c)
(d)
###***
AM-404 + Cocaine AM-404 + Cocaine
ThresholdP
re-d
rug
val
ue
(%)
Pre
-dru
g v
alu
e (%
)
Pre
-dru
g v
alu
e (%
)P
re-d
rug
val
ue
(%)
0
40
80
120
160
200Asymptote
0
40
80
120
160
200
240
SR141716A + AM-404 + Cocaine SR141716A + AM-404 + Cocaine
Threshold
###***
###***
###***
0
40
80
120
160
200Asymptote
Vehicle + Saline
Key: panels (a), (c)
Vehicle + CocaineAM10 + SalineAM10 + Cocaine
Veh + Veh + SalVeh + Veh + CocVeh + AM10 + SalVeh + AM10 + CocSR0.02 + Veh + SalSR0.02 + Veh + CocSR0.02 + AM10 + SalSR0.02 + AM10 + Coc
Key: panels (b), (d)
Figure 2. Changes in self-stimulation threshold (expressed as percentage of pre-drug values) (a, b) and asymptotic rate
of responding (c, d), following acute i.p. AM-404 (30 mg/kg) and cocaine (5 mg/kg) or SR141716A (0.02 mg/kg), AM-404
(10 mg/kg) and cocaine (5 mg/kg) administration. Vertical bars represent the S.E.M. The asterisks (*) signify an intracranial
self-stimulation (ICSS) threshold significantly different from the control group: *** p<0.001, ### p<0.001 compared to the other
groups.
CB1R stimulation counteracts cocaine’s behavioural actions 911

presented in Figure 5(a, c), respectively. WIN55,212-2
(1 mg/kg) did not produce any significant change
on motor activity. Cocaine (5 mg/kg) produced
a statistically significant increase in spontaneous
motor activity [F(1, 28)=5.030, p=0.033] and rearing
[F(1, 28)=4.067, p=0.053], compared to the vehicle
group. WIN55,212-2 (1 mg/kg) completely blocked
this effect on locomotion [F(1, 28)=3.820, p<0.05] and
rearing [F(1, 28)=14.005, p=0.001], while it had no
effect per se.
Experiment 4. Reversal of the action of WIN55,212-2
on cocaine-induced hyperlocomotion (5 mg/kg) with
SR141716A. Figure 5(b, d) presents the reversal of the
Vehicle
0.8 1.0 1.2 1.4 1.6 1.8
0
40
80
120
160
(n = 124) (n = 124)
(n = 119)
(n = 119)
(n = 113)
(n = 104)
Log n pulses/train
Log n pulses/train Log n pulses/train
Log n pulses/trainLog n pulses/train
Log n pulses/train
Leve
r p
ress
ing
/min
Leve
r p
ress
ing
/min
Leve
r p
ress
ing
/min
Leve
r p
ress
ing
/min
Leve
r p
ress
ing
/min
Leve
r p
ress
ing
/min
AM30
0.8 1.0 1.2 1.4 1.6 1.8
0
40
80
120
160
Vehicle + Cocaine
0.8 1.0 1.2 1.4 1.6 1.8
0
40
80
120
160 AM10 + Cocaine
0.8 1.0 1.2 1.4 1.6 1.8
0
40
80
120
160
SR0.02 + Vehicle + Cocaine
0.8 1.0 1.2 1.4 1.6 1.8
0
40
80
120
160 SR0.02 + AM10 + Cocaine
0.8 1.0 1.2 1.4 1.6 1.8
0
40
80
120
160
Figure 3. Rate-frequency functions (rate of lever pressing as a function of stimulation frequency) taken from representative
animals for each drug treatment. Each plot represents data from a single animal under pre-drug and drug conditions.
Rate frequency functions were obtained by logarithmically decreasing the frequency of the stimulation pulses
from a value that sustained maximal lever pressing to one that failed to sustain lever pressing. –#–, Pre-injection ;
–$–, post-injection.
912 S. Vlachou et al.
action of WIN55,212-2 on the cocaine-induced hyper-
locomotion and rearing, respectively, by SR141716A.
Cocaine (5 mg/kg) produced a statistically significant
increase in spontaneous motor activity [F(1, 64)=110.959, p<0.0001] and rearing [F(1, 64)=31.443,
p<0.0001]. SR141716A and WIN55,212-2 had no effect
per se on either motor activity or rearing. SR141716A
(0.02 mg/kg) completely reversed the effect of
WIN55,212-2 on the cocaine-induced hyperlocomotion
[F(1, 64)=7.596, p=0.008] and increased rearing
[F(1, 64)=6.182, p=0.016].
Experiment 5. Effect of WIN55,212-2 on cocaine-induced
hyperlocomotion (10 mg/kg). The effect of WIN 55,212-2
on cocaine-induced hyperlocomotion and rearing is
presented in Figure 6(a, c), respectively. Two-way
ANOVA showed that cocaine (10 mg/kg) produced a
statistically significant increase in spontaneous motor
activity [F(1, 28)=27.049, p<0.0001], compared to the
vehicle group, while it did not affect rearing [F(1, 28)=3.604, p=0.068]. WIN55,212-2 (1 mg/kg) completely
blocked this effect on locomotion [F(1, 28)=7.515,
p=0.011] and rearing [F(1, 28)=4.855, p=0.036], while
it had no effect per se [F(1, 28)=3.510, p=0.071].
Experiment 6. Reversal of the action of WIN55,212-2
on cocaine-induced hyperlocomotion (10 mg/kg) with
SR141716A. Figure 6(b, d) presents the reversal of the
action of WIN55,212-2 on the cocaine-induced hyper-
locomotion and rearing, respectively, by SR141716A.
Cocaine (10 mg/kg) significantly increased spon-
taneous motor activity [F(1, 56)=186.407, p<0.0001]
and rearing [F(1, 56)=46.455, p<0.0001]. SR141716A
and WIN55,212-2 had no effect per se on either
locomotor activity [F(1, 56)=3.141, p=0.082 and
F(1, 56)=2.761, p=0.102, respectively] or rearing
WIN 55,212-20
1000
2000
3000
4000
5000
6000
(a) (c)
(d)(b)
**
Mo
bili
ty c
ou
nts
WIN 55,212-20
400
800
1200
1600
2000
Vehicle WIN0.3 WIN1 WIN3
****
*
Rea
rin
g c
ou
nts
SR141716A + WIN 55,212-20
1000
2000
3000
4000
5000
6000
Veh + Veh Veh + WIN3 SR0.02 + Vehicle SR0.02 + WIN3
++###***
Mo
bili
ty c
ou
nts
SR141716A + WIN 55,212-20
400
800
1200
1600
2000
+++###***
Rea
rin
g c
ou
nts
Figure 4. Effects of WIN55,212-2 (0, 0.3, 1 and 3 mg/kg) on locomotor activity and effect of SR141716A (0.02 mg/kg) on the
WIN55,212-2 (3 mg/kg)-reduced locomotion. Histograms represent the photocell disruptions caused by the animals’ locomotion
(a, b) and rearing (c, d) (mean¡S.E.M.). The asterisks (*) show a statistically significant effect compared to the vehicle group:
* p<0.05, ** p<0.01, *** p<0.001 ; ### p<0.001 compared to the Veh+WIN3 group; ++ p<0.01, +++ p<0.001 compared to the
SR0.02+Veh group.
CB1R stimulation counteracts cocaine’s behavioural actions 913

[F(1, 56)=1.122, p=0.294 and F(1, 56)=3.067, p=0.085,
respectively]. SR141716A (0.02 mg/kg) completely
reversed the effect of WIN55,212-2 on the cocaine-
induced hyperlocomotion [F(1, 56)=7.170, p=0.010]
and increased rearing [F(1, 56)=4.855, p=0.036].
Study 2
Experiment 1. Effect of AM-404 on locomotor activity.
AM-404 did not affect significantly locomotion
[F(3, 32)=0.701, p=0.559] or rearing [F(3, 32)=0.995,
p=0.408], although there was a tendency for a re-
duction in both indices at all doses tested, as shown in
Figure 7(a, b), respectively.
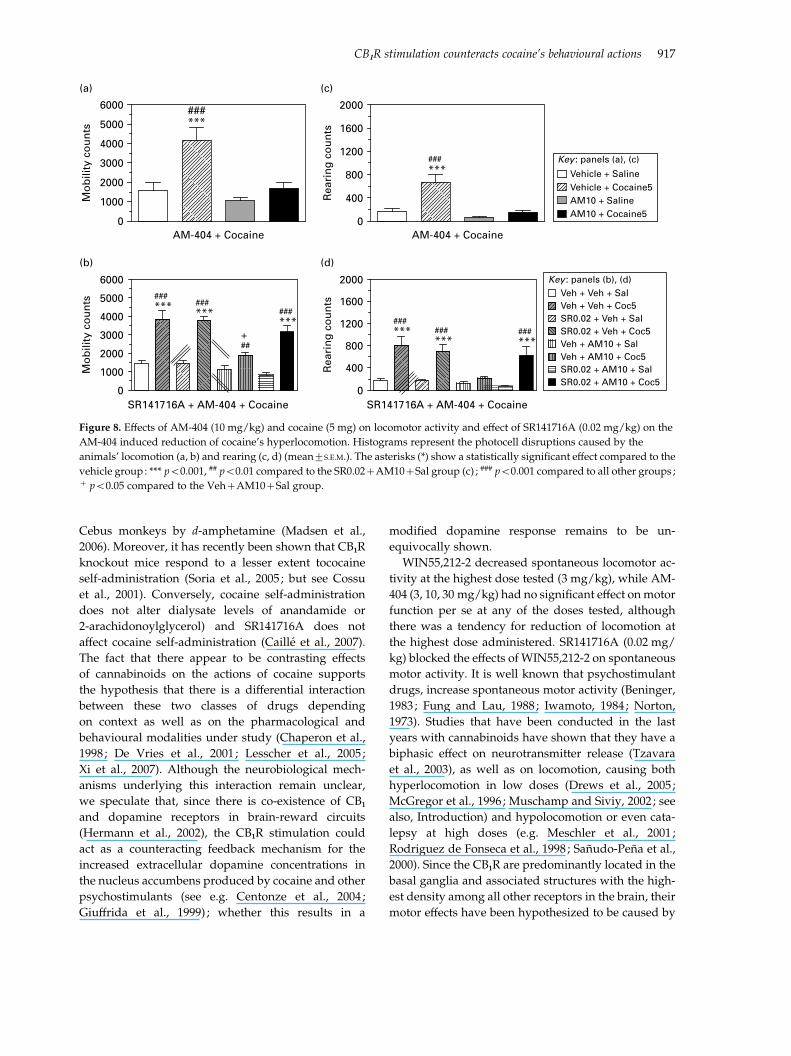
Experiment 2. Effect of AM-404 (10 mg/kg) on cocaine-
induced hyperlocomotion (5 mg/kg). Cocaine (5 mg/kg)
produced a statistically significant increase in spon-
taneous motor activity [F(1, 28)=14.535, p=0.001]
and rearing [F(1, 28)=14.141, p=0.001]. AM-404
(10 mg/kg) significantly blocked this effect on
locomotion [F(1, 28)=5.319, p=0.029] and rearing
[F(1, 28)=6.974, p=0.013], as presented in Figure 8(a,
c), respectively.
Experiment 3. Reversal of the action of AM-404 (10 mg/kg)
on cocaine-induced hyperlocomotion (5 mg/kg) with
SR141716A. Cocaine (5 mg/kg) produced a significant
increase in motor activity [F(1, 72)=119.549, p<0.0001] and rearing [F(1, 72)=48.882, p<0.0001].
SR141716A (0.02 mg/kg) did not have any effect
on motor activity. There was a statistically signif-
icant interaction between AM-404 and cocaine in
motor activity [F(1, 72)=4.846, p=0.031] and rearing
[F(1, 72)=4.065, p=0.048]. This was due to the fact
that, AM-404 blocked the stimulatory effects of
cocaine. SR141716A (0.02 mg/kg) completely reversed
the effect of AM-404 on cocaine-induced hyper-
locomotion [F(1, 72)=5.439, p=0.022] and increased
rearing [F(1, 72)=5.188, p=0.026], as shown in
Figure 8(b, d), respectively.
Experiment 4. Effect of AM-404 (10 mg/kg) on cocaine-
induced hyperlocomotion (10 mg/kg). Figure 9(a, d)
shows the effect of AM-404 on cocaine-induced
hyperlocomotion and rearing, respectively. Cocaine
(10 mg/kg) produced a statistically significant
increase in locomotor activity [F(1, 28)=22.912, p<0.0001] and rearing [F(1, 28)=12.646, p=0.001].
WIN 55,212-2 + Cocaine0
1000
2000
3000
4000
5000
6000
(a) (c)
(d)(b)
+##**
Mo
bili
ty c
ou
nts
WIN 55,212-2 + Cocaine0
400
800
1200
1600
2000
+++##***
Rea
rin
g c
ou
nts
SR141716A + WIN 55,212-2 + Cocaine0
1000
2000
3000
4000
5000
6000
###***
###***
*
###***
Mo
bili
ty c
ou
nts
SR141716A - WIN 55,212-2 + Cocaine0
400
800
1200
1600
2000
##**
##**
###***
Rea
rin
g c
ou
nts
Veh + Veh + Sal
Veh + Veh + Coc5
SR0.02 + Veh + Sal
SR0.02 + Veh + Coc5
Veh + WIN1 + Sal
Veh + WIN1 + Coc5
SR0.02 + WIN1 + Sal
SR0.02 + WIN1 + Coc5
Vehicle + Saline
Vehicle + Cocaine5
WIN1 + Saline
WIN1 + Cocaine5
Key: panels (a), (c)
Key: panels (b), (d)
Figure 5. Effects of WIN55,212-2 (1 mg/kg) and cocaine (5 mg) on locomotor activity and effect of SR141716A (0.02 mg/kg)
on the WIN55,212-2-induced reduction of cocaine’s hyperlocomotion. Histograms represent the photocell disruptions caused
by the animals’ locomotion (a, b) and rearing (c, d) (mean¡S.E.M.). The asterisks (*) show a statistically significant effect
compared to the vehicle group: ** p<0.01, *** p<0.001, ## p<0.01, ### p<0.001 compared to all other groups ; ++ p<0.01,+++ p<0.001 compared to the WIN1+Coc5 group.
914 S. Vlachou et al.

AM-404 (10 mg/kg), which had no effect per se, failed
to affect either the hyperlocomotion [F(1, 28)=0.106,
p=0.747] or the enhanced rearing [F(1, 28)=1.427,
p=0.242] induced by cocaine, while there was a tend-
ency for reduction in the increased rearing induced by
cocaine.
Experiment 5. Effect of AM-404 (30 mg/kg) on cocaine-
induced hyperlocomotion (10 mg/kg). Figure 9(b, e) shows
the effect of AM-404 (30 mg/kg) on cocaine-induced
hyperlocomotion (10 mg/kg) and rearing, respect-
ively. Two-way ANOVA showed that cocaine (10 mg/
kg) produced a statistically significant increase in
locomotor activity [F(1, 32)=175.820, p<0.0001] and
rearing [F(1, 32)=49.002, p<0.0001]. AM-404 (30 mg/
kg) significantly blocked the hyperlocomotion
[F(1, 32)=9.333, p=0.005] and rearing [F(1, 32)=5.231,
p=0.029] induced by cocaine.
Experiment 6. Reversal of the action of AM-404 (30 mg/kg)
on cocaine-induced hyperlocomotion (10 mg/kg) with
SR141716A. Cocaine (10 mg/kg) produced a signifi-
cant increase in motor activity [F(1, 64)=158.361,
p<0.0001] and rearing [F(1, 64)=86.983, p<0.0001].
There was a statistically significant effect of cocaine
after the administration of SR141716A and AM-404
on motor activity [F(1, 64)=4.599, p=0.036 and
F(1, 64)=6.600, p=0.013, respectively] and rearing
[F(1,64)=4.579, p=0.034 and F(1, 64)=5.381, p=0.024,
respectively], although it is clearly shown that AM-
404 reduced the hyperlocomotor effect of cocaine.
SR141716A (0.02 mg/kg) completely reversed the ef-
fect of AM-404 on the cocaine-induced hyperlocomo-
tion [F(1, 64)=5.153, p=0.027], while there seemed to
be no interaction on rearing [F(1, 64)=2.742, p=0.103],
as shown in Figure 9(c, f), respectively.
Discussion
The present study showed that the endocannabinoid
neurotransmission enhancer AM-404 at relatively
low doses (1, 3 and 10 mg/kg) did not affect in-
tracranial self-stimulation behaviour, whereas at
the highest dose (30 mg/kg) increased brain reward
thresholds. These changes were not accompanied
by significant changes in asymptotic rates of respond-
ing. The anhedonic-like effect of AM-404, observed
at the highest dose, was completely abolished by
pretreatment with the CB1R antagonist SR141716A.
However, since it is not clear whether AM-404 at the
WIN 55,212-2 + Cocaine0
1000
2000
3000
4000
5000
6000***
(a)
+###
#
Mo
bili
ty c
ou
nts
Mo
bili
ty c
ou
nts
WIN 55,212-2 + Cocaine0
400
800
1200
1600
2000 ##**
(c)
SR141716A + WIN 55,212-2 + Cocaine0
1000
2000
3000
4000
5000
6000 ###***
(b)
###***
###***
###***
SR141716A + WIN 55,212-2 + Cocaine0
400
800
1200
1600
2000
++###***
(d)
++###*** +
###***
Rea
rin
g c
ou
nts
Rea
rin
g c
ou
nts
Veh + Veh + SalVeh + Veh + Coc10SR0.02 + Veh + SalSR0.02 + Veh + Coc10Veh + WIN1 + SalVeh + WIN1 + Coc10SR0.02 + WIN1 + SalSR0.02 + WIN1 + Coc10
Vehicle + SalineVehicle + Cocaine10WIN1 + SalineWIN1 + Cocaine10
Key : panels (b), (d)
Key : panels (a), (c)
Figure 6. Effects of WIN55,212-2 (1 mg/kg) and cocaine (10 mg) on locomotor activity and effect of SR141716A (0.02 mg/kg)
on the WIN55,212-2-induced reduction of cocaine’s hyperlocomotion. Histograms represent the photocell disruptions caused
by the animals’ locomotion (a, b) and rearing (c, d) (mean¡S.E.M.). The asterisks (*) show a statistically significant effect compared
to the vehicle group: ** p<0.01, *** p<0.001, # p<0.05, ## p<0.01, ### p<0.001 compared to all other groups ; + p<0.05,++ p<0.01 compared to the WIN1+Coc10 (a) or Veh+WIN1+Coc10 group (d).
CB1R stimulation counteracts cocaine’s behavioural actions 915

dose of 30 mg/kg can still selectively prevent ana-
ndamide elimination, the observed effects could be
due to a non-selective increase of extracellular con-
centrations of endocannabinoids other than ananda-
mide or due to unselective actions of this drug, such as
direct stimulation of the CB1R, the latter been more
probable since AM-404 action was reversed by
SR141716A (Dickason-Chesterfield et al., 2006). It is
worth noting that in our study SR141716A did not af-
fect ICSS threshold by itself, suggesting lack of a tonic
regulation of reward/reinforcement processes via
CB1R. The present results are in agreement with a re-
cent study from our laboratory (Vlachou et al., 2006),
in which the endocannabinoid neurotransmission
modulators PMSF, URB-597 and OMDM-2, depending
on the dose administered, either do not affect or de-
crease brain reward function. The finding that AM-404
lacks appetitive properties also agrees with other stu-
dies, where anandamide and URB-597 do not produce
conditioned place preference in male Wistar rats
(Gobbi et al., 2005 ; Mallet and Beninger, 1998).
However, it should be mentioned that anandamide
and methanandamide have been shown to be self-
administered by squirrel monkeys (Justinova et al.,
2005) and AM-404 induces conditioned place pre-
ference in rats living in an enriched environment
(Solinas et al., 2005). These seemingly contrasting re-
sults could be attributed to differences in the pharma-
cological properties and the dose range of the tested
compounds, the species and the strain of the animals
used or the methods followed; in addition, the route
of administration might be a determinant factor in
assessing appetitive actions of cannabinoids, as has
previously been argued (Vlachou et al., 2003, 2005,
2006, 2007).
Acute administration of AM-404 counteracted the
reward-facilitating effect of systemic cocaine at a dose
that by itself was ineffective in altering brain stimu-
lation reward thresholds. Cocaine (5 mg/kg) pro-
duced a significant reduction in self-stimulation
threshold, without altering maximal rates of respond-
ing, in agreement with previous studies (Maldonado-
Irrizary et al., 1994 ; Matthews et al., 1996; Panagis
et al., 2000; Panagis and Spyraki, 1996; Ranaldi et al.,
1997 ; Vlachou et al., 2003). AM-404 (10 mg/kg) atten-
uated the reward-facilitating action of cocaine, an
effect which was reversed by pretreatment with
SR141716A (0.02 mg/kg). The present results are in
agreement with a number of studies, indicating in-
teractions of cannabinoids with psychostimulants
(Braida et al., 2001 ; Braida and Sala, 2002 ; Cadoni et al.,
2001 ; De Vries et al., 2001 ; Fattore et al., 1999;
Gallate et al., 1999 ; Ghozland et al., 2002 ; Gonzalez
et al., 2002a,b ; Filip et al., 2006 ; Gonzalez-Cuevas et al.,
2007 ; Higuera-Matas et al., 2007 ; Lamarque et al.,
2001 ; Navarro et al., 2001 ; Pontieri et al., 2001a,b;
Tanda, 2007 ; Vlachou et al., 2003). We have recently
shown that the CB1R agonist WIN55,212-2 at a dose
that does not affect brain stimulation reward per se
counteracts the reward-facilitating effects of cocaine
(Vlachou et al., 2003). Fattore and colleagues have
shown that WIN55,212-2 reduces the intravenous
self-administration of cocaine (Fattore et al., 1999), a
finding also consistent with the most recent study
by Panlilio and colleagues, in which D9-tetra-
hydrocannabinol (THC) was found to reduce the self-
administration of cocaine under a progressive ratio
schedule of reinforcement (Panlilio et al., 2007), and
with another recent study by Cippitelli and colleagues,
in which AM-404 was found to reduce ethanol
self-administration (Cippitelli et al., 2007). On the other
hand, it has been found that the potent CB1R agonist
HU-210 induces relapse to cocaine-seeking after a pro-
longed withdrawal period (De Vries et al., 2001), and
CP55,940 does not seem to affect the arousal caused to
00 3
AM-404 (mg)
AM-404 (mg)
10 30
0 3 10 30
1000
2000
3000
4000
5000
6000
Mo
bili
ty c
ou
nts
0
400
800
1200
1600
2000
(b)
(a)
Rea
rin
g c
ou
nts
Figure 7. Effects of AM-404 [0 (vehicle), 3, 10 and 30 mg] on
locomotor activity and rearing. Histograms represent the
photocell disruptions caused by the animals’ locomotion (a)
and rearing (b) (mean¡S.E.M.).
916 S. Vlachou et al.
Cebus monkeys by d-amphetamine (Madsen et al.,
2006). Moreover, it has recently been shown that CB1R
knockout mice respond to a lesser extent tococaine
self-administration (Soria et al., 2005 ; but see Cossu
et al., 2001). Conversely, cocaine self-administration
does not alter dialysate levels of anandamide or
2-arachidonoylglycerol) and SR141716A does not
affect cocaine self-administration (Caille et al., 2007).
The fact that there appear to be contrasting effects
of cannabinoids on the actions of cocaine supports
the hypothesis that there is a differential interaction
between these two classes of drugs depending
on context as well as on the pharmacological and
behavioural modalities under study (Chaperon et al.,
1998 ; De Vries et al., 2001 ; Lesscher et al., 2005 ;
Xi et al., 2007). Although the neurobiological mech-
anisms underlying this interaction remain unclear,
we speculate that, since there is co-existence of CB1
and dopamine receptors in brain-reward circuits
(Hermann et al., 2002), the CB1R stimulation could
act as a counteracting feedback mechanism for the
increased extracellular dopamine concentrations in
the nucleus accumbens produced by cocaine and other
psychostimulants (see e.g. Centonze et al., 2004 ;
Giuffrida et al., 1999) ; whether this results in a
modified dopamine response remains to be un-
equivocally shown.
WIN55,212-2 decreased spontaneous locomotor ac-
tivity at the highest dose tested (3 mg/kg), while AM-
404 (3, 10, 30 mg/kg) had no significant effect onmotor
function per se at any of the doses tested, although
there was a tendency for reduction of locomotion at
the highest dose administered. SR141716A (0.02 mg/
kg) blocked the effects of WIN55,212-2 on spontaneous
motor activity. It is well known that psychostimulant
drugs, increase spontaneous motor activity (Beninger,
1983 ; Fung and Lau, 1988 ; Iwamoto, 1984 ; Norton,
1973). Studies that have been conducted in the last
years with cannabinoids have shown that they have a
biphasic effect on neurotransmitter release (Tzavara
et al., 2003), as well as on locomotion, causing both
hyperlocomotion in low doses (Drews et al., 2005 ;
McGregor et al., 1996 ; Muschamp and Siviy, 2002; see
also, Introduction) and hypolocomotion or even cata-
lepsy at high doses (e.g. Meschler et al., 2001 ;
Rodriguez de Fonseca et al., 1998 ; Sanudo-Pena et al.,
2000). Since the CB1R are predominantly located in the
basal ganglia and associated structures with the high-
est density among all other receptors in the brain, their
motor effects have been hypothesized to be caused by
AM-404 + Cocaine0
1000
2000
3000
4000
5000
6000
0
1000
2000
3000
4000
5000
6000
###***
(a)M
ob
ility
co
un
tsM
ob
ility
co
un
ts
AM-404 + Cocaine0
400
800
1200
1600
2000
###***
(c)
Rea
rin
g c
ou
nts
Rea
rin
g c
ou
nts
SR141716A + AM-404 + Cocaine
###***
+##
(b)
###*** ###
***
SR141716A + AM-404 + Cocaine0
400
800
1200
1600
2000
###***
(d)
###***
###***
Vehicle + SalineVehicle + Cocaine5AM10 + SalineAM10 + Cocaine5
Veh + Veh + SalVeh + Veh + Coc5SR0.02 + Veh + SalSR0.02 + Veh + Coc5Veh + AM10 + SalVeh + AM10 + Coc5SR0.02 + AM10 + SalSR0.02 + AM10 + Coc5
Key : panels (b), (d)
Key : panels (a), (c)
Figure 8. Effects of AM-404 (10 mg/kg) and cocaine (5 mg) on locomotor activity and effect of SR141716A (0.02 mg/kg) on the
AM-404 induced reduction of cocaine’s hyperlocomotion. Histograms represent the photocell disruptions caused by the
animals’ locomotion (a, b) and rearing (c, d) (mean¡S.E.M.). The asterisks (*) show a statistically significant effect compared to the
vehicle group: *** p<0.001, ## p<0.01 compared to the SR0.02+AM10+Sal group (c) ; ### p<0.001 compared to all other groups ;+ p<0.05 compared to the Veh+AM10+Sal group.
CB1R stimulation counteracts cocaine’s behavioural actions 917

an interplay of actions on the dopaminergic, gluta-
matergic and GABAergic systems in these brain areas
(Sanudo-Pena et al., 1999, 2000). Therefore, the re-
duction in locomotor activity observed in our studies
could be attributed to stimulation of CB1R in the
striatum, which increases GABAergic neurotransmis-
sion in the globus pallidus or substantia nigra (Maneuf
et al., 1997). It could also be due to trans-synaptic
stimulation of GABAergic neurons, which project to
the substantia nigra and cause reduced stimulation of
the nigrostriatal dopaminergic neurons (Gueudet et al.,
1995).
Our results are in agreement with previous studies
showing that anandamide, THC or CB1R agonists re-
duce motor function (Darmani, 2001 ; Ferrari et al.,
1999 ; Romero et al., 2002 ; Singh et al., 2005 ; Valjent
et al., 2002). However, other studies have shown dif-
ferent results (Drews et al., 2005; Muschamp and
Siviy, 2002 ; Przegalinski et al., 2005). Przegalinski and
colleagues have shown that administration of 3 mg
and 6 mg WIN55,212-2 does not affect locomotion,
while in our study there was a clear hypolocomotion
and reduced rearing after administration of 3 mg
WIN55,212-2. Our results also differ from Muschamp
and Siviy (2002), where there was an increase in
locomotion with 1 mg/kg WIN55,212-2, although this
was shown in Lewis rats and only in the first 15 min
of the experimental session. It is probable that the
different results could be attributed to different ex-
perimental conditions.
Cocaine (5, 10 mg/kg) caused a significant, dose-
dependent increase in locomotor activity, which was
AM-404 + Cocaine0
1000
2000
3000
4000
5000
6000##**
(a)
##**
AM-404 + Cocaine0
400
800
1200
1600
2000
+###**
(d)
AM-404 + Cocaine0
1000
2000
3000
4000
5000
6000 ###***
(b)
+++###***
AM-404 + Cocaine0
400
800
1200
1600
2000###***
(e)
+++##**
Rea
rin
g c
ou
nts
Rea
rin
g c
ou
nts
SR141716A + AM-404 + Cocaine0
100020003000400050006000 ###
***
(c)
###***
###***
+++##**
Mo
bili
ty c
ou
nts
Mo
bili
ty c
ou
nts
Mo
bili
ty c
ou
nts
Rea
rin
g c
ou
nts
SR141716A + AM-404 + Cocaine0
400
800
1200
1600
2000
###***
(f)
###***###
*** +++#*
Vehicle + SalineVehicle + Cocaine10AM10 + SalineAM10 + Cocaine10
Vehicle + SalineVehicle + Cocaine10AM30 + SalineAM30 + Cocaine10
Veh + Veh + SalVeh + Veh + Coc10SR0.02 + Veh + SalSR0.02 + Veh + Coc10Veh + AM30 + SalVeh + AM30 + Coc10SR0.02 + AM30 + SalSR0.02 + AM30 + Coc10
Key: panels (c), (f)
Key: panels (b), (e)
Key: panels (a), (d)
Figure 9. Effects of AM-404 (10 and 30 mg/kg) and cocaine (10 mg) on locomotor activity and effect of SR141716A (0.02 mg/kg)
on the AM-404 (30 mg)-induced reduction of cocaine’s hyperlocomotion. Histograms represent the photocell disruptions caused
by the animals’ locomotion (a–c) and rearing (d–f) (mean¡S.E.M.). The asterisks (*) show a statistically significant effect
compared to the vehicle group: * p<0.05, ** p<0.01, *** p<0.001, # p<0.05, ## p<0.01, ### p<0.001 compared to AM10 or
AM30+Sal (a, b, d, e), or compared to all other groups (c, f) ; + p<0.05, compared to the AM10+Coc10 group; +++ p<0.001
compared to the Veh+Coc10 (e) or all the cocaine groups (c, f).
918 S. Vlachou et al.

reduced by WIN55,212-2 (1 mg/kg) and AM-404 (10,
30 mg/kg, respectively). AM-404 (10 mg/kg) did not
affect the hyperolocomotor effects of cocaine (10 mg/
kg), although it did reduce cocaine’s stimulatory ac-
tion on rearing. A higher dose of AM-404 (30 mg/kg)
was found to reduce cocaine (10 mg/kg)-induced hy-
peractivity. SR141716A (0.02 mg/kg) blocked the ef-
fects of WIN55,212-2 and AM-404 on spontaneous
motor activity and on the hyperlocomotion induced by
cocaine, while it had no effect on motor function per
se. The above finding shows that WIN55,212-2 and
probably AM-404 exhibited their actions through
CB1R stimulation. Ferrari and colleagues have also
shown that HU-210 reduces the increased locomotion
and rearing caused by cocaine (Ferrari et al., 1999), as
it was also recently shown for morphine and alcohol
(Hagues et al., 2007). Similarly, Muschamp and Siviy
(2002) have shown that acute administration of
WIN55,212-2 (1 mg/kg), in Lewis rats inhibits the
hyperlocomotion and increased rearing induced by
cocaine. Our results also agree with other studies
involving THC and its counteracting action on am-
phetamine-induced hyperlocomotion (Gorriti et al.,
1999). Importantly, in a very recent study Rodvelt and
colleagues (2007) have shown that WIN55,212-2 at-
tenuates nicotine-induced hyperactivity. It is probable
that WIN55,212-2 and AM-404 alter the hyperactivity
produced by cocaine through an indirect mechanism,
such as alteration in endocannabinoid tone of motor
circuits. A possible mechanism could involve a nega-
tive feedback mechanism induced by cannabinoids in
the striatum, as mentioned above (see Centonze et al.,
2004, 2007 ; Giuffrida et al., 1999). A second alternative
mechanism of action of at least WIN55,212-2 could be
the inhibition of dopamine transporter, which has
been shown in the case of WIN55,212-2 resulting in
competition with cocaine that also inhibits the dop-
amine transporter (Steffens and Feuerstein, 2004).
In conclusion, acute systemic administration of the
direct CB1R agonist WIN55,212-2 and the endo-
cannabinoid neurotransmission enhancer AM-404,
even at high doses, did not have reinforcing effects,
but rather reversed the reward-facilitating effects of
cocaine in the ICSS paradigm. Furthermore, the same
compounds either did not affect, or (at high doses)
decreased locomotor activity. Interestingly, both
WIN55,212-2 and AM-404 in doses that did not affect
spontaneous motor activity reduced the hyperloco-
motion induced by cocaine. These effects are mediated
through CB1R stimulation, since they were reversed by
pretreatment with the CB1R antagonist SR141716A.
Therefore, cannabinoids clearly play a modulatory
role in cocaine’s psychostimulant actions. Based upon
the present findings, we speculate that drugs increas-
ing the cannabinergic tone could be further tested as a
potential treatment for various aspects of psycho-
stimulant addiction and psychomotor excitability. In
this regard, it is worth-noting that Tzavara et al. (2006)
have recently shown that endocannabinoids acting
through vanilloid1 receptors reduce hyperdopamin-
ergia-related hyperactivity in a relevant genetic animal
model. Thus, endocannabinoid neurotransmission
modulators appear to offer the potential of a dual
receptor action through both CB1R and non-CB1R
counteracting overt psychomotor stimulation without
affecting basal function and with seemingly low
overall psychotropic liability.
Acknowledgements
This study was supported by a grant from the
Research Committee (KA 2303) and the Department
of Psychology of the University of Crete. Styliani
Vlachou was supported by a scholarship from Pro-
pondis Foundation.
Statement of Interest
None.
References
BeltramoM, Stella N, Calignano A, Lin SY, Makriyannis A,
Piomelli D (1997). Functional role of high-affinity
anandamide transport, as revealed by selective inhibition.
Science 277, 1094–1097.
Beninger RJ (1983). The role of dopamine in locomotor
activity and learning. Brain Research 287, 173–196.
Bortolato M, Campolongo P, Mangieri RA, Scattoni ML,
Frau R, Trezza V, La Rana G, Russo R, Calignano A,
Gessa GL, Cuomo V, Piomelli D (2006). Anxiolytic-like
properties of the anandamide transport inhibitor AM404.
Neuropsychopharmacology 31, 2652–2659.
Braida D, Pozzi M, Cavallini R, Sala M (2001). Conditioned
place preference induced by the cannabinoid agonist CP
55,940 : interaction with the opioid system. Neuroscience
104, 923–926.
Braida D, Sala M (2002). Role of the endocannabinoid system
in MDMA intracerebral self-administration in rats.
British Journal of Pharmacology 136, 1089–1092.
Braida D, Iosue S, Pegorini S, Sala M (2005). 3,4-
methylenedioxymethamphetamine-induced conditioned
place preference (CPP) is mediated by endocannabinoid
system. Pharmacological Research 51, 177–182.
Cadoni C, Pisanu A, Solinas M, Acquas E, Di Chiara G
(2001). Behavioural sensitization after repeated exposure to
delta 9-tetrahydrocannabinol and cross-sensitization with
morphine. Psychopharmacology 158, 259–266.
CB1R stimulation counteracts cocaine’s behavioural actions 919

Caille S, Alvarez-Jaimes L, Polis I, Stouffer DG, Parsons LH
(2007). Specific alterations of extracellular
endocannabinoid levels in the nucleus accumbens by
ethanol, heroin, and cocaine self-administration. Journal of
Neuroscience 27, 3695–3702.
Centonze D, Battista N, Rossi S, Mercuri NB, Finazzi-Agro
A, Bernardi G, Calabresi P, Maccarrone M (2004). A
critical interaction between dopamine D2 receptors and
endocannabinoids mediates the effects of cocaine on
striatal gabaergic transmission. Neuropsychopharmacology
29, 1488–1497.
Centonze D, Rossi S, De Chiara V, Prosperetti C, Battista N,
Bernardi G, Mercuri NB, Usiello A, MaccarroneM (2007).
Chronic cocaine sensitizes striatal GABAergic synapses to
the stimulation of cannabinoid CB1 receptors. European
Journal of Neuroscience 25, 1631–1640.
Chaperon F, Soubrie P, Puech AJ, Thiebot MH (1998).
Involvement of central cannabinoid (CB1) receptors in
the establishment of place conditioning in rats.
Psychopharmacology 135, 324–332.
Chaperon F, Thiebot MH (1999). Behavioral effects of
cannabinoid agents in animals. Critical Reviews in
Neurobiology 13, 243–281.
Cippitelli A, Bilbao A, Gorriti MA, Navarro M, Massi M,
Piomelli D, Ciccocioppo R, Rordiguez de Fonseca F
(2007). The anandamide transport inhibitor AM404
reduces ethanol self-administration. European Journal of
Neuroscience 26, 476–486.
Compton DR, Martin BR (1997). The effect of the enzyme
inhibitor phenylmethylsulfonyl fluoride on the
pharmacological effect of anandamide in the mouse model
of cannabimimetic activity. Journal of Pharmacology and
Experimental Therapeutics 283, 1138–1143.
Cossu G, Ledent C, Fattore L, Imperato A, Bohme GA,
Parmentier M, Fratta W (2001). Cannabinoid CB1
receptor knock out mice fail to self-administer morphine
but not other drugs of abuse. Behavioural Brain Research 118,
61–65.
Coulombe D, Miliaressis E (1987). Fitting intracranial
self-stimulation data with growth models. Behavioral
Neuroscience 101, 209–214.
Darmani NA (2001). The cannabinoid CB1 receptor
antagonist SR141716A reverses the antiemetic and motor
depressant actions of WIN 55,212-2. European Journal of
Pharmacology 430, 49–58.
de Lago E, Ligresti A, Ortar G, Morera E, Cabranes A,
Pryce G, Bifulco M, Baker D, Fernandez-Ruiz J,
Di Marzo V (2004). In vivo pharmacological actions of
two novel inhibitors of anandamide cellular uptake.
European Journal of Pharmacology 484, 249–257.
De Vries TJ, Shaham Y, Homberg JR, Crombag H,
Schuurman K, Dieben J, Vanderschuren LJ,
Schoffelmeer AN (2001). A cannabinoid mechanism
in relapse to cocaine seeking. Nature Medicine 7,
1151–1154.
Del Arco I, Navarro M, Bilbao A, Ferrer B, Piomelli D,
Rodriguez de Fonseca F (2002). Attenuation of
spontaneous opiate withdrawal in mice by the
anandamide transport inhibitor AM404. European Journal
of Pharmacology 454, 103–104.
Dickason-Chesterfield AK, Kidd SR,Moore SA, Schaus JM,
Liu B, Nomikos GG, Felder CC (2006). Pharmacological
characterization of endocannabinoid transport and fatty
acid amide hydrolase inhibitors. Cellular and Molecular
Neurobiology 26, 407–423.
Drews E, Schneider M, Koch M (2005). Effects of the
cannabinoid receptor agonist WIN 55,212-2 on operant
behavior and locomotor activity in rats. Pharmacology,
Biochemistry and Behaviour 80, 145–150.
Fattore L, Martellotta MC, Cossu G, Mascia MS, Fratta W
(1999). CB1 cannabinoid receptor agonist WIN 55,212-2
decreases intravenous cocaine self-administration in rats.
Behavioural Brain Research 104, 141–146.
Ferrari F, Ottani A, Giuliani D (1999). Influence of the
cannabinoid agonist HU-210 on cocaine and
CQP 201-403-induced behavioral effects in rat. Life Sciences
65, 823–831.
Filip M, Golda A, Zaniewska M, McCreary AC, Nowak E,
Kolasiewicz W, Przegalinski E (2006). Involvement of
cannabinoid CB1 receptors in drug addiction : effects of
rimonabant on behavioral responses induced by cocaine.
Pharmacological Reports 58, 806–819.
Foltin RW, Fischman MW, Pippen PA, Kelly TH (1993).
Behavioral effects of cocaine alone and in combination with
ethanol or marijuana in humans. Drug and Alcohol
Dependence 32, 93–106.
Fung YK, Lau YS (1988). Receptor mechanisms of
nicotine-induced locomotor hyperactivity in chronic
nicotine-treated rats. European Journal of Pharmacology 152,
263–271.
Gallate JE, Sharov T, Mallet PE, McGregor IS (1999).
Increased motivation for beer in rats following
administration of a cannabinoid CB1 receptor agonist.
European Journal of Pharmacology 370, 233–240.
Ghozland S, Matthes HWD, Simonin F, Filliol D,
Kieffer BL, Maldonado R (2002). Motivational effects of
cannabinoids are mediated by m-opioid and k-opioid
receptors. Journal of Neuroscience 22, 1146–1154.
Giuffrida A, Parsons LH, Kerr TM, Rodriguez de Fonseca F,
Navarro M, Piomelli D (1999). Dopamine activation of
endogenous cannabinoid signaling in dorsal striatum.
Nature Neuroscience 2, 358–363.
Gobbi G, Bambico FR, Mangieri R, Bortolato M,
Campolongo P, Solinas M, Cassano T, Morgese MG,
Debonnel G, Duranti A, et al. (2005). Antidepressant-like
activity and modulation of brain monoaminergic
transmission by blockade of anandamide hydrolysis.
Proceedings of the National Academy of Sciences USA 102,
18620–18625.
Gonzalez S, Cascio MG, Fernandez-Ruiz J, Fezza F, Di
Marzo V, Ramos JA (2002a). Changes in endocannabinoid
contents in the brain of rats chronically exposed to
nicotine, ethanol or cocaine. Brain Research 954, 73–81.
Gonzalez S, Fernandez-Ruiz J, Sparpaglione V, Parolaro D,
Ramos JA (2002b). Chronic exposure to morphine, cocaine
or ethanol in rats produced different effects in brain
920 S. Vlachou et al.

cannabinoid CB1 receptor binding and mRNA levels. Drug
and Alcohol Dependence 66, 77–84.
Gonzalez-Cuevas G, Aujla H, Martin-Fardon R,
Lopez-Moreno JA, Navarro M, Weiss F (2007). Subchronic
cannabinoid agonist (WIN 55,212-2) treatment during
cocaine abstinence alters subsequent cocaine seeking
behavior. Neuropsychopharmacology 32, 2260–2266.
Gorriti MA, Rodriguez de Fonseca F, Navarro M, Palomo T
(1999). Chronic (-)D9-tetrahydrocannabinol treatment
induces sensitization to the psychomotor effects of
amphetamine in rats. European Journal of Pharmacology 365,
133–142.
Gueudet C, Santucci V, Rinaldi-Carmona M, Soubrie P,
Le Fur G (1995). The CB1 cannabinoid receptor antagonist
SR141716A affects A9 dopamine neuronal activity in the
rats. Neuroreport 6, 1421–1425.
Hagues G, Costentin J, Duterte-Boucher D (2007).
Modulation of morphine and alcohol motor stimulant
effects by cannabinoid receptors ligands. Behavioural Brain
Research 178, 274–282.
Hansson AC, Bermudez-Silva FJ, Malinen H, Hyytia P,
Sanchez-Vera I, Rimondini R, Rodriguez de Fonseca F,
Kunos G, Sommer WH, Heilig M (2007). Genetic
impairment of frontocortical endocannabinoid
degradation and high alcohol preference.
Neuropsychopharmacology 32, 117–126.
Hermann H, Marsicano G, Lutz B (2002). Coexpression of
the cannabinoid receptor type 1 with dopamine and
serotonin receptors in distinct neuronal subpopulations
of the adult mouse forebrain. Neuroscience 109, 451–480.
Higuera-Matas A, Soto-Montenegro ML, del Ormo N,
Miguens M, Torres I, Vaquero JJ, Sanchez J,
Garcia-Lecumberri C, Desco M, Ambrosio E (2007).
Augmented acquisition of cocaine self-administration and
altered brain glucose metabolism in adult female but not
male rats exposed to a cannabinoid agonist during
adolescence. Neuropsychopharmacology. Published online : 6
June 2007. doi:10.1038/sj.npp.1301467.
Huestis MA, Gorelick DA, Heishman SJ, Preston KL,
Nelson RA, Moolchan ET, Frank RA (2001). Blockade of
effects of smoked marijuana by the CB1-selective
cannabinoid receptor antagonist SR141716. Archives of
Genetics in Psychiatry 58, 322–328.
Iwamoto ET (1984). An assessment of the spontaneous
activity of rats administered morphine, phencyclidine,
or nicotine using automated and observational methods.
Psychopharmacology 84, 374–382.
Justinova Z, Solinas M, Tanda G, Redhi GH, Goldberg SR
(2005). The endogenous cannabinoid anandamide and its
synthetic analog R(+)-methanandamide are intravenously
self-administered by squirrel monkeys. Journal of
Neuroscience 25, 5645–5650.
Kathuria S, Gaetani S, Fegley D, Valino F, Duranti A,
Tontini A, Mor M, Tarzia G, La Rana G, Calignano A,
et al. (2003). Modulation of anxiety through blockade
of anandamide hydrolysis. Nature Medicine 9, 76–81.
Lamarque S, Tghzouti K, Simon H (2001). Chronic treatment
with Delta (9)-tetrahydrocannabinol enhances the
locomotor response to amphetamine and heroin.
Implications for vulnerability to drug addiction.
Neuropharmacology 41, 118–129.
Lesscher HMB, Hoogveld E, Burbach JPH, van Ree JM,
Gerrits MAFM (2005). Endogenous cannabinoids are not
involved in cocaine reinforcement and development of
cocaine-induced behavioural sensitization. European
Neuropsychopharmacology 15, 31–37.
Lukas SE, Sholar M, Kouri E, Fukuzako H, Mendelson JH
(1994). Marijuana smoking increases plasma cocaine
levels and subjective reports of euphoria in male
volunteers. Pharmacology, Biochemistry and Behaviour 48,
715–721.
Lupica CR, Riegel AC (2005). Endocannabinoid release from
midbrain dopamine neurons : A potential substrate for
cannabinoid receptor antagonist treatment of addiction.
Neuropharmacology 48, 1105–1116.
Madsen MV, Peacock L, Verge T, Andersen MB (2006).
Effects of the cannabinoid CB1 receptor agonist CP 55,940
and antagonist SR141716A on d-amphetamine-induced
behaviours in Cebus monkeys. Journal of
Psychopharmacology 20, 622–628.
Maldonado-Irrizary CS, Stellar JR, Kelley AE (1994). Effects
of cocaine and GBR-12909 on brain stimulation reward.
Pharmacology, Biochemistry and Behaviour 48, 915–920.
Mallet PE, Beninger RJ (1998). D9-tetrahydrocannabinol,
but not the endogenous cannabinoid receptor ligand
anandamide, produces conditioned place avoidance. Life
Sciences 62, 2431–2439.
Maneuf YP, Croosman AR, Brotchie JM (1997). The
cannabinoid receptor agonist WIN 55,212-2 reduces D2,
but not D1, dopamine receptor-mediated alleviation of
akinesia in the reserpine-treated rat model of Parkinson’s
disease. Experimental Neurology 148, 265–270.
Martin M, Ledent C, Parmentier M, Maldonado R,
Valverde O (2000). Cocaine, but not morphine, induces
conditioned place preference and sensitization to
locomotor responses in CB1 knock out mice. European
Journal of Neuroscience 12, 4038–4046.
Matthews K, Baldo BA, Markou A, LownO, Overstreet DH,
Koob GF (1996). Rewarding electrical brain stimulation:
similar thresholds for flinders sensitive line
hypercholinergic and flinders resistant line
hypocholinergic rats. Physiology and Behaviour 59,
1155–1162.
McGregor IS, Issakidis CN, Prior G (1996). Aversive
effects of the synthetic cannabinoid CP 55,940 in
rats. Pharmacology, Biochemistry and Behaviour 53,
657–664.
Meschler JP, Howlett AC, Madras BK (2001). Cannabinoid
receptor agonist and antagonist effects onmotor function in
normal and 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine
(MPTP)-induced non-human primates. Psychopharmacology
156, 79–85.
Muschamp JW, Siviy SM (2002). Behavioral sensitization to
amphetamine follows chronic administration of the CB1
agonist WIN 55,212-2 in Lewis rats. Pharmacology,
Biochemistry and Behaviour 73, 835–842.
CB1R stimulation counteracts cocaine’s behavioural actions 921

Navarro M, Carrera MR, Fratta W, Valverde O, Cossu G,
Fattore L, Chowen JA, Gomez R, del Arco I, VillanuaMA,
et al. (2001). Functional interaction between opioid and
cannabinoid receptors in drug self-administration. Journal
of Neuroscience 21, 5344–5350.
Norton S (1973). Amphetamine as a model for hyperactivity
in the rat. Physiology and Behavior 11, 181–186.
Pacher P, Batkai S, Kunos G (2006). The endocannabinoid
system as an emerging target of pharmacotherapy.
Pharmacological Reviews 58, 389–462.
Panagis G, Spyraki C (1996). Neuropharmacological
evidence for the role of dopamine in ventral pallidum
self-stimulation. Psychopharmacology 123, 280–288.
Panagis G, Kastellakis A, Spyraki C, Nomikos G (2000).
Effects of methyllycaconitine (MLA), an a7 nicotinic
receptor antagonist, on nicotine- and cocaine-induced
potentiation of brain stimulation reward.
Psychopharmacology 149, 388–396.
Panlilio LV, Solinas M, Matthews SA, Goldberg SR (2007).
Previous exposure to THC alters the reinforcing efficacy
and anxiety-related effects of cocaine in rats.
Neuropsychopharmacology 32, 646–657.
Parker LA, Burton P, Sorge RE, Yakiwchuk C, Mechoulam
R (2004). Effect of low doses of Delta(9)-
tetrahydrocannabinol and cannabidiol on the extinction
of cocaine-induced and amphetamine-induced
conditioned place preference learning in rats.
Psychopharmacology 175, 360–366.
Paxinos G, Watson C (1998). The Rat Brain in Stereotaxic
Coordinates, 4th edn. San Diego : Academic Press.
Pontieri FE, Monnazzi P, Scontrini A, Buttarelli FR,
Patacchioli FR (2001a). Behavioral sensitization to heroin
by cannabinoid pretreatment in the rat. European Journal of
Pharmacology 421, R1–R3.
Pontieri FE, Monnazzi P, Scontrini A, Buttarelli FR,
Patacchioli FR (2001b). Behavioral sensitization to WIN
55,212-2 in rats pretreated with heroin. Brain Research 898,
178–180.
Przegalinski E, Gothert M, Frankowska M, Filip M (2005).
WIN 55,212-2-induced reduction of cocaine
hyperlocomotion : Possible inhibition of 5-HT3 receptor
function. European Journal of Pharmacology 517, 68–73.
Ranaldi R, Bauco P, Wise R (1997). Synergistic effects of
cocaine and dizocilpine (MK-801) on brain stimulation
reward. Brain Research 760, 231–237.
Rinaldi-Carmona M, Barth F, Heaulme M, Shire D,
Calandra B, Congy C, Martinez S, Maruani J, Neliat G,
Caput D, et al. (1994). SR141716A, a potent and selective
antagonist of the brain cannabinoid receptor. FEBS Letters
350, 240–244.
Rodriguez de Fonseca F, Del Arco I, Bermudez-Silva FJ,
Bilbao A, Cippitelli A, Navarro M (2005). The
endocannabinoid system: physiology and pharmacology.
Alcohol and Alcoholism 40, 2–14.
Rodriguez de Fonseca F, Del Arco I, Martin-Calderon JL,
Gorriti MA, Navarro M (1998). Role of the endogenous
cannabinoid system in the regulation of motor activity.
Neurobiology and Disease 5, 483–501.
Rodvelt KR, Bumgarner DM, Putnam WC, Miller DK
(2007). WIN-55,212-2 and SR-141716A alter nicotine-
induced changes in locomotor activity, but do not alter
nicotine-evoked [3H]dopamine release. Life Sciences 80,
337–344.
Romero J, Lastres-Becker I, De Miguel R, Berrendero F,
Ramos JA, Fernandez-Ruiz J (2002). The endogenous
cannabinoid system and the basal ganglia : biochemical,
pharmacological and therapeutics aspects. Pharmacology
and Therapeutics 95, 137–152.
Sanudo-Pena MC, Romero J, Seale GE, Fernandez-Ruiz JJ,
Walker JM (2000). Activational role of cannabinoids
on movement. European Journal of Pharmacology 391,
269–274.
Sanudo-Pena MC, Tsou K, Walker JM (1999). Motor actions
of cannabinoids in the basal ganglia output nuclei. Life
Sciences 65, 703–713.
Schenk S, Partridge B (1999). Cocaine-seeking produced by
experimenter-administered drug injections : dose-effect
relationships in rats. Psychopharmacology 147, 285–290.
Singh ME, McGregor IS, Mallet PE (2005). Repeated
exposure to D9-tetrahydrocannabinol alters
heroin-induced locomotor sensitization and
Fos-immunoreactivity. Neuropharmacology 49, 1189–1200.
Solinas M, Panlilio LV, Tanda G, Makriyannis A,
Matthews SA, Goldberg SR (2005). Cannabinoid agonists
but not inhibitors of endogenous cannabinoid transport or
metabolism enhance the reinforcing efficacy of heroin in
rats. Neuropsychopharmacology 30, 2046–2057.
Soria G, Mendizabal V, Tourino C, Robledo P, Ledent C,
Parmentier M, Maldonado R, Valverde O (2005).
Lack of CB1 cannabinoid receptor impairs cocaine
self-administration. Neuropsychopharmacology 30,
1670–1680.
Steffens M, Feuerstein TJ (2004). Receptor-independent
depression of DA and 5-HT uptake by cannabinoids in rat
neocortex–involvement of Na(+)/K(+)-ATPase.
Neurochemistry International 44, 529–538.
Sulcova E, Mechoulam R, Fride E (1998). Biphasic effects of
anandamide. Pharmacology Biochemistry and Behavior 59,
347–352.
Tanda G (2007). Modulation of the endocannabinoid system:
therapeutic potential against cocaine dependence.
Pharmacological Research 56, 406–417.
Tzavara ET, Li ET, Moutsimilli L, Bisogno T, Di Marzo V,
Phebus LA, Nomikos GG, Giros B (2006).
Endocannabinoids activate transient receptor potential
vanilloid 1 receptors to reduce hyperdopaminergia-related
hyperactivity : therapeutic implications. Biological
Psychiatry 59, 508–515.
Tzavara ET, Wade M, Nomikos GG (2003). Biphasic effects
of cannabinoids on acetylcholine release in the
hippocampus: site and mechanism of action. Journal of
Neuroscience 23, 9374–9384.
Valjent E, Mitchell JM, Besson M, Caboche J, Maldonado R
(2002). Behavioural and biochemical evidence for
interactions between D9-tetrahydrocannabinol and
nicotine. British Journal of Pharmacology 135, 564–578.
922 S. Vlachou et al.

Vigano D, Valenti M, Cascio MG, Di Marzo V, Parolaro D,
Rubino T (2004). Changes in endocannabinoid levels in a
rat model of behavioural sensitization to morphine.
European Journal of Neuroscience 20, 1849–1857.
Vinklerova J, Novakova J, Sulcova A (2002). Inhibition of
methamphetamine self-administration in rats by
cannabinoid receptor antagonist AM 251. Journal of
Psychopharmacology 16, 139–143.
Vlachou S, Nomikos GG, Panagis G (2003). WIN 55,212-2
decreases the reinforcing actions of cocaine through CB1
cannabinoid receptor stimulation. Behavioural Brain
Research 141, 215–222.
Vlachou S, Nomikos GG, Panagis G (2005). CB1
cannabinoid receptor agonists increase intracranial
self-stimulation thresholds in the rat. Psychopharmacology
179, 498–508.
Vlachou S, Nomikos GG, Panagis G (2006). Effects of
endocannabinoid neurotransmission modulators
on brain stimulation reward. Psychopharmacology 188,
293–305.
Vlachou S, Nomikos GG, Stephens DN, Panagis G
(2007). Lack of evidence for appetitive effects of
D9-tetrahydrocannabinol in the intracranial
self-stimulation and conditioned place preference
procedures in rodents. Behavioural Pharmacology 18,
311–319.
Xi ZX, Spiller K, Pak AC, Gilbert J, Dillon C, Li X, Peng XQ,
Gardner EL (2007). Cannabinoid CB1 receptor antagonists
attenuate cocaine’s rewarding effects : experiments with
self-administration and brain-stimulation reward in rats.
Neuropsychopharmacology. Published online : 29 August
2007. doi: 10.1038/sj.npp.1301552.
CB1R stimulation counteracts cocaine’s behavioural actions 923
Top Related
Copyright © 2022 FDOKUMEN

