
ZIERITZ, A. 2010. Variability, function and phylogenetic significance of unionoid shell characters....
191
VARIABILITY, FUNCTION AND PHYLOGENETIC SIGNIFICANCE OF UNIONOID SHELL CHARACTERS by ALEXANDRA ZIERITZ ST CATHARINE’S COLLEGE SUBMITTED FOR THE DEGREE OF DOCTOR OF PHILOSPHY UNIVERSITY OF CAMBRIDGE,JUNE 2010 SUPERVISOR: DR.DAVID CHRISTOPHER ALDRIDGE
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of ZIERITZ, A. 2010. Variability, function and phylogenetic significance of unionoid shell characters....

VARIABILITY, FUNCTION AND
PHYLOGENETIC SIGNIFICANCE OF
UNIONOID SHELL CHARACTERS
by
ALEXANDRA ZIERITZ
ST CATHARINE’S COLLEGE
SUBMITTED FOR THE DEGREE OF DOCTOR OF PHILOSPHY
UNIVERSITY OF CAMBRIDGE, JUNE 2010
SUPERVISOR:
DR. DAVID CHRISTOPHER ALDRIDGE


iii
PREFACE
This dissertation is submitted for the degree of Doctor of Philosophy. It is the result of my
own work and includes nothing which is the outcome of work done in collaboration except
as stated in the following:
Dr. Aldridge (PhD supervisor) provided general scientific guidance and advice, and helpful
comments on previous drafts of all chapters.
Dr. Hoffman and Prof. Amos supervised AFLP analysis. They also provided helpful advice and
comments regarding statistical analysis of AFLP data, and helpful comments on a previous draft of
Chapter 3.
Dr. Bogan provided some helpful comments and discussion on fossil unionoids, and helpful
comments on a previous draft of Chapter 5.
Dr. Harper and Prof. Checa provided invaluable expertise and discussion on bivalve
microsculptures. Dr. Harper furthermore provided supervision during scanning electron
microscopy and helpful comments on a previous draft of Chapter 6.
No part of this dissertation has been or is being concurrently submitted for a degree,
diploma or other qualification at any other university.
Permission is granted to consult or copy the information contained herein for the purpose
of private study, but not for publication.
This thesis does not exceed the limit prescribed by the Degree Committee of Biology as
stated in the Memorandum to Graduate Students.
Alexandra Zieritz


CONTENTS
Title page iPreface iiiContents vAcknowledgements ixSummary xiGlossary xiii Chapter 1 - Introduction 1
1.1 A brief introduction to the uniqueness of unionoids 41.1.1 Unionoid phylogeny, diversity and distribution 41.1.2 Importance and conservation status 71.1.3 Life cycle and morphological variation 7
1.2 Importance of understanding morphological patterns in unionoid shells
8
1.2.1 Systematics, phylogeny and evolution 91.2.2 Ecology, conservation and environmental reconstruction 101.2.3 Reconstruction of population parameters 11
1.3 Aims and questions addressed in this thesis 111.3.1 “Gross shell morphology”: size, shape, inflation and
thickness 12
1.3.2 Umbonal sculpture 121.3.3 Periostracal microprojections 13
1.4 Style of thesis 13 Chapter 2 - Identification of ecophenotypic trends within three European
freshwater mussel species (Bivalvia: Unionoida) using traditional and modern morphometric techniques
15
2.1 Abstract 172.2 Introduction 172.3 Materials and methods 19
2.3.1 Sampling of individuals 192.3.2 Sampling of habitat parameters 212.3.3 Growth measurements, and age and sex determination of
individuals 21
2.3.4 Morphological analysis 212.3.4.1 Fourier shape analysis 212.3.4.2 Analysis of traditional shell and anatomical
measurements 22

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
vi
2.3.5 Statistical analysis 232.4 Results 24
2.4.1 Habitat parameters 242.4.2 Species composition 242.4.3 Growth 252.4.4 Morphological analysis 26
2.4.4.1 Morphological trends in U. pictorum of five paired sites and influence of habitat and non-habitat factors
26
2.4.4.2 Morphological trends in all three unionoid species at Abingdon
28
2.5 Discussion 302.5.1 Patterns across all species 302.5.2 Patterns within single species 322.5.3 Utility of the patterns observed 33
Chapter 3 - Phenotypic plasticity and genetic isolation-by-distance in the
freshwater mussel Unio pictorum (Mollusca: Unionoida) 35
3.1 Abstract 373.2 Introduction 373.3 Materials and methods 39
3.3.1 Sampling 393.3.2 Morphological analysis 403.3.3 Genetic analysis 413.3.4 Quantification of the genotyping error rate 433.3.5 Genetic data analysis 43
3.4 Results 443.4.1 Morphological analysis 443.4.2 Population structure 46
3.5 Discussion 493.5.1 Genetic population structure 503.5.2 Phenotypic plasticity of shell form 51
Chapter 4 - Sexual, habitat-constrained and parasite-induced dimorphism in
the shell of a freshwater mussel (Anodonta anatina, Unionidae) 55
4.1 Abstract 574.2 Introduction 574.3 Materials and methods 60
4.3.1 Sampling, sex determination and trematode infection 604.3.2 Size and growth rates 614.3.3 Shell shape, thickness and density 61

Glossary
vii
4.4 Results 634.4.1 Comparison of the five study populations 634.4.2 Sex ratios 644.4.3 Size and growth rates 644.4.4 Shell shape, thickness and density 654.4.5 Trematode parasitism 70
4.5 Discussion 714.5.1 Size and hermaphroditism 714.5.2 Sexual dimorphism in sagittal shell shape 724.5.3 Sexual dimorphism in relative shell width 724.5.4 Sexual dimorphism in shell thickness and density 734.5.5 Trematode parasite-induced dimorphism 744.5.6 Habitat-constrained dimorphism 744.5.7 Application of the patterns observed 75
Chapter 5 - Variability and a new model for character evolution of umbonal sculptures in the Unionoida
77
5.1 Abstract 795.2 Introduction 795.3 Materials and methods 82
5.3.1 Classification of beak sculpture types 825.3.2 Development of model of character evolution, and
identification of homologies and homoplasies 83
5.4 Results 845.4.1 Morphological types of unionoid beak sculpture 845.4.2 Intermediate forms and implications for character evolution
of unionoid beak sculpture 88
5.5 Discussion 895.5.1 (In)validity of previous models of character evolution 895.5.2 Plesiomorphic character state and the fossil record 90
5.5.2.1 Smooth vs. sculptured plesiomorphic character state 905.5.2.2 Most likely plesiomorphic beak sculpture type 91
5.5.3 A new model of beak sculpture character evolution in the Unionoida
91
5.5.3.1 Implications for unionoid phylogeny and evolution 945.5.3.2 Convergences and implications for probable functional
morphologies 94
Chapter 6 - Variability, function and phylogenetic significance of periostracal
microprojections in palaeoheterodont bivalves 97
6.1 Abstract 996.2 Introduction 99

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
viii
6.3 Materials and methods 1046.4 Results 105
6.4.1 Occurrence and morphology of periostracal microprojections in Palaeoheterodonta
105
6.4.2 Variation in spike morphology, abundance and distribution across the shell
108
6.4.3 Mineralisation status of periostracal microprojections 1106.4.4 Trigonioid ‘bosses’ 1126.4.5 Influence of periostracal thickness on presence/absence of
microprojections 112
6.5 Discussion 1136.5.1 Occurrence of periostracal microprojections across the
unionoid phylogeny 113
6.5.2 Morphological variation and possible functional morphologies of structures
115
6.5.3 Comparison to spikes of other bivalve groups and implications for bivalve phylogeny
118
Chapter 7 - Conclusions 119
7.1 What can we learn from a unionoid shell? 1227.1.1 Systematics, phylogeny and evolution 124
7.1.1.1 “Gross shell morphology”: size and form 1247.1.1.2 Umbonal sculpture 1257.1.1.2 Periostracal microprojections 126
7.1.2 Ecology, conservation and environmental reconstruction 1267.1.2.1 “Gross shell morphology”: size, shape, inflation,
thickness, density and adductor scar sizes 127
7.1.2.2 Umbonal sculpture 1287.1.2.3 Periostracal microprojections 129
7.1.3 Determination of sex and trematode loads 1297.1.3.1 “Gross shell morphology”: size, shape, inflation and
thickness 129
7.2 Future directions 1307.2.1 Testing for consistency of the patterns observed 1317.2.2 Open questions 1317.2.3 Application of methods to non-unionoid taxa 132
Appendix 133
Table A.1 Umbonal sculpture types of unionoid taxa examined from shell material, photographs and/or drawings
135
Bibliography 149

ix
ACKNOWLEDGEMENTS
First and foremost I thank my supervisor David Aldridge for his guidance and
support over the past three and a half years. I am particularly grateful to him for giving me
the freedom to pursue my own scientific interests and for encouraging me throughout. My
secondary advisors Elizabeth (“Liz”) Harper and Richard Preece provided invaluable
additional malacological expertise and were always there when I needed them. Special
thanks go to Richard for showing me around the museum collection and to Liz for
spending hours with me on the SEM, teaching me about bivalve microstructure and life.
My subsistence costs during this PhD were predominantly covered by studentships
of the Austrian Federal Ministry of Science and Research and the Cambridge European
Trust. Additional financial support was kindly provided by the Ministry of Lower Austria,
the Siegfried-Ludwig Fund, the Balfour Fund and the St Catharine’s College. Genetic
analyses of mussel populations (Chapter 3) were funded by a research grant of the
Conchological Society of Great Britain & Ireland.
I could not have asked for a more pleasant and stimulating working environment
than the Aquatic Ecology Group. Thank you so much for all your help and support, both in
the field and in the office – Anna McIvor, Beccy Mant, Du Lina, Gawsia Chowdhury,
Holly Barclay, Line zu Ermgassen, Matthew Oreska and Nicole Spann. Numerous
additional people from the Department of Zoology and other Cambridge University
departments have dedicated some of their valuable time to teach me. Bill Amos, Joe
Hoffman and the whole Molecular Ecology Group gave me a “molecular” home for
several months and introduced me into AFLP analysis. John Parker offered much
appreciated advice on how to develop a model of character evolution. I also wish to thank
Andrea Manica for his statistical advice, Keith Gray for teaching me how to prepare shells
for SEM, André Sartori for his help in the field and inspiring bivalve discussions, our
technical assistants Ian Goldstone and Ben Taylor, our graduate secretary Linda Wheatley,
and Clair Castle and Jane Acred for being the most helpful librarians I have ever met.
Generous travel grants by the Malacological Society of London, the Freshwater
Mollusk Conservation Society and the St Catharine’s College gave me the opportunity to
present and discuss my work with scientists from other parts of the world. Some of these,
and in particular, Antonio Checa, Art Bogan and James Crampton have offered some

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
x
greatly appreciated expert’s advice and insight. Daniel Graf, Kevin Cummings and Keith
Walker aided in identification of tricky specimens.
Investigation of interspecific variation in shell morphology would not have been
possible without the generous donations of shell material by following people and
institutions: The Bivalve Tree-of-Life Project, Dan Hua (Freshwater Mollusk Conservation
Center, VA, USA), Nathan Eckert (Virginia Department of Game and Inland Fisheries,
VA, USA), Manuel Lopez-Lima (University of Porto), Dr. Ellinor Michel and Dr.
Jonathan Todd (both Natural History Museum, London), Keith Scriven (Maerdy and
Mawddach Hatchery) and Conor Wilson (Queen’s University Belfast).
I also wish to thank Abingdon Boat Marina, Thames & Kennet Marina, Harleyford
Estate & Marina, Racecourse Marina Windsor and Saxon Moorings, and all the friendly
people there for allowing access to the marina study sites.
Finally, I am extremely grateful to my parents Michael and Renate, and my whole
family for their love, support, letters, emails and visits. Many thanks also go to my friends
both at home and in Cambridge, who always knew how to put a smile on my face and
helped me through this whole process. Last but definitely not least, I want to thank my
partner Roman, who has been putting up with me for over a decade now, developed a
computer program called “schlexihexi” that saved me days of copy-paste, and knows
exactly when it is time to treat me with some chocolate and an episode of ‘Columbo’.

xi
SUMMARY
Freshwater mussels of the order Unionoida show a wide variability in shell features
but an understanding of which factors determine which trends in shell morphology is poor.
This thesis investigates inter- and intraspecific patterns in unionoid shell characters and
their potential use for reconstructing (1) environments, (2) characteristics of populations,
and (3) evolutionary trends and phylogenies.
Investigation of morphological patterns within three unionoid species from two
habitat types (marinas and river) of the River Thames, UK, elucidated consistent
ecophenotypic trends in maximum shell size, relative adductor size and shape of the dorso-
posterior shell margin. These shell characters may thus have broad ecological significance
and could have considerable utility to palaeontologists, taxonomists and conservation
biologists.
Molecular analyses using Amplified Fragment Length Polymorphisms suggested
that pronounced differences in shell morphology between populations of the same species
were caused by phenotypic plasticity. Observed genetic differences along the River
Thames, on the other hand, were consistent with a pattern of isolation by distance and
probably reflect limited dispersal via host fish species upon which unionoid larvae are
obligate parasites.
While relative shell width was a poor indicator of environment, this character was
significantly influenced by allometric growth, sexual dimorphism and trematode
parasitism. Detailed investigations on Anodonta anatina revealed that differences in the
degree of sexual dimorphism between populations may reflect the overarching effect of
habitat on morphology. In addition, other non-habitat related dimorphic patterns in sagittal
shape and thickness of shells were observed.
Interspecific morphological trends and their potential use for reconstructing
phylogenies were investigated with regard to two types of shell sculptures. First, a new
model of character evolution of umbonal sculptures in the Unionoida was developed by
examination of over 150 extinct and modern species. Second, investigation of shell
surfaces from specimens of all extant palaeoheterodont (unionoid + trigonioid) families
using scanning electron microscopy revealed the presence of three types of periostracal
microprojections. These possibly aid in the stabilisation and/or orientation of the mussel
within the sediment. Observations on both umbonal and periostracal sculptures indicated
considerable phylogenetic value of these two shell features.


xiii
GLOSSARY
Adductor muscle: One of usually two large muscles (one anterior, one posterior) that contract to close the shell and maintain it in that condition.
Alae (adj. alate): Anterior and/or posterior winglike projections of the valves that extend dorsally above the hinge line; also called wings; bivalves exhibiting alae are also called symphynote.
Allometric growth: The variation in the relative rates of growth of various parts of the body.
Annual ring: Compact line of temporarily arrested growth or rest period appearing on the shell surface as a raised or darker comarginal line; also called annulus.
Annulus (pl. annuli): See annual ring.
Anterior: Front or forward; head end.
Apomorphy (adj. apomorphic): A derived characteristic of a clade; any feature novel to a species and its descendants.
Beak: See umbo.
Beak sculptures: See umbonal sculptures.
Character state: The specific “value” taken by a character in a specific taxon (e.g. for character ‘colour’, character states ‘red’, ‘green’…).
Clade: A monophyletic group of two or more species.
Comarginal: Parallel to the shell margin; also sometimes referred to as concentric.
Concentric: See comarginal.
Crown group: A group consisting of living representatives, and their ancestors, back to the most recent common ancestor of that group.
Convergence (adj. convergent): Independent evolution of similar traits in unrelated lineages; also called homoplasy.
Corrugated: Marked by wrinkles or ridges and grooves.
Ctenidium: A thin, platelike paired organ within the mantle cavity which serves as an organ of respiration and food-gathering in unionoids, and at least partly, as marsupium in female unionoids; usually suspended by tissue or cilial junctions from the dorsal region of the animal and typically comprising a W-shaped doubly folded lamella on each side of the visceral mass; each side typically consisting of an inner and outer demibranch.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
xiv
Demibranch: See ctenidium.
Deposit feeding: Feeding type during which organic particles are harvested from sediments; see also suspension feeding.
Divaricate: Branching, usually in reference to external sculpture.
Distal: Situated away from point of attachment or origin; terminal end.
Dorsal: The hinge side of a bivalve; opposite of ventral.
Edentulous: Without hinge teeth.
Eutrophic: Nutrient-enriched and consequently highly productive (water body).
Exhalant siphon/aperture: Aperture or siphon that controls water outflow from the mantle cavity.
Filter feeding: Feeding type involving the filtering of organic particles from water by the gills, after which appropriately sized particles are transported to the mouth; see also suspension feeding, deposit feeding.
Foot: Muscular organ at the ventral part of the visceral mass; used by contraction and expansion for locomotion, burrowing and/or anchoring a bivalve.
Gill: See ctendidium.
Glochidium (pl. glochidia): The bivalve larvae of freshwater mussels in the superfamily Unionoidea which are generally parasitic on fish.
Hermaphrodite (adj. hermaphroditic): Sexually mature animal in which male and female gametes are produced by the same individual, either simultaneously or in sequence; see also protandric and protogynous.
Hinge: Dorsal border of the articulated valves, including the ligament, hinge teeth, and other structures that function to permanently unite the two valves; also called hinge line.
Hinge dentition: See hinge teeth.
Hinge line: See hinge.
Hinge teeth: A series of calcified dorsal interlocking teeth and sockets that allow alignment of the valves to be maintained during opening and closing.
Homology (adj. homologous): Similarity between characteristics of organisms that is due to their shared ancestry.
Homoplasy (adj. homoplasic): See convergence.
Infauna (adj. infaunal): Organisms which live in soft sediment and are large enough to displace sediment.

Glossary
xv
Inhalant siphon/aperture: Aperture or siphon, usually posterior, that controls water intake into the mantle cavity.
Interstitium (adj. interstitial): Pore spaces between the grains of rock and sediments.
Lateral: Situated at or extending to the side.
Lentic: Standing water environment.
Ligament: Elastic structure that connects the two bivalve shells at the hinge line and functions as a spring to open the valves when the adductor muscles relax.
Lotic: Running water environment.
Mantle: Fleshy outer tissue surrounding the organs of a molluscan body and secreting the periostracum and shell; consisting of two lobes in a bivalve, one lining each shell, and at the ventral edge several folds that may have different functions or features.
Mantle cavity: Chamber between the mantle lobes and interior visceral mass.
Marsupium (pl. marsupia): In unionoids, a brood pouch for eggs and developing glochidia, formed by the complete ctenidia or a restricted portion of the ctenidia.
Monophyletic: Containing a common ancestor and all of its descendants (pertaining to a taxonomic group); defined by synapomorphies.
Muscle scar: Impression on the shell interior that indicates the attachment position of a muscle.
Nonindigenous: Not native to an area.
Obesity: See relative shell width.
Ontogeny (adj. ontogenetic): The developmental history of an organism.
Oocyte: Immature egg cell.
Oogonia: Earliest recognizable form of the egg.
Palaeoenvironment: The environment of a former period of geologic time.
Pedal: By means of the foot.
Periostracum: Outermost layer of the shell; secreted by the mantle.
Plesiomorphy (adj. plesiomorphic): Ancestral state of a character in an evolutionary analysis.
Polyphyletic: Taxon that does not contain the most recent common ancestor of its members.
Posterior: Hind or rear; anal end.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
xvi
Prismatic: Shell microstructural variety consisting of parallel columnar prisms of calcium carbonate.
Protandric/protandrous: A form of hermaphroditism in which the male phase precedes the female phase during the life cycle of the same individual; see also hermaphrodite.
Protogynous: Condition in a sequential hermaphrodite in which female gonads mature before male gonads.
Radial: In this thesis referring to external sculptural features that originate at a central point at the umbo and fan outward toward the margins.
Random genetic drift: The process of change in the genetic composition of a population due to chance or random events rather than by natural selection.
Relative shell length: Degree of elongation across the antero-posterior axis.
Relative shell width: Degree of inflation across the lateral shell axis.
Rib: An elongated sculptural element that is raised above the surrounding shell surface; also called ridge.
Ridge: See rib.
Riverine: Characteristic of rivers.
Rugae: See umbonal sculptures.
Sagittal: A sagittal plane is an imaginary plane that travels vertically from the top to the bottom of the body, dividing it into left and right portions.
Sculpture: Ornament or markings on the shell surface.
Semi-infaunal: Partially infaunal.
Sexual dimorphism: Condition in which males and females of the same species are morphologically different.
Shell height: Maximum sagittal shell diameter perpendicular to shell length (usually across the dorso-ventral axis).
Shell length: Maximum sagittal shell diameter (usually across the antero-posterior axis).
Shell width: Maximum lateral shell diameter.
Siphon: Posterior extension (usually two) of the mantle, made tubular by either tissue fusion or ciliary junctions of the mantle folds, through which water, waste products and gametes are directed in and out of the body.
Sister clades: Taxa with a common ancestor and no additional descendents.
Spermatozoa: The male reproductive cell.

Glossary
xvii
Suspension feeding: Feeding type during which organic particles are harvested from the water column.
Symphynote: See alate.
Synapomorphy (adj. synapomorphic): A shared, derived, taxon-defining trait or characteristic.
Teeth: See hinge teeth.
Transverse: Situated or lying across; crosswise.
Umbo (pl. umbos or umbones; adj. umbonal): The raised portion of the dorsal margin of a shell that reflects the early growth stage (= oldest part of the shell); formed by the embryonic shell around which the rest of the shell develops distally in a concentric manner; also called beak.
Umbonal sculptures: Sculptures on the umbo; also called beak sculptures or rugae.
Valve: The right or left half of a bivalve shell.
Ventral: The underside or foot-side of a bivalve; opposite of dorsal.
Visceral mass: Region of the bivalve body containing most of the digestive, excretory, circulatory, and nervous systems, that is suspended dorsally between the gills and that usually terminates ventrally as the foot.
Wings: See alae.


CHAPTER 1
INTRODUCTION

3
CHAPTER 1
INTRODUCTION
Freshwater mussels of the order Unionoida (also known as pearly mussels or naiads)
show an extremely wide range in shell characters. The notable differences between
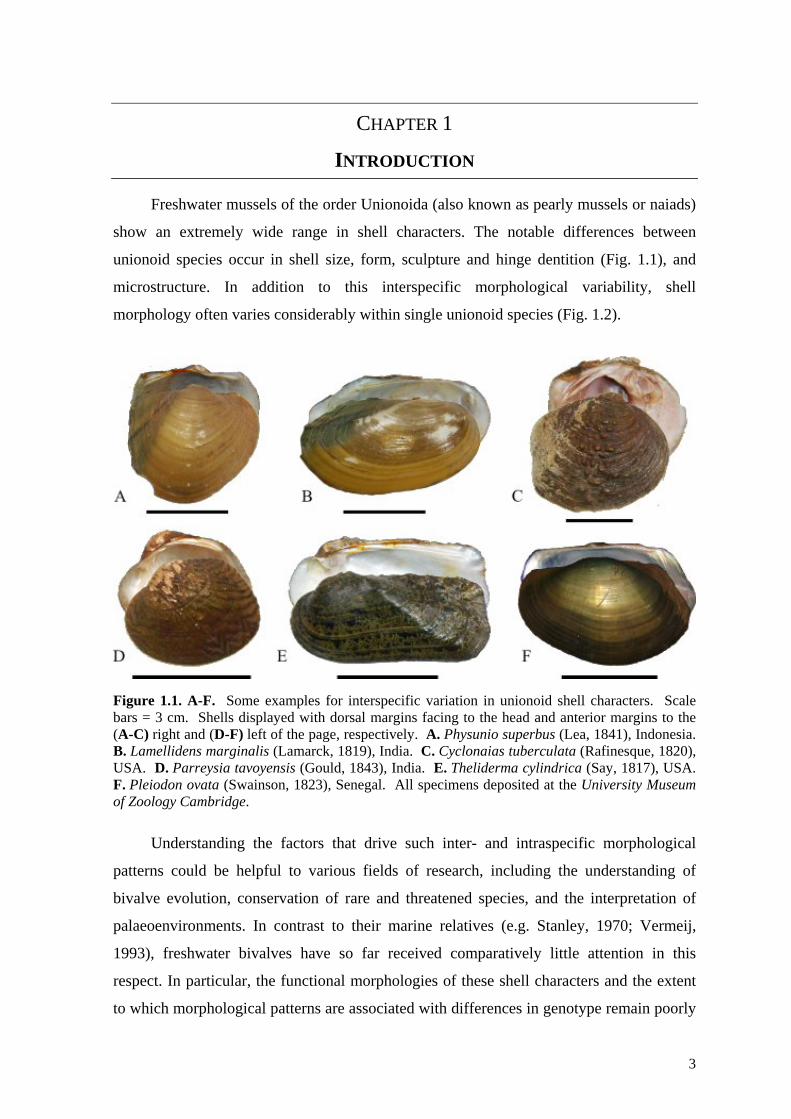
unionoid species occur in shell size, form, sculpture and hinge dentition (Fig. 1.1), and
microstructure. In addition to this interspecific morphological variability, shell
morphology often varies considerably within single unionoid species (Fig. 1.2).
Figure 1.1. A-F. Some examples for interspecific variation in unionoid shell characters. Scale bars = 3 cm. Shells displayed with dorsal margins facing to the head and anterior margins to the (A-C) right and (D-F) left of the page, respectively. A. Physunio superbus (Lea, 1841), Indonesia. B. Lamellidens marginalis (Lamarck, 1819), India. C. Cyclonaias tuberculata (Rafinesque, 1820), USA. D. Parreysia tavoyensis (Gould, 1843), India. E. Theliderma cylindrica (Say, 1817), USA. F. Pleiodon ovata (Swainson, 1823), Senegal. All specimens deposited at the University Museum of Zoology Cambridge.
Understanding the factors that drive such inter- and intraspecific morphological
patterns could be helpful to various fields of research, including the understanding of
bivalve evolution, conservation of rare and threatened species, and the interpretation of
palaeoenvironments. In contrast to their marine relatives (e.g. Stanley, 1970; Vermeij,
1993), freshwater bivalves have so far received comparatively little attention in this
respect. In particular, the functional morphologies of these shell characters and the extent
to which morphological patterns are associated with differences in genotype remain poorly

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
4
understood. It is the aim of this thesis to fill some of these gaps in our understanding of
unionoid shell morphologies and the mechanisms involved in determining interspecific
and intraspecific variation.
Figure 1.2. A-I. Some examples for intraspecific variation in shell shape of Unio pictorum (Linnaeus, 1758) in the UK. Scale bars = 3 cm. Left valves displayed with anterior margins facing to the left and dorsal margin to the head of the page. A. Cut-off Channel near Hilgay, Norfolk. B. River Meden, Nottinghamshire. C. Lake Llangorse, Wales (NMW.1920.393.001). D. Welford Reservoir, Northamptonshire. E. Canal near Bath, Somerset. F. Kennet and Avon Canal, Somerset. G. Wateringbury Stream, Kent. H. River Lee, Essex. I. River Frome, Dorset (NMW.1955.158). Specimens from (A) private collection, (B,D,E,G,H) the Natural History Museum London, (C,I) the National Museum of Wales and (F) the University Museum of Zoology Cambridge.
1.1. A BRIEF INTRODUCTION TO THE UNIQUENESS OF UNIONOIDS
1.1.1. UNIONOID PHYLOGENY, DIVERSITY AND DISTRIBUTION
Unionoids are filter feeding freshwater bivalves with a distribution spanning all
continents except Antarctica. Besides their wide spatial distribution, this group of
freshwater mussels is also an ancient clade, with the oldest known representatives of their
extant crown group dating from the Triassic (Haas, 1969b; Watters, 2001). Though the
phylogenetic origin of the Unionoida is currently unknown, they possibly descended from

Chapter 1 - Introduction
5
ancient members of the marine Trigonioida (Newell & Boyd, 1975; Giribet & Wheeler,
2002).
Current unionoid diversity is still poorly recorded but latest global estimates range
from about 800 species (Bogan, 2008; Bogan & Roe, 2008) to approximately 900 species
(Graf & Cummings, 2006b, 2007b). Based on phylogenetic analyses by Graf & Cummings
(2006b; Fig. 1.3A) using combined morphology + mitochondrial (COI) DNA + nuclear
ribosomal (28S) DNA data, Graf & Cummings (2007b) represents the most recent attempt
of a global checklist of freshwater mussel species. These authors list a total of 840
described unionoid species, which they classify into 166 genera, six families and two
superfamilies (Table 1.1). The large number of incertae sedis taxa (i.s., i.e. "of uncertain
placement”) in Graf & Cummings’ (2007b) classification reflects our incomplete
understanding of unionoid systematics and phylogeny, which is further evidenced by
contradictory phylogenetic cladograms obtained by other authors. For example, analyses
based on mitochondrial (COI) DNA (Bogan & Hoeh, 2000) and combined mitochondrial
(COI) DNA + morphology data (Hoeh, Bogan & Heard, 2001) (Fig. 1.3B) recovered a
polyphyletic status of both the “Unionidae” and the freshwater oysters (“Etheriidae”). The
respective positions of the Hyriidae and Margaritiferidae within the Unionoida are further
issues currently under intense scientific debate (see Graf & Cummings, 2006b; Hoeh et al.,
2009; Graf & Cummings, 2010a).
1according to Graf & Cummings (2006b, 2007b) member of the Etheriidae
Figure 1.3. A-B. Two contrasting phylogenetic cladograms of the Unionoida (both redrawn from Hoeh et al. (2009)). A. Combined evidence phylogeny based on morphology + mtDNA (COI) + nuclear ribosomal DNA (28S) data (Graf & Cummings, 2006b: fig. 4). B. Combined summary tree based on mtDNA (COI) (Bogan & Hoeh, 2000) and morphology + mtDNA (COI) (Hoeh, Bogan & Heard, 2001).

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
6
Table 1.1. Unionoid systematics after Graf & Cummings (2007b).
Superfamily UNIONOIDEA Genera
Family UNIONIDAE Subfam. UNIONINAE
Tribe UNIONINI Acuticosta, Arconaia, Cafferia, Cuneopsis, Inversiunio, Lanceolaria, Lepidodesma, Nodularia, Rhombuniopsis, Schistodesmus, Unio
Tribe ANODONTINI Alasmidonta, Anemina, Anodonta, Anodontoides, Arcidens, Cristaria, Lasmigona, Pegias, Pseudanodonta, Pyganodon, Simpsonaias, Simpsonella, Sinanodonta, Strophitus, Utterbackia
Subfam. AMBLEMINAE Tribe AMBLEMINI Amblema Tribe LAMPSILINI Actinonaias, Arotonaias, Cyprogenia, Cyrtonaias,
Delphinonaias, Disconaias, Dromus, Ellipsaria, Epioblasma, Friersonia, Glebula, Hamiota, Lampsilis, Lemiox, Leptodea, Ligumia, Medionidus, Obliquaria, Obovaria, Potamilus, Ptychobranchus, Toxolasma, Truncilla, Venustaconcha, Villosa
Tribe PLEUROBEMINI Cyclonaias, Elliptio, Elliptoideus, Fusconaia, Hemistena, Lexingtonia, Plethobasus, Pleurobema, Popenaias, Uniomerus
Tribe QUADRULINI Amphinaias, Megalonaias, Plectomerus, Quadrula, Quincuncina, Theliderma, Tritogonia
Tribe GONIDEINI Gonidea i. s. Mesoamerican Ambleminae Barynaias, Martensnaias, Micronaias, Nephritica, Nephronaias,
Pachynaias, Psoronaias, Psorula, Reticulatus, Sphenonaias i.s. UNIONIDAE
Western Palearctic: Leguminaia, Microcondylaea, Potomida, Pseudodontopsis Afrotropical: Brazzaea, Coelatura, Grandidieria, Mweruella, Nitia,
Nyassunio, Prisodontopsis, Pseudospatha Indotropical: Arcidopsis, Caudiculatus, Chamberlainia, Contradens,
Ctenodesma, Diaurora, Discomya, Elongaria, Ensidens, Haasodonta, Harmandia, Hyriopsis, Inversidens, Lamellidens, Lamprotula, Modellnaia, Oxynaia, Parreysia, Physunio, Pilsbryoconcha, Pressidens, Prohyriopsis, Protunio, Pseudobaphia, Pseudodon, Ptychorhynchus, Rectidens, Scabies, Schepmania, Solenaia, Trapezoideus, Unionetta
Family MARGARITIFERIDAE Margaritifera
Superfamily ETHERIOIDEA
Family HYRIIDAE Subfam. HYRIINAE
Tribe HYRIINI Prisodon Tribe CASTALIINI Callonaia, Castalia, Castaliella Tribe RHIPIDODONTINI Diplodon, Rhipidodonta Tribe HYRIDELLINI Cucumerunio, Echyridella, Hyridella, Virgus
Subfam. VELESUNIONINAE Alathyria, Lortiella, Microdontia, Velesunio, Westralunio
Family ETHERIIDAE Acostaea, Bartlettia, Etheria, Pseudomulleria
Family MYCETOPODIDAE Subfam. MYCETOPODINAE Mycetopodella, Mycetopoda Subfam. ANODONTITINAE Anodontites, Lamproscapha Subfam. LEILINAE Leila Subfam. MONOCONDYLAEINAE Diplodontites, Fossula, Haasica, Iheringella, Monocondylaea,
Tamsiella
Family IRIDINIDAE Subfam. IRIDININAE Chelidonopsis, Mutela, Pleiodon Subfam. ASPATHARIINAE Aspatharia, Chambardia, Moncetia

Chapter 1 - Introduction
7
1.1.2. IMPORTANCE AND CONSERVATION STATUS
Our insufficient understanding of unionoid evolution and diversity is particularly
unfortunate given that freshwater mussels are amongst the most critically endangered
groups of animals worldwide (Lydeard et al., 2004; Strayer, 2006). In fact, in North
America alone, about 36 unionoid species (10%) are already presumed extinct (Neves et
al., 1997). Most important causes for their decline include habitat loss and degradation,
loss of host fishes (see 1.1.3) and the introduction of nonindigenous species (Bogan, 1993;
Williams et al., 1993; Neves et al., 1997). As a result, two of the six unionoid species
inhabiting freshwater habitats in the UK, are considered to be a national conservation
priority, appearing on the UK Biodiversity Action Plan priority list (Aldridge, 2004).
At the same time, Unionoida have important ecosystem functions such as particle
filtration and processing, nutrient release and sediment mixing (Vaughn & Hakenkamp,
2001). As a consequence, freshwater mussels are increasingly recognised as keystone
fauna (Aldridge, Fayle & Jackson, 2007), and their decline can profoundly affect
ecosystem processes in aquatic habitats (Geist & Auerswald, 2007). In addition, unionoids
are used by people, both as a food source and for pearl production (Anthony & Downing,
2001). Finally, these shell-bearing organisms can be used as environmental indicators,
thereby providing critical data for monitoring habitat changes (e.g. Mutvei & Westermark,
2001).
1.1.3. LIFE CYCLE AND MORPHOLOGICAL VARIATION
Besides a practical interest due to their conservation status, and ecological and
economic importance, freshwater mussels are particularly interesting from a biological
perspective. Unionoids exhibit a unique life cycle incorporating both parental care (i.e.
brooding) and, in most species, obligate larval parasitism upon freshwater fishes (Kat,
1984; Wächtler, Mansur & Richter, 2001) (Fig. 1.4). The parasitic stage, lasting from a
few weeks to several months, is believed to have evolved as a means of distribution of
these otherwise relatively immobile animals (Kat, 1984). As a further consequence of their
hosts’ mobility, however, habitat conditions in which the juvenile mussel excysts from the
host will be rendered highly unpredictable. This unpredictability of the “new” habitat(s) of
a given mussel’s offspring is believed to be a major source of the wide morphological
variability characteristic of many unionoid species.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
8
Figure 1.4. A-E. The unionoid life cycle. A. Schematic drawing of a typical unionoid life cycle. Sperm released by an adult male via his exhalant siphon is infiltrated by the female through her inhalant siphon. In the female gills (marsupia), fertilised eggs develop into mature larvae (glochidia), which are released into the water column. Glochidia attach and encyst into the gills, fins or scales of freshwater fishes. After this parasitic phase, the then fully transformed juvenile mussel drops off the fish and begins its life in the sediment. B. Glochidial shell of Anodonta anatina (Linnaeus, 1758). C. Teeth on the external surface of the glochidial hook in A. anatina. D. Glochidia of Sinanodonta woodiana (Lea, 1834) encysted into fins of the Golden Line Fish (Sinocyclocheilus grahami Regan, 1904) (photograph by Holly Barclay); shell lengths approx. 330 µm. E. 2 year-old juvenile of Margaritifera margaritifera (Linnaeus, 1758); shell length approx. 2 mm.
1.2. IMPORTANCE OF UNDERSTANDING MORPHOLOGICAL PATTERNS IN UNIONOID SHELLS
The morphological makeup (phenotype) of a given mussel is a result of two main
mechanisms: (1) its genetic composition (genotype) and (2) environmental or other non-
genetic factors triggering changes in morphology.
On the species and higher taxonomic levels, ‘genetic inheritance’ results in similar
traits of related taxa due to common ancestry (‘homologies’). Similarities in life habit,
environmental conditions and/or other factors, on the other hand, can lead to independent

Chapter 1 - Introduction
9
evolution of similar morphological solutions (‘homoplasies’ or ‘convergences’) in
unrelated taxa.
Equally, morphological differences within single species can be a result of either
differences in the genotype or due to plasticity of the phenotype (Via et al., 1995).
Intraspecific patterns in shell morphology lacking a genetic basis may directly or indirectly
be caused by differences in, for example, sex, levels of parasitic infestation or
environmental conditions. The ‘habitat’ factor is regarded of particular importance to
intraspecific patterns in unionoid shell morphologies. However, though such intraspecific
ecomorphotypes may simply reflect phenotypic plasticity, these could also be a result of
selective survival of only those genotypes that are most adequately equipped for the
respective environmental conditions. The unique unionoid life cycle potentially allows
both natural selection and phenotypic plasticity to operate, but the relative importance of
these two mechanisms is currently unknown.
In conclusion, shell morphology may reflect both the (phylo)genetic history and
other characteristics, such as life habit and habitat, of a given species, population or
individual. Whereas a morphological characteristic that has predominantly been influenced
by inheritance can tell us about a mussel’s genetic composition and/or phylogenetic
position, morphological convergences between or within species can contain information
on the character’s function. An accurate understanding of inter- and intraspecific patterns
in shell morphology can thus be valuable for various fields of research, some of which are
discussed in the following.
1.2.1. SYSTEMATICS, PHYLOGENY AND EVOLUTION
Early unionoid taxonomists (e.g. Lea, 1870; Simpson, 1900; Simpson, 1914; Modell,
1942) based their classifications predominantly on shell traits and, in particular, shell form,
sculpture and hinge dentition. By including anatomical, life history and molecular
characters in their phylogenetic analyses, other and subsequent authors (e.g. Ortmann,
1912; Prashad, 1931; Haas, 1969a, b; Davis, 1983, 1984), however, recognised widespread
convergences in shell morphologies of unrelated unionoid taxa. In addition, the large
extent of intraspecific variation, most notably in shell form and sculpture, has in the past
led to considerable synonymy (e.g. Rossmässler, 1835-1837; Küster, 1848; Locard, 1890)
and still causes taxonomic confusion. The high degree of interspecific convergence and

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
10
intraspecific variability renders these shell characters unsuitable for addressing questions
regarding unionoid taxonomy and phylogeny.
The hard part of unionoid and other bivalves generally represents, on the other hand,
the only source of information available for the study of fossil taxa. A good understanding
of present homologies, convergences and the extent of intraspecific variation in shell
morphologies is therefore a prerequisite for resolving ancient freshwater mussels’
systematics and phylogeny. Knowledge on the evolution of a given shell character, and
thereby, the morphological characteristics of basal (plesiomorphic) and derived
(apomorphic) character states, can be particularly useful in this respect. At the same time,
this information can help us answer unresolved questions regarding modern unionoids’
evolution.
1.2.2. ECOLOGY, CONSERVATION AND ENVIRONMENTAL RECONSTRUCTION
Identifying habitat characteristics associated with particular morphological traits of a
given unionoid species, population or individual, can provide valuable information about
its ecology. This knowledge can in further course help identify habitat requirements of
endangered species and assist in their management. In addition, comparing
ecomorphotypes of recent shells with similar fossilised ones can enable the reconstruction
of palaeoenvironments. The application of unionoids to these purposes has so far been
limited due to a lack of an adequate understanding of which shell morphologies are
triggered by which environmental conditions.
At species and higher taxonomic levels, certain convergences in shell morphologies
of unrelated taxa sharing particular ecological characteristics, are relatively well
understood. For example, ‘symphynote’ species, characterised by a dorsal, “wing-shaped”
extension of the shell, termed ‘alae’ or ‘wings’, can typically be found in standing/slow-
flowing, fine sediment water bodies. Alae have independently arisen several times within
the Unionoida, are believed to prevent sinking into soft substrates and are commonly
associated with a laterally compressed form, reduced dentition and a lack of sculpture of
the shell. Unionoid species with well developed shell sculpture (such as macroscopic
‘ridges’ or ‘pustules’), on the other hand, are often associated with fast-flowing habitats
(Watters, 1994).
Whereas identification of interspecific convergences requires knowledge of the
extent of genetic inheritance of the respective shell trait, this does not apply to consistent

Chapter 1 - Introduction
11
ecophenotypic trends within a species. The often extreme degree of intraspecific variation
in unionoid shell morphology could thus be particularly valuable for the aforementioned
fields of research. Despite the considerable amount of literature on habitat-shell
morphology associations within unionoid species (e.g. Wetherby, 1882; Ortmann, 1920;
Agrell, 1948), most of these studies suffer from one or more caveats, including (1) a lack
of statistic evidence, and (2) failure to consider influence of non-habitat factors that might
influence shell morphology.
1.2.3. RECONSTRUCTION OF POPULATION PARAMETERS
In addition to those concerning a mussel’s environmental surroundings, other factors
can influence the morphology of a given specimen (Seed, 1980). These include (1) sexual
dimorphism, (2) allometric growth during ontogeny, and (3) morphological alteration due
to parasitic infestation (Roper & Hickey, 1994; Scholz, 2003). While it is crucial to
consider the relative influence of such non-habitat factors when attempting to elucidate
ecophenotypic patterns, knowledge on how morphology changes with various parameters
can also be helpful in its own right. Identification of sexual shell dimorphisms can, for
example, facilitate quick sex determination in the field and/or the reconstruction of ancient
populations’ sex ratios.
The importance of the respective non-habitat and habitat factors on particular
morphological trends in freshwater mussel species are still poorly understood. Sexual shell
dimorphism in Unionoida is known predominantly from the North American tribe
Lampsilini, whereas patterns in European groups remain unresolved. The extent to which
allometric growth and/or parasitic infestation influence morphology within unionoid
species remains almost completely unresolved.
1.3. AIMS AND QUESTIONS ADDRESSED IN THIS THESIS
The aim of this thesis is to improve our understanding of the factors that determine
inter- and intraspecific patterns in unionoid shell morphology. Such an understanding can
ultimately be used in the reconstruction of evolutionary trends and phylogenies,
environments and habitat requirements of species, and other characteristics of individuals
and populations such as their sex ratios or parasitic loads. Broadly, three types of shell
characters were studied: (1) gross shell morphology (i.e. size, shape, inflation and
thickness), (2) umbonal (‘beak’) sculpture, and (3) periostracal microsculpture.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
12
1.3.1. “GROSS SHELL MORPHOLOGY”: SIZE, SHAPE, INFLATION AND THICKNESS
Due to a high degree of interspecific convergences and intraspecific variability, the
shell characters of ‘size’ (at a given age), ‘shape’, ‘inflation’ and ‘thickness’ are
considered to be of poor taxonomic and phylogenetic value. However, an accurate
understanding of which factors trigger such trends within a given species could yield
considerable information.
Our lack of knowledge in this respect is reflected by conflicting observations of
habitat-morphology associations reported by previous authors. In particular, descriptions
of how shell form changes with certain habitat parameters and consequentially, the
functional explanations given, are often contradictory. Reasons for such inconsistencies
may include inappropriate measurements of habitat, failure to consider influence of non-
habitat factors and random genetic drift, and inadequate choice of morphological features.
Applying traditional and modern morphometric techniques, Chapter 2 investigates
intraspecific trends in gross shell morphology and internal characters within three
European unionoid species. The main objectives were to (1) detect consistent, truly
ecophenotypic intraspecific trends in unionoid shell shape, (2) determine the importance of
various habitat and non-habitat factors on an individual’s morphology, (3) elucidate
internal morphological characteristics associated with certain changes in shell morphology,
and (4) discuss probable functional advantages of shell ecomorphotypes found.
In Chapter 3, I use Amplified Fragment Length Polymorphisms (AFLPs) to ask
whether intraspecific morphotypes identified in Chapter 2 are due to genetic differences or
phenotypic plasticity. Chapter 4 investigates the importance of non-habitat parameters in
determining intraspecific trends in shell morphology. In particular, the roles of sexual and
parasite-induced dimorphism are investigated.
1.3.2. UMBONAL SCULPTURE
Sculptures restricted to the early shell region (‘umbonal sculptures’, ‘beak
sculptures’ or ‘rugae’) have long been used for identification of unionoid species and,
occasionally, genera. Specificity of this shell character at higher taxonomic levels and
consequently, its phylogenetic value and adequacy for classification of fossil taxa, is still
under considerable scientific debate. To a large part, this is due to a lack of understanding
of unionoid beak sculpture character evolution. Knowledge on synapomorphic,
plesiomorphic and convergent character states can help address unresolved questions on

Chapter 1 - Introduction
13
unionoid phylogeny and systematics, but might additionally help formulate likely
functional hypotheses of this shell character. Ultimately, this could give novel insights on
ecological aspects of this poorly understood, but crucial life stage.
Based on examination of over 150 ancient and modern species, Chapter 5 discusses
patterns of interspecific variability and presents a new model of character evolution of
umbonal sculptures in the Unionoida.
1.3.3. PERIOSTRACAL MICROPROJECTIONS
Periostracal microprojections, observable only by high magnification (electron)
microscopy, have been little studied in palaeoheterodont (i.e. unionoid and trigonioid)
bivalves. However, recent studies on other bivalve groups (Glover & Taylor, 2010; Checa
& Harper, in press) indicate that these structures are probably of considerable phylogenetic
value. In Chapter 6, I analysed specimens covering all six unionoid families and the only
extant genus of the Trigonioida. This enabled me to provide the first comprehensive
review of periostracal microprojections in the Palaeoheterodonta, and to discuss their
possible functional morphologies and importance in phylogenetic reconstruction.
1.4. STYLE OF THESIS
Chapters 2-6 are written in paper-style and slightly altered versions are either
published (Zieritz & Aldridge, 2009; Zieritz et al., 2010; Zieritz et al., in press) or
currently under review (Zieritz & Aldridge, in review; Zieritz, Bogan & Aldridge, in
review) by a peer-reviewed scientific journal.


CHAPTER 2
IDENTIFICATION OF ECOPHENOTYPIC TRENDS WITHIN THREE EUROPEAN
FRESHWATER MUSSEL SPECIES (BIVALVIA:UNIONOIDA) USING TRADITIONAL AND
MODERN MORPHOMETRIC TECHNIQUES

“Rafinesque collected the Unionidae extensively in Kentucky and
published a large number of genera, minor groups, and
species…His figures are more like those made by children, or the
caricatures drawn by aboriginal tribes, than the creations of an
intelligent naturalist…”
Simpson (1900)

17
CHAPTER 2
IDENTIFICATION OF ECOPHENOTYPIC TRENDS WITHIN THREE EUROPEAN FRESHWATER MUSSEL SPECIES (BIVALVIA:
UNIONOIDA) USING TRADITIONAL AND MODERN MORPHOMETRIC TECHNIQUES
2.1. ABSTRACT
Most species of freshwater mussels (Unionoida) show a wide variability in shell
form and size but an understanding of which factors determine unionoid morphology is
poor. We identified ecophenotypic trends in shell and internal characters within three
unionoid species from two habitat types (marinas and river) of the River Thames, UK,
using traditional and modern morphometric techniques. In marinas, all species grew to
larger maximum sizes than in the river, which might be a result of higher temperatures and
phytoplankton densities in marinas. Unio pictorum in marinas was more elongated than in
the river and Fourier shape analysis revealed a trend from dorsally arched river specimens
to straight dorsal and pointed posterior margins in marina individuals. The degree of shell
elongation and shape of dorso-posterior margin were not associated with sediment
composition, but were associated with the different hydrological characters of the two
habitat types. Relative shell width was a poor indicator of collection site and influenced by
allometric growth. Unlike U. pictorum, a difference in shell elongation of marina and river
mussels could not be detected in Unio tumidus and Anodonta anatina. However, all three
species showed the same trends regarding the shape of the dorso-posterior shell margin.
This shell character may thus have broad ecological significance and could have
considerable utility to palaeontologists, taxonomists and conservation biologists.
2.2. INTRODUCTION
Freshwater mussels of the order Unionoida display a wide range of intraspecific
morphological variability (Fig. 1.2), which has in the past led to considerable synonymy
(Küster, 1848; Lea, 1870; Locard, 1890). Besides the obvious value for taxonomic
research, identifying habitat characteristics associated with given phenotypes of a species
can provide valuable information about its ecology, assist in the management of

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
18
endangered species (Bogan, 1993), and enable the reconstruction of palaeoenvironments.
In addition to those concerning a mussel’s habitat, other factors such as its sex or
ontogenetic stage might influence the morphology of a given specimen (Seed, 1980;
Tevesz & Carter, 1980). However, the importance of the respective non-habitat and habitat
factors on particular morphological trends in freshwater mussel species and the
mechanisms involved are still poorly understood.
The relationship between morphotype and habitat has been studied in unionoids for
over 100 years (Hazay, 1881; Buchner, 1910; Israel, 1910; Haas & Schwarz, 1913;
Ortmann, 1920; Grier & Mueller, 1926; Bloomer, 1938). However, such descriptions,
usually based on measurements of the three shell dimensions (length, height and width),
are often contradictory. For example, in their study on 18 unionoid species at the
Mississippi River system, USA, Grier & Mueller (1926) found that in a slow flowing, fine
sediment river-lake, some species displayed more elongated shells (decreased shell height
to length ratio) than in the main channel, whereas others showed the exact opposite
pattern. Sometimes even populations within the same species are apparently “reacting” in
an opposing manner. Cvancara (1972) found Lampsilis radiata (Gmelin, 1791) to grow
relatively longer shells in finer substrates of Long Lake, Minnesota, whereas in Lake Erie,
Hinch, Bailey & Green (1986) observed shells of the same species to be more elongated in
sand than in mud.
Three factors may explain the apparent contradictions in the relationship between
shell form and habitat. First, the characters under study might simply not be associated
with habitat. For example, unionoids are known to switch from an interstitial deposit
feeding to a suspension feeding mode of life at a certain point in their early life (Yeager,
Cherry & Neves, 1994), which might be accompanied by a change in allometric growth.
Other non-habitat factors potentially influencing growth and morphology in unionoids are
a shift of metabolism at sexual maturity, sexual dimorphism or genetic differences of
geographically distant populations possibly caused by random genetic drift. Although
infestation by certain parasites has shown to induce abnormal shell growth in New Zealand
unionoids (Roper & Hickey, 1994), no such growth-altering parasites are currently known
from European waters (but see Chapter 4). Second, inappropriate measurements of habitat
may have been made with the key environmental determinant being overlooked. Finally,
the choice of morphological features (typically the three shell dimensions) may not
provide sufficient description of morphology. Ecologically more significant trends are
likely to be elucidated by analysing more accurate morphological descriptions, such as the

Chapter 2 – Ecophenotypic trends in freshwater mussels
19
whole outline of a shell. Statistical analysis of the degree of difference of, for example,
sagittal shell outlines has been made feasible only recently by the development of
advanced morphometric methods such as Fourier shape analysis. These tools are widely
used by palaeontologists but, unfortunately, have found only sporadic application in
ecological studies on recent freshwater unionoids (Scholz, 2003).
Intraspecific shell morphotypes that are consistently correlated to particular habitat
conditions could potentially be adaptations to the same. For example, short shells might be
advantageous under high current velocities. Alternatively, a given shell shape might
simply be a result of certain adaptive internal characteristics of the respective mussel
(Kauffman, 1969; Stanley, 1970); for example, it might be that, in a given species, larger
feet are advantageous in providing better anchorage in fast flows; this could result in
relatively large anterior parts of the shell. Thus, detecting changes in internal characters
associated with those in shell forms could help explain why certain shell forms are present
in certain habitats.
In the present study, we combined an ecological study design (i.e. five replicate
paired sites) with traditional and advanced morphometric techniques aiming to statistically
estimate morphological patterns within three European unionoid species. The important
study aims were to: (1) compare traditional and advanced morphometric methods in
detecting morphological patterns in the species; (2) elucidate internal morphological
characteristics associated with certain changes in shell morphology; (3) determine the
importance of various habitat factors (i.e. sediment, water movement, temperature, food
availability) and non-habitat factors (i.e. ontogenetic factors (age and size), sex,
geographic distance of populations) on an individual’s morphology; and (4) estimate
consistence of morphological patterns over replicates and species.
2.3. MATERIALS AND METHODS
2.3.1. SAMPLING OF INDIVIDUALS
From May to September 2007, freshwater mussel populations at five marina and five
adjacent river sites of the River Thames from Abingdon to Old Windsor were surveyed
(Fig. 2.1). Marinas were directly connected to the river, and paired sampling sites were no
more than 800 m apart. Marinas were characterised by areas in the approximate range 10-
80 ha and maximum water depths in the approximate range 1.5-4 m, and were constructed
at least 30 years ago (i.e. longer than the longevity of any of the mussel species studied;

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
20
Aldridge (1999b)). The River Thames in the stretch studied has an average width of 50 m,
and flow is regulated by a series of weirs, such that mean summer and winter discharge
values are approximately 40 m³s-1 and 80 m³s-1 (data from the UK Environment Agency),
respectively .
Figure 2.1. Sampling area and the five sampling locations at the River Thames. At each location one marina and one river site was sampled. Species sampled for morphological analysis at respective locations were Unio pictorum (grey circles), Unio tumidus (filled circle) and Anodonta anatina (open circle). Map © Crown Copyright/database right 2008. An Ordnance Survey/EDINA supplied service.
Relative mussel densities and species abundances at each site were estimated by
counting the number of individuals per species present in 15 replicate dredges
(approximately 5 m in length, dredge aperture 46 x 21 x 7.5 cm, mesh size = 2.5 cm).
To elucidate (dis-)similarities in morphological trends, at least 15 Unio pictorum
(Linnaeus, 1758) individuals were collected by hand at each marina and river site and
brought back to the laboratory for subsequent morphological analysis. Additionally, the
two other unionoids present in the study area, Anodonta anatina (Linnaeus, 1758) and
Unio tumidus Philipsson, 1788, were sampled at one of the five locations (Abingdon;
Fig. 2.1). Overall, a total of 181 U. pictorum, 39 U. tumidus and 39 A. anatina specimens
were investigated regarding their morphological characters. To minimise morphological
differences between populations resulting from differences of ontogenetic stages of their
individuals, all mussels collected were more than 4 years old based on external annual
rings (Haskin, 1954), and sagittal shell areas between populations of the same species were
not significantly different (ANOVA: U. pictorum: F9,181 = 1.868, P = 0.060; U. tumidus:
F1,38 = 0.236, P = 0.630; A. anatina: F1,38 = 0.735, P = 0.397). All specimens analysed have
been deposited at the University Museum of Zoology Cambridge.

Chapter 2 – Ecophenotypic trends in freshwater mussels
21
2.3.2. SAMPLING OF HABITAT PARAMETERS
At each site, one measurement of water temperature was made using a digital
thermometer. Five replicate measurements of chlorophyll a concentrations were also taken
using an in vivo fluorometer (Aquafluor Handheld Fluorometer, Turner Designs,
Sunnyvale, California). Measurements at marina and river pairs were made within a period
of 1h.
At each site, five random sediment cores were taken down to a depth of 10-15 cm.
Sediment samples were dried to constant weight and sieved through a sieving tower of six
sieves with a mesh size in the range 2-31.5 mm. Sediment fractions <2 mm grain size were
analysed in a Malvern Mastersizer 2000 giving relative volume-proportions of grain size
classes to the 0.01% level. Assuming a similar specific density for all grain sizes, the
weighing of the total respective <2 mm- and >2 mm-grain size proportions allowed the
conversion of volume-proportions to weight-proportions. Finally, median grain sizes of
samples were calculated from their cumulative weight percentage plots. The organic
matter content of each sediment sample was estimated by calculating weight loss on
ignition (24h at 600°C).
2.3.3. GROWTH MEASUREMENTS, AND AGE AND SEX DETERMINATION OF INDIVIDUALS
The age of each U. pictorum specimen was estimated by counting annual winter
rings displayed on the shell (Aldridge, 1999b). For shape-independent analysis of growth,
lengths and heights of individuals at each of these annuli were measured with a digital
calliper, and sagittal shell areas (SaA) at each year of age were estimated by applying the
standard formula for an ellipse. Square root of sagittal shell area (SaA1/2) per age plots
were produced for each site by calculating the mean SaA1/2 at each year. Growth
parameters were determined using the Walford plot model (Walford, 1946) assuming von
Bertalanffy growth curves (von Bertalanffy, 1938). All specimens were sexed by
microscopic inspection of gonadal tissue sensu Heard (1975).
2.3.4. MORPHOLOGICAL ANALYSIS
2.3.4.1. Fourier shape analysis
To compare two-dimensional shell outlines of the individuals, we used Fourier shape
analysis sensu Crampton & Haines (1996). This multivariate morphometric tool analyses

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
22
the whole sagittal outline of the shell, making it ideal for bivalves that lack the sufficient
number of homologous landmarks needed for landmark-based morphometric techniques.
In Fourier shape analysis, an outline contour is decomposed into a number of basic
waves, termed harmonics, with each of them in turn explained by two respective Fourier
coefficients. The more harmonics, and therefore Fourier coefficients, calculated, the more
accurately the description of the outline will be. Each shell outline is so described by a set
of Fourier coefficients that can be statistically treated as any usual variable. Because
Fourier coefficients contain no size information, a standardisation of the outlines to the
same size is not necessary.
For analysis, digital photographs of right valves of all specimens were taken. After
enhancing contrast between shell and background, shell outlines were digitised with
program ImageJ (Rasband, 2008). These digitised outlines were then used for Fast Fourier
Transform using the HANGLE software (Crampton & Haines, 1996). Before calculating
Fourier coefficients, a smoothing normalisation of 10 was applied for reduction of high-
frequency pixel noise resulting from the digitisation process. Preliminary analyses showed
that the first eight harmonics explained the shell outlines with sufficiently high precision,
whereby the first harmonic does not contain any shape information and, thus, is discarded
from the analysis by the software. This resulted in a set of 14 Fourier coefficients per
individual. Finally, HMATCH software was used to rotate all outlines treated in one
statistical analysis (e.g. all outlines of one species) for maximum overlap. The sets of
Fourier coefficients obtained this way were subsequently used in statistical analysis
described below.
2.3.4.2. Analysis of traditional shell and anatomical measurements
Figure 2.2 illustrates shell and anatomical measurements taken. For traditional shell
shape analysis the three shell dimensions length, height and width were measured at each
specimen. Additionally, several anatomical measurements were taken to elucidate changes
in internal characters associated with those in shell forms. Sagittal cross-sectional areas of
the adductors and unfused “siphons” (apertures) were estimated by measuring their dorso-
ventral and antero-posterior radii and applying the standard formula for the area of an
ellipse. Gills were dissected from the mussel and areas measured. Excised tissue (dissected
foot and remaining soft parts separately) was dried to constant mass (60°C for 48h).
Sagittal shell area (SaA) and shell volume (V) were estimated by applying the standard

Chapter 2 – Ecophenotypic trends in freshwater mussels
23
formulas for an ellipse and ellipsoid, respectively, using the measurements of the three
shell dimensions.
Figure 2.2. Shell and anatomical measurements taken at each specimen. Abbreviations: aA, anterior adductor area; eS, exhalant ‘siphon’ area; F, dry foot weight; H, shell height; iG, inner gill area; iS, inhalant “siphon” area; L, shell length; oG, outer gill area; pA, posterior adductor area; W, shell width. Additionally, the dry weight of all soft tissues of the animal was measured.
2.3.5. STATISTICAL ANALYSIS
Statistical analyses were performed using PAST (Hammer, Harper & Ryan, 2001;
Hammer & Harper, 2006) and MINITAB for Windows, Version 14 (Minitab Inc.).
Altogether eight separate principal component analyses (PCA) were carried out
using either sets of 14 Fourier coefficients or five internal morphological ratios for four
respective groups of populations: U. pictorum from all ten sites together, and each of the
three unionoid species from the Abingdon location separately. The number of principal
components (PCs) to be retained was determined by use of the broken stick model of the
scree plot. Only the first two PCs were used for graphical representation in scatter plots.
Synthetic outlines of “extreme shell forms” were drawn using HCURVE software sensu
Crampton & Haines (1996).
To estimate importance of non-habitat factors in relation to the habitat factor on
morphological trends, three three-way-analyses of covariance (ANCOVAs) were
performed on relative shell length, relative shell width, and the first PC obtained by PCA
on Fourier coefficients on all ten U. pictorum populations using factors habitat, location
and either (1) sex, (2) covariate size, or (3) covariate age. Eta-squared values calculated
this way are a measure of proportion of variance explained by each factor or covariate,
respectively.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
24
2.4. RESULTS
2.4.1. HABITAT PARAMETERS
Marinas were approximately 0.5°C warmer than adjacent river sites (paired: t =
3.627, P = 0.022, N = 5; Table 2.1). No significant differences could be observed between
marina and river sites in chlorophyll a concentration (Wilcoxon: z = 1.753, P = 0.080, N =
5), median grain size (paired: t = 1.989, P = 0.117, N = 5) and percent organic matter
content of the sediment (paired: t = 1.867, P = 0.135, N = 5).
Table 2.1. Chlorophyll a, median grain sizes, organic matter content of sediment (means ± SD) and water temperature at the ten sites. Capital letters indicate the ‘location’; lower case the ‘habitat’ of the site: A, Abingdon; C, Caversham; M, Marlow; W, Windsor; O, Old Windsor; m, marina; r, river.
Site Temperature (°C) Chl a [μg L-1] Q50 (grain size, µm) % Organic matter Am 13.9 7.6 ± 0.8 4433 ± 3001 4.23 ± 0.37 Ar 13.2 6.6 ± 0.3 3780 ± 5823 4.46 ± 2.65 Cm 14 5.0 ± 0.1 9191 ± 6705 3.89 ± 1.32 Cr 13.1 5.8 ± 0.2 13724 ± 11847 3.06 ± 1.66 Mm 13.5 8.4 ± 0.8 5162 ± 6997 13.26 ± 7.36 Mr 13 5.8 ± 1.3 11453 ± 9321 2.71 ± 2.05 Wm 13.7 24.4 ± 1.1 1160 ± 2037 5.87 ± 5.79 Wr 13.5 5.8 ± 0.5 12845 ± 3471 1.56 ± 0.68 Om 13.7 434.5 ± 11.0 1379 ± 1525 8.31 ± 6.22 Or 13.5 5.7 ± 0.3 1668 ± 3404 5.97 ± 2.05
2.4.2. SPECIES COMPOSITION
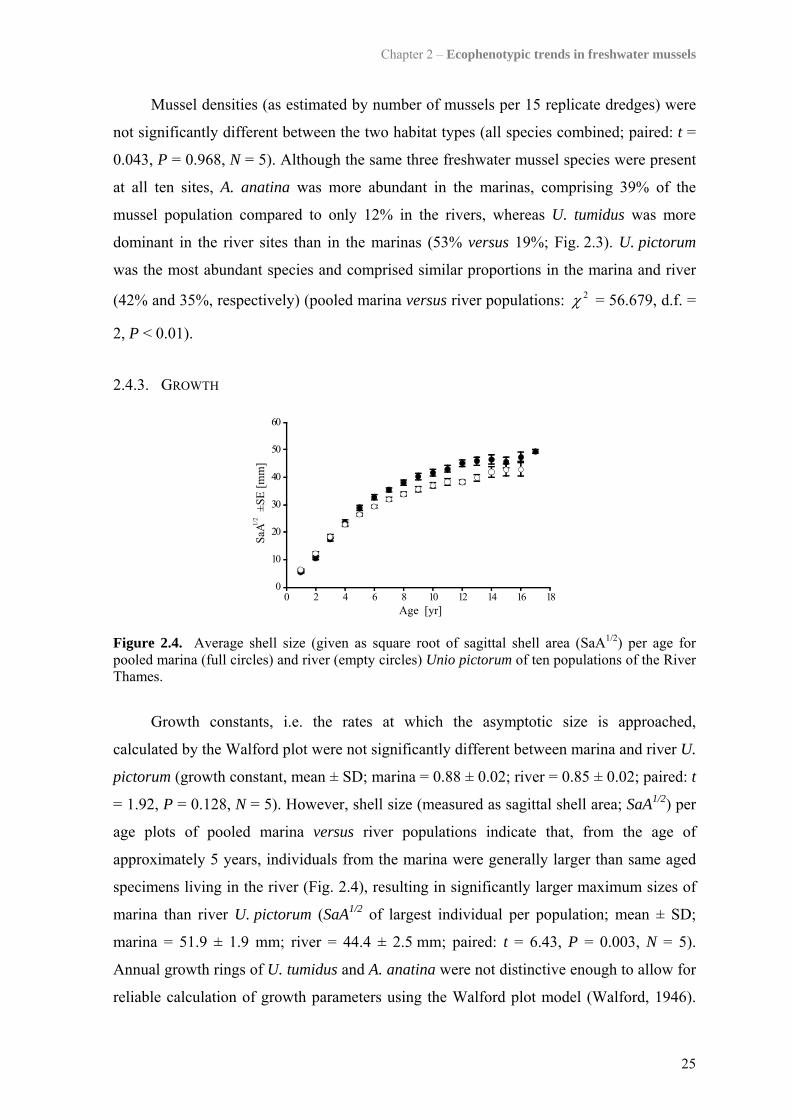
Figure 2.3. Relative abundances of the three unionoid species present at the ten sites. Numbers above each column correspond to overall number of specimens dredged at each site. Abbreviations: m, marina site; r, river site.
Chapter 2 – Ecophenotypic trends in freshwater mussels
25
Mussel densities (as estimated by number of mussels per 15 replicate dredges) were
not significantly different between the two habitat types (all species combined; paired: t =
0.043, P = 0.968, N = 5). Although the same three freshwater mussel species were present
at all ten sites, A. anatina was more abundant in the marinas, comprising 39% of the
mussel population compared to only 12% in the rivers, whereas U. tumidus was more
dominant in the river sites than in the marinas (53% versus 19%; Fig. 2.3). U. pictorum
was the most abundant species and comprised similar proportions in the marina and river
(42% and 35%, respectively) (pooled marina versus river populations: 2χ = 56.679, d.f. =
2, P < 0.01).
2.4.3. GROWTH
Figure 2.4. Average shell size (given as square root of sagittal shell area (SaA1/2) per age for pooled marina (full circles) and river (empty circles) Unio pictorum of ten populations of the River Thames.
Growth constants, i.e. the rates at which the asymptotic size is approached,
calculated by the Walford plot were not significantly different between marina and river U.
pictorum (growth constant, mean ± SD; marina = 0.88 ± 0.02; river = 0.85 ± 0.02; paired: t
= 1.92, P = 0.128, N = 5). However, shell size (measured as sagittal shell area; SaA1/2) per
age plots of pooled marina versus river populations indicate that, from the age of
approximately 5 years, individuals from the marina were generally larger than same aged
specimens living in the river (Fig. 2.4), resulting in significantly larger maximum sizes of
marina than river U. pictorum (SaA1/2 of largest individual per population; mean ± SD;
marina = 51.9 ± 1.9 mm; river = 44.4 ± 2.5 mm; paired: t = 6.43, P = 0.003, N = 5).
Annual growth rings of U. tumidus and A. anatina were not distinctive enough to allow for
reliable calculation of growth parameters using the Walford plot model (Walford, 1946).

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
26
However, in both species, maximum shell sizes were larger in the respective marina
populations (A. anatina: SaA1/2max = 70.4 mm at marina, SaA1/2
max = 56.0 mm at river; U.
tumidus: SaA1/2max = 53.8 mm at marina, SaA1/2
max = 51.2 mm at river).
2.4.4. MORPHOLOGICAL ANALYSIS
2.4.4.1. Morphological trends in U. pictorum of five paired sites and influence of habitat and non-habitat factors
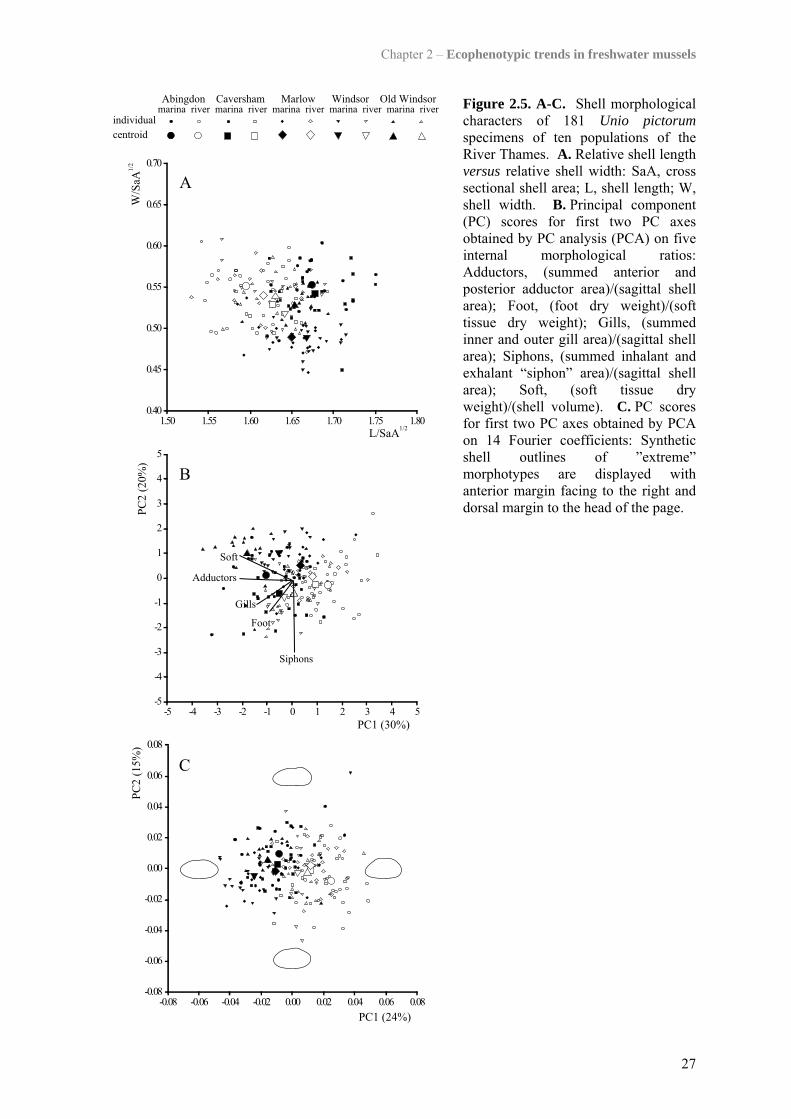
Figure 2.5 displays scores of 181 U. pictorum specimens for (A) relative shell
elongation and obesity, and the first two PCs obtained by analyses using (B) five internal
morphological ratios and (C) 14 Fourier coefficients, respectively. All three scatter plots
show a clear clustering of river versus marina individuals along their first respective axes,
indicating that, across all five paired sites, U. pictorum from the same habitat were
morphologically more similar to each other than those from the same location.
Synthetic outlines (Fig. 2.5C) and photographs (Fig. 2.6) visualise sagittal shapes of
“extreme” habitat forms. Marina shells typically displayed a pointed posterior and straight
dorsal margin, whereas those from the river tended to be dorsally arched. Additionally,
marina U. pictorum were comparatively more elongated than river forms, which is shown
both by modern (Fig. 2.5C) and traditional (Fig. 2.5A) morphometric techniques. These
trends in sagittal shell shape were accompanied by relatively higher dry soft tissue weight
and larger adductor muscles in the marina mussels compared to those in the river
(Fig. 2.5B). Scatter along subsequent PC axes retained by the broken stick model, but not
displayed in Figure 2.5 (i.e. PC3 of Fourier coefficients explaining 11% of variance),
reflected variation within populations and did not discriminate marina from river mussels.
Eta-squared values obtained by three three-way-ANCOVAs indicate that the shape
of the dorso-posterior margin (PC1 of Fourier shape analysis) was mainly influenced by
the habitat the mussel was living in, and that neither ontogenetic stage nor sex of an
individual had any considerable effect in this respect (Fig. 2.7). The degree of shell
elongation was significantly influenced by factors habitat and location but also the
interaction of these two factors. This indicates that ecophenotypic trends in this shell
character differed between locations. Although shell obesity was significantly influenced
by several habitat and non-habitat factors/covariates and their interactions, this character
was especially associated with the habitat-location interaction factor (Fig. 2.7). This is also
indicated in Figure 2.5A, demonstrating that mussels of the Marlow and Windsor marinas
displayed generally lower relative shell width values than those from all other sites.
Chapter 2 – Ecophenotypic trends in freshwater mussels
27
Figure 2.5. A-C. Shell morphological characters of 181 Unio pictorum specimens of ten populations of the River Thames. A. Relative shell length versus relative shell width: SaA, cross sectional shell area; L, shell length; W, shell width. B. Principal component (PC) scores for first two PC axes obtained by PC analysis (PCA) on five internal morphological ratios: Adductors, (summed anterior and posterior adductor area)/(sagittal shell area); Foot, (foot dry weight)/(soft tissue dry weight); Gills, (summed inner and outer gill area)/(sagittal shell area); Siphons, (summed inhalant and exhalant “siphon” area)/(sagittal shell area); Soft, (soft tissue dry weight)/(shell volume). C. PC scores for first two PC axes obtained by PCA on 14 Fourier coefficients: Synthetic shell outlines of ”extreme” morphotypes are displayed with anterior margin facing to the right and dorsal margin to the head of the page.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
28
Figure 2.6. A-B. Photographs of ‘extreme’ (A) marina (Windsor), and (B) river (Abingdon) morphotypes of Unio pictorum.
Figure 2.7. A-C. Proportions of total variance in the shell morphologic parameters relative shell length (rel. L; given as L/SaA1/2), relative shell width (rel. W; given as W/SaA1/2), and shape of dorso-posterior margin (PC1 Fourier; given as PC1 scores obtained by principal component analysis on 14 Fourier coefficients) of ten Unio pictorum populations, respectively, as a result of variation in the factors habitat (Hab), location (Loc) and (A) sex (Sex), (B) covariate sagittal shell area (Size; i.e. measurement for size), or (C) covariate age (Age). Only significant interactions are shown as vertical bars.
2.4.4.2. Morphological trends in all three unionoid species at Abingdon
In all three unionoid species at Abingdon, scatter along the first two axes of PCAs on
14 Fourier coefficients show similar patterns with regard to a pronounced clustering of
individuals from the same habitat, and a trend from relatively straight dorsal and pointed
posterior margins in marina mussels to downwardly curved dorso-posterior margins in
river forms (Fig. 2.8G,H,I). However, the elongation of marina forms was more
pronounced for U. pictorum than for A. anatina and could not be observed in U. tumidus
(Fig. 2.8A,B,C,G,H,I). Marina specimens of A. anatina show a trend to more pronounced
wing development than river individuals (Fig. 2.8I). Finally, all three species showed the
same trend for relatively larger adductor muscles and higher soft tissue weight per shell
volume in marina mussels compared to those from the main channel of the River Thames
(Fig. 2.8D,E,F). Subsequent significant principal components retained but not displayed in
Figure 2.8 (i.e. PCA on Fourier coefficients; U. pictorum: PC3 11%; U. tumidus: 12% of
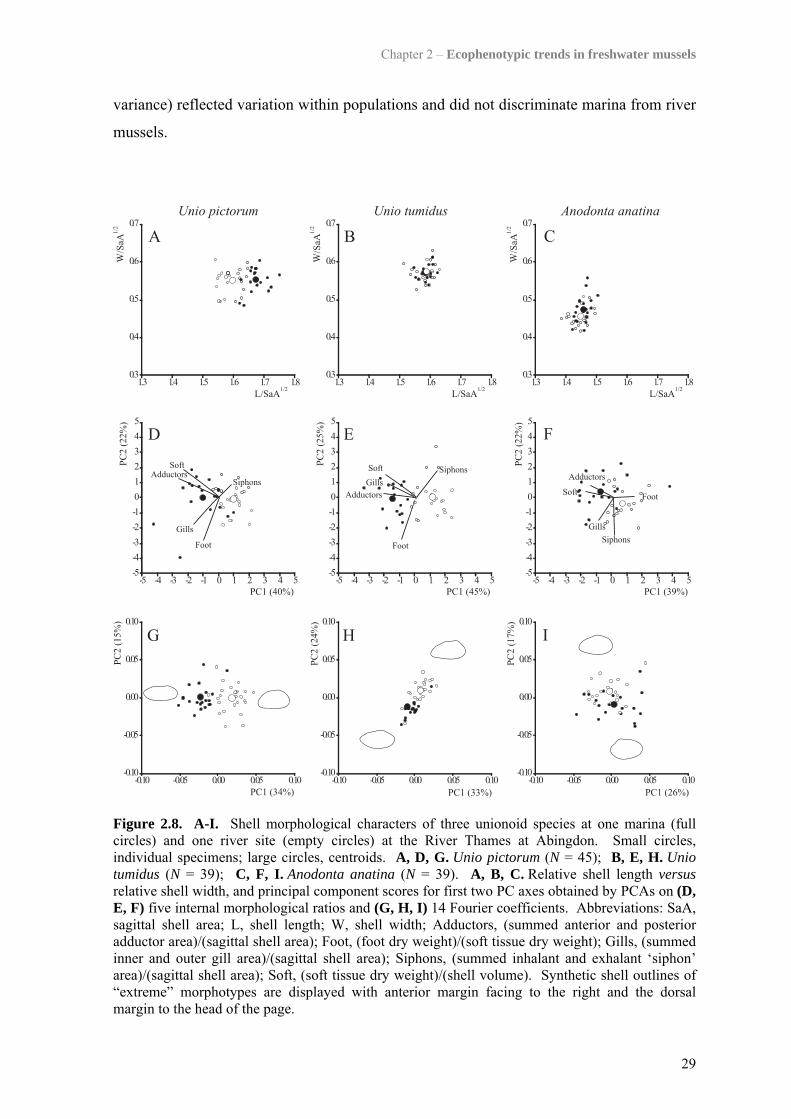
Chapter 2 – Ecophenotypic trends in freshwater mussels
29
variance) reflected variation within populations and did not discriminate marina from river
mussels.
Figure 2.8. A-I. Shell morphological characters of three unionoid species at one marina (full circles) and one river site (empty circles) at the River Thames at Abingdon. Small circles, individual specimens; large circles, centroids. A, D, G. Unio pictorum (N = 45); B, E, H. Unio tumidus (N = 39); C, F, I. Anodonta anatina (N = 39). A, B, C. Relative shell length versus relative shell width, and principal component scores for first two PC axes obtained by PCAs on (D, E, F) five internal morphological ratios and (G, H, I) 14 Fourier coefficients. Abbreviations: SaA, sagittal shell area; L, shell length; W, shell width; Adductors, (summed anterior and posterior adductor area)/(sagittal shell area); Foot, (foot dry weight)/(soft tissue dry weight); Gills, (summed inner and outer gill area)/(sagittal shell area); Siphons, (summed inhalant and exhalant ‘siphon’ area)/(sagittal shell area); Soft, (soft tissue dry weight)/(shell volume). Synthetic shell outlines of “extreme” morphotypes are displayed with anterior margin facing to the right and the dorsal margin to the head of the page.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
30
2.5. DISCUSSION
The main aim of the present study was to identify ecophenotypic trends in unionoid
shell and internal morphology that could find application in taxonomy, palaeontology, and
conservation of rare species. We revealed a number of consistent morphological patterns,
both within and between species, which characterise mussels collected from river or
marina sites.
2.5.1. PATTERNS ACROSS ALL SPECIES
In general, marina-collected specimens grew larger, had relatively higher soft tissue
dry weight, larger adductors, were less dorsally arched and had a more pointed posterior
than river-collected specimens. Shell size at a given age in freshwater mussel species has
often been shown to be associated with changes in water temperature and/or food
(especially phytoplankton) availability (Grier, 1920; Negus, 1966; Reigle, 1967; Ghent,
Singer & Johnson-Singer, 1978). In the present study, in marinas, all species grew to larger
maximum sizes than those in the river. This could be explained by the higher water
temperatures in marinas, which can support a longer growing season and faster metabolism
(Mann, 1965). Mussel growth might additionally have been enhanced by higher
phytoplankton densities that would be expected in the standing, relatively warm water
bodies of marinas compared to the flowing, colder river. Lack of a complete dataset on
seasonal patterns may explain why we were unable to detect significant differences in
chlorophyll a levels between the two habitats. The higher quality of growing conditions in
the marinas is also supported by the mussels’ higher soft tissue dry weight per shell cavity
volume ratios, comprising a measurement commonly used as bivalve body condition index
(Crosby & Gale, 1990).
The observation of relatively large adductors in marina specimens compared to those
in the river may be a side-effect of differences in relative soft tissue mass at the two
habitats. Under such a scenario, adductor size may therefore be considered as non-
adaptive. Adductors are known to play an important role in bivalve burrowing and
anchorage (Trueman, 1966a, b, 1968), and larger and therefore stronger adductors would
be expected to be more valuable in stronger water movement. Our observation of
proportionately larger muscles in slower flowing habitats is clearly in conflict with this
theory and other observations (Sell, 1907-1908; Balla & Walker, 1991). It is quite possible
that internal anatomy is not independent of shell morphology, and so adductor size might
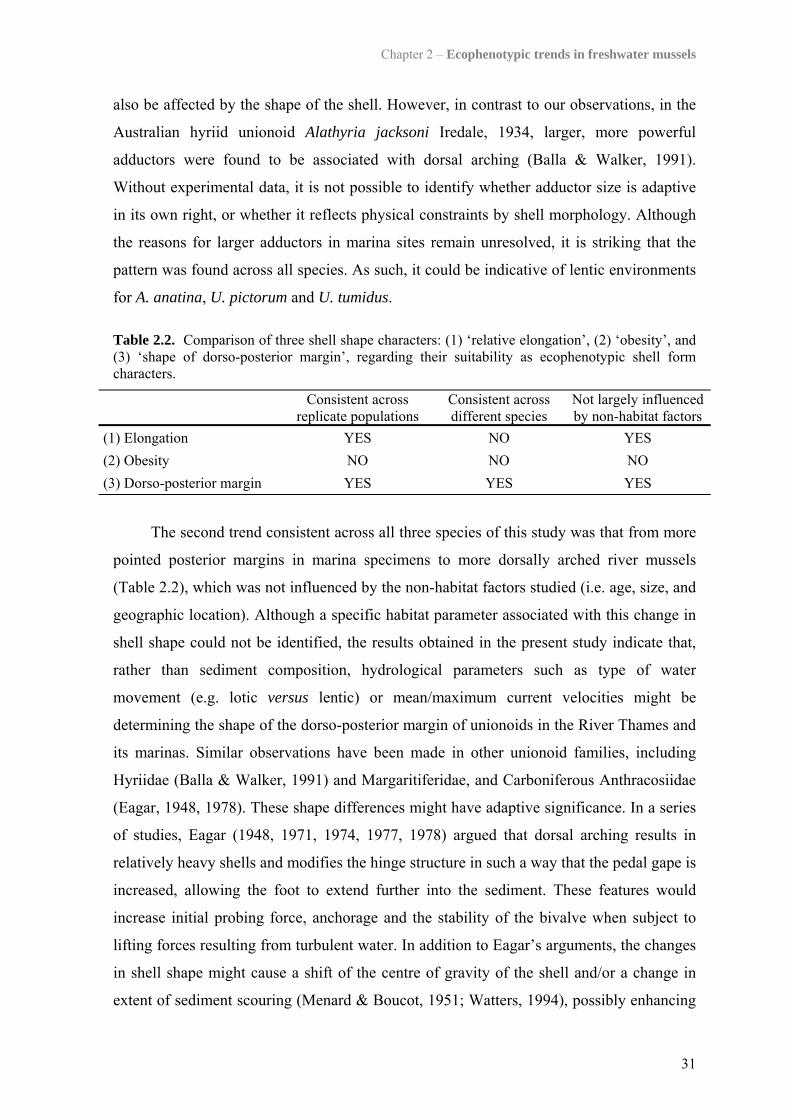
Chapter 2 – Ecophenotypic trends in freshwater mussels
31
also be affected by the shape of the shell. However, in contrast to our observations, in the
Australian hyriid unionoid Alathyria jacksoni Iredale, 1934, larger, more powerful
adductors were found to be associated with dorsal arching (Balla & Walker, 1991).
Without experimental data, it is not possible to identify whether adductor size is adaptive
in its own right, or whether it reflects physical constraints by shell morphology. Although
the reasons for larger adductors in marina sites remain unresolved, it is striking that the
pattern was found across all species. As such, it could be indicative of lentic environments
for A. anatina, U. pictorum and U. tumidus.
Table 2.2. Comparison of three shell shape characters: (1) ‘relative elongation’, (2) ‘obesity’, and (3) ‘shape of dorso-posterior margin’, regarding their suitability as ecophenotypic shell form characters.
Consistent across replicate populations
Consistent across different species
Not largely influenced by non-habitat factors
(1) Elongation YES NO YES (2) Obesity NO NO NO (3) Dorso-posterior margin YES YES YES
The second trend consistent across all three species of this study was that from more
pointed posterior margins in marina specimens to more dorsally arched river mussels
(Table 2.2), which was not influenced by the non-habitat factors studied (i.e. age, size, and
geographic location). Although a specific habitat parameter associated with this change in
shell shape could not be identified, the results obtained in the present study indicate that,
rather than sediment composition, hydrological parameters such as type of water
movement (e.g. lotic versus lentic) or mean/maximum current velocities might be
determining the shape of the dorso-posterior margin of unionoids in the River Thames and
its marinas. Similar observations have been made in other unionoid families, including
Hyriidae (Balla & Walker, 1991) and Margaritiferidae, and Carboniferous Anthracosiidae
(Eagar, 1948, 1978). These shape differences might have adaptive significance. In a series
of studies, Eagar (1948, 1971, 1974, 1977, 1978) argued that dorsal arching results in
relatively heavy shells and modifies the hinge structure in such a way that the pedal gape is
increased, allowing the foot to extend further into the sediment. These features would
increase initial probing force, anchorage and the stability of the bivalve when subject to
lifting forces resulting from turbulent water. In addition to Eagar’s arguments, the changes
in shell shape might cause a shift of the centre of gravity of the shell and/or a change in
extent of sediment scouring (Menard & Boucot, 1951; Watters, 1994), possibly enhancing

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
32
the effects mentioned above. Nevertheless, the differences in shell shapes could have no
adaptive value whatsoever but be caused by a non-functional reaction of the mussel to the
environment (e.g. via distortion of the shell secreting mantle margin as a result of water
movement). Intraspecific variation in life position such as burrowing depth or angle
(Amyot & Downing, 1991; Scholz, 2003) could further influence shell growth and should
be considered in this respect. Although the functional significance of posterior pointing
remains unresolved, the consistency of this shell form trend indicates that the shape of the
dorso-posterior margin could be a powerful ecophenotypic character in freshwater
mussels, and be valid for a wide range of species and habitats.
2.5.2. PATTERNS WITHIN SINGLE SPECIES
Some shell form trends, such as that regarding the degree of wing development in
A. anatina, were observed only in one of the three species studied. Wings (alae) are
considered to act as secondary ligaments or hinge teeth making up for the reduced
dentition of most symphynote species (Savazzi & Peiyi, 1992), whereas, as a result of their
well developed dentition, members of the genus Unio are not known to increase their
dorsal margin surface in such a manner. Watters (1994) argued that enlargement of wings
within a given species might enhance the “snowshoe-effect” (i.e. preventing sinking and
providing increased stability in soft or unstable substrates), which is a probable
explanation for our observations of bigger alae in marina specimens. The degree of shell
elongation, on the other hand, was markedly different between populations in the two
habitats for U. pictorum but could not be detected in the other two unionoid species.
Similar observations on inconsistent ecophenotypic trends of unionoid species are
common in the literature. For example, several studies reported that, in the genus Unio,
more elongated shells were associated with slower flowing habitats (Bridgeman, 1875;
Hey, 1882; Israel, 1910; March, 1910-1911; Tudorancea, 1972), although contradictory
observations have been made on Margaritifera margaritifera (Linnaeus, 1758) (Altnöder,
1926). Such inconsistencies of intraspecific patterns might root in both ecological and
behavioural differences between individuals or populations. For example, microhabitat can
affect the degree of embedding in the sediment (Agrell, 1948), which could lead to
differences in physical forces acting on the mussels and, in turn, influence their shell
formation. Additionally, dominance of A. anatina and U. tumidus at marina and river sites,
respectively, indicates that these two species show stronger habitat specificity than the

Chapter 2 – Ecophenotypic trends in freshwater mussels
33
more generalist U. pictorum. On this basis, generalists would be expected to show a higher
degree of phenotypic variability, which could explain our failure to detect ecophenotypic
trends in shell height to length ratios in A. anatina and U. tumidus.
By contrast to patterns in sagittal shell shape, relative shell width (obesity) of
U. pictorum proved to be considerably influenced by allometric growth. Similar alterations
in growth pattern with size and/or age have commonly been described in both marine
(Ohba, 1959; Johannessen, 1973) and freshwater mussels (Ball, 1922; Scholz, 2003).
Nevertheless, relative shell width has often been used as an ecophenotypic character,
which has even led to the formulation of the ‘Law of Stream Distribution’ (Ortmann,
1920) with numerous studies supporting (Grier, 1920; Ball, 1922; Grier & Mueller, 1926;
Reigle, 1967; Cvancara, 1972; Anderson & Ingham, 1978; Hinch, Bailey & Green, 1986;
Hinch, Kelly & Green, 1989) as well as contradicting the same (March, 1910-1911; Grier,
1920; Bailey & Green, 1988; Hinch & Bailey, 1988). An additional cause for this apparent
contradiction in pattern of obesity is likely to stem from the non-independence of the shell
measurements (length, width and height) used in traditional morphometric studies. A
consideration of allometric growth patterns can overcome such problems.
2.5.3. UTILITY OF THE PATTERNS OBSERVED
Knowing the range of morphological variation within a species is doubtlessly
important for unionoid taxonomy. Furthermore, understanding the factors certain
morphological trends are associated with can help reconstructing palaeoenvironments
(Good, 2004; Scholz & Hartmann, 2007a, b; Scholz, Tietz & Büchner, 2007) and aid in
conservation of rare or threatened species. These are often known predominantly from
shell material, with their habitat requirements poorly understood. Perhaps a comparison of
morphology with commoner congenerics can be informative, provided that features are
consistent across species (e.g. degree of arching). This may help understand reasons for a
decline (e.g. habitat loss) and also identify habitat where surveys for rare species can be
focused.
At least in temperate latitudes where annual growth rings are usually well
established, maximum sizes of shells are likely to be a good indicator of mean/maximum
water temperatures and phytoplankton densities at the habitat, and could thus be used for
palaeoenvironmental reconstructions and estimation of condition of the freshwater mussel
population at the site. Adductor scar sizes in shells could further help estimate the

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
34
condition of an individual, although other factors, such as current velocity, should be taken
into account as well. According to Claassen (1998), shell shape is perhaps the best method
for reconstructing habitat with freshwater molluscs. The results obtained in the present
study indicate that the degree of dorsal arching is one of the most generally valid
characters in unionoids and, thus, modern rather than traditional morphometric techniques
should be used in future studies of this kind. The shape of the posterior dorsal margin
appears to be associated with hydrological character of the habitat and could thus be used
in both palaeontological reconstructions and identification of habitat requirements of (rare)
species.

CHAPTER 3
PHENOTYPIC PLASTICITY AND GENETIC ISOLATION-BY-DISTANCE IN THE
FRESHWATER MUSSEL UNIO PICTORUM(MOLLUSCA: UNIONOIDA)

“In 1892 Arnould Locard, one of the great lights of the new school,
stated that there were 208 species of Unios and 250 Anodontas in
France alone. Life is too short and valuable to be wasted in any
attempt at deciphering such nonsense, and I have not even
cumbered the pages of this work with a list of these new species.
Those interested can find them in the works of Westerlund and
Kobelt.”
Simpson (1900)

37
CHAPTER 3
PHENOTYPIC PLASTICITY AND GENETIC ISOLATION-BY-DISTANCE IN THE FRESHWATER MUSSEL UNIO PICTORUM
(MOLLUSCA: UNIONOIDA)
3.1. ABSTRACT
Freshwater mussels (Unionoida) show high intraspecific morphological variability,
and some shell morphological traits are believed to be associated with habitat conditions. It
is not known whether and which of these ecophenotypic differences reflect underlying
genetic differentiation or are the result of phenotypic plasticity. Using 103 amplified
fragment length polymorphism (AFLP) markers, we studied population genetics of three
paired Unio pictorum populations sampled from two different habitat types (marina and
river) along the River Thames. We found genetic differences along the Thames which
were consistent with a pattern of isolation by distance and probably reflect limited
dispersal via host fish species upon which unionoid larvae are obligate parasites. No
consistent genetic differences were found between the two different habitat types
suggesting that morphological differences in the degree of shell elongation and the shape
of dorso-posterior margin are caused by phenotypic plasticity. Our study provides the first
good evidence for phenotypic plasticity of shell shape in a European unionoid and
illustrates the need to include genetic data in order properly to interpret geographic
patterns of morphological variation.
3.2. INTRODUCTION
Many species of freshwater mussels of the order Unionoida display great variability
in shell morphology (Fig. 1.2), some of which appear to be associated with differences in
habitat (Chapter 2; Ortmann, 1920; Agrell, 1948; Eagar, 1978; Watters, 1994; Zieritz &
Aldridge, 2009). These so-called ecomorphotypes may arise through two mechanisms.
First, they may simply reflect phenotypic plasticity, the ability to change phenotype in
response to variation in the environment (Via et al., 1995). The alternative mechanism
involves genetic variability. If a species produces very large numbers of offspring, it is
possible that those that manage to settle in any given habitat represent a biased subset of

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
38
all offspring, specifically those able to do best under those conditions. The unionoid life
cycle potentially allows for both mechanisms to operate. Thus, the larvae of almost all
unionoid species are obligate fish parasites, making the habitat in which the juvenile
mussel excysts from the host highly unpredictable, and hence putting a premium on the
ability to thrive under diverse habitats. Equally, females brood eggs in marsupia which can
be fertilised by several males via the inhalant current, potentially producing genetically
diverse offspring (e.g. Kat, 1984). Which of these two mechanisms dominates in nature
remains open to debate.
Few genetic studies have explicitly investigated the role of genetic versus phenotypic
variability on morphological variation in unionoids, and these have typically used
relatively conservative DNA markers that lacked adequate resolution to detect genetic
differences among populations (e.g. Soroka & Zdanowski, 2001). Nevertheless, previous
publications have generated mixed support for the two hypotheses. For example, reciprocal
transplant experiments on two unionoid species between Canadian lakes showed that shell
height to length ratios usually change as a function of the environment, indicating
phenotypic plasticity of shell form. On the other hand, overall growth rates appear to be
under genetic control (Hinch, Bailey & Green, 1986; Hinch & Green, 1989).
Transplant experiments offer a direct test of whether phenotypic plasticity operates,
but are often undesirable ecologically. Alternative tests are based on searches for a
correlation between phenotype and genotype. For example, Buhay et al. (2002) and
Machordom et al. (2003), studied morphologically distinct Epioblasma and Margaritifera
“subspecies” respectively, but failed to find differences based on allozymes and/or
mitochondrial genes. On the other hand, Serb, Buhay & Lydeard (2003) found that
mtDNA sequence data supported the validity of several Quadrula taxa that had originally
been classified on the basis of shell morphology but were later lumped with other species
on the basis that this simply reflected phenotypic plasticity. Moreover, in their study on
genetics of central European freshwater pearl mussels (Margaritifera margaritifera
Linneaus, 1758), Geist & Kuehn (2005) observed that, while in some cases
morphologically atypical mussels showed a strong genetic divergence to other populations,
in other populations a link between genetic status and shell shape was not evident.
Such inconsistent findings hint at a deeper problem. In looking for a correlation
between phenotype and genotype it is natural to select individuals from opposite ends of
the observed morphological range. These will often be drawn from habitats that are not
only divergent in ecological properties, but also widely separated geographically.

Chapter 3 – Phenotypic plasticity in Unio pictorum
39
Consequently, the resulting samples may appear genetically different simply through a
long-term lack of gene flow, rather than the presence of discrete lineages adapted to each
habitat. Since most of the work on unionoid population genetics has so far focused on
systematics and phylogeny (e.g. Davis & Fuller, 1981; Serb, Buhay & Lydeard, 2003;
Källersjö et al., 2005), conservation genetics (e.g. Mulvey et al., 1997; Buhay et al., 2002;
Geist & Kuehn, 2005) and evolutionary history (e.g. Nagel, 2000; Huff et al., 2004;
Elderkin et al., 2007), morphological differences of populations in these studies were
usually accompanied with geographic distance. This makes it difficult to assess if genetic
differences between such populations are merely a result of isolation by distance (i.e.
evidence for phenotypic plasticity) or actually reflect differences in morphology (i.e.
evidence for genetically induced morphological differences).
Here, we attempt to distinguish between phenotypic plasticity and genetic adaptation
by exploiting an unusual system in which large habitat differences can be found over very
short geographic distances. Unio pictorum (Linnaeus, 1758) is an abundant species in the
River Thames and can be found living both in the river itself, and in adjacent marinas (Fig.
3.1). By sampling from paired sites along the river, we are able to ask whether any genetic
differences are due primarily to a simple isolation by distance model, or to the existence of
two genetically distinct morphs, one adapted to the marina habitat and one to the river.
Genetic analysis was conducted using Amplified Fragment Length Polymorphisms
(AFLPs) (Vos et al., 1995; Blears et al., 1998), a technique that reveals large numbers of
variable traits capable of resolving finescale differentiation within a population (Mueller &
Wolfenbarger, 1999).
3.3. MATERIAL AND METHODS
3.3.1. SAMPLING
From May to October 2007, a total of 146 U. pictorum specimens were collected by
hand from three river and three adjacent marina sites of the River Thames (Fig. 3.1). A
foot tissue sample was removed from each mussel and stored in 96% ethanol for
subsequent genetic analysis. All specimens and genetic samples have been deposited at the
University Museum of Zoology Cambridge.
Marinas were directly connected to the river and the paired sampling sites were no
more than 800 m apart, whereas geographic distances between the three sampling locations
ranged from about 30-100 km. The River Thames in the stretch studied has an average

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
40
width of ca. 50 m, and maximum depth of ca. 4 m. Flow is regulated by a series of weirs,
such that mean summer and winter discharges approximate 40 to 80 m3 s-1, respectively
(data from UK Environment Agency). Marina sites were typical lentic systems with no
flow, surface areas of 10 to 80 ha and maximum depths up to 4 m. The river sites typified a
UK lowland lotic system. Chapter 2 and Zieritz & Aldridge (2009) showed that water
temperatures and phytoplankton densities in River Thames marinas were consistently
higher than in the adjacent main river channel.
Figure 3.1. Sampling locations along the River Thames. Sample sizes are shown for each marina and river site (see methods section for details). Map © Crown Copyright/database right 2008, an Ordnance Survey/EDINA supplied service.
3.3.2. MORPHOLOGICAL ANALYSIS
Shell length (L; maximum diameter) and shell height (H; maximum diameter
perpendicular to L) were measured in each individual to ±0.1 cm using a vernier calliper.
To estimate relative influence of habitat and location on the degree of shell elongation
(H:L-ratio), we constructed a general linear model (GLM; Minitab 15 for Windows). For
this, H was the response variable with L and geographic distance (measured as distance
from most upstream site, i.e. Abingdon Marina, to ±10 m) fitted as covariates and habitat
as a factor with two levels. To find the minimum adequate model, the full model was fitted
and then simplified by sequentially removing non significant predictor variables (Crawley,
2002).
For a more detailed analysis of shell morphology, we used Fourier shape analysis, as
developed and explained by Crampton & Haines (1996). This method decomposes xy-

Chapter 3 – Phenotypic plasticity in Unio pictorum
41
coordinates of an outline into a number of harmonics, each of which is in turn explained by
two Fourier coefficients, which can be analysed statistically like any other traits. Digital
photographs of all specimens were taken and digitised using the program IMAGEJ
(Rasband, 2008). The digitised outlines were then subjected to fast Fourier transformation
using the program HANGLE, applying a smoothing normalisation of 20 to eliminate high-
frequency pixel noise. Preliminary analysis indicated that the first 10 harmonics described
the outlines with sufficiently high precision. Discarding of the first harmonic, not
containing any shape information, by the program resulted in a set of 18 Fourier
coefficients per individual. After rotating outlines to maximum overlap by program
HMATCH, principal component analysis (PCA) was performed on the 18 Fourier
coefficients using program PAST (Hammer & Harper, 2006). Number of principal
components to be retained was determined by use of the broken stick model of the scree
plot. Synthetic outlines of “extreme shell forms” were drawn using program HCURVE as
explained in Crampton & Haines (1996).
To test for statistical significance of habitat and location on sagittal shell shape,
GLMs of all significant principal components were carried out, fitting habitat (as a factor
with two levels) together with geographic distance as predictors, and sequentially
dropping insignificant factors from the model (Crawley, 2002). Morphological distances in
overall sagittal shell outline between the six populations were estimated by performance of
discriminant function analysis (DFA) and subsequent calculation of Mahalanobis’s D²
distances of each population pair using Minitab 15 for Windows. Age and sex were not
included as predictors in GLMs because sexual dimorphism and allometric growth during
ontogeny exert a negligible influence on relative shell elongation and overall sagittal shell
shape in the six populations studied (Chapter 2; Zieritz & Aldridge, 2009). Similarly, the
degree of shell inflation (relative shell width) was not considered, in this case because, in
contrast to shell elongation and sagittal shell shape, this morphological character is
strongly influenced by ontogenetic growth in unionoids (Chapter 2; Zieritz & Aldridge,
2009).
3.3.3. GENETIC ANALYSIS
Total genomic DNA was extracted from a small piece of foot tissue using a high salt
method (Watts, 2001). AFLP genotyping was based on the protocol of Vos et al. (1995)
and is described in detail by Dasmahapatra, Hoffman & Amos (2009). Briefly, 100-400 ng
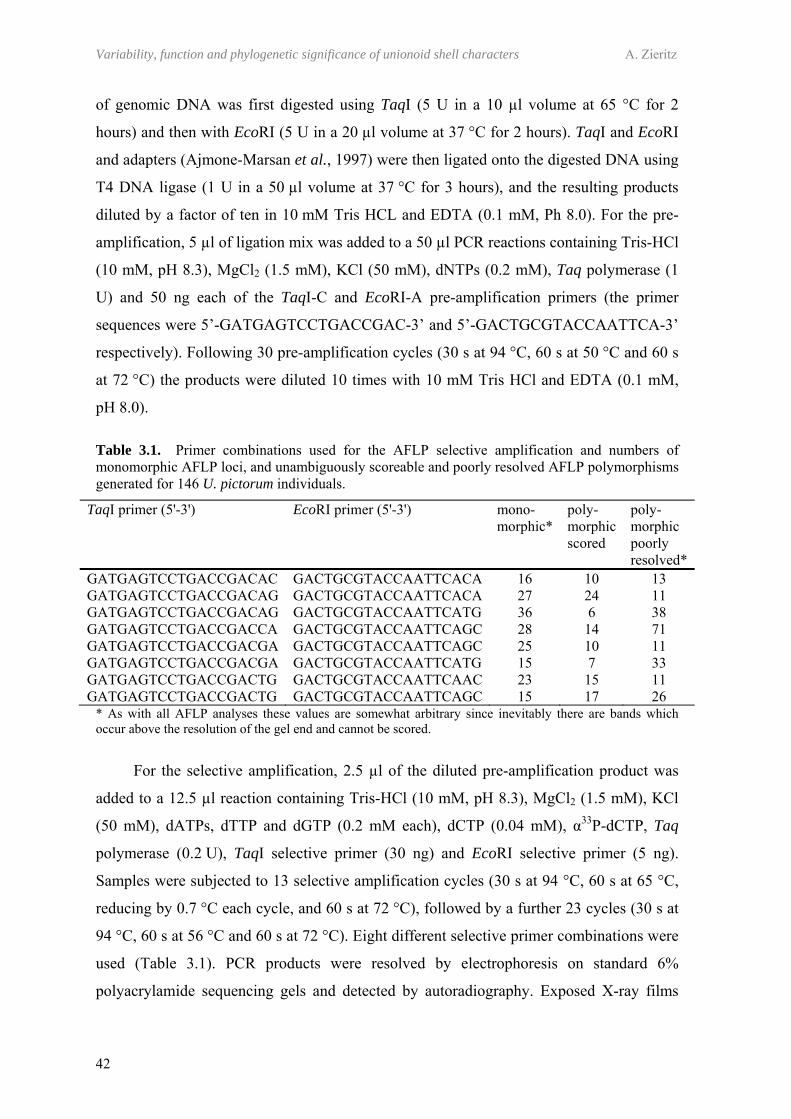
Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
42
of genomic DNA was first digested using TaqI (5 U in a 10 µl volume at 65 °C for 2
hours) and then with EcoRI (5 U in a 20 µl volume at 37 °C for 2 hours). TaqI and EcoRI
and adapters (Ajmone-Marsan et al., 1997) were then ligated onto the digested DNA using
T4 DNA ligase (1 U in a 50 µl volume at 37 °C for 3 hours), and the resulting products
diluted by a factor of ten in 10 mM Tris HCL and EDTA (0.1 mM, Ph 8.0). For the pre-
amplification, 5 µl of ligation mix was added to a 50 µl PCR reactions containing Tris-HCl
(10 mM, pH 8.3), MgCl2 (1.5 mM), KCl (50 mM), dNTPs (0.2 mM), Taq polymerase (1
U) and 50 ng each of the TaqI-C and EcoRI-A pre-amplification primers (the primer
sequences were 5’-GATGAGTCCTGACCGAC-3’ and 5’-GACTGCGTACCAATTCA-3’
respectively). Following 30 pre-amplification cycles (30 s at 94 °C, 60 s at 50 °C and 60 s
at 72 °C) the products were diluted 10 times with 10 mM Tris HCl and EDTA (0.1 mM,
pH 8.0).
Table 3.1. Primer combinations used for the AFLP selective amplification and numbers of monomorphic AFLP loci, and unambiguously scoreable and poorly resolved AFLP polymorphisms generated for 146 U. pictorum individuals.
TaqI primer (5'-3') EcoRI primer (5'-3') mono-morphic*
poly-morphic scored
poly-morphic poorly resolved*
GATGAGTCCTGACCGACAC GACTGCGTACCAATTCACA 16 10 13 GATGAGTCCTGACCGACAG GACTGCGTACCAATTCACA 27 24 11 GATGAGTCCTGACCGACAG GACTGCGTACCAATTCATG 36 6 38 GATGAGTCCTGACCGACCA GACTGCGTACCAATTCAGC 28 14 71 GATGAGTCCTGACCGACGA GACTGCGTACCAATTCAGC 25 10 11 GATGAGTCCTGACCGACGA GACTGCGTACCAATTCATG 15 7 33 GATGAGTCCTGACCGACTG GACTGCGTACCAATTCAAC 23 15 11 GATGAGTCCTGACCGACTG GACTGCGTACCAATTCAGC 15 17 26
* As with all AFLP analyses these values are somewhat arbitrary since inevitably there are bands which occur above the resolution of the gel end and cannot be scored.
For the selective amplification, 2.5 µl of the diluted pre-amplification product was
added to a 12.5 µl reaction containing Tris-HCl (10 mM, pH 8.3), MgCl2 (1.5 mM), KCl
(50 mM), dATPs, dTTP and dGTP (0.2 mM each), dCTP (0.04 mM), α33P-dCTP, Taq
polymerase (0.2 U), TaqI selective primer (30 ng) and EcoRI selective primer (5 ng).
Samples were subjected to 13 selective amplification cycles (30 s at 94 °C, 60 s at 65 °C,
reducing by 0.7 °C each cycle, and 60 s at 72 °C), followed by a further 23 cycles (30 s at
94 °C, 60 s at 56 °C and 60 s at 72 °C). Eight different selective primer combinations were
used (Table 3.1). PCR products were resolved by electrophoresis on standard 6%
polyacrylamide sequencing gels and detected by autoradiography. Exposed X-ray films

Chapter 3 – Phenotypic plasticity in Unio pictorum
43
were assessed and if required, a second exposure was made for an adjusted time period.
Gels were scored manually.
3.3.4. QUANTIFICATION OF THE GENOTYPING ERROR RATE
Although AFLPs tend to be reproducible due to the use of highly specific restriction
endonucleases coupled with stringent PCR conditions (Vos et al., 1995; Mueller &
Wolfenbarger, 1999; Bonin et al., 2004), genotyping errors can nevertheless accrue, with
potential sources of error including DNA contamination (Dyer & Leonard, 2000),
restriction artefacts (Polisky et al., 1975), human error (Bonin et al., 2004) and variation in
DNA quality among samples. Consequently, we estimated the genotyping error rate for
our dataset by independently re-extracting, re-genotyping and blind-scoring 14 (9.6%) of
the samples following Hoffman & Amos (2005). To ensure broad coverage of the dataset
in this regenotyping exercise, two to three individuals were selected at random from each
of the six populations. The error rate per reaction was then quantified as the number of
mismatching genotypes divided by the number of polymorphic bands compared (Bonin et
al., 2004).
3.3.5. GENETIC DATA ANALYSIS
All gels were independently scored by two observers and genotypes recorded as ‘1’
= band present or ‘0’ = band absent. Data were entered into a spreadsheet. We used the
program AFLP-SURV v1.0 (Vekemans, 2002) to calculate estimated heterozygosity
values for each population and pairwise FST values between the six populations (Weir &
Cockerham, 1984) following the approach of Lynch & Milligan (1994). This program was
also used to conduct a permutation test for overall genetic differentiation using 10, 000
permutations of the dataset.
In order to test whether any of the AFLP loci scored could be subject to divergent
selection relating to habitat type and thus, potentially associated with shell morphology,
we used the Dfdist program package (Beaumont & Balding, 2004; Beaumont, 2008)
following the approach of Beaumont & Nichols (1996). Dfdist implements the Bayesian
method of Zhivotovsky (1999) to estimate allelic frequencies from the proportion of
recessive phenotypes (absent bands) and then compares FST values estimated for each
locus against a theoretical null distribution of genetic differentiation conditional upon
heterozygosity in a subdivided population using the coalescent and an island model of

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
44
migration. Loci that fall outside specified confidence limits can be identified as having
significantly elevated or reduced FST values relative to expectations under the assumption
of selective neutrality. This analysis was conducted separately on four population pairs: (1)
Pooled marina vs. river, (2) Abingdon marina vs. Abingdon river, (3) Marlow marina vs.
Marlow river and (4) Old Windsor marina vs. Old Windsor river.
For each analysis, parameters a = 0.25 and b = 0.25 were used for the beta-
distributed prior of the Bayesian allele frequency estimator and the trimmed mean FST was
computed by removing the 30% highest and lowest of the observed FST values. A null
distribution was generated based on 50 000 simulated loci, the parameter 4Nµ set to 0.5
and the target-neutral FST determined using the program pv2. Simulation models were run
using different small baseline FST values until the “correct” FST for the simulations was
obtained, i.e. half the neutral points were greater than/less than the median and no trend in
the proportion of p-values greater than 0.5 with increasing heterozygosity was observable
(Beaumont, 2008). The robustness of the results to variations in the simulation parameters
was further evaluated by repeating the analyses with varying values of 4Nµ (i.e. 0.2 and
1.0). Loci with FST values above the 99% quantile were inferred as being potentially under
directional selection. This approach bypasses the joint problem that trimmed mean FST (as
recommended by Caballero, Quesada & Rolan-Alvarez (2008) was slightly negative in all
cases, while a zero FST cannot be used because it prevents the coalescent from determining
a common ancestor (see Miller et al., 2007).
3.4. RESULTS
3.4.1. MORPHOLOGICAL ANALYSIS
We began by constructing general linear models to explore which factors are
significantly associated with shell shape, summarised in Table 3.2. When shell height was
the response variable, habitat but not geographic distance was retained as a significant
predictor, indicating a strong morphological difference between mussels from the marina
and linked river habitats. This difference is also evidenced by the pronounced clustering of
marina and river individuals in a log height-length plot (Fig. 3.2). To learn about other
aspects of shape we summarised the 18 Fourier coefficients using a PCA analysis and then
fitted four further GLMs, one each using the first four principal components as the
response variable (Table 3.2). These four components were selected using the broken stick
model and together explain 57.1% of the total variance in sagittal shell shape.

Chapter 3 – Phenotypic plasticity in Unio pictorum
45
Table 3.2. Results of general linear models (GLMs) of shell height and the four significant principal components obtained by Fourier shape analysis, comparing the six U. pictorum populations. Values correspond to the minimal adequate models (Crawley, 2002).
Response variable Source d.f. SS F P
Shell height Habitat 1 0.621 39.04 <0.0001 R² = 0.91 Geographic distance - - - n.s. Shell length 1 19.421 1219.98 <0.0001 Error 143 2.276 PC1 (Fourier) Habitat 1 0.033 133.03 <0.0001 R² = 0.49 Geographic distance 1 0.002 9.34 0.003 Error 143 0.035 PC2 (Fourier) Habitat - - - n.s. R² = 0.02 Geographic distance - - - n.s. Error - - PC3 (Fourier) Habitat - - - n.s. R² = 0.28 Geographic distance 1 0.010 57.44 <0.0001 Error 143 0.025 PC4 (Fourier) Habitat - - - n.s. R² = 0.00 Geographic distance - - - n.s. Error - -
Figure 3.2. Log shell length vs. shell height scatterplots of six U. pictorum populations sampled along the River Thames. Upper case indicates the location, lower case the habitat of the site: A, Abingdon; M, Marlow; O, Old Windsor; m, marina; r, river.
PC1 scores were significantly influenced by both habitat type and geographic
distance (Table 3.2). However, whereas fitting the GLM without the term ‘geographic
distance’ resulted in a decrease of only 2.6% of variance explained by the model, 46% of
the variance explained was lost following exclusion of the term ‘habitat’. This strong
habitat-shell shape association is also reflected by the pronounced clustering of river and

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
46
marina individuals in Fig. 3.3, which shows that marina mussels tended to have more
pointed posterior margins whereas river shells displayed more arched dorso-posterior
margins. While the second and fourth principal components were not significantly
influenced by any of the two factors, PC3 was significantly correlated with geographic
distance but not habitat (Table 3.2). With respect to extreme shell outlines along this axis
(Fig. 3.3), this suggests that the posterior region of U. pictorum from the Marlow and Old
Windsor populations was broader than those from the two Abingdon sites.
Figure 3.3. Principal component scores for the first four PC axes obtained by PCA on 18 Fourier coefficients. Synthetic shell outlines of “extreme” morphotypes are displayed with the anterior margin facing to the right and the dorsal margin to the top of the page.
3.4.2. POPULATION STRUCTURE
By genotyping a total of 146 individuals at eight different selective primer
combinations, we obtained 103 AFLP loci that could be scored unambiguously across
most of the samples (Table 3.1). The AFLP dataset analysed consisted of 13, 917 binary
characters representing the presence and absence of bands. The genotyping error rate was
2.4% (35 mismatches observed out of 1442 comparisons), which is broadly consistent with
a range of previously reported values for studies using AFLPs (Bensch & Åkesson, 2005;
Bonin, Ehrich & Manel, 2007).
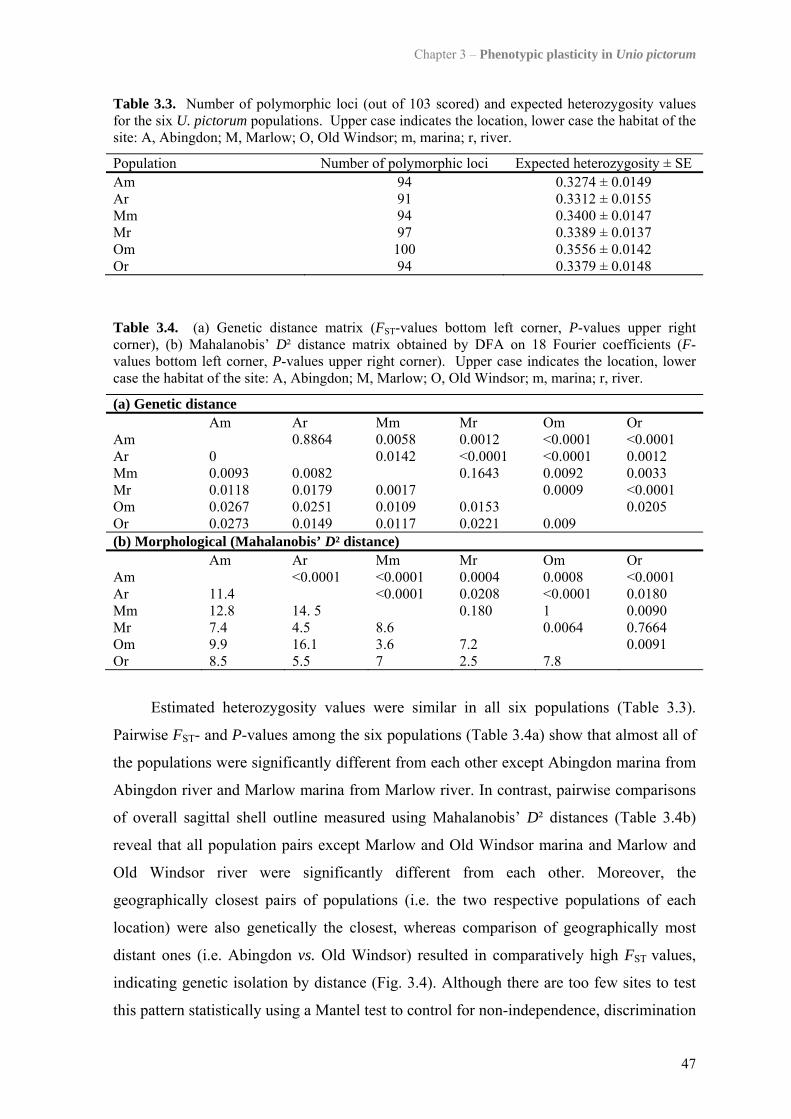
Chapter 3 – Phenotypic plasticity in Unio pictorum
47
Table 3.3. Number of polymorphic loci (out of 103 scored) and expected heterozygosity values for the six U. pictorum populations. Upper case indicates the location, lower case the habitat of the site: A, Abingdon; M, Marlow; O, Old Windsor; m, marina; r, river.
Population Number of polymorphic loci Expected heterozygosity ± SE Am 94 0.3274 ± 0.0149 Ar 91 0.3312 ± 0.0155 Mm 94 0.3400 ± 0.0147 Mr 97 0.3389 ± 0.0137 Om 100 0.3556 ± 0.0142 Or 94 0.3379 ± 0.0148
Table 3.4. (a) Genetic distance matrix (FST-values bottom left corner, P-values upper right corner), (b) Mahalanobis’ D² distance matrix obtained by DFA on 18 Fourier coefficients (F-values bottom left corner, P-values upper right corner). Upper case indicates the location, lower case the habitat of the site: A, Abingdon; M, Marlow; O, Old Windsor; m, marina; r, river.
(a) Genetic distance Am Ar Mm Mr Om Or Am 0.8864 0.0058 0.0012 <0.0001 <0.0001 Ar 0 0.0142 <0.0001 <0.0001 0.0012 Mm 0.0093 0.0082 0.1643 0.0092 0.0033 Mr 0.0118 0.0179 0.0017 0.0009 <0.0001 Om 0.0267 0.0251 0.0109 0.0153 0.0205 Or 0.0273 0.0149 0.0117 0.0221 0.009 (b) Morphological (Mahalanobis’ D² distance) Am Ar Mm Mr Om Or Am <0.0001 <0.0001 0.0004 0.0008 <0.0001 Ar 11.4 <0.0001 0.0208 <0.0001 0.0180 Mm 12.8 14. 5 0.180 1 0.0090 Mr 7.4 4.5 8.6 0.0064 0.7664 Om 9.9 16.1 3.6 7.2 0.0091 Or 8.5 5.5 7 2.5 7.8
Estimated heterozygosity values were similar in all six populations (Table 3.3).
Pairwise FST- and P-values among the six populations (Table 3.4a) show that almost all of
the populations were significantly different from each other except Abingdon marina from
Abingdon river and Marlow marina from Marlow river. In contrast, pairwise comparisons
of overall sagittal shell outline measured using Mahalanobis’ D² distances (Table 3.4b)
reveal that all population pairs except Marlow and Old Windsor marina and Marlow and
Old Windsor river were significantly different from each other. Moreover, the
geographically closest pairs of populations (i.e. the two respective populations of each
location) were also genetically the closest, whereas comparison of geographically most
distant ones (i.e. Abingdon vs. Old Windsor) resulted in comparatively high FST values,
indicating genetic isolation by distance (Fig. 3.4). Although there are too few sites to test
this pattern statistically using a Mantel test to control for non-independence, discrimination
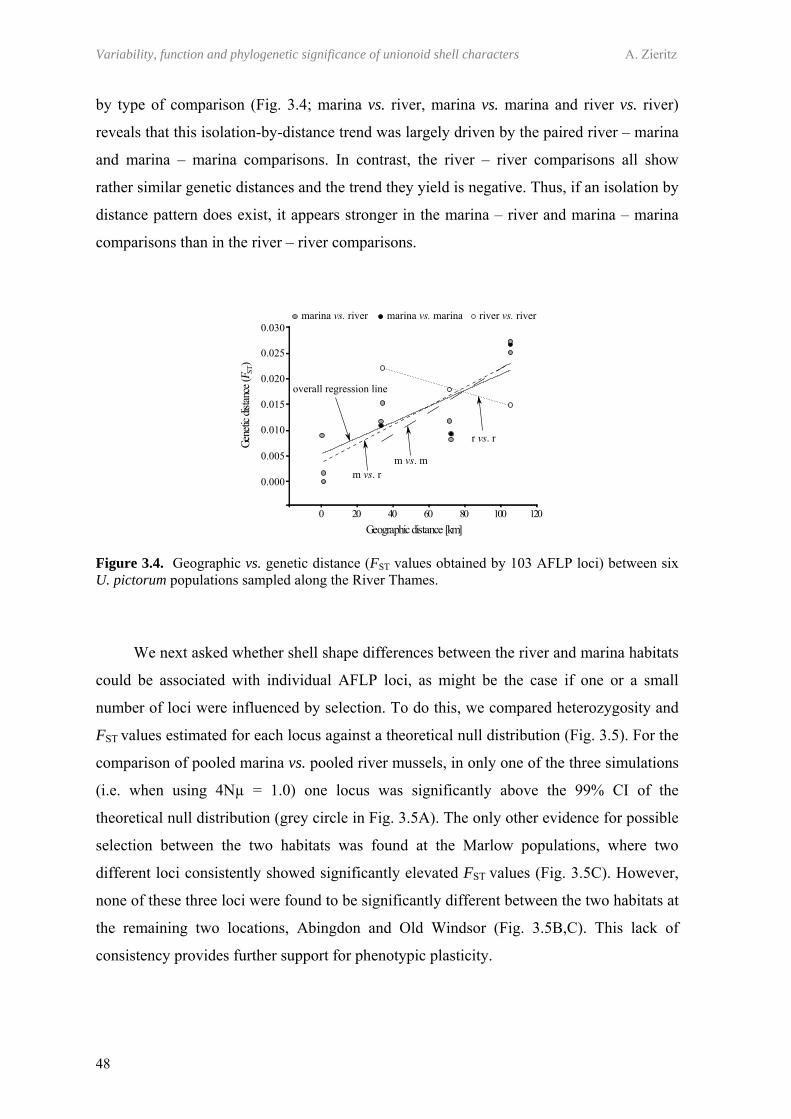
Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
48
by type of comparison (Fig. 3.4; marina vs. river, marina vs. marina and river vs. river)
reveals that this isolation-by-distance trend was largely driven by the paired river – marina
and marina – marina comparisons. In contrast, the river – river comparisons all show
rather similar genetic distances and the trend they yield is negative. Thus, if an isolation by
distance pattern does exist, it appears stronger in the marina – river and marina – marina
comparisons than in the river – river comparisons.
Figure 3.4. Geographic vs. genetic distance (FST values obtained by 103 AFLP loci) between six U. pictorum populations sampled along the River Thames.
We next asked whether shell shape differences between the river and marina habitats
could be associated with individual AFLP loci, as might be the case if one or a small
number of loci were influenced by selection. To do this, we compared heterozygosity and
FST values estimated for each locus against a theoretical null distribution (Fig. 3.5). For the
comparison of pooled marina vs. pooled river mussels, in only one of the three simulations
(i.e. when using 4Nµ = 1.0) one locus was significantly above the 99% CI of the
theoretical null distribution (grey circle in Fig. 3.5A). The only other evidence for possible
selection between the two habitats was found at the Marlow populations, where two
different loci consistently showed significantly elevated FST values (Fig. 3.5C). However,
none of these three loci were found to be significantly different between the two habitats at
the remaining two locations, Abingdon and Old Windsor (Fig. 3.5B,C). This lack of
consistency provides further support for phenotypic plasticity.
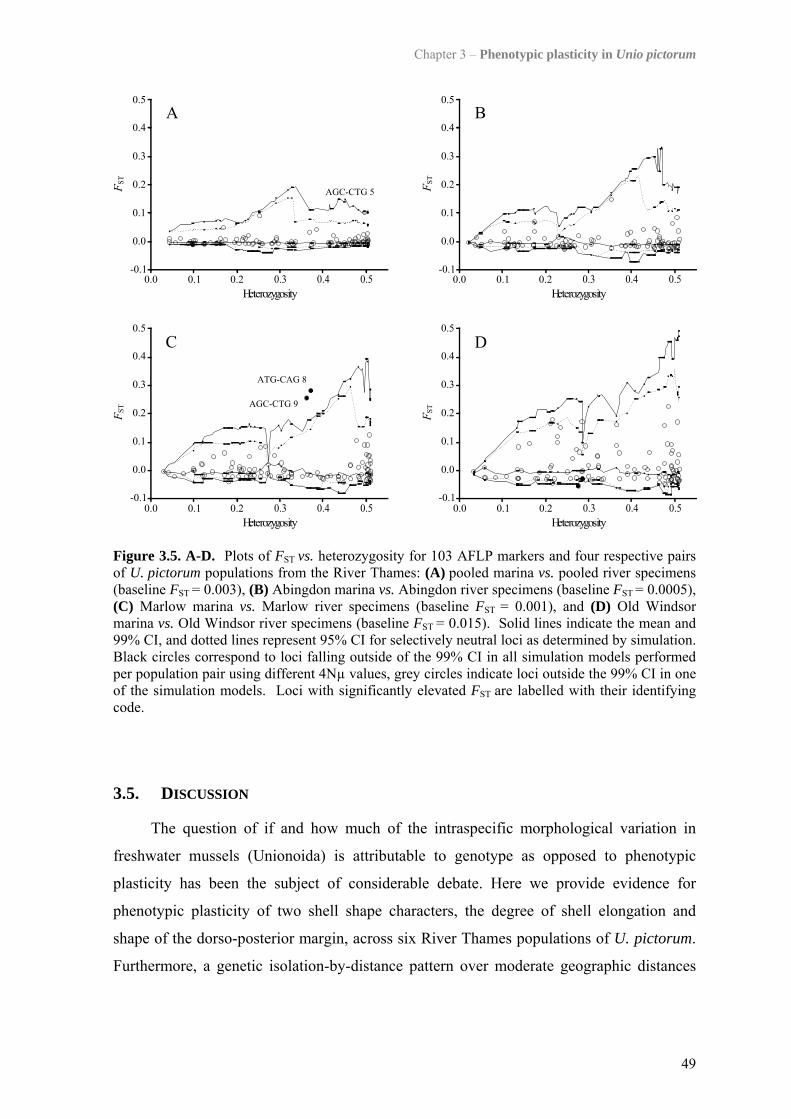
Chapter 3 – Phenotypic plasticity in Unio pictorum
49
Figure 3.5. A-D. Plots of FST vs. heterozygosity for 103 AFLP markers and four respective pairs of U. pictorum populations from the River Thames: (A) pooled marina vs. pooled river specimens (baseline FST = 0.003), (B) Abingdon marina vs. Abingdon river specimens (baseline FST = 0.0005), (C) Marlow marina vs. Marlow river specimens (baseline FST = 0.001), and (D) Old Windsor marina vs. Old Windsor river specimens (baseline FST = 0.015). Solid lines indicate the mean and 99% CI, and dotted lines represent 95% CI for selectively neutral loci as determined by simulation. Black circles correspond to loci falling outside of the 99% CI in all simulation models performed per population pair using different 4Nµ values, grey circles indicate loci outside the 99% CI in one of the simulation models. Loci with significantly elevated FST are labelled with their identifying code.
3.5. DISCUSSION
The question of if and how much of the intraspecific morphological variation in
freshwater mussels (Unionoida) is attributable to genotype as opposed to phenotypic
plasticity has been the subject of considerable debate. Here we provide evidence for
phenotypic plasticity of two shell shape characters, the degree of shell elongation and
shape of the dorso-posterior margin, across six River Thames populations of U. pictorum.
Furthermore, a genetic isolation-by-distance pattern over moderate geographic distances

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
50
(i.e. 30-100 km), indicates that the AFLP markers offered sufficient genetic resolution to
detect subtle differences in genetic composition within the populations under study.
3.5.1. GENETIC POPULATION STRUCTURE
Overall our data suggest a trend of increasing genetic distance with increasing
geographic distance over a scale of about 100 km, which is presumably driven by
appreciable numbers of larvae (glochidia) settling out close to where they were born.
However, an isolation by distance pattern was stronger across marina – marina
comparisons of sites than across river – river comparisons. One possibility is that such a
pattern could arise entirely from chance. Alternatively however, it might reflect a key
aspect of the unionoid life cycle in which the prevalent means of dispersal is thought to be
via parasitic larvae that attach to fish (Kat, 1984). U. pictorum exploits a variety of
different host fish species (Berrie & Bioze, 1985; Aldridge, 1997; Blažek & Gelnar, 2006).
In the River Thames, Berrie & Bioze (1985) observed ten fish species to be infested by
glochidia of the two British Unio species, but the three-spined stickleback (Gasterosteus
aculeatus Linnaeus, 1758) and perch (Perca fluviatilis Linnaeus, 1758) together carried
about 90% of all Unio glochidia. While both fish species are widespread and common
within the River Thames (UK Environment Agency, pers. comm.), the relatively small-
sized three-spined stickleback is more commonly associated with lentic localities such as
marinas (Maitland & Campbell, 1992) from which populations will show predominantly
localised dispersal (Bolnick et al., 2009). Localised dispersal is also implied from a study
of lentic (lake) perch (Bergek & Björklund, 2007) where cryptic barriers to dispersal
restricted gene flow. Conversely, host fishes, especially perch, within the river can be
expected to be both actively and passively more mobile (Tudorache et al., 2008). If the
dominant host fishes within the marinas show relatively low dispersal compared with the
river, this would explain the weaker isolation by distance observed for riverine mussel
samples, with their vagile hosts dispersing excysted juvenile mussels some distance from
their parents. Thus, the striking difference in IBD trends between marina and river
populations demonstrates that the evolutionary consequences of a parasite-host interaction
(genetic divergence of parasite populations) could be dependent on the habitat where the
interaction takes place.
A pattern of genetic isolation-by-distance has been observed in other species of
freshwater mussel, for example, the North American species Quadrula quadrula

Chapter 3 – Phenotypic plasticity in Unio pictorum
51
Rafinesque, 1820 (Berg et al., 1998), but is by no means universal. Thus, other studies
have failed to find evidence of isolation by distance across similar or even greater
geographic distances in, for example, Lexingtonia dolabelloides (Lea, 1840) (Grobler et
al., 2006), M. margaritifera (Machordom et al., 2003; Bouza et al., 2007) and Amblema
plicata (Say, 1817) (Elderkin et al., 2007). Reasons for such differences between species
and/or water bodies are likely to be complicated and, besides differences in the vagility of
their fish hosts, probably include confounding patterns of pre- and postglacial colonisation
that may “overshadow” recent dispersal events and anthropogenic influences relating to
fish stocking measures.
3.5.2. PHENOTYPIC PLASTICITY OF SHELL FORM
By comparing geographically distant, replicate population pairs of two respective
habitats, the present study has shown that individuals sampled from the same habitat were
morphologically more similar but genetically more distinct from one another compared
with those from different but geographically adjacent habitats. This suggests that the
degree of relative elongation, i.e. shell height to length ratio, and the shape of the dorso-
posterior margin, two shell characters that are strongly and consistently different between
marina and river sites, are unlikely to be under genetic control in U. pictorum. Further
evidence for phenotypic plasticity of shell shape in U. pictorum is provided by the fact that
not a single locus could be identified that showed convincing evidence of natural selection
among the different habitats we sampled, and thus, could potentially be associated with
shell shape differences of ecomorphotypes.
Our findings support observations by Hinch, Bailey & Green (1986), who, on the
basis of reciprocal transplant experiments, found that relative shell height to length growth
in the North American unionoid Elliptio complanata (Lightfoot, 1786) was determined by
phenotypic plasticity. Besides providing molecular evidence for the same pattern in the
European species U. pictorum, we also show that the shape of the dorso-posterior margin,
which is a more consistent ecophenotypic shell character in unionoids than relative shell
elongation (Chapter 2; Zieritz & Aldridge, 2009), is also probably not under genetic
control. This knowledge fills a gap in the scarce molecular evidence for phenotypic
plasticity of shell morphology in unionoids, which has so far consisted mainly of studies
reporting a lack of match of intraspecific morphological and genetic patterns (Buhay et al.,
2002; Machordom et al., 2003; Molina, 2004; Geist & Kuehn, 2005). However, since most

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
52
of these previous studies were not primarily focused on determining the basis of
phenotypic plasticity, they failed to include a joint analysis of morphology, genetics and
geographic distance of populations, and therefore generally fail to make a convincing case.
Evidence for phenotypic plasticity of shell morphology has been found in a number
of other mollusc taxa that occupy heterogenous habitats and have high dispersal potential,
most notably marine gastropods with planktonic larvae (e.g. Littorina striata (King &
Broderip, 1832), de Wolf, Backeljau & Verhagen, 1998; Nodilittorina australis (Gray,
1826), Yeap, Black & Johnson, 2001; Nacella concinna (Strebel, 1908), Hoffman et al.,
2010). Conversely, local adaptation in other gastropod species might be capable of driving
genetic divergence even within continuous populations (e.g. Tregenza & Butlin, 1999;
Doebeli & Dieckmann, 2003). Potential examples for such ecological nonallopatric
speciation include Littorina saxatilis (Olivi, 1792) (Johannesson & Johannesson, 1996;
Panova, Hollander & Johannesson, 2006; Conde-Padín et al., 2007) and Nucella lapillus
(Linnaeus, 1758) (Guerra-Varela et al., 2009). However, extensive gene flow is predicted
to prevent local adaptation (Johannesson, 2003) so that these species are usually direct
developers with restricted larval dispersal. Moreover, despite some evidence for divergent
selection acting on marine broadcast spawners with long-lived pelagic larvae (Luttikhuizen
et al., 2003; de Aranzamendi et al., 2008), the dispersal of unionoid larvae by their fish
hosts apparently leads to a relatively even redistribution of larval genotypes across
different habitats. Consequently, morphological adjustments are possible only through a
plastic phenotype.
The unionoid shell ecophenotypes observed might have adaptive significance which,
along with their possible functional morphologies, are discussed in detail in Chapter 2,
Zieritz & Aldridge (2009) and references therein. In short, hydrological parameters such as
water movement (e.g. mean and/or maximum current velocities) are probably the main
factors determining the sagittal shape of these unionoids’ shells. Dorso-posterior arching
of river morphotypes, for example, results in heavier weight and increased pedal gape of
the shell, allowing the foot to extend further into the sediment. These features would
increase initial probing force, anchorage, and the stability of the bivalve when subject to
lifting forces resulting from turbulent water (Eagar, 1978). Alternatively, differences in
shell shapes could have no adaptive value whatsoever but be caused by a non-functional
reaction of the mussel to the environment (e.g. via distortion of the shell secreting mantle
margin as a result of water movement). If the observed phenotypic plasticity of shell form

Chapter 3 – Phenotypic plasticity in Unio pictorum
53
is adaptive (i.e. results in an improvement in growth, survival or reproduction; Stearns,
1989), nonadaptive or even maladaptive remains to be tested.
Finally, our study potentially has implications for mussel classification. Previous
work has debated the extent to which morphotypes reflect phenotypic plasticity versus
genuine differentiation leading to speciation (e.g. Mulvey et al., 1997; Serb, Buhay &
Lydeard, 2003). This problem is exacerbated by the problem that contrasting morphologies
were often sampled from geographically distant sites, increasing the chance that
morphological and genetic isolation will be correlated (Davis et al., 1981; Davis, 1983,
1984; Baker et al., 2003). A further issue is that, over and above any phenotypic plasticity,
some molluscan shell characteristics are known to be under genetic control (e.g. smoothed
versus ribbed shell sculpture in the gastropod Oncomelia hupensis Gredler, 1881; Davis &
Ruff, 1974). Although limited in range, our study illustrates the value of looking at
genetics and morphology over a range of geographic scales in order to interpret which
factors are most important in driving divergence and provides what we believe is the first
good evidence for phenotypic plasticity of shell shape in U. pictorum. Further similar
studies on other unionoid species using similar markers will be needed in order to establish
the generality of what we have found.


CHAPTER 4
SEXUAL, HABITAT-CONSTRAINED AND PARASITE-INDUCED DIMORPHISM IN THE
SHELL OF A FRESHWATER MUSSEL (ANODONTA ANATINA, UNIONIDAE)

“I’m not obese, it’s all mussel!”
Slogan by Holly Barclay

57
CHAPTER 4
SEXUAL, HABITAT-CONSTRAINED AND PARASITE-INDUCED DIMORPHISM IN THE SHELL OF A FRESHWATER MUSSEL
(ANODONTA ANATINA, UNIONIDAE)
4.1. ABSTRACT
We investigated to what extent sex, trematode infection and habitat effects determine
shell dimorphisms in five populations of the freshwater mussel Anodonta anatina.
Significant sexual shell width dimorphism was displayed by three of the five study
populations. Here, shells of females were significantly wider than males, probably as a
result of altered shell growth to accommodate marsupial gills. In two of these populations
female shells were additionally significantly thinner than those of males, which could be a
result of resource depletion by offspring production. No significant dimorphic patterns
were found in two other populations of the same species. The different degree of sexual
dimorphism between populations may reflect the overarching effect of habitat on
morphology. Thus, populations in the most favourable habitat might exhibit faster growth
rates, attain larger maximum sizes and produce more offspring, which results in more
swollen gills and consequently more inflated shells of gravid females compared to less
fecund populations. None of the populations showed any evidence for sexual dimorphism
in any of the shell characters ‘overall size’, ‘growth rate’, ‘sagittal shape’ and ‘density’. In
addition to sexual dimorphisms, infestation by trematode parasites significantly altered
sagittal and lateral shell shape of A. anatina in one of the populations, with infected
specimens being wider and more elongated.
4.2. INTRODUCTION
The interpretation of diverse patterns in the morphology of animal species has long
been a matter of scientific interest. Understanding which factors determine morphological
characteristics of individuals within a species can provide useful information concerning
the biology and ecology of species, populations or even individuals. Taxa producing hard
parts that persist in sedimentary deposits can additionally be used to reconstruct
characteristics of ancient species, populations and palaeoenvironments. Shell morphology

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
58
of freshwater mussels (Unionoida) could be of particular value in these respects.
Unionoids are common and widespread, inhabiting all continents except the Antarctic
(Graf & Cummings, 2006b). Their oldest known representatives stem from the Triassic
(Watters, 2001), and their typically high degree of intraspecific variation in shell
morphology potentially contains a range of information about an individual’s biological
and ecological characteristics. However, due to a lack of understanding of which factors
cause which trends in shell form, such intraspecific morphological differences in unionoids
are rarely used for redrawing characteristics of both the present and the past.
Probably the best studied, though still insufficiently understood, type of intraspecific
variation in unionoid shell morphology is that associated with habitat conditions (i.e.
ecophenotypic) (see Chapter 2; Chapter 3; Zieritz & Aldridge, 2009; Zieritz et al., 2010;
and references therein). Here we investigate two other, largely ignored sources of
intraspecific variation in freshwater mussels; sexual dimorphism and parasite induced
change in shell morphology.
Numerous striking examples of sexual dimorphism are known from the animal
kingdom and in particular mammals (e.g. Thom, Harrington & Macdonald, 2004), birds
(e.g. Owens & Hartley, 1998), fish (e.g. Kitano, Mori & Peichel, 2007), reptiles (e.g.
Brecko et al., 2008) and insects (e.g. Stillwell & Fox, 2007). All these are examples for
well developed sexual differences in external structures and are caused by (1) preference
of one sex for particular traits of the other sex, (2) intra-sexual competition (sexual
selection) or (3) niche divergence between the sexes (Shine, 1989; Parker, 1992;
Andersson, 1994). Though none of these three forms of sexual dimorphism would be
expected in broadcast spawners such as freshwater mussels, sexual dimorphism in this
group may arise as a direct result of differences in their sexes’ internal anatomy.
Protandric or protogynous consecutive hermaphroditism results in size dimorphism
of sexes in several marine bivalve families (e.g. Veneridae (Loosanoff, 1936), Astartidae
(Saleuddin, 1965), Galeommatidae (Morton, 1976)). While hermaphroditism is rare in
unionoids (e.g. Bloomer, 1934; McIvor & Aldridge, 2007), protandry and consequential
sexually dimorphic size has been documented in the unionid Elliptio complanata
(Lightfoot, 1786) (Downing et al., 1989).
Sexual dimorphism may also arise from the unique reproductive mode of unionoids.
Their larvae mature in certain parts of the female gills (marsupia), and in some of these
cases the resulting swelling of gravid marsupia is believed to alter growth in female shells.
A particularly pronounced sexual shape dimorphism is exhibited by most members of the

Chapter 4 – Shell dimorphism in Anodonta anatina
59
tribe Lampsilini (subfamily Ambleminae; phylogeny after Graf & Cummings, 2006b) (e.g.
Kirtland, 1834; Heard & Guckert, 1970; Kotrla & James, 1987). Lampsilines are unique in
using only the posterior portions of their outer gill demibranchs as marsupia (heterogenous
condition), which enlarge both laterally and ventrally when gravid, resulting in female
shells with more evenly rounded posterior margins and more inflated ventro-posterior
regions than males. More subtle patterns of shape dimorphism have been found in species
with gills marsupial across their entire lengths (homogenous condition) (e.g. Kirtland,
1834; Heard & Guckert, 1970).
European freshwater mussels of the unionid subfamily Unioninae have homogenous
marsupia and are generally believed not to display any considerable sexual shell
dimorphism. The occasional observations on sex-related differences in shell morphology,
and in particular shell inflation (width), are mostly anecdotal (e.g. Noll, 1869; Hazay,
1881; Sterki, 1903). Brander (1954) described male shells of Pseudanodonta complanata
(Rossmässler, 1835) as more elongated and ovally shaped than the more trapezoid females.
Contrary to this, Siebold (1837) found females of Anodonta anatina (Linnaeus, 1758) and
Anodonta cygnea (Linnaeus, 1758) to be more elongated than males. Other unionid shell
characters that have been considered to be sexually dimorphic include shell colour (Israel,
1910), thickness (Jass & Glenn, 2004) and composition of shell material (Cameron,
Cameron & Paterson, 1979).
Infection by parasites is another factor that may cause alteration in shell size and
shape. Freshwater mussels are known to host a number of parasites including copepods
(e.g. Saarinen & Taskinen, 2004), mites (e.g. Mitchell & Pitchford, 1953), chironomids
(e.g. Forsyth & McCallum, 1978), fish (e.g. Aldridge, 1999a) and trematodes (e.g. Jokela,
Uotila & Taskinen, 1993). The effects of parasitism on unionoid morphology are
understudied but shell growth in A. anatina has been shown to be slowed during heavy
infection by the bucephalid trematode Rhipidocotyle fennica Gibson, Valtonen et
Taskinen, 1992 (Taskinen & Valtonen, 1995). Additionally, trematodes can be associated
with reduced reproductive output (Gangloff, Lenertz & Feminella, 2008), which may
affect the development of dimorphic characters.
This study represents the first attempt to investigate if and to what extent sex,
trematode infection and indirect habitat effects induce shell dimorphisms in five
populations of the European unionoid A. anatina. A modern morphometric method
(Fourier shape analysis) was employed to describe shell shape. Morphological characters
under study were (1) size and growth rates, (2) relative elongation, (3) overall sagittal shell

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
60
shape, (4) relative width, (5) thickness and (6) density of the shell. Possible reasons for the
dimorphic patterns observed and why certain sexual differences are present in some but
not in other populations are discussed.
4.3. MATERIALS AND METHODS
4.3.1. SAMPLING, SEX DETERMINATION AND TREMATODE INFECTION
From May 2007 to September 2009, five A. anatina populations were investigated
for dimorphic patterns in shell morphology (Fig. 4.1). To test if habitat conditions
contributed to dimorphic trends, we selected five sites that differed in both environmental
conditions and characteristics of their A. anatina populations (such as population densities
and growth rates). The River Thames in the stretch studied is a typical UK lowland lotic
system whereas the marina sites are lentic habitats with almost no flow. Quidenham Mere
is a eutrophic lake system.
Figure 4.1. Locations of the five UK Anodonta anatina populations of the study. Abbreviations: Q, Quidenham Mere; AM, Abingdon Boat Marina (River Thames); AR, River Thames, Abingdon/Culham; CM, Thames & Kennet Marina (River Thames); CR, River Thames, Caversham. Map © Crown Copyright/database right 2009. An Ordnance Survey/EDINA supplied service.
A. anatina abundances at each site were estimated by counting the number of
individuals present in 15 replicate dredges (approx. 5 m in length, dredge aperture 46 cm x
21 cm x 7.5 cm, mesh size = 2.5 cm). In addition, 15-40 individuals per population,
depending on A. anatina abundance at the site, were sampled by hand and taken to the
laboratory for further morphological investigation.

Chapter 4 – Shell dimorphism in Anodonta anatina
61
In the laboratory, the sex of each individual was determined by macroscopic
inspection of the demibranchs for swelling of the outer gills, indicating
female/hermaphroditic sex, and examination of gonadal fluid under a light microscope.
Mussels presenting oogonia, oocytes or larvae (glochidia) of any developmental stage
were assigned as females; occurrence of spermatozoa and/or sperm-morulae indicated
male sex (Heard, 1975). Individuals exhibiting both male and female gonad characteristics
were classified as hermaphrodites. State of trematode parasitism (‘parasitised’ or ‘not
parasitised’) was also determined by examining gonadal fluid under a light microscope. An
individual mussel was considered parasitised if digenean trematode sporocysts and/or
cercariae were present in one or more of three gonadal samples.
4.3.2. SIZE AND GROWTH RATES
Age of 15-35 specimens per site was estimated by counting annual winter rings
(Aldridge, 1999b). Maximum length at each annulus was measured with a digital calliper
to ± 0.1 mm, and length-age plots were produced for each population by calculating the
mean shell length at each year. Growth parameters were determined using the Walford plot
model (Walford, 1946) assuming von Bertalanffy growth curves.
Differences between populations in size and growth rates were statistically tested
using the measurements of shell length (L; across the maximum sagittal diameter) and
shell length at the third annulus (L3) for each aged mussel. The same two measurements
were used to test for sexual size dimorphism (consecutive hermaphroditism) and
differences in growth rates between sexes within each of the five populations.
4.3.3. SHELL SHAPE, THICKNESS AND DENSITY
For analysis of shell shape the three shell dimensions length (L), height (H;
maximum sagittal diameter perpendicular to L) and width (W; maximum lateral diameter)
were measured to ± 0.1 mm using a digital calliper. Using these measurements, sagittal
area (SaA) and surface area (SuA) of each mussel were estimated by applying the formulas
for an ellipse and ellipsoid, respectively. Shell dry weight (DW) and shell volume (V;
measured as water displaced by the shell using method of Rodhouse (1977)) were recorded
to ± 0.01 cm³. These parameters were subsequently used in general linear models (GLMs;
Minitab 15 for Windows) to test for statistical significance of sexual dimorphism in degree
of shell elongation, inflation, thickness and density. For this purpose, GLMs were carried

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
62
out on H, W, V and DW, fitting L, SaA1/2, SuA and V as covariates, respectively, population
as a factor with five levels, sex as a factor with two levels, and the covariate*sex,
covariate*population and sex*population interaction factors. Insignificant factors were
then sequentially dropped from the model (Crawley, 2002). For those parameters that
proved to be significantly influenced by sex, the sex*covariate and/or the sex*population
interaction factor, GLMs were then carried out for each population separately in order to
elucidate which population(s) exhibited significant sexual dimorphism in the respective
shell character.
For reasons explained in the results section, trematode parasite-induced alteration in
shell morphology was tested only in the Quidenham Mere population regarding all four
characters as described above. For this purpose, GLMs were carried out including the
respective covariate, both trematode (parasitism) and sex as factors with two levels, and
the covariate*sex-, covariate*trematodes- and sex*trematodes-interaction factors.
To examine if sexually dimorphic and/or parasite-induced patterns were present with
respect to overall sagittal shape of the shell, we carried out Fourier shape analyses as
developed and explained by Crampton & Haines (1996) on two groups of populations: (1)
all five populations combined and (2) the Quidenham Mere population alone. This method
decomposes xy-coordinates of an outline into a number of harmonics, each of which is in
turn explained by two respective Fourier coefficients, which can be statistically treated as
any usual variable. For this analysis digital photographs of specimens were taken, and
outlines were digitised using the program IMAGEJ (Rasband, 2008). The xy-coordinates
of digitised outlines were then subjected to fast Fourier transformation using program
HANGLE (Crampton & Haines, 2007) applying a smoothing normalisation of 20 to
eliminate high-frequency pixel noise. Preliminary analysis indicated that the first ten
harmonics described the outlines with sufficiently high precision. Discarding of the first
harmonic, which does not contain any shape information, resulted in a set of 18 Fourier
coefficients per individual. After rotating outlines to maximum overlap by program
HMATCH (Crampton & Haines, 2007), principal component analysis (PCA) was
performed on the 18 Fourier coefficients using program PAST (Hammer & Harper, 2006).
Number of significant principal components was determined by the broken stick model of
the scree plot. Subsequently, GLMs were carried out on significant PC axes obtained by
PCA on (1) all five populations including factors population and sex and the
sex*population interaction factor, and (2) Quidenham Mere population including factors
sex and trematodes, and their interaction.
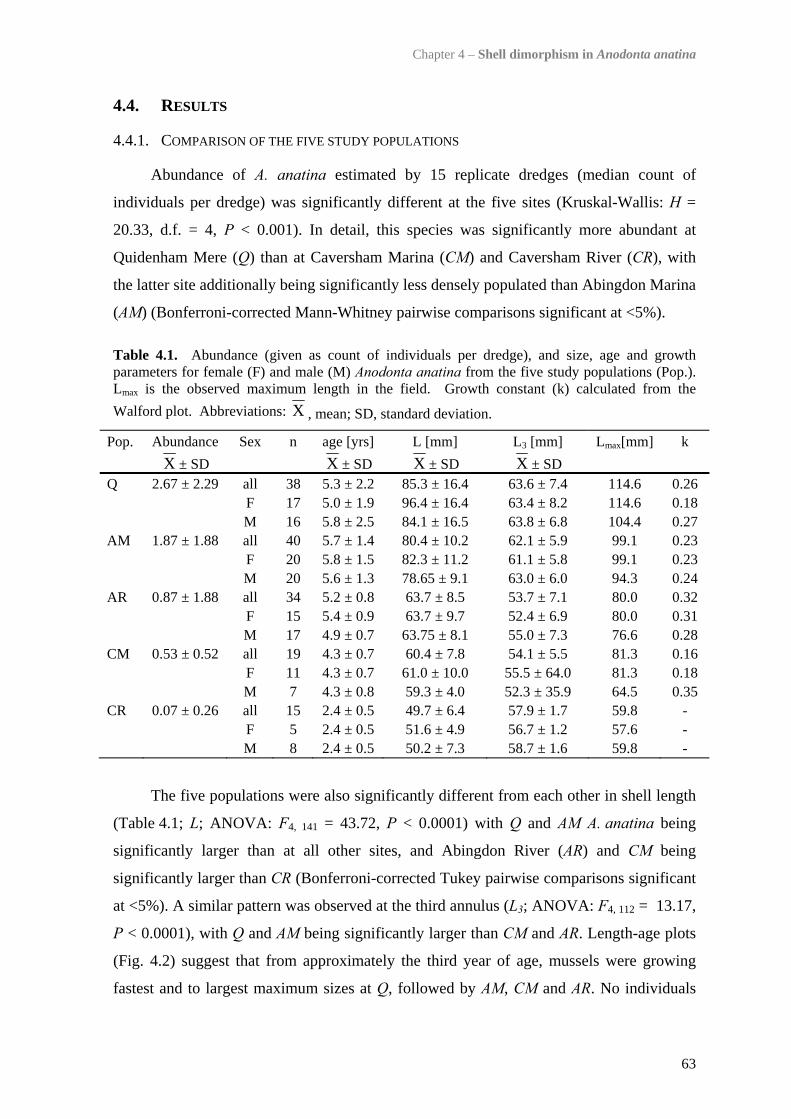
Chapter 4 – Shell dimorphism in Anodonta anatina
63
4.4. RESULTS
4.4.1. COMPARISON OF THE FIVE STUDY POPULATIONS
Abundance of A. anatina estimated by 15 replicate dredges (median count of
individuals per dredge) was significantly different at the five sites (Kruskal-Wallis: H =
20.33, d.f. = 4, P < 0.001). In detail, this species was significantly more abundant at
Quidenham Mere (Q) than at Caversham Marina (CM) and Caversham River (CR), with
the latter site additionally being significantly less densely populated than Abingdon Marina
(AM) (Bonferroni-corrected Mann-Whitney pairwise comparisons significant at <5%).
Table 4.1. Abundance (given as count of individuals per dredge), and size, age and growth parameters for female (F) and male (M) Anodonta anatina from the five study populations (Pop.). Lmax is the observed maximum length in the field. Growth constant (k) calculated from the Walford plot. Abbreviations: X , mean; SD, standard deviation.
Pop. Abundance Sex n age [yrs] L [mm] L3 [mm] Lmax[mm] k X ± SD X ± SD X ± SD X ± SD Q 2.67 ± 2.29 all 38 5.3 ± 2.2 85.3 ± 16.4 63.6 ± 7.4 114.6 0.26 F 17 5.0 ± 1.9 96.4 ± 16.4 63.4 ± 8.2 114.6 0.18 M 16 5.8 ± 2.5 84.1 ± 16.5 63.8 ± 6.8 104.4 0.27 AM 1.87 ± 1.88 all 40 5.7 ± 1.4 80.4 ± 10.2 62.1 ± 5.9 99.1 0.23 F 20 5.8 ± 1.5 82.3 ± 11.2 61.1 ± 5.8 99.1 0.23 M 20 5.6 ± 1.3 78.65 ± 9.1 63.0 ± 6.0 94.3 0.24 AR 0.87 ± 1.88 all 34 5.2 ± 0.8 63.7 ± 8.5 53.7 ± 7.1 80.0 0.32 F 15 5.4 ± 0.9 63.7 ± 9.7 52.4 ± 6.9 80.0 0.31 M 17 4.9 ± 0.7 63.75 ± 8.1 55.0 ± 7.3 76.6 0.28 CM 0.53 ± 0.52 all 19 4.3 ± 0.7 60.4 ± 7.8 54.1 ± 5.5 81.3 0.16 F 11 4.3 ± 0.7 61.0 ± 10.0 55.5 ± 64.0 81.3 0.18 M 7 4.3 ± 0.8 59.3 ± 4.0 52.3 ± 35.9 64.5 0.35 CR 0.07 ± 0.26 all 15 2.4 ± 0.5 49.7 ± 6.4 57.9 ± 1.7 59.8 - F 5 2.4 ± 0.5 51.6 ± 4.9 56.7 ± 1.2 57.6 - M 8 2.4 ± 0.5 50.2 ± 7.3 58.7 ± 1.6 59.8 -
The five populations were also significantly different from each other in shell length
(Table 4.1; L; ANOVA: F4, 141 = 43.72, P < 0.0001) with Q and AM A. anatina being
significantly larger than at all other sites, and Abingdon River (AR) and CM being
significantly larger than CR (Bonferroni-corrected Tukey pairwise comparisons significant
at <5%). A similar pattern was observed at the third annulus (L3; ANOVA: F4, 112 = 13.17,
P < 0.0001), with Q and AM being significantly larger than CM and AR. Length-age plots
(Fig. 4.2) suggest that from approximately the third year of age, mussels were growing
fastest and to largest maximum sizes at Q, followed by AM, CM and AR. No individuals
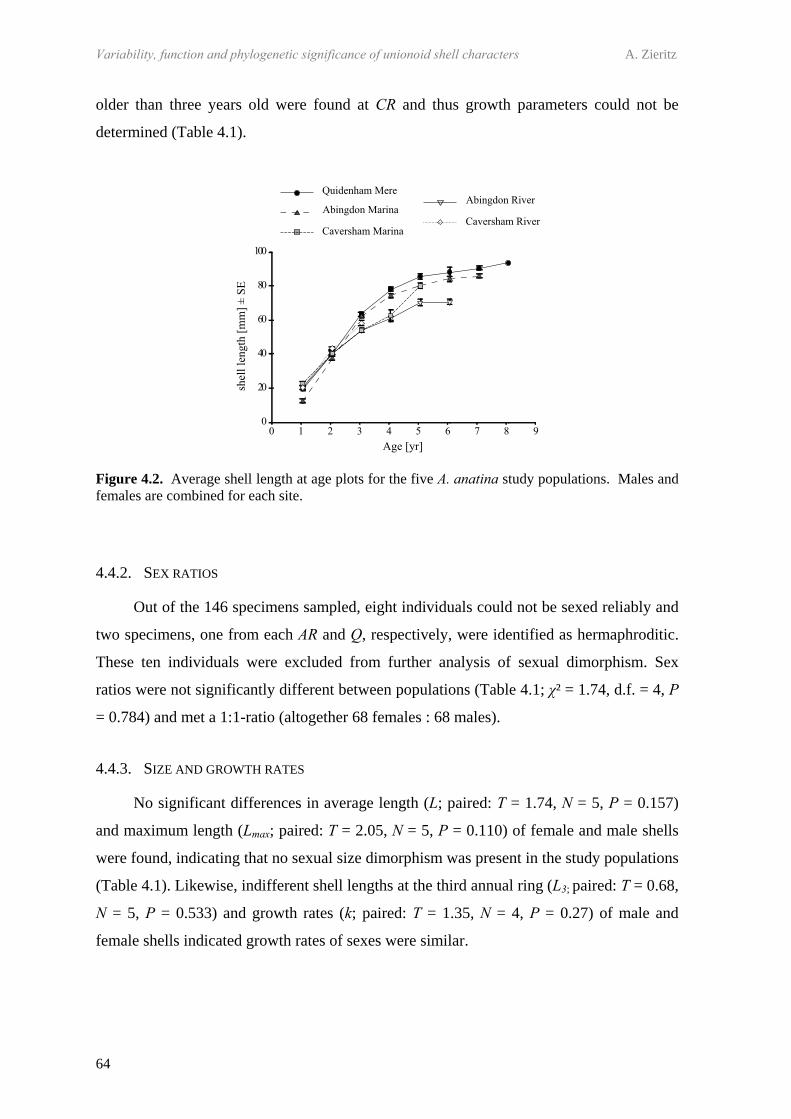
Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
64
older than three years old were found at CR and thus growth parameters could not be
determined (Table 4.1).
Figure 4.2. Average shell length at age plots for the five A. anatina study populations. Males and females are combined for each site.
4.4.2. SEX RATIOS
Out of the 146 specimens sampled, eight individuals could not be sexed reliably and
two specimens, one from each AR and Q, respectively, were identified as hermaphroditic.
These ten individuals were excluded from further analysis of sexual dimorphism. Sex
ratios were not significantly different between populations (Table 4.1; χ² = 1.74, d.f. = 4, P
= 0.784) and met a 1:1-ratio (altogether 68 females : 68 males).
4.4.3. SIZE AND GROWTH RATES
No significant differences in average length (L; paired: T = 1.74, N = 5, P = 0.157)
and maximum length (Lmax; paired: T = 2.05, N = 5, P = 0.110) of female and male shells
were found, indicating that no sexual size dimorphism was present in the study populations
(Table 4.1). Likewise, indifferent shell lengths at the third annual ring (L3; paired: T = 0.68,
N = 5, P = 0.533) and growth rates (k; paired: T = 1.35, N = 4, P = 0.27) of male and
female shells indicated growth rates of sexes were similar.
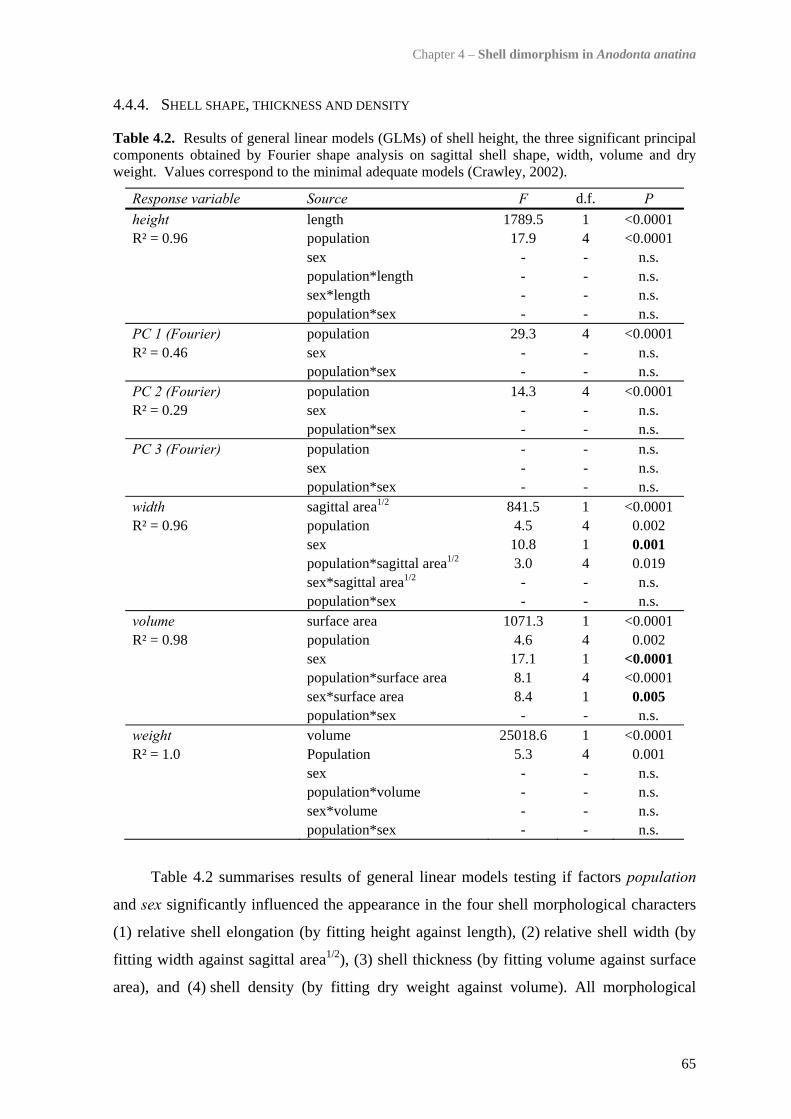
Chapter 4 – Shell dimorphism in Anodonta anatina
65
4.4.4. SHELL SHAPE, THICKNESS AND DENSITY
Table 4.2. Results of general linear models (GLMs) of shell height, the three significant principal components obtained by Fourier shape analysis on sagittal shell shape, width, volume and dry weight. Values correspond to the minimal adequate models (Crawley, 2002).
Response variable Source F d.f. P height length 1789.5 1 <0.0001 R² = 0.96 population 17.9 4 <0.0001 sex - - n.s. population*length - - n.s. sex*length - - n.s. population*sex - - n.s. PC 1 (Fourier) population 29.3 4 <0.0001 R² = 0.46 sex - - n.s. population*sex - - n.s. PC 2 (Fourier) population 14.3 4 <0.0001 R² = 0.29 sex - - n.s. population*sex - - n.s. PC 3 (Fourier) population - - n.s. sex - - n.s. population*sex - - n.s. width sagittal area1/2 841.5 1 <0.0001 R² = 0.96 population 4.5 4 0.002 sex 10.8 1 0.001 population*sagittal area1/2 3.0 4 0.019 sex*sagittal area1/2 - - n.s. population*sex - - n.s. volume surface area 1071.3 1 <0.0001 R² = 0.98 population 4.6 4 0.002 sex 17.1 1 <0.0001 population*surface area 8.1 4 <0.0001 sex*surface area 8.4 1 0.005 population*sex - - n.s. weight volume 25018.6 1 <0.0001 R² = 1.0 Population 5.3 4 0.001 sex - - n.s. population*volume - - n.s. sex*volume - - n.s. population*sex - - n.s.
Table 4.2 summarises results of general linear models testing if factors population
and sex significantly influenced the appearance in the four shell morphological characters
(1) relative shell elongation (by fitting height against length), (2) relative shell width (by
fitting width against sagittal area1/2), (3) shell thickness (by fitting volume against surface
area), and (4) shell density (by fitting dry weight against volume). All morphological

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
66
characters under study, including overall sagittal shape of the shell as determined by
Fourier shape analysis, showed significant differences between populations (Table 4.2,
Fig. 4.3).
Figure 4.3. Scatterplots of PC1 and PC2 values obtained by principal component analyses on 18 Fourier coefficients of five A. anatina populations. Synthetic outlines of extreme sagittal shapes obtained using method of Haines & Crampton (2000), displayed with anterior margin facing to the right and dorsal margin to the head of the page. Values for the third significant principal component, explaining 10.4% of the variation in shell outline, are not displayed.
Factor sex was retained as a significant predictor only for relative shell width and
thickness (Table 4.2), which were therefore analysed for each population separately.
Though females of all populations tended to have relatively wider shells than males
(Fig. 4.4), GLMs revealed that this sexual difference in relative shell width was significant
only in populations AM and CR (Table 4.3). Shell thickness, on the other hand, was
significantly influenced by sex only in populations Q and AM (Table 4.3), with females
exhibiting thinner shells than males (Fig. 4.5).
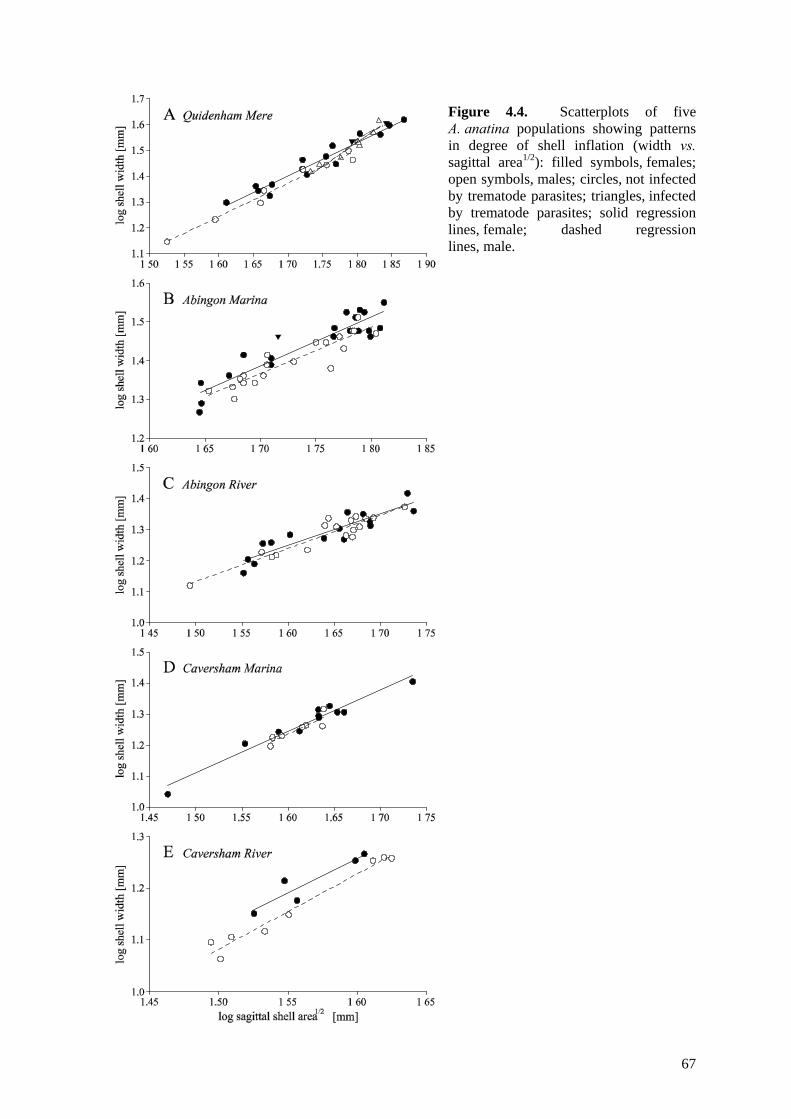
67
Figure 4.4. Scatterplots of five A. anatina populations showing patterns in degree of shell inflation (width vs. sagittal area1/2): filled symbols, females; open symbols, males; circles, not infected by trematode parasites; triangles, infected by trematode parasites; solid regression lines, female; dashed regression lines, male.
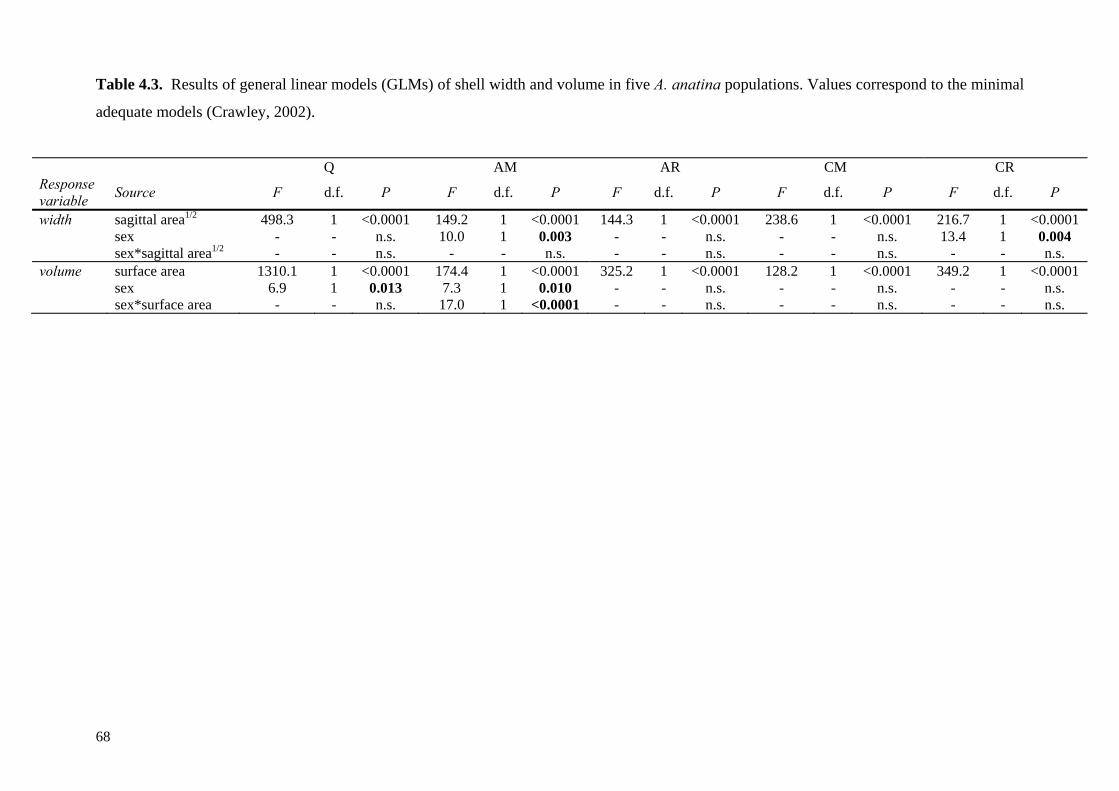
68
Table 4.3. Results of general linear models (GLMs) of shell width and volume in five A. anatina populations. Values correspond to the minimal
adequate models (Crawley, 2002).
Q AM AR CM CR Response variable Source F d.f. P F d.f. P F d.f. P F d.f. P F d.f. P
width sagittal area1/2 498.3 1 <0.0001 149.2 1 <0.0001 144.3 1 <0.0001 238.6 1 <0.0001 216.7 1 <0.0001 sex - - n.s. 10.0 1 0.003 - - n.s. - - n.s. 13.4 1 0.004 sex*sagittal area1/2 - - n.s. - - n.s. - - n.s. - - n.s. - - n.s. volume surface area 1310.1 1 <0.0001 174.4 1 <0.0001 325.2 1 <0.0001 128.2 1 <0.0001 349.2 1 <0.0001 sex 6.9 1 0.013 7.3 1 0.010 - - n.s. - - n.s. - - n.s. sex*surface area - - n.s. 17.0 1 <0.0001 - - n.s. - - n.s. - - n.s.

Chapter 4 – Shell dimorphism in Anodonta anatina
69
Figure 4.5. Scatterplots of five A. anatina populations showing patterns in shell thickness (volume vs. surface area): filled circles and solid regression lines, females; open circles and dashed regression lines, males.
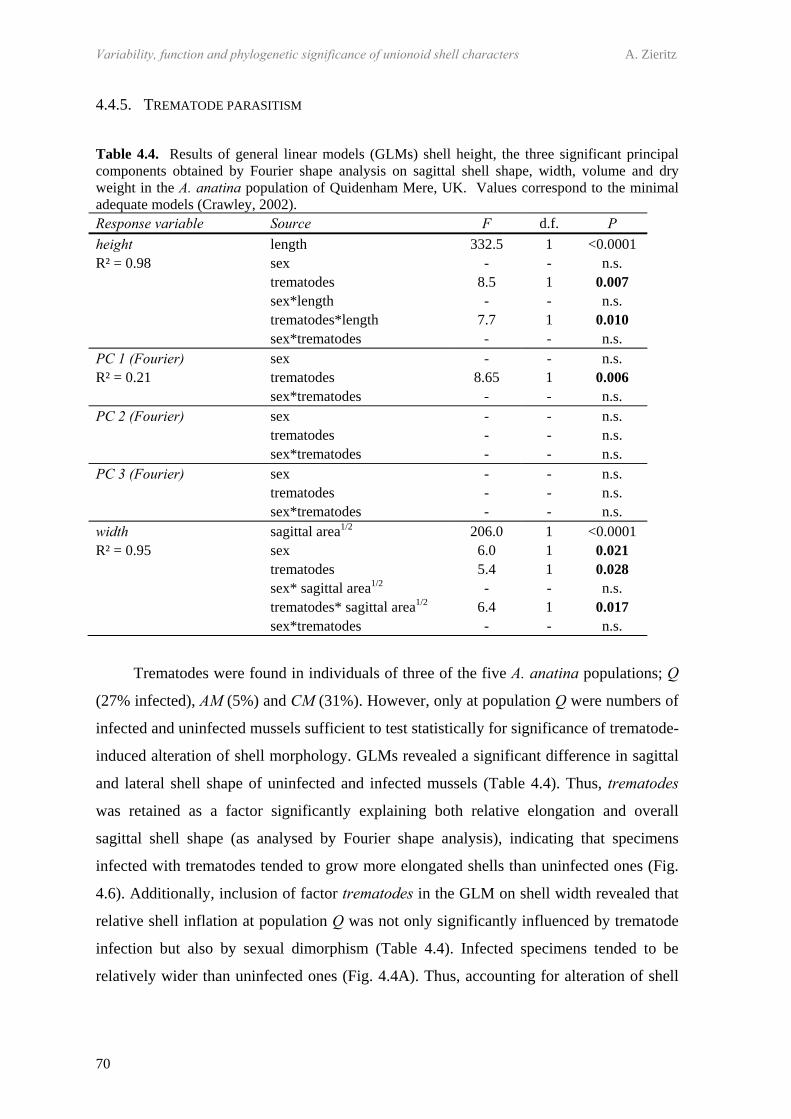
Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
70
4.4.5. TREMATODE PARASITISM
Table 4.4. Results of general linear models (GLMs) shell height, the three significant principal components obtained by Fourier shape analysis on sagittal shell shape, width, volume and dry weight in the A. anatina population of Quidenham Mere, UK. Values correspond to the minimal adequate models (Crawley, 2002). Response variable Source F d.f. P height length 332.5 1 <0.0001 R² = 0.98 sex - - n.s. trematodes 8.5 1 0.007 sex*length - - n.s. trematodes*length 7.7 1 0.010 sex*trematodes - - n.s. PC 1 (Fourier) sex - - n.s. R² = 0.21 trematodes 8.65 1 0.006 sex*trematodes - - n.s. PC 2 (Fourier) sex - - n.s. trematodes - - n.s. sex*trematodes - - n.s. PC 3 (Fourier) sex - - n.s. trematodes - - n.s. sex*trematodes - - n.s. width sagittal area1/2 206.0 1 <0.0001 R² = 0.95 sex 6.0 1 0.021 trematodes 5.4 1 0.028 sex* sagittal area1/2 - - n.s. trematodes* sagittal area1/2 6.4 1 0.017 sex*trematodes - - n.s.
Trematodes were found in individuals of three of the five A. anatina populations; Q
(27% infected), AM (5%) and CM (31%). However, only at population Q were numbers of
infected and uninfected mussels sufficient to test statistically for significance of trematode-
induced alteration of shell morphology. GLMs revealed a significant difference in sagittal
and lateral shell shape of uninfected and infected mussels (Table 4.4). Thus, trematodes
was retained as a factor significantly explaining both relative elongation and overall
sagittal shell shape (as analysed by Fourier shape analysis), indicating that specimens
infected with trematodes tended to grow more elongated shells than uninfected ones (Fig.
4.6). Additionally, inclusion of factor trematodes in the GLM on shell width revealed that
relative shell inflation at population Q was not only significantly influenced by trematode
infection but also by sexual dimorphism (Table 4.4). Infected specimens tended to be
relatively wider than uninfected ones (Fig. 4.4A). Thus, accounting for alteration of shell
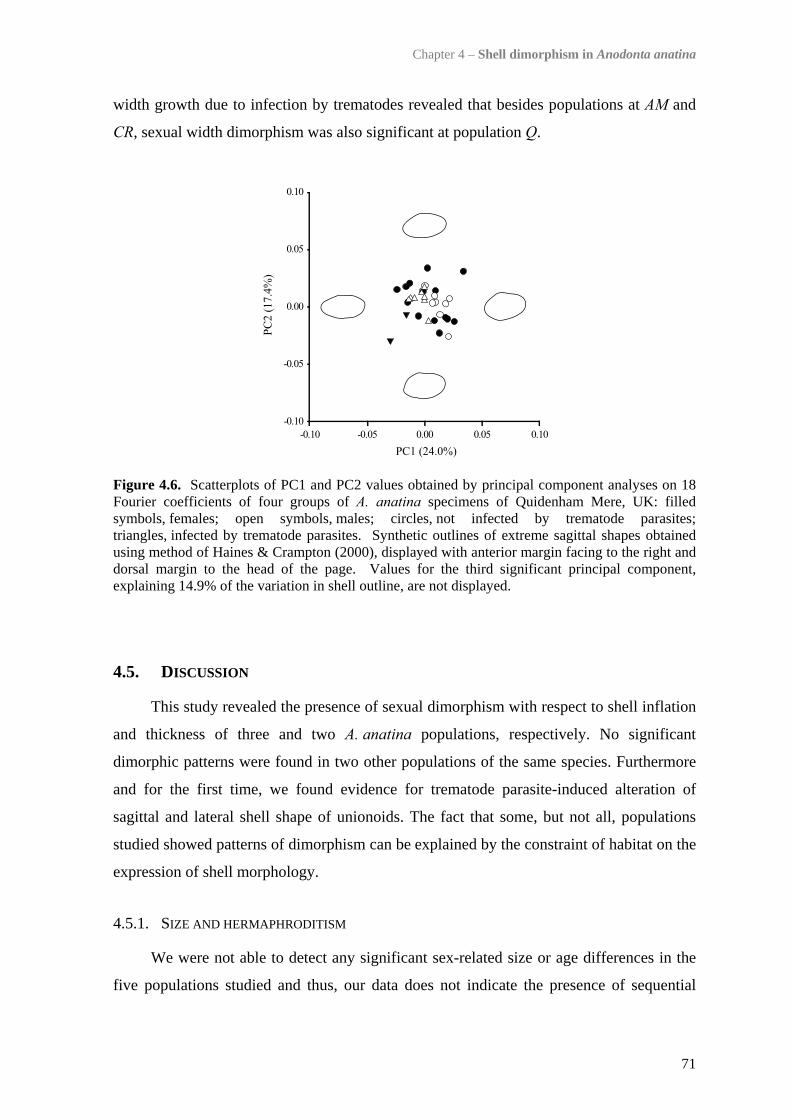
Chapter 4 – Shell dimorphism in Anodonta anatina
71
width growth due to infection by trematodes revealed that besides populations at AM and
CR, sexual width dimorphism was also significant at population Q.
Figure 4.6. Scatterplots of PC1 and PC2 values obtained by principal component analyses on 18 Fourier coefficients of four groups of A. anatina specimens of Quidenham Mere, UK: filled symbols, females; open symbols, males; circles, not infected by trematode parasites; triangles, infected by trematode parasites. Synthetic outlines of extreme sagittal shapes obtained using method of Haines & Crampton (2000), displayed with anterior margin facing to the right and dorsal margin to the head of the page. Values for the third significant principal component, explaining 14.9% of the variation in shell outline, are not displayed.
4.5. DISCUSSION
This study revealed the presence of sexual dimorphism with respect to shell inflation
and thickness of three and two A. anatina populations, respectively. No significant
dimorphic patterns were found in two other populations of the same species. Furthermore
and for the first time, we found evidence for trematode parasite-induced alteration of
sagittal and lateral shell shape of unionoids. The fact that some, but not all, populations
studied showed patterns of dimorphism can be explained by the constraint of habitat on the
expression of shell morphology.
4.5.1. SIZE AND HERMAPHRODITISM
We were not able to detect any significant sex-related size or age differences in the
five populations studied and thus, our data does not indicate the presence of sequential

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
72
hermaphroditism in these populations of A. anatina. This is further supported by the fact
that only two out of 138 sexed specimens were identified as hermaphroditic.
4.5.2. SEXUAL DIMORPHISM IN SAGITTAL SHELL SHAPE
None of the A. anatina populations studied showed any sex-related difference with
regard to sagittal shape of shells. This is not surprising given that when gravid, marsupia in
this and other Unioninae primarily expand laterally (i.e. along the shell width axis) and not
ventrally as in lampsilines, known for their well developed sexual dimorphism in sagittal
shell shape (e.g. Jass & Glenn, 2004). Although some authors of the 19th century claimed
the presence of sex-related differences regarding shell elongation of European unionids
(e.g. Kirtland, 1834; Siebold, 1837; Buchner, 1900; Brander, 1954), most of these
observations are merely anecdotal. Fallacious unionid taxonomy and doubtful sex
identification at that time further add to the fact that such documentations should be treated
with caution. More recently, however, more subtle differences in sagittal shell shape were
observed in several Indotropical unionids. Agrawal (1974) and Hamai (2004) found the
anterior part of female Parreysia wynegungaensis (Lea, 1859) and Inversidens japonensis
(Lea, 1859) respectively, to be more developed than in males. Unfortunately, these authors
did not discuss possible explanations for the patterns they observed.
4.5.3. SEXUAL DIMORPHISM IN RELATIVE SHELL WIDTH
There has been considerable anecdotal documentation of differences in shell width
of male and females in several unionoid species (e.g. Kirtland, 1834; Hazay, 1881; Sterki,
1903; Hayashi, 1935) but this study provides the first statistically significant evidence for
shell width dimorphism in a European unionoid. As expected, in all study populations,
females were more inflated than males. It is likely that this is a result of altered width
growth of females during the period of gravidity. For example, gravid marsupial gills in
the long-term brooding Pyganodon cataracta (Say, 1817) swell to nearly thirty times their
non-brooding width (Tankersley & Dimock, 1992). However, sexual width dimorphism
was significant in only three of the five populations studied and similar inconsistencies in
dimorphic patterns across populations of the same species have been reported in the past
(Weisensee, 1916; Brander, 1954). Possible reasons for such interpopulational
discrepancies have, however, not been discussed before, leaving one to question why
dimorphic patterns occur in some populations but not in others.

Chapter 4 – Shell dimorphism in Anodonta anatina
73
Our study provides some evidence for an indirect effect of environmental condition
on sexual shell width dimorphism in a unionoid. Thus, two of the three populations
exhibiting significant dimorphic patterns in this character (i.e. Q and AM) were those with
the highest population abundances, fastest growth rates and largest maximum sizes. All
these parameters indicate that habitat conditions at these two sites were more favourable
for A. anatina than at the remaining sites. Under such “good” habitat conditions, a
female/hermaphroditic mussel could be expected to be able to be more fecund, resulting in
more swollen gills and consequently more inflated shells.
Favourable conditions for growth are also indicated by the large shell length at the
third annulus in the third population displaying significant shell width dimorphism (CR).
The low density of mussels at this site, coupled with unusually young maximum age in this
population (< 4 years) could be the result of a catastrophic event rather than reflecting
unfavourable conditions. In this population, some of the gravid specimens were only two
years old, which is especially young for maturity in anodontines (Heard, 1975) but may
help explain the sexual dimorphism recorded. Thus, interpopulational variation of life
history traits such as size/age of maturity, incubation length and/or timing of glochidial
release (e.g. Bauer, 2001; Hochwald, 2001; Haag & Stanton, 2003) might also influence
female shell bulging and should be considered when depicting sexual dimorphic patterns
in unionoid populations.
4.5.4. SEXUAL DIMORPHISM IN SHELL THICKNESS AND DENSITY
Shell thickness and density have rarely been considered as potentially sex-
influenced. Nevertheless, in two of the five A. anatina populations a significant sex-related
difference in shell thickness was found, with female shells tending towards thinner shells
than males.
It seems likely that this type of sexual dimorphism could be triggered by costs of
producing and nourishing offspring, depleting resources that otherwise would be used for
shell production. Brooding female unionoids are known to contribute both nutrients
(Wood, 1974) and calcium (Silverman, Steffens & Dietz, 1985; Silverman, Kays & Dietz,
1987) to their eggs and glochidia, formed in water channels of their marsupia. Reduced
nutrient and/or calcium availability during the gravid period of females have been argued
to slow down or even cause negative growth (Downing & Downing, 1993; McIvor &
Aldridge, 2007). Though we did not detect any differences in perimeter growth rates of

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
74
male and female mussels, thickness growth could be affected in a similar way. What is
more, investing energy in growing larger on the expense of growing thick could be
advantageous given that fecundity has repeatedly been shown to increase with size in
unionoids (e.g. Bauer, 1998; Hochwald, 2001; Haag & Stanton, 2003).
4.5.6. TREMATODE PARASITE-INDUCED DIMORPHISM
Based on one A. anatina population of Quidenham Mere, our study suggests that
unionoid shell shape can be significantly influenced by trematode parasitism. This
represents the first evidence of parasite-induced shape alteration in a European unionoid
though previous authors have shown that overall growth rates can be slowed down in
mussels due to parasitism by trematodes (Taskinen & Valtonen, 1995; Taskinen, 1998).
Other studies have found that trematode parasitism can additionally reduce reproductive
output and physiological condition of unionoids (Jokela, Uotila & Taskinen, 1993;
Gangloff, Lenertz & Feminella, 2008). While reduced fecundity might be expected to
result in narrower shells, our data suggest that infected mussels of both sexes tended to be
relatively more inflated than uninfected ones. One possible explanation for this pattern
could be that reproduction of trematodes within the mussel’s gonad might cause bulging of
the mussel’s soft tissue, ultimately resulting in more inflated shell growth. Further work is
needed to understand the effects of trematodes on shell swelling and indeed on our
observation that infected A. anatina were especially elongated.
4.5.6. HABITAT-CONSTRAINED DIMORPHISM
While this study has found pronounced patterns in shell dimorphism in A. anatina,
which can be attributed to sex and parasite infection, no pattern was universal across all
five populations in terms of its presence and/or degree of expression. This variation in
expression of morphology can be explained by habitat constraints (e.g. Chapter 2; Zieritz
& Aldridge, 2009) which may in some circumstances have a dominating effect on
morphology. Similar patterns of environmentally-constrained differences between
populations in the degree of, for example, sexual dimorphisms have been observed in other
animal taxa such as gastropods (Goodwin & Fish, 1977) and beetles (Stillwell & Fox,
2007).

Chapter 4 – Shell dimorphism in Anodonta anatina
75
4.5.7. APPLICATION OF THE PATTERNS OBSERVED
Freshwater mussels are some of the most imperilled fauna in the world’s freshwaters
(Bogan, 1993; Lydeard et al., 2004). Understanding the trade-offs between reproductive
potential and habitat-imposed constraints may assist in identifying optimal habitats for
particular species and selecting appropriate management activities.
In some populations, shell shape may provide a tool for the rapid identification of
sex in the field, which may be especially useful in propagation programmes, where only
gravid females are typically required. Interspecific differences in male and female shell
morphologies may reveal species-specific patterns in brooding, fecundity and size at
maturity. An understanding of the biotic and abiotic factors that influence shell shape may
be particular valuable in inferring habitat requirements for very rare, extinct or fossil
species, where little or no data can be collected from living populations. Our data also
suggest that a change in morphology of a population over time could reflect a change in
parasite load, which could prove a useful tool for managers of closely-monitored
populations, such as those used in pearl cultivation or those of high conservation value.


CHAPTER 5
VARIABILITY AND A NEW MODEL FOR CHARACTER EVOLUTION OF UMBONAL
SCULPTURES IN THE UNIONOIDA

“The fossils are giving me indigestion over where the generic lines
are drawn and how long the genera persist in the fossil record.”
Arthur Bogan (pers. comm.)

79
CHAPTER 5
VARIABILITY AND A NEW MODEL FOR CHARACTER EVOLUTION OF UMBONAL SCULPTURES IN THE UNIONOIDA
5.1. ABSTRACT
‘Umbonal sculptures’ (also known as ‘beak sculptures’ or ‘rugae’) of freshwater
mussels (Unionoida) are restricted to the early shell region and have long been used for
species identification. Specificity of beak sculptures to higher taxonomic levels, and their
value for phylogenetic reconstructions and classification of fossil taxa is still under
considerable scientific debate. This is due to our lack of understanding of character
evolution and consequently, knowledge of synapomorphies, plesiomorphies and
convergences in beak sculpture character states. Based on examination of over 150
species, covering five of the six unionoid families, this study presents a new model of
character evolution of umbonal sculptures in this bivalve group. Presence of ‘V-shaped’
sculpture in Triassic unionoids and fossil members of their closest extant relatives, the
Trigonioida, indicates that this type represents the original character state of unionoid beak
sculpture. From this plesiomorphic ‘V-shaped’ type, ‘pseudo-radial’ sculpture probably
arose in one line of development, whereas ‘W-shaped’ and ultimately ‘double-looped’
sculpture was developed in other unionoid groups. All remaining beak sculpture types
observed in the study material (i.e. ‘pseudo-concentric’, ‘single-looped’, ‘nodulous’,
‘wrinkled’ and ‘smooth’) represent derivates of the ‘V-shaped’, ‘W-shaped’ and/or
‘double-looped’ type. Implications of this new model of character evolution for unionoid
phylogeny and functional morphology of beak sculpture are discussed.
5.2. INTRODUCTION
Most juvenile freshwater mussels (Unionoida) exhibit surface protrusions, which at a
certain life stage (usually within the first years of age) either completely cease to be
produced or change into a different type of sculpture. Morphological characteristics of
these so called ‘umbonal sculptures’ (also ‘beak sculptures’ or ‘rugae’) have long been
recognised to be species-specific and are thus widely used as an identification tool (e.g.
Marshall, 1890; Simpson, 1900; Kennard, Salisbury & Woodward, 1925; Bloomer, 1937;

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
80
Dolmen & von Proschwitz, 1999; Killeen, Aldridge & Oliver, 2004; Watters, Hoggarth &
Stansbery, 2009). While some authors discussed unifying characteristics of these structures
at the genus and occasionally higher taxonomic levels (Frierson, 1909b, a; Ortmann,
1912), distribution of beak sculpture morphotypes across the unionoid phylogeny and
consequently, character evolution of beak sculpture remains poorly understood.
Knowing if a given character state of unionoid beak sculpture is plesiomorphic
(basal), apomorphic (derived) or homoplasic (convergent) may help determine
phylogenetic positions of taxa and resolve open questions on these bivalves’ systematics
and phylogeny. This is particularly important with regard to the fossil record, for which
taxonomic classification is based largely on beak sculpture and a few other shell characters
(e.g. Haas, 1969b; Sha, 1993; Guo, 1998a).
By recognising ecological similarities of convergent but unrelated taxa, a good
understanding of present homologies and homoplasies of beak sculptures can additionally
help identify probable functional morphologies of this shell character. Several possible
functions have been attributed to umbonal sculptures, including predator deterrence
(Clarke, 1986), anchoring (Watters, 1994) and shell reinforcement (Checa & Jiménez-
Jiménez, 2003a), but function remains poorly resolved. Being most pronounced during the
early postlarval period of unionoid life, understanding the functional morphology of
umbonal sculptures could give vital insights on ecological aspects of this crucial but poorly
understood life stage.
Attempts have been made to reconstruct the appearance of different types of rugae in
the course of unionoid evolution, but proposed models of character evolution are largely
contradictory (Fig. 5.1). Simpson (1900) believed the original character state of unionoid
beak sculpture to be ‘radial’ (Fig. 5.1A). This author argued that curving and coalescation
of one or more of the middle pairs of radial bars would result in a ‘divaricate’ sculpturing,
which would ultimately have developed in ‘concentric’ and ‘double-looped’ types,
exhibited by what he regarded as the most derived unionoid species. Largely agreeing with
Simpson’s model of character evolution, Modell (1942) (Fig. 5.1C) considered the
phylogenetic signal of umbonal sculptures of utmost importance and in fact based a whole
phylogenetic tree predominantly on this shell character. Ortmann (1912) (Fig. 5.1B), on
the other hand, not only disagreed with the phylogenetic importance of rugae above the
genus level but also believed that all types of umbonal sculpturing originated from a
‘concentric’ type. He considered the ‘divaricate’ (zig-zag) and ‘radial’ sculpture as the

Chapter 5 – Umbonal sculptures of Unionoida
81
most extreme conditions of two lines of development via intermediate ‘double-looped’ and
‘V-shaped’ stages, respectively.
Figure 5.1. A-C. Previously suggested models for character evolution of unionoid umbonal sculptures, simplified after descriptions by (A) Simpson (1900), (B) Ortmann (1912) and (C) Modell (1942).
Over the past 30 years or so, modern techniques including the use of molecular
markers, have drastically altered our view on unionoid phylogeny and evolution, which,
nevertheless remains a matter of intense scientific debate (see 1.1.1; Graf & Cummings,
2006b; Hoeh et al., 2009; Graf & Cummings, 2010a). Character evolution of umbonal
sculpture has not been revisited in this new context but this shell trait has repeatedly been
included in modern phylogenetic analyses (e.g. Hoeh, Bogan & Heard, 2001; Graf &
Cummings, 2006b). Based largely on one of the three aforementioned models of character
evolution (i.e. Simpson, 1900; Ortmann, 1912; Modell, 1942), these attempts have,
however, been rather unsuccessful. Following Ortmann’s (1912) model of character
evolution, Graf & Cummings (2006b), for example, classified beak shell sculpture into
three character states (1) ‘simple concentric’ or ‘absent’, (2) ‘radial’ and (3) ‘double-
looped’ or ‘zigzag’. Using this system of character state classification, Graf & Cummings’
(2006b) phylogenetic analysis resulted in a relatively low consistency index of only 0.222
for beak sculpture, indicating an unexpectedly high rate of change in this shell feature’s
morphology during unionoid evolution. Alternatively, the three character states used by

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
82
these authors might be inadequate in describing the morphological appearances of
unionoid umbonal sculptures.
Another caveat of almost all previous studies dealing with umbonal rugae
morphology is their general disregard of the morphogenetic aspect of sculpture formation.
According to Checa & Jiménez-Jiménez (2003b), bivalve ribs can be classified into three
categories, which differ in their fabricational morphology and consequently, are likely to
be of considerable phylogenetic value. (1) ‘Radial’ ribs, characteristic for e.g. cardiids, are
produced by particular portions of the mantle specialised for rib secretion and
consequently, are represented as helicospirals that converge towards the umbo (Checa,
2002; Checa & Jiménez-Jiménez, 2003b). (2) ‘Comarginal’ (‘concentric’) ribs are secreted
by periodic extension of the mantle all along its length and thus, always appear completely
parallel to the margin (Checa, 2002; Checa & Jiménez-Jiménez, 2003b). This ribbing
pattern is displayed for example in tellinids. (3) The ‘oblique’ (‘divaricate’) pattern is less
commonly found in bivalves. Ribs of this type have directions that are intermediate
between radial and comarginal, and are produced by an elaborate contact-guidance
behaviour of the mantle (Checa, 2002). Watters (1994) argued that all types of unionoid
beak sculpture are divaricate, with all other forms such as ‘double-looped’, ‘single-looped’
and ‘barred’ representing derivations of the same.
This study uses the beak sculpture from over 150 unionoid species, covering all
families but the Etheriidae, to (1) develop an up-to-date model of character evolution using
recent advances in understanding of bivalve phylogeny and rib formation, (2) identify
convergence of ecologically similar species, and on that basis, (3) discuss phylogenetic
significance and probable functional morphologies of these structures.
5.3. MATERIALS AND METHODS
5.3.1. CLASSIFICATION OF BEAK SCULPTURE TYPES
Where possible, beak sculpture morphotype was determined directly on shell
material of the University Museum of Zoology Cambridge (UMZC) and private collections
(p.c.). To maximise number of taxa included in the analysis, beak sculpture morphology
was additionally examined on the basis of photographs and/or drawings from following
publications and websites: Unionoida from the Nearctica (Marshall, 1890; Coker et al.,
1921; Clarke, 1973, 1981, 1985; Frank & Lee, 1998; Grabarkiewicz & Todd, 2010;
Klocek, Bland & Barghusen, 2010), Neotropica (Mansur & Pereira, 2006; Mansur &

Chapter 5 – Umbonal sculptures of Unionoida
83
Pimpão, 2008; Pimpão, Rocha & de Castro Fettuccia, 2008; Cummings & Mayer, 2009;
Kohl, 2010), Afrotropica (von Martens, 1897; Pilsbry & Bequaert, 1927; Haas, 1929;
Scholz & Glaubrecht, 2004; Graf & Cummings, 2006a, 2007a), Palearctica (Germain,
1922; Kennard, Salisbury & Woodward, 1925; Killeen, Aldridge & Oliver, 2004; Araujo
et al., 2009), Indotropica (Savazzi & Peiyi, 1992; Fischer, 2007), Australasia (McMichael
& Hiscock, 1958; Playford & Walker, 2008) and worldwide (Modell, 1942; Haas, 1969b;
Graf & Cummings, 2010b). Further photographs of Indotropical unionids and Nearctical
margaritiferids were kindly provided by Ms Du Lina (Kunming Institute of Zoology,
China) and Ms Mary Sollows (University of New Brunswick, Canada), respectively.
Overall, beak sculpture of 156 modern unionoid species (about 300 specimens), covering
80 of the 161 currently described unionoid genera (according to Graf & Cummings,
2007b), was examined on the basis of shell and/or picture material (Table 5.1; Appendix
Table A.1).
For some taxa, only a small number of shells/pictures of well preserved umbonal
surface areas could be obtained. In these cases, additional information on beak sculpture
morphology was gathered from descriptions in the aforementioned and additional
publications (Simpson, 1900, 1914; Connolly, 1939; Modell, 1949; Dell, 1953; Parodiz &
Bonetto, 1963; Modell, 1964; Bonetto, 1966; Haas, 1969a; Ponder & Bayer, 2004). This
was the case particularly for taxa with apparently smooth umbos, i.e. Velesunioninae,
Mycetopodidae and Iridininae.
5.3.2. DEVELOPMENT OF MODEL OF CHARACTER EVOLUTION, AND IDENTIFICATION OF HOMOLOGIES AND HOMOPLASIES
Umbonal sculpture of each individual analysed was classified into at least one of
nine easily distinguishable morphological types. Single specimens that exhibited
intermediate forms and/or more than one form simultaneously were assigned to several
beak sculpture types. Based on this data on main and intermediate types of rugae observed
in the unionoid taxa examined, a model of beak sculpture evolution within the Unionoida
was subsequently developed. This model was further used as basis to identify homologies
and homoplasies (convergences) of sculpture morphotypes across the unionoid phylogeny.
This was done on the two currently best accepted cladograms on interfamiliar relationships
of the Unionoida, i.e. (1) Graf & Cummings (2006b) and (2) Bogan & Hoeh (2000) +
Hoeh, Bogan & Heard (2001). Although beak sculpture morphology had been included as
a character in both Graf & Cummings’ (2006b) and Hoeh, Bogan & Heard’s (2001)

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
84
phylogenetic analyses, non-independence of our analysis from these two studies can be
considered negligible for two reasons: (1) both Graf & Cummings’ (2006b) and Hoeh,
Bogan & Heard’s (2001) phylogenies are based on a large number of DNA sequences and
morphological data, and (2) in both cases, classification of beak sculpture character states
differs from the one applied in the present study.
5.4. RESULTS
5.4.1. MORPHOLOGICAL TYPES OF UNIONOID BEAK SCULPTURE
Beak sculptures of the more than 150 unionoid species analysed could be classified
into nine well distinguishable types (Fig. 5.2; Table 5.1):
(1) ‘V-shaped’ sculpture, characterised by one central “V” and at least one pair of
oblique lateral ribs, was found in some Afrotropical and Indotropical incertae sedis
unionids (e.g. Parreysia triembola (Benson, 1855); Fig. 5.2A) and members of the
Hyriinae (e.g. Prisodon corrugatus (Lamarck, 1819); Fig. 5.2B). Although no V-sculpture
could be found in any specimens of the family Iridinidae examined, Haas (1969a: p.583)
described this beak sculpture type from the iridinid species Mutela hargeri (Smith, 1908).
(2) In two of the four hyriid tribes and a few Indotropical unionids, predominantly of
the genus Parreysia, an almost radial beak sculpture is exhibited. However, in all of these
cases, ribs either (a) originate from several points near the umbo (e.g. Diplodon chilensis
(Gray, 1828); Fig. 5.2C) and/or (b) show at least one V- or Λ-shaped
convergence/divergence (e.g. Castalia ambigua Lamarck, 1819; Fig. 5.3A). Since such
irregularities would not occur in truly radial ribs sensu Checa (2002) and Checa &
Jiménez-Jiménez (2003b), we propose the term ‘pseudo-radial’ rather than ‘radial’ for this
kind of unionoid beak sculpture.
(3) ‘W-shaped’ (’zigzag’) sculpture, representing two or more series of connected
“V”s, was identified in several unionine, quadruline (e.g. Quadrula quadrula (Rafinesque,
1820); Fig. 5.2D) and incertae sedis unionids (e.g. Scabies crispata (Gould, 1843); Fig.
5.2E).
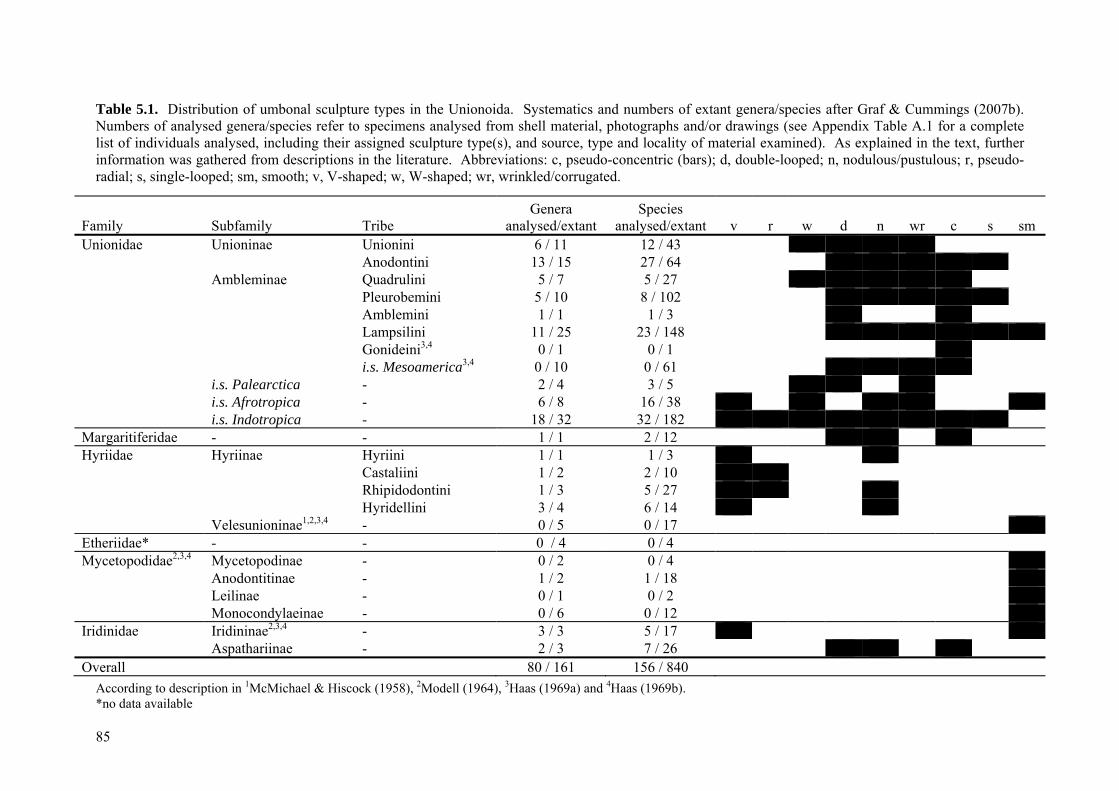
85
Table 5.1. Distribution of umbonal sculpture types in the Unionoida. Systematics and numbers of extant genera/species after Graf & Cummings (2007b). Numbers of analysed genera/species refer to specimens analysed from shell material, photographs and/or drawings (see Appendix Table A.1 for a complete list of individuals analysed, including their assigned sculpture type(s), and source, type and locality of material examined). As explained in the text, further information was gathered from descriptions in the literature. Abbreviations: c, pseudo-concentric (bars); d, double-looped; n, nodulous/pustulous; r, pseudo-radial; s, single-looped; sm, smooth; v, V-shaped; w, W-shaped; wr, wrinkled/corrugated.
According to description in 1McMichael & Hiscock (1958), 2Modell (1964), 3Haas (1969a) and 4Haas (1969b). *no data available
Family Subfamily Tribe Genera
analysed/extant Species
analysed/extant v r w d n wr c s sm Unionidae Unioninae Unionini 6 / 11 12 / 43
Anodontini 13 / 15 27 / 64 Ambleminae Quadrulini 5 / 7 5 / 27
Pleurobemini 5 / 10 8 / 102 Amblemini 1 / 1 1 / 3 Lampsilini 11 / 25 23 / 148 Gonideini3,4 0 / 1 0 / 1 i.s. Mesoamerica3,4 0 / 10 0 / 61
i.s. Palearctica - 2 / 4 3 / 5 i.s. Afrotropica - 6 / 8 16 / 38 i.s. Indotropica - 18 / 32 32 / 182
Margaritiferidae - - 1 / 1 2 / 12 Hyriidae
Hyriinae
Hyriini 1 / 1 1 / 3 Castaliini 1 / 2 2 / 10 Rhipidodontini 1 / 3 5 / 27 Hyridellini 3 / 4 6 / 14
Velesunioninae1,2,3,4 - 0 / 5 0 / 17 Etheriidae* - - 0 / 4 0 / 4 Mycetopodidae2,3,4
Mycetopodinae - 0 / 2 0 / 4 Anodontitinae - 1 / 2 1 / 18 Leilinae - 0 / 1 0 / 2 Monocondylaeinae - 0 / 6 0 / 12
Iridinidae Iridininae2,3,4 - 3 / 3 5 / 17 Aspathariinae - 2 / 3 7 / 26
Overall 80 / 161 156 / 840
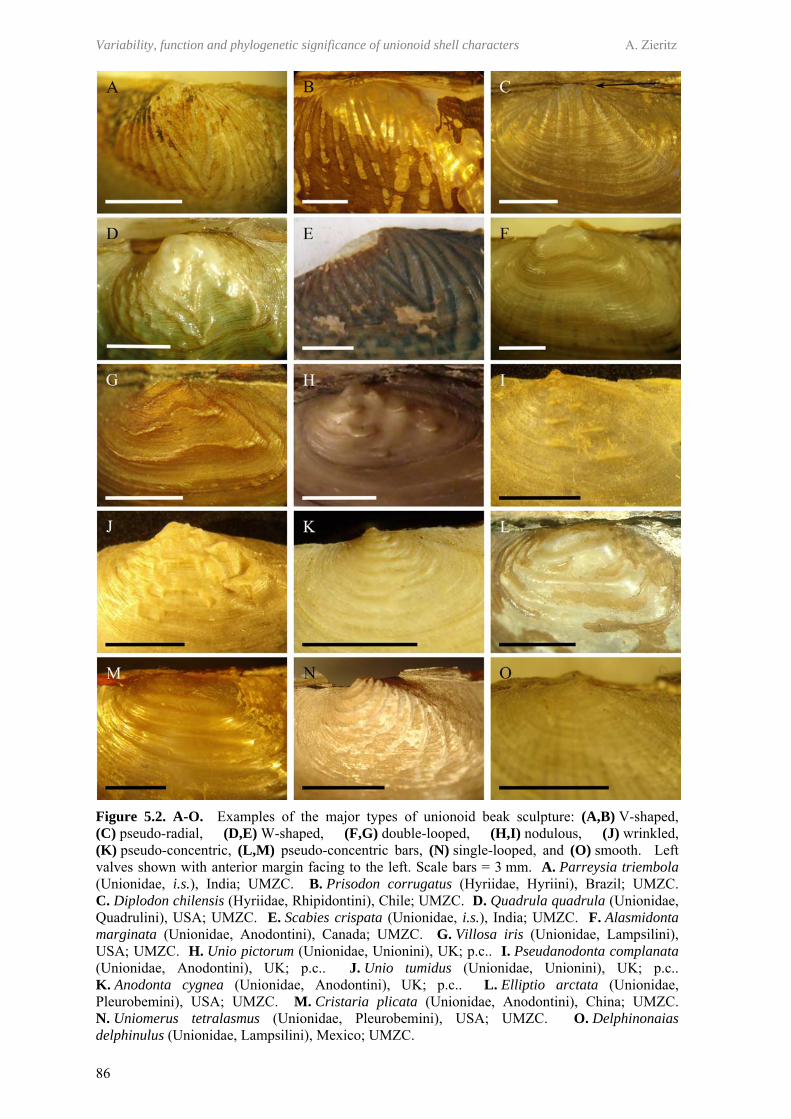
Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
86
Figure 5.2. A-O. Examples of the major types of unionoid beak sculpture: (A,B) V-shaped, (C) pseudo-radial, (D,E) W-shaped, (F,G) double-looped, (H,I) nodulous, (J) wrinkled, (K) pseudo-concentric, (L,M) pseudo-concentric bars, (N) single-looped, and (O) smooth. Left valves shown with anterior margin facing to the left. Scale bars = 3 mm. A. Parreysia triembola (Unionidae, i.s.), India; UMZC. B. Prisodon corrugatus (Hyriidae, Hyriini), Brazil; UMZC. C. Diplodon chilensis (Hyriidae, Rhipidontini), Chile; UMZC. D. Quadrula quadrula (Unionidae, Quadrulini), USA; UMZC. E. Scabies crispata (Unionidae, i.s.), India; UMZC. F. Alasmidonta marginata (Unionidae, Anodontini), Canada; UMZC. G. Villosa iris (Unionidae, Lampsilini), USA; UMZC. H. Unio pictorum (Unionidae, Unionini), UK; p.c.. I. Pseudanodonta complanata (Unionidae, Anodontini), UK; p.c.. J. Unio tumidus (Unionidae, Unionini), UK; p.c.. K. Anodonta cygnea (Unionidae, Anodontini), UK; p.c.. L. Elliptio arctata (Unionidae, Pleurobemini), USA; UMZC. M. Cristaria plicata (Unionidae, Anodontini), China; UMZC. N. Uniomerus tetralasmus (Unionidae, Pleurobemini), USA; UMZC. O. Delphinonaias delphinulus (Unionidae, Lampsilini), Mexico; UMZC.

Chapter 5 – Umbonal sculptures of Unionoida
87
(4) Almost all tribes of the Unionidae, the Margaritiferidae and one genus
(Aspatharia) of the iridinid subfamily Aspathariinae show a ‘double-looped’ sculpture
(e.g. Alasmidonta marginata Say, 1818; Fig. 5.2F; and Villosa iris (Lea, 1829); Fig. 5.2G).
A considerable degree of variation was observed in this beak sculpture type, which can
predominantly be attributed to differences in thickness of the ribs, in the angle of the two
loops and in the angle at the point of convergence of the two loops.
(5) Partially or completely ‘nodulous’ protrusions were found in members of almost
all families and subfamilies. These protuberances are often arranged in two or more
radiating rows (e.g. Unio pictorum (Linnaeus, 1758); Fig. 5.2H; Pseudanodonta
complanata (Rossmässler, 1835); Fig. 5.2I; Parreysia corrugata (Müller, 1774); Fig. 5.3B;
Coelatura aegyptiaca (Cailliaud, 1827); Fig. 5.3D; Unio tumidus Philipsson, 1788; 5.3F)
but can also appear irregularly distributed (e.g. Diplodon multistriatus (Lea, 1831); Fig.
5.3C; Tritogonia verrucosa (Rafinesque, 1820); Fig. 5.3E).
(6) Ribs in the shape of irregular zigzags, loops or corrugations were classified as
‘wrinkled’ (‘corrugated’) and were exhibited by almost all unionid subfamilies and tribes
(e.g. U. tumidus; Fig. 5.2J).
(7) In several unionid, margaritiferid and aspathariine specimens, ribs of nearly
concentric outline but never strictly following growth rings were found and classified as
‘pseudo-concentric’ (e.g. Anodonta cygnea (Linnaeus, 1758); Fig. 5.2K). The ventral part
of such ribs sometimes tends away from this curved structure towards a more linear “bar”
shape (e.g. Elliptio arctata (Conrad, 1834); Fig. 5.2L). In some species, these ‘pseudo-
concentric bars’ are completely free standing (e.g. Cristaria plicata (Leach, 1815); Fig.
5.2M).
(8) Another type of sculpture that has been misleadingly described as “concentric”
was exhibited by only a few genera of unionids (e.g. Toxolasma, Uniomerus). Loops of
such ‘single-looped’ rugae are sometimes orientated somewhat oblique to the growth
direction and usually originate from one point at the umbo (e.g. Uniomerus tetralasmus
(Say, 1831); Fig. 5.2N).
(9) The Mycetopodidae, the hyriid subfamily Velesunioninae and most members of
the iridinid subfamily Iridininae are all characterised by completely ‘smooth’ beaks. Apart
from these three taxa, specimens examined belonging to three species of the family
Unionidae (i.e. Pseudospatha tanganyicensis (Smith, 1880), Actinonaias pectorosa
(Conrad, 1834) and Delphinonaias delphinulus (Morelet, 1849); Fig. 5.2O) did not exhibit
any apparent beak sculpture.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
88
5.4.2. INTERMEDIATE FORMS AND IMPLICATIONS FOR CHARACTER EVOLUTION OF UNIONOID BEAK SCULPTURE
(1) At least at one point of the shell of most ‘pseudo-radial’ forms, radiating bars
either converge or diverge to/from each other, forming a V- or Λ-shaped pattern (e.g.
C. ambigua; Fig. 5.3A). ‘Pseudo-radial’ sculpture may therefore be regarded as a derivate
of the ‘V-shaped’ and possibly also the ‘W-shaped’ type, potentially arising from a change
in angle at the points of convergence/divergence of the V-bars.
(2) In a few cases (e.g. U. tumidus; Fig. 5.3F), beak sculpture apparently represents
an intermediate form between ‘W-shaped’ and ‘double-looped’.
(3) ‘Nodulous’ protrusions were found to be associated with ‘V-shaped’ (e.g.
P. corrugata; Fig. 5.3B; D. multistriatus; Fig. 5.3C), ‘W-shaped’ (C. aegyptiaca;
Fig. 5.3D; T. verrucosa; Fig. 5.3E), ‘W-shaped/double-looped’ (e.g. U. tumidus; Fig. 5.3F)
and ‘double-looped’ sculpture.
(4) Several species presented irregular ‘wrinkles’ or corrugations associated with
either ‘V-shaped’, ‘W-shaped’ or ‘double-looped’ sculpture (e.g. Lasmigona compressa
(von Martens, 1860); Fig. 5.3G).
(5) Gradual alteration from ‘double-looped’ to ‘pseudo-concentric (barred)’ rugae
from the umbonal to the distal shell part within a specimen was commonly observed (e.g.
Elliptio dilatata (Rafinesque, 1820); Fig. 5.3H).
(6) ‘Single-looped’ rugae were present only in those higher taxa that also exhibited
‘double-looped’ sculpture, suggesting that this sculpture developed from the ‘double-
looped’ type (Table 5.1). While a gradual reduction, and ultimately omission of one of the
two loops from umbonal to distal shell parts was observed in several unionid species (e.g.
Pegias fabula (Lea, 1838); Fig. 5.3I), none of the shells examined displayed an umbonal
sculpture that represented an unambiguously intermediate form between the ‘double-
looped’ and the ‘oblique single-looped’ type of, for example, U. tetralasmus (Fig. 5.2N). It
is therefore unclear if the ‘regular single-looped’ type (of e.g. P. fabula; Fig. 5.3I) and the
‘oblique single-looped’ type (of e.g. U. tetralasmus (Fig. 5.2N)) represent
morphogenetically identical or different forms of beak sculpture.
(7) Although no specimens displayed clear intermediate stages between ‘V-shaped‘
and ‘W-shaped’ sculpture, both these forms were found within single genera (e.g.
Parreysia), indicating that these are very closely “related” forms.

Chapter 5 – Umbonal sculptures of Unionoida
89
Figure 5.3. A-I. Intermediate types of unionoid beak sculpture. Left valves shown with anterior margin facing to the left. Scale bars = 3 mm. A. Castalia ambigua (Hyriidae, Castaliini), Brazil; V-shaped – pseudo-radial. B. Parreysia corrugata (Unionidae, i.s.), India; V-shaped – pseudo-radial – nodulous. C. Diplodon multistriatus (Hyriidae, Rhipidontini), Brazil; V-shaped – pseudo-radial. D. Coelatura aegyptiaca (Unionidae, i.s.), Egypt; W-shaped – nodulous. E. Tritogonia verrucosa (Unionidae, Quadrulini), USA; W-shaped – nodulous. F. Unio tumidus (Unionidae, Unionini), UK; W-shaped – double-looped – nodulous. G. Lasmigona compressa (Unionidae, Anodontini), USA; double-looped – wrinkled. H. Elliptio dilatata (Unionidae, Pleurobemini), USA; double-looped – pseudo-concentric bars. I. Pegias fabula (Unionidae, Anodontini), USA; double-looped – single-looped. All specimens UMZC. Abbreviations: cb, concentric bars; d, double-looped; n, nodulous; r, pseudo-radial; s, single-looped; v, V-shaped; w, W-shaped; wr, wrinkled.
5.5. DISCUSSION
5.5.1. (IN)VALIDITY OF PREVIOUS MODELS OF CHARACTER EVOLUTION
Confirming observations by Watters (1994), beak sculptures of all shells examined
in this study can be assigned to the oblique (divaricate) type of sculpture formation.
Although some unionoids exhibit seemingly “radial” or “concentric (i.e. comarginal)” ribs,
and despite ubiquitous use of these terms by previous authors, none of the sculptures
examined in this study can be characterised as such in a morphogenetic sense. In fact, such

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
90
‘pseudo-radial’ and ‘pseudo-concentric’ sculptures should be regarded as derived forms of
oblique ribs. On this basis, previous models for character evolution of unionoid umbonal
sculptures proposed by Simpson (1900), Ortmann (1912) and Modell (1942), which
presume an original radial or concentric (comarginal) state, respectively, have to be
rejected. Occurrence of both ‘V-shaped’ and ‘W-shaped’ sculptures in very closely related
taxa, e.g. species within the genera Parreysia and Coelatura, provides a further argument
falsifying Ortmann (1912), who regarded these beak sculpture types as part of two separate
lineages of divergence (Fig. 5.1B).
5.5.2. PLESIOMORPHIC CHARACTER STATE AND THE FOSSIL RECORD
5.5.2.1. Smooth vs. sculptured plesiomorphic character state
All unionoid beak sculptures are of the divaricate type, which involves a
sophisticated contact-guidance mechanism of the mantle and has evolved only a few times
within the whole Bivalvia (Checa, 2002; Checa & Jiménez-Jiménez, 2003a). Against this
background, several facts indicate a common ancestor of the extant Unionoida exhibiting a
sculptured as opposed to a smooth beak:
(1) Oblique/divaricate sculpture is also present in many members of the closest
extant relatives of the Unionoida, the marine Trigonioida (e.g. Cox, 1969; Hoeh et al.,
1998). It is unlikely that oblique rib formation would have evolved independently in these
two very closely related groups. Other authors (e.g. Carter, Campbell & Campbell, 2006),
however, argued that Unionoida evolved from the Pachycardiidae, a Permian/Triassic
family with smooth beaks and currently unresolved systematic position (Haas, 1969b;
Newell & Boyd, 1975). This fact and the presence of apparently smooth beaks in some
Triassic unionoids, such as a “Mycetopoda-like” specimen described by Wanner (1921),
might indicate a smooth common ancestor of the Unionoida.
(2) If the plesiomorphic character state was a smooth umbo, beak sculpture would
have evolved several times within the Unionoida. Switching off or losing the ability for
sculpture production when “not needed” would involve fewer changes in genotype and
would thus be more likely to happen than repeated independent evolution of oblique
sculpture within a single bivalve order.
(3) Finally, morphologies of beak sculptures that would have evolved independently
from each other in case of a ‘smooth’ plesiomorphic scenario, are sometimes strikingly

Chapter 5 – Umbonal sculptures of Unionoida
91
similar (e.g. ‘V-shaped’ and/or ‘pseudo-radial’ sculpture of some Hyriinae and
Indotropical Unionidae).
5.5.2.2. Most likely plesiomorphic beak sculpture type
Presuming a sculptured plesiomorphic character state, the following facts indicate
that the basal condition was of the ‘V-shaped’ type:
(1) Of all unionoid beak sculptures observed in this study, the ‘V-shaped’ type is the
only one also exhibited by several ancient trigonioid genera, such as Asiatotrigonia,
Iotrigonia, Korobkovitrigonia, Orthotrigonia and Vaugonia (Cox, 1969; Cooper, 1991;
Checa & Jiménez-Jiménez, 2003a).
(2) Plesiomorphy of ‘V-shaped’ sculpture for the Unionoida is further supported by
similar sculpture patterns in early representatives of this group. For example, shells from
the North American Triassic, some of the oldest fossil records generally accepted to
belong to this group of bivalves, commonly exhibit ‘V-shaped’ and/or ‘pseudo-radial’
sculpture (Simpson, 1896; Wanner, 1921; Reeside, 1927; Modell, 1957; Good, 1989,
1998). Strikingly similar ‘V-shaped’, ‘W-shaped’ and ‘pseudo-radial’ sculptures can also
be found in the Cretaceous Trigonioidoidea of Eurasia (Gu, 1998; Guo, 1998b, a; Sha,
2007). Trigonioidoids are believed to be an extinct superfamily of the Unionoida (Sha,
2006) and may represent an independent parallel-evolution from a primitive ‘V-shaped’
type towards ‘W-shaped’ and ‘pseudo-radial’ sculpture (Guo, 1998a). It should be noted,
however, that rugae resembling the ‘double-looped’ type of modern unionoids are
exhibited by another relatively early (probably Jurassic) group of freshwater mussels, the
vetulonaians (Branson, 1935; Watters, 2001).
5.5.3. A NEW MODEL OF BEAK SCULPTURE CHARACTER EVOLUTION IN THE UNIONOIDA
Assuming a plesiomorphic ‘V-shaped’ umbonal sculpture and based on our
observations on intermediate forms of beak sculptures, we propose a new model of
character evolution of umbonal rugae in the Unionoida (Fig. 5.4), on which basis
homologies and homoplasies can be identified (Figs 5.5A,B). Two lines of development
originate from a primitive ‘V-shaped’ sculpture, which is retained in some Afrotropical
and Indotropical unionids, and members of the hyriid subfamily Hyriinae and iridinid
subfamily Iridininae: (1) ‘Pseudo-radially’ sculptured umbos developed at least twice, i.e.
in the Hyriinae and Indotropical Unionidae. (2) In the remaining Unionidae, but also the
Margaritiferidae and aspathariine Iridinidae, ‘W-shaped’ and ultimately ‘double-looped’

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
92
sculpture is exhibited. Assuming strictly unidirectional evolution from the ‘W-shaped’ to
the ‘double-looped’ character state, this change would have occurred at least five times
within the Unionoida (Fig. 5.5A,B). However, reciprocal evolution from ‘double-looped’
to ‘W-shaped’ beak sculpture cannot be excluded, particularly since these two sculpture
types differ only in the speed at which the double-looped waves travel in the longitudinal
direction (i.e. slower in the ‘double-looped’ than in the ‘W-shaped’ pattern). The
possibility of bi-directional evolution from ‘W-shaped’ to ‘double-looped’ beak sculpture
and vice versa is indicated by a double arrow in Figure 5.4 and merging of w- and d-
symbols in Figure 5.5.
‘Pseudo-concentric’ and possibly also ‘single-looped’ forms represent derivates of
the ‘double-looped’ type, and evolved several times within the Unionoida. Equally, the
‘nodulous’ and ‘wrinkled’ character states arose independently numerous times and may
be derived from ‘double-looped’, ‘W-shaped’ and ‘V-shaped’ sculpture. It is rarely
possible to determine which of these three types was the ancestral ribbing pattern to a
given ‘nodulous’ or ‘wrinkled’ form, as the underlying patterning is usually not retained in
specimens exhibiting these sculpture types. Lastly, ‘smooth’ forms can potentially develop
from any other character state.
Figure 5.4. A new model of character evolution of unionoid umbonal sculptures. In addition, completely smooth beaks may develop at any stage. Continuous arrows indicate evolutionary connection between two beak sculpture types evidenced by presence of *intermediate stages between two sculpture types, or #both sculpture types in species of the same genus. Dashed arrows indicate possible – but unproven – connection between two beak sculpture types. For discussion of directionality of character evolution see main text.
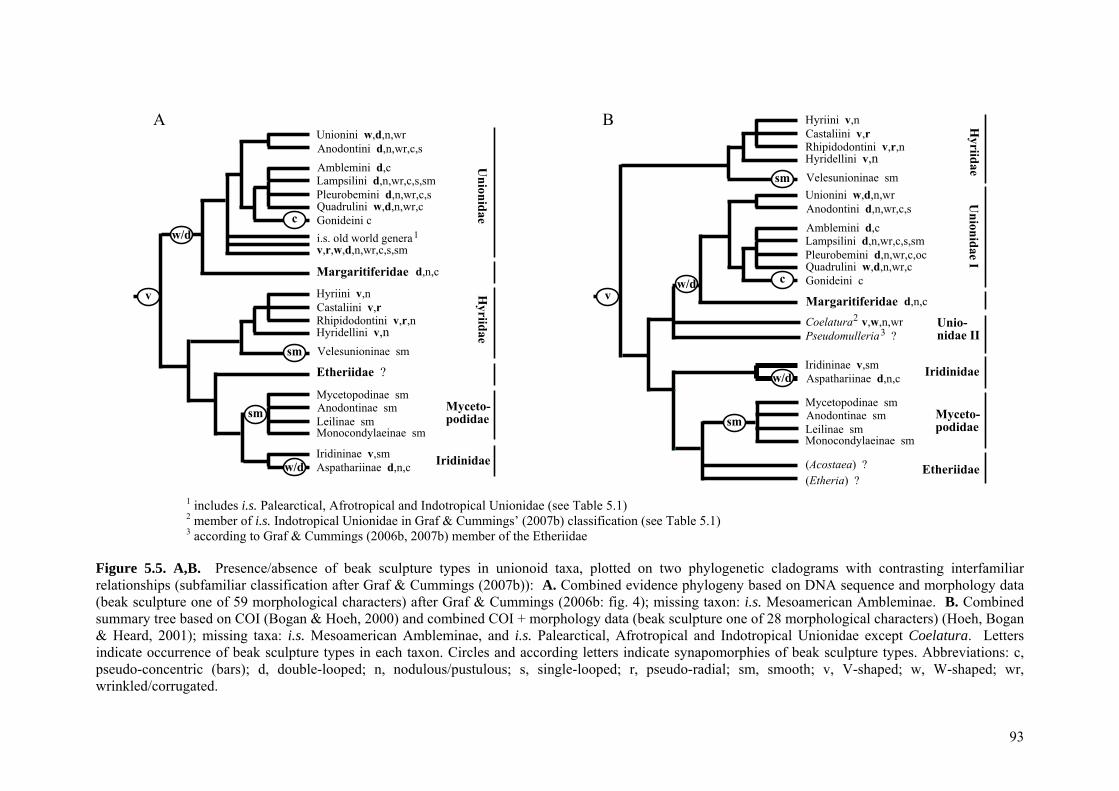
93
1 includes i.s. Palearctical, Afrotropical and Indotropical Unionidae (see Table 5.1) 2 member of i.s. Indotropical Unionidae in Graf & Cummings’ (2007b) classification (see Table 5.1) 3 according to Graf & Cummings (2006b, 2007b) member of the Etheriidae
Figure 5.5. A,B. Presence/absence of beak sculpture types in unionoid taxa, plotted on two phylogenetic cladograms with contrasting interfamiliar relationships (subfamiliar classification after Graf & Cummings (2007b)): A. Combined evidence phylogeny based on DNA sequence and morphology data (beak sculpture one of 59 morphological characters) after Graf & Cummings (2006b: fig. 4); missing taxon: i.s. Mesoamerican Ambleminae. B. Combined summary tree based on COI (Bogan & Hoeh, 2000) and combined COI + morphology data (beak sculpture one of 28 morphological characters) (Hoeh, Bogan & Heard, 2001); missing taxa: i.s. Mesoamerican Ambleminae, and i.s. Palearctical, Afrotropical and Indotropical Unionidae except Coelatura. Letters indicate occurrence of beak sculpture types in each taxon. Circles and according letters indicate synapomorphies of beak sculpture types. Abbreviations: c, pseudo-concentric (bars); d, double-looped; n, nodulous/pustulous; s, single-looped; r, pseudo-radial; sm, smooth; v, V-shaped; w, W-shaped; wr, wrinkled/corrugated.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
94
5.5.3.1. Implications for unionoid phylogeny and evolution
The new model of beak sculpture evolution developed in this study (Fig. 5.4) has
profound implications on our view on homologies vs. homoplasies of umbonal sculptures
within the unionoids. For example, while Graf & Cummings (2006b) considered the
smoothness of umbos of the Velesunioninae as the plesiomorphic state, results of the
present study indicate synapomorphy of this character state for this hyriid subfamily. On
the other hand, morphological similarity of ‘V-shaped’ and ‘pseudo-radial’ sculpture of
some Indotropical unionids (e.g. Harmandia) and Neotropical hyriids (e.g. Castalia) are
considered as superficial (i.e. homoplasic) by Graf & Cummings (2006b). Our
observations, however, indicate the ‘V-shaped’ character state to be the plesiomorphic one
Knowledge on morphological characteristics of primitive and derived beak
sculptures can further be used to hypothesise on the phylogenetic position of unresolved
taxa, such as Parreysia and Coelatura. Thus, the primitive ‘V-shaped’ or ‘W-shaped’
sculpture exhibited by these genera suggests a basal position within the Unionidae.
Another example is the poorly understood phylogeny within the Ambleminae, consisting
of the tribes Amblemini, Lampsilini, Pleurobemini, Quadrulini and Gonideini (Table 1.1).
Several phylogenetic analyses (Lydeard, Mulvey & Davis, 1996; Campbell et al., 2005;
Graf & Cummings, 2006b), found the Quadrulini to be the basal tribe to all other tribes of
this subfamily. Based on the primitive ‘W-shaped’ sculpture present in most quadruline
specimens analysed in the present study, a basal quadruline tribe would conform with the
present model of character evolution. However, other authors obtained cladograms that did
not place the Quadrulini as the sole basal tribe of the Ambleminae (Bogan & Hoeh, 2000;
Graf & Foighil, 2000; Hoeh, Bogan & Heard, 2001). It will be interesting to see if future
phylogenetic studies on such unresolved taxa will support or undermine our proposed
model of character evolution.
5.5.3.2. Convergences and implications for probable functional morphologies
The pronounced difference between juvenile and adult shell sculpture implies that
rugae are unlikely to play an important role in adults but may be extremely important in
juvenile stages. Other differences between juvenile and adult shell morphology have been
observed, for example, in microsculpture (Chapter 6; Zieritz et al., in press) and lime
content (Coker et al., 1921). Such morphological discrepancies of the two life stages are
likely to be associated with differences in their ecology and/or behaviour, e.g. feeding

Chapter 5 – Umbonal sculptures of Unionoida
95
habit, burrowing depth, and susceptibility to dislodgement and predation (Coker et al.,
1921; Yeager, Cherry & Neves, 1994). In addition, there is some evidence of juveniles
requiring different environments than adults do (Hruska, 1999; Geist & Auerswald, 2007),
but due to their small size and low abundances, optimal habitat conditions for individual
species are generally unknown.
Understanding the functional morphology of beak sculpture could potentially be
helpful for reconstructing differences in juvenile habitat requirements of unionoid species.
A number of possible functions has been proposed for rugae, but most likely functional
morphologies are (1) anchoring, (2) shell reinforcement, and/or (3) burrowing (Watters,
1994; Checa & Jiménez-Jiménez, 2003a). All these three hypotheses are supported by our
observation that species characteristic of fast flowing, coarse sediment habitats often
exhibit ‘wrinkled’ or ‘nodulous’ umbos (e.g. Unio spp. (Killeen, Aldridge & Oliver, 2004),
Quadrula spp. (Parmalee & Bogan, 1998)). Repeated convergences to completely
‘smooth’ beaks occurred mainly in either (a) taxa characteristic of standing or slow
flowing habitats with soft-sediment (e.g. D. delphinulus, Velesunioninae), or (b)
“lanceolate” taxa which, in general, are deep and occasionally fast burrowers (e.g.
P. tanganyicensis, most Mycetopodidae and Iridininae). Whereas both these types of
convergences conform with an anchoring and/or reinforcing function of rugae, absence of
umbonal sculpture in several deeply burrowing unionoid species contradicts a burrowing
function of umbonal rugae.


CHAPTER 6
VARIABILITY, FUNCTION AND PHYLOGENETIC SIGNIFICANCE OF
PERIOSTRACAL MICROPROJECTIONS IN PALAEOHETERODONT BIVALVES

“Mrs Jalin: George?
Mr Jalin: Yes, Gladys.
Mrs Jalin: There's a man at the door with a moustache.
Mr Jalin: Tell him I've already got one. All right, all right. What's he want
then?
Mrs Jalin: He says do we want a documentary on molluscs.
Mr Jalin: Molluscs?!
Mrs Jalin: Yes.
Mr Jalin: What's he mean, molluscs?
Mrs Jalin: MOLLUSCS!! GASTROPODS! LAMELLIBRANCHS!
CEPHALOPODS!
Mr Jalin: Oh molluscs, I thought you said bacon.”
Monty Python

99
CHAPTER 6
VARIABILITY, FUNCTION AND PHYLOGENETIC SIGNIFICANCE OF PERIOSTRACAL MICROPROJECTIONS IN PALAEOHETERODONT
BIVALVES
6.1. ABSTRACT
Microprojections in the Palaeoheterodonta (Trigonioida and Unionoida) have been
little studied but could be important characters for resolving questions on phylogeny and
ecology of these bivalves. By investigating 26 unionoid and one trigonioid species using
scanning electron microscopy, we identified three types of periostracal microprojections.
(1) Microridges were present only in one respective species of the two unionoid families
Mycetopodidae (Anodontites trapesialis) and Iridinidae (Chambardia bourguignati), and
may represent a synapomorphy for the mycetopodid-iridinid clade. In A. trapesialis,
microridges were additionally equipped with (2) flag-like projections (microfringes),
possibly a synapomorphic character for the Mycetopodidae. Examination of partially
bleached specimens indicated that both microridges and microfringes are predominantly or
purely organic. In contrast, previously undescribed (3) spicule-like spikes represent
calcifications within the periostracum. These were found in 20 of the 27 species and four
of the six unionoid families. Particularly large and abundant spikes were exhibited in
umbonal (juvenile) parts of the shell, and species characteristic of fast flowing habitats.
These structures may thus serve in protecting the periostracum and shell underneath,
and/or stabilising life position by increasing shell friction. Microfringes and microridges,
on the other hand, possibly aid in the orientation of the mussel within the sediment.
6.2. INTRODUCTION
The outermost layer of bivalve shells, the periostracum, is a multilayered, quinone-
tanned protein (Beedham, 1958), primarily serving as a substrate for growth of the
calcified shell and allowing the necessary conditions for crystallisation to be achieved and
maintained (e.g. Taylor & Kennedy, 1969; Wilbur & Saleuddin, 1983; Checa, 2000). A
further periostracal function is that of a waterproof covering, preventing corrosion of the
underlying shell by acidic or poorly buffered waters (Taylor & Kennedy, 1969; Tevesz &

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
100
Carter, 1980) and therefore of particular importance in freshwater mussels (Unionoida).
Finally, Harper (1997) has shown that differences in periostracal thickness can constrain
formation of various shell morphologies and in particular, sculpture.
Unionoids comprise an estimated 840 species from six families and inhabit
freshwater habitats on all continents except Antarctica (Graf & Cummings, 2007b). Their
adults are suspension feeders and, with exception of the freshwater oysters (Etheriidae),
spend most of their lives in a (partially) buried position, though most species are also
capable of considerable horizontal movement across the sediment (Balfour & Smock,
1995; Amyot & Downing, 1997). Most taxonomic classifications consider the marine,
monogeneric order Trigonioida as the closest living relatives of the Unionoida, and group
these two bivalve orders into the subclass Palaeoheterodonta (e.g. Bieler & Mikkelsen,
1996; Hoeh et al., 1998; Graf & Cummings, 2006b). While structure, composition,
mineralogy and calcification processes of palaeoheterodont shells are relatively well
understood (e.g. Taylor & Kennedy, 1969; Taylor, Kennedy & Hall, 1969; Tevesz &
Carter, 1980; Carter, 1990; Checa, 2000; Checa & Rodríguez-Navarro, 2001; Marie et al.,
2007), surprisingly little is known about surface ‘microprojections’ in trigonioids and
unionoids. These structures usually are of merely a few µm in size and thus, can be made
visible only by high magnification microscopy.
Periostracal microsculptures are known to occur in several marine bivalve orders.
Some of these structures are not calcified (e.g. in Arcoida) or are secreted over the exterior
of the periostracum and thus, are not periostracal in origin (e.g. "periostracal" hairs of
some Mytiloidea) (Bottjer & Carter, 1980). Nevertheless, though initially believed to be a
purely organic layer (Gray, 1825), calcifications “within” the periostracum have since been
reported from members of various bivalve orders: Praenuculida, Mytiloida,
Modiomorphida, Trigonioida, Carditoida, Anomalodesmata, Veneroida and Myoida
(Aller, 1974; Carter & Aller, 1975; Newell & Boyd, 1975; Carter, 1978; Bottjer & Carter,
1980; Carter, 1990; Schneider & Carter, 2001; Glover & Taylor, 2010; Checa & Harper, in
press). Such calcifications are known to take up various shapes and sizes, up to more than
400 µm long needles in venerids (Glover & Taylor, 2010). However, detailed
investigations of the morphology and formation of these calcified structures are restricted
to Aller (1974), Glover & Taylor (2010) and Checa & Harper(in press). Thus, for the
majority of the taxa mentioned above, we do not know if these structures are actually
periostracal in origin or part of the calcified shell underneath.

Chapter 6 – Periostracal microprojections in Palaeoheterodonta
101
With respect to the Unionoida, we are aware of merely a single paper using high
magnification (electron) microscopy to describe periostracal microprojections in two
species of Anodontites, belonging to the unionoid family Mycetopodidae (Callil &
Mansur, 2005). Based on their observations on this one genus, these authors classified
periostracum projections into four size-related types: (1) macroscopically visible ‘folds’,
(2) mesoscopically visible ‘corrugations’, (3) ‘microridges’, and (4) ‘microfringes’ and
‘spikes’. Structures of type 1 and 2 have been described from several South American,
Chinese and African taxa of the mycetopodid (e.g. Anodontites), unionid (e.g. Cuneopsis)
and iridinid (e.g. Pleiodon) families (Marshall, 1926). However, apart from the
aforementioned mycetopodid taxa, nothing is known about occurrence of periostracal
microridges, microfringes and/or spikes in any other palaeoheterodont family and species.
What is more, we do not know anything about the mineralisation status (i.e. calcified or
organic) and formation process of these structures.
As a result of this lack of knowledge, periostracal microsculptures have so far not
been considered in phylogenetic analyses of Palaeoheterodonta (e.g. Hoeh, Bogan &
Heard, 2001; Graf & Cummings, 2006b). Including presence/absence data of homologous
types of microprojections in such studies might, however, help resolving questions
regarding taxonomic relationships and character evolution within this group of bivalves,
both issues currently under considerable debate (see Graf & Cummings, 2006b; Hoeh et
al., 2009; Graf & Cummings, 2010a). Furthermore, understanding the functional
morphologies of sculptures could potentially lead to new insights on habitat preferences
and behavioural characteristics of certain bivalve species and/or life stages. This
knowledge is of particular importance for developing conservation measures of the many
endangered freshwater bivalve taxa (Bogan, 1993; Williams et al., 1993; Lydeard et al.,
2004). All this necessitates, however, a basic knowledge on occurrence, morphology and
distribution of periostracal microprojections in the palaeoheterodont bivalves; data we are
currently lacking.
This study aims to fill this gap of knowledge. Incorporating species of all extant
palaeoheterodont families, we provide a first dataset on (1) morphology and mineralisation
status of their periostracal microprojections, (2) presence/absence of the different types of
microprojections across their phylogeny, and (3) inter- and intraspecific variability of
morphology, abundance and distribution of their microprojections. Potential implications
on palaeoheterodont phylogeny, character evolution and functional morphologies of the
structures are discussed.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
102
(Right page) Table 6.1. Classification, collection locality and measurements of microprojections of the specimens examined. Phylogeny after Graf & Cummings (2007b). Abbreviations in brackets indicate source of specimens if not from private collection of one of the authors: UMZC, University Museum of Zoology Cambridge; NHML, Natural History Museum London. Other abbreviations: b, boss; mf, microfringe; mr, microridge; n.d., no data; s, spike; SD, standard deviation; X , mean.
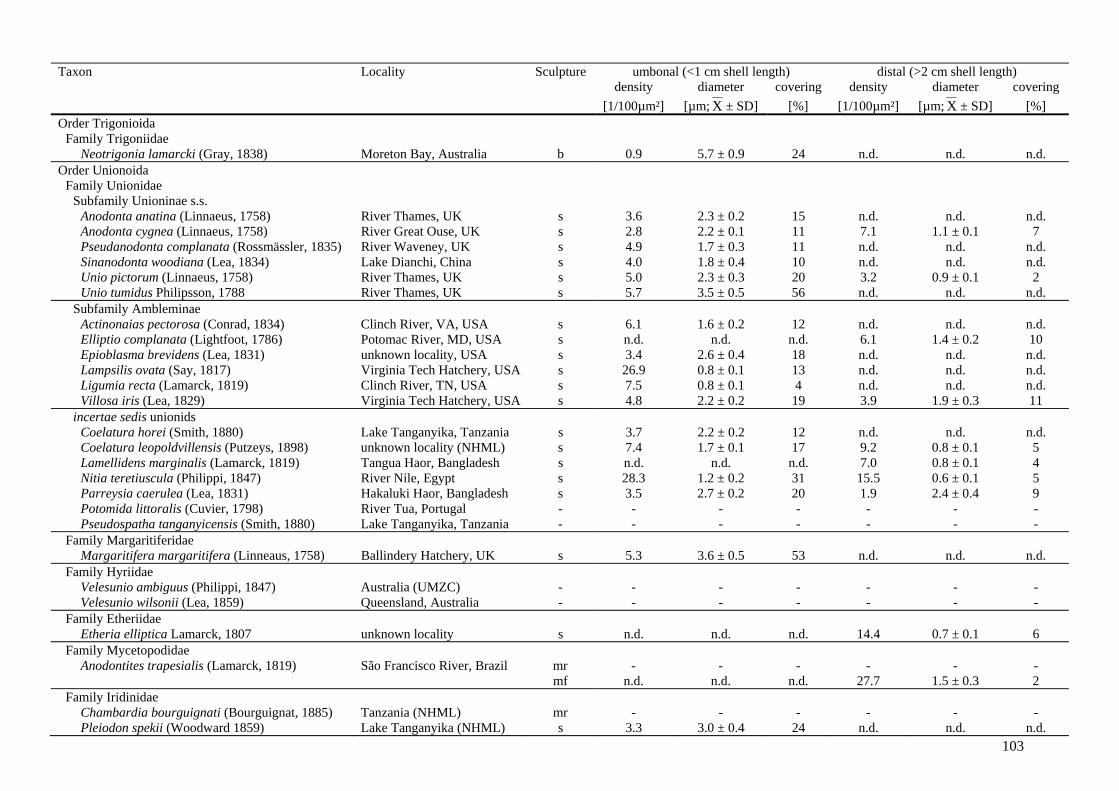
103
Taxon Locality Sculpture umbonal (<1 cm shell length) distal (>2 cm shell length) density diameter covering density diameter covering [1/100µm²] [µm; X ± SD] [%] [1/100µm²] [µm; X ± SD] [%] Order Trigonioida
Family Trigoniidae Neotrigonia lamarcki (Gray, 1838) Moreton Bay, Australia b 0.9 5.7 ± 0.9 24 n.d. n.d. n.d.
Order Unionoida Family Unionidae
Subfamily Unioninae s.s. Anodonta anatina (Linnaeus, 1758) River Thames, UK s 3.6 2.3 ± 0.2 15 n.d. n.d. n.d. Anodonta cygnea (Linnaeus, 1758) River Great Ouse, UK s 2.8 2.2 ± 0.1 11 7.1 1.1 ± 0.1 7 Pseudanodonta complanata (Rossmässler, 1835) River Waveney, UK s 4.9 1.7 ± 0.3 11 n.d. n.d. n.d. Sinanodonta woodiana (Lea, 1834) Lake Dianchi, China s 4.0 1.8 ± 0.4 10 n.d. n.d. n.d. Unio pictorum (Linnaeus, 1758) River Thames, UK s 5.0 2.3 ± 0.3 20 3.2 0.9 ± 0.1 2 Unio tumidus Philipsson, 1788 River Thames, UK s 5.7 3.5 ± 0.5 56 n.d. n.d. n.d.
Subfamily Ambleminae Actinonaias pectorosa (Conrad, 1834) Clinch River, VA, USA s 6.1 1.6 ± 0.2 12 n.d. n.d. n.d. Elliptio complanata (Lightfoot, 1786) Potomac River, MD, USA s n.d. n.d. n.d. 6.1 1.4 ± 0.2 10 Epioblasma brevidens (Lea, 1831) unknown locality, USA s 3.4 2.6 ± 0.4 18 n.d. n.d. n.d. Lampsilis ovata (Say, 1817) Virginia Tech Hatchery, USA s 26.9 0.8 ± 0.1 13 n.d. n.d. n.d. Ligumia recta (Lamarck, 1819) Clinch River, TN, USA s 7.5 0.8 ± 0.1 4 n.d. n.d. n.d. Villosa iris (Lea, 1829) Virginia Tech Hatchery, USA s 4.8 2.2 ± 0.2 19 3.9 1.9 ± 0.3 11
incertae sedis unionids Coelatura horei (Smith, 1880) Lake Tanganyika, Tanzania s 3.7 2.2 ± 0.2 12 n.d. n.d. n.d. Coelatura leopoldvillensis (Putzeys, 1898) unknown locality (NHML) s 7.4 1.7 ± 0.1 17 9.2 0.8 ± 0.1 5 Lamellidens marginalis (Lamarck, 1819) Tangua Haor, Bangladesh s n.d. n.d. n.d. 7.0 0.8 ± 0.1 4 Nitia teretiuscula (Philippi, 1847) River Nile, Egypt s 28.3 1.2 ± 0.2 31 15.5 0.6 ± 0.1 5 Parreysia caerulea (Lea, 1831) Hakaluki Haor, Bangladesh s 3.5 2.7 ± 0.2 20 1.9 2.4 ± 0.4 9 Potomida littoralis (Cuvier, 1798) River Tua, Portugal - - - - - - - Pseudospatha tanganyicensis (Smith, 1880) Lake Tanganyika, Tanzania - - - - - - -
Family Margaritiferidae Margaritifera margaritifera (Linneaus, 1758) Ballindery Hatchery, UK s 5.3 3.6 ± 0.5 53 n.d. n.d. n.d.
Family Hyriidae Velesunio ambiguus (Philippi, 1847) Australia (UMZC) - - - - - - - Velesunio wilsonii (Lea, 1859) Queensland, Australia - - - - - - -
Family Etheriidae Etheria elliptica Lamarck, 1807 unknown locality s n.d. n.d. n.d. 14.4 0.7 ± 0.1 6
Family Mycetopodidae Anodontites trapesialis (Lamarck, 1819) São Francisco River, Brazil mr - - - - - - mf n.d. n.d. n.d. 27.7 1.5 ± 0.3 2
Family Iridinidae Chambardia bourguignati (Bourguignat, 1885) Tanzania (NHML) mr - - - - - - Pleiodon spekii (Woodward 1859) Lake Tanganyika (NHML) s 3.3 3.0 ± 0.4 24 n.d. n.d. n.d.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
104
6.3. MATERIALS AND METHODS
Shell samples with well preserved periostracum from 27 species, representing at
least one species from each of the six unionoid families and one species of the Trigonioida,
were collected or obtained from various sources (Table 6.1). During the initial stages of
this study, it became apparent that, if present at all, microsculptures were more abundant in
the umbonal (juvenile) region of the shell and thus, preferably young specimens were
studied. However, in some cases only large individuals could be obtained. These were
fragmented, and fragments with well preserved periostracum were selected from as close
to the umbo as possible. One to three individuals per species were analysed for occurrence
of periostracal microprojections using scanning electron microscopy (SEM).
To investigate if periostracal projections were calcified or purely organic, one to two
shells of Anodonta anatina, Unio tumidus, Lampsilis ovata and Anodontites trapesialis
were immersed in commercial bleach for 10-120 minutes (exact time given in respective
Figure legends) until the periostracum was partly digested, and subsequently washed in
distilled water.
One valve of A. anatina, Sinanodonta woodiana, Unio pictorum and U. tumidus was
prepared for visualisation of periostracal projections in transverse sections. For this
purpose, shells were embedded in resin, radially sectioned with a circular rock saw,
polished with a series of grit sizes, etched for 15 seconds in 5% HCl and subsequently
washed in distilled water.
Last, to examine a possible association of periostracal thickness and
presence/absence of microprojections, shell fragments of 12 unionoid species from five
families were selected for determination of periostracum thickness values.
All samples except those that had been bleached and those prepared for
determination of periostracal thickness were cleaned in an ultrasonic bath for
approximately 20 seconds. All samples were then dried at room temperature and
subsequently fixed on stubs with carbon adhesive tape, super glue or silver glue. After
sputter-coating the samples with gold the shell surface area was scanned, and images were
obtained using a JEOL 820 scanning electron microscope.
Measurements were made on image files obtained by SEM using program ImageJ
(Rasband, 2008). Images were scaled using scale bars obtained by SEM. Maximum and
minimum thickness of the periostracum was measured in one picture of a fragment in
transverse perspective per species. Densities and average sizes (diameter and height) of

Chapter 6 – Periostracal microprojections in Palaeoheterodonta
105
discrete periostracal microprojections (i.e. spikes, bosses and microfringes) were estimated
for one specimen of each species exhibiting the respective structures. Dependent on the
availability of preserved periostracum, this was done on either (1) only the umbonal region
(i.e. <1 cm shell length), (2) only the distal region (i.e. >2 cm shell length) or (3) both the
umbonal and distal region. For this purpose, microprojections were counted on a randomly
selected area of one SEM image, and abundance per 100 µm² shell area calculated. On the
same images, 10 structures of each type present were selected randomly, their basal
maximum diameter measured, and average and standard deviation of diameters calculated.
Average proportion of shell area covered by the given microsculpture was estimated using
average basal diameter and density values.
All SEM specimens examined have been deposited at the Field Museum Chicago
(Neotrigonia lamarcki), the University Museum of Zoology Cambridge (Velesunio
ambiguus (Philippi, 1847): UMZC 2010.25) or the Natural History Museum London (all
remaining species: NHML Aq Zoo-2010-115).
6.4. RESULTS
6.4.1. OCCURRENCE AND MORPHOLOGY OF PERIOSTRACAL MICROPROJECTIONS IN PALAEOHETERODONTA
Periostracal microprojections present in the studied shell material can generally be
classified into three types: (1) microridges (Fig. 6.1A), (2) flap-like microfringes (Fig.
6.1A,B), and (3) spicule-like periostracal projections (spikes) (Fig. 6.2).
Figure 6.1. A-B. Examples of microfringes and microridges in two unionoid families. A. Anodontites trapesialis (Mycetopodidae). B. Chambardia bourguignati (Iridinidae). Abbreviations: mf, microfringe; mr, microridge.
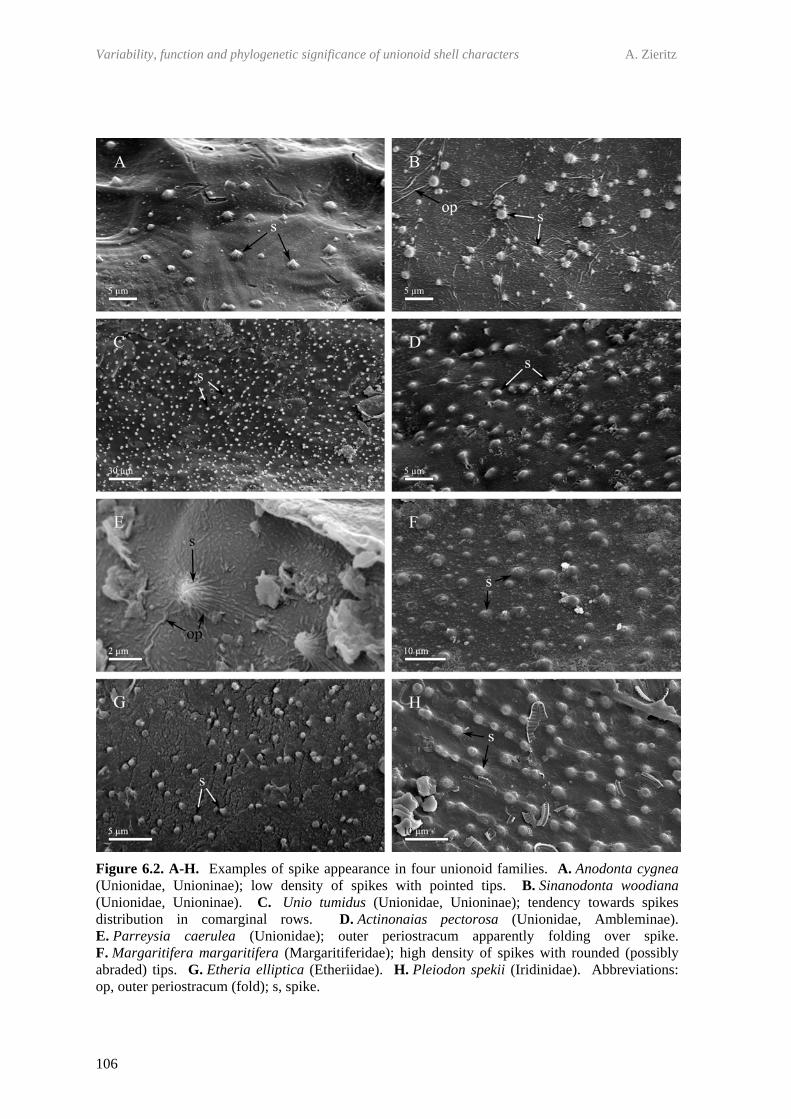
Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
106
Figure 6.2. A-H. Examples of spike appearance in four unionoid families. A. Anodonta cygnea (Unionidae, Unioninae); low density of spikes with pointed tips. B. Sinanodonta woodiana (Unionidae, Unioninae). C. Unio tumidus (Unionidae, Unioninae); tendency towards spikes distribution in comarginal rows. D. Actinonaias pectorosa (Unionidae, Ambleminae). E. Parreysia caerulea (Unionidae); outer periostracum apparently folding over spike. F. Margaritifera margaritifera (Margaritiferidae); high density of spikes with rounded (possibly abraded) tips. G. Etheria elliptica (Etheriidae). H. Pleiodon spekii (Iridinidae). Abbreviations: op, outer periostracum (fold); s, spike.
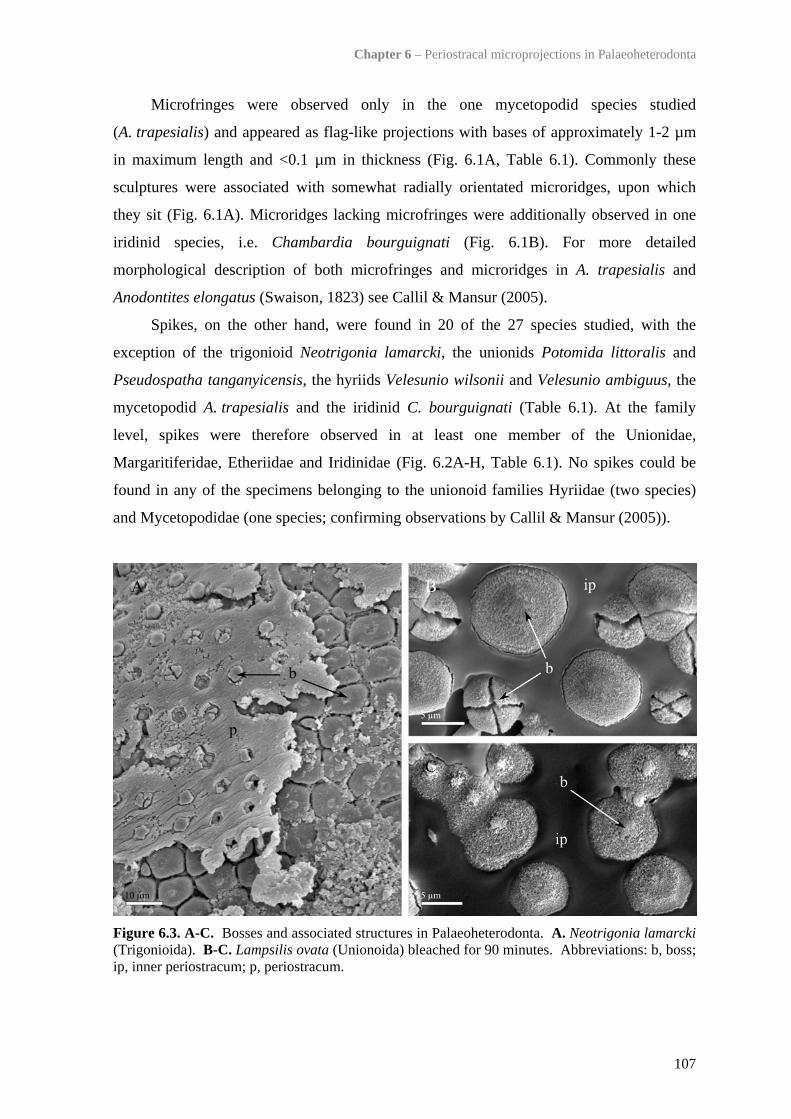
Chapter 6 – Periostracal microprojections in Palaeoheterodonta
107
Microfringes were observed only in the one mycetopodid species studied
(A. trapesialis) and appeared as flag-like projections with bases of approximately 1-2 µm
in maximum length and <0.1 µm in thickness (Fig. 6.1A, Table 6.1). Commonly these
sculptures were associated with somewhat radially orientated microridges, upon which
they sit (Fig. 6.1A). Microridges lacking microfringes were additionally observed in one
iridinid species, i.e. Chambardia bourguignati (Fig. 6.1B). For more detailed
morphological description of both microfringes and microridges in A. trapesialis and
Anodontites elongatus (Swaison, 1823) see Callil & Mansur (2005).
Spikes, on the other hand, were found in 20 of the 27 species studied, with the
exception of the trigonioid Neotrigonia lamarcki, the unionids Potomida littoralis and
Pseudospatha tanganyicensis, the hyriids Velesunio wilsonii and Velesunio ambiguus, the
mycetopodid A. trapesialis and the iridinid C. bourguignati (Table 6.1). At the family
level, spikes were therefore observed in at least one member of the Unionidae,
Margaritiferidae, Etheriidae and Iridinidae (Fig. 6.2A-H, Table 6.1). No spikes could be
found in any of the specimens belonging to the unionoid families Hyriidae (two species)
and Mycetopodidae (one species; confirming observations by Callil & Mansur (2005)).
Figure 6.3. A-C. Bosses and associated structures in Palaeoheterodonta. A. Neotrigonia lamarcki (Trigonioida). B-C. Lampsilis ovata (Unionoida) bleached for 90 minutes. Abbreviations: b, boss; ip, inner periostracum; p, periostracum.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
108
Finally, shell surfaces of specimens of the trigonioid N. lamarcki were covered with
regularly distributed, pronounced bosses, apparently associated with the prismatic layer
underneath (Fig. 6.3A). As described by Taylor, Kennedy & Hall (1969) and Newell &
Boyd (1975), these structures probably represent the initial calcification centres of prisms.
6.4.2. VARIATION IN SPIKE MORPHOLOGY, ABUNDANCE AND DISTRIBUTION ACROSS THE SHELL
Spikes typically appeared in a conical shape with a somewhat circular to elliptical
base ranging from less than 1 µm to approximately 5 µm in diameter, and protruded less
than 1 µm to approximately 2 µm above the periostracum (Fig 6.2A-H). Pointedness of
tips varied both between and within species. Anodonta cygnea had, for example,
predominantly rather pointed spikes (Fig. 6.2A), whereas Margaritifera margaritifera
spikes appeared uniformly blunt across the whole shell area (Fig. 6.2F). With regard to
variation within single individuals, a more blunted appearance was especially common in
older parts of shells.
Maximum spike diameter usually varied considerably within a single specimen
(Table 6.1) and a range of different sizes could even be seen on only a few µm² of shell
area (e.g. Fig. 6.2A,B,F). Nevertheless, in all six species for which available shell material
allowed us to compare umbonal and distal regions, average spike diameter was larger in
the umbonal compared to the distal part of the shell (Table 6.1). This implies a tendency of
spikes to become smaller towards the shell margin. Besides this variation within species
and individuals, some interspecific differences in spike size seemed striking. For example,
L. ovata and Ligumia recta spikes were on average less than 1 µm in diameter, despite the
fact that juvenile shells were analysed (Table 6.1). Equally, on the four, though relatively
distal fragments of two freshwater oyster (Etheria elliptica) specimens studied, only spikes
< 1 µm in diameter and height, were found (Fig. 6.2G, Table 6.1). In contrast, some of the
largest spikes with basal diameters of about 4-4.5 µm were observed on shells of
M. margaritifera (Fig. 6.2F), Unio tumidus (Fig. 6.2C) and Pleiodon spekii (Fig. 6.2H)
(Table 6.1).
Besides this variation in spikes size, our data suggest that spike count per unit area
differs considerably between species. Partly this appears to be inversely related to average
spike size, i.e. spikes of relatively small diameters regularly appeared at relatively high
densities (Table 6.1; e.g. in L. ovata, Coelatura leopoldvillensis and Actinonaias
pectorosa). Nevertheless, in some species, high spike densities were observed despite large

Chapter 6 – Periostracal microprojections in Palaeoheterodonta
109
spike sizes (Table 6.1; e.g. M. margaritifera, U. tumidus). In addition to this interspecific
variation, spike density was also highly variable within the same specimen. In some
species, spikes were more abundant at the umbonal region when compared to more
recently formed shell areas (Table 6.1; i.e. U. pictorum, V. iris and Parreysia caerulea),
while other species showed the opposite pattern (Table 6.1; i.e. A. cygnea and
C. leopoldvillensis).
Figure 6.4. A-B. Occurrence of comarginal growth bands with and without spikes in Anodonta cygnea (Unionidae, Unioninae). A. Low magnification of shell surface (inset shown in B). B. High magnification of bands with and without spikes. Abbreviation: s, spike.
By combining our data on average spike diameters and densities we were able to
investigate trends in the proportion of the shell segment covered by spikes. On a species
level, percentage of umbonal areas covered by spikes was over five times higher in
U. tumidus and M. margaritifera than in L. recta and S. woodiana (Table 6.1). As with
spike size, a clear pattern emerged when looking at distribution across single shells.
Interestingly, in all the six specimens where both umbonal and distal shell fragments were
analysed, relative spike coverage was always higher in the umbonal compared to the distal
region (Table 6.1). What is more, at later parts of the shell a pattern of comarginal bands
lacking spikes intercepted by bands carrying spikes was found in several species (Fig.
6.4A,B). Such spike-free growth bands were observed in A. cygnea, V. iris, Epioblasma
brevidens and C. leopoldvillensis. Late shell fragments of even larger sized individuals of

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
110
some species (e.g. U. pictorum, A. cygnea) completely lacked spikes, indicating a decrease
of spike density from early to late shell parts.
A consistent pattern regarding spike density or size distribution along the anterior-
posterior axis could not be found. However, in some individuals such as the one available
L. recta specimen, spikes were more abundant at the posterior compared to the anterior
region of the shell.
Last, in the palaeoheterodont species studied, spikes usually appear to be randomly
distributed across the shell surface (Fig. 6.2A,B,D,F,G,H). However, in one individual of
U. tumidus, a tendency to a distribution in comarginal rows was observed (Fig. 6.2C). We
were not able to detect any consistency of this distribution pattern however, i.e. spikes in
other areas of the same individual were distributed randomly.
6.4.3. MINERALISATION STATUS OF PERIOSTRACAL MICROPROJECTIONS
Though mostly of a somewhat circular shape, in some cases, spikes exhibited an
almost hexagonal outline (Fig. 6.5), suggesting that these represent singular or twinned
aragonitic crystals. Further evidence for a calcified nature of these structures is provided
by spike skeletons that remained after partly bleaching organic shell parts of A. anatina,
U. tumidus and L. ovata specimens (Fig. 6.6B). On the other hand, no projections but only
the bases of microfringes were to be seen after bleaching of A. trapesialis shell fragments
known to carry microridges and –fringes (Fig. 6.6A). This implies that apart from a
possibly mineralised core of microfringes, these structures consist of organic material
which was oxidised during the bleaching process.
Figure 6.5. Periostracal spikes in Unio pictorum (Unionidae, Unioninae). Abbreviation: s, spike.
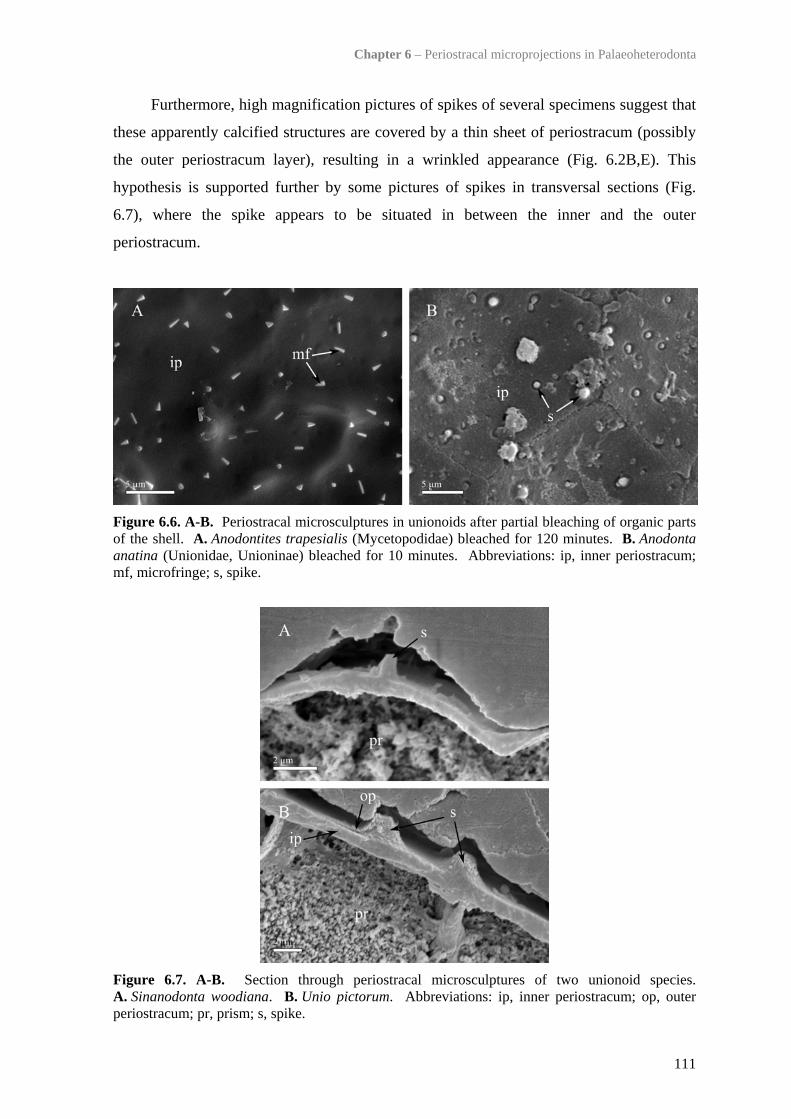
Chapter 6 – Periostracal microprojections in Palaeoheterodonta
111
Furthermore, high magnification pictures of spikes of several specimens suggest that
these apparently calcified structures are covered by a thin sheet of periostracum (possibly
the outer periostracum layer), resulting in a wrinkled appearance (Fig. 6.2B,E). This
hypothesis is supported further by some pictures of spikes in transversal sections (Fig.
6.7), where the spike appears to be situated in between the inner and the outer
periostracum.
Figure 6.6. A-B. Periostracal microsculptures in unionoids after partial bleaching of organic parts of the shell. A. Anodontites trapesialis (Mycetopodidae) bleached for 120 minutes. B. Anodonta anatina (Unionidae, Unioninae) bleached for 10 minutes. Abbreviations: ip, inner periostracum; mf, microfringe; s, spike.
Figure 6.7. A-B. Section through periostracal microsculptures of two unionoid species. A. Sinanodonta woodiana. B. Unio pictorum. Abbreviations: ip, inner periostracum; op, outer periostracum; pr, prism; s, spike.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
112
6.4.4. TRIGONIOID ‘BOSSES’
The one trigonioid studied, N. lamarcki, possessed large (i.e. average diameter of 5.7
µm), regularly-spaced bosses (Table 6.1, Fig. 6.3A). These bosses were largest and best
developed in troughs between ribs. The fact that one boss always originates from the
centre of one prism (Fig. 6.3A) indicates that these structures represent the calcification
centres of prisms. Although no bosses could be observed in any untreated unionoid
specimens, similar structures appeared after partial bleaching of organic shell parts in
L. ovata (Fig. 6.3B,C) and three other unionoid species that were examined in this respect.
Unionoid bosses equally represent calcification centres of prisms but never protrude above
the periostracum as trigonioid bosses do.
6.4.5. INFLUENCE OF PERIOSTRACAL THICKNESS ON PRESENCE/ABSENCE OF MICROPROJECTIONS
No correlation could be observed between presence/absence of microprojections and
thickness of periostracum across 12 palaeoheterodont species analysed (Table 6.2). Thus,
spikes were present in both species with extremely thin (e.g. S. woodiana) and extremely
thick (e.g. M. margaritifera) periostraca.
Table 6.2. Minimum and maximum periostracum thickness values measured for 15 palaeoheterodont species. Sorting order from lowest to largest maximum thickness. Abbreviations: b, boss; mf, microfringe; mr, microridge; s, spike.
Species Type(s) of
microprojection present
Approximate periostracum thickness [µm]
Source (if not this study)
Sinanodonta woodiana s 0.5 – 1 Unio pictorum s 0.5 – 2 Pleiodon spekii s 1.5 – 2.5 Potomida littoralis - 2 – 3 Checa (2000) Anodonta anatina s 2.5 – 3.5 Lampsilis ovata s 4 – 5 Unio tumidus s 4.5 – 6 Pseudospatha tanganyicensis - 4.5 – 6.5 Velesunio ambiguus - 5 – 6.5 Anodontites trapesialis mr, mf 4 – 7 Neotrigonia spp. b 3 – 12* Harper (1997) Chambardia bourguignati mr 10 – 12.5 Etheria elliptica s 14* Harper (1997) Coelatura horei s 10 – 15.5 Margaritifera margaritifera s 16.5 – 20.5
*values represent mean values (for four species in the case of Neotrigonia spp.)

Chapter 6 – Periostracal microprojections in Palaeoheterodonta
113
6.5. DISCUSSION
6.5.1. OCCURRENCE OF PERIOSTRACAL MICROPROJECTIONS ACROSS THE UNIONOID PHYLOGENY
In the 27 palaeoheterodont taxa studied, three different types of periostracal
microprojections were identified. Non-calcified ‘microridges’ were present only in one
mycetopodid (A. trapesialis) and one iridinid (C. bourguignati) species. In A. trapesialis,
these were additionally accompanied by flag-shaped ‘microfringes’. Though calcified
periostracal ‘spikes’ were present in most shells examined, we were not able to find these
structures in the trigonioid N. lamarcki and six species from four of the six unionoid
families, including all hyriid and mycetopodid species analysed.
A further type of microprojection was observed only in the one trigonioid species
studied. These ‘bosses’ protrude from the prismatic layer of the shell itself and are thus
possibly not periostracal in origin.
The main objective of this study was to provide a first overview on periostracal
protrusions exhibited across the palaeoheterodont phylogeny. As a consequence, only a
limited number of specimens per species (usually one) could be investigated, which
precluded investigation of intraspecific variability of these structures. Nevertheless, despite
the limited and preliminary nature of our dataset, there appears to be some phylogenetic
signal in the distribution of features recorded.
Bosses, found only in N. lamarcki, most likely represent the calcification centres of
the aragonitic prisms (as previously suggested by Taylor, Kennedy & Hall (1969) and
Newell & Boyd (1975)), which ultimately “pierce” through the overlaying periostracum.
Although similar structures can be found after bleaching of the organic periostracum of
unionoid shells, bosses of freshwater palaeoheterodonts examined were never as well
developed and always covered by the periostracum. This and the fact that Newell & Boyd
(1975) also found bosses in several fossil trigonioid taxa indicates that this character could
be synapomorphic for the Trigonioida.
Despite the highly debated and insufficient understanding of most interfamilial
relationships in the Unionoida, most modern phylogenies consider the Mycetopodidae and
Iridinidae as closely related (Fig. 6.8). The fact that microridges were present exclusively
in members of these two families could therefore indicate a homologous nature of this
morphological trait, which potentially evolved only after divergence of the mycetopodid-
iridinid branch (Fig. 6.8A; phylogeny after Graf & Cummings (2006b)) or the iridinid-

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
114
mycetopodid-etheriid branch (Fig. 6.8B; phylogeny after Bogan & Hoeh (2000) and Hoeh,
Bogan & Heard (2001)). Similarly, exclusiveness of microfringes to the mycetopodids
could indicate evolution of this shell character just before this family branched off from
the remaining unionoids. Our dataset can be extended by observations on one additional
mycetopodid species by Callil & Mansur (2005), who found similar microridges and
microfringes in A. elongatus. As these conclusions are drawn from just two iridinid and
two mycetopodid species of only one genus, respectively, it will be important and
interesting to determine whether or not other genera of these unionoid families exhibit the
same morphological traits.
1 according to Graf & Cummings (2006b, 2007b) member of the Etheriidae
Figure 6.8. A-B. Presence/absence of microprojections in the palaeoheterodont families plotted on two contrasting phylogenetic cladograms (both redrawn from Hoeh et al. (2009)). A. Combined evidence phylogeny after Graf & Cummings (2006b: fig. 4). B. Combined summary tree based on COI (Bogan & Hoeh, 2000) and combined evidence data (Hoeh, Bogan & Heard, 2001). Letters indicate occurrence of microprojections in each taxon: b, bosses; mf, microfringes; mr, microridges; s, spikes. Grey circles and associated letters indicate synapomorphies of the four characters.
Due to their distinct morphology and mineralised nature, spikes, observed in all
unionoid families except hyriids and mycetopodids, should be considered as a different,
possibly non-homologous character state to the predominantly un-mineralised
microfringes. As recently discussed by Hoeh et al. (2009) and Graf & Cummings (2010a),
the position of the Hyriidae in the unionoid phylogeny is of particular uncertainty. Several
analyses based on morphology (Graf, 2000; Hoeh, Bogan & Heard, 2001), combined
morphology + mtDNA (COI) (Roe & Hoeh, 2003), and combined morphology + mtDNA
(COI) + nuclear ribosomal DNA (28S) (Graf & Cummings, 2006b) (Fig. 6.8A) support a
close relationship of hyriids and “etherioids” (i.e. Etheriidae + Mycetopodidae +

Chapter 6 – Periostracal microprojections in Palaeoheterodonta
115
Iridinidae). Alternative phylogenetic analyses based on COI sequences (Bogan & Hoeh,
2000; Hoeh et al., 2002; Walker et al., 2006), and combined morphology + COI data
(Hoeh, Bogan & Heard, 2001) (Fig. 6.8B) indicate that the Hyriidae is the most basal
family of unionoids. Against this background, absence of spikes in the hyriids could
potentially be interesting. That is, if the second phylogenetic scenario was correct, the
ability to produce spikes might have evolved only after the split of hyriids from the rest of
the unionoids.
On the other hand, based on the data available, spikes are also absent in the
Mycetopodidae, regarded as one of the most derived unionoid families. Thus, we failed to
detect spikes in A. trapesialis, and though Callil & Mansur (2005) mention the presence of
“spikes” in this species and A. elongatus, closer inspection of their figures suggests that
what they observed in fact represented merely the tips of microfringes in a situation where
the remaining part of the fringe was covered by extraneous material. Spike production
might therefore have been secondarily “switched off” in the mycetopodids. Interestingly,
all species exhibiting microridges (that is the mycetopodid Anodontites spp. and iridinid
C. bourguignati) also lacked spikes, which might imply that these two structures are never
produced at the same time. However, spikes were found in the second iridinid species
under study. Similarly, in the Unionidae both species with and without spikes were found,
though it should be noted that the phylogenetic positions of both spike-free unionid species
(P. littoralis and P. tanganyicensis) within this family are unclear (incertae sedis). In any
case, these inconsistencies in spike formation within evolutionary entities show that
expression of this character has been secondarily lost on several occasions, indicating that
spike production is probably easy to be “switched off”.
Further studies on other palaeoheterodont species will be needed to support or
contradict our preliminary hypotheses on character evolution of palaeoheterodont
microprojections. Indeed, in older shells with partly damaged periostracum,
microprojections were often very difficult to find and thus, future studies might well
identify these structures in species we now describe as “free of microprojections”.
6.5.2. MORPHOLOGICAL VARIATION AND POSSIBLE FUNCTIONAL MORPHOLOGIES OF STRUCTURES
The morphological range of palaeoheterodont spikes appeared as rather uniform,
especially when compared to spikes in the Anomalodesmata, which are known to exhibit
morphologies ranging from pointed-conical spikes to flat plaques (Checa & Harper, in

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
116
press). Nevertheless, a certain amount of variation was observed in the degree of
pointedness, size and abundance of spikes, both between species and across the shell of
single specimens belonging to the Palaeoheterodonta.
A trend of pointed spikes at newly produced parts of the shell towards gradually
more blunt spikes closer to the umbonal area was observed in several specimens and
indicates that originally pointed spikes become abraded by the surrounding sediment.
Further support for this hypothesis is provided when comparing spikes of the freshwater
pearl mussel M. margaritifera with that of the swan mussel A. cygnea. The former species
is known to live in oligotrophic streams and rivers with coarse substrate and often high
current velocities (Altnöder, 1926; Killeen, Aldridge & Oliver, 2004). These habitat
features would be expected to result in a fast abrasion and blunt appearance of spikes
across the whole of the shell. The spikes of the A. cygnea specimen, which was collected
from a slow-flowing, muddy habitat, typical for this species (Stone et al., 1982; Killeen,
Aldridge & Oliver, 2004), on the other hand, were almost continuously “fresh” and
pointed. Consequently, one function of spikes of the (semi)infaunally living unionoids
could be a mechanical protection of the (inner) periostracum, which is of particular
importance to freshwater mussels. Additionally or alternatively, a spiky shell surface will
also increase friction of the shell within the sediment. Thus, Aller (1974) hypothesised that
spikes in the infaunal marine anomalodesmatan genus Laternula function either as active
burrowing aids or stabilisers of life position (by increasing friction force).
Several additional observations support two of the aforementioned functions of
palaeoheterodont spikes, i.e. (1) protection of the periostracum and underlying shell, and
(2) stabilising the mussel’s life position. Firstly, a particularly large proportion of shell
area was covered by comparatively large and densely packed spikes in U. tumidus and
M. margaritifera. Both these species prefer fast flowing stream and river habitats (Killeen,
Aldridge & Oliver, 2004), and an increased friction would improve anchoring of the shell
in the sediment and thus, withstanding dislodgement by high current velocities. A. cygnea
and S. woodiana, on the other hand, occupy slower flowing rivers and/or lakes (Killeen,
Aldridge & Oliver, 2004) and appeared at the other end of the scale regarding spike size
and shell area covered by spikes. Only very small spikes were also found in E. elliptica,
which can perhaps be explained by this species’ unique life habit. Freshwater oysters are
cemented to hard substrate by one valve (Yonge, 1962), which results in the reduction of
shell abrasion by sediment and consequently, the hypothetical advantage a shell with
increased friction would bring. Last, an absolutely smooth periostracum was presented by

Chapter 6 – Periostracal microprojections in Palaeoheterodonta
117
the African unionid P. tanganyicensis. Although almost nothing is known about the
ecology of this bivalve, it has been observed to be an extremely fast burrower (E. Michel,
personal communication); secondary loss of spike production might aid this capability.
In addition to differences between species, spike size varied considerably within
single specimens. Partly this is probably due to variability in ‘spike age’ (developmental
stage of spikes). Nevertheless, we did find a consistent trend of spikes becoming smaller
and occupying a smaller proportion of the surface area from umbonal to distal shell parts.
On a functional level, this pattern would equal greater shell friction for young individuals
compared to older ones. Juvenile unionoids are known to live interstitially as deposit
feeders up to a certain life stage, when they change to a semi-infaunal, suspension feeding
life habit (Yeager, Cherry & Neves, 1994). Higher spike density would thus increase
friction and consequently, stability of the juvenile within the sediment. Additionally or
alternatively, a spiky surface would be expected to alter flow patterns around the shell,
which could be of particular importance to small/juvenile mussels. Last, the burrowing
habit of trigonioids and unionoids makes the umbonal area the shell part most liable to
erosion (Trueman, 1966a). Thus, a higher abundance and size of spikes in this shell region
would be expected if spikes serve as a protection of the shell underneath.
Interestingly, Callil & Mansur (2005) found a similar pattern of higher abundance of
microfringes in juvenile compared to later parts of A. trapesialis and A. elongatus shells.
These authors suggest an association of these structures with increased sensitivity to water
flow, possibly aiding the juvenile in orientating itself within the sediment. Given their
extremely thin (<0.1 µm) and mostly uncalcified morphology, mycetopodid microfringes
are unlikely to offer any notable frictional improvements. Their function is thus likely to
differ from that of other unionoids’ spikes. Although Callil & Mansur (2005) do not give
any suggestion for functional morphology of microridges, their morphological
characteristics, i.e. parallel rows of ridges, may be effective in aiding the orientation of
mussels.
Apart from the functional explanations given above, presence/absence and
development of certain types of periostracal microprojections might be associated with
other morphological characteristics of the shell, in particular with thickness and
composition of the periostracum. For example, spike production might be ceased below a
certain threshold of periostracal thickness. However, we failed to detect any obvious
correlation of occurrence of microprojection and thickness of periostracum (Table 6.2).

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
118
6.5.3. COMPARISON TO SPIKES OF OTHER BIVALVE GROUPS AND IMPLICATIONS FOR BIVALVE PHYLOGENY
Periostracal calcifications such as the unionoid spikes described in this study have
been reported from several other bivalve groups, most notably the Veneroida (Glover &
Taylor, 2010), Anomalodesmata (Checa & Harper, in press) and Myoida (Carter, 1978).
According to Glover & Taylor (2010) and Checa & Harper (in press), present evidence
suggests that calcification of the periostracum has occurred independently in several
bivalve clades. Checa & Harper (in press) hypothesise, however, that spikes of the closely
related Anomalodesmata and Palaeoheterodonta might be homologous structures. The
latter theory is supported by our observation that, similarly to those in anomalodesmatans,
unionoid spikes are probably formed immediately below the outer periostracum. On the
other hand, morphology of anomalodesmatan spikes differs somewhat from their
palaeoheterodont relatives. In particular, anomalodesmatan spikes are generally much
larger (i.e. about 20 µm in diameter; Checa & Harper, in press) and often arranged in
comarginal rows (e.g. in Laternula flexuosa Reeve, 1863 (Aller, 1974)). Furthermore, we
could not identify spikes in the Trigonioida, generally considered as a sister clade of the
Unionoida (Newell & Boyd, 1975; Healy, 1989; Hoeh et al., 1998; Watters, 2001). If
anomalodesmatan and unionoid spikes were homologous structures, this could therefore
mean a loss of character development in the trigonioids. Alternatively, spikes could be a
synapomorphic character of the Unionoida.
The matter is further complicated by the unresolved formation process of the
protruding trigonioid bosses. Though we observed bosses directly at the surface of
N. lamarcki shells, it is not clear that these protrusions were not initially covered by a
pellicle-like outer periostracum. Trigonioid bosses may in fact develop in those parts of the
prisms embedded within the periostracum, which could indicate a common ability of
intraperiostracal calcification, synapomorphic to Palaeoheterodonta, Anomalodesmata and
possibly other heterodont bivalve taxa. An improved understanding of the production
mode of spikes and protruding bosses in the Palaeoheterodonta will resolve these open
questions.

CONCLUSIONS


121
CHAPTER 7
CONCLUSIONS
In this thesis I aimed to better understand the factors which determine the
morphology of freshwater mussels (Unionoida). The morphological makeup (phenotype)
of a given freshwater mussel’s shell can be influenced by a range of factors and
mechanisms. These include (1) the specimen’s genetic composition, (2) its life habit and
surrounding environmental conditions, and (3) other characteristics of the individual such
as its sex, age, size and level of parasitic infestation. The results of this thesis show that all
these factors may play a part, but that their relative importance varies considerably
between different shell features, different populations and different species. By
understanding the factors that determine shell morphology, we have an opportunity to use
inter- and intraspecific patterns for the reconstruction of environments and habitat
requirements of species, determination of sex ratios or parasitic loads of individuals and
populations, and gain new insights into evolutionary trends and phylogenies.
Due to the great intraspecific variability in shell ‘size’, ‘shape’, ‘inflation’ and
‘thickness’, these characters were investigated with respect to patterns within single
unionoid species (Chapter 2-4). Chapter 5 and 6 took a broader look at unionoid shell
morphology to consider the taxonomic value and functional importance of macroscopic
(umbonal sculpture) and microscopic (periostracal microprojections) shell features.
Historically, considerable attention has been paid to unionoid shell morphology and
its functional and taxonomic significance. However, the absence of clear patterns and
numerous conflicting interpretations mean that the last major contribution to this field was
made over 30 years ago (Eagar, 1978). The opportunity to gain new insights into the
factors that determine unionoid shell morphology has been made possible by the
availability of modern statistical and analytical techniques. These tools have enabled me to
reconcile the apparent contradictions in the literature and identify new features of
taxonomic importance.
Fourier shape analysis provided a new and more powerful tool for the investigation
of shell shape in freshwater mussels, yielding consistent patterns across taxa which could
not be detected using traditional morphometric methods. Amplified fragment length
polymorphism (AFLP) analysis provided an opportunity to investigate intraspecific
variation in genotype within a small geographic scale and revealed that phenotypic

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
122
plasticity was of overriding importance in this instance. Scanning electron microscopy,
combined with access to extensive and well catalogued shell collections enabled me to
conduct studies of macro- and microscopic shell sculptures and identify new features of
considerable taxonomic and phylogenetic value.
7.1. WHAT CAN WE LEARN FROM A UNIONOID SHELL?
Based on results of this thesis and previous publications, Table 7.1 summarises the
potential utility of 12 unionoid shell characters for taxonomic classification and
phylogenetic reconstructions, environmental reconstructions, and the determination of sex
and parasite loads. The relative importance of these features for different applications is
considered below.
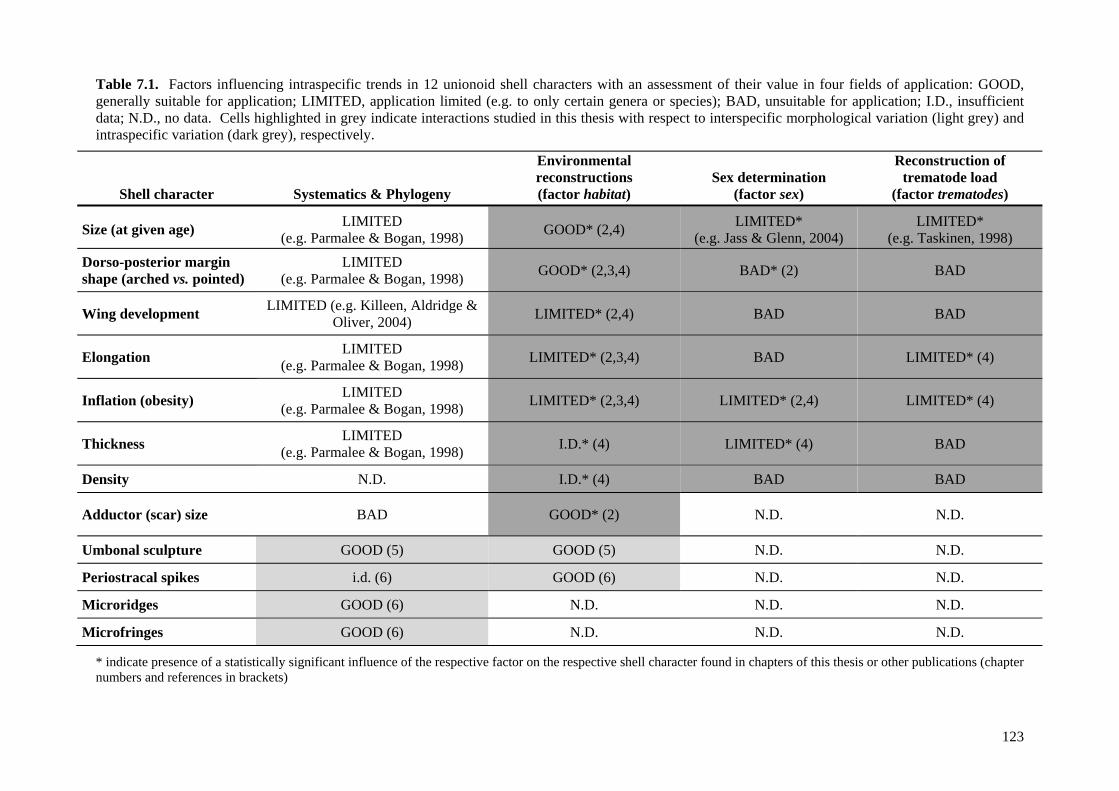
123
Table 7.1. Factors influencing intraspecific trends in 12 unionoid shell characters with an assessment of their value in four fields of application: GOOD, generally suitable for application; LIMITED, application limited (e.g. to only certain genera or species); BAD, unsuitable for application; I.D., insufficient data; N.D., no data. Cells highlighted in grey indicate interactions studied in this thesis with respect to interspecific morphological variation (light grey) and intraspecific variation (dark grey), respectively.
Shell character
Systematics & Phylogeny
Environmental reconstructions (factor habitat)
Sex determination
(factor sex)
Reconstruction of trematode load
(factor trematodes)
Size (at given age) LIMITED (e.g. Parmalee & Bogan, 1998) GOOD* (2,4) LIMITED*
(e.g. Jass & Glenn, 2004) LIMITED*
(e.g. Taskinen, 1998)
Dorso-posterior margin shape (arched vs. pointed)
LIMITED (e.g. Parmalee & Bogan, 1998) GOOD* (2,3,4) BAD* (2) BAD
Wing development LIMITED (e.g. Killeen, Aldridge & Oliver, 2004) LIMITED* (2,4) BAD BAD
Elongation LIMITED (e.g. Parmalee & Bogan, 1998) LIMITED* (2,3,4) BAD LIMITED* (4)
Inflation (obesity) LIMITED (e.g. Parmalee & Bogan, 1998) LIMITED* (2,3,4) LIMITED* (2,4) LIMITED* (4)
Thickness LIMITED (e.g. Parmalee & Bogan, 1998) I.D.* (4) LIMITED* (4) BAD
Density N.D. I.D.* (4) BAD BAD
Adductor (scar) size BAD GOOD* (2) N.D. N.D.
Umbonal sculpture GOOD (5) GOOD (5) N.D. N.D.
Periostracal spikes i.d. (6) GOOD (6) N.D. N.D.
Microridges GOOD (6) N.D. N.D. N.D.
Microfringes GOOD (6) N.D. N.D. N.D.
* indicate presence of a statistically significant influence of the respective factor on the respective shell character found in chapters of this thesis or other publications (chapter numbers and references in brackets)

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
124
7.1.1. SYSTEMATICS, PHYLOGENY AND EVOLUTION
Despite recent methodological and analytical advances, including the use of
molecular markers, unionoid phylogeny is still poorly understood. Under particularly
intense debate are the phylogenetic positions of the Hyriidae within the Unionoida (e.g.
Graf & Cummings (2006b) vs. Hoeh et al. (2009)), several tropical genera within the
Unionidae (e.g. Graf & Cummings, 2006b) and the Quadrulini within the unionid
subfamily Ambleminae (e.g. Campbell et al. (2005) vs. Hoeh, Bogan & Heard (2001)).
These unresolved questions are, to a great extent, the result of our lack of understanding of
which shell features are suitable for the use in reconstructions of bivalve phylogenies. That
is, a shell character is of high phylogenetic value if it does not exhibit a high degree of
intraspecific variation and if interspecific patterns in this shell feature reflect bivalve
evolution rather than convergences of unrelated taxa. What is more, with respect to those
characters that currently are used in phylogenetic analyses, we usually lack a proper
understanding of character evolution and thus, cannot discriminate between
phylogenetically informative (homoplasic) and uninformative (convergent) character
states.
7.1.1.1. “Gross shell morphology”: size and form
To a limited degree, gross shell morphology can be useful for taxonomic
classification and phylogenetic reconstructions. Shell features that are to some extent
informative and reliable at the species and higher taxonomic levels include ‘shell size’
(e.g. Anodonta cygnea (Linnaeus, 1758) attains much larger maximum shell sizes than
Anodonta anatina (Linnaeus, 1758)) and ‘presence/absence of wings’ (e.g. wings are
present in Anodonta spp. but not in Unio spp.) (e.g. Parmalee & Bogan, 1998; Killeen,
Aldridge & Oliver, 2004). However, due to the high degree of interspecific convergences
and intraspecific variability in shell ‘size’, ‘form’ and ‘thickness’, the use of these shell
characters in higher taxonomic classifications or phylogenetic reconstructions is generally
regarded as problematic (e.g. Ortmann, 1912; Prashad, 1931; Haas, 1969a, b; Davis, 1983,
1984).
The fact that shell ‘shape’ of a unionoid is a poor indicator of its genetic composition
was demonstrated in Chapter 3. Thus, genetic comparisons of three paired Unio pictorum
(Linnaeus, 1758) populations from two habitats failed to identify consistent genetic
differences between two shell ecomorphotypes. This suggests that pronounced

Chapter 7 - Conclusions
125
morphological differences in the degree of shell elongation and the shape of the dorso-
posterior margin in this unionid species are caused by plasticity of the phenotype and
provides the first good evidence for phenotypic plasticity of shell shape in a European
freshwater mussel. Genetic differences along the River Thames, on the other hand, were
consistent with a pattern of isolation by geographic distance and probably reflect limited
dispersal via host fish species upon which unionoid larvae are obligate parasites.
7.1.1.2. Umbonal sculpture
In contrast to the extreme phenotypic plasticity in, for example, shell ‘size’ and
‘shape’, the morphological appearance of ‘umbonal sculptures’ within a species is much
more conservative. As a result, ‘umbonal sculpture’ is one of the few shell features used in
modern phylogenetic analyses of the Unionoida (e.g. Hoeh, Bogan & Heard, 2001; Graf &
Cummings, 2006b). What is more, these sculptures are one of the few features available
for taxonomic classification of both modern and fossil taxa, which in turn is the basis for
improving our surprisingly poor knowledge on early unionoid evolution. Unfortunately,
these fields of research suffer from our lack of understanding of homoplasies and
homologies of umbonal sculpture types across the unionoid phylogeny. Thus, previously
suggested models of character evolution of beak sculptures (Simpson, 1900; Ortmann,
1912; Modell, 1942) are contradictory and based on outdated assumptions on unionoid
evolution and sculpture formation.
Based on examination of over 150 modern and extinct species, Chapter 5 presented a
new model of character evolution of umbonal sculptures in the Unionoida. Challenging all
previous models of character evolution, my observations indicated that the plesiomorphic
characters state of unionoid beak sculpture is ‘V-shaped’. ‘Pseudo-radial’ sculpture
probably arose in one line of development, whereas ‘W-shaped’ and ultimately ‘double-
looped’ sculpture was developed in other unionoid groups. All other forms of umbonal
sculpture present in this bivalve group represent derivates of one of these four main types.
The study confirmed the suitability of umbonal sculptures for reconstructing
phylogenies and classifying unknown specimens to higher unionoid taxonomic levels.
Furthermore, examination of the distribution of beak sculpture types across the unionoid
phylogeny revealed new insights into unresolved questions on freshwater mussel
evolution. For example, presence of ‘V-shaped’ and ‘pseudo-radial’ sculpture in the
Hyriidae and some incertae sedis unionid genera (e.g. Coelatura and Parreysia) suggested
a basal position of these taxa within the Unionoida. A basal position of the Quadrulini

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
126
within the unionid subfamily Ambleminae was supported by the ‘W-shaped’ umbonal
sculpture of many quadruline species examined. On the other hand, I showed that some
beak sculpture types, such as ‘nodulous’, ‘wrinkled’ and ‘smooth’, have evolved
independently numerous times within the Unionoida and thus, represent phylogenetically
uninformative, convergent character states.
7.1.1.3. Periostracal microprojections
In Chapter 6, the phylogenetic significance of a further, previously unrecognised
shell character was investigated for the first time in the Unionoida and their presumed
closest living relatives, the Trigonioida. Such periostracal microprojections had been
described in other bivalve groups (e.g. Aller, 1974; Glover & Taylor, 2010; Checa &
Harper, in press).
Investigation of shell surfaces of specimens covering all extant palaeoheterodont
(unionoid + trigonioid) families using scanning electron microscopy, revealed three types
of periostracal microprojections; microridges, microfringes and spicule-like spikes.
Examination of the distribution of these three distinct morphologies across the
palaeoheterodont phylogeny indicated a considerable value for the reconstruction of
bivalve evolution. Thus, microridges were present only in one respective species of the
two unionoid families Mycetopodidae (Anodontites trapesialis (Lamarck, 1819)) and
Iridinidae (Chambardia bourguignati (Bourguignat, 1885)), and may represent a
synapomorphy for the mycetopodid-iridinid clade. In A. trapesialis, microridges were
additionally equipped with flag-like microfringes, which are possibly a synapomorphic
character for the Mycetopodidae.
Calcified spikes were widespread across the unionoid phylogeny but were not found
in any hyriid and mycetopodid species examined. Given the relatively well established,
derived position of the Mycetopodidae within the Unionoida, it is likely that spike
production has secondarily been lost or switched off in this family. Absence of spikes in
the Hyriidae, on the other hand, could represent the plesiomorphic unionoid character
state. As in the case of beak sculptures (7.1.1.2), this would suggest a basal position of the
Hyriidae within the Unionoida.
7.1.2. ECOLOGY, CONSERVATION AND ENVIRONMENTAL RECONSTRUCTION
Most (palaeo)environmental reconstructions using unionoid shells have so far been
based on relative abundance data of species with known habitat requirements (i.e. species

Chapter 7 - Conclusions
127
assemblages) (e.g. Matteson, 1960; Morey & Crothers, 1998; Radley & Barker, 1998).
Naturally, these exclude most endangered and extinct taxa, restricting utility of such
assemblage analyses to the reconstruction of only relatively recent palaeoenvironments.
Knowledge on inter- and intraspecific trends in shell morphologies that are reliable
indicators of certain environmental conditions and exhibited across a wide taxonomic
range can overcome this problem. At the species and higher taxonomic levels, this can be
attained by identification of unifying characteristics in habitat requirements of unrelated
taxa exhibiting convergent morphologies. Currently, our knowledge in this respect is
restricted to only a small number of unionoid shell morphologies, in particular wing
development (i.e. symphynote species usually indicate soft sediment habitats) and adult
shell sculpture (i.e. well sculptured species indicate fast-flowing habitats) (Watters, 1994).
Information about the environment can also be obtained using intraspecific trends in
morphology that are consistently associated with changes in habitat condition. Probably
the most widely used unionoid shell character in this respect has so far been maximum or
average ‘shell size’ of populations, which is generally regarded to be positively correlated
with temperature and/or food availability (e.g. Grier, 1920; Mann, 1965). Additionally, the
extreme variability in shell form of many unionoid species could be particularly useful for
redrawing characteristics of the flow or sediment conditions of the habitat. Unfortunately,
reconstruction of environments and habitat requirements of species on the basis of
intraspecific patterns in unionoid shell shape has so far been limited by the lack of
identification of consistent ecophenotypic trends that are not considerably confused by
non-habitat factors such as sexual dimorphism or ontogenetic growth (Tevesz & Carter,
1980; Claassen, 1998).
7.1.2.1. “Gross shell morphology”: size, shape, inflation, thickness, density and adductor scar sizes
Partially confirming observations by Grier (1920) and Mann (1965), investigation of
morphological patterns within three unionid species from two habitat types (marinas and
river) of the River Thames, UK, in Chapter 2 showed that larger maximum shell sizes (and
probably also relatively large sizes of adductor muscle scars) can be indicative of warmer
temperature and/or greater phytoplankton abundances. What is more, the study elucidated
an intraspecific ecophenotypic trend in shell form that was consistently exhibited across
replicate populations and species. This trend from dorsally arched river specimens to
straight dorsal and pointed posterior margins in marina individuals was always associated

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
128
with differences in the hydrological condition of the habitat and could have an adaptive
value of stabilising the mussel’s life position in fast flowing habitats. Similar observations
by Eagar (1948, 1971, 1974, 1977, 1978) on margaritiferid unionoids and Carboniferous
anthracosiid species suggest that this ecophenotypic trend is present across a wide range of
freshwater mussel taxa. Intraspecific differences in the shape of the dorso-posterior margin
of unionoid shells could thus be used, for example, in the determination of habitat
requirements of endangered species and reconstruction of flow regimes of ancient
freshwater habitats.
Relative ‘elongation’ and ‘wing development’ were two further shell characters that
showed significant habitat-associated trends. However, these patterns were restricted only
to single species, limiting their potential utility for environmental reconstructions to only
these species (or possibly genera) where a particular trend has been elucidated.
Shell ‘inflation’ should generally be regarded as a poor indicator of habitat since it
was shown to be considerably influenced by allometric growth, sexual dimorphism and
trematode parasites (Chapter 2 and 4). These observations not only help explain
contradictory observations in ecophenotypic trends in this character by previous authors
(e.g. Ortmann, 1920) but also demonstrate the importance of considering non-habitat
factors when attempting to elucidate truly ecophenotypic trends.
Statistically significant differences between populations in two further shell
characters, i.e. ‘thickness’ and ‘density’ of the shell, was demonstrated in Chapter 4
(4.4.4). However, which habitat conditions are associated with these changes in shell
morphology and the potential value of these characters for environmental reconstructions
remains unclear.
7.1.2.2. Umbonal sculpture
The new model of character evolution of unionoid rugae developed in Chapter 5
elucidated interspecific convergences in sculpture types. For example, though species with
relatively faint or completely ‘smooth’ umbos were found in several unrelated taxa, almost
all of them were characteristic of lentic habitats. Species with well developed rugae, and
particularly of the ‘wrinkled’ or ‘nodulous’ type, on the other hand, were typically
inhabitants of turbulent habitats. These three types of umbonal sculptures could thus be
used to for assigning a species to a typically standing or flowing habitat.
Provided a good understanding of their function(s) is available, beak sculptures
could ultimately be used for determination of habitat requirements of juveniles of different

Chapter 7 - Conclusions
129
unionoid species. Such knowledge could be of particular value for developing optimal
habitat recreation measures or conditions in artificial propagation of endangered
freshwater mussels, thereby increasing the usually extremely low rate of survival in the
first months of the postparasitic life stage (Wächtler, Mansur & Richter, 2001). However,
though my observations indicated that umbonal rugae probably serve in increasing
reinforcement and/or anchoring of the juvenile shell in the sediment, these functional
hypotheses remain to be tested in experiments.
7.1.2.3. Periostracal microprojections
Particularly large and abundant periostracal spikes were observed in species
characteristic of fast flowing, coarse sediment habitats and possibly serve to protect the
periostracum and underlying shell, and/or to stabilise life position by increasing shell
friction (Chapter 6). Average spike size and abundance of a given unionoid shell of
unknown origin may thus be used to redraw information about flow and/or sediment
conditions of the habitat it once inhabited.
Microfringes and microridges possibly aid in the orientation of the mussel within the
sediment, but information on the patterns of these microsculptures across different habitats
currently remains too sparse to enable any ecophenotypic generalisations to be made.
7.1.3. DETERMINATION OF SEX AND TREMATODE LOADS
7.1.3.1. “Gross shell morphology”: size, shape, inflation and thickness
Intraspecific patterns in some shell features were considerably affected by other
factors than the habitat, rendering such characters unsuitable for application in
environmental reconstructions. Such morphological trends may, on the other hand, help in
the reconstruction of other characteristics of an individual, population or species. This is
particularly true for changes in unionoid shell morphology that are induced by sexual
dimorphism and parasitic infestation. Apart from the well known differences in sagittal
shell shape between male and female lampsilines (e.g. Kotrla & James, 1987), both these
types of intraspecific shell dimorphism are currently poorly understood in the Unionoida.
Chapter 4, which investigated five A. anatina populations, represents the first study
revealing statistically significant sexual and parasite-induced dimorphic trends in the shells
of a European unionid species.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
130
More specifically, differences between sexes of A. anatina were observed in shell
‘inflation’ and ‘thickness’, and probably are a result of altered female shell growth due to
accommodation of marsupial gills and resource depletion by offspring production,
respectively. My results further indicated that the extent of morphological differences
between sexes was determined by the environmental conditions to which the respective
population is subjected. This observation helps explain the so far inconclusive reports on
sexual shell dimorphism in the unionid subfamily Unioninae and answers a long debated
question; why some populations are apparently dimorphic and others are not. In general,
sexual dimorphic trends were more pronounced in populations from “high quality” habitats
compared to those living at less favourable conditions. Thus, only in populations of high
reproductive output could differences in shell width and/or thickness between sexes be a
useful tool, providing rapid sex determination in the field or enabling determination of a
fossil population’s sex ratio.
The same study also showed for the first time that trematode parasites can
significantly alter both inflation and sagittal shape of freshwater mussel shells. The
underlying reasons and mechanisms involved in these parasite-induced changes in shell
morphology are currently unclear. Nevertheless, provided a better understanding of
trematode-mussel interactions, this or similar dimorphic patterns could potentially become
a useful tool for detecting parasite infections, that can lead to castration or even death of
unionoids (e.g. Jokela, Uotila & Taskinen, 1993; Gangloff, Lenertz & Feminella, 2008;
Grizzle & Brunner, 2009). As such, monitoring for changes in morphology within
threatened populations of unionoids may provide important indications of changes in
harmful parasite burdens.
7.2. FUTURE DIRECTIONS
This thesis provides several new insights into the suitability of 12 unionoid shell
characters for phylogenetic and environmental reconstruction, and determination of sex
and trematode loads. In doing so, it also raises many new questions which are worthy of
further investigation.

Chapter 7 - Conclusions
131
7.2.1. TESTING FOR CONSISTENCY OF THE PATTERNS OBSERVED
Investigations on intraspecific ecophenotypic trends and the degree of phenotypic
plasticity in shell morphology were based on populations of just one river catchment area,
and involved only three and one common unionoid species, respectively. In order to
establish the generality of what was found in these studies, similar experiments on other
unionoid species involving populations of various river catchments and lake habitats are
needed. The distribution of genetic diversity of a given mussel species/population would,
for example, be expected to be strongly dependent on the mobility of the fish species it is
parasitising. This could be tested by looking at several unionoid species using different
host fish species.
The dimorphic patterns in shell morphology observed and discussed in Chapter 4
were based merely on a small number of populations of only a single unionoid species.
Further studies involving populations from a wide range of habitats and additional species
are recommended in order to test if the hypotheses developed here hold true. If and to what
extent the degree of sexual dimorphisms is influenced by the “quality” of the habitat and
thus, mussel fecundity at the site, could be further investigated on the basis of
measurements of glochidia production per female.
Although a large number of species were included in the analysis of umbonal
sculptures in Chapter 5, some unionoid families were underrepresented or, in the case of
the Etheriidae, completely omitted due to a lack of available material. Consideration of
additional material could help test the robustness of my model and could help to illuminate
the taxonomy of fossil taxa.
The main purpose of Chapter 6 was to provide an initial overview on periostracal
microprojections present in the palaeoheterodont families. Further studies are needed to
confirm or reject the synapomorphies and other hypotheses proposed in this study.
7.2.2. OPEN QUESTIONS
One of the main objectives of this thesis has been the identification of consistent
ecophenotypic trends in unionoid shell features. The next big challenge in this respect will
be to test the adaptive significance of the patterns observed and the functional hypotheses
developed in this thesis. This will require an experimental approach. For example, the
adaptive significance of the intraspecific ecomorphotypes of marina and river habitats of
the River Thames could be investigated by means of reciprocal transplant experiments.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
132
Flume experiments could be used to test the hypothesis that arched “river” shells are able
to withstand higher current velocities than “marina” morphotypes. Equally, laboratory
experiments on juvenile unionoids could investigate the functional significance of
unionoid umbonal sculptures.
To what extent sex and parasites influence unionoid shell morphology remains
poorly understood. This is particularly so for most Afrotropical and Indotropical unionid
species, for which we often lack even the most basic biological data, such as the time of
brooding or which parts of the female gills are used as marsupia. With respect to European
unionoids, it would be interesting to test if and to what extent other freshwater mollusc
parasites, such as parasitic fish (e.g. Bitterling (Rhodeus spp.)) or mites (Unionicolidae),
alter unionoid shell growth.
As discussed in Chapter 6 (6.5.3), investigation of the formation process of
trigonioid bosses and unionoid spikes is needed to resolve the question if these structures
represent a common ability to periostracal calcification in the two palaeoheterodont orders.
Study of intraspecific variation in both unionoid microprojections and umbonal sculpture
including specimens from a range of habitats will possibly elucidate ecophenotypic trends
useful for reconstruction of habitat requirements and environments of juvenile mussels.
7.2.3. APPLICATION OF METHODS TO NON-UNIONOID TAXA
While this thesis has focused on the use of new tools to better understand
morphology in unionoid mussels, the approach could also provide new and important
insights into other taxa. Many of the patterns observed in my studies may also hold true for
other freshwater and marine bivalves, while a closer inspection of aquatic gastropods is
likely to reveal consistent intra- and interspecific patterns in shell shape which can be
attributed to genotype, habitat type, sex and parasite load. Indeed, the marine limpets
(Patella spp.) and periwinkles (Littorina spp.) provide classic examples of consistent
ecophenotypes that have been linked with trade-offs between factors such as reproductive
capacity, risk of displacement by wave action, and vulnerability to predation (e.g. Vermeij,
1973; Johannesson, 2003). Revisiting these examples using more powerful descriptive and
analytical tools may generate an enhanced understanding in these already intensively
studied systems. Moreover, as tools continue to become increasingly powerful (e.g. 3D-
imaging), we can expect to continue to gain an increasingly greater understanding of the
factors that determine morphology of a particular organism.

APPENDIX

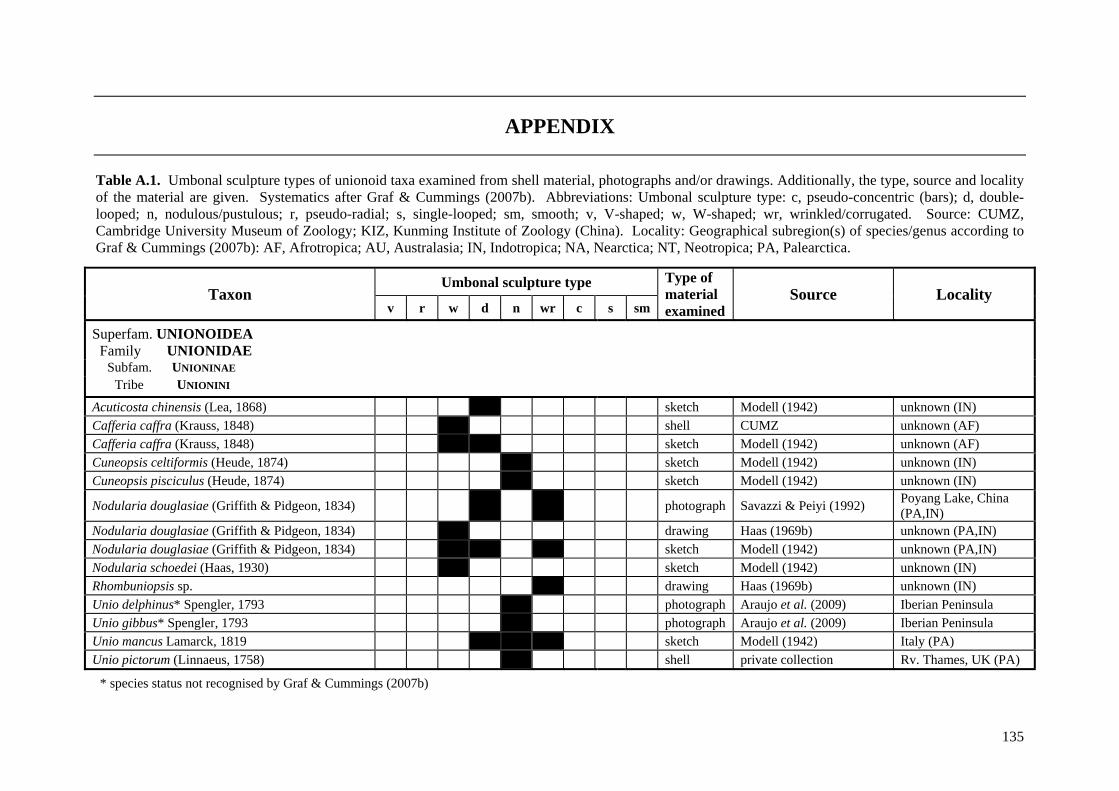
135
APPENDIX
Table A.1. Umbonal sculpture types of unionoid taxa examined from shell material, photographs and/or drawings. Additionally, the type, source and locality of the material are given. Systematics after Graf & Cummings (2007b). Abbreviations: Umbonal sculpture type: c, pseudo-concentric (bars); d, double-looped; n, nodulous/pustulous; r, pseudo-radial; s, single-looped; sm, smooth; v, V-shaped; w, W-shaped; wr, wrinkled/corrugated. Source: CUMZ, Cambridge University Museum of Zoology; KIZ, Kunming Institute of Zoology (China). Locality: Geographical subregion(s) of species/genus according to Graf & Cummings (2007b): AF, Afrotropica; AU, Australasia; IN, Indotropica; NA, Nearctica; NT, Neotropica; PA, Palearctica.
Taxon Umbonal sculpture type Type of
material examined
Source Locality v r w d n wr c s sm
Superfam. UNIONOIDEA Family UNIONIDAE
Subfam. UNIONINAE Tribe UNIONINI
Acuticosta chinensis (Lea, 1868) x sketch Modell (1942) unknown (IN) Cafferia caffra (Krauss, 1848) x shell CUMZ unknown (AF) Cafferia caffra (Krauss, 1848) x x sketch Modell (1942) unknown (AF) Cuneopsis celtiformis (Heude, 1874) x sketch Modell (1942) unknown (IN) Cuneopsis pisciculus (Heude, 1874) sketch Modell (1942) unknown (IN)
Nodularia douglasiae (Griffith & Pidgeon, 1834) x x photograph Savazzi & Peiyi (1992) Poyang Lake, China (PA,IN)
Nodularia douglasiae (Griffith & Pidgeon, 1834) x drawing Haas (1969b) unknown (PA,IN) Nodularia douglasiae (Griffith & Pidgeon, 1834) x x x sketch Modell (1942) unknown (PA,IN) Nodularia schoedei (Haas, 1930) x sketch Modell (1942) unknown (IN) Rhombuniopsis sp. x drawing Haas (1969b) unknown (IN) Unio delphinus* Spengler, 1793 x photograph Araujo et al. (2009) Iberian Peninsula Unio gibbus* Spengler, 1793 x photograph Araujo et al. (2009) Iberian Peninsula Unio mancus Lamarck, 1819 x x x sketch Modell (1942) Italy (PA) Unio pictorum (Linnaeus, 1758) x shell private collection Rv. Thames, UK (PA)
* species status not recognised by Graf & Cummings (2007b)
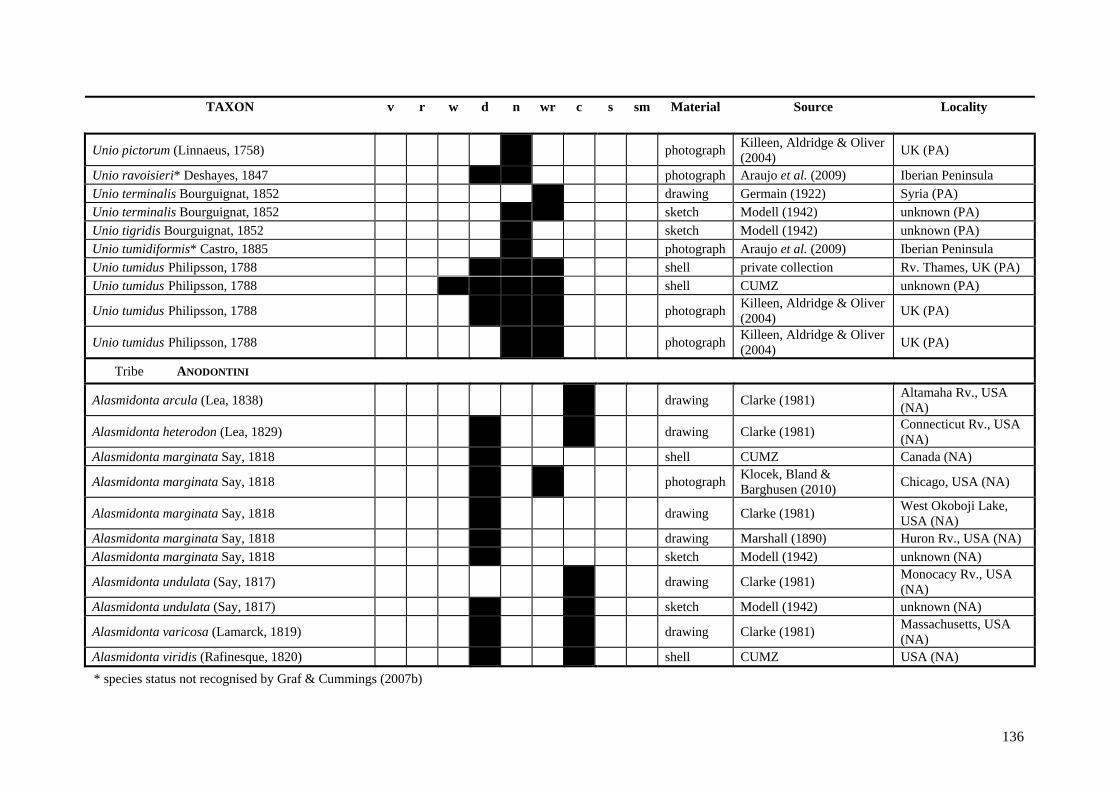
136
TAXON v r w d n wr c s sm Material Source Locality
Unio pictorum (Linnaeus, 1758) photograph Killeen, Aldridge & Oliver (2004) UK (PA)
Unio ravoisieri* Deshayes, 1847 x x photograph Araujo et al. (2009) Iberian Peninsula Unio terminalis Bourguignat, 1852 drawing Germain (1922) Syria (PA) Unio terminalis Bourguignat, 1852 x x sketch Modell (1942) unknown (PA) Unio tigridis Bourguignat, 1852 x sketch Modell (1942) unknown (PA) Unio tumidiformis* Castro, 1885 x photograph Araujo et al. (2009) Iberian Peninsula Unio tumidus Philipsson, 1788 x x x shell private collection Rv. Thames, UK (PA) Unio tumidus Philipsson, 1788 shell CUMZ unknown (PA)
Unio tumidus Philipsson, 1788 photograph Killeen, Aldridge & Oliver (2004) UK (PA)
Unio tumidus Philipsson, 1788 photograph Killeen, Aldridge & Oliver (2004) UK (PA)
Tribe ANODONTINI
Alasmidonta arcula (Lea, 1838) drawing Clarke (1981) Altamaha Rv., USA (NA)
Alasmidonta heterodon (Lea, 1829) drawing Clarke (1981) Connecticut Rv., USA (NA)
Alasmidonta marginata Say, 1818 x shell CUMZ Canada (NA)
Alasmidonta marginata Say, 1818 photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Alasmidonta marginata Say, 1818 drawing Clarke (1981) West Okoboji Lake, USA (NA)
Alasmidonta marginata Say, 1818 x drawing Marshall (1890) Huron Rv., USA (NA) Alasmidonta marginata Say, 1818 x sketch Modell (1942) unknown (NA)
Alasmidonta undulata (Say, 1817) drawing Clarke (1981) Monocacy Rv., USA (NA)
Alasmidonta undulata (Say, 1817) x sketch Modell (1942) unknown (NA)
Alasmidonta varicosa (Lamarck, 1819) drawing Clarke (1981) Massachusetts, USA (NA)
Alasmidonta viridis (Rafinesque, 1820) x x shell CUMZ USA (NA) * species status not recognised by Graf & Cummings (2007b)
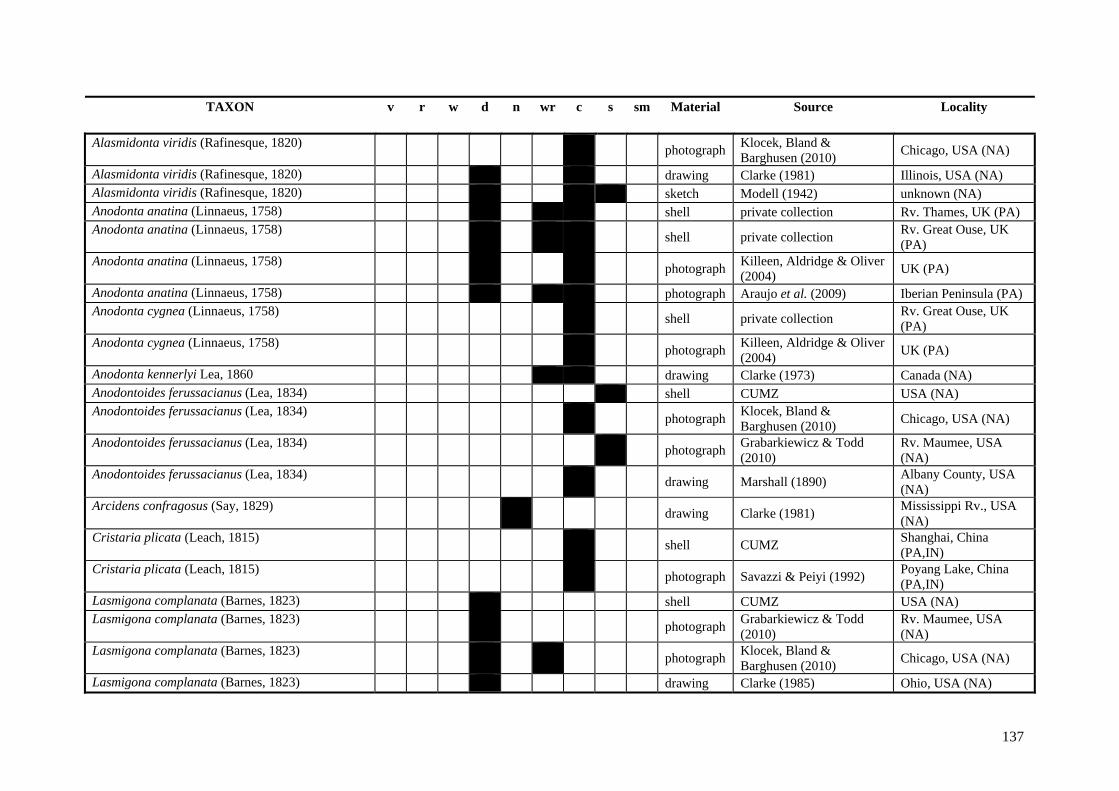
137
TAXON v r w d n wr c s sm Material Source Locality Alasmidonta viridis (Rafinesque, 1820) photograph Klocek, Bland &
Barghusen (2010) Chicago, USA (NA)
Alasmidonta viridis (Rafinesque, 1820) drawing Clarke (1981) Illinois, USA (NA) Alasmidonta viridis (Rafinesque, 1820) x x x sketch Modell (1942) unknown (NA) Anodonta anatina (Linnaeus, 1758) x x x shell private collection Rv. Thames, UK (PA) Anodonta anatina (Linnaeus, 1758) shell private collection Rv. Great Ouse, UK
(PA) Anodonta anatina (Linnaeus, 1758) photograph Killeen, Aldridge & Oliver
(2004) UK (PA)
Anodonta anatina (Linnaeus, 1758) photograph Araujo et al. (2009) Iberian Peninsula (PA) Anodonta cygnea (Linnaeus, 1758) x shell private collection Rv. Great Ouse, UK
(PA) Anodonta cygnea (Linnaeus, 1758) photograph Killeen, Aldridge & Oliver
(2004) UK (PA)
Anodonta kennerlyi Lea, 1860 x x drawing Clarke (1973) Canada (NA) Anodontoides ferussacianus (Lea, 1834) x shell CUMZ USA (NA) Anodontoides ferussacianus (Lea, 1834) photograph Klocek, Bland &
Barghusen (2010) Chicago, USA (NA)
Anodontoides ferussacianus (Lea, 1834) x photograph Grabarkiewicz & Todd (2010)
Rv. Maumee, USA (NA)
Anodontoides ferussacianus (Lea, 1834) x drawing Marshall (1890) Albany County, USA (NA)
Arcidens confragosus (Say, 1829) drawing Clarke (1981) Mississippi Rv., USA (NA)
Cristaria plicata (Leach, 1815) x shell CUMZ Shanghai, China (PA,IN)
Cristaria plicata (Leach, 1815) x photograph Savazzi & Peiyi (1992) Poyang Lake, China (PA,IN)
Lasmigona complanata (Barnes, 1823) x shell CUMZ USA (NA) Lasmigona complanata (Barnes, 1823) x photograph Grabarkiewicz & Todd
(2010) Rv. Maumee, USA (NA)
Lasmigona complanata (Barnes, 1823) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Lasmigona complanata (Barnes, 1823) x drawing Clarke (1985) Ohio, USA (NA)
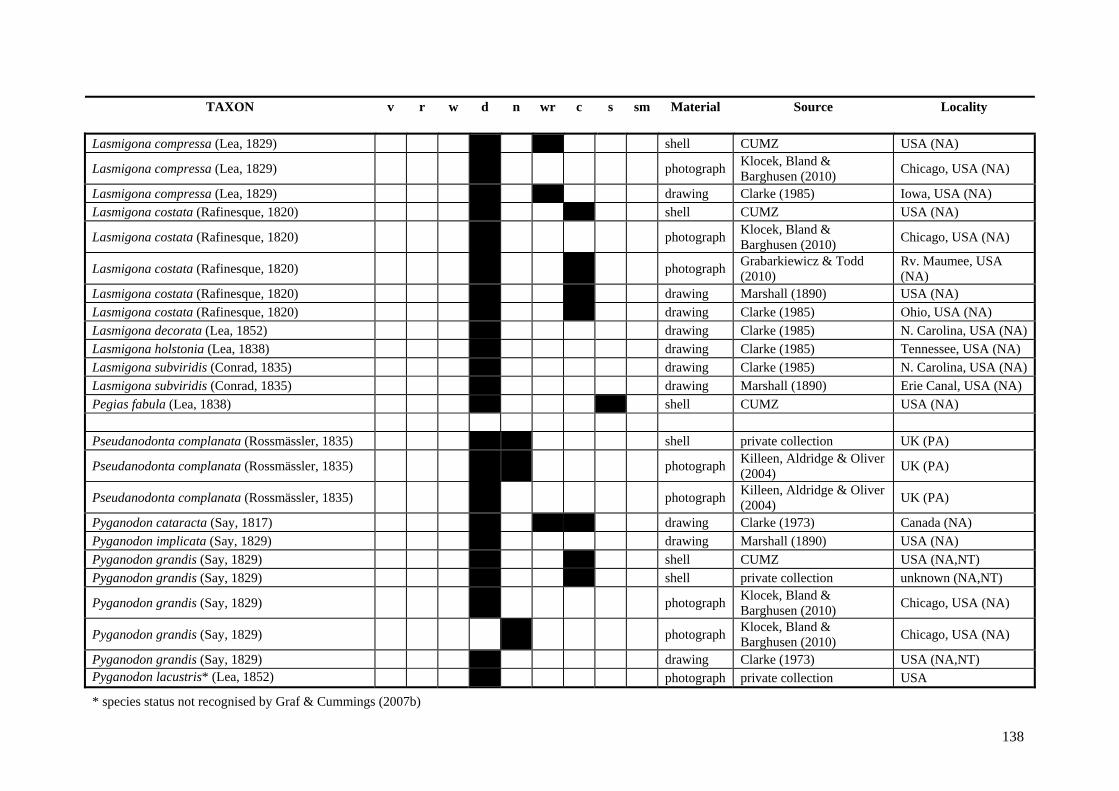
138
TAXON v r w d n wr c s sm Material Source Locality Lasmigona compressa (Lea, 1829) x x shell CUMZ USA (NA)
Lasmigona compressa (Lea, 1829) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Lasmigona compressa (Lea, 1829) x x drawing Clarke (1985) Iowa, USA (NA) Lasmigona costata (Rafinesque, 1820) x x shell CUMZ USA (NA)
Lasmigona costata (Rafinesque, 1820) x photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Lasmigona costata (Rafinesque, 1820) x photograph Grabarkiewicz & Todd (2010)
Rv. Maumee, USA (NA)
Lasmigona costata (Rafinesque, 1820) x x drawing Marshall (1890) USA (NA) Lasmigona costata (Rafinesque, 1820) x x drawing Clarke (1985) Ohio, USA (NA) Lasmigona decorata (Lea, 1852) x drawing Clarke (1985) N. Carolina, USA (NA) Lasmigona holstonia (Lea, 1838) x drawing Clarke (1985) Tennessee, USA (NA) Lasmigona subviridis (Conrad, 1835) x drawing Clarke (1985) N. Carolina, USA (NA) Lasmigona subviridis (Conrad, 1835) x drawing Marshall (1890) Erie Canal, USA (NA) Pegias fabula (Lea, 1838) x x shell CUMZ USA (NA) Pseudanodonta complanata (Rossmässler, 1835) x x shell private collection UK (PA)
Pseudanodonta complanata (Rossmässler, 1835) photograph Killeen, Aldridge & Oliver (2004) UK (PA)
Pseudanodonta complanata (Rossmässler, 1835) photograph Killeen, Aldridge & Oliver (2004) UK (PA)
Pyganodon cataracta (Say, 1817) x x drawing Clarke (1973) Canada (NA) Pyganodon implicata (Say, 1829) x drawing Marshall (1890) USA (NA) Pyganodon grandis (Say, 1829) x shell CUMZ USA (NA,NT) Pyganodon grandis (Say, 1829) x x shell private collection unknown (NA,NT)
Pyganodon grandis (Say, 1829) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Pyganodon grandis (Say, 1829) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Pyganodon grandis (Say, 1829) x drawing Clarke (1973) USA (NA,NT) Pyganodon lacustris* (Lea, 1852) x photograph private collection USA
* species status not recognised by Graf & Cummings (2007b)
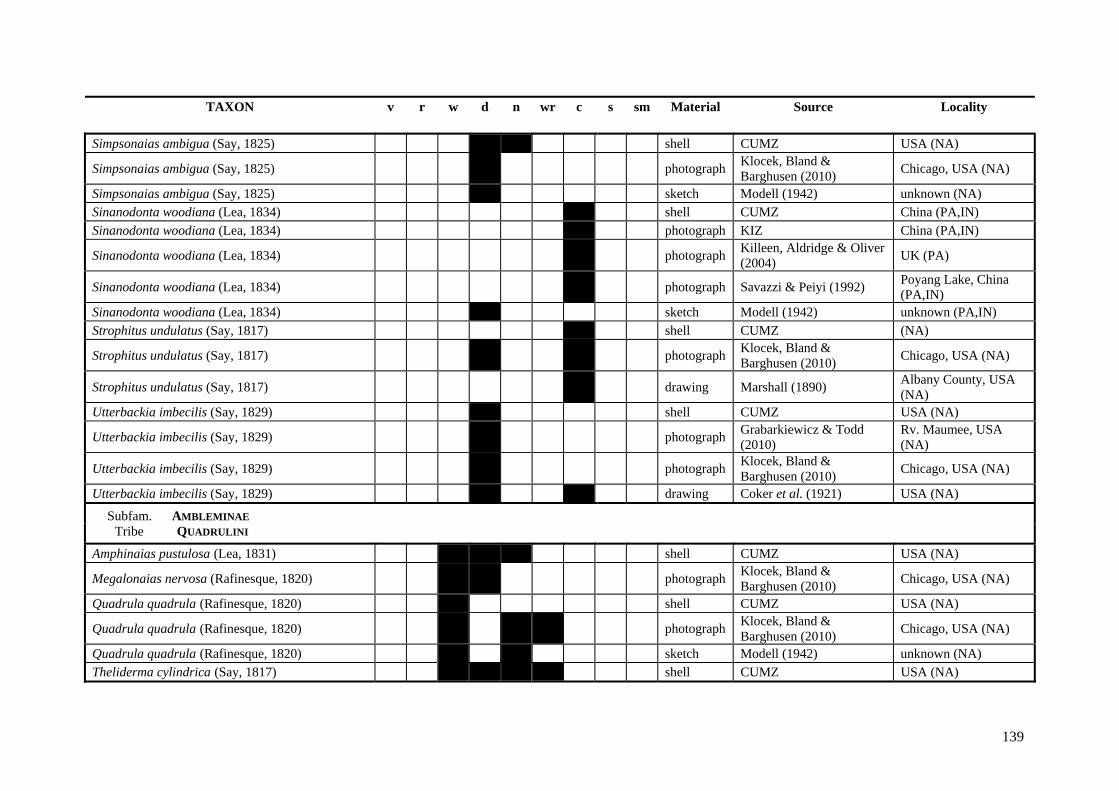
139
TAXON v r w d n wr c s sm Material Source Locality Simpsonaias ambigua (Say, 1825) x x shell CUMZ USA (NA)
Simpsonaias ambigua (Say, 1825) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Simpsonaias ambigua (Say, 1825) x sketch Modell (1942) unknown (NA) Sinanodonta woodiana (Lea, 1834) x shell CUMZ China (PA,IN) Sinanodonta woodiana (Lea, 1834) x photograph KIZ China (PA,IN)
Sinanodonta woodiana (Lea, 1834) photograph Killeen, Aldridge & Oliver (2004) UK (PA)
Sinanodonta woodiana (Lea, 1834) x photograph Savazzi & Peiyi (1992) Poyang Lake, China (PA,IN)
Sinanodonta woodiana (Lea, 1834) x sketch Modell (1942) unknown (PA,IN) Strophitus undulatus (Say, 1817) x shell CUMZ (NA)
Strophitus undulatus (Say, 1817) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Strophitus undulatus (Say, 1817) x drawing Marshall (1890) Albany County, USA (NA)
Utterbackia imbecilis (Say, 1829) x shell CUMZ USA (NA)
Utterbackia imbecilis (Say, 1829) x photograph Grabarkiewicz & Todd (2010)
Rv. Maumee, USA (NA)
Utterbackia imbecilis (Say, 1829) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Utterbackia imbecilis (Say, 1829) x x drawing Coker et al. (1921) USA (NA)
Subfam. AMBLEMINAE Tribe QUADRULINI
Amphinaias pustulosa (Lea, 1831) x x shell CUMZ USA (NA)
Megalonaias nervosa (Rafinesque, 1820) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Quadrula quadrula (Rafinesque, 1820) x shell CUMZ USA (NA)
Quadrula quadrula (Rafinesque, 1820) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Quadrula quadrula (Rafinesque, 1820) x x sketch Modell (1942) unknown (NA) Theliderma cylindrica (Say, 1817) x x x x shell CUMZ USA (NA)
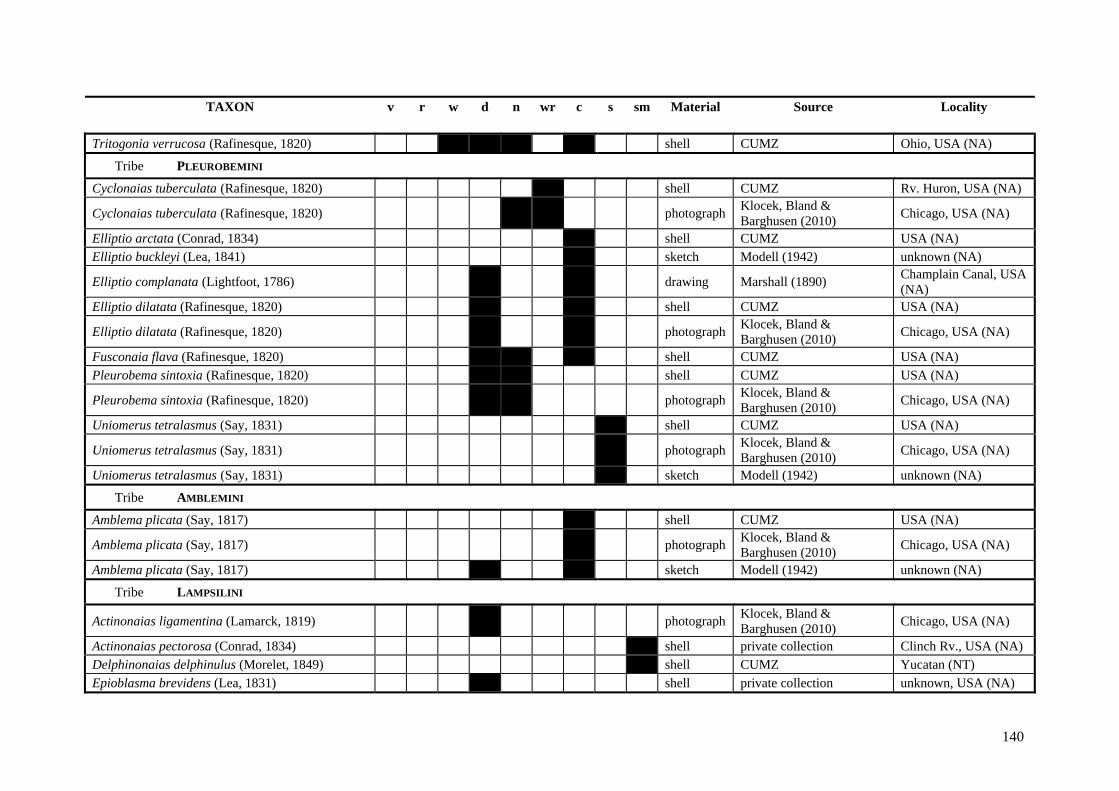
140
TAXON v r w d n wr c s sm Material Source Locality Tritogonia verrucosa (Rafinesque, 1820) x x x x shell CUMZ Ohio, USA (NA)
Tribe PLEUROBEMINI
Cyclonaias tuberculata (Rafinesque, 1820) x shell CUMZ Rv. Huron, USA (NA)
Cyclonaias tuberculata (Rafinesque, 1820) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Elliptio arctata (Conrad, 1834) x shell CUMZ USA (NA) Elliptio buckleyi (Lea, 1841) x sketch Modell (1942) unknown (NA)
Elliptio complanata (Lightfoot, 1786) x x drawing Marshall (1890) Champlain Canal, USA (NA)
Elliptio dilatata (Rafinesque, 1820) x x shell CUMZ USA (NA)
Elliptio dilatata (Rafinesque, 1820) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Fusconaia flava (Rafinesque, 1820) x x x shell CUMZ USA (NA) Pleurobema sintoxia (Rafinesque, 1820) x x shell CUMZ USA (NA)
Pleurobema sintoxia (Rafinesque, 1820) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Uniomerus tetralasmus (Say, 1831) x shell CUMZ USA (NA)
Uniomerus tetralasmus (Say, 1831) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Uniomerus tetralasmus (Say, 1831) x sketch Modell (1942) unknown (NA)
Tribe AMBLEMINI
Amblema plicata (Say, 1817) x shell CUMZ USA (NA)
Amblema plicata (Say, 1817) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Amblema plicata (Say, 1817) x x sketch Modell (1942) unknown (NA)
Tribe LAMPSILINI
Actinonaias ligamentina (Lamarck, 1819) x photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Actinonaias pectorosa (Conrad, 1834) x shell private collection Clinch Rv., USA (NA) Delphinonaias delphinulus (Morelet, 1849) x shell CUMZ Yucatan (NT) Epioblasma brevidens (Lea, 1831) x shell private collection unknown, USA (NA)
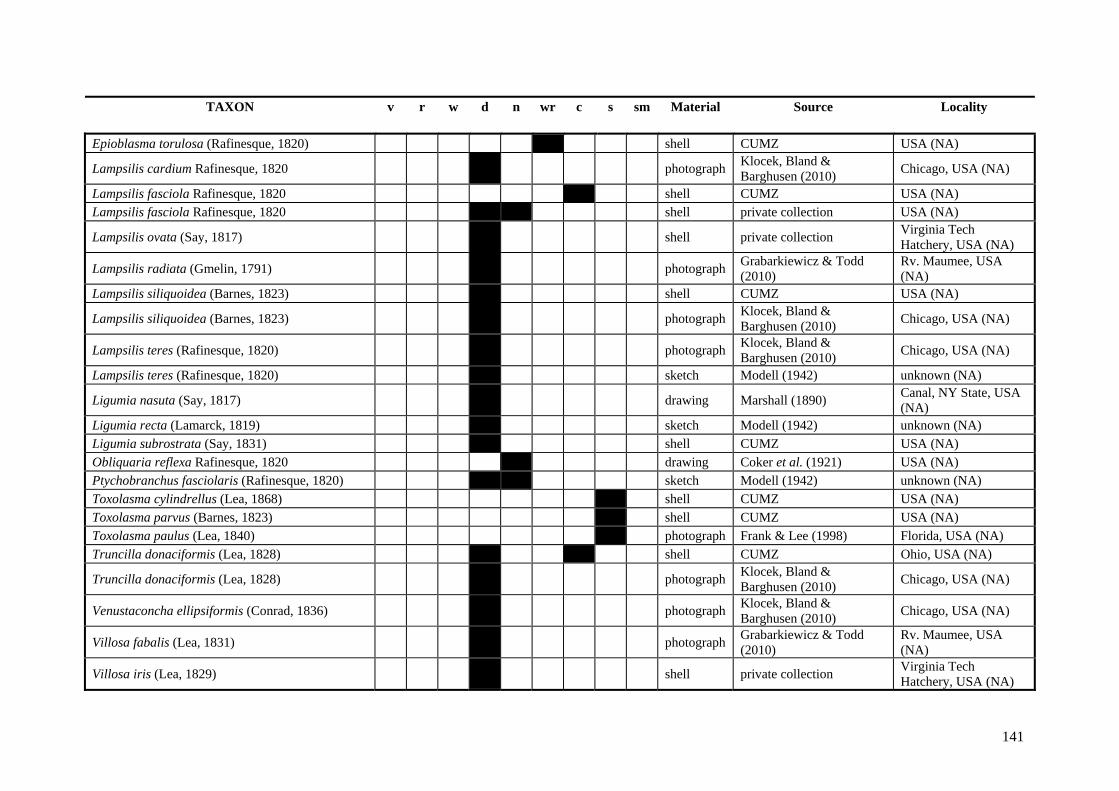
141
TAXON v r w d n wr c s sm Material Source Locality Epioblasma torulosa (Rafinesque, 1820) x shell CUMZ USA (NA)
Lampsilis cardium Rafinesque, 1820 photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Lampsilis fasciola Rafinesque, 1820 x shell CUMZ USA (NA) Lampsilis fasciola Rafinesque, 1820 x x shell private collection USA (NA)
Lampsilis ovata (Say, 1817) x shell private collection Virginia Tech Hatchery, USA (NA)
Lampsilis radiata (Gmelin, 1791) x photograph Grabarkiewicz & Todd (2010)
Rv. Maumee, USA (NA)
Lampsilis siliquoidea (Barnes, 1823) x shell CUMZ USA (NA)
Lampsilis siliquoidea (Barnes, 1823) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Lampsilis teres (Rafinesque, 1820) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Lampsilis teres (Rafinesque, 1820) x sketch Modell (1942) unknown (NA)
Ligumia nasuta (Say, 1817) x drawing Marshall (1890) Canal, NY State, USA (NA)
Ligumia recta (Lamarck, 1819) x sketch Modell (1942) unknown (NA) Ligumia subrostrata (Say, 1831) x shell CUMZ USA (NA) Obliquaria reflexa Rafinesque, 1820 drawing Coker et al. (1921) USA (NA) Ptychobranchus fasciolaris (Rafinesque, 1820) x x sketch Modell (1942) unknown (NA) Toxolasma cylindrellus (Lea, 1868) x shell CUMZ USA (NA) Toxolasma parvus (Barnes, 1823) x shell CUMZ USA (NA) Toxolasma paulus (Lea, 1840) x photograph Frank & Lee (1998) Florida, USA (NA) Truncilla donaciformis (Lea, 1828) x x shell CUMZ Ohio, USA (NA)
Truncilla donaciformis (Lea, 1828) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Venustaconcha ellipsiformis (Conrad, 1836) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Villosa fabalis (Lea, 1831) x photograph Grabarkiewicz & Todd (2010)
Rv. Maumee, USA (NA)
Villosa iris (Lea, 1829) shell private collection Virginia Tech Hatchery, USA (NA)
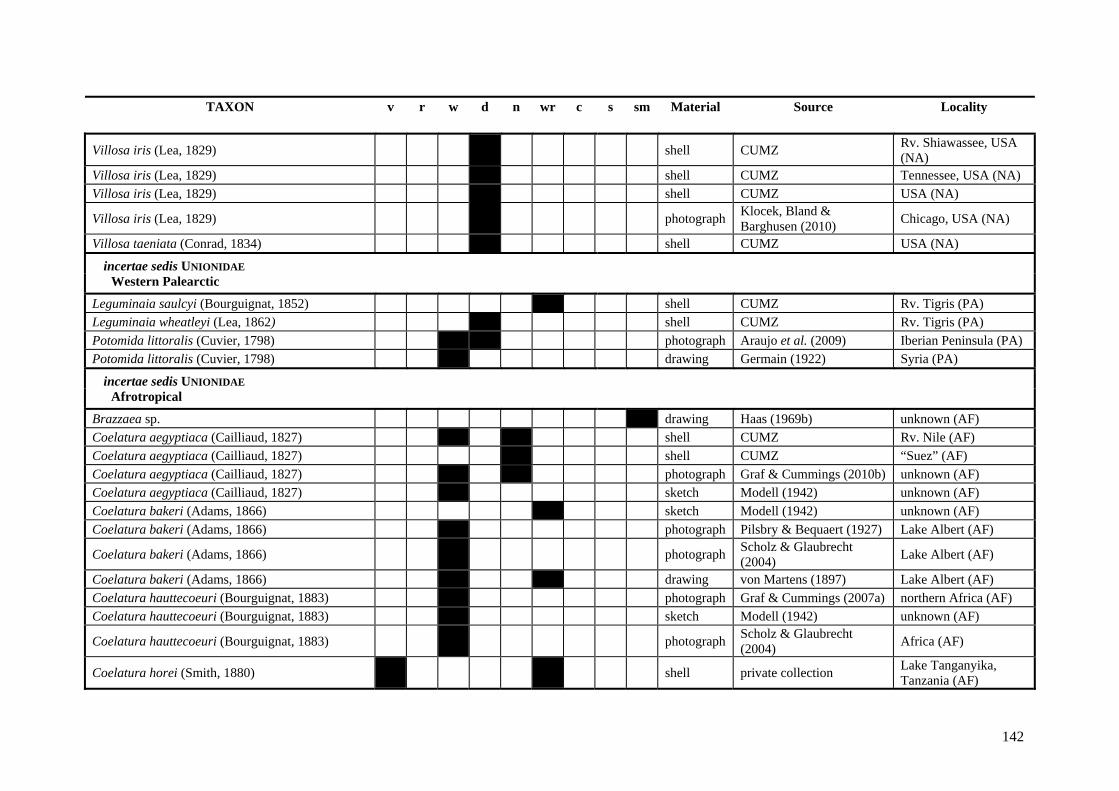
142
TAXON v r w d n wr c s sm Material Source Locality
Villosa iris (Lea, 1829) x shell CUMZ Rv. Shiawassee, USA (NA)
Villosa iris (Lea, 1829) x shell CUMZ Tennessee, USA (NA) Villosa iris (Lea, 1829) x shell CUMZ USA (NA)
Villosa iris (Lea, 1829) photograph Klocek, Bland & Barghusen (2010) Chicago, USA (NA)
Villosa taeniata (Conrad, 1834) x shell CUMZ USA (NA)
incertae sedis UNIONIDAE Western Palearctic
Leguminaia saulcyi (Bourguignat, 1852) x shell CUMZ Rv. Tigris (PA) Leguminaia wheatleyi (Lea, 1862) x shell CUMZ Rv. Tigris (PA) Potomida littoralis (Cuvier, 1798) x x photograph Araujo et al. (2009) Iberian Peninsula (PA) Potomida littoralis (Cuvier, 1798) drawing Germain (1922) Syria (PA)
incertae sedis UNIONIDAE Afrotropical
Brazzaea sp. x drawing Haas (1969b) unknown (AF) Coelatura aegyptiaca (Cailliaud, 1827) x x shell CUMZ Rv. Nile (AF) Coelatura aegyptiaca (Cailliaud, 1827) x shell CUMZ “Suez” (AF) Coelatura aegyptiaca (Cailliaud, 1827) photograph Graf & Cummings (2010b) unknown (AF) Coelatura aegyptiaca (Cailliaud, 1827) x sketch Modell (1942) unknown (AF) Coelatura bakeri (Adams, 1866) x sketch Modell (1942) unknown (AF) Coelatura bakeri (Adams, 1866) x photograph Pilsbry & Bequaert (1927) Lake Albert (AF)
Coelatura bakeri (Adams, 1866) x photograph Scholz & Glaubrecht (2004) Lake Albert (AF)
Coelatura bakeri (Adams, 1866) x x drawing von Martens (1897) Lake Albert (AF) Coelatura hauttecoeuri (Bourguignat, 1883) x photograph Graf & Cummings (2007a) northern Africa (AF) Coelatura hauttecoeuri (Bourguignat, 1883) x sketch Modell (1942) unknown (AF)
Coelatura hauttecoeuri (Bourguignat, 1883) x photograph Scholz & Glaubrecht (2004) Africa (AF)
Coelatura horei (Smith, 1880) x x shell private collection Lake Tanganyika, Tanzania (AF)
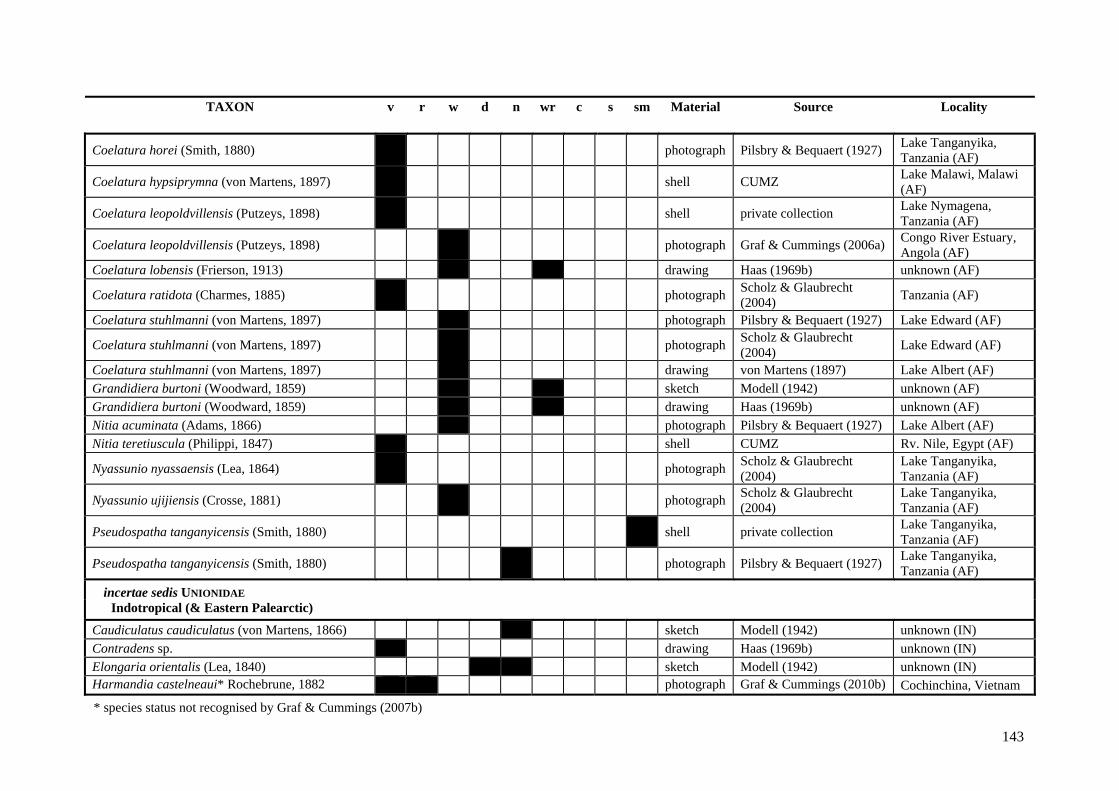
143
TAXON v r w d n wr c s sm Material Source Locality
Coelatura horei (Smith, 1880) x photograph Pilsbry & Bequaert (1927) Lake Tanganyika, Tanzania (AF)
Coelatura hypsiprymna (von Martens, 1897) x shell CUMZ Lake Malawi, Malawi (AF)
Coelatura leopoldvillensis (Putzeys, 1898) x shell private collection Lake Nymagena, Tanzania (AF)
Coelatura leopoldvillensis (Putzeys, 1898) x photograph Graf & Cummings (2006a) Congo River Estuary, Angola (AF)
Coelatura lobensis (Frierson, 1913) x x drawing Haas (1969b) unknown (AF)
Coelatura ratidota (Charmes, 1885) x photograph Scholz & Glaubrecht (2004) Tanzania (AF)
Coelatura stuhlmanni (von Martens, 1897) x photograph Pilsbry & Bequaert (1927) Lake Edward (AF)
Coelatura stuhlmanni (von Martens, 1897) x photograph Scholz & Glaubrecht (2004) Lake Edward (AF)
Coelatura stuhlmanni (von Martens, 1897) x drawing von Martens (1897) Lake Albert (AF) Grandidiera burtoni (Woodward, 1859) x x sketch Modell (1942) unknown (AF) Grandidiera burtoni (Woodward, 1859) x x drawing Haas (1969b) unknown (AF) Nitia acuminata (Adams, 1866) x photograph Pilsbry & Bequaert (1927) Lake Albert (AF) Nitia teretiuscula (Philippi, 1847) x shell CUMZ Rv. Nile, Egypt (AF)
Nyassunio nyassaensis (Lea, 1864) x photograph Scholz & Glaubrecht (2004)
Lake Tanganyika, Tanzania (AF)
Nyassunio ujijiensis (Crosse, 1881) x photograph Scholz & Glaubrecht (2004)
Lake Tanganyika, Tanzania (AF)
Pseudospatha tanganyicensis (Smith, 1880) x shell private collection Lake Tanganyika, Tanzania (AF)
Pseudospatha tanganyicensis (Smith, 1880) x photograph Pilsbry & Bequaert (1927) Lake Tanganyika, Tanzania (AF)
incertae sedis UNIONIDAE Indotropical (& Eastern Palearctic)
Caudiculatus caudiculatus (von Martens, 1866) x sketch Modell (1942) unknown (IN) Contradens sp. x drawing Haas (1969b) unknown (IN) Elongaria orientalis (Lea, 1840) x x sketch Modell (1942) unknown (IN) Harmandia castelneaui* Rochebrune, 1882 x photograph Graf & Cummings (2010b) Cochinchina, Vietnam
* species status not recognised by Graf & Cummings (2007b)
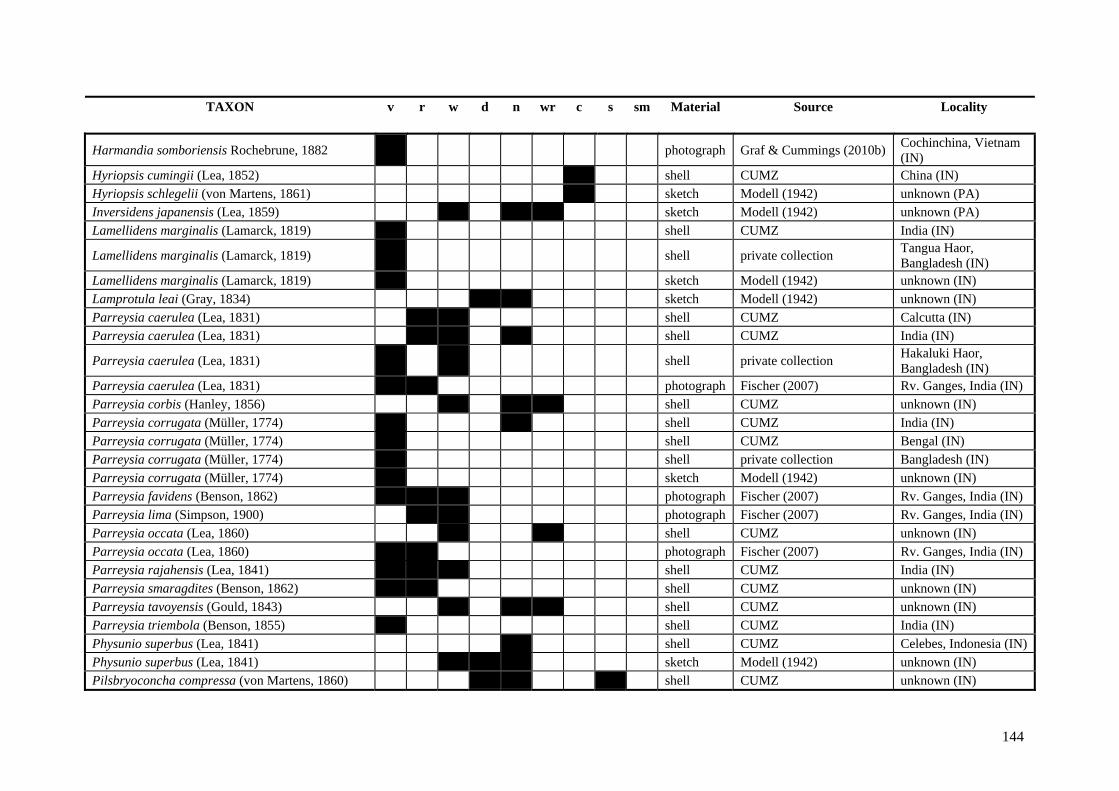
144
TAXON v r w d n wr c s sm Material Source Locality
Harmandia somboriensis Rochebrune, 1882 x photograph Graf & Cummings (2010b) Cochinchina, Vietnam (IN)
Hyriopsis cumingii (Lea, 1852) x shell CUMZ China (IN) Hyriopsis schlegelii (von Martens, 1861) x sketch Modell (1942) unknown (PA) Inversidens japanensis (Lea, 1859) x x x sketch Modell (1942) unknown (PA) Lamellidens marginalis (Lamarck, 1819) x shell CUMZ India (IN)
Lamellidens marginalis (Lamarck, 1819) x shell private collection Tangua Haor, Bangladesh (IN)
Lamellidens marginalis (Lamarck, 1819) x sketch Modell (1942) unknown (IN) Lamprotula leai (Gray, 1834) x x sketch Modell (1942) unknown (IN) Parreysia caerulea (Lea, 1831) x x shell CUMZ Calcutta (IN) Parreysia caerulea (Lea, 1831) x x x shell CUMZ India (IN)
Parreysia caerulea (Lea, 1831) x shell private collection Hakaluki Haor, Bangladesh (IN)
Parreysia caerulea (Lea, 1831) x x photograph Fischer (2007) Rv. Ganges, India (IN) Parreysia corbis (Hanley, 1856) x x shell CUMZ unknown (IN) Parreysia corrugata (Müller, 1774) x x shell CUMZ India (IN) Parreysia corrugata (Müller, 1774) x shell CUMZ Bengal (IN) Parreysia corrugata (Müller, 1774) x shell private collection Bangladesh (IN) Parreysia corrugata (Müller, 1774) x sketch Modell (1942) unknown (IN) Parreysia favidens (Benson, 1862) x x x photograph Fischer (2007) Rv. Ganges, India (IN) Parreysia lima (Simpson, 1900) x x photograph Fischer (2007) Rv. Ganges, India (IN) Parreysia occata (Lea, 1860) x shell CUMZ unknown (IN) Parreysia occata (Lea, 1860) x photograph Fischer (2007) Rv. Ganges, India (IN) Parreysia rajahensis (Lea, 1841) x x x shell CUMZ India (IN) Parreysia smaragdites (Benson, 1862) x x shell CUMZ unknown (IN) Parreysia tavoyensis (Gould, 1843) x x shell CUMZ unknown (IN) Parreysia triembola (Benson, 1855) x shell CUMZ India (IN) Physunio superbus (Lea, 1841) x shell CUMZ Celebes, Indonesia (IN) Physunio superbus (Lea, 1841) x x x sketch Modell (1942) unknown (IN) Pilsbryoconcha compressa (von Martens, 1860) x x x shell CUMZ unknown (IN)
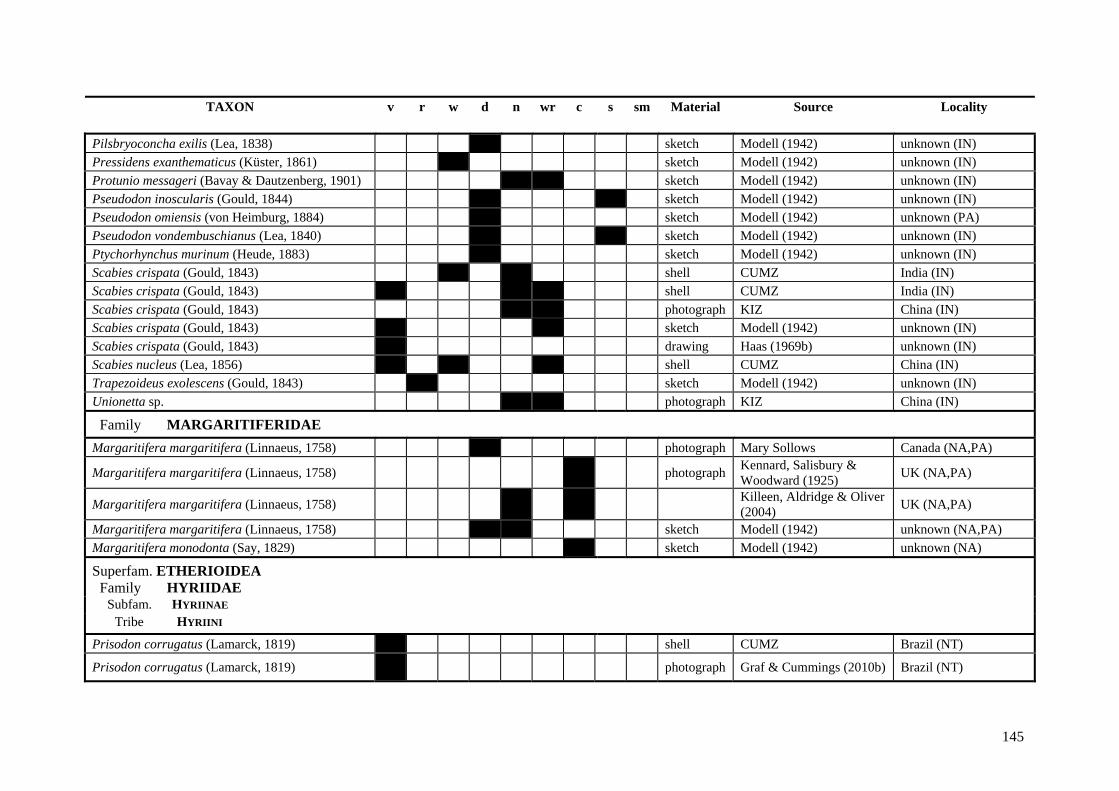
145
TAXON v r w d n wr c s sm Material Source Locality Pilsbryoconcha exilis (Lea, 1838) x sketch Modell (1942) unknown (IN) Pressidens exanthematicus (Küster, 1861) x sketch Modell (1942) unknown (IN) Protunio messageri (Bavay & Dautzenberg, 1901) x x sketch Modell (1942) unknown (IN) Pseudodon inoscularis (Gould, 1844) x x sketch Modell (1942) unknown (IN) Pseudodon omiensis (von Heimburg, 1884) x sketch Modell (1942) unknown (PA) Pseudodon vondembuschianus (Lea, 1840) x x sketch Modell (1942) unknown (IN) Ptychorhynchus murinum (Heude, 1883) x sketch Modell (1942) unknown (IN) Scabies crispata (Gould, 1843) x x shell CUMZ India (IN) Scabies crispata (Gould, 1843) x x shell CUMZ India (IN) Scabies crispata (Gould, 1843) photograph KIZ China (IN) Scabies crispata (Gould, 1843) x x sketch Modell (1942) unknown (IN) Scabies crispata (Gould, 1843) x drawing Haas (1969b) unknown (IN) Scabies nucleus (Lea, 1856) x x x shell CUMZ China (IN) Trapezoideus exolescens (Gould, 1843) x sketch Modell (1942) unknown (IN) Unionetta sp. x photograph KIZ China (IN)
Family MARGARITIFERIDAE
Margaritifera margaritifera (Linnaeus, 1758) x photograph Mary Sollows Canada (NA,PA)
Margaritifera margaritifera (Linnaeus, 1758) x photograph Kennard, Salisbury & Woodward (1925) UK (NA,PA)
Margaritifera margaritifera (Linnaeus, 1758) Killeen, Aldridge & Oliver (2004) UK (NA,PA)
Margaritifera margaritifera (Linnaeus, 1758) x x sketch Modell (1942) unknown (NA,PA) Margaritifera monodonta (Say, 1829) x sketch Modell (1942) unknown (NA)
Superfam. ETHERIOIDEA Family HYRIIDAE
Subfam. HYRIINAE Tribe HYRIINI
Prisodon corrugatus (Lamarck, 1819) x shell CUMZ Brazil (NT)
Prisodon corrugatus (Lamarck, 1819) x photograph Graf & Cummings (2010b) Brazil (NT)
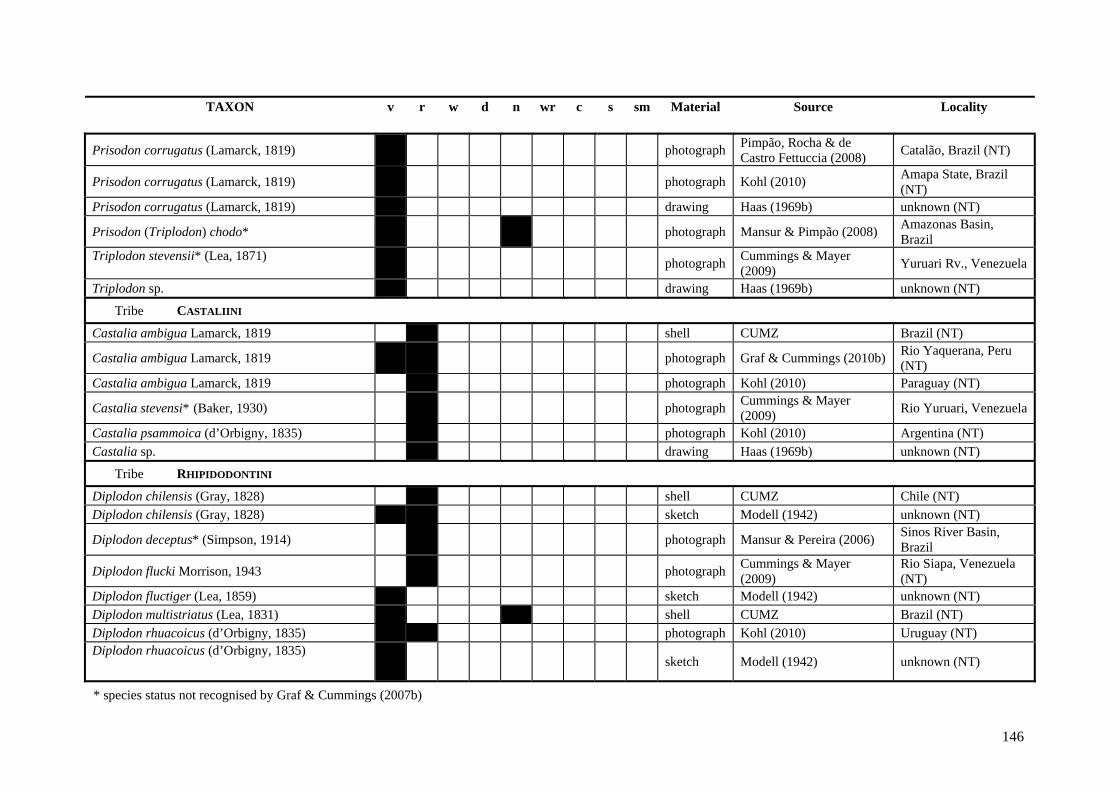
146
TAXON v r w d n wr c s sm Material Source Locality
Prisodon corrugatus (Lamarck, 1819) x photograph Pimpão, Rocha & de Castro Fettuccia (2008) Catalão, Brazil (NT)
Prisodon corrugatus (Lamarck, 1819) photograph Kohl (2010) Amapa State, Brazil (NT)
Prisodon corrugatus (Lamarck, 1819) x drawing Haas (1969b) unknown (NT)
Prisodon (Triplodon) chodo* x x photograph Mansur & Pimpão (2008) Amazonas Basin, Brazil
Triplodon stevensii* (Lea, 1871) photograph Cummings & Mayer
(2009) Yuruari Rv., Venezuela
Triplodon sp. x drawing Haas (1969b) unknown (NT)
Tribe CASTALIINI
Castalia ambigua Lamarck, 1819 x shell CUMZ Brazil (NT)
Castalia ambigua Lamarck, 1819 x x photograph Graf & Cummings (2010b) Rio Yaquerana, Peru (NT)
Castalia ambigua Lamarck, 1819 photograph Kohl (2010) Paraguay (NT)
Castalia stevensi* (Baker, 1930) photograph Cummings & Mayer (2009) Rio Yuruari, Venezuela
Castalia psammoica (d’Orbigny, 1835) x photograph Kohl (2010) Argentina (NT) Castalia sp. x drawing Haas (1969b) unknown (NT)
Tribe RHIPIDODONTINI
Diplodon chilensis (Gray, 1828) x shell CUMZ Chile (NT) Diplodon chilensis (Gray, 1828) x x sketch Modell (1942) unknown (NT)
Diplodon deceptus* (Simpson, 1914) photograph Mansur & Pereira (2006) Sinos River Basin, Brazil
Diplodon flucki Morrison, 1943 x photograph Cummings & Mayer (2009)
Rio Siapa, Venezuela (NT)
Diplodon fluctiger (Lea, 1859) x sketch Modell (1942) unknown (NT) Diplodon multistriatus (Lea, 1831) x x shell CUMZ Brazil (NT) Diplodon rhuacoicus (d’Orbigny, 1835) x photograph Kohl (2010) Uruguay (NT) Diplodon rhuacoicus (d’Orbigny, 1835)
x sketch Modell (1942) unknown (NT)
* species status not recognised by Graf & Cummings (2007b)
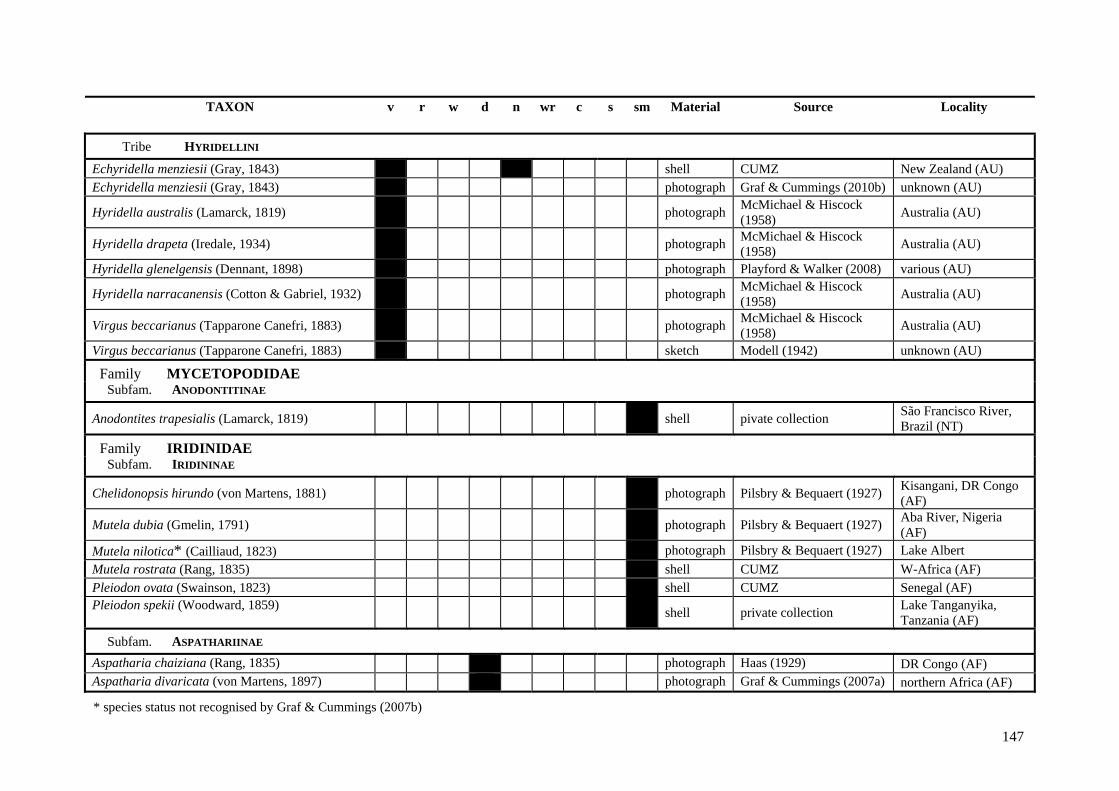
147
TAXON v r w d n wr c s sm Material Source Locality
Tribe HYRIDELLINI
Echyridella menziesii (Gray, 1843) x shell CUMZ New Zealand (AU) Echyridella menziesii (Gray, 1843) x photograph Graf & Cummings (2010b) unknown (AU)
Hyridella australis (Lamarck, 1819) x photograph McMichael & Hiscock (1958) Australia (AU)
Hyridella drapeta (Iredale, 1934) x photograph McMichael & Hiscock (1958) Australia (AU)
Hyridella glenelgensis (Dennant, 1898) x photograph Playford & Walker (2008) various (AU)
Hyridella narracanensis (Cotton & Gabriel, 1932) x photograph McMichael & Hiscock (1958) Australia (AU)
Virgus beccarianus (Tapparone Canefri, 1883) x photograph McMichael & Hiscock (1958) Australia (AU)
Virgus beccarianus (Tapparone Canefri, 1883) x sketch Modell (1942) unknown (AU)
Family MYCETOPODIDAE Subfam. ANODONTITINAE
Anodontites trapesialis (Lamarck, 1819) shell pivate collection São Francisco River, Brazil (NT)
Family IRIDINIDAE Subfam. IRIDININAE
Chelidonopsis hirundo (von Martens, 1881) x photograph Pilsbry & Bequaert (1927) Kisangani, DR Congo (AF)
Mutela dubia (Gmelin, 1791) x photograph Pilsbry & Bequaert (1927) Aba River, Nigeria (AF)
Mutela nilotica* (Cailliaud, 1823) x photograph Pilsbry & Bequaert (1927) Lake Albert Mutela rostrata (Rang, 1835) x shell CUMZ W-Africa (AF) Pleiodon ovata (Swainson, 1823) x shell CUMZ Senegal (AF) Pleiodon spekii (Woodward, 1859) x shell private collection Lake Tanganyika,
Tanzania (AF) Subfam. ASPATHARIINAE
Aspatharia chaiziana (Rang, 1835) x photograph Haas (1929) DR Congo (AF) Aspatharia divaricata (von Martens, 1897) x photograph Graf & Cummings (2007a) northern Africa (AF)
* species status not recognised by Graf & Cummings (2007b)
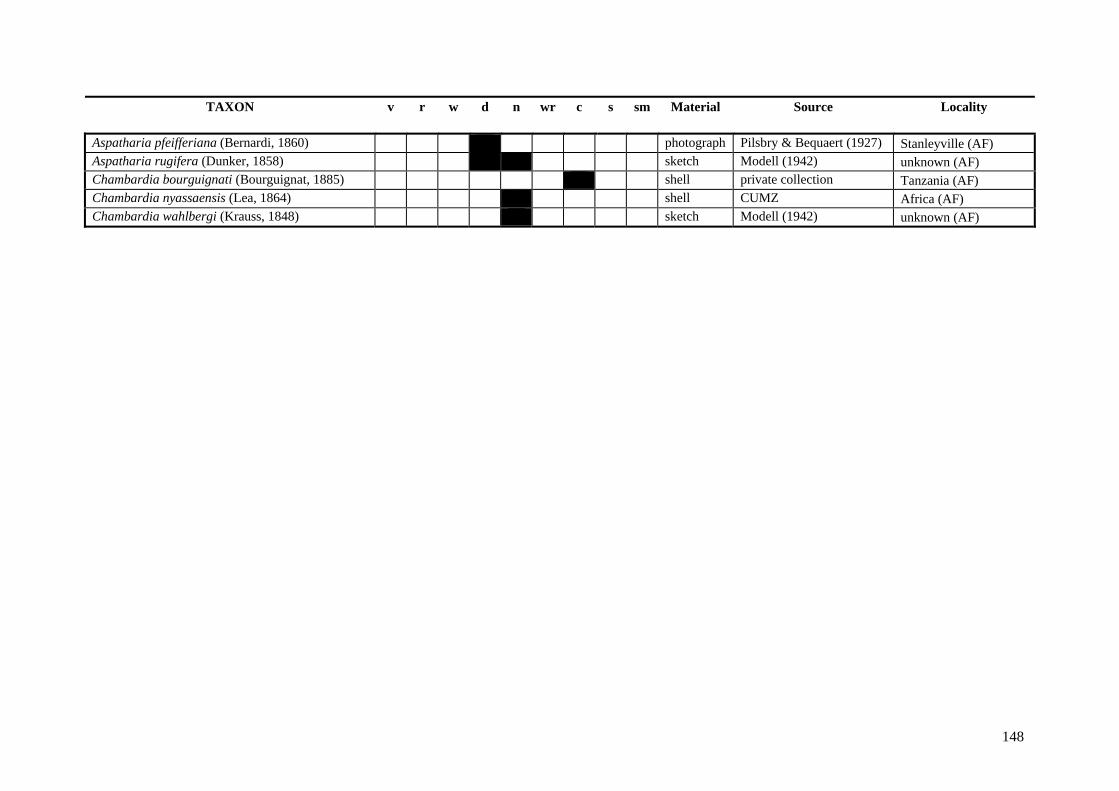
148
TAXON v r w d n wr c s sm Material Source Locality Aspatharia pfeifferiana (Bernardi, 1860) x photograph Pilsbry & Bequaert (1927) Stanleyville (AF) Aspatharia rugifera (Dunker, 1858) x sketch Modell (1942) unknown (AF) Chambardia bourguignati (Bourguignat, 1885) x shell private collection Tanzania (AF) Chambardia nyassaensis (Lea, 1864) x shell CUMZ Africa (AF) Chambardia wahlbergi (Krauss, 1848) x sketch Modell (1942) unknown (AF)

BIBLIOGRAPHY


151
BIBLIOGRAPHY
Agrawal, H.P. 1974. Sexuality of relative growth in the freshwater mussel, Parreysia wynegungaensis (Lea) (Pelecypoda: Unionidae). Indian Journal of Zoology 2: 29-31.
Agrell, I. 1948. The shell morphology of some Swedish unionides as affected by ecological conditions. Arkiv för Zoologi 41A: 1-30.
Ajmone-Marsan, J., Valentini, A., Cassandro, M., Vecchiotti-Antaldi, G., Bertoni, G. & Kuiper, M. 1997. AFLPTM markers for DNA fingerprinting in cattle. Animal Genetics 82: 418-426.
Aldridge, D.C. 1997. Reproductive ecology of bitterling (Rhodeus sericeus Pallas) and unionid mussels. PhD thesis, University of Cambridge, UK. 154 pp.
Aldridge, D.C. 1999a. Development of European bitterling in the gills of freshwater mussels. Journal of Fish Biology 54: 138–151.
Aldridge, D.C. 1999b. The morphology, growth and reproduction of Unionidae (Bivalvia) in a fenland waterway. Journal of Molluscan Studies 65: 47-60.
Aldridge, D.C. 2004. Conservation of freshwater unionid mussels in Britain. Journal of Conchology Special Publication 3: 81-90.
Aldridge, D.C., Fayle, T. & Jackson, N. 2007. Freshwater mussel abundance predicts biodiversity in UK lowland rivers. Aquatic Conservation: Marine and Freshwater Ecosystems 17: 554-564.
Aller, R.C. 1974. Prefabrication of shell ornamentation in the bivalve Laternula. Lethaia 7: 43-56.
Altnöder, K. 1926. Beobachtungen über die Biologie von Margaritana margaritifera und über die Ökologie ihres Wohnorts. Archiv für Hydrobiologie 17: 423-447.
Amyot, J.-P. & Downing, J.A. 1991. Endo- and epibenthic distribution of the unionid mollusc Elliptio complanata. Journal of the North American Benthological Society 10: 280-285.
Amyot, J.-P. & Downing, J.A. 1997. Seasonal variation in vertical and horizontal movement of the freshwater bivalve Elliptio complanata (Mollusca: Bivalvia). Freshwater Biology 37: 345-354.
Anderson, R.V. & Ingham, R.E. 1978. Character variation in mollusc populations of Lampsilis Rafinesque, 1820. Transactions of the Illinois State Academy of Science 71: 403-411.
Andersson, M. 1994. Sexual selection. Princeton University Press: Princeton, NJ. 624 pp.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
152
Anthony, J.L. & Downing, J.A. 2001. Exploitation trajectory of a declining fauna: a century of freshwater mussel fisheries in North America. Canadian Journal of Fisheries and Aquatic Sciences 58: 2071-2090.
Araujo, R., Reis, J., Machordom, A., Toledo, C., Madeira, M.J., Gómez, I., Velasco, J.C., Morales, J., Barea, J.M., Ondina, P. & Ayala, I. 2009. Las náyades de la península Ibérica. Iberus 27: 7-72.
Bailey, R.C. & Green, R.H. 1988. Within-basin variation in the shell morphology and growth rate of a freshwater mussel. Canadian Journal of Zoology 66: 1704-1708.
Baker, A.M., Bartlett, C., Bunn, S.E., Goudkamp, K., Sheldon, F. & Hughes, J.M. 2003. Cryptic species and morphological plasticity in long-lived bivalves (Unionoida : Hyriidae) from inland Australia. Molecular Ecology 12: 2707-2717.
Balfour, D.L. & Smock, L.A. 1995. Distribution, age structure, and movement of the freshwater mussel Elliptio complanata (Mollusca: Unionidae) in a headwater stream. Journal of Freshwater Ecology 10: 225-268.
Ball, G.H. 1922. Variation in freshwater mussels. Ecology 3: 93-121.
Balla, S.A. & Walker, K.F. 1991. Shape variation in the Australian freshwater mussel Alathyria jacksoni Iredale (Bivalvia, Hyriidae). Hydrobiologia 220: 89-98.
Bauer, G. 1998. Allocation policy of female freshwater mussels. Oecologia 117: 90-94.
Bauer, G. 2001. Life history variation on different taxonomic levels of naiads. In: Bauer, G. & Wächtler K. (eds.). Ecology and Evolution of the Freshwater Mussels Unionoida. Berlin: Springer-Verlag. pp. 83-91.
Beaumont, A.R. 2008. Dfdist package. Available at http://www.rubic.rdg.ac.uk/~mab/stuff/ (accessed April 2009).
Beaumont, M.A. & Balding, D.J. 2004. Identifying adaptive genetic divergence among populations from genome scans. Molecular Ecology 13: 969–980.
Beaumont, M.A. & Nichols, R.A. 1996. Evaluating loci for use in the genetic analysis of population structure. Proceedings of the Royal Society of London, Series B 263: 1619-1626.
Beedham, G.E. 1958. Observations on the mantle of the Lamellibranchia. Quarterly Journal of Microscopical Science 99: 181-197.
Bensch, S. & Åkesson, M. 2005. Ten years of AFLP in ecology and evolution: Why so few animals? Molecular Ecology 14: 2899-2914.
Berg, D.J., Cantonwine, E.G., Hoeh, W.R. & Guttman, S.I. 1998. Genetic structure of Quadrula quadrula (Bivalva: Unionidae): little variation across large distances. Journal of Shellfish Research 17: 1365-1373.
Bergek, S. & Björklund, M. 2007. Cryptic barriers to dispersal within a lake allow genetic differentiation of Eurasian perch. Evolution 61: 2035-2041.

Bibliography
153
Berrie, A.D. & Bioze, B.J. 1985. The fish hosts of Unio glochidia in the River Thames. Verhandlungen der Internationalen Vereinigung für theoretische und angewandte Limnologie 22: 2712-2716.
Bieler, R. & Mikkelsen, P.M. 1996. Bivalvia – a look at the Branches. Zoological Journal of the Linnean Society 148: 223–235.
Blažek, R. & Gelnar, M. 2006. Temporal and spatial distribution of glochidial larval stages of European unionid mussels (Mollusca: Unionidae) on host fishes. Folia Parasitologica 53: 98–106.
Blears, M.J., De Grandis, S.A., Lee, H. & Trevors, J.T. 1998. Amplified fragment length polymorphism (AFLP): a review of the procedure and its applications. Journal of Industrial Microbiology & Biotechnology 21: 99-114.
Bloomer, H.H. 1934. On the sex, and sex-modification of the gill, of Anodonta cygnea. Proceedings of the Malacological Society of London 21: 20-29.
Bloomer, H.H. 1937. On distinguishing the shell of Anodonta cygnea from that of Anodonta anatina. Journal of Conchology 20: 321-327.
Bloomer, H.H. 1938. The British species of Anodonta Lamarck, and their varieties. Journal of Conchology 21: 33-48.
Bogan, A.E. 1993. Freshwater bivalve extinctions (Mollusca: Unionoida): A search for causes. American Zoologist 33: 599-609.
Bogan, A.E. 2008. Global diversity of freshwater mussels (Mollusca, Bivalvia) in freshwater. Hydrobiologia 595: 139–147.
Bogan, A.E. & Hoeh, W.R. 2000. On becoming cemented: evolutionary relationships among the genera in the freshwater bivalve family Etheriidae (Bivalvia: Unionoida). In: Harper, E.M., Taylor J.D. & Crame J.A. (eds.). The evolutionary biology of the Bivalvia. London: The Geological Society of London. pp. 31-46.
Bogan, A.E. & Roe, K. 2008. Freshwater bivalve (Unioniformes) diversity, systematics, and evolution: status and future directions. Journal of the North American Benthological Society 27: 349-369.
Bolnick, D.I., Snowberg, L.K., Patenia, C., Stutz, W.E., Ingram, T. & Lau, O.L. 2009. Phenotype-dependent native habitat preference facilitates divergence between parapatric lake and stream stickleback. Evolution 63: 2004-2016.
Bonetto, A.A. 1966. Especies de la subfamilia Monocondylaeinae en las aguas del sistema del Rio de La Plata (Moll. Mutelacea). Archiv für Molluskenkunde 95: 3-14.
Bonin, A., Bellemain, E., Bronken Eidesen, P., Pompanon, F., Brochmann, C. & Taberlet, P. 2004. How to track and assess genotyping errors in population genetic studies. Molecular Ecology 13: 3261-3273.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
154
Bonin, A., Ehrich, D. & Manel, S. 2007. Statistical analysis of amplified fragment length polymorphism data: a toolbox for molecular ecologists and evolutionists. Molecular Ecology 16: 3737-3758.
Bottjer, D.J. & Carter, J.G. 1980. Functional and phylogenetic significance of projecting periostracal structures in the Bivalvia (Mollusca). Journal of Paleontology 54: 200-216.
Bouza, C., Castro, J., Martínez, P., Amaro, R., Fernández, C., Ondina, P., Outeiro, A. & San Miguel, E. 2007. Threatened freshwater pearl mussel Margaritifera margaritifera L. in NW Spain: low and very structured genetic variation in southern peripheral populations assessed using microsatellite markers. Conservation Genetics 8: 937-948.
Brander, T. 1954. Über Geschlechtsdimorphismus bei europäischen Unionazeen. Archiv für Molluskenkunde 83: 163-172.
Branson, C.C. 1935. Fresh-water invertebrates from the Morrison (Jurassic?) of Wyoming. Journal of Paleontology 9: 514-522.
Brecko, J., Huyghe, K., Vanhooydonck, B., Herrel, A., Grbac, I. & Van Damme, R. 2008. Functional and ecological relevance of intraspecific variation in body size and shape in the lizard Podarcis melisellensis (Lacertidae). Biological Journal of the Linnean Society 94: 251-264.
Bridgeman, J.B. 1875. A variety caused by a locality. Quarterly Journal of Conchology 1: 70.
Buchner, O. 1900. Beiträge zur Formenkenntnis der einheimischen Anodonten unter besonderer Berücksichtigung der württembergischen Vorkommnisse. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 56: 60-223.
Buchner, O. 1910. Ueber individuelle Formverschiedenheiten bei Anodonten. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 65: 50.
Buhay, J.E., Serb, J.M., Dean, C.R., Parham, Q. & Lydeard, C. 2002. Conservation genetics of two endangered unionid bivalve species, Epioblasma florentina walkeri and E. capsaeformis (Unionidae: Lampsilini). Journal of Molluscan Studies 68: 385-391.
Caballero, A., Quesada, H. & Rolán-Alvarez, E. 2008. Impact of amplified fragment length polymorphism size homoplasy on the estimation of population genetic diversity and the detection of selective loci. Genetics 179: 539-554.
Callil, C.T. & Mansur, M.C.D. 2005. Ultrastructural analysis of the shells of Anodontites trapesialis (Lamarck) and Anodontites elongatus (Swaison) (Mollusca, Bivalvia, Etherioidea) from the Mato Gross Pantanal Region, Brazil. Revista Brasileira de Zoologia 22: 724-734.
Cameron, C.J., Cameron, I.F. & Paterson, C.G. 1979. Contribution of organic shell matter to biomass estimates of unionid bivalves. Canadian Journal of Zoology 57: 1666-1669.

Bibliography
155
Campbell, D.C., Serb, J.M., Buhay, J.E., Roe, K.J., Minton, R.L. & Lydeard, C. 2005. Phylogeny of North American amblemines (Bivalvia, Unionoida): prodigious polyphyly proves pervasive across genera. Invertebrate Biology 124: 131-164.
Carter, J.G. 1978. Ecology and evolution of the Gastrochaenae (Mollusca, Bivalvia) with notes on the evolution of the endolithic habitat. Peabody Museum Bulletin 41: 1-92.
Carter, J.G. 1990. Evolutionary significance of shell microstructures in the Palaeotaxodonta, Pteriomorpha and Isofilibranchia (Bivalvia: Mollusca). In: Carter, J.G. (ed.) Skeletal biomineralization: patterns, processes and evolutionary trends. New York: Van Nostrand Reinhold. pp. 136–271.
Carter, J.G. & Aller, R.C. 1975. Calcification in the bivalve periostracum. Lethaia 8: 315-320.
Carter, J.G., Campbell, D.C. & Campbell, M.R. 2006. Morphological phylogenetics of the early Bivalvia. In Malchus, N. & Pons J.M., (eds.). Organic Diversity and Evolution Supplement. Abstracts and Posters of the “International Congress on Bivalvia” at the Universitat Autònoma de Barcelona, Spain, 22-27 July 2006, 20-21.
Checa, A.G. 2000. A new model for periostracum and shell formation in Unionidae (Bivalvia, Mollusca). Tissue & Cell 32: 405-416.
Checa, A.G. 2002. Fabricational morphology of oblique ribs in bivalves. Journal of Morphology 254: 195-209.
Checa, A.G. & Harper, E.M. in press. Spikey bivalves: intra-periostracal crystal growth in anomalodesmatans. Biological Bulletin.
Checa, A.G. & Jiménez-Jiménez, A.P. 2003a. Evolutionary morphology of oblique ribs of bivalves. Palaeontology 46: 709-724.
Checa, A.G. & Jiménez-Jiménez, A.P. 2003b. Rib fabrication in Ostreoidea and Plicatuloidea (Bivalvia, Pteriomorphia) and its evolutionary significance. Zoomorphology 122: 145-159.
Checa, A.G. & Rodríguez-Navarro, A. 2001. Geometrical and crystallographic constraints determine the self-organization of shell microstructures in Unionidae (Bivalvia: Mollusca). Proceedings of the Royal Society of London, Series B 268: 771-778.
Claassen, C. 1998. Shells. Cambridge University Press: Cambridge. 266 pp.
Clarke, A.H. 1973. The fresh water mollusks of the Canadian Interior Basin. Malacologia 13: 1-509.
Clarke, A.H. 1981. The tribe Alasmidontini (Unionidae: Anodontinae), part I. Pegias, Alasmidonta, and Arcidens. Smithsonian Contributions to Zoology 326: 1-101.
Clarke, A.H. 1985. The tribe Alasmidontini (Unionidae: Anodontinae), part II. Lasmigona and Simpsonaias. Smithsonian Contributions to Zoology 399: 1-75.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
156
Clarke, A.H. 1986. The mesoconch: a record of juvenile life in Unionidae. Malacology Data Net 1: 21-36.
Coker, R.E., Shira, A.F., Clark, H.W. & Howard, A.D. 1921. Natural history and propagation of fresh-water mussels. Bulletin of the Bureau of Fisheries (U.S.) 37: 75-181.
Conde-Padín, P., Carvajal-Rodríguez, A., Carballo, M., Caballero, A. & Rolán-Alvarez, E. 2007. Genetic variation for shell traits in a direct-developing marine snail involved in a putative sympatric ecological speciation process. Evolutionary Ecology 21: 635-650.
Connolly, M. 1939. A monographic study of South African non-marine Mollusca. Annals of the South African Museum 33: 1-660.
Cooper, M.R. 1991. Lower Cretaceous Trigonioida (Mollusca, Bivalvia) from the Algoa Basin, with a revised classification of the order. Annals of the South African Museum 100: 1-52.
Cox, L.R. 1969. Superfamily Trigoniacea. In: Moore, R.C. (ed.) Treatise on invertebrate paleontology. Lawrence, Kansas: Geological Society of America & University of Kansas Press. pp. N471-N488.
Crampton, J.S. & Haines, A.J. 1996. Users' manual for programs HANGLE, HMATCH and HCURVE for the Fourier shape analysis of two-dimensional outlines Institute of Geological and Nuclear Sciences Science Report. 1-28.
Crampton, J.S. & Haines, A.J. 2007. Programs HANGLE, HMATCH and HCURVE for the Fourier shape analysis of two-dimensional outlines.
Crawley, M.J. 2002. Statistical computing, an introduction to data analysis using S-plus. John Wiley and Sons Ltd: Chichester.
Crosby, M.P. & Gale, L.D. 1990. A review of bivalve condition index methodologies with a suggested standard method. Journal of Shellfish Research 9: 233-239.
Cummings, K.S. & Mayer, C.A. 2009. The Freshwater Mussels (Bivalvia: Etherioidea) of Venezuela. Available at http://www.inhs.illinois.edu/animals_plants/mollusk/SA/Ven.html (accessed 2010).
Cvancara, A.M. 1972. Lake mussel distribution as determined with SCUBA. Ecology 53: 154-157.
Dasmahapatra, K.K., Hoffman, J.I. & Amos, W. 2009. Pinniped phylogenetic relationships inferred using AFLP markers. Heredity 103: 168-177.
Davis, G.M. 1983. Relative roles of molecular genetics, anatomy, morphometrics and ecology in assessing relationships among North American Unionidae (Bivalvia). In: Oxford, G.S. & Rollinson D. (eds.). Protein polymorphism: adaptive and taxonomic significance. London and New York: Academic Press. pp. 193-222.

Bibliography
157
Davis, G.M. 1984. Genetic relationships among some North American Unionidae (Bivalvia): Sibling species, convergence, and cladistic relationships. Malacologia 25: 629-648.
Davis, G.M. & Fuller, S.L.H. 1981. Genetic relationships among recent Unionacea (Bivalvia) of North America. Malacologia 20: 217-253.
Davis, G.M., Heard, W.H., Fuller, S.L.H. & Hesterman, C. 1981. Molecular genetics and speciation in Elliptio and its relationships to other taxa of North American Unionidae (Bivalvia). Biological Journal of the Linnean Society 15: 131-150.
Davis, G.M. & Ruff, M. 1974. Oncomelania hupensis (Gastropoda: Hydrobiidae): hybridization, genetics, and transmission of Schistosoma japonicum. Malacological Review 6: 181-197.
de Aranzamendi, M.C., Sahade, R., Tatian, M. & Chiappero, M.B. 2008. Genetic differentiation between morphotypes in the Antarctic limpet Nacella concinna as revealed by inter-simple sequence repeat markers. Marine Biology 154: 875-885.
de Wolf, H., Backeljau, T. & Verhagen, R. 1998. Spatio-temporal genetic structure and gene flow between two distinct shell morphs of the planktonic developing periwinkle Littorina striata (Mollusca: Prosobranchia). Marine Ecology progress series 163: 155–163.
Dell, R.K. 1953. The freshwater Mollusca of New Zealand. Transactions of the Royal Society of New Zealand 81: 221-237.
Doebeli, M. & Dieckmann, U. 2003. Speciation along environmental gradients. Nature 421: 259-264.
Dolmen, D. & von Proschwitz, T. 1999. Searching for the swan mussel Anodonta cygnea in Norway: And the use of characters to distinguish it from the duck mussel A. anatina. Fauna 52: 82-88.
Downing, J.A., Amyot, J.-P., Pérusse, M. & Rochon, Y. 1989. Viceral sex, hermaphroditism and protandry in a population of the freshwater bivalve Elliptio complanata. Journal of the North American Benthological Society 8: 92-99.
Downing, W.L. & Downing, J.A. 1993. Molluscan shell growth and loss. Nature 362: 506.
Dyer, A.T. & Leonard, K.J. 2000. Contamination, error, and nonspecific molecular tools. Phytopathology 90: 565-567.
Eagar, R.M.C. 1948. Variation in shape of shell with respect to ecological station. A review dealing with Recent Unionidae and certain species of the Anthracosiidae in Upper Carboniferous times. Proceedings of the Royal Society of Edinburgh, Section B 63: 130-148.
Eagar, R.M.C. 1971. Variation in shape of shell in relation to palaeoecological station in some non-marine Bivalvia of the Coal Measures of south-east Kentucky and Britain. Compte rendu te Congres International de Stratigraphie et de Geologie du Carbonifere, Krefeld 2: 387-413.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
158
Eagar, R.M.C. 1974. Shape of shell of Carbonicola in relation to burrowing. Lethaia 7: 219-238.
Eagar, R.M.C. 1977. Shape of shell in relation to weight of Margaritifera margaritifera. Journal of Conchology 29: 207-218.
Eagar, R.M.C. 1978. Shape and function of the shell: a comparison of some living and fossil bivalve mussels. Biological Reviews of the Cambridge Philosophical Society 53: 169-210.
Elderkin, C.L., Christian, A.D., Vaughn, C.C., Metcalfe-Smith, J.L. & Berg, D.J. 2007. Population genetics of the freshwater mussel, Amblema plicata (Say 1817) (Bivalvia: Unionidae): Evidence of high dispersal and post-glacial colonization. Conservation Genetics 8: 355-372.
Fischer, W. 2007. The Bivalvia of the Ganga River. Available at http://ipp.boku.ac.at/private/wf/Bivalvia_India/Bivalvia_Ganga_River.html (accessed 2010).
Forsyth, D.J. & McCallum, I.D. 1978. Xenochironomus canterburyensis (Diptera: Chironomidae), an insectan inquiline commensal of Hyridella menziesi (Mollusca: Lamellibranchia). Journal of Zoology 186: 331-334.
Frank, B. & Lee, H.G. 1998. The Jacksonville Shell Club. Available at http://www.jaxshells.org/ (accessed 2010).
Frierson, L.S. 1909a. Notes on Oriental Unionidae. Nautilus 24: 97-98.
Frierson, L.S. 1909b. Remarks on the subfamilies Hyriinae and Unioninae. Nautilus 22: 106-107.
Gangloff, M.M., Lenertz, K.K. & Feminella, J.W. 2008. Parasitic mite and trematode abundance are associated with reduced reproductive output and physiological condition of freshwater mussels. Hydrobiologia 610: 25-31.
Geist, J. & Auerswald, K. 2007. Physicochemical stream bed characteristics and recruitment of the freshwater pearl mussel (Margaritifera margaritifera). Freshwater Biology 52: 2299-2316.
Geist, J. & Kuehn, R. 2005. Genetic diversity and differentiation of central European freshwater pearl mussel (Margaritifera margaritifera L.) populations: implications for conservation and management. Molecular Ecology 14: 425-439.
Germain, L. 1922. Mollusques terrestres et fluviatiles de Syrie. Part 2: Pelecypodes. J.-B. Baillere et Fils: Paris.
Ghent, A.W., Singer, R. & Johnson-Singer, L. 1978. Depth distributions determined with SCUBA, and associated studies of the freshwater unionid clams Elliptio complanata and Anodonta grandis in Lake Bernard, Ontario. Canadian Journal of Zoology 56: 1654-1663.

Bibliography
159
Giribet, G. & Wheeler, W. 2002. On bivalve phylogeny: a high-level analysis of the Bivalvia (Mollusca) based on combined morphology and DNA sequence data. Invertebrate Biology 121: 271-324.
Glover, E.A. & Taylor, J.D. 2010. Needles and pins: acicular crystalline periostracal calcification in venerid bivalves (Bivalvia: Veneridae). Journal of Molluscan Studies 76: 157-179.
Good, S.C. 1989. Nonmarine mollusca in the Upper Triassic Chinle Formation and related strata of the western Interior: systematics and distribution. In: Lucas, S.G. & Hunt A.P. (eds.). Dawn of the Age of Dinosaurs in the American South-west. Albuquerque, New Mexico: New Mexico Museum of Natural History. pp. 233-248.
Good, S.C. 1998. Freshwater bivalve fauna of the Late Triassic (Carnian-Norian) Chinle, Dockum, and Dolores Formations of the Southwest United States. In: Johnston, P.A. & Haggart J.W. (eds.). Bivalves: an eon of evolution - palaeobiological studies honoring Norman D. Newell. Calgary: University of Calgary Press.
Good, S.C. 2004. Paleoenvironmental and paleoclimatic significance of freshwater bivalves in the Upper Jurassic Morrison Formation, Western Interior, USA. Sedimentary Geology 167: 163-176.
Goodwin, B.J. & Fish, J.D. 1977. Inter- and intraspecific variation in Littorina obtusata and L. mariae (Gastropoda: Prosobranchia). Journal of Molluscan Studies 43: 241-254.
Grabarkiewicz, J. & Todd, C. 2010. Freshwater Mussels of the Maumee Drainage. Available at http://www.farmertodd.com/musselguide/ (accessed 2010).
Graf, D.L. 2000. The Etherioidea revisited: A phylogenetic analysis of hyriid relationships (Mollusca: Bivalvia: Paleoheterodonta: Unionoida). Occasional Papers of the Museum of Zoology University of Michigan: 1-21.
Graf, D.L. & Cummings, K.S. 2006a. Freshwater mussels (Mollusca: Bivalvia: Unionoida) of Angola, with description of a new species, Mutela wistarmorrisi. Proceedings of the Academy of Natural Sciences of Philadelphia 155: 163-194.
Graf, D.L. & Cummings, K.S. 2006b. Palaeoheterodont diversity (Mollusca: Trigonioida + Unionoida): what we know and what we wish we knew about freshwater mussel evolution. Zoological Journal of the Linnean Society 148: 343-394.
Graf, D.L. & Cummings, K.S. 2007a. Preliminary review of the freshwater mussels (Mollusca: Bivalvia: Unionoida) of northern Africa, with emphasis on the Nile. Journal of the Egyptian German Society for Zoology 53D: 89-118.
Graf, D.L. & Cummings, K.S. 2007b. Review of the systematics and global diversity of freshwater mussel species (Bivalvia: Unionoida). Journal of Molluscan Studies 73: 291-314.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
160
Graf, D.L. & Cummings, K.S. 2010a. Comments on the value of COI for family-level freshwater Mussel systematics: a reply to Hoeh, Bogan, Heard & Chapman. Malacologia 52: 191-197.
Graf, D.L. & Cummings, K.S. 2010b. The MUSSEL Project — Home Page. Available at http://mussel-project.ua.edu (accessed 2010).
Graf, D.L. & Foighil, D.O. 2000. The evolution of brooding characters among the freshwater pearly mussels (Bivalvia : Unionoidea) of North America. Journal of Molluscan Studies 66: 157-170.
Gray, J.E. 1825. Conchological observations, being an attempt to fix the study of conchology on a firm basis. Journal of Zoology 1: 204-223.
Grier, N.M. 1920. Morphological features of certain mussel-shells found in Lake Erie, compared with those of the corresponding species found in the drainage of the Upper Ohio. Annales of the Carnegie Museum 13: 145-182.
Grier, N.M. & Mueller, J.F. 1926. Further studies in correlation of shape and station in fresh water mussels. Bulletin of the Wagner Free Institute of Science 1: 11-28.
Grizzle, J.M. & Brunner, C.J. 2009. Infectious Diseases of Freshwater Mussels and Other Freshwater Bivalve Mollusks. Reviews in Fisheries Science 17: 425-467.
Grobler, P.J., Jones, J.W., Johnson, N.A., Beaty, B., Struthers, J., Neves, R.J. & Hallerman, E.M. 2006. Patterns of genetic differentiation and conservation of the Slabside Pearlymussel, Lexingtonia dolabelloides (Lea, 1840) in the Tennessee drainage. Journal of Molluscan Studies 72: 65-75.
Gu, Z. 1998. Evolutionary trends in non-marine Cretaceous bivalves of northeast China. In: Johnston, P.A. & Haggart J.W. (eds.). Bivalves: an eon of evolution - palaeobiological studies honoring Norman D. Newell. Calgary: University of Calgary Press. pp. 267-276.
Guerra-Varela, J., Colson, I., Backeljau, T., Breugelmans, K., Hughes, R.N. & Rolán-Alvarez, E. 2009. The evolutionary mechanism maintaining shell shape and molecular differentiation between two ecotypes of the dogwhelk Nucella lapillus. Evolutionary Ecology 23: 261-280.
Guo, F. 1998a. Origin and phylogeny of the Trigonioidoidea (Non-Marine Cretaceous Bivalves). In: Johnston, P.A. & Haggart J.W. (eds.). Bivalves: an eon of evolution - palaeobiological studies honoring Norman D. Newell. Calgary: University of Calgary Press. pp. 277-289.
Guo, F. 1998b. Sinonaiinae, a new Subfamily of Asian Non-Marine Cretaceous Bivalves. In: Johnston, P.A. & Haggart J.W. (eds.). Bivalves: an eon of evolution - palaeobiological studies honoring Norman D. Newell. Calgary: University of Calgary Press. pp. 291-294.
Haag, W.R. & Stanton, J.L. 2003. Variation in fecundity and other reproductive traits in freshwater mussels. Freshwater Biology 48: 2118-2130.

Bibliography
161
Haas, F. 1929. Die von der Zweiten Deutschen Zentral-Afrika-Expedition 1910-11 mitgebrachten Süßwassermuscheln. Senckenbergiana 11: 110-116.
Haas, F. 1969a. Superfamilia Unionacea. In: Mertens, R. & Henning W. (eds.). Das Tierreich (Berlin). Berlin: de Gruyter & Co. pp. 1-663.
Haas, F. 1969b. Superfamily Unionacea. In: Moore, R.C. (ed.) Treatise on invertebrate paleontology. Lawrence, Kansas: Geological Society of America & University of Kansas Press. pp. N411-N470.
Haas, F. & Schwarz, E. 1913. Die Unioniden des Gebietes zwischen Main und deutscher Donau in tiergeographischer Hinsicht. Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften, mathematisch-physikalische Klasse 26 1-34.
Haines, A.J. & Crampton, J.S. 2000. Improvements to the method of Fourier shape analysis as applied in morphometric studies. Palaeontology 43: 765-783.
Hamai, I. 2004. Sexuality of relative growth in the freshwater mussel, Inversidens japonensis (Lea). Science reports of Tohoku Imperial University (Biology) 167: 163-176.
Hammer, Ø. & Harper, D.A.T. 2006. PAST version 1.57. Available at http://folk.uio.no/ohammer/past/ (accessed April 2009).
Hammer, Ø., Harper, D.A.T. & Ryan, P.D. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologica Electronica 4: 9 pp.
Harper, E.M. 1997. The molluscan periostracum: an important constraint in bivalve evolution. Palaeontology 40: 71-97.
Haskin, H.H. 1954. Age determination in molluscs. Transactions of the New York Academy of Sciences 16: 300-304.
Hayashi, K. 1935. On the convexity of the shell of Anodonta. Venus 5: 23-25.
Hazay, J. 1881. Die Mollusken-Fauna von Budapest. Malakozoologische Blätter 4: 132-208.
Healy, J.M. 1989. Spermiogenesis and spermatozoa in the relict bivalve genus Neotrigonia: relevance to trigonioid relationships, particularly with Unionoidea. Marine Biology 103: 75-85.
Heard, W.H. 1975. Sexuality and other aspects of reproduction in Anodonta (Pelecypoda: Unionidae). Malacologia 15: 81-103.
Heard, W.H. & Guckert, R.H. 1970. A re-evaluation of the recent Unionadea (Pelecypoda) of North America. Malacologia 10: 333-355.
Hey, W.C. 1882. Freshwater mussels in the Ouse and the Fosse. Journal of Conchology 3: 263.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
162
Hinch, S.G. & Bailey, R.C. 1988. Within- and among-lake variation in shell morphology of the freshwater clam Elliptio complanata (Bivalvia: Unionidae) from south-central Ontario lakes. Hydrobiologia 157: 27-32.
Hinch, S.G., Bailey, R.C. & Green, R.H. 1986. Growth of Lampsilis radiata (Bivalvia: Unionidae) in sand and mud: a reciprocal transplant experiment. Canadian Journal of Fisheries and Aquatic Sciences 43: 548-552.
Hinch, S.G. & Green, R.H. 1989. The effects of source and destination on growth and metal uptake in freshwater clams reciprocally transplanted among south central Ontario lakes. Canadian Journal of Zoology 67: 855-863.
Hinch, S.G., Kelly, L.J. & Green, R.H. 1989. Morphological variation of Elliptio complanata (Bivalvia: Unionidae) in differing sediments of soft-water lakes exposed to acidic deposition. Canadian Journal of Zoology 67: 1895-1899.
Hochwald, S. 2001. Plasticity of life-history traits in Unio crassus. In: Bauer, G. & Wächtler K. (eds.). Ecology and Evolution of the Freshwater Mussels Unionoida. Berlin: Springer-Verlag. pp. 127-141.
Hoeh, W.R., Black, M.B., Gustafson, E., Bogan, A.E., Lutz, R.A. & Vrijenhoek, R.C. 1998. Testing alternative hypotheses of Neotrigonia (Bivalvia: Trigonioida) phylogenetic relationships using Cytochrome C Oxidase Subunit I DNA sequences. Malacologia 40: 267-278.
Hoeh, W.R., Bogan, A.E., Cummings, K.S. & Guttman, S.I. 2002. Evolutionary relationships among the higher taxa of freshwater mussels (Bivalvia: Unionoida): inferences on phylogeny and character evolution from analyses of DNA sequence data. Malacological Review 31/32: 117-141.
Hoeh, W.R., Bogan, A.E. & Heard, W.H. 2001. A phylogenetic perspective on the evolution of morphological and reproductive characteristics in the Unionoida. In: Bauer, G. & Wächtler K. (eds.). Ecology and Evolution of the Freshwater Mussels Unionoida. Berlin: Springer-Verlag. pp. 257-280.
Hoeh, W.R., Bogan, A.E., Heard, W.H. & Chapman, E.G. 2009. Palaeoheterodont phylogeny, character evolution, diversity and phylogenetic classification: a reflection on methods of analysis. Malacologia 51: 307-317.
Hoffman, J.I. & Amos, W. 2005. Microsatellite genotyping errors: detection approaches, common sources and consequences for paternal exclusion. Molecular Ecology 14: 599-612.
Hoffman, J.I., Peck, L.S., Hillyard, G., Zieritz, A. & Clark, M.S. 2010. No evidence for genetic differentiation between Antarctic limpet Nacella concinna morphotypes. Marine Biology 157: 765-778.
Hruska, J. 1999. Nahrungsansprueche der Flussperlmuschel und deren halbnatuerliche Aufzucht in der tschechischen Republik. Heldia 4: 69-79.
Huff, S.W., Campbell, D., Gustafson, D.L., Lydeard, C., Altaba, C.R. & Giribet, G. 2004. Investigations into the phylogenetic relationships of freshwater pearl mussels

Bibliography
163
(Bivalvia: Margaritiferidae) based on molecular data: implications for their taxonomy and biogeography. Journal of Molluscan Studies 70: 379-388.
Israel, F.B. 1910. Die Najadeen des Weidagebietes. Nachrichtenblatt der Deutschen Malakozoologischen Gesellschaft 42, Supplement 4: 49-58.
Jass, J. & Glenn, J. 2004. Sexual dimorphism in Lampsilis siliquoidea (Barnes, 1823) (Bivalvia: Unionidae). American Malacological Bulletin 18: 45-47.
Johannessen, O.H. 1973. Population structure and individual growth of Venerupis pullastra (Montagu) (Lamellibranchia). Sarsia 52: 97-116.
Johannesson, B. & Johannesson, K. 1996. Population differences in behaviour and morphology in the snail Littorina saxatilis: Phenotypic plasticity or genetic differentiation? Journal of Zoology 240: 475-493.
Johannesson, K. 2003. Evolution in Littorina: ecology matters. Journal of Sea Research 49: 107-117.
Jokela, J., Uotila, L. & Taskinen, J. 1993. Effects of the castrating trematode parasite Rhipidocotyle fennica on energy allocation of fresh-water clam Anodonta piscinalis. Functional Ecology 7: 332-338.
Källersjö, M., von Proschwitz, T., Lundberg, S., Eldenäs, P. & Erséus, C. 2005. Evaluation of IST rDNA as a complement to mitochondrial gene sequences for phylogenetic studies in freshwater mussels: an example using Unionidae from north-western Europe. Zoologica Scripta 34: 415-424.
Kat, P.W. 1984. Parasitism and the Unionacea (Bivalvia). Biological Reviews of the Cambridge Philosophical Society 59: 189-208.
Kauffman, E.G. 1969. Form, function and evolution. In: Moore, R.C. (ed.) Treatise on invertebrate paleontology. Part N. Mollusca 6. Bivalvia. Lawrence, Kansas: Geological Society of America & University of Kansas Press. pp. N129-205.
Kennard, A.L.S., Salisbury, A.E. & Woodward, B.B. 1925. Notes on the British Post-Pliocene Unionidae, with more especial regard to the means of identification of fossil fragments. Proceedings of the Malacological Society of London 16: 267-285.
Killeen, I., Aldridge, D.C. & Oliver, G. 2004. Freshwater Bivalves of Britain and Ireland. FSC Publications: Shrewsbury.
Kirtland, J.P. 1834. Observations on the sexual characteristics of the animals belonging to Lamarck's family of Naiades. American Journal of Science and Arts 26: 117-220.
Kitano, J., Mori, S. & Peichel, C.L. 2007. Sexual dimorphism in the external morphology of the threespine stickleback (Gasterosteus aculeatus). Copeia 2: 336-349.
Klocek, R., Bland, J. & Barghusen, L. 2010. A Field Guide to the Freshwater Mussels of Chicago Wilderness. Available at www.fieldmuseum.org/chicagoguides (accessed 2010).

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
164
Kohl, M. 2010. Freshwater Molluscan Shells. Available at http://mkohl1.net/FWshells.html (accessed 2010).
Kotrla, M.B. & James, C.F. 1987. Sexual dimorphism of shell shape and growth of Villosa villosa (Wright) and Elliptio icterina (Conrad) (Bivalvia: Unionidae). Journal of Molluscan Studies 53: 13-23.
Küster, H.C. 1848. Die Flussperlmuscheln (Unio et Hyria). Bauer und Raspe: Nürnberg.
Lea, I. 1870. A synopsis of the family Unionidae. Fourth Edition: Philadelphia, Pennsylvania.
Locard, A. 1890. Contributions á la faune malacologique française. 14. Catalogue des espėces françaises appartenant aix genres Pseudanodonta et Anodonta connues jusqu' á ce jour. Annales de la Société Linnéenne de Lyon 36: 50-285.
Loosanoff, V. 1936. Sexual phases in the quohog. Science 83: 287-288.
Luttikhuizen, P.C., Drent, J., Van Delden, W. & Piersma, T. 2003. Spatially structured genetic variation in a broadcast spawning bivalve: quantitative vs. molecular traits. Journal of Evolutionary Biology 16: 260–272.
Lydeard, C., Cowie, R.H., Ponder, W.F., Bogan, A.E., Bouchet, P., Clark, S.A., Cummings, K.S., Frest, T.J., Gargominy, O., Herbert, D.G., Hershler, R., Perez, K.E., Roth, B., Seddon, M., Strong, E.E. & Thompson, F.G. 2004. The global decline of nonmarine mollusks. BioScience 54: 321-330.
Lydeard, C., Mulvey, M. & Davis, G.M. 1996. Molecular systematics and evolution of reproductive traits of North American freshwater unionacean mussels (Mollusca: Bivalvia) as inferred from 16S rRNA gene sequences. Philosophical Transactions (Series B): Biological Sciences 351: 1593-1603.
Lynch, M. & Milligan, B.G. 1994. Analysis of population genetic structure with RAPD markers. Molecular Ecology 3: 91-99.
Machordom, A., Araujo, R., Erpenbeck, D. & Ramos, M.A. 2003. Phylogeography and conservation genetics of endangered European Margaritiferidae (Bivalvia: Unionoidea). Biological Journal of the Linnean Society 78: 235-252.
Maitland, P.S. & Campbell, R.N. 1992. Freshwater Fishes of the British Isles. Harper Collins: London.
Mann, K.H. 1965. Heated effluents and their effects on the invertebrate fauna of rivers. Proceedings of the Society for Water Treatment and Examination 14: 45-53.
Mansur, M.C.D. & Pereira, D. 2006. Limnic bivalves of the Sinos river basin, Rio Grande do Sul, Brazil (Bivalvia, Unionoida, Veneroida And Mytiloida). Revista Brasileira de Zoologia 23: 1123-1147.
Mansur, M.C.D. & Pimpão, D.M. 2008. Triplodon chodo, a new species of pearly fresh water mussel from the Amazon Basin (Mollusca : Bivalvia : Unionoida : Hyriidae). Revista Brasileira de Zoologia 25: 111-115.

Bibliography
165
March, M.C. 1910-1911. Studies in morphogenesis of certain Pelecypoda. Manchester Memoirs 4: 1-18.
Marie, B., Luquet, G., De Barros, J.-P.P., Guichard, N., Morel, S., Alcaraz, G., Bollache, L. & Marin, F. 2007. The shell matrix of the freshwater mussel Unio pictorum (Paleoheterodonta, Unionoida). FEBS Journal 274: 2933-2945.
Marshall, W.B. 1890. Beaks of Unionidae inhabiting the vicinity of Albany, N.Y. Bulletin of the New York State Museum 2: 169-189.
Marshall, W.B. 1926. Microscopic sculpture of pearly freshwater mussel shells. Proceedings of the United States National Museum 67: 1-14.
Matteson, M.R. 1960. Reconstruction of prehistoric environments through the analysis of molluscan collections from shell middens. American Antiquity 26: 117-120.
McIvor, A.L. & Aldridge, D.C. 2007. The reproductive biology of the depressed river mussel, Pseudanodonta complanata (Bivalvia: Unionidae), with implications for its conservation. Journal of Molluscan Studies 73: 259-266.
McMichael, D.F. & Hiscock, I.D. 1958. A monograph of freshwater mussels (Mollusca: Pelecypoda) of the Australian region. Australian Journal of Marine and Freshwater Research 9: 372-508.
Menard, H.W. & Boucot, A.J. 1951. Experiments on the movement of shells by water. American Journal of Science 249: 131-151.
Miller, N.J., Ciosi, M., Sappington, T.W., Ratcliffe, S.T., Spencer, J.L. & Guillemaud, T. 2007. Genome scan of Diabrotica virgifera virgifera for genetic variation associated with crop rotation tolerance. Journal of Applied Entomology 131: 378–385.
Mitchell, R.D. & Pitchford, G.W. 1953. On mites parasitizing Anodonta in England. Journal of Conchology 23: 365-370.
Modell, H. 1942. Das natürliche System der Najaden. Archiv für Molluskenkunde 74: 161-191.
Modell, H. 1949. Das natürliche System der Najaden. 2. Archiv für Molluskenkunde 78: 29-48.
Modell, H. 1957. Die fossilen Najaden Nordamerikas. Archiv für Molluskenkunde 86: 183-200.
Modell, H. 1964. Das natürliche System der Najaden. 3. Archiv für Molluskenkunde 93: 71-126.
Molina, R.A. 2004. Morphological and genetic description of the freshwater mussel, Elliptio complanata (Lightfoot, 1786) in the Cape Fear River system, NC. PhD thesis, North Carolina State University

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
166
Morey, D.F. & Crothers, G.M. 1998. Clearing up clouded waters: Palaeoenvironmental analysis of freshwater mussel assemblages from the Green River shell middens, western Kentucky. Journal of Archaeological Science 25: 907-926.
Morton, B. 1976. Secondary brooding of temporary dwarf males in Ephippodonta (Ephippodontina) oedipus sp. nov. (Bivalvia: Leptonacea). Journal of Conchology 29: 31-39.
Mueller, U.G. & Wolfenbarger, L.L. 1999. AFLP genotyping and fingerprinting. Trends in Ecology and Evolution 14: 389-394.
Mulvey, M., Lydeard, C., Pyer, D.L., Hicks, K.M., Brim-Box, J., Williams, J.D. & Butler, R.S. 1997. Conservation genetics of North American freshwater mussels Amblema and Megalonaias. Conservation Biology 11: 868-878.
Mutvei, H. & Westermark, T. 2001. How environmental information can be obtained from naiad shells. In: Bauer, G. & Wächtler K. (eds.). Ecology and Evolution of the Freshwater Mussels Unionoida. Berlin: Springer-Verlag. pp. 367-379.
Nagel, K.O. 2000. Testing hypotheses on the dispersal and evolutionary history of freshwater mussels (Mollusca: Bivalvia: Unionidae). Journal of Evolutionary Biology 13: 854-865.
Negus, C.L. 1966. A quantitative study of growth and reproduction of unionid mussels in the River Thames at Reading. Journal of Animal Ecology 35: 513-532.
Neves, R.J., Bogan, A.E., Williams, J.D., Ahlstedt, S.A. & Hartfield, P.D. 1997. Status of aquatic mollusks in the southeastern United States: a downward spiral of diversity. In: Benz, G.W. & Collins D.E. (eds.). Aquatic Fauna in Peril: The Southeastern Perspective. Special Publication No. 1. Decatur, GA: Southeast Aquatic Research Institute, Lenz Design and Communications. pp. 43–86.
Newell, N.D. & Boyd, D.W. 1975. Parallel evolution in early trigonioidean bivalves. Bulletin of the American Museum of Natural History 154: 55-162.
Noll, F.C. 1869. Unsere Flußmuscheln (Najaden). Ihre Entwicklung und ihre Beziehungen zur übrigen Thierwelt. Bericht über die Senckenbergische naturforschende Gesellschaft in Frankfurt am Main 70: 33-44.
Ohba, S. 1959. Ecological studies in the natural population of a clam, Tapes japonica, with special reference to seasonal variations in the size and structure of the population and to individual growth. Biological Journal of Okayama University 5: 13-42.
Ortmann, A.E. 1912. Notes upon the families and genera of the Najades. Annals of the Carnegie Museum 8: 222-364.
Ortmann, A.E. 1920. Correlation of shape and station in freshwater mussels (Naiades). Proceedings of the American Philosophical Society 59: 268-312.
Owens, I.P.F. & Hartley, I.R. 1998. Sexual dimorphism in birds: why are there so many different forms of dimorphism? Proceedings of the Royal Society B-Biological Sciences 265: 397-407.

Bibliography
167
Panova, M., Hollander, J. & Johannesson, K. 2006. Site-specific genetic divergence in parallel hybrid zones suggests nonallopatric evolution of reproductive barriers. Molecular Ecology 15: 4021-4031.
Parker, G.A. 1992. The evolution of sexual size dimorphism in fish. Journal of Fish Biology, Supplement B 41: 1-20.
Parmalee, P.W. & Bogan, A.E. 1998. The freshwater mussels of Tennessee. The University of Tennessee Press: Knoxville. 328 pp.
Parodiz, J.J. & Bonetto, A.A. 1963. Taxonomy and zoogeographic relationships of the South American naiades (Pelecypoda: Unionacea and Mutelacea). Malacologia 1: 179-213.
Pilsbry, H.A. & Bequaert, J.C. 1927. The aquatic mollusks of the Belgian Congo: with a geographical and ecological account of Congo malacology. Bulletin of the American Museum of Natural History 53: 69-602.
Pimpão, D.M., Rocha, M.S. & de Castro Fettuccia, D. 2008. Freshwater mussels of Catalão, confluence of Solimões and Negro rivers, state of Amazonas, Brazil. Check List 4: 395–400.
Playford, T.J. & Walker, K.F. 2008. Status of the endangered Glenelg River Mussel Hyridella glenelgensis (Unionoida: Hyrudae) in Australia. Aquatic Conservation-Marine and Freshwater Ecosystems 18: 679-691.
Polisky, B., Greene, P., Garfin, D.E., McCarthy, B.J., Goodman, H.M. & Boyer, H.W. 1975. Specificity of substrate recognition by the EcoRI restriction endonuclease. Proceedings of the National Academy of Sciences of the United States of America 72: 3310-3314.
Ponder, W.E. & Bayer, M. 2004. A new species of Lortiella (Mollusca : Bivalvia : Unionoidea : Hyriidae) from northern Australia. Molluscan Research 24: 89-102.
Prashad, B. 1931. Some noteworthy examples of parallel evolution in the molluscan faunas of South-eastern Asia and South America. Proceedings of the Royal Society of Edinburgh 51: 42-53.
Radley, J.D. & Barker, M.J. 1998. Palaeoenvironmental analysis of shell beds in the Wealden Group (Lower Cretaceous) of the Isle of Wight, southern England: an initial account. Cretaceous Research 19: 489-504.
Rasband, W. 2008. ImageJ. Image processing and analysis in Java. Available at http://rsbweb.nih.gov/ij (accessed April 2009).
Reeside, J.B. 1927. Two new unionid pelecyopds from the Upper Triassic. Journal of the Washington Academy of Science 17: 476-478.
Reigle, N.J. 1967. An occurrence of Anodonta (Mollusca, Pelecypoda) in deep water. American Midland Naturalist 78: 530-531.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
168
Rodhouse, P.G. 1977. An improved method for measuring volume of bivalves. Aquaculture 11: 279-280.
Roe, K.J. & Hoeh, W.R. 2003. Systematics of freshwater mussels (Bivalvia: Unionoida). In: Lydeard, C. & Lindberg D. (eds.). Molecular Systematics and Phylogeography of Mollusks. Washington: Smithsonian Inst. Press. pp. 91-122.
Roper, D.S. & Hickey, C.W. 1994. Population structure, shell morphology, age and condition of the freshwater mussel Hyridella menziesi (Unionacea: Hyriidae) from seven lake and river sites in the Waikato River system. Hydrobiologia 284: 205-217.
Rossmässler, E.A. 1835-1837. Iconographie der Land- und Süßwasser-Mollusken mit vorzüglicher Berücksichtigung der europäischen noch nicht abgebildeten Arten. Arnoldische Buchhandlung: Dresden und Leipzig.
Saarinen, M. & Taskinen, J. 2004. Aspects of the ecology and natural history of Paraergasilus rylovi (Copepoda, Ergasilidae) parasitic in unionids of Finland. Journal of Parasitology 90: 948-952.
Saleuddin, A.S.M. 1965. The gonads and reproductive cycle of Astarte sulcata (Da Costa) and sexuality in A. elliptica (Brown). Proceedings of the Malacological Society of London 36: 229-257.
Savazzi, E. & Peiyi, Y. 1992. Some morphological adaptations in freshwater bivalves. Lethaia 25: 195-209.
Schneider, J.A. & Carter, J.G. 2001. Evolution and phylogenetic significance of cardoidean shell microstructure (Mollusca, Bivalvia). Journal of Paleontology 75: 607–643.
Scholz, H. 2003. Taxonomy, ecology, ecomorphology, and morphodynamics of the Unionoida (Bivalvia) of Lake Malawi (East-Africa). Beringeria 33: 1-86.
Scholz, H. & Glaubrecht, M. 2004. Evaluating limnic diversity: Toward a revision of the unionid bivalve Coelatura Conrad, 1853 in the Great Lakes of East Africa and adjacent drainage systems (Mollusca, Bivalvia, Unionidae). Mitteilungen des Museums für Naturkunde in Berlin, Zoologische Reihe 80: 89-121.
Scholz, H. & Hartmann, J.H. 2007a. Fourier analysis and the extinction of unionoid bivalves near the Cretaceous-Tertiary boundary of the Western Interior, USA: Pattern, causes, and ecological significance. Palaeogeography, Palaeoclimatology, Palaeoecology 255: 48-63.
Scholz, H. & Hartmann, J.H. 2007b. Paleoenvironmental reconstruction of the Upper Cretaceous Hell Creek formation of the Williston Basin, Montana, USA: Implications from the quantitative analysis of Unionoid bivalve taxonomic diversity and morphologic disparity. Palaios 22: 24-34.
Scholz, H., Tietz, O. & Büchner, J. 2007. Unionoid bivalves from the Miocene of Berzdorf (eastern Germany): taxonomic remarks and implications for palaeoecology and

Bibliography
169
palaeoclimatology. Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen 244: 43-51.
Seed, R. 1980. Shell growth and form in the Bivalvia. In: Rhoads, D.C. & Lutz R.A. (eds.). Skeletal Growth of Aquatic Organisms. New York: Plenum Press. pp. 23-67.
Sell, H. 1907-1908. Biologische Beobachtungen an Najaden. Archiv für Hydrobiologie 3: 179-180.
Serb, J.M., Buhay, J.E. & Lydeard, C. 2003. Molecular systematics of the North American freshwater bivalve genus Quadrula (Unionidae: Ambleminae) based on mitochondrial ND1 sequences. Molecular Phylogenetics and Evolution 28: 1-11.
Sha, J.G. 1993. Trigonioides from early Cretaceous Hekou Formation of Hekou Basin, Ninghua, Fujian, China, with special remarks on classification of Trigonioididae. Acta Palaeontologica Sinica 32: 285-302.
Sha, J.G. 2006. Ontogenetic variations of the Early Cretaceous non-marine bivalve Trigonioides (T.) heilongjiangensis and their phylogenetic significance. In Malchus, N. & Pons J.M., (eds.). Organic Diversity and Evolution Supplement. Abstracts and Posters of the “International Congress on Bivalvia” at the Universitat Autònoma de Barcelona, Spain, 22-27 July 2006, 69-70.
Sha, J.G. 2007. Cretaceous trigonioidid (non-marine Bivalvia) assemblages and biostratigraphy in Asia with special remarks on the classification of Trigonioidacea. Journal of Asian Earth Sciences 29: 62-83.
Shine, R. 1989. Ecological causes for the evolution of sexual dimorphism: a review of the evidence. The Quarterly Review of Biology 64: 419-461.
Siebold, C.T.v. 1837. Über den Unterschied der Schalenbildung der männlichen und weiblichen Anodonten. Archiv für Naturgeschichte 3: 415-416.
Silverman, H., Kays, W.T. & Dietz, T.H. 1987. Maternal calcium contribution to glochidial shells in freshwater mussels (Eulamellibranchia: Unionidae). Journal of Experimental Zoology 242: 137-146.
Silverman, H., Steffens, W.L. & Dietz, T.H. 1985. Calcium from extracellular concretions in the gills of freshwater unionid mussels is mobilized during reproduction. Journal of Experimental Zoology 236: 137-147.
Simpson, C.T. 1896. Description of four new Triassic Unios from the Staked Plains of Texas. Proceedings of the United States National Museum 18: 381-385.
Simpson, C.T. 1900. Synopsis of the naiades, or pearly fresh-water mussels. Proceedings of the United States National Museum 22: 501-872.
Simpson, C.T. 1914. A Descriptive Catalogue of the Naiads, or Pearly Fresh-water Mussels. Part I-III. Bryant Walker: Detroit, Michigan.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
170
Soroka, M. & Zdanowski, B. 2001. Morphological and genetic variability of the population of Anodonta woodiana (Lea, 1834) occurring in the heated Konin lakes system. Archives of Polish Fisheries 9: 239-252.
Stanley, S.M. 1970. Relation of shell form to life habits in the Bivalvia (Mollusca). Memoirs of the Geological Society of America 125: 1-296.
Stearns, S.C. 1989. The evolutionary significance of phenotypic plasticity. BioScience 39: 436-445.
Sterki, V. 1903. Notes on the Unionidae with their classification. American Naturalist 37: 103-113.
Stillwell, R.C. & Fox, C.W. 2007. Environmental effects on sexual size dimorphism of a seed-feeding beetle. Oecologia 153: 273-280.
Stone, N.M., Earll, R., Hodgson, A., Mather, J.G., Parker, J. & Woodward, F.R. 1982. The distributions of three sympatric mussel species (Bivalvia: Unionidae) in Budworth Mere, Cheshire. Journal of Molluscan Studies 48: 266-274.
Strayer, D.L. 2006. Challenges for freshwater invertebrate conservation. Journal of the North American Benthological Society 25: 271-287.
Tankersley, R.A. & Dimock, R.V., JR. 1992. Quantitative analysis of the structure and function of the marsupial gills of the freshwater mussel Anodonta cataracta. Biological Bulletin 182: 145-154.
Taskinen, J. 1998. Influence of trematode parasitism on the growth of a bivalve host in the field. International Journal for Parasitology 28: 599-602.
Taskinen, J. & Valtonen, E.T. 1995. Age-, size- and, sex-specific infection of Anodonta piscinalis (Bivalvia: Unionidae) with Rhipidocotyle fennica (Digenea: Bucephalidae) and its influence on host reproduction. Canadian Journal of Zoology 73: 887-897.
Taylor, J.D. & Kennedy, W.J. 1969. The influence of the periostracum on the shell structure of bivalve molluscs. Calcified Tissue Research 3: 274-283.
Taylor, J.D., Kennedy, W.J. & Hall, A. 1969. The shell structure and mineralogy of the Bivalvia. Introduction. Nuculacea-Trigonacea. Bulletin of the British Museum (Natural History), Zoology. Supplement 3: 1-125.
Tevesz, M.J.S. & Carter, J.G. 1980. Environmental relationships of shell form and structure in Unionacean bivalves. In: Rhoads, D.C. & Lutz. R.A. (eds.). Skeletal Growth of Aquatic Organisms. New York: Plenum Press. pp. 295-322.
Thom, M.D., Harrington, L.A. & Macdonald, D.W. 2004. Why are American mink sexually dimorphic? A role for niche separation. Oikos 105: 525-535.
Tregenza, T. & Butlin, R.K. 1999. Speciation without isolation. Nature 400: 311-312.

Bibliography
171
Trueman, E.R. 1966a. Bivalve mollusks: Fluid dynamics of burrowing. Science 152: 523-525.
Trueman, E.R. 1966b. The fluid dynamics of the bivalve molluscs Mya and Margaritifera. Journal of Experimental Biology 45: 369-382.
Trueman, E.R. 1968. The locomotion of the freshwater clam Margaritifera margaritifera (Unionacea: Margaritiferidae). Malacologia 6: 401-410.
Tudorache, C., Viaene, P., Blust, R., Vereecken, H. & De Boeck, G. 2008. A comparison of swimming capacity and energy use in seven European freshwater fish species. Ecology of Freshwater Fish 17: 284-291.
Tudorancea, C. 1972. Studies on Unionidae populations from the Crapina-Jijila complex of pools (Danube zone liable to indulation). Hydrobiologia 39: 527-561.
Vaughn, C.C. & Hakenkamp, C.C. 2001. The functional role of burrowing bivalves in freshwater ecosystems. Freshwater Biology 46: 1431-1446.
Vekemans, X. 2002. AFLP-SURV version 1.0. Distributed by the author. Bruxelles, Belgium: Laboratoire de Génétique et Ecologie Végétale, Université Libre de Bruxelles.
Vermeij, G.J. 1973. Morphological patterns in high-intertidal gastropods - adaptive strategies and their limitations. Marine Biology 20: 319-346.
Vermeij, G.J. 1993. A natural history of shells. Princeton University Press: Princeton, NJ. 216 pp.
Via, S., Gomulkiewicz, R., Scheiner, S.M., Schlichting, C.D. & Van Tienderen, P.H. 1995. Adaptive phenotypic plasticity: consensus and controversy. Trends in Ecology and Evolution 10: 212-217.
von Bertalanffy, L. 1938. A quantitative theory of organic growth. Human Biology 10: 181-213.
von Martens, E. 1897. Beschalte Weichthiere Deutsch-Ost-Afrikas. Dietrich Reimer (Ernst Vohsen): Berlin.
Vos, P., Hogers, R., Bleeker, M., Reijans, M., van de Lee, T., Hornes, M., Frijters, A., Pot, J., Peleman, J. & Kuiper, M. 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Research 23: 4407-4414.
Wächtler, K., Mansur, M.C.D. & Richter, T. 2001. Larval types and early postlarval biology in naiads (Unionoida). In: Bauer, G. & Wächtler K. (eds.). Ecology and Evolution of the Freshwater Mussels Unionoida. Berlin: Springer-Verlag. pp. 93-125.
Walford, L.A. 1946. A new graphic method of describing the growth of animals. Biological Bulletin 90: 141-147.

Variability, function and phylogenetic significance of unionoid shell characters A. Zieritz
172
Walker, J.M., Curole, J.P., Wade, D.E., Chapman, E.G., Bogan, A.E., Watters, G.T. & Hoeh, W.R. 2006. Taxonomic distribution and phylogenetic utility of gender-associated mitochondrial genomes in the Unionoida (Bivalvia). Malacologia 48: 265-282.
Wanner, H.E. 1921. Some faunal remains from the Trias of York County, Pennsylvania. Proceedings of the Academy of Natural Sciences of Philadelphia 73: 25-37.
Watters, G.T. 1994. Form and function of unionoidean shell sculpture and shape (Bivalvia). American Malacological Bulletin 11: 1-20.
Watters, G.T. 2001. The evolution of the Unionacea in North America, and its implications for the worldwide fauna. In: Bauer, G. & Wächtler K. (eds.). Ecology and Evolution of the Freshwater Mussels Unionoida. Berlin: Springer-Verlag. pp. 281-307.
Watters, G.T., Hoggarth, M.A. & Stansbery, D.H. 2009. The Freshwater Mussels of Ohio. The Ohio State University Press: Columbus.
Watts, P.C. 2001. Extraction of DNA from tissue: High salt method. Available at http://www.genomics.liv.ac.uk/animal/RESEARCH/ISOLATIO.PDF (accessed April 2009).
Weir, B.S. & Cockerham, C.C. 1984. Estimating F-statistics for the analysis of population structure. Evolution 38: 1358–1370.
Weisensee, H. 1916. Die Geschlechtsverhältnisse und der Geschlechtsapparat bei Anodonta. Zeitschrift für wissenschaftliche Zoologie 115: 262-335.
Wetherby, A.G. 1882. On the geographical distribution of certain fresh-water mollusks of North America and the probable causes of their variation. American Journal of Science 23 (3rd series): 203-212.
Wilbur, K.M. & Saleuddin, A.S.M. 1983. Shell formation. In: Saleuddin, A.S.M. & Wilbur K.M. (eds.). The Mollusca, Volume 4: Physiology. London and New York: Academic Press. pp. 235-287.
Williams, J.D., Warren, M.L., Cummings, K.S., Harris, J.L. & Neves, R.J. 1993. Conservation status of freshwater mussels of the United States and Canada. Fisheries 18: 6-22.
Wood, E.M. 1974. Development and morphology of the glochidium larva of Anodonta cygnea (Mollusca: Bivalvia). Journal of Zoology 173: 1-14.
Yeager, M.M., Cherry, D.S. & Neves, R.J. 1994. Feeding and burrowing behaviors of juvenile rainbow mussels, Villosa iris (Bivalvia, Unionidae). Journal of the North American Benthological Society 13: 217-222.
Yeap, K.L., Black, R. & Johnson, M.S. 2001. The complexity of phenotypic plasticity in the intertidal snail Nodilittorina australis. Biological Journal of the Linnean Society 72: 63-76.

Bibliography
173
Yonge, C.M. 1962. On Etheria elliptica Lam. and the course of evolution, including assumption of monomyrianism, in the family Etheriidae (Bivalvia: Unionacea). Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 244: 423-458.
Zhivotovsky, L.A. 1999. Estimating population structure in diploids with multilocus dominant DNA markers. Molecular Ecology 8: 907–913.
Zieritz, A. & Aldridge, D.C. 2009. Identification of ecophenotypic trends within three European freshwater mussel species (Bivalvia: Unionoida) using traditional and modern morphometric techniques. Biological Journal of the Linnean Society 98: 814–825.
Zieritz, A. & Aldridge, D.C. in review. Sexual, habitat-constrained and parasite-induced dimorphism in the shell of a freshwater mussel (Anodonta anatina, Unionidae).
Zieritz, A., Bogan, A.E. & Aldridge, D.C. in review. Variability and a new model for character evolution of umbonal sculptures in the Unionoida.
Zieritz, A., Checa, A.G., Aldridge, D.C. & Harper, E.M. in press. Variability, function and phylogenetic significance of periostracal microprojections in unionoid bivalves. Journal of Zoological Systematics and Evolutionary Research.
Zieritz, A., Hoffman, J.I., Amos, W. & Aldridge, D.C. 2010. Phenotypic plasticity and genetic isolation-by-distance in the freshwater mussel Unio pictorum (Mollusca: Unionoida). Evolutionary Ecology 24: 923-938.




















