







Bahasa
Halaman
Hukum


Hum Genet (2010) 128:261–267
DOI 10.1007/s00439-010-0846-zORIGINAL INVESTIGATION
An ancient autosomal haplotype bearing a rare achromatopsia-causing founder mutation is shared among Arab Muslims and Oriental Jews
Lina Zelinger · Alex Greenberg · Susanne Kohl · Eyal Banin · Dror Sharon
Received: 6 April 2010 / Accepted: 2 June 2010 / Published online: 13 June 2010© Springer-Verlag 2010
Abstract Numerous cultural aspects, mainly based onhistorical records, suggest a common origin of the Middle-Eastern Arab Muslim and Jewish populations. This is sup-ported, to some extent, by Y-chromosome haplogroup anal-ysis of Middle-Eastern and European samples. Up to date,no genomic regions that are shared among Arab Muslimand Jewish chromosomes and are unique to these popula-tions have been reported. Here, we report of a rare achro-matopsia-causing CNGA3 mutation (c.1585G>A) presentsin both Arab Muslim and Oriental Jewish patients. A haplo-type analysis of c.1585G>A-bearing chromosomes fromMiddle Eastern and European origins revealed a sharedMuslim–Jewish haplotype, which is diVerent from thosedetected in European patients, indicating a recurrent muta-tion stratiWed by a Jewish–Muslim founder eVect. Compre-hensive whole-genome haplotype analysis using 250 Ksingle nucleotide polymorphism arrays revealed a largehomozygous region of »11 Mbp shared by both Arab Mus-lim and Oriental Jewish chromosomes. A subsequentmicrosatellite analysis of a 21.5 cM interval includingCNGA3 and the adjacent chromosome 2 centromererevealed a unique and extremely rare haplotype associated
with the c.1585G>A mutation. The age of the sharedc.1585G>A mutation was calculated using the microsatel-lite genotyping data to be about 200 generations ago. Asimilar analysis of mutation age based on the Arab Muslimdata alone showed that the mutation was unlikely to be theproduct of a recent gene Xow event. The data present heredemonstrate a large (11 Mbp) genomic region that is likelyto originate from an ancient common ancestor of Middle-Eastern Arab Muslims and Jews who lived approximately5,000 years ago.
Introduction
The Arab Muslim and Jewish populations in Israel and thePalestinian territories are speculated, mainly by historicalrecords, to originate from a single founder population, withan estimated divergence time of a few thousand years. Sup-porting the common origin are many shared aspectsbetween the two languages (Arabic and Hebrew) and reli-gions (Islam and Judaism). Aiming to provide scientiWcevidence for the common origin of these populations, aseries of genetic studies have been conducted, mainly usinggenetic markers on the Y chromosome (Hammer et al.2000; Nebel et al. 2000, 2001), mitochondrial DNA analy-sis (Bonne-Tamir et al. 1986), and haplotyping of speciWcgenomic regions (Niell et al. 2003; Peretz et al. 1997; Sha-hin et al. 2002). In most of these studies, the frequencies ofdiVerent haplotypes that are common in many populationsworldwide have been determined in Middle-Eastern popu-lations followed by construction of phylogenetic trees.While these studies, and mainly the Y-chromosome haplo-group analysis, provided valuable data regarding migrationof individuals, they suVer from a relatively low number ofgenotyped markers (up to 19) and the inability to distinguish
Electronic supplementary material The online version of this article (doi:10.1007/s00439-010-0846-z) contains supplementary material, which is available to authorized users.
L. Zelinger · A. Greenberg · E. Banin · D. Sharon (&)Department of Ophthalmology, Hadassah-Hebrew University Medical Center, Jerusalem, Israele-mail: [email protected]
S. KohlMolecular Genetics Laboratory, Centre for Ophthalmology, Institute for Ophthalmic Research, Tübingen University, Tübingen, Germany
123

262 Hum Genet (2010) 128:261–267
between founder eVect and gene Xow since the studiedhaplogroups are panethnic and present in most of the stud-ied populations.
A more accurate conclusion might be obtained by com-paring genomic regions in which a sequence change hasoccurred before and in proximity to the divergence of thestudied populations (and their descended sub-populations)and will, therefore, exhibit a haplotype that is unique tothese populations. This will eliminate the need for assump-tions regarding the evolution of shared haplotypes and canprovide clear-cut evidence for a founder eVect. Since the fre-quency of large shared haplotypes between populations sep-arated for a few thousand years is predicted to be very low,we assume that some of these haplotypes will be easier todetect if they include a disease-causing mutation. Therefore,one might search for a rare Mendelian phenotype occurringin both studied populations and use the phenotype or diseaseas a marker for identifying rare shared haplotypes. We reporthere a relatively large genomic region harboring an achro-matopsia-causing mutation that is shared by Arab Muslimand Oriental Jewish individuals. Oriental Jews are a closedMiddle Eastern Jewish population (originating mainly fromthe Middle East and North Africa) due to their relative isola-tion from adjacent populations since the time of the Babylo-nian exile in the sixth century BC until their immigration toIsrael in the 1950s. The mutation age calculation and haplo-type analysis strongly suggests a founder mutation andexcludes a possible recent gene Xow event.
Materials and methods
Patient recruitment
The tenets of the Declaration of Helsinki were followed andinformed consent was obtained from all patients who par-ticipated in this study prior to donation of a blood sample.Blood samples were obtained from the index patient, otheraVected and unaVected family members for DNA analysis.Genomic DNA was extracted from peripheral blood usingthe FlexiGene DNA kit (QIAGEN).
Clinical evaluation
A full ophthalmologic examination was performed in allpatients. Subsequently, color vision testing, full-Weld elec-troretinography (ERG), Goldman visual Welds, and colorvision testing were performed.
Mutation analysis
Primers Xanking all coding CNGA3 exons were designedusing Primer3 (http://frodo.wi.mit.edu/cgi-bin/primer3/
primer3_www.cgi). The primer sequences are available byrequest. PCR was performed in a reaction volume of 30 �lwith 35 cycles. Mutation analysis was performed by directsequencing of PCR products.
SNP and microsatellite analysis
Genotyping of single nucleotide polymorphism (SNP)markers within CNGA3 was performed by restriction analy-sis (rs3754901 and rs2055593 with BccI and rs3769754with HpaII) or sequencing analysis (rs3769756). Wholegenome SNP analysis was performed by the 250 K AVyme-trix system. Criteria for hybridization quality were set top < 0.05 and a region of homozygosity was determined by aminimal number of 100 consecutive homozygous markers.Whole-genome SNP data were analyzed by the IBDWnder(http://dna.leeds.ac.uk/ibdfinder/) with a genotyping errorrate of 1% and a minimal homozygosity run of 30 SNPs.Genotyping of nine microsatellite markers (D2S2180,D2S289, D2S2333, D2S2216, D2S2159, D2S113,D2S2311, D2S2209, and D2S2972) was performed by ABIPRISM® 3100 Genetic Analyzer. The data were analyzedusing the Peak Scanner™ Software v1.0, both by AppliedBiosystems Inc. Haplotypes were reconstructed from homozy-gous microsatellite markers. The data were analyzed by theNETWORK 4.5.1.6 software under the STR format and thenetwork construction was done based on the median joining(MJ) model (Bandelt et al. 1999).
Estimation of mutation age
Analysis of mutation age was performed by the DMLE+program (version 2.2) (Reeve and Rannala 2002). The pop-ulation growth (r) was estimated by the following formula:T1 = T0e
(gr), where T1 represents the current population size,T0 represents the ancestral population size, and g representsthe number of generations. The estimated current popula-tion size (T1) was determined as 500,000 for the OrientalJewish population and 230,000 for the Arab Muslim popu-lation in the vicinity of Jerusalem (the data are based on theIsraeli Central Bureau of Statistics). The initial size of thetwo studied populations can be only roughly estimated byhistorical records and, therefore, we used a range of 5,000–50,000 for all calculations. Using the above-mentioned for-mula, Oriental Jewish population growth (r) was calculatedas 0.039, 0.033, 0.028 (for T0 = 10,000 and a variable num-ber of 100, 120, and 140 generations, respectively) or0.030, 0.025, 0.021 (for T0 = 25,000 and a variable numberof 100, 120, and 140 generations, respectively). Muslimpopulation growth was calculated as 0.038, 0.032, 0.027(for T0 = 5,000 and variable number of 100, 120, and 140generations, respectively) or 0.031, 0.026, 0.022 (forT0 = 10,000 and variable number of 100, 120, and 140
123

Hum Genet (2010) 128:261–267 263
generations, respectively). Based on these parameters, themutation age was calculated using DMLE+. Since theresults of the most extreme parameters were within the 95%conWdence interval of the average parameters, we used forour analysis only the DMLE+ data based on an average of25,000 individuals. For all calculations we used a 25 yearsinterval per generation.
Results
As part of our study of the genetics of retinal disease in theIsraeli and Palestinian populations, we performed mutationanalysis in a set of over 700 families with inherited retinaldiseases and characterized the ethnic distribution of theidentiWed mutations. As expected, the majority of identiWedmutations were rare and restricted to a single family (73%,58 out of 80 mutations). The remaining mutations werespeciWc to a single ethnic group (24%), identiWed withinJewish or Muslim subpopulations sharing a geographi-cal origin (i.e., Jews of Moroccan descent). Three excep-tions were identiWed, in which a disease-causingmutation appeared in both the Muslim and Jewish popu-lations: c.119-2A>C in the NR2E3 gene (MIM *604485),c.5882G>A (p.G1961E) in the ABCA4 gene (MIM *601691),and c.1585G>A (p.V529M) in the CNGA3 gene (MIM*600053). The Wrst two mutations are panethnic and repre-sent the most common mutations identiWed in the corre-sponding genes in diVerent populations (Briggs et al. 2001;Haider et al. 2000; Sharon et al. 2003; Webster et al. 2001).Therefore, they represent either ancient founder mutationsoccurring early in human evolution or recurrent base substi-tutions due to mutation hotspots. The third mutation,c.1585G>A in CNGA3, has been identiWed in 10 out of 30families with achromatopsia, an autosomal recessive retinaldisorder characterized by the absence of cone photoreceptor
function. Patients with achromatopsia suVer from reducedvisual acuity, nystagmus, photophobia, and congenital severelyimpaired or complete color blindness, usually with minimalfunduscopic Wndings.
The c.1585G>A mutation has been reported so far as thecause of disease in a single European family (Kohl et al.1998) and two American families (Nishiguchi et al. 2005).In the current study, we identiWed the mutation in 6 out of19 Arab Muslim families with achromatopsia residing indiVerent East Jerusalem villages and in all 4 Oriental Jewishfamilies with achromatopsia. We estimated the mutationcarrier frequencies in Arab Muslims and Oriental Jews as0.8% (one carrier out of 121 control individuals, 95% conW-dence interval: 0.2–4.5%) and 1% (one carrier out of 100control individuals, 95% conWdence interval: 0.2–5%),respectively. In addition, we identiWed the mutation in threeChristian European families that have not been previouslyreported. All European patients were heterozygous forc.1585G>A and carried a diVerent CNGA3 mutation on thecounter allele. On the other hand, most of the Arab Muslimand Jewish patients were homozygous for the c.1585G>Amutation (Table 1 and Online Resource 1), mainly due tothe relatively high consanguinity levels in these populations(Goldschmidt et al. 1960; Jaber et al. 1994; Zlotogora 2002).This enables us to accurately determine the c.1585G>A-associated haplotype in these patients without the need forhaplotype reconstruction.
Aiming to shed a light on the history of the CNGA3c.1585G>A mutation in these populations, we initially per-formed a haplotype analysis along a 28,564 bp fragmentwithin the CNGA3 gene using a set of four SNP markers.For each marker, we genotyped individuals from 11 fami-lies (3 Arab Muslim, 4 Oriental Jewish, and 4 Europeanfamilies) in which the c.1585G>A mutation had been iden-tiWed, and reconstructed the haplotype in heterozygousindividuals. The analysis clearly indicated that both Arab
Table 1 Families with achromatopsia due to the CNGA3 c.1585G>A mutation
a The number of paternal and maternal generations to the common ancestor in a consanguineous family
Family number Origin Number of patients
Level of consanguinitya
Mutation(s)
MOL0010 Iranian Jew 2 3:2 c.1585G>A/c.1585G>A
MOL0013 Iraqi Jew 1 None c.1585G>A/c.1642G>A
MOL0176 Arab Muslim, Jerusalem 5 2:1 c.1585G>A/c.1585G>A
MOL0183 Arab Muslim, Jerusalem 5 3:3 c.1585G>A/c.1585G>A
MOL0254 Arab Muslim, Jerusalem 1 2:2 c.1585G>A/c.1585G>A
MOL0328 Jew (Iraq, Kurdistan/Iran) 1 None c.1585G>A/c.1585G>A
MOL0340 Jew (Kurdistan, Iraq/Iran) 2 None c.1585G>A/c.1294delG
MOL0529 Arab Muslim, Jerusalem 3 2:2 c.1585G>A/c.1585G>A
MOL0738 Arab Muslim, Jerusalem 2 3:3 c.1585G>A/c.1585G>A
MOL0834 Arab Muslim, Jerusalem 1 2:2 c.1585G>A/c.1585G>A
123
264 Hum Genet (2010) 128:261–267
Muslim and Oriental Jewish c.1585G>A-bearing chromo-somes share an identical haplotype, which is diVerent fromthe haplotypes identiWed in the European families (OnlineResource 2). The haplotype phase could not be solved intwo of the European families due to non-informative SNPmarkers, but microsatellite analysis indicated a diVerenthaplotype compared to the shared Muslim–Jewish haplo-type.
Aiming to accurately determine the borders of the sharedMuslim–Jewish c.1585G>A-associated haplotype and com-pare it to other loci throughout the genome, we performed a250 K whole-genome SNP analysis in six patients, all
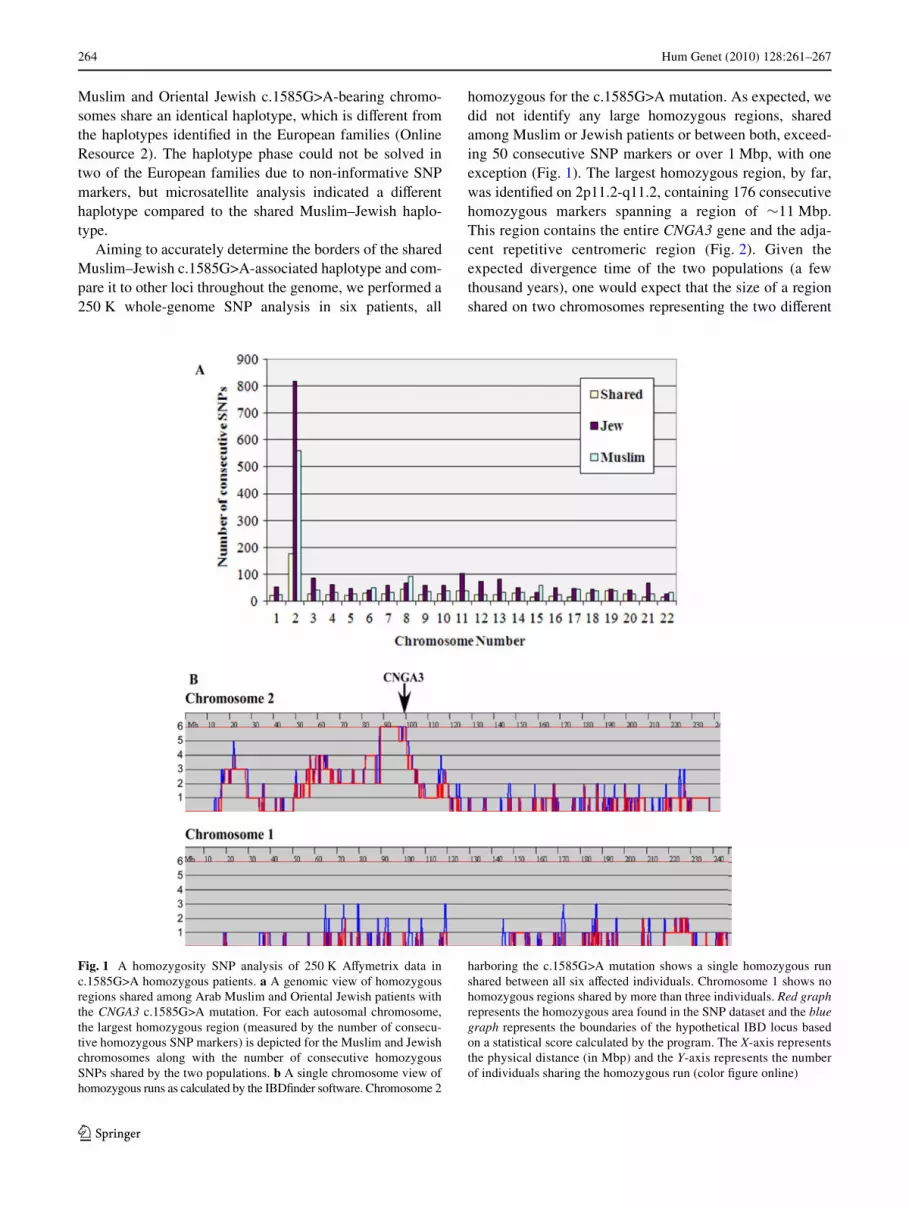
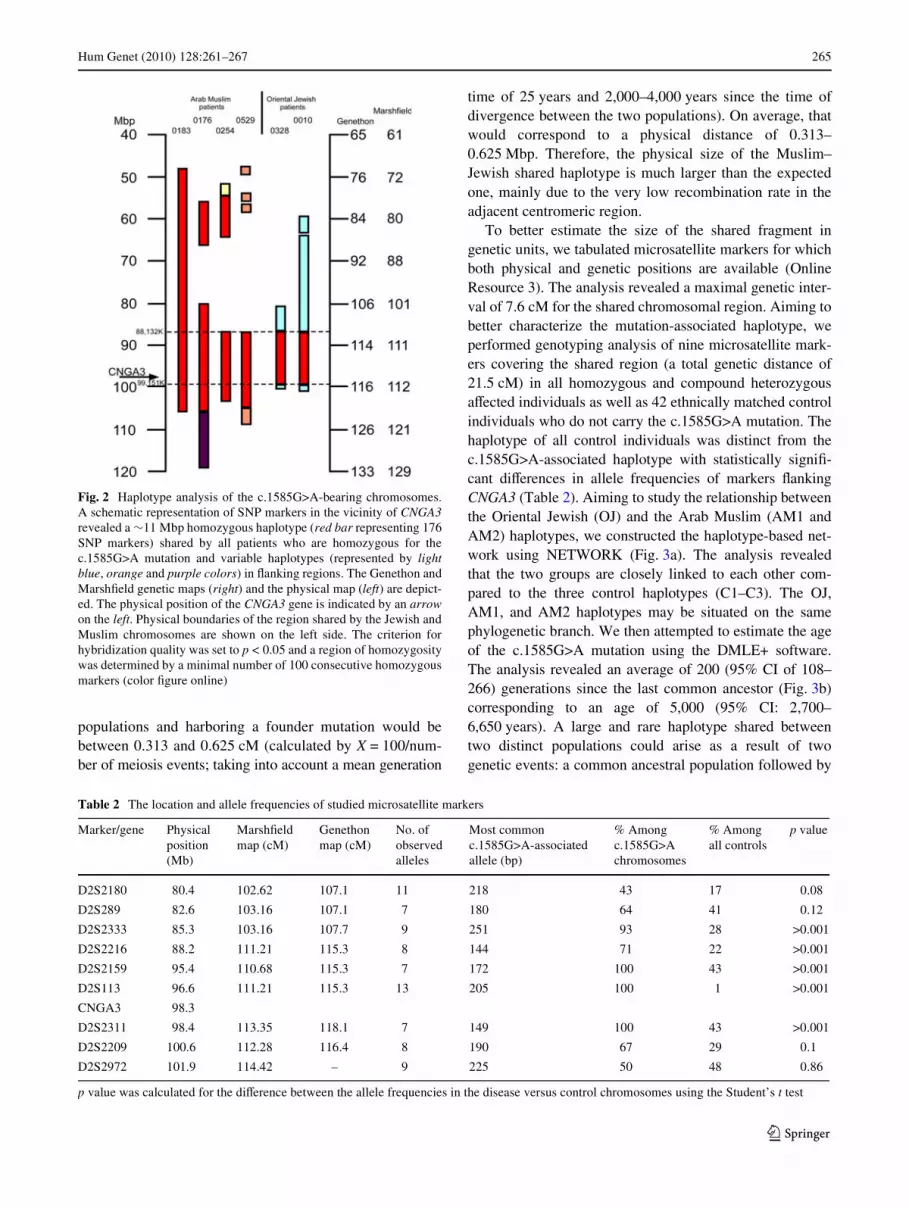
homozygous for the c.1585G>A mutation. As expected, wedid not identify any large homozygous regions, sharedamong Muslim or Jewish patients or between both, exceed-ing 50 consecutive SNP markers or over 1 Mbp, with oneexception (Fig. 1). The largest homozygous region, by far,was identiWed on 2p11.2-q11.2, containing 176 consecutivehomozygous markers spanning a region of »11 Mbp.This region contains the entire CNGA3 gene and the adja-cent repetitive centromeric region (Fig. 2). Given theexpected divergence time of the two populations (a fewthousand years), one would expect that the size of a regionshared on two chromosomes representing the two diVerent
Fig. 1 A homozygosity SNP analysis of 250 K AVymetrix data inc.1585G>A homozygous patients. a A genomic view of homozygousregions shared among Arab Muslim and Oriental Jewish patients withthe CNGA3 c.1585G>A mutation. For each autosomal chromosome,the largest homozygous region (measured by the number of consecu-tive homozygous SNP markers) is depicted for the Muslim and Jewishchromosomes along with the number of consecutive homozygousSNPs shared by the two populations. b A single chromosome view ofhomozygous runs as calculated by the IBDWnder software. Chromosome 2
harboring the c.1585G>A mutation shows a single homozygous runshared between all six aVected individuals. Chromosome 1 shows nohomozygous regions shared by more than three individuals. Red graphrepresents the homozygous area found in the SNP dataset and the bluegraph represents the boundaries of the hypothetical IBD locus basedon a statistical score calculated by the program. The X-axis representsthe physical distance (in Mbp) and the Y-axis represents the numberof individuals sharing the homozygous run (color Wgure online)
123
Hum Genet (2010) 128:261–267 265
populations and harboring a founder mutation would bebetween 0.313 and 0.625 cM (calculated by X = 100/num-ber of meiosis events; taking into account a mean generation
time of 25 years and 2,000–4,000 years since the time ofdivergence between the two populations). On average, thatwould correspond to a physical distance of 0.313–0.625 Mbp. Therefore, the physical size of the Muslim–Jewish shared haplotype is much larger than the expectedone, mainly due to the very low recombination rate in theadjacent centromeric region.
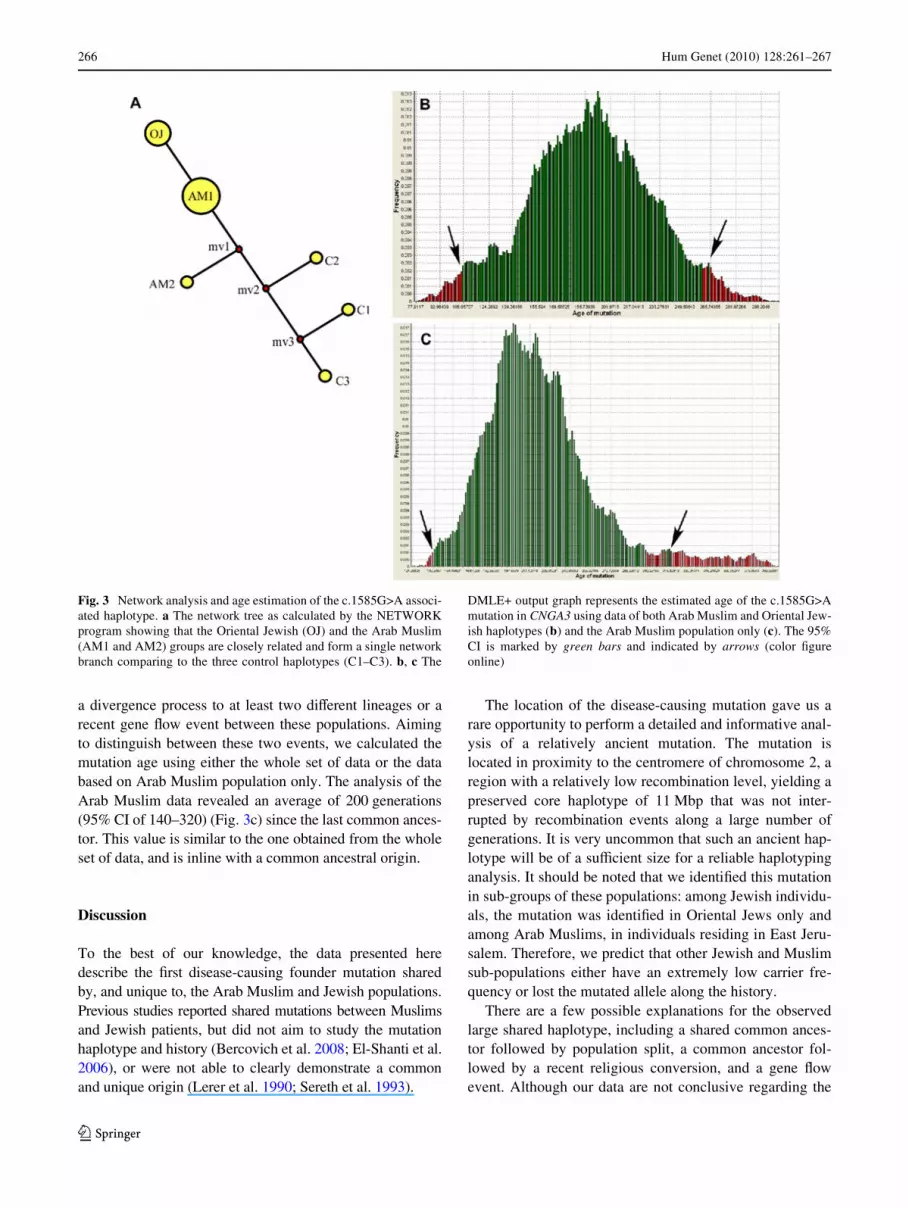
To better estimate the size of the shared fragment ingenetic units, we tabulated microsatellite markers for whichboth physical and genetic positions are available (OnlineResource 3). The analysis revealed a maximal genetic inter-val of 7.6 cM for the shared chromosomal region. Aiming tobetter characterize the mutation-associated haplotype, weperformed genotyping analysis of nine microsatellite mark-ers covering the shared region (a total genetic distance of21.5 cM) in all homozygous and compound heterozygousaVected individuals as well as 42 ethnically matched controlindividuals who do not carry the c.1585G>A mutation. Thehaplotype of all control individuals was distinct from thec.1585G>A-associated haplotype with statistically signiW-cant diVerences in allele frequencies of markers XankingCNGA3 (Table 2). Aiming to study the relationship betweenthe Oriental Jewish (OJ) and the Arab Muslim (AM1 andAM2) haplotypes, we constructed the haplotype-based net-work using NETWORK (Fig. 3a). The analysis revealedthat the two groups are closely linked to each other com-pared to the three control haplotypes (C1–C3). The OJ,AM1, and AM2 haplotypes may be situated on the samephylogenetic branch. We then attempted to estimate the ageof the c.1585G>A mutation using the DMLE+ software.The analysis revealed an average of 200 (95% CI of 108–266) generations since the last common ancestor (Fig. 3b)corresponding to an age of 5,000 (95% CI: 2,700–6,650 years). A large and rare haplotype shared betweentwo distinct populations could arise as a result of twogenetic events: a common ancestral population followed by
Fig. 2 Haplotype analysis of the c.1585G>A-bearing chromosomes.A schematic representation of SNP markers in the vicinity of CNGA3revealed a »11 Mbp homozygous haplotype (red bar representing 176SNP markers) shared by all patients who are homozygous for thec.1585G>A mutation and variable haplotypes (represented by lightblue, orange and purple colors) in Xanking regions. The Genethon andMarshWeld genetic maps (right) and the physical map (left) are depict-ed. The physical position of the CNGA3 gene is indicated by an arrowon the left. Physical boundaries of the region shared by the Jewish andMuslim chromosomes are shown on the left side. The criterion forhybridization quality was set to p < 0.05 and a region of homozygositywas determined by a minimal number of 100 consecutive homozygousmarkers (color Wgure online)
Table 2 The location and allele frequencies of studied microsatellite markers
p value was calculated for the diVerence between the allele frequencies in the disease versus control chromosomes using the Student’s t test
Marker/gene Physical position (Mb)
MarshWeld map (cM)
Genethon map (cM)
No. of observed alleles
Most common c.1585G>A-associated allele (bp)
% Among c.1585G>A chromosomes
% Among all controls
p value
D2S2180 80.4 102.62 107.1 11 218 43 17 0.08
D2S289 82.6 103.16 107.1 7 180 64 41 0.12
D2S2333 85.3 103.16 107.7 9 251 93 28 >0.001
D2S2216 88.2 111.21 115.3 8 144 71 22 >0.001
D2S2159 95.4 110.68 115.3 7 172 100 43 >0.001
D2S113 96.6 111.21 115.3 13 205 100 1 >0.001
CNGA3 98.3
D2S2311 98.4 113.35 118.1 7 149 100 43 >0.001
D2S2209 100.6 112.28 116.4 8 190 67 29 0.1
D2S2972 101.9 114.42 – 9 225 50 48 0.86
123
266 Hum Genet (2010) 128:261–267
a divergence process to at least two diVerent lineages or arecent gene Xow event between these populations. Aimingto distinguish between these two events, we calculated themutation age using either the whole set of data or the databased on Arab Muslim population only. The analysis of theArab Muslim data revealed an average of 200 generations(95% CI of 140–320) (Fig. 3c) since the last common ances-tor. This value is similar to the one obtained from the wholeset of data, and is inline with a common ancestral origin.
Discussion
To the best of our knowledge, the data presented heredescribe the Wrst disease-causing founder mutation sharedby, and unique to, the Arab Muslim and Jewish populations.Previous studies reported shared mutations between Muslimsand Jewish patients, but did not aim to study the mutationhaplotype and history (Bercovich et al. 2008; El-Shanti et al.2006), or were not able to clearly demonstrate a commonand unique origin (Lerer et al. 1990; Sereth et al. 1993).
The location of the disease-causing mutation gave us arare opportunity to perform a detailed and informative anal-ysis of a relatively ancient mutation. The mutation islocated in proximity to the centromere of chromosome 2, aregion with a relatively low recombination level, yielding apreserved core haplotype of 11 Mbp that was not inter-rupted by recombination events along a large number ofgenerations. It is very uncommon that such an ancient hap-lotype will be of a suYcient size for a reliable haplotypinganalysis. It should be noted that we identiWed this mutationin sub-groups of these populations: among Jewish individu-als, the mutation was identiWed in Oriental Jews only andamong Arab Muslims, in individuals residing in East Jeru-salem. Therefore, we predict that other Jewish and Muslimsub-populations either have an extremely low carrier fre-quency or lost the mutated allele along the history.
There are a few possible explanations for the observedlarge shared haplotype, including a shared common ances-tor followed by population split, a common ancestor fol-lowed by a recent religious conversion, and a gene Xowevent. Although our data are not conclusive regarding the
Fig. 3 Network analysis and age estimation of the c.1585G>A associ-ated haplotype. a The network tree as calculated by the NETWORKprogram showing that the Oriental Jewish (OJ) and the Arab Muslim(AM1 and AM2) groups are closely related and form a single networkbranch comparing to the three control haplotypes (C1–C3). b, c The
DMLE+ output graph represents the estimated age of the c.1585G>Amutation in CNGA3 using data of both Arab Muslim and Oriental Jew-ish haplotypes (b) and the Arab Muslim population only (c). The 95%CI is marked by green bars and indicated by arrows (color Wgureonline)
123

Hum Genet (2010) 128:261–267 267
history of the studied haplotype, it does not support a recentgene Xow event between the two populations, since the agecalculated based on data obtained from a single populationwas not diVerent from the one obtained from both popula-tions. It is impossible, however, to exclude an ancient geneXow event that has occurred in proximity or within the timeframe of the divergence of these two populations. It is rea-sonable to assume that ancient gene Xow events do occuralong such process of separation and are therefore diYcultor even impossible to distinguish between the two. A recentreligious conversion can not be excluded, but this is notsupported by the familial history as reported by the patientsand their family members.
In-line with previously reported data based on the mito-chondrial and Y-chromosome studies (Bonne-Tamir et al.1986; Hammer et al. 2000; Nebel et al. 2000, 2001), ourresults suggest a shared origin between the two Middle-Eastern populations. Based on this Wnding, we propose thatthe shared mutation arose before the divergence of the Mid-dle-Eastern Arab Muslim and Jewish populations andtherefore represent a true disease-causing founder mutationin these populations.
Acknowledgments We thank the patients and their families for theirparticipation in the study, Prof. Bernd Wissinger, Dr. Marina Faermanand Prof. Moshe Sharon for advice, and Liat Ben-Avi and Dr. IsraelaLerer for technical support. This work was supported by the Founda-tion Fighting Blindness (FFB-Grant number BR-GE-0607-0395-HUJ); the Chief Scientist OYce of the Israeli Ministry of Health (Grantnumber 3000003241); and the Yedidut research grant.We declare that the experiments were in compliance with the HelsinkiDeclaration and were approved by the Institutional Helsinki commit-tee.
ConXict of interest statement The authors declare that they have noconXict of interest.
References
Bandelt HJ, Forster P, Rohl A (1999) Median-joining networks forinferring intraspeciWc phylogenies. Mol Biol Evol 16:37–48
Bercovich D, Elimelech A, Yardeni T, Korem S, Zlotogora J, Gal N,Goldstein N, Vilensky B, Segev R, Avraham S, Loewenthal R,Schwartz G, Anikster Y (2008) A mutation analysis of the phen-ylalanine hydroxylase (PAH) gene in the Israeli population. AnnHum Genet 72:305–309
Bonne-Tamir B, Johnson MJ, Natali A, Wallace DC, Cavalli-SforzaLL (1986) Human mitochondrial DNA types in two Israeli popu-lations—a comparative study at the DNA level. Am J Hum Genet38:341–351
Briggs CE, Rucinski D, Rosenfeld PJ, Hirose T, Berson EL, Dryja TP(2001) Mutations in ABCR (ABCA4) in patients with Stargardtmacular degeneration or cone-rod degeneration. Invest OphthalmolVis Sci 42:2229–2236
El-Shanti H, Abdel Majeed H, El-Khateeb M (2006) Familial Mediter-ranean fever in Arabs. Lancet 367:1016–1024
Goldschmidt E, Ronen A, Ronen I (1960) Changing marriage systemin the Jewish communities in Israel. Ann Hum Genet 24:191–204
Haider NB, Jacobson SG, Cideciyan AV, Swiderski R, Streb LM,Searby C, Beck G, Hockey R, Hanna DB, Gorman S, Duhl D,Carmi R, Bennett J, Weleber RG, Fishman GA, Wright AF, StoneEM, SheYeld VC (2000) Mutation of a nuclear receptor gene,NR2E3, causes enhanced S cone syndrome, a disorder of retinalcell fate. Nat Genet 24:127–131
Hammer MF, Redd AJ, Wood ET, Bonner MR, Jarjanazi H, Karafet T,Santachiara-Benerecetti S, Oppenheim A, Jobling MA, Jenkins T,Ostrer H, Bonne-Tamir B (2000) Jewish and Middle Eastern non-Jewish populations share a common pool of Y-chromosome biall-elic haplotypes. Proc Natl Acad Sci USA 97:6769–6774
Jaber L, Bailey-Wilson JE, Haj-Yehia M, Hernandez J, Shohat M(1994) Consanguineous matings in an Israeli-Arab community.Arch Pediatr Adolesc Med 148:412–415
Kohl S, Marx T, Giddings I, Jagle H, Jacobson SG, Apfelstedt-Sylla E,Zrenner E, Sharpe LT, Wissinger B (1998) Total colour blindnessis caused by mutations in the gene encoding the alpha-subunit ofthe cone photoreceptor cGMP-gated cation channel. Nat Genet19:257–259
Lerer I, Cohen S, Chemke M, Sanilevich A, Rivlin J, Golan A, YahavJ, Friedman A, Abeliovich D (1990) The frequency of the deltaF508 mutation on cystic Wbrosis chromosomes in Israeli families:correlation to CF haplotypes in Jewish communities and Arabs.Hum Genet 85:416–417
Nebel A, Filon D, Weiss DA, Weale M, Faerman M, Oppenheim A,Thomas MG (2000) High-resolution Y chromosome haplotypesof Israeli and Palestinian Arabs reveal geographic substructureand substantial overlap with haplotypes of Jews. Hum Genet107:630–641
Nebel A, Filon D, Brinkmann B, Majumder PP, Faerman M, OppenheimA (2001) The Y chromosome pool of Jews as part of the geneticlandscape of the Middle East. Am J Hum Genet 69:1095–1112
Niell BL, Long JC, Rennert G, Gruber SB (2003) Genetic anthropol-ogy of the colorectal cancer-susceptibility allele APC I1307 K:evidence of genetic drift within the Ashkenazim. Am J HumGenet 73:1250–1260
Nishiguchi KM, Sandberg MA, Gorji N, Berson EL, Dryja TP (2005)Cone cGMP-gated channel mutations and clinical Wndings inpatients with achromatopsia, macular degeneration, and otherhereditary cone diseases. Hum Mutat 25:248–258
Peretz H, Mulai A, Usher S, Zivelin A, Segal A, Weisman Z, Mittel-man M, Lupo H, Lanir N, Brenner B, Shpilberg O, Seligsohn U(1997) The two common mutations causing factor XI deWciencyin Jews stem from distinct founders: one of ancient Middle East-ern origin and another of more recent European origin. Blood90:2654–2659
Reeve JP, Rannala B (2002) DMLE+: Bayesian linkage disequilibriumgene mapping. Bioinformatics 18:894–895
Sereth H, Shoshani T, Bashan N, Kerem BS (1993) Extended haplo-type analysis of cystic Wbrosis mutations and its implications forthe selective advantage hypothesis. Hum Genet 92:289–295
Shahin H, Walsh T, Sobe T, Lynch E, King MC, Avraham KB, KanaanM (2002) Genetics of congenital deafness in the Palestinian pop-ulation: multiple connexin 26 alleles with shared origins in theMiddle. East Hum Genet 110:284–289
Sharon D, Sandberg MA, Caruso RC, Berson EL, Dryja TP (2003)Shared mutations in NR2E3 in enhanced S-cone syndrome, Gold-mann-Favre syndrome, and many cases of clumped pigmentaryretinal degeneration. Arch Ophthalmol 121:1316–1323
Webster AR, Heon E, Lotery AJ, Vandenburgh K, Casavant TL, OhKT, Beck G, Fishman GA, Lam BL, Levin A, Heckenlively JR,Jacobson SG, Weleber RG, SheYeld VC, Stone EM (2001)An analysis of allelic variation in the ABCA4 gene. InvestOphthalmol Vis Sci 42:1179–1189
Zlotogora J (2002) What is the birth defect risk associated with consan-guineous marriages? Am J Med Genet 109:70–71
123

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
Top Related
Copyright © 2022 FDOKUMEN

