







Bahasa
Halaman
Hukum


Chironomid (Diptera: Chironomidae) communitiesin six European glacier-fed streams
B. LODS-CROZET,* V. LENCIONI,² J . S . OÂ LAFSSON,³ D. L. SNOOK,§ G. VELLE,±J . E . BRITTAIN,** E. CASTELLA* and B. ROSSARO²²
*Laboratoire d'Ecologie et de Biologie Aquatique, University of Geneva, Geneva, Switzerland
²Department of Invertebrate Zoology and Hydrobiology, Museum of Natural Science of Trento, Trento, Italy
³Institute of Biology, University of Iceland, Reykjavik, Iceland
§School of Geography and Environmental Sciences, University of Birmingham, Birmingham, U.K.
±Museum of Zoology, University of Bergen, Bergen, Norway
**Freshwater Ecology and Inland Fisheries Laboratory, University of Oslo, Oslo, Norway
²²Department of Biology, Section Ecology, University of Milano, Milano, Italy
SUMMARY
1. A study on glacial stream ecosystems was carried out in six regions across Europe, from
Svalbard to the French Pyrenees. The main aim was to test the validity of the conceptual
model of Milner & Petts (1994) with regard to the zonation of chironomids of glacier-fed
rivers along altitudinal and latitudinal gradient.
2. Channel stability varied considerably, both on the latitudinal and altitudinal scale,
being lowest in the northern regions (Svalbard, Iceland and Norway) and the Swiss Alps.
Water temperature at the upstream sites was always <2 °C.
3. There was a prominent difference in taxonomic richness between the Alpine and the
northern European regions, with a higher number of taxa in the south. In all regions, the
chironomid community was characterized by the genus Diamesa and the subfamily
Orthocladiinae. Of a total of 63 taxa recorded, two (Diamesa bertrami and Orthocladiusfrigidus) were common in all the regions except Svalbard.
4. On the basis of cluster analysis, seven distinct groups of sites were evident amongst
glacial-fed systems of the ®ve regions (Pyrenees excluded). This classi®cation separated
the glacier-fed streams on geographical, latitudinal and downstream gradients.
5. Canonical Correspondence Analysis (CCA) of environmental variables was carried out
using 41 taxa at 105 sites. Slope, water depth, distance from source, water temperature and
the Pfankuch channel stability index were found to be the major explanatory environ-
mental variables. The analysis separated Diamesinae and typical upstream orthoclads
from the other chironomids by low temperature and high channel instability.
6. In all six regions, Diamesa was present closest to the glacier. Within 200 m of the glacier
snout, other genera of Diamesinae were found together with Orthocladiinae. Pioneer taxa
like Diamesa species coexisted with later colonizers like Eukiefferiella minor/®ttkaui in
relatively unstable channels.
7. The longitudinal succession of chironomid assemblages across altitudinal and latitu-
dinal gradients in glacial streams followed the same pattern, with similar genera and
groups of species. The general aspects of the conceptual model of Milner & Petts (1994)
were supported. However, Diamesa species have wider temperature limits than predicted
and other Diamesinae as well as Orthocladiinae colonize metakryal habitats.
Correspondence: Brigitte Lods-Crozet, Laboratoire d'Ecologie et de Biologie Aquatique, University of Geneva, 18, ch. des
Clochettes, CH-1206 Geneva, Switzerland. E-mail: [email protected]
Freshwater Biology (2001) 46, 1791±1809
Ó 2001 Blackwell Science Ltd 1791

Keywords: alpine, arctic, glacial stream, Chironomidae, latitudinal distribution
Introduction
The cold temperatures and marked seasonality of
Alpine and arctic rivers limit the potential types of
insect life cycles (Rossaro, 1991a, b; Danks, 1999).
Detailed information about life cycles in these regions
is still fragmentary. Glacial meltwater streams, de®ned
as kryal biotopes by Steffan (1971), are characterized
by very cold (near 0 °C), highly turbid and fast
¯owing waters with low channel stability and sea-
sonal ¯ow regimes with high diel peaks in summer.
Most previous research on glacier-fed streams has not
primarily focused on chironomid communities
(Thienemann, 1936; Dorier, 1937; Saether, 1968;
Steffan, 1971; Serra-Tosio, 1973; Kownacki, 1991) and
only a few studies have compared streams in different
regions (Kownacka & Kownacki, 1975; Kawecka,
Kownacki & Kownacka, 1978; Milner & Petts, 1994;
Lindegaard & Brodersen, 1995). The subfamily Diam-
esinae `and especially the genus Diamesa' are typically
the ®rst taxa to colonize streams immediately down-
stream of source glaciers. Diamesinae are by virtue of
their narrow ecological niche (mainly cold-stenother-
mal species) and widespread distribution through the
Palaearctic region, is a very pertinent group for
zoogeographic and ecological research (Serra-Tosio,
1973; Rossaro, 1995).
The gradient of environmental conditions that
occurs as a function of altitude or latitude within
European streams offers excellent opportunities to
investigate factors which in¯uence the diversity,
composition and abundance of stream organisms
(Ward, 1986). Stream habitats that are frequently
and/or intensely disturbed are expected to exhibit
low species richness, because few species are able to
colonize them during the brief periods between
disturbance events or to tolerate high disturbance
intensity (Townsend, Scarsbrook & DoleÂdec, 1997).
Likewise, Townsend (1989) suggested that even
where competition or predation have been shown to
play a role of shaping stream communities, the tem-
poral phenomena of disturbance and colonization are
invariably of fundamental importance. Stream benthic
communities occurring under frequent disturbances
comprise species that have been variously referred
to as fugitive or r-selected species (Pianka, 1978).
The rapid colonization ability of chironomids enables
them to dominate early in colonization processes
(Ladle et al., 1985; Townsend, 1989; Ruse, 1994).
In an attempt to explain the primary physical,
chemical and biological factors that determine macro-
invertebrate distributions downstream of glaciers
across Europe, Milner & Petts (1994) proposed a con-
ceptual model to explain variations in the zoobenthic
community structure within and between glacier-fed
streams. This model assumed that the primary
physical variables in¯uencing the macroinvertebrates
succession in glacial streams are maximum water
temperature and channel stability, which both increase
downstream from the glacial margins. As chironomids
are a major element in the model, we have focused our
study on two objectives: (i) to test the sensitivity of
Milner & Petts' conceptual model concerning the
zonation of stream chironomids in glacier-fed rivers
and (ii) to examine whether longitudinal succession of
the chironomid assemblages in glacial streams follows
the same pattern across altitudinal and latitudinal
gradient. This is the ®rst investigation of chironomid
assemblages based upon uni®ed sampling and ana-
lytical procedures at a range of sites along a wide
latitudinal gradient.
Methods
Study sites
The study was carried out in six glacier-melt dominated
streams, the Taillon (TAI, French Pyrenees), the Conca
(CON, Italian Alps), the Mutt (MUT, Swiss Alps),
Daleleva (BRI, Western Norway), W-JoÈkulsa (WJO,
Iceland) and Bayelva (BAY, Svalbard). These streams
form a latitudinal and altitudinal gradient across
Europe from the Pyrenees in the south to Svalbard in
the north. A detailed description of their location and
characteristics is given in other papers in this volume
(Brittain et al., 2001a; Gislason et al., 2001; Lods-Crozet
et al., 2001; Maiolini & Lencioni, 2001; Snook & Milner,
2001) but their principal characteristics are summar-
ized in Table 1. In general the glacial systems studied
were smaller in the southern part of Europe (Pyrennes
and the Alps) than in the northern countries, both
with respect to glacial area and discharge.
In each stream, four permanent study transects
were established within 15-m long sites representing
1792 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

characteristic reaches that were identi®ed on the basis
of valley and channel geomorphology and the com-
position of aquatic communities. The ®rst site was as
close to the glacial snout as possible, and the second
one within 1000 m of the snout and upstream of any
major tributaries. The furthermost downstream site
was chosen where, within zoogeographical con-
straints, a fully developed invertebrate community
occurred [as de®ned in the conceptual model of
Milner & Petts (1994)], i.e. where stone¯ies, caddis¯ies
and may¯ies families were represented together with
chironomids (Orthocladiinae and Chironominae) and
other dipteran families. Four to eight sites were
located in each study catchment to represent the
different channel sectors.
In each river system, except Bayelva on Svalbard,
®eld surveys were carried out during three periods in
both 1996 and 1997: immediately postspring snow-
melt (June), in mid-summer during the ice melt
(August) and at low water level (September). The
snowpack prevented some investigations at upstream
sites in June. As the ice free season on Svalbard is
short, Bayelva was sampled only during early July
and late August 1997.
Geomorphological and environmental variables
Geomorphological descriptions (width of valley
¯oor, width of all active channels, slope) were
carried out at each site. The stream bottom com-
ponent of Pfankuch's index (Pfankuch, 1975) was
used to assess channel stability by scoring ®ve
variables (rock angularity, bed-surface brightness,
particle packing, percentage stable materials, scour-
ing and aquatic vegetation), with high total scores
representing unstable channels at the site scale.
This index gave through a rapid and simple
assessment, a real measure of stream disturbance
because it is strongly correlated both with ¯ood
frequency and degree of bed movement (Townsend
et al., 1997).
During a 5 day sampling period during each ®eld
survey, water temperature, water level, discharge,
conductivity and suspended solids were monitored at
minimum and maximum ¯ows at the upstream and
downstream sites. Discharge was determined with a
¯ow meter (depth±velocity transects), at a gauging
station or by the salt dilution method (Hongve, 1987).
Water temperature was monitored continously atTab
le1
Lo
cati
on
and
char
acte
rist
ics
of
the
gla
cier
-fed
riv
ers
inv
esti
gat
edin
the
AA
SE
Rp
roje
ct
Riv
er
syst
emC
od
eR
egio
nC
o-o
rdin
ates
Max
.al
titu
de
of
catc
hm
ent
(ma.
s.l.
)
Cat
chm
ent
area
at
do
wn
stre
am
reac
h(k
m2)
Gla
cier
area
(km
2)
Pre
cip
itat
ion
1996
/97
(mm
)
Nu
mb
er
of
stu
dy
reac
hes
Dis
tan
ceo
f
reac
hes
fro
m
gla
cier
(m)
Alt
itu
din
al
ran
ge
of
riv
er
stu
die
d
(ma.
s.l.
)
Dis
char
ge
ran
ge
(m3
s)1)
Tre
elin
e
alti
tud
e
(ma.
s.l.
)
Tai
llo
nT
AI
Py
ren
ees
43°0
6'N
,0°
01'W
2975
6.4
0.2
²4
50±1
500
1870
±250
00.
3±0.
416
00
Co
nca
CO
NS
.E.
Alp
s46
°06'
N,
10°3
6'E
3463
4.2
0.2
1497
735
0±46
0013
00±2
830
0.3±
2.1
2000
Mu
ttM
UT
N.
Alp
s46
°33'
N,
8°24
'E30
997
0.6
1595
55±
3600
1800
±260
00.
6±1.
520
00
Dal
elv
aB
RI
W.
No
rway
61°4
0'N
,6°
50'E
1915
25.6
22*
1270
510
0±71
0010
±340
0.5±
3560
0
W±J
ok
u Èls
aÂW
JOC
entr
alIc
elan
d64
°50'
N,
18°4
5'W
1800
840
6866
211
5±45
000
160±
860
25±3
1N
otr
ees³
Bay
elv
aB
AY
Sp
itsb
erg
en,
Sv
alb
ard
78°5
5'N
,11
°50'
E74
230
.912
483
430
0±29
005±
500.
01±1
5.6
No
tree
s
*Par
to
fa
larg
ep
late
aug
laci
erw
ith
anar
eao
f48
7k
m2.
²c.
300
mm
reco
rded
du
rin
gJu
lyan
dA
ug
ust
.³T
he
nat
ura
ltr
eeli
ne,
wit
ho
ut
anth
rop
og
enic
imp
acts
,w
ou
ldb
e30
0±40
0m
a.s.
l.
Chironomid communities in glacial streams 1793
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

most river systems by digital loggers (details in other
papers in this volume).
At each of the four transects within each site, the
wetted channel width to a maximum depth of 0.5 m
and the depth/velocity pro®le were assessed. The
bed-sediment composition was assessed visually or
by contact at each point of the depth/velocity
pro®les.
Biological sampling
Within each sampling site, ten standardized kick
samples (30 s) were collected for invertebrates using a
standard pond net (30 cm ´ 30 cm) with a mesh size of
250 lm. In the case of the Icelandic system, ten stones
were sampled at each site and macroinvertebrates
rinsed off, the surface area of each stone was traced on a
paper and its area used when calculating the inverteb-
rate density (Gislason, OÂ lafsson & Adalsteinsson,
1998). At the kick sample scale, the dominant sediment
particle size, water depth and six consecutive measures
of ¯ow velocity were taken. In addition, three stones
were collected at random in each site and benthic algae
were scraped off on the upper surface (area
3 cm ´ 3 cm) and washed onto a Whatman GF/C ®lter
paper. Chorophyll a was determined spectrophoto-
metrically in the laboratory (APHA, 1992).
The chironomid material used for this study ori-
ginates only from larvae and pupae collected by kick
or stone samples in the streams. The animals from ®ve
to ten kick samples and all stone samples were sorted,
counted and identi®ed. Random subsampling was
adopted when chironomid abundance exceeded 30
and 100 for Norwegian and Icelandic rivers, respect-
ively.
Within each subfamily, a taxonomic limit was
imposed by the larvae and further identi®cation with
pupae was ignored, unless all pupae belonged to one
species. In that case, the larvae were also ascribed to
this species. On account of taxonomic problems with
Diamesa larvae, it was decided to establish eight dis-
crete groups which were: (1) Diamesa aberrata gr.; (2)
D. bertrami Edwards; (3) D. damp® gr.; (4) D. davisi gr.;
(5) D. latitarsis gr.; (6) D. steinboecki Goetgh.; (7)
D. zernyi gr./cinerella gr.; (8) Diamesa gr. A:. which
included the ®rst and second instars of D. bertrami,
D. davisi gr., D. latitarsis gr. and D. steinboecki. Iden-
ti®cation keys and species descriptions were selected
from the European literature (Thienemann, 1952;
Serra-Tosio, 1967; Saether, 1968; Rossaro, 1980, 1981,
1982; Ferrarese & Rossaro, 1981; Cranston, 1982;
Ferrarese, 1983; Wiederholm, 1983; Wiederholm,
1986; Makarchenko, 1985; Nocentini, 1985; Langton,
1991; Schmid, 1993; Janecek, 1998). The chironomids
from the Pyrenees system were not identi®ed to the
same extent as at the other sites and were therefore not
included in multivariate analyses. Abbreviations of
species names were used according to a standardized
coding system developed by Schnell et al. (1999).
Data analysis
The elementary units used in all the analyses were
site data on a given date, i.e. the average of 5±10
replicate kick or stone samples. The quantitative
faunal data, expressed as the number of individuals
m±2, were processed by correspondence analysis
(CA), which provides a reciprocal ordination of the
species and the sampling units (106 selected sites). All
the data on chironmid densities were [log10(x + 1)]
transformed prior to analysis. A cluster analysis by
Ward's method (Ward, 1963), which grouped samp-
ling sites according to the similarity of their chiron-
omid assemblages, was carried out using the factorial
scores of the sites in the previous CA as a summary of
the faunal data. Following Roux (1991), the contribu-
tion (CV(j,p)) of each taxon (j) to each site cluster (p)
was calculated as:
�CV�j;p�� � �Zpj ÿ Zj�2=Rj�Zpj ÿ Zj�2
with Zpj � average of taxon j in cluster p, Zj � overall
average of taxon j.
Canonical correspondence analysis (CCA) (ter
Braak, 1986) was used to determine relationships
between the environmental properties of each site
and their respective chironomid composition. In the
CCA biplots, axes represent the most important
environmental gradients along which the chirono-
mid fauna was distributed. The signi®cance of the
CCA axes was determined by Monte Carlo permu-
tation testing (1000 permutations) of the eigenvalues
(Fraile, Escou®er & Raibaut, 1993). The variations in
abundance of the most characteristic chironomid
taxa in relation to temperature were described using
second order polynomial regression curves. All cal-
culations were carried out using the ADE-4 software
(Thioulouse et al., 1997).
1794 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809
Results
Habitat characteristics
A summary of the physico-chemical and biological
data in the six glacial streams is given in Table 2.
The site slope of the six systems decreased with the
altitude range of the glacial rivers and with the
latitudinal gradient. Pfankuch's Index of channel
stability varied from 58 (highly unstable stream
bottom) to 18 (stable stream bed). Conductivity was
very low for the Italian and Norwegian systems
(£13 and £10 lS cm±1, respectively), whereas the four
other rivers had conductivities up to 80 and even
140 lS cm±1 (Swiss Alps). Algal biomass, measured as
chlorophyll a concentration, reached maximum values
of 10.0 mg m±2 in most of the glacial systems except
in Dalelva and Bayelva where the concentrations
reached 30.6 (BRI02) and 19.3 (BAY04) mg m±2,
respectively. The Alpine systems (Pyrenees and the
Alps) were dominated by boulders and cobbles while
the Dalelva, W-JoÈkulsa and Bayelva sites were char-
acterized by gravel, sand and silt.
Mean current velocities were comparable in the
Alpine and Pyrenean sites (Taillon, Conca and Mutt:
Table 2 Summary of physical, chemical and periphyton variables at the site scale (�SD: interdate variation) for the study streams
Site
no.
Distance
from the
snout (m)
Site
slope
(m m)1)
Water
width
(m)
Water
depth
(m)
Pfankuch
Index
Speci®c
conductivity
(lS cm)1 at 20 °C)
Chlorophyll a
(mg m)2)
TAI01 50 0.079 1.8 (0.03) 0.14 (0.01) 41 72.2 (24.7) 0.91
TAI02 900 0.085 2.0 (0.25) 0.09 (0.02) 30 80.6 (21.9) 1.91 (0.85)
TAI03 1425 0.041 3.3 (0.06) 0.14 (0.03) 32 77.3 (20.0) 3.59 (1.05)
TAI35 1500 0.03 4.7 (2.0) 0.16 (0.07) 21 84.0 (41.5) 1.23 (1.23)
CON00 310 0.40 0.7 0.07 18 7.0 (4.4) 2.12 (1.10)
CON01 710 0.13 1.5 0.06 18 7.9 (5.8) 6.77 (3.55)
CON02 1070 0.28 1.8 0.07 32 7.4 (2.6) 6.66 (5.85)
CON03 1520 0.22 2.3 0.07 25 8.5 (2.6) 1.06 (0.56)
CON04 2600 0.02 6.3 0.14 34 10.1 (0.9) 2.60 (1.59)
CON05 2740 0.07 4.8 0.14 27 10.0 (0.9) 2.93 (0.75)
CON08 4600 0.13 15.7 0.13 19 10.2 (2.1) 2.34 (1.28)
MUT01 5 0.1 3.5 (3.4) 0.05 (0.02) 58 27.4 (0.4) 0.16 (0.14)
MUT02 200 0.24 5.1 (3.37) 0.04 (0.01) 38 66.8 (37.8) 0.57 (0.68)
MUT03 350 0.3 7.9 (1.94) 0.06 (0.01) 37 70.6 (24.1) 1.87 (1.07)
MUT04 1700 0.17 5.2 (0.55) 0.16 (0.04) 33 139.0 (49.6) 1.86 (0.22)
MUT05 3600 0.18 4.6 (1.6) 0.21 (0.06) 29 118.6 (36.5) 3.54 (0.15)
BRI01 100 0.034 18 0.25 (0.22) 36 8.2 (3.3) 28.16 (31.74)
BRI02 700 0.015 7 0.30 (0.20) 55 6.6 (4.8) 30.64 (7.67)
BRI03 2000 0.070 8.5 0.31 (0.13) 39 7.0 (4.9) 20.75 (6.41)
BRI04 3300 0.021 15.1 0.31 (0.13) 51 9.3 (4.6) 12.01 (6.39)
BRI05 7100 0.006 9.3 0.49 (0.14) 41 9.1 (4.2) 22.62 (21.10)
WJO01 5 0.005 3.5 (1.0) 0.07 (0.029) 55 10.1 (0.3) 0.09
WJO15 500 0.013 5.1 (0.14) 0.18 (0.03) 34 8.1 (0.2) 0.46 (0.44)
WJO1c 3500 0.004 8.5 (0.71) 0.20 (0.07) 51 28.6 (26.4) 0.44 (0.47)
WJO02 1400 0.006 6.0 (1.4) 0.19 (0.07) 38 16.3 (8.8) 0.78 (0.55)
WJO2b 3000 0.005 12.7 (3.16) 0.44 (0.06) 47 49.6 (6.1) 0.78 (1.32)
WJO2c 22 000 0.007 8.2 (3.9) 0.25 (0.07) 44 77.5 (13.1) 0.05 (0.02)
WJO03 4500 0.012 8.4 (2.1) 0.39 (0.09) 28 27.5 (15.5) 0.34 (0.08)
WJO35 7500 0.008 16.8 (0.28) 0.23 (0.04) 41 17.3 (5.7) 0.36 (0.38)
WJO04 22 500 0.006 36.0 (2.5) 0.44 (0.07) 39 56.3 (17.4) 0.08 (0.03)
WJO05 42 000 0.014 27.5 (1.0) 0.55 (0.07) 21 81.4 (12.8) 1.24 (0.93)
WJO06 45 000 0.009 34.0 (1.0) 0.60 (0.10) 35 72.1 (20.2) 0.31 (0.21)
BAY01 300 0.024 10.2 (5.0) 0.21 (0.02) 38 37.6 (10.4) 0.12 (0.002)
BAY02 1200 0.006 31.3 (1.66) 0.18 (0.01) 54 49.2 (12.4) 0.96 (0.96)
BAY03 1800 0.006 38.1 (21.0) 0.17 (0.04) 58 63.5 (23.3) 5.3 (4.7)
BAY04 2900 0.008 22.2 (7.42) 0.17 (0.01) 44 81.7 (28.6) 19.3 (8.7)
TAI: Taillon (Pyrennees); CON: Conca (Southern Alps); MUT (Northern Alps); BRI: Dalelva (W-Norway); WJO: W-JoÈkulsa (Iceland);
BAY: Bayelva (Svalbard).
Chironomid communities in glacial streams 1795
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

0.60 � 0.22 m s±1); higher current velocities were
recorded in Bayelva (0.82 � 0.14 m s±1). Low values
of suspended sediment were recorded in the Italian
stream (mean of 8.1 mg L±1), followed by the Nor-
wegian (30.6 mg L±1), Pyreneean (45.2 mg L±1) and
Svalbard (86.7 mg L±1) streams, but much higher
values were found in the Swiss and Icelandic streams
(797.5 and 457.1 mg L±1, respectively).
At the upstream sites, the mean and maximum
water temperature was almost always below 2 °C.
The water temperature recorded during the faunal
surveys was signi®cantly correlated with the maxi-
mum temperature (R2 � 0.61, P < 0.001); it was the
only temperature variable retained in the mutivariate
analysis as it was available for all sites and dates.
Latitudinal distribution of chironomid assemblages
A decrease in chironomid taxa richness with latitude
from the Alps to the Svalbard was observed (Table 3).
The most taxa rich subfamilies in the six glacier-fed
rivers were Diamesinae and Orthocladiinae (12 and 42
taxa, respectively). In terms of richness, the Diame-
sinae : Orthocladiinae ratio was relatively constant
(30±40%) across the European latitudinal gradient.
At the generic and species level, the highest taxa
richness was recorded in the Italian Alps (42 taxa) and
the lowest in Bayelva (seven taxa). From a total of 63
taxa (Appendix 1), two species, D. bertrami and
Orthocladius (O.) frigidus (Zetter.), had the widest
latitudinal distribution, only being absent in Bayelva
(Svalbard). In terms of mean chironomid abundance,
no distinct patterns were apparent along the latitudi-
nal gradient (Table 3). The highest mean chironomid
densities were recorded in the Northern Alps (Mutt),
followed by the Central Icelandic river, the Western
Norwegian one, the Pyrenees (Taillon), the Italian
Alps and Svalbard. Diamesinae accounted for 83±97%
of the total abundance except in the Italian and
Icelandic systems where they remained below 50%.
The contribution of Chironominae and Tanypodinae
was low, both in terms of density and species
richness.
Ordination and classi®cation of river systems basedon their chironomid assemblages
Five river systems were considered in the following
analyses (Conca, Mutt, Dalelva, W-JoÈkulsa and
Bayelva). Ordination of the 63 taxa by 106 sites/dates
data matrix using CA demonstrated a strong stream-
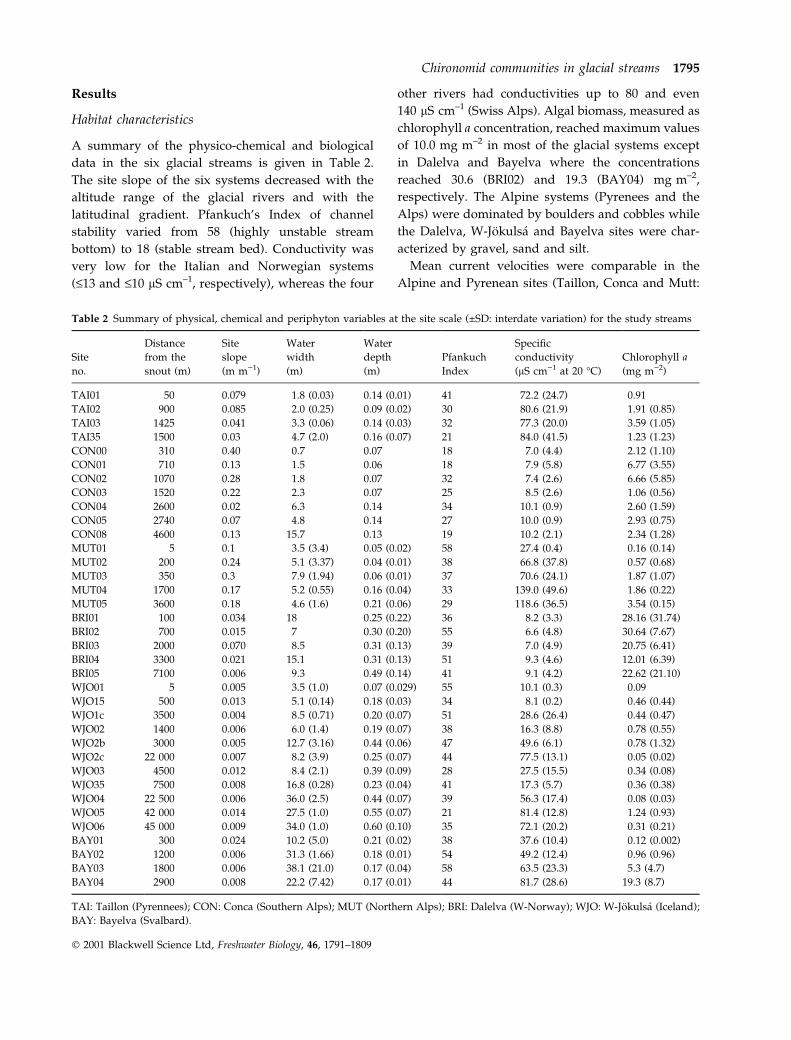
speci®c effect along the ®rst two factorial axes (Fig. 1).
The species-poor Svalbard system occupied the central
position of the ordination. The ®rst axis divided the
Italian system from the three others while the second
axis spread the Swiss, Norwegian and Icelandic ones.
Table 4 summarizes the seven groups of sites
resulting from the cluster analysis with typical taxa
found in each group. This classi®cation separated
the glacial systems on both a geographical-latitudi-
nal and a downstream gradient. Diamesa steinboecki,
D. latitarsis gr. and Diamesa gr. A (group 1) character-
ized the uppermost sites (c. the ®rst 350 m) of the
Alpine streams. Two groups (2 and 3) split the Italian
system into its upstream and downstream sites. In
group 2, O. rivicola gr., D. steinboecki, Pseudokiefferiella
parva (Edwards) were the most dominant taxa bet-
ween 750 and 1500 m from the glacier snout. Group 3
comprised the downstream chironomid communities
characterized by taxa such as Micropsectra atrofasciata
(Kieff.) Tvetenia spp. and Eukiefferiella brevicalcar
(Kieff.)/tirolensis Goetgh. Group 4 comprised four
Table 3 Taxonomic richness and mean abundance expressed as individuals m±2 � 95% Con®dence Level (number of kick
samples) of Chironomidae in the six glacial streams
TAI CON MUT BRI WJO BAY Total
Taxonomic richness 27 42 29 20 20 7 63
Diamesinae 6 9 8 5 4 2 12
Orthocladiinae 16 30 20 14 13 5 42
Chironominae 4 2 1 0 2 0 6
Tanypodinae 1 1 0 1 1 0 3
Mean abundance 1306 � 469 (100) 636 � 106 (209) 2569 � 597 (216) 2010 � 526 (66) 2163 � 511 (233) 114 � 67 (23)
Diamesinae 1085 � 379 215 � 79 2457 � 589 1944 � 520 1005 � 237 100 � 69
Orthocladiinae 192 � 68 301 � 60 111 � 44 65 � 34 1100 � 390 14 � 7
Chironominae 29 � 22 114 � 32 0.5 � 0.3 0 58 � 41 0
1796 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809
taxa characteristic of the Mutt system in the Swiss
Alps: Diamesa latitarsis gr., Orthocladius spp., O. (E.)
luteipes Goetgh. and Paraphaenocladius spp. Six taxa
predominated group 5, which included the whole
Dalelva glacial system, and the upstream sites of the
W-JoÈkulsa (c. from 5 m to 7.5 km distance downstream
from the snout) with D. bertrami, D. latitarsis gr. and
D. bohemani Goetgh./zernyi Edwards. Group 6 encom-
passed the downstream Icelandic sites where E. minor
(Edwards), E. claripennis gr., Orthocladius (O.) frigidus
and Diamesinae juveniles dominated the chironomid
assemblages. The Svalbard glacial stream, Bayelva,
formed the seventh group with three distinct taxa:
Thienemannia spp., Cricotopus fuscus gr. and D. aberrata
gr. However, two sites from Bayelva (BAY02 in July
and BAY03 in August 1997) belong to groups 2 and 3
because of their unindenti®ed juvenile Orthocladiinae.
Longitudinal patterns of chironomid assemblages
downstream of glacial margins
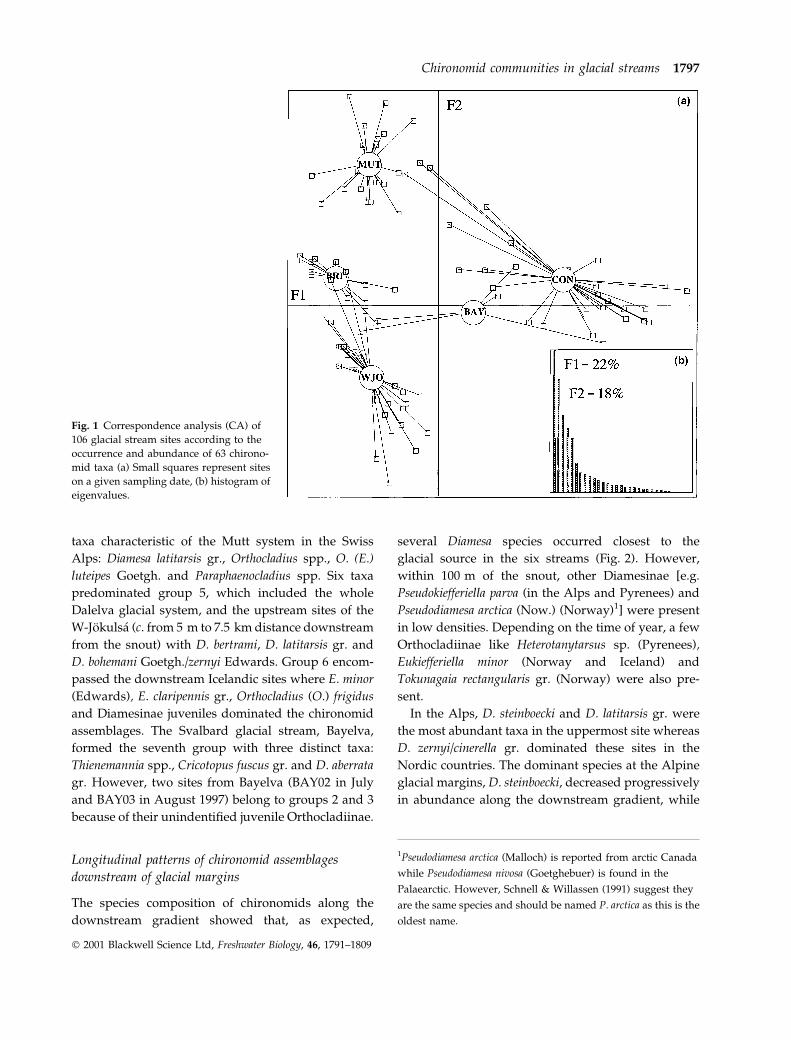
The species composition of chironomids along the
downstream gradient showed that, as expected,
several Diamesa species occurred closest to the
glacial source in the six streams (Fig. 2). However,
within 100 m of the snout, other Diamesinae [e.g.
Pseudokiefferiella parva (in the Alps and Pyrenees) and
Pseudodiamesa arctica (Now.) (Norway)1] were present
in low densities. Depending on the time of year, a few
Orthocladiinae like Heterotanytarsus sp. (Pyrenees),
Eukiefferiella minor (Norway and Iceland) and
Tokunagaia rectangularis gr. (Norway) were also pre-
sent.
In the Alps, D. steinboecki and D. latitarsis gr. were
the most abundant taxa in the uppermost site whereas
D. zernyi/cinerella gr. dominated these sites in the
Nordic countries. The dominant species at the Alpine
glacial margins, D. steinboecki, decreased progressively
in abundance along the downstream gradient, while
Fig. 1 Correspondence analysis (CA) of
106 glacial stream sites according to the
occurrence and abundance of 63 chirono-
mid taxa (a) Small squares represent sites
on a given sampling date, (b) histogram of
eigenvalues.
1Pseudodiamesa arctica (Malloch) is reported from arctic Canada
while Pseudodiamesa nivosa (Goetghebuer) is found in the
Palaearctic. However, Schnell & Willassen (1991) suggest they
are the same species and should be named P. arctica as this is the
oldest name.
Chironomid communities in glacial streams 1797
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809
the other Diamesa species were still found up to 4 km
downstream from the glacier.
In the subfamily Orthocladiinae, the colonists to
become established closest to the glacier were
O. frigidus (Pyrenees, Swiss Alps, Iceland), O. rivicola gr.
(Italian Alps, Svalbard), Heleniella sp. (Italian Alps),
Eukiefferiella minor/®ttkaui and claripennis gr. (Italian
Alps, Norway, Iceland), Parametriocnemus stylatus Kieff.
(Pyrenees), Tvetenia sp. (Italian and Swiss Alps) and
Thienemannia sp. (Svalbard).
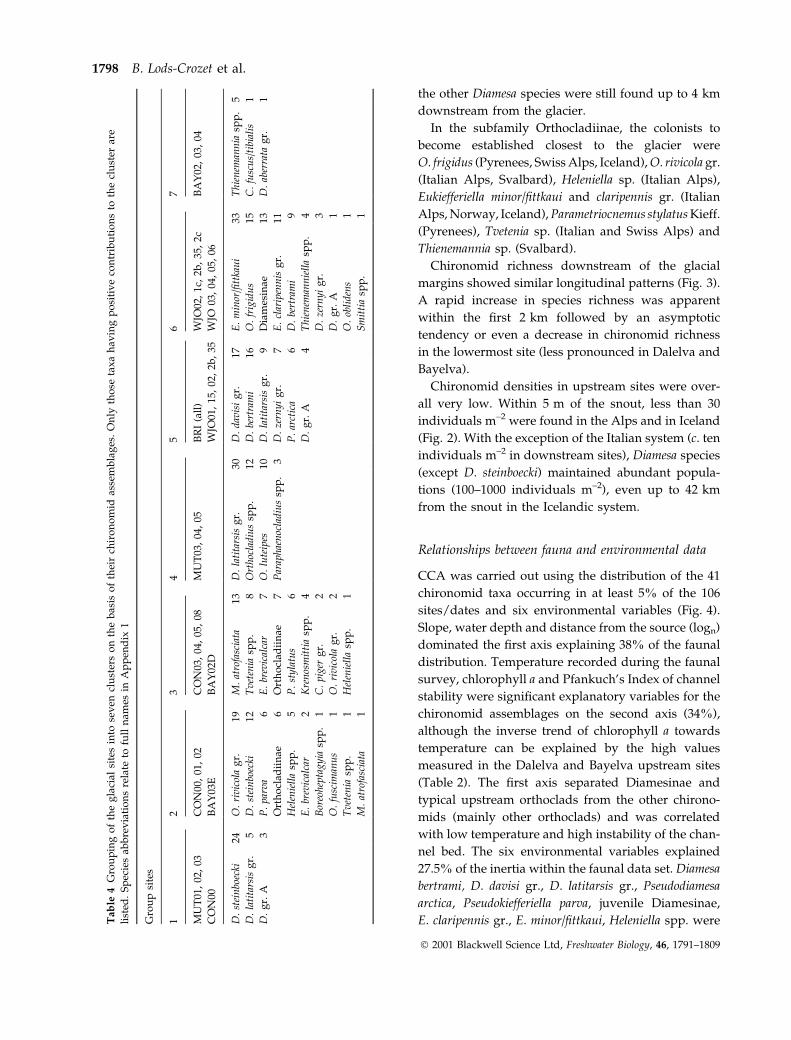
Chironomid richness downstream of the glacial
margins showed similar longitudinal patterns (Fig. 3).
A rapid increase in species richness was apparent
within the ®rst 2 km followed by an asymptotic
tendency or even a decrease in chironomid richness
in the lowermost site (less pronounced in Dalelva and
Bayelva).
Chironomid densities in upstream sites were over-
all very low. Within 5 m of the snout, less than 30
individuals m±2 were found in the Alps and in Iceland
(Fig. 2). With the exception of the Italian system (c. ten
individuals m±2 in downstream sites), Diamesa species
(except D. steinboecki) maintained abundant popula-
tions (100±1000 individuals m±2), even up to 42 km
from the snout in the Icelandic system.
Relationships between fauna and environmental data
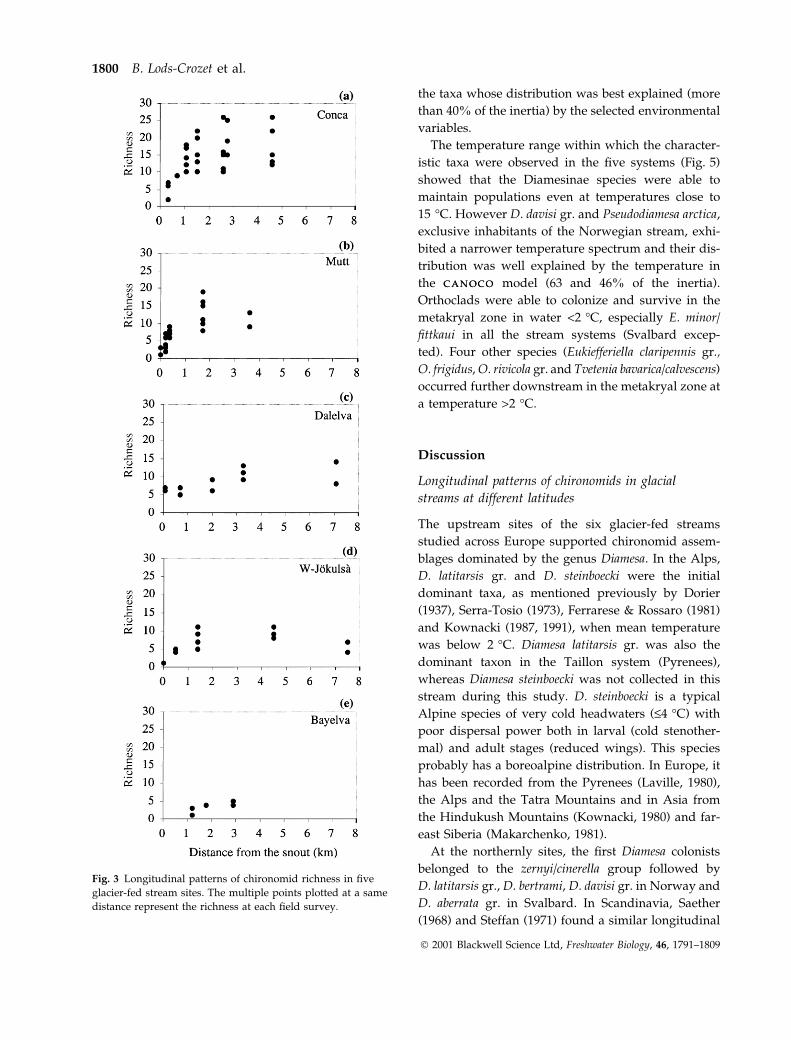
CCA was carried out using the distribution of the 41
chironomid taxa occurring in at least 5% of the 106
sites/dates and six environmental variables (Fig. 4).
Slope, water depth and distance from the source (logn)
dominated the ®rst axis explaining 38% of the faunal
distribution. Temperature recorded during the faunal
survey, chlorophyll a and Pfankuch's Index of channel
stability were signi®cant explanatory variables for the
chironomid assemblages on the second axis (34%),
although the inverse trend of chlorophyll a towards
temperature can be explained by the high values
measured in the Dalelva and Bayelva upstream sites
(Table 2). The ®rst axis separated Diamesinae and
typical upstream orthoclads from the other chirono-
mids (mainly other orthoclads) and was correlated
with low temperature and high instability of the chan-
nel bed. The six environmental variables explained
27.5% of the inertia within the faunal data set. Diamesa
bertrami, D. davisi gr., D. latitarsis gr., Pseudodiamesa
arctica, Pseudokiefferiella parva, juvenile Diamesinae,
E. claripennis gr., E. minor/®ttkaui, Heleniella spp. wereTab
le4
Gro
up
ing
of
the
gla
cial
site
sin
tose
ven
clu
ster
so
nth
eb
asis
of
thei
rch
iro
no
mid
asse
mb
lag
es.
On
lyth
ose
tax
ah
avin
gp
osi
tiv
eco
ntr
ibu
tio
ns
toth
ecl
ust
erar
e
list
ed.
Sp
ecie
sab
bre
via
tio
ns
rela
teto
full
nam
esin
Ap
pen
dix
1
Gro
up
site
s
12
34
56
7
MU
T01
,02
,03
CO
N00
CO
N00
,01
,02
BA
Y03
E
CO
N03
,04
,05
,08
BA
Y02
D
MU
T03
,04
,05
BR
I(a
ll)
WJO
01,
15,
02,
2b,
35
WJO
02,
1c,
2b,
35,
2c
WJO
03,
04,
05,
06
BA
Y02
,03
,04
D.
stei
nbo
ecki
24O
.ri
vico
lag
r.19
M.
atro
fasc
iata
13D
.la
tita
rsis
gr.
30D
.da
visi
gr.
17E
.m
inor
/®tt
kau
i33
Thi
enem
ann
iasp
p.
5
D.
lati
tars
isg
r.5
D.
stei
nbo
ecki
12T
vete
nia
spp
.8
Ort
hocl
adiu
ssp
p.
12D
.be
rtra
mi
16O
.fr
igid
us
15C
.fu
scu
s/ti
bial
is1
D.
gr.
A3
P.
parv
a6
E.
brev
ical
car
7O
.lu
teip
es10
D.
lati
tars
isg
r.9
Dia
mes
inae
13D
.ab
erra
tag
r.1
Ort
ho
clad
iin
ae6
Ort
ho
clad
iin
ae7
Par
apha
enoc
ladi
us
spp
.3
D.
zern
yig
r.7
E.
clar
ipen
nis
gr.
11
Hel
enie
lla
spp
.5
P.
styl
atu
s6
P.
arct
ica
6D
.be
rtra
mi
9
E.
brev
ical
car
2K
ren
osm
itti
asp
p.
4D
.g
r.A
4T
hien
eman
nie
lla
spp
.4
Bor
eohe
ptag
yia
spp
.1
C.
pige
rg
r.2
D.
zern
yig
r.3
O.
fusc
iman
us
1O
.ri
vico
lag
r.2
D.
gr.
A1
Tve
ten
iasp
p.
1H
elen
iell
asp
p.
1O
.ob
lide
ns
1
M.
atro
fasc
iata
1S
mit
tia
spp
.1
1798 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

Fig. 2 Longitudinal patterns of chironomid assemblages in glacial streams during two summer periods. Species abbreviations
relate to full names in Appendix 1.
Chironomid communities in glacial streams 1799
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809
the taxa whose distribution was best explained (more
than 40% of the inertia) by the selected environmental
variables.
The temperature range within which the character-
istic taxa were observed in the ®ve systems (Fig. 5)
showed that the Diamesinae species were able to
maintain populations even at temperatures close to
15 °C. However D. davisi gr. and Pseudodiamesa arctica,
exclusive inhabitants of the Norwegian stream, exhi-
bited a narrower temperature spectrum and their dis-
tribution was well explained by the temperature in
the CANOCOCANOCO model (63 and 46% of the inertia).
Orthoclads were able to colonize and survive in the
metakryal zone in water <2 °C, especially E. minor/
®ttkaui in all the stream systems (Svalbard excep-
ted). Four other species (Eukiefferiella claripennis gr.,
O. frigidus, O. rivicola gr. and Tvetenia bavarica/calvescens)
occurred further downstream in the metakryal zone at
a temperature >2 °C.
Discussion
Longitudinal patterns of chironomids in glacialstreams at different latitudes
The upstream sites of the six glacier-fed streams
studied across Europe supported chironomid assem-
blages dominated by the genus Diamesa. In the Alps,
D. latitarsis gr. and D. steinboecki were the initial
dominant taxa, as mentioned previously by Dorier
(1937), Serra-Tosio (1973), Ferrarese & Rossaro (1981)
and Kownacki (1987, 1991), when mean temperature
was below 2 °C. Diamesa latitarsis gr. was also the
dominant taxon in the Taillon system (Pyrenees),
whereas Diamesa steinboecki was not collected in this
stream during this study. D. steinboecki is a typical
Alpine species of very cold headwaters (£4 °C) with
poor dispersal power both in larval (cold stenother-
mal) and adult stages (reduced wings). This species
probably has a boreoalpine distribution. In Europe, it
has been recorded from the Pyrenees (Laville, 1980),
the Alps and the Tatra Mountains and in Asia from
the Hindukush Mountains (Kownacki, 1980) and far-
east Siberia (Makarchenko, 1981).
At the northernly sites, the ®rst Diamesa colonists
belonged to the zernyi/cinerella group followed by
D. latitarsis gr., D. bertrami, D. davisi gr. in Norway and
D. aberrata gr. in Svalbard. In Scandinavia, Saether
(1968) and Steffan (1971) found a similar longitudinal
Fig. 3 Longitudinal patterns of chironomid richness in ®ve
glacier-fed stream sites. The multiple points plotted at a same
distance represent the richness at each ®eld survey.
1800 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

pattern to the Alps (e.g. D. lindrothi and D. valkanovi
both in the latitarsis gr and D. davisi gr., a group of
species close to D. steinboecki).
Few studies provide quantitative data on Diamesa
abundance downstream from glacial margins (Sae-
ther, 1968; Kownacki, 1991). A striking result of our
study was the abundant Diamesa populations (100±
1000 individuals m±2) even far from the snout and
even when several other invertebrates had become
abundant (Fig. 2). The Conca system was different in
that mean chironomid density (636 individuals m±2)
was low compared with the other stream systems
(except Svalbard) with a net decrease of Diamesa
species at the downstream sites. Similar densities (550
individuals m±2) were recorded by Kownacki (1991) in
South Tyrolian streams. One other difference high-
lighted by the cluster analysis was the splitting of the
Conca system into distinct upstream (C0, C1, C2) and
downstream (C3, C4, C5, C8) chironomid communi-
ties (Table 4). This re¯ected signi®cant differences in
water temperature (minimum, maximum and mean),
slope, water depth and discharge (Lencioni, 2000;
Maiolini & Lencioni, 2001).
Two others Diamesinae species were found in cold
headwaters with a mean temperature range of
0±10 °C: Pseudokiefferiella parva (in the Alps and
Fig. 4 C A N O C OC A N O C O model of the distribution of 41 chironomid taxa and six environmental variables (a) The histogram displays the
inertia accounted for by each factorial axis (b) Species abbreviations relate to full names in Appendix 1.
Chironomid communities in glacial streams 1801
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809
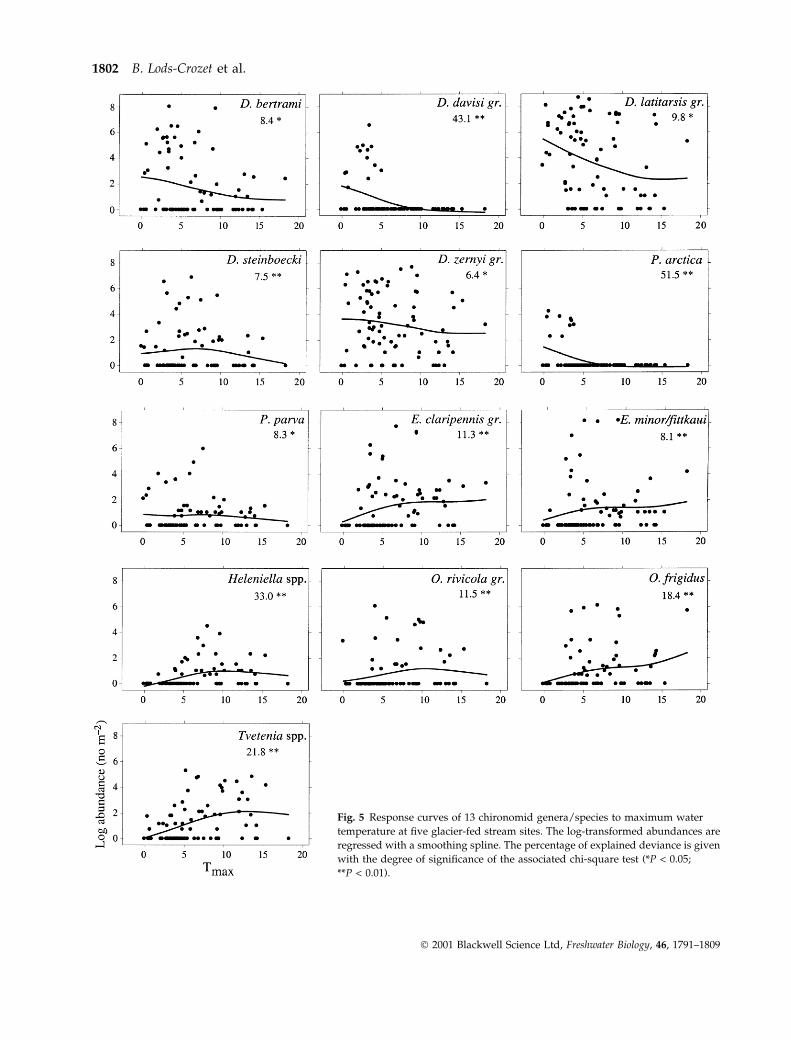
Fig. 5 Response curves of 13 chironomid genera/species to maximum water
temperature at ®ve glacier-fed stream sites. The log-transformed abundances are
regressed with a smoothing spline. The percentage of explained deviance is given
with the degree of signi®cance of the associated chi-square test (*P < 0.05;
**P < 0.01).
1802 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

Pyrenees) and Pseudodiamesa arctica (Norway). They
have not been previously reported in kryal sites close
to glacial snouts, but have mostly been collected in
spring-fed streams where P. parva colonizes mosses
(Serra-Tosio, 1973; Kownacki, 1987; Ilg et al., 2001).
Pseudodiamesa arctica/nivosa is also characteristic of
high mountain lakes (Kownacki, 1987; Schnell &
Willassen, 1991).
The presence of Orthocladiinae species in associ-
ation with Diamesa at low temperature (<2 °C) and
very close to the snout (metakryal zone) has not been
previously documented. Eukiefferiella species and
O. rivicola gr. were the ®rst Orthocladiinae taxa
collected at mean temperature above 4 °C and about
1000 m from the snout (Saether, 1968; Kawecka et al.,
1978; Kownacki, 1991). Ward (1994) stated that in the
hypokryal (where Tmax exceeds 2 °C) other chirono-
mid genera and simuliids may occur. In our six
systems, Eukiefferiella, Orthocladius and Tvetenia spe-
cies were able to colonize the ®rst 200 m at mean
temperatures close to 0 °C. A similar pattern was
observed in glacial streams of Alaska by Milner (1994)
where an Orthocladius species was present at 2 °C.
Mountain tops may be viewed as islands in the
context of island biogeography theory (McArthur &
Wilson, 1967; Ward, 1994). Alpine streams are often
small isolated habitats compared with northern
European and arctic glacial streams originating from
extensive ice-caps. This evolutionary isolation in
combination with Ice Age refugia in the Alps (espe-
cially the south), is perceptible in the decrease of the
chironomid richness along the latitudinal gradient
although many species of Diamesinae and Orthocla-
diinae have broad geographical distribution patterns.
The successional sequence of chironomid assem-
blages along latitudinal gradients in glacial streams
follows the same pattern, with similar genera, groups
of species and species involved. Diamesa bertrami was
the most widespread species, collected in all studied
river systems, but previously recorded only on Sval-
bard in the surroundings of Ny-AÊ lesund (Serra-Tosio,
1973). These results are consistent with the trend of
colonization and successional sequence as suggested
by Milner & Petts (1994) and observed by Milner
(1994) in Alaska. These apparently predictable pat-
terns are, at least in part, a re¯ection of the harsh
physical conditions that greatly limit the numbers and
types of species that form the colonizing species pool
for glacial streams at different latitudes (Milner, 1994).
Life strategies
Chironomids of glacier-fed streams tend to display
melanism in all stages (mostly black or dark brown)
and some species of Diamesa (e.g. D. steinboecki gr.,
D. davisi gr.) have developed other strategies for cold
environments such as wing reduction, reduction in
the size of the male antennal plume and antennal ratio
and enlargement of the legs (Saether, 1968; Herrmann
et al., 1987). Brachypterous specimens (i.e. with
reduced wings) of D. steinboecki were only collected
in the Swiss Alps during this study.
Recently, the prevalence of life-cycle ¯exibility
or opportunism has been emphasized for polar
arthropods (Danks, 1999), allowing growth and
development whenever conditions are favourable.
Chironomids are one of the taxa that have devel-
oped strategies to survive the harsh conditions
prevailing in these extreme environments, such as
acceleration of the emergence and reproduction pro-
cesses, resting stages (diapause) or temperature
thresholds for developmental control (Danks &
Oliver, 1972a, b). Evidence of short life cycles was
emphasized by Serra-Tosio (1973), documenting that
in cold Alpine streams above 1800 m a.s.l., D. aberrata
and D. zernyi completed their life cycle in less than
40 days. Diamesa tonsa emerged throughout the winter
at 0 °C in an western Norwegian river (Jonsson &
Sandlund, 1975). Diamesa incallida, a krenal species,
lays eggs throughout the year and produces 8±10
generations depending on water temperature (Nolte
& Hoffmann, 1992). Experimental in situ growth
chambers used at an altitude of 2200 m in the Taillon
stream (average water temperature of 4.5 °C) demon-
strated that egg hatching to pupation of D. zernyi/
bohemani took between 39 and 54 days in June±July.
Analysis of larval head capsule widths suggested that
D. bertrami and D. latitarsis was bivoltine with emer-
gence periods in early June and August/September
(Snook, 2000).
Milner (1994) suggested that Diamesa species are
eliminated through competition with other species in
relatively stable channels, and dominate in unstable
channels because of the scarcity of competitors. In
Alaska, D. davisi was considered a fugitive species,
restricted to areas of low density of other taxa, either
because of low temperatures or frequent disturbance,
rather than being physiologically inhibited by high
temperatures (Flory & Milner, 1999). However, in
Chironomid communities in glacial streams 1803
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

Dalelva (Norway), D. davisi gr., occurred with species
such as E. claripennis gr. at relatively high densities
(cf. Fig. 2) in unstable channels (high values of
Pfankuch's index). Similar patterns are apparent in
the longitudinal faunal gradients in the Swiss Alps
(Mutt) and in Iceland (W-JoÈkulsaÂ) for D. latitarsis gr.
and O. frigidus. It is possible that spatial or temporal
segregation may exist (e.g. hyporheic refugia, differ-
ent periods of development). Alternatively, interspec-
i®c competition may not be a key factor in these
dynamic streams.
Relationships between chironomids
and environmental factors
The Pyreneean, Alpine and Icelandic glacier-fed
streams were characterized by low chlorophyll a
concentrations in the upstream sites compared with
those recorded for Western Norway (Dalelva) and the
downstream site of Bayelva (Western Spitzbergen).
Similarly low values of algal biomass were found in
the Eastern Swiss Alps by Uehlinger, Zah, & Buergi
(1998). In Dalelva, a proglacial lake has almost been
covered by the advancing glacier. This may have
caused nutrients release from the lake sediments and
the surrounding land areas as result of erosion by ice,
thus promoting algal growth in the outlet stream.
Another characteristic that partly applies to Dalelva
but especially to Bayelva, is the closeness to the sea.
Northern and arctic areas also have a signi®cantly
longer daylength during summer. These points cou-
pled with a rather moderate channel stability index in
the upstream site, could explain the presence of the
orthoclads E. minor (1996) and E. claripennis gr.,
E. minor, Limnophyes, Tokunagaia rectangularis gr. and
Orthocladius cf. dentifer in the metakryal zone of
Dalelva during June 1997.
The conceptual model of Milner & Petts (1994)
relates zoobenthic gradients to water temperature and
geomorphological instability, as a function of distance
from the glacial margins. The present study examined
chironomid distributions in relation to geomorpho-
logical and physico-chemical variables and provided
further documentation bearing on the conceptual
model. Fig. 6 summarizes this longitudinal distribu-
tion of key chironomid of the kryal zone of glacier-fed
streams taxa in relation with maximum water tem-
perature. Diamesa species were predicted to decrease
when maximum temperature exceeded 4 °C where
channel stability increased. In the European glacial-
fed streams studied, Diamesa maintained abundant
populations in warmer environments (Tmax 6 15 °C).
Fig. 6 Schematic representation of longitudinal succession of characteristic chironomid taxa in relation to water temperature and
distance from snout in six European glacier-fed streams (Pyrenees to Svalbard).
1804 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

Other Diamesinae (Pseudokiefferiella parva, Pseudodia-
mesa arctica) and the orthoclad species E. minor/®ttkaui
were also able to colonize the metakryal zone
(Tmax 6 2 °C) in the ®rst 100 m from the glacier.
The orthoclads Eukiefferiella claripennis gr., O. frigidus,
O. rivicola gr. and Tvetenia came later downstream at
Tmax > 2 °C.
Algal ®laments such as diatoms and the chryso-
phyte Hydrurus foetidus (Vill.), common in the Alps
and Northern Scandinavia (Ward, 1994; Kawecka et al.,
1978; Lods-Crozet et al., 2001; Maiolini & Lencioni,
2001) provide resources and protection (low shear
stress) from currents and abrasive sediments and can
play a role as refugia for Diamesinae and Orthocladii-
nae in the upper reaches of glacial streams (Fig. 7).
Modi®ers to the conceptual model such as pro-
glacial lakes that reduce turbidity and increased
temperature, channel stability and nutrients induced
a shift in the sequence of occurrence of the chirono-
mid taxa as predicted by Milner & Petts (1994). This
was illustrated in the river Dalelva (Western Norway).
Another predicted in¯uence is the effect of tributaries.
This impact can be very different depending on their
size and discharge, their hydrological origin (snow-
melt, spring-fed or glacial melt) and the distance from
the source. Examples from the Swiss Alps and
western Norway from small snowmelt tributaries
indicate a minor contribution to the main channel
benthos (Petts & Bickerton, 1994; Haug, Salveit &
Brittain, 2001; Ilg et al., 2001).
In conclusion, despite very harsh conditions (low
temperature, narrow thermal heterogeneity, high
turbidity and high channel instability), glacial streams
support a relatively rich chironomid community
(a total of 63 taxa in the six streams studied) with
increasing densities and diversity downstream of the
glacier snout as predicted by Milner & Petts (1994).
Nevertheless, chironomid richness reaches 150 species
if all the headwaters (glacial meltwater, snowmelt and
spring-fed stream) are considered (Brittain et al.,
2001b). This represents about 15% of the total number
of species of Chironomidae known from Europe
(Ashe & Cranston, 1990). In addition, a wider
geographical and ecological distribution of several
species has been documented in relation to previous
studies. Future studies, should also address the need
for progress in chironomid taxonomy. Keys for the
genus Diamesa, are a prerequisite for a better under-
standing of chironomid community dynamics in
glacial headwaters.
Acknowledgments
The project, Arctic and Alpine Stream Ecosystem
Research (AASER) was supported by the European
Commission (No ENV-CT95-0164) and the Swiss
Federal Of®ce for Education and Science. We
are grateful to G.A. Halvorsen, P. Langton and
E. Willassen for their advice on chironomid taxon-
omy. The authors thank also A.M. Milner and an
anonymous reviewer for helpful comments on an
early version of the manuscript.
References
APHA (1992) Standard Methods for the Examination of
Water and Wastewater, 18th edn. American Public
Health Association, NW, Washington, DC, USA.
Fig. 7 Range of glacier-fed rivers studied showing (a) the Taillon stream (site 3) in the French Pyrenees and (b) Bayleva on Svalbard.
Chironomid communities in glacial streams 1805
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

Ashe P. & Cranston P.S. (1990) Family Chironomidae. In:
Catalogue of Palaearctic Diptera (Eds A. SooÁs & L. Papp),
pp. 113±355. Akademiai KiadoÁ : Budapest and Elsevier
Science Publishers, Amsterdam, The Netherlands.
ter Braak C.J.F. (1986) Canonical correspondence analy-
sis: a new eigenvector technique for multivariate direct
gradient analysis. Ecology, 67, 1167±1179.
Brittain J.E., Salveit S.J., Castella E., Bogen J., Bùsnes T.,
Bremnes T. & Velle G. (2001a) The macroinvertebrate
communities of two contrasting Norwegian glacial
rivers in relation to environmental variables. Freshwater
Biology, 46, 1723±1736.
Brittain J.E., Adalsteinsson H., Castella E., Gislason G.M.,
Lencioni V., Lods-Crozet B., Maiolini B., Milner A.M.,
Petts G.E. & Saltveit S.J. (2001b) Towards a conceptual
understanding of arctic and Alpine streams. Verhandl-
ungen der Internationalen Vereinigung fuÈ r Theoretische
und Angewandte Limnologie, 27, 740±743.
Cranston P.S. (1982) A key to the larvae of the British Ortho-
cladiinae (Chironomidae). Freshwater Biological Associ-
ation, Scienti®c Publication no. 45, Ambleside, UK.
Danks H.V. (1999) Life cycles in polar arthropods ±
¯exible or programmed? European Journal of Entomol-
ogy, 96, 83±102.
Danks H.V. & Oliver D.R. (1972a) Seasonal emergence of
some arctic Chironomidae (Diptera). Canadian Ento-
mologist, 104, 661±686.
Danks H.V. & Oliver D.R. (1972b) Diel periodicities
of emergence of some high arctic Chironomidae
(Diptera). Canadian Entomologist, 104, 903±916.
Dorier A. (1937) La faune des eaux courantes alpines.
Verhandlungen der Internationalen Vereinigung fuÈ r Theo-
retische und Angewandte Limnologie, 8, 33±41.
Ferrarese U. (1983) Chironomidi 3 (Diptera Chironomidae:
Tanypodinae). Guide per il riconoscimento delle specie
animali delle acque interne italiane, 26. Consiglio Nazio-
nale delle Ricerche, Verona, Italy.
Ferrarese U. & Rossaro B. (1981) Chironomidi 1 (Diptera
Chironomidae: GeneralitaÁ, Diamesinae, Prodiamesinae).
Guide per il riconoscimento delle specie animali delle acque
interne italiane, 12. Consiglio Nazionale delle Ricerche,
Verona, Italy.
Flory E.A. & Milner A.M. (1999) The role of competition
in invertebrate community development in a new
stream in Glacier Bay National Park, Alaska. Aquatic
Ecology, 33, 175±184.
Fraile L., Escou®er Y. & Raibaut A. (1993) Analyse des
correspondances de donneÂes plani®eÂes: Etude de la
cheÂmotaxie de la larve infestante d'un parasite. Bio-
metrics, 49, 1142±1153.
Gislason G.M., Adalsteinsson H.O., Hansen I., OÂ lafsson
J.S. & Svavarsdottir K. (2001) Longitudinal changes in
macroinvertebrate assemblages along a glacial river
system in central Iceland. Freshwater Biology, 46, 1737±
1751.
Gislason G.M., OÂ lafsson J.S. & Adalsteinsson H. (1998)
Animal communities in Icelandic rivers in relation
to catchment characteristics and water chemistry ±
preliminary results. Nordic Hydrology, 29, 129±148.
Herrmann S.J., Sublette J.E. & Sublette M. (1987) Mid-
winter emergence of Diamesa leona Roback in the
Upper Arkansas River, Colorado, with notes on other
Diamesinae (Diptera: Chironomidae). Entomologica Scan-
dinavica Suppl., 29, 309±322.
Hongve D. (1987) A revised procedure for discharge
measurement by means of salt dilution method.
Hydrological Processes, 1, 267±270.
Ilg C., Castella E., Lods-Crozet B. & Marmonier P. (2001)
Invertebrate drift and physico-chemical variables in
the tributaries of the Mutt, a Swiss glacial stream.
Archiv fuÈr Hydrobiologie, 151, 335±352.
Janecek B. (1998) Chironomidae (ZuckmuÈ cken) ± Larven.
Fauna Aquatica Austriaca, Taxonomie und OÈ kologie
aquatisher wirbelloser Organismen. Teil V. UniversitaÈt
fuÈ r Bodenkultur, Abteilung Hydrobiologie, Vienna,
Austria.
Jonsson B. & Sandlund O.T. (1975) Notes of winter
activity of two Diamesa species (Diptera, Chironomi-
dae) from Voss, Norway. Norwegian Journal of Ento-
mology, 22, 1±6.
Kawecka B., Kownacki A. & Kownacka M. (1978) Food
relations between algae and bottom fauna communi-
ties in glacial streams. Verhandlungen der Internationalen
Vereinigung fuÈr Theoretische und Angewandte Limnologie,
20, 1527±1530.
Kownacka M. & Kownacki A. (1975) Gletscherbach-
ZuckmuÈ cken der OÈ tztaler Alpen in Tirol (Diptera:
Chironomidae: Diamesinae). Entomologica Germanica,
2, 35±43.
Kownacki A. (1980) Ecology and biogeography of the
Diamesa steinboecki Group. Acta Universitatis Carolinae ±
Biologica, 1978, 95±102.
Kownacki A. (1987) Benthic invertebrate fauna of high
mountain streams in Caucasus. Acta Universitatis Lod-
ziensis, Folia Caucasica, 2, 89±99.
Kownacki A. (1991) Zonal distribution and classi®cation
of the invertebrate communities in high mountain
streams in South Tirol (Italy). Verhandlugen der Inter-
nationale Vereinigung fuÈ r Theoretische und Angewandte
Limnologie, 24, 2010±2014.
Ladle M., Cooling D.A., Welton J.S. & Bass J.A.B. (1985)
Studies on Chironomidae in experimental recirculating
stream systems. II. The growth development and
production of a spring generation of Orthocladius
(Euorthocladius) calvus Pinder. Freshwater Biology, 15,
243±255.
1806 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

Langton P.H. (1991) A Key to Pupal Exuviae of West
Palaearctic Chironomidae. Privately published by P.H.
Langton, 5 Kylebeg Av., Mountsandel, Cleraine Co.
Londonderry, Northern Ireland.
Laville H. (1980) Inventaire 1980 des Chironomides
(Diptera) connus des PyreÂneÂes. Annales de Limnologie,
16, 211±223.
Lencioni V. (2000) Chironomid (Diptera: Chironomidae)
assemblages in three alpine glacial systems. PhD Thesis,
The University of Innsbruck, Innsbruck, Austria.
Lindegaard C. & Brodersen K.P. (1995) Distribution of
Chironomidae (Diptera) in the river continuum. In:
Chironomids: from Genes to Ecosystems (Ed. P. Crans-
ton), pp. 257±271. CSIRO Publications, Melbourne,
Australia.
Lods-Crozet B., Castella E., Cambin D., Ilg C., Knispel S.
& Mayor-SimeÂant H. (2001) Macroinvertebrate com-
munity structure in relation to environmental variables
in a Swiss glacial stream. Freshwater Biology, 46, 1641±
1661.
Maiolini B. & Lencioni V. (2001) Longitudinal distribu-
tion of macroinvertebrate assemblages in a glacially
in¯uenced stream system in the Italian Alps. Freshwa-
ter Biology, 46, 1625±1639.
Makarchenko E.A. (1981) Taksonomyia i rasprostranenie
nekotorykh vidov khironomid podsemeõÈtva Diamesi-
nae (Diptera, Chironomidae) dal'nego Vostoka SSSR.
(Taxonomy and distribution of some chironomid
species of the subfamily Diamesinae (Diptera, Chiro-
nomidae) of the Far East USSR). Bespozvonochnye
zhivotnye v zkosistemakh losostevykh rek dal'nego. Vo-
stoka, Vladivostok, USSR, pp. 83±112.
Makarchenko E.A. (1985) Chironomids of the Soviet Far
East. Subfamilies Poodonominae, Diamesinae and Prodiam-
esinae (Diptera, Chironomidae). DVNC AN SSSR, Vlad-
ivostok, USSR.
McArthur R.H. & Wilson E.O. (1967) The Theory of Island
Biogeography. Princeton University Press, Princeton,
NJ, USA.
Milner A.M. (1994) Colonisation and succession of inver-
tebrate communities in a new stream in Glacier Bay
National Park, Alaska. Freshwater Biology, 32, 387±400.
Milner A.M. & Petts G.E. (1994) Glacial rivers: physical
habitat and ecology. Freshwater Biology, 32, 295±307.
Nocentini A. (1985) Chironomidi 4 (Diptera Chironomidae:
Chironominae, Larve). Guide per il riconoscimento delle
specie animali delle acque interne italiane, 29. Consiglio
Nazionale delle Ricerche, Verona, Italy.
Nolte U. & Hoffmann T. (1992) Fast life in cold water.
Diamesa Incallida (Chironomidae). Ecography, 15, 25±30.
Petts G.E. & Bickerton M.A. (1994) In¯uence of water
abstraction on the macroinvertebrate community
gradient within a glacial stream system: La Borgne
d'Arolla, Valais, Switzerland. Freshwater Biology, 32,
375±386.
Pfankuch D.J. (1975) Stream Reach Inventory and Channel
Stability Evaluation. US Department of Agriculture
Forest Service, Region 1, Missoula, MT, USA.
Pianka E.R. (1978) Evolutionary Ecology. Harper & Row,
New York, USA.
Rossaro B. (1980) Description of some unknown larvae of
Diamesa genus and corrections to previous descriptions
(Diptera: Chironomidae). Archiv fuÈ r Hydrobiologie, 90,
298±308.
Rossaro B. (1981) Analisi delle larve delle Diamesinae
europee, con descrizione di alcune specie (Diptera
Chironomidae). Bollettino del Museo Civico di Storia
Naturale di Verona, VIII, pp. 29±43.
Rossaro B. (1982) Chironomidi 2 (Diptera, Chironomidae:
Orthocladiinae). Guide per il riconoscimento delle specie
animali delle acque interne italiane, 16. Consiglio Nazio-
nale delle Ricerche, Verona, Italy.
Rossaro B. (1991a) Chironomids and water temperature.
Aquatic Insects, 13, 87±98.
Rossaro B. (1991b) Factors that determine Chironomidae
species distribution in fresh waters. Bollettino Zoologico,
58, 281±286.
Rossaro B. (1995) The distribution of Palaearctic Diame-
sinae (Insecta, Diptera, Chironomidae). Spixiana, 18,
177±186.
Roux M. (1991) Interpretation of hierarchical clustering.
In: Applied Multivariate Analysis in SAR and Environ-
mental Studies (Eds J. Devillers & W. Karcher), pp. 137±
152. Kluwer Academic Publishers, Dordrecht, The
Netherlands.
Ruse L.P. (1994) Chironomid microdistribution in gravel
of an English chalk river. Freshwater Biology, 32, 533±551.
Saether O.A. (1968) Chironomids of the Finse Area,
Norway, with special reference to their distribution in
a glacier brook. Archiv fuÈ r Hydrobiologie, 64, 426±453.
Saltveit S.J., Haug I. & Brittain J.E. (2001) Invertebrate
drift in a glacial river and its non-glacial tributary.
Freshwater Biology, 46, 1777±1789.
Schmid P.E. (1993) A key to the larval Chironomidae and
their instars from Austrian Danube region streams and
rivers. Part 1: Diamesinae, Prodiamesinae and Ortho-
cladiinae. Wasser und Abwasser Supplementband, 3/93,
1±513.
Schnell é.A., Rieradevall M., Granados I. & Hanssen O.
(1999) A chironomid taxa coding system for use in
ecological and palaeoecological databases. Report SNO
3710±97, Mountain Lake Research (MOLAR), ISBN
82-577-3276-1.
Schnell é.A. & Willassen E. (1991) Fjñrmyggarten
Pseudodiamesa arctica (Malloch) i to Hùyfjellsreservoa-
rer (the Chironomid species Pseudodiamesa arctica
Chironomid communities in glacial streams 1807
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809

(Malloch) in two high montain reservoirs). Report
number 76, Freshwater Ecology and Inland Fisheries
Laboratory (LFI), Department of Zoology, University
of Bergen, Bergern, Norway.
Serra-Tosio B. (1967) Taxonomie et eÂcologie des Diamesa
du groupe latitarsis (Diptera, Chironomidae). Travaux
Du Laboratoire d'Hydrobiologie et de Pisciculture de
Grenoble, 57/58, 65±91.
Serra-Tosio B. (1973) Ecologie et biogeÂographie des
Diamesini d'Europe (Diptera, Chironomidae). Travaux
Du Laboratoire d'Hydrobiologie et de Pisciculture de
Grenoble, 63, 5±175.
Snook D.L. (2000) Macroinvertebrate communities in Alpine
glacier-fed streams: The Taillon catchment in the French
PyreÂneÂes. PhD Thesis, The University of Birmingham,
Birmingham, UK.
Snook D.L. & Milner A.M. (2001) The in¯uence of glacial
runoff on stream macroinvertebrate communities in
the Taillon catchment, French PyreÂneÂes. Freshwater
Biology, 46, 1609±1623.
Steffan A.W. (1971) Chironomid (Diptera) biocoenoses in
Scandinavian glacier brooks. Canadian Entomologist,
103, 477±486.
Thienemann A. (1936) Alpine Chironomiden (Ergebnisse
von Untersuchungen in der Gegend von Garmisch-
Partenkirchen, Oberbayern). Archiv fuÈ r Hydrobiologie,
30, 167±262.
Thienemann A. (1952) Bestimmungstabelle fuÈ r die Lar-
ven der mit Diamesa naÈchsten verwandten Chironom-
iden. BeitraÈge Zur Entomologie, 2, 244±256.
Thioulouse J., DoleÂdec S., Chessel D. & Olivier J.M. (1997)
ADE-4: a multivariate analysis and graphical display
software. Statistics and Computing, 7, 75±83.
Townsend C.R. (1989) The patch dynamics concept of
stream community ecology. Journal of the North Ameri-
can Benthological Society, 8, 36±50.
Townsend C.R., Scarsbrook M.R. & DoleÂdec S. (1997)
Quantifying disturbance in streams: alternative
measures of disturbance in relation to macroinver-
tebrate species traits and species richness. Journal
of the North American Benthological Society, 16, 531±
544.
Uehlinger U., Zah R. & Buergi H. (1998) The Val Roseg
project: temporal and spatial patterns of benthic algae in
an Alpine stream ecosystem in¯uenced by glacier
runoff. In: Hydrology, Water Resources and Ecology in
Headwaters (Eds K. Kovar, N.E. Tappeiner, N.E. Petters
& R.G. Craig), pp. 419±424. IAHS Press, Wallingford,
UK.
Ward J.H. (1963) Hierarchical grouping to optimize an
objective function. Journal of the American Statistical
Association, 58, 236±244.
Ward J.V. (1986) Altitudinal zonation in a Rocky
Mountain stream. Archiv fuÈ r Hydrobiologie Suppl., 74,
133±199.
Ward J.V. (1994) Ecology of Alpine streams. Freshwater
Biology, 32, 277±294.
Wiederholm T. (1983) Chironomidae of the Holarctic
region. Keys and Diagnoses. Part 1. Larvae. Entomologica
Scandinavica Suppl., 19, 1±449.
Wiederholm T. (1986) Chironomidae of the Holarctic
region. Keys and Diagnoses. Part 2. Pupae. Entomologica
Scandinavica Suppl., 28, 1±482.
(Manuscript accepted 5 September 2001)
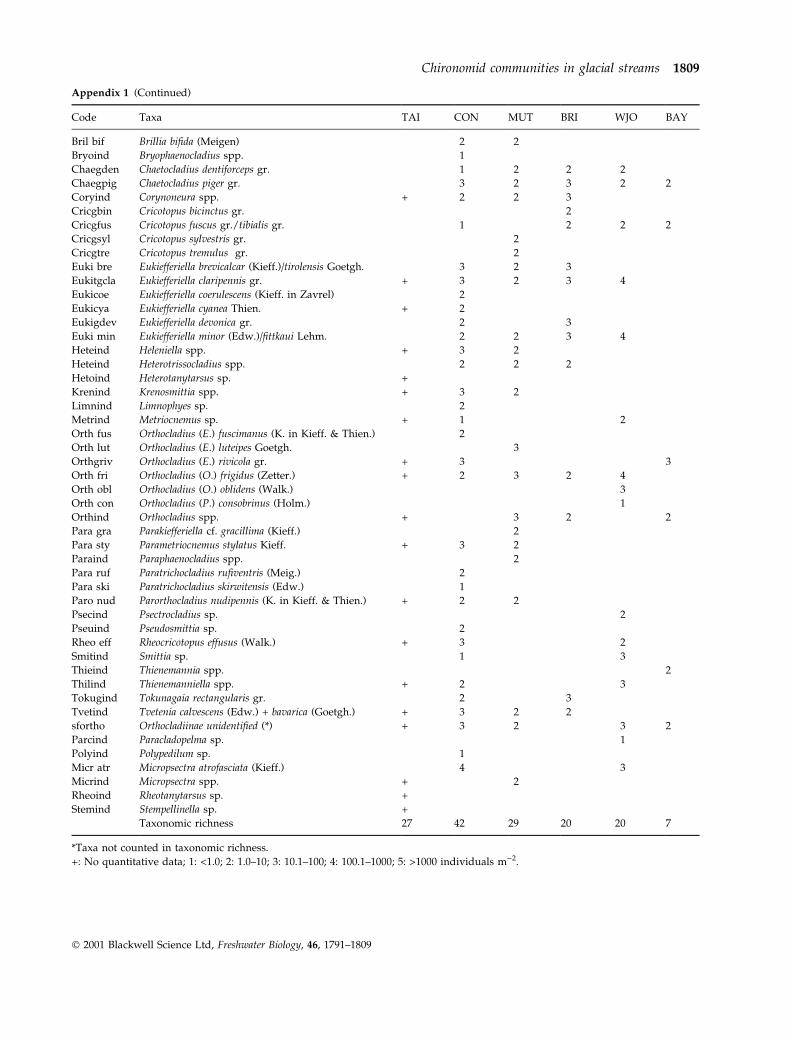
Appendix 1 Chironomidae species composition and mean abundance (expressed in classes) of in the six glacial stream
Code Taxa TAI CON MUT BRI WJO BAY
Concind Conchapelopia sp. 2
Macrind Macropelopia sp. + 1
Zavrind Zavrelimyia sp. 3
Boreind Boreoheptagyia 2 2
Diamgabe Diamesa aberrata gr. + 2 2
Diam ber Diamesa bertrami Edwards + 3 2 4 4
Diamgdam Diamesa damp® gr. 2
Diamgdav Diamesa davisi gr. 4
Diamglat Diamesa latitarsis gr. + 3 5 4 4
Diam ste Diamesa steinboecki Goetgh. 4 3
Diamgzer Diamesa zernyi gr./cinerella gr. + 4 4 4 4 4
DiamgA Diamesa gr. A(*) + 4 5 3 4
Pseu arc Pseudodiamesa arctica (Mall.) 3
Pseu bra Pseudodiamesa branickii (Now.) + 3 2
Pseu par Pseudodiefferiella parva (Edwards) + 3 3
Syndind Syndiamesa sp. 2 2
sfdiame Diamesinae unidenti®ed (*) + 2 4 2
1808 B. Lods-Crozet et al.
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809
Appendix 1 (Continued)
Code Taxa TAI CON MUT BRI WJO BAY
Bril bif Brillia bi®da (Meigen) 2 2
Bryoind Bryophaenocladius spp. 1
Chaegden Chaetocladius dentiforceps gr. 1 2 2 2
Chaegpig Chaetocladius piger gr. 3 2 3 2 2
Coryind Corynoneura spp. + 2 2 3
Cricgbin Cricotopus bicinctus gr. 2
Cricgfus Cricotopus fuscus gr./tibialis gr. 1 2 2 2
Cricgsyl Cricotopus sylvestris gr. 2
Cricgtre Cricotopus tremulus gr. 2
Euki bre Eukiefferiella brevicalcar (Kieff.)/tirolensis Goetgh. 3 2 3
Eukitgcla Eukiefferiella claripennis gr. + 3 2 3 4
Eukicoe Eukiefferiella coerulescens (Kieff. in Zavrel) 2
Eukicya Eukiefferiella cyanea Thien. + 2
Eukigdev Eukiefferiella devonica gr. 2 3
Euki min Eukiefferiella minor (Edw.)/®ttkaui Lehm. 2 2 3 4
Heteind Heleniella spp. + 3 2
Heteind Heterotrissocladius spp. 2 2 2
Hetoind Heterotanytarsus sp. +
Krenind Krenosmittia spp. + 3 2
Limnind Limnophyes sp. 2
Metrind Metriocnemus sp. + 1 2
Orth fus Orthocladius (E.) fuscimanus (K. in Kieff. & Thien.) 2
Orth lut Orthocladius (E.) luteipes Goetgh. 3
Orthgriv Orthocladius (E.) rivicola gr. + 3 3
Orth fri Orthocladius (O.) frigidus (Zetter.) + 2 3 2 4
Orth obl Orthocladius (O.) oblidens (Walk.) 3
Orth con Orthocladius (P.) consobrinus (Holm.) 1
Orthind Orthocladius spp. + 3 2 2
Para gra Parakiefferiella cf. gracillima (Kieff.) 2
Para sty Parametriocnemus stylatus Kieff. + 3 2
Paraind Paraphaenocladius spp. 2
Para ruf Paratrichocladius ru®ventris (Meig.) 2
Para ski Paratrichocladius skirwitensis (Edw.) 1
Paro nud Parorthocladius nudipennis (K. in Kieff. & Thien.) + 2 2
Psecind Psectrocladius sp. 2
Pseuind Pseudosmittia sp. 2
Rheo eff Rheocricotopus effusus (Walk.) + 3 2
Smitind Smittia sp. 1 3
Thieind Thienemannia spp. 2
Thilind Thienemanniella spp. + 2 3
Tokugind Tokunagaia rectangularis gr. 2 3
Tvetind Tvetenia calvescens (Edw.) + bavarica (Goetgh.) + 3 2 2
sfortho Orthocladiinae unidenti®ed (*) + 3 2 3 2
Parcind Paracladopelma sp. 1
Polyind Polypedilum sp. 1
Micr atr Micropsectra atrofasciata (Kieff.) 4 3
Micrind Micropsectra spp. + 2
Rheoind Rheotanytarsus sp. +
Stemind Stempellinella sp. +
Taxonomic richness 27 42 29 20 20 7
*Taxa not counted in taxonomic richness.
+: No quantitative data; 1: <1.0; 2: 1.0±10; 3: 10.1±100; 4: 100.1±1000; 5: >1000 individuals m)2.
Chironomid communities in glacial streams 1809
Ó 2001 Blackwell Science Ltd, Freshwater Biology, 46, 1791±1809
Top Related
Copyright © 2022 FDOKUMEN

