







Bahasa
Halaman
Hukum


i
DIFERENSIASI GENETIK MELIACEAE
PADA REGION Second Internal Transcribed Spacer (ITS2)
dan Maturase-K (mat-K)
MIRA NOVIANTI
DEPARTEMEN SILVIKULTUR
FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR
2012
ii
DIFERENSIASI GENETIK MELIACEAE
PADA REGION Second Internal Transcribed Spacer (ITS2)
dan Maturase-K (mat-K)
MIRA NOVIANTI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Kehutanan pada
Departemen Silvikultur
DEPARTEMEN SILVIKULTUR
FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR
2012
iii
RINGKASAN
MIRA NOVIANTI Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K) Di bawah bimbingan
ISKANDAR Z SIREGAR
Anggota famili Meliaceae seperti mahoni daun besar (Swietenia
macrophylla) mahoni daun kecil (Swietenia mahagoni) mindi (Melia
azedarach) mimba (Azadirachta indica) dan khaya (Khaya anthotheca) termasuk
ke dalam jenis penting baik secara ekonomi maupun ekologi Pada umumnya
anggota Meliaceae dimanfaatkan untuk penghasil kayu buah atau kandungan
bahan kimianya sehingga permintaan terhadap jenis ini sangat tinggi yang
menyebabkan penurunan populasi alami Untuk itu diperlukan upaya pelestarian
jenis yang sebaiknya berbasis scientific yang salah satunya adalah melalui
pemanfaatan penanda genetik (DNA)
Penanda DNA dapat digunakan untuk melihat adanya variasi antar
(diferensiasi) dan di dalam jenis dengan akurat melalui deteksi perubahan urutan
basa nukleotida Penelitian ini bertujuan untuk mengetahui variasi genetik dan
jarak kekerabatan dari lima jenis Meliaceae sehingga pembagian taksonominya
menjadi lebih jelas DNA sekuen untuk kedua wilayah diekstraksi dengan metode
CTAB diikuti dengan proses PCR dan sekuen (httpbase-asiacom) Pengamatan
ada tidaknya pola pita polimorfik dilakukan dengan enzim restriksi secara in
silico Analisis data dilakukan dengan bantuan perangkat lunak BioEdit
ClustalW2 EMBL-EBI TreeViewX PopGene NTSYs dan pDRAW32 Hasil
perhitungan jarak genetik pada sekuens DNA pada wilayah ITS2 menunjukkan
bahwa jenis yang memiliki jarak genetik yang paling jauh adalah antara S
mahagoni dengan S macrophylla dan jenis dengan jarak genetik terdekat adalah
antara A indica dengan M azedarach Hasil ini sesuai dengan analisis yang
dilakukan pada wilayah gen mat-K
Kata kunci DNA enzim restriksi ITS2 mat-K Meliaceae
iv
ABSTRACT
MIRA NOVIANTI Genetic Differentiation of Meliaceae based on Second
Internal Transribed Spacer (ITS2) dan Maturase-K (Mat-K) Regions Supervised
by ISKANDAR Z SIREGAR
Members of Meliaceae (Swietenia macrophylla Swietenia mahogany
Melia azedarach Azadirachta indica dan Khaya anthotheca) play significant
roles both economically and ecologically The species are used for timber and
non-timber purposes like fruits chemicals and herbal products and demands are
therefore high In their natural stands the populations are depleting and
conservation of the species is urgently called To formulate the sound stratgy for
species conservation further information at molecular levels ie DNA sequences
are required DNA markers can be used to assess intraspecific genetic
differentiation based on particular gene regions or DNA fragments The aim of
this study was to determine genetic differentiation of five important species of
Meliaceae following current taxonomy classification
Two DNA sequences ie ITS2 and mat-K were used in the analysis ITS2
sequences were revealed following DNA extraction (CTAB) PCR amplification
and sequencing (httpbase-asiacom) while mat-K sequences were obtained from
the genbank database (httpncbinlmnihgovgenbank) Differentiation analysis
was performed using following softwares namely BioEdit ClustalW2 EMBL-
EBI TreeViewX PopGene NTSYs and pDRAW32 Results showed that the
aligned sequences based on ITS2 and mat-K regions were also in agreement with
previous findings showing that K anthoteca S mahogany and S macrophylla
were grouped in one cluster while the another consisted of A indica and M
azedarach Average nucleotide diversities at ITS2 and mat-K were 027398 and
018441 respectively
Keywords DNA ITS2 mat-K Meliaceae restriction enzymes
v
PERNYATAAN
Dengan ini saya menyatakan bahwa skripsi yang berjudul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (Mat-K)rdquo adalah benar-benar hasil karya sendiri dengan bimbingan
dosen pembimbing yang belum pernah digunakan sebagai karya pada perguruan
tinggi atau lembaga manapun Sumber informasi yang berasal atau dikutip dari
karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan
dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini
Bogor November 2012
Mira Novianti
E44080023
vi
LEMBAR PENGESAHAN
Judul Skripsi Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K)
Nama Mira Novianti
NIM E44080023
Menyetujui
Dosen Pembimbing
Prof Dr Ir Iskandar Z Siregar MForSc
NIP 19660320 199002 1 001
Mengetahui
Ketua Departemen Silvikultur
Fakultas Kehutanan IPB
Prof Dr Ir Nurheni Wijayanto MS
NIP 19601024 198403 1 009
Tanggal Lulus
vii
KATA PENGANTAR
Alhamdulillah segala puji dan syukur bagi Allah SWT yang telah
memberikan rahmat dan hidayahNya sehingga penulis dapat menyelesaikan
skripsi ini sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
Institut Pertanian Bogor Dalam penelitian yang dilaksanakan sejak bulan
September 2011 sampai dengan Juli 2012 penulis memilih judul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (mat-K)rdquo
Keragaman genetik memiliki peranan sangat penting dalam program
pemuliaan pohon Pengetahuan keragaman genetik ditunjang melalui pengetahuan
biologi molekuler Selanjutnya pengetahuan ini dapat digunakan untuk program
konservasi dan pemanfaatannya
Penulis menyadari bahwa karya tulis ini masih jauh dari kesempurnaan
oleh karena itu saran dan kritik sangat penulis butuhkan Penulis berharap semoga
hasil penelitian ini dapat memberikan manfaat bagi penulis dan semua pihak yang
membutuhkan
Bogor November 2012
Penulis
viii
UCAPAN TERIMA KASIH
Puji dan syukur bagi Allah SWT karena berkat rahmat dan hidayahNya
penulis dapat menyelesaikan skripsi ini Dalam kesempatan ini penulis
mengucapkan terima kasih kepada
1 Prof Dr Ir Iskandar Z Siregar MForSc atas bimbingan dan ilmu yang telah
diberikan
2 Resti Meilani Shut MSi selaku dosen penguji dan Dr Ir Iwan Hilwan MS
selaku ketua sidang yang telah memberikan sarannya untuk penulisan skripsi
ini
3 Ibunda Juswaidar Ayahanda Hasymi Kamaruddin saudaraku Ulfa Umami
dan keluarga besar yang telah mendoakan menyemangati dan membantu
penulis
4 Senior-senior dan teman satu bimbingan di Laboratorium Analisis Genetika
Bagian Silvikultur Fakultas Kehutanan IPB (Laswi Irmayanti Shut Azizah
Shut Elviana Shut Fifi Gus Dwiyanti Shut dan Argha AC Nugraha)
5 Adinda beserta keluarga yang selalu memberikan dukungan dan bantuan bagi
penulis
6 Essy Harnelly PhD Kokom Komariah SE MM Arida Susilowati Shut
MSi Rima HS Siburian Shut MSi Dr Utut Widyastuti Dr Ir T M Oemijati
R MS yang telah memberikan saran dan motivasi bagi penulis
7 Teman-teman penulis (Marrsquoahthul Ishlah Felix Julian Aji P Alm Syahrul
Isnaini Osmond Vito Eliazar SP Intan Fajar Kemala Evi Rumindah
Sinaga)
8 Rekan-rekan mahasiswa Mayor Silvikultur Angkatan 45
Penulis berharap karya ini dapat bermanfaat dan berguna bagi semua pihak
dan mohon maaf atas segala kekurangan
Bogor November 2012
Penulis
ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

ii
DIFERENSIASI GENETIK MELIACEAE
PADA REGION Second Internal Transcribed Spacer (ITS2)
dan Maturase-K (mat-K)
MIRA NOVIANTI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Kehutanan pada
Departemen Silvikultur
DEPARTEMEN SILVIKULTUR
FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR
2012
iii
RINGKASAN
MIRA NOVIANTI Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K) Di bawah bimbingan
ISKANDAR Z SIREGAR
Anggota famili Meliaceae seperti mahoni daun besar (Swietenia
macrophylla) mahoni daun kecil (Swietenia mahagoni) mindi (Melia
azedarach) mimba (Azadirachta indica) dan khaya (Khaya anthotheca) termasuk
ke dalam jenis penting baik secara ekonomi maupun ekologi Pada umumnya
anggota Meliaceae dimanfaatkan untuk penghasil kayu buah atau kandungan
bahan kimianya sehingga permintaan terhadap jenis ini sangat tinggi yang
menyebabkan penurunan populasi alami Untuk itu diperlukan upaya pelestarian
jenis yang sebaiknya berbasis scientific yang salah satunya adalah melalui
pemanfaatan penanda genetik (DNA)
Penanda DNA dapat digunakan untuk melihat adanya variasi antar
(diferensiasi) dan di dalam jenis dengan akurat melalui deteksi perubahan urutan
basa nukleotida Penelitian ini bertujuan untuk mengetahui variasi genetik dan
jarak kekerabatan dari lima jenis Meliaceae sehingga pembagian taksonominya
menjadi lebih jelas DNA sekuen untuk kedua wilayah diekstraksi dengan metode
CTAB diikuti dengan proses PCR dan sekuen (httpbase-asiacom) Pengamatan
ada tidaknya pola pita polimorfik dilakukan dengan enzim restriksi secara in
silico Analisis data dilakukan dengan bantuan perangkat lunak BioEdit
ClustalW2 EMBL-EBI TreeViewX PopGene NTSYs dan pDRAW32 Hasil
perhitungan jarak genetik pada sekuens DNA pada wilayah ITS2 menunjukkan
bahwa jenis yang memiliki jarak genetik yang paling jauh adalah antara S
mahagoni dengan S macrophylla dan jenis dengan jarak genetik terdekat adalah
antara A indica dengan M azedarach Hasil ini sesuai dengan analisis yang
dilakukan pada wilayah gen mat-K
Kata kunci DNA enzim restriksi ITS2 mat-K Meliaceae
iv
ABSTRACT
MIRA NOVIANTI Genetic Differentiation of Meliaceae based on Second
Internal Transribed Spacer (ITS2) dan Maturase-K (Mat-K) Regions Supervised
by ISKANDAR Z SIREGAR
Members of Meliaceae (Swietenia macrophylla Swietenia mahogany
Melia azedarach Azadirachta indica dan Khaya anthotheca) play significant
roles both economically and ecologically The species are used for timber and
non-timber purposes like fruits chemicals and herbal products and demands are
therefore high In their natural stands the populations are depleting and
conservation of the species is urgently called To formulate the sound stratgy for
species conservation further information at molecular levels ie DNA sequences
are required DNA markers can be used to assess intraspecific genetic
differentiation based on particular gene regions or DNA fragments The aim of
this study was to determine genetic differentiation of five important species of
Meliaceae following current taxonomy classification
Two DNA sequences ie ITS2 and mat-K were used in the analysis ITS2
sequences were revealed following DNA extraction (CTAB) PCR amplification
and sequencing (httpbase-asiacom) while mat-K sequences were obtained from
the genbank database (httpncbinlmnihgovgenbank) Differentiation analysis
was performed using following softwares namely BioEdit ClustalW2 EMBL-
EBI TreeViewX PopGene NTSYs and pDRAW32 Results showed that the
aligned sequences based on ITS2 and mat-K regions were also in agreement with
previous findings showing that K anthoteca S mahogany and S macrophylla
were grouped in one cluster while the another consisted of A indica and M
azedarach Average nucleotide diversities at ITS2 and mat-K were 027398 and
018441 respectively
Keywords DNA ITS2 mat-K Meliaceae restriction enzymes
v
PERNYATAAN
Dengan ini saya menyatakan bahwa skripsi yang berjudul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (Mat-K)rdquo adalah benar-benar hasil karya sendiri dengan bimbingan
dosen pembimbing yang belum pernah digunakan sebagai karya pada perguruan
tinggi atau lembaga manapun Sumber informasi yang berasal atau dikutip dari
karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan
dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini
Bogor November 2012
Mira Novianti
E44080023
vi
LEMBAR PENGESAHAN
Judul Skripsi Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K)
Nama Mira Novianti
NIM E44080023
Menyetujui
Dosen Pembimbing
Prof Dr Ir Iskandar Z Siregar MForSc
NIP 19660320 199002 1 001
Mengetahui
Ketua Departemen Silvikultur
Fakultas Kehutanan IPB
Prof Dr Ir Nurheni Wijayanto MS
NIP 19601024 198403 1 009
Tanggal Lulus
vii
KATA PENGANTAR
Alhamdulillah segala puji dan syukur bagi Allah SWT yang telah
memberikan rahmat dan hidayahNya sehingga penulis dapat menyelesaikan
skripsi ini sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
Institut Pertanian Bogor Dalam penelitian yang dilaksanakan sejak bulan
September 2011 sampai dengan Juli 2012 penulis memilih judul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (mat-K)rdquo
Keragaman genetik memiliki peranan sangat penting dalam program
pemuliaan pohon Pengetahuan keragaman genetik ditunjang melalui pengetahuan
biologi molekuler Selanjutnya pengetahuan ini dapat digunakan untuk program
konservasi dan pemanfaatannya
Penulis menyadari bahwa karya tulis ini masih jauh dari kesempurnaan
oleh karena itu saran dan kritik sangat penulis butuhkan Penulis berharap semoga
hasil penelitian ini dapat memberikan manfaat bagi penulis dan semua pihak yang
membutuhkan
Bogor November 2012
Penulis
viii
UCAPAN TERIMA KASIH
Puji dan syukur bagi Allah SWT karena berkat rahmat dan hidayahNya
penulis dapat menyelesaikan skripsi ini Dalam kesempatan ini penulis
mengucapkan terima kasih kepada
1 Prof Dr Ir Iskandar Z Siregar MForSc atas bimbingan dan ilmu yang telah
diberikan
2 Resti Meilani Shut MSi selaku dosen penguji dan Dr Ir Iwan Hilwan MS
selaku ketua sidang yang telah memberikan sarannya untuk penulisan skripsi
ini
3 Ibunda Juswaidar Ayahanda Hasymi Kamaruddin saudaraku Ulfa Umami
dan keluarga besar yang telah mendoakan menyemangati dan membantu
penulis
4 Senior-senior dan teman satu bimbingan di Laboratorium Analisis Genetika
Bagian Silvikultur Fakultas Kehutanan IPB (Laswi Irmayanti Shut Azizah
Shut Elviana Shut Fifi Gus Dwiyanti Shut dan Argha AC Nugraha)
5 Adinda beserta keluarga yang selalu memberikan dukungan dan bantuan bagi
penulis
6 Essy Harnelly PhD Kokom Komariah SE MM Arida Susilowati Shut
MSi Rima HS Siburian Shut MSi Dr Utut Widyastuti Dr Ir T M Oemijati
R MS yang telah memberikan saran dan motivasi bagi penulis
7 Teman-teman penulis (Marrsquoahthul Ishlah Felix Julian Aji P Alm Syahrul
Isnaini Osmond Vito Eliazar SP Intan Fajar Kemala Evi Rumindah
Sinaga)
8 Rekan-rekan mahasiswa Mayor Silvikultur Angkatan 45
Penulis berharap karya ini dapat bermanfaat dan berguna bagi semua pihak
dan mohon maaf atas segala kekurangan
Bogor November 2012
Penulis
ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

iii
RINGKASAN
MIRA NOVIANTI Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K) Di bawah bimbingan
ISKANDAR Z SIREGAR
Anggota famili Meliaceae seperti mahoni daun besar (Swietenia
macrophylla) mahoni daun kecil (Swietenia mahagoni) mindi (Melia
azedarach) mimba (Azadirachta indica) dan khaya (Khaya anthotheca) termasuk
ke dalam jenis penting baik secara ekonomi maupun ekologi Pada umumnya
anggota Meliaceae dimanfaatkan untuk penghasil kayu buah atau kandungan
bahan kimianya sehingga permintaan terhadap jenis ini sangat tinggi yang
menyebabkan penurunan populasi alami Untuk itu diperlukan upaya pelestarian
jenis yang sebaiknya berbasis scientific yang salah satunya adalah melalui
pemanfaatan penanda genetik (DNA)
Penanda DNA dapat digunakan untuk melihat adanya variasi antar
(diferensiasi) dan di dalam jenis dengan akurat melalui deteksi perubahan urutan
basa nukleotida Penelitian ini bertujuan untuk mengetahui variasi genetik dan
jarak kekerabatan dari lima jenis Meliaceae sehingga pembagian taksonominya
menjadi lebih jelas DNA sekuen untuk kedua wilayah diekstraksi dengan metode
CTAB diikuti dengan proses PCR dan sekuen (httpbase-asiacom) Pengamatan
ada tidaknya pola pita polimorfik dilakukan dengan enzim restriksi secara in
silico Analisis data dilakukan dengan bantuan perangkat lunak BioEdit
ClustalW2 EMBL-EBI TreeViewX PopGene NTSYs dan pDRAW32 Hasil
perhitungan jarak genetik pada sekuens DNA pada wilayah ITS2 menunjukkan
bahwa jenis yang memiliki jarak genetik yang paling jauh adalah antara S
mahagoni dengan S macrophylla dan jenis dengan jarak genetik terdekat adalah
antara A indica dengan M azedarach Hasil ini sesuai dengan analisis yang
dilakukan pada wilayah gen mat-K
Kata kunci DNA enzim restriksi ITS2 mat-K Meliaceae
iv
ABSTRACT
MIRA NOVIANTI Genetic Differentiation of Meliaceae based on Second
Internal Transribed Spacer (ITS2) dan Maturase-K (Mat-K) Regions Supervised
by ISKANDAR Z SIREGAR
Members of Meliaceae (Swietenia macrophylla Swietenia mahogany
Melia azedarach Azadirachta indica dan Khaya anthotheca) play significant
roles both economically and ecologically The species are used for timber and
non-timber purposes like fruits chemicals and herbal products and demands are
therefore high In their natural stands the populations are depleting and
conservation of the species is urgently called To formulate the sound stratgy for
species conservation further information at molecular levels ie DNA sequences
are required DNA markers can be used to assess intraspecific genetic
differentiation based on particular gene regions or DNA fragments The aim of
this study was to determine genetic differentiation of five important species of
Meliaceae following current taxonomy classification
Two DNA sequences ie ITS2 and mat-K were used in the analysis ITS2
sequences were revealed following DNA extraction (CTAB) PCR amplification
and sequencing (httpbase-asiacom) while mat-K sequences were obtained from
the genbank database (httpncbinlmnihgovgenbank) Differentiation analysis
was performed using following softwares namely BioEdit ClustalW2 EMBL-
EBI TreeViewX PopGene NTSYs and pDRAW32 Results showed that the
aligned sequences based on ITS2 and mat-K regions were also in agreement with
previous findings showing that K anthoteca S mahogany and S macrophylla
were grouped in one cluster while the another consisted of A indica and M
azedarach Average nucleotide diversities at ITS2 and mat-K were 027398 and
018441 respectively
Keywords DNA ITS2 mat-K Meliaceae restriction enzymes
v
PERNYATAAN
Dengan ini saya menyatakan bahwa skripsi yang berjudul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (Mat-K)rdquo adalah benar-benar hasil karya sendiri dengan bimbingan
dosen pembimbing yang belum pernah digunakan sebagai karya pada perguruan
tinggi atau lembaga manapun Sumber informasi yang berasal atau dikutip dari
karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan
dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini
Bogor November 2012
Mira Novianti
E44080023
vi
LEMBAR PENGESAHAN
Judul Skripsi Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K)
Nama Mira Novianti
NIM E44080023
Menyetujui
Dosen Pembimbing
Prof Dr Ir Iskandar Z Siregar MForSc
NIP 19660320 199002 1 001
Mengetahui
Ketua Departemen Silvikultur
Fakultas Kehutanan IPB
Prof Dr Ir Nurheni Wijayanto MS
NIP 19601024 198403 1 009
Tanggal Lulus
vii
KATA PENGANTAR
Alhamdulillah segala puji dan syukur bagi Allah SWT yang telah
memberikan rahmat dan hidayahNya sehingga penulis dapat menyelesaikan
skripsi ini sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
Institut Pertanian Bogor Dalam penelitian yang dilaksanakan sejak bulan
September 2011 sampai dengan Juli 2012 penulis memilih judul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (mat-K)rdquo
Keragaman genetik memiliki peranan sangat penting dalam program
pemuliaan pohon Pengetahuan keragaman genetik ditunjang melalui pengetahuan
biologi molekuler Selanjutnya pengetahuan ini dapat digunakan untuk program
konservasi dan pemanfaatannya
Penulis menyadari bahwa karya tulis ini masih jauh dari kesempurnaan
oleh karena itu saran dan kritik sangat penulis butuhkan Penulis berharap semoga
hasil penelitian ini dapat memberikan manfaat bagi penulis dan semua pihak yang
membutuhkan
Bogor November 2012
Penulis
viii
UCAPAN TERIMA KASIH
Puji dan syukur bagi Allah SWT karena berkat rahmat dan hidayahNya
penulis dapat menyelesaikan skripsi ini Dalam kesempatan ini penulis
mengucapkan terima kasih kepada
1 Prof Dr Ir Iskandar Z Siregar MForSc atas bimbingan dan ilmu yang telah
diberikan
2 Resti Meilani Shut MSi selaku dosen penguji dan Dr Ir Iwan Hilwan MS
selaku ketua sidang yang telah memberikan sarannya untuk penulisan skripsi
ini
3 Ibunda Juswaidar Ayahanda Hasymi Kamaruddin saudaraku Ulfa Umami
dan keluarga besar yang telah mendoakan menyemangati dan membantu
penulis
4 Senior-senior dan teman satu bimbingan di Laboratorium Analisis Genetika
Bagian Silvikultur Fakultas Kehutanan IPB (Laswi Irmayanti Shut Azizah
Shut Elviana Shut Fifi Gus Dwiyanti Shut dan Argha AC Nugraha)
5 Adinda beserta keluarga yang selalu memberikan dukungan dan bantuan bagi
penulis
6 Essy Harnelly PhD Kokom Komariah SE MM Arida Susilowati Shut
MSi Rima HS Siburian Shut MSi Dr Utut Widyastuti Dr Ir T M Oemijati
R MS yang telah memberikan saran dan motivasi bagi penulis
7 Teman-teman penulis (Marrsquoahthul Ishlah Felix Julian Aji P Alm Syahrul
Isnaini Osmond Vito Eliazar SP Intan Fajar Kemala Evi Rumindah
Sinaga)
8 Rekan-rekan mahasiswa Mayor Silvikultur Angkatan 45
Penulis berharap karya ini dapat bermanfaat dan berguna bagi semua pihak
dan mohon maaf atas segala kekurangan
Bogor November 2012
Penulis
ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

iv
ABSTRACT
MIRA NOVIANTI Genetic Differentiation of Meliaceae based on Second
Internal Transribed Spacer (ITS2) dan Maturase-K (Mat-K) Regions Supervised
by ISKANDAR Z SIREGAR
Members of Meliaceae (Swietenia macrophylla Swietenia mahogany
Melia azedarach Azadirachta indica dan Khaya anthotheca) play significant
roles both economically and ecologically The species are used for timber and
non-timber purposes like fruits chemicals and herbal products and demands are
therefore high In their natural stands the populations are depleting and
conservation of the species is urgently called To formulate the sound stratgy for
species conservation further information at molecular levels ie DNA sequences
are required DNA markers can be used to assess intraspecific genetic
differentiation based on particular gene regions or DNA fragments The aim of
this study was to determine genetic differentiation of five important species of
Meliaceae following current taxonomy classification
Two DNA sequences ie ITS2 and mat-K were used in the analysis ITS2
sequences were revealed following DNA extraction (CTAB) PCR amplification
and sequencing (httpbase-asiacom) while mat-K sequences were obtained from
the genbank database (httpncbinlmnihgovgenbank) Differentiation analysis
was performed using following softwares namely BioEdit ClustalW2 EMBL-
EBI TreeViewX PopGene NTSYs and pDRAW32 Results showed that the
aligned sequences based on ITS2 and mat-K regions were also in agreement with
previous findings showing that K anthoteca S mahogany and S macrophylla
were grouped in one cluster while the another consisted of A indica and M
azedarach Average nucleotide diversities at ITS2 and mat-K were 027398 and
018441 respectively
Keywords DNA ITS2 mat-K Meliaceae restriction enzymes
v
PERNYATAAN
Dengan ini saya menyatakan bahwa skripsi yang berjudul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (Mat-K)rdquo adalah benar-benar hasil karya sendiri dengan bimbingan
dosen pembimbing yang belum pernah digunakan sebagai karya pada perguruan
tinggi atau lembaga manapun Sumber informasi yang berasal atau dikutip dari
karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan
dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini
Bogor November 2012
Mira Novianti
E44080023
vi
LEMBAR PENGESAHAN
Judul Skripsi Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K)
Nama Mira Novianti
NIM E44080023
Menyetujui
Dosen Pembimbing
Prof Dr Ir Iskandar Z Siregar MForSc
NIP 19660320 199002 1 001
Mengetahui
Ketua Departemen Silvikultur
Fakultas Kehutanan IPB
Prof Dr Ir Nurheni Wijayanto MS
NIP 19601024 198403 1 009
Tanggal Lulus
vii
KATA PENGANTAR
Alhamdulillah segala puji dan syukur bagi Allah SWT yang telah
memberikan rahmat dan hidayahNya sehingga penulis dapat menyelesaikan
skripsi ini sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
Institut Pertanian Bogor Dalam penelitian yang dilaksanakan sejak bulan
September 2011 sampai dengan Juli 2012 penulis memilih judul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (mat-K)rdquo
Keragaman genetik memiliki peranan sangat penting dalam program
pemuliaan pohon Pengetahuan keragaman genetik ditunjang melalui pengetahuan
biologi molekuler Selanjutnya pengetahuan ini dapat digunakan untuk program
konservasi dan pemanfaatannya
Penulis menyadari bahwa karya tulis ini masih jauh dari kesempurnaan
oleh karena itu saran dan kritik sangat penulis butuhkan Penulis berharap semoga
hasil penelitian ini dapat memberikan manfaat bagi penulis dan semua pihak yang
membutuhkan
Bogor November 2012
Penulis
viii
UCAPAN TERIMA KASIH
Puji dan syukur bagi Allah SWT karena berkat rahmat dan hidayahNya
penulis dapat menyelesaikan skripsi ini Dalam kesempatan ini penulis
mengucapkan terima kasih kepada
1 Prof Dr Ir Iskandar Z Siregar MForSc atas bimbingan dan ilmu yang telah
diberikan
2 Resti Meilani Shut MSi selaku dosen penguji dan Dr Ir Iwan Hilwan MS
selaku ketua sidang yang telah memberikan sarannya untuk penulisan skripsi
ini
3 Ibunda Juswaidar Ayahanda Hasymi Kamaruddin saudaraku Ulfa Umami
dan keluarga besar yang telah mendoakan menyemangati dan membantu
penulis
4 Senior-senior dan teman satu bimbingan di Laboratorium Analisis Genetika
Bagian Silvikultur Fakultas Kehutanan IPB (Laswi Irmayanti Shut Azizah
Shut Elviana Shut Fifi Gus Dwiyanti Shut dan Argha AC Nugraha)
5 Adinda beserta keluarga yang selalu memberikan dukungan dan bantuan bagi
penulis
6 Essy Harnelly PhD Kokom Komariah SE MM Arida Susilowati Shut
MSi Rima HS Siburian Shut MSi Dr Utut Widyastuti Dr Ir T M Oemijati
R MS yang telah memberikan saran dan motivasi bagi penulis
7 Teman-teman penulis (Marrsquoahthul Ishlah Felix Julian Aji P Alm Syahrul
Isnaini Osmond Vito Eliazar SP Intan Fajar Kemala Evi Rumindah
Sinaga)
8 Rekan-rekan mahasiswa Mayor Silvikultur Angkatan 45
Penulis berharap karya ini dapat bermanfaat dan berguna bagi semua pihak
dan mohon maaf atas segala kekurangan
Bogor November 2012
Penulis
ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

v
PERNYATAAN
Dengan ini saya menyatakan bahwa skripsi yang berjudul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (Mat-K)rdquo adalah benar-benar hasil karya sendiri dengan bimbingan
dosen pembimbing yang belum pernah digunakan sebagai karya pada perguruan
tinggi atau lembaga manapun Sumber informasi yang berasal atau dikutip dari
karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan
dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini
Bogor November 2012
Mira Novianti
E44080023
vi
LEMBAR PENGESAHAN
Judul Skripsi Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K)
Nama Mira Novianti
NIM E44080023
Menyetujui
Dosen Pembimbing
Prof Dr Ir Iskandar Z Siregar MForSc
NIP 19660320 199002 1 001
Mengetahui
Ketua Departemen Silvikultur
Fakultas Kehutanan IPB
Prof Dr Ir Nurheni Wijayanto MS
NIP 19601024 198403 1 009
Tanggal Lulus
vii
KATA PENGANTAR
Alhamdulillah segala puji dan syukur bagi Allah SWT yang telah
memberikan rahmat dan hidayahNya sehingga penulis dapat menyelesaikan
skripsi ini sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
Institut Pertanian Bogor Dalam penelitian yang dilaksanakan sejak bulan
September 2011 sampai dengan Juli 2012 penulis memilih judul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (mat-K)rdquo
Keragaman genetik memiliki peranan sangat penting dalam program
pemuliaan pohon Pengetahuan keragaman genetik ditunjang melalui pengetahuan
biologi molekuler Selanjutnya pengetahuan ini dapat digunakan untuk program
konservasi dan pemanfaatannya
Penulis menyadari bahwa karya tulis ini masih jauh dari kesempurnaan
oleh karena itu saran dan kritik sangat penulis butuhkan Penulis berharap semoga
hasil penelitian ini dapat memberikan manfaat bagi penulis dan semua pihak yang
membutuhkan
Bogor November 2012
Penulis
viii
UCAPAN TERIMA KASIH
Puji dan syukur bagi Allah SWT karena berkat rahmat dan hidayahNya
penulis dapat menyelesaikan skripsi ini Dalam kesempatan ini penulis
mengucapkan terima kasih kepada
1 Prof Dr Ir Iskandar Z Siregar MForSc atas bimbingan dan ilmu yang telah
diberikan
2 Resti Meilani Shut MSi selaku dosen penguji dan Dr Ir Iwan Hilwan MS
selaku ketua sidang yang telah memberikan sarannya untuk penulisan skripsi
ini
3 Ibunda Juswaidar Ayahanda Hasymi Kamaruddin saudaraku Ulfa Umami
dan keluarga besar yang telah mendoakan menyemangati dan membantu
penulis
4 Senior-senior dan teman satu bimbingan di Laboratorium Analisis Genetika
Bagian Silvikultur Fakultas Kehutanan IPB (Laswi Irmayanti Shut Azizah
Shut Elviana Shut Fifi Gus Dwiyanti Shut dan Argha AC Nugraha)
5 Adinda beserta keluarga yang selalu memberikan dukungan dan bantuan bagi
penulis
6 Essy Harnelly PhD Kokom Komariah SE MM Arida Susilowati Shut
MSi Rima HS Siburian Shut MSi Dr Utut Widyastuti Dr Ir T M Oemijati
R MS yang telah memberikan saran dan motivasi bagi penulis
7 Teman-teman penulis (Marrsquoahthul Ishlah Felix Julian Aji P Alm Syahrul
Isnaini Osmond Vito Eliazar SP Intan Fajar Kemala Evi Rumindah
Sinaga)
8 Rekan-rekan mahasiswa Mayor Silvikultur Angkatan 45
Penulis berharap karya ini dapat bermanfaat dan berguna bagi semua pihak
dan mohon maaf atas segala kekurangan
Bogor November 2012
Penulis
ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

vi
LEMBAR PENGESAHAN
Judul Skripsi Diferensiasi Genetik Meliaceae pada Region Second Internal
Transcribed Spacer (ITS2) dan Maturase-K (mat-K)
Nama Mira Novianti
NIM E44080023
Menyetujui
Dosen Pembimbing
Prof Dr Ir Iskandar Z Siregar MForSc
NIP 19660320 199002 1 001
Mengetahui
Ketua Departemen Silvikultur
Fakultas Kehutanan IPB
Prof Dr Ir Nurheni Wijayanto MS
NIP 19601024 198403 1 009
Tanggal Lulus
vii
KATA PENGANTAR
Alhamdulillah segala puji dan syukur bagi Allah SWT yang telah
memberikan rahmat dan hidayahNya sehingga penulis dapat menyelesaikan
skripsi ini sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
Institut Pertanian Bogor Dalam penelitian yang dilaksanakan sejak bulan
September 2011 sampai dengan Juli 2012 penulis memilih judul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (mat-K)rdquo
Keragaman genetik memiliki peranan sangat penting dalam program
pemuliaan pohon Pengetahuan keragaman genetik ditunjang melalui pengetahuan
biologi molekuler Selanjutnya pengetahuan ini dapat digunakan untuk program
konservasi dan pemanfaatannya
Penulis menyadari bahwa karya tulis ini masih jauh dari kesempurnaan
oleh karena itu saran dan kritik sangat penulis butuhkan Penulis berharap semoga
hasil penelitian ini dapat memberikan manfaat bagi penulis dan semua pihak yang
membutuhkan
Bogor November 2012
Penulis
viii
UCAPAN TERIMA KASIH
Puji dan syukur bagi Allah SWT karena berkat rahmat dan hidayahNya
penulis dapat menyelesaikan skripsi ini Dalam kesempatan ini penulis
mengucapkan terima kasih kepada
1 Prof Dr Ir Iskandar Z Siregar MForSc atas bimbingan dan ilmu yang telah
diberikan
2 Resti Meilani Shut MSi selaku dosen penguji dan Dr Ir Iwan Hilwan MS
selaku ketua sidang yang telah memberikan sarannya untuk penulisan skripsi
ini
3 Ibunda Juswaidar Ayahanda Hasymi Kamaruddin saudaraku Ulfa Umami
dan keluarga besar yang telah mendoakan menyemangati dan membantu
penulis
4 Senior-senior dan teman satu bimbingan di Laboratorium Analisis Genetika
Bagian Silvikultur Fakultas Kehutanan IPB (Laswi Irmayanti Shut Azizah
Shut Elviana Shut Fifi Gus Dwiyanti Shut dan Argha AC Nugraha)
5 Adinda beserta keluarga yang selalu memberikan dukungan dan bantuan bagi
penulis
6 Essy Harnelly PhD Kokom Komariah SE MM Arida Susilowati Shut
MSi Rima HS Siburian Shut MSi Dr Utut Widyastuti Dr Ir T M Oemijati
R MS yang telah memberikan saran dan motivasi bagi penulis
7 Teman-teman penulis (Marrsquoahthul Ishlah Felix Julian Aji P Alm Syahrul
Isnaini Osmond Vito Eliazar SP Intan Fajar Kemala Evi Rumindah
Sinaga)
8 Rekan-rekan mahasiswa Mayor Silvikultur Angkatan 45
Penulis berharap karya ini dapat bermanfaat dan berguna bagi semua pihak
dan mohon maaf atas segala kekurangan
Bogor November 2012
Penulis
ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

vii
KATA PENGANTAR
Alhamdulillah segala puji dan syukur bagi Allah SWT yang telah
memberikan rahmat dan hidayahNya sehingga penulis dapat menyelesaikan
skripsi ini sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan
Institut Pertanian Bogor Dalam penelitian yang dilaksanakan sejak bulan
September 2011 sampai dengan Juli 2012 penulis memilih judul ldquoDiferensiasi
Genetik Meliaceae pada Region Second Internal Transcribed Spacer (ITS2) dan
Maturase-K (mat-K)rdquo
Keragaman genetik memiliki peranan sangat penting dalam program
pemuliaan pohon Pengetahuan keragaman genetik ditunjang melalui pengetahuan
biologi molekuler Selanjutnya pengetahuan ini dapat digunakan untuk program
konservasi dan pemanfaatannya
Penulis menyadari bahwa karya tulis ini masih jauh dari kesempurnaan
oleh karena itu saran dan kritik sangat penulis butuhkan Penulis berharap semoga
hasil penelitian ini dapat memberikan manfaat bagi penulis dan semua pihak yang
membutuhkan
Bogor November 2012
Penulis
viii
UCAPAN TERIMA KASIH
Puji dan syukur bagi Allah SWT karena berkat rahmat dan hidayahNya
penulis dapat menyelesaikan skripsi ini Dalam kesempatan ini penulis
mengucapkan terima kasih kepada
1 Prof Dr Ir Iskandar Z Siregar MForSc atas bimbingan dan ilmu yang telah
diberikan
2 Resti Meilani Shut MSi selaku dosen penguji dan Dr Ir Iwan Hilwan MS
selaku ketua sidang yang telah memberikan sarannya untuk penulisan skripsi
ini
3 Ibunda Juswaidar Ayahanda Hasymi Kamaruddin saudaraku Ulfa Umami
dan keluarga besar yang telah mendoakan menyemangati dan membantu
penulis
4 Senior-senior dan teman satu bimbingan di Laboratorium Analisis Genetika
Bagian Silvikultur Fakultas Kehutanan IPB (Laswi Irmayanti Shut Azizah
Shut Elviana Shut Fifi Gus Dwiyanti Shut dan Argha AC Nugraha)
5 Adinda beserta keluarga yang selalu memberikan dukungan dan bantuan bagi
penulis
6 Essy Harnelly PhD Kokom Komariah SE MM Arida Susilowati Shut
MSi Rima HS Siburian Shut MSi Dr Utut Widyastuti Dr Ir T M Oemijati
R MS yang telah memberikan saran dan motivasi bagi penulis
7 Teman-teman penulis (Marrsquoahthul Ishlah Felix Julian Aji P Alm Syahrul
Isnaini Osmond Vito Eliazar SP Intan Fajar Kemala Evi Rumindah
Sinaga)
8 Rekan-rekan mahasiswa Mayor Silvikultur Angkatan 45
Penulis berharap karya ini dapat bermanfaat dan berguna bagi semua pihak
dan mohon maaf atas segala kekurangan
Bogor November 2012
Penulis
ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

viii
UCAPAN TERIMA KASIH
Puji dan syukur bagi Allah SWT karena berkat rahmat dan hidayahNya
penulis dapat menyelesaikan skripsi ini Dalam kesempatan ini penulis
mengucapkan terima kasih kepada
1 Prof Dr Ir Iskandar Z Siregar MForSc atas bimbingan dan ilmu yang telah
diberikan
2 Resti Meilani Shut MSi selaku dosen penguji dan Dr Ir Iwan Hilwan MS
selaku ketua sidang yang telah memberikan sarannya untuk penulisan skripsi
ini
3 Ibunda Juswaidar Ayahanda Hasymi Kamaruddin saudaraku Ulfa Umami
dan keluarga besar yang telah mendoakan menyemangati dan membantu
penulis
4 Senior-senior dan teman satu bimbingan di Laboratorium Analisis Genetika
Bagian Silvikultur Fakultas Kehutanan IPB (Laswi Irmayanti Shut Azizah
Shut Elviana Shut Fifi Gus Dwiyanti Shut dan Argha AC Nugraha)
5 Adinda beserta keluarga yang selalu memberikan dukungan dan bantuan bagi
penulis
6 Essy Harnelly PhD Kokom Komariah SE MM Arida Susilowati Shut
MSi Rima HS Siburian Shut MSi Dr Utut Widyastuti Dr Ir T M Oemijati
R MS yang telah memberikan saran dan motivasi bagi penulis
7 Teman-teman penulis (Marrsquoahthul Ishlah Felix Julian Aji P Alm Syahrul
Isnaini Osmond Vito Eliazar SP Intan Fajar Kemala Evi Rumindah
Sinaga)
8 Rekan-rekan mahasiswa Mayor Silvikultur Angkatan 45
Penulis berharap karya ini dapat bermanfaat dan berguna bagi semua pihak
dan mohon maaf atas segala kekurangan
Bogor November 2012
Penulis
ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

ix
RIWAYAT HIDUP
Penulis lahir di Batusangkar Sumatera Barat pada tanggal 22 November
1989 sebagai putri dari Hasymi Kamaruddin dan Juswaidar Penulis merupakan
anak pertama dari dua bersaudara
Tahun 2008 penulis lulus dari SMAN 1 Sungayang dan pada tahun yang
sama lulus seleksi masuk IPB jalur Undangan Seleksi Masuk IPB (USMI)
Penulis diterima sebagai mahasiswa di Departemen Silvikultur Fakultas
Kehutanan
Selama perkuliahan penulis mengikuti Praktek Penganalan Ekosistem
Hutan (P2EH) yang dilaksanakan di Sancang Timur Papandayan Praktek
Pengolahan Hutan (P2H) yang dilaksanakan di Hutan Pendidikan Gunung Walat
(HPGW) Sukabumi Praktek Kerja Profesi (PKP) dilaksanakan di pertambangan
nikel PT Vale Indonesia Tbk Sorowako Sulawesi Selatan Untuk memperoleh
gelar Sarjana Kehutanan IPB penulis meyelesaikan skripsi dengan judul
Diferensiasi Genetik Meliaceae pada Region Second Internal Transcribed Spacer
(ITS2) dan Maturase-K (mat-K)
x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

x
DAFTAR ISI
Halaman
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiv
I PENDAHULUAN
11 Latar Belakang 1
12 Tujuan 2
13 Manfaat Penelitian 2
II TINJAUAN PUSTAKA
21 Deskripsi Spesies 3
22 Keragaman Genetik Hutan 4
23 PCR (Polymerase Chain Reaction) 4
24 Sequencing DNA 6
25 Second Internal Transcribed Spacer (ITS2) 6
26 Maturase K (mat-K) 7
III BAHAN DAN METODE
31 Waktu dan Tempat Penelitian 9
32 Bahan dan Alat Penelitian 9
321 Koleksi Contoh Uji Daun 9
322 Analisis Genetik 9
33 Prosedur Penelitian 10
331 Analisis Sekuen ITS2 10
332 Analisis Fragmen ITS2 secara In Silico 13
333 Analisis Fragmen mat-K 13
IV HASIL DAN PEMBAHASAN
41 Analisis DNA 15
411 Ekstraksi DNA 15
412 Analisis Hasil PCR 16
42 Analisis Sequence DNA 17
421 Analisis Runutan Nukleotida 17
422 Keragaman Antar Spesies 19
xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

xi
V KESIMPULAN DAN SARAN
51 Kesimpulan 25
52 Saran 25
DAFTAR PUSTAKA 26
LAMPIRAN 28
xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

xii
DAFTAR TABEL
Halaman
1 Alat dan bahan untuk teknik analisis genetik 9
2 Primer untuk amplifikasi DNA 12
3 Komposisi PCR 12
4 Tahapan proses PCR 12
5 Spesies pada wilayah mat-K 13
6 Rataan komposisi nukleotida 17
7 Spesies enzim yang hanya memotong pada satu jenis 19
8 Rata-rata substitusi nukleotida per sites antar spesies pada ITS2 23
9 Klaster jenis berdasarkan ITS2 dan mat-K 24
xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

xiii
DAFTAR GAMBAR
Halaman
1 Ilustrasi PCR 5
2 Gen tiga genom pada tanaman sebagai kandidat untuk barcoding 6
3 Diagram ITS 7
4 Wilayah mat-K 8
5 Alur penelitian 11
6 Pola band DNA 15
7 Hasil amplifikasi ITS2 16
8 Runutan nukleotida ITS2 18
9 Dendrogram in silico ITS2 20
10 Dendrogram in silico mat-K 20
11 Dendrogram ITS2 21
12 Dendrogram ITS 21
13 Dendrogram mat-K 22
14 Dendrogram mat-K (Meullner 2003) 22
xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

xiv
DAFTAR LAMPIRAN
Halaman
1 Electropherogram enzim restriksi secara in silico pada ITS2 29
2 Electropherogram enzim restriksi secara in silico pada mat-K 31
3 Runutan sekuen mat-K 33
4 Polimorfik ITS2 35
5 Polimorfik mat-K 37
BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

BAB I
PENDAHULUAN
11 Latar Belakang
Meliaceae merupakan suku yang secara umum terdiri dari pohon pada
ordo Sapindales Spesies yang termasuk anggota Meliaceae di antaranya mahoni
daun lebar (Swietenia macrophylla K) mahoni daun kecil (Swietenia mahagoni
(L) Jacq) mindi (Melia azedarach L) mimba (Azadirachta indica Juss) dan
khaya (Khaya anthotheca (Welw) C DC) Spesies pada famili ini termasuk ke
dalam spesies yang sangat penting di dunia baik dalam aspek ekonomi maupun
aspek ekologi Pada umumnya anggota Meliaceae dimanfaatkan untuk penghasil
kayu buah atau kandungan bahan kimianya Beberapa spesies penghasil kayu
yang bernilai ekonomi adalah mahoni (S macrophylla) mimba (A indica) dan
mindi (M azedarach) yang mengandung zat-zat yang bisa dijadikan sebagai
bahan pestisida Tetapi dalam perkembangannya sekarang ini persediaan tegakan
alami Meliaceae sudah mulai menurun drastis (Muellner et al 2011) Dengan
demikian diperlukan upaya pelestarian dan pemuliaan anggota Meliaceae seiring
dengan peningkatan permintaan terhadap spesies ini Penggunaan teknologi
molekuler dalam bidang kehutanan di Indonesia pada saat ini umumnya masih
diarahkan untuk konservasi genetik dan pemuliaan pohon dari spesies-spesies
yang dianggap penting Saat ini arahan bagi penyusunan strategi konservasi
genetik pohon hutan dapat dilakukan dengan lebih baik melalui pemanfaatan
informasi genetik dari suatu spesies untuk melengkapi data morfologi yang ada
(buah daun batang dll) Oleh karena itu penelitian ini dilakukan yang diarahkan
untuk mendapatkan informasi lebih lengkap pada tingkat molekuler yang
selanjutnya juga dapat digunakan untuk melengkapi informasi taksonomi spesies
Setiap individu memiliki sifat atau karakter yang berbeda Variasi genetik
yang dimiliki oleh pohon-pohon hutan termasuk pada anggota famili Meliaceae
tidak selalu sama Ketidaksamaan genetik pada spesies ini kemudian memberikan
sifat-sifat yang berbeda pada setiap pohonnya Perbedaan sifat yang dibawa oleh
genetik dapat dilihat dari perbedaan susunan nukleotida yang terdapat pada DNA
suatu individu Untuk mengetahui perbedaan runutan nukleotida ini dapat dilihat
melalui teknik analisis sekuen Analisis sekuen merupakan suatu teknik yang
2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

2
dianggap paling baik untuk melihat keanekaragaman hayati suatu kelompok
organisme Teknik ini berkembang setelah diciptakannya mesin DNA sequencer
Pada prinsipnya polimorfisme dilihat dari urutan atau sekuen DNA dari fragmen
tertentu dari suatu genom organisme Kekerabatan dan keragaman genetik dari
spesies yang digunakan dapat dilihat melalui hasil sekuen yang diperoleh
12 Tujuan
Tujuan dari penelitian ini adalah untuk menduga variasi dan diferensiasi
genetik antar spesies (interspecific variation) lima spesies pohon anggota
Meliaceae yaitu S macrophylla S mahagoni M azedarach A indica dan K
anthotheca
13 Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar mengenai
kekerabatan lima spesies anggota famili Meliaceae Informasi kekerabatan yang
diperoleh tersebut dapat digunakan sebagai referensi dalam penanganan
silvikultur spesies untuk kepentingan konservasi sumberdaya genetik dan
pemuliaan spesies
3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

3
BAB II
TINJAUAN PUSTAKA
21 Deskripsi Spesies
Azadirachta indica memiliki nama lokal mimba atau nimbi Tanaman
mimba dapat beradaptasi di daerah tropis Di Indonesia tanaman mimba dapat
tumbuh dengan baik di daerah dataran rendah dengan ketinggian sekitar 800 m
dpl Di Indonesia tanaman mimba banyak terdapat di Jawa Timur Jawa Tengah
Jawa Barat Bali dan Nusa Tenggara Barat (NTB) Pada umumnya tanaman
mimba ditanam sebagai tanaman peneduh jalan (Rukmana dan Oesman 2002)
Melia azedarach memiliki nama lokal mindi atau mindi berbuah kecil
Tumbuh pada daerah dataran rendah hingga dataran tinggi pada ketinggian
sampai dengan 1200 m dpl dapat tumbuh pada suhu minimum Himalaya pada
ketinggian 1800minus2200 m dpl dapat tumbuh pada suhu minimum -5oC suhu
maksimum 39oC dengan curah hujan rata-rata per tahun 600minus2000 mm (Ahmed
dan Idris 1997 dalam Dephut 2001) Pohon mindi memiliki persebaran alami di
India dan Burma kemudian banyak ditanam di daerah tropis dan subtropis
termasuk Indonesia (Mabberley 1984 dalam Dephut 2001) Untuk Indonesia
sudah banyak ditanam di daerah Sumatera Jawa Nusa Tenggara dan Irian Jaya
(Whitmore dan Tantra 1986)
Swietenia mahagoni memiliki nama lokal mahoni daun kecil Spesies ini
secara alami dijumpai pada iklim dengan curah hujan tahunan 580minus800 mm
Hasil pertanamannya lebih rendah dibandingkan dengan S macrophylla tetapi
pada tapak yang kering tumbuh sangat baik dan kualitas kayunyapun lebih baik
Secara komersial spesies ini tidak berarti apabila tersedia dalam skala kecil Akan
berpotensi bila ditanam dalam skala besar khususnya di daerah kering terutama
untuk memperoleh kayu berkualitas tinggi Spesies ini juga digunakan pada
agroforestry untuk meningkatkan kualitas tanah dan tanaman hias (Dephut 2001)
Swietenia macrophylla memiliki nama lokal mahoni daun lebar Tanaman
ini tumbuh pada ketinggian 0-1500 m dpl suhu tahunan 11minus36oC dan curah
hujan tahunan 1524minus5085 mm (BPT 1986) Tanaman ini mempunyai peranan
yang cukup penting secara ekonomi karena kayunya dapat digunakan untuk kayu
bangunan dan perkakas Saat ini tanaman mahoni merupakan salah satu tanaman
4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

4
prioritas untuk pembangunan Hutan Tanaman Industri dan reboisasi hutan
produksi
Khaya anthotheca memiliki nama lokal khaya Penyebaran alami di daerah
Afrika tropis di daerah tersebut spesies ini merupakan spesies kayu perdagangan
utama dengan nama perdagangan internasional mahoni Afrika dan di Indonesia
mempunyai nama perdagangan mahoni Uganda (Burhaman 2004) Kayu spesies
ini mudah dikerjakan mudah dikupas tanpa direbus terlebih dahulu serta
perekatannya baik dan secara umum memenuhi persyaratan Kayunya dapat
dipergunakan untuk keperluan bahan baku kayu lapis bahan baku pembalutan
mebel dan perkakas rumah tangga lainnya (Burhaman 2004)
22 Keragaman Genetik Tanaman Hutan
Keragaman genetik adalah suatu tingkatan biodiversitas yang merujuk
pada jumlah total karakteristik genetik dalam genetika keseluruhan spesies
Keragaman genetik suatu spesies tanaman dapat dievaluasi pada dua tingkatan
yaitu keragaman dalam spesies (intra-species) dan keragaman antar spesies (inter-
species) (Finkeldey 2005)
Secara umum ada dua sebab utama yang menyebabkan keragaman yaitu
perbedaan lingkungan (enviromental variation) dan perbedaan susunan genetik
yang diturunkan dari tetua kepada keturunannya (genetic variation) Adanya
keragaman dalam suatu spesies perlu diketahui terlebih dahulu sebelum memulai
pemuliaan pohon karena adanya keragaman genetik merupakan prasyarat mutlak
dalam pemuliaan yaitu memungkinkan seleksi dan untuk mencegah
dihasilkannya tanaman yang tidak bermutu (Soerianegara dan Djamhuri 1979)
23 PCR (polymerase chain reactions)
Tahapan dalam proses PCR meliputi denaturasi pada suhu tinggi
penempelan DNA pada cetakan (tahap annealing) serta pemanjangan primer
dengan melakukan reaksi polimerisasi nukleotida untuk membentuk rantai DNA
baru (tahap extention) (Saiki et al 1998 dalam Mahfira 2010) Tahapan dari PCR
adalah sebagai berikut
(1) Tahap peleburan atau denaturasi
Denaturasi rantai DNA berlansung pada suhu 94oC atau pada suhu 95
oC (Saiki et
al 1998) dengan selang waktu antara 15 detik sampai 2 menit Dalam proses
5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

5
denaturasi dua rantai DNA akan terpisah dan masing-masing rantai DNA akan
digunakan sebagai cetakan pada proses PCR DNA yang memiliki struktur
kompleks dapat didenaturasi pada suhu 100oC selama beberapa menit namun
kemampuan aktivitas enzim Taq DNA polymerase menjadi turun
(2) Tahap penempelan atau annealing
Penempelan primer pada DNA cetakan disebut annealing Besarnya suhu
annealing tergantung pada panjang dan jumlah basa G dan C dalam primer serta
konsentrasi garam dalam buffer
(3) Tahap pemanjangan atau extention
Reaksi polimerisasi nukleotida oleh enzim Taq DNA polymerase (extention)
dimulai dari ujung 5rsquoα-fosfat dan berakhir pada ujung 3rsquo gugus hidoksil (H) Suhu
extention yang digunakan berkisar antara 70-74oC karena pada selang suhu
tersebut enzim Taq DNA polymerase bekerja optimum Lamanya tahap extention
1minus2 menit Waktu extention yang terlalu lama akan menghasilkan produk
amplifikasi yang tidak spesifik (Saiki et al 1998) Secara terperinci tahapan PCR
disajikan pada Gambar 1
Gambar 1 Ilustrasi siklus PCR (1) denaturasi (2) annealing (3) extention (4) siklus ke-1
selesai (Rice 2009)
24 Sequencing DNA
Sequencing DNA adalah proses penentuan urutan dari basa A T G dan C
dalam sepotong DNA Pada intinya DNA digunakan sebagai cetakan untuk
6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

6
menghasilkan serangkaian fragmen yang panjangnya berbeda satu sama lain oleh
satu basa Fragmen kemudian dipisahkan berdasarkan ukuran dan basis di akhir
diidentifikasi menciptakan urutan asli DNA (Twyman 2003 dalam Elviana 2010)
Aplikasi utama dari sequencing DNA dalam studi sistematik adalah i)
evolusi gen termasuk studi dari proses yang menghasilkan level variasi sequence
(urutan basa) studi asal-muasal alel baru atu lokus baru serta investigasi
pemusatan (convergence) dan seleksi ii) studi intraspesifik populasi termasuk
pelacakan organisme dan genealogi alel dalam spesies dan variasi geografik
aliran gen (gen flow) hibridasi serta konservasi genetika dan iii) studi
interspesifik populasi seperti rekonstruksi filogenetik untuk mengevaluasi pola
dan proses evolusi makro (Hillis et al 1996b dalam Elviana 2010)
25 Second Internal Transcribed Spacer (ITS2)
Daerah pada ITS2 dipilih untuk kandidat DNA barcode karena sekuen dari
ITS2 berpotensi untuk penanda filogenetik dan secara luas digunakan untuk
rekonstruksi filogenetik pada kedua genus dan pada tingkat spesies (Chen 2010)
Secara terperinci dapat dilihat pada Gambar 2 bahwa posisi ITS2 termasuk dari
kandidat untuk barcoding namun masih dalam proses penelitian
Gambar 2 Gen dari tiga genom pada tanaman sebagai kandidat untuk barcoding (Chen
2010)
ITS merupakan penanda pada DNA yang bersifat universal penanda
tersebut mampu teramplifikasi pada semua organisme tidak termasuk pada
vertebrata Keuntungan dari ITS adalah dalam membandingkan penanda plastid
7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

7
pada keturunan biparental memiliki tingkat penggandaan yang tinggi daripada
keturunan dari induk betina (Muir et al 2001 dalam Koch et al 2008)
ITS terus menjadi daerah yang lazim digunakan untuk rekonstruksi
filogenetik angka publikasi sekuen ITS meningkat tiga kali lipat semenjak tahun
2003 (Koch et al 2008) daerah ITS terdiri dari ITS1 dan ITS2 yang terletak di
antara ekson 58S dan ujung ITS1 dibatasi oleh ekson 18S sedangkan ujung ITS2
dibatasi oleh ekson 26S daerah tersebut merupakan daerah yang memiliki tingkat
conserved yang tinggi (Koch et al 2008)
Tingkat penggandaan yang tinggi merupakan sebuah alasan bagi aplikasi
ITS yang luas dalam sistem molekular (Rogers dan Bendich 1987 dalam Koch et
al 2008) Seleksi dari daerah kandidat barcoding pada daerah conserved dapat
diselesaikan melalui wilayah ITS Jika dibandingkan dengan gen 58S ITS1 dan
ITS2 lebih tersedia pada sekuen utama Selain itu ITS2 lebih conserved daripada
ITS1 dan dapat memenuhi untuk perunutan pada tingkat genus (Hershkovitz dan
Lewis 1996 dalam Koch et al 2008) Daerah ITS secara terperinci disajikan pada
Gambar 3
Gambar 3 Diagram daerah internal transcribed spacer (Koch 2008)
26 Maturase-K (mat-K)
Mat-K dapat mengamplifikasi gen pada DNA kloroplas (cpDNA) Gen
kloroplas mat-K merupakan sebagian besar variable gen coding dari angiospermae
dan telah diusulkan untuk menjadi barcode pada tanaman (Yu et al 2011) Secara
terperinci daerah mat-K disajikan pada Gambar 4
8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

8
Gambar 4 Wilayah MaturaseK (mat-K) (Wicke 2009)
Wilayah coding mat-K menunjukkan hasil yang sangat bagus dilihat dari
pembagian yang hampir sama pada substitusi nukleotida di posisi kodon satu
kedua dan ketiga Dengan demikian gen mat-K menyusun lebih cepat secara
kontras di gen plastid lainnya walaupun turunan dasar dan fungsi yang terbatas
(Hilu dan Liang 1997 dalam Wicke dan Quandt 2009)
Gen Mat-K telah menunjukkan salah satu dari penggantian nukleotida yang
tertinggi diantara 20 gen plastid yang terbaik dalam panjang 1 kilobase (kb) dan
saat ini digunakan untuk menduga hubungan filogenetik dalam tingkat famili
(Koch et al 2001 dalam Muellner 2003)
9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

9
BAB III
BAHAN DAN METODE
31 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2011 sampai dengan Juli
2012 Kegiatan ekstraksi DNA sampai PCR-RFLP dilakukan di laboratorium
Analisis Genetik Bagian Silvikultur Fakultas Kehutanan Institut Pertanian
Bogor Sequencing DNA contoh dikirim ke PT Genetika Science di Singapura
(httpbase-asiacom)
32 Bahan dan Alat Penelitian
321 Koleksi Contoh Uji Daun
Contoh daun terdiri dari Smacrophylla Smahagoni A indica M
azedarach dan M excelsa yang digunakan untuk analisis PCR dan sequencing
DNA Contoh daun berasal dari tiga lokasi yaitu sekitar kampus Fakultas
Kehutanan IPB (Smacrophylla Smahagoni K anthotheca dan M azedarach)
dan Pusat Penelitian dan Pengembangan Kehutanan Bogor (A indica)
322 Analisis Genetik
Alat dan bahan yang digunakan untuk analisis genetik dengan penanda
PCR terbagi dalam beberapa tahap pekerjaan yaitu tahapan ekstraksi DNA uji
kualitas DNA visualisasi DNA serta analisis data Alat dan bahan yang
digunakan pada teknik PCR disajikan pada Tabel 1
Tabel 1 Alat dan bahan untuk teknik analisis genetik
Analisis Tahapan Kerja
Ekstraksi Uji kualitas DNA PCR
PCR
Alat sarung tangan
tube 2ml mortar sudip
pestel tips mikropipet
mesin centrifugase
vorteks waterbath
freezer desikator
Alat sarung tangan timbangan
analitik gelas ukur tabung
erlenmeyer cetakan agar
microwavemikropipet mesin
elektroforesis fisher
scientificbak EtBr kameraalat
foto DNA mesin UV
Transiluminatorlaptop
Alat sarung tangan
tube 02ml
rakmikrotube alat
tulis mikropipet tips
mesin centrifugasi
mesin PCR PTC - 100
Bahan PVP 1
Chloroform IAA
Isopropanol dingin
NaCl Etanol 100
buffer TE
Bahan agarosee bufer TAE
1x DNA ekstraksi Blue juice
10x EtBr
Bahan DNA primer
spesifik forward dan
reverse Green Go
Taq master mix
(promegaM7122)
10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

10
33 Prosedur Penelitian
Metode yang digunakan dalam penelitian ini adalah teknik PCR Secara
umum prosedur penelitian teknik PCR disajikan pada Gambar 5 Tahap penelitian
terdiri dari tiga tahap analisis yaitu analisis sekuen ITS2 analisis fragmen ITS2
dan analisis fragmen mat-K secara in silico
331 Analisis Sekuen ITS2
Ekstraksi DNA
Kegiatan ekstraksi dan isolasi DNA dari daun dilakukan dengan
menggunakan metode CTAB (cetyl trimethyl ammonium bromide) yang
dimodifikasi untuk mendapatkan DNA yang cukup murni Contoh daun (2x2 cm2)
digerus di dalam pestel bersih lalu dipindahkan ke dalam tube 15 mL dan
ditambahkan 500minus700 larutan penyangga ekstrak dan 100 microL PVP 2 Hasil
gerusan kemudian dikocok dengan vortex agar menjadi homogen selanjutnya
diinkubasi dalam waterbath selama 60 menit pada suhu 650C Untuk mengikat
DNA ditambahkan kloroform 500 microL dan fenol 10 microL lalu dikocok sampai
homogen Sentrifugasi dilakukan untuk memisahkan fase air dan fase bahan
organik (supernatan) pada kecepatan 11000 rpm selama 10 menit
Supernatan diambil dan dipindahkan ke dalam tube baru lalu ditambahkan
isopropanol dingin 500 microL dan NaCl 300 microL Campuran supernatan dengan
isopropanol dingin dan NaCl disimpan dalam freezer selama 60 menit untuk
mendapatkan supernatan DNA Kegiatan selanjutnya adalah pencucian DNA
dengan menambahkan etanol 100 sebanyak 300 microL lalu disentrifugasi pada
kecepatan 11000 rpm selama 10 menit Cairan etanol dibuang dengan hati-hati
agar supernatan DNA tidak ikut terbuang pelet DNA disimpan di dalam desikator
selama plusmn15 menit selanjutnya ditambahkan larutan penyangga TE 20 microL setelah
itu dikocok dan disentrifugasi kembali Langkah tersebut dilakukan sampai
mendapatkan DNA yang murni jika belum mendapatkan DNA yang bersih dan
masih memiliki banyak kotoran perlu diulangi kembali langkah-langkah seperti
yang telah dijelaskan di atas DNA yang bersih dapat diuji melalui tahap
elektroforesis pada gel agarose yang dilakukan pada setiap akhir dari suatu teknik
analisis genetik Agarose yang digunakan memiliki konsentrasi yang berbeda pada
setiap pengujian kualitas DNA sesuai pada teknik yang sedang dilakukan
11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

11
Sampel daun
Ekstraksi DNA
Ya
Elektroforesis
agarose 1
PCR
Sekuen
Restriksi enzim
secara in silico
Analisis Data
Tidak
Ya Tidak
Elektroforesis
agarose 2
5 jenis S mc S
mg M az A in
dan K an
CTAB (Doyle and
Doyle 1990)
-ITS2
PCR (Banks 1985)
5 jenis sampel
ITS2
18 enzim (sesuai
pada 12)
-BioEdit v709
-ClustalW2 EBI-EMBL
-TreeviewX
-PopGene NTSYs dan
pDRAW32
Gambar 5 Bagan alur penelitian di laboratorium (S mc = S macrophylla S mg = S
mahagoni K an = K anthotheca A in = A indica M az =M azedarach)
Pengujian Kualitas DNA
DNA hasil ekstraksi kemudian uji kualitas dengan menggunakan teknik
elektroforesis agarose 1 (wv) Gel ini dibuat dengan melarutkan agarose
sebanyak 033 g ke dalam 33 microL larutan TAE (tris HCl-acetic acid-EDTA)
Kemudian campuran dipanaskan di dalam microwave sampai mendidih Larutan
gel kemudian dituangkan ke dalam cetakan gel Cetakan gel tersebut telah
dipasang sisircomb yang berfungsi untuk membuat sumur gel Setelah gel
membeku sisir dicabut dan gel beserta cetakannya diletakkan di dalam bak
elektroforesis yang telah berisi larutan penyangga TAE DNA hasil ekstraksi
diambil sebanyak 3 microL dan ditambah 2 microL blue juice 10X selanjutnya dilakukan
proses elektroforesis dengan menggunakan aliran listrik dengan tegangan 100 V
selama 1 jam
Gel yang sudah dielektroforesis dilakukan pewarnaan dengan
merendamkan gel di dalam larutan Ethidium Bromida (EtBr) yaitu campuran 10
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2
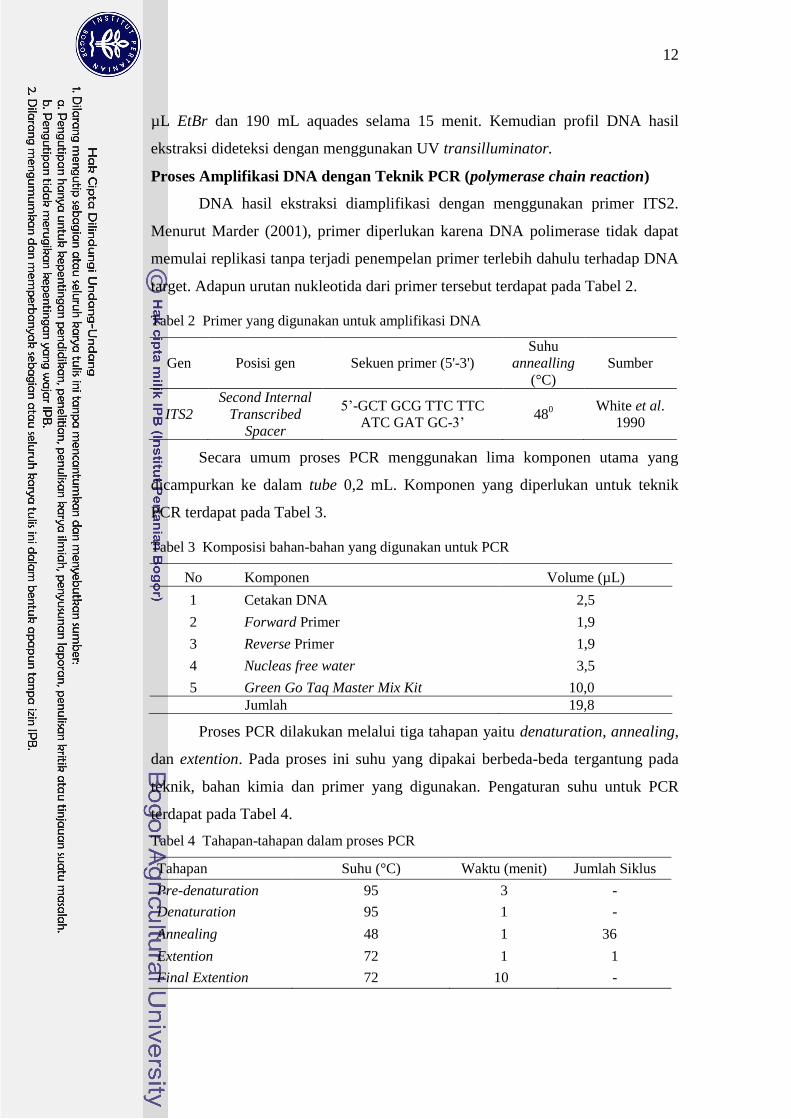
12
microL EtBr dan 190 mL aquades selama 15 menit Kemudian profil DNA hasil
ekstraksi dideteksi dengan menggunakan UV transilluminator
Proses Amplifikasi DNA dengan Teknik PCR (polymerase chain reaction)
DNA hasil ekstraksi diamplifikasi dengan menggunakan primer ITS2
Menurut Marder (2001) primer diperlukan karena DNA polimerase tidak dapat
memulai replikasi tanpa terjadi penempelan primer terlebih dahulu terhadap DNA
target Adapun urutan nukleotida dari primer tersebut terdapat pada Tabel 2
Tabel 2 Primer yang digunakan untuk amplifikasi DNA
Gen Posisi gen Sekuen primer (5-3) Suhu
annealling
(degC) Sumber
ITS2 Second Internal
Transcribed
Spacer
5rsquo-GCT GCG TTC TTC
ATC GAT GC-3rsquo 48
0 White et al
1990
Secara umum proses PCR menggunakan lima komponen utama yang
dicampurkan ke dalam tube 02 mL Komponen yang diperlukan untuk teknik
PCR terdapat pada Tabel 3
Tabel 3 Komposisi bahan-bahan yang digunakan untuk PCR
No Komponen Volume (microL)
1 Cetakan DNA 25
2 Forward Primer 19
3 Reverse Primer 19
4 Nucleas free water 35
5 Green Go Taq Master Mix Kit 100 Jumlah 198
Proses PCR dilakukan melalui tiga tahapan yaitu denaturation annealing
dan extention Pada proses ini suhu yang dipakai berbeda-beda tergantung pada
teknik bahan kimia dan primer yang digunakan Pengaturan suhu untuk PCR
terdapat pada Tabel 4
Tabel 4 Tahapan-tahapan dalam proses PCR
Tahapan Suhu (degC) Waktu (menit) Jumlah Siklus
Pre-denaturation 95 3 -
Denaturation 95 1 -
Annealing 48 1 36
Extention 72 1 1
Final Extention 72 10 -
13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

13
Hasil PCR kemudian dianalisis dengan melakukan elektroforesis gel agarose 2
(wv) dalam larutan buffer TAE 1x Setelah running selesai dilakukan distaining
dalam larutan Ethidium Bromide (EtBr)
Sequencing DNA
Proses sequencing dilakukan setelah DNA hasil amplifikasi diukur
kualitasnya dengan cara dielektroforesis pada gel agarose 2 DNA hasil
amplifikasi tersebut kemudian dikirim ke PT Genetika Science Indonesia di
Singapura bersama primer yang telah digunakan dalam proses amplifikasi
Kemudian dianalisis dengan program ClustalW2 EMBL-EBI
(httpebiacukToolsmsaclustal-w2) software BioEdit dan TreeViewX (Hall
2007) serta analisis keanekaragaman menggunakan software DNAsp versi 35
(Rozas dan Rozas 1999) digunakan untuk mencari nilai keragaman nukleotida
332 Analisis Fragmen ITS2 secara In Silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
333 Analisis Fragmen matK
Data Mining Meliaceae
Data mining tidak dilakukan melalui prosedur kerja pada laboratorium
Data ini diambil dari database GenBank (httpwwwncbinlmnihg-ovgenbank)
untuk lima spesies yang sama pada region ITS2 ukuran sekuen yang digunakan
berbeda pada kedua region untuk eksplorasi marker pad mat-K digunakan sekuen
DNA dengan pangjang plusmn 800 bp secara terperinci dapat dilihat seperti disajikan
pada Tabel 5
Tabel 5 Spesies yang digunakan untuk region mat-K (GenBank 2012)
Spesies Sumber GenBank
Accession Panjang
basa (bp)
Swietenia macrophylla
Swietenia mahagoni
Azadirachta indica
Melia azedarach
Khaya anthotheca
Muellner et al (2003)
Clayton et al (2007)
Muellner et al (2003)
Abbott et al (2009)
Muellner et al (2003)
AY128200
EU042907
AY128180
GU134981
AY128190
857
918
863
827
857
14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

14
Sequence DNA
Runutan DNA dari data mining dirunutkan dengan menggunakan program
perangkat lunak Perangkat lunak yang digunakan adalah ClustalW2 software
BioEdit TreeViewX
Analisis Fragmen matK secara in silico
Sekuen DNA dimasukkan pada program pDRAW32 yang dikembangkan
oleh AcaClone Software (httpwwwacaclonecom) Masing-masing sekuen
DNA kemudian dipotong menggunakan 18 enzim restriksi Setelah dipotong
kemudian ditampilkan dengan elektroforesis gel agarose 2
15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

15
BAB IV
HASIL DAN PEMBAHASAN
41 Analisis DNA
411 Ekstraksi DNA
Ekstraksi DNA merupakan langkah awal dalam analisis molekuler
Masalah-masalah yang timbul dalam ekstraksi DNA merupakan hal yang penting
untuk diperhatikan dan perlu diatasi Kualitas dan kuantitas hasil dari ekstraksi
DNA tergantung dari spesies tanaman yang digunakan dan mempengaruhi analisis
lanjut seperti pada proses PCR maupun pemotongan DNA dengan enzim restriksi
Gambar 6 Pola band DNA A) K anthotheca B) S macrophylla C) S mahagoni D) A
indica E) M azedarach 1) sumur DNA 2) band DNA
Hasil ekstraksi DNA yang disajikan pada Gambar 6 ada beberapa band
yang tidak muncul dan kurang jelas Pola tersebut dapat disebabkan tidak adanya
atau kurang mencukupinya DNA yang terisolasi untuk uji elektroforesis selain itu
kemungkinan masih adanya DNA di dalam tube karena campuran tidak homogen
jumlah ekstrak yang sedikit atau terbuang pada saat pencucian Hal lain yang bisa
terjadi adalah terkontaminasinya DNA atau memiliki tingkat kemurnian yang
rendah
Kondisi DNA dengan tingkat kemurnian dan berat molekul yang tinggi
sangat diupayakan agar dapat melakukan analisis genetika molekuler Ekstraksi
A B C E D
2
1
16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

16
DNA pada jaringan tanaman dengan kemurnian yang tinggi seringkali sulit
diperoleh untuk itu diberi perlakuan pengenceran DNA yang berbeda-beda
dilihat dari segi kualitasnya Pengenceran ini bertujuan untuk memudahkan saat
melakukan PCR primer dapat teramplifikasi pada band DNA dengan baik Jika
DNA terlalu banyak kotoran maka hasil PCR tidak bagus karena primer tidak
dapat teramplifikasi
412 Analisis Hasil PCR
PCR merupakan salah satu teknik analisis DNA yang dapat digunakan
untuk mempelajari proses evolusi tanaman (Elviana 2010) Kesesuaian dalam
menggunakan primer sangat tergantung pada teramplifikasinya DNA dengan baik
sehingga mengasilkan pola band yang bagus Primer universal yang berhasil
teramplifikasi untuk beberapa spesies dalam famili Meliaceae di antaranya terdiri
dari S macrophylla S mahagony M azedarach A indica dan K anthotheca
adalah second internal transcribed spacer (ITS2) Menurut Chen (2010) region
untuk ITS2 cukup pendek yakni ada pada selang 160-320bp Panjang fragmen
hasil amplifikasi primer ITS2 adalah 307 bp seperti yang disajikan pada Gambar
7
Gambar 7 Hasil amplifikasi PCR ITS2 (1278 M az 34 Smc 56 S mg 910 A in
1112 K an)
Hasil dari amplifikasi secara umum tidak memperlihatkan band yang
sangat jelas ada beberapa band yang kurang jelas sehingga menimbulkan
keraguan saat menganalisis band Ada beberapa faktor yang menyebabkan hal
tersebut di antaranya tingkat kemurnian DNA yang rendah dan campuran bahan
yang kurang tepat Konsentrasi DNA sampel ukuran panjang primer komposisi
307 bp
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2
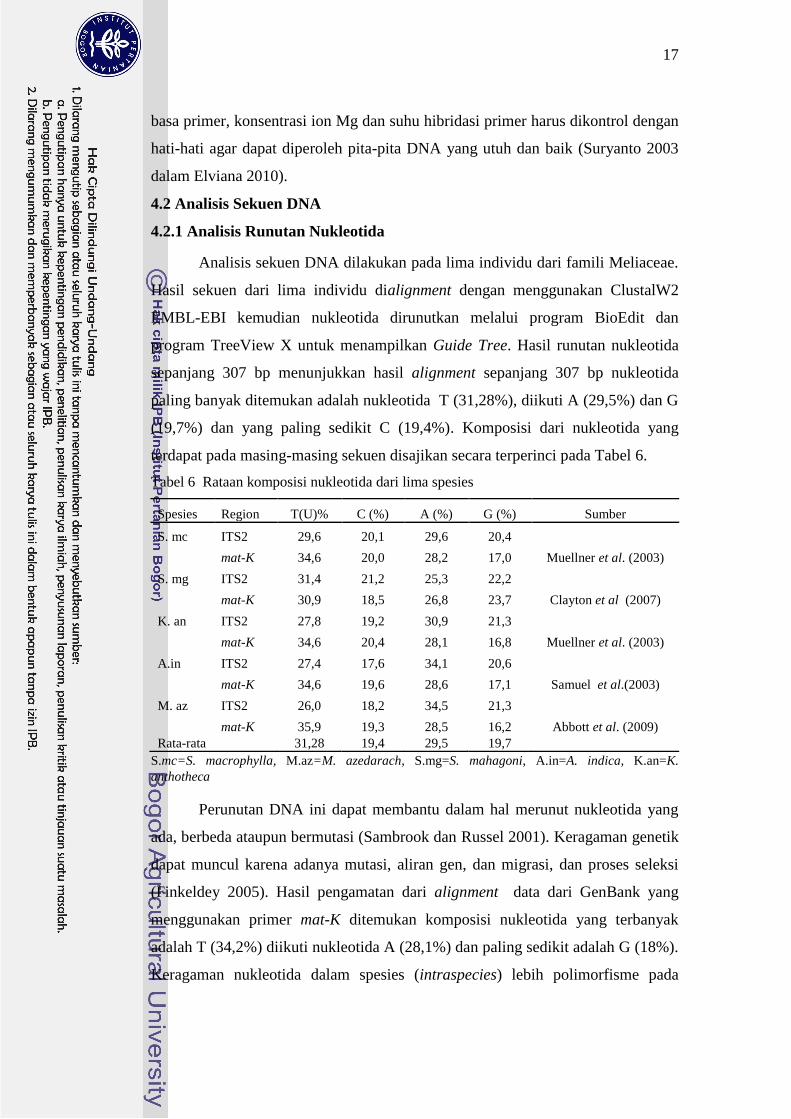
17
basa primer konsentrasi ion Mg dan suhu hibridasi primer harus dikontrol dengan
hati-hati agar dapat diperoleh pita-pita DNA yang utuh dan baik (Suryanto 2003
dalam Elviana 2010)
42 Analisis Sekuen DNA
421 Analisis Runutan Nukleotida
Analisis sekuen DNA dilakukan pada lima individu dari famili Meliaceae
Hasil sekuen dari lima individu dialignment dengan menggunakan ClustalW2
EMBL-EBI kemudian nukleotida dirunutkan melalui program BioEdit dan
program TreeView X untuk menampilkan Guide Tree Hasil runutan nukleotida
sepanjang 307 bp menunjukkan hasil alignment sepanjang 307 bp nukleotida
paling banyak ditemukan adalah nukleotida T (3128) diikuti A (295) dan G
(197) dan yang paling sedikit C (194) Komposisi dari nukleotida yang
terdapat pada masing-masing sekuen disajikan secara terperinci pada Tabel 6
Tabel 6 Rataan komposisi nukleotida dari lima spesies
Spesies Region T(U) C () A () G () Sumber
S mc ITS2 296 201 296 204
mat-K 346 200 282 170 Muellner et al (2003)
S mg ITS2 314 212 253 222
mat-K 309 185 268 237 Clayton et al (2007)
K an ITS2 278 192 309 213
mat-K 346 204 281 168 Muellner et al (2003)
Ain ITS2 274 176 341 206
mat-K 346 196 286 171 Samuel et al(2003)
M az ITS2 260 182 345 213
mat-K 359 193 285 162 Abbott et al (2009)
Rata-rata 3128 194 295 197
Smc=S macrophylla Maz=M azedarach Smg=S mahagoni Ain=A indica Kan=K
anthotheca
Perunutan DNA ini dapat membantu dalam hal merunut nukleotida yang
ada berbeda ataupun bermutasi (Sambrook dan Russel 2001) Keragaman genetik
dapat muncul karena adanya mutasi aliran gen dan migrasi dan proses seleksi
(Finkeldey 2005) Hasil pengamatan dari alignment data dari GenBank yang
menggunakan primer mat-K ditemukan komposisi nukleotida yang terbanyak
adalah T (342) diikuti nukleotida A (281) dan paling sedikit adalah G (18)
Keragaman nukleotida dalam spesies (intraspecies) lebih polimorfisme pada
18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

18
region mat-K sebab dipengaruhi oleh panjang bp yang dimiliki mat-K lebih besar
jika dibandingkan dengan region ITS2 Jika dibandingkan dengan persentase
keragaman berdasarkan nukleotida pada spesies Shorea laevis dari famili
Dipterocarpaceae pada region yang sama yaitu mat-K sepanjag 844 bp diperoleh
hasil untuk nukleotida T (352) diikuti A (326) G (178) dan C (145)
(Elviana 2010) dapat dilihat komposisi nukleotidanya tidak memiliki selisih
komposisi yang cukup jauh Komposisi nukleotida dari manusia juga dapat dilihat
yaitu nukleotida A (309) nukleotida T (294) nukleotida G (199) dan
nukleotida C (198) dengan DNA normal dari spesies yang berbeda
menunjukkan adanya keteraturan yaitu di antaranya komposisi basa dari DNA
suatu organisme adalah tetap pada semua selnya dan mempunyai karakterisitik
tertentu selain itu jumlah adenin dengan timin selalu sama dalam DNA suatu
organisme hal yang sama juga terjadi pada jumlah guanin dan sitosin (OCW
2009) Perunutan DNA dari spesies yang digunakan secara terperinci disajikan
pada Gambar 8
Gambar 8 Runutan nukleotida dari lima species(mc_4 S macrophylla ma_I M
azedarach mg_32 S mahagoni K3 K anthotheca mb_3 A indica)
yang di alignment dari EBL-EBI Tanda bintang menunjukkan nukleotida
yang sama
Hubungan kekerabatan antara dua individu atau lebih dalam suatu famili
dapat diukur dari kesamaan beberapa sifat dengan asumsi bahwa karakter yang
19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

19
berbeda disebabkan oleh perbedaan genetik yang dimiliki oleh masing-masing
individu Runutan hasil sekuen dari lima spesies bila dibandingkan dengan
kombinasi data sekuen dari GenBank yaitu runutan DNA menggunakan primer
mat-K dengan species yang sama dari famili Meliaceae diantaranya S macrophylla
(Muellner et al 2002) S mahagoni (Clayton et al 2007) K anthotheca (Muellner et al
2002) M azedarach (Abbott et al 2009) dan A indica (Muellner et al 2002) secara
teperinci disajikan pada Lampiran 3
422 Keragaman antar spesies
Keragaman situs restriksi secara In Silico
Hasil sekuen diberi perlakuan pemotongan fragmen dengan 18 spesies
enzim melalui program pDRAW32 Hasil pemotongan kemudian
diinterpretasikan pada pada 3 gel agarose marker Promega 100 bp DNA ladder
seperti yang disajikan pada Lampiran 1 berdasarkan wilayah ITS2 dan mat-K
dapat dilihat pada Lampiran 2 dari analisis menggunakan primer ITS2 dan mat-K
yang dipotong dengan 18 enzim secara in silico terjadi banyak pemotongan Hasil
pemotongan tersebut menunjukkan variasi DNA (polimorfisme) pada setiap
spesies pola pemotongan yang dihasilkan berbeda untuk masing-masing spesies
Setiap spesies dipotong oleh enzim yang berbeda dan memiliki enzim tertentu
yang hanya memotong pada satu spesies dapat dilihat secara terperinci pada
Tabel 7
Tabel 7 Spesies enzim yang hanya memotong pada satu spesies berdasarkan dua region
Region Kode Spesies Enzim ITS2 M az M azedarach BstUI A in Aindica HpaII
K an K anthotheca HhaI
S mc S macrophylla MseI
S mg S mahagoni AluI
mat-K M az M azedarach BamHI BfaI EcoRI HaeIII HinfI MseI
RsaI SspI TaqI A in Aindica BstUI HpaII
S mg S mahagoni AluI HindIII HpaII PstI
S mc S macrophylla EcoRI HpaII
K an K anthotheca HaeIII
Hasil pemotongan kembali dilihat jarak kekerabatan antara semua individu
yang diamati yang diinterpretasikan pada dendrogram Dendrogram yang
disajikan menunjukkan spesies yang satu klaster atau berada dalam klaster yang
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2
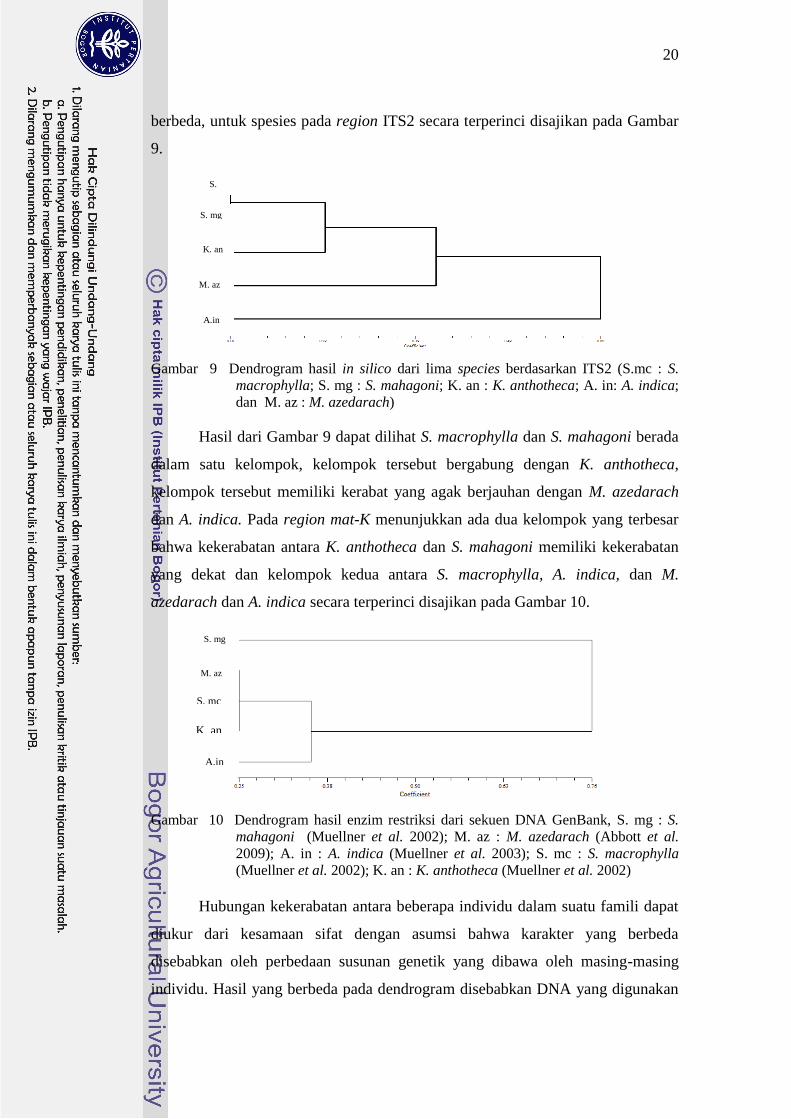
20
berbeda untuk spesies pada region ITS2 secara terperinci disajikan pada Gambar
9
Gambar 9 Dendrogram hasil in silico dari lima species berdasarkan ITS2 (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach)
Hasil dari Gambar 9 dapat dilihat S macrophylla dan S mahagoni berada
dalam satu kelompok kelompok tersebut bergabung dengan K anthotheca
kelompok tersebut memiliki kerabat yang agak berjauhan dengan M azedarach
dan A indica Pada region mat-K menunjukkan ada dua kelompok yang terbesar
bahwa kekerabatan antara K anthotheca dan S mahagoni memiliki kekerabatan
yang dekat dan kelompok kedua antara S macrophylla A indica dan M
azedarach dan A indica secara terperinci disajikan pada Gambar 10
Gambar 10 Dendrogram hasil enzim restriksi dari sekuen DNA GenBank S mg S
mahagoni (Muellner et al 2002) M az M azedarach (Abbott et al
2009) A in A indica (Muellner et al 2003) S mc S macrophylla
(Muellner et al 2002) K an K anthotheca (Muellner et al 2002)
Hubungan kekerabatan antara beberapa individu dalam suatu famili dapat
diukur dari kesamaan sifat dengan asumsi bahwa karakter yang berbeda
disebabkan oleh perbedaan susunan genetik yang dibawa oleh masing-masing
individu Hasil yang berbeda pada dendrogram disebabkan DNA yang digunakan
Ain
M az
K an
S mg
S
mc
S mg
K an
M az
S mc
Ain
21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

21
berbeda dari segi primer sehingga DNA yang dihasilkan juga berbeda untuk
ITS2 merupakan wilayah nukleus sedangkan mat-K merupakan wilayah cpDNA
Keragaman nukleotida
Salah satu pengelompokan dari kelima individu pada region ITS2
berdasarkan perbedaan runutan nukleotida melalui analisis dendrogram yang
disajikan pada Gambar 11 menunjukkan jarak kekerabatan yang terdiri tiga klaster
yaitu A indica dan M azedarach disusul K anthotheca dan S mahagoni dan
terakhir S macrophylla
Gambar 11 Dendrogram dari lima species berdasarkan ITS2 (Smc S macrophylla S
mg S mahagoni K an K anthotheca A in A indica dan M az M
azedarach)
Jika dibandingkan dengan daerah ITS berdasarkan hasil penelitian Muellner
(2011) jarak genetiknya menunjukkan hasil yang sesuai dengan region ITS2 K
anthotheca dan S macrophylla satu klaster serta A indica dan M azedarach
berada dalam satu klaster seperti yang disajikan pada Gambar 12
Gambar 12 Dendrogram region ITS (Muellner 2011)
K anthotheca
S mahagoni
S macrophylla
A indica
M azedarach
000005010015020
Smg
Smc
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2
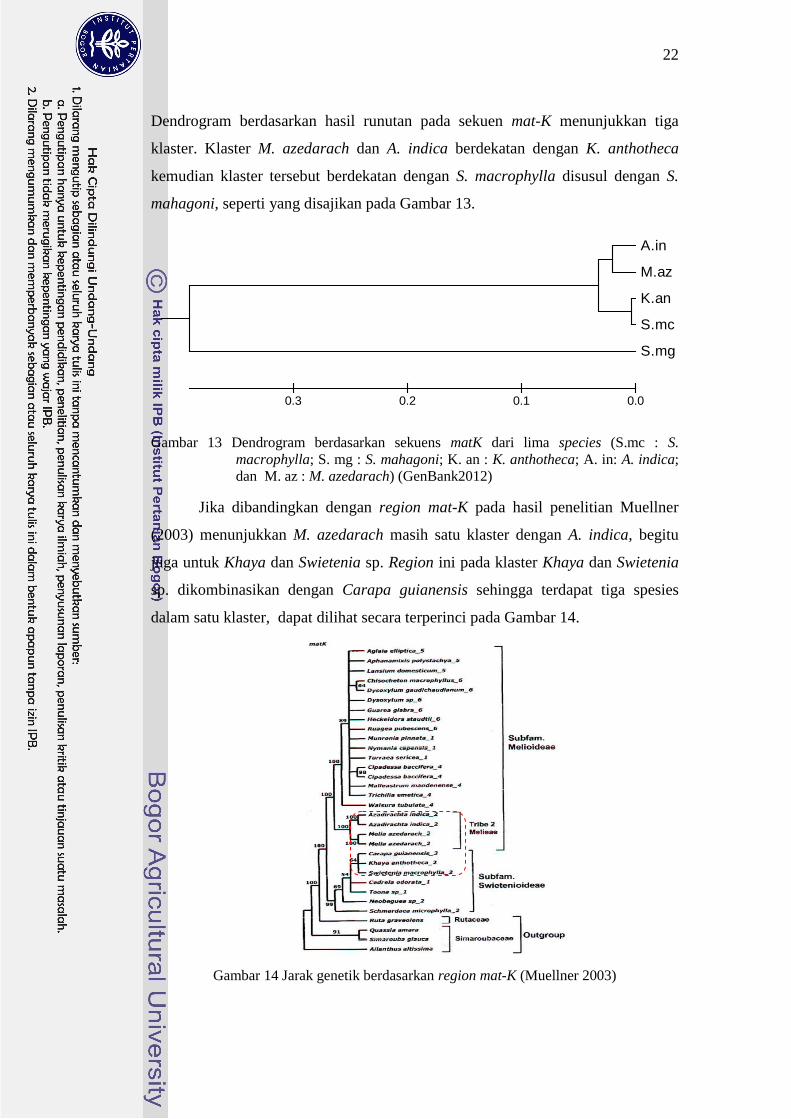
22
Dendrogram berdasarkan hasil runutan pada sekuen mat-K menunjukkan tiga
klaster Klaster M azedarach dan A indica berdekatan dengan K anthotheca
kemudian klaster tersebut berdekatan dengan S macrophylla disusul dengan S
mahagoni seperti yang disajikan pada Gambar 13
Gambar 13 Dendrogram berdasarkan sekuens matK dari lima species (Smc S
macrophylla S mg S mahagoni K an K anthotheca A in A indica
dan M az M azedarach) (GenBank2012)
Jika dibandingkan dengan region mat-K pada hasil penelitian Muellner
(2003) menunjukkan M azedarach masih satu klaster dengan A indica begitu
juga untuk Khaya dan Swietenia sp Region ini pada klaster Khaya dan Swietenia
sp dikombinasikan dengan Carapa guianensis sehingga terdapat tiga spesies
dalam satu klaster dapat dilihat secara terperinci pada Gambar 14
Gambar 14 Jarak genetik berdasarkan region mat-K (Muellner 2003)
Ain
Maz
Kan
Smc
Smg
00010203
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2
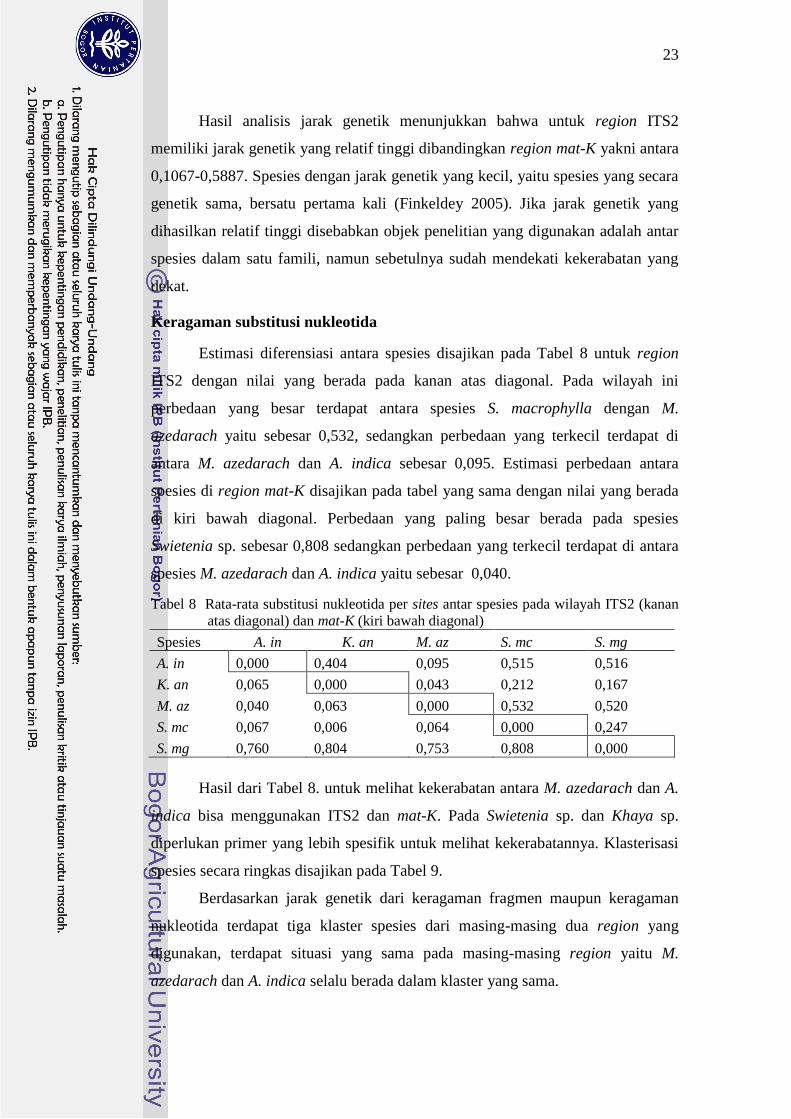
23
Hasil analisis jarak genetik menunjukkan bahwa untuk region ITS2
memiliki jarak genetik yang relatif tinggi dibandingkan region mat-K yakni antara
01067-05887 Spesies dengan jarak genetik yang kecil yaitu spesies yang secara
genetik sama bersatu pertama kali (Finkeldey 2005) Jika jarak genetik yang
dihasilkan relatif tinggi disebabkan objek penelitian yang digunakan adalah antar
spesies dalam satu famili namun sebetulnya sudah mendekati kekerabatan yang
dekat
Keragaman substitusi nukleotida
Estimasi diferensiasi antara spesies disajikan pada Tabel 8 untuk region
ITS2 dengan nilai yang berada pada kanan atas diagonal Pada wilayah ini
perbedaan yang besar terdapat antara spesies S macrophylla dengan M
azedarach yaitu sebesar 0532 sedangkan perbedaan yang terkecil terdapat di
antara M azedarach dan A indica sebesar 0095 Estimasi perbedaan antara
spesies di region mat-K disajikan pada tabel yang sama dengan nilai yang berada
di kiri bawah diagonal Perbedaan yang paling besar berada pada spesies
Swietenia sp sebesar 0808 sedangkan perbedaan yang terkecil terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
Tabel 8 Rata-rata substitusi nukleotida per sites antar spesies pada wilayah ITS2 (kanan
atas diagonal) dan mat-K (kiri bawah diagonal)
Spesies A in K an M az S mc S mg
A in 0000 0404 0095 0515 0516
K an 0065 0000 0043 0212 0167
M az 0040 0063 0000 0532 0520
S mc 0067 0006 0064 0000 0247
S mg 0760 0804 0753 0808 0000
Hasil dari Tabel 8 untuk melihat kekerabatan antara M azedarach dan A
indica bisa menggunakan ITS2 dan mat-K Pada Swietenia sp dan Khaya sp
diperlukan primer yang lebih spesifik untuk melihat kekerabatannya Klasterisasi
spesies secara ringkas disajikan pada Tabel 9
Berdasarkan jarak genetik dari keragaman fragmen maupun keragaman
nukleotida terdapat tiga klaster spesies dari masing-masing dua region yang
digunakan terdapat situasi yang sama pada masing-masing region yaitu M
azedarach dan A indica selalu berada dalam klaster yang sama
24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

24
Tabel 9 Klaster spesies berdasarkan ITS2 dan mat-K
Klaster ITS2 Mat-K
1 M azedarach dan A indica A indica dan M Azedarach
2 S mahagoni dan K anthotheca K anthotheca dan (A indica dan M
azedarach) 3 S macrophylla S macrophylla S mahagoni
25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

25
BAB V
KESIMPULAN DAN SARAN
51 Kesimpulan
Kesimpulan dari penelitian ini adalah sebagai berikut i) aplikasi enzim
restriksi dapat mendeteksi adanya keragaman untuk semua spesies pada sebagian
dari dua wilayah gen ii) berdasarkan variasi nukleotida keragaman antar spesies
dengan memiliki klaster yang sama yakni antara M azedarach dan A indica Pada
kedua wilayah DNA terdapat tiga kelompok kekerabatan famili Meliaceae
Perbedaan yang terkecil berdasarkan komposisi nukleotida terdapat di antara
spesies M azedarach dan A indica yaitu sebesar 0040
52 Saran
Eksplorasi primer yang lebih spesifik perlu dilakukan lebih lanjut untuk
menduga variasi DNA pada wilayah-wilayah gen yang konservatif (cpDNA)
26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

26
DAFTAR PUSTAKA
Ahmed S Idris S 1997 Melia azedarach L (Meliaceae) auxiliary plants plants
resources of South-East Asia Prosea 11187-190
[BTP] Balai Teknologi Perbenihan 1986 Petunjuk Teknis Penangan dan
Pengujian Mutu Benih mahoni Bogor (ID) BTP
Burhaman 2004 Atlas Benih Tanaman Hutan Indonesia Jilid 2 Bogor Balai
Penelitian dan Pengembangan Kehutanan
Chen S Yao H Han J Liu C Song J Sji L Zhu Y Ma X Gao T Pang X et al
2010 Validation of the ITS2 region as a novel DNA barcode for identifying
medicinal plant species Plos One 5(1)1-8
[Dephut] Departemen Kehutanan 2001 Mindi (Melia azedarach L) Jakarta
Badan Penelitian dan Pengembangan Kehutanan
Elviana 2010 Keragaman genetik kloroplas (cpDNA) bangkirai (Shorea laevis
Ridl) di Kalimantan berdasarkan penanda PCR-RFLP dan sekuens sebagian
wilayah trnl-f [skripsi] Bogor (ID) Fakultas Kehutanan Institut Pertanian
Bogor
Finkeldey R 2005 Pengantar Genetika Hutan Tropis Djamhuri E Siregar IZ
Siregar UJ Kertadikara AW penerjemah Goettingen Institute of Forest
Genetics and Forest Tree Breeding Georg-August-University-Goettingen
Terjemahan dari An Introduction to Tropical Forest Genetics
Hall T 2007 Populations genetics alignment tools [terhubung berkala]
httpwwwmbioncsueduBio_Editbio_edit html [25 April 2012]
Koch MA Calonje M Gong W 2008 Non-coding nuclear DNA markers in
phylogenetics reconstruction Plant Syst Evol 282257-280
Mahfira UO 2010 Variasi kloroplas Shorea laevis Ridl di Kalimantan
berdasarkan penanda mikrosatelit [skripsi] Bogor (ID) Fakultas
Kehutanan Institut Pertanian Bogor
Muellner AN Samuel R Johnson SA Cheek M Pennington TD 2003 Molecular
phylogenetics of Meliaceae (Sapindales) based on nuclear and plastid DNA
sequences American Journal of Botany 90(3)471-480
Muellner AN Schaefer H Lahaye R 2011 DNA barcoding-evaluation of
candidate DNA barcoding loci for economically important timber species of
the mahogany family (Meliaceae) Molecular Ecology Resources 11450-
460
Nei M 1987 InMolecular Evolutionary Genetics New York Columbia
University Press
27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

27
OCW 2009 Asam nukleat (nucleic acid) [terhubung berkala]
httpocwusuacidbbc115_slide_asam_nukleat_atau_nucleic_acidpdf
[10 Okt 2012]
Putri KP Yulianti B 2003 Info Benih Pemilihan Spesies Eksotik Sebagai
Alternatif Bogor (ID) Badan Penelitian dan Pengembangan Kehutanan
Rice G 2009 Polymerase chain reaction (PCR) [terhubung berkala]
httpserccarletonedumicrobe-liferesearches-methodsgenomics-pcrhtml
[27 Juli 2012]
Rukmana R Oesman YY 2002 Nimba Tanaman Penghasil Pestisida Alami
Yogyakarta Kanisius
Siregar IZ 2000 Genetic aspects of the reproductive system of Pinus merkusii
Jungh et de Vriese in Indonesia [disertasi] Gottingen Faculty of Forest
Sciences and Forest Ecology Georg-August University of Gottingen
Suryanto D 2001 Selection and characterization of bacterial isolates for
monocyclic aromatic degradation [disertasi] Bogor (ID) Institut Pertanian
Bogor
Soerianegara I Djamhuri E 1979 Pemuliaan Pohon Hutan Bogor Departemen
Manajemen Kehutanan Fakultas Kehutanan Institut Pertanian Bogor
Whitmore TC Tantra IGM Tree Flora of Indonesia Bogor (ID) Sumatra check
list FRDC
Wicke S Quandt D 2009 Universal primers for the amplification of the plastid
trnKmatK region in land plants Anales Jard Bot Madrid 66(2)285-288
Yu J Xue JH Zhou SL 2011 New universal matK primers for DNA barcoding
angiosperms Journal of Systematics and Evolution 49176-181
28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

28
LAMPIRAN
29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

29
Lampiran 1 Elektroferogram enzim restriksi secara in silico pada ITS2 Ploting hasil
analisis in silico menggunakan 18 enzim restiksi (HindIII HinfI PstI AluI
BamHI BfaI BstUI DraI EcoRI HaeIII HhaI HpaI HpaII KpnI
Sau3AI MseI RsaI SspI TaqI) terhadap 5 sekuen DNA yang berbeda
spesies pada 3 gel agarose marker Promega 100bp DNA step ladder
S macrophylla 294 bp
M azedarach 307 bp
S mahagoni 302 bp
b
)
a
c
30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

30
Lanjutan Lampiran 1
A indica 307 bp
K anthotheca 305 bp
d
e
31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

31
Lampiran 2 Elektroferogram enzim restriksi secara In Silico pada mat-K Ploting
hasil analisis in silico menggunakan 18 enzim restiksi (HindIII
HinfI PstI AluI BamHI BfaI BstUI DraI EcoRI HaeIII HhaI
HpaI HpaII KpnI Sau3AI MseI RsaI SspI TaqI) terhadap 5
sekuen DNA yang berbeda spesies pada 3 gel agarose marker
Promega 100bp DNA step ladder
S mahagoni (Clayton et al 2007) 918 bp
M azedarach (Abbott et al 2009) 827bp
A indica (Muellner et al 2002b) 863 bp
1
2
3
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2
32
Lanjutan Lampiran 2
S macrophylla (Muellner et al 2002) 857 bp
K anthotheca (Muellner et al 2002) 857 bp
4
5
33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

33
Lampiran 3 Runutan sekuen mat-K Alignment runutan nukleotida dari lima species
dari GenBank (httpwwwncbinlmnihgov) kemudian di alignment dari
EBL-EBI (httpwwwebiacuk) Tanda bintang menunjukkan nukleotida
yang sama
34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

34
Lanjutan Lampiran 3
35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

35
Lampiran 4 Polimorfik pada lima spesies Meliaceae di region mat-K
36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

36
Lanjutan Lampiran 4
37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2

37
Lampiran 5 Polimorfik pada lima spesies Meliaceae di region ITS2
Top Related
Copyright © 2022 FDOKUMEN

