







Bahasa
Halaman
Hukum


www.elsevier.com/locate/jmicmeth
Journal of Microbiological Methods 57 (2004) 197–206
Amplified ribosomal DNA restriction analysis for the
characterization of Azotobacteraceae: a contribution to the study of
these free-living nitrogen-fixing bacteria
Lucia Aquilantia,*, Ilaria Mannazzua, Roberto Papaa,Lucia Cavalcab, Francesca Clementia
aDipartimento di Scienze degli Alimenti, Universita Politecnica delle Marche, via Brecce Bianche, 60131 Ancona, ItalybDipartimento di Scienze e Tecnologie Ambientali e Microbiologiche, Universita di Milano, via Celoria 2, 20133 Milan, Italy
Received 2 December 2003; received in revised form 8 January 2004; accepted 8 January 2004
Abstract
A 16S rRNA gene-based fingerprinting method was developed for the identification of Azotobacteraceae and tested onto 48
soil isolates and 28 reference strains belonging to the free-living nitrogen-fixing bacterial group and to the most common
species found in soil samples. According to this method, the 16S rRNA gene was amplified using universal primers for
Eubacteria and PCR products were subsequently digested with RsaI, HhaI, HpaII, FnuDII, and AluI. The analysis of the
restriction profiles obtained showed that the method is able to define a unique species-specific phylotype (SSP) for each of the
eight Azotobacteraceae species tested. Cluster analysis was successfully employed for the identification of members of the
family Azotobacteraceae, being assignation into species of the isolates confirmed by means of partial 16S rRNA gene
sequencing.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Azotobacter; Azotobacteraceae; ARDRA; Cluster analysis
1. Introduction within this family: Azotobacter and Azomonas. The
The family Azotobacteraceae belongs to the g-
subclass of the Proteobacteria (Tchan, 1984) and is
composed of free nitrogen-fixing bacteria (FNFB)
commonly living dispersed in soil, water and sedi-
ments. Studies on the rRNA cistrons (De Smedth et
al., 1980) and the application of molecular taxonomic
techniques led to the identification of two genera
0167-7012/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.mimet.2004.01.006
* Corresponding author. Tel.: +39-712204985; fax: +39-
712204858.
E-mail address: [email protected] (L. Aquilanti).
genus Azotobacter comprises seven species, namely
Azotobacter chroococcum, Azotobacter vinelandii,
Azotobacter beijerinckii, Azotobacter paspali, Azoto-
bacter armeniacus, Azotobacter nigricans (Tchan and
New, 1984a), and Azotobacter salinestris (Page and
Shivprasad, 1991), while genus Azomonas comprises
three species, namely Azomonas macrocytogenes,
Azomonas agilis, and Azomonas insignis (Tchan and
New, 1984b).
The positive interactions between members of the
Azotobacteraceae family and a variety of field grown
crops (Jackson et al., 1964; Rovira, 1965; Denarie and

L. Aquilanti et al. / Journal of Microbiological Methods 57 (2004) 197–206198
Blanchere, 1966), their capability to synthesize anti-
biotics and growth-promoting substances (Harper and
Lynch, 1979; Pandey and Kumar, 1990) and their
ability to fix atmospheric nitrogen non-simbiotically
(Gouri and Jagasnnatathan, 1995; Mrkovacki et al.,
1996; Zahir and Arshad, 1996; Zahir et al., 1996)
were widely taken under study. At present, strains
belonging to the species A. vinelandii and A. chroo-
coccum are proficiently employed as soil inoculants in
rainy areas and in warm and alkaline soils (Pandey et
al., 1998). Moreover, some species of Azotobactera-
ceae were exploited for the production of compounds
of commercial interest, such as polysaccharides
(Clementi, 1997; Sabra et al., 2001), pigments and
vitamins (Mishustin and Shilnikova, 1971; Pandey et
al., 1998).
All these evidences justify the interest recently
arisen on the development of analytical tools suitable
for a rapid and reliable identification of these bacteria
and for their study in soil microbial communities.
In this context, the potential of amplified ribosomal
DNA restriction analysis (ARDRA) for the character-
ization and differentiation of Azotobacteraceae was
evaluated, and the more appropriate tool for the elab-
oration of data was assessed. This approach allowed a
sound method for the unambiguous identification of
Azotobacteraceae to be developed, also in view of
deepening knowledge on the ecology of these micro-
organisms and their biotechnological exploitations.
2. Materials and methods
2.1. Bacterial strains and culture conditions
We analysed 76 cultures, 28 of which were
reference strains. Among these: A. vinelandii
DSM576, DSM2289, DSM2290, DSM87; A. chroo-
coccum DSM2286, DSM377; A. armeniacus
DSM2284, A. beijerinkii DSM378, Azorhizophilus
paspali DSM2283, A. macrocytogenes DSM721, A.
agilis DSM375, A. insignis DSM1845, Azospirillum
brasilense DSM1690, Azospirillum lipoferum
DSM1691, Azospirillum amazonense DSM2787,
Azospirillum halopraeferens DSM3675, Beijerinckia
indica DSM1715, Beijerinckia mobilis DSM2326,
Beijerinckia fluminensis DSM2327, and Beijerinckia
derxii DSM2329 were obtained from Deutsche
Sammlung von Mikrorganismen und Zellkulturen
(Braunschweig, Germany); Agrobacterium tumefa-
ciens AT, Agrobacterium rhizogenes AR, Agrobacte-
rium radiobacter DCBA10, Sinorhizobium meliloti
DCBA8, Rhizobium leguminosarum DCBA11, Pseu-
domonas corrugata PC, Pseudomonas fluorescens
DCBA14, and Pseudomonas tolasii DCBA4 were
provided by the Department of Chemistry and Agri-
cultural Biotechnologies, University of Pisa, Italy.
The remaining 48 cultures were soil isolates that
originated from central Italian soils, preliminarily
characterized as described by Aquilanti et al. (sub-
mitted for publication).
Bacterial strains were cultured overnight on Tryptic
Soy Agar (TSA) or broth (TSB) (Oxoid, Basingstoke,
Hampshire, England) at 30 jC and stored frozen in
50% (w/v) glycerol in TSB at � 80 jC.
2.2. ARDRA
In order to choose the restriction enzymes to be used
for ARDRA, the 16S rRNA sequences of Azotobacter
and Azomonas retrieved from the GenBank were vir-
tually restricted using the Genetool software (Version
2.0) (Wishart et al., 2000). Crude DNAwas extracted as
previously described (Aquilanti et al., submitted for
publication). The 16S rRNA gene was amplified by
means of universal primers 27f and 1495r (Weisburg et
al., 1991) in 50 Al reaction mixture containing 1 U of
Taq DNA polymerase (Amersham Biosciences,
Uppsala, Sweden), 1� reaction buffer (10 mM Tris–
HCl pH 8.8, 1.5 mMMgCl2, 50 mM KCl, 0,1% Triton
X-100), 200 AM of the four dNTPs, and 2 AM of each
primer. The PCR reactions were run for 35 cycles on a
Applied Biosystem Gene Amp PCR System 9700
(Foster City, CA, USA) as follows: denaturation at 94
jC for 1 min, annealing at 55 jC for 1 min, elongation
at 72 jC for 2 min. An initial denaturation step at 95 jCfor 4 min and a final extension step at 72 jC for 15 min
were also performed. The PCR products were digested
with the tretrameric restriction enzymes (TRE) RsaI,
HhaI, HpaII, AluI, and FnuDII (Roche Diagnostics,
Mannheim, Germany). Five units of each enzyme were
added to 12.5 Al of the amplification product and
incubated for 5 h at 37 jC in a total volume of 25 Al.The digests were resolved by electrophoresis on a 2.5%
(w/v) agarose gel in 1� Tris–borate–EDTA buffer
(TBE, 0.09MTris base, 0.09M sodium borate, 2.5mM

L. Aquilanti et al. / Journal of Microbiological Methods 57 (2004) 197–206 199
EDTA, pH 8.3) at 3.2 V/cm for 4 h, against a 50-bp
DNA ladder (Amersham Pharmacia, Uppsala, Swe-
den). Gels were stained in ethidium bromide for 15 min
and rinsed for 5 min in distilled water. Gel electronic
images were visualized by means of ImageMaster VDS
(Amersham Pharmacia), captured with LISCAP soft-
ware v.1.0 (Amersham Pharmacia), and stored as TIFF
files. Digital images were normalized with the above
mentioned 50-bp DNA size marker and analyzed with
GelCompar software v. 4.0 (Applied Maths, Kortrijk,
Belgium).
2.3. Statistical analyses of ARDRA patterns
For data scoring only fragments z 50-bp were
considered. A binary data matrix was constructed on
the basis of the presence or absence of each band
which was scored 1 or 0, respectively. The different
restriction patterns obtained were defined as different
haplotypes for each enzyme and all the strains char-
acterized by the same combination of haplotypes
obtained with the five enzymes were grouped togeth-
er, defining a phylotype, which is often used to term
the 16S rRNA clones differentiated by restriction
Table 1
Tetrameric restriction enzymes (TRE) employed in the ARDRA analysis
TRE Restriction fragments H Azotobacteraceae
No. %
RsaI 21 16 1.798 a 650–350–220–1
b 470–360–190–1
c 900–340–140–1
HhaI 27 24 2.496 a1 410–280–180–1
b1 580–410–280–2
e1 580–410–280–1
d1 580–410–280–1
c1 410–370–280–2
f1 410–380–280–2
g1 560–410–280–2
h1 410–370–280–2
AluI 27 23 2.347 b3 630–240–210–1
HpaII 27 22 2.374 a2 550–400–130–1
d2 600–400–130–1
c2 680–400–130–8
f2 550–450–1130–
g2 680–500–130–8
h2 680–500–170–1
FnuDII 23 15 2.298 c4 400–300–190–1
No.: number of restriction fragments obtained; %: percentage of polymorp
Shannon–Weaver index.
enzymes (Tiedje et al., 1999). Haplotype diversity
(H) was estimated using the Shannon–Weaver index
(Shannon and Weaver, 1949; Ludwig and Reynolds,
1988) which was calculated as: H =�Spilnpi, wherepi is the frequency of the ith haplotype.
All the strains characterized by the same combina-
tion of haplotypes obtained with the five enzymes
were grouped together, defining a phylotype.
Genetic distance between phylotypes was calculat-
ed using the Nei and Li (Nei and Li, 1979) coefficient
and the resulting distance matrix was used for the
UPGMA cluster analysis (Sokal and Sneath, 1963)
and the Neighbor-joining mid-point analysis (Rohlf,
1993; Saitou and Nei, 1987) using TREECON for
Windows (Version 1.2) (Van Der Peer and DeWachter,
1994) and NTSYS-PC (Version 1.6) (Rohlf, 1993),
respectively. The consistency of each node was esti-
mated by bootstrapping over markers (Felsenstein,
1985) using 1000 pseudoreplications.
2.4. Sequence analyses
The nearly complete sequence of the 16S rRNA
gene of A. vinelandii DSM576, A. chroococcum
haplotypes Species
50–120-bp Azotobacter spp.; Azomonas macrocytogenes
40–120-bp Azomonas agilis
20-bp Azomonas insignis
60–120-bp Azotobacter vinelandii
60-bp Azotobacter chroococcum
90-bp Azotobacter beijerinckii
80–160-bp Azorizhophilus paspali
60–180–160-bp Azotobacter armeniacus
60–200-bp Azomonas macrocytogenes
60-bp Azomonas agilis
60–220-bp Azomonas insignis
60–80-bp Azotobacter armeniacus
10–80-bp Azotobacter vinelandii
10–80-bp Azorhizophilus paspali
0-bp Azotobacter armeniacus
110–80-bp Azomonas macrocytogenes
0-bp Azomonas agilis
40–80-bp Azomonas insignis
50–120-bp Azotobacter armeniacus
hic restriction fragments; H: diversity calculated on the basis of the
L. Aquilanti et al. / Journal of Microbiological Methods 57 (2004) 197–206200
DSM2286, A. beijerinckii DSM378, and A. macro-
cytogenes DSM721 was amplified using universal
primers 27f and 1495r (Weisburg et al., 1991) and the
internal primers Pi-1 (5V-AGC AGT GGG GAA TAT
TGG AC-3V) and Pi-2 (5V-GCTAAC TTC GTG CCA
GCA G-3V). A rRNA gene fragment of about 300-bp
was amplified with universal primers Y1 and/or Y2
(Young et al., 1991) from one isolate for each isolate
phylotype (IP) selected. PCR products were purified
using the GFX PCR DNA and Gel Band Purification
Kit (Amersham Pharmacia Biotech, Uppsala, Sweden)
according to the instructions and sent to MWG Biotech
(Ebersberg, Germany) for sequencing.
The partial rRNA gene sequences of the isolates
were preliminarily compared with those present in
the Basic BLAST search (Altschul et al., 1997),
and then aligned using the ClustalW program
(Higgins et al., 1992) available at the European
Bioinformatic Institute website (http://www.ebi.a-
c.uk/clustalw/) with the sequences retrieved from
the GenBank database (Benson et al., 2003) avail-
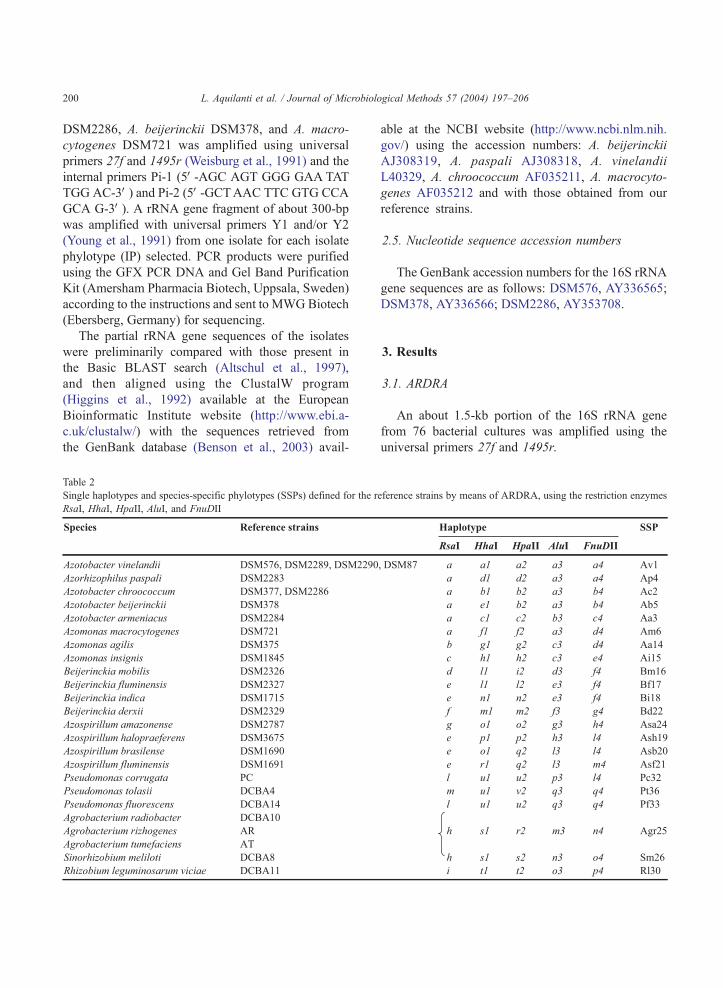
Table 2
Single haplotypes and species-specific phylotypes (SSPs) defined for the r
RsaI, HhaI, HpaII, AluI, and FnuDII
able at the NCBI website (http://www.ncbi.nlm.nih.
gov/) using the accession numbers: A. beijerinckii
AJ308319, A. paspali AJ308318, A. vinelandii
L40329, A. chroococcum AF035211, A. macrocyto-
genes AF035212 and with those obtained from our
reference strains.
2.5. Nucleotide sequence accession numbers
The GenBank accession numbers for the 16S rRNA
gene sequences are as follows: DSM576, AY336565;
DSM378, AY336566; DSM2286, AY353708.
3. Results
3.1. ARDRA
An about 1.5-kb portion of the 16S rRNA gene
from 76 bacterial cultures was amplified using the
universal primers 27f and 1495r.
eference strains by means of ARDRA, using the restriction enzymes

Table 3
Haplotypes and total isolates phylotypes (IPs) defined for soil
isolates by means of ARDRA, using the five restriction enzymes
RsaI, HhaI, HpaII, AluI, and FnuDII
Soil isolates Haplotype IP
RsaI HhaI HpaII AluI FnuDII
BB4, 6, 8;
AX16; AZ2, 8
a b1 b2 a3 a4 L7
BN16 a f1 b2 a3 a4 L35
AY7, 14 a b1 b2 b3 a4 L9
BN9; BZ55 a f1 b2 w3 a4 L12
AX8, BB3 a b1 b2 a3 b4 Ac2
BG8,18; BP11 a f1 b2 a3 d4 L13
BG10,13, 14, 22 a f1 b2 y3 l4 L10
BL46 h s1 ae2 m3 n4 L28
BD2; CH10 h w1 aa2 m3 t4 L27
CD20 l y1 ab2 r3 t4 L34
M1 r w1 ac2 z3 t4 L41
CM8 a b1 b2 a3 t4 L8
CH13 p j1 w2 u3 t4 L39
CH30 f v1 y2 t3 t4 L23
BH16 o x1 ad2 a3 u4 L38
BH23 a b1 ad2 j3 u4 L11
BH15 o x1 ad2 j3 u4 L37
BF15 s aa1 b2 k3 v4 L42
BY3, CA24 I ab1 b2 x3 z4 L31
BL22; BD2; BM33;
BO7, 18, 27, 41;
BQ7; BY25;
BV62; BT1; CA28
h s1 r2 m3 n4 Agr25
BH27, 29, 34 h s1 r2 m3 n4-I Agr25-I
BO18; BQ7;
BT25; BY25
h s1 r2 m3 n4-II Agr25-II
L. Aquilanti et al. / Journal of Microbiological Methods 57 (2004) 197–206 201
The results of virtual restriction with the Genetool
software onto the sequences of A. paspali, A. chroo-
coccum, A. vinelandii, A. beijerinckii, and A. macro-
cytogenes (accession numbers: 18072914, 2661011,
790695, 18072421, and 2661012, respectively) led
to the selection of five restriction enzymes, namely
RsaI, HhaI, HpaII, AluI, and FnuDII, to be employed
for the characterization of members of the family
Azotobacteraceae. Thus, the amplification products
obtained by using the above mentioned primers
on all the cultures under study were subjected to
restriction analysis yielding a total of 125 fragments.
The percentages of polymorphic fragments obtained
with each enzyme are shown (Table 1).
The sum of the sizes of the restriction fragments
was consistent with the size of the undigested ampli-
fication products. Slight discrepancies were likely due
to the production of small restriction fragments (V 50
bp), undetectable on the agarose gels.
3.2. Haplotype analysis
In order to assess the existence of species-specific
restriction patterns the analysis of haplotypes was
performed for each of the enzymes utilized.
As reported (Table 1) the enzyme RsaI showed the
lowest diversity (H = 1.80). Among RsaI haplotypes,
a, b, and c referred uniquely to members of the family
Azotobacteraceae (Table 2). In particular, all the
Azotobacter species tested plus A. macrocytogenes
produced the haplotype a, while A. agilis and A.
insignis produced two species-specific haplotypes, b
and c, respectively (Table 1). Reference strains not
belonging to the Azotobacteraceae family produced
either species specific or nonspecific RsaI haplotypes
(Table 2).
Among the 48 soil isolates, 22 showed the RsaI
haplotype a, while none showed the haplotypes b or c
(Table 3).
HhaI and HpaII were characterized by high Shan-
non–Weaver index and showed the highest level of
resolution within the family Azotobacteraceae (Table
1).HhaI allowed the individuation of a species-specific
haplotype for each of the eight species within the
family of interest, while HpaII allowed six species-
specific haplotypes to be obtained (Table 1), being the
haplotype b2 common to the species A. chroococcum
and A. beijerinckii (Table 2). Both these two enzymes
revealed a high discriminatory power also within the
other genera considered (Table 1).
By contrast, AluI and FnuDII revealed to be the
less discriminating. In particular, restriction analysis
of the 16S rRNA gene of the reference strains with
AluI permitted to obtain 27 DNA restriction frag-
ments, combined in 14 haplotypes. Nine of them
were species-specific but only b3 was related to the
family Azotobacteraceae, being species-specific for
A. armeniacus (Table 1). Similarly, FnuDII produced
a total of 14 haplotypes, 2 of which, c4 and e4,
specific for A. armeniacus and A. insignis, respec-
tively (Table 2).
3.3. Phylotype analysis
Phylotypes were built up grouping together the
strains with the same combination of haplotypes
L. Aquilanti et al. / Journal of Microbiological Methods 57 (2004) 197–206202
(Table 2). The reference strains tested, belonging to 24
species, produced 22 different phylotypes. Of these,
all but one were species-specific, whereas the phylo-
type Agr25 was genus-specific for Agrobacterium,
including the three species A. tumefaciens, A. rhizo-
genes, and A. radiobacter (Table 2). Interestingly,
within the family Azotobacteraceae, eight SSPs were
found. More in detail, Av1, Ac2, Aa3, Ap4, and Ab5
defined the species belonging to the genus Azotobac-
ter, namely A. vinelandii, A. chroococcum, A. arme-
niacus, A. paspali, and A. beijerinckii, respectively,
while Am6, Aa14, and Ai15 were specific for the
three species of the genus Azomonas. On the basis of
the phylotype analysis, 2 out of the 48 isolates could
be ascribed to the species A. chrococcum and 12 to the
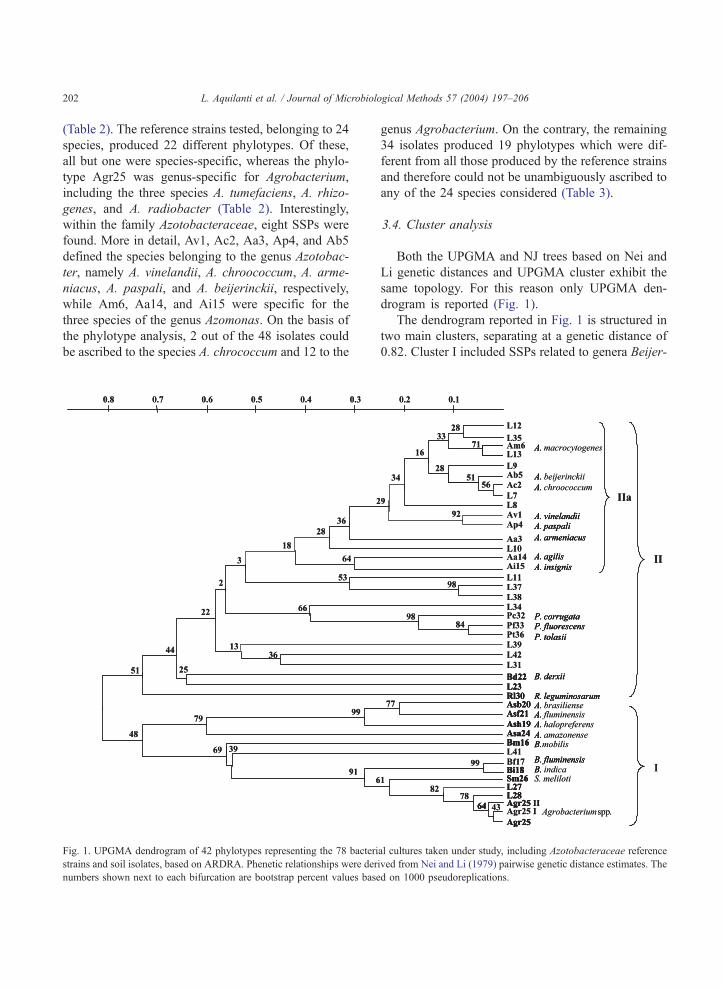
Fig. 1. UPGMA dendrogram of 42 phylotypes representing the 78 bacteri
strains and soil isolates, based on ARDRA. Phenetic relationships were der
numbers shown next to each bifurcation are bootstrap percent values base
genus Agrobacterium. On the contrary, the remaining
34 isolates produced 19 phylotypes which were dif-
ferent from all those produced by the reference strains
and therefore could not be unambiguously ascribed to
any of the 24 species considered (Table 3).
3.4. Cluster analysis
Both the UPGMA and NJ trees based on Nei and
Li genetic distances and UPGMA cluster exhibit the
same topology. For this reason only UPGMA den-
drogram is reported (Fig. 1).
The dendrogram reported in Fig. 1 is structured in
two main clusters, separating at a genetic distance of
0.82. Cluster I included SSPs related to genera Beijer-
al cultures taken under study, including Azotobacteraceae reference
ived from Nei and Li (1979) pairwise genetic distance estimates. The
d on 1000 pseudoreplications.

L. Aquilanti et al. / Journal of Microbiolo
inckia, Agrobacterium, Sinorhizobium, and Azospir-
illum along with three IPs, namely L41, L27, and L28.
Cluster II included all SSPs related to the family
Azotobacteraceae, and phylotypes related to genera
Pseudomonas (Pc32, Pf33, Pt36), B. derxii (Bd22),
and R. leguminosarum (Rl30). Fifteen soil IPs were
also included in this cluster (Fig. 1).
The eight Azotobacteraceae SSPs and seven IPs
were grouped in a single subcluster, named IIa
(d = 0.42; boot = 18%) (Fig. 1), separating from all
the other materials analysed. The Azotobacteraceae
subcluster was divided into two branches: one
(d = 0.30; boot = 64%) constituted by the two A. agilis
and A. insignis SSPs (Aa14, Ai15) and the other
(d = 0.35 boot = 28%) constituted by seven IPs (L7,
L9, L13, L12, L35, L8, and L10) plus all the Azoto-
bacter and A. macrocytogenes SSPs. Solidity within
the Azotobacteraceae subcluster was highlighted by
bootstrap values z 50% for half of the nodes and
50>xz 20 for most of the remaining ones.
The distance matrix, computed using the Nei and
Li coefficient on the basis of ARDRA restriction
profiles, was used to evaluate the genetic distance
between IPs and reference strains phylotypes. Seven
of the 19 phylotypes comprising soil isolates showed
a genetic distance V 0.20 to SSPs related to the
family under study (Table 4) and the corresponding
17 isolates were therefore ascribed to three species of
Azotobacteraceae. In particular, 13 isolates were
recognized as A. chroococcum, 2 as A. macrocyto-
genes, and 2 as A. beijerinckii.
Table 4
Assignation into species of soil isolates clustered in IPs included in
the Azotobacteraceae subcluster IIa
IP Soil strains Closest SSP Identification
Ac2 BB3, AX8 Ac2 (0.00) A. chroococcum
L7 BB4, BB6, BB8,
AX16, AZ2, AZ8
Ac2 (0.02) A. chroococcum
L8 CM8 Ac2 (0.12) A. chroococcum
L9 AY7, AY14 Ab5 (0.15) A. beijerinckii
L10 BG13 Am6 (0.26) A. macrocytogenes
L12 BN9, BZ55 Ac2 (0.17) A. chroococcum
L13 BG8, BG18, BP11 Am6 (0.04) A. macrocytogenes
L35 BN16 Ac2 (0.08) A. chroococcum
The closest SSP and the corresponding assigned species are shown.
Strains selected for sequencing are in bold, while in brackets are
shown the genetic distance values calculated on the basis of the Nie
and Li coefficient.
3.5. Sequence alignment
In order to confirm the efficacy of the identification
hypothesized by combining ARDRA and cluster anal-
ysis, we proceeded with the sequence analysis of one
representant for each IP assigned to the Azotobacter-
aceae family (Ac2, L7, L8, L9, L10, L12, L13, and
L35). Thus, an about 300-bp 16S rRNA gene fragment
was sequenced with universal primers Y1 and Y2 from
the eight isolates selected (Table 4). Due to the low
number of Azotobacteraceae rRNA gene sequences in
GenBank and EMBL databases, the 16S rRNA gene of
reference strains belonging to the four species identi-
fied in the population of soil isolates was also se-
quenced. Thus almost the entire rRNA gene of A.
vinelandii DSM576, A. chroococcum DSM2286, A.
beijerinckii DSM378, and A. macrocytogenes
DSM721 was amplified. The sequences obtained were
deposited in GenBank. The rRNA partial gene sequen-
ces of isolates and reference strains were subjected to
BLAST analysis and multiple alignment by means of
ClustalW. Assignation into species obtained by means
of ARDRA and cluster analysis was confirmed for
each of the eight isolates investigated, with a percent-
age of homology z 97% to the species A. chroococ-
cum, A. beijerinckii, and A. macrocytogenes. Indeed,
even though there is no threshold value of 16S rRNA
homology for species recognition (Stackebrandt and
Goebel, 1994), organisms sharing more than 97%
rRNA similarity are considered as belonging to a
single species (Vandamme et al., 1996).
gical Methods 57 (2004) 197–206 203
4. Discussion
Seventy-six bacterial cultures were taken under
study by means of ARDRA analysis. Of these, 48
constitute a collection of soil strains isolated from
different areas of central Italy (L. Aquilanti, F. Favilli,
and F. Clementi, submitted for publication). The
remaining 28 were reference strains of either species
considered within the family Azotobacteraceae or
species of the most common soil bacterial genera,
among which: (i) Azospirillum and Beijerinckia,
which comprise FNFB commonly living in soil; (ii)
Rhizobium whose members are symbiotic N-fixers but
can occur in soil also as free living forms; (iii) Agro-
bacterium which, surprisingly, constituted more than

L. Aquilanti et al. / Journal of Microbiological Methods 57 (2004) 197–206204
50% of the isolates retrieved on free nitrogen media in
our previous study (L. Aquilanti, F. Favilli, and F.
Clementi, submitted for publication). In addition,
three strains ascribed to the genus Pseudomonas were
included in this study, being easily isolated from soil
samples and strictly related to the genus Azotobacter
for what concerns the 16S rRNA sequence (De
Smedth et al., 1980).
ARDRA is commonly utilized as an alternative to
more laborious and expensive methods for the
identification of eubacteria, being the analysis of
the rRNA cistron a good criterion for microbial
classification at both genus and species level (Gri-
mont and Grimont, 1986; Massol-Deya et al., 1995).
However, as far as we know, this molecular tool has
never been utilized on Azotobacteraceae whose
identification, traditionally based on conventional
biochemical tests, is often tentative. Therefore, the
development of a simple identification method yield-
ing reliable and unambiguous results appears to be
useful, even necessary, for azotobacters assignation
into species.
In the present work, we propose a biphasic ap-
proach to Azotobacteraceae spp. identification. We
shall demonstrate that this approach, consisting of
haplotypes definition and cluster analysis, can be
utilized as an alternative to more complex and time-
consuming methods such as Single Strain Conforma-
tion Polymorphism (SSCP) (Schwieger and Tebbe,
1998) and Denaturing Gradient Gel Electrophoresis
(DGGE) (Lovell et al., 2001; Rosado et al., 1998)
recently applied to FNFB identification.
The first step in developing this method was the
selection of the enzymes to be used for the 16S
rRNA gene restriction analysis. In order to avoid
time-consuming and low-yielding experimental
work, the sequences of Azotobacteraceae, available
in ribosomal databases, were subjected to computer
analysis of digestion sites. Other studies have shown
that at least four enzymes are necessary to resolve
the 16S rRNA gene of different species (Tchan,
1984; Moyer et al., 1996). It is well known that
the combined use of four or more tetrameric restric-
tion enzymes has the effect of increasing the degree
of taxa discrimination rendering the type of TRE
used of no importance. All these evaluations promp-
ted us to select and utilize five enzymes, namely,
AluI, HpaII, FnuDI, RsaI, and HhaI.
On the other hand, it has been reported that
identification at the species level by ARDRA can be
achieved even with three or fewer TREs (Cheneby et
al., 2000; Ventura et al., 2001; Jang et al., 2002),
provided that a careful choice of the TRE is prelim-
inary performed. Our results showed that the use of
RsaI, HpaII, and HhaI on reference strains leads to the
clear discriminate between the different Azotobacter-
aceae species. This is in agreement with the results
obtained by Tiedje et al. (1999) which evidenced that
HhaI, RsaI, and HpaII gave the greatest resolution in
terms of philogenetical dissection of soil natural
communities, and by Moyer et al. (1996) which
described HhaI and RsaI as optimal for the detection
and differentiation of bacterial taxa, on the basis of the
average number of restriction sites.
However, utilization of RsaI, HhaI, and HpaII on
our soil isolates resulted not sufficient for their
unequivocal identification, possibly due to the pres-
ence of one or more restriction sites on all of the
rRNA gene copies present on a single genome, or to
the presence of rRNA gene alleles with different
sequences within a single genome, as described by
Vaneechoutte et al. (1992). In fact, only a limited
number of isolates could be identified on the basis of
the combination of the haplotypes obtained with the
three enzymes. Therefore, we considered advisable
to analyze the population of soil isolates with the
complete set of enzymes in order to identify them on
the basis of cluster analysis and assess whether this
statistic tool could disclose the data obtained by
ARDRA.
The similarity dendrograms, constructed by the
UPGMA method, confirmed the phylogenetic rela-
tionships between the genera Azotobacter, Azomo-
nas, Azospirillum, and Beijerinckia proposed by De
Smedth et al. (1980). Moreover, all Azotobactera-
ceae SSPs were clustered together, thus confirming
the phylogenetic relatedness of species within this
taxon. The closer genetic relatedness of the Pseudo-
monadaceae rRNA cistrons to those of Azotobacter
and Azomonas other than to those of the remaining
reference species was also shown. This is in accor-
dance with the results of De Smedth et al. (1980),
who described these three groups as close branches
of the same phylogenetic tree. The ARDRA-based
dendrogram obtained confirmed also that A. vinelan-
dii and A. paspali were the closest species within

L. Aquilanti et al. / Journal of Microbiological Methods 57 (2004) 197–206 205
genus Azotobacter (De Smedth et al., 1980). Within
genus Azomonas, A. insignis and A. agilis clustered
at a 0.32 genetic distance, forming a well-defined
subcluster separated both from A. macrocytogenes
and the other Azotobacter species. The closer relat-
edness observed for A. macrocytogenes with respect
to the genus Azotobacter, rather than Azomonas
argues in favour of the classification proposed by
Skerman et al. (1980) in contrast to that of New and
Tchan (1982).
Finally, the analysis of ribosomal sequences con-
firmed the identification obtained for 17 isolates by
the combination of ARDRA and cluster analysis.
Thus the biphasic approach proposed proved to be
suitable for the identification of members of the
Azotobacteraceae family at the species level and
represents a contribution to the disclosure and study
of the microbial diversity, also in view of biotech-
nological exploitation of free-living nitrogen-fixing
bacteria.
References
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, Z., Miller, W.,
Lipman, D.J., 1997. Gapped BLAST and PSI-BLAST: a new
generation of protein database search programs. Nucleic Acids
Res. 25, 3389–3402.
Aquilanti, L., Favilli, F., Clementi, F., Assessment of a deliable
strategy for the isolation of Azobacter from soil samples. Soil
Biology and Biochemistry (submitted for publication).
Benson, D.A., Karsch-Mizrachi, I., Lipman, D.J., Ostell, J., Rapp,
B.A., Wheeler, D.L., 2003. GenBank. Nucleic Acids Res. 31,
23–27.
Cheneby, D., Philippot, L., Hartmann, A., Henault, C., Germon,
J.C., 2000. 16S rRNA gene analysis for characterization of
denitrifying bacteria isolated from three agricultural soils.
FEMS Microbiol. Ecol. 34, 121–128.
Clementi, F., 1997. Alginate production by Azotobacter vinelandii.
Crit. Rev. Biotechnol. 4, 327–361.
Denarie, J., Blanchere, H., 1966. Inoculation de graines de vegetaux
cultives a l’aide de souches bacteriennes. Ann. Inst. Pasteur. 3,
57–74.
De Smedth, J., Bauwens, M., Tytgat, R., De Ley, J., 1980. Intra- and
intergeneric similarities of ribosomal ribonucleic acid cistrons of
free-living, nitrogen-fixing bacteria. Int. J. Syst. Bacteriol. 30,
106–122.
Felsenstein, J., 1985. Confidence limits on phylogenies: an ap-
proach using the bootstrap. Evolution 39, 783–791.
Gouri, P.S.V.M., Jagasnnatathan, R., 1995. Biotechnology in organ-
ic farming. Biotechnol. Dev. Rev. 5, 34–47.
Grimont, F., Grimont, P.A.D., 1986. Ribosomal ribonucleic acid
gene restriction patterns as potential taxonomic tools. Ann. Inst.
Pasteur., Microbiol. 137B, 165–175.
Harper, H.T., Lynch, J.M., 1979. Effects of Azotobacter chroococ-
cum on barley seed germination and seedling development.
J. Gen. Microbiol. 112, 45–51.
Higgins, D., Blasby, A., Fuchs, R., Clustal, V., 1992. Improved
software for multiple sequence analysis. Comput. Appl. Biosci.
8, 88–192.
Jackson, R.M., Brown, M.E., Burlingham, S.K., 1964. Similar
effects on tomato plants of Azotobacter inoculation and appli-
cation of gibberellins. Nature 203, 851–852.
Jang, J., Bongjoon, K., Jongho, L., Jeongho, K., Jeong, G., Han, H.,
2002. Identification of Weissella species by the genus-specific
amplified ribosomal DNA restriction analysis. FEMS Microbiol.
Ecol. 212, 29–34.
Lovell, C.R., Friez, M.J., Longshore, J.W., Bagwell, C.E., 2001.
Recovery and phylogenetic analysis of nifH sequences from
diazotrophic bacteria associated with dead aboveground bio-
mass of Spartina alterniflora. Appl. Environ. Microbiol. 67,
5308–5314.
Ludwig, J.A., Reynolds, J.F., 1988. Statistical Ecology Wiley, New
York, NY.
Massol-Deya, A.A., Odelson, D.A.M., Hickey, F., Tiedje, J.M.,
1995. Bacterial community fingerprinting of amplified 16S
and 16–23S ribosomal DNA gene sequences and restriction
endonuclease analysis (ARDRA). Microb. Ecol. 3.3.2, 1–8.
Mishustin, E.N., Shilnikova, V.K., 1971. Biological Fixation of
Atmospheric Nitrogen Macmillan, London, UK.
Moyer, C.L., Tiedje, J.M., Dobbs, F.C., Karl, D.M., 1996. A com-
puter-simulated restriction fragment length polymorphism anal-
ysis of bacterial small-subunit rRNA genes: efficacy of selected
tetrameric restriction enzymes for studies of microbial diversity
in nature. Appl. Environ. Microbiol. 62, 2501–2507.
Mrkovacki, N., Mezei, S., Kovacev, L., 1996. Effect of Azotobacter
inoculation on dry matter mass and nitrogen content in the
hybrid varieties of sugar beet. Period. Sci. Res. Field Veg. Crops
25, 107–113.
Nei, M., Li, W.H., 1979. Mathematical model for studying genetic
variation in terms of restriction endonucleases. Proc. Natl. Acad.
Sci. U. S. A. 76, 5269–5273.
New, P.B., Tchan, Y.T., 1982. Azomonas macrocytogenes (ex Bail-
lie, Hodgkiss, and Norris 1962, 118) nom. rev. Int. J. Syst.
Bacteriol. 32, 381–382.
Page, W.J., Shivprasad, S., 1991. Azotobacter salinestris sp. nov., a
sodium-dependent, microaerophili, and aeroadaptive nitrogen-
fixing bacterium. Int. J. Syst. Bacteriol. 41, 369–376.
Pandey, A., Kumar, S., 1990. Inhibitory effects of Azotobacter
chroococcum and Azospirillum brasiliense on a range of rhizo-
sphere fungi. Indian J. Exp. Biol. 28, 52–54.
Pandey, A., Sharma, E.S., Palni, L.M., 1998. Influence of bacterial
inoculation on maize in upland farming systems of the Sikkim
Himalaya. Soil Biol. Biochem. 30, 379–384.
Rohlf, F.J., 1993. NTSYS. PC. Numerical Taxonomy and Multivar-
iate Analysis System, Version 1.8 Applied Biostatistics, New
York, NY.
Rosado, A.S., Duarte, G.F., Seldin, L., Van Elsas, J.D., 1998. Ge-
netic diversity of nifH gene sequences in Paenibacillus Azoto-

L. Aquilanti et al. / Journal of Microbiological Methods 57 (2004) 197–206206
fixans strains and soil samples analyzed by denaturing gradient.
Appl. Environ. Microbiol. 64, 2770–2779.
Rovira, A.D., 1965. Interactions between plant roots and soil micro-
organisms. Ann. Rev. Microbiol. 19, 241–266.
Sabra, W., Zeng, A.P., Deckwer, W.D., 2001. Bacterial alginate:
physiology, product quality and process aspects. Appl. Micro-
biol. Biotechnol. 56, 315–325.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new
method for reconstructing phylogenetic trees. Mol. Biol. Evol.
4, 406–425.
Schwieger, F., Tebbe, C.C., 1998. A new approach to utilize PCR-
single-strand-conformation polymorphism for 16S rRNA gene-
based microbial community analysis. Appl. Environ. Microbiol.
64, 4870–4876.
Shannon, C.E., Weaver, W., 1949. The Mathematical Theory of
Communication University of Illinois Press, Urbana, IL.
Skerman, V.B.D., McGowan, V., Sneath, P.H.A., 1980. Approved
lists of bacterial names. Int. J. Syst. Bacteriol. 30, 225–420.
Sokal, R.R., Sneath, P.H.A., 1963. Principle of Numerical Taxon-
omy. Freeman, San Francisco, pp. 181–185.
Stackebrandt, E., Goebel, B.M., 1994. Taxonomic note: a place for
DNA–DNA reassociation and 16S rRNA sequence analysis in
the present species definition in bacteriology. Int. J. Syst. Bac-
teriol. 44, 846–849.
Tchan, Y.T., 1984. Family II Azotobacteraceae Pribram 1933, 5AL.
In: Krieg, N.R., Holt, J.G. (Eds.), Bergey’s Manual of System-
atic Bacteriology, vol. 1. Williams & Wilkins, Baltimore, MD,
pp. 219–220.
Tchan, Y.T, New, P.B., 1984a. Genus I Azotobacter, Beijerinck
1901, 567AL. In: Krieg, N.R., Holt, J.G. (Eds.), Bergey’s Man-
ual of Determinative Bacteriology, vol. 1. Williams & Wilkins,
Baltimore, MD, pp. 220–229.
Tchan, Y.T., New, P.B., 1984b. Genus II Azomonas, Winogradsky
1938, 391AL. In: Krieg, N.R., Holt, J.G. (Eds.), Bergey’s Man-
ual of Determinative Bacteriology, vol. 1. Williams & Wilkins,
Baltimore, MD, pp. 230–234.
Tiedje, J.M., Asuming-Brempong, S., Nusslein, K., Marsh, T.L.,
Flynn, S.J., 1999. Opening the black box of soil microbial di-
versity. Appl. Soil Ecol. 13, 109–122.
Vandamme, P., Pot, B., Gillis, M., De Vos, P., Kersters, K., Swings,
J., 1996. Polyphasic taxonomy, a consensus approach to bacte-
rial systematics. Microbiol. Rev. 60, 407–438.
Van Der Peer, Y., De Wachter, R., 1994. TREECON for Windows:
a software package for the construction and drawing of evolu-
tionary trees for the Microsoft Windows environment. Comput.
Appl. Biosci. 10, 570–596.
Vaneechoutte, M., Rossau, R., De Vos, P., 1992. Rapid identifica-
tion of the Comamonadaceae with amplified ribosomal restric-
tion analysis (ARDRA). FEMS Microbiol. Lett. 93, 227–234.
Ventura, M., Elli, M., Reniero, R., Zink, R., 2001. Molecular
microbial analysis of Bifidobacterium isolates from different
environments by the species-specific amplified ribosomal
DNA restriction analysis (ARDRA). FEMS Microbiol. Ecol.
36, 113–121.
Young, J.P.W., Downer, H.L., Eardly, B.D., 1991. Phylogeny of the
phototrophic Rhizobium strain BTAil by polymerase chain reac-
tion-based sequencing of a 16S rRNA gene segment. J. Bacter-
iol. 173, 2271–2277.
Weisburg, W.G., Barns, S.M., Pelletier, D.A., Lane, D.J., 1991. 16S
ribosomal DNA amplification for phylogenetic study. J. Bacter-
iol. 173, 697–703.
Wishart, D.S., Stothard, P., Van Domselaar, G.H., 2000. PepTool
and GeneTool: platform-independent tools for biological se-
quence analysis. Methods Mol. Biol. 132, 93–113.
Zahir, Z.A., Arshad, M., 1996. Effectiveness of Azotobacter inoc-
ulation for improving potato yield under fertilised conditions.
Pak. J. Agric. Sci. 33, 1–5.
Zahir, Z.A., Arshad, M., Hussain, A., Sarfraz, M., 1996. Improving
wheat yield by inoculation with Azotobacter under optimum
fertiliser application. Pak. J. Agric. Sci. 11, 129–131.
Top Related
Copyright © 2022 FDOKUMEN

