







Bahasa
Halaman
Hukum


ELSEVIER Virus Research 39 (1995) 165-179
Virus Research
Characterization of revertants of a Sindbis virus 6K gene mutant that affects proteolytic processing
and virus assembly
Lidia Ivanova, Lam Le 1, M i l t o n J. S c h l e s i n g e r *
Department of Molecular Microbiology, Washington University School of Medicine, Box 8230, 660 So. Euclid St., St. Louis, MO 63110, USA
Received 4 May 1995; revised 30 May 1995; accepted 1 August 1995
Abstract
Alphaviruses of the Togaviridae encode a small hydrophobic polypeptide of 55 amino acids, noted as the 6K protein, that is synthesized as part of a polyprotein containing the sequences of the two major transmembranal viral structural glycoproteins. Mutations, insertions and deletions in the 6K appear to selectively interfere with the final stages of virus assembly and budding, producing aberrant, multi-cored infectious viruses. In addition, some of these mutations were pleiotropic and much more inhibitory to virus formation. One of the latter, a substitution of alanine in the wild-type Sindbis virus 6K gene by arginine, has been studied further and shown to interfere with normal proteolytic processing of the polyprotein. Cells infected with this mutant but not the wild-type virus also displayed viral antigens in nuclear membranes and released fragments of membranes into the cell culture media. A revertant, obtained by enrichment for a faster growing strain, 'suppressed' these defects and genetic mapping showed that the arginine codon had been modified to encode a methionine. However, the sequence of the 6K protein in this revertant was not wild-type and the revertant was still defective in assembly as demonstrated by formation of aberrant particles. A complete restoration of wild-type particle formation for this revertant could be effected by modifying the E2 glycoprotein sequence.
Keywords: Sindbis virus; Site-mutation; Membrane protein; Assembly; Revertant; Prote- olytic processing; Glycoprotein
* Corresponding author. Tel.: + 1 (314) 362-2762; Fax: + 1 (314) 362-1232. 1 Present address: Department of Biology, Hanoi University, Hanoi, Vietnam.
0168-1702/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0168-1702(95)00083-6

166 L. Ivanova et al. /Virus Research 39 (1995) 165-179
1. Introduction
Encoded in a gene between the two glycoprotein structural genes of the alphaviruses of the Togaviridae is a small hydrophobic polypeptide called 6K that initially is synthesized as part of the polyprotein that forms the spike glycoproteins of the infectious virion (Strauss et al., 1984; Strauss and Strauss, 1994). During expression of the alphavirus subgenomic 26S mRNA, the 6K protein is proteolyti- cally released from the nascent polyprotein in two stages. First, its amino terminus is separated from the carboxyl-terminus of the p62, the precursor of the E2 glycoprotein, by the signal peptidase localized to the lumen of the infected cell's endoplasmic reticulum (ER) after the p62 carboxyl terminus has been translocated into the ER membrane (Liu and Brown, 1993). Second, the carboxyl terminus of the 6K is cleaved from the nascent polyprotein by signal peptidase acting on the carboxyl terminal domain of 6K, which has been inserted into the ER membrane (Liljestrom and Garoff, 1991). This processing scheme requires that the 6K protein be topologically oriented in the membrane with two transmembrane domains: one with the amino terminus of the 6K in the ER lumen and the second with the carboxyl terminus in the ER lumen. In this orientation, some amino acids in the central part of the 6K would be positioned at the cytoplasmic face of the ER membrane. Included in this latter part of the 6K sequence are a cluster of cysteines, which are modified by covalently attached long-chain fatty acids (Gaedigk-Nitschko et al., 1990). Thus, the 6K protein is highly lipophilic and strongly embedded in the membrane bilayer. After synthesis, the 6K protein is transported along with the two glycoproteins through the secretory vesicles of the cell and appears at the plasma membrane of the infected cell (Gaedigk-Nitschko and Schlesinger, 1990; Lusa et al., 1991). The amounts of 6K protein initially formed are equivalent (on a molar basis) to those of the p62 and E1 glycoproteins; however, only a small fraction of the 6K proteins appears in extracellular infectious virions. On average, there are roughly 10 moles of glycoprotein to 1 of the 6K protein in these particles.
Our laboratory has been investigating the role of the 6K protein in alphavirus replication and has reported previously that site-directed mutations in the 6K gene can perturb virus assembly both qualitatively and quantitatively, often producing a significant fraction of the infectious virus in the form of multi-cored particles. (Gaedigk-Nitschko et al., 1990; Ivanova et al., 1995). While most of the 6K gene mutations had relatively little overall effect on the amount of infectious particles formed during a single-cycle growth curve, others were more defective in virus replication and growth suggesting that these latter mutations were pleiotropic in their effects.
In this communication, we describe studies of one of these pleiotropic mutants. We have been able to identify the molecular basis for this more severe defect by isolating and characterizing a genetic revertant that corrects for this defect. This revertant, however, still contained a mutationally altered 6K protein and retained the phenotypic trait displayed for alterations in the 6K protein itself, namely the release of aberrant, multi-cored particles. Additional genetic manipulations involv-

L. Ivanova et al. /Virus Research 39 (1995) 165-179 167
ing the insertion of mutations in the ectodomain of the E2 glycoprotein led to restoration of a wild-type phenotype for this latter characteristic.
2. Materials and methods
2.1. Cell cultures and virus strains
Secondary cultures of chicken embryo fibroblasts (CEF) were grown in Earle's minimal medium (MEM) supplemented with 3% fetal bovine serum (FBS). Unless specified otherwise, cells were grown at 37°C. Mutant strains were derived from a cDNA encoding the complete genome for Sindbis virus; it is noted as Toto 1101 (Rice et al., 1987).
2.2. Isolation of the mutant and the revertant
Site-directed mutagenesis was carried out to change the codon for alanine (GCU) at position 28 of the 6K protein to one for arginine (AGG) using procedures and reagents identical to those previously described (Gaedigk-Nitschko et al., 1990; Ivanova and Schlesinger, 1993). Recombinant bacteriophage M13mpl8, carrying a 1496 nucleotide Sal I fragment of the Sindbis virus full length cDNA clone (provided by C. Rice) was used for the mutagenesis. The oligonucleotide 5' AACGATGAACCTGGCCAAAGG 3' was used. A BssH II/Spl I fragment was subcloned back into the Sindbis virus full-length cDNA (Toto 1101) and the presence of the correct mutation was verified again by sequencing. Linearized plasmid DNA was transcribed into infectious mRNA using SP6 polymerase and the RNA transfected into secondary chicken embryo fibroblast cells (CEF) using lipofectin. The RNA transcribed from the mutagenized cDNA yielded small plaques of infectious virus, which displayed a significantly slower growth than wild-type virus on CEF.
We took advantage of the slower growth of the A28R mutant in CEF cells to isolate a revertant. CEF cells were infected with the A28R mutant at an moi of 100. At 3 h post-infection, medium from cells was used to infect another CEF monolayer and this process was continued for another 8 passages. Early released virus from this 8th passage was plaque-purified and isolates had growth rates that were faster than that of the original mutant and similar to the wild-type virus. In addition, their plaque size was larger than the mutant and similar to the wild-type virus. To determine the genotype of this new strain, total RNA was isolated from BHK cells infected with a plaque-purified revertant and used as a template for cDNA synthesis. A set of specific primers (Ivanova and Schlesinger, 1993) covering the 6K gene was employed for PCR amplification. A BssH II /Spl I fragment was subcloned into the parental Toto 1101 cDNA and the DNA transcribed in mRNA, which was used to transfect CEF cells. Virus emerging from this transfection was plaque-purified and analyzed for phenotypic properties, which were found identi-

168 L. Ivanova et al. / l, qrus Research 39 (1995) 165-179
cal to those of the revertant. Subsequent sequencing of the replaced DNA fragment led to identification of a codon change in the 6K gene.
2.3. Construction of recombinant sindbis viruses
Three reconstructed genetic variants of Sindbis virus were made. For the first, noted as A28Mcon, the BssH II/Spl I fragment from the revertant virus was subcloned into the Toto 1101 cDNA. For the second, noted as A28M:E2 Q55H, H333Q, the BssH II/Spl I fragment from the revertant virus was subcloned into a strain noted RB (Ivanova et al., 1995). This construct contained the 6K change of A28M and two changes (from Toto 1101) in the E2 glycoprotein: Q55H and H333Q. For the third, noted A28M:E2 H333Q, a Bcl I/BssH II fragment which contained only one change in the E2 glycoprotein: H333Q (Ivanova et al., 1995) was subcloned into the A28M revertant strain. This construct had two differences from Toto 1101; the 6K A28M and the E2 glycoprotein H333Q. All replaced regions were sequenced and the new mutant cDNAs transcribed and mRNAs transfected into CEF cells.
2.4. Immunofluorescence and electron microscopy
CEF cells were grown on coverslips to low confluency and infected at an moi of 20 with wild-type virus, A28R mutant, A28M revertant, and A28Mcon strains. At 5 h post-infection, cells were fixed with 4% paraformaldehyde. Cellular membranes were permeabilized with 0.5% Triton X-100. Viral antigens were visualized using rabbit polyclonal antibodies that had been pre-adsorbed to uninfected CEF. The primary antibodies were those directed against intact Sindbis virus, purified Sindbis virus glycoproteins and a peptide corresponding to the amino-terminal 15 amino acids of the 6K protein (Gaeidigk-Nitschko and Schlesinger, 1990). Goat- anti-rabbit antibody conjugated with rhodamine (Capped was used as the second antibody.
For visualizing particle morphology by electron microscopy, virus was harvested from infected CEF at 12 h post-infection and concentrated by ultracentrifugation. Samples were applied to carbon-coated grids and stained with 1% uranyl acetate as previously described (Ivanova et al., 1995).
2.5. Determination of amounts of 6K protein in virus
CEF (4 x 106 cells) were infected with an moi of 10 and labeled with 35S-cy- steine (1300 Ci/mM, Amersham) at 5 h post-infection (at 4.5 h, the MEM was changed to one lacking cysteine). 80/zCi were added and an additional 80 /~Ci were added at 6.5 h. At 8 h post-infection, media were collected, clarified by a centrifugation at 3000 rpm for 10 min and virus pelleted by 1 h centrifugation in a Beckman Ti70 rotor at 40,000 rpm in Oak-Ridge-type centrifuge tubes. The pelleted virus was resuspended in 0.5 ml of PBS containing 1% FBS overnight at 4°C, and this concentrated fraction was layered onto 15-40% sucrose gradients

L. Ivanova et al. / Virus Research 39 (1995) 165-179 169
containing TNE. The gradients were centrifuged at 36,000 rpm in a Beckman SW41 rotor for 1 h at 4°C and fractions collected and analyzed for cpm. Peaks were pooled and incubated with Cellufine resin (300/zl of a 1:15 diluted suspen- sion in PBS) for 6 h with rocking. The virus binds to this resin and after brief centrifugation, the resin was washed twice with PBS at 4°C. Virus proteins were released from the resin by addition of 50/zl of SDS-gel loading buffer and boiling for 4 min. Resin was removed by centrifugation and denatured samples analyzed by SDS/PAGE on 5-20% gradient acrylamide gels. Samples of virus that had not been separated on sucrose gradients were boiled with SDS-gel loading buffer and also analyzed. After electrophoresis, the gels were treated for fluorography. The radioactive bands were excised from the dried gels and radioactivity determined by scintillation counting.
2.6. Pulse-chase analysis
Cultures of CEF (10 6 cell/dish) were infected with virus at an moi of 20 and labeled for 10 min at 37°C with either 20/~Ci of 35S-methionine or 35S-cysteine at 4.5 h post-infection. The cell culture media (MEM with 3% FBS) had been changed to one lacking either methionine or cysteine at 4 h. Media were removed and one set of cells (pulse-labeled) were immediately harvested. Two others were treated with fresh media containing 0.3 mM cycloheximide and incubated for an additional 30 or 60 min (chase samples). Cell monolayers were washed twice with 1 ml cold PBS and washes added to the media. Cells were scraped with PBS (three times with 0.5 ml each), collected by centrifugation and frozen. They were lysed with 50/zl of SDS-gel loading buffer, frozen and thawed three times and boiled for 4 min. Samples were loaded onto 10% acrylamide gel for SDS/PAGE. Virus from the media was collected by incubating media plus washes with Cellufine resin (400 /~1 of a 1:15 dilution of stock) for 12 h at 4°C. The resin was washed several times with PBS and virus protein removed with SDS-gel loading buffer and then analyzed by SDS/PAGE.
3. Results
3.1. Phenotype of the original site-mutation
The site-directed mutation studied here was constructed in order to analyze a hydrophobic amino acid sequence at the amino terminus of the 6K that was postulated to function as one of the 'stop-transfer' signals during nascent polypep- tide translocation of the Sindbis virus polyprotein into the ER membrane and also as one of two transmembrane domains for the 6K polypeptide (Hashimoto et al., 1981; Melancon and Garoff, 1986; Liljestrom and Garoff, 1991). The amino-termi- nal hydrophobic sequence consists of amino acids 9-33 of the 6K protein and the hydrophobic alanine at position 28 was replaced with the hydrophilic amino acid arginine. The prediction was made that this substitution would shorten the hydrophobic sequence by 6 amino acids and perturb the 'stop-transfer' activity.
170 L. Ivanova et aL/ I/irus Research 39 (1995) 165-179
o
0
,,,,,n i i
10
. . o - O . . . . . . . . . . . . . . . •
9 '
.o ,"
• ° ~ . . . . O 8 °o° .s ..O - ~
°° s"
7 ,D~//.,';" s SS"
e" ss
/ - - o - - r .t
6
5 i~, "J . . . . • .... rev R28M
. . . . ,o-.- con R28M
~. wt Tolo 1101
4 i m m m
0.0 2.0 4 .0 6.0 8.0
hours post infection
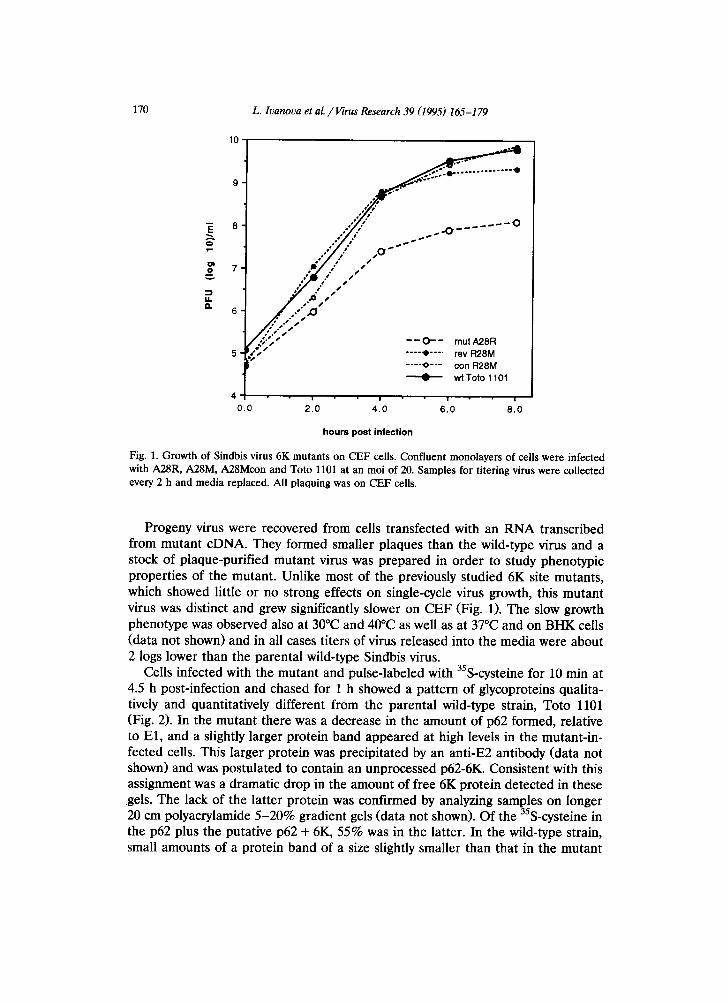
Fig. 1. Growth of Sindbis virus 6K mutants on CEF cells. Confluent monolayers of cells were infected with A28R, A28M, A28Mcon and Toto 1101 at an moi of 20. Samples for titering virus were collected every 2 h and media replaced. All plaquing was on CEF cells.
Progeny virus were recovered from cells transfected with an RNA transcribed from mutant cDNA. They formed smaller plaques than the wild-type virus and a stock of plaque-purified mutant virus was prepared in order to study phenotypic properties of the mutant. Unlike most of the previously studied 6K site mutants, which showed little or no strong effects on single-cycle virus growth, this mutant virus was distinct and grew significantly slower on CEF (Fig. 1). The slow growth phenotype was observed also at 30°C and 40°C as well as at 37°C and on BHK cells (data not shown) and in all cases titers of virus released into the media were about 2 logs lower than the parental wild-type Sindbis virus.
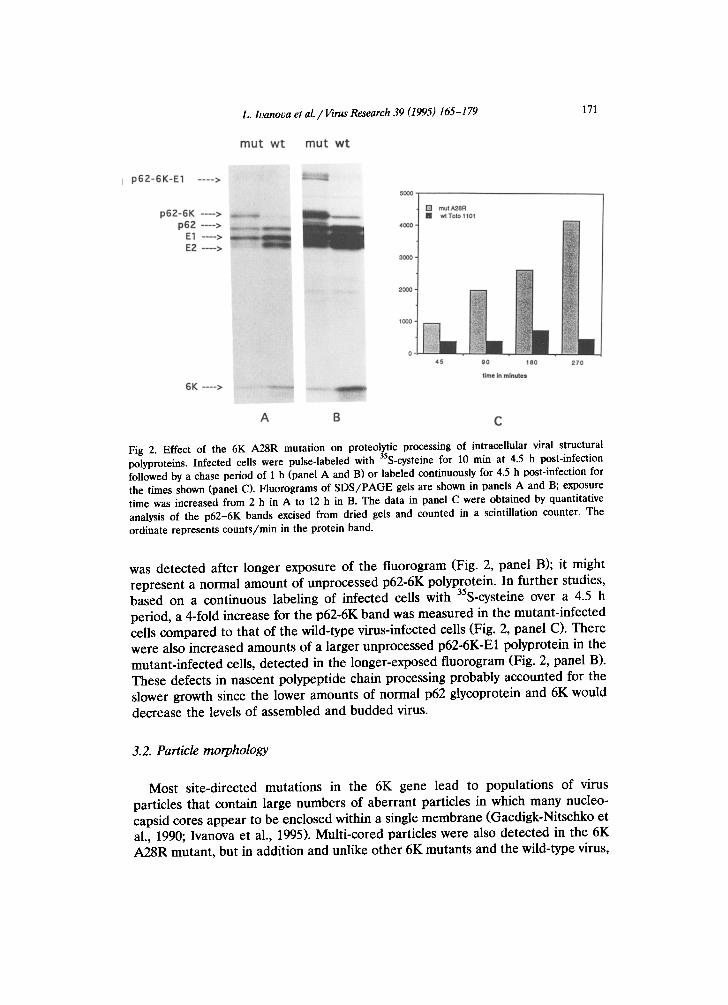
Ceils infected with the mutant and pulse-labeled with 35S-cysteine for 10 min at 4.5 h post-infection and chased for 1 h showed a pattern of glycoproteins qualita- tively and quantitatively different from the parental wild-type strain, Toto 1101 (Fig. 2). In the mutant there was a decrease in the amount of p62 formed, relative to El, and a slightly larger protein band appeared at high levels in the mutant-in- fected cells. This larger protein was precipitated by an anti-E2 antibody (data not shown) and was postulated to contain an unprocessed p62-6K. Consistent with this assignment was a dramatic drop in the amount of free 6K protein detected in these .gels. The lack of the latter protein was confirmed by analyzing samples on longer 20 cm polyacrylamide 5-20% gradient gels (data not shown). Of the 35S-cysteine in the p62 plus the putative p62 + 6K, 55% was in the latter. In the wild-type strain, small amounts of a protein band of a size slightly smaller than that in the mutant
L. luanova et al. / l/irus Research 39 (1995) 165-179 171
mut w t rout w t
p 6 Z - 6 K - E 1 . . . . >
p 6 Z - 6 K - - - - > p62 - - - ->
E1 - - - ->
E2 - - - ->
6K - - - ->
[ ] rout A28R • wt Toto 1101
45 90 180
time in minutes
A B C
270
Fig 2. Effect of the 6K A28R mutation on proteolytic processing of intracellular viral structural polyproteins. Infected cells were pulse-labeled with 35S-cysteine for 10 min at 4.5 h post-infection followed by a chase period of 1 h (panel A and B) or labeled continuously for 4.5 h post-infection for the times shown (panel C). Fluorograms of SDS/PAGE gels are shown in panels A and B; exposure time was increased from 2 h in A to 12 h in B. The data in panel C were obtained by quantitative analysis of the p62-6K bands excised from dried gels and counted in a scintillation counter. The ordinate represents counts/rain in the protein band.
was detected after longer exposure of the fluorogram (Fig. 2, panel B); it might represent a normal amount of unprocessed p62-6K polyprotein. In further studies, based on a continuous labeling of infected cells with 35S-cysteine over a 4.5 h period, a 4-fold increase for the p62-6K band was measured in the mutant-infected cells compared to that of the wild-type virus-infected cells (Fig. 2, panel C). There were also increased amounts of a larger unprocessed p62-6K-E1 polyprotein in the mutant-infected cells, detected in the longer-exposed fluorogram (Fig. 2, panel B). These defects in nascent polypeptide chain processing probably accounted for the slower growth since the lower amounts of normal p62 glycoprotein and 6K would decrease the levels of assembled and budded virus.
3.2. Particle morphology
Most site-directed mutations in the 6K gene lead to populations of virus particles that contain large numbers of aberrant particles in which many nucleo- capsid cores appear to be enclosed within a single membrane (Gaedigk-Nitschko et al., 1990; Ivanova et al., 1995). Multi-cored particles were also detected in the 6K A28R mutant, but in addition and unlike other 6K mutants and the wild-type virus,
172 L. lvanova et al. /Virus Research 39 (1995) 165-179
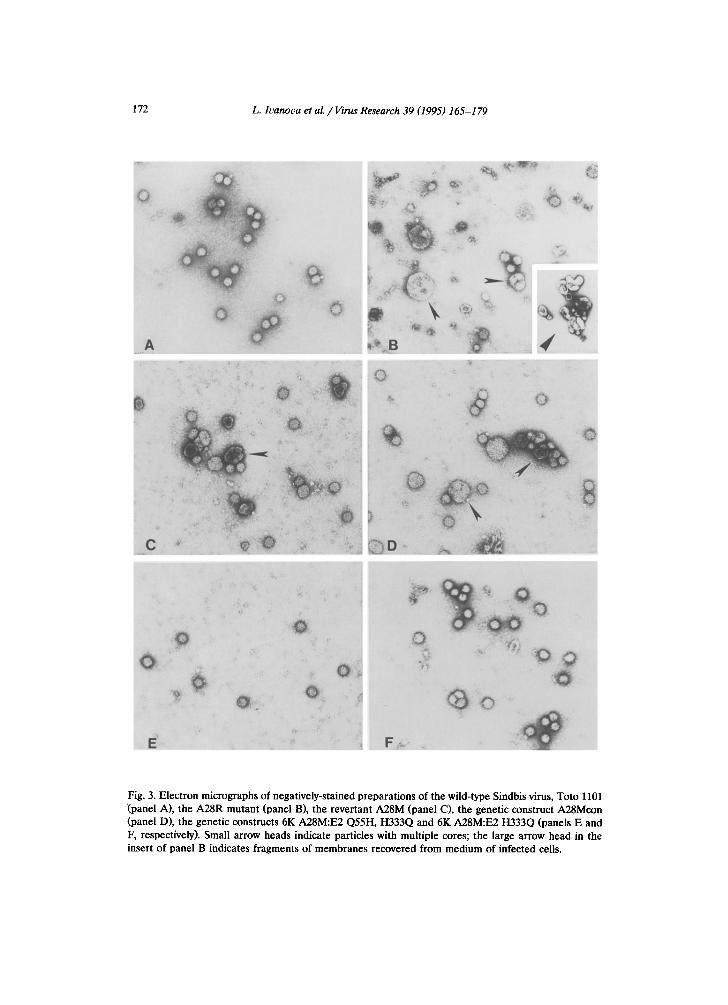
Fig. 3. Electron micrographs of negatively-stained preparations of the wild-type Sindbis virus, Toto 1101 (panel A), the A28R mutant (panel B), the revertant A28M (panel C), the genetic construct A28Mcon (panel D), the genetic constructs 6K A28M:E2 Q55H, H333Q and 6K A28M:E2 H333Q (panels E and F, respectively). Small arrow heads indicate particles with multiple cores; the large arrow head in the insert of panel B indicates fragments of membranes recovered from medium of infected cells.
L. lvanova et al. / Virus Research 39 (1995) 165-179 173
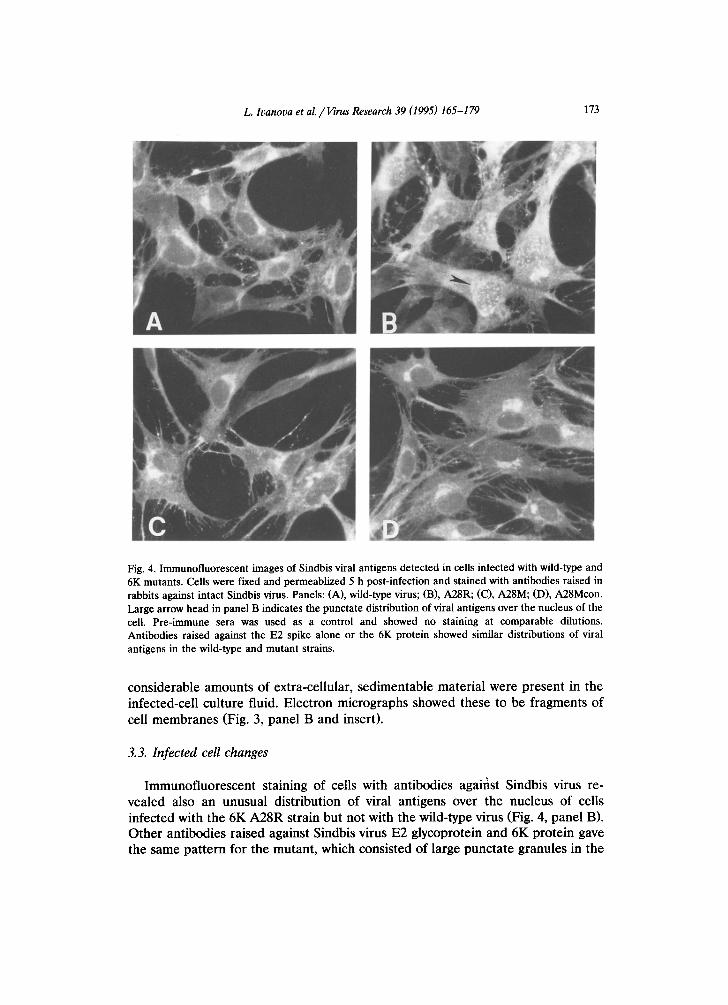
Fig. 4. Immunofluorescent images of Sindbis viral antigens detected in cells infected with wild-type and 6K mutants. Cells were fixed and permeablized 5 h post-infection and stained with antibodies raised in rabbits against intact Sindbis virus. Panels: (A), wild-type virus; (B), A28R; (C), A28M; (D), A28Mcon. Large arrow head in panel B indicates the punctate distribution of viral antigens over the nucleus of the cell. Pre-immune sera was used as a control and showed no staining at comparable dilutions. Antibodies raised against the E2 spike alone or the 6K protein showed similar distributions of viral antigens in the wild-type and mutant strains.
cons ide rab le amoun t s of ext ra-cel lu lar , s e d ime n ta b l e ma te r i a l were p r e se n t in the infec ted-ce l l cu l tu re fluid. E lec t ron mic rographs showed these to be f r agmen t s of cell m e m b r a n e s (Fig. 3, pane l B and insert) .
3.3. Infected cell changes
I m m u n o f l u o r e s c e n t s ta ining of cells wi th an t ibod ies agaifist S indbis virus re- vea led also an unusua l d i s t r ibu t ion of viral an t igens over the nucleus of cells in fec ted with the 6K A 2 8 R s t ra in bu t not with the wi ld- type virus (Fig. 4, p a n e l B). O t h e r an t ibod ie s r a i sed aga ins t Sindbis virus E2 g lycopro te in and 6K p ro t e in gave the same p a t t e r n for the mutan t , which cons is ted of la rge punc t a t e g ranu les in the

174 L. Ivanova et al. / I/irus Research 39 (1995) 165-179
nuclear region and stronger signals throughout the cytoplasm. The precise compo- sition of these granules has not been examined, however, their detection by antibodies that selectively bind to sequences in the 6K protein and by antibodies that selectively bind to sequences in the p62 protein indicate that sequences from both p62 and 6K are part of these aggregates. This pattern may be indicative of a block in normal intracellular transport with accumulation in endoplasmic reticular membranes and subsequent diffusion into nuclear membranes.
3.4. Isolation and phenotypic traits of a revertant
To gain further insight into the effect of the arginine substitution at position 28 of the 6K protein, we took advantage of the slower-growth phenotype to isolate a revertant, which would grow like wild-type virus and display normal plaque size. Several isolates were plaque-purified and their phenotype analyzed. One putative revertant grew faster than the mutant on CEF with kinetics much like wild-type (Fig. 1) and, importantly, had a pattern of intracellular virus glycoproteins more like the wild-type (Fig. 5, lane 4) and distinct from the mutant (Fig. 5, lane 3). Electron micrographs of virus particles released from ceils infected with the revertant showed very few membrane fragments but there were still significantly more large, aberrant multi-cored viruses than detected with wild-type Sindbis virus (Fig. 3, panel A). Immunofluorescent staining of cells infected with the revertant showed a distribution of viral antigens comparable to that observed in cells infected with wild-type virus (Fig. 4, panel A). Thus, this revertant corrected for several defects in the original mutant yet remained 'mutant' with respect to aberrant particle formation.
3.5. Genetic mapping of the revertant
The faster-growing revertant as well as several others with 'wild-type' plaque morphologies and growth kinetics was subjected to a genetic analysis. Infected cells were treated to isolate RNA and appropriate primers were used to reverse transcribe the RNA into a cDNA that covered the region of the genome encoding the original 6K mutation and small portions of the E2 and E1 genes. PCR was used to amplify the DNA which was then sequenced. Only one codon sequence was changed in all of the isolates. The arginine codon in the mutant was changed from AGG to AUG, leading to substitution of a methionine at this position. To insure that it was this new codon that accounted for the phenotype of the revertant, a new Sindbis virus mutant, A28Mcon, was constructed. The cDNA carrying the methionine codon was ligated into a cDNA for the wild-type Toto 1101 that had been cut with restriction enzymes to remove the 6K gene. This reconstructed cDNA was then transcribed into RNA which was transfected into cells to obtain progeny virus. Plaques of the A28Mcon strain were normal in size and several were picked for analysis of phenotypic traits. Like the revertant, the reconstructed strain was almost normal in growth on CEF (Fig. 1) and its pattern of intracellular viral proteins resembled the wild-type and the original revertant
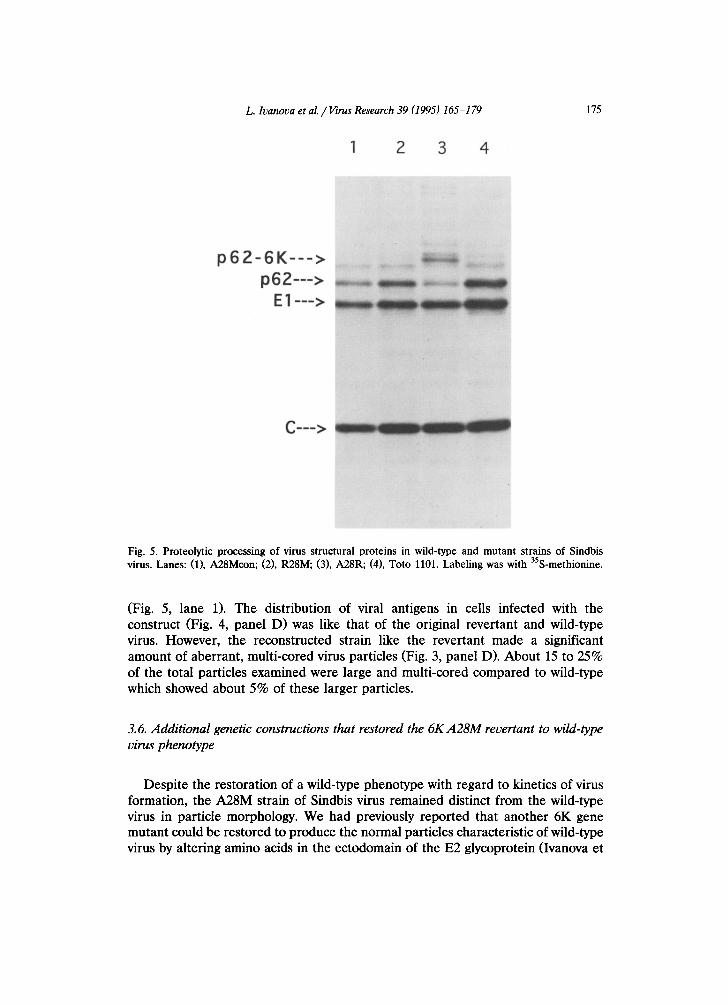
L. Ivanova et al. /I/irus Research 39 (1995) 165-179
I 2 3 4
175
p62-6K---> p62--->
El--->
C-__>
Fig. 5. Proteolytic processing of virus structural proteins in wild-type and mutant strains of Sindbis virus. Lanes: (1), A28Mcon; (2), R28M; (3), A28R; (4), Toto 1101. Labeling was with 35S-methionine.
(Fig. 5, lane 1). The distribution of viral antigens in cells infected with the construct (Fig. 4, panel D) was like that of the original revertant and wild-type virus. However, the reconstructed strain like the revertant made a significant amount of aberrant, multi-cored virus particles (Fig. 3, panel D). About 15 to 25% of the total particles examined were large and multi-cored compared to wild-type which showed about 5% of these larger particles.
3.6. Addit ional genetic constructions that restored the 6K A 2 8 M revertant to wild-type virus phenotype
Despite the restoration of a wild-type phenotype with regard to kinetics of virus formation, the A28M strain of Sindbis virus remained distinct from the wild-type virus in particle morphology. We had previously reported that another 6K gene mutant could be restored to produce the normal particles characteristic of wild-type virus by altering amino acids in the ectodomain of the E2 glycoprotein (Ivanova et

176 L. Ivanova et al. / Virus Research 39 (1995) 165-179
al., 1995). We tested whether these E2 mutations might also restore normal virus assembly to the 6K A28M strain by appropriate genetic constructs. Two new constructs were prepared: one was noted 6K A28M:E2 Q55H, H333Q, and the other was 6K A28M:E2 H333Q. Both of these new constructs resembled wild-type Sindbis virus in the morphology of virus secreted into the cell culture medium (Fig. 3, panels E and F). Thus, identical changes in the E2 glycoprotein could revert two different mutations in the 6K gene.
3. 7. Presence of the 6K protein in virus particles
We have previously reported that virions from cells infected with the wild-type Sindbis and Semliki Forest viruses incorporate low levels of 6K protein (Gaedigk- Nitschko and Schlesinger, 1990; Loewy et al., 1995). To determine if the virions produced by cells infected with those 6K mutants that formed aberrant, multi-cored particles also contained 6K proteins, we labeled infected cells with 35S-cysteine for 2 h at 4 h post-infection and collected the medium, which was then fractionated by rate-zonal sedimentation on a sucrose gradient. The peak corresponding to virus was pooled and mixed with cellufine resin to trap virus. The cellufine was washed several times and virus released by SDS-gel loading buffer. Samples were subjected to SDS/PAGE using a 5-20% acrylamide gel gradient. After autoradiography, the bands corresponding to the glycoproteins and the 6K protein were excised and counted in a scintillation counter. The results showed that the ratio of 6K: E2 + E1 radioactivity was 12% (+3%) of the value expected for a 1:1 stoichiometry for wild-type virus from two different Sindbis strains, Sin A and Toto 1101, and from two strains containing mutant forms of the 6K protein (Table 1). A second preparation was analyzed also. From these results, we concluded that 6K was a normal component in all infectious virus particles. However, we still do not know if these few proteins (24-30/ particle) are clustered in one domain of the lipid
Table 1 Ratio of 6K to E1 + E2 in virions formed by wild-type and 6K mutant strains
Virus strain Glycoprotein 6K protein Percent (cpm) cpm of 6K b
Sin A a 24,500 340 10 Sin A 34,700 500 10 Toto 1101 55,000 1350 17 Toto 1101 18,400 320 12 6K A28M 29,600 320 8 6K A28M 32,700 740 16 6K A28Mcon 22,000 470 15 6K A28Mcon 51,500 820 I1
a Another strain of wild-type Sindbis virus present in this laboratory. b Based on a stoichiometric 1:1 ratio of 7 (34 cysteines in E1 +E2 and 5 in 6K), for every E l / E 2 heterodimer, this number would be 100.
i.e. if there were 1 6K

L. lvanova et al. / Virus Research 39 (1995) 165-179 177
Normal NHa
6K
Lumen
- - ER Membrane
Cytoplasm
Mutant
Signslesa
I 6K
~ E 1 Lumen
- -%~-- -- ER Membrane
Cytoplasm
Fig. 6. Model indicating topology of normal and mutant 6K A28R in the ER membrane of infected cells.
envelope or distributed throughout the particle, perhaps at regions of very high curvature in the lipid envelope.
4. Discussion
The genetic analyses presented in this report provide strong support for the topological model of the 6K protein which has both an amino and carboxyl segment serving as transmembrane domains for this small polypeptide in both the virus and the infected cell. It has been postulated that the first one third of the 6K amino acid sequence serves as a stop transfer sequence during synthesis and translocation of the alphavirus polyprotein encoding the carboxyl terminal portion of the p62 and amino terminal portion of the 6K (see Fig. 6). The two new protein bands appearing in cells infected with the mutant indicated that replacement of the very hydrophilic arginine for the normal alanine residue interfered with the signal peptidase activity responsible for proteolytic processing that separates the 1962 and 6K as well as affecting to a smaller extent the subsequent separation of the 6K from the El. The slower growth phenotype of the mutant provided a means for selecting a revertant and several clones were found to have a methionine at position 28 of the 6K sequence. Thus, restoration of a normal proteolytic process-

178 L. Ivanova et al. /Virus Research 39 (1995) 165-179
ing of the Sindbis virus structural polyprotein was effected by a change from the arginine of the mutant to a hydrophobic amino acid in the revertants.
This result argues for the importance of hydrophobicity in this domain of the 6K protein's sequence and provides experimental support for the model presented earlier on topological distribution of the 6K protein's sequences during biosynthe- sis and proteolytic processing of the polyprotein. Restoration of the hydrophobic domain required the single G --, U base change, which for an RNA virus can occur at a relatively high mutation frequency. Other single-base changes leading to substitution at other 6K sites were not detected in a screen of 10 different revertants, suggesting that the selection pressure employed here, which was based on more rapid growth, enriched for a genetic alteration that removed the hy- drophilic residue. This result supports our contention that the growth defect arose from deficient proteolytic processing in the mutant of the polyprotein at the p62-6K junction.
To restore the full phenotypic properties of the wild-type Sindbis virus, the A28M revertant was genetically manipulated to contain additional mutations in the E2 glycoprotein. In a recent report we showed that a Sindbis virus mutant with a different substitution in the 6K gene, which led to aberrant particle formation, could revert to a wild-type phenotype by mutations in the ectodomain of the Sindbis virus E2 glycoprotein (Ivanova et al., 1995). In that study, we found that the revertant contained two changes in the E2 but, based on genetic reconstruc- tions, either one of these was sufficient. We employed two of these strains to make new constructs with the 6K A28M variant and, as shown here, both could alter the assembly process to lead to single-cored particles. One might therefore consider these E2 changes as general suppressors of the 6K mutant phenotype. However, a change has recently been detected in the E1 glycoprotein that appeared to suppress another 6K mutation (Ivanova and Schlesinger, unpublished data). The molecular basis for a mutant 6K protein's effect on particle assembly and the restoration of normal virus release by changes in the ectodomain of the glycopro- tein has yet to be determined.
Acknowledgements
We thank Marilyn Aach Levy (Dept. Cell Biol. and Physiol. Wash. U. School of Medicine) for invaluable assistance with the electron microscopic studies. This research was supported by a grant from the U.S. Public Health Service AI9494.
References
Gaedigk-Nitschko, K. and Schlesinger, M.J. (1990) The Sindbis virus 6K protein can be detected in virions and is acylated with fatty acid. Virology 175, 274-281.
Gaedigk-Nitschko, K., Ding, M., Levy, M.A. and Schlesinger, M.J. (1990) Site-directed mutations in the

L. Ivanova et al. /Virus Research 39 (1995) 165-179 179
Sindbis virus 6K protein reveal sites for fatty acylation and the underacylated protein affects virus release and virion structure. Virology 175, 282-291.
Hashimoto, K., Erdei, S., Deranen, S., Saraste, J. and Kaariainen, L. (1981) Evidence for a separate signal sequence for the carboxy-terminal envelope glycoprotein E1 of Semliki Forest Virus. J. Virol. 38, 34-40.
Ivanova, L. and Schlesinger, M.J. (1993) Site-directed mutations in the Sindbis virus E2 glycoprotein identify paimitoylation sites and affect virus budding. J. Virol. 67, 2546-2551.
Ivanova, L., Lustig, S. and Schlesinger, M.J. (1995) A pseudo-revertant of a Sindbis virus 6K protein mutant, which corrects for aberrant particle formation, contains two new mutations that map to the ectodomain of the E2 glycoprotein. Virology 206, 1027-1034.
Liljestrom, P. and Garoff, H. (1991) Internally located cleavable signal sequences direct the formation of Semliki Forest virus membrane proteins from a polyprotein precursor. J. Virol. 65, 147-154.
Liljestrom, P., Lusa, S., Huylebroeck, D. and Garoff, H. (1991) In vitro mutagenesis of a full-length cDNA clone of Semliki Forest virus: the small 6,000 molecular weight membrane protein modulates virus release. J. Virol. 65, 4107-4113.
Liu, N. and Brown, D.T. (1993) Transient translocation of the cytoplasmic (endo) domain of a type I membrane glycoprotein into cellular membranes. J. Cell Biol. 120, 877-883.
Loewy, A., Smyth, J., von Bonsdorff, C-H., Liljestrom, P. and Schlesinger, M.J. (1995) The 6-kilodalton membrane protein of Semliki Forest virus is involved in the budding proces. J. Virol. 69, 469-475.
Lusa, S., Garoff, H. and Liljestrom, P. (1991) Fate of the 6K membrane protein of Semliki Forest virus during virus assembly. Virology 185, 843-846.
Melancon, P. and Garoff, H. (1986) Reinitiation of translocation in the Semliki Forest virus structural polyprotein: identification of the signal for the E1 glycoprotein. EMBO J. 5, 1551-1560.
Rice, C.M., Levis, R., Strauss, J.H. and Huang, H.V. (1987) Production of infectious RNA transcripts from Sindbis virus cDNA clones: Mapping of lethal mutations, rescue of a temperature-sensitive marker and in vitro mutagenesis to generate defined mutants. J. Virol. 61, 3809-3819.
Strauss, J.H. and Strauss, E.G. (1994) The atphaviruses: gene expression, replication, and evolution. Microbiol. Rev. 59, 491-562.
Strauss, E.G., Rice, C.M. and Strauss, J.H. (1984) Complete nucleotide sequence of the genomic RNA of Sindbis virus. Virology 133, 92-110.
Top Related
Copyright © 2022 FDOKUMEN

