Male Choice and Male-male Competition in Idotea baitica (Crustacea, Isopoda
12
Ethology 96, 46-57 (1994) 0 1994 Paul Parey Scientific Publishers, Berlin and Hamburg ISSN 0179-1613 Satakunta Environmental Research Centre, University of Turku, Pori, and Department of Ecology, Theoretical Ecology, Ecology Building, Lund University, Lund Male Choice and Male-male Competition in Idorea balrica (Crustacea, Isopoda) VEIJO JORMALAINEN, SAMI MERILAITA & JUHA TUOMI JORMALAINEN, V., MERILAITA, S. & TUOMI, J. 1993: Male choice and male-male competition in Idoteu bultrca (Crustacea, Isopoda). Ethology 96, 4657. Abstract Sexual selection in mate-guarding Crustacea may involve several processes: male choice, male- male competition, and female choice. T o evaluate the relative importance of the different processes in mate choice of the aquatic isopod I. baltica we studied 1) the mate-choice criteria of males, 2) effects of sex ratio on the outcome of the mating contest, and 3) the role of size in male-male interactions. When given a choice between a small and a large female, males most often chose the one that matured earlier for parturial ecdysis. Maturity was a more important choice criterion than female size, but these also correlated positively. Large males had a mating advantage in both male- and female-biased sex ratios; pairing was size-assortative only in the male-biased ratio where guarding was also longer. If an extra male was placed with a precopulatory pair, 30 % take-overs occurred, large males surpassing. Present and earlier work suggests that male size is an asset in both intra- and intersexual interactions. There is little or no direct phenotypic sexual selection on female size: sexual selection for large males presumably contributes to the evolution of sexual size dimorphism in I. baftzca. Corresponding author: Veijo JORMALAINEN, Department of Biology, Lab. of Ecological Zoology, University of Turku, FIN-20500 Turku. Introduction In aquatic mate-guarding crustaceans (e.g. Isopoda, Gammaroidea), males have, in addition to structural enhancements for fighting (CONLAN 1991), behavioural adaptations improving their success in mating contest. These include high activity needed in mate search (JORMALAINEN & TUOMI 1989), ability to assess female quality (e.g. RIDLEY & THOMPSON 1985; DICK & ELWOOD 1989), and ability to take-over females from other males (e.g. MANNING 1975; WARD 1984). In species in which fertilization is possible only during a short span after female moult, mate-guarding is a plausible male competitive strategy (PARKER 1974). A male’s decision whether to guard a particular female depends on his U.S. Copyright Clearance Center Code Statement: 01 79- 1613/94/9601 -0046$1 o.oo/o
Transcript of Male Choice and Male-male Competition in Idotea baitica (Crustacea, Isopoda

Ethology 96, 46-57 (1994) 0 1994 Paul Parey Scientific Publishers, Berlin and Hamburg ISSN 0179-1613
Satakunta Environmental Research Centre, University of Turku, Pori, and Department of Ecology, Theoretical Ecology,
Ecology Building, Lund University, Lund
Male Choice and Male-male Competition in Idorea balrica (Crustacea, Isopoda)
VEIJO JORMALAINEN, SAMI MERILAITA & JUHA TUOMI JORMALAINEN, V., MERILAITA, S. & TUOMI, J. 1993: Male choice and male-male competition in Idoteu bultrca (Crustacea, Isopoda). Ethology 96, 4 6 5 7 .
Abstract
Sexual selection in mate-guarding Crustacea may involve several processes: male choice, male- male competition, and female choice. T o evaluate the relative importance of the different processes in mate choice of the aquatic isopod I . baltica we studied 1) the mate-choice criteria of males, 2) effects of sex ratio on the outcome of the mating contest, and 3) the role of size in male-male interactions. When given a choice between a small and a large female, males most often chose the one that matured earlier for parturial ecdysis. Maturity was a more important choice criterion than female size, but these also correlated positively. Large males had a mating advantage in both male- and female-biased sex ratios; pairing was size-assortative only in the male-biased ratio where guarding was also longer. If an extra male was placed with a precopulatory pair, 30 % take-overs occurred, large males surpassing. Present and earlier work suggests that male size is an asset in both intra- and intersexual interactions. There is little or no direct phenotypic sexual selection on female size: sexual selection for large males presumably contributes to the evolution of sexual size dimorphism in I . baftzca.
Corresponding author: Veijo JORMALAINEN, Department of Biology, Lab. of Ecological Zoology, University of Turku, FIN-20500 Turku.
Introduction
In aquatic mate-guarding crustaceans (e.g. Isopoda, Gammaroidea), males have, in addition to structural enhancements for fighting (CONLAN 1991), behavioural adaptations improving their success in mating contest. These include high activity needed in mate search (JORMALAINEN & TUOMI 1989), ability to assess female quality (e.g. RIDLEY & THOMPSON 1985; DICK & ELWOOD 1989), and ability to take-over females from other males (e.g. MANNING 1975; WARD 1984).
In species in which fertilization is possible only during a short span after female moult, mate-guarding is a plausible male competitive strategy (PARKER 1974). A male’s decision whether to guard a particular female depends on his
U.S. Copyright Clearance Center Code Statement: 01 79- 16 13/94/9601 -0046$1 o.oo/o

Mate Choice in Idorea baltica 47
expectation of (a) the number of fertilized eggs, and (b) the guarding time, inversely related to female maturity. The number of offspring per guarded female can be maximized by choosing a large female. However, provided that there are receptive females available, the potential number of mates is limited by the guarding time, and can be maximized by choosing females that mature sooner for copulation. Female size (MANNING 1975; SHUSTER 1981; WARD 1984) and matu- rity (BIRKHEAD & CLARKSON 1980; THOMPSON & MANNING 1981; DUNHAM et al. 1986), or both features jointly (MANNING 1980; ELWOOD et al. 1987; DICK & ELWOOD 1990) have been found to function as male mate choice criteria. But critical discrimination between these two variables is onerous, especially as they may be correlated (THOMPSON & MANNING 1981). Nevertheless, male mate choice criteria may be an important aspect of sexual selection of female traits.
In addition to male choice, sexual selection in aquatic crustaceans often involves directly male-male competition, usually favouring large males because of their ability to take over females from smaller males (RIDLEY & THOMPSON 1979; VEUILLE 1980; SHUSTER 1981; WARD 1983, 1988; ELWOOD et al. 1987). Moreover, females intensively resist precopulatory seizure, which may lead to female choice for male size and vigour (RIDLEY & THOMPSON 1979; WARD 1984; ELWOOD et al. 1987; JOKMALAINEN & MERILAITA 1994).
As sex ratio is an important determinant of the intensity of sexual selection (MANNING 1980; SUTHERLAND 1987), it can be manipulated to test some predic- tions about sexual selection and precopula duration. Firstly, the outcome of mating contest in different sex ratios should differ according to the mechanism of sexual selection. If the mating advantage of large males is due to direct male-male competition, the size difference between precopulatory and solitary males should be more pronounced in a male-biased sex ratio where male-male competition is more severe. But if the mating advantage of large males is due to their better ability to overcome female resistance (JORMALAINEN & MERILAITA 1994), they should have an equal advantage in both male- and female-biased sex ratios. Secondly, theoretical analyses predict that the optimal guarding time, from the male’s point of view, varies with the proportion of males (GRAFEN & RIDLEY 1983). Males should guard longer when their probability of finding a female decreases, and the precopulatory phase should be prolonged in the male-biased as compared to the female-biased sex ratio.
In preliminary experiments with an aquatic isopod Idotea bafticu (Pallas), we found that I ) males are choosy when selecting a precopula partner; but the experimental setup could not critically differentiate between the various male choice criteria, and 2) precopulatory males were larger than non-precopulatory males, indicating either large male advantage in male-male competition or female choice for male size WOKMALAINEN et al. 1992). We next studied the male’s mate choice criteria for the first time by manipulating both female maturity and the male’s previous experience of the female population (exp. 1). In exp. 2 we manipulated sex ratio and measured its effect on the outcome of the mating contest and the duration of precopulatory guarding. Further, we studied the role of take-overs and the effect of male size in direct male-male competition (exp. 3).

48 VEIJO JORMALAINEN, SAMI MERILAITA & JUHA TUOMI
Methods
Experiment 1
We first manipulated the maturity stage, i.e. the closeness of parturial ecdysis of females before subjecting them to male choice. For this, females were reared in two temperatures, 10 "C and 15 "C.
Females were sampled on 18-25 Apr. 1991 from the Gulf of Bothnia (61" 21'N, 21" 26'E), W- Finland, well before the actual breeding period (May-Jul.). The manipulation was initiated by gradually increasing the temperature from the natural 3 "C to 10" or 15 "C, and continued for 18 to 25 d during which the females, reared in groups of 20-30, underwent two ecdyses and were in premoult stage before the parturial moult. Water was exchanged and new bladderwrack (Fucus vesiculosus) was offered for food weekly.
To study the effects on his choices of a male's former experience of a female population, males were reared in two kinds of conditions for 23 to 28 d before the choice experiment. One group was reared in two 100-1 aquaria with 120 males in each, the other group in two aquaria with sex ratio of 60 males to 60 females in each.
Choices were staged in 1-1 containers where a male was offered two females differing at least 1 mm in length (cf. Fig. 1 in JORMALAINEN et al. 1992). We arranged three different choice situations where either the smaller or larger female was reared in 15 "C and the other in 10 "C, or both females were reared in either 10" or 15 "C. A male from one or the other male treatment group was assigned randomly to each container.
Containers were monitored daily until both females had undergone parturial ecdysis or one of them had died. Precopulatory guarding, ecdysis and the presence of eggs in marsupiurn after ecdysis were checked. A female was considered the male's first choice when either 1 ) she was the first precopula partner for over two successive monitoring intervals, or 2) she was the second consistent guarding partner after one observation of the precopula with the other female. Further, the female was considered as the first choice if 3) she was the first one to undergo ecdysis and carried eggs afterwards, or 4) if the females underwent ecdysis simultaneously she alone carried eggs. The female was considered as mated (cases 3 and 4) only if she laid eggs into marsupium within 24 h after ecdysis, but not if eggs were laid later, as unfertilized females delay egg-laying for some days (BOROWSKY 1987). About half the cases were deleted from further analysis because the male's first choice could not be judged, or one of the animals died before precopula was observed, or the male did not precopulate and neither female laid eggs shortly after ecdysis.
Expectations about male choice in different choice situations differ depending on the choice criterion of the male. If female size is the primary criterion, males should choose large females in all choice situations (Table 1). But if males use female maturity as a choice criterion, and size and maturity are independent of each other, no size selection is expected in the group in which both females were reared in the same temperature (Table 1) . Because the females reared in 15 "C should be,
Tuble 1: Expected first choice of male allowed to choose between a large and small female in different choice situations in exp. 1. Either size or maturity of the female is assumed as a male mate choice criterion. Expectations are given for situations in which size and maturity are mutually
independent (A) or correlate positively (B)
Male criteria
A B A B Choice situation Size Maturity
Both females reared in same temperature
Large in 15 "C, small in 10 "C
Small in 15 "C, large in 10 "C
large large no size large
large large large large
selection
large large small small

Mate Choice in Idotea baltica 49
on average, more mature we expect choice of large females in the group in which the large female was reared in 15 "C and the small one in 10 "C, and a choice of small females in the group in which the small female was reared in 15 "C. However, assuming that maturity and size correlate positively, choice of large females is expected also in the group in which both females were reared in the same temperature (Table 1). The difference in maturity is most pronounced when the large female is reared in 15 "C (Fig. I), and accordingly, the choice of large females is expected to be most distinct in this group.
I Cold culture Warm culture
Fig. I: Relationship between female size (L) and maturity (M) in different choice situations in exp. 1 assuming that size and ma- turity are positively correlated and that the rearing temperature accelerates maturity develop- ment. 1: large, s : small females, c: cold, w: warm rearing temper- atures. Line segments give expec- tations for maturity differences between large and small females
in different choice situations
M Mcs ws Mwl
cs-CL, ws-WL s- I S I cs-WL S I
CL-ws I S
Experiment 2
Males and females were allowed to form pairs in 12- and 20-1 aquaria with sex ratios of 5 females : 15 males, and 15 females : 5 males. Animals for the three 5 : 15 and three 15 : 5 aquaria were sampled from the Gulf of Bothnia (61" 21'N, 21" 26'E), W-Finland, and reared in Reposaari Island, and for the rest of the aquaria from the Archipelago, SW-Finland (60" 14'N, 21" 60'E), and reared in Seili Island. Thus we tested animals from two different populations. Both sexes differed in size between Seili and Reposaari populations (cf. Tables 3, 4). However, the animals selected randomly in two sex ratio treatments did not differ in size (cf. Tables 3, 4).
When starting the aquaria females were in premoult stage, their next moult being the parturial one. Animals were measured at the beginning, and precopulatory pairs were removed, measured, and replaced once a day with randomly chosen individuals, to keep the sex ratio constant. Dead females, and those that had undergone ecdysis were replaced with premoult females. Tests continued for 5 to 1 1 d in different aquaria. Maturity of precopulatory females was studied by moving the newly formed precopulatory pairs from the aquaria into 1-1 containers, where time to the female's parturial moult was checked.
To analyze the size difference between paired and non-paired animals, we calculated the mean sizes of precopulatory and non-precopulatory isopods for every observation period. These paired measures (pre- versus non-precopulatory) were used in an hierarchical ANOVA model. We had 4 to 5 measures from most aquaria, but in one aquarium we had only one observation for males and in two aquaria two observations for both males and females.
Experiment 3
Precopulatory pairs, formed in different sex ratio aquaria in Seili Island (exp. 2) and in two extra aquaria with 15 : 15 sex ratio, were removed daily and placed in 1-1 containers with stones and some bladderwrack. Each pair was separated for length measurement, and when the identical pair had formed again one extra male was added. The new male was randomly either (at least 2 mm) larger or smaller than the resident.
Ethology, Vol. 96 ( I ) 4

50 VEIJO JORMALAINEN, SAMI MERILAITA & JUHA TUOMI
Precopulatory partners were identified once a day until the female moulted. The criterion for take-over was that the female was seen in precopula with the extra male before the start of her ecdysis. After one take-over, the resident male managed to regain the female. But possibly there were more cases where pairs switched between observations. Moulting occurred within 1 to 5 d after adding the extra male.
16
- 15
E 14
E ,
5 ' 0, 13 c a -I 1 2 -
11
10
Results
Experiment 1 : Manipulation of Female Maturity and Male Experience
The manipulation of the timing of parturial ecdysis was effective. Females reared at 15 "C underwent parturial ecdysis over 6 d earlier than those reared at 10 "C (X = 23.1 d, SD = 5.4, n = 90, and X = 29.4 d, SD = 4.4, n = 46, from the beginning of the manipulation period; t = 6.71, p < 0.001). Size and matu- rity of the females were positively related, since female length correlated nega- tively with the time left to ecdysis. This correlation was significant within the group reared at 15 "C (Fig. 2), but not in that reared at 10 "C (r = -0.028, n = 46, ns), although the latter may partly be an artefact. Observations of the latest moults are sparse in this group, because fewer females survived the longer rearing time needed to observe parturial ecdysis as compared to the group reared in 15 "C.
0 n=90 P=<O.Ol
- 0 . 0 0
- 0 0 b b ( 0 0 0 0 b 0
ob: 8 0 8 - 0 0 0 0 8 # b 0 0 b
0.1: 0s.: 0
0 0 0 0 1 0 0 0 0 8 - 0 . I . : 0
0 0 0 0 0 0
b 0
- 0 0 .
Fig. 2: Correlation between I I I 1 I I female length and time left to partu-
r=-0.323 l7 r
As the males' former experience had no statistically significant effects on their choices, male treatment groups were pooled (Table 2). But in choosing between females reared in the same temperature, males with former experience of females tended to be choosier than males without (Table 2).
Where both females were reared in the same temperature, males selected randomly between large and small females (Table 2). When only the larger females were reared at 15 "C, males chose large females. But males tended to prefer small females when only these were reared at 15 "C (Table 2). We pooled

Mate Choice in ldoteu balticu 51
Table 2: Frequencies of cases in which either the large or small female was the first mating partner of the male in three choice situations and in males with different experience of female population. G-test statistics tests whether males choose randomly, and whether there is heterogeneity between the
choices of males with different experience
Choice Male Selected females situation experience large small G df P
~~~ ~~
Both females reared only males 8 10 0.223 1 ns in same 1 : 1 sex ratio 7 2 2.942 1 0.086 temperature pooled 15 12 0.334 1 ns
heterogeneity 2.831 1 0.092
Large in 15 "C, only males 13 2 9.014 1 <0.01
pooled 27 3 22.461 1 <0.001 small in 10 "C 1 : 1 sex ratio 14 1 13.447 1 <0.001
heterogeneity 0.337 1 ns
Small in 15 "C, only males 3 5 0.505 1 ns large in 10 "C 1 : 1 sex ratio 5 11 2.306 1 ns
pooled 8 16 2.718 1 0.099 heterogeneity 0.093 1 ns
the results from choice situations where the females were reared in different temperatures, to test the null-hypothesis that males choose randomly between females cultured at 15 "C and 10 "C. The test revealed that males significantly preferred females cultured at 15 "C (pooled G = 20.267, df = 1, p < O.OOI) , and hence males chose females near parturial ecdysis. However, there was significant heterogeneity between the choice situations (heterogeneity G = 4.535, df = I , p < 0.05), indicating that the preference for mature females was stronger when larger females were reared at 15 "C. The results fit quite well in the expectations of female maturity as a male mate choice criterion (cf. Table 1, Fig. 1). According to expectations, choice of small females was confirmed when reared in warmer conditions, and the most distinct choice of large females was confirmed when these were reared in warmer conditions. Thus a male's guarding decision was based rather on female maturity than on size.
Once formed, a precopulatory pair did not separate before ecdysis in 27 cases, in which precopulatory guarding time did not differ either between males reared with males (X = 3.0 d, SD = 2.3, n = 16) and 1 : 1 sex ratio (X = 2.3 d, SD = 1.0, n = 11; Kruskal-Wallis test, 2 appr. = 0.31, df = 1, ns), or between choice situations (both females in the same temp.: .? = 3.0 d, SD = 1.5, n = 9; large reared in warm: X = 2.1 d, SD = 1.04, n = 11; small reared in warm: X = 3.3 d, SD = 3.1, n = 7; Kruskal-Wallis test, 2 appr. = 1.87, df = 2, ns). There was no indication of differences in precopula duration between choices of the larger (X = 2.4 d, SD = 1.2, n = 18) or smaller (2 = 3.3 d, SD = 2.9, n = 9) female (Kruskal-Wallis test, 2 appr. = 0.31, df = 1, ns).
52 VEIJO JORMALAINEN, SAMI MERILAITA & JUHA TUOMI
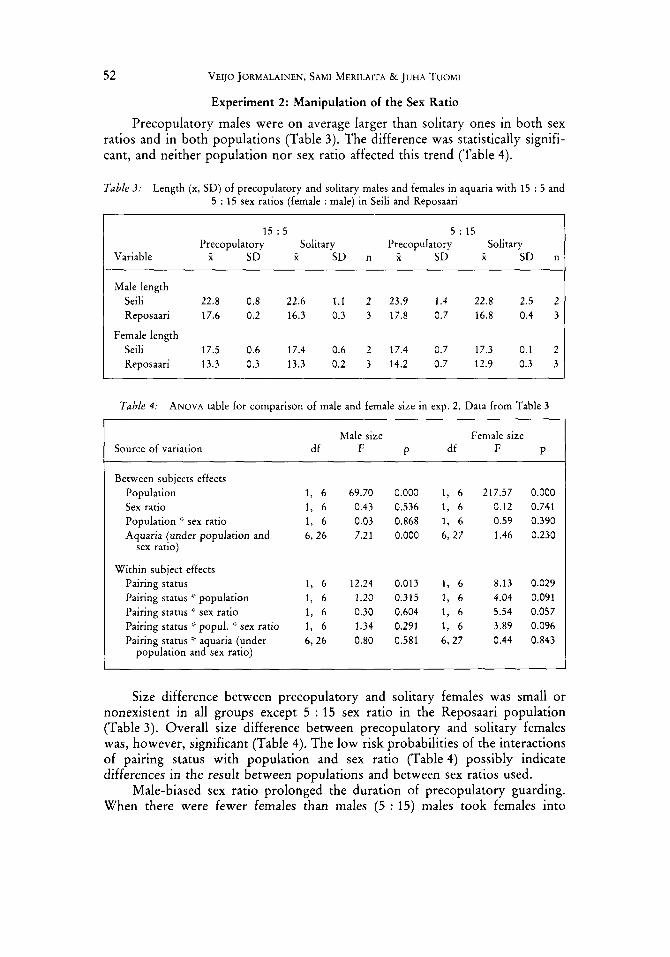
Experiment 2: Manipulation of the Sex Ratio Precopulatory males were on average larger than solitary ones in both sex
ratios and in both populations (Table 3). The difference was statistically signifi- cant, and neither population nor sex ratio affected this trend (Table 4).
Table 3: Length (x, SD) of precopulatory and solitary males and females in aquaria with 15 : 5 and 5 : 15 sex ratios (female : male) in Seili and Reposaari
1 5 : 5 5 : 15 Solitary Precopulatory Solitary
Variable X SD X SD n X SD X SD n Precopulatory
Male length Seili 22.8 0.8 22.6 1.1 2 23.9 1.4 22.8 2.5 2 Reposaari 17.6 0.2 16.3 0.3 3 17.8 0.7 16.8 0.4 3
Female length Seili 17.5 0.6 17.4 0.6 2 17.4 0.7 17.3 0.1 2 Reposaari 13.3 0.3 13.3 0.2 3 14.2 0.7 12.9 0.3 3
Table 4: ANOVA table for comparison of male and female size in exp. 2. Data from Table 3
Source of variation Male size Female size
df F P df F P
Between subjects effects Population Sex ratio Population ::. sex ratio Aquaria (under population and
sex ratio)
Within subject effects Pairing status Pairing status ’: population Pairing status ’:. sex ratio Pairing status :f popul. ’> sex ratio Pairing status ‘f aquaria (under
population and sex ratio)
69.70 0.43 0.03 7.21
12.24 1.20 0.30 1.34 0.80
0.000 1, 6 217.57 0.536 1, 6 0.12 0.868 1, 6 0.59 0.000 6, 27 1.46
0.013 1, 6 8.13 0.315 1, 6 4.04 0.604 1, 6 5.54 0.291 1, 6 3.89 0.581 6, 27 0.44
0.000 0.741 0.390 0.230
0.029 0.091 0.057 0.096 0.843
Size difference between precopulatory and solitary females was small or nonexistent in all groups except 5 : 15 sex ratio in the Reposaari population (Table 3). Overall size difference between precopulatory and solitary females was, however, significant (Table 4). The low risk probabilities of the interactions of pairing status with population and sex ratio (Table 4) possibly indicate differences in the result between populations and between sex ratios used.
Male-biased sex ratio prolonged the duration of precopulatory guarding. When there were fewer females than males (5 : 15) males took females into

Mate Choice in Idotea baltica 53
precopula earlier (X = 2.00 d before ecdysis, SD = 1.22, n = 17) than in female- biased sex ratio (X = 1.23 d before ecdysis, SD = 1.34, n = 22; Kruskal-Wallis; 2 appr. = 7.23, df = 1, p < 0.01).
The correlation between the residuals (aquaria) of male and female size in precopula was significant in 5 : 15 sex ratio (r = 0.48, n = 37, p < 0.01) but not in 15 : 5 sex ratio (r = 0.03, n = 44, p = 0.864). Consequently, pair formation was size assortative only in male-biased sex ratio.
Experiment 3: Male Size and Take-overs
Take-overs happened in 10 cases of 30, most within a day after adding the extra male. Male size affected take-over probability. Take-overs happened in 69 % of the cases in which the resident male was smaller, and in 14 % in which he was larger. Mean length of resident males was 23.1 mm (SD = 2.81, n = 30), of extra males 23.5 mm (SD = 2.44, n = 30).
Extra males involved in take-overs were on average 2.85 mm (SD = 2.24, n = 10) longer than resident males. Resident males retaining females were on average 0.80 mm (SD = 3.54, n = 20) longer than extra males (Fig. 3). The size difference of resident and extra males between take-over and retaining cases was statistically significant (ANOVA; df = 1, 28; F = 8.77; p < 0.01). There was no difference in female length between cases with or without take-overs (Fig. 3).
Fig. 3: Length (i, SD) of females, first pre- copulatory males and added males in cases without take-over (white bars: n = 20) or 0
take-over occurred (black bars; n = 10) Female First male Added male
Discussion
Female Maturity and Size
When given a choice between two females I. bulticu males chose first the female that matured earlier to parturial ecdysis. The males were thus able to assess the interval to female parturial ecdysis. We have no clear evidence that males were able to choose females on the size-basis directly, although we cannot completely exclude such a possibility either. However, female size was of minor importance, as in most cases males preferred more mature females irrespective of size.

54 VEIJO JORMALAINEN, SAMI MERILAITA & JUHA TUOMI
Furthermore, there was no indication of differences in precopula duration between small and large females. Thus results in 1. baltica do not support the idea that males were able to balance the cues of maturity and size in order to optimize guarding duration, as suggested in Aseflus (THOMPSON & MANNING 1981) and Gammarus pulex (ELWOOD et al. 1987).
We propose two working hypotheses that might explain the preference for females close to parturial ecdysis. First, a male can increase his current reproduc- tive success by choosing a large female, but by choosing a female near parturial ecdysis he may increase the number of mates, and hence his future reproductive success (JORMALAINEN et al. 1992). If the number of mates is a more crucial factor than female size in terms of male fitness, selection should favour female maturity as the primary mate-choice criterion of males.
Second, the start of guarding may not be purely a male decision problem. I . baltica females resist precopulatory attempts heavily, and usually succeed in escaping several times before the permanent precopula begins (JORMALAINEN & MERILAITA 1994). In our first experiment the use of the rearing temperature in maturity manipulation most probably also affected other traits that may covary with maturity, including the level of female resistance. If resistance diminishes with the time left to ecdysis (JORMALAINEN & MERILAITA 1994), it is easier for a male to start precopula with a female near parturial ecdysis. Moreover, intense female resistance may constrain males to accept the best they can, instead of striving for an optimum. Especially the ability of small males to affect precopula duration may be limited. Thus female behaviour may have a more prominent role in the mate choice of Z. baltica than generally assumed in aquatic crustaceans.
Male-male Competition
Direct male-male competition occurs when males try to take over a guarded female. Take-overs were frequent, and occurred in most cases where the resident was smaller than the intruder male. Female size had no effect on take-over probability. Take-over may benefit successful males, especially as a female already guarded is probably relatively near her parturial moult and no longer resists when faced by a new male (JORMALAINEN & MERILAITA 1994). Assuming that single and guarded females are equally available, and that guarded females do not resist, males profit from take-overs (other things being equal) if (I) there is a lower cost of subduing a guarding male than a single female, (2) the mean value of guarded females exceeds the mean value of single females, or (3) the odds for a take-over are higher than for encountering and achieving precopula with a single female. Thus take-over may be a profitable strategy, especially for large males with a high probability of success. Furthermore, due to the male-biased sex ratio during the most active breeding season (JORMALAINEN & TUOMI 1989), take-over may often be the only option to get a mate.
If the larger size of precopulatory males (exp. 2) were a result of direct male- male competition alone, we would expect more intense competition and more pronounced size differences between solitary and precopulatory males in male- biased as compared to female-biased sex ratio. But the mating advantage of large

Mate Choice in Idotea baltica 55
males was evident in both sex ratios, presumably indicating that intersexual interactions augmented the better mating success of large males. For example, when a male and a female are reared in pairs large males are able to perform longer precopulas than small males, apparently because of their better ability to over- come female resistance (JORMALAINEN & MERILAITA 1994). Our result that pre- copulatory females were larger than solitary ones may be due to the correlation between maturity and size. If males choose mates primarily on the basis of maturity, then on the average they choose larger females.
Pairing was size-assortative only in male-biased sex ratio. A similar ten- dency, though not statistically significant, was also found in an earlier experiment (JORMALAINEN et a]. 1992). The results emphasize the importance of male-male competition in the emergence of assortative pairing (for other factors see e.g. ELWOOD et al. 1987; WARD & PORTER 1993). Size assortative pairing in I . baltica may well be a combined outcome of direct male-male competition and female resistance, both favouring large male size, and male choice for mature, and so on average larger females.
Guarding Duration
Precopulatory guarding was prolonged in male-biased sex ratio as compared to female-biased sex ratio, a similar result as in Asellus (MANNING 1980). This variation in precopula duration can be interpreted either by sex-ratio dependent changes in (1) male decision to start guarding or (2) female resistance. The precondition for ( I ) is that males acquire information about the available female population and adjust their guarding duration accordingly (BIRKHEAD & CLARK- SON 1980; DUNHAM & HURSHMAN 1990). For this there is experimental evidence in Gammavus (WARD 1983; DUNHAM & HURSHMAN 1990). In I . baltica this adjustment requires that males are able to intensify their precopulatory attempts to overcome female resistance. As such, the result fits in with the predictions of mate guarding models (PARKER 1974; GRAFEN & RIDLEY 1983): larger investment in guarding is expected as the encounter rate diminishes. The other possibility (2) is that female resistance decreases faster with male-biased sex ratio, so that longer precopula is allowed. For example, females may be prepared to resist only until a certain threshold, determined by costs and benefits of resistance, that is exceeded quicker in male-biased sex ratio where precopulatory attempts are obviously more numerous.
In conclusion, phenotypic selection on large male size in I . baltica operates through both intra- and intersexual interactions. On the other hand, there is only weak or no direct sexual selection on female size through male choice, as females are taken in precopula according to maturity. A phenotypic correlation between female size and maturity was found. Presuming that this also represents genetic correlation, indirect selection on female size is expected (ARNOLD 1987). Further- more, sexual selection is likely to be stronger among males than among females, as the operational sex ratio is highly male biased because of the mating system. Sexual selection is, therefore likely to contribute to the evolution of sexual size dimorphism in I . baltica.

56 VEIJO JORMALAINEN, SAMI MERILAITA & JUHA TUOMI
Acknowledgements
We are grateful to Archipelago Research Institute of the University of Turku for providing research facilities, and to personnel of the Satakunta Environmental Research Centre for their help during experiments. V. JORMALAINEN enjoyed a grant from the KONE Foundation during the course of the study.
Literature Cited
ARNOLD, S. J. 1987: Genetic correlation and the evolution of physiology. In: New Directions in Ecological Physiology. (FEDER, M. E., B E N N E ~ , A. F., BURGGREN, W. W. & HUEY, R. B., eds.) Cambridge Univ. Press, New York, pp. 189-215.
BIRKHEAD, T. R. & CLARKSON, K. 1980: Mate selection and precopulatory guarding in Gammarus pulex. 2. Tierpsychol. 52, 365-380.
BOROWSKY, B. 1987: Laboratory studies of the pattern of reproduction of the isopod Crustacean Idotea baltica. Fish. Bull. 85, 377-380.
CONLAN, K. E. 1991: Precopulatory mating behavior and sexual dimorphism in the amphipod Crustacea. Hydrobiol. 223, 255-282.
DICK, J. T. A. & ELWOOD, R. W. 1989: Assessments and decisions during mate choice in Gummarus pulex (Amphipoda). Behaviour 109, 235-246.
-- & - - 1990: Symmetrical assessment of female quality by male Gammarus pulex (Amphipoda) during struggles over precopula females. Anim. Behav. 40, 877-883.
DUNHAM, P., ALEXANDER, T. & HURSHMAN, A. 1986: Precopulatory mate guarding in an amphipod, Gammarus lawrencianus Bousfield. Anim. Behav. 34, 1680-1686.
_ - & HURSHMAN, A. 1990: Precopulatory mate guarding in the amphipod, Gammarus luwren- cianus: effects of social stimulation during the post-copulation interval. Anim. Behav. 39, 9 7 6 9 7 9 .
ELWOOD, R. W. & DICK, J. T. A. 1990: The amorous Gammarus: the relationship between precopula duration and size-assortative mating in G. pulex. Anim. Behav. 39, 828-833.
_ - , GIBSON, J. & NEIL, S. 1987: The amorous Gammarus: size assortative mating in G . pulex. Anim. Behav. 35, 1-6.
GRAFEN, A. & RIDLEY, M. 1983: A model of mate guarding. J. Theor. Biol. 102, 549-567. JORMALAINEN, V. & MERILAITA, S. 1994: Female resistance and precopulatory guarding in isopod
Idotea baltica (Pallas). Behaviour, in press. - - & TUOMI, J. 1989: Sexual differences in habitat selection and activity of the color polymorphic
isopod Idotea baltica. Anim. Behav. 38, 5 7 6 5 8 5 . ---- , & MERILAITA, S. 1992: Mate choice for male and female size in aquatic isopod Idotea
baltica. Ann. 2001. Fennici 29, 161-167. MANNING, J. T. 1975: Male discrimination and investment in Asellus aquaticus (L.) and A. meridianus
Racovitsza (Crustacea: Isopoda). Behaviour 55, 1-14. -- 1980: Sex ratio and optimal male time investment strategies in Asellus aquaticus (L.) and
A. meridianus Racovitza. Behaviour 74, 264-273. PARKER, G. A. 1974: Courtship persistence and female guarding as male time investment strategies.
Behaviour 48, 157-184. RIDLEY, M. & THOMPSON, D. 1979: Size and mating in Asellus aquaticus (Crustacea: Isopoda). 2.
Tierpsychol. 51, 380-397. -- & - - 1985: Sexual selection of population dynamics in aquatic Crustacea. In: Behavioural
Ecology. (SIBLY, R. M. & SMITH, R. H., eds.) Blackwell, Oxford, pp. 409-422. SHUSTER, S. M. 1981 : Sexual selection in the socorro isopod Thermosphaeroma thermophilum (Cole)
(Crustacea: Peracarida). Anim. Behav. 29, 698-707. SUTHERLAND, W. J. 1987: Random and deterministic components of variance in mating success. In:
Sexual Selection: Testing the Alternatives. (BRADBURY, J. W. & ANDERSSON, M. B., eds.) John Wiley & Sons, Avon, p. 307.
THOMPSON, D. J. & MANNING, J. T. 1981: Mate selection by Asellus (Crustacea: Isopoda). Behaviour 78, 178-187.

Mate Choice in Idotea bulticu 57
VEUILLE, M. 1980: Sexual behaviour and evolution of sexual dimorphism in body size in laera (Isopoda Asellota). Biol. J. Linn. SOC. 13, 89-100.
WARD, P. I. 1983: Advantages and a disadvantage of large size for male Gammaruspulex (Crustacea: Amphipoda). Behav. Ecol. Sociobiol. 14, 69-76.
-- 1984: The effects of size on the mating decisions of Gammarus pulex (Crustacea, Amphipoda). Z . Tierpsychol. 64, 174-184.
- _ 1988: Sexual selection, natural selection, and body size in Gammarus p u l a (Amphipoda). Am. Nat. 131, 348-359.
_ _ & PORTER, A. H. 1993: The relative roles of habitat structure and male-male competition in the mating system of Gammarus pulex (Crustacea; Amphipoda): a simulation study. Anim. Behav. 45, 119-133.
Received: August 12, 1993
Accepted: October 4, 1993 (W. Wickler)