







Bahasa
Halaman
Hukum


Marine Chemistry 121 (2010) 132–144
Contents lists available at ScienceDirect
Marine Chemistry
j ourna l homepage: www.e lsev ie r.com/ locate /marchem
Comparative evaluation of sediment trap and 234Th-derived POC fluxes from theupper oligotrophic waters of the Gulf of Mexico and the subtropical northwesternPacific Ocean
Chin-Chang Hung a,b, Chen Xu a, Peter H. Santschi a,⁎, Sai-Jin Zhang a, Kathleen A. Schwehr a,Antonietta Quigg a, Laodong Guo c, Gwo-Ching Gong b, James L. Pinckney d, Richard A. Long e, Ching-LingWei f
a Department of Oceanography and Marine Sciences, Texas A&M University, 5007 Ave. U, Galveston, TX 77551, USAb Institute of Marine Environmental Chemistry and Ecology, National Taiwan Ocean University, Keelung, 20224, Taiwanc Department of Marine Science, University of Southern Mississippi, 1020 Balch Blvd., Stennis Space Center, MS 39529, USAd Estuarine Ecology Lab, Marine Science Program and Department of Biological Sciences, University of South Carolina, Columbia, SC 29208, USAe Department of Biological Sciences and Marine Science Program, University of South Carolina, Columbia, SC 29208, USAf Institute of Oceanography, National Taiwan University, Taipei, 10617, Taiwan
⁎ Corresponding author.E-mail address: [email protected] (P.H. Santschi)
0304-4203/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.marchem.2010.03.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 June 2009Received in revised form 23 March 2010Accepted 23 March 2010Available online 1 April 2010
Keywords:POC flux234Th/POC ratiosSediment trapsHaptophytesDiatomsGulf of MexicoEast China SeaNorthwestern Pacific Ocean
To better understand the inter-relationships between particulate organic carbon (POC) fluxes andphytoplankton and bacteria biomass and production, we compared POC fluxes determined in sediment trapsand approaches based on size-fractionated (1–10, 10–50, 50–150 and N150 μm) suspended particulate 234Thand POC concentrations in oligotrophic sections of the Gulf ofMexico during August 2005 andMay 2006 and inthe oligotrophic northwestern Pacific Ocean during 2009. In 2005, the sediment trap POC flux near the bottomof the euphotic zone (120 m) ranged from 71 to 94 mg C m−2 day−1, while 234Th-derived POC fluxes usingPOC/234Th ratios in the 10–50 µm and 50–150 µm varied from 71 to 150 mg C m−2 day−1. In 2006, thesediment trap POC flux at 120 m ranged from 24 to 67 mg C m−2 day−1, while the 234Th-derived POC fluxes inthe 10–50 µm fraction were comparable or somewhat higher, ranging from 71 to 119 mg C m−2 day−1. ThePOC fluxes in 2006, calculated by using POC/234Th ratios in the 1–10 µm and the 50–150 µm fractions weremuch higher, ranging from 847 to 1369 mg C m−2 day−1. Correlations with biological and chemicalparameters support a likely mechanism of sinking aggregates of haptophytes (0.2–20 µm) of higher densityheld together by Th-complexing and uronic acid containing exopolymeric substances. The observations that234Th (and POC) is mainly associated with medium-sized (10–15 µm) suspended particles rather than larger(50–150 µm) ones may be caused by the use of a one-filter method and standard filtration and processingprocedures that were applied here for collecting suspended particles. This then raises the question of whatconstitutes representative material from the ocean that settles on the characteristic time scale of 234Th. As acomparison, size-fractionated trap-collected particles in the oligotrophic northwestern Pacific Ocean showedthat the 10–50 μm fraction contained the largest proportion of POC (22–41%), followed by the 50–150 μm(22–37%), the N150 μm (15–27%), and the 1–10 μm (17–23%) fraction. The partitioning of 234Th in trap-collectedparticles was slightly different from that of POC, with the 1–10 μm fraction representing the largest proportion(27–48%) of 234Th flux. Together, the b50 μmparticles contributed, on average, 52±6% of POC, which suggeststhat the POC/234Th ratios traditionally derived from large (N50 µm) pump-collected particles may notaccurately reflect themajority of sinking particles. Therefore, estimated POC fluxesmay be significantly biasedusing a conventional 234Th based approach, i.e., using POC/234Th ratios from a single filter obtained from large(N50 µm) pump-collected particles.
.
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The “biological pump”, i.e., the removal of CO2 from the surfaceocean via formation and sinking of biological remains, determined as
particulate organic carbon (POC), plays a crucial role in the globalcycling of carbon, nutrient and particle reactive elements. Therefore,an accurate estimate of POC concentrations and export fluxes from theeuphotic zone is crucial for a better understanding of the biogeo-chemical cycling of carbon in the ocean. Despite possible biases byhydrodynamic and swimmer effects, POC fluxes are often determinedby sediment traps (Gardner, 1980; Buesseler, 1991). As an alternative,234Th/238U disequilibrium has been increasingly used for estimating

133C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
POC fluxes in different marine environments (e.g., Buesseler et al.,1992a, 1995; Coale and Bruland, 1985; Cochran et al., 1995; Murray etal., 1996; Buesseler et al., 2006, and references therein). In the latterapproach, POC fluxes out of the euphotic zone are determined by theproduct of the POC/234Th ratio in sinking particles and the depth-integrated 234Th flux from the euphotic zone (Buesseler et al., 2006and references therein). For a reliable POC export flux estimate, boththe 234Th flux and POC/234Th ratio of sinking particles are required. Asfor the choice of the appropriate POC/234Th ratio, it should berepresentative of the assemblages of particles that sink over asignificant vertical distance (at least tens to hundreds of meters) onthe characteristic time scale of 234Th. However, recent studies haveshown that ratios of POC/234Th could vary with water depth, particlesize fractions and different hydrographic regimes (e.g., Cochran et al.,1995; Buesseler, 1998, Santschi et al., 1999, 2003; Moran et al., 2003;Buesseler et al., 2006). While the ratio of POC/234Th is a crucialquestion for marine biogeochemistry, our understanding of themechanisms that control the POC/234Th ratios is still evolving (Baconet al., 1996; Murray et al., 1996; Buesseler, 1998; Ducklow et al., 2001;Benitez-Nelson et al., 2001; Coppola et al., 2002; Chen et al., 2003;Santschi et al., 2003, 2006).
In most studies, filtered large particles (e.g., N53 or N70 μm,collected via large volume filtration) are assumed to be representativeof the majority of sinking particles (Buesseler et al., 1995; Bacon et al.,1996; Cochran et al., 2000) and the POC/234Th ratio of such largeparticles is often adopted to estimate the POC flux in different marineenvironments (Buesseler et al., 1995; Bacon et al., 1996; Cochran et al.,2000; Benitez-Nelson et al., 2001; Moran et al., 2003; Hung et al.,2004; Buesseler et al., 2006). If this assumption is not universallyaccepted, we must continue to examine the question: are the filteredlarge suspended particles really representative of major settlingparticles in the oligotrophic ocean, where normally pico- and nano-plankton dominate (Campbell et al., 1994; Li, 1994; Moon-van derStaay et al., 2000). Currently, there is no consensus to this question.For example, Coppola et al. (2002) compared POC/234Th ratios onsinking particles and suspended particles (N0.6 μm) in the BarentsSea. These authors concluded that the POC flux estimates weremore reliable if the POC/234Th ratios determined from sedimenttrap material were used. Hung et al. (2004) found that POC fluxesmeasured by traps and 234Th derived fluxes (using POC/234Th on largeparticles, e.g. N53 µm, with POC and 234Th measured separately ondifferent filters) in the Gulf of Mexico were comparable. However,Hung and Gong (2007) reported that the POC export fluxes out of theeuphotic zone estimated by sediment traps and 234Th approaches(using the POC/234Th ratio on medium-sized particles, e.g., the 10–50 μm) in the Kuroshio Current were in best agreement. Lalande et al.(2008) and Lepore et al. (2009) have estimated POC export by usingsediment trap and size-fractionated POC/234Th ratios on suspendedparticles and found that the estimated POC export by sediment trapsand 234Th/238U disequilibrium can each be biased, albeit, in differentways. Moreover, Richardson and Jackson (2007) reported that smallphytoplankton plays an important role in driving carbon export in thesurface ocean. Based on these so far inconsistent results, more fieldobservations are required to investigate which part of the suspendedparticle spectrum represents sinking particles, and if any discrepancyin results could be produced by the collection method, i.e., filtration.Since sinking particles in the ocean likely settle out as largeaggregates, as shown by in situ camera based methods (e.g., Santschiet al., 1999), handling of these aggregates by filtration could producealterations of the original particle sizes. For this purpose, one needs tocompare POC fluxes calculated from POC/234Th ratios of size-fractionated particles with those from sediment traps to determinewhich ones are optimal for estimating the POC flux from the euphoticzone.
Surface-tetheredfloating sediment trapsweredeployedon the samedays as size-fractionated (b1 µm, 1–10 µm, 10–50 µm, 50–150 µm
and N150 µm) particulate 234Th and POC samples were collected bylarge volume in situ pumps from two contrasting (but isolated)oceanographic settings, i.e., cold core rings (CCR) vs. warm core rings(WCR) in the Gulf of Mexico during August 2005 and May 2006.Generally, CCRs contain upwelling nutrient-rich waters with higherprimary productivity and higher organic matter fluxes. Conversely,WCRs normally contain more nutrient-depleted waters with lowerproductivity or biomass, as well as lower organic matter fluxes (Biggs,1992;Wormuthet al., 2000;Guoet al., 2002a; Santschi et al., 2003). Ringor eddy systems such as these are ideal for addressing our question on234Th and POC, as their long-term (∼1 year) isolation assures a steady-state system, thus not requiring time-series measurements (Guo et al.,2002a; Santschi et al., 2003; Hung et al., 2004). Given that Station 1waslocated in a near-coastal upwelling system and Station 2 was locatedbetween a WCR and CCR, the steady-state assumption would not bevalid for these stations, and horizontal and vertical transport processesmay be important (e.g., Savoye et al., 2006). Our investigation thusallows a comparison between sediment trap-collected POC exportfluxes and POC fluxes calculated from an assessment of 234Thdeficiencies combined with POC/234Th ratios from size-fractionatedsuspended particles or sediment trap materials. Detailed results ofaccompanying data sets on particle size distribution of 234Th, POC, aswell as biochemical and biological (i.e., phytoplankton and bacteria)composition andbiomass concentration,will be reported elsewhere (Xuet al., in press).
Furthermore, because the 234Th approach relies on POC/234Thratios determined for particles collected by large volume in situ pumpson 50 µm (or similar-sized) mesh, we therefore used a similarsequential filtration system to measure, on the same filter, theabundance of POC and 234Th concentration in size-fractionated (the1–10, 10–50, 50–150 and N150 µm) sinking particles (collected byfloating sediment traps) from the oligotrophic waters of thesubtropical northwestern Pacific in 2009 to examine what are therepresentative settling particles and their contribution to the settlingflux through the euphotic zone. Besides, we verified the particle sizedistribution of sinking particles in selected samples from thenorthwestern Pacific Ocean via scanning electron microscopy (SEM).The POC and 234Th data from trap-collected particles in the East ChinaSea and northwestern Pacific Oceanwere then comparedwith the POCand 234Th data from both pump-collected and trap-collected particles(without size-fractionation) from the Gulf of Mexico.
2. Materials and methods
2.1. Gulf of Mexico
Seawater, sediment trap-collected and pump-collected size-frac-tionated particulate matter samples from the Gulf of Mexico wereobtained aboard the R/V Pelican during August 1–14, 2005 and the R/VSeward Johnsonduring April 30 toMay 11, 2006, in the vicinity of CCRsand WCRs in the northwest Gulf of Mexico (Fig. 1). In 2005, station 1(28°59′N, 89°43′W, water depth 2000 m), with a surface watertemperature of 28.8 °C, was located within a coastal upwelling region,whereas station 2 (28°48′N, 88°29′W, water depth 1000 m) waslocated on the northern edge of a WCR, with a surface watertemperature of 29.4 °C. In 2006, the sampling locations also consistedof contrasting hydrographic regimes, with stations 3 (25°52′N, 92°31′W, water temperature at 25 m=25.0 °C, water depth 1980 m) and 5(27°51′N, 88°38′W,water temperature at 25 m=24.5 °C, water depth2076 m) inside CCRs, and station 4 (26°55′N, 90°00′W, watertemperature at 25 m=25.4 °C, water depth 2080 m) within a WCR.
Suspended particles, defined as the particles retained by filtrationof different size classes, were sequentially collected by Nitex screens(i.e., 150, 50, and 10 μm) with an in situ filtration system (Guo et al.,2002a;Hung et al., 2004). Particulatematter collected onNitex screenswas subsequently resuspended into GF/F filtered seawater by
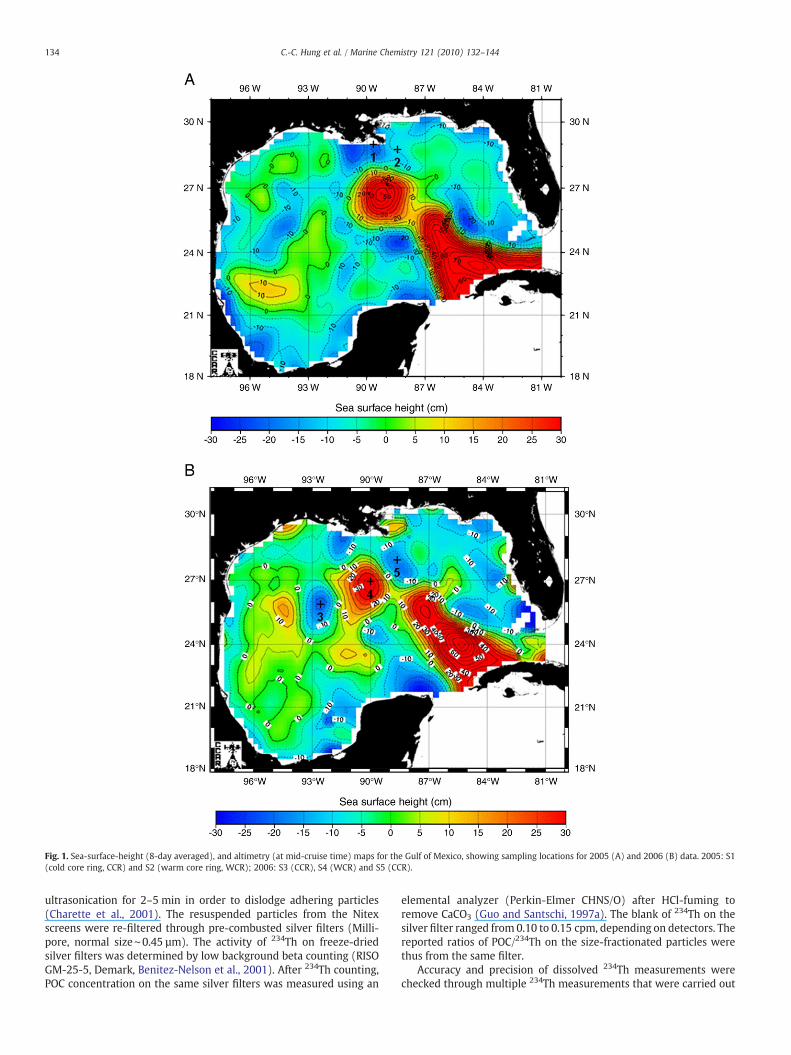
Fig. 1. Sea-surface-height (8-day averaged), and altimetry (at mid-cruise time) maps for the Gulf of Mexico, showing sampling locations for 2005 (A) and 2006 (B) data. 2005: S1(cold core ring, CCR) and S2 (warm core ring, WCR); 2006: S3 (CCR), S4 (WCR) and S5 (CCR).
134 C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
ultrasonication for 2–5 min in order to dislodge adhering particles(Charette et al., 2001). The resuspended particles from the Nitexscreens were re-filtered through pre-combusted silver filters (Milli-pore, normal size∼0.45 µm). The activity of 234Th on freeze-driedsilver filters was determined by low background beta counting (RISOGM-25-5, Demark, Benitez-Nelson et al., 2001). After 234Th counting,POC concentration on the same silver filters was measured using an
elemental analyzer (Perkin-Elmer CHNS/O) after HCl-fuming toremove CaCO3 (Guo and Santschi, 1997a). The blank of 234Th on thesilver filter ranged from 0.10 to 0.15 cpm, depending on detectors. Thereported ratios of POC/234Th on the size-fractionated particles werethus from the same filter.
Accuracy and precision of dissolved 234Th measurements werechecked through multiple 234Th measurements that were carried out

135C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
for selected samples. 234Th was calibrated using equilibrated 238U/234Th solutions for gamma counting, and comparing results with 234Thfrom these solutions by low level beta counting. Backgrounds for thedissolved 234Th measurements were determined using empty count-ing vials for gamma counting. Additionally, we have participated in thetotal and particulate Th-234 intercalibration organized by GEOTRACESprogram (Maiti et al., 2010). So far, 234Th activities determined in ourlab compare favorably to those determined in other labs.
Concentrations of dissolved 234Th were determined using proce-dures described in Guo et al. (2002a) and Hung et al. (2004), and itsaccuracy and precision recently validated by Hung et al. (2008).Briefly, large volumes of seawater (up to 1000–4000 l) were collectedby a submersible pump from the upper water column (up to 140 m)that had been successfully used in a number of previous studies(Baskaran et al., 1993; Guo et al., 1995, 1997; Santschi et al., 1999). Thesubmersible pump was equipped with three outlets: the first outletwas used to provide aliquots for POC and 234Th concentration on size-fractionated (N150 µm, 50–150 µm, 10–50 µm, and 1–10 µm) parti-cles (as depicted above). The second outlet was used to control theflow rate of seawater. The third one contained four sequentialcartridge filters (polypropylene, Sparkling Clear Industries) withdecreasing pore sizes of 150 µm, 50 µm, 10 µm, and 0.5 µm, and twoMnO2-impregnated cartridgefilters (pore size=0.5 µm). Thefirst fourconsecutive cartridge filters were designed to collect 234Th in size-fractionated suspended particles, while the last two MnO2-impreg-nated cartridge filters were intended to extract the b0.5 µm dissolved234Th (Buesseler et al., 1992b; Cochran et al., 1995). The six cartridgefilters resulted in five different size fractions of 234Th, including theN150 µm, 50–150 µm, 10–50 µm, 0.5–10 µm, and b0.5 µm beingcollected. The activity of 234Th was determined by either lowbackground beta counting (see above) or gamma counting in aCanberra ultra high purity Germanium well detector (Santschi et al.,1999; Guo et al., 2002a). Cartridge-based 234Th data on suspendedparticles were, though comparable, slightly higher than data frompump-based 234Th (Xu et al., in press).
Sinking particles, defined as the particles collected by sedimenttraps, were collected with a surface-tethered floating sediment traparray consisting of six cylindrical plastic core tubes (6.8 cmdiameter and 1:10 aspect ratio) filled with filtered seawater(Polypropylene cartridge filter, nominal size 0.5 μm, defined asunpoisoned traps) with a grid (grid spacing 1 cm×1 cm) coveringthe trap mouth. Tubes were attached to a surface buoy anddeployed for 1 to 1.3 days, at depths of 65, 90, 120 and 140 m.Before analysis, swimmers on all filters were carefully removedusing forceps under a microscope. Sinking particles were filteredthrough silver filters for the determination of 234Th and POCconcentrations (methods same as above). In 2006, POC and 234Thfluxes were corrected for the amounts lost into the overlying waterduring exposure by adding amounts of dissolved organic carbonand 234Th (determined by the small volume method, Benitez-Nelson et al., 2001) in solution to those on particles. DOC wasdetermined according to Guo and Santschi (1997b). While thiscorrection increased 234Th fluxes, on average, by a factor of 4, POCfluxes increased by about 100%.
High performance liquid chromatography (HPLC) was used todetermine chemosystematic photosynthetic pigments for phyto-plankton. Filters were lyophilized for 12 h at −50 °C, placed in 90%acetone (1 ml), sonicated, and extracted at −20 °C for 18–20 h.Filtered extracts (200 µl) were injected into a Shimadzu HPLCequipped with a monomeric (Rainin Microsorb-MV, 0.46×10 cm,3 µm) and a polymeric (Vydac 201TP54, 0.46×25 cm, 5 µm) reverse-phase C18 column in series. A nonlinear binary gradient was used forpigment separations (Pinckney et al. 1996). Absorption spectra andchromatograms (440±4 nm) were acquired using a Shimadzu SPD-M10av photodiode array detector. Pigment peaks were identified bycomparison of retention times and absorption spectra with pure
standards (DHI, Denmark). The synthetic carotenoid β-apo-8′-car-otenal was used as an internal standard. ChemTax, a matrixfactorization program, was used to calculate the absolute and relativeabundances of major algal groups from concentrations of chemosyste-matic photopigment biomarkers (Mackey et al. 1996; Wright et al.1996).While haptophytes are composed of a mixed group of algae thatare quite diverse, and include prymnesiophytes, we report onhaptophytes separately from prymnesiophytes. The carotenoid pigment(19′-butanoyloxyfucoxanthin) is found in most members of thehaptophytes but not in prymnesiophytes, such that the pigment profileallows ChemTax to separate the two groups.
Uronic acid (URA) concentrations were determined according toHung et al. (2003a,b) modified by Xu et al. (in press). Primaryproductivity (PP) was derived by an empirical SeaWiFS (or MODIS)chlorophyll–temperature algorithm (Behrenfeld and Falkowski,1997). The PP values were averaged adopting eight-day mean values,expressed as mg C m−2 day−1. Bacterial abundance in the watercolumn during the 2005 and 2006 expeditions was determined byDAPI staining (Feig and Porter, 1980). Bacterial biomass was thenestimated with a carbon conversion factor of 2×10−14 gcell−1 (Leeand Fuhrman, 1987). In 2001, bacterial biomass was determined byusing the acridine orange direct count method (Hobbie et al., 1977),and results are therefore not necessarily comparable.
2.2. Northwestern Pacific Ocean
Samples were collected aboard the R/V Ocean Researcher II in thenorthwestern Pacific Ocean on 15–17 May and 16 June 2009. Sinkingparticles were obtained by sediment traps deployed at 70 m depth atstation 9 (25°30′ N, 121°58′E, water depth 130 m) and at 120 and150 m at station 13 (25°12′ N, 121°56′E, water depth 1651 m) insurface-tethered drifting sediment trap arrays and attached to asurface buoy fitted with a global positioning system antenna (TGB-500, TAIYO, Japan). Each trap array consisted of 12 6.8-cm-diametercylindrical plastic core tubeswith honeycomb baffles covering the trapmouths (Walsh, 1990; Santschi et al., 2003; Hung et al., 2003a; Hunget al., 2009). The plastic core tubes were filled with filtered seawater(after 0.5 μm pore size and then 0.1 μm).
After recovery, the tubes containing trapped particles were placedupright for 3 h at approximately 4 °C (in a refrigerator) to allow theparticles to settle. The upper water layer in the trap tube was thenreleased through a predrilled hole, and the trapped particles in thebottom layer were immediately sequentially wet-sieved throughNitex screens into three size fractions with cut-offs of 150, 50 and10 μm. The size-fractionated (N150, 50–150, 10–50, and b10 μm) trap-collected particles were rinsed with filtered seawater (b0.5 μm poresize) and then re-filtered through pre-combusted quartz filters (QMA,pore size∼1 μm). The quartz filters were rinsed with Milli-Q waterafter filtration. Selected samples contained two quartz filters; thesecond was treated as a blank in the POC and 234Th analyses.Swimmers on the quartz filters were detected microscopically andcarefully removed using forceps. The activity of 234Th was measuredusing a low background (approximately 0.15–0.33 cpm), dependingon the beta detector (containing five detectors, RISO GM-25-5,Denmark, Benitez-Nelson et al., 2001). After 234Th counting the POCconcentration on the quartz filters was measured using an elementalanalyzer (Elementa, Germany) following HCl-fuming to remove anyCaCO3. The POC blanks ranged from 1.5 to 2.1 μmol per QMA filter. Theanalytical uncertainty (±1SD) for POC was 2–5% for duplicatemeasurements. The size-fractionated POC and 234Th fluxes werecalculated as the average of values (mg Cm−2 day−1) from the samedepth for the two individual trap arrays.
To compare POC and 234Th fluxes collected by unpoisoned traps tothose in poisoned traps, a set of sediment traps with four core tubeswas deployed in an oligotrophic setting of the northwest Pacific Ocean,similar to that in the Gulf of Mexico: two traps filled with filtered
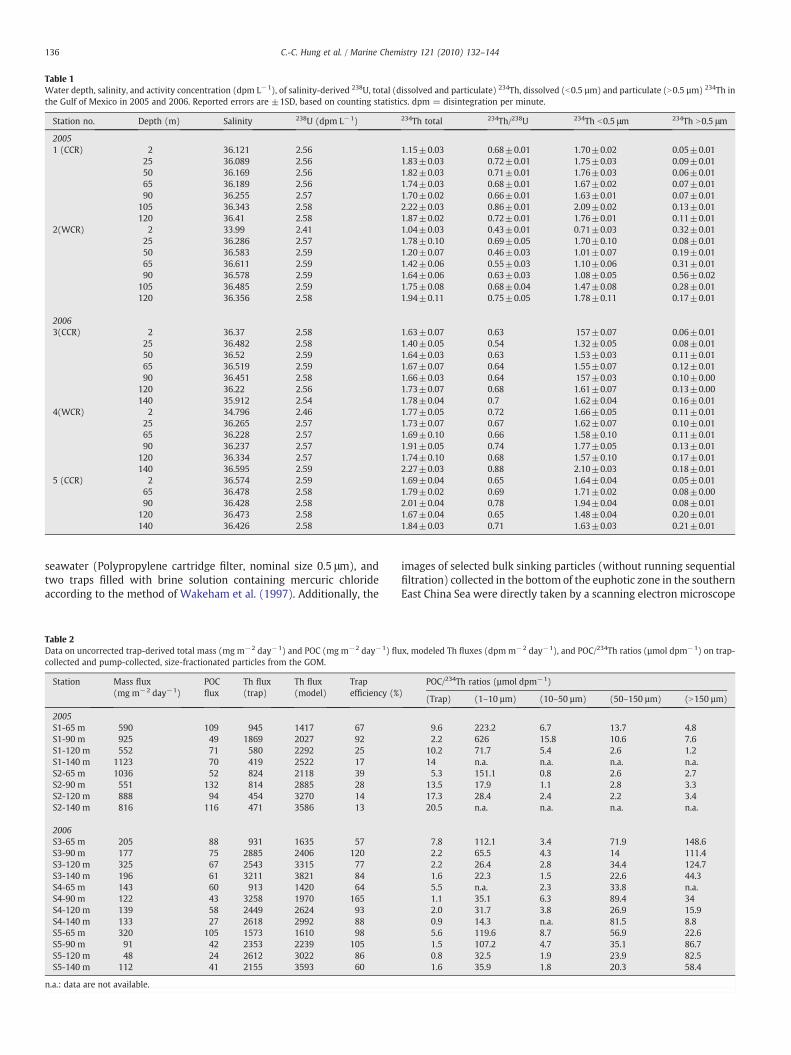
Table 1Water depth, salinity, and activity concentration (dpm L−1), of salinity-derived 238U, total (dissolved and particulate) 234Th, dissolved (b0.5 µm) and particulate (N0.5 µm) 234Th inthe Gulf of Mexico in 2005 and 2006. Reported errors are ±1SD, based on counting statistics. dpm = disintegration per minute.
Station no. Depth (m) Salinity 238U (dpm L−1) 234Th total 234Th/238U 234Th b0.5 μm 234Th N0.5 μm
20051 (CCR) 2 36.121 2.56 1.15±0.03 0.68±0.01 1.70±0.02 0.05±0.01
25 36.089 2.56 1.83±0.03 0.72±0.01 1.75±0.03 0.09±0.0150 36.169 2.56 1.82±0.03 0.71±0.01 1.76±0.03 0.06±0.0165 36.189 2.56 1.74±0.03 0.68±0.01 1.67±0.02 0.07±0.0190 36.255 2.57 1.70±0.02 0.66±0.01 1.63±0.01 0.07±0.01
105 36.343 2.58 2.22±0.03 0.86±0.01 2.09±0.02 0.13±0.01120 36.41 2.58 1.87±0.02 0.72±0.01 1.76±0.01 0.11±0.01
2(WCR) 2 33.99 2.41 1.04±0.03 0.43±0.01 0.71±0.03 0.32±0.0125 36.286 2.57 1.78±0.10 0.69±0.05 1.70±0.10 0.08±0.0150 36.583 2.59 1.20±0.07 0.46±0.03 1.01±0.07 0.19±0.0165 36.611 2.59 1.42±0.06 0.55±0.03 1.10±0.06 0.31±0.0190 36.578 2.59 1.64±0.06 0.63±0.03 1.08±0.05 0.56±0.02
105 36.485 2.59 1.75±0.08 0.68±0.04 1.47±0.08 0.28±0.01120 36.356 2.58 1.94±0.11 0.75±0.05 1.78±0.11 0.17±0.01
20063(CCR) 2 36.37 2.58 1.63±0.07 0.63 157±0.07 0.06±0.01
25 36.482 2.58 1.40±0.05 0.54 1.32±0.05 0.08±0.0150 36.52 2.59 1.64±0.03 0.63 1.53±0.03 0.11±0.0165 36.519 2.59 1.67±0.07 0.64 1.55±0.07 0.12±0.0190 36.451 2.58 1.66±0.03 0.64 157±0.03 0.10±0.00
120 36.22 2.56 1.73±0.07 0.68 1.61±0.07 0.13±0.00140 35.912 2.54 1.78±0.04 0.7 1.62±0.04 0.16±0.01
4(WCR) 2 34.796 2.46 1.77±0.05 0.72 1.66±0.05 0.11±0.0125 36.265 2.57 1.73±0.07 0.67 1.62±0.07 0.10±0.0165 36.228 2.57 1.69±0.10 0.66 1.58±0.10 0.11±0.0190 36.237 2.57 1.91±0.05 0.74 1.77±0.05 0.13±0.01
120 36.334 2.57 1.74±0.10 0.68 1.57±0.10 0.17±0.01140 36.595 2.59 2.27±0.03 0.88 2.10±0.03 0.18±0.01
5 (CCR) 2 36.574 2.59 1.69±0.04 0.65 1.64±0.04 0.05±0.0165 36.478 2.58 1.79±0.02 0.69 1.71±0.02 0.08±0.0090 36.428 2.58 2.01±0.04 0.78 1.94±0.04 0.08±0.01
120 36.473 2.58 1.67±0.04 0.65 1.48±0.04 0.20±0.01140 36.426 2.58 1.84±0.03 0.71 1.63±0.03 0.21±0.01
136 C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
seawater (Polypropylene cartridge filter, nominal size 0.5 μm), andtwo traps filled with brine solution containing mercuric chlorideaccording to the method of Wakeham et al. (1997). Additionally, the
Table 2Data on uncorrected trap-derived total mass (mg m−2 day−1) and POC (mg m−2 day−1) flucollected and pump-collected, size-fractionated particles from the GOM.
Station Mass flux(mg m−2 day−1)
POCflux
Th flux(trap)
Th flux(model)
Trapefficiency (%
2005S1-65 m 590 109 945 1417 67S1-90 m 925 49 1869 2027 92S1-120 m 552 71 580 2292 25S1-140 m 1123 70 419 2522 17S2-65 m 1036 52 824 2118 39S2-90 m 551 132 814 2885 28S2-120 m 888 94 454 3270 14S2-140 m 816 116 471 3586 13
2006S3-65 m 205 88 931 1635 57S3-90 m 177 75 2885 2406 120S3-120 m 325 67 2543 3315 77S3-140 m 196 61 3211 3821 84S4-65 m 143 60 913 1420 64S4-90 m 122 43 3258 1970 165S4-120 m 139 58 2449 2624 93S4-140 m 133 27 2618 2992 88S5-65 m 320 105 1573 1610 98S5-90 m 91 42 2353 2239 105S5-120 m 48 24 2612 3022 86S5-140 m 112 41 2155 3593 60
n.a.: data are not available.
images of selected bulk sinking particles (without running sequentialfiltration) collected in the bottom of the euphotic zone in the southernEast China Sea were directly taken by a scanning electron microscope
x, modeled Th fluxes (dpm m−2 day−1), and POC/234Th ratios (µmol dpm−1) on trap-
)POC/234Th ratios (µmol dpm−1)
(Trap) (1–10 µm) (10–50 µm) (50–150 µm) (N150 µm)
9.6 223.2 6.7 13.7 4.82.2 626 15.8 10.6 7.6
10.2 71.7 5.4 2.6 1.214 n.a. n.a. n.a. n.a.5.3 151.1 0.8 2.6 2.7
13.5 17.9 1.1 2.8 3.317.3 28.4 2.4 2.2 3.420.5 n.a. n.a. n.a. n.a.
7.8 112.1 3.4 71.9 148.62.2 65.5 4.3 14 111.42.2 26.4 2.8 34.4 124.71.6 22.3 1.5 22.6 44.35.5 n.a. 2.3 33.8 n.a.1.1 35.1 6.3 89.4 342.0 31.7 3.8 26.9 15.90.9 14.3 n.a. 81.5 8.85.6 119.6 8.7 56.9 22.61.5 107.2 4.7 35.1 86.70.8 32.5 1.9 23.9 82.51.6 35.9 1.8 20.3 58.4
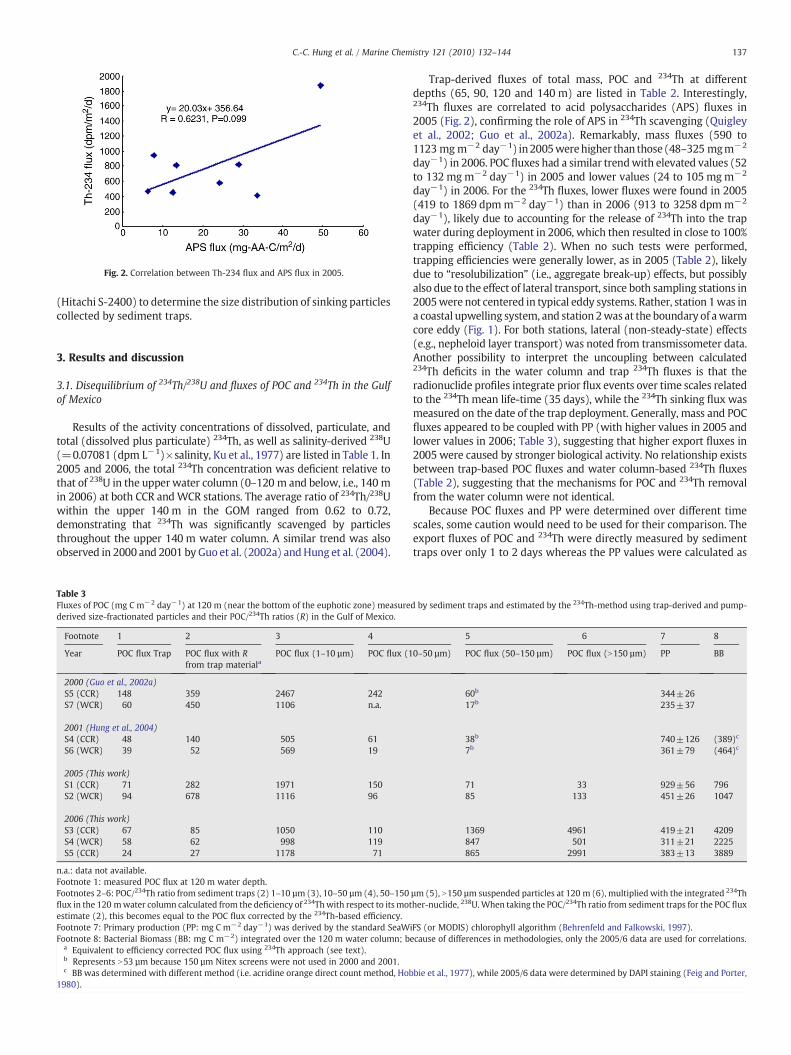
Fig. 2. Correlation between Th-234 flux and APS flux in 2005.
137C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
(Hitachi S-2400) to determine the size distribution of sinking particlescollected by sediment traps.
3. Results and discussion
3.1. Disequilibrium of 234Th/238U and fluxes of POC and 234Th in the Gulfof Mexico
Results of the activity concentrations of dissolved, particulate, andtotal (dissolved plus particulate) 234Th, as well as salinity-derived 238U(=0.07081 (dpm L−1)×salinity, Ku et al., 1977) are listed in Table 1. In2005 and 2006, the total 234Th concentration was deficient relative tothat of 238U in the upper water column (0–120 m and below, i.e., 140 min 2006) at both CCR andWCR stations. The average ratio of 234Th/238Uwithin the upper 140 m in the GOM ranged from 0.62 to 0.72,demonstrating that 234Th was significantly scavenged by particlesthroughout the upper 140 m water column. A similar trend was alsoobserved in 2000 and 2001 by Guo et al. (2002a) andHung et al. (2004).
Table 3Fluxes of POC (mg C m−2 day−1) at 120 m (near the bottom of the euphotic zone) measurederived size-fractionated particles and their POC/234Th ratios (R) in the Gulf of Mexico.
Footnote 1 2 3 4
Year POC flux Trap POC flux with Rfrom trap materiala
POC flux (1–10 µm) POC flux (1
2000 (Guo et al., 2002a)S5 (CCR) 148 359 2467 242S7 (WCR) 60 450 1106 n.a.
2001 (Hung et al., 2004)S4 (CCR) 48 140 505 61S6 (WCR) 39 52 569 19
2005 (This work)S1 (CCR) 71 282 1971 150S2 (WCR) 94 678 1116 96
2006 (This work)S3 (CCR) 67 85 1050 110S4 (WCR) 58 62 998 119S5 (CCR) 24 27 1178 71
n.a.: data not available.Footnote 1: measured POC flux at 120 m water depth.Footnotes 2–6: POC/234Th ratio from sediment traps (2) 1–10 µm (3), 10–50 µm (4), 50–150flux in the 120 mwater column calculated from the deficiency of 234Thwith respect to itsmotestimate (2), this becomes equal to the POC flux corrected by the 234Th-based efficiency.Footnote 7: Primary production (PP: mg C m−2 day−1) was derived by the standard SeaWFootnote 8: Bacterial Biomass (BB: mg C m−2) integrated over the 120 m water column; b
a Equivalent to efficiency corrected POC flux using 234Th approach (see text).b Represents N53 µm because 150 µm Nitex screens were not used in 2000 and 2001.c BB was determined with different method (i.e. acridine orange direct count method, Hob
1980).
Trap-derived fluxes of total mass, POC and 234Th at differentdepths (65, 90, 120 and 140 m) are listed in Table 2. Interestingly,234Th fluxes are correlated to acid polysaccharides (APS) fluxes in2005 (Fig. 2), confirming the role of APS in 234Th scavenging (Quigleyet al., 2002; Guo et al., 2002a). Remarkably, mass fluxes (590 to1123 mgm−2 day−1) in2005werehigher than those (48–325 mgm−2
day−1) in 2006. POC fluxes had a similar trendwith elevated values (52to 132 mgm−2 day−1) in 2005 and lower values (24 to 105 mgm−2
day−1) in 2006. For the 234Th fluxes, lower fluxes were found in 2005(419 to 1869 dpmm−2 day−1) than in 2006 (913 to 3258 dpmm−2
day−1), likely due to accounting for the release of 234Th into the trapwater during deployment in 2006, which then resulted in close to 100%trapping efficiency (Table 2). When no such tests were performed,trapping efficiencies were generally lower, as in 2005 (Table 2), likelydue to “resolubilization” (i.e., aggregate break-up) effects, but possiblyalso due to the effect of lateral transport, since both sampling stations in2005were not centered in typical eddy systems. Rather, station 1was ina coastal upwelling system, and station 2was at the boundary of awarmcore eddy (Fig. 1). For both stations, lateral (non-steady-state) effects(e.g., nepheloid layer transport) was noted from transmissometer data.Another possibility to interpret the uncoupling between calculated234Th deficits in the water column and trap 234Th fluxes is that theradionuclide profiles integrate prior flux events over time scales relatedto the 234Th mean life-time (35 days), while the 234Th sinking flux wasmeasured on the date of the trap deployment. Generally, mass and POCfluxes appeared to be coupled with PP (with higher values in 2005 andlower values in 2006; Table 3), suggesting that higher export fluxes in2005 were caused by stronger biological activity. No relationship existsbetween trap-based POC fluxes and water column-based 234Th fluxes(Table 2), suggesting that the mechanisms for POC and 234Th removalfrom the water column were not identical.
Because POC fluxes and PP were determined over different timescales, some caution would need to be used for their comparison. Theexport fluxes of POC and 234Th were directly measured by sedimenttraps over only 1 to 2 days whereas the PP values were calculated as
d by sediment traps and estimated by the 234Th-method using trap-derived and pump-
5 6 7 8
0–50 µm) POC flux (50–150 µm) POC flux (N150 µm) PP BB
60b 344±2617b 235±37
38b 740±126 (389)c
7b 361±79 (464)c
71 33 929±56 79685 133 451±26 1047
1369 4961 419±21 4209847 501 311±21 2225865 2991 383±13 3889
µm (5), N150 µm suspended particles at 120 m (6), multiplied with the integrated 234Thher-nuclide, 238U.When taking the POC/234Th ratio from sediment traps for the POC flux
iFS (or MODIS) chlorophyll algorithm (Behrenfeld and Falkowski, 1997).ecause of differences in methodologies, only the 2005/6 data are used for correlations.
bie et al., 1977), while 2005/6 data were determined by DAPI staining (Feig and Porter,
138 C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
weekly averages (∼eight-day averaged values); however, because ofnear-steady-state conditions during the 2006 study periodwithin ringsystems, this difference might not be significant within these systems,but likely would be if outside ring systems. Near-steady-stateconditions in ring systems facilitate the comparison of the trap-based POC flux approach to the 234Th-based POC flux approach. Thus,the complementary use of the 234Th approach and the POC fluxdetermination by sediment traps required further, more detailed,investigations and is discussed below.
3.2. POC flux and 234Th flux in the East China Sea and the northwesternPacific based on sediment traps
Short-term (∼a half day exposure) POC fluxes in both traps(unpoisoned and poisoned) were comparable (POC flux=187±9 mgm−2 day−1, with the 234Th flux=3826±470 dpmm−2 day−1
for unpoisoned traps, and POC flux=167±3 mgm−2 day−1, with234Th flux=3513±137 dpmm−2 day−1 for poisoned traps). The POCflux in the poisoned traps seems to be only slightly higher than that inunpoisoned traps, suggesting that POC could be consumed in theunpoisoned traps by swimmers or the release of POC into the poisonedtraps by defecating swimmers (Buesseler et al., 2007 and referencestherein). However, both 234Th fluxes lie within the measureduncertainties. Overall, these results suggest that 234Th fluxes inunpoisoned traps with short-term exposures are not significantlyinfluenced by particle degradation when the deployed time is short(hours to about 1 day).
Sinkingmaterial from some of the sediment traps wasmeasured forPOC and 234Th on the bulk material in the East China Sea whereasmaterial from the remaining traps was size-fractionated before runningPOC and 234Th analysis on the fractionatedmaterial. Comparisons of thedata from the bulk material with the respective sums of the data fromthe fractionated material suggest that the recovery of POC and 234Thduring size-fractionation was quantitative. A detailed comparison of
Table 4Fluxes of POC and 234Th via size-fractionated (N150, 50–150, 10–50, and 1–10 µm) trapped
Sta. Bottom(m)
Ze(m)
Trap(m)
Size(μm)
POC flux(mg C m−2 day
9-1 130 39 70 N150 11.5±1.650–150 27.7±1.410–50 21.3±1.11–10 14.1±0.7N1 74.5±2.4
9-2 130 41 70 N150 18.1±0.950–150 22.4±1.110–50 38.0±1.91–10 22.7±1.1N1 101.2±2.6
9-3 130 49 70 N150 44.8±2.250–150 51.7±2.610–50 91.2±1.71–10 37.7±1.9N1 225.4±4.2
13 1651 85 120 N150 12.1±1.150–150 18.7±0.910–50 19.5±1.01–10 10.3±0.5N1 60.6±1.8
13 1651 85 150 N150 13.1±0.950–150 13.5±0.710–50 10.7±0.51–10 11.0±0.5N1 48.3±1.4
Note: N1 µm=sum of N150, 50–150, 10–50, and 1–10 µm.Sta. 9-1, 9-2 and 9-3 representing sampling date on 15 May, 17 May and 16 June in 2009.Sta. 13 representing sampling date on 16 May in 2009.Ze: depth of the euphotic zone (1% of surface light intensity).
sequentialfiltrationwithnon-sequentialfiltration is reportedelsewhere(Hung et al., in press).
Fluxes of POC and 234Th in size-fractionated (1–10, 10–50, 50–150,and N150 μm) sinking particles with three individual deployments inthe upwelling region (i.e., station 9) of the southern East China Sea andin the oligotrophic ocean (station 13) of the subtropical northwesternPacific are shown in Table 4. The POC flux in bulk sinking particles in theEast China Sea and the northwest Pacific Ocean ranged from 74.5 to225.4 mgm−2 day−1 and 48–61 to 60.6 mgm−2 day−1, respectively.In comparison, our POC flux data in both upwelling regions andoligotropic waters are in agreement with reported values by Hung andGong (2007) and Hung et al. (2009, in press). Within the trap-collectedPOC pool in the East China Sea, the 10–50 μm fraction contained thelargest proportion of POC (29–41%), followed by the 50–150 μm (22–37%), the 1–10 μm(17–22%), and the N150 μm(15–20%) fraction. In thesubtropical northwestern Pacific, the distribution pattern of POC in thetrap-collected POC pool is analogous to that of POC in the East China Seaand the 10–50 μm fraction contained the largest portion of POC (22–32%), followed by the 50–150 μm fraction (28–31%), the N150 μmfraction (20–27%) and the 1–10 μm fraction. The content of 234Th intrap-collected particles was slightly different from that of POC, with the10–50 μm fraction representing the largest proportion (27–48%) of234Th flux. For the 234Th fluxes in the East China Sea, the 10–50 μmfraction had the largest values (1203–2262 dpmm−2 day−1), followedby the 1–10 μm (594–1334 dpmm−2 day−1), the 50–150 μm (376–1049 dpmm−2 day−1), and the N150 μm (156–557 dpmm−2 day−1).In the northwestern Pacific, the 10–50 μm fraction contained the largestvalues (486–775 dpmm−2 day−1), followed by the 1–10 μm (391–478 dpmm−2 day−1), the N150 μm (459–477 dpmm−2 day−1), andthe 50–150 μm (222–315 dpmm−2 day−1). The distribution pattern ofsize-fractionated POC on the trap-collected particles is slightlydifferent from that of 234Th in both the upwelling region and theoligotrophic water. However, small trap-collected particles were stillthe predominant size class, in both the POC and the 234Th, and inboth the upwelling region and the oligtrophic water. We found that
particles in the East China Sea (sta. 9) and the northwestern Pacific (sta. 13) in 2009.
−1)
234Th flux(dpm m−2 day−1)
POC/234Th(μmol dpm−1)
POC(%)
234Th(%)
156±8 6.14±0.89 15.4 6.0376±13 6.13±0.38 37.1 14.6
1203±26 1.47±0.08 28.6 46.6594±18 1.97±0.12 18.9 23.0
2583±37 2.40±0.09198±9 7.60±0.52 17.9 5.5537±16 3.48±0.20 22.1 14.8
1737±30 1.82±0.10 37.6 47.8930±23 2.03±0.11 22.4 25.6
3637±44 2.32±0.07557±22 6.71±0.43 19.9 10.7
1049±31 4.11±0.24 22.9 20.22262±46 3.36±0.09 40.5 43.51334±36 2.35±0.13 16.7 25.75201±69 3.61±0.08477±14 2.11±0.20 20.0 23.0315±10 4.94±0.30 30.8 15.2775±17 2.10±0.11 32.1 37.3391±13 2.20±0.13 17.0 18.8
2078±28 2.43±0.08459±14 2.38±0.18 27.1 25.7222±8 5.08±0.32 28.0 12.4486±14 1.84±0.11 22.2 27.2478±14 1.92±0.11 22.8 26.7
1788±26 2.25±0.07

139C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
the small (i.e., 1–10 μm) and the medium-sized particles (10–50 μm) had the largest share (45–60%) of POC contents, indicatingthat it is the smaller particles that seem to represent the majority ofthe settling particles in the surface ocean, in agreement with thethree possible sinking mechanisms proposed by Richardson andJackson (2007, and references therein) and the reported values (53–87%) by Hung et al. (in press).
Fig. 3 shows images of bulk sinking particles collected from thebottom of the euphotic zone, demonstrating that sinking particlescontain diatom cells, fecal pellets, detritus and small particleaggregates. The size classes on panels A and B of Fig. 3 also showthat even the largest particles were less than 50 µm, and some ofparticles are even less than 10 µm. Overall, the images show thatparticles smaller than 50 µm indeed accounted for a considerablefraction of bulk sinking particles in the upwelling region. The resultsnot only support our field observations in the Gulf of Mexico, with amarine system dominated by diatoms in 2005, suggesting that thesediatom-rich particles may have been responsible at that time for themajority of sinking particles, but also provide convincing evidence thatsmaller particles can sink through the euphotic zone. As a conse-quence, it should not be assumed that one could adopt the POC/234Thratios from large (N50 µm) pump-collected particles without furtherevidence.
3.3. 234Th approach derived POC flux in the Gulf of Mexico
Trap-derived POC export fluxes in 2005 and 2006 ranged from 24to 132 mg m−2 day−1, with higher values at the CCR and lower valuesat the WCR stations (except for S4 in 2006) (Table 2). The POC fluxderived from the 234Th/238U disequilibrium approach can be estimat-
Fig. 3. SEM images of bulk sinking particles collected from the bottom of the euphotic zone ishow sinking particles containing diatom cells, fecal pellets, detritus and aggregates, with a lasome of particles b10 μm.
ed by Eq. (1), assuming steady-state conditions and neglectingadvective and diffusion terms of 234Th:
½POC flux� = ðPOC=234ThÞp × ½Fluxof 234Th� ð1Þ
where (POC/234Th)p represents the ratio of POC/234Th (µmol dpm−1,dpm=disintegration per minute) on potentially sinking particlescollected at a given depth (usually at the bottom of the euphoticzone), [Flux of 234Th] is the calculated flux of 234Th derived from thedeficiency of 234Th relative to its parent 238U in the water column atthat depth, as [238U–234Th]tot⁎λ⁎h, with 238U, 234Th in dpm/m3,λ=decay constant of 234Th (0.03 day−1), and h=height (m) of thewater column over which the flux is integrated.
Uncertainties in 234Th-based POC fluxes include uncertainties andbiases in themeasurements of POC (e.g., Gardner et al., 2003), biases ofsediment traps (Gardner, 1980; Buesseler et al., 2007), ignoringhorizontal and vertical transport processes (e.g., Savoye et al., 2006),assumptions with regards to what particles are actually settling(Szlosek et al., 2009), and biases in determining POC/234Th ratios (e.g.,Buesseler et al., 2006).
Theoretically, the POC/234Th ratio should be from particles thatsignificantly sink on the characteristic 234Th time scale (i.e., 1 month).Often, the logical assumption is made that large particles arerepresentative of major settling particles (e.g., Buesseler et al., 2006,and references herein). However, this assumption has also beenquestioned in the literature (Bacon et al., 1996; Cochran et al., 2000).An examination of POC/234Th ratios from different sized particles isrequired because the actual size spectrum of sinking particles is verydifficult to obtain and is usually not known in most previous 234Th-derived carbon export investigations.
n the southern East China Sea. The scale bar is 10 μm in panels A, B, C and D. The figuresrge fraction of the particles in the bulk sinking particles with dimensions of b50 μmand

Fig. 4. POC/234Th versus average size of filtered particles from 120 m, near the bottom ofthe euphotic zone, in the Gulf of Mexico in 2005 and 2006.
140 C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
The POC/234Th ratios in different size suspended fractions (1–10,10–50, and 50–150 μm) are shown in Table 2 and Fig. 4 for the2005 and 2006 cruises, and are in broad agreement with previouslyreported results (Guo et al., 2002a, Santschi et al., 2003; Hung etal., 2004). The ratios of POC/234Th from four depths (65, 90, 120and 140 m) in small (1–10 μm), medium-sized (10–50 μm), large(50–150 μm) and largest particles (N150 μm) in the GOM in 2005ranged from 18 to 626, 1.1 to 15.8, 2.2 to 13.7 and 1.2 to7.6 μmol dpm−1, and 14.3 to 119.6, 1.5 to 8.7, 14 to 89 and 9 to149 μmol dpm−1 in 2006, respectively. Lower ratios of POC/234Thwere found in intermediate and large particles in 2005, whilelower ratios were observed only in intermediate sized particles in2006, with one to two orders of magnitude difference between theextreme values, which ranged from 1 to 626 μmol dpm−1. Thetrend in the 2005 (but not 2006) data is similar to that previouslyreported (Guo et al., 2002a; Hung et al., 2004), showing that POC/234Th ratios increase with decreasing particle size, i.e. with anaverage of 37, 3.6, and 3.1 μmol dpm−1 for small, intermediate, andlarge particles, respectively. The 2006 data, however, show aminimum for the 10–50 µm size particles.
The mechanisms that control the ratios of POC/234Th insuspended and sinking particles are not fully understood. Besidesthe fact that different collection methods for large suspendedparticles (using bottle collection or in situ pumping system) andsinking particles can give different POC concentrations and POC/234Th ratios, differences also occur due to the unique chemicalcomposition of particles (e.g., acidic polysaccharide content andother organic carrier compounds), and phytoplankton assemblages(Quigley et al., 2002; Guo et al., 2002b; Santschi et al., 2003; Hungand Gong, 2007; Zhang et al., 2008). Details on how the POC/234Thratio is influenced by the chemical composition of particles arediscussed in Xu et al. (in press). Regardless of the exact mechanismsthat control 234Th adsorption by particles, the POC/234Th ratio onlarge particles (N50 µm) is usually used to estimate the POC flux inmost studies (Buesseler et al., 1995; Bacon et al., 1996; Cochran etal., 2000; Benitez-Nelson et al., 2001; Moran et al., 2003; Hung etal., 2004; Hung and Gong, 2007). Contrary to these studies, smallerparticles seem to be representative of the major carbon flux in theEast China Sea, within upwelling systems and the oligotrophicocean, based on recent investigations (Hung et al., in press). Therepresentativeness of large particles has also been questionedpreviously by Bacon et al. (1996), who maintained that thisassumption has not been proven. Since these size fractions aredetermined by filtration, it is also possible that the ratios in settlingaggregates can be altered by filtration-induced disaggregation (or“solubilization”) effects, as discussed further below.
Even though it is possible that larger aggregates composed ofsmaller particles are disrupted by sequential filtration, it should bepossible to compare the results from both approaches, if similarfiltration methods (i.e., sequential filtration of at least two differentsizes, Buesseler et al., 2006, and references therein) used for sinkingparticles are also applied to suspended particles. Previous observa-tions indicated that smaller size fractions of suspended particles(b50 µm) had the largest shares (N70%) of 234Th and POC (Guo et al.,2002a,b; Hung et al., 2004; Hung and Gong, 2007), when compared toother fractions (N50 µm).
In a recent paper by Lepore et al. (2009), they reported for oneof their study sites on the New England continental shelf that 60–99% of 234Th and POC was contained in the 10–50 µm fraction, anobservation which was different from their other study sites, andwhich they explained by different settling rates for 234Th and POC.According to De La Rocha and Passow (2007), suspended calciumcarbonate particles, e.g., from coccolithophorids, a haptophyte, havebeen observed to both fragments and increase the density ofaggregates, and thus, may increase their sinking velocities by up tohundreds of times.
Thus, it is plausible that these smaller particles also represent amajor portion in sinking particles composed of aggregates. Further-more, if it is indeed the case that smaller particles (e.g., via differentaggregation and sinking mechanisms, as described by Richardson andJackson, 2007) represent the predominant fraction of sinkingparticles, then the choice of the appropriate particle size and POC/234Th ratio becomes very important for calculating the POC flux usingthe 234Th/238U disequilibrium approach.
The POC/234Th ratios in trap-derived particles observed over thetwo years (2005/6) ranged from 0.8 to 20 μmol dpm−1, which is anarrower range than those observed for the different size classes ofpump-derived particles. This difference reflects the fact that theparticles captured by the sediment traps are largely distinct from thesuspended particles and suggests that it is important to choose thevalues of POC/234Th in size-fractionated particles by considering thepredominant phytoplankton species (such as nanoplanktonic hapto-phytes) rather than becoming rigidly focused on a fixed size class ofsuspended particles, such as the N50 µm size class.
In other words, the main objective for using ratios from size-fractionated particles is to use them as proxies for the “dominantsinking particles”. As noted earlier, autotrophic picoplankton (b2 µm)are often the predominant phytoplankton produced in oceanicenvironments (Michaels and Silver, 1988; Legendre and LeFevre,1995). Furthermore, field data and models suggest that smallerparticles may be more important in exporting carbon into the interiorof the deep ocean via potential sinking processes such as mineralballasting, aggregation of small particles and grazing (Armstrong et al.,2001; Jackson 1990, 2001; Barber, 2007; Richardson and Jackson, 2007and references therein). Therefore, there is no a priori reason for thePOC/234Th ratio in large (N50 µm) suspended particles to be chosen asa proxy for the actual “sinking particles”, because these larger particlesare often composed of smaller particles weakly held together inaggregates (Burd et al., 2007) thatmay easily break-up or disaggregateand revert into smaller sizes (b50 µm) upon filtration. In fact, theimages of bulk sinking particles in Fig. 3A–D and POC data in size-fractionated sinking particles (Table 4) indeed show that sinkingparticles contained many small particles (b50 µm) in both upwellingregion in the East China Sea and oligotrophic ocean in the northwestPacific. In other words, the observed SEM images on bulk sinkingparticles reflect the fact that large particles only account for a minorportion of sinking particles. Furthermore, based on the carbon and234Th content in size-fractionated sinking particles (Table 4), largeparticles (N50 µm) only constituted 40–56% for carbon and 21–38% for234Th. The inconsistent carbon and 234Th content in large sinkingparticles reflect the fact that carbon and 234Th did not exist in sinkingparticles in equal proportion.
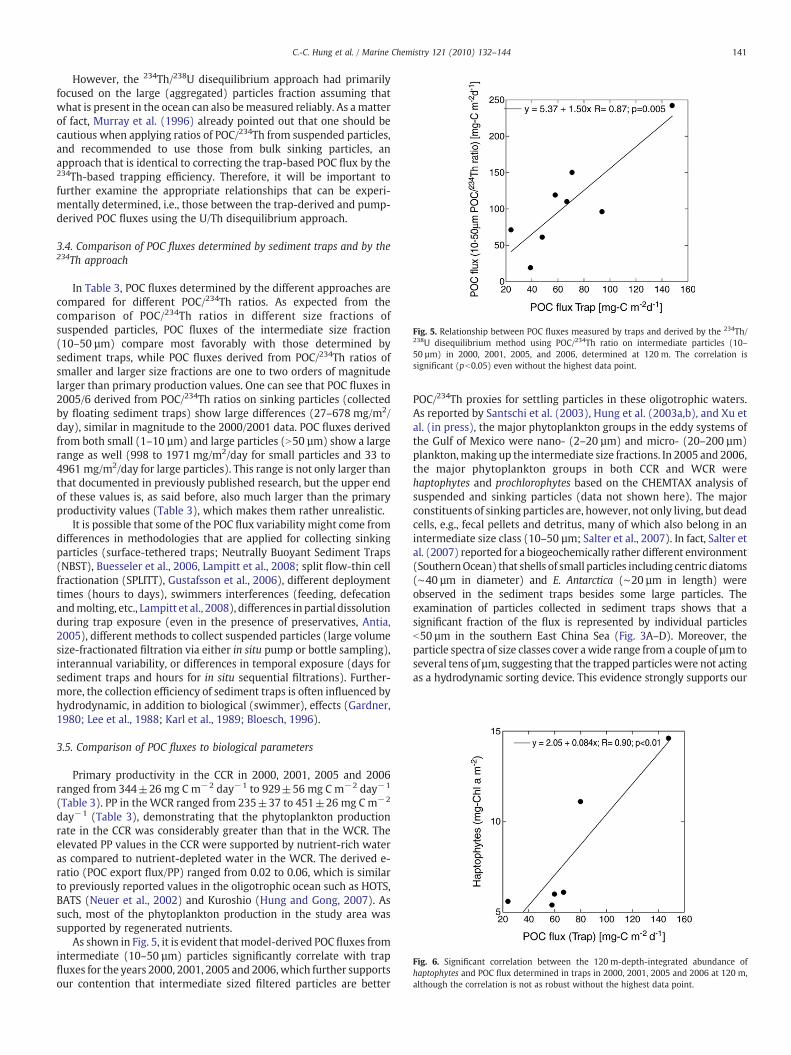
Fig. 5. Relationship between POC fluxes measured by traps and derived by the 234Th/238U disequilibrium method using POC/234Th ratio on intermediate particles (10–50 µm) in 2000, 2001, 2005, and 2006, determined at 120 m. The correlation issignificant (pb0.05) even without the highest data point.
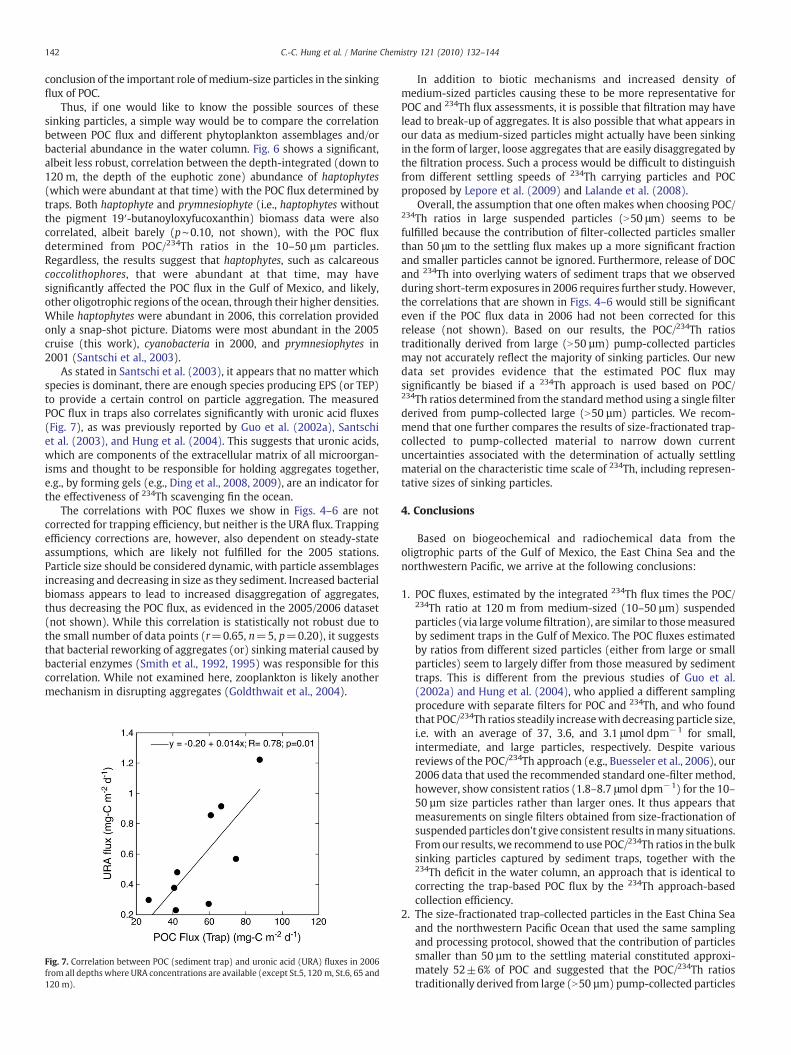
Fig. 6. Significant correlation between the 120 m-depth-integrated abundance ofhaptophytes and POC flux determined in traps in 2000, 2001, 2005 and 2006 at 120 m,although the correlation is not as robust without the highest data point.
141C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
However, the 234Th/238U disequilibrium approach had primarilyfocused on the large (aggregated) particles fraction assuming thatwhat is present in the ocean can also bemeasured reliably. As amatterof fact, Murray et al. (1996) already pointed out that one should becautious when applying ratios of POC/234Th from suspended particles,and recommended to use those from bulk sinking particles, anapproach that is identical to correcting the trap-based POC flux by the234Th-based trapping efficiency. Therefore, it will be important tofurther examine the appropriate relationships that can be experi-mentally determined, i.e., those between the trap-derived and pump-derived POC fluxes using the U/Th disequilibrium approach.
3.4. Comparison of POC fluxes determined by sediment traps and by the234Th approach
In Table 3, POC fluxes determined by the different approaches arecompared for different POC/234Th ratios. As expected from thecomparison of POC/234Th ratios in different size fractions ofsuspended particles, POC fluxes of the intermediate size fraction(10–50 µm) compare most favorably with those determined bysediment traps, while POC fluxes derived from POC/234Th ratios ofsmaller and larger size fractions are one to two orders of magnitudelarger than primary production values. One can see that POC fluxes in2005/6 derived from POC/234Th ratios on sinking particles (collectedby floating sediment traps) show large differences (27–678 mg/m2/day), similar in magnitude to the 2000/2001 data. POC fluxes derivedfrom both small (1–10 µm) and large particles (N50 µm) show a largerange as well (998 to 1971 mg/m2/day for small particles and 33 to4961 mg/m2/day for large particles). This range is not only larger thanthat documented in previously published research, but the upper endof these values is, as said before, also much larger than the primaryproductivity values (Table 3), which makes them rather unrealistic.
It is possible that some of the POC flux variability might come fromdifferences in methodologies that are applied for collecting sinkingparticles (surface-tethered traps; Neutrally Buoyant Sediment Traps(NBST), Buesseler et al., 2006, Lampitt et al., 2008; split flow-thin cellfractionation (SPLITT), Gustafsson et al., 2006), different deploymenttimes (hours to days), swimmers interferences (feeding, defecationandmolting, etc., Lampitt et al., 2008), differences in partial dissolutionduring trap exposure (even in the presence of preservatives, Antia,2005), different methods to collect suspended particles (large volumesize-fractionated filtration via either in situ pump or bottle sampling),interannual variability, or differences in temporal exposure (days forsediment traps and hours for in situ sequential filtrations). Further-more, the collection efficiency of sediment traps is often influenced byhydrodynamic, in addition to biological (swimmer), effects (Gardner,1980; Lee et al., 1988; Karl et al., 1989; Bloesch, 1996).
3.5. Comparison of POC fluxes to biological parameters
Primary productivity in the CCR in 2000, 2001, 2005 and 2006ranged from 344±26 mg C m−2 day−1 to 929±56 mg C m−2 day−1
(Table 3). PP in theWCR ranged from 235±37 to 451±26 mg C m−2
day−1 (Table 3), demonstrating that the phytoplankton productionrate in the CCR was considerably greater than that in the WCR. Theelevated PP values in the CCR were supported by nutrient-rich wateras compared to nutrient-depleted water in the WCR. The derived e-ratio (POC export flux/PP) ranged from 0.02 to 0.06, which is similarto previously reported values in the oligotrophic ocean such as HOTS,BATS (Neuer et al., 2002) and Kuroshio (Hung and Gong, 2007). Assuch, most of the phytoplankton production in the study area wassupported by regenerated nutrients.
As shown in Fig. 5, it is evident thatmodel-derived POC fluxes fromintermediate (10–50 µm) particles significantly correlate with trapfluxes for the years 2000, 2001, 2005 and 2006,which further supportsour contention that intermediate sized filtered particles are better
POC/234Th proxies for settling particles in these oligotrophic waters.As reported by Santschi et al. (2003), Hung et al. (2003a,b), and Xu etal. (in press), the major phytoplankton groups in the eddy systems ofthe Gulf of Mexico were nano- (2–20 µm) and micro- (20–200 µm)plankton,making up the intermediate size fractions. In 2005 and 2006,the major phytoplankton groups in both CCR and WCR werehaptophytes and prochlorophytes based on the CHEMTAX analysis ofsuspended and sinking particles (data not shown here). The majorconstituents of sinking particles are, however, not only living, but deadcells, e.g., fecal pellets and detritus, many of which also belong in anintermediate size class (10–50 µm; Salter et al., 2007). In fact, Salter etal. (2007) reported for a biogeochemically rather different environment(SouthernOcean) that shells of small particles including centric diatoms(∼40 µm in diameter) and E. Antarctica (∼20 µm in length) wereobserved in the sediment traps besides some large particles. Theexamination of particles collected in sediment traps shows that asignificant fraction of the flux is represented by individual particlesb50 µm in the southern East China Sea (Fig. 3A–D). Moreover, theparticle spectra of size classes cover awide range from a couple of µm toseveral tens of µm, suggesting that the trapped particleswere not actingas a hydrodynamic sorting device. This evidence strongly supports our
142 C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
conclusion of the important role of medium-size particles in the sinkingflux of POC.
Thus, if one would like to know the possible sources of thesesinking particles, a simple way would be to compare the correlationbetween POC flux and different phytoplankton assemblages and/orbacterial abundance in the water column. Fig. 6 shows a significant,albeit less robust, correlation between the depth-integrated (down to120 m, the depth of the euphotic zone) abundance of haptophytes(which were abundant at that time) with the POC flux determined bytraps. Both haptophyte and prymnesiophyte (i.e., haptophytes withoutthe pigment 19′-butanoyloxyfucoxanthin) biomass data were alsocorrelated, albeit barely (p∼0.10, not shown), with the POC fluxdetermined from POC/234Th ratios in the 10–50 µm particles.Regardless, the results suggest that haptophytes, such as calcareouscoccolithophores, that were abundant at that time, may havesignificantly affected the POC flux in the Gulf of Mexico, and likely,other oligotrophic regions of the ocean, through their higher densities.While haptophytes were abundant in 2006, this correlation providedonly a snap-shot picture. Diatoms were most abundant in the 2005cruise (this work), cyanobacteria in 2000, and prymnesiophytes in2001 (Santschi et al., 2003).
As stated in Santschi et al. (2003), it appears that no matter whichspecies is dominant, there are enough species producing EPS (or TEP)to provide a certain control on particle aggregation. The measuredPOC flux in traps also correlates significantly with uronic acid fluxes(Fig. 7), as was previously reported by Guo et al. (2002a), Santschiet al. (2003), and Hung et al. (2004). This suggests that uronic acids,which are components of the extracellular matrix of all microorgan-isms and thought to be responsible for holding aggregates together,e.g., by forming gels (e.g., Ding et al., 2008, 2009), are an indicator forthe effectiveness of 234Th scavenging fin the ocean.
The correlations with POC fluxes we show in Figs. 4–6 are notcorrected for trapping efficiency, but neither is the URA flux. Trappingefficiency corrections are, however, also dependent on steady-stateassumptions, which are likely not fulfilled for the 2005 stations.Particle size should be considered dynamic, with particle assemblagesincreasing and decreasing in size as they sediment. Increased bacterialbiomass appears to lead to increased disaggregation of aggregates,thus decreasing the POC flux, as evidenced in the 2005/2006 dataset(not shown). While this correlation is statistically not robust due tothe small number of data points (r=0.65, n=5, p=0.20), it suggeststhat bacterial reworking of aggregates (or) sinking material caused bybacterial enzymes (Smith et al., 1992, 1995) was responsible for thiscorrelation. While not examined here, zooplankton is likely anothermechanism in disrupting aggregates (Goldthwait et al., 2004).
Fig. 7. Correlation between POC (sediment trap) and uronic acid (URA) fluxes in 2006from all depths where URA concentrations are available (except St.5, 120 m, St.6, 65 and120 m).
In addition to biotic mechanisms and increased density ofmedium-sized particles causing these to be more representative forPOC and 234Th flux assessments, it is possible that filtration may havelead to break-up of aggregates. It is also possible that what appears inour data as medium-sized particles might actually have been sinkingin the form of larger, loose aggregates that are easily disaggregated bythe filtration process. Such a process would be difficult to distinguishfrom different settling speeds of 234Th carrying particles and POCproposed by Lepore et al. (2009) and Lalande et al. (2008).
Overall, the assumption that one often makes when choosing POC/234Th ratios in large suspended particles (N50 μm) seems to befulfilled because the contribution of filter-collected particles smallerthan 50 μm to the settling flux makes up a more significant fractionand smaller particles cannot be ignored. Furthermore, release of DOCand 234Th into overlying waters of sediment traps that we observedduring short-term exposures in 2006 requires further study. However,the correlations that are shown in Figs. 4–6 would still be significanteven if the POC flux data in 2006 had not been corrected for thisrelease (not shown). Based on our results, the POC/234Th ratiostraditionally derived from large (N50 µm) pump-collected particlesmay not accurately reflect the majority of sinking particles. Our newdata set provides evidence that the estimated POC flux maysignificantly be biased if a 234Th approach is used based on POC/234Th ratios determined from the standardmethod using a single filterderived from pump-collected large (N50 µm) particles. We recom-mend that one further compares the results of size-fractionated trap-collected to pump-collected material to narrow down currentuncertainties associated with the determination of actually settlingmaterial on the characteristic time scale of 234Th, including represen-tative sizes of sinking particles.
4. Conclusions
Based on biogeochemical and radiochemical data from theoligtrophic parts of the Gulf of Mexico, the East China Sea and thenorthwestern Pacific, we arrive at the following conclusions:
1. POC fluxes, estimated by the integrated 234Th flux times the POC/234Th ratio at 120 m from medium-sized (10–50 µm) suspendedparticles (via large volumefiltration), are similar to thosemeasuredby sediment traps in the Gulf of Mexico. The POC fluxes estimatedby ratios from different sized particles (either from large or smallparticles) seem to largely differ from those measured by sedimenttraps. This is different from the previous studies of Guo et al.(2002a) and Hung et al. (2004), who applied a different samplingprocedure with separate filters for POC and 234Th, and who foundthat POC/234Th ratios steadily increasewith decreasing particle size,i.e. with an average of 37, 3.6, and 3.1 μmol dpm−1 for small,intermediate, and large particles, respectively. Despite variousreviews of the POC/234Th approach (e.g., Buesseler et al., 2006), our2006 data that used the recommended standard one-filter method,however, show consistent ratios (1.8–8.7 μmol dpm−1) for the 10–50 µm size particles rather than larger ones. It thus appears thatmeasurements on single filters obtained from size-fractionation ofsuspendedparticles don't give consistent results inmany situations.Fromour results, we recommend to use POC/234Th ratios in the bulksinking particles captured by sediment traps, together with the234Th deficit in the water column, an approach that is identical tocorrecting the trap-based POC flux by the 234Th approach-basedcollection efficiency.
2. The size-fractionated trap-collected particles in the East China Seaand the northwestern Pacific Ocean that used the same samplingand processing protocol, showed that the contribution of particlessmaller than 50 µm to the settling material constituted approxi-mately 52±6% of POC and suggested that the POC/234Th ratiostraditionally derived from large (N50 µm) pump-collected particles

143C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
may not accurately reflect the majority of sinking particles. Ournew data set thus provides evidence that the 234Th approachbased on POC/234Th ratios from large (N50 µm) pump-collectedparticles that is common in the oceanographic communitymight significantly bias the estimated POC flux obtained bythis approach.
3. The observation that 234Th is mainly associated with medium-sizedparticles rather than larger ones (that would be expected to sinkmore rapidly) does not mean to imply that these particles actuallysank in their disaggregated state. Rather, the observed size spectrummight be a result of the commonly used handling and filtrationprocedures for collection of suspended particles. We suggest that, inorder to define the biological and chemical conditions that lead tosuch observations, correlations with biological and chemical para-meters should be explored as much as possible.
4. POCfluxes in 2005/6 significantly and positively correlatedwith theintegrated haptophytes biomass and the uronic acid flux, while234Thflux also correlatedwith the APSflux. In addition, a less robustand negative correlation with bacterial biomass was found. Theserelationships strongly suggest that at that time not only was thisnanoplanktonic group themain driver for the vertical POC flux, andresponsible for the observed concentration of 234Th in themedium-sized particles, but also confirmed the role of acid-polysaccharide-rich EPS in holding the loosely-held aggregates together, suggestingthat a role for bacteria in reworking particle aggregates thatcounteract efficient removal from the water by sinking.
5. We therefore recommend that if size-fractionated suspendedparticles are used for the POC/234Th ratio, this ratio needs to befurther examined in different oceanic regimes and appropriatelyselected for estimating POC export fluxes because particle size(resulting from different phytoplankton assemblages) in variousoceanic regimes may be different. If possible, the sinking particles(e.g., those collected by sediment traps) should also be further size-fractionated in order to verify the dominant size fractions of sinkingparticles.
Acknowledgements
We appreciate the assistance of the captain, crew and scienceparties of the R/V Pelican and the R/V Seward Johnson. This researchwas supported by NSF grants (BES-0210865, OCE-0351559, and OCE-08511191) to P. H. Santschi and J. Pinckney, the Welch Foundation(Grant BD-0046) to C. Xu, and NSF OCE-0627820 and OCE-0850957 toL.D. Guo. We are also grateful for the support of the NSC of Taiwan(NSC97-2745-M-019-001, NSC98-2611-M-019-014 and NSC98-2628-M-019-011) and the Center of Marine Bioenvironment andBiotechnology (CMBB) at National Taiwan Ocean University. Wewantto give our special thanks to two anonymous reviewers for theirconstructive comments, which greatly improved the manuscript.
References
Antia, A.N., 2005. Particle-associated dissolved elemental fluxes: revising thestochiometry of mixed layer export. Biogeosci. 2, 189–204.
Armstrong, R.A., Lee, C., Hedges, J.I., Honjo, S., Wakeham, S.G., 2001. A new, mechanisticmodel for organic carbon fluxes in the ocean based on the quantitative associationof POC with ballast minerals. Deep Sea Research II 49, 219–236.
Bacon, M.P., Cochran, J.K., Hirschberg, D., Hammar, T.R., Fleer, A.P., 1996. Export flux ofcarbon at the equator during the EqPac time-series cruises estimated from 234Thmeasurements. Deep-Sea Res. 43, 1133–1154.
Barber, R.T., 2007. Picoplankton do some heavy lifting. Science 315, 777–778.Baskaran, M., Murphy, D.J., Santschi, P.H., Orr, J.C., Schink, D.R., 1993. Amethod for rapid
in situ extraction of Th, Pb and Ra isotopes from large volumes of seawater. Deep-Sea Res. 40, 849–865.
Behrenfeld, M.J., Falkowski, P.G., 1997. Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol. Oceangr. 42, 1–20.
Benitez-Nelson, C., Buesseler, K.O., Karl, K., Andrews, J., 2001. A time-series study ofparticulate matter export in the north Pacific Subtropical Gyre based upon234Th:238U disequilibrium. Deep-Sea Res. 48, 2595–2611.
Biggs, D.C., 1992. Nutrients, plankton, and productivity in a warm-core ring in thewestern Gulf of Mexico. J. Geophys Res. 97 (C2), 2143–2154.
Bloesch, J., 1996. Towards a new generation of sediment traps and a bettermeasurement/understanding of settling particle flux in lakes and oceans: a hydro-dynamical protocol. Aqu. Sci. 58, 283–296.
Buesseler, K.O., 1991. Do upper ocean sediment traps provide an accurate record ofparticle flux? Nature 353, 420–423.
Buesseler, K.O., 1998. The decoupling of production and particulate export in thesurface ocean. Glob. Biogeochem. Cycles 12, 297–310.
Buesseler, K.O., Bacon, M.P., Cochran, J.K., Livingston, H.D., 1992a. Carbon and nitrogenexport during the JGOFS North Atlantic Bloom Experiment estimated from234Th:238U disequilibria. Deep-Sea Res. 39, 1115–1137.
Buesseler, K.O., Cochran, J.K., Bacon, M.P., Livingston, H.D., Casso, S.A., Hirschberg, D.,Hartman, M.C., Fleer, A.P., 1992b. Determination of thorium isotopes in seawater bynon-destructive and radiochemical procedures. Deep-Sea Res. 39, 1103–1114.
Buesseler, K.O., Andrews, J.A., Hartman, M.C., Belastock, R., Chai, F., 1995. Regionalestimates of the export flux of particulate organic carbon derived from thorium-234 during the JGOFS EQPAC program. Deep-Sea Res. II 42, 777–804.
Buesseler, K.O., Benitez-Nelson, C.R., Moran, S.B., Burd, A., Charette, M., Cochran, J.K.,Coppola, L., Fisher, N.S., Fowler, S.W., Gardner, W.D., Guo, L.D., Gustafsson, O.,Lamborg, C., Masque, P., Miquel, J.C., Passow, U., Santschi, P.H., Savoye, N., Stewart,G., Trull, T., 2006. An assessment of particulate organic carbon to thorium-234ratios in the ocean and their impact on the application of 234Th as a POC flux proxy.Mar. Chem. 100, 213–233.
Buesseler, K.O., Antia, A.N., Chen, M., Fowler, S.W., Gardner, W.D., Gustafsson, O.,Harada, K., Michaels, A.F., Rutgers van der Loeff, M., Sarin, M., Steinberg, D.K., Trull,T., 2007. An assessment of the use of sediment traps for estimating upper oceanparticle fluxes. J. Mar. Res. 65, 345–416.
Burd, A.B., Jackson, G.A., Moran, S.B., 2007. The role of the particle size spectrum inestimating POC fluxes from Th-234/U-238 disequilibrium. Deep-Sea Res. I 54,897–918.
Campbell, L., Nolla, H.A., Vaulot, D., 1994. The importance of Prochlorococcus tocommunity structure in the central North Pacific Ocean. Limnol. Oceanogr. 39,954–961.
Charette, M.A., Moran, S.B., Pike, S.M., Smith, J.N., 2001. Investigation the carbon cycle inthe Gulf of Maine using the natural tracer thorium-234. J. Geophys. Res. 106,11553–11579.
Chen, M., Huang, Y., Cai, P., Guo, L., 2003. Particulate organic carbon export fluxes in theCanada Basin and Bering Sea as derived from 234Th/238U disequilibria. Arctic 56,32–44.
Coale, K.H., Bruland, K.W., 1985. 234Th:238U disequilibria within the California Current.Limnol. Oceangr. 30, 22–33.
Cochran, J.K., Barnes, C., Achman, D., Hirschberg, D.T., 1995. Thorium-234/Uranium-238disequilibrium as an indicator of scavenging rates and particulate organic carbonfluxes in the Northeast Water Polynya, Greenland. J. Geophys. Res. 100 (C3),4399–4410.
Cochran, J.K., Buesseler, K.O., Bacon, M.P., et al., 2000. Short-lived thorium isotopes (Th-234, Th-228) as indicators of POC export and particle cycling in the Ross Sea,Southern Ocean. Deep-Sea Res. II 47, 3451–3490.
Coppola, L., Roy-Barman, M., Wassmann, P., Mulsow, S., Jeandel, C., 2002. Calibration ofsediment traps and particulate organic carbon export using 234Th in the BarentsSea. Mar. Chem. 80, 11–26.
De La Rocha, C.L., Passow, U., 2007. Factors influencing the sinking of POC and theefficiency of the biological carbon pump. Deep-Sea Res. II 54, 639–658.
Ding, Y.-X., Chin, W.-C., Rodriguez, A., Hung, C.-C., Santschi, P.H., Verdugo, P., 2008.Amphiphilic exopolymers from Sagitulla stellata induce self-assembly of DOMpolymers and the formation of marine microgels. Mar. Chem. 112, 11–19.
Ding, Y.-X., Hung, C.-C., Santschi, P.H., Verdugo, P., Chin, W.-C., 2009. Spontaneousassembly of exopolymers from phytoplankton. Terrestrial, Atmospheric andOceanic Sciences (TAO) 20 (5), 741–747.
Ducklow, H.W., Steinberg, D.K., Buesseler, K.O., 2001. Upper ocean carbon export andthe biological pump. Oceanography 14, 50–58.
Feig, Y.S., Porter, K.G., 1980. The use of DAPI for identifying and counting aquaticmicroflora. Limnol. Oceanogr. 25, 943–948.
Gardner, W.D., 1980. Field assessment of sediment traps. J. Mar. Res. 38, 41–52.Gardner, W.D., Richardson, M.J., Carlson, C.A., Hansell, D., Mishonov, A.V., 2003.
Determining true particulate organic carbon: bottles, pumps and methodologies.Deep-Sea Res. II. 50, 655–674.
Goldthwait, S., Yen, J., Brown, J., Alldredge, A., 2004. Quantification of marine snowfragmentation by swimming Euphausiids. Limnol. Oceangr. 49, 940–952.
Guo, L., Santschi, P.H., 1997a. Isotopic and elemental characterization of colloidalorganic matter from the Chesapeake Bay and Galveston Bay. Mar. Chem. 59, 1–15.
Guo, L., Santschi, P.H., 1997b. Measurements of dissolved organic carbon (DOC) inseawater by the high temperature combustion method. Acta Oceanol. Sin. 16 (2),59–73.
Guo, L., Santschi, P.H., Baskaran, M., Zindler, A., 1995. Distribution of dissolved andparticulate 230Th and 232Th in seawater from the Gulf of Mexico and off CapeHatteras as measured by SIMS. Earth Planet. Sci. Lett. 133, 117–128.
Guo, L., Santschi, P.H., Baskaran, M., 1997. Interaction of thorium isotopes with colloidalorganic matter in oceanic environments. Colloids Surf. A 120, 255–271.
Guo, L., Hung, C.-C., Santschi, P.H., Walsh, I., 2002a. 234Th scavenging and itsrelationship to acid polysaccharide abundance in the Gulf of Mexico. Mar. Chem.78, 103–119.
Guo, L., Chen, M., Gueguen, C., 2002b. Control of Pa/Th ratio by particulate chemicalcomposition in the ocean. Geophys. Res. Lett. 29 (20), 1960. doi:10.1029/2002GL015666.

144 C.-C. Hung et al. / Marine Chemistry 121 (2010) 132–144
Gustafsson, O., Larsson, J., Andersson, P., Ingri, J., 2006. The POC/Th-234 ratio of settlingparticles isolated using split flow-thin cell fractionation (SPLITT). Mar. Chem. 100,314–322.
Hobbie, J.E., Daley, R.J., Jasper, S., 1977. Use of nuclepore filters for counting bacteria byfluorescence microscopy. Appl. Environ. Micorbiol. 33, 1225–1228.
Hung, C.-C., Gong, G.C., 2007. Export flux of POC in the main stream of the Kuroshio.Geophys. Res. Lett. 34, L18606. doi:10.1029/2007GL030236.
Hung, C.-C., Guo, L., Schultz Jr., G.E., Pinckney, Santschi, P.H., 2003a. Production and fluxof carbohydrate species in the Gulf of Mexico. Glob. Biogeochem. Cycles 17.doi:10.1029/2002GB001988.
Hung, C.-C., Guo, L., Santschi, P.H., Alvarado-Quiroz, N., Haye, J.M., 2003b. Distributionsof carbohydrate species in the Gulf of Mexico. Mar. Chem. 81, 119–135.
Hung, C.-C., Guo, L., Roberts, K., Santschi, P.H., 2004. Upper ocean carbon fluxdetermined by 234Th and sediment traps in the Gulf of Mexico. Geochem. J. 38,601–611.
Hung, C.-C., Gong, G.-C., Chung, W.-C., Kuo, W.-T., Lin, F.-C., 2009. Enhancement ofparticulate organic carbon export flux induced by atmospheric forcing in thesubtropical oligotrophic northwest Pacific Ocean. Mar. Chem. 113, 19–24.
Hung, C.-C., Gwo-Ching Gong, G.-C., Santschi, P.H., in press. Ratios of POC/234Th indifferent size classes of sediment-trap-collected particles from different oceanicenvironments. J. Geophys. Res.
Jackson, G.A, 1990. A model of the formation of marine algal flocs by physicalcoagulation processes. Deep-Sea Res. A 37/8, 1197–1211.
Jackson, G.A., 2001. Effect of coagulation on amodel planktonic foodweb. Deep-Sea Res.I 48, 95–123.
Karl, D.M., Knauer, G.A., Martin, J.H., 1989. Downward flux of particulate organic matterin the ocean — a particle decomposition paradox. Nature 332, 438–441.
Ku, T.L., Knauss, K.G., Matthew, G.G., 1977. Uranium in the open ocean: concentrationand isotopic composition. Deep-Sea Res. 24, 1005–1017.
Lalande, C., Moran, S.B., PaulWassmann, P., Jacqueline,M., Grebmeier, J.M., Cooper, L.W.,2008. 234Th-derived particulate organic carbon fluxes in the northern Barents Seawith comparison to drifting sediment trap fluxes. J. Mar. Syst. 73, 103–113.
Lampitt, R.S., Boorman, B., Brown, L., Lucas, M., Salter, I., Sanders, R., Sawa, K., Seeyave,S., Thomalla, S.J., Turnewitsch, R., 2008. Particle export from the euphotic zone:estimates using a novel drifting sediment trap, 234Th and new production. Deep-Sea Res. I 55, 1484–1502.
Lee, S., Fuhrman, J.A., 1987. Relationships between biovolume and biomass of naturallyderived marine bacterioplankton. Appl. Environ. Microbiol. 53, 1298–1303.
Lee, C., Hedges, J.I., Wakeham, S.G., Zhu, N., 1992. Effectiveness of poisons andpreservatives in retarding bacterial activity in sediment trap material. Limnologyand Oceanography 37, 117–130.
Legendre, L., LeFevre, J., 1995. Microbial food webs and the export of biogenic carbon inocean. Aquat. Microb. Ecol. 9, 69–77.
Lepore, K., Moran, S.B., Burd, A.B., Jackson, G.A., Smith, J.N., Kelly, R.P., Kaberi, H.,Stavrakakis, S., Assimakopoulou, G., 2009. Sediment trap and in-situ pump size-fractionated POC/234Th ratios in the Mediterranean Sea and Northwest Atlantic:implications for POC export. Deep-Sea Res. I 56, 599–613.
Li, W.K.W., 1994. Primary productivity of prochlorophytes, cyanobacteria, andeukaryotic ultraphytoplankton: measurements from flow cytometric sorting.Limnol. Oceanogr. 39, 169–175.
Mackey, M., Mackey, D., Kiggins, H., Wright, S., 1996. CHEMTAX-A program forestimating class abundances from chemical markers: application to HPLC mea-surements of phytoplankton. Mar. Ecol. Prog. Ser. 144, 265–283.
Maiti, K., Buesseler, K., Benitez-Nelson, C.R., Cochran, J.K., Dai, M., Dehairs, F., Masque, P.,Miller, L.A., Moran, B., Morris, P.J., Miquel, J., Peine, F., Planchon, F., RutgersvanderLoeff, M.P.H., Santschi, P.H., Turnewitsch, R., Waples, J.T., 2010. Total and particulatethorium-234 results from GEOTRACES intercalibration cruises. 2010 OceanSciences Meeting, Feb. 22–26, Portland, Oregon. Abstract CO23A-02.
Michaels, A.F., Silver, M.W., 1988. Primary production, sinking fluxes and the microbialfood web. Deep-Sea Res. 35, 473–490.
Moon-van der Staay, S.Y., vanderStaay, G.W.M., Guillou, L., Vaulot, D., Claustre, H.,Medlin, L.K., 2000. Abundance and diversity of prymnesiophytes in the picoplank-ton equatorial Pacific Ocean inferred from 18S rDNA sequences. Limnol. Oceanogr.45 (1), 98–109.
Moran, S.B., Weinstein, S.E., Edmonds, H.N., Smith, J.N., Kelly, R.P., Pilson, M.E.Q.,Harrison, W.G., 2003. Does 234Th/238U disequilibrium provide an accurate record ofthe export flux of particulate organic carbon from the upper ocean? Limnol.Oceanogr. 48, 1018–1029.
Murray, J.W., Young, J., Newton, J., Dunne, J., Chapin, T., Paul, B., 1996. Export flux ofparticulate organic carbon from the central Equatorial Pacific determined using acombined drifting trap — 234Th approach. Deep-Sea Res. 43, 1095–1132.
Neuer, S., Davenport, R., Freudenthal, T.,Wefer, G., Llinas, O., Rueda,M.-J., Steinberg, D.K.,Karl, D.M., 2002. Differences in the biological carbon pump at three subtropicalocean sites. Geophy. Res. Lett. 29, 1885. doi:10.1029/2002GL015393.
Pinckney, J.L., Millie, D.F., Howe, K.E., Paerl, H.W., Hurley, J., 1996. Flow scintillationcounting of 14C-labeled microalgal photosynthetic pigments. J. Plankton Res. 18,1867–1880.
Quigley, M.S., Santschi, P.H., Hung, C.-C., Guo, L., Honeyman, B.D., 2002. Importance ofacid polysaccharides for 234Th complexation to marine organic matter. Limnol.Oceanogr. 47, 367–377.
Richardson, T., Jackson, G.A., 2007. Small phytoplankton and carbon export from thesurface ocean. Science 315, 838–840.
Salter, I., Lampitt, R.S., Sanders, R., Poulton, A., Kemp, A.E.S., Boorman, B., Saw, K., Pearce, R.,2007. Estimating carbon, silica and diatom export from a natural fertilizedphytoplankton bloom in the Southern Ocean using PELAGRA: a novel driftingsediment trap. Deep-Sea Res. II 54, 2233–2259.
Santschi, P.H., Guo, L., Quigley, M.S., Baskaran, M., Walsh, I., 1999. Boundary exchangeand scavenging of radionuclides in continental margin waters of the MiddleAtlantic Bight: implications for organic carbon fluxes. Cont. Shelf Res. 19, 609–636.
Santschi, P.H., Hung, C.-C., Guo, L., Schultz, G.G., Pinckney, J., Walsh, I., 2003. Control ofacid polysaccharide production, 234Th and particulate organic carbon export flux bymarine organisms. Geophy. Res. Lett. 30. doi:10.1029/2002GL015727.
Santschi, P.H., Murray, J.W., Baskaran, M., Benitez-Nelson, C.R., Guo, L.D., Hung, C.-C.,Lamborg, C., Moran, S.B., Passow, U., Roy-Barman, M., 2006. Thorium speciation inseawater. Mar. Chem. 100, 250–268.
Savoye, N., Benitez-Nelson, C., Burd, A.B., Cochran, J.K., Charette, M., Buesseler, K.O.,Jackson, G.A., Roy-Barman, M., Schmidt, S., Elskens, M., 2006. 234Th sorption andexport models in the water column: a review. Mar. Chem. 100, 234–249.
Smith, D.C., Simon, M., Alldredge, A., Azam, F., 1992. Intense hydrolytic enzyme activityon marine aggregates and implications for rapid particle dissolution. Nature 359,139–142.
Smith, D.C., Steward, G.F., Long, R.A., Azam, F., 1995. Bacterial mediation of carbonfluxes during a phytoplankton bloom in a mesocosm. Deep-Sea Res. 42, 75–97.
Szlosek, J., Cochran, J.K., Miquel, J.C., Masqué, P., Armstrong, R.A., Fowler, S.W., Gasser,B., Hirschberg, D.J., 2009. Particulate organic carbon–234Th relationships in particlesseparated by settling velocity in the northwest Mediterranean Sea. Deep-Sea Res. II56, 1519–1532.
Wakeham, S.G., Hedges, J.I., Lee, C., Peterson, M.L., Hernes, P., 1997. Compositions andtransport of lipid biomarkers through the water column and surficial sediments ofthe equatorial Pacific Ocean. Deep-Sea Res. II 44, 2131–2162.
Walsh, I.D., 1990. Project CATSTIX: Camera, Transmissometer and Sediment TrapIntegration Experiment. Ph.D. Dissertation. Texas A&M University, College Station,TX 77843.
Wormuth, J.H., Ressler, P.H., Cady, R.B., Harris, E.J., 2000. Zooplankton and micronektonin cyclones and anticyclones in the Northeast Gulf of Mexico. Gulf of MexicoScience, pp. 23–34.
Wright, S., Thomas, D., Marchant, H., Higgins, H., Mackey, M., Mackey, D., 1996. Analysisof phytoplankton of the Australian sector of the Southern Ocean: comparisons ofmicroscopy and size frequency data with interpretations of pigment HPLC datausing the CHEMTAX matrix factor-isolation program. Mar. Ecol. Prog. Ser. 144,285–298.
Xu, C., Santschi, P. H., Hung, C.-C., Zhang, S., Schwehr, K, A., Roberts, K.A., Guo, L.D., Gong,G.-C., Quigg, A., Long, R., Pinckney, J., Duan, S.W., Amon, R., Wei, C.-L., in press.Controls of Th-234 removal from the oligotrophic ocean by polyuronic acids andmodified by microbial activity. Marine Chemistry.
Zhang, S., Xu, C., Santschi, P.H., 2008. Chemical characterization of extracellularpolymeric substances produced by the marine diatom Amphora sp., and theirinteractions with 234Th (IV). Marine Chemistry 112, 81–92.
Top Related
Copyright © 2022 FDOKUMEN

