
ýýýý .ý ,-ýi ý: - QMRO Home
246
/- i ýýýý .ý ,-ýi ý: rtpia, IZI- r r`. tJIL Q C. YCIJ. 3, OF Ok, ýW" fý°' T II Alv Yý"'ý rte r" OF ;'., w,. ýJ FtC.: )i., CLJ. x)11 OF PL . t_ýE'. i. )ýýIC BT, ý. ý.; _; --GI{E_: ý1 X1: 1 Gi IL sei º` 1' LOPI. M iiÄ1+: Rý. ý$ 0 . 4. thesis -muhmitted by J'ULCTIIT"1 . AUGJi392 for the degree of Ph. D. in the University of London. Department of Bo ta i-y anti 13iýci_erzi., tr; 7 ýteýt*"ý_eld Col. ee, I1o2. d. ýri . June 1977 U'IV. LONDON 0ýi Ot LS- ßp f--R- I
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of ýýýý .ý ,-ýi ý: - QMRO Home

/-
i
ýýýý .ý
,-ýi ý:
rtpia, IZI- r r`. tJIL Q C. YCIJ. 3, OF Ok, ýW"
fý°' T II Alv Yý"'ý rte r" OF ;'., w,. ýJ FtC.: )i., CLJ. x)11 OF
PL . t_ýE'. i. )ýýIC BT, ý. ý.; _; --GI{E_: ý1 X1: 1 Gi IL sei º` 1' LOPI. M iiÄ1+: Rý. ý$
0
. 4. thesis -muhmitted by
J'ULCTIIT"1 . AUGJi392
for the degree of Ph. D. in the University
of London.
Department of Bo ta i-y anti 13iýci_erzi., tr; 7 ýteýt*"ý_eld
Col. ee, I1o2. d. ýri .
June 1977
U'IV.
LONDON 0ýi Ot LS-
ßp f--R- I

14T
The p'i ica1 end, chomical factors ti hick affect
sporu at oil, and tu e ý-ro 'th and. eur\rivai of vegetative
populations of blue-, green algae were examined in two
eutrophic kataglacial lakes of the Ellesmere (Worth Salop)
group of meres. "A'he im . aortance of individual factors were
examined under controlled conditions using natural
populations or uni-algal isolates from the mores.
The induction of akinete formation, which, in all the
cases examined, proceeds once the elga f. ori: is a surface bloom,
is thought to be due to the extreme conditions, iparticularly
of solar radiation, which prevail at the surface during; the
summer. Winter blooms did not sporuj. ate. Nutrient
deficiency, particularly of orthophosphate, is not
considered to be a critical factor. This is supported by
the finding that blooming ulgal material is not depleted of
phoriphorue, that alkaline phospheta, e activity was low, and
that nutrient doficicncy under controlled conditions did
not induce sporulation of the species pre6 omi. nc±nt in the
Mores.
Nitrogenase activity (acetylene reduction) and the
photosynthetic rate (oxygen evolutiozi) were observed to
fall in the period subsequent to bloom formation. The
reduction of rnoaaured not photosynthesis may be correlated
with the increase of bacterial numbers. The activity of
light-damaged material was lower than that of unaffected
populations. It appears that there must be a low cellular
carbon to nitrogen ratio before akinete differentiation
xithetic can begin, indicating that a reduction of photos-y
activity relative to that of nitrogon assimilation is
r. equiýred . Akinete:: probably do not form a significant over-
wintoring r: iechanism in the lakes investigated. Numbers of
ak. inetes found during the winter were low. The bloom
PPO:; t:. _ations prob>_bly develop frort growth of the over-
wintering planktonic vegetative popul. ationa during the
"sprina- and early E3uarier. Cornparisoni3 of planktonic,
sedirmentinf; and benthic algal material indicates that

-3- germination of akinetes ohortly after their maturation
may give rise to the greater part of the ov erwinteiiing
vegetative populutioni.
Physical rather than chemical faetoro appear to
control the population sizes and 11Th cycles in the
enviroiirnents ; studied.

Iý" tw etw ". . rý1}spy. .
. i(. I. ýýtI. 1ý. j. iýý... i....,.. 3
X trinh to a. j: roau r3' nincoro thºjrikti to Dr 3? utox Tny
Par hits r; uitlana0 and oolntinuouti unaourni(. anont throughout
this study.
:u cry aoUunglioU at Waatfiold i: o: +, 1o o, London, IA
nlno erntotul :. ̀or diuounnion and help. In ; ptirtiouln I
would li3cu to than}: Brondu Thur'ro, Itr It. Plitoliall nncl 'Dr 11. hvnnr.
Vor nn iaitroduation to thcc : 3, cloppicm ruircaa, rind or
mcny holp ul owmrýc ntaý nrua tt ', r.. : tons ý ý. wa inc1tbted to
1)r t:. 13. Itayaio1d&.
In ithlttiuix, I rin inclobtocl to t ho :. 'rýictkooti or tho
Ororvonor l: ntatuo for pormintiion to work tit Lthito Morn, to
Nr L, It. Jobb for allowing, no0an0 to thu oliorcº and for
utorcaco fuuilit . ua, t, nd to Hr it. I:. Pininwcirinr sind ?! t Jor
K. U. 1'ottorcircco £or purrtiuuicm to work nt F: uttlo Moro,
B1t ku I uru and Newton r. oru. I nm ciltan grrntofui to thu
Prunhw: ttor Iiiologicn1 Aonoointion for uoo of thu Moran
Laborittory it Truoton Pýontfoxd Viuld Cuntru und to
1)r A. Ilrtyluy, thu itnrdun tit 3'r,, titon iºont1'orct.
Thin Fork hers buoix oup1 ortocl by it i; rtint from tho
Itetturni J: nvi, onriont 1(tn tj, trah Council, whocio nncintlinao
.ýj ctttýSýz. i.? nckrou?. odj; o.

('t' 'rr J.. S91 .
Page
....... Otto .................... ' ..... 2
ACr:; IOi; 7ýI: ýGEI ::::; ä ...........:.................:..... 4 JOIST U. a. ' 1'AD7:, I: 5 ........:............................ g
.................................... 12
Tlil: FIELD DATA
Introduction.... ' ....... ............................ 15 . The Mer. er ............. .................:..:........ `. 24 Licit of : ir)ecies Encountered in the tier. ee During the 29 ', er. iod of thý. s nve ßtß. rr . tion... ...................: . Zoonlankton ........................................ 31 The äitec Znyea:; tir-nted
l: ettlo Aiero .............................. 32
White 11er. e ............................... 32
23lake here and Newton ilex© :.............. 33 fateritis and P"Tc iiio(. f3
Field Sampling
k'Tankton..... ".,.. ." ."............... 36"
De thor.... ".......... "......... ". "" 36
äedimentation ....................... 39
The Enumeration of Field Material
Planktonic .... """"""".... """""...... 42
Benthic ................... "....... "" 44 Total particulate sedimentation ..... 45 sedimentation of algal material..... 45
Th e Chemical Ant. lyeis of Field Jctmples
Orthophosphate ................. **0*** 47 2litVate 1.11 d 11itritc7) " ....... " 48 ....... . UXgE)ll»..... ". ". "",.. ". "......... " 51
.......... p11
...... .. . 52 . ............. .
....... .. ". ""..: Conductivity 53 ..... .. " The Iie; -zsu2i o naht of l'Iiysical_ Par. Fir. i tero
J tý : E. pe 2' ilttLre. "..... . .... .".. . ..... """ 53 l+ i g- lit ...... .... ". """... ". ".. " ......
53
Total particulate euz3lperndcd solids.. 53

-6-
i
Re su? _t. s
The: Cheriica and 21r. oical l nvironrient `temperature .........................
Lieht ............................... Orthophosphate ...................... I'; itrat e ........ " ..................... .
T! ie Annual Cycle of Growth arwd Succession
of the Blue-green Algae
The plankton ....... ................. Sedinentation.......................
)enthic populations ................. Dismission
The Growth and Abundance of Planktonic
Populations .., ............................ Sporu]ation and Germination........... '... Ov rw i nt or ing
""""""""""" . 000000 """"""""""
JLC ION 2
GROWTH AND CELL I)IPIJREIITIATZUTN UNDER A VARIETY OF
PHY ICAL AND CHL'I. ZCAL GO DITzozrs I21tý"pduct?
_021. .... "..... """""".. "...................
I1 ter. ia? _s and P"'ethodB
Isolation ............................... Culture D: edict ............................ Preparation of Utock-. 'jjolutionr, and Media.
The Culture of ýil gae """""""""""""""""""""
3ui±ors..... """"""""""""""""""""""""". """
L .L ht
High intensity ....................
Nonochroriatic, action spectra.......
Ultra-violet ........................
Asse crent of Al al Materiale """""
F1 E: ül I ! -, -E;
tTI"Oý'7 h, """""""""""""""""""""". ".
Page
54 54 65
71 71
71
83 95
104
110
115
121
122
125
125
1 27
128
129
129
130
132
134
135
136
1DI 7

-7--
Page
iTutrients and `or. ulatioii ................ 139
3ýz lit Quantity and $porulation'........... 147
Light Quality and üporulation............ 143
Ultra-violet Light and Sporulation....... 149
Diseus:, ion
Growth and Sporulation .................... 152
'Inorganic Nutrients and 2poruilation...... 153
Other Compounds and Sporulation........ .. 155
Light Quantity and Sporu? ration........... 156
Light Quality and Sporulation.... ........ 157
" Ultra-violet Light and. : 3porulation....... 158
SBCÄICPJ 3 T1ii 7rHY3IOLC(fl CAL ACTIVITY LP IU TUI: A,, L POPULATIONS
OF ALGAL Introductic, n. "... """"".. ".... """" """""""""""""" """ 160
Materials RflcI. Methods
Collection of 'Material and Preliminary 7. reEitrliP. llt """"""""""""""""""""""""""""""""
165.
Alkal. ne Phosphatage..................... 165
Labile Orthoj)hosphato.................... 167
Nitrogena5e (Acetylene fleduction)........ 169
Photosynthesis (Oxy-en Production)....... 170
Carbon, Hydrogen and Nitrogen Analysis... 171
Assessment of Bacterial Plumbers in Algal
Samples .... ..... 171
Results
Yho phorus Availability .................. 172 NitroL ezi Metabolism...... .... ............ 177 Oxygen Production.... 09... ............... 185
Discussion
Yhosphorus ............................... 192
Nitrogen and Carbon ...................... 193
Change of Tletaboliom and üporulatior_..... 198

Page
............... ......:.............. 204 t] rý"-l :; T` trrcr
" 1i. L' : ý. rte. 4L 1ý a3 """"""""""""""": """"""""""""""""". """". "" 213
11
Reprint of published paper bound in at back: -
Rather, J. A. and Fay, P-0977) Oporu? ation and the
developne: nt of planktonic blue-green algae in two
Jalopian rneres. Yroc. R. Soc. ?. rond. B. 195,317 - 332.
i

LL3 Ute' 1ABIL J
; ýo . 1'ago, Title
1 28 Uhan ; es of nutrient concentration of surface
samples from the merest
2 66 Depth (m) of 10;; and 1 of sub-surface radiation.
Concentration of orthophosphate-in the terse.
3 6ü Kettle I: ere and White I-Tere, 1975.
4 69 Kettle Mere and White Piere, 1976.
5 70 Concentration' of nitrate in the mores (Kettle
Tiere and White Mere: ) , 1975 and 1976.
6 71 Conductivity, transpar9ncy and pH of the surface
waters of the mores, 1976.
Amount of algal material in the* plankton.
,7 73 Kettle Here, 1 974 "
8 74 White Mere, ' 197 . 9 75 Kettle Tiere, 1975. -
10 76 White More, 1975.
11 . 77 Kettle Mere, 1976"
12 78 White Mere, 1976.
Akinote percentage of algae from the meres.
13. ß0 Kettle Mere and White Mere, 1974.
14 81 Kettle Mere and White Mere, 1975.
15 82 Kottle Tiere and ýIhite : lore, 1976. *
Integrated algal standing crop.
16 105 Kettle bier. e, 1974. - 17 106 White Piere, 1974.
18 107 Kettle Tiere and White Fiere, 1976.
: IECTýUI3 2
19 124 List of the species obtained and maintained in
culture, their source and method of isolation.
20 126 Cocaposition of ASI'I 4 medium. 21 136 Growth rates of isolates of AI)h. flus-mono.
22 138 :. 'f3 ect of iI11ji'E. i buffer on the gro%rth of
planktonic t3.1 Etl isolates,

-10-
To o. Page itlc y
23 140 Effect of orthophosphate, and nitx'ate on cell
differentiation. of a White I: ere strain of AT)h. f1 or, --n2uao.
24 141 Effect of orthophosphate and nitrate on cell
differentiation of a I: ettle Here otrain of
Aph. flos-ncruae.
25 143 Effect of HI: PES buffer on heterocyst frequency.
26 144 Effect of I3I: YES buffer on sporulation. 27 146 Nporulation of . 4nabaena sp. ' (? ) under different
nutrient conditions.
28 149 Effect of u. v. light-'on algal growth.
29 '150 Lfi'ect of short u. v. doses on the sporulation of
! Dh. flos-HQUHO isolates from Kot-', -, le Mere and
White Mere. '
30 151 Effect of u. v., and fluorescent light inteneity,
on the sporulation of A. flo s-tequoe collected
from MJhi to More, spring 1976.
SECTION 3
Labile orthophosphate (boiling water extracts). 31 173 Kettle Mere, 1975 and 1976.
32 174 White Mere, 1975 and 1976.
3; 175 Newton Mere, 1975 and 1976.
34 176 Alkaline phoophataEe activity of algae from the
raeres, 1975 and 1976.
Nitrogenase activity (ac. etylene reduction).
35 173 Kettle Mere , 1976.
36 179 White Mere, 1976. 37 1F30 No'iton here , 1976.
Het er. ocyst frequency of algae from the rneres. 38 181 Kettle Here and White Here, 1974.
39 182 Kettle Mere and White , "fiere, 1975. 40 183 Kettle here and shite Mere, 1976.
41 184 Carbon : nitrogen : hydrogen ratios of bloo-i-
forr_iing algae, 1976.

- 11 -
No. Pago Title " Oxygen production.
42 l a6 Fettle 1 976 . 43 1 F37 : ihitc T". ore, 1976.
. 44 1813 Newton Mere, 1976.
45 190 Bacterial numbers in algal samples, 1976.

No. ýaý o
1 25 2 37 3 40 4 '41 5 50
Vertical profiles of tern] erature.
6 55 Zettle IJere, 1974. 7 56 White Mere, 1974.
8 57 Isettle AZere, 1975. 9 58 White Mere, 1975.
10 59 Kettle Mere, 1976#' 11 60 White Mere, 1976.
Vertical distribution of oxygen. 12 61 Kettle Ilere, 1 975 " 13 62 White Ziere, 1975.
14 63 Kettle I"lore, 1976.
15 54 White. Mere, 1976.
16 67 Absorption of filtered water samples from
nettle Here and White Mere.
Total particulate sedimentation.
17 84 Kettle IIore, 1974.
18 85 kettle T. ere and White liiere, 1 975 "
, . 19 86 Kettle D? ere and White here, 1976.
Rate of sedimentation of algal riaterial.
20 " 88 Kettle Ilore, 1974.
21 89 Kettle Iiere, 1975.
22 90 white Mere, 1975.
23 91 Kettle here, 1976.
24 92 White *1ere, 1976.
"u: 3I)ended sot id. 3.
25 93 Kettle IIere nnci White hero, 1975.
26 94 Kettle here and #+ýiitu Liere, 19lß"
LIJT ul" i a.
GUt.: U
i1t]. e
The Pieres of the ' 11esriere Region.
: JaxipLing sites on I: ettle llorc and ý3hito Mere.
Di asrara of sedimentation trap.
Positioning of r; ediraent traps.
Column used for reducing nitrate to nitrite in
waiter samples.

-13-
Iyo. Page Title
Amount of algal riaterial in the surface eedir-cuts. 27 96 Kettle Nere, 1974.
28 97 1Jhite : Tere, 1 974 . 29 98 Kett? e here, 1 975 " 30 99 White here, 1975. 31 100 Kettle Mere, 1976.
32 101 ºlhite Mere, 1976.
SEC'T'ION 2 33 130 Spectral distribution of energy output of
"Daylight" fluorescent tubes.
SECTION 3
34 168 Digestion apparatus for the extraction of labile
orthophosphate.
35 169 Partial removal of as phase frogs serum bottles
using "Vacutainers".

SECTION 1
THE FILLD D. ýTA

IIýTIWUDUCýICI4'
The succession and growth of algal species has been
the subject of much research. The potbntial object of this
research is the control of excessive growths of the "nuisance
species" (Fogg, 1969) of algae. - The effect of this excessive
-growth, commonly termed "blooms" or "water blooms" vary
from unsightly scums on the surface of the water or an un-
wholesome soupy appearance, through the clogging of filters
in water treatment works,, to the poisioning of livestock
through the release of toxins. Fish mortality may result
from deoxygenation of water as the algal mass succumbs to
bacterial decay.. The contamination of water supplies is
produced through the release of organic compounds which the
blue-green algae excrete in abundance (Fogg, 1952). Although
recently brought into prominence through concern over the
environment, algal blooms have long been recorded. One of
the first printed records appeared in Smith and Sowerby's
"English Botany", 1808,. in which Gloeotrichia echinulata (Conferva echinuleta) was observed in a lako in Anglesey.
I1uch earlier is the record by Giraldus Cambrensis, who in a
tour of Wales during the twelfth century, writes of what,
' from the description of colour, probably was a Cyanophycean
bloom in Llangorse lake, Breconshire (Griffiths, 1938).
The problem has become increasingly apparent with the
enrichment of lakes and rivers through extensive urbanizat-
ion and the proliferation of the use of inorganic -
fertilizers. This process of eutrophication has produced ''a
general concern over the health of water bodies, particularly
with increasing demand for supply and recreation.
It is apparent that any effective control over the
ecology of aquatic organisms must be based on a comprehens-
ive lwowledge of the behaviour of the separate species to
the range of conditions which are likely to be met, the
interactions between species, and the effect that each
species may have on the environment. Of equal importance,
and only slightly less complexity, is a detailed compre- hension of the ph; 'sical and chemical factors of the system
under investigation.

-16-
The dominance of blue-green algae in many lakes and
reservoirs in both temperate and tropical regions has been
studied in relation tb many factors. Of particular importance in temperate regions appears to be the amelior-
ation of conditions during the. spring. Increasing insolation
results in a warming up of waters and a more favourable
light climate for algal growth . (Tailing, 1971). A 'decrease
in'the amount of wind will result in an increasing stability
of the water column, which favours motile and buoyant forms
(Moss, 1969) at the expense of the denser algae, particularly
diatoms, which rely on turbulence to maintain the cells in
suspension (Lund, 1'966; Knoechel and I: alff, 1975). The
transition from the spring to the summer assemblages is the
result of complex interactions between temperature,
turbulence, solar radiation and the physiological capacities
of the species concerned. The amount of time spent below
the photic zone will affect' different species to varying
extents as a result of the compensation point requirement
for active growth. Temperature may have a similar effect in
the selection of different species through differences in
the optima of photosynthesis and' respiration.
The close correlation between temperature and the
ascendancy of the blue-green algae has been demonstrated by
Hammer (1964,1969) in Saskatchewan lakes. ITinimum temp-
eratures have been determined for the growth of different
species which, together with varying optima for the concen-
tration of orthophosphate, determine the order of occurrence
of different species. A similar relationship has been
observed by Reynolds (1971) for the Shropshire (Salopian)
meres, but with variations in the order of the species
common to both areas. Lin (1972) ex, mined the iYifluence of
a reduction of nitrate concentration on chdnge of populat-
ions of algae, from a diatom community to a blue-green
algal one. -Interrelationships between the blue-green algal
species have also been investigated with a view to.
explaining the sequence of succession. It was suggested
-(Lin, - 1972; ilacuner, 1964) that the decay of the previous
species stimulated the growth of subsequent species. Vance (1965) also stresses the importance of extracellular

-17- metabolites in the control of growth of natural, populations.
The inverse relationship,, the inhibition of growth of the
proceeding species by the succeeding' species has been
demonstrated by Keating (1975) in cultures and also using
filtrates of lake water collected when the various species
were dominant. - With the relative, stagnation of the epilimnion,
nutrient supply and deficiency produce, further selective
pressures. Available carbon dioxide has recently been
discovered to be important (Ruent. el, 1969; King, 1970). A
supply of gaseous carbon dioxide results in a change of the
'dominant species from the, blue-green algae, which are more
efficient at removing carbon dioxide at low concentrations,
to. green algae (Shapiro, 1973). A similar effect has been
"observed by , Haynes (1975)
. on the artificial destratification
of a small lake. However, ,
Schindler et al. (1972) has shown
that sufficient carbon dioxide enters the water from the
atmosphere ' through diffusion to permit eutrophication, ifý
concentrations of nitrates and phosphates are sufficiently
high. Destratification would also supply other nutrients
from the, hypolimnion and stimulate growth (Iverson at al.,
1974). If completely mixed, however, _a nutrient limited
bloom will be replaced by a light limited one of smaller
size (Lorenzen and Mitchell, 1973). The depth available for
mixing and the attenuation of, light, the "critical depth"
of Sverdrup (1953 would be decisive.
The ability of, luxury consumption and storage of
phosphates by blue-green algae (Battex-ton, and Van Baalen,
1968), the enzyme mediated relea8e of orthophosphate from
organic phosphorus compounds. (Fitzgerald, _and
Nelson, 1966;,
Heath and Cooke, 1975) and the-ability of aerobic nitrogen
fixation in the heterocystous species (Fogg, 1971) are
probably important factors in supporting bloom densities of
algae. Although the rapid appearence of a dense scum of algal
material gives an impression of a vast bionass and a
phenomenal growth rate, this is often nisleading.. Reynolds (1971,1972,1973a) has shown that a bloom is merely the
accumulation of plan,: tonic material at the surface , of the

_is_ water, "t ie suspended 'material becomes' 'crammed' within a few
centime tiers at the surface. This buoyancy is imparted by
the pr es: enoc of gas vex iclos within the cell leading to a
reduction of density. 'The relationships between cell turgor
pressure, photosynthesis, 'gas vacuole number and buoyancy
are described by Walsby (1 971) and Dinsdale and 4. alsby (1972).
With the incredise of cell turgor pressure brought about
through the accumulation of osmotically active compounds
produced by' photosynthetic activity, the weaker gas vacuoles
collapse. The density of the alga thus increases, and'the
cells'sink through the water column to a region of lower
light intensity. ,A
reduced rate of photosynthesis, ' which
may also be brought about by carbon dioxide deficiency
(Waleby and Booker; " 1 976) , and 'the conversion 'of - simple
sugars. to osmotically inactive storage pröducte, results in
a reduction of cell" turgor pressure and enables the further
de novo synthesis of vesicles. These factors were also
investigated by Reynolds (1975a) for a natural population
of Änabaene circinnlis. Surface blooms result from an over-
reaction of the potentially sensitive and accurate buoyancy
control mechanism (Walsby and Klern. er, 1974) with a sudden
decrease of turbulence (Reynolds, 1972). Thu' overbuoyant
algal material rises rapidly to the surface, where it
becomes--trapped by surface tension. Gentle wind then removes
the scum to the lee shore, but mixing into the water is not
achieved unless the. wind is, sufficiently strong to break
the surface of the" water.
In this position the algal populations are highly
vulnerable to extremes of temperature and light. Bacterial
decay would proceed at an optimal rate at the temperatures
generally' encountered at the surfpLce-during calm conditions
in the summer. Alga-lysing bacteria were found in many
eutrophic freshwater lakes. Though growth requirements of
bacteria and condition, for lysir are similar to that
which prevail in stagnant surface waters, -extensive lysis
is rare in natural ecosystems (Daft et al., 1975). The
"nue bore of bacteria found in the natural environment was
never enough to lyse an algal bloom within 60 hours. If
there is an input of more bacteria (eg. from a sewage works)

-19-
or if the' alga-is attacked by other pathogens, the conditions
would favour lysis by the bacteria.
The depletion of-carbon dioxide and the supersaturation
of oxygen, produced, as a"result of the high rate of. photo-
synthesis in productive waters and the-increase of
temperature, leads to photo-oxidativo-conditions (Abeliovich
and Shilo, 1972; Boyd et al., 1975). Photo-oxidation has
been shown to occur under natural conditions even at
relatively low temperatures (Shilo,,, 1975), though, natural
populations, have a greater resistance against photo-oxidation
than laboratory. grown material. This resistance is, conferred
by high levels of super-oxide disrnutase (Eloff et al. ,, 1 976) . Since the"greater portion-of the planktonic algal
population is compressed within a shallow depth of water at
the surface during blooms, the survival of some of this is
critical for the development of future populations.,
A , factor of considerable importance for the periodicity
and development of large planktonic populations, which has
so far received scant attention when compared with the
number of investigations performed while algal material is
abundant, is the distribution, abundance and behaviour of
the algal populations during the greater part of-the year
when they are not an important component of the phyto-
plankton.
An important adaptation of many heterocystous blue-
green algae toadverse conditions is the formation of
akinetes, resistant "resting spores". which are protected by
an elaborate envelope (Fogg of a., 1973). Each akinete is
produced by the transformation of. a single vegetative cell,
and-this is accompanied by cell enlargement,. deposition of
storage material . and` envelope layers, and in the planktonic
species, by the disappearance of Gas vacuoles. The increase of cytoplasmic density resulting from the
loss of gas vacuoles and the massive deposition of cyano-
phycin (structured) granules permits the akinetes to sink
and ovorwinter on the mud sediments (Wildman et a1.., 1975)-
Thin-has been observed in many water bodies, particularly the deeper continental lakes, and it is generally assumed that the tikinetes form the : 3tock for future vegetative

-20-
growth of the Anabaenn species . (Vladimirova, 1968). A
similar pattern has been ob6erved for G. echänulata (Hoelofs
and Oglesby, 1970; than, 1974) and 4DhRnisomenon floe-accuse
Rose, 1934; Chernusova-et al., 1968). -Some doubt has,
however, been expressed as-to the relative importance of
sedimented overwintering akinetes, and whether they
represent a significant source of vegetative material. Although germination has been observed to occur with improv-
ing conditions of. light and temperature in the. spring while the lakes are, still unstratified (Reynolds, 1 971 , 1972)t
the numbers encountered are never very large and probably do not contribute significantly to the population maxima (Reynolds, 1972,1975a). Sufficient vegetative material
survives during the winter within the. plankton, and the
. increase of populations can be^ccounted. for by the,
vegetative gröwth of this planktonic material. Reif and Fine (1967) also observed planktonic populations of an Anabaena species.
-. Non-sporulating species, particularly Microcystis, may
survive the winter, in a viable vegetative , state in the
bottom muds (: iirenko et al., .1 969; Reynolds and Rogers,
1976). An increase in the depth of mixing, as summer
stratification breaks down, initiates the hydrostatic
collapse of gas vacuoles and the sedimentation of colonies. Investigations by-Reynolds and Rogers. (1976) on Rostherne
Piere, one of, the deepest of the meree, have clearly shown
the sedimentation of, colonies into the'mud, where'they
remain during the winter.,, At the onset of summer stagnation in the following year a marked migration of colonies takes
place from the sediments to the epilimnion. The transition of a vegetative population to a resting
stage may be expected to be initiated by a deterioration of the conditions neccessary for vegetative growth. Nutrient
deficiency, particularly that of orthophosphate (Wolk, 1965)
and nitrate (Glade, 1914) have been reported to be control- ling factors of akinete_formation in laboratory cultures. Experiments of Fisher and- tlolk (1976) showed that extra- cellular products taken from an old stationary phase culture stimulated the immediate formation of akinetes when. added

-21-
in small amounts to juvenile cultures-of Cy lindrospermun
licheniforme. This finding may be- corroborated by the
observation of Rose (1934) on natural. populations of. Aph. flos-aquae where the decay of-surface blooms during
the summer induces eporulation, of filaments below the
surface. This'is probably a coincident'-rather than a
causal relationship, as this species has been observed to
produce akinetes during the winter months (Lemmerman, 1900;
Wildman et al., 1975). Akinete production in G. echinulata does not appear to be stimulated by adverse conditions (Roelofs and Oglesby, 1970), as they were observed in
rapidly growing cultures and in planktonic populations before the population maximum was reached. Fritsch (1945)
suggests that the filament size in this genus and in
Cylindrospermum may determine akinete production, =but no
indication of potentially-respon&ýble factors were given. With few exceptions, akinete production is'limited to
heteroc. -stous (nitrogen fixing) species. Singh (1942a),
Hill (1972) and Cronberg (1973) have described species of
the genus Raphidopsis, a non-heterocystouc, alga in-which
akinetes have been regularly observed..
From experiments in which the differentiation ofla
vegetative cell into an akinete is inhibited by. surgical
rehoval of the adjacent heterocyst, 'lolk (1966) postulates
a control over sporulation by heterocyzts. This is supported
by general observations of a consistent relationship
between the relative positions of heterocysts and akinetes
within a filament. The relationship appears tobe species
specific. Departures from-this were. also described. No
regular relationship was found between the position of
akinetes, and heterocysts of iLnnbaena planktonica (Burger,
1974). Sinha and-Kumar (1973) obtained noii-heterocystous
mutants of Annbnonn doliolurn which were capable of
sporulation. Tyagi (1974) has demonstrated an inhibition of
sporulation by inorganic nitrogen compounds and a stimulation
, by glucose in the wild-type strain of the same species. A
similar inhibitory effect of nitrate was shown by Singh and Srivastava (1968). These observations' tend to suggest that the position of akinotes within the filament depends on

-22- criticMMl concentration levels within the cell of the
prothicts of nitrogen and carbon fixation.
Factors which could be of importance in the control of developmental stages of akinetes are light quality and
quantity. A light induced' developmental cycle has been
described for I"Jostoc muscorum '(Lazaroff and Vishniac, 1 961 )
in which low light intensity, or the addition of culture filtrate from light grown cultures initiates the formation
of the typical-Itostocacean morphology from the aseriate
stage of dark grown cultures. The effects of light quality
on the Nostocacean developmental cycle was inveotigated by
Lazaroff (1966). A non-photosynthetic control mech"anism"
appears to function. Red light (650 nm)'stimulates the
development of'filaments from the aseriate stage. This
effect may be photo-reversed byý exposure of the alga to a
broadband in the green region of'the spectrum. The action
spectrum'of the photo-induction. of the developmental cycle displays a single sharp. peak of activity at 650 nm (Lazaroff
and Schiff, 1962).
Light is also a factor required'for akinete germination.
Harder (1917a) established a linear correlation between
light flux and the rate of germination. A similar relation-
ship was observed between light intensity and germination. No germination occurred under dark conditions. The same was
also observed with A. doliolurl by Singh and Sunita (1974).
Red light was found to be most effective in the stimuj. ation
of germination (Harder, 1917b; *Reddy et al., 1975). The
effect of red light can be reversed by far-red. The green
and blue regions of the spectrum have no effect (Reddy
et a?., 1975). Kauchik and Kumar (1970) found that
germination could occur in all regions of the spectrum, indicating a requirement for non-photosynthetic light and
an absence of developmental photo-control of the type
described by Lazaroff (1966).
No light requirement was observed for the germination
of a'_; inetes of d:. i1_on-neune although this species appeared to require cold pretreatment before germination (.: ose, 1934). Germination immediately after akinete maturat- ion has been observed under favourable conditions with many

-23-
species of Anabnena (Fritsch, 1904; 3pratt, 1911; Fay, 1969a).
Glade (1914) also observed the imrriediata'germinatiori'of
akinetes of Cý, indrosi)brmum on the re-addition of 'fresh
medium, and Singh and Sunita (1974) of akinetes produced in.
light grown cultures of A. doliolun.
Although some data have been gathered on factors
affecting akinete production and germination, these are
almost exclusively confined to'laboratory studies and to
algal species obtained from culture collections. The species
represented are originally and principally coil-borne algae,
the planktonic forms being relatively-difficult to culture
successfully until recently. ' Little work has been done on
the importance of-akinetes as overwintering or resting
structures in natural planktonic populations.
In this study an attempt was made at'the'assesment of
the relative importance of akinetes and vegetative material
in the'perennation and prdpagation of planktonic species of
blue-green algae in two contrasted lacustrine environments,.
and also of factors which affect population development.
This was coupled with laboratory experiments; -using isolates
fron the study-sites and natural plankton samples, designed
to isolate'the potential factors responsible for the
induction of sporulation and akinete germination.

THE P"i1: RE
The mere: of the Salop-Cheshire Plain are a series
of water-filled hollows in the mantle of glacial drift
which covers the solid rock of the region. The thickness of the drift varies, but in the Ellesmere" region the Triassic
sandstones and marls lie under about 100 rm of the recent deposits (Sinker, 1962). The hollows may be either enclosed between hummocks of glacial deposits, or kettle holes formed
by the melting of large blocks of ice left during the
retreat of the ice front in the last (Weicheelian)
glaciation. Some of the merest particularly in Cheshire,
may have resulted from subsidence through the dissolution
of the underlying strata. The mores have a clustered distribution, one of the most important of which lies in
the region around illesmere (Fig. 1). This also marks the
southern boundary of a minor re-advance of ice, resulting
in a terminal moraine connecting Wrexham, Whitchurch and Bar Hill in a bilobate form (Boulton and Woreley, 1965).
An a result of their mode of formation, many of the
meres have neither natural: inflows or outflowe of any
significance. In some cases the mores are connected to each
other and to the adjacent low lying areas by ditches, such
as the Crose P'Iero-Sweat I"lore complex (Sinker, 1962). The
water is supplied principally by percolation through the
drift, the water level marking the intersection of the
water table and the surface of the ground. Reynolds (1975b)
concluded from studies on the supply of silica in Crose
Mere that the inflow of water is through the rim of the
viere, no overflow,, _ rather than through the lake basin in
general, which in rendered impermeable through the
deposition of secondary sediments. Despite the buffering effect that a phreatic water
supply would have during periods of drought and on the
chemical environment of. the mores, considerable fluctuations
of water levelo in the meree have been produced as a result
of drainage scheme during the lust century. Hardy (1939)
"r!. entione a tall of 2.5 in in the level of Crone here and further instances tAre given by Sinker (1962). That
fluctuations in levels have occurred before written records

-25-
Fig. 1 The Peres of the Ellesmere Region.
SCALE: - One'inch to one statute mile = 1: 63360
Iianmer, Mere
Grid'
north
The More
Ellesmere
Newton Mere
Blake Mere Kettle 1Mere
Cole Mere
White Here
0 Sweat Mere
Croce Mere ,.

-26-
may be inferred froca the sE. parati6n 'of 'open bodies- of`
water by peat deposits, such-as the separation of Blake
Mere and- I. ettle- Mere by a peaty bank. Fluctuations of
water level, as well as the steepness of, many'of the banks
and a lack of-input'of'. sediments in enclosed' lakes, may-ý°'
account for the general paucityýof marginal vegetation (Sinker 1962). ,
Through the leaching of dissolved' compounds in the
glacial drift by the ground water which supplies°the meres,
these lakes became eutrophic. - However a range of
concentrations of dissolved material'exist in the different
meres, which Gorham (1957) correlates with differences of
local geology. He also attributes the high' level of sodium
and chlorine ions to evaporationresulting'in a-concentrat- ion ofthese ions. Although thV. 'Ellesmere area has a
precipitation to evaporation ratio of 1.4: 1 (Sinker, 1962)
this would' result". in a 3.5-fold inoreäse'of dissolved
compounds when compared'with the input through precipitation" (Gorham, 1957).
As discussed above, blue-green algal blooms are not
wholly'a recent ' phenomenon: Neither are they to the.
inhabitants of the-shores'of'the meres. "The manifestation
of surface blooms, -eloquently described by : linker (1962),
has been preserved in the local folk-lore, -which has
produced its own term for blooms: - the "breaking of the
nier. es". Another colloquial expression, although less well known, is-the "cruddling of the Tieres" , by analogy with
the separation of-milk into curd and whey on the addition
of rennet (Wardle, 1897). Phillips (1884,1893) was the
first who described the Cyanophycean origin of the breaking
of the meres around Ellesmere. Further descriptions are
given by Griffiths (1925), and by Reynolds (1973b, 1973c)
who also compares the annual periodicity of Grose Viere with
that of the other mores and of the Cheshire mores described
by Lind (1944). The similarity of the composition of
species of algae in many mores led to the concept of a
-'regional type' of succession (Reynolds, 1973b). Departures from this are discussed in terms.. of the typo of agricultural
ac tivityy in the area bordering the mores, taxe nature of
I

-27- the catchment area, morphometry of the mores and ionic
composition in relation -to the ecological ranges of the
predominant species.
Although the merea represent, a chemically stable habitat, as indicated b; r the absence of major qualitative
chances in the phytopl anl: ton, an increase in the use of inorganic fertilizers on the farmland within the catchment
areas are probably the cause df the largo increases of
nitrates in recent years (Table 1). An increase of 4 to'
10 - fold has been observed between the investigations of Gorhan (1957) and Reynolds (1973b). This is not restricted to the meres in'Salop, an Rostherne Mere in Cheshire also
appears to have undergone increasing eutrophication (Pearsall, 1923; Lind, 1944; -Grimshaw and Hudson, 1970),
although the increase of nitrates is leas marked.
Concentration of orthophosphato, already high when first
analysed in 1955 in samples of water from the mares (Gorham. 1957), have changed little when compared wlthýthose of
nitrates over the same period, (Reynolds,. 1973b).
The monitoring of changes in the phytoplankton
assemblages which result, from changes in the nutrient
concentration and-composition of the mares, particularly'an
-alteration from limiting concentrations of nitrate to those
of phosphate, would be of Great value in determining the
environmental requirements of planktonic algae, and their
interrelationships. The Salopian mer. es, with a wide variety
of sizes and morphometry within a relatively small area,
appear an ideal site for such a study. The mares are deliberately referred to in the singular; their importance
lies in their value as a group of lakes, and it is the
Croup which should be studied and conserved.

0 4) U U
H
"0
F4 O0 El
O H
H El 0
N N
EHI (11 1i Uý ^
a^
Uy
O"
Or a ßr-4
to
C) $1 U'
N 4) i
"ri "r1 El St P'O
cl UO
O U] ý, ) v C)
9 GS 4
C. )
x p4 C) .n r-3 E+
t4 0 4.3
F-4
. -. -. OD o c- c, ON
1 f
.0 L(\ rn
C) Q>
Lr\ i S4
r £ ý
Q) " rl "r1 X 3 t1 .
0 0 0 ö. 1 1
to o
fJ i+
Co Co
0 01
oo .o v- , +ý o
i N
e O
O O
"I
-Co N ul O c- N
11
Lý Nrr
t(1 O
11 tz
, -.
A
.' N" "ý ý\ r C .' '-, r 't. )
v C! ) 7 ti . 4. ) r1 Iß
cO
Q. Rr E"t
-28-
.ý ... CC) %lo 'n N ON ON
ON a (IN Ir- T-
... . V,
ßs1 G) U tA3 13
r J ý v t! J cd
S4 G\ U c'
Q) ,0 "ri "r1
ý? F c3 ß
'co CJý N' ao rr\ i " r O O O Q O t
O 'D 0
O O aý U N Sa ý ý 9) Q fV " " ;.; u1 0 O r 0 0 Q) t !
+) O O "r1 r O
N
0 0
Ill o N CT NN
wO rN
N
C I. .Ii
. ý i hl
ON v
ro 4) r. ri t7 cj 0
r
O r' C3
. W-4 E4
a'
u-ý a3 v LA G
v1 "a 0) r
[, - C-
C)
ö ýrt 1-3 :;: ti
r1 U1 O0O 0Ö11
O
NO w Kl%
N 0
LA OO M OOii
co co 1O
ci O
rn a' tt11
oo
03 "11
o-% 1 11 Co
C'
0
0 O 0'
0 0 ä
CO
U .U o
O
co Q N N t\ C1 Q " "
:: r- U CJ ý
ß) " ! 1 1 U + 0 0
U%
0
M +11
Co O
'i C' CO
ti ON N C" u' G' r-
'- f L1 cr T-
. .. I r- "a r- -
2I r-{
.4-) Lý V r"1
4-1 GZ
Ci .Z
0 1.0
rii .4
P Q
4 0 G 0
4
) r
a R: E+ 1 0 Pý Eý i

LIST Uff' . ̀i''1 Lei r)-yCOU1 T1: HLV IN TH I'ii 1ti 3 DURIIdG THE
PERIOD OF THIS INVI. "'iSTIGATICN. (1974 - 1976 ).
Kettle White Newton Blake Mere Mere Agiere Mere
MYXOPÜYCEAB: Chroococcalea
Coeloq haeriuri ku. tzing; nnuri Ilttiieti. +
Coel osphaeritun naeglianum Unger. + +
I'? icrocystis aeruj±nosa K{itzing. + +
i"Iicrocystis floc-Eigurte ('Iittrock) +
Kirchner
I"iicrocystis wesenbergii Komarek + + ":
' Hormogoniales ,
Anabaena circinalis (Kiltz. ) + +
Hansgirg - Anabaena flos-aquae (Lyngb. ) Ereb. + + *#
Anabaena solitaria-Kleb. + +
Anabaena spiroides Kleb. .+
+
Aphanizonenon flos-aquae (L. ) Ralfe. + +
Gloeotrichia echinulata +
(J. E. _Srith)
Richt.
Oscillator. is Opp. + +.
Predominant genera of other Classes. .
13ACILLARIUYHYGEAE
Melosira. spp. +
Stephanodi. scus epp. +
Asterione- 1a formosa- Hass. - + +
Fragilaria cr-otonensis Kitton
" CHZUF. O1HYC1! AE: Volvocales
Ludorina spp. +
YPnd. orina App. +
Volvox , l, )bator Ehrenb. +
Chiorococcales
Anl: istrodesrlus spp. +
Scenedesmits upp. + +
Yedirnstrum boryranur"º (j'urp. ) Heneeh. +
I'edinstrt: m (1111-)l -nx Iisyen +

-30-
Kettle White Iiewton Blake Mere' fiere : Tare Here
Conjugaloa
Clo3teriurn spy' +
DINlOP: HXCI.. B
Ceratium hi. rundinella 0. F. I-I. +
YericUniur' sp. +
CRY. T UPI YCEA
Cryrptorzona , spp. +t
Notes.
Presence recorded. , Recorded, but collections mad©
only in limited periods during the eu ier.
No record does not indicate absence of the species
concerned in Blake Mere and Neuton Mere.
The M xopiayceae were identified and named from Geitler
(1925) and Komarek (1958).
Where information is given, in the descriptions of the
raeres investigated, of . the species which were found by
other workers, the full name an3. authority is given of that
species quoted in the paper. cited. This is because the
authorities used by the various investigators are different (including the authorities quoted-here). However, where the
same species was found by. the same author indifferent
meres abbreviations are used, after the full name and
authorit; ts are given where first encountered. Where
further reference (e. g. in the Results to Section 1) to
the species found in the meres during; this study are made,
abbreviations are also used.
hh-

-31-
ZOuPLANKTOII
The data given here were supplzed by rosemary J.
Nicholson (perscnal' corzmunictition) froi work done on''
Kettle Mere and White Mere during 1974.
Predominant speci0s:
CLADOCERA
Daphnia hyaline, Loydig
Daphnia lonpisnir 0. F. fuller
COPEI'ODA
Cyclone strenuus Fischer.
It was observed that the population maxima , of both
the cladoceran and copepod species tends to co-incide with
high densities of Aphanizomenori and Anabaenx species. There
is no indication, however, of significant grazing of the
filamentous blue-green algae by the zoopJ. ankton. Changes of
zooplaxkton numbers may be associated' with increases of
bacterial populations during summer blooms.
From studies using algal strains from the Cambridge
Culture Collection it was found that logarithmic phase
cultures of A. floc-aquae, Anh, floc-F duce and M. aeruginosa (non colonial form) were capable of supporting Daphnia
Growth. Aphanizomenon gracile Lemm. and G. echinulata
were not grazed under similar laboratory conditions.
khý-

THi: Sl'S`. E 3 INUM2IGATiil)
Kettle. i"iere.
Kettle Iiere (National Grid reference SJ 418 341)#-
which, as the name suggests, is probably a kettle hole,
is one of the smaller mares, having;, a surface area'of 2 ha
and a maximum depth of 7 m. It is well sheltered from winds
in all directions, `being `at 'the bottom of a deep 'conical"
depression and fringed with' woodland'. The form-of the lake
basin is bowl-shaped, with steep banks, 1 to 2m high except
where eroded by the livestock which drink at the mere and
probably contribute to the already hi, gh'concentration'of
dissolved compounds. Iargin'al vegetation is poorly developed,
and is. -limited to a small area of Nurpher, there being-no -
change since the investigation of Griffiths (1925). Due to`
the surrounding. deciduous woodlands and to the peat, banks-,
around the mere the input of organic material'is considerable.
she water is coloured a peaty-brown and the=bottom mud is
principally an unstructured. black organic-ooze (gyttia).
Although having a relatively low conductivity when
compared with other mores, the concentrations of orthophos-
phate and'nitrate are high. -The concentration. of a range
of ions are given by Gorham . 0957).
This mere. has - long been famous for ' "breaking"
(Yhillips, 1 854), `the dominant species at that time being
Anahaeria cirdinalis Rabh. Griffiths (1925) records.
Anhan: izomenon f1_os-sguoe (L. ) Ralfe as the- dominant
together with Anabaena a'ffinig Lemri:, T4 icroc, %stis floc-aquae
(Uittr. ) Kirch. was rare. The predominant species found by
Reynolds (1973b) between 1966 and 1968 were Anfibaena
circin ilis °Rabh. , ex Born. et Plah., An. abnena f'! os-acturne
Breb. ex . Born. - et Plah. -' and Anhanioonenon floc, -aaune Ralfs
ex Born. et Plah. IIicro'c stir or coccoid species wore not
recorded.
Uh to Toro.
White f'iere' (Rational Grid reference : 3J 415 330), with a stü., face area of 26 ha and a. muxiczum depth of 15 n, is
considerably rlora exposcd than 1: ettle ! ier. o. The deeper
uree. s are relatively enal:! and are collfiried to a steeply

-33-
shelving portion öf the east end of the. mere. In general
the depth is betw'en 4 and 8 m, gradually deepening from
the shallow west end -up to the lip of the trough in the
east. Marginal vegetation; `predominantly `iýenyanthý s and Nuphrar., is confined to the shallow areas with gentle
gradients in the leeward regions of the mere. Fine organic
sedimcits accumulate in this region, the sediments in the
more'exposed areas consisting mostly of sand. This is
probably a result of wave action, which can be considerable,
removing the smaller'particulate deposits.
The concentration of dissolved salts in higher than in
Kettle Ziere and a wider range 'of algal species is found.
Only A. circinaiis was observed by Phillips (1884). Of the
filamentous species"a variety of A. "affinia was recorded as
dominant and Anabaena cir. cinaDis Kuetm. Hansg., Aph. flos-
aquae and Rivularia echinulata l?. Richter as rare by
Griffiths (1925). No I"iicrocystis species were recorded.
Reynolds (1973b) records A. circinalis, A. flos-aquae,
Anaheena Espiroides Klebe, Anabaena solitaries Klebe and
Alph. flos-saune as well an Microcystis aeruginosa Rutz.
emend Blenk. and Coelosphaaeer. Turn -r_aegelianum (Unger. ) Lemu.
Blake 'Piere and Newton 'Mere.
During 1976 a'! gal material from Blake P"Iere and more
espocially from Newton Acre, was collected and used for
expeririental purposes (see Section 3). Thus brief xV
descriptions of these meres would be useful, although no
regular field-work was done at these sites.
Make I Piere. (National Grid " reference SJ 416 339).
This ziere, which occupies the same basin as Kettle
Mere and is separated from the latter by a-peaty bank, is
as sheltered as Kettle Viere through being completely
surröurided by woodland. The two meres are connected by a
shallow ditch. The surface area (8 ha) of iOlakle ; -: ere is
considerably treater than that of : settle T"lero, and has a
rnrxin m depth of 13 n. : ": arginal vegetation is relatively
sparse although is abundant in the shallow bays.
slic; rare aquatic p_ ant, Lunhrgr puniest, was reported to

-34-
occur in Blake Ilere (+3in1: er,, 1962).
Apart from Nowton Nero, Blake Piere is the most dilute
of the meres examined by Reynolds (1'973b). The boatmen of
the Shropohire Union Canal, which runs along the western
chore of the mere, were aware of this aspect of chemistry.
The soft water of Blake I-? ore was used-in preference to that.
of other eueres on account of it making such good tea
(Phillips, 1884). No significant increase of dissolved
orthophosphate was observed between Reynolds' (1973b)
investigation and this study (Table 1).
As well as the softness of the water, Blake Mere was
also distinguished by the absence of breaking (Phillips,
1884). Griffiths (1925). stated that the mere was not known
to break until the. last few years. He records A. affinis,
and A-. circinal_is in smaller quantity.. The following species
were recorded by Reynolda (19-73b): - C. naej elianuri,
A. circinalis, A. floc-Hnuae and Anh. floc- ice.
Newton P-sore. (National Grid reference SJ 425 343).
In terms of the degree of shelter this more is
intermediate between Kettle Mere and jihite Mere. It is of a
similar size and depth as Blake Mere (i. e. a surface area
of 9 ha and a maximum depth of 10 m) but is more exposed
due to the sparsity of surrounding woodland. Marginal
vegetation is also very scarce. Newton I'"lere is the most dilute of the series examined
by Reynolds (1973b) and, like Blake Fiere, no increase in
the concentration of orthophosphate was observed (Table 1).
Attempts to reduce the breaking-of this mere by the
addition of copper sulphate were made for several years (Griffiths, 1925).
The formation of an early bloom (during February) has
been reported by Phillips (1884), the species involved
being A. circinglis and Cocaosj)ti, -, ieriun klit int ianum NL1C.
Doli. cro! -, nernurt ralfo ii (Ktltz,. ) [= A. affinis according to
Griffiths (1925)] was observed during the summer. Species
recorded by Griffiths (1925) were: - Rticrocy! stis aerut-iinosaV Kfltz. (dominant) and in smaller quantities A. circa rialis
and t; nrttýlict: ýplýaý, rý tzrt 1ict ý .L rt:: ur1 (Unger) Lem-r-1. (rare).

-35-
Reyno? _ds
(1973b) z. "ecord - A. cir. cinali s and Anh. los-&iine,
but none of the coccoid : ipecies.

3L.
MA ER GALS AND METHODS
Field 'Sampling.
Plankton. - Plankton samples were collected at regular intervals
of depth, generally of 1 m, using a Ruttner sampler of 11
or 21 capacity. The sites from which samples were collected
are indicated on fig. 2. When first lowered into the water
the sampler fills with surface water, which, due to the
lids which obstruct the free flow of water through the
sampler tube, will be carried down to deeper layers. To
reduce contamination the sampler is raised and lowered
through 0.25 ma few times to flush the sampler at the depth
from which the sample is to be collected. Although this is
undesirable as far, as stratification is concerned, it
appears to be effective, in that little blue-green algal
material is collected in regions where none would be-
expected to occur, such as below the thermocline in stably
stratified lakos dur: iný7. surface blooms. This has also been
verified using alternative sampling methods (such as a pump)
which are not prone to this error.
After bringing the sample to the surface the water.
was partitioned for various analyses described below.
When blue-green algal material was sufficiently
abundant, large volumes (5 to 10 1) of water were collected
for analytical and experimental purposes and for the assay
of physiological activity.
As far as possible, -preparation and preservation of
the samples were carried out in the field. Material for
visual exam4ý. nation and counting were preserved by the
addition of Lugol's iodine to 'a final concentration of 1i",.
Samples for chemical analysis (dissolved com-ounds) were
filtered sterile through 0.45 µm Millipore filters, or
analysed immediately after collection.
Benthos.
Mud samples were collected using the Buttner sampler, or, during 1975, a Jenkiii surface mud sampler (Mortimer,
19, t2). It was realised that the Kiuttner sampler is not the
most suitable instrument for accurate ouraplirig; of muds, as

-37-
Sampling position
0 Sedimentztion trap
200 m'

-38- the bottom lid disturbs the surface sediments as it is
lowered. The type of sediments generally encountered in the
lal: es-studied was a soft organic ooze within which the
material is zairlry evenly nixed'by turbulence (Davis, 1968).
An exception to this would'be the recent sedimentation of
algal material during periods of thermal stratification. The-softness-of the surface deposits enabled apparantly
undisturbed samples to be collected. The weight of-the
sampler alone is-sufficient to'make it sink into the mud,
once lowered gently'to the bottom and kept vertical by
maintaining, "tension in the line. -These problems were avoided by the use: of the Jenkin sampler. This sampler is designed
to minimize the disturbance of-surface sediments produced- by turbulence as the sampler is. lowered or as-the sample is
collected. An un-obstrtiicted. Perspex tube is>held vertically
in a four legged frame, thus enabling the sampler-to be
stood on the'lake bottom after slowly lowering itt. through
the water. The lids which seal the top and,. bottom'of the
tube are. held back, by levers. Powerful springs clamp the
lids onto the tube once the catch-is released. The movement
of the lids is damped, using a dashpot, to reduce disturbance
of the core sample. The cores were separated into-three portions: - the
water just above the mud surface, the surface sediments,
and the deeper-sediments. -Bach portion represented-approx-
imately 50 mm depth of the core and were seperated by 50 mm
from-each other to prevent cross-contamination. The most
effective method of separation of the core was also the
crudest. The sampler was carefully tilted and, the excess
supernatant water first decanted off., hnown volumes of each
portion were'- collected by continually pouring into calibrated
sample bottles. The loose and flocculent nature of the mud
-did not allow the cutting of sections, and the presence of large numbers of3: leaves and other pieces of organic material
made collection of mud by pipetting difficult. The samples
were preserved in formaldehyde, sufficient being added to
produce a final concentration of 4. %

-39- Sedimentation.
The rate of sedimentation of the suspehded particulate
= material was determined by the use of sediment traps. Each
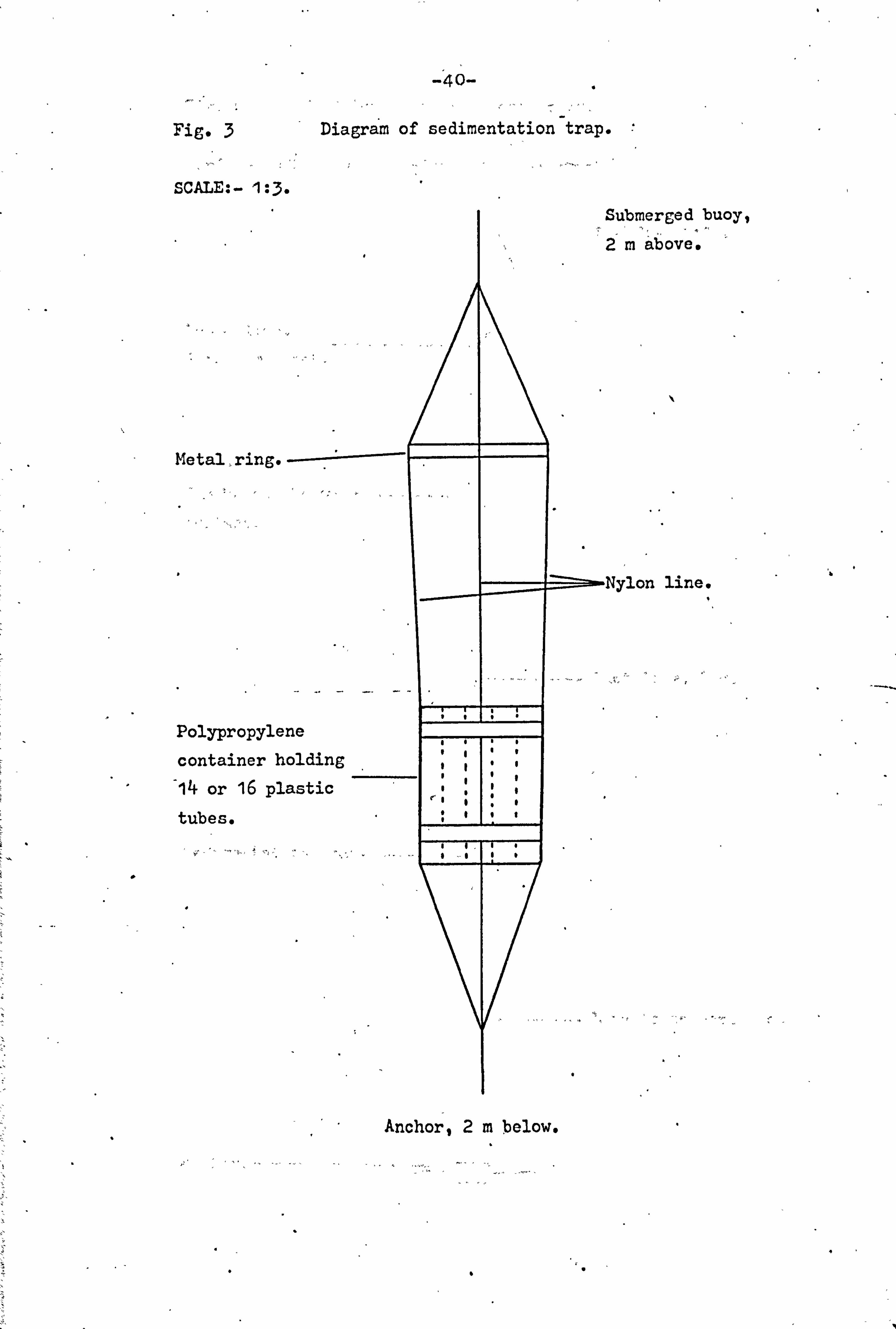
trap (Fig. 3) consisted of a polythene oontairier. which held
14 or 16 smaller plastic tubes of 14 mm internal diameter
" and 100 ram high. Each tube acted as a small sediment trap
and also served to reduce turbulence and erosion of the
sedimented material within the trap. The traps were anchored 2m above the mud surface in the positions indicated in
fig. 2, with a submerged buoy 2 ra. above the trap itself.
The trap was located by a small surface buoy attached to
the submerged buoy by a loose., line-. (Fig. 4). The use of a
submerged buoy reduced the effect of surface wave action to
a minimum. The positioning of the trap 2m above the mud
reduces excessive eontarninatiorl from the surface deposits.
As each trap was placed at least 4m below the water surface (ie. in a total depth of waterýof'6 m)'and`below the summer
thermocline, there was no growth of epiphytes on the traps.
When samples were collected the entire trap was lifted
out of the water and the tubes, containing the sediment,
removed. They were replaced with-n set of clean tubes, thus
reducing the potential accumulation of bacteria to a
minimum. Significant interference of this kind was not
observed, but on some 'occasions during the autumn a leaf
may come-to rest on top of the tubes. The now tubes were
filled with'alga-free (tap) water and the trap lowered, for
a further period of collection, gently in position so as
not to disturb the sediments. The trapped material was
preserved in Lugol's iodine.
The rates of sedimentation, using this type of trap
with straight parallel walls, has been found by Kirchner (1975) and Pennington (1974) not to be significantly different for traps of different sizes, unless the surface
area to depth ratio is high (i. e. in the case of shallow
traps). During isothermal conditions resuspension (erosion)
of material in thene traps is higher than in traps with a
collar, such as the Tauber trap (Pennington, 1974). The
linear correlation between the amount of particulate matter trapped and the total mouth area indicates that the "snow
-40-
Fig. 3 Diagram of sedimentation trap.
SCALE: - 1: 3. Submerged buoy,
2m above.
Metal > ring. ---=-
Polypropylene
container holding
14 or 16 plastic tubes.
Anchor, 2m below.
N
line. 'a
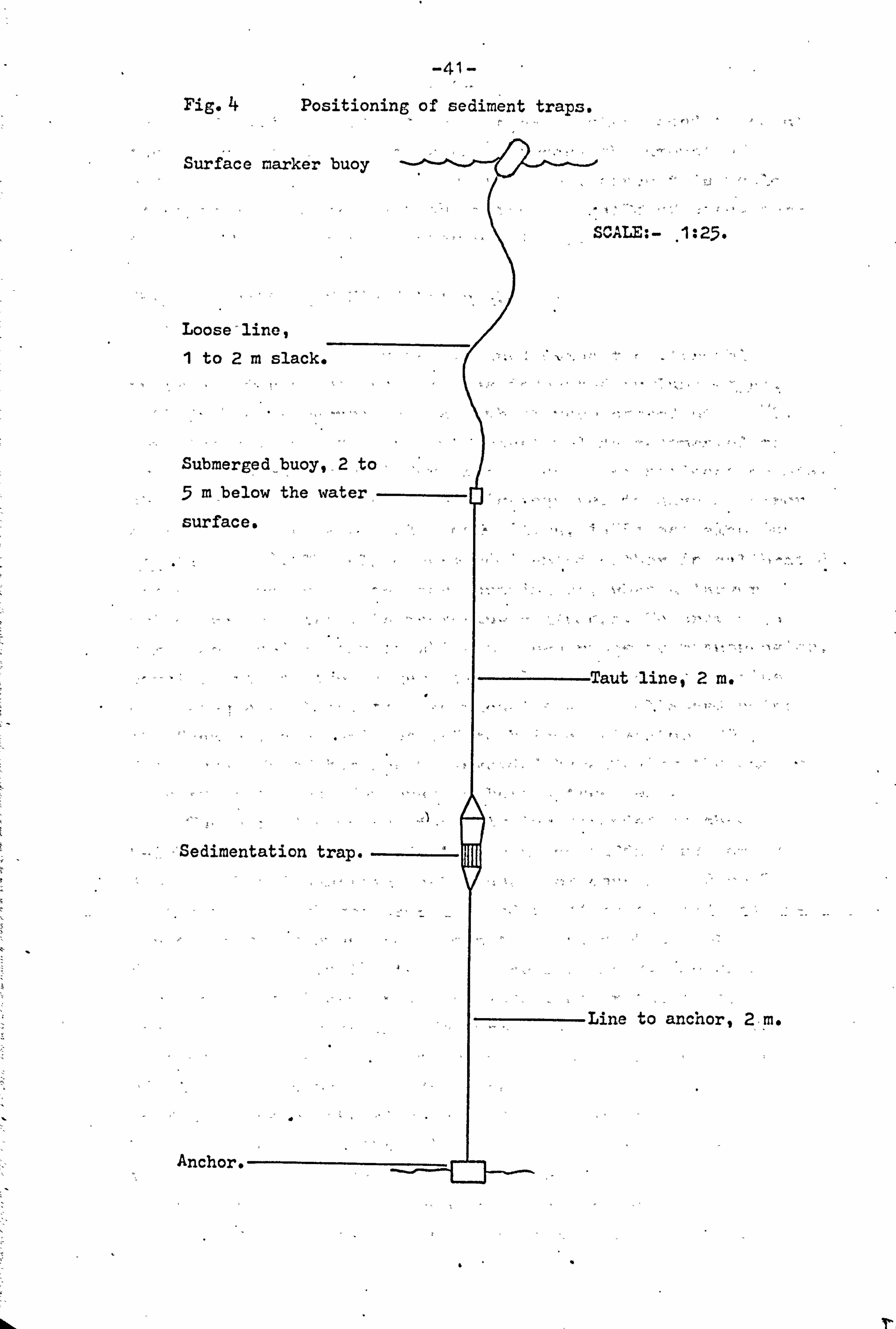
-41- Fig. + Positioning of sediment traps.
Surface marker buoy
SCALE: - , 1: 25.
Loose line, 1 to 2m slack. ti .
Submerged . buoy, 2 to
mbelow the water, - surface.
Taut 'line, 2 m.
'Sedimentation trap. -----
Line to anchor, 2, m.
Anchor.
ýý i"'

-42- fence effect" (the formation of non-turbulent conditions at the rim of thetrap which would increase the amount of -
material sediuented) is not'sißnificant, since this would.
be expected to increase with increasing ratio of cirounfer-,
ence to-surface'ar. ea (Pennington, '1974).
The Enumeration Of Field Material. '
Planktonip. Plankton samples were examined using the-Utermohl",
inverted-microscope technique as described-by-Lund et a3,0-' % (1958). The preserved samples, which were"storedrat, 10 0C,
were thoroughly mixed and sufficient volume sedimented to
provide enough material for statistically significant counts.
About 100 organisms per sample'is required-to"give an--error
of not more than' 20h" (Hobro and" Willen, 1975;, -see also Lund
at al., '1958). The algae were'sedimented either in"calibrated
sedimentation tubes used' for''oounting"or, when a larger
volume was required, in measuring`cylinders. To provide a
more rapid sedimentation which is'less prone-to resuspension,
particularly of the larger colonial gas""vacuolate, 'speoies
of blue-green algae, the gas vacuoles were collapsed"-uoing
the""häimer, cork'and bottle" technique (Klebahn, 1895).
The formation of bubbles was avoided by-allowing the sample
to warm up to room temperaturg before treatment.
The period required for settling-depended on the--
height of the vessel used, '1 or 2'h was sufficient for the
5 or 10 ml sedimentation, cells used for counting, " 2 or 3
days für a-250 ml'measuring* cylinder. After the sedimentat-
ion of large volumes the supernatant was siphoned off using
fine bore glass tubing to limit the flow rate. The siphon
was carefully placed in the cylinder, after the material
had sediriented, such that no turbulence was produced. The
loss of blue-green algal material tend'other large species
of algae was 'found to be low (less than "1 iö) , as determined
by filtration of the siphoned off supernatant through a Millipore membrane filter, followed by drying, clearing in
immersion (cedar wood) oil and e:: amination as described by
Mc llabb (1 960). A known proportion of the settled material
was then re-sedirientod in a cedimcjntation cell. A problem
kh, I

-43-
often encountered iihen algal material. had to be sedimented
in this way was the relative scarcity of blue-green algae
. in relation to other particulate material. Thus it was
sometimes neccessary to use several .` sedimentation cells per
sample so that what. was being counted was not obscured by'''
-other material. -Separation by differential buoyancy, '' before
the gas vacuoles were collapsed', did, -not provide consistent
results.
The technique. of sedimentine a , known amount of algal
materialland the scanning of the-entire surface area of the
sedimentation cell avoided the-problem of non-random distribution. on the-floor of the sedimentation cell. =The time required td examine'each sample was not excessive-
since the low power objective was sufficient-to identify
and enumerate the blue-green algal species studied.
- The results are expressed as-numbers of. filaments or
cells per ml. The standing crop per unit area (1. m2) of
water was calculated, by trapezoidal integration of the depth
profile of algal distribution (Lyon, 1970)°. - Frequencies of heterocysts, and akinetes were also determined. For this`a
minimum of 1000 cells were examined sequentially along "
filaments within' randomly chosen, transects-across-the sedim-
entation cell. Frequencies were expressed as i-2 of the total
cell number. 'a
After-use: thecells were rinsed with distilled water
andthe_internal surfaces were brushed, using a small paint
brush, to remove any adhering algal material-. - This method
was efficacious, as examination of cleaned cells, after
being filled with water and allowed to sediment in the
usual manner, did not , reveal the press ence of any algal
material. I. lore severe cleaning techniques, such as the use
of concentrated nitric. acid,. have been used sparingly when
required, since the cover glasses used for the base of the
cells were sealed with the epoxy recin''Araldite (Ciba-Geigy
Ltd. ), which appears to be resistant to acids for a short
period.
hhý

k -44-
Benthic.
The quantitative examination of mud samples was Rosenthal'!
performed by means of "a ° Mod-Fuchs,, ýhae. mocytometer. To
achieve any degree of precision a large number of sub-
samples had to be examined, due principally to the obscuring
of algal material, particularly akinetes, by detritus.
The samples were thoroughly mixed before subsampling
and diluted if necessary. Gas vacuolso in a small volume of
sample could be collapsed by filling a 10 or 20 nil plastic
disposable syringe and exerting pressure while holding the
thumb over the spigot. Enough pressure can be generated by
pressing with the palm of the hand'(Walsby, 1971):. Usually
it was necessary'to examine up to 10 to 15 ' complete grid
areas (18 to 27 mm3) to find sufficient material. Clumps of
akinetes were broken up by-shaking by hand a small sample in
-a serum bottle with 10 small (2 mm) glass beads for 30 s.
This relatively mild treatment did not damage the cell'
material present. '
In order to facilitate the identification of algal
material against the background of detritus, a Zeiss Standard
18 microscope with epi-fluoreecence condenser Model IV%F
was used, utilising the natural fluorescence of chlorophyll
. -a. This was effective; but many akinetes, particularly
those which appear to have been in the mud for a long
period, did not fluoresce' brightly. This may be explained {
by the observation that chlorophyll may be replaced by {
phaeophytin in akinetee of A. cylindricrti (Fay, 1969b).
Transmitted light microscopy was used to enumerate non-
fluorescent material such as empty akinete envelopes.
The larger algae, such no the colonial coccoid'species,
were counted using a Sedgwick Rafter cell, the use of which
is described below (Page 46 ). Although the mud had to be rs
considerably diluted a large volume can be scanned quickly
with low power magnification. Due to the difficulty in sampling from a particular
stratum of the mud core, the results are express>ed as
numbers per ml o± the sample. The linear correlation between
the amount of algal material and the proportion of solid
material in the sample enables a correction factor to be

-45-
applied. This permits a more precise comparison between
sanples. collected at different times. The proportion of
solid material was determined by settling the samples under
gravity for two or, three days in tapered, graduated,
centrifuge tubes.
Total Particulate Sedimentation.
Ten tubes from each sediment trap were randomly
selected and their contents tipped into a beaker. The tubes
were rinsed. out with distilled water and the'combined sample
filtered through a coarse gauze (1 mm'mesh size)`to remove
the chironomids which were sometimes present. These organi=-
sms are benthic, "the traps, which are below the thermocline,
constitute a suitable habitat. The filtrate, after rinsing
the gauze, was washed into a 250 ml centrifuge tube and the
gas vacuoles collapsed by a sudden increase of pressure as-
described above (Page 42 ). The material was then centrifuged
at 1000 xg for 15 min. At the and of'this time the speed
was reduced gradually over a period of 10 min to avoid
resuspension due to the vortex produced if the speed is
reduced too quickly. The supernatant was decanted and the
residue carefully transferred into small preweighed
porcelain crucibles. Ito'further'washings of the sediment'
were done, since the loss of''material becomes significant.
The remaining liquid zwar evaporated using a hotplate and
then the crucibles, öontäining'the dried sediment,, were
dried at 110 °C to constant weight. After weighing the
sediment was ached at 500 °C for 24 h in a muffle furnace
to determine the loss on ignition (ash weight). Conventional
methods for accurately weighing hygroscopic substances were
used throughout. 'Yreparation for the next batch then
commenced with-the cleaning of the 'crucibles, first in
distilled water and then in concentrated nitric acid. After
drying in the specified conditions they were weighed.
Sedimentation of Alea1 'Material. The. -remainder-of the tubas (4 or'6, depending on the
size of the trap) wore used for the assoosment of algal
material. The contents of t1ie' tubes were mixed, producing a

-46-
single sample-for each trap. The concentration of algae-was determined using a Sedgwick Rafter cell. Like. othexTvisual
examinations the gas vacuoles were collapsed to speed
sedimentation, using the syringe technique. The Sedgwick
Rafter cell, which had previously been washed in distilled
witer and thoroughly dried, was filled with slightly more
than 1.0 ml of the ' sample .' The, chamber was covered with the
cover glass such that no air bubbles were trapped. After
allowing the material to settle for 1 min sufficient
transects were examined so that at least 100 individuals of
the most abundant species were encountered. The-type of cell (Maxta Graticules, London) used is engraved with a grid of 1 mm squares, each square representing a volume of 1 mms- (1 Ili) which' facilitated rapid counting and simple
calculation at whatever magnification or microscope was
used. This was particularly useful when the field counting
technique (Woelkerling et "a ., 1,976)
. was used when algal
material was relatively, abundant.
It is assumed in the methods used above that the
entire content -of the sediment trap tubes was in the
process of sedimentation. Thua, in the assessment, of.
particulate sedimentation the entire-volume was centrifuged; and in the enumeration of algal-material a known proportion
of the contents of the tube was examined. The total amount
of material within the trap can thus be calculated, and
knowing the area of the'trap"mouthrand the time interval
, between samples, the rate was calculated in, terms'of
numbers (or g) per m2 per day.
The Chemical Analysis Of Field Snnnlec.
Those water samples which were to be used for the
analysis of dissolved'subctances were sterile filtered
through washed 0.45 Lm Millipore membrane filters either at the site immediately after collection or shortly afterwards (the maxima L delay of 5h had little effect on the
concentration of analysed compounds if the algal material
was not abundant). If these filtered samples were not to be
analyued within 4 days they were deep frozen, otherwise
khý

-47- they were kept at 10 °C until analysis: Using this method
the effects of algal and bacterial uptake of orthophosphate (Heron, 1962) or the release of orthophosphate through the
use of chloroform as a preservative (Fitzgerald and Faust,
1967) were minimised. All containers and glassware were
soaked in nitrated sulphuric acid (100 ml saturated
solution of sodium nitrate in 11 concentrated sulphuric
acid) and rinsed several time$ in distilled water.
Orthophosphate.
The, method of-analysis in that described by Golterman
(1971 ). In a hit,. hl. y acidic environment orthophosphate forma
a yellow complex with molybdate
to a blue compound by ascorbic
by antimon'y'.
(Reagents')
ions which is then reduced,
acid in a reaction catalysed
All reagents were made up from Analar (BDH, 'Poole,
Dorset) grade chemicals.,
Orthophosphate standard solution; ý40 µg`ml P04-P.
0.1757 g of dry (1 h at 1150 0C) potassium di-hydrogen
orthophosphate was dissolved in distilled water and diluted
to 1 1. A few drops of chloroform were added as a
preservative. "Fresh dil. utions. from this stock were prepared
for each calibration curve or standard test solution.
I1olyjbdate - antimony solution.
4.8 g ammonium molybdat e, (NH4) 6No7024.412O,
and 0.1 g % sodium amtimony tartrate, NaSbOC4H4O6, were dissolved in
400 ml 4 11 sulphuric `acid and made up to 500 rml with the
same acid.
Ascorbic acid, 0.1 M.
2.0 g ascorbic acid dissolved
water. Solutions were not used for
(Procedure)
Into a 50 ml volumetric flask
the filtered sample diluted with
nece: sary. 5 ml of the r o1'bdate - 2 ml bf the ascorbic ¬ cid solution
in 1 CU ml distilled
rsore than one week.
wao pilpetted 40 ml of
listilled water if
antimony solution and
were added and the
volume made up to 50 nl. The contents were thoroughly mixed
and the absorbance of the sample and standards measured at

s
-43-
least 10 min after -mixing , at . 882 nm against reagent blanks
using 10. min cuvettes.
she absorbance of the. lake «ater, zeroed against
distilled water, was insignificant at the wavelength used.
No interfering corpoundh3 (organic compounds, i arsenate)-
appeared to be. present, as determin ed by using internal
standards. The. concentrations of orthophosphate met with
were generally high enough for direct determinations,
requiring no, further concentration.
Nitrate and Nitrite.
Nitrite concentrations. wera determined by the
sulphanilamide and It-(1-näpthyl)ethylenediacline diazotizat-
ion reaction described-by Bendschneid*er. and Robinson-. (1952).
Nitrate was reduced-to'-nitrite usinC. the cadmium - copper
-reduction column method described Eby : Wood ct-a. (1967) .
(Realteints, nitrite analysis)
Sulphanilamide..,
5g sulphanilamide, I1i2S02C6H4NII2, were dissolved in
500 ml 1.2N hydrochloric acid and stored in an amber bottle.
= N-(1-napthyl)ethylenediamine dih. ydrochloride.. s -
0.5 g-N-(1-napthyl)ethylenedi6mine dihydrochioride,
C10H7NHCII CH2NH2.2HC1, " was. dissolved in' 500 ml distilled, 2
water and stored in the dark in an amber bottle.
Standard nitrite solution', 70 µC N02-M ml-1.
0.345 g of, dry (1 h at 110 °C) sodium nitrite was,
dissolved in distilled water and a few drops of chloroform
added as preservative. The volume was made up to 1 1...
Dilutions were prepared as required. (Procedure, nitrite analysis)
1 ml of the sulphanilamide reagent Was added to 50 ml
of sample in a 100 mzl stoppered measuring cylinder. This
was followed ' within 2 to 6 min, by 1 ml, of I1- (1-napthyl)
ethylenediamine dihydroohlöride and the absorbance of
standards and samples measured- at 543 nrn in 10 cup cuvettes.
A Bausch and Lomb Spectronic 700 spectrophotometer was used. (Reagents, nitrate reduction)
Copper sulphate solution, 0.08M.
20 g cop,. -or cull. hate in 1.1 distilled water.

--4 9-
Ethylenediaminetetraacetic acid (MTA) solution,. 0.11,1. 38'g of the tetrasodiura salt of EDTA dissolved in
500 ml diestil led water, diluted to 1'1.
Hydrochloric acid, 211.
85 ul of 12N hydrochloric acid diluted to 500 ml with distilled water.
Nitric acid, 0.311.
10 ml of 15.411 nitric acid diluted to 500 ml with distilled water.
Hydrochloric acid, 0-001511-
0.125 ml of 12I1 hydrochloric acid diluted to 11 with 'distilled water.
Column wash solution.
1 ml EllTA solution in 50 ml 0.001511 hydrochloric acid.
-Mitrate standard solution, 70 Ilia 1403-H. ml-1. 0.1264 g dried (1 h at 110 °C)
potassium nitrate
dissolved in distilled water and diluted to 250 ml. A few
drops of chloroform were added as a preservative. Dilutions-
were prepared as required. (Procedure, nitrate reduction)
Coarse cadmium turnings, already prepared, were washed
in 211 hydrochloric acid and rinsed with distilled water, "
washed in 0.311 nitric acid, rinsed in distilled water, and finally washed in 211 hydrochloric acid and rinsed in
distilled water again. The filings were treated with 100 ml
of the copper sulphate solution per 40 g of the filings in
a 250 ml conical f -lask stoppered with a bunk; through which
passed a wide bore glass tube. The treated filings were
washed with distilled water such'that they were not exposed
to the air. she column (Fig. 5), plugged at the bottom with Pyrex
glass wool, was filled with distilled water and the filings
tipped in via the tube, again so that they were not exposed to the air. During this process the column was tapped to
settle the contents. 50 rnl of the wash solution was run through the column and was then allowed to stand for 24 h,
renewing the wash solution several tines. Before use the
prepared colurina were conditioned by passing through 31 of
solution coiitnininj; 840 Eta; NO3-N 1- and 20 ml 3DT,, solution

-50-- Fig. 5 Column used to reduce nitrate to nitrite in water samples.
Redrawn from ', food et al. (1067).
SCALE: - 1: 2
50 ril reservoir
Metering /stopcock
" .-..
spy
Copperized cadmium
filing3
-ý Measuring
cylinder
Capillary Pyrex glass wool plug tubing

-51 -
1_1 To each 50 ml of-sample to be reduced iras added 1 ml
EDTA solution. Excess"wach solution was"-removed from the
colurin by a pipette to about 5' mra above the top of the
filings. A small-aliquot of the sample was added to the
reservoir to rinse out the previous sample, and"1 to 2 ml'
allowed to pass through the column before removing the
rest using apipette. The remainder of the sample was
added and ameasuring cylinder put under the discharge-tip.
About 5 ml was allowed to pass through the column to rinse
the cylinder, and 151m1"ýcollected. A consistant flow rate
of approximately-1-drop per second, adjusted using a stop
clock, was maintained for a seriesofsarnples, standards
...,. and ý blanks.,
The reduced samples were gnalysed for nitrite as
described above. 'Por each"eeries'of samples standards of
known concentration of nitrate were also run through'the
column to determine the efficiency-'of reduction.
It was found-that the ' nitrite` levels -were very low in
the water"bodies sampled and'therefore analyses of nitrite
were only done-when the concentration"would be expected to
be higher 'than, normal, such- as *, in the "anoxic reducing
conditions in the hypolimnion. Contamination of water
samples"'by nitrate contained-in cigarette tobacco (Taylor,
1975) would be minimal.
Oxygen.
Oxygen concentrations-were determined using the
Winkler uethud described by Golterrian (1971 ) or by using
an Electronic instruments Ltd. (Chertsey, Surrey) model
1520 oxygen probe. (Reagents, Winkler)
Potassitun iodate, 0.100: 1.
3.567 g, dry (1 h at 110 °C) potassium iodate dissolved
in 11 distilled water.
Uodium thiosulphate, 0.02511.
6.2 C sodium thiosurphate dissolved in distilled
water. A pellet of sodiun hydroxide wan added to stabilise the solution and the solution diluted to 1l und stored in

-52-
an. ariber bottle.
Alkaline iodide solution. 250 c sodium hydroxide dissolved in distilled water
and 75 L; potassium iodide added and diluted to'-500 ml.
Polyviol indicator.
A saturated solution of polyvinyl alcohol in distilled
water. Manganous sulphate, 50iß.
250 g : iancanous oulphate dissolved in 500 ml distilled
water..
Orthophosphoric acid, a. g. = 1.75.
(Procedure,. Winkler) "' Samples were collected in calibrated 60 ml stoppered
glass bottles such that no contamination by air or trapping
of air bubbles occurred. 1 ml" of the manganous sulphate
solution and 1 ml of the alkaline iodide solution were added
in rapid succession to each bottle using syringes. -The-
stoppers were replaced and the contents mixed, and mixed
again after the precipitate-had-settled-to the lower third
of the bottle. Before titration 2 ml orthophosphorio acid
was added and the bottle resealed such that none of the
precipitate was lost. The entire contents of each bottle
was speedily titrated against 0.02521 thioe-ulphate. using a
rzicroburette (10 ril) and Polyviel as the indicator. A few
drops of the indicator (orange to colourless) were added
as the end-point (pale straw colour) was approached. The thiosulphate spas standardised weekley by titration
against the standard potassium iodate solution. 0.02511
thiosulphate appeared to be relatively stable over n
period of up to two months.
Calculation: - U
8000 x rlthio x titre
2 Vol of bottle -2 nil
Nomograms (Hart, 1967) were used to determine ý--, saturation.
pH. she pI-i of the surface waters was measured on site
using aI etrohm battery operated" pII meter. Olotrohm. Type
L'280 n, IlerKoau, 1,4witoorl-rid j.

-53-
Conductivity.
An Electronic Switcht. aar Ltd (iiitchin, Englund) type
r: C-1 conductivity bridge was used and the conductivity of
the surface water expressed as µciho cm-1. - .
During' the summer of 1975 a Bach (Ames, Iowa, t7. S. A. )
Direct Reading Engineers' laboratory, model D: l- L/2, was
used in the field for the chemical analysis of: water
samples. The results were comparable but less precise than
those obtained using laboratory methods. The analytical
procedures are derived from those described in-the American
Public Health Association ; tandn, rd'rilethods (1971) .
The Measurement Of Physical Parameters.
Temperature.
The Ruttner sampler contained an accurate mercury in
glass thermometer and the oxygen probe was automatically
temperature compensated; the temperature readout could also be obtained but to a lower degree of precision than the
mercury thermometer.
Light.
Light penetration was estimated uainCj a Griffin
Environmental Comparator (Wembley, IIiddlesex, England), the
sensitive element of which comprised yf a cadmium Sulphide
photocell encapsulated in a glass bead. It is most
sensitive in the red region of the spectrum. During the
summer of 1976 a 300 rim diameter matt surface aluminium
Secchi disc was used to determine transparancy.
¶olal Particulate Suspended Solids.
An aliquot, t eneral? y 100 to 400 m_l, of the preserved
plankton -sample was filtered through a washed, pre-weighed
25 cam Gr/C 4hatnen (made by It &R Baleton, Englund) Glass fibre disc and washed with 100 nl distilled water. These
were dried at 110 00 to constant weight. The length of
drying period did not affect the weight significantly once
constant wei ht had been refached.

RESULTS
The Chemical Aid Physical :: nvironr, Lent.
`. temperature. - Distinct effects on thermal stratification and on
water temperature of the different degrees of exposure and
of the different surface area: depth ratios, in thetwo
meres studied, are shown in figs. 6 to 11* Thermal
stratification in Kettle Mere is already well marked-, by
April and renains-stabily, stratified until October (rigs.
6,8 and 10). The period of stratification i"s considerably
curtailed in White Mere where it prevails generally' only
between June and September (Fins. 7,9 and 11). The
difference in stability of the, water'column, between the two
meres is also refldcted by the higher surface temperatures
reached and the more sharply defined thermocline in Kettle
Mere. Further, the epilimnion'is shallower than in White
Mere. Although both'meres are exposed. ,. to, periods of, windy
weather causing the cooling of the surface layers of water,
mixing . in White-Mere generally extends deeper, -, and may
sometimes disrupt stratification during the summer, -as
happened for example' in August, -1975 (rig. ' 9). --
The water temperature during isothermal periods is
similar in both meres, although on the breakdown of
, stratification in the autumn, there is a more marked
cooling in Kettle lýaere when compared' with White More, due
to the mixing of a relatively large volume of cold
hypolimnetic water with the warmer waters of the shallow
epilimnion.
Oxygen.
A similar relationship between exposure, turbulence
and the vertical distribution of oxygen is also obtained
for the two years (1975 and 1976) that data are available (Figs. 12 to 15). The water below the thermocline in nettle
Paare remained alriost totally anoxic for several months
during the sLUiver (Figs. 12 and 14). The 1,; ) saturation
"isopleth may approach to within 2m of the surface during
calm, periods, which, will result in . considerable
, oxygen
depletion of the surface waters, should a relatively minor

-5! -
co
C' r1 " (U c- ". l C) 0
4-t tr 0 i g
Cri ii 4- "rl P
CJ C, C4 '. 0 C) V E CL)
r1 ". -t G) -W 4J 4-)
tý aý S 0 "r1 Q) Cu 4ý C7
ý1! 1f
ocý
NO CN co
.
tu "uI(I0Q
O
H Ü
C7
ti
a:
tl u.

-56-
o'
co
c- $..
H C)
lam- vC 4.3 +) +) +3
"ri . i: 4-4
iW :=:. 0 H
:2
J
G7 41 4) r. 0
7
Y H N
2
H
}
a zý r4
CIZ

rn -
V1
r" co
r"
Mr
cc
c- C: r
r- 00 I'D
It cl-
/ Lr, L! C'-- u C\ r4. " C) r "-l a) 0 ti
C) F+ 4- "rt s4 w r n) $.., c)
Ca r . "r4
" 4-) . a> . 4-) - #.. 5 .. ._.. t: . 43 S. 0 ". -I CI C L4 U. W :4 . 0 N
-a--
CN ýl s\
v
rid
uj
tiv
. C:
0 1
r=.
ý7
-D

-58-
N1c"
Lrl%
0
N
N--
O
" C7
- 0 c - " tJ
T- t«. to O : $3
o G. S
Q) r4 U E
C3 "ri $ý n
.ý. c w Q :4y ' 0 I-i
Ia
.. a
U
F+ U
C: J
4-1
0
F:
. =7 w
0 c- c" iM -2- U1 %D L` Co C" cz-

4) t9
m r4 " Ca c- "ý U0
G ;3 C) S4 d-) "r4
C) S, to r4 N £3
Cl v C) r-i "rf +C) .C
b: +ý Lr O "r4 4) G> 4ý L7 GV 's : -' OH
A ýO rO co Cý! RI CJ rr
Nr
Lrl%
O 00
L'\
0 T- (IQ 1 .*
u
-59-
1
a
0 <s
a
ti a
a
cý
c1 ö
7 ti
:L
ct

ýi
0
Pi
rD
0
9"
w 11
c^ý
Fý. P)
CN

a
io try
PQ
ýBJI \ý
o u> N
0 L3 "r4 .r CW
C r. -+ a a, ". T- fm 0 (0 u; '
.r Li +) 0 - " to H r.
ý4 IC " 0= ºP4 r. C P P4 O
r-1 C) Q> "'i cis 13 . 4a £: . º)
r W 0 u cn
t. 3 Z. r+ Cý Z, 4+ 0 Z' cm.
0
U O
O
G=,
O e- N tr1 .ý LrN lG
-61-

-62-
0 n
0
0
o
Lr\
o to "-, .1 cý m y r) h Z r a 4-4 co
0 0
r 4-) i-) 0 ce >C U
ý 2' r 0 r' 0 + F s " . 0
C) r+ C C) "r4 C C) KN «_ C L 0 -) r U U C) C3 t, c0 U C3 Cj r+ X C) V L. U c3 L.
+) 41 0 i: ". a 4. ", S. c a zý ti u ±. Q) 44 0 X cri a C) s .) (L4 > 0 U 4) Li 4 " -4 LC to
i
i
a
c c
v W 0
a z
s, a
5 t
0
ac r
co w LL-
Z ti
orn: - rn a .. ý uý ;. cx C. c z-

-63-
L
o W . r4 . C) .. 3 .. 3 co
o, r3 p 0 C'
. o co C) aý U; U
Cj "ri "r1 C) "
c, r_ P. o E c t +31
- 4) 0 r r. 4 "rq
X C) ý + f
4. ) +0 0 S. Lo
I
V. a)
. rj 44 0 s'.
0 o
o
ýÜ
C r
0 0
n
C \` Jý
71- W
i
N
41 r- 0
. T!
C7ý W
a ._
a H
ti
QrN trl I--" tC
U' %4U n

-64-
Q
" 4) 4, c0 Q> tai lý 7 ri 0 4) A Cb
ON i4 0 CO a) t/! £i 0 r 4) 6) +. U <. G) "r"ý N
L! N 0 f+ N ,> U "" "rA "ri Q) " a) Cs R. "
C) 'D " 0 l. v f:. , C: E Q) fr P. 0 +) P, O 0) H N 1 "rl V. 4) rl
hfl :z tý hJ +. 3 U - O +> d 4-1
" 4) +3 O 1:: c. "'i 44 0 V. ) "rl St c; C) +) 'L7 F. 4-)
"ri C N 4-+ 0 . S. cß Z 0. 0 cd j : i: > 0 0 i-) Q "ri "rl U) to
m
a- h
c
_i x
0 00
; -ý .ý ý*.
ýý
0 r fU týl -t (. vJ ß" c
r,
I
H y
L

-65-
amount -of mixing occur. This is also observed during the
breakdown of stratification in the autumn, when a large
volume of oxy`en depleted water is mixed with n relatively
small- volume of saturated or dul y-satursited water, which
results in a low percentage saturation of the water body
as a whole. Saturation gradually increases during the
winter º1onths to about 7U; j', before stratification is renewed. Periods of anoxia in White !, ere are limited to the
deepest areas during the most calm conditions (Figs. 13 and 15). The depth of water column which is fully saturated is
greater than in Kettle Mere, and the reduction of saturation during the breakdown of stratification is less marked.
Although periods of supersaturation occur in both
mores during the summer, in White Mere these periods are
longer, a greater depth of water is affected and the
maximum levels of supersaturation are higher.
Light.
There are considerable differences in the penetration
of light between Kettle More and White Mere even during
periods when populations of planktonic algae are low (Table
2, Secchi disc data for 1976 in table 6). The considerable
light absorbance of the water in Kettle Tiere is principally
due to the high level of dissolved organic compounds, which
impart a brown colour to the water. The absorbance is
particularly high in the u. v. and blue regions (200 to 400
nm) of the spectrum and is twice as high in Kettle Hero
than in White Mere (Fig. 16). Kirk (1976) reports that
. dissolved yellow organic compounds ("gelbstoff") greatly
reduces the amount of photosynthetically active radiation
and that the blue region of the spectrum is extinguished at
moderate de, »ths. The reduction of the depth of penetration of light
during periods of algal abundance is more marked in White
Tiere than in Bettle !,: ere. The greater absorblince of the
Kettle ere water tends to damp down any further fluctuations
of light penetration due to chancef-, in tilt-, till,
which, in any cane, are generally higher in White Fiere.
kh,

-66-
.". ""s
4l
"r+ -NNN \O .'r
l c- N, \ MN K1 N N, % N --d" n ON rN
fý N . 1- ON 00 _: t u1 ttl O 0
N c- NN K1 N. - Ký
N
+) O 00 O tt'N 0% O- ur . a-) ". """"""
C) rrOrrrr r- r "
0 . r4 43. ct4
. rq
CIS 4-) Cl
\"\N\Ö
N 0. ̂ N t11 N 00 N a) C. ) cö 4i Q) P4 f+
. LrN
t0 C` ONN0
:3 a) 0n ***"tt1 ýO t! 1 N'NNN
a) co +2
a' u1 O rý OOOO1 O- c- N -7 M Kl rrrrrr IA N
a\ C- ONO Ln ON co ulý -t ao OnN
f: rMNM4 c- 4NN. NNN N
OO .- . I- ON OOOO u1 OOO o".. NrrNNr r- O r- -r4 cV "r
4-4 0
E vd N1 ýO N- co co CO 00 O cam- N . s: 4-3 \ ý1 ýO \ C` \\\\\ r- \\ ý- 4) Cl -I- \\ Ul \N 03 t+1 \` u1 \
AN IA NNNN c- c- NN /O LN
03
N $4 IA co 0M0N u1 0"0
N GD ý: "t""". """"I" Ul U. ' U. ' Kl N. ' ýO %Z Uý
H C) v 4-1 8 u1 0 L1 [- t, 00 L` Q01! 1 Elr- Pý N c- rr- ý- rNr
Q. ' r 0)
u1 N0N0OO UN ýý ""t"""t"t""" c- N e- 1\ ý' NNNý
Q: H 4) 6 to OOOOO u1 OL-\ Q', OO
rrr c- 1- c- c- 41:
rO c- c-
o e- cv
A r' rN Qý r K\ r (V Q ýN Nr
0 N 0 0 rl v4
a

-67-
O C)
O O Co
U 0
0 0
8 .+1 E'
F:
. L: ; -,
s~
.l o ^J C) : -' _:: t. :J
p 0
0 0 n1
C)
kh,
ý! ) ý,,; -r, ' ". 10! -, "tIy

-68-
U1
a'
4-
H to rz w
to N Si 4)
C)
. Li i) 0
. r{
C) 4J
A
G7 O
,Si
O
P4 O
4-4 O
91. O
cd
M 0
co H
I
O1r C'" r 00 OOO LIl r %. 0 Co [` C' M0 K\ N [` Ol%
8 Co Co
4) OOOrrNN -3 Nc r' O
OMO ul CO cO \. O O %, O - Kl ONr N Ln 03 ire rn 0 ý-o 00 (Di CO c- 4.3 :
". -1 OOc; O c; O. ONOr l"- l l! O j2 s
CU CO w co N rin CO Z- N . ̀moo 0 0ý 00 ,0
OOoOOO r" rrrrOO
OOWO Co OOON %IU u1 L` i~ NMr [- L` U1 N 10 Q' NN Lý N
Nrr c- N Pal -OcfrOO
C) 0 U1 O\ C% 4- 0r0 (` \O %IO U-\ L'-
+ý rOOOOr-rrJOOO C) '
OO- U\ O oO "ý- OL\ ýU l` C'- LI\ öN K1 r\r\- O' C- ýO uý ul C-N
rr (5 OOOc; 2r r" CGOO n e-% v
cs- rl t
H9 Ä+
"H1
ru O LI- AS 0v ti
9-4 Ei 4-4 d) 0 O -P S
, sl O
, ll Äi 0y
N0 AS
rti -rf
11.0 C`- CO CO CO W 0' Or cr- r
d) 41 WN. %Z ce \ \,
M\Nc h\1 \\
c ul ON C- NrrNNNNr 03 ,t pr

-69-
N rn
4-
H to E a ca O
(D
n) 43
a) 4-3
P, ca 0 p4 0
s-4 0
0 0 0 4) Cd
0) U
O U
_: t 0 H
cn N
Nz
., l
e
a) C)
4-3
C)
0
0 H C4 s
4i 0
. c: N
"r{ 0
W\ 0 U1 NM -t 03 00 N- M0 r' t\ NN _e LA r4, \ _e n -i- UN
000O00000 c- c- %- r- c-
Lr\ N c- 0 ý, O WN /D 000 Lr\ %, D 0 L- 00 _t .: ' -i' In NNN- -4- Lf f\ t! 1 C` \-C co OOOOOOOOOOOO3O6
- C\ N c- ur n00N (v 0 h' t4\ M r1 rn NNN -4- -7- -L U\ NN 'N OOOO0OOOOOOOO88
ON Cý LII- _zt- 0 Nl 0 [` - CO Co rn C' cv ýD \. 0 w �O \. 0 N r- CO ý CO CO CO rn rn
............... 00000000000000 v2
n ul N L1 N1 NnU 'N 0N IN .-0 ý-O C` 00 *1O N I'0 N ,D'. O %Z C` \O [` Lam- '. O '. O '. O
" .0.0.0.0...
0.0.0.0.0.0". 0 0000
cl-O 'O c- OO co *r NOI '1 00 O w CL) t` ur ul% '. o L\ u\ u-N 4- -I- . -- . .0.0.0.0. o. O. O. O. O. o. O. O. O. O 0
CO NWN1O CO KN
cam- N
u1
I-N
0
H o el- U
E d) 0
{'3 30
"0 4-i o N0 Id 10 (is ti
w

4
f t
-70-
n cý uý ýo '. o rn 0 0 GO
O O 1
0 1
O 1
L w Oý N N r O '. O Co r , z, r O
1 0
1 "O 0 O O O
Z "ri
co ýD O Co £ O
O "0 1 1 1 " O
H
EO N O Ö Ö 0
E O O 0 O O
O N
ý' N
- I\ N Ö
N Q) N\ " " " 1 1 1 " O O O 0
x N N ý r
N F
O . " 1 1 P
"
r
. " O O O O O
s, a) H 4 O
ýi U) O
"-I +H
"F) O cj
-P " Q) - G) "r4 d) U cQ Ga O
U
\ - 'ý
n "0 N N
Co r- N
rl tý \ Ul
\ o - , cd
S] Q N c N r CO
H
OD 1= ö 0 " " I 1 1 1 " i " O O O O
N CM r ý " 1 1 1 1 1 " 1 " .. 0 O O O c-
CO CO CO -I" r4, \ r- c- N r- c; r O O O O 0 O
O
0 A M CO N N N
u n �
Ul Kl [- O O 1 O O O O O O O
0
M tr D
0 0 0 Q O w
r n N ý N tt $
i 1 ÜIN O O O O O O O
I+l
fü
O U
iy G
.r
4-1 O
O
rd
7
\ M \ N \ C cý oc, O" N c0 J ! rl N - N c r ýý
k

i
-71-
t i N. rn u1 ur\ CO O N - t: c y\ I 1 1
C` ýJ Cý 00 Co all Co Co 03 CC (J\ Cý
U L` 0. ON >-a
vi r U r SI
" M ßr1 O 1. tr\ c'- N N r d Cý CU vý N' CI ýO N d) S-i rP I 1 1 1 I I I I " " " " " " " zý S-( N U) , PCl 4- 4- PA N r r g
O v 0) -P H
S: "ri
G) O "r1 tý ýt -d- l- L` N- O '. O 0 O oo 0 l! l\ 4- P. o 4-1 0 Cl- CO O' tS\ M O' O\ l` 1 CO l- I CO lý- U 0 N N N N N cat N N N N N N N
H N O U rý
4H t- F"t
1 Ul
9 N Ul Nl 00 O tf N U1 r4, \ - o 4 1 1 1 I + N Cl- lý l- Cl- C'- CO lZ O CO CO t7 ý O 4-1
C o v
'T' SI
p"+ Q: 7i O S-i N- tI\ ü1 r- u' U"\ lý-
++ - c3 O\ '. D L'\ Co 4- 0 lý "ri SI d) t. 1 I I 1 1 1 1 I " " " " " " "
W :, c c- c N N Cd c r{
U)
-p G) U U H i"i
u) +' H 2S "ri i-> $ q aý o x Y,
U "rl -P . L' "rt
b ri u1 ul cam, ) N to a. ) N N h N lc\ U'
\Jp 0 +-2 (x) o O" O N UN '-0 C'- 1 .O 1 %Z I lý- (0 c-- N r N c- v- r- c- r r 'c U
0 H cý 'ü
ci ?a O H .D U
" L) O \ N 03 N lt1 CV IJl r N ui c- % N N Ul

-72- Orthophospii. at e.
Concentrations of orthophosphate were'high throughout
the period of investigation, although during periods of
stratification there were signs of depletion in the surface layers of water (Tables 3 and 4). Concentrations in general
were slightly higher in Kettle Mere than in rihite Mere. The
high concentrations sometimes measured near the bottom,
particularly during 1975 (Table 3), are probably a result
of contamination by sediments of the sample taken.
%
Nitrate.
Nitrate (Table 5), like orthophosphate, appearsto be
present in abundance, and no marked depletion was observed during population maxima. The concentrations, however, are
generally lower, and since mdrq nitrogen than phosphorus is required for growth, particularly of non nitrogen-fixing
organisms, this element is more likely to become limiting
than phosphorus (Gerloff at al., 1952).
The Annual Cycle Of Growth And 3ucceeeion Of The Blue-Green
Algae
The Plankton.
The sequence of appearance of the principal bloom
forming species of blue-green algae (abler, 7 to 12)
generally conforms to that observed in other temperate
eutrwohic lakes (Hammer, 1964; Reynolds, 1'971 ).
A. circinalis and A. floc-a u. ̂ Eo frequently produce large
populations, particularly in White Ziere, in the period
immediately preceeding thermal stratification (Tables 8,10
and 12). There is some indication that the populations of
A. cireinalia develop before those of A. floc-aquae. This is
particularly noticeable in 'Iihite ;. ere with the collapse in
mid :,: ay, 1976, of the unusually large overwintering populat-
ion of . º. ci rci nalis (: `nable 12). . 24nall differences in the
succes:; ion of algal species are frequently obscured by the
bimodal distribution of algal numbers with time, a recovery
of p? nnktonic ponu1ati onH oec: t: rri ný; a month or so after the
initial oollFajlse of a hloori population. The increase of
numbers in Kettle Nero npT ears to be halted with the onset
i6,

-73-
Pl w `
"ý 4-3 cý rýl
v 0
/i 0
W U r1 ci O 0j
rl O M O rl U F4 " O
d N
-N U]
N 04 d) £
rAi c$ d %lD r1 H " vý
4H (u c3 4-+ r1 +3 0 4-H S., "r+ O 0 0 M fA 1 x- Q) i-4 0 UJ
U .Q . "
:J 4-1 1 Q 0 0
z ý- " fti C` N ON
0 1 O C.
Pl P4 H 4%
O O a) Pi Q) f-4 M
P-1 ý rt H
4-3 E N +) O -p 01
H Z to 0 "r-4 0 a) s4 4-3 'LS r M R r+ 0
. z. ro
co 0 4-i H O 4-i
4-) "
O O
Q r-i
%10
N1
to . rq
U kj
Ui
s
ni r- _zr CO OOOOOOO c- r t- O
ONOOr cU c 000 c- OOO c- N r-
o0 u\ 000 cv o .f -J-
C(O)N COO ö ni
OOO%. p NrM c- COO00000
80 NNOOrOOOOOOO
M Cam- u01 r Iý1 N000000
000000000000
N 000O, 00000O00
000000000000
o00000O00000
OOOOOOOOOO ,OO
OO9O -f OOO 0-: I-0 CO r 4- ON r t! 1 00
U-N T- o0000o0Ooöc; o
co No oOOOOOO O- OOO
ý1 t! 1 O OOOOOOO0ONO6
0 N %lO QOOOOOOOOOO
OO 1 ý'
rOOOOOOOOO
O Cý Ü tIl r ý: . C)
cU ul O N Pal 0 C0 Q c- 0 0
ý - c- N CO co Q-, r j \ \ \ \ Oý \ \ \ a) \ \ ('D C) N' t: A \ C-. Q V) C) r r N CTS r I' c- Chi Oti N N r

-74-
Im r-H E N C)
0 H 0 U
0
Q) ii r C)
"r{ " Qi "r 4 Ui
4-i N 4-i ri 0 tM ä.
O 0 i H N $4 0
"0 Q) U
0 0 . - z
O ". .o 4-) a4 E n c co
$4 a, H I ý.
P, 0 ri P. 4) E U) r1 0
c' ri "r1 "(l
c ,^ VJ
H . c, RS P U rl O
Q) 'C7 U d- "r-1 Cd d
: r-I
Q
co c^
4.1 <4 0
-FP
0
00 cl, H . fl cj Eý
ý. O t<l try o C. -
to M O" O N O c- 0 0 O c- N r O
u, cT ac 0 ýD 7
U) CO m " . 4. O . .% O O NN O a (\ 0 cv -ct O 0
U 0
O c " L` N w . c ;V K%
O O O o0 -ý N O c-- c- 1 t+\ O O
aý f o N O
pq O O O O O O O 0 o %Z 0 C0
ü; 0 -d- --J- o N
Z O O O O O O O 0 u\ O 4-1
0 rý1 N 0 O O O O O O O O cC rý1
Ö
O O u1 110
cd pq 0 0 0 0 0 0 0 (z 4 O O O
N O O O O \O O
r+ O O O O O N w u\ c" O ý- O
0 O 0 O N 1.0 K\ a 0 o 0 o O o N N. CO U\ c 0 0 4
ul\ N N ý-
a) PQ 0 0 0 0 0 0 0 0 0 0 0 0
"r1 v " O ý-4 0 O C3
O O O O O N c- r \O O O O O O
(ý rl r o
l` O O O N Cý 1
O O O O O N
w N- V- O O O "
3 Ö 0 0 0 N- 0 4)
. . . . " %o cl\ 4) "1. ) O Gq O O O . '- c- CO O Lr, \ O O O cu . 4) ")
4-1 v 0 c. Lo
1 "' + ;. O O O rN r-
O r
c r 4 N N
O H c1) 4
i Lr\ 10 0 r- rl(N 0 -H
" N- O O 0 " r tý r t6 O O O O V- M N O N U1 L`\ C- C'J
(D .) "i") r
O lr O w Q O . ' " M
"14 m 0 0 c- c- N ß" 0 0" \, D O" N\ r 4i 4.4 H
Irj 23 k-0 " Cý w
N %lD N- N. r UJ 4-, 4. ) v M 0 C. Gý N\ cc 4' N 0 " N O U ý-i . i: 0 0 l\J VD CO c- c- N 4. CC .± H 0 C>
0. r-i H U E- H H
pý co 0 0 " LI\ lD r(\ 00 i) '. D O\ lf\ G) U U
0 rTh 'O o - o' o r: O 0 O O -a rý tý\ \-O r\ -Z N c c c° -, t U ü] Q U (1) (L) m H H
[-_ co CO C) G1 CQ C` ý \ \ \ Oý '\ \ \
ý C) cj \ . .
L-1 i-a c- r- N
U" c- N
l r i1 G\ N Cv r
0 Pd

-75-
t- oo P4 00 _: t N Uh N r U
' C(D ý Ä c- O O O r 1 N
c N O O O
` 11-0 N N +p C N of c- n C D" c - 0) " " . " CO C% -j- c- c- " "
r+l O O O O c- -t M rýl rfl K\ O O O O U O
0 r- 0 rn 0 " ". n . - -t t4, \ 00 0
>; O r 0 c c- N u\ -4' C` M cJ O O O O
r-1 E
cn ýo a o 0 0 o a o ýn o o O o o O Ü N
0 : 0 1,0 0 c- 0 ö
rrý c- O O O O O Ö rýý O O O O O O
U
ö ,ý O M
ý uý v) ý: G cý c-
N 'O G a O O
RJ N O O O O
a) N
N r r-i . 1. *ri
)U r4 '. 0 0 0 0 0 0 0 0 0 0 0 0 0
CH CH
w 4-1 r 0 0 0 ". + M cV ý- O
fý 0 c
rn O O Ö O O O O Ö o M O O O O Q) Q) U C7
Z 0 ýý M Cý S N O i1 ý
z N l 1
lf l >~ A 0 O O c- r- -t c- N Z- N N O O O O
O - N Pl
C) 1 '-O O O O O O O O O O O O O O O 1 A °
a) u> c M " (A O M o o cý ö 0 0 o rý ö v o o O o N
gi r1 ri C) M " CO
M U , 0 Orn M 0 N U1 ä (D O O N O M O Ul O W O c- O O O
Cl) 'i3 V
Z %
\Z O c- d O O 0 0 0 0 0 o 0 0 O r-4 9: Z
C) 0 r M O ul O O O O O O 0 O O O O G
4a rI 0 4t
+> " C- Q
O O n 15 O N G O O O d O O O O 0
v1 " "-+ w 0 0 0 0 0 0 0 0 0 0 0 0 0 0
O' C)
r-I "ri ýO c- N
vi $
M O O O O 0 O O O O O O O O O H
U
O c- cm CJ G` 0 O O N O G O d O t7--
, "i. 0 +' C) \D (ý. CO Cr 03 CO C) ` r" CV
ca " t Cff G" (\! C"- ;u %- c- iv tv tG N w

" -7E-
p4 ... . . . .. . UI pq OOO O- O OM c- lý ýD O
4 T- O .. 7' O O L1 4. O
OOOO '- O M Oý ßf1 N O O U
o N 4. O O %D M
;i0 O0O0O 0O M C` a0 O O 0
r
to 4) Q
O to r-4 0
Ö
t! ) " d +-
ri c4 cd rq r4 4.4 -r-4 to i4 4-4 a) Cd
4.4 ri "1-) O O H
f~ rl O N iq O 1A
, fl N U
4.4 ,:
5 0 IA
Z " .l ON
1 N c- O "" Sa +1 9 "ý n
u i z
M O r
1 Z
' L' P, 0 ". ., ý
ä ' 0
aý 4. ' $-l ul PL " 0 N to to ý' "rl ri
cJ N d
S 0
C) 'C7 U "4) cl "r4 c
Cd (d N c
cd CZ 40 O 0
4-i d r°"1 0 N"+
"+. "
0 0
0 "j
E-4 "rl v
o ö as 0 0 0 0 0 _e 0 0 0 0 0 0 0
M tom. U O O O O O 'D O O O O O O O O O
UN O 0 O
O O O O N -
O N O O O O O O N- v
E O
Cq O O O O O c- 0 0 0 0 0 0 0 +ý O
%. O W N a) 0 0 0 0 O -
0 O r (3 0 0 0 0 4)
UN 0 ,D ö
O O O O O tt1 lt1 Ö c- O o O O
i
Lr Pý i I
W O O O O o r o O O O O O O "
r- OOOOÖ-00000
,OOO �0 \O _e -t M
-01 O O O O O %0 r O O O O O O O O
0 M O O N
Ö
n Fq M c; O O cc r O O r O O O
Lr\ ' J (3 (Z z1- c; 0 0 A
0
eý * N
`: 0 a -1- o N N ° 0. 0" t- ý 00 0 4-4 O r N %M [` N1 N O, 2 Cam. Ö N O e- 0 0
ri N
u1 O O "ºi PQ c- O O O O 0 O O O O O O (3
a N -d O co O O O M 00 O O O O O O O H
a Ei N l- 00 Co -
O O O N r
O O O O O O O O 0 U
a) M lýD N. CO 00 00 00 O r r
N to Cai cý \ 1 \ N 1 N- A
A A \ º CO t \ l ". N u1 Q' N C- N r- - Cu N '. D N ý- CO O

-77-
U) N . r{ 0
H 0 O
0
4J 4.11
U)
4-1 4.1 N N
4-i 4-4 "r4 O O 0
0 Si fa H N N 0
,Q U
, EI 4-1 ;t ; z. 0
" Fi '. O 0 O .O 0" tý Q, F
1 pi
d) N P.. . 54 c9 ü) +) 0 r+ N tQ +p
r1 "ri -I-) ". ý +. P c) f1 U
"r-C --t O $4 N ^o O
U
td H SI fQ N EO ai
SI 4-t cý O
-I-3.
O F
a, r1 03 Ci
fi
Uff. \O
U)
U) M U O
U . ri 0
c3
0 ri t'\
p . 41
c4 "r ý, o
4) M
0
"ri iý
Qo
vw C:
O
4-1
O
. r-, H n; r
. r{ U S-4
0
i'ý u\ uý UN ti l% OO -2-r- OOOOOO rn 0 ........... 0. OOOOOOO-NOO r2 000
00 -ý c- CO N c-- C` O tll c- OOO V- 0O 00 ,NN 00
OOOOOOOOOOOOOOOr
T- IA ltd c- N w0 uý r -t 0rr . d- Nl I'D N\ 00C.
0088600 t- 000 I'D c-
N K' OO0000000000O0OO
NNNN
OOOOOOOOOO8O O" O N\ O
ONN Ký Co 0OU0O00000N uo KN
N L- r-
0000000000000000
0000000000000000
N " OOOOOOOOOOOUOOOO
4 0000000000000000
NNN
00000000000000000
NM ýO 000000000000OM _ä
%lo 0
0000o00oOÖaQoo0o
r- OOOOOOO0O6NOOOOO
0o co OOOOOOOOON8 OOOOO
OÖ0 NO
0 ul% Kl 0NN %lo OOOOOOOOO'-9OOOOO
T- T" G T- 0000u, N CO tt\ N OOOO0OOOO c- OO c- O Uý C
c '- OOvc0N T- N "D O 4-
OOO O) OO c- U c- NO CU O jt c-
%. D
rn
0 . -.: E
aý 4)
C) (ii ýý A
N -S UN u\ ý10 kOw 'U rv \ r+1 \ ýf \L\\'\\\\
Cxlý N CO (\l u\ J P\\ cl, )N)cNN
r\ 4-

-78-
t(N P. O N- 0 oo O UN
to q O O M o Ö Ö Ö O O O O O O to i - Co N
0 0 U1 '. O 4- '. D
Z: O O O O O O O 40 O O O - O O Ö O
0
t O O -* r O O O O O O O a- O r O O N . ,
1 , -I 0 %0
d c d q O O O O O O O O O O O O O O K1
tý O d
544 1
M C- O N "
0 to r-i Ö " O O O O O O O O O O O O c- -e O c- Q
4 v -ý O
00 t- -2" 0 O
- 0 O O O O O O O O O O O O N
- 4
c ya v
d 0 I r cý d q O O O O O O O O O O O O O O O O +1
ý tH rl tý
f. 0
w U . to 1ý 4.4 "r+
93 0 0 ö Z O O O O O O O O O O O O O O O O e
$4 r-4 G) $4 0 1Q
ý 4-+ " a o o 0 0 0 0 0 0 0 o 0 0 0 0 0. o o O ° N- to
(ä Z 9.4 W q O o 0 o O O o 0 0 o O o 0 0 0 4 cä r-A $Z 1 :Z 10 P. Ö _ r.
P. d N _' la 9) (D p4 JL4
E W r4 O O O O O O O O O O O O O O O N L('
O u
3 U] 0
gl el ri 4j . -p "
l 03 O
N -: g am ä O O O O O O O O O O O O O O O
"0 ö
94 to o
4) $ r4 t+\ O 00 00 O O O ri d d fr., " . 1 . " " " " O
N M O O '- N 0 0 0 0 0 0 c; 0 c- 0 rý U 0 d 92 G) 0 to O 4- 4- " Co 0 P r4 cS cn z O O N N1 O O O O O O O c-
r L-
N N r O 0
4-4 d, ri 47 0 M +3
0 0 0 N O 4- O uN +? " " " " " " " . '. O Ký N- r N
4 O O O N1 \ O c- O C r tN c- c- N N C` 4-4 0
0 E m
b Co r
0 %0 1 ' i = N\ r 0 0 IM N O O O O O O O O O E N
IK- t Z 0 P e- N
r- ' N O Ü U'\ M c-
� . " (0
� s. r' ºý1 ý- N ý- r 0 0 0 0 0 0 0 0 0 H ý
0 C) a %Z am m
8 ' O - c Ö
M c 4 ý %M ' 93 O 0 q- M e- r+\ N c - C' O O O O O O O O .ý C)
U d
4r -P
d t+1 l1ý l! ý lfý \D IU IO %o % D O p 41 cm NN + \ Lr\ \ \ \ '. D \ \ \ \ \ ti N d \ \ \ O \ QO r \ r OD 4- 00 N-
A a 00 CO N to cm t+\ r N N -ý - c- ý- N N U Ö

-79-
of thermal stratification (able:; 7,9 and 11). Accumulation
at the surface tends to be more rapid and intense, a
reflectioxi of the greater stability of the water column of Kettle Tiere.
The initial appearance of A. st)iroides and A. solitaria is typically during mid summer and there is some evidence to support Reynolds (1971) contention that A. solit=; rim is
a summer forra while A. Bpiroicles f. spiroides Kleb. may be
classed as a spring form, together with A. circinalis. These differences are more clearly visible in Kettle Here.
The growth of Anh. gIos-naguAe generally mirrors that of A. cir. cinal: ie. This similarity. - is more distinctly seen in
Kettle 1-? ere, the appearance of populations in . Ihite Here
tends to be more variable. - The last of the blue-green algae to form blooms of any
significant extent is Microgstia (principally M. aerufrinosa,
DI. wese*ert*ii is also present in small amounts in both
meres). These blooms are formed before thermal stratification
breaks down in the autumn. Relatively high overwintering
planktonic populations of Microc;, stis are found.
Although not included in tables 7 to 12, G_ ochinulqta
made brief appearances in the phytoplankton of White Mere
during June and July. Surface blooms of this alga were
observed to occur almost siruultaz, eously in Newton Nere,
where it frequently becomes dominant, and in White Mere.
No occurrGncco were recorded for Iýettle Piere.
During periods of surface bloom formation, with a
large proportion of the algal population within a few
millimeters of the surface, a sudden change in the
physiology: ical activity of the heterocystous species occurs.
This change is revealed by the extensive formation of
akinetee, and is often so rapid, particularly in Kettle
Mere, that sovie stages are eaoily missed altogether (Tables
13 to 15).
Akinete production (sporulati(On) is initiated at the
formation of a surface bloom of the t; pecieo concerned, irre: pent the of the size of the pop; i1, ati. sri. 6: ': q11
po,, u1 tic: tis of . ý. _ iroi, i.: ý Ae_ f oi±tFi rý, 14it}7 little
e113e in the P1ci1::: ton st the ý: u: re tzr.: e, Esporulý, to riss rorad7ly

-80- -
CT t i t
to
" I 1 I
(D rj UN U'% ' Ö
co c N (D 4-) ri t
10 H 4. ) 4-) H co
O F3 N
1 1 1 1 1 1 N 0 4-4
y N O r l0 " . --ti�yý
a c d
Q 3 O F\ - - 1 1
ý rl (D ö "
0 o t 1 O 1 cff " CT' ýi r
4.3 V F ö
" 4-4 IL4
c N O N
N rte. " " " "r4
P4 o O r- r 1 + a Q a)
" a 4-3 to 10 2 0 0 4i
0 X 4-4
`ý 1 t N I I + tý
Q A W "ri 0
Kl N ý{ ý "r i r r ý lý H
H cJ 3 I M O O p 'd 4)
"rl N LO -P + I t O 1 + 0 CO d)
" Ö
(D C) it G) rq 4-ý 4)
to
4.3 4-4 -H r,
0 O -H 4-) IH -Y4 H 4)
C- \
C- \ \
00 \ Oý " .)
4 Z DC 3 Ä - O r N Q
N l N Z
+ 1 0 x 3

-81-
Lf's
C-
C,
4) El 0
r4
CH 0 N m co 4, ss U
C) P,
d
N
. r{
C) r1 H
H 0)
1 I I 1 1 0 t I 1 1 0 I " 0
p 4)
CH
" 1 1 O 1 1 I 1 O O 1 1 1 1 1 0
Ir: N
4-) cd
C: 0 b O I I I 1 O I 1 1 t 1 1 1 1 P a)
4) H H
r i O 0 ". "
4 t I O I O O O O N 1 1 1 t I N " k O t
t o O
N rI C I I I I I I I I I I I I I I
. rj (D r-4 0 > to P4
f\l O N
1 O O 1 O 1 1 1 1 1 1 1 I 1 O C
4-)
1 O O 1 1 O O O r O O O I O 0) 4J
I T3 N ýi W Hi
x RS H
LLO (Z 1 O O t 1 1 1 1 1 1 t 1 1 1 0
" H 0 U
! L, t -I CO +) r-I 1
N y 0 t H "rl
" H
a) H 4) 54 SL4 "
. s . O' ro
_ 1 O O 1 1 1 1 I 1 1 t I 1 t E+
O C) +) +s G)
to 4-) 0) O 4-3
4 O ++
ri C) K\ '-0 C- 00 CO CO 00 O r N 4) H G)
c5 \ \ ºn \ N V3 C trl .D \ N L\ \ O + A N L\ ' N N- N c- v- N ý N T- CO Z " l .a

-82-
N a'
d C)
13 0 w m to 1
4-4 0 a) bo c0 -p U
C) P4 Q) +) a) . r.,
u*%
a) ri Cd H
0)
c6 r, ý' 111t1tt1I111OOO
tO O
O
"1t1I11$1I1IOOONN . ct
cß
11111i1111111111 li§ O N111111111111II11
. 41
to d
1IIIIII1OO1 t\ O 1r
. r1 co
111111I11I 1OOOON
Q
O
"5 1OO1Oi0000000001
O H
t1111IOON111111O
tr] wOOOOOOOOII1I1OOO
U
v1f1111O1OO1OOO11
O O K1 -f' !! 'ý ltd ýO ýO ý ýD c\0 r
c3 \\N\O\ 00 ý' \I I-
ý' 00 \ cV C` Q o0 CC) N U\ Nr ý- N /O NN4rN
b
ti
m
d ri A2 cat
H
0

_C j }ý
as the gene: rally 1nnr[, er populations of A; circinrt1is and
A. flosýaclizze, although for these species too, population
size is no requirement for sporulation. The clearly
demarkated succession of species in i, ottle Ller. e leads to
blooms predominantly of a single species, and this situation
further results in a distinct succession of sporulation
throughout the summer. The more or less simultaneous
blooming of several species in White Mere, though present
in different proportions, results in a more or leas
simultaneous sporulation. Unlike the AnabHona species,
AJ)h. - flea-Rnuae rarely produces akinetee in large numbers.
An absence of akinetes of thin species in Kettle Mere was
reported by Griffiths (1925), and none were observed either
during 1974 or 1975" Under the unusual weather conditions
of the summer of 1 976 , however, large numbers were produced
in Kettle : era during October.
Sedirientation.
With the differentiation of akinutes the alga concerned
is effectively removed as a component of the phytoplankton.
This is through rapid sedimentation rather than a
redistribution within the water column. The loss of gas
vacuoles and the formation of cyanophycin granules as
akinetes mature leads to an increase in. density.
Sedimentation is also has-toned by the clumping of large
numbers of ak., i. netes and fragmented filaments in a matrix of
mucilage incorporating bacteria. Rates of sedimentation of
total particulate natter are given in figs. 17 to 19.
Although there is some correlation of this with the
eed _mentation of algal material (figs. 20 to 24), this is
frequently obscured by the greater contribution of resuspen-
ded matter, particulsrlj in isothermal conditions. This
source of interference would be lower in Kettle fiere than
in White i"iere. This may be reflected by t; ', e lower values
for siwpended solid:,, except during periods of surface
blooming, in 1 976 25 and 26) . Tli(: total ctrnount of
riaterial trapped in Kettle Mere is consiratently lower,
even during periods of atzblo otrutifiontion. she
coiltributxwic of the, differ : nt i; ec: ieEi to the total

-84-
Cý
7 0
f". 0 C)
C7 '
ký
cý
J
a
.ý ýS v
Y ). w
c
i
_iU
Ic cl lrU
n) iV t, r ýý
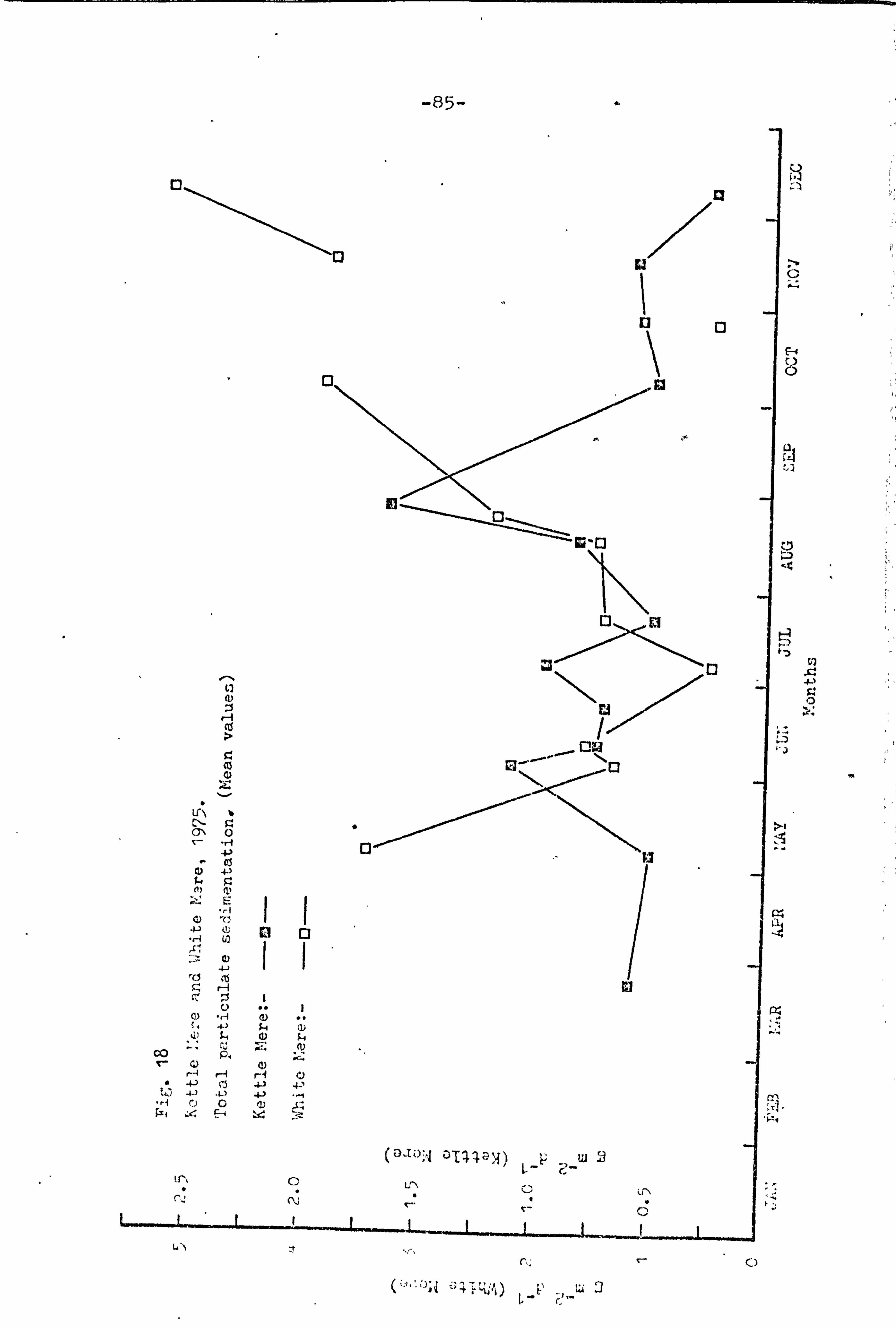
-85-
13
13
A.
a
nQ
"oý
rý I c3m
co ®1%Q
tfý " ON Ö" r- "ri Q
R) tja
Q1 7ý
cQ
-3: 0
H c 4-3 a
:_ cc «c. N 00 Pi
r ri r-H C)
:ü4. ) 4) . +. ) ".. i
(hto °T44a; d) Lr, U u\
Ll r.. r,; T--
cy
c
8
U L-ý1
G7
. 94 4-i
O
C)
; T. Li) 2

-86-
r)
b
U U
a sI
U)
0 Y="
r,
H
i
(9. Aalq 'I
L, " _-r

-87-
sedimentation rates are clearly seperated, which reflects
the succession of the species in the water and of the
sequence of sporulation'(Figs. 2u and 21). The contribution
of Ilicroerstis to the total sedimentation rates in the
autumn is particularly marked.
Resuspended detritus forms the main component of the
trapped material in White E ere during; the winter months.
Sedimentation of diatoms, mostly Asterionella, as the
water column becomes less turbulent during the spring,
forms the major proportion of sedinented material during
April and May. There follows a marked decline in the amount
of material trapped. Although this decline marks a five-
fold reduction of the amount of sedimenting material, the
values are twice as high as those in Kettle Mere. This is
mainly due to the sedimentation of larger populations of
algal material, but the greater degree of turbulence within
the whole water body may be an important factor.. The
sedimentation rates obtained (between 0.5 and 2.0 U m-2
day-1 for kettle Mere and 1.0 to 5.0 gm2 day-1 for. White
ilere) are similar to those reported for other small hard-
water lakes (White and Wetzel, 1975). The inorganic residue
left after combustion was higher in material sedirnented from White Mere than from Kettle Mere. In the case caf White Mere the percentage ash varied from 45,,, to 60w of the
total weight, in Kettle Mere from 20, to 30/0. The greater
proportion of organic material in the seston of Kettle
Mere is probably a reflection of the differences between the
sediments of the two reres. , The rates of sedimentation of the principal forms of
blue-green algal material are given in figs. 20 to 24.
Peaks of sedimentation of algal material correspond with
periods of sporulation and/or the decline of . surface blooms.
nigh rates of sedimentation of i iicrucyritis colonies,
particularly in Kettle More (Figs. 20,21 and 23), may have
been brought about bfr a greater depth of mixing, as
indicated by the deepened epi?. imn: i. on in the autumn. A
similar response of i iicroc-:: tis 1, rß: 7 - been observed in
Rostheriiu 1%. ere b; i Reynolds and Ro,; ers (1976). The high
rates of ssedi mentation of 1Lr. ihT iizý in 4i'lhite tiers in Apr. il

-88-
8
1-`
113
:$
~J
I I
,y
, I ( co s.
c LZ "r1 ý-"
iJ k Q, O r-i C: S U
"r4 x 4-4 cl, ni
C9 G7 V J N N CJ
aý 4 J Ll
C' L
'A in r-4 r-4 ýA " 4-1 r4
". ý! C c, O O O
. f, .ý
r-
ý) W (ý ý" .1 aC: m rt j

-89-
D
ýr
O "
91
13
ýD rn
L1
L7
[ F ý'
i
1
rf
G
il
r-1
V
'e'1
l
. -ý-
fý
"r"t
Cam : c7 E
4.4 E- U v
U .1
cZ
C,
fý U (-)
ýý c"ý
ä .ý
ýý r. U
F. '_-ý
ý, ý , S,
ýr ý'. )
1. ̂. C,
t- ý
D
". ýDý`
V .ý t
1. \
()

I
-go-
®\I %-
v
p,
1
ti uý Dq
D cý
CIO
Dn
r4 co . ri 4-3 rt Cl D DK ra ro V H tom: 4. c: t
o c) F4 to U
+i C. 11 N P. DPa. ® r;
. d) :J1O N u] 1 i. 1
N G=i tU U] rI t' W r-i
ri () O0 Fi C. 4 ai C-. a
L. ý_ý fl
(`j nl ) r'+ l' li
p :a ýýL". z. t: i,: n'j
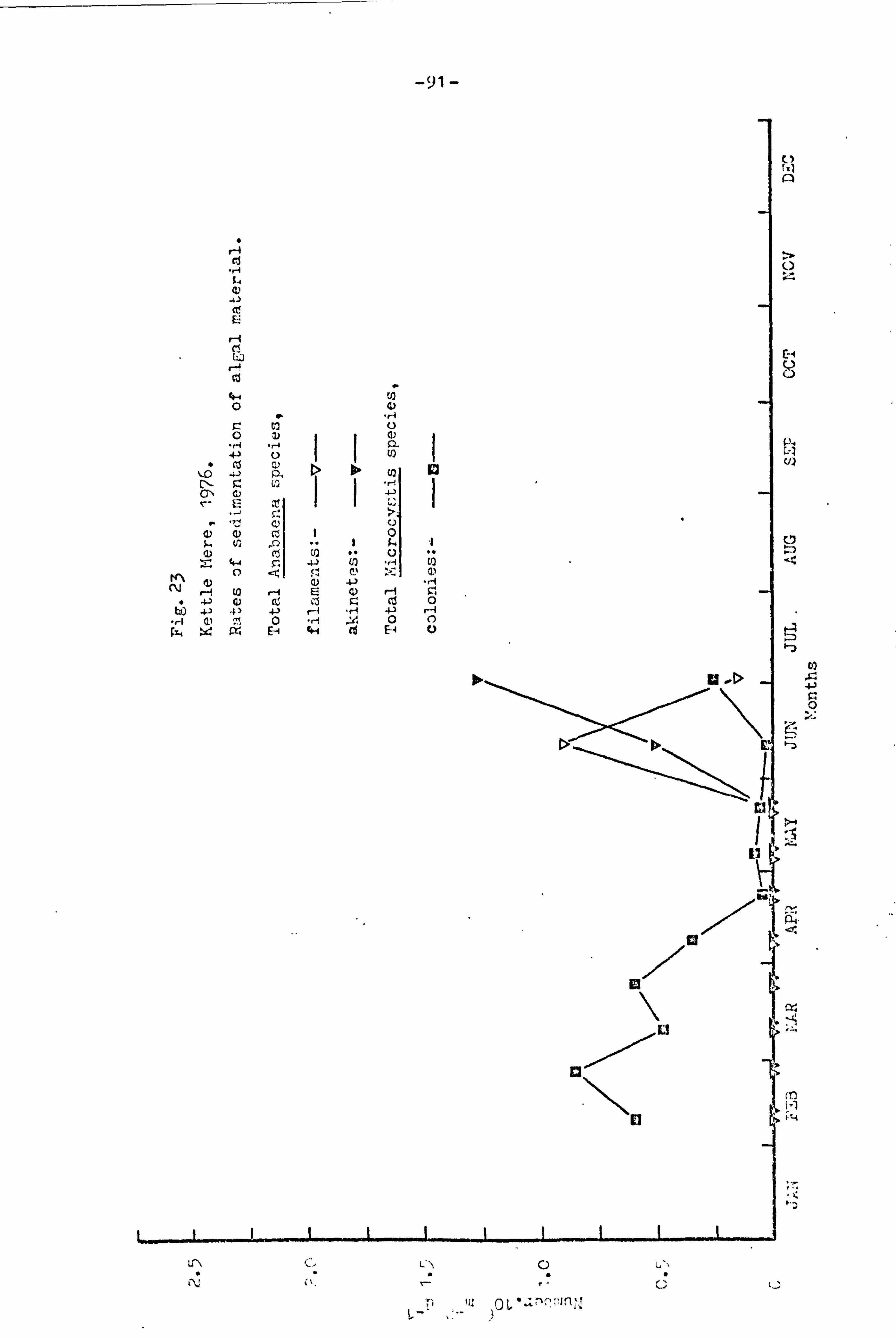
-91-
U f. ý
0
E4 C-)
Cl)
c,
ti to
0
ti
'3
N
ä
cý
in c ýa t-ý
71 'L r..
-'s i; �
PJ (cG c_)

-92-
co U, t4
4-4 0
Ci
fti
UV
41 0
Qf "r4 ý U) U
N . 14
%10 i: i It Q In 9), C- c -s
Ii+
"" Cý c'y 1O
V. C)
C> !n .ý-! i. !
ý ýN r-1 - +ý Li "n Lý '
NOU "'
. 1"
N W' rA r-1 t= CJ riC. ' " -0 G CZ 0 .0 tý: MO
P4 = !ýO E-4 41 tb EU
aD
F'j
I--
i
f 'el
Ll a
Dp
(sue : ý, s - ý -t. ýrý Luýý: 1ý, üý, ) p uº Oi. ". zaýý: nTi
(Sv'rýi: lýýtii L': 1,. . i{t:.. j
5Ti)'LUt_ý UJ ý~`_ý ~.. ý"ý `,
I, LI IT 1

-93-
B Dý7
a cl
A®D
G, G7 "-"
Z 0 Lr\ U ýJ N
r1 4) ij ri O
cý x ii) cn <ý c: a
bý
DQ
Q
\I/D` i9
pD
®ý p Uýp
ýlnj `N:. 1
w 1/ D tJ , ýDý 0
ot
1 äý©D ^
C- C .
1 1 ti
ý, 't a3 C V v t'
c; :, r-1 ti
.V G1 YY {ý
,4 co
'iJ ., L
r)
r! 2
4-1
ti
fýG

-94-
0 L> F3
( ©
I D ! 3
I I ( N
" C) 1 v
4-4
r N
ri Q u;
1 ..
r.
1 ""
. - )N c- r-'
0 (C
Ll
1
'
I
0
C) hJ
N
t
N H
+ý
'(C s.: C)
N V 0. i
N H 'cý
£ 0
U
Q) C)
J
Q)
QI C6
r1 El 0
V
W
Vl
f`I
to 14 CL)
ý: .' u) (C H 0 'H . C;
l '' ul
1-1
cn J) -4 > 0 i
0 `` D of C'i ýD
/7
ri D0 \\
Eý D41 ""'Aoc>
Q
N
rý
0
4ý
U/ 0
r
Cl)
N
Gý
®a ýQ ý"
® Dý
D ti/ýý
L-- \,..; Li \ _r '''i fJ r- Cý \ ,_r
qi
Ea
. 1..
.u U
: r:
>,
r, ý, s
r, .ý
t__-1 ; ý..
4ý

-95-
Land May of 1 976 (Fig, 24) marks the rapid decline of the
planktonic population during mid May.
From the large amounts of material sedimenting through
the water column it would appear that most of the population,
including gas vacuolate filaments and colonies, had
suffered a reduction in buoyancy. It has been suggested
that the activity of proteolytic enzymes in crude cell
extracts allowed to stand increases the fragility of gas
vacuoles (Jalsby, 1971), thus a slight increase of turbulence
could bring about hydrostatic collapse of the vaouoles and
induce sedimentation. This is particularly noticeable in the
1976 White Mere population of A. circinalis (Fig. 24) which
had not undergone sporulation at the time of sedimentation.
In the winter, and also-at intervals between the
sedimentation of surface blooms, little algal material is
trapped. This indicates an absence of resuspension from. the
sediments of large amounts of algal material in the period
subsequent to sedimentation, as well as the lack of
further large planktonic populations of the species
concerned.
Benthic Populations.
Fluctuations in the amount of algal material in the
surface sediments are given in figs. 27,29 and 31 for
Nettle Mere and figs. 26,30 and 32 for. White Tiere. Large
increases in the numbers of intact akixietes were found in
both meres in 1974 and 1975. The peak in early June is
produced by the sedir_ientation of ak. inotes of A circina_li^
and that in August mainly by akinetes of A. floc-amuse. The
late sure-mer forms contribute to akinete numbers to a
smaller extent, particularly in Kettle Mere, where akinetes
of A. F$1)iroidos are found in early Au,; u. -3t and of
A. solitaries in late August or early Lieptember. A
particularly large: number of akinetes of A. s, piro: ide was
observed in the surface mud of tiihite I"iere in $epteriber,
1 974E (Fig. 28). The unusually hot and drys weather during
the surlrier of 1976 produced a different pattern of
succession and spor. ulation. 1 cirý:: ýýi Lio Was aiccomp . trtik: d
by an e: urJ_y bloom of A. s lon- 3ý ýr-w at Lhe c? nd of T: Fty in

-96-
C3 tU ý' : -y rý
(J
r. ý
a rg ynA
Qri
r.. ®0
0
..:.
n-ý H U: t " 4) (j
. aý TJ ." ® rj ai G uý c ý
41 v (to " ti 'O CJ I -ý Q% rI n
Q Rý
(r: C) CJ C1 G. l ci
r f. I J' "r "1
111) 171 N y' +) rl .
J
H C. C C) ri U H4 Ný " f61 .ý
+ý J u L
E: "S & )
^3 :, 6 0 et L % :s . Q U)
4- H Q0
Nl ýl U1 Cý U1 c- c-

-97-
0
0
0
UC
Q
ý3
7 C ýA ýý
0
Ci
00
h4
4.3
I I co
( ! . f I C
a-1 13
Ö 1- r
ýý. -c ";, w a., E' ci
L1\
a.
a .y
ýý 'ýý tý.,
ý;
iý
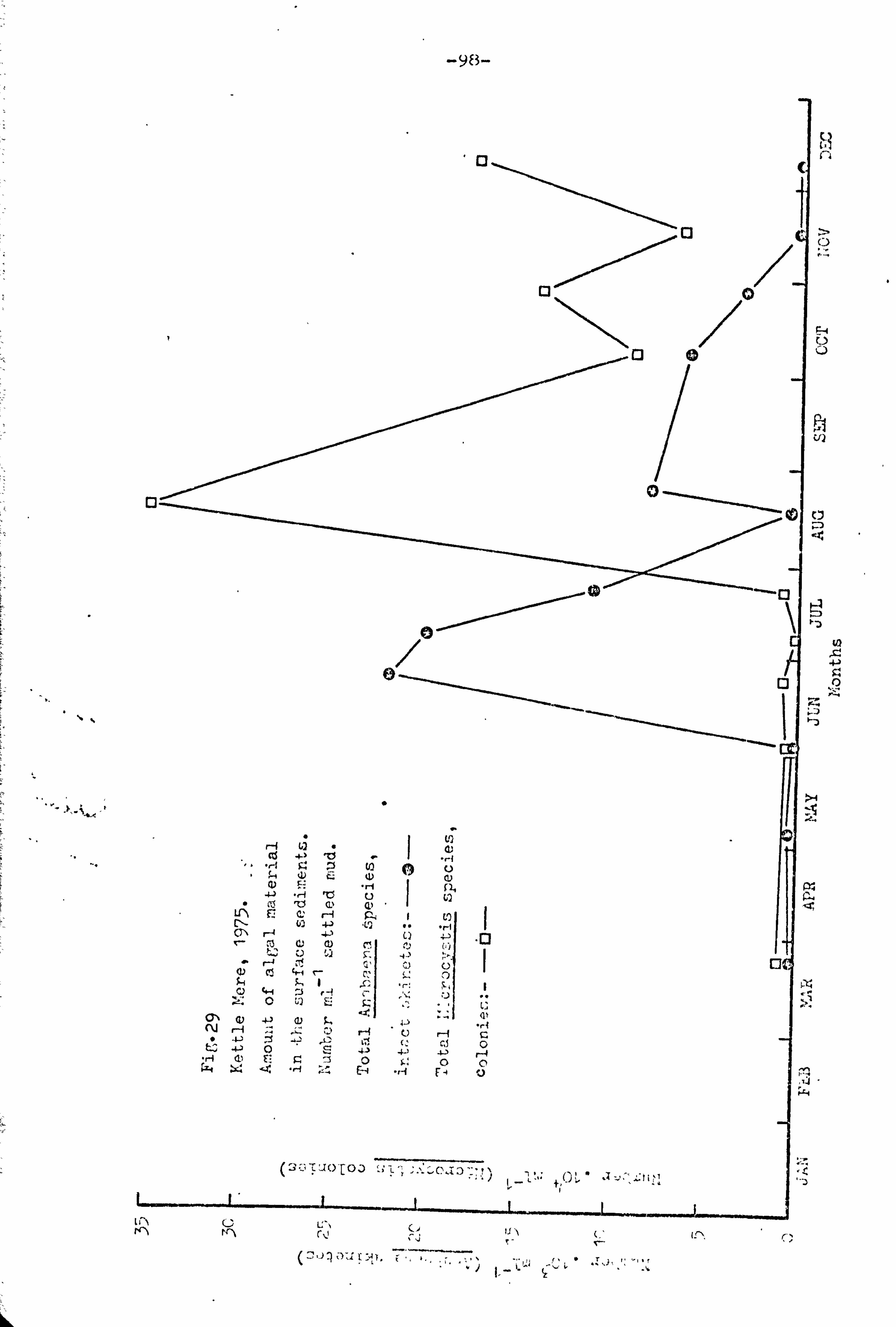
-98-
i3
13
I
/1 o
00
lpl
0 -. `ý
. r`_jý
(J.. L;. zo IO3
el, ý
tit C r4l NIN
T-c, Ir-
,ý\ V
Q
6ý ri L7 "
I cd 4) H
" :+ a) C lU °' b 'ä cý
" Q
u £ 0 4 I L - rn o u ý r1 C -
r-+ - c) 4
Q T C: w c! :. )
s ,ý Cr r I
4-1 r- 0
nr r{ N r-i i J rS U ri G
4J Q r t,
j CJ 0
ri al F !~ O 4 i:
4-ý- 0 r-4
0 ter : ýc: o .a N ". -i E -4 0
to
4-4
0
i a w
0 M rý
nj . i4
.r
If', C:

-99-
? "ý _ý
ý 9%
C3
0 c» K-- D
O
a 0
a0 b>
ý: /: .ý
ýý <ý
A '-'
0 r1 4.. )'. "C ^J cc 0 i> . 14 U S. U t' N Q) Cr t.: "r4
E i ý2,
. -, .., d c, I J G; " i ý' r' f'. 1 ýý
I
a c, c a) c ) i;
rl c; ra I
c4 j, r c' 1 .. -. ý. 1 _`4 U
O Pu O v: _ v .J ,. 1 t4 VI A
N f~ :> H C) t H C"
e:: " ra C) 11 1 ,; f ., -' H Ö 0 0 f_
Lt \ C) !. \ tf
tý
Y t=1
J-y
fJ
6L

-100-
' ý.
.. j, ... ; ý,
N- c"
K1 tý
w
S " co
--I tr
i, t v N
4-) -r1 'LC U ýT J CJ Q)
"1 4-3
cL` 0) N C, G) t', J "
' A C3 W Cil t. )
as 4-, r c: c
U rQ E C c; ", -i r')
1ý Q) ý4 -1ý "r 1 S; .L 4) H V H
"0 cL n3 Ci O 43 -0 Aý3 r-A
El $4 11 0 C, 0 . "r1 f. E4 r+ H u
r-ý
c-) u
U)
c5 -r
0Q
13
13
C, 13
Cl''"r 1
tf\ wl t' RI ý-
t_ ll -()ä t
N
. t: 4J P., 0
;., .r
r
. r,
rz:
;ý ,. I
j': '

-101-
p "1
0 ® ̂ . ýa t-I : " C)
"-3 - 'i U] U N tý C) C)
" }. a) H P. a) W t, "N 0 4) -r i C- r-, -P CI + c\ ') c) U ;t a C) "1 U r r1 .. ý
ri ' Q; 4; ýiJ V
C) z+ 1 S' fti <U S4 4-I 71 r! 0. Cl <. "r1
4-3 N 1. ý' C) r! 1 ri ý-i J
-4. ) . -, 1) of ri 4a C c)
Gv -t --1 E-
;a 0 F -.
Cý C, J 0
0 a
rý
iA .C Fý O
Cý
G$
0ý
iýý Qý t'3
,n .' rt\ ýý c: J' ýý U114 'l C_)
L

-1 U2-
Kettle Mere. The other f3Pecief3 eniunierated did not start to
develop largr populations until the end of June. The only
contribution to Hkinete numbers in the mud, until the
sporuln. tion of A. spiroides in October, was the sporulation
of a relatively small population of A. fflos-a7uae at the
end of May. Iihite Mere, like Kettle fiere, had relatively
small populations during June, following; the collapse of
the A. cir. cinalis population in Hay. No akinetes were
produced by the latter population and none were observed
until mid July and August, when the planktonic populations
had recovered and again produced surface blooms.
Although vegetative material of the sporulating
species was identified in the sediment traps, the length of
survival in the anoxic mud of Kettle Mere is short.
Filaments are only observed in the period immediately after
sedimentation. They appear to persist longer in the mud of
White-Mere, but here too there is a rapid decline in
numbers, particularly during periods of stratification when
the hypolimnion is depleted of oxygen. A decrease of
filament numbers in the mud during the winter may be the
result of mixing into the plankton. Thus the numbers found
in the mud result from an interchange of filaments between
the mud and the plankton. Despite the rapid input of akinetes into the sediments
of both meres, there is an equally rapid decline in their
numbers. This rapidity may be riasked by the sedimentation
of akinetes of another Anribaena species, which would
increase the total numbers plotted in fibs. 27 to 32. Each
species exhibits a similar pattern, and the whole process
of sporulation and sedimentation is generally completed
within the period of stable stratification. Resuspension is
therefore unlikely to be the major cause of decreased
akinete numbers in either mere, which is corroborated by
the low numbers of akinctes found in the plankton during;
the Iautuyul and printer. After the decline in akinet¬, numbers
of thc: last ap.: ciuo to "poruJ. fate (generally A. voll', tnria or
A. ^ iroidt 3) the nurahrr. s remain. low throughout the winter
ancl the following An exc e}! tiun to tlhi:; is the l LLr,; e
muib ;r Of fljýa71(ýEý: i found in Kottl(. ' sture bes. -wo? n ", arcli and

-103-
976. in contrast to the vegetative cells of Anabaena, there
are large overwinterinj; populations of I-Iiqrocncitie-ý in the
surface sediments, particularly in 1t tle '; ere. Lise the Anabaeni species, however, an increase of numbers in the
mud is reflected by a decrease' in the plankton, within
which during the winter few colonies are to be found. The
reverse situation can be seen in Kettle Tiere where an increase of numbers in the plankton during August and September (1974) and a smaller increase in June (1976) is
paralleled by a decrease of numbers in the sedirments. The numbers of overwintering; colonies of Microcystis
on the sediments of White Mere are smaller, which, like in
the case of filamentous species, is probably a consequence
of the greater degree of turbulence. The relatively high
number of pl-snktonic colonies, as compared with that in
Kettle Mere, supports this view. Decrease in numbers of material sodim©nted during
periods of severe oxygen depletion of the hypolimnion are
also appar t, although less precipitous than that of filamentous material and akinetes. Benthie population levels are supplemented by further sedimentation of
planktonic colonies during destratificatjon in the autumn.

DL CU axOl4 The C, rowtl-!.. And Abu ahce Q Plan oni, c Pot Actions.
Despite the similarities in nutrient composition and
concentration of the two mores investijated, the dominant
blue-green algal species and also the population levels
attained are significantly different. This difference is
more clearly seen in tables 16 to 18 in. which integrated
algal standing crops for 1974 and 1976 are given, than in
tables 7 to 12 which give algal abundance at selected depths.
The large and actively growing population o±
A. circinalis in White More during the winter of 1975 to
1976 supports Reynolds (1971) data on the lower temperature
tolerance for growth of this species, which favours its
early dominance in relation to other species. The most
rapid period of growth was, however, when the water had
warmed to above 9 °C. The response of pro-bloom populations
of Aph. fos-aquae is also similar to that reported by
Reynolds (1971), the increase of numbers being moot rapid
once the water temperature had reached 12 °C. The 1974
occurrence of Ajh. flos-aquae in White Mere was unusual in
that a planktonic population did not develop until the
termination of thermal stratification, possibly the result
of an increase of nutrient concentration in the surface
waters.
Although occurring in both meres, Anh. _
floe-ýzcýuae is
atypical of the filamentous species in that larger
populations were present in Kette Nero for the years
investigated (except 1976) than in White Mere. hammer
(1964) reports that this species did not increase until
the temperature had exceeded 20 0C. A high temperature
requirement for growth may explain the differences between
these two meres, with a more rapid wariaing-up and higher
temperatures reached by the epilit: uiion of Kettle Berl
encouraging more rapid gror:: tli. This inay also account for
the large populations found in White [,, ere during; the hot
weather of 1 97o . Large po, )ul atiozny Of this specie", eitre,
hoviever, reported for White here in 1967 and 1968 and also
for. other : LarE, e mc: xý. ý ýlieyzioJ_c? ,1 971 )
Gen lie Fand I,: uloney (1 969) reportod that a Phosphorus
L

-105-
N "r1 4-) C7 c- co
N c ` O 0 0 c - O 0 N
i cC
J
r f. e " U
N N
O C H 0 O " cy- IL) U) cö
0 0 N fý
si ) N
O Q E H
O N KN
U Cd 4-4 t! 1 r (`J r" v c- Q O O Q +- rl C 44
w r-I o " "r1 Cl) 4.1 S, a) cd
N "rl "rl 0 0
cC H +) S4 Z 0 -f rI N U Cam- H
ON 0 E 1 4-+ c- u) O O O Q O Q O Q O Q
"" 0 "rr P1 w "
" U] (1) j-i Pl .d C) 0 f-I El: (a S-r O 0 U f~ F. N 'Z3
(2) H r t0 r. 4") O SI 0 I 4-3 fti
-* O c- I'D U, \ C- 'Ü
i
0 0 p, vJ 0 0 a0 N Ctl O N O\ _; I- u1 0 l ý
-P r + SI " z4 -rl
'-1
O cý f-1 RS
r s~ cý !J rI Ul
co 0 4 O O O O O O O O O
r
Lo r. =4 "
0 H
U) "r1 H ýO z C-
"rl C 00 Q1 0 c- c- Vi " .. 7- Q "
r-{ ! -a C'N r- " VIN " ,a ri c- c- Pal c- c- 0 c- c- 0 T--
Q) w ". 0 Co cxi . ö \ \ a. '1, 0 c- r c- (T c- N r N
0' N
f

-106. -
N
0 V-
O ", i 0 0
r- O üQ
ON
to cd
4) H 9
CH HO"
H f-1 O 4) "ri
CH Q O "I~ 0
Si 20 d) U -4 C- E1 4+ Q\
Or
ß, " 01 00
ýo $-4
O F; ¬"; tU 0:
Ucj U G)
h7 4) 01 "ri
rl N ý' '0 -ri P-0
Pr c0 cß u)
y "rl ri 4)
cü O
TJ V JN
-i-3 co .. ý
to .
t, --
r4 c1 H
ä Pl
U) r1
U O Fa U
. r{
O
U) O
H 4i
. t;
Cd 4) . r. { H 0 to
Q
,d 0
. rj P-
L;
CT
0
H co ri
"r-I U
U
rn CO Kl% c- c
OONrrr N\ NN
OOOOOOO O- O
O 0000 U\
rNNOO
Mr Iý
ý: 0000 c-' c- VD Nr0
"C 00 tNr\ v- N\ O\ N rc\ c- c- O00 V- N O*N Oti rNN
u U1 l- c--
N N CC) UN Z- Lfl%
-N ýO K1 cam-- C`
Q) "0 "0 LI- Cl- 00 CO ol\ ý3 U ßc7 \\a; O r\ Cý \ t` 2 c-' N (7" r fý c- cU O\ N

-107-
P,
to .ý N 43 C.
a' N N . 00
ý" C) .t N r N T- O\ O o
" U ", 1
U] , c. d)
F, ' G) O co H 0 O " V Ui
4-) 1 0 1A
N chi 4
O O Q r
º3 ri -1 C) 4 -1 9
4-1 o .
.rj C 4-1 t-l ) C cd
ö ö 0 H " +)
z 0 n H C)
° 4 T o 0 0 0 0 0 P4 0. 0 P4 $4
o C m U Z O 'L3
N H ". 4 -P 0
0 1 ri 4 M " " CO r P P
(11 O O O M cV C`
W P
v] 4 ct "rý
9 > Ui V.
i fi Ui 10 0 o 4) i a " rn
H
6
: : : : : :
00 rg
p r1 Lr\ O r r r co V
H d
M ýD Cý" iM ONOONiN
00 - 0 cO cam- CZ L['
rn 0 0 0 0 0 0 0 r
C, ti C, a) 43 . rj
C o 0 0 0 0 0 0 rn
N ýO r Q'
N oO %0 L-
O N1 c- 1 C, - c-
0 U'S C7\
%0 %110 %10
MNNN
L

-108-
concentration of 1 .0 LM supports a population of 105 cells
r_il of Anh. floc-aquae and that low concentrations of
phosphorus can support bloom populations. Flowcver, in
continuous cultures of diatoms and bacteria Fuhs at al. (1972) found that growth is limited below 3 ý1g 1-1
phosphorus and that the atomic ratio of nitrogen to
phosphorus required for maximum growth is 16: 1. A ratio of
about 55: 1, determined from analysis of water samples from
Canadarago Lake, leads the authors to suggest that
phosphorus will be the limiting nutrient in the lake
studied (Fulis e: t al. , 197-2) . atomic ratio of nitrogen
to phosphorus was found to be much lower in the meres
studied (from 6: 1 to 1: 1), which would indicate that
nitrogen rather than phosphorus would become the first
limiting nutrient. Studies on Yhaeodactylum by Goldman
(1976) showed that enrichment of phosphorus had no effect
on growth, whereas increasing concentrations of nitrate
produced a linear response. Concentrations of phosphate and.
nitrate used by Goldman (1976) were high in comparison to
those found in the rneres. Despite this, however, no
significant depletion of phosphorus or nitrogen was observed
in the surface waters of the meres. Studies on the mineral
nutrition of M. aeriginosa by Gerloff and Skoog (1957) also
indicate that the requirement of nitrogen is higher than
that of phosphorus, while the converse nay be true for
those taxes which are capable of nitrogen fixation. The
concentration of nitrate in the mere.:: was not observed to
fall to very low levels (i. e. below 0.1 to 0.05 me 1-1),
even during periods when non nitrogen-fixing species were
most abundant. The contribution of nitrogen-fixing species
to the nitrogen budget of the lake may be important in
maintaining the concentration of nitrate during stratificat-
ion and surface b?. oorls (Fog; and Stewart, 1965). Nutrient
depletion, particularly of nitrate, occasionally approaching
limiting levels, was reported to occur in cruse : sere
o7 dts, 1971 ). In Kettle I, iere and White iiere nutrient
deficiency does not appear to limit the growth of the
planktonic re(n al,; uc.
J,. factor of particular : imj)ortnnc; J in 1 hc,, control uf

-109-
growth of planktonic populations in the meres is the amount
of light which is received by the orga: nisrn. Large
quantities of dissolved organic matter in Kettle here are
supplied principally through the autumnal leaf fall of the
surrounding woodland and its subsequent decomposition. This
material strongly absorbs photosynthetically active
radiation (Kirk, 1976), and therefore its presence in the
water will reduce the depth of the photic zone considerably. The potential for photosynthetic growth, particularly during turbulent conditions, is therefore greatly diminished in comparison to that of White Moro.
With the reduction of turbulence during the early
summer the buoyant algal material floats towards the
surface and frequently forms surface scums. This placoc the
alga under completely diff037ent conditions as compared with those that exist while suspended within the water. Light
intensity is high and carbon dioxide may become depleted in
the surface layers, factors which will tend to reduce algal
growth (Tailing, 1976). The earlier stratification of Kettle
Mere places a further imposition on population sizes. The
observed increase of pH values suggests that the proportion of freely available carbon dioxide to that of other inorganic carbon ions is decreased as a result of the
abundance of algae in the surface layers (Table 6).
The speed at which an algal species disappears as a
component of the phytoplankton is frequently more rapid than its appearance at the water surface. Under conditions
of depletion of free carbon dioxide as the pH increases,
and supersaturatioii of oxygen in the surface: waters and
under a high degree of insolation, photo-oxidative death of bloom forming algae may be one of the main factors causing their rapid deteý'. oration (Abeliovich and : ýhilo, 1972; Shilo, 1975). Although the temperature of the surface
waters in temperate lakes may approach, for limited periods during the suruier, that in the Israeli fish ponds examined by the above authors, high water temperature is not it requirement for photo-oxidmtionn (Abc: liovich and 6hilo, 1972). Photo-oxidation of blooming; blue-E; re,, ni algae under natural ooud±tjo"13 has been reported by Bluff et fll..
(1976) and by

-110-
Boyd et al. (1975).
Photo-oxidative death may not be the only cause for
the collapse of blooming population. of algae. Vaeil' eve.
and Levitin (1974) suggest a combination of factors,
including changes in the partial pressure of carbon dioxide
and oxygen, nutrient deficiency and bacterial and/or viral
infection. Algae-lysing bacteria have been isolated from
eutrophic lakes by Daft et al. (1975) and found that their
numbers are directly related to the algal biomass. Lycis
may also result from a change in environmental conditions
affecting the balance between the algae and bacteria.
Although bacteria were observed in large numbers within
clumps of senescent algal material, their appearance is
probably a consequence of algal breakdown rather than the
cause of it. The rate of breaks'own increased considerably
when algal material collected from natural bloom populations
was transferred and kept in the laboratory.
Although vegetative cell material of the summer
surface blooms frequently disintegrates and disappears
almost-without trace, this is not always the case. The
sudden disappearance of a large population of A. circinalis
in White Mere during mid May (1976) appears to be the
result of sedimentation of intact vegetative filaments onto
the sediments, This material did not reappear in the
plankton until July, and meanwhile filament numbero in the
sediments had greatly diminished. Populations of r1i'crooystis
are also secliraented and remained in the riud in a relatively
intact state, being more resistant than the filamentous
species to bacterial attack.
orulation And Germ-' n,.,., t ion.
: 3evcr. al rý:. )orts ro1rted the production of ak. in©tes in
1aborator. y batch culturoo to conditions of nutrient
depletion. is o1 : (1965) found that phot3phate deficiuncy
induced c orulation of A. c-j_zn«ricn. Deficicncif: o of both
phom-rhate and nitrate stir., ulated sI>orulution in
n (Glad, --, . 191, i). Observatimis on the
s )orulrit1on of planktonic sp, iciea in the mores, however, do
not indicatt: that nutrient rx critical role.
61

-111-
The very rapidity of the phenomenon, when compared with
algal growth, suggests that depletion of nitrate and
phosphate is not the main factor. These compounds are
always in abundant cuDply even in the water filtered from
algal scums. Other nutrients, such as the trace elements,
which are in sub-optimal concentrations in some lakes, may be of importance (Goldman, 1965). Ixtracellular products,
which accumulate in batch cultures, have been shown to
stimulate sporulation of C. licheniforrie (Fisher and Wolk,
1976). This could also be of importance in the natural
environment, when lyric and decay of part of the population,
enhanced by the activity of bacteria at elevated temperatur-
es, will result in an increased concentration of extra-
cellular products within the' scion.
However, when considerizlg. the range of species present
and the differing population sizes accumulated in a bloom
at various times of the summer period in two chemically different meres, and the similarity between the response of
organisms, it is likely that other factors, not present in
a laboratory batch culture, may also be involved.
The regular coincidence of akinete formation with the
occurrence of surface blooms for all the species studied implies that conditions at the water surface are the
stimulatory agents. A sudden increase of light intensity as
the algal material reaches the surface would explain the
observed suddenness of the onset of ak. inete production and its limitation to b__oo. je produced during the sur: imer.
Treatment with u. v. light (253 nn at U. 62 m'r; cm-2) has been
shown to induce earlier sporulation of A. _
doliolur (LAngh
et cl., 1967). Supporting evidence for the role of high
light intensity is found in the absence of sporulation of
surface blovris formed in on-In periods during the winter.
The density of the overwinterink; planktonic population
of A. ci rcinalis in White vIere during late 1 975 and early 197() r ,: £ic!: ed thoso at which thi ß specios would Eýporulat..
duri : L; the suii er, and considerably exceedod the ýo, ýu1 tiori: >
nt which the other -3peci, c_s would r3porul_ette, 3wrticular_l_. y in
Kettle ITnfortunutely this population had oudrienly dccr, oti. -ý,.
d iii i umboro (:, er .:, 'o '1110) bt iforo t}a normui1
L

-112-
"sur.: mer spurulation period" had begun.
Filamentous vegetative material aeldoni survives for
long period:. alter its 3Odinentation from e. declining bloom.
Although blue-green algae (i.. muscorun and A. ? i. ncir. icý-a. )
have been found to be more resistant to anaerobic
conditions than other taxa, the reported naxirlum length of
survival is ten days (Dokulil, 1971). It hao been suggested
that the resistance of planktonic species which form
surface blooms to anoxic conditions is the result of a
metabolic change from aerobic to anaerobic respiration (13ud' ina et a!., 1973). However, the unhealthy state of
post-bloori populations would curtail the length of survival. This frequently' results in the reduction of vegetative
material to an indetectable level, particularly in Kettle
Mere. Akinetes appear to be much more resistant structures
to the attack of bacteria and to anaerobic conditions
prevailing; in the lake sediments. This may be inferred by
the presence of appara. itly healthy and intact ekineterý in
the mud throughout the year, although often in considerably
reduced numbers when compared with the input during
sporulation. The rapid decline of akinete numbers in the sediments
would suggest that this was not so, that in fact akinetes
were as susceptible to the effects of anoxic coldi. tions and bacterial attack as vegetative material. Apart from the
presence of akinetes during periods when nortL-tlity would be
expected to be high, such as the interval between the
onset of stratification and the sedimerntation of a fresh
crop o Fan-f. nctee, the reduction in iiurnbere iu equally
rapid where alcine. tes have uedineiited in the period
immediately following this brcahdown of thermal. etratil: i c-gtion.
This can be seen in White I: erc , 'here the: . f1_uctutºtion. 3 in
nusabber. u of aki netei of thooc specie: 3 which , 3porul_atc 1,1te
4n the uum ner follow a :; iL�ilar pattern to that in I. ettle.
T": ere, wi: icii is still L; trrltified at that tir: e . fºprtrt i rcri bacteria other mßy 1)t3 of
`1 ()21; )) 0I-): ervE cI the, j: ri?: Jc ict. of : 1,
c';: 'tr. id on the rnl: _net es of f: _t cý_l'i tf. n1 , rt ; r_ 1ton i ci
., )--i i iiii la" (_ ffJ____ Lt: iu. " viir. J? )t: t11' tut Üt, f
L

-113-
I. ortarek, 1958) in : settle : Iere, White i.: ere and Blake Mere.
)? reserved mater. ial from these mores was examined l by Canter (1 954) who plact s this fungus in a new fillec1 es, Rhi2 o 'hon
akinetimm. This species, and also a similar one which
attacks the akinetes of e 'k. sr. iro: icie s were observed in
Kettle :: ere and White here. On some occasions up to ß01
of the akinctes of these species were infected by the fungi.
The infection appears to be limited to material which is
still in the plankton, those akinetea, which have sedinented
were found to be' free from attack, protected, perhaps, by
the anoxic conditions. Although fungal disease may be an important factor controlling akinete numbers of the above
species, and nay in part account for the small populations
of these algae found in the rieres, other An b enct species
seem not to be affected.
Hence, it is likely that the main cause of the rapid
reduction of akinete numbers is germination shortly after
maturation. Germination, without a resting period has been
observed in cultures of many blue-green algae, in the
original growth medium (Pay, 1969m; Khan, 1974), or on
transfer to fresh medium or following replenishment of
deficient nutrients (Glade, 1914). Immediate germination
would account for the observed recovery of many sporulating
planktonic po ulation: s about a month after the collapse of
a bloom. Al: illete germination is probably more important in
Kettle Fiere than in Whito ! lern, wheru the nur beres of
surviving veg.. tative filaments are generally higher . Although few actual instances of the proces,; of gornination
have bean obsc, "vcd, it was fou. zld th; tt up to 6O, of the
total number of skinlete:.; were empty while : till in the
plankton. ft_luments were sometimes ol)6ervod during
those periods, l, ut these could equally ro ; alt from
fi_l_ar: eiit f raga&; lltatioll and the roueding-off. of the terminal
cells . she germination of alk: inotes tmmuy be . stirultated by a
charts e of comc. itions in the water as th,; y t}lrou,, '. i the
water column,. 1htrtrieiit doneeirtratir'n: r could be of
ir.., ortnrce, b. zt r. -ctin t--(-t -r. iilr, rj_tioF- l; Eotlre(. nn t11H
2' : ý;; UIlU( 3 Of 'L. rLJ t;, 1) chei icaJ_Ly t,, nd f'11i211i1CýIi', r cmJ_1"ý ]. ±verI; L
L

-114-.
neres suggests that light intensity is, the critical factor.
Also the similarity between a range of species, admittedly
of a single genus, indicate that environmental rather than
genetically determined factors control the response of
akinetes. With the sedimentation of akinetes onto the surface
sediments in Kettle Mere and the deeper areas of White Mere,
the light intensity to which the material is exposed is
greatly reduced. Light has been shown to be a requirement
for germination (Harder, 1917a., b; üingh and :; unites, 1974),
and also that light of the red region of the spectrtun in most effective (Harder, 1917b; Reddy et al., 1975).
Although light does penetrate to the sediments in both
meres, the wavelengths above 600 nm are absorbed more
rapidly than the shorter wavelengths by water, the most
penetrating wavelength being at 560 nm (Kawana, 1975).
Wavelengths below 500 nrn, in the blue and violet regions of
the spectrum, are also strongly absorbed by wster (Nestlake, 1966). I. aushik and Kumar (1970) found that
germination of two blue-green algae, A. dolioliun and Fischerella mucicola, could proceed in light of all wave-
lengths. No light requirement was apparant for the
germination of kx)h. floc-anuag (Rose, 1934). The range of
responses encountered suggests that although light is
required, the quality and quantity of light needed by the
various species-is different. However, it would not be
prudent to extrapolate from observations made in the
laboratory to possible events in the field, or from one
species. to another. Conditions, whether they be light,
chemical or other, appear to be conducive to the germination
of those akinetes which have not done so while. stil"_ in
the plant-: ton. This would be a considerable . portion since
the length of time spent in the plankton appears to be
relatively short. The non-germination of a small proportion
of akinetes in the mud leads one to speculate that light
may be the critical factor. Some of the material would be
subjected to conditions of total darkness by being covered
with sediments through d sturbance of. the surface layers
during tli& auturmnnl overturn.

-115-
Resusp©nsion of recently sediraented material is not - considered to be an important factor in the reduction of
akinete numbers, which is Lenerali., º complete while the
meres are still stratified. Fuw anirietes were observed in
the plankton once sedimentation had occurred, even during
periods of maximum decline in numberä. Those akinetea which have sedirnented onto the mud surface in Kettle Mere are not lifted into the plankton during the winter, but appear to be mixed more deeply into the mud. In White Mere,
particularly in areas where the bottom is relatively hard,
very few akinetes are observed during the winter. This may be the result of mixing upwards into the plankton, possibly followed by germination under increased light intensity,
contributing thereby to the overwinterina vegetative
. populations. Numbers in the so'ter mud are generally higher,
which may be due to a thicker covering of sediments hindering resuspension, to a greater depth of sediments in
which akirietes are found as well as to the redeposition of
suspended akinetes removed from other areas of the mere.
Overwinterinj.
Viable akinetes of Anabaena and Alhianizomenon species
have been observed in the mud of many lakes during the
winter after the mass death and subsequent sedimentation of
algal populations (Rose, 1934; Chernousova et al., 1968;
Vladimirova, 1968; Wildman et alo, 1975). The presence of
akinetes, which are generally assumed to be formed under
unfavourable conditions, leads to the conclusion that these
structures will produce by germination new vegetative
populations once more favourable conditions return in the
following spring. Reynolds (1971,1972) observed germination
of akinetes of A. cir. cinalis in Crose Mere while the lake
was still unstratified in the spring. The, inj)ortance of this for population development is considered to be
doubtful (Reynolds, 1972,1973a) since numbers were never
very large and the incr. ease of the plnnktonic populations could be accouated for by the growth of the ovorwintering stock of planktonic vegetative; material.
Fror-i the present invustiL; Fition: s oil I: ottle 1-, ere and
L.

-11 6-
White Mere the same conclusions might be drawn. The
germination of akinetes appears to take place soon after their maturation rather than in the sprint, or early summer
of the following year. The filaments produced may contribute to the revival of the planktonic vedetative population,
particularly in Kettle Mere where little vegetative
material survives a surface bloom. It is this population
which overwinters in a vegetative state in the plankton
and is particularly abundant in white Mere. Further
germination of akinetes may occur during the winter and
spring, resulting in a gradual diminution in numbers of
akinetes in the mud. This pproc'escs appears to be insignificant,
the changes involved are small and may be attributed to
other factors such as attack by pathogens or the mixing into deeper sediments. No sianzficant increase in the
evidence of germination was observed during the spring in
either mere. If the vernal germination of akinetee
contributes at all to the plý-mnktonic populations, this
contribution would be expected to be greater in Kettle Afore,
where the overwintering vegetative populations are much
smaller, than in White Mere.
From this it would appear that akinetes, rather than
acting as an overwintering mechanism, seem to protect the
algal populations from extinction during the extreme conditions of a surface bloom. The thick envelope would
protect there from bacterial attack and they are resistant
to extremes of oxygen tension, light intensity and
temperature. Thus the response of the algal species to
surface blooming and the fate of the akinetes in the
subsequent period appear to be critical for the development
of future populations.
A few comparisons between the different specie, may illustrate the point further. In Kettle 1'ý; ere during 1974 a large population of A h. flog-ngune bloomed in early June.
The numbers ra,. idly declined thereafter and no akinutes
were ob: erved. Only a very minor recovery of the oluxiktonic
population was observed,, in corit'rf,.:, t to those species which did oporitl te, and the , )opulý, itjon levels in 1975 were los:;
than 1, j of the mn:: ii. lur1 Produced in 1974- yTklu, 1976 pC)j)Ll_'it1o11

-117-
had increased sor. iowhat when compared to that pr. - sesrt In
1975, and also, unusually for Kettle 1,: er. e, alcinetes were
produced. ilthough it. is3 not possible to draw firm
conclusions from this, particularly sinco other species
undergo similar fluctuations in the comparatively extreme
conditions of Kettle Fiere, it appears that species which
sporulate regularly on blooming are "buffered" against
further great fluctuations of their abundance due to a deterioration of conditions (Lund, 1965). This can be seen
in White I"Iere, where the population abundance of the main
akinete forming species A. circinalis and A. flora-aquae
exhibits a relatively regular pattern throughout the three
years of the investigation. This does not, however, remove
the influence of environmental factors on the growth of the
vegetative population, on the production of akineten and on
the subsequent development and abundance of future
populations.
Although resistant, akinetes are not totally immune
from attack. As mentioned earlier, a large proportion of
the akinetee of A. solitaries and A. spiroides were observed
to be infected with a chytrid fungus. This would reduce' the
potential overwintering stock bf either akinetes or
filaments (produced by. germination of these species when
con pared with unaffected algal species. If such a tenuous
extrapolation is-to be trusted, then one might expect
fucdre populations to be reduced in comparison to that
which might have developed in the absence of pathogens. The
Marge fluctuations observed in the numbers of these species
in both 1; ettle here and 'white Nero, is certainly in marked
contrast to those of other sporulating species of . 1inabfiena,
. and tends to support the idea of a reduction of the
nbuf ferine; " capacity of the ahinctes due tQ attack by
pathogenic organisms. This, however, is only one factor in
a complex interaction of riany factors, and thus must not be
viewed in isolation froii the rest of the ecology of the
species cor_('ernnc: d .
she colonies o Iiicrc)c-:, tip appear to be mach more
resistant : -cructurns than the vegetative stages of the
ti:; ii cn:: pccio" to the extr"e: ico of conditions, 1711. ieii prevail

-118-
during surface blooming and subsequent sedimcntfutiou. This i-iould be expected if riicroc sties Species are to
survive at all, since-: no Morphologically distinct resting structures are produced. Changes of enzyme coz: iposition and
activity of I'i. aer. ugiiiosa were found as populations
declined in the post-blood phase (: 'ud' ina et fei., 1973).
These changes resulted in a transfer from aerobic to
anaerobic respiration, which would enable colonies to
survive in the mud. Other species, including ADh. f. J. os-
a uae and A. variebilis were also found to be capable of
anaerobic respiration in pure cultures (tiud' ina at rel. , 1973).
Viable colonies, as determined by their fJ. uoresoenee
and their ability to grow in-the laboratory from riud
samples, were found in the Qedjments of both mores between
the time of sedimentation in the autumn and the reappuarence in the plankton during the late summer. A distinct annual
migration of colonies was observed in the deeper lakes: (Sirenko et al., 1969; iopachevskiy et al., 1969; Reynolds
and Rogers, 1976) in which populations of I"Iicrocystis which
sedimented in the autumn to the mud have overwintered there,
and then a proportion of these migrated upwards through the
water column to the epilimnion the following year. The
stimulus to upward migration is probably an increase in the
reducing conditions resulting in a stimulation of
physiological activity (Topachevskiy et a)., 1969) and gas
vacuole formation (Reynolds and Rogers, 1976). A similar
pattern of the distribution of colonies was observed in
Kettle Pifere, which, due to its sheltered position and low
surface area to depth ratio, will result in relatively
undisturbed sediments. she generally lower number of colonies in the mud
during the winter in dhite IIere is probably the result of
turbult. %nce maintaining a larger proportion of the
population in suspension. he same was observed by iýeynolds
(197a) for Grose Mere, which is a. 1 so a relatively shallow body of water in relation to its surface area. In both
Kettle hic-re and White i ere, Irowever, there is an increase
of colonies in the ; nud during; the autumn prod iced by

-119-
sedimentation. Later reduction in numbers in the mud of
Kettle here is mainly due to the mixing of "colonie: i deposited on the . iurface of the mud into the deeper layers
by turbulence insufficient to lift material into the
plankton.
S

SECTION 2
GO , IT"AND CELL DIFFEi3ETJ IATIUI1 UI DER A VARIETY
OF FHY$ICAIý AIM cm iICAL CONDITIONS
hL

TI: ': WDUCýTON
Contrary to previous. investigations, which are discussed fully in the Introduction to Section 1, nutrients,
particularly orthophosphate and nitrate, do not appear to be critical factors in the control of sporulation of
natural populations. Although deficiency of orthophosphate
or nitrate may induce sporulation of the planktonic species
studied, such conditions were not encountered in the meres,
even during bloom formation. The coincidence of akinete
production with the formation of a surface bloom indicates
that the conditions prevalent at, the surface during the
summer may conceal the critical factors. such factors would include a high degree of insolation, depletion of dissolved
carbon dioxide and the accumulation of extracellular
products. A change of light quality as the algal material floats towards the water surface could also be an important
factor. These factors may act directly on the mechanism
controlling akinete di ferentiation, or indirectly through
producing a change of the growth rate or physiological
activity of the algal cells.
Although some aspects of the physiological activity of natural populations will be examined in Section 3, some problems will occur in the intepretation of this type of experimental work. Despite the fact that the incubations
themselves were performed under controlled conditions, the
environment to which the material was previously subjected in the field was only approximately known. ]: ach iiicubc. tion is thus an individual experiment as the conditions and Material chances.
Despite the inherent problems of the selection of
strains adapted to growth in the laboratory environment, it
was considered that the constancy of the conditions of,
growth and the homogeneity of the algal material would
produce useful insight into the process. of growth and differentiation.
It was to toot the validity of the suf, Gestion. s made in the of ýiectiorr 1 that the experiments on isolates of Iplz: nktoniic al, trre from the nuru2r3 herein described
wore aevioed.

rIATnr IAL; i AND MiiTiIODS
Isolation.
The samples of algal material which were used for the
attempts at . c; colation were collected, if sufficiently
abundant, from the plankton, or from mud samples. The use
of damaged or unhealthy material, for example that present during surface blooms, was avoided. Large inoculations (about 10iß volume) of the natural populations were riade into 100 ml conical flasks containing 50 ml dilute culture
media (See pace 126 for composition). The medium used was free of combined nitrogen, which suppressed the growth of
non nitrogen-fixing species of'algae. The rate of growth of bacteria was also initially reduced. in the early phases of
growth after inoculation into the media free of combined
nitrogen. ümall amounts of the mud samples were also
enriched with nutrient media. Although growth would oocur in the natural, unenriched, samples, this was stimulated in
enriched cultures. In the. early stages of growth of these
mixed cultures, all the heterocystous epecies which were
originally present in the sample were still represented,
and this enabled attempts to be made to isolate and grow them all.
As well as the selection of nitrogen-fixing blue-green
algae through the use of media free of combined nitrogen,
temperature treatments were also used in some cases to the
selective disadvantage of other algae (Allen and itanier,
1968). This was only applied to the mud samples collected during the late autumn and winter of 1974, from which a
, wide range of algal taxa were cultivated. The samples were heated at 30 0C for 30 rain in a water bath and then
returned to normal culture conditions. This treatment
appeared to be effective in killing off most of the algae
apart from the blue-green species. Wierinea (1968) has
used this technique to remove non-sporing bacteria from
cultures of akinete forming blue-green algae. However, due
to the potential selection of resistant, strains this
"technique was not generally used here. Sirlilarly, chemical treatments such as the use of aiitibioties, and treatment
with u. v. light were also avoided. Despite these precautions

-123-
selective pressures would be applied as a result of the
choice of media and the culture conditions used.
Further purification was by means of micropipetting
and sterile washing, (Eberly, 1964; Gentile and, Maloney,
1969). From the culture flask in which the algae have been
growing a drop of the suspension was placed on a sterile Petri dish. The suspension was diluted with sterile medium if rendered necessary by the density of the algal filaments.
On the same Petri dish were placed up to six drops of
sterile medium. A micro-pipette was produced by the heating
of the tip of a sterilised plugged Pasteur pipette. Once
the glass had become soft it was rapidly pulled out, thus
producing a fine tube. This was broken to a suitable length
by bending.
Transfers of small volumes of algal suspension were
made from drop to drop. A Bausch and Lomb (Rochester., 11. Y.,
U. S. A. ) stereozoom binocular microscope (up to x 60) was
used to identify and remove single filaments. After five or six washings, a single filament was transferred to a sterile 7 ml serum bottle containing 1 ml medium. This was then
incubated under the conditions described below (page 128 ).
this procedure was repeated for each species which was isolated. Also for each species, at least 20 separate filaments were isolated and incubated into serum bottles.
The manipulations were carried out under aseptic conditions in a Iii croflow Limited (Fleet, runts. ) inoculation chamber. Because of the continuous current of sterile air which
passes out through the ckbinet, care was taken that the
drops of liquid did not dry out. When necessary additions
of sterile distilled water were made. In those isolations in which growtrl occurred further
transfers were made into sterile media in 1 C)U nl conical
flasks. The material was also es. +mined microscopically for
Purity. Altholzßli with this isolation procedure unial. gal isolates were obtained, no axenic cultures were produced. gable 19 gives n list of strains isolated and cul turod.
While st: veral species survived for liraited periods in
laboratory culture, it was nut pe.:: 3ib]. e to maintain them for lcrig periods in culture. The mo: 5t uL1 ptc:, ble L;, pcciuo,
.L

-124-
Table 19. List of the species obtained and maintained in
culture, their source and method of isolation.
Collected and isolated in 1974.
Species Site Source : Micro- manip.
E'utrient enrichment
Temperature treatment
Anh. f1 os-Flr}uae !P1 Mud + + +
Anh. floc-educe WTI Mud + + +
Anahaerin up. (? ) Wm Mud + + -
A. floc-nguae 111.1 Pl + + -
A. : niroides NM Pl + + - A. circinalis WPI Pl + + - A. sol itaria E. ß Pl + + -
Sites: - aI = Kettle Mere, WN White Mero, rIT-I = Newton More,
EM = The Mere , Ellesmere.
Source: - Pl = Plankton. (? ) = tent ativo ident ification.
which grew well under the conditions used, appeared to be
Aph. floc-aquae, followed by the smaller celled Anabasna
species. This is reflected in the range of species that it
was possible to maintain for more than two years in culture. The larger celled Anahaena species, particularly A. soiroides
and A. solitaria, but also A. circinalis were, by contrast, difficult to maintain for any length of time. This type of
planktonic algae has, however, been successfully isolated
and grown. Volk and Phinney (1968) report the culture of
A. mpiroides in Gerloff et al. 's (1950) modification of Chu's (1942) number. 10 medium. Mc Lachlan and Gorham's (1961) ASTA medium was also successfully used by these
authors.
A factor which may be of importance in the survival
of single filaments of planktonic algae is their buoyancy.
Eberly (1964) suggests that the trapping of single filaments
in the meniscus and the subsequent drying of the medium may be a factor contributing to the low percentage of survival
of inocula of single filament: 3. The effect of this factor
was reduced in the isolation inocula by twice or thrice
daily mixirr;; continuous agitation rapidly breaks up the
filaments of the planktonic species Inver tieatcd . These
factors may contribute to the low degreo of survival of the

-125-
large celled species. These species, however., appear to be
inherently more fragile in any case, filaments and cells
were often observed to break up during the process of
micromanipulation and sterile washing, despite careful handling. Gerloff et al. (1950) observed that cultures of
planktonic blue-green algae are only established with large
inocula. The importance of optimal concentrations of
glycollic acid, an extracellular product, in the medium
for the rapid growth of cultures is discussed by Fogg (1965).
Culture Media.
Of the various media tested, ASPI 4 supported the
active growth of the greatest range of the planktonic
species. Other media tried, in decreasing, order of success,
were: - Allen and Arnon's (1955) medium, A. floh-actuaa
medium (Slalsby and ichelbarger, 1968), ABM 1 (Gorham at R_ ., 1964) and G©rloff at al. 's (1950) modification of Chu' E; (1942) number 10 medium.
The composition of the ASPI 4 medium used in this
investigation is basically similar to the ASPI 1 medium
used by Gorham at al. (1964), (Table 20). The major ions
are present in the same concentrations but the trace
elements are generally increased four-fold. Ferric chloride is increased ten-fol in concentration and sodium molybdate is an extra constituent. Sodium EDTA is also increased
five-fold in concentration to the AUM 1 medium.
Preparation Of Stock Solutions And-r-ledia.
All the conetituente of the medium were of analytical
grade cheriicale and were dissolved in Pyrex giae distilled
water. Sufficient of each salt was dissolved in one litre
of distilled water
over that used in
make up 11 of the
dic3eolved in water
hydrochloric acid.
dark.
to produce a thou$and-fold concentration the medium. Thus 1 r1 of each stock would
medium. The ferric chloride wac
acidified to a Iii of about 1 .5 with
The stock solutions were stored in the
äufl, i c" mt volume: of each of the otock -; Olutions t7t r. o added to hair the final volume of distilled ': r+ti. r to

-126-
Table 20. Composition of AB:.. 4 medium.. (Smith and Foy, 1 974 )
Compound
A Full-ytrenj; th rid 1 µrao,.
1
I)iluýe mg 1-
i3 (10,. ) E1rnol
1-T
Podium nitrate (NaII03) ** 850 10000 85 1000
Magnesium sulphate 49 200 4.9 20
(I"IgSO1 . 71120)
Magnesium chloride 41 200 4.1 20 0"IgC12.611
20) Calcium chloride 29 200 2.9 20 (CaC12.2H20)
Di-potassiurz hydrogen ortho- 17.4 100 1.74 10
phosphate (K2IiPO4)
Di-sodium hydrogen ortho- 14.2 100 1.42 10
HPO ) phosphate (Na 2 4
Ethylobinediaminetetraaeetie 35 100 3.5 10
acid, disodiuri salt (Ila2 EDTA)
Ferric chloride (FeC13)' 6.5 40 0.65 4.0 Boric acid (H3BO3) 8.0 16U 0.8 16
Manganous chloride . 4.0 28 0.4 2.8
(MnOl2 . 411 20)
Zinc chloride (ZnCl2) 1.2 12.8 0.12 1.28
Cobaltouß chloride 0.08 0.32 0.008 0.032 (CoCl2.6H20)
Cupric chloride 0.004 0.0032/0.0004 0.00032 (CuCl2.21I20 )
Sodium molybdate 2.4 10 0.24 1.0 (ITa2DIo04.2h20)
**, Sodium nitrate was generally omitted.
produce the final concentration of the constituents ne
given in table 20. Section A of the table (full otrenCtti)
givos the concentrations usually uuecl for this rmedium,
which is similar to that often uced for laboratory batch
cultures. The concentrations given in section B of the
table, which are 10, j of the full , strength conct ntrrations,
WFG:: that , nenernlly used for the culture of algae j, -, plated

-127-
from the raerr: e. The dilute raediura WEM used throughout for
most of the experimental work on there isolates.
The additions of stock solutionb were made in the
order given in the table, and between each addition the
medium was mixed. The orthophosphate salts were not
generally added at this stage except when dilute media were
prepared. After making up almost to the final volume with
distilled water, the pH was checked using a Pye Unicam
Limited (Cambridge, England) Model 290 pH meter and
adjusted to a value of 7.5 with dilute sodium hydroxide
and hydrochloric acid solutions. Initially the pH of the
medium was low, due to the acidified solution of ferric
chloride. After adjustment of the pH the medium was made
up to the final volume, taking into account any further
-additions which may be required.
50 ml of the medium was dispensed into 100 ml conical
flasks (or 100 ml in 250 ml flasks), which were then
plugged with non-absorbent cotton wool. The plugs were
capped with aluminium foil. These small volumes of medium,
and other apparatus, were steam sterilized at 120 oC
(15 p. s. i. g., = 10.35 kN m-2) for 15 min. Larger volumes of
medium (above 1 1) were autoclaved for 25 min at the above
pressure.
After cooling, appropriate additions of orthophosphate
were made: This was done to avoid the precipitation of
polyvalent cations which would occur if the whole medium
had been autoclaved (Good and Izawa, 1972). Precipitation
was particularly noticeable in the full strength medium. In
the dilute medium, however, precipitation was not apparent,
or only of ight, if all the constituents were mixed before
autoclaving. The prepared sterile medium was not used for at least
24 h after <: utoelaving, and before use the p11 of a spare
aliquot was checked.
The Culture Of Algae.
1t) ml conical ftn: skc were used for the maintenance of
stock cultures and also for some of the experiment:, i1 work.
lnoculnti, -n$ into frw:: iii raediu111 were carried out under aseptic

-128-
conditions usin sterile plugged Pasteur pipettes. An
inoculurm of about 2,, volume was used . Stoclc " cultures were
grown under "Daylight" fluorescent tubes (Ecko, I"'. azda or
Phillips) at a luminous flux of 1000 to 2000 lux. (For
further details on light conditions, see the section on "Light, Fluorescent" below, page 129 ). The temperature was
ambient laboratory temperatures, which fluctuated during
the year from 16 to 26 0C. The'naterial was grown under
static conditions, as it was found that many of the species
isolated did not grow when agitated. This was also observed
for A. sxpi roides by Volk and Phinney (1968). Many blue-green
algae do not Crow if shaken tog- vigorously (Pay and Fogg,
1962) and -others show optimal shaking speeds (1'ocg and Than Tun, 1960). A degree of mixing is, however, generally
considered beneficial for rapid growth, and an evenly
suspended culture and homogeneity of treatment, particularly
of light, of all the cells, The species which were isolated
appear to be highly sensitive to agitation, but are
generally maintained in a suspended, evenly distributed
condition in static cultures duo to the presence of gas
vacuoles within the cells and an absence of large quantities
of mucilage. A limited degree of mixing was supplied
through gentle swirling of the cultures three or four times
a day. The isolates of Aph. f_los-ßryuae appear to be an
exception to the above. Although the filaments were evenly
suspended in static culture, this species could tolerate
mixing, shaking or gentle aeration through a sinter. This
enabled these isolates to be grown in the type of
continuous culture apparatus described by Pty and
Kulasooriya (1973).
Buffers.
hero buffered nt dia were used , the buffer was added
after adjusting the p: i of the medium before fiutoclavinC.
Buffers used ', are tris (} ydroxynýeth;, ý1} acainornethene (THIS)
sied ýý-2-hydro yothylpitýera7ine-Tý' -? _-ethailot. ulýýhvnic acid T}le cation binding cajtci. ties of these compounds
are ne 1if, ibý_ý: (Good e al., 1. x)66) and they apppear, through
not beirr; effected by autocl"iv: iur-, to be heut eteble (smith

-129-
and Foy, 1 971,4). Three dipeptide- buffers, DL-alai l- lycine,
DL-alanrl DL-alrtnine and T, -glycil-g? ycine, were also
te. ited. A ranee of concentration:, from 0.5 to 4.0
were used for =iiýPi::: 3 and : iltlä, and a range of 0.05 to 0.5
g 1-1 for the peptide buffers. All the buffers except TC{IÜ
were obtained from Signa Chemical Co. Ltd., äurrey. ri"IZIS
was obtained from B. D. 11. Ltd.
Teml2erature.
Much of the initial experimental work was done under
ambient temperature conditions in the laboratory. Although
the temperature varies-significantly throughout the year,
fluctuations were generally small for the duration of a
short-term experiment of a few hours. Ranges are given with
results of each experiment where controlled conditions were
not used and the fluctuations significant.
Experiments on growth rates were carried out under
controlled conditions in a Grant (Cambridge, LnCland)
thermostatic water bath. A refrigeration coil was required
to maintain a constant temperature below 25 °C, particularly
during the summer months. Experiments were carried out at
20 ±1 °C .
During 1975 a Gallenkamp (London) cooled illuminated
incubator, model IIi 287 was used for the storage of stock
cultures and some experimäntal work. Either continuous
illumination or a 12 h light - 12 h dark cycle was used.
For the maintenance of stock cultures a temperature of
10 0C was selected. for other experimental work 15 oC
was
used.
Li t; ht
Fluorescent.
stock cultures were grown under lateral illumination
by, "Daylight" fluorescent tubes. she light output had a
peak e1lergy at 58? 0 nn, falling off rapidly at increasing
wavelen, -t11s to 25, -' of the peak cutout at 650 nr. 1 and 5/, ') at
7Ui, rill. At shorter wavelengths the decline i": ras l e. 3,; rapid,
with a shoulder at 4 30 I'm of 40,, of the peak output. Energy
at ý: a\re t: ". t; t}13 below 3 j0 In was norligib e \ý'ig. 3--J)
ý.

-1: 5U-
Fig. 33. Spectral distribution of energy output of
"Daylight" fluorescent tubes. Redrawn from Thorn Ltd.
Technical Pocket Book.
100
75
50
25
0 300 400 500 600 700
Wavelength, nm
Vertical scale: - mll nm band width for a 1500 mm tube at 65 W
The light intensity used for stock cultures was 1000 to
2000 lux, and for general experimental purposes 2500 lux.
For some experiments using fluorescent tubes intensities
of up to 10000 lux (10 klux) were obtained. Data will be
given where relevant. An Lvans Electroseleniwn Ltd. (Halstead, Essex portafle photoelectric photometer was
used- to measure luminous flux.
High Intensity.
tilhen a very high light intensity was required to
illuninatu algal cultures, a G. L. C. Ltd. (London) tungsten
halogen lamp (from I"Ialhari Photographic, London) was used.
The energy output from the lamp, which was housed in a
reflector, was 1415 W m-2 at 500 ram from the filament. A
value of 45 klux was obtained under the same conditions. The wattage was measured with a Kip-) and Zonen (Delft,
Holland) model AL 4 F: icrova galvanometer and a cocnpen:; ated
therr: onile type CA 1.
Due to the high output of heat from this lamp, the
culture ve ; el i were protected by a water, cooled Per: pev
shield internored Bett een lartip Ctwi cultures "1 11:, was in

-131-
the form of a covered shallow tray with provieiori for a
rapid flow of cooling water between the eurfucess. The upper
surface of the shield was not lamm than 500 rrrr from the
lamp fi. lamexit, to prevent local overheating;. Cultures for
incubation were placed immediately below the shield. A
mall fan was used to circulate the air between the shield
and the cultures and also above the shield. Temperature
control was effective, with no more than a2 0C rise in
the temperature of water placed under the shield during a 5h period of the maximum intensity used.
Approximately 7O, 'of the total power output of the
lamp is absorbed by the water-filled shield. That this was
primarily in the red and longer wavelengths of the energy
spectrum can be determined from the greater degree of
transmission of luminous fluX., About 4Uij of the light
measured using a photometer is absorbed. The maximum
luminous flux at the liquid culture surface was 15 klux,
and the wattage 270 11 m 2. The cultures were about 600 mm.
from the filament.
To produce a more even distribution of light throughout
the culture and a more precis Ly determined light climate,
the incubations were done in plastic Petri dishes. 15 ml of
the algal suspension was pipetted into each dish, producing
a shallow pool 2 mm deep. The cultures were covered with
the plastic lids to reduce evaporation and the volumes were
made up to a pre-calibrated mark with distilled water if
required.
A12h light - 12 11 dark cycle was used. The temperature
did not vary by more than 1 °C (from 20 °C) throughout the
incubation period. This was for a maxiriur1 of one week. As well as using natural populationo of planktonic
algal material collected from the mere., cultures of the
laboratory gr. own i solateoo were used. This material war, not
buffered by using any of the compounds described in the
section on buffers (above, page 123 ). The algal isolates
were initially pr. epL red as described in the section on the
culture of algae (Pace 1,27 ) and for each strain throe
replicate-, were treated. Two "stages of 1'rowth of the stock
Culture?:; wc.: re'. u feel LAB U Zporinnertal ciatorial. These were

-1ý2-
actively groFzing material in the mid logarithmic phc. aso ,
and post logarithmic, stationary phase cultures. The
re;; pect-i ve ages of the cultures, under the conditions used
for the growth of stock cultures, were 8 and 17 days. From
cultures of these algae material was harvested and
incubated further under the conditions of strong irradiation
described above.
Small volumes (about 1.0 ml) of the incubations were
collected daily from each replicate for microscopic
examination.
A luminous flux of up to 10 klux wa obtained using
fluorescent tubes without heating of the cultures. A fan
was sufficiently effective in mni. ritainin¬; the ambient room
temperature in the vicinity of the culture area, so that
no further precautions were required when fluorescent tubes
were used as the light source. Apart fron this detail the
procedure for high light intensity experiments was the
same as that described above. Due to the reduced -fire risk,
experiments were also performed under continuous illumination.
Monochromatic, Action Spectra. . To obtain light of a sufficiently high level of energy
and of a precisely known and narrow wave-band, a range of
interference filters were used in conjunction with a Rank
Aldis Tutor 2 transparency projector. The interference
filters, of 47 mm effective diameter, were obtained from
Carl Zeiss, Jena (C. Z. Scientific Instruments Ltd., London).
The half width (the range of wavelengths transmitted at
half the peak energy tratnsm_zission) for the range of filters
used was 6 to 9 nni. A li Lhtpr. oof filter holder was fitted to
the condenser of the projector such that the rim of the
filter was in contact with the concen er. This ensured that
the filter was normal to the incident light. The condenser
was adjusted so that it would be focussed at the maximum
distance allowed. oiith this Bettir. the beam would be
approximately parallel. Shift of the peak wavelenCths to a
shorter wavelencth would be minimal for angles of tilt up
to 1 Uv. Thhi s shift in proportioria]_ to the square of the
tilt, fi71gle and would be F bout , of the wuveleIl1 th for
I

-1_ý
; lf-10C:. nor: l_t1 to t}'; (E: inc . de :t
light hC: am, an Ct': t. rý, ure of 2". C t'i7"UCýuce r c'" ., no rlý_ý... ýý,: 1r1ko
c'r_:. _ ge in the on.
f1J_ ýer c ial'i? Cte: ri Ectico was obtained fro t}'. e I':: f. 21i1 i(ic. t: ltrc ': 3
data :.. }1<ii: ts '_. Ct Cýit£iý Uf; ue (and,
u?.: -, o from of
Gr(t105 1. lrJY . Yet'"(, 'iu. J
'-ý upon f?
/ 1I( ý' C4 11,. _....
c". 4 V_ upon
The Y)o`. Jkex, of the p-)Y") j ector Li: of a. 3 Vi i) !! Fi'+; ý.: 3 Ai1. J_G}1
produced an en: -, rte,:, flu, ": of 3 ': 1 ri-` at 5C() to 6C'() r. il: l from
the light source with the filters in. plcac . : °hE3 J>o^: i Lion of
the incuý)atio_i vessels was varied . io cis to maintain a vu
cono trtnt energy flux throuühJ, the experiment for each
wave? _e}zg; th. 7)u(: to the fru*ittmcy of lamp
automatic record was ýýept of the length of bu. rn_ý. ný;. An
electric ctoc.: we.,, -, svvlitc_1(. d on or Off vile :t : L: it; }lt oy)c: r: ýteti
relay. A cadImiuam üulp''}tide photo-conductive cell wee used
and the circuit wired no that the relay was activated when
the light ' eveý_ increased above pan. tarbiý: ar,, value set u: 3in
a T)otentior! eter. The recorded tii, c in therefore the time
which has elapsed since the projector was switched on to
the time when -witched off or t' r. ou h bu:! 1) failure. All l_ the
componexit a were obtained through Radio Oý1),, Arct3 Cor1poI'? Hnt S
Ltdd' London.
The a ý_ t^ilter7 :ý ý_ used were the algal . solute grown
under the condition dC: EscribeCL in the section on the
Pn -fe 1 27 . T"! --o .: cat: x_ : 11 was tii:; e: t when in
the early o; iri t: rzic pha e of growth. Three re;, lici tern of
each _E?: 3 at each wavelength wter(. -, incl. ii:, Itede The
incubation Yt:: E 1; W -rc 10 mm UZF±�; ic .:? iE: (: . rO1)h )t 01 :1 ter
This E3?.: 'ü? ^ed call even iIIII. -lirlatiou
over the , >11rface
of the ctl-Iture. Although -the cul tsrc. s w(. 1vu not C'<; 1 ß. 1f i Clý
tue Ut most of the isolates : icl. ectod for this
evo: L7_yv sus, ? 7t'. nd dl " 2)4Me, ho-o(-, vor, became
})L1U; '_1: _ý; aid floated to the Hltr1aco of the ']i : ý1Lf[1" N: l: 'tý 1.
to h": II. -ICdI due to the f=!: all area of il on.
. '. ra11.. v'. rý_rl
wrmý poSitiuno d nor. ýr. -i1_ to t1iE-e I. nciüc2ilt l_. a betr: a aad- : ueh

-1: )-
-11'3: a "Ic; rL.; ' flu, -, ö.! 1Lta r: a rucuiv"d. Y', '-. Cl.
waE 'L-,. ouseci in a 1±. F; ht 'tight bo; c tiritli t11(:, projector sIrii ii. rij,
t}: rough :a holt it onto ý--n . 2hß to:,, pcrrituro dIid not e: --reed
the ambi nt tý . "Ytt. ]Rýitu ? out:; i d6 the ox, and the e: c. -crir, ent. ^. a
Wore executed at c: ter 7F? rature of 23 OC. (, nu. W r. "o' eng th Q
l4'7xýnj :a alr$o dolle n; 10 1 C. 5) C, thfý npp ra us, at
that time, being houst. c. in n C(lü-roori.
mt the termination L )-f incubation ;. period, t_unorally
after about 7ý h, the reJ-licrýter were cýxrý. i±ncci ciicroz copica--
1ýy.
Ultra-violet.
or irradi&itiorl of ulaa1 raut.: ria1 withi u. v. 1i lit, tý 0
Hank-via (31our; h, Berk,: hiru) fiuorcscenco lamp I: Qdul 19 way
used. Tki ru-tcd to-ýu l out put at 5C0 im i fror ti, i: -2 -2 lamp Was l; vi ra , o" which 1V 'ki m vi w-3 in t. ho rci; iori of
3'o 370 27x2. di, flor3nce i: l the rt:: )1.1ý. t of heatink, up
of the ? amp during, operation,, which in also measured 'by the
-therrio ie us-AA
she cu? tares word prepared in the swne way as the,
material used for the action spectrum (above), i. e. they
were in the early logarithr iic phc ie of growth. 15 ml I.
al_± quots were dispensed into pla. Jt: Lc Ye. tr: i. dishes ac!. d. were.
thee: Of 't-i,. -, U iI'"i. i i to
2 lr. Dur- rrf the z) r aci of ixr.: d"on the eovrs 'Jere 2 -(,
ft
Ci "1'" tº__tc r the i: Y' at 1e 1. ý, ]. Et J`± d t': G Cam:, t? C :. r:: p-lc.. Ci 'E'lld
she prates p: :c":: 1 under
'For .. QC'ig - er x Uji to 1 l) ;! of lo 1,. -vE; '_: i of
L1. V. Fl : 3inmi1_rir ilrOCC. 1. ure was i. tcio tool ,
h1. C1^It, VUT for
t jý C7 1C ýU ;L ý_ciý "QZ_.
i ý'ae'_^C' l1: if Cý" . ý_
Il j- .,, @X r'2 rý `-lE: iv I. ýit11]^2i1
'2ho atlUtlilt on 011(2.119.
V'1^f tiT COVF, 2'. __r c the E
of layers of 0,2mc _, of nittji1, '. A "(-a"iccd t1
t3; 'tG11: 11; O rncli tiori by . boll- 5U.,,. A' iii. i
YE? `ý. j ? (_ý Cü2. ttL1 t? s were further incubated tind (? r normatl.
i-"_1.1-uaJ. n. at 3_U11.
iýL1L' tw'7C? Tc !i C11. J 7i( LLtJ: t 1()Z1 Ci(3: iCr. l )l': ý i! L)C)VC tJ41`k?
perfor!,. ed at I-oi':, ': L11 lilÜZt Zlý ý. '! ti L)7 : story 1 C'; 1Y t; Z'; l't t1]'L'.; of

-135-
Lxper. i hunt a at controlled wt; ro dont3 in :. l
water bath at 22 0C. In t1ies"e experir,: entE the norc', zl
f uoreocent 1. J_. 11r1inaticni of 800 lux was ll?? iý_f; C: i?. ̂ . tOCl with -2
u. v. radiation of-10 ,+m. The culture-, were; crown izi
250 : "11 conioul 1a. 31: s which were otopperc: 2 wi to non
absorbent cotton wool harms. The tray or. vwhiah the f1s0lts
were put in the crater bath was moved : slowly -uh rough itbout 15 linear oruci]. 1 Ltions per in co that rý1Jr_ the fla., Acs
rece-5_ved an equal Eiuaunt of light but thu cooaitcnts were not
unduly' Sha1zon" ý''he'' algal $u$poncii0 n: i v ere illuminated
continually throughout the duration of t; he ex1jar. irnent .
x a_ Iýýiteri ? Asse :: anent Of
T: icroscopic ex imination Of each of the three repl is teD
were made at (usually daily) interv: tiF; during nn experiment
and at the end of the experä^zeýýtý7_ period.. 'mints were
made of heterociyst and akinete frc cjuency Fis described in
the, Methods to 2oc tion 1 (rage 43 ;. Obuervatio: s were
also made on the appearence of the r-illral cultures and of
the filaments and cells.
Determinations of Cell density, and from this
calculat: +. ono of the growth rate, were made uuing an L`vci. nm
klectroc; eien±um Ltd (Halstead, Essex) colorimeter. A red
filter was used. The mean doubling time, G, in hours, was
calculated fron the relative growth constant, k (Fogg,
1965). The relationship between o. )t-i_cal density readings
and cell density ', zcs found to be linear (Nef±crt, 1 971 ).
loG1 v ODt -1 log10 ÜJ0
t ihere t=t Lrn intorwil in }hcau. rG )etween
r*ieasurcr1ent of OD vag u ,: ̂ .
ÜD Optic il don Ly a±ter. t hours.
0. UJ() - Optical cien:; ityy when t --
To cnlou1at' G: - G-0.301
16,

X36
Rl: jULT:;
Growth.
'the 'pattern of growth exi: ibiteci by the algal isolates
is of tree lc;, rpi (. EL]. . '"i. E C: 'ioidal form. The two strains of
Arph. isolated re:; pectively from Kettle Mere
and ýI_hite ',.. ere, we. 'e the crost extensively used of the
iF, o_! atoms from the rieros for growth experiments. This wren
due to the ease of culture., rapid growth, t)od suspending
qualities and lack of clumping, and a close correlation
between cell nurý. 'aers and optical dentmit4-. Data for the
grow l; h constant, k, in log10 units day-1 and the mean
doiib7 in g time, G, in clays, for different conditions; fire
given in table 21 . The data presented here, and in other
tables, are the mean of three replicates.
Table 21 . Growth rates of isolates of .4hf. los-aS uae.
leraperature: - 20 0C. Light: - 3700 lux, continuouf.; i11,. r ±natinn. 10jo inoculuri.
Piedium (ASM 4) Kettle
k hero
G Wh: itcj
k Mora
G
10; x, +1,103, +P04 0.207 1.45 0.128 2.36
" -I' J3 " 0.1 60 1.88 0.164 1.83
50iÄ, +110 ," 0.156 1.93 0.196 1.56
to _1; 03, it 0.119 2.54. 0.252 1.19
Teinperatuuro :- 23 to 28 qC (Moan = 25
continuous illumiiiý-ition. 2ý) i. rioculum.
10iß, +I? C: 3, +PU4 0.176 1.71
_1iO3 11 0.261 1.15
5C,; +11t 3, it C). ', U1 1 . G0
,: -I]0 ,"0.272 1.11
00) . Licht: - 2500 lux,
0.207 1.45
0.176 1.71
0.326 0.92
0.301 1.00
: note: - Values represent the maximum rates achieved, over a
two day period.
ýýhr. i rates obtained were similar to tLo. -e:, of other
fi? _'':. lentoucý blue-green algae under similar conditions (l''ny
and F: ul asooriya, 1 97,45; Smith and 2P'oy, 1')74; Foy et 11.. , 1 ; 76). Gro r li in riedie co.: t: sir _rýt a :; ourcc: of co ibined
the 7 : ý7 i1- nt. ý_y lower, than

-137-
groiith in media free of combined nit. °ogen in the Kettle
.. ere ct ' in. The reverse tends to be true for the '. hhite i-iero strain ,
--0_
thcupJI the d ffereiice! 3 in both otrn r, were sr alý_. : 3oth strz"i.: _ns e h1 bit Q similar response, to an
increase of 'CE; L1perature, with an increase of growth rate,
despite the lo or light intensity used. Increase of biorna: s
of both strains proceeds at a sir filar. rate. Cultures in dilute medium (1Q,; full strength, column
B, table 20) had similar maximum rates as those in the more
concentrated (5C;
' of full strength) media. The , row, h
curves in the two media follow similar patterns until the
'10th to 14th day after inoculation, when nutrients
orthophooDhate) in the dilute mediuri are exhausted. Tho
final yield is from. t: ro to three times higher in the
concentrated media than in the dilute. $irailarities of
gro. ith rmtos in media of d;. ff. er. ent concentrations was shown
by Shubert (1974) for äconede: 3rrius.
'ºi±th a 2; c inoculurn, u lag phase of 5 or 6 days wen
observed. This was reduced to 4 or 5 days when the size of
the inoculum was increased to 10,,, ". No change in the runximum
growth rate or final yield was produced.
Buffers.
During the period of active growth there was a marked. increase of pH. Several buffers were used to maintain a
constant ph throughout the growth cycle. Of those tested,
only TR: f proved to be to:: ic to the two strains of
Apl,. flo-R. , Iu ,e used. High concentrations (above 0.02,: ß w/v)
of the dipoppti'e buffers were also dctrimmýmtal to grswth. In the case of the dä- ýeý_, +; ide buf. ' ̀ 'ers this was probably not
a result of --direct toxicity, but through the encourcgement
of excessive bacterial growth. Concentrations of below
O. O1; ý w/v provod to no effective in maintaining a stable p-'I
throujllout the. growth of the alga and also reduced the
amount of bacterial g;. rowth. Macro:: copically the cultures
Zookerl healthy, and viewc: d micro;;, copical7. y the mater.? n1
ýaýppeared nor: n. al. Filaments in cultures buf'fcsred with
were, however, unwiually long. This v. a;,
probably due to a reduction of frc1; _. 'nentation as a renul. t of

-1 33-
the absence of h. eterocy. sts, even in media
which suggests that U 1ycyl-ý 1, ºcine, the
di-popticle , is utilised by the filga as a
combined nitrogen. This is interesting in
that relatively high concentretiona of ni-
free of nitrate, F: i:! lplet3t of
Source of
view of the fact
Irate do not
completely inhibit differentiation.
HEPI. S buffer, used by ämith and Foy (1974) was
observed to have the least c! ffect on algal growth and
morphology of the range of buffers tooted (But see table 26).
Growth constants and mean doubling times in 10/'; AM4
medium buffered with a range of concentrations of lIET i
are given in table 22. "
Table 22. Effect of IL: Yl'J. i buffer on the growth of
planktonic algal isolates.
Temperature: - 24 to 25 0C. Light: - 3500 lux, continuous fluorescent. k in log, 0 units day-1, G in days.
Ash* f]os-a ime Anaba'ena sp (7 IL PB 1I isolate WZ1 isolate ttilhito More g 1-ý kGkGkG
0 0.261 1.15 0.238.1.26 0.249 1.21
0.5 0.261 1.15 0.261 1.15 0.318 0.94
1.0 0.162 1.85 0.219 1.37 0.382 0.78
2.0 0.21 21 1.41 0.1 98 1.51 0.301 1.00
4.0 0.199 1.51 0.131 2.29 0.471 0.64*
8.0 0.079 3.76 Very little 0.500 0.60*
*, Of doubtful accuracy duo to the low population densities.
The molecular weicht of 230.3, and a concentration
of 0.0211, used by Smith and Foy (1974), is equivalent to
4.77 g l-1.
With increasing concentration.; of F1EY;?.: ý up to 4a 1-
the maximum growth rate of the Aph. floc-a iiae: was reduced
slightly. Thies spociei appears to be nensitive to higher
concentrationO. by contrast, strains of A. flos-LIauae and Ap, h. flo. 9-F3_n_Liae exactined by 3riitti and Foy (1974) rho: ed a r -duct-3-on of tho clean douralizig tic: io in raediti buffered with
corpared with unbuffered controls. The 'White:
, --,, re icio_ati3 of A'. 1}'1. iýý Oý-ýt( llflt? Wa; i riore Cnol1F31ti. ve to

I 'Z
-19-
concentrations of buffer above 2g 1-1 than the Kettle Ii ere
strain, which showed a Tradual decline of growth rýto with
increasing concentrations of At 2g 1-1 and below,
there was ? ittle effect on the growth rate throughout the
growth cycle, when compared to control cultures. In the
Kettle Isere strain, however, the final yield was increased
by up to 501, of the control (unbuffered) cultures, for
cultures buffered with 0-5 or 1.0 g 1- 1 HEPL: 3.
Unlike the A, h. flos-a uae strains, the growth rate of
the Anabaena sp (? ) showed a marked increase with increasing
buffer concentrations. With increasing concentrations of
'HEP: sS the lag phase was lengthened. At 8g 1-1 significant
growth did not occur until after 12 drys in culture,
whereas in the unbuffered controls and in the cultures with
-low concentrations of buffer, the lag phase was lec, 3 than
three days. A similar extension of the lag phrase of growth
was not observed in the Aph. floh-a uao cultures. The
yields obt ned in the buffered and unbuffered cultures
were similar at the same stage of growth in the different
treatments except at the highest concentrations of buffer.
pH control using this buffer was effective. The pH did
not fluctuate by more than 0.2 unite from the pä value of
7.55.
Tautrients And t$: iorulntion.
The response of the Kettle : lere and 'dhitß I4ere strain.,
of Aýýh. f. 1n: ý"'quae to varf. ous concentrations of nitrate and
orthophosphate are given in tables 23 and 24. No akinetea
were produced by the White : ere 'strain under any of the
conbinations of trea, t:: ients, even after the cultures had
l! orig passed into the stationary p}ia: ie of growth (Thblo 23).
Sj; orulation did proceed in the Kettle here strain,
part icul r: ly in old cultures (29 days after inoculation,
table 24). Akinetes were observed in younger cultures,
particularly in those with low coitcentrationo, of nitrate
(anca normal ortho: tho:. phate concentrations). !. *umbers vere
. generally low in terns of porcentaýe frequency, and only
one in 10 or one in 20 h©teroc: 'o-cc, were oboorved to have an
Ke. mciatecl ak: tnete. : L± iLer 1)ureuiitlt; e spw7uiatiOn HI. C&
ý. -

-140-
ca E
U UI "
O O
.N
ci -' cý r" 1
4i 4. ) O O 5
Fw Or r 9.1 "ý U O r-I H ö
1ý_ c3 U tO c$ Ll v
. rl ay " 4-3 0 rd co a) 4-) r Pi " i. Pi
C) d) U ü) 4-+ z U 0 O 4-4 :5 (1 "r 1 C d) Pi O C c3 0
1 0 rl 4l +) a) H O U 4.1
U1
O ,O O O
N Z z .N ". i " O cd 4.4 +3 V- F-4 0 54 i q
0 " -1 " r O c- t
cd 0
>~ +ý Pr c'3 U) r. uZ I O
r1 N L -P N N I
Q 4) "0 ri c- 0 4 1 "
"r4 C) " O
- tJ O (\j . 4. ) r4 0 " V L, M O cJ " O
4-t 0 s3 O O fti
4-) r"
U N ý. O Q)
4.4 $ O W C)
N 0
Q t: 0 a E-4 E-4
O O O O O
O O O " O O O
0 .0
0 0 0 0
IIIIII
rn OOO. O0
o N O O N
O O
O O to ý" O O O O
(v r-
M %lo Ü
N O O " O O rd H 0
Ö r4
0 0 0 0 H
U +3 " -5 Q
H
Ö U O U
t) 0
H pý
0
nc 4 .) V 4. ) N O O
, n V1ý, Y)
J\ r. lý)
U
Y'Z V1
UR L N\ c) 0 0 r11 U c) c)
. - "i
m 0 s4I L 0 c0 C4 0
.C z" 4 y 4 4'
:z "x ": i =: 's ý

-141-
H
WIN c to ;s 41 O " O O " O O " O O o c- N c- O
., -1 +) Cd o C 0 1 " " O O " O O " " + I. I) () WN N h1 N G O
o z.
0 c c- v7 U1 N - 0 -d" N
rd A I - U1 O O U -: I- " (NJ U1 rl W H " " " " " O 0 " - ýý O N O O -. t N c- O 4 z t0 a) " 4-3 -1,
V-
;4 a a) cc a ) 0 U1 (\J 0
0 0 4-( C 41 O C) " O O 1 1 1 -rf N O P-s 0 N N 'd I 1-( 0
u) 0 , c: i es
H H H F -, 0) 4+ 4-1 0 U
" C] 41 All o 0
Q ý
4-1 4-1 -H 0 c O --) t- t4r\ O
' ON 0 ö . -I N O o O O I 1 1
ti "d ,o .. ý C ct H u: 1 0
Q p O O 1 1
O 0 9 '- N N O O "
U ý ;. ( 4 I 0 c2 0 O ýn ( ý c o .. 1 0 " O 0 0 O ti
, S-ý a. ) a) cO O tll c- O O N P. { 4; +' U ti -I-' O h O
Is" O N 0 0 o ä
r\ ö 00 ül 0 (D O O " C) C) O Cl) rd
41 ýýl N
4.1 O 0 O O O iq
4-31 t- 0 4) rl O u1 O cV co O in, O O r C) N Z " 7 N . c- ON - O O r"
4-1 1 O rý n O "- o O 0 04 r-+ w c) a) fi U
. j" "N 4) +ý +ý r-1 C\J c3 .L G) U) 4ý Cw
0 " 0 'H ri P, U Ul C 0 0 0 t1 .Q £ f.. L It, C3 4) Q) h C) 4., E-4 £-+ a-) 4 -) D I1 n ,
4-a I 4. a N" O Uý I U I 4 z. e;
N -a C: V- r; C: i. , 1a c.. - V" V Q) U C", U N C. ('ý j U C) ti)
S"{ 0 f-(
-P 0) Q)
1 S (ý iý 4) U i+
0 ý+
4-' CJ -C-) 4+
Q)
C) 4) c: c; I ro r; { ýu . m r- 4 41 rJ r: "Y ý-, '
_ý 4ýj 4)
L

-142-
heterocyst frequenc-11111 ere observed at 17 and 29 days after
inoculation. The data for 21 days after inoculation,
however, does not show a similar pattern. Despite the
presence of akinotes, no increase in percentage frequency
was observed and the ratio between heterocyst and akin©te
numbers remained relatively constant throughout the
duration of the e°p-),.: ri. ment.
A similar pett:: rn was observed when the orthophosphate
concentration was varied, with a normal concentration of
nitrate. Few or no akinetes were found until 29 days after inoculation, when a marked increase was observed. Cultures
with low orthophosplint e concentrations had stopped growing before this time with no production of akinetes, Fand none
were observed in cultures grown at 10 mg 1-1 orthophosphorue. hIowever, for orthophoeT.; hate concentratione between 0.1 and 1.0 mg 1-1 about 1 of the total cell nudger produced
akinetes. A stimulation of sporulation was observed on several
occasions in media containing nitrate nitrogen, but this
response wee not consistent. Cultures grown in media
deficient in orthophosphate and nitrate, or at low
concentrations of these two ions, did not sporulrite. This
was true also of natural populations of planktonic algae (A. circinaalis and Aj, )h. floe-sause collected during the
early summers of 1 975 and 1 976) as well an the A ph.
uae isolates.
Other components of the medium, particularly trace
elements aid iron (che lated and ur_chelat(-:., d), were
px-amined in a similar manner to orthophosphate and nitrate.
:: one were found to induce sporulation to any greater
extent than that of control cultures, although growth was
occasionally greatly reduced.
The results described above were obtained from batch
cultures in unbuffered media. As mentioned in the section
on Results (Page 137) some of the di-peptide bu_°fer. o
tested had an effect on heterocy: st dif. 'erentiation. It was
thus t}iouj; ht unadvicable to use such buffers for experimento
in which the presence and frequency of hats>roc ya. ts could be
critical, is ich as the forr; ation of a'" irroter; Gent}:, 1965,

-143-
1966). ". i: ýP: ý: ý buffer was found to elir; htly ine "easo
heter. ocyst frequency during the period of active growth (Table 25). This was particularly noticai)ie for the
Table 25. Effect of HAPAJ buffer on hotorocyet frequency.
Frequency of heterocysto given arg a percentage of the total
cell number. Observations made 10 and 16 days after
inoculation. See table 22 for culture condition c.
. flos-acixr-v: Anabac. ne op. (?
HEP1 3 Kettle ; 'giere White Mere White Mere
g 1-1 10 days 16 (lays 10 days 16 days, 10 days 16 days
0 4.9 3.4 3.3 * 6.8 +ý*
0.5 6.0 4.5 4.6 2.2 6.2 5.1
1.0 6.6 .
3.6 5.0 2.1 7.1 5.4
. 2.0 5.9 5.4 - 3. ) 6.3 7.2
4.0 7.4 5.0 .-*5.6 4.2
8.0 - 4.3 - 2.0 - 6.5
** , Filaments fragment-ed. *, Breakdown of ale,, al material (lysis? ).
-, No data, little growth at time of sampling.
Aph-. flos-adac isolates, the differences between. the
Anab_zenn sp. (? ) cultures being less marked. A possible
factor may be the lower rate of growth in the buffered
cultures, although with the reduction of growth rate as the
buffer concentration increases there was no parallel
increase in heterocy: 3t frequency. Thus, after 10 days, the
buffered culture will be at an earlier, stage of the growth
cycle. This was more clearly seen in the nettle Mere strain
of Anh. floc-Lo: cluae, where, at similar stages of growth (measured by optical density), the heter. ocyot frequencies
were similar. The unbuffered culture after 10 days growth
would be comparable to the buffer, , -d cultures after 16 days.
This difference in growth pattern was not so clearly marked
in the White Mere isolate, in which the growth of unbuffered
cultures closely matched that of cultures with low
concentrations of buffer (to 1g 1-1
When akinete frequency was assessed a more distinct
-pattern was observed, particularly for the Kettle Nero
isolate of froh . f? _os-n. yune (Table 26).

-144-
Table 26. Effect of buffer on sporulation.
Frequency of aki_netes given an numbers per 100 heterocysts.
Observations made 10, "16 and 21 days after inoculation.
See table 22 for culture conditions.
g 1-1 os-
10 daIV76 16 acaua day6 21 days 10 da. -, 7o 16 days
? 21 days
0 0 0 0 13() 200 200+
0.5 0 0 18 31 65 200+
1.0 0 10 24 33 134 133
2.0 0 6 13 21 57 98
4.0 0 2 7 7 63 154 8.0 0 1 24 0 2 70
Notes. The White Miere strain of ATh. flo:. -rinuae did not
produce akinetes in any of the cultures at any time during
the experiment.
Due to the clumping arid- fragmentation of filaments,
accurate enumeration of chains of akinetee Vans; difficult.
M-1 = Kettle Mere.
Unbuffered cultures did not produce any akinotev in the
Kettle Mere strain of Aph. floc--pause throughout the period
of examination. However, even in the low concentrations of
buffer a marked stimulation of sporulation was observed
after 16 days. The density of algal material in this case
war, similar to that
culture. The latter was, by this ti
phase whereas the buffered cultures
active growth up to 21 days. Higher
buffer,. although producing cultures
in the unbuffered
ne, in the stationary
etili. showed Figno of
concentrations of
of lower bioraues at
16 days, still )roduces a stimulation of sporulatic)ri. The
pil of the media at this stage were similar (7.5 in the
buffered culture, 6.6 in the unbuf, 'ered culture, after the,
fall from a maximum of 9.5 at 12 days) . At 21 days after
_y similar, inoculation the pattern of response WHO basicall
but with ,; reatc r differences between the buffered and
unbuffered cultures. The optical density of the cultures
grown in ° .OL; 1 was only one : sixth of that of the
unbuffered cultures and the P11 Values were similar (7.6 in

-145-
the unbuffered culture, 7.7 in the buffered culture). A more "normal" pattern of growth anci sporuleition was
exhibited by the Eiiabaena ep. (? ) used. This algal strain,
of ur_de t ermined significance in the natural plar_ktonie
environment, sporulates prolifically in normal unbuffered batch culture after 5 to 7 days growth (i. e. at the end of
the logarithmic phase). In this respect it differs from the
Aph. floe-aquae isolates, which, in corirlon with many other
isolates of planktonic algae, only produces akinetes in old
cultures long in the stationary phase of growth.
After 10 days growth the unbuffered cultures, which had reached the statioziary phase, had produced large
numbers of akiretes. Akinetes. were also produced in the
buffered cultures, but to a lesser extent. The amount of
algal material in these cultt1res was less than in the
unbuffsred cultures. By the tie the buffered cultures had
reached a similar density, and also stage of growth, a
similar number of akinetes was observed to that produced in
the unbuffered cultures. Th; " effect of the retardation of
rapid growth can be clearly seen in the maximum concentration
of buffer used (8 g 1-1). In this culture little growth had
occurred by 10 days and no akinetes were observed. After. 16
days some growth had occurred and a few akinetos were
observed and by 21 days a similar number of akinetes were found as in the early stages of sporulation of the other.
cultures. The influence of culture filtrates was also examined on
the progress of sporulation in these algae (Fisher and Wolk,
1976). The data for the "baena ep. (? ) is given in table 27. Ho marked stimulation of akinete production was observed, sporulr-! lion proceeding in an approximately normal manner
after about 1 wevk of growth. Earlier differentiation of
akinetes occurred in media deficient of orthhophosphorus.
Earlier differentiation would also be expected to occur if
extracellular products produced by old cultures stimulated
sporulation. A greater, but not significantly earlier,
differentiation was observed in material inoculated into
older culture filtrates. This declined as the rrowortioii of
addition of fresh media increased. The extent of spor. iz_lation
L

-146-
Table 27. Sporu ration of Anabz<: nft tap. (7) under diffcrent
nutrient conditions.
Temperature: - 23 °C C. Light:! 4 klux, "I)aylieht" fluorescent,
continuous illumination. Tledin and addition:, sterilized by
aseptic membrane filtration.
Frequency of akinetes expressed as numbers per 100
heterocysts.
Unbuffered Buffered media
media (1 g 1- 1 1I}: 1, ES)
Media plus additions 5 days 7 days 5 days 7 days
ASM 4 (+PO4, -NO3) .0 30 0 0
ASM 4 (-P04 , -1103) 27 47 0 39
100.1 Aph. f-a (FNNI) filt. 0 88 0 50
50iä ++ It If 0 2 0 0
1iG it it It 0 0 0 U
oo, ü to (uri) If 0 32 0 0
500 it to 0 25 0 1 If it " 0 27 0 26
100j1 Anabaana sp. (? ) flit. 0 69 0 69
50io it .0 40 0 24
IS if 0 19 0 31
100iä Kettle Mere water " 0 0 0 6
50, o +4 0 19 0 9
1 ýu nu ýr 0 33 0 11
100iä v+hite Mere Water 0 79 17 65
500 to u If 0 30 0 40
1 j'ý "nn 0 24 0 20
20, E Kettle I"Iere water (-P04) 0 113 0 21
20, ä White Fiere water 0 128 0 0
20`, 0 ti h. f-a U11) flit., -1'04 0 25 0 1U5
20, E it (UM) "" It 0 18 .0 72
20; ö Arirabaena sp. (? )" " 59 E30 29 119
Notes. Where mixtures were use d, propo rtions were m ade up
; aitiz A; X-i 4 (10iQ) . This mediun containe d ort hophosph ate but
comlýý.. nF 11itrogen as nitrvate w as omitt ed. ( -Po4) ortho-
phospI: fate omitted fro!. medium. (-NO j) = nit rate omi tted.
iß4 = Kettle Mere. WM = White M ere. flit, = culture filtrate.

- 147-
in un--enriched culture., filtrates was similar to that
! produced in orthophosphate deficient media. Differences
between culture filtrates were possibly due to differences
in nutrient depletion by the different algae, although no
analy.. e: were done to test this a: -sunption.
Filtered water samples from the mores (Kettle Iere and
in the lowest (1'* %) White Here) produced similar responses
concentration used. Higher concentrations of Kettle fare
water produced a lower degree of sporulnntion than those
cultures grown in White Mere water. This, like the
differences between culture filtrates, was possibly A.
-result of the differing orthophofz?, hate concentrations. The
concentration of orthophosphate in these water samples was
0.7 and 0.4 mg 1-1 PO4-P respectively for Kettle Niere ctn.
White I ere.
When, instead of using normal orthophosphate
containing medium, phosphorus deficient media were used to
supplement-the cultures, a general increase of spor-ulation
was observed. Values we're lower in buffered cultures,
possibly due to the reduction of growth observed in cultures
buffered with H1, PES' (see above). Cultures which contained a
proportion of Anabeena op. (? ) culture filtrate produced a
considerable increase of sporulation, even after 5 days
of growth when the other cultures had not yet sporulated.
Similar experiments were also performed with the Kottlo
Meru strain of 1: Ah. . 'los-t. Lq e but no oporulation was
obeerved beyond that normally encountered in old cultures
or in media buffered with The White i Jere strain
typically did not sporulate.
Lit-, ht c izantityArid b-poruý_at. ion.
As described above, sore strains of alrac will produce
akinetes under certain nutrient conditions. The source of
light in all cases was continuous fluorescent illumination
by "Daylight" '! amps at betveen 2 and 4 klux. some cultures
were exposed to a cyclic light environment of 12 h light - 12 h da. rl: at 15 0C but otherwise r, rnilar conditions to that
describ: ad above. These conditions dich not produce an incrt: ase of akineto production when compared to a similar

-14 i-
length of e; xpooure, to continuous light. Under the
intt n: sitie: 3 used no increase of growth in the li4,, ht period
was observed over that produced under cent: ý. nuou. $ iJ lwiinat-
ion. The ariount of material produced 'war proportional to
the length of illumination.
Cultures of Anabaena cy'lindrica L. which were grown
under normal light conditions and then transferred to the
dark ceased to grow and did not sporulate. port term
re-illumination (fron 30 s to 2 h) of culturco kept in the
dark for 3 days and then returned to darknens produced. no
effect on gr-owth or sporulation. These experiments were
done in inorganic media without a source of combined
nitrogen. A similar cessation of growth and absence of
sporulation was observed when cultures wore : pubjected to
high light intensities. Cultures of the species listed in
table 19 did not grow when'placed under fluorescent
illumination of 8 to 10 klux. Exposure to high intensity
light from the tungsten, halogen lamp had a similar affect.
Return to normal light intensities produced normal growth.
A similar response was also noted in natural planktonic
populations. This material, particularly for experiments at
high light intensity, was collected from White Mere during
the early months of 1976. The dominant alga was
A. circi. na. lio and some A. . floc-aquae was Hlao present.
No increase of sporulation, apart from those akinetco
already present, was observed when post-lo; arithmic
cultures were incubated under high light intensity (also
8 klux). A similar lull of growth and other activities, to
that found when juvenile cultures tiyere used, was observed.
Cultures of all ages became bleached when exposed to high
light intensity.
Tai. ; 1Lt t'uolity And Jnorul-ition.
At the wavelengths used (475,575, E25,675 725 and
875 nri) only one case of induction of sporulation bias
observed under the conditions applied and for the list of
specit. ý: 3 given in table 19. At the end of the incubation
period of about 7> h coat±nuou.; 7.7 liar ination the nic-al

-149-
material was in an apparantly healthy state. Little growth had occurred, but no determinations were made.
Of the material used only the Anaba nn op. (? )
produced any akinetes. A omall number, 29 per 100 heterocysts and 19 per 100 heterocysts, were observed in incubations at 625 and 675 n.: i respectively. This was
probably due to growth and develoymeiit during the incubation
period since the stock material used for these oxperimente
was in the early logarithmic phase of growth (4 days old). Thus at the time of examination (i. e. after a total of
about 7 days) sporulation would have occurred anyway in
normal cultures.
Ultra-Violet ? d, ht And porultttion.
One of the most noticeable effects of continuous irradiation with relatively low energies of u. v. was a
marked diminution of growth (Table 28).
Table 28. Effect , of u. v. on algal growth.
Temperature: - 22 '! ' 0.5 °C. Light: - 800 lux "Daylight"
fluorescent and 1.0 w m-2 u. v., continuous. Optical
density measured after. 10 days ¬; owth.
Control U. D. Treated U. D. Algal material (- u. v. ) (+ u. v. )
A13h. fl oa-aquae (El; ) 205 ý.. w.
0.9 " (wI2) 2.0 0.2
A. floc-aquae (V{; 1) 1.0 O A. circinalis (Tai) 0.6 0.1
12"I, Kettle Mere isolate. UN, White Piere isolate.
In common with other factors (such as high or low light
intensity) which reduce the growth rite, the treatments
used with u. v. light did not stimulate sporti?. ation. she effects of short term doses of higher levels of
u. v. were investigated for the two otraino of A )h. floc-aguac (Table 29). Although there was a slight increase in numbers with increasing doses of radiation, this was relatively small-and the result;, were variable. Via effect of u. v. was not consi: steat between experim,. ntc;. äim filar experiments

-15U-
Table 29. Effect of short u. v. doses on the eporuiation of Animo. f? os-ac,, zae isolates from Kettle i"Iere and White Nere.
Dosage rate :-1U 6i rj-2 . After treatment the cultures wure
grown under the conditions described in the section on The Culture of Algae (Pate 12.7). Observations were made 2
wee s after treatment (no sporulation before). A: cinete frequency expressed as numbers Per 1 UL) 2ietorocycrts.
Length of u. v. dose (min)
Akinete frequency
Iongth of u. v. Akinete dose (rain) frequency
0 72 8
0.5 45, 16 83 1 132 32 64 2 70 64 171 4 102 128 194
Note. ho akinetes were observed i n any of the treatrtento
for the White Mere strain of Aiph.
performed with the other isolates did riot stimulate akinete
production. Both short and long term exposures to high and low levels, respectively, of u. v. light also failed to
produce a response in the natural A. cireinalis populations
collected from White Piere during the early months of 1976.
Observations were generally made at daily intervals in
the period after treatment. In those species which normally
sporulate (such as the Anabaerna sp. (? ) from White Mere)
after a period in culture, akinetes were observed in the
treated cultures after the normal period of time. In the
case of the Anrbaena sp. - (? ) this would be after about 1
week. However, in those treatments which had been subjccted to a greater dose of u. v. (30 min and above) fewer akinetee
were observed. he amount of material produced by these
cultures throu. 'ri growth ciao aluo lower.
These culzures, includinni; the natural nlariktonic Populations of ý ci rcir a1_i from White i"lere, when transferred to low light (10 to 100 lux) Titer incubation
with u. v., did not eporulate. At 3 and 10 klux. increasing,
numbers of akinete. s were observed, particularly in the
untreated cultures of A. o---, 'able 30) . Although the. IL. flc `3ýlCll1,
ýst , which was, F1 minor compollont of the
L

-151-
gb1. e 3C). Effect of u. v., and fluorescent ? _J-,
ht intensity,
on the sporulation of A. =los-Hgtije collected from White
Mere, spring 1976.
u. v. treatment: - 5 rein at 10 w m- 2. Temperature: - 23 to
25 0C. Observations made 5 days after treatment. After
treatment with u. v. the cultures were grown under the light
intensities indicated. Akinete frequency expressed ao
numbers per 100 heterocycte.
Light intensity lux Control u. v. treated
10 to 100 00
3t)OO 51
10000 32 7
Note. No akinctes before 3 days after treatment.
phytoplankton at the time of collection, produced akinetee, the dominant alga, A. cir. cinalis, did not. The material
appeared healthy, although the amount of growth in the u. v. treated cultures was less than that of the control. The
cultures, particularly in the high light intensity treatment,
were taken over by A. flos-ftguae. The filaments of A. oireinatis became fragmented after only 1 or 2 days
exposure to light of 3 and 10 klux intensity.

DI$CUS3ION
Growth. And Bporulation.
'With few exceptions, sporulation of laboratory cultures,
was not observed until the late stationary phase of growth.
This, or the complete absence of sporulation, appears to be
a feature of isolates of heterocystous planktonic algae.
Eberly (1966) did not find akinetes of t h. flos-£«iuae
after a period in laboratory culture although Wildman et al.
" (1975) observed akinetes in old cultures of the same
species. Schwabe and . strange - Bursche (1964) observed that
. although akinetes were produced in natural populations of
Aphanizotnenon gracile Lerrn. during the sutntner, there was
generally no sporulation in laboratory culture.
A eigrificar_t departure from the eenoral behaviour was
observed in cultures of the Kettle hero strain of Aph. floc-
aquae buffered with HEPES. 5porulation started earlier in
the buffered cultures, while no akinetes were observed in
the unbuffered control-cultures after 3 weeks of growth.
Although the control culture had, by this time, reached the
stationary phase of growth, the buffered cultures were
still actively Crowing. Culture density did not appear to
significantly affect the differentiation of akinetes. Those
cultures gro n __. the proecnce of higher concentrations of
which had suffered an extended lag phase qnd
cane=eq, zent: ya lower biomass after a giver p(-. rind of tirle,
had sporti_fiýL- G_ý to an equal extort as the denser. cultures >d.
in l_o,, rcr conccntration. c of
A1. thour h it would appear that IIEYES has some
stimulating effect on the sporulation of Aph. floS-Rguae,
other factors need to be considered. Nutrient depletion is
expected to be similar in cultures of similar densities and
the n. r during the period of akinete formation and
immediately prior to it were similar in the control and
buffered culture . Also, in eulturu.: e buffered at a similar
p11 using other buffers, no stimulation of sporulation was
obeorved. Thus, in those cultures in which little growth had occurred, such ne iin high co: cetvations of a
lower deL; reo of snorulation would he expecte(i if nutrient
deficiency i3 tile rinin caua factor.

-153-
These observations, however, are relavrnnt to only one
strain of one species. The other strain of Anh. flog=-aaune,
from Weite Were, did not produce any akinetea at all, in
any of the treatments. The Anabaena op. (? } provided a more
typical response. Oporulation was lower in cultures in
which there was less growth. This was the case in cultures
buffered with increasing concentrations of HEUS, in which,
although the growth rate tended to increase, the lag phase
was extended. Ater a period of growth subsequent to
inoculation the amount of algal material in the buffered
cultures was lower than"in the control culture. This was
reflected in the lower degree of sporulrstion, which,
however, increased to an amount equivalent to that observed
in control cultures. This occurred once the optical density
had increased to a similar leve*. in the buffor. ed cultures
as that of the control cultures. Like with the Aph. fl. oe-
actuaae strain, no akinetes Were observed before the
stationary phase, although sporul_ation in this isolate
proceeded at a somewhat earlier phase of the growth cycle
than in the Aph. floe-ac uaQ cultures.
Inorfanic Nutrients And Sporulation.
Cultures which were inoculated into full inorganic
media (except of the omission of combined nitrogen
compounds) and allowed to grow under normal conditions
occasionally produced akinetes at some stage in the poet-
logarithmic phase of growth. It is generally considered
that eporulation is induced by nutrient deficiency. Ortho-
phosphate deficiency has been found to induce early
differentiation in cultures of A. cvlirdrica (t1olk, 1965)
and Aph. ! os-aquae (Gentile and Maloney, 1969). Although
orthophosphate was reduced to indetectable concentrations
(below 5.0 N; 1-1) in old cultures of Anh. floe-Aquae
(whether or not they have sporulated), no induction of
sporulation was observed in those cultures which were
inoculated into orthophoc: phate deficient media. The
Aniibaena : p. (? ) on the other hand did chow a significantly
earlier production of akinotee in those media deficient in
orthophosphate. This strain of alEa, although isolated from

-154-
White hors, is not 'one of thu typical planktonic forms. Its
response to nutrient deficiency more resembles that of the
terrestrial isolates of algae (such as A. Rorliidrica,
A. doliolum, Cy' indrospernum etc. ), general? _y used by
previous investigators, than that of the planktonic species. While low orthophosphate concentrations did not
stimulate the sporu1r: tion of Arch. floc-c une, high
concentrations (10 mg 1-1) appeared to be inhibitory. Thus
it would appear that orthophosphate plays some role,
although the response was not as marked as observations on
other species. The failure of induction of aporulation of
cultures inoculated directly into phosphate deficient
media suggests that other nutrients may be of importance.
Another factor may be the requirement of a certain amount
of growth and development per se before differentiation of
vegetative cells into akinetee can occur.
Ariong the other inorganic nutrients tested was nitrate. Previous work has shown that nitrate deficiency induces
sporulation in C lindrosZermum (Glade, 1914) and that in
A. doliolum sporulation is inhibited by nitrate (Tyagi,
1974). Singh and Srivastava (196¬3) also suggest that
combined nitrogen is the controlling factor of morphogenesis, including sporulation, -of A. doliolum. In the Ah. floe-
aquae cultures there was an inverse relationship between
nitrate concentration and *heterocyst frequ mcy, which was
more marked in the White Mere strain. This is typical of the heterocystous species (Fogg, 1949; Kr1le et 1970).
Akinete formation appeared to be neither stimulated nor
inhibited by the presence of nitrate, although at the stage
of akinete production the concentration of nitrate would be
expected to be low. (This, however, was not measured).
Despite these observations, however, it was found that on
some occasions a marked stimulation of eporulation was
produced in cultures of Aph. floh-aeutiey (from Nettle Here)
which were grown in media coutaininf, nitrates.
From these observations, and also from the failure to
find -another single nutrient element (such ßc iron or any
of the trace elements) which would. cif*, nificrintly stimulate
sporulation, it would app(--ur that thto factors involved are

-155-
rnoro cor: l: lu than that as., ociated with the depletion of a
single nutri ent. The respoima of the terrestrial forms,
however, a, ->>ears to be, somewhat more' straight-tor award than
that of the pltinktoriic species. Although nitrogen and
phoophorus metabolisn appear to be critical factors, the
stimulation of akinote differentiation in the planktonic
species is probably a complex interaction between these and
some other factors. Possj, bl© candidates for ouch other
factors may be other nutrients, the age and development of
the cultures, and the presence, of extraeellular. products.
-Other Coripound: a And . norulation.
Organic co. pounds, either as extracellular products
produced by the algal cells (Fogg, 1965; Huntsman and
. Barber, 19'75) or through the activity of aseoeiated
bacteria (raunt, 1 961 ; smith, 1973) are often essential for
the'growth of algae in cultures. This is particularly true
when small- inocula are used and during the early phaeos of
growth. Apart from the stimulatory effect on the early
phases of growth, extracellular products have been found to
stimulate the production of akin. etos in C. licheniformo (Fisher and 'lolk, 1976). Although a similar type of response
was not observed for the strains of algae isolated from the
meres, one compound, the buffer k LPL6, was found to
stimulate siorulation in the Kettle Here strain of
Ash. floc-a(iuae. This process seemed to be independant of
the concentration of the compound (HOPES) for the rang:
tested.
Sometimes other morphological effects were also
observed in cultures to which certain organic compounds
were added. The complete suppression of heterocysts of
Aph. flus-nauae was noticed in cultures buffered with
N glycyl-gI. -vcine. (A similar effect was observed in
Anabaena o. -cil! ý,, rioides Bory by Demeter (1956) in a culture
media which contained sodium Elutamate). This is in
contrast to the finding that in cultures of Anh. flr.: -aquýio heterocysts were still present, even at high concentrations
of nitrate. It is pos. sLble that 11 E;? _ycy1--(; lycine is taken
up and assimilated b, - the exlE; rj, producing; an inhibit on of

-156-
heterocyst differentiation. That this is more complete than
the suppression of differentiation in nitrate containing
media is probably a reflection of the lower energy required
for di-peptide uptake and assimilation than that which is
required for nitrate reduction. However, since the cultures
were not axenic this must remain a hypothetical conclusion
as bacterial metAbolicm of N glycyl-glycine could have
produced conditions responsible for the described
inhibition of heterocyst differentiation.
As expected from the relationship' between hoterocysts
and akinete differentiation (Yolk, 1966) no akinetea were
found in cultures in which heterocyst differentiation had
been suppressed. This is in contrast to the observations of
Demeter (1956) that in the cultures growing with sodium
glutamate akinetes were observed, despite the absence of
heterocysts. This appears to be a direct stimulation of
sporulation by this compound, such as that observed with
HEYES buffer. Demeter (1956) found that both heterocysts
and akinetes were rare in cultures grown in media from
which sodium glutamate was omitted. The medium used by
Demeter (1956) was based on that developed by Allen (1952)
and included potassium nitrate, ammonium chloride and
di-potassium hydrogen orthophosphate. In media free of
combined nitrogen both het©rooysts and akinetes were present
in abundance (Demeter, 1956).
Li, -ht Quantity And Sporulation.
Contrary to the expectations from observations of
natural ponulations sampled during the summer, high light
intensity did not stimulate sporulation. \olk (1965)
observed a reduction of sporulation of A. c yiin. iri en, at
light intensities above 800 lux, and above 2500 lux few
akinetes were formed. A reduction of sporulation was also
observed under low light conditions. The observation by
Ylolk (1965) were made after a fixed period of time, and thus
it is possible that the cultures under different light
treatments had. reached different staken of growth at the
time of sar. °, pling.
Cu1. tu-" e: which were exposed to high light intennsity
l

-157-
did not grow. This was also true of cultures kept in the
dark or'unc? er low light interisity. As well as cell division
and enlargement, other metabolic processes api)ear to be
halted or reduced under these conditions. The result i5 an
absence of akinetes, even in the natural materiel used for
some of the experiments. It is, therefore, appar. Ant that
sporulation in laboratory cultures will only proceed after
a certain amount of growth, and if that growth be reduced
sporulation will be retarded. However, even when post- logarithmic phase laboratory cultures were used no
sporulation was observed.
The natural material used, collected during the spring
of 1976, probably represents actively growing, healthy,
logarithmic-phase culture3. If this was the o ee, than a
-similar response to that of the laboratory cultures used
while in the logarithmic phase of growth, can be expected.
Light Quality And Sporulstion.
Illumination of algal material with light of different
wavelengths produced a similar response to that discussed
above. When there was little or insufficient growth, no
akinetes were produced. The type of photo-inducable
developmental cycle observed for IT. muscorum by Lazaroff
and Vishniac (1961,1962) and for N. commune by Robinson
and Miller (1970) was not apparant. Induction of the
I4ostocacean developmental cycle was induced by a brief
exposure of the dark grown material to light of 650 um and
reversal brought about by exposure to greon light. Although
light of these spectral regions was used in the experiments
described here, the absence of response possibly indicates
a major difference between the life cycles and developmental
stimulikf the terrestial Nostoc species and the planktonic
Anabaena and. Aphanizor"; enon species. The a1 eence of a
response of the planktonic isolates to illumination with a
tungsten filament bulb after 2 days of dark trontment
supports this supposition.
Kau., hik and Kumar (1970) found that, apart from white licht, red light supported the creatost rate of growth of A. d liolum. The growtL-h in blue light wn lower, and jr.

-158-
green l id; hht miriirul. The transfer of cultures, grown in
white (tungsten) light for up to 4 days, into Green or blue
lig: it conditions produced no sporulation. IU: i netes were
observed, however, when cultures grown in white: light for
6 to 8 days were transferred to blue or green light. This
response is similar to that encountered with the Anabaonr-t
sp. (? ) used in this study, although Kau:; hilc rand Kumnr
(1970) did not indicate the age of cultures in which
akinetes would be observed when maintained in white light
only.
Ultra-Violet Light And-S; por. ulation.
Treatment with u. v. light was reminiscent of the other
light treatments in that extensive sporulation was not
observed in cultures in which growth was inhibited. Singh
et al. (1967) observed an earlier differentiation of
akinetes of A. doliolum and Qylindr. or; permum maius Kuetz.
when treated with u. v. light. In these e;: perirnc: nts, and also
in others in which u. v. was used to produce mutant strains
of blue-green algae (Singh, 1967; Singh and Dikchit, 1976;
Sinha and Kumar, 1973) a shorter-wavelength of u. v. light
(about 254 nm) w. s used than that applied in the experiments
described here.
The proportion in solar radiation of wavelengths below
300 nrl which reaches the earth's surface is very small (Collingbourne, 1966) when compared with that of other
wavelengths in the near u. v. and visible spectrum. A
considerable proportion of energy at a wavelength of 365 nm,
which was that used in this otud,, reaches the earths
surface during clear weather. Thus the u. v. wavelength
tested bears some resemblance to that encountered during
sunny days when algal material has formed surface blooms.

SECTION 3
THIS PHYSIOLOGICAL ACTIVITY OF NATUI A3
POPULATIONS of ALGAE)

II PRODUCTION
Although some physical and chemical factors have been
described and diocues d in t; cction 1 in terns of their
relevance to algal growth and the life cycle, the measured
values of these factors represent the external environment
to which the populations as a whole were exposed, rather
than the environment within a cell or for a particular
species. Knowledge of the concentration of certain disoolved
compounds gives little information of their availability to
the organieem. Marked local differences of the concentration
of nutrients may occur, particularly during a relatively
stagnant period. Such conditioiis may impose the limitation
of diffusive supply. Although plentifully present in the
general eater body, depletion of nutrients may occur in the
vicinity of an algal cell or colony, or within the cell
itself. Other factors could also produce local depletion of
nutrients. Bacteria, through their more favourable surface
area to volume ratio, take up and utilize nutrients ouch as
phosphates more rapidly than algae (Fuhr at n3-. 0 1972; Rhee,
1972).
Nutrient, particularly phosphate, availability may be
affected indirectly by the presence of trace elements.
Shapiro and Glass (1975) reported that additions of manganesee
as well as phosphates had a very marked effect on algal
growth when compared with phosphate additions alone.
Various other trace elements have been shown to be required
by blue-green algae, including cobalt (Holm-Hansen at al.,
1954) and copper (Telitchen:, o et al., 1970). Iron
concentration was found to determine which becomes the
dominant algal species in mixed batch cultures ([Torton and
Leo, 1974). Enzyme activity is often dependant on the
presence of trace elements. Molybdenum is a requirement for
the synthesis of both nitrogenase and nitrate reductn. se (Pay and Vasconcelos, 1974) and cobalt for the formation of
vitamin B1 2
(Holm-iiansen et al., 1954). As well as their
importance: in ; physiological processes, some tract) elecmontu:
have a vi: j4ble effect on the morphology of the species.
i: hitton et _. (1975) observed that terminal hairs are
produced in filaments of Ctllothrjx yiinlieri in iron

-161-
deficient culturfes. A similar effect was observed in
filaments of Gloeotrichin when grown in a medium free from
combined nitrogen. ün addition of an ammonium salt, the
filaments appeared uniform throughout (}: 'ay et al., 1963).
Phosphorus has been shown to have a controlling influence
on the colonial morphology of $cened. osrmus (Trainor and Shubert, 1974; Shubert and Trainor, 1974). The shape and
size of the "packets" in which algal cells are grouped may be expected to produce nutrient limitations through an increase in the length of the diffusive pathway in large
colonies or clumps. it has been suggested, on the other hand, that the gelatinous sheath of blue-green algae may form a favourable microhabitat for algal growth through the
concentration of limiting nutrients (Lange, 1976).
Alternative sources of potentially limiting nutrients
may be important during periods of stratification and
surface blooms. The hydrolysis of organic phosphorus
compounds in natural water samples has been shown to occur (Reichardt, 1971; Herbes et al., 1975), and an inverse
relationship between alkaline phosphatase activity and
orthophosphate concentration has' been demonstrated (Heaney,
1973; Healey, 1973; Ihlenfeldt and Gibson, 1975). Heath and Cooke (1975) observed up to a 30-fold increase in alkaline
phosphatase activity during a bloom of ADh. floe-a use in a
eutr: ophic lake. The authors suggest that alkaline
phosphatase activity could be an important source of
orthophosphate provided that a sufficient concentration of
phosphornonoesters are present in the waters. Fitzgerald
and Nelson (1966) also observed a large increase in enzyme
activity in phosphorus depleted cultures of natural
material over that found in culture: which were supplemented
with phosphates. This inverse relationship is only
applicable to phosphorus starved coils. In continuous
culture experiments using A. floc-figur-, e, hone (1971 ) ums
unable to determine any relationship between alkaline
phosphata; se: activity and labile call phosphates. In this
case phosphorus would be limiting au a result of activo and
rapid g, rowth, rather than the culls being starved. L itratu
was observed to stimulate enzyme activity in these
S

-162-
exper. j. r.: ents, as a consequence of the creator yield
resulting in an increased phosphorus demand. In the natural
environment no correlation between enzyme activity and
phoaphorus uptake or cell phosphorus was found during the
course of a bloom of xphh. floc-aquae (ilealey and Hend . cal,
1976). levels of activity were variabl. e, despite the
nitrogen and phosphorus deficient nature of the algal
material at the end of the bloom.
On the restoration of phosphorus sufficient conditions,
a rapid uptake of phosphorus occurs which is in excess of
requirements (Ketchum, 1939; Batterton and Van Ballen,
1968; Sicko-Goad and Jensen, 1976). Sufficient is
accumulated to support two further generation:; of growth
when the material is transferred to phosphorus deficient
medium (Batterton and Van Baalen, 1968). Stewart and
Alexander (1971) found that sufficierit phosphorus is
assimilated and stored within the cells of nitrogen-fixing
blue-green algae during one day to supply the requirements
of growth at 25 0C for one week. A long period of growth
of A. spiroides at the expense of stored reserves of
phosphorus has been observed by Volk and Yhixiney (1968).
These reserves are stored as polyphosphate (volutin) bodies
and may contain the major fraction of the total cell
phosphorus (Sicl: o-Goad et nl., 1975). The phosphorus is
stored in an osmotically inactive state as long chain
inorganic polymers (polyphosphates) (. >icko-Goad and Jensen,
1974).
In addition to the effects on growth, phosphoru.,
" deficient nay also affect the availability of other
nutrients. Stewart et ale (1970) and Stewart and Alexander
(1971) observed a rapid increase in nitrogenase activity on
the addition of phosphates to phosphorus starved cello.
Fixation of atrio: spheric nitroeen by blue-green algae
can be an important source of oombiiivd nitrogen. The
contribution of nitrogen fixation to the nitrogon bud ; et is
often nigh in tropical lake. (Hiom e sind Viner, 1971). high
rates of activity were found in many temperate lakem;,
including,, those xn the BritiE3h 10le, 3 (Iiorlio F1nd Pore;, 1 970;
itewart, 197.2). Go3iitile and IN, 'ulonoS' (1969) hove found t1l tt

-163-
that nitrog; on fixation under optimal conditions wao
sufficient to supply the nitrogen requirerleiits of
floc-aquae. A positivE: corrt; l, -ition botwe. an nitrogen
fixation and concentration of disoolvod or , rnic nitro, 3un
compounds uns observed (Horne and : ogg, 1 97C; home 'ýt ºl_. , 1972) and a negative (h orne at al. ,1 972) or not ET: r}J1ifjcant (Horne and Fogg, 1970) correlation between nitrato
concentration and nitrogonase activity.
Many of the metabolic procesces described above
require a source. of energy which, in photoautotrophic
organisms, is supplied through photosynthesis. Photo-
synthetic activity in turn is äffected by the availability
of the compounds mentioned above and thus any aspect of
physiological activity cannot be viewed without considerat-
ion of the whole physiology cif the organism. Many
observations have been made on the reduction of the rate of
photosynthesis in the surface layers of wFator under
-conditions of intense sunlight (Baker et el., 1969;
Reynolds, 1971; Tailing, 1971). Under prolonged exposure to
high solar radiation the rates of photosynthesis fell
(Harris ans? Lott, 1973). These authors suggest that
photorespiration is the controlling mechanism which governs
photosynthetic behaviour under continuous high light
intensity.
Thus during the formation of a surface bloom many
physiological changes could occur which may only be
apparant from the resulting alteration and differentiation.
Healey and Hendzel (1976) followed the course of changes
of cell nitrogen and phosphorus, chlorophyll n, ULNA content
and the protein to carbohydrate ratio of bloomo os
Anh. flos-Ha2uae. During the early otages of bloom formation
Growth was exponential and cell contents of RNA,
chlorophyll a, phosphorus and nitrogen were high. This was
reversed prior to the collapse of a bloom. Nevertheless,
the authors conclude that increauud nitrogen Mnd pho&iphoruL
deficiency is not the cause of bloom collapse. The
physiological state of the blooming algal population, is
reminiscent of the stationary phase of batch cultures.
Lhttnt; eE observed in the j)hySio oL; ie£il activity wero" t': oro

-1 Cuj
marked than those of the concentration of dit. iso v'-d ioni in
the water.
Changes of enzyrie compliment and activity were also
observed by Laud' ina et al. (1973) during the courco of
water blooms in the Dnepir reservoirs. such changes may
reflect an adaptation to chancing conditions or a change
from one stage of the life cycle to the next. Kostlan and
Nesterova (1975) suggest that the accumulation of storage
products during the process of sporulation in the final
stages of surface blooms indicates a decrease of the total
viability (activity) of the population.
In order to obtain further information of the critical
factors governing growth and development of the planktonic
blue-green algae of the meree some aspects of the
physiological activity of algal material were monitored
during the period of bloom formation. Through this it was
hoped to determine the effects of large scale environmental
factors on the activity of blue-green algal populations,
and of the changes of physiological activity on further
development of populations.

ids.
i"iATLR1ALä AND IlliT. IUD 3
Collection Of iiriterial And Yrolirliniirj Treatment.
Bulk living sarfples collected from the üur. face waters
of the meres were allowed to stand for 2h in '2 J. wide
mouth polypropylene containers or 25U ml mea: iurina cylinderes. When the blue-green algal material had rioen to the surface it could be easily collected from the meniscus by gentle
euction. By taking up different amounts of water, any desired concentration of algal material could be obtained. The larger zooplankton and particulate material was removed before the separation of the blue-green algae by pouring the water sample through a coarse net of 1 mm square mash.
Those which were not removed tend to swim rapidly to tho
surface of the water on standing for a few minutes. Thece
were rdpidly and efficiently 'removed by dieplacement ox'
100 to 200 ml of the sample with a beaker before the blue-
green algae had started to rise. These treatments resulted
in a relatively clean suspension of the buoyant blue-green
algae. Little was observed to be non-buoyant except
akinetes, dead material and samples which were beginning to
decay. The delay between collection in the field and start
of the assay incubations was ea, maximum of 2h if the
incubations were carried out at Preston PIontford, 6h if in
Lo21d oil.
Alkaline Phosnhatase e The D-nitrophenol colourirletric technique for the
determination of alkaline phosphatase activity was as described in :: iei,; a Technical i3ullctin, .i umber 1 04 (1974).
Reagents.
The reagents used, except soditua hydroxide and
hydrochloric acid, were obtained from Uig: ria Clieraicai Co.
Ltd, Kingston upon Thames, surrey, tnL; lund.
Alkaline buffer solution (glycine, pH 10.5) U. 1i":, with
chloroform preservative. Phor3phata e iubstrate (y-nitrophonol
phosphate, d. it3odiu ).
. jiipp ied me tablets, encu-; h . t'or 2 tests. One
tablet di cool ved i,: 1.2 ml ciiatillocä water .
; ý-zi ýtropk1L: nol : stwisurcl : ýolu viý: i. (16 Ilrnol rnl-1)

-166-
Dilutions (0.5 cal diluted to 100 ml with 0.02N sodium
hydroxide) prepared as required.
Sodium hydroxide (u. 02N).
0.8 g dissolved in 11 of solution.
Hydrochloric acid (concentrated., 12N).
Procedure.
Aliquots of intact cell suspensions were incubated at
37 oC. The enzyme alkaline phospha. tase is located at the
cell wall of a marine pseudornonad (Tho-ip , on and Nacl. eod,
1974) and within-the multilayered cell wall of a Gram
negative bacterium (Costerton, 1973). Berman (1970) has
shown that the activities measured in intact and disrupted
cells of Peredinim are the same. Bone (1971) used intact
material of A. flos-aC, uae. Chloroform, used as a
preservative, had no effect on the activity of the enzyme (Berman, 1970).
0.5 ail of the buffer and 0.5 ml of the substrate were
pipetted into 15 ml I-1cCartney bottles and allowed to
equilibrate thermally. The incubation was started by the
addition of 0.2 ml algal suspension or a blank of 0.2 ml
distilled water. After 1h5 ml of 0.02N sodium hydroxide.
was added to stop the reaction.
The absorbance of test, blank and standard samples was
measured at 410 na in 10 mm silica cuvettes. To determine
the absorbance of the sample alone (the algae were removed
before measurement by filtration through 1lhatman GIS/C
filters) a drop of concentrated hydrochloric acid was added
to each sample, which renders the P-nitrophenol colourless.
Calculation.
Due to the relatively low levels of activity in field
samples the method adopted provided an eightfold increase
of measured activity as compared with the original assay
description. This was achieved by the doubling of the
incubation period (from 30 min to 1 h), a doubling of the
volume of inoculurn (from 0.1 ml to 0.2 ml) and a halving of
the volume of the sodium hydroxide solution used (from 10
ml to 5 ml). This was taken into account when Preparing
standard curves as deecribed in the original method (sigma,
1 974) . The äncrkia t- in the amotult of L-n1. ýIýo, ýherzol

-167-
libi. rated appears to be linear up to 2h (see also Herman, 1970).
An aliquot of the concentrated sarnplo was diluted by a factor of ten and the dry weight deterriired as described in Total Particulate Suspended Solids (page 53 ). Activity of
alkaline phosphatase was expressed as äigma units per mg dry weight. (One jigma unit - the liberation of 1 µmol
Z-nitrophenol per h under the stated test conditions).
Labile Orthotphosn hate.
Boiling water extractable orthophosphate clot erminations
were performed according to the method described by
Fitzgerald and . +, elson 1966) .5 to 10 mg of filtered and washed material was used. 1 mg is the minimum quantity
required for a sufficiently accurate determination using the orthophosphate analysis method described above (Page 47 ).
Each filter disc (or several if necessary) was put in
a 30 ml flat bottomed glass tube together with 1U ml
distilled water. The mouth of the tube was sealed with a glass sphere (Fig. 34). This prevented contamination and drying out of the digestion mixture. The tubes were held by
a grid in a boiling water bath filled with distilled water.
After boiling for 60 min the' samples were cooled. Any
fibres detached. from the filter dimes were removed by
filtration through 25 mm diameter GF/C discs held in a
Swinnox (Millipore, Wembley, Middlesex, England) membrane
filter holder fitted with a 20 ml syringe. The filtrate was
collected in a 25 ml volumetric flask and analer; ed for
orthophosphate as described in Uection 1 (Page 47 )
reducing additions of reagents pro rata.
Contamination of the samples was found to be minimal. The orthophosphate content of Whatman F/. 0 filters (Pyrex
glass fibre filters with no binding agents) is low oven if
unwashed. No significant increase was noted when-several blank filters were boiled in the same tube.
Hydrolysis of organic phosphorus compounds, which
would occur in acid solutions, was not apparent. Phosphate-
starved algal Liat,. rial and lake water rich in organic
phocphoruc3 compounds produced no detectable orthophor. presto
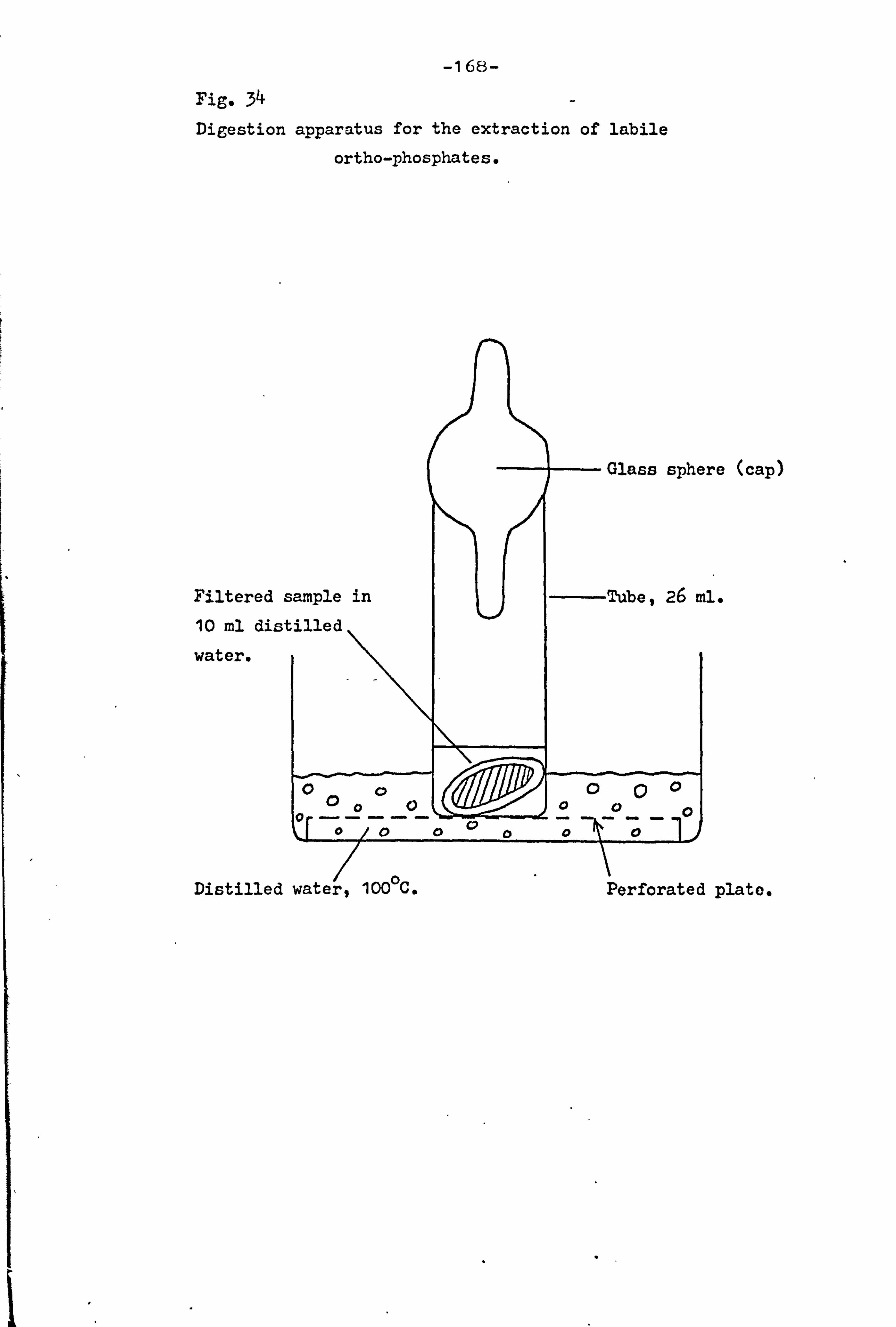
-168-
Fig. 34 Digestion apparatus for the extraction of labile
ortho-phosphates.
Glass sphere (cap)
Filtered sample in
10 ml distilled
water.
000 ýý fl o70
Distilled water, 100°C.
Tube, 26 ml.
ffl-o CP
Perforated plate.

-169-
after digestion, which t. culd be expected if those were
hydrolysed.
ýi tlt3 ar ou.. t of labile orthophosphate is givon as mg;
orthophosphate-Pilo 0: thoruti per mg dry weight of alga.
I'litrof: enase, Acetylene Reduction 2.
Nitro en fixation was estimated ufiin. lg; the acetylene
reduction technique of Stewart at RI. (1 967) and. Hardy
e". ale (1968).
3 ml of the suspension was pipetted into a 7.5 ml
serum bott_e and the incubation started bfr injecting an
amount of acetylene equivalent to about 1O of the gas
phase through the rubber septum. Prior to the addition an
equal volume of air was removed so that the incubation was
carried out under atmospheric pressure. The suspensions
were continually mixed on a rotating wheel and the incubati-
ons terminated after 30 min by partial removal of the gas
phase urizig pre-evacuated Vacutainers (Reckton and
Dickinson Co., Rutherford, New Jersey, U. S. A. ) (Schell
and Alexander, 1970, see fig. 35). After collection the
F: i. g. 35. Partial removal of the as phase from a serum
bottle using Vacutainers. (From üchell and Alexundor, 1970).
Vacutainer Double.
Yerfori
Septum
Iricuba-
E'it? MrIe
iio_'_der
Thick rubber septum
)orur bottle
Vacutainere were equilibrated to atn, o ppiieric pros.. ure. The
incubations were carried out at laboratory temperature$
. (18 to 22 °c) which did not vary by more than ± 0.5 °C
throughout the incubation period. The light izitenuity at thc: ouri'ace of the incubation vec3. sG1 was 1 QUU lux, the

-1'l0-
source being an egiza1 mixture of tungFAtin -2i). }tmentt and fluorescent lamps.
The ethylene concentration was mere
chrorlatoCraphy u. 3ina a Varian AeroL; raph
Thames, Surrey, England) with a Toropak
each sample was removed an equal volume
1.0 ml) of distilled water was injected
s: urcncl by C as 2ueO (ýia1ton=on-
Ft aolurºn. BcjJA'ore (general? y 0.5 or
to maintain the as
sample at atmospheric pressure on removal of a sample. Estimations were made of dry weight of algal material
and the data are given as nrnol ethylene produced per rng per h. Counts were made of heterocyet and akiuete frequency (as jv' of the total cell number) of intact filarnante and
also the concent2^ation of heterocystss in the incubation
suspension was established. Determination of hoterocyst
density was aided and a more randorn distribution produced
within the hae. ^ºocytometer after breaking algal clumpo by
shaking for 1 min in a serum bottle with 10 small glass beads.
Calculation: -
Total nmol produced = x. a+b
b
X=b. nmol Hample volume used
For aizalysia a= Volume of gas phase of serum bottle
b= Volume of Vdcutainer (= 5 ml)
Photoc: vnthesi a (Üx7ri en : c'roduction
Gross oxygen production was measured using the light
and dark bottle Winkler techniqu e (Vollenucider, 1974).
Blue-green algal suapenc-ions were prepared as described
above (page 165 ). The concentrate was diluted . with
filtered lake water from the site of colleotion to within the range of 5 to 20 mg 1-1 dry weight. Light and dark (covered with aluminium foil) 60 ml bottles, together with bottles for the determination of initial oxyGen concentration,
were carefully filled with weil. mixed (by Btirrin(, not
shr-eking) ou3pension and iný, ubFited under the narre conditions
as cte cribed for the nit,, ogenase aeeave (pa(-, e 169 ). Tho
zero time bottle wao fixed at the start oz the: incurton.

-1 71-
An incubation period of 2 or 3 li way to
obtain significant differences in concuntrultion.
Gross o:;, i; en production (the diift ro, ico in o: cygcrn
concentration cIiangeL3 between light and. (dark bottles) was
e: _prassed as me oxygen -produced per xi dry weight of alga
per hour. Counts were also made of absolute call nunbers
and relative abundances of the species prccont. Not
photosynthetic activity was calculated from the difference
between the oxygen concentration in the light bottle and
the initial oxygen concentration.
Carbon, Hvdr. orýen And TL. itroggen Anl lvuis.
Analysis of total carbon, hydrogen and nitrogen wore
performed by the Chemistry Depart:: ient of the school of
Pharmacy, London. The instrument used, a Carlo irba model
1104, had an absolute error of about 0.3>ö.
Assessment Of Bacterial (lumbers In Algal Samnle . Bacterial numbers in algal samples were determined by
counting the number of colonies which grew on agar plates
from a known quantity of sample (and hence a known mass of
algal material). A small volume (2.0 ml) of the sample was
first homogenized by vigourous shaking for 5 ruin in a
sterile 7 ml serum bottle containing 10 small glans beads.
Dilutions were then made into sterile diluted AUI-I 4
medium. A known quantity, generally 0.5 or 1.0 ml of
suitable dilutions (10-3 to 10-6) were thouroughJ. y mixed
with about 15 ml of sterilized agar while the agar was
still molton, but not hot, in 1etri dishes.
The bacterial agar medium used contained 20 g iincto
nutrient agar (Difco Laboratories, Michigan, U2A), 1. U g
glucose, 0.5 c sodium caseinate, 1.0 g yeast extract and 50 ml soil extract. The constituents were made up to 11
with dilute ASii 4 medium and the p1-I adjusted to 7.3 . The plates were incubated for 4 days under lt+boratory
temperatures (about 24 00).

I i, 1 ýýiJ JJyiJ
]'hoo, horus lývailýhility.
ý'he concentrations of labile orthophosplia"te Oxtrr-toted from the wa: 3-zed algal r; aterial are given in tables 31 to
33. This component of cell phoe; phor. us represents the
stored surplus, taken up through luxury consumption (Fitzgerald and Nelson, 1966). The amounts present are
consistently high in all the mer. e. s studiud throughout the
period of blooming. Little or no orthophouphate is
extracted from cultures which have been grown in the
laboratory under phosphorus-starved conditions. There is no indication of depletion of stored orthophosphate, even during periods of intense blooming and epor-ulation, such as
of A. floc-aquae in 'Kettle T-lore (Table 31). Dil. Oerences
between the Anabaena species are small and the levels found
are similar for all three mores from which material was
collected. The amount extracted from A )Ii i. fl os-ac line (Table
31) is similar to that of the Anabaena species, but the
stored surplus in. G. echinulatf3, particularly in Newton
Mere (Table 33), is generally higher.
The concentrations r3presented are similar to those
found by Fit ziTerald and Nelson (1966) for natural
planktonic populations. The amount of labile orthophosphate increases in cultures supplemented with phosphate and
becomes raarkedly- depleted under phosphorus-sstarved
conditions. The concentrations found in natural phyto-
plank ton populations are intermediate between these
extremes but are closer to the values of the phosphate-
. enriched Culture: than to that mea snxred nttor phosphorus-
starved ti atraun s.
The abundance of phosphorus ir, also reflected in the
low levels of ttlkaline phoaphatftse activity (ýOublo 34).
Again there are no significant changen during the aura:, ier in
the amour meres from which material was collected. Ociao
species appear to have slightly higher levels of activi
such as G. cliiiiuifita in Ilowton hare, F; ol. ý. tttiria rind A. ýpiroid ''iýc: Activities obtained are i imi: tr to t11okue
reported !: or cultures frown in cuurpIus phoop1u tes
}'itzgeraid and ivle lson, 1966; Borne, 1 971 ).

-173-
p. a 0
m C) OOOOOOOOOOOOOOOO 0 $1
rn C)
"o ca cC t
UN v2 0 aOO oo [- N ai CH OO0000000 C% T- N C% v- rr, \
T- " .n +) B"
aýi "aýi
r-i ý
ý3 r-1
jam' cC
H 9-4 td
0M
"EÖOfOOOOOOOOO T-
OOOO 0 (0
vý Q +'
$4 Co "r4 O to
O0p
$4 g f. -H d ) to +' 0
p 4.3 0 52 9.4 R 0 -1-4 3 ß. URoOOOOOOOOOO
t- O 000
n 00 to
cu (C $ 0 a) d
vd $$4
ci
(Q d) C (D ma0 p f' rOO8O O$
ggOOOOOoOO
0 A9 00 CO , C: A
+) 4) "r1 54 92 O it cd
"ý v
$4 OOOooOOO O\ 0\
ON000
(C O C) r-i U
i-I 60 .a8 Ee CO N CO .# .7n Q' C' N tt\ O' c- WM 8
cm Q\ 0 CO /0 CO X1\ C- . -2 N cm t+l 0
_e R T- 0 r- 0NO0 c- N v' e- 'r NO r' 0. OOOOOOOOOOOOOOOO dD
Cý ýD C) 0
\= z27 \ cý NN rn ýN\\\ Qrr ýf OGH rNNM :ý
0 H 0
U
0 to
U U
U U P4 Co 43
. r.,

-174-
gd 43
rn rZ o `- . O O O O O O O O O O O O O O 'd U b CO
L r\ c' CO C)
00 " ý
ý N
i a 3
ý 04 0
O O O O O O O O O w O O O O O
O p
- 41 V
ii
42 0
c: "1
0 x
°' 0 ) ß4
P o 0 0 0 0 0 0 0 0 . ° o 0 $y Pt N G) W .0 +) 0 M1 " cö . r; $ 4
o C q
. 11 E t i
W ý 0
.0 m b +)
00
1 t0
O O O O O rn a\
C\ Co ( g
e- . : p
ýD p CT r r
CO 4
k ä ý p
t0 Ö
" Ö fr
i $V+ O " o 0 o
N r N O O e O O O O
r1 o " o
!m
N F-
raj
S r
N" O O
0. ul K\
0 u'\ c-
0 00 e- e-
rn 0' r
0 N
N g M N "-
Cý N r
CO CO N
0 00 K1
n r
r O r
A. o 0 0 0 0 0 o O o o a o 0 0 0
C)
N 0ý0
\ N1 u1 '. D
0 = coo N ems-
- N
N N N N
..
o C Y% a% o v- . 11
om ' "r4 V to () CL) a ., 4 to U Cl +s
10
E 0 b
O A V]
" r"

-175-
Cd 41
" cý
8 8 T-
A 0 O O O O O O O O O
E- ;4 fr cö
4J (D 0 .. .n
to 3
>a H 4-4 0 O O O O O O O O O O O O
ö U 3
oo- H z 1" . a oä 4 )
" El 0 .. .N ö 4
0 ) V 'vo 4 b
° O t , g X
14 -r-4 (1) 0 41 V- O O O O O O O O O O O 0
A to 14 P, 10 CO CO "d " -P 0 a ä
o am t b "ri C) H CO tO c4 "r1 tII ( I O to % ' 0 W ° 0 0 ": t 0 0 0 0 0 0 0 0 0 0 CO () 44 O CO Si
d-) () a) " C3 $4 a a w C) w o "r4
.O (0 H 9 o o o o
O ö T- S
O S
O a 0 ON T- T- O
- O O ö t i +. U
4) F' H N " "rl t) 6 .0 0
r"-1 U T-
M E K l 0 O' 0 0 M ON ko U"\ 0 U, \ C\j N
&
H ß, O O O O O O O O O O O O O bo
u1 'D
O + ' Ä
\
sue-
\ N N
\
M lý r
t` tý lý n = = = Lam.

-176-
+ t° to Cd to es to co rq "4 ( as 0 ro (0 ý
r. 0 r. :s 0 "ý V. " r.
d C. f+ "r1 U "rl "ý U
d L7 r4 "rl V
f"'. "rl fA U U U O
Ö 0 "¢ t I 1 1 1 I d C7 Q C7
4) A s 4
0 N v 0 [. S
" 4 a I I 1 1 1 1 O O O
r0 f 1 Ui 4-) " 4
r1 q " 4-) V. " " +ý C) U "r4 CO U "
" "ý :ý "fi
v W
N U
d 1 4 Ä d .. a 4 1 1
14 1
d a Lfl% ;4 CO L%- r .. . 0 ON 150 4 . 1
d " . 8 " a 14 Ö Ö ON " t0 " 4) 0 " to v a a o 0 o I 1 1 0
cd 1) to a co or o cc Fl co $4
a n 9 - " w c n .i n) t , (ö v
N ý C -9.4 0 r4 to ) o 0 r-i . rh
C) co Cd = "ý to 4i U 4ý H 0 R
m 4a) I 1 C 1 1 1 4 4 4 I o
a co , -i 0
ý ý r -I 3
N º
1 O " 1
41 ä
1 p
a I o ö ö J 1 CO +
rl 4.3
-H 0 c a "r1 G) 4) O +J I c0 0 O O C ( ý r to to m Q ) "r -1 e 0
i
4J H O c " t 0 -i 4 " (p £'
l
.3 1 (U 4 ß 1 44
4-) 49 49
"r CC 4. N. O O O 4 ti ý' 3 3 ý o Ö CO
c ä " ºý ö ö 1 O ö . O
° ' - Ä ý ý ý
N U,
.p .0 to N. N. T- T-
N tr
b 4) tJ r4
'CJ
W d
.ý V d
a CO a,
4-4 0
ti
4-4 0 CO 4.3
CO w 0 U d
e-i p4
9 CO
v
+

-177-
Nitr. oren . letabolism.
The activity of nitrogonase of algal material
collected from the mares, as determined by the reduction of
acetylene, was measured during the summer of 1976 and the
results are given in tables 35 to 37. In Kettle More
(Table 35) activities are generally higher than thorsco
measured in the other mares. Activity falls as the
population of any species a _-os, but this reduction in
obscured by the increased activity of other species. Rates
of activity of A. circinalis appear to be comparitively 0
high andýA. fl os-aoua low, although on surface bloom
formation the influence of bacteria may be significant.
Ethylene production by soil bacteria has been reported by
Primrose and Dilworth (1976) although in anaerobic
environments. Like A. circinalia, A. snir. oiden appcre to
exhibit a high activity. During late Juno, while the
population of rah. flog-a: iixae was increasing, nitr. ogenase
activity of this species was fairly low, despite a
relatively high heterocyot frequency (Table 40). Within a
week activities had increased to near those of the Ane. bnona
species, although the heterocyet frequency had deolined.
Activities of material from white Mere (Table 36) aro
lower than those recorded for material from Kettle More.
The dominant algal species throughout the period of
measurements was A. flos-aquae, and the effects of the
increasing age of this population can be clearly soon.
Despite the increasing heterocyyst frequency (Table 40) the
total activity (per rig dry weight) and also the activity
" expressed in terms of hoterocyst numbers declines. Although
heterocysts were observed to undergo morphological changes,
no akinetes were observed (except. 3/7i 76) . Fluctuations in heterocyst frequency of algal material
collected throughout the year from Kettle Mere and Villite
Mere are given in tables 38 (data for 1974) and 39 (data
for 1975).
Although ni. trogenase activities of material collected
from Newton Mere (Table 37) are generally lowor than thorax
of Kettle: ; Iere, a different succession of species was
recorded In this mere. Algal material did not become

-178-
d cU 1f a0
" Pl .1c( 1$ 11111000OOOOOO >a 0 3°"
ö -4 to i IA
41 Aao000000 Co rn lD T- r- (- CO
ä
a) o >Z $4 N0
C) C) a r-i
0 aý 111 11 111101oOOo
a) "r w
"OOOOOOOOOO c2 Prr
to Ö" 't2 "ri d +ý 3
'. +a a) Ea
cd (O
LQ` ý- 1 ý- s- 1I11111
(D 8 r-i 0 c2
4-4 pp 1 NOroOOOOOOOOO
E)
"r4 'LS "rl O
H Pf vä c6 Co o "1 OOo110001011I1
Co 00 m (Lm "r", a. H 0" (U o IA NgOo vr4" m rn ONoOo0 r4N
0 cQ
1~ 4. ) f4 0 C) :a 4- zp
E>N 'D LD N. N. (M 0O'.
O '. O I T- T- L` 0O M "rý Lf%ö IA . ýD Mr ON ý' O"N IA N CO NN M 9:
0-OONO""""M"" Qr
N"v %D "
Co "
O" """"
rý M it
Hm
$. yN ýM
Co : r- C\ Ö (X3 Ö
CO V% N- r
CO N.
EOON r- N %Z MN r(" OM1. N
ýp M 4AN-==NNNN fV t! 1 ýO
4J .rj. r-4 W o . a3 LO a)
F3 . 14
w 0
a) +J +. 3 co to "rl as U
U0 0o
w
. ai (0 o o ti w +ý apý ä 4J 4-1
to ", co ö0
o Lý 0 b aý 4) +) ýa a)
nS RS
0
0.4 43 11-4 ;40 C) 0
U +1 co 0 ýS
"r4 U '"-4 +3 0 00 $4
.n C7
Co ö 4+ v0 ao
.ýý ono
VU ri 4N n! C) C) "P+ aa
cJ F. .: 7 (0
* ö4 3Q

-179-
d Co $. o a
oÖ ý1
1' 0 1 O 1 0 1 1 1 1 1 1 1
Co o o w 4 o aý "
x o rn o o O o O o O o 1 a p leIll
CY% (1) «4 10 93
:, r sf b 0 C ä Z to i a 0 10 to Cd to 0 4) p4 1 1 1 1 1 1 1 1 O o O 1 1 a) ö r4
ö 9.1 o m e
c o PL 4. ) lu 13 0
" r. N -t A " 0 0 0 0 0 0 0 0 M O 1 91
ý w 4) o m 0 0 o N r t t
3 q l CO 9 N 94 ý f" cd 1 >b cö R cu 0 4)
<» 54 A to o N -1 r 1.4 Ici cr - 9.9 %- cö
? 1 0 0 0 0 0 0 0 0 0 O 1 b
ß 0 4.3
9: Im to ri H
19 Ei 4-4 a)
, M 8
N 8
n 8
0 g 8 ri 3
43 A. 0 CO r Q. c- CO t- 0 to CO
t, . 1
CO k 3
43 Ü 1 Q
O r ý. {
4- in tq
c6 J ".. Cd 10 U d v 4-1 0 - 'd ti 0.. 14 O 4)) U C i4 .e
; .. i
p.
. ri 0 (Z Al
o P4 ö a: " "" 4-) Cl)
ý N o C) 5 " . .. 3 + w w 0) fi 0) * " /o 0 ` ' j, - * W, ) ° 3 > . CO O 4. o r M O O O o 0 1 . ý CO ö ä to a 0 Ü 94 . r
N
W 5 Z 43 0 Z
I -0 0 E
0) 0 * >
" 11 S ýO N
CO 0 CO -
ON CO
M N
N O c- M
M K1 `
H c0 t
4 0 10 r N [ M L cC o 4) M -0 0
O v 1 "
N "
-- "
r- " "
e- " O " O " O " O "
O N1 " 1 o
0 ý
to 9 0 ge 0) $Z
10 C) 10 to H 0 >a " p, ß +> +3
try 9, y
H 'C1 r
4- -
%0 O% 00
0 N
M f
%. 0 KI
8 4 -
e- N. -
N1 .. C. -
+1 4+ o
tom) P, - I \ N e t c 4- ON T P $4 F+ x .N bO " " . " " " " " " " " " " cti r1 G) G) U v O r M O tf1 L` r O O O O O 93 0 a s . 0 4
w w 4-5 ýi
, ri "" (. ý {. 1 (Q Is
> "ri
+1 Qi
i-N 1A
\ \ '. Q
\ '0
\ \ . -.
t\
-"
ti
"
n
A
n "
43 Q)
c ä
U m Co L. N \ \ \ \ \ " A c- e- N N N N N . -r fV M UA ?C 9 -

-180-
43 Co U 0
"y
dr
P4
Ö $4
+) O 3 O"
43 "
1 2110 s' "rI O C)
"º4 3
U Ps
, goo
av f'f
r-I Pi V- -P N .b C
vd U
4. ) "rl 0
"r4 P
U C) Cd 0
C, O ri U Cd Iz $4 4> 9ý a, 0N
4D o zý
N K Co d Cd
H to to a, IL, P, x (1)
'Ta
"rI
U
c1 Si 41 0 Co a
`Ö ä 0 0 0 0 0 0 0 0 0 0 a' .o w
rn 0 0 0 CY% 0 0 0 Co O
0
cu $4 :1 0 Pt to O O O O O O O O O O
to a, 'O r 0 0 0 0 0 0 0 0 0
CY
00 I '- r O O N O O O O O
to O t0 H 43 tH m
N tll O O Kl O O O O O
"rl 0
r N
t+1 O r N r *
O
0 " O
8 o a -
+ N
- t(\ - a'. a
t11
to 44rI
.O- co e- p. L r\ t! ý 0
U\ -Y öN NN - r+ c0 öLrö
0 r- AOOO -- OoOOO 4
n ?a"
3 -yC
u1 ON N C\j
e- r- OwN ., t "0 n1 co "N. N. C" N 4-) LO C) 0 vENOMOOO
v
ANN tC\ r
fý( M
t\ W
0
4J Cd
0 43 to
"H
0
0 4)
-j ýs 0U i ý9
t) rj ,., "ä 4'
co 4.3 0 V
"° 0
0 4)
.N o0
"rl 0 4-3 4, p 4. ) 0 -rl (D
0 4-4 0or U
tao
cz 4-) 4-31 0Ö
týA 0) V
aw "rý-j
tÖ ±"1 y 4-3 0 a) P4 Co
1t1ýý

-181- -
0 4 cd y 1 1 1 I O O
In . - O E- r-i ON 41
" 1 I 1 1 1 1 1 1
In d
r 0 "r l N K1 CO ON r4
a) .1 I M N A I 1 O O 8 X. 4-1 4) "r1
rh 0 Ea
0 O O I 1 1 1 I 1 1
4.4 " U 4
U 4-3
N r-I N C` 141 CO " .) " am
. rj 3 I t(\
N r I 1 I 1 Ci
G 4-1 0 0 -r
"rI a 0
Cd 1 1 ö M
1 1 1 I 4 + 0 4) " U 0 +) o 93 Cl) U N O $4 CD ri " r4 " C) cd O ON O' C- -4' N U U P-4 O = " " " " " " rI O
r 1 N N N N 1 O O 4-4 W 4. ) CH ý Cl) 1 0
0 (A 4) U 0 t1
-I O "r4 +> 0 (D I 1 N I I I 1 1 4.3 4-4
4-) " 0 ýC .o w
b ri
10 0 "rl H 4- N1 N I! 1 LA M N f4
co ý, y i t! ý rýl rn týl 1ý1 O ý' (1) 4J SI
" .ý ¬3 0 U O
H " K" a ) C O
. + C ) Gi
1 O M 1 1 1 I V] "r1 C) fti " ýa U : c. N
0 44 U
Li H r1 4) G) O 4) +3
r. - a) 0 (D Q) N 00 CEO 0' d x H 9
cd 0 Kl N O Z0 0 A M r ciý N N r s 1

-182-
ur% C- rn
N a) a,
a)
6 0
a) ca rl Cd 4-4 0 aý 9b c)
a) U a) P4
4-3 ra U 0 a) m
rn ON
12 H
e- Il'
cý I I I 1 N v 1 1 1 1 I 1 1 O N O
H CH T; O O
" 1 O O 1 1 1 1 M N I 1 1 1 P
c r c 0
3 1 1 1 1 K\ 1 I 1 1 1 I 1 1
"rl 0
N L` M C` r N- O\ N
1 1 O 1 O O O M M M I 1 1 1
"ý :. I 1 1 1 N 1 1 M I I 1 1 1 1
O 1 1 Ö 1 c- I 1 1 1 1 r 1
ce N M O' e- M M U1 0 UN N " " " " " " " " " " " O O ý' N N N N M M r 1 1
1 N O
H CO C-
1 O O 1 1 1 1 1 1 1 1 1 1 1 "
in ,H _e N e- 00
ý cS 3 O 1 I N N r 1 1 I 1 1 1 " O H
U O
"r1 N N U 4
1 eý " 1 1 I 1 I I 1 I 1 1 1
N H A
- H v K1 ý0 [` CO CO o0 CO 0 r N 0 . M \
' N '. D CO M ýD \ Lý tt1 \ O
A N t1' O\ N L, N c" - N N '. 0 N T- Co C/2

-183-
N. rn
U)
0
0)
E 0 ti
N Cd ea H Cd 44 0 4) bu c)
w a N
4-31 U)
U 0 d a)
0 C) r4
H
a)
N 0
Q
cý,
"r4
O
N U
ro 0
d)
cd }
4
K% 00 u1 e- 11I1I111I11ttN c- -! 1-0
.o ao UN o a. 111111111111: NN4O
1111111111111II1I
"1 11111111111I11I1
QN N 111111111111". N1
Oý Qý Oý t1ý 1111II1111I+NMNrM
OOOt! 1 O a' NM 11s+1r1f- c- rNNM -ý- r1
O co 1111I111 cý I11I11N
" b
_r 0NI! \ N Ltd -4- r .ý -t .* hO O ý- rO c- r c- rriI1IiMMMH
to 0
W LL NON
1I11I11O1"1NNr11 org
H
cti NM .ý tf1 t! ý ýO ýD vý ý 00
O
N oO
NLN M- NWrNN t\ to

-184-
4) Ö
.O r to U1 0% r' Cl- -e N e- C, "
ý "
ýp o" ' "
K\ "
N I 1 1 "
- "
N "
-- 1 T: " "
- 4) O
Cd -1
4. ) O
O' N
O `- [
ýO O
a 04 " N O O O O O O O N 1 O O
b0 X4ý
0 "r4
4)
6
H j2
Ä
2 g e-
0 e-
g c-
p 0 q . 2) 94 N C' .* .
t` CO
rr % sz M
O l`
o M
4-4 1491
o 1 0
O H 0
cö
K7 Q) to O N
4)
4)
O Rý ö
U
to C)
"i4 0
p, 4
O
1 p O
r
"
1
d
R 1
r lO
"
Cý
RS 1 U) O
r-1 .4
"
O R
1
rl 4-i
"
to r
l
P ". a U
r1 O
"
i N ei
-i
q "rl U t+ U r1
1
"
O Cd U0 O
"
p2 r1 mi
rl U
"r1 U
"
O R
C 1
IO O
r-I a
"
N M
r-i
ri U
"rl U
"
N
(0 1
(A O
S"l1
"
t0 ti
U f,
"rý U
"
4)
1 O
f1 `4'i
" vý a a s a s a ¢ 'C -s a 'c a
to 0 Ö ö ö ö 0
0 0 o 0 8 " " " "
. ah Z r- c- r r r r r ". .. .. ". ." .. ." ." of ..
93 "0
cm r*I %D
CM ON
cm Co Co
Z
0 ö ö ö ö ö ö ö C'
S. to ." ." .. .. .. ..
C' N L1- '. O e- '. D T- _e
%Z 9 %D
CY% Co ;Z C' rdl%
\o s7 cad
*lO %iö %D %0
q v oO N N Pal Pn 'lD
d O
C) r-1 Q) O x _
"rI H 4) . Ci c, cn ca se :ý z

-1c5-
abundant until m-d June and the species which bloome-d wirst,
A. circinalis, remained so until mid July, in contrast to
its early dominance and equally early disappearance in
White I"Iere and its small populations in Kettle. Nero.
Activities of A. circinalin, although showing a decreasing
trend, are more variable than that observed for A. f). os-
aquae in White ! -ere. There is no significant chunge with
the onset of sporulation. Colonies of G. echinulatn, which
were rapidly separated from the rest of the phytoplankt-on
by filtering through gauge and by purification using the
technique of differential buoyancy, also showed a declinjnc,
activity.
The low activity on nitrogenase of White Iiera algae is
also reflected in the higher-carbon to nitrogen ratios (Table -4s). An increase in the carbon to nitrogen ratio (C: 11) is also apdarant in the
. kettle Mere po.; )ulation of
A. flus-%auae which had undergone extensive sporulation in
early June. This change suggests a relative decrease in the
amount of nitrogen fixed or a relative increase in the rate
of carbon fixation as akinetes mature. There is comparitively
little change in the C: 21 ratio in Newton Mere algae, which
was characterised by the continuous dominance of a oinglb
species.
Ox r en i1ralucticn.
complexity of considerations to that which
is involved in the interpretation of the nitrodenase assays
is appNrax.;; with data on the photo-production of oxygen in
natural populations of algae incubated in the laboratory
(Tables 42 to 44). In general the patterns are E, im-EUtr.,
with hier gross photosyxithetia ret( in Kettle Niere than
in the other raeres, and a general climinution of SIctivi. t. 7 Ha
the sum . er proce, do. Both gro; 3s and net rates of oxy; can
production: fall, the net values becoming negative :t the
end of ýw:. e tie respiratory uptake exceeds photo:; , nthetic
evolution of oxy �eil.
In the Kettle ; "iere i: laterial ('fable 42) ,; rosa rat..:. of
oxygen production are low durjne, early Jurte, whc n A.
`! _. (><; -t1C' 1'! t? (tu e doC1=211i21t : -&l(*a)
is 1L71Cýi? 2^t; n]_I1ý? F3ý7C)7Ll . tit'1U21.

-186-
ca
v C)
'. O 0
94 Q)
" P,
Cý C, m0
C) H
C) zs
" c- o to
vr 1
U it 'CS
U
C äJ C
U t\;
Nv
C M
tr) E-1 O
U)
S" iJ
Z7 C)
10 ýý ca o0o8000000 LN -t "a 1t\ N rn r.
to Q) O00000O0O C0 N0OOr
pr c- -rNrr
P. N
a
11------- . :; rZ 0 r+ U ri "ri
U y
1d Sý
O ct3
oovvvvv Li u0O0NNN co "o C0 U. Cý nl c- c-- N Kl r\ 'U 4i c- FS O"
"rj All 4-5 P
p "ri to
ýi "ri O VOOOOOOONO CO IA M lý %, 0 ON c- Nr Cý CT L(\ U -t _l tV m ýo vi
cS 0 LI Sti "rl C) r-! ýý c3
000000000n C1\ OC
v
it
r-A
U
ON CG CO N Cam- co r - O Cam- 11II11., `
Cr
OI111II" U" "Ia
O C: UO c0- t^
ttl ,t III
Raj v
0
4-3 0j \U te\ 0 C7 J U'\ C; ) G> "N c- N t. c1 ONG'C. UO\
K\ 0N \O (D L\ Cý CJý hlN c- CT C7 -1I cI '11
C7
O ý` UO LC\ O 'O 1\ C; ̀
(` tf \L7 }P\ ý_) iU 9ý 111111O C`ý O 47 ?
U 'D `- N\ C) -t C rN týl cJ` J0NN
O
tý Ö" [z) NMQ ir
O O' 0
C) -t
Cam`-- 0CJ C7 U-\.? O` ýý to N4rr'. O N^\ c- b.., N c- N c- c (`1 N
fl c- CU NNI\
ä
0
II 1 i
m
ý-1
it
"
o 0 N o 2ý
aJ JJ H
" ý7 C. ý
"r1
Gý " N r7 aý
+' 4 1 I 1 "rl ý:
4J "ri ra ý
Th 1 - t i1 C U
. )
1 1 1 "i
aý C) dJ
[- l"i ý: "1 tom{
r{ W
rH FI c.
4. ) C) o
-H N U1
1 V-, c- c- r ti ,.,

-1 t37-
c0 C)
-0 J O OOOOrWN Cý r Nl pr 4
O r v. +) (6 H
R OV
L7 w U' "r' I
10 a) Q0 00000o0 u) º+ý cý rý "" 0 ,
0 A.
in C) Q\ U aý N Ö
lý cC c F I 110 I 0 U "F-> O += u ýý a"**" 0H C"% 0 0 0 0 0 0 o O C) :; P, 4-
O r` O -P Ili Q) CYN Ls
r) EO r-:. . ý
C) .. G -
-H 4-1 Fl
t-4 I; C) "^ 0 Si OOOO O O -t O Cý O O O\ O
C H
O G C7 S+ O 4. ) i7 t) (l± "0
Cj 4-)
N C) ( V1 " C- O w
! G! 1 w O to CJ t: 3 "
CO 8 'iJ " 'T,
rye r'l i * ** L^ "i= (3y ýý
o 4 i O Id
1' "r ö
R+ N 1 H <tl
O y Ll 0 to , -I . PP-1, r-l
O S-, r-l N- rf\ N ý, O CJ O CO c- '. O FOB.
V
N ÖÜ! ! 11 C7 Ü r ("IN 0
U "r+ I I II " " " " 0
Fý7 tj 0 O O O
" " .3 4.3
0
; J. Zv 1 1 I 1 1 ! P O 43
O 1 º) Gi rq (a 0 1.14 14 EA 10 y 'b 1 l`J
N (C
F= OP\ C) " CO
C co UN -r
Cc 0 C
Ul 0 '
C`_ (
O 0 L(", r- c 4'
4.1 - "rl . t N Cam- ON o . C, c-, , 10 ,
f+ - CJ M G, t C) ci
tQ ý
c U
c 1 1
f -N x 1 1 1 1 , 'N } N H
". ) 0 to 0 4) C) rf i
0 -iý c- tý U1 C: 7 1ý1 C'- N h1 L^ Lý N
CV N I'0 (V Ifs Lý ý- "ri 4-). ( ! 11 " " "
O C O O " O " r
" h1
" N\ "
C'- " hl
N 17 0 (u
tC tý
ri N
-i S
-, t: P, w
U) O O" `(1
Cl -7'
OO ^\ C"
L` ' 0
C) C\
M U-\ r1 C` \ N" " . r '. 0 ' 10 rh 4- Hc
Fat " . - r- (\l G Cý ý. ýil U. r+ r
\ 41 \` \\ f - ý> N" ý r ý c r..
' C- V " ci hý
iý t ` tip (ý , .` \ i\I i+\
\ iIý \ ; t
ý ( v (sJ C\1 (V (\! Y1
. J- ! ;y C/ M t,

-188-
U) a) p
OOOOOO J4 Iý
C9 U
" e-1 Q
of cl
H0 t, J `li
VOC. rMNr ý' N r+l Cý Q rid +) HrlIn
0 4-+ c- 0 94 rHl C
" ¢ý '3 CJ ,i
CG4: HO . F)
'" >~ fti 1 ý110 cl- C11% 110 N-U C7 O\ '. 0 Wo CT CJ'. l0ý- N- '°' CJ tUv;
ctý H L) Vý ro
Z" cu "''1
"r 1~ 1 6) O ö) UO
", 4 eH "`ý r{ c- r r- N If\ C7 '. O -. I" rfl
0" Or '= Or CU N C' oo iOV -ý ttl rl ul NNr Ca N 03 O `ý F '-D OOOO r- 0" OOOA0 . 4y
C' 10
11I1 U "zý
p 1: 14
0 :j
tti .: J 0 P, " "''ý ti
P' a b3 C P+ L` NNr lý CO r cm T!
a4""" CO Cj 10 Q
" U\ H £11tI r- 0
a)
"rl C;
44 " C .ý0 4) H 'd $: H"
v0 , -i L- CO to - ßr1 N-\ N as '. 0 ä0 N- LO Cýn. i. n N
ný H U)
va 0 ice- LO C, 04""""Q: N f4 ýi '. Cý OOr0rr c-" ON
Lv 4)
0 co
Pl U9 n ''
+5 N
11 CO rl i==. L: 'U a""""
o tt lOC c- oO N--\ n h\ ,i I-
rr c- trr c:
Cr \, D '0 \, O N- N- N C' C-
ýa N ýt rJl o ri

-189-
Net photosynthesis is only slightly loiter thz-in gross ratter.,
indicating a low rate of respiration. With the decline of
the A. x'7.0. =, -,, ciu<<e j)O )Ula. tion and an ineref. se, of (rc*_-n algae (including Volvo, ) there is an increase of j_)hotosynjt}jetic
activity. The relationship between gross and net rates
appear to be cior. e norrial for the iiiorocy, itin and.
A. circinalis populations monitored in mid Juno. Activities
were high when compared with the A. f7. c. )s-ncWae population,
but A. circinalis appears to have undergone a slight
decline in activity towards the latter }calf of June.
From late June there was a marked fa"L. I., particularly
of the net photosynthetic rate. Although there is a
downward trend of gross rates of oxygen production, the
increase in respiratory activity appears to be greater.
This leads to a large difference between ßroc; s and net
rates of oxygen production. This period coincides with a
marked' increase in temperature of the surface waters (Fig.
10), which results not only in the increase of algFil
respiration, but also enhanced bacterial Growth. Accumulation
at the water surface of algal material during this calm
hot weather, and its subsequent breakdown, would represent
the source of nutrients required for rapid bacterial
growth. Bacterial respiration is probably an important
factor in the uptake of oxygen. Increasing numbers of
bacteria were found attached to the algal material during
June Hnd July (Table 45). This may explain the low rates of
net oxygen production (i. e. the high respircation rates)
observed on some occasions during early July'.
Distinct differences in the rate of oxygen evolution
also occur with the char_r; e of domi %ance from one species to
another. A high gros, uptake was observed for alf; al
material, principally Volvox and other crec; n algae, on the
27th and 30th of June. Activities in terms of cell numbers
were also high. On the 29th of June the incubation
eam1) eJ were composed almost totally of A01. floO-ricuae,
and it was observed that production in terms of cell
nunbers was much lower than in the incuibationt: of gree-ri
algae . This is probably due to the smaller volunin uf the
cu? J_:: Of : lIpl. r1_0 -U uno. When OX 7 (. n production its

-190-
expressed in terms of amount per mg dry weight the values
are similar. she respiration rate of the green algae
appears to be higher than that of Ah. i'l os--r IF , under the
conditions applied. in contrast to Arh, fý-orj-rac Llae, A. s; siroidee appears to be as active an the green algae
described above, but a high rate of oxygen uptake reduces
net production to a negative value. Further comparisons
between different species cannot be made during early July,
due to the mixture of a number of species in more or lens
equal proportions. %
Table 45. Bacterial numbers in algal samples, 1976.
Numbers x 10-3 mg-1 (dry weicht).
Dates, "Kettle ilere White Mere Newton More 19,16 iso Alga No
" Alga No Alga,
6/6 1105 A. floc-aauao ----
22/6 -- 84 A. flor-nguan -
3/7 6281 A. sniroides 4330 " of 1622 A. circa. nrQ ici .
5/7 5085 and 938 " 10676
6/7 ---- 193501 Mixture
27671 A. circinnlio
2625 G. echinulata
A similar response to changing conditions (for exarriple
temperature, Fig. 1 1) , to that found in Kettle More, was
also observed in White Mere (Table 43). Gross rates of
oxygen production appear to be fairly high throughout the
period of observation although net production declines.
Thus although photosZ; ynthesis remains relatively constant,
respiratory activity increases, producing negative values
for net oxygen production. Like Kettle More, bacterial
numbers in White Mere increased during, this period (Table
45). Both photosynthetic and respiratory ectivity appear to
be inhibited in material collected from a dense surface
scum on the 29th of June. It is probable that this is a
reý--u_lt of d1ýrnaý; o to the 'alga_l cells rather than nutrient
deppl. etion, nines both photo; s; jnthcisis und ror31Dirotiun were
affected. '11i material WELD also diluted with filt*. rt: d lnktý

-191-
water to a concentration oinilar to that which was u.., ually
used in these incubations.
tiiLi rates for both not and gross oxyj; en production,
calculated as a function of cell numbers, wat3 observed on
the 5th of July, when a mixture of A. oriroid¬'s and
colonial green algae (Scenedesmus and was
present. It is possible that the green pleite are re, 3pono blo
for the increa e, although when next day the o::, on
production of a population predoriinaintly of green algae war
measured, it was found to be much lower. Ae vir.. tller
population of A. spiroicles did not Show a siriiiar light
photosynthetic activity. However, this was with a DaMple
obtained from a dense scum and therefore probably coffered
considerable damage. Oxygen production exprosoed as a, function of dry weight had also increased, Wt to a emallor
extent.
Gross oxygen production was loss variable during the
period of sampling In late Juno and early Ju: Ly in Newton
I1ere. (Table 44) than in Kettle Mere and White Mere. Thin
may be a reflection of the stability of the population of A. circina? _is, which remained thn dominant alga throughout
the period of blooming in this mere. Rates calculated as an expression of dry weight were similar to those estimated in
the other rzeres, but when calculated in turm: -e of cell
numbers, lower rates were sometimes obtained. However, for
species of similar size (ouch as the A. floes-a uae_ in White
Mere) similar rates were recorded. The difference in rates
shown by algae of different size is clear when compared
with samples taken from Mettle Ner. e during the dominance of A. spiroidos, a large celled alga.
Although variable, rates of net oxygen production in
Newton Teere tended to be lower and neýEitlve values were
obtained on fewer occasions, than in Kettle Mere and White
Mere. Towards the end of the sampling period an increase in
respiratory activity was observed, which is probably
correlated with the increase in numbers of bacteria (Table
45). -

ý cý ýý .
D12cussIOll Phosphorus.
The abundance of orthophosphate, which prevails
throughout the thermally stratified periods in the meros,
is reflected in the high levels of intracellular phosphorus
of natural algal populations. Intracellular phosphorus was
found to be high in the early phäees of bloom formation of
Ah. flos-aciune (Haley and HHendoel, 1976) and of Perid3, nittm
cinctum (jerruya and Berman, 1975). In contrast to the
observations made on algae from the mores, the level of
intracellular phosphorus declined at the end of growth and
prior to the collapse of blooms in both the investi. gationt;
quoted. Gerloff and : 3koog (1954) found that the cellular
levels of phosphorus and nitrogen in natural. blooms of
M. aeruginosa are comparable to laboratory cultures grown
in an excess of these nutrients. Concentrations of nitrogen
and phosphorus in the lakes from which the materiel was
collected were not, however, stated. Healey and Hendzel
(1976) found changes of cell nitrogen and phosphorus much
larger than the change of concentration of nutrients in the
water and suggested that the former are more accurate
indications of the progress of a bloom than the latter.
The low levels of alkaline phoaphataso observed in
bloom populations of algae from the meres also indicate a
sufficiency of phosphorus. An increase of alkaline
phosphatase activity (Healey, 1973; Beath and Cooke, 1975)
and enzyme synthesis (Ih_lenfeldt and Gibson, 1975) have
been demonstrated under conditions of phosphorus depletion.
However, Healey and Hiendzel (1 976) found that alkaline
phosphatase activity was variable and could not be related
to changes in phosphate uptake and cell phosphorus. bone (1971) suggests that an inverse relationship between coil
phosphorus and alkaline phosphatese activity is exhibited
only in phosphorus starved cells rather than in actively
growing phosphorus limited cells.
The indications frort the results discussed in this
section are that there is a plentiful supply of biologics iiy
available phosphorus as orthophoe -, hate and that the algal

-19: 5-
material which accumulates as a surface scum does not
become phosphorus depleted.
Nitrogen And Carbon.
The nitrogenase enzyme has been the subject of much
study, and many factors have been investigated which affect
its synthesis and activity in a range of biological
systems. Cell-free extracts of the enzyme are highly
sensitive to oxygen (Pay and Cox, 1967), and in the non- heterocystous species enzyme activity is only detected in
micro-aerophilic conditions (Rippka et aJ., 1971; Kenyon
et al., 1972). An exception appears to be Glococapas app. (Wyatt and Silvey, 1969), and apparently the marine blue-
green alga Trichodeernium, which can reduce acetylene to
ethylene in both aerobic and "anaerobio conditions ('. I ay1er
et al., 1973). It has been suggested that the colonial
nature of the alga protects the inner cello from oxygen (Carpenter and Price, 1976).
Despite the conditions of a high oxygen partial
pressure-in the surface waters of the merea during the
summer, nitrogenase activity was detected in all the
samples which contained h©terocystous blue-green algae, indicating that heterocysts provide effective protection
against oxygen inactivation of the enzyme. It is possible
that the colonial organization of some of the species, such
as G. echinalata, may give additional protection to the
central heterocysts from the inhibitory effect of oxygen. However the nitrogenase activity of G. echinulata, expressed in terns of heterocyst efficiency, was not significantly
greater than that measured for A. circinnl: in collected from
the same site at the same time.
Nevertheless the observed activities of algae from the
meres may be below the pötential activity. Stewart and Pearson (1970) have shown that nitrogen fixation and growth
of A. floc--p. 'uae and N. muscorum in laboratory cultures is
greatly increased at partial pressures of oxygen below 0.2
atmospheres. Data presented by Stewart (1971) for natural
populations of Gi oeotricM. a and Rivu2,, aria demonstrated that
nitrogen fixation can proceed oven, at elevate. }d oxygen

-194-
partial pressures. "A doubling from 2U to 40 reduces the
the naximurn activity (at 2U; ß) by 5U, ß. Inhibition of
activity by oxygen is-rapidly reversible (Stewart and Pearson, 1970). This factor may be of importance in the
markedly stratified environment of the mores, whore oxygen is greatly depleted a metre or two below the surface.
As mentioned in the Discussion to Section 1, the
soluble inorganic nitrogen to phosphorus atomic ratio in
the mores investigated was about 5: 1. Although nitrogen
will therefore become limiting before phosphorus, the
conce ntratio no 'of nitrate in the surface samples eollooted during periods of bloom formation was not significantly
reduced. Its presence would, however, be expected to have
an effect on nitrogenase activity. Horns and Fogg (1970)
" observed that although there was no direct correlation
between the rate of nitrogen fixation and nitrate
concentration, fixation was generally limited to periods
when the amount of dissolved nitrate nitrogen was below
0.3 mg l- A similar limitation of nitrogen fixation to
periods of low nitrate concentration was also reported by
Dugdale and Dugdale (1962) and Horne at n : L. (19'j2).
Additions of nitrate and ammonia were found to have an inconsistent effect on'fixation, sometimes even increasing
the rate (Goering and Neess, 1964). Although the oonoontrat- ion of nitrate in the mores occasionally exceeds 0.3 mg 1-1
it is poss. hl. e that the concentration within the algal
clump, and colonies is reduced. Since the requirement for
nitrogen is higher than that for phosphorus, depletion of nitrogen within the cell is nor© likely to occur than
depletion of phosphorus given similar external concentrations. The fixation of atmospheric nitrogen requires a supply
of energy and reductant. Energy is supplied, in the light,
through the photosynthetic production of ATP. Reductant,
in theory, may be supplied directly by photosynthetic
electron transport processes, that is, by a direct photo-
reduction of nitrogen (Fogg and Than Tun, 1953). Fiore
recent work, reviewed by Fogg It al. (1973), indicates that dark processes are the main source of roductant to
nitrogemnLc> in aerobic heterocyutoua all;.,; wherc ti. otcrocy:; ts

-195-
are the nitrogen-fixing sites.
Duggdal e and I)ugdnle (1 962) and Gooring and Neese (1964) have shown that nitrogen fixation in plankton
samples is mainly light dependent, although some fixation
may occur in the dark (Dugdale and Dugdale, 1962; Pay,
1976). The depth profile of enzyme activity in 6±indormere (the "other" Lake District) shows a similar pattern to the
primary productivity/depth curve (Horne and Fogg, 1970).
The incubations of natural populations performed during
the summer of 1976 were done at a constant light intensity
of 1000 lux. However, during the period of sampling, from
May to July, the activity recorded in blooming populations decreased. This was also matched by a fall in the rote of
photosynthesis, although gross rates were generally
maintained. Although the fluctuations in the rate of
photosynthetic oxygen production and of acetylene reduction
were considerable, a decreasing trend was observable for
each of the species of algae to form a surface bloom. Thiu
was particularly marked in Newton Mere in which A. circinuijo
was the dominant alga throughout the period of sampling, The reduction of nitrogenase activity may be the
result of the reduction of the photosynthetic rate, or both
may be the consequence of the reduction of growth and thud the increasing average ago of the algal cells in a surface bloom. Weare and Benemann (1973) observed an increasing
sensitivity of ni trogenase activity to oxygen as brrtoh
cultures of A. ctriindricn increased in age. Algal material
collected from surface scums in areas sheltered from wind
, and turbulence appears to be less active than that collected from the more exposed and thoroughly mixed areas of the
mere. This indicate: i that the high light intensity at the
surface of the water during the summer is the probable
primary cause.
This reduction of nitroeenase activity was observed despite the increasing, heterocyest frequency which generally
occur.: during the sunurer. Heterocyct frequency, considered
alune, is not an accurate indication of activity. The
frequency of heterocy: 3ts, however, in general roflc. ctn the cellular nitrogen status, (Iul_noouri. -a et n-I 197-2) . Pay

-196-
and Vasconcelos (1974) found that in cultured of j1. cylirtdri.. r
grown in the absence of vanadium and molybddnum hoterocyst
differentiation was induced although there wao no increase
of nitrogenase activity. Kale et al. (1973) obberved a reduction of heterocyst numbers in cultures of A. r1ribigun
grown without trace elements, although no measurements of
nitrogenase activity were made. Horne and Goldman (1974)
observed that trace additions of copper to natural bloom
populations produced a marked fall of nitrogenase activity
which was more marked than that of photosynthesis or
chlorophyll a levels.
Other factors may also have an' effect on hotorocy©t
differentiation and nitrogenase activity. Fogg (1949) found
that a source of readily assimilable combined nitrogen
. inhibits heterocyst formation'wjiile a source of organic
carbon increases heterocyst frequency. Although nitrate wan
present in appreciable quantities during the summer in the
meres, suppression of heterocyst differentiation by this
compound is incomplete. Ogawa and Carr (1969) found
heterocysts to be present in filaments of A. variabilin
grown in media containing 28 mg 1-1 nitrate. In cultured of N. muscorurn transferred to media containing 100 mg J, ̂ 1
nitrate, heterocyst frequency fell from 6` to 1iß after
which there was no further decline (Stewart et e., 1968).
Organic nitrogen compounds were: observed to completely inhibit the differentiation of heterocyste of Aph. floan-
aa uas (see Section 2).
A sufficiency of phosphorus is also required. Stewart
et a?. (1970) and Stewart and Alexander (1971) demonstrated
a stimulation of nitrogenase activity in phosphor. ue
starved cultures to which phosphate war-: added. The ineroa©o
of activity occurs within 15 min, although the increase in
size of the ATP pool proceeds less rapidly (Stewart and
Alexander, 1971). A further characteristic of phosphorus
deficiency of A. variabilis observed by hiecley (1973) is a
reduction of heterocyst frequency from 4; u to U. 5, o within 60 h. Readdition of phosphate produced an increase of heterocyst numbers to a normal level within a similar
period. It is considered that the supply o]' Eihot; phL, rue to

-1 97-
the u1igai cells under the conditions encountered in the
mares is cuf-'icient, and that other factors' riny be
nrimari1y rc:: ponc ble for the reduction of nitroeannse
activity'.
Heterocysts display a limited period of physio1. oCical
activity (Tischer, 1957) and appear to have a limited life (Pay, 1973). Senescent heterocysts become vacuolato and
appear devoid of cell contents. A decline of nitrogenaso
activity of U. muscorum after 4 days in batch culture was
observed by Stew, =art et al. (1968) and a reduction of heterocyst frequency was also found. However, when the
acetylene reducing efficiency of the hetoroeysts was
calculated, it was found to have fallen to a third of the
maximum value after a week in batch culture (Stewart eat al., 1968). '.
Probably the most significant factor affecting the
change of nitrogenase activity of algal material from the
Tieres is the high light intensity to which the bloom is
exposed. The formation of a surface bloom probably marks the termination of growth of the species concerned, and thus any further differentiation of physiologically active heterocysts may be reduced. The decline of net rates of
photosynthesis may also affect the rate of nitrogen fixation. The generally low-ratio of carbon to nitrogen may be a reflection of previous nitrogen fixing activity. (or a
result of photosynthesis being more sensitive to high light
intensity than nitrogen fixation), an abundr; nco of
assimilable combined nitrogen or of a diminished photo-
synthetic activity. An increase of heterocyst differantiat-
ion and nitrogenase activity in A. cylinclricF has been
shown by Uulasooriya et al. (1972) to occur only when the
cellular C: N ratio increases from 4.5: 1 to about 6: 1.
Values of about 4.5: 1 were found in material from the mares, even in heterocystous filaments. Changes in the C: I1 ratio
can be rapid (Kulasooriya et n., 1972) and thus the
heterocyc3ts, which will persist in unbroken filaments, may be a legacy of an earlier deficiency of nitrogen produced as the flan, is actively growi. nt;. .

-198-
Chant*e; s Of 1: (-. tabo3. isn And jporulation.
A consistent feature of the A-nnbaeria species oboorvod
in the meres is the association between the . )osition of
akinetes in the filament in relation to the hoter. oo, st. In
the species encountered the first akinete to be formed
differentiates from the third vegetative cell from the
heterocyst. Small variations in nunbers of undifferentiated
cells between the akinete and the heteroc; rnt were found to
be due to further cell divisions subsequent to akineto
formation. Further. differentiation of akinetes, producing a
short chain of two or three akinetes, proooede ountripetally,
away from the heterocyst.
A similar relationship between hat©rocynts and akinetas has been observed for other species of blue-green a. iiae.
The akinetes of A. cylindrica d4fferentiato from cells
adjacent to the intercalary heterocysts (dolle, 1965; 1966).
In OTlindro - permum and Glo"eotriohia also akinetee are
produced next to the terminal hetorocyst.
This relationchi) led to the suggestion, first made
by Carter (1856) that the h©terocysts exert some control
over the formation of akinetes (Fritsch, 1904; Bharadwaju,
1933; Singh, 1942b). Experimental evidence for this .
interaction was obtained by Volk (1966), who found that
heterocysts could inhibit as well as stimulate akinote
differentiation under some conditions.
Despite the apparent relationship between akinetos rind
the presence of heterocysts, some exceptions exist. in come
species, such as ! ý'odularia (Fritsch, 1951) and A. doliolttn
(Tyagi, 1974), akinete differentiation starts in the cells
equidistant between two heterocycts. burger (1974) observed
a considerable decree of variability in the position of the
akinetes in the filament in natural populations of A. als nhtonica, although 6O;, of the akineten wore found
four or less cells away from the heterocyst. äi^tultaneoue
development of all the veC, etative cells into a_tinetes,
without apptlrant influence. of heteroe;; Fata, Was observed in
A. doliolutn by 3iný; h acid Srivaotava (1968). In a: tudy of
mutant : strsins of t1u stime species $in(; li (and Dikr31hit
c011cJ. ude; that il eterC)Cyst and akineto differw: Jitiatiuit sr4ý

-199-
seperate processes regulated by different genes. Also
mutant strains of A. doliolum were produced which lacked
heterocti'sts but could sporulate (: iinha and Kumar, 19 7: %) . A natural example of akinete production in a non-
heterocystous filamentous blue-green alga is provided by
the 1. aphidopsis species (Singh, 1942a; Hill, 1972; Croribcrg,
1973).
According to an alternative concept the recent history
of the potential akinete-forming cell may be of importance.
Mitchison and Wilcox (1972) observed assymetric call
division in Anabaena catenula and the differentiation of the
smaller cells only into heterocysts. In Cylinc rosnermuui Eiji. both heterocysts and akinetes are differentiated from
recently divided sub-terminal calla (Clark and Jansen, 1969),
which indicates that the metabolism of the ochs must be in
a susceptible phase before differentiation can proceed.
Although the pattern of response of different taxe
varies, the cellular C: 11, ratio is probably an important
indicator of the factors governing akinet© as well an
heterocyst development. Fritsch (1951) suggested that
heterocysts secrete growth stimulating subotanons, the
nature of which changes in the later stages of development
which will result in the stimulation of akineto difforent-
iation. Alternatively he suggests that the reaction of the
vegetative cells to the excreted compounds may change. The
substances involved are probably nitrogen compounds. It
was suggested by Fogg O951) that heterocysts release
nitrogenous substances to adjacent cells when the nitrogen
in the rnediur. l becomes low. Large amounts of fixed nitrogon
compounds are released by growing cultures of blue-Lroon
algae (%`og , 1952. ).
An inverse relationship, that of heterocysts acting as
a sinkt for carbon compounds, was demonstrated by Uolk
(1968). From this it may be suggested that a reduction of
carbon compounds in the cells adjacent to the heturocyi-It
nay initiate alrinote development. '. folk (1965) indicates
that pho_. phorus starvation, nlthouz -li it induces kiporuý atiori,
: iW c}rily sezieý_ti:: c: the vc3�etri-tivO CU11. s to the iýýflu 'nc: c of
the hetc: rocyst; s. It is also j)os ibl. c that a reduction c): ' tliu

It
-200-
sugar phosphate levels in the vegetative cells, as a result
of phosphorus starvation and the transfer of carbon
compounds from the vej; etative cells to. the hetorocysto racy
be the stimulus to sporulation (i+olk, 1975). A concentration
gradient of the relevant compound along the algal filament,
and the requirement of a critical concentration may account
for the generally consistent position of the akinetes
within the filament. however, in thoso tpecios in which
akinetes are found adjacent to the heter. ocyst, the root of
the filament is then isolated from the influence of the
heterocyst due to the dense and intact akinete envelope. A
suitable physiological state may have already boon reached
by any further cells which undergo differentiation before
being completely cut off from the hetorocyst. This may
explain the occurrence in many species of only short chains
of akinetes. Alternatively, external factors may asour, ie
importance.
These factors may act directly on the vegetative cello,
without the intervention of the hetorooysts, or, au in the
case of phosphorus starvation, E3ensitice the cells to the
influence of further factors. It *is pouc2ble that the
factors which stimulate svvorulation operate throut; h the
mir. iicking of the interaction between the vegetative cell
which may differentiate into an akinete, and the lieterocyst.
Although extracellulur products, probably organic nitrogen
compounds, have been shown to stimulate oporulation of
C. lichenif orme (Fisher and 4aolk, 1976) nitrate was found
to be inhibitory to the sporulation of A. dolioJum (Singh
and. Srivastava, 1968; Tyyagi, 1974). Glucose was found to
stimulate sporulation and, moreover, the reducing activity
of heterocysts was reduced in sporulatin ; cultures (Tyaj,
1974). A more vigorous nitrogenase activity was observed in
non-oporulatir. C mutants of A. dolio1_1um than in the normal
wild-type or in akincte-forming mutant strains (:; inch and
Dikshit, 1976).
The factors involved are therefore very cormp>l. e. x, and
insufficient data are available to Lpoytulato anythine more ti: rýn a c; er: ral : _iec}innism of the induction of f>porn Fatjcon in
t:: c r< res .he cellular C: IJ 13ocil ratio in probably of

-201-
considerable importance in the control of differentiation
in the populations collectod from the n roc .A ratio of C: 11
of about 4.5: 1 was found in sporulating r: iater. ial of A. floc-aau e in 'Kettle ziere and i31aI e : zero and also in
aporulating A. cireinalis in I; ewton Mere. Although the nsarao
ratio was observed in material collected on 3U Juno 1976
fron Newton Piere in which no al: irietoo were found,
sporul ation was immenant and in fact akinetes war c observed in other samples collected from the same more at the same time.. In contrast the material collected from White More
during the same period had a higher ON ratio of about 5,5: 1 to 6: 1. T1o akinetes were observed in this material (A. flus-aquae) throughout Juno or July. Although
comparisons between material from different sitee, even
when of similar taxa, are not strictly legitimate, it would
appear that a greater diminution of photosynthetic activity
than nitr"ogenase activity is a prerequisite of sporulation.
To suggest which would be affected to the greater extent,
nitrogenase by the high levels of omygen present in the
surface waters or photosynthesis by the depletion of carbon
dioxide for example or by the intense solar radiation,
would require further investigation. The idea, developed in
the previous sections, that nutrient deficiency (particularly
phosphate) is not the main and direct factor governing the
sportlation of algae from the mores is supported by the
evidence discussed here.

GENERAL SUN NARY

GUNERAL 8U 12 :: CRY
From June 1974 to August 1976 a study was made of the
abundance and distribution within the water column of a
range of planktonic blue-green algal species. Special
emphasis was laid on the early growth of bloom populations
and the method of survival during adverse conditions,
particularly during the winter. The sites used for the
investigation were the meres around Ellesmere, north Salop,
England. Two, Kettle Mere and White Mere, were investigated
throughout the project, and two others, Blake Moro and
Newton Mere, were occasionally studied.
The meres are a series of water-filled kettlo holoo
and hollows in the mantle of glacial drift which covers the
region. The water supply is phreatie, and thus the mores
represent a relatively stable physical and chemical
environment, although changes in water level have been
produced in some mores as a result of drainage schemes. The
meres investigated are naturally eutrophie, and have
probably always been so. The regular occurrence of
surface blooms of blue-green algae ("the breaking of the
meres") is well established in the local folk-lore (Jackson, 1883; Phillips, 1884).
A typical succession of blue-green algae (Reynolds,
1971) was found in Kettle Mere and White More. A. ciroinalis
was generally the first to form surface blooms, followed
by A. floe-aquae. Other species, including A. aniroidos,
A. solitaries and Aph. floc-aquae bloomed later in the
summer. Microcystis generally did not become abundant
until the late summer or autumn. This pattern of succession
appears to bG consistant throughout the temperate lakes in
which blue-green algae occur. Apparent optimal temperatures
for the occurrence of planktonic algae in Lake Michigan (Stoermer and Ladewski, 1976) are similar to those observed
for the meres. Populations were generally smaller in Kettle
Mere than in White Mere. This may be the result of earlier
bloom formation due to the very sheltered position of Kettle Mere and its low surface area to depth ratio. Another factor may be the strong absorption of light by the

-204-
peat coloured water of Kettle t. ere, which may be of
critical importance in comparivons between the two mares. Kirk (1976) observed that dissolved organic compounds
greatly increased the attenuation of photosynthetically
active radiation when compared with that of other
wavelengths. Differonces in nutrient composition and
concentration were, by comparison, arall. Although
concentrated at the surface during stagnant periods, algril blooms did not become nutrient (nitrate and phosphate) depleted.
With the formation of a surface bloom during the
summer, many of the heterocystous species of blue-green
algae sporulated. This was observed to occur whether the
various species formed surface blooms in a distinct
succession, which occurred in Kettle Tiere, or when several
species bloomed simultaneously, as frequently happened in
White Mere. It has been suggested that sporulation in a
response to unfavourable nutrient conditions (Glade, 1914;
Wolk, 1965) or a result of the ageing of the culture and the concomitant increase in the concentration of extra-
cellular products (Fisher and Wolk, 1976). However, in the
meres studied, sporulation was not accompanied with any
significant change in the concentration of essential inorganic nutrients. A corroboratory observation wan made
on natural populations of G. echinu_lata, in which akineteo
were formed during active growth and in juvenile ooloinie© (Roelofs and Oglesby, 1970). It wee thorefore considered that the change of conditions encountered as the alga floats towards the surface of the water, rather than
direct nutrient deficiency, stimulates the differentiation
of akinetos. A factor of apparent iniportanee, which wan
consistent for all the instances of aporulation examined, is a high degree of insolation. This consideration is
supported by the observation that blooms of hetorocyatou©
algae which form during calm periods in the winter did
not epcrulate. However, no indication of the type of photo-inducible life cycle described by Lazaroff and Vishniac (1961,1962) for IT. muscorum was found in either natural or laboratory populations of the algal species

-205-
present in the rleres. This suggests that the factors
involved in the induction of sporulation act through
changes of physiological activity rather than controlling
directly the process of akinete differentiation.
During the period of bloom formation and sporulation,
the conditions at the water surface are optimal for
photo-oxidation (Abeliovich and $hilo, 1972) and may cause
the breakdown of surface blooms. The vegetative cells of
the filamentous species appear to be particularly sensitive
to such conditions. After the formation of akinetes the
mass of algal material descends through the water column
and settles over the surface sediments. Vegetative material
also sediments out, but little of this survives an
intensive surface bloom, particularly in Rettle Mere. The
process of sedimentation is essentially complete within a
few days following sporulation. Thus, in Kettle Mere, Fs
distinct succession of sedimenting algal material is
observed. While akinetes generally persist in the surface
sediments, vegetative material does not, particularly in
Kettle Mere, probably because of the anoxic conditions in
its hypolimnion.
In the period subsequent to sedimentation, during the
late summer and autumn, there is a marked decline in the
number of akinetee in the mud. This decline is probably not,
or scarcely due to ressuspension of sedimented material,
since the reduction in numbers sometimes preceeda the
breakdown of thermal stratification. Also very few akinetes
were found in the plankton after sedimentation, during the
winter period. It is noteworthy for comparison that
sedimented pollen grains are readily resuspended during
the autumn and winter in a lake of similar area and depth
as LJhite : ere (Davis, 1968). At this time, however, a
revival of the vegetative populations was observed. It is
suggested that marry of the akinetes germinate during late
summer and in the autumn and contribute to this population. That this immediate germination after sporulation occurs in
the natural environment is supported by the observation of large numbers of empty akinete envelopes in the plankton before sedimentation. Germination of akinetes, without a

-206-
resting period, in the original culture medium has been
observed in labcratory cultures (Fay, 1969a). The
contribution of germlings to the planktonic populations
is more important in Kettle Mere, where much less
vegetative material survives a surface bloom then in White
Mere. It is this vegetative population which ovorwiritoro in
the plankton. Extensive vernal germination of akinotea was
not observed, although numbers of akinetee in the sediments
gradually decreased throughout the winter and the following
spring. Previous investigations on the overwintering of
sporulating algae suggested that the nedimontod akinoton
remain in the mud during the winter and germinate in the
spring (Rose, 1934; Vladimirova, 1968; Wildman et n1., 1975).
The contrary observations reported here are in support of
the conclusions made by Reynolds (1972,1975a) in his
previous investigations on Crose Mere, in which he
suggested that akinete germination during the spring
probably did not contribute significantly to the population
maxima. Therefore it is considered that akinetes do not
represent a significant overwintering device but rather
serve to protect the population against the extreme
conditions prevalent during a surface bloom. Thus akinotes do act as a "buffer" (Lund, 1965) although not in the
manner which was suggested by Vladinirova (1968) to occur
in the deep Russian reservoirs, in which the ovorwintering
akinetes preserve the populations during the winter until
germination occurs under more favourable conditions. There
are indications that those filamentous heteroeystous
species which do not always produce akinetos (Aph fl, oo..
a uae. in Kettle Ziere), or those whose akinetea are prone
to attack by pathogens (A. solitaria and A. spiroiden) do
not develop such large populations end undergo greater
fluctuations in abundance from year to year than those
species which regularly produce large numbers of akinetee (A. circinalis and A. floe-aquae),
In contrast to the fragile filamentous species, the
coccoid colonial species, especially Mieroevs tin, survive
a surface bloom intact and are sedimented in an apparantly

-207-
healthy state. in a relatively non-turbulent lake the
colonies remain in the sediments during the winter, as was found in the sheltered Kettle Mere. During the summer the colonies migrate u; pwHrds through the water column to
form a surface bloom. This pattern of migration of i'icrocvatis is thus similar to that described for deeper
lakes (Sirenko et ei., 1969; Reynolds and Rogers, 1976). A low degree of turbulence appears to be critical for the
formation of clearly distinct migration patterns. Reynolds (1973a), in investigations on a relatively shallow lake (Crose Mere) found no such pattern. A similar masking of migrations by mixing was also observed in the larger of the two meres (White Mere) investigated here.
The impression gained from the field work, that
nutrient deficiency was not the stimulus for sporulation,
was confirmed by experiments performed under controlled
conditions using uni-algal isolates of the species
predonir_ant in the mores. It was found, however, that high
concentrations (above 10 rig 1-1) of orthophosphate could inhibit akinete formation. No inorganic nutrient (of those tested) acting singly, was found to consistently' induce
sporulation by its absence or presence. The converse relationship, that of cellular
differentiation being stimulated by extracellular compounds
accumulating in ageing cultures, such as was observed by
Fisher and Nolk (1976) in experiments with C. licheniforme,
was not found for any of the planktonic species grown in
the laboratory. However, some organic compounds, which
were supplied to the medium as buffering agents, were found to affect the morphology of the filament. The presence of the simple chi-peptide, N glycyl-glycine, totally inhibited
heterocyst formation. Cultures buffered with HHF. YES werd found to undergo earlier spcrulation than that observed in
control cultures. This behaviour was, however, only
observed for a single strain of Anh. floe-aquae. Otherwise HEPES did not affect the growth of the culture, except for an extension of the lag phase which was directly
proportional to the concentration of the compound in the growth medium.

-208-
$porulation in laboratory cultures of planl. tonio
isolates generally proceeds in the stationary phase of
the growth cycle. If the alga was prevented from rec+ohing
that stage, no akinetes would be produced. A delay of
sporulation may be achieved by limiting a critical
nutrient (ea. iron) required for growth, or by altering
another factor. Cultures exposed to very high or low
light intensity, or kept in the dark, grew very slowly,
if at all. In consequence no sporulation was observed
under these treatments. However, in studies on
heterotrophic cultures of Anabaenornin, Watanabe and
Yamamoto (1967) found that this alga will produce akinetes
in cultures grown in the dark with a supply of glucose.
A similar delay or absence of eporulation to that found in
cultures kept in the dark was observed when the cultures
were grown under u. v. light, or monochromatic light of a
wavelength which did not support growth. Also algae Crown
under continuous culture conditions did not sporulate. Thus
the formation of akinetes can equally proceed at the
expence of photosynthetic fixation of carbon dioxide or
through heterotrophic uptake of organic carbon compounds. In some species of algae, however, light seems to be a direct requirement for the induction of sporulation (3jazaroff and Vishriiac, 1962).
Sporulation marks a change in the physioloeic, Ll
e. ctivity of the algal population, a change from active logarithmic growth to a senescent phase in which overall
physiological activity declines.
Some support for these suggestions was obtained from
incubations of material collected from the field during
the course of surface blooms. Analysis of labile
orthophosphate and assays of alkaline phosphatase activity indicate that phosphorus does not become limiting during a
surface bloom. Nitrogenase activity and photosynthesis of
algal material incubated under controlled conditions were found to decline with increasing age of the bloom
population. In general material taken from an algal scum, previously exposed to high solar radiation, was loss
active than planktonic material which was disporaed in the

-209-
water or which occurred at the surface in shaded areaa, of
the mere. Values of the net rates of photosynthesis declined into negative figures, indicating increased rates
of oxygen uptake by respiration. Part of that respiration
may be bacterial, as the numbers of bacteria attached on
algal filaments, were found to increase during a bloom.
Blooms of a uingle species which survive for longer periods
undergo a less rapid decline of nitrogenace activity and
photosynthesis. This indicates a greater physiological
resistance of the alga to the conditions prevalent at the
surface during blooming, and thus a potential method of
assessing the state and possible future behaviour of the
bloom. In studies of blooms of Aph. fl_os-aquae, Healey
and Hendzel (1976) found changes of cellular nitrogen
and phosphorus composition comparable to those occurring
when batch cultures become nutrient deficient. Dissolved
nutrients in the weter, which were available to the natural
populations were, however, in abundant supply during a bloom (Healey and Hendzel, 1976). This abundance of
nutrients was reflected in this study, but signs of
nutrient deficiency were not observed.
Analysis of cellular carbon and nitrogen reveals that
sporulating populations have a low C: N ratio. Although not fully confirmed, it would appear that a greater reduction
of photosynthetic activity when compared to that of
nitrogen assimilation is a precondition of akinete
differentiation. Therefore, although the production of
akinetes may be indicative of a general decline of
physiological activity, the changes involved must follow
a controlled and pre-determined pattern.
The process of akinete maturation involves a
significant in rease of cell mass (growth) when conpared
with undifferentiated vegetative cells (Simon, 1977).
Sporulation thus represents a considerable investment in
terms of biosynthetic activity and is accompanied by
cellular re-organization, all of which cocur at the expense fand with the contribution of the other cells (vegetative
and hetorocysts) of the filamentous organism.

-210-
An important aspect of water quality control,
particularly with increasing outrophication and utilization
oý water resources, is the ability to predict likely,
population densities of algae under a given aeries of
conditions. Of equal importance, however, is the size of
algal populations in the period prior to. rapid growth and
the ability of these populations to undergo rapid
multiplication once conditions become favourable.
This "inoculu&" population, for the species of algae
and the environments investigated, probably bears some
relationship to the size of the surviving post-bloom
populations. The conditions at the water surface during the
summer have been shown to be unfavourable for the survival
of vegetative populations. Although there are differences
in the sines of the surviving and overwintering vegetative
populations between the two meres, the basic patterns of
akineie formation and germination, and vegetative growth,
are similar. The importance of akinete production for the
survival cf algal populations is also shown by both mores,
and there is some indication that those species which do
not produce abundant numbers of viable aknetes are
relatively limited in population development in future
years.
These findings are of critical importance in the
investigation of blue-green algal population changes in
response to eutrophication. That population levels of
future years may, in part, depend on the response of the
previous years population to adverse conditions suggests
that this approach could be used as an indicator of likely
maximal development of algal populations under postulated
conditions. However, before a quantitative relationship
can be established, it is essential that algal numbers,
their distribution and behaviour be monitored over an
extended period to test these assumptions.
Although oome factors, such as a high level of
inE: olation, were established as possible decisive factors
controlling differentiation, further work on these aspects is required to elucidate the mechanism involved. Of
particular importance is the monitoring of physiological

-21 1-
changes of algae in the period prior to blooms, and during
bloom formation and breakdown. Healey and Hendoel (1976)
found that changes of cellular phosphorus and nitrogen
content were Uretter than the changes of the same nutrients
in the water during the formation and collapse of a bloom.
They probably arise from a significant change of metabolism
during this period. A similar situation was observed in the
meres, where large changes of nitrogenase activity and
carbon fixation in the blooming algal population were
recorded against a background of a dramatically changing
physical and a relatively stable external chemical
environment. It thus appears that changes of physiological
activity are a' more accurate reflection of the state of the
algal population and more suitable indicators of its
likely future behaviour, including the induction of
sporulation.
0

REFERENCES
f

REFERENCES
Abeliovich, A. and Shilo, Pi. (1972) Photo-oxidative death
in blue-green bacteria. J. Bact. 111,682 - 689.
Allen, ii. 13. (1952) The cultivation of Iiyxophyceae. Arch.
Mikrobiol. 1,34 - 53.
and Arnon, D. I. (1955) Studies on nitrogen-fixi:: g
blue-green algae I. Growth and nitrogen fixation by
Anabaena cylindrica Lemm. Pl. Physiol.. Lancaster Q,
366 - 372.
Allen, M. Ii. and Stanier, R. Y. (1968) Selective isolation
of blue-green algae from water and soil. J. ren.
Microbiol. 51,203 - 209.
American Public Health Association (1971) Standard Methods
for the Examination of Water and Wastewater, 13 th ed.
Am. Publ. Health Assoc. Inc. New York.
Baker, A. L., Brook, A. J. and Ilemer, A. R. (1969) Some
photosynthetic characteristics of a naturally occurring
population of Oscillatoria atardhii Gomont. limnol.
Oceanogr. 14,327 - 333.
Batterton, J. C. and Van Baalen, C. (1968) Phosphorus
deficiency and phosphate uptake in the blue-Green alga Anacystie nidulans. Can. J. hiicrobiol. 1A, 341 - 348.
Bendschneider, X. and Robinson, R. J. (1952) A new opeotro-
photometric method for the determination of nitrite in
sea water. J. mar. Res. 11,87 - 96.
Berman, T. (1970) Alkaline phosphatases and phosphorus
availability in Lake Kinneret. Limnol. Oceanojr. J,
663 - 674.
Bharadwaja, Y. (1933) Contributions to our knowledge of the
lyxophyceae of India. Ann. Bot. A,, 117 - 143.
Bone, D. H. (1971) Relationship between phosphates and
alkaline phosphatase of Annbaena flos-a use in
continuous culture. Arch. Mikrobiol. 80,147 - 153.
Boulton, G. S. and Woreley, P. (1965) Late Weichselian
glaciation of the Cheshire - Shropshire basin. Nature,
Lond. '207,,
704 - 706. Boyd, C. E., Prather, E. E. and Parks, R. W. '(1975) Sudden
mortality of a massive phytoplankton bloom. Weed. Soi. 223,61 - 67.

-214-
Burt, J. S. (1961) Isolation of bacteria-free cultures- from horrnogone-producing blue-green algae. V'ature.
Lond. 192,1275 - 1276.
Burger, J. (1974) A study of two populations of Annbaenn
lnktonica Brunnth. (Cyanophyta) from Minnesota.
Phycolog; ia 11,125 - 129.
Canter, H. M. (1954) Fungal parasites of the phytoplankton
III. Studies on British Chytrids, XIII. Trans. Br.
mycol. Soc. 2Z, 111 - 133.
Carpenter, E. J. and Price, C. C. (1976) Marine Onoillr; toria (Trichodeemium): Explanation for aerobic nitrogen
fixation without heterocysts. Science. N. Y. 191.
1278 - 1280.
Carter, H. J. (1856) Notes on the freshwater infusoria of
the island of Bombay.. No. 1. Organisation. Ann. I-Inc.
nat. Hist. (Ser. II) 18,115 - 132 and 221 - 249.
Chernusova, V. ICI., Sirenko, C. A. and Arendarahuk, V. V. (1968) Localization and physiological condition of mass
species of blue-green algae during late autumn and
spring periods. In "Tsvetnie"Vody ýBloominr. " Water)
(ed. Topachevsky, A. V. ) pp 81 - 91. Izdat "Naukova"
Dumka, Kiev U. S. S. R. [English tranalationJ.
Chu, S. P. (1942) The influence of the mineral composition
of the medium on the growth of planktonic algae. Part
I. Methods and culture media. J. Ecol. 0,284 - 325.
Clark, R. L. and Jensen, T. E. (1969) Ultrastruoture of
akinete development in a blue-green alga, Cvindronpermum
ap. Cyto? oEia , 439 - 448.
Collingbourne, R. H. (1966) General principals of radiation
rneterology. In Light as an Boo? ogical Factor (ode:
Bainbridge, R., Evans, G. C. and Rackham, 0. ) pp 1- 15. .
Blackwell Scientific Publishers, Oxford and Edinburgh.
Costerton, J. W. F. (1973) Relationship of a wall
associated enzyme with specific layers of the cell wall
of a Gram-negative bacterium. J. Bact. 1-14,1281 - 1293.
Cronberg, G. (1973) Development and ecology of Rnrhidopnin
mediterranes Skuja in the Swedish Lake Trummen. Svens bot. Tidskr. Al, 59 - 64.
Daft, N. J., McCord, S. B. and Stewart, V1. D. P. (1975)

-215-
Ecological studies on algal-lysing bacteria in fresh-
waters. Freshwater Biol. a, 577 - 596.
Davis, M. B. (1968) Pollen grains in lake sediments:
redeposition caused by seasonal water circulation.
Science. N. Y. 162,796 - 799.
Demeter, C. (1956) Uber Ilodificationen bei Cyanophyoeon.
Arch. I'Iikrobiol. 24,105 - 133.
Dinsdale, N. T. and Walsby, A. E. (1972) The interrelations
of cell turgor pressure, gas-vacuolcition and buoyancy
in a blue-green alga. J. -exp. --Bot.
22,561 - 570.
Dokulil, M. (1971) Atmung und Anaerobioaerenteno von
Sußwasseralgen. Int. Revue tree. Hydrobioll.., 56,751 - 768.
Dugdale, V. A. and Dugdale, R. C. (1962) Nitrogen
metabolism in lakes: role of nitrogen fixation in
Sanctuary Lake, Pennsylvania. Limnol. Oceanojr.
170 - 177.
Eberly, V1. R. (1964) Preliminary results in the laboratory
culture of planktonic blue-green algae. Proo. Ind ans Acad. Sci. , 165 - 168.
(1966) rotes on some new and rare IIyxophyceao in
laboratory culture. Trans, Am. microsc. Boo.
130 - 133.
Eloff, J. N., Steinitz, Y. and Shilo, U. (1976) Photo-
oxidation of cyanobacteria in natural conditions. Appl,
Environ. r i. crobiol. 1,119 - 126.
Fay, P. (1969a) Metabolic activities of isolated spores of
Anabeena ovlindrica. J. exp. Bot. 20,100 - 109.
(1969b) Cell differentiation and pigment composition
in Anabaena cylindrica. Arch. ISikrobiol. 67,62 - 70.
(1973) The heterocyst. In The I3iolorv of the Bluo-
Green AlRao. (eds: Carr, N. G. and Whitton, B. A. ) Ch.
12, pp. 238 - 259. Blackwell Scientific Publications,
Oxford and Edinburgh.
(1976) Factors influencing dark nitrogen fixation in
a blue-green alga. Appl. Environ. Microbio.. J, 376 - 379.
and Cox, R. U. (1967) Oxygen inhibition of nitrogen fixation in cell-free preparations of blue-green algae.
loo

-216-
Biochem. bioph>is. Acta 142,562 - 569.
and Fogg, G. E. (1962) Studies on nitrogen fixation
by blue-green algae III. Growth and nitrogen fixation in
Chlorogloea fritschii Mitra. Arch. Mikrobiol. . 2,
310 - 321.
and Kulasooriya, S. A. (1973) A simple apparatus for
the continuous culture of photosynthetic micro-
organisms. Br. phycol J. 8,51 - 57.
, Stewart, W. D. P., Walsby, A. D. and Fogg, G. B.
(1966) Is the hoterocyst the site of nitrogen fixation
in blue-green algae? Nature, Lond. 220,810 - 812.
and Vasconcelos, L. (1974) Nitrogen metabolism and
ultrastructure in Anabaena cylindrica II. The effect of
molybdenum and vanadium. Arch. 14icr. obiol.. . 22,221 - 230.
Fisher, R. W. and Wolk, C. P. (1976) Substance stimulating
the differentiation of spores of the blue-green alga
C3, l indrospermum licheniforme. Nature. Lond. 259,
394 - 395" Fitzgerald, G. P. and Faust, S. L. (1967) Effects of water
sample preservation methods on the release of phosphorus from algae. Limnol. Oceanorr. 12,332 - 334"
and Nelson, T. C. (1966) Extractive and enzymatic
analyses for limiting or surplus phosphorus in algae. J. Phycol.
_29 32 - 37.
Fogg, G. E. (1949) Growth and heterocyst production in
Anabaena cylindrioa Lemm. II. In relation to carbon
and nitrogen metabolism. Ann. Bot.. N. S. 1J, 241 - 259-
(1951) Growth and heterocyst production in Antbaena
cylindrica Lenzen. III. The cytology of heteroeysts. Ann.
Bot., N. Ü. 11i 23 - 35.
(1952) The production of extracoellular nitrogenous
substances by a blue-green alga. Proo. R. Boo. B 139,
372 - 397. (1965) Algal Cultures and Phytopiankton Eeolorw.
University of Wisconcin Press, Madison and Milwaukee,
U. S. A.
(1969) The physiology of an algal nuisance. Proc. R. Soc. B jM, 175 - 189.
(1971) I'litrogen fixation in lakes. In Biolorioa1_

-217-
Nitroren Fixation in Natural and Af; riculteral Habitats
(eds: Lie, T. A. and Mulder, E. G. ) pp. 393 - 401. ELL
soil, Special volume.
and Stewart, N. D. P. (1965) Nitrogen fixation in
blue-green algae. Sci. Progr. ri, 191 - 201.
Pay, P. and 1? alsby, A. H. (1973) The Blue-
Green Algae. Academic Press, London and New York.
and Than Tun (1958) Photochemical reduction of
elementary nitrogen in the blue-green alga Anabeena
c. ylindrica. Biochem. _biophys.
Acta Q, 209 - 210.
and (1960) Interrelations of photosynthesis and
assimilation of elementary nitrogen in a blue-green
alga. Proc. R. Soc. B, 111 - 127.
Foy, R. H., Gibson, C. B. and Smith, R. V. (1976) The
influence of daylength, light intensity and temperature
on the growth rate of planktonic blue-Green algae. Br.
phycol J. 11,151 - 163.
Fritsch, F. E. (1904) Studies on Cyanophyceae III. Sore
points in the reproduction of Anabaena. New Phytol.
216 - 228. (1945) The Structure and Reproduction of the A1P, te
Vol. 2. University Press, Cambridge.
(1951) The heterocyst: a botanical enigma. Proc. Linn.
Soc. Lond. 162,194 - 211.
Fuhs, G. W., Demrierle, S. D. 9 Canelli, B. and Chen, 11. (1972) Characterization of phosphorus-limited plankton
algae (with reflections on the limiting-nutrient
concept). In Nutrients and Butrophieatinn (ed:
Likens, G. 'E. ) Am. Soc. Limnol. Oceanogr. Special
symposium 1. pp. 113 - 133.
Geitler, L. (1925) Cyanophyceae. Vol. 12 of Die Süsewasser-
flora Deutchlandn, 6sterreiche und der Schweiz (ad:
Pascher, A. ). Gustav Fischer, Jena.
Gentile, J. H. and Maloney, T. E. (1969) Toxicity and
environmental requirements of a strain of Anhanizomenon
floe-aquae (L. ) Ralfe. Can. J. Microbiol. j, 165 - 173. Gerloff, G. C., Fitzgerald, G. P. and Skoog, F. (1950) The
isolation, purification and culture of blue-croon algae. Am. J. Bot. 27,216 - 218.

-218-
and , _�_, (1952) The mineral nutrition of
r1icrocystis aeruFinosa. Am. J. Dot. ; LQ, 26 - 32.
and Skoog, F. (1954) Cell contents of nitrogen and
phosphorus as a measure of their availability for the
growth of Microcystis aeruginosa. Ecolofv ,,
348 - 353.
and (1957) Nitrogen as a limiting factor for
the growth of Micr. ocystis aeruginosa in Southern
Wisconcin lakes. Ecology M, 556 - 561.
Glade, R. (1914) Zur kenntnis der Gattung Clindrospermum.
Beitr. Biol. Pfl. 12,295 - 344.
Goering, J. J. and Neess, J. C. (1964) Nitrogen fixation
in two Wisconcin lakes. Limnol. Ocennogr. y, 535 - 539.
Goldman, C. R. (1965) Micronutrient limiting factors and
their detection in natural phytoplankton populations.
Mern. Ist. Ital. Idrobiol. 18 (Suppl. ), 121 - 135.
Goldman, J. C. (1976) Identification of nitrogen as a
growth limiting nutrient in wastewaters and coastal
marine waters through continuous culture algal assays.
Wat. Res. 10,97'- 104.
Golterman, H. L. (1971) Methods for Chemical Analysis of
Fresh Waters. International Biological Programme
Handbook No. 8. Blackwell Scientific Publications,
Oxford, London and Edinburgh.
Good, N. E. and Izawa, S. (1972) Hydrogen ion buffers. In
Methods in Enzymology Vol. 24 B (ed: San Pietro, A. )
PP- 53 - 68. Academic Press, New York and London.
, Winget, G. D., Winter, W., Connolly, T. N., Izawa, S.
and Singh, R. N. M. (1966) Hydrogen ion buffers for
biological research. Biochem. N. Y. !,, 467 - 477.
Gorham, E. (1957) The chemical composition of some waters
from lowland lakes in Shropshire, England. Tel us S,
174 - 179.
Gorham, P. R., McLachlan, J. S., Hammer, U. T. and Kim, W. K. (1964) Isolation and culture of toxic strains of Anrtbaena flos-aquae (Lyngb. ) de Breb. Veh_int. Verein. theor. anaew Limno . ,
1-ý, 796 - 004.
Griffiths, B. M. (1925) Studies in the phytoplankton of the lowland waters of Great Britain ICI. The phytoplankton
of Shropshire, Cheshire and Staffordshire. J. Linn.

-219-
. Soc. (IZot. ) 4,75 - 98 (1938) Early references to waterbloouis in British
lakes. Proc. Linn. Soc. fond. 151,12 - 19.
Grimshaw, H. 14. and Hudson, M. J. (1970) Some mineral
nutrient studies of a lowland more in Chenhiro,
England. FIydrobioloi 36,329 - 341.
Hammer, U. T. (1964) The succea:; ion of "bloom" species of
blue-green algae and some causal factors. Verh. int.
Verein. theor. sn, ew. Limnol. 829 - 836.
(1969) Blue-green algal blooms in Saskatchewan lakes.
Verh. int. Verein. theor. anRew. Limno_. j, 116 - 125.
Harder, R. (1917a) Uber die Beziehung der Keimung von
Cyanophyceensporen zum Licht. Ber. dt. bot. Gen. 2.5
58 - 64. (1917b) Uber die Bezeihung de© Lichtes our Keimung
von Cyanophyceensporen. Jb. Wise. Bot. 58,237 - 294.
Hardy, E. F. (1939) Studies on the poet glacial history of
the British vegetation V. The Shropshire and Flint
t1aelor mouses. New Phytol., 364 - 396.
Hardy, R. W. F., Holeten, R. D., Jackson, B. K. and
Burns, It. C. (1969) The acetylene-ethylene assay for
112 fixation: laboratory and field evaluation.
Physiol., Lancaster . 42,1 185 - 1207.
Harris, G. P. and Lott, J. N. A. (1973) Lieht intensity and
photosynthetic rates in phytoplankton. J. Fish. Res.
Bd Can. 0,1771 - 1778.
Hart, I. C. (1967) Nomograms to calculate dissolved-oxygen
contents and exchange (mass-transfer) ooeffioiontß. Wat. Res. 1,391 - 395.
Haynes, R. C. (1975) Some eoologioal effects of artificial
circulation on a small eutrophic lake with particular
emphasis on phytoplankton II. Kezar Lake experiment,
1969. H , drobiolo, iia -4-6,141 - 170.
Healey, P. P. (1973) Characteristics of pho©phorue
deficiency in Anabaena. J. Phvcol. , U, 383 - 394.
and Hendzel, L. L. (1976) Phyeioloßical changes during the course of blooms of A, Lhanizornenon floe-aquae.
J. Fish. RAs. Bd Can. 22,36 - 41.
Heaney, Be 1. (1973) Problems related to the growth of

-220-
blue-green algal populations. Ph. L. Thesis, New
University, Ulster, Coleraine.
Heath, R. T. and Cooke, G. D. (1975) The aignifieanoe of
alkaline phosphataee in a eutrophic lake. Verh, int.
Verein. theor. angew. Lirmnol. 19,959 - 965.
Herbes, S. E., Allen, H. E. and Mancy, X. H. (1975)
Enzymatic characterization of soluble organic phosphorus
in lake water. Science, N. Y. 18' , 432 - 434.
Heron, J. (1962) Determination of phosphate in water after
storage in polyethylene. Limnol. Ooeanoj3r. 3,, 316 - 321.
Hill, H. (1972) A new Rrnphidonsis species (Cyanophyta,
Rivulariaceae) from Ninnesota lakes. Phyaoloaia j1,
213 - 215.
Hobro, R. and YJillen, E. (1975) Phytoplankton counting and
volume calculations for the Baltic -a Method
comparison. Vatten j, 317 - 326.
Holm-Hansen, O., 'Gerloff, G. C. and Skoog, F. (1954) Cobalt
as an essential element for blue-green algae.
Physiologia Pl. 3,665 - 675.
Horne, A. J., Dillard, J. E., Fujita, D. X. and Goldman, C. R.
(1972) Nitrogen fixation in Clear Lake, California II.
Synoptic studies on the autumn Anabaenn bloom. Limno . Oceanogr.
_ 11,693 - 703.
and Fogg, G. B. (1970) Nitrogen fixation in some English lakes. Proo. R. Soc. B J, 351 - 366.
and Goldman, C. R. (1974) Suppression of nitrogen
fixation by blue-green algae in a eutrophic lake with
trace additions of copper. Science. N. Y. 1,409 - 410.
and Viner, A. B. (1971) Nitrogen fixation and its
significance in tropical Lake George, Uganda. Nature.
Lond_ 22,417 - 418.
Huntsman, S. A. and Barber, R. T. (1975) Modification of
phytoplankton growth by excreted compounds in low
_ density populations. J. Phvcol. 11,10 - 13.
Ihlenfeldt, M. J. A. and Gibson, J. (1975) Phosphate
utilisation and alkaline phosphataoe activity in
Anacystis nidulans (Synechocoocus). Arch. I"iicrobiol.
102,23 - 28.

-221-
Iverson, R. L. , Curl, H. C. , O' Cozinoro, 11. B. , , Kirk, D. - and
Zakar, K. (1974) Summer ph. 'toplankton blooms in Auke
Bay, Alaska, driven by wind mixing of the water column.
Lirno'_. Oceanopr. _
12,271 - 278.
Jackson, G. F. (1883) Shron5hire Folk-Lore (ed: Burne, C. S.,
from the collection of G. F. Jackson). Trubner, London.
Kale, S. R., Balial, M. and Talpai3ayi, E. R. U. (1970)
Heterocyst differentiation in Anabaena nnbirua Rao I.
Growth of the alga. Indian Biologist 2,30 - 40.
and -
(1973) Heterocyst development in
. Anabaena ambigua Rao IV. Effects of some mineral salts.
Phycos 12,11 - 17.
Kaushik, M. and Kumar, H. D. (1970) The effect of licht on
growth and development of two nitrogen-fixing blue-
green algae. Arch. Mikrobiol. , 52 - 57.
Kawana, K. (1975) Spectral distribution of underwater
irradiance. Bull. Fac. Fish. Hokkaido Univ. 26,
235 - 248.
Keating, K. I. (1975) Algal allelopathic involvement in
bloom sequence in freshwater. J. Phycol. 11, (Suppl. )
12 - 13. (Abotr. ).
Kenyon, C. IJ., Rippka, R. and Stanier, H. Y. (1972) Fatty n
acid composition and physiolog cal proportiea of some
filamentous blue-green algae. Arch. Mikr. ohio P2,
216 - 236.
Ketchum, B. H. (1939) The ievelopment and restoration of
deficiencies in the phosphorus and nitrogen composition
of unicellular plants. J. cell. oomp. Phvºsiol. 1,
373 - 381. Khan, K. R. (1974) Overwintering mechanisma in Gloeotrichirt
echinulata (J. M. Smith) Richt. J. Phycol. 10, (Surpl. )
4 only. (Abstr. ).
King, D. L. (1970) The role of carbon in eutrophication.
1 J. Wat. Pollut. Control Fed. 42,2035 - 2051.
Kirchner, W. B. (1975) An evaluation of sediment trap
methodology. Limnol. Oceano, r. 20,657 - 660.
Kirk, J. T. 0. (1976) Yellow substance (Gelbstoff) and its
contribution to the attenuation of photosynthetically active radiation in some inland and coastal south-

-222-
eastern Australian waters. Aust. J. mar. Freohwat. Res.
21,61 - 71.
Klebahn, H. (1895) Gasvakuolen, ein Be©tandteil der Zellen
der wasserblütenbildenden Phycochromaoeen. Flore. Jena
80,241 - 282.
I: noechel, I. and Kalff, J. (1975) Algal sedimentation: the
cause of a diatom - blue-green succession. Verh. int.
Verein. theor. angew. Limnol. 12-, 745 - 754.
Komarek, J. (1958) Die taxonomische Revision der Plankton-
ischen Blaualgen der Techechlowakei. In Algoliecho
Studien (Kou. irek, J. and Ettl, H. ) pp. 10 - 286. Czech
Academy of Sciences, Prague.
Kostlan, N. V. and Necterova, D. U. (1975) Change in
content of phosphorus compounds in Aphinizomenon floo-
ac uaG (L. ) Ralfs during vegetation. Ukr. Bot. Zh. ý2,
355 - 358- [In Ru©uifui, Lngliuh abotrcuot,.
Kuentzel, L. E. (1969) Bacteria, carbon dioxide and algal
blooms. J. Wat. Pollut. Control Fed. 'Al, 1737 - 1747.
Kulasooriya, S. A., Lang, N. J. and-Pay, P. (1972) The
heterocysts of blue-green algae Ill. Differentiation
and nitrogenase activity. Proc. R. Boo. B 181,199 - 209.
Lange, W. (1976) Speculations on a possable essential
function of the gelatinous sheath of blue-green algae.
Can. J. Mi_crobio_l_ 22,1181 - 1185.
Lazaroff, N. (1966) Photoinduotion and photoreversal of the
Nostooacean developmental cycle. J. Phyool. ,? � 7- 17.
and Schiff, J. (1962) Action spectrum for developmental
photo-induction of the blue-green alga Nostoo muncorum.
Science, N. Y. IM, 603 - 604.
and Vishniae, W. (1961) The effect of light on the
developmental cycle of Ilostoc muscortxm, a filamentous
blue-green alga. J. non. Nicrobiol. 25,365 - 374.
and (1962) The participation of filament
anastomosis in the developmental cycle of Nontoe
mus, a blue-green alga. J. non. TSiorobiol. 20,
203 - 210.
Lemmermann, E. (1900) Beiträge zur Kenntnis der Planktonalgen VII. Des Yhytoplankton den Zwiohenahner

-223-
I""Ieeres. Der. dt. bot. Ges. 18,135 - 143.
Lin, C. K. (1972) Phytoplankton succestiion in a eutrophic
lake with special reference to blue-green algal blooms.
Hydrobiologin 29,321 - 334.
Lind, H. M. (1944) The phytoplankton of some Cheshire mares.
Diem. Proc. Pianchr lit. phil. Soc. 86,83 - 105-
Lorenzen, M. and Mitchell, R. (1973) Theoretical offoote of
artificial destratification on algal production in
impoundments. Envir. Sei. Technol. Z, 939 - 944.
Lund, J. W. G. (1965) The ecology of the freshwater phyto-
plankton. Biol. Rev. 4,0,231 - 293. (1966) The importance of turbulence in the periodicity
of certain freshwater species of the genus Me os i_}r . Bot. Zhurn.
. 51 ,1 76 - 187. [In Rizssian] .,
, Kipling, C. and Le Cron, E. D. (1958) The inverted
microscope method of estimating algal numbers and the
statistical basis of estinations by counting.
Iiydrobiologia 11,143 - 170.
Lyon, A. J. (1970) Dealing with Data. Pergamon, Oxford.
McLachlan, J. and Gorham, P. R. (1961) Growth of Miorocyntip
aerum inosa I: utz. in a precipitate-free medium buffered
with TRIS. Can. J. Microbiol. . 7.869 - 882.
McNabb, C. D. (1960) Enumeration of freshwater plankton
concentrated on the membrane filter. Limnol.. Oonanor! r, E, 57 - 61.
Meffert, N. B. (1971) Cultivation and growth of two
planktonic Osci1_latorin species. Mitt, int. Vc, rein,
theor. angew. Limnol. 1, j, 189 - 205.
Diitchison, G. J. and Wilcox, M. (1972) Rule governing cell
division in Anabaena. iTature, Lond. ga, 110 - 111.
Mortimer, C. H. (1942) The exchange of din solved substanooo between mud and water in lakes III and IV. Summary and
references. J. Ecol. 0,147 - 201.
Morton, S. D. and Lee, T. H. (1974) Algal blooms - possible effects of iron. Envir. Sei. Technol. A, 673 674"
Moss, B. (1969) Vertical heterogeneity in the water column of Abbots Pond II. The influence of physical and chemical conditions on the spatial and temporal distribution of the phytop? ankton and of a coiamitnity of
4

-224-
epipelic algae. J. Ecol. . jZ, 397 - 414.
Ogawa, R. L. and Carr, J. F. (1969) The influence of
nitrogen on heterocyst production in blue-green algae. L mnol. 0ceanoir. 14,342 - 351.
Pearsall, U. 11. (1923) The phytoplankton of Rostherne More.
lem. Proc. Nanchr lit. Phil. Soc. U, 45 - 55.
Pennington, W. (1974) Seston and sediment formation in five
Lake District lakes. J. Boni. 62,215 - 251.
Phillips, W. (1384) The breaking of the Shropshire Lorca.
Trans. Shrops. Ar. chaeol. nat. Hießt. Soc. Z, 277 - 300. (1893) The breaking of the Shropshire Nerea. Mid j.
Nat. 16,1 - 15.
Primrose, S. B. and Dilworth, N. J. (1976) Ethylene
production by bacteria. J. Pen. T'linrohiol. , 177 -
182.
Reddy, P. N., Rao, P. S. N. and Talpasayi, E. R. 2. (1975)
Effect of red and far-red illuminations on the
germination of spores of two blue-croon algae. Cures
Sci. 44,678 - 679.
Reichardt, U. (1971) Catalytic mobilisation of phosphate in lake water and by Cyanophyta. Hydrobiolof; in 377 - 394.
Reif, C. B. and Fine, D. L. (1967) Two winter blue-green
algal populations. Ph'colotlia 6,105 -'106. Reynolds, C. S. (1971) The ecology of the planktonic blue-
green algae in the North Shrop; hire mores, Lngland. Fld
Stud. 2,409 - 432. (1972) Growth, gas vacuolation and buoyancy in a
natural population of a planktonic blue-croon alga. Freshwat. Biol. 2, E37 - 106.
(1973a) Growth and buoyancy of Ilicrocyatia aorupi. noaa Kütz. emend Elenkin in a shallow eutrophic lake. 'roc,
R. Soc. B 18A, 29 - 50. (1973b) Ph.,, ytoplankton periodicity of some ;: orth
Shropshire meres. Br. phN�ycol J. 8,301 - 32U. (1973o) The phytoplankton of Croce More, Shropshire.
Br. phycol J. - 8,153 - 162. (1975a) Interrelations of photosynthetic behaviour
and buoyancy regulation in a natural population of a

-225- blue-green alga. Freshwat. Biol. ý, 323 - 338.
(1975b) Temperature and nutrient concentration in the
characterization of the water supply to a small
kataglacial lake basin. Freshwat. Biol. ,.,
339 - 356"
and Rogers, D. A. (1976) Seasonal variations in the
vertical distribution and buoyancy of Microcyptin
aeruginosa K1tz. emend. Elenkin in Rostherne Mere,
'E'ngland. Hydrobiologin A. 8-, 17 - 23.
Rhee, G-Y. (1972) Competition between an alga and an
aquatic bacterium for phosphate. JLimnol. Ooenno, r. , l,
505 - 514.
Rippka, R., Nielson, A., Kunisawa, R. and Cohen-Bazire, G.
(1971) Nitrogen fixation by unicellular blue-green
algae. Arch. Mikrobiol_ 2-6,341 - 348.
Robinson, B. L. and Miller, J. H. (1970) Photomorphogenesia
in the blue-green alga Nostoo commune 584. Physio1oRin,
P1., 461 - 472.
Roelofs, T. D. and Oglesby, R. T. (1970) Ecological
observations on the planktonic cyanophyto Gloeotrichi3
echinulata. Limnol. Oceanofr_ j5,224 - 229.
Rose, E. T. (1934) Notes on the life history of A�3hanizomonon
flos-aquae. Univ. ' Iowa Utud. Nat. Hist. 16,129 - 141.
Schell, P. M. and Alexander, V. (1970) Improved incubation
and gas sampling techniques for nitrogen fixation
studies. Limnol. Oceanoir. , 961 -'962.
Schindler, D. W., Brunskill, G. J., Emerson, 2.,
Broecker, W. S. and Pang, T. -H. (1972) Atmospheric
carbon dioxide; its role in maintaining phytoplankton
standing crops. Science. N. Y. U, 1192 - 1194.
Schwabe, G. H. and Strange-Bursche, E. (1964) Ubor
A hanizomenon gracile Lemm. (Blaualgen und Lebensraum,
IX). Arch. Hydrobiol. 60,249 - 277.
Serruya, C. and Berman, T. (1975) Phosphorus, nitrogen and the growth of algae in Lake Kinneret. J. Phycol. 11,
155 - 162. Shapiro, J. (1973) Blue-green algae: why they become
dominant. Science. N. Y. 1 9,382 - 384.
and Glass, G. L. (1975) Synergistio effects of phosphate and manganese on growth of Lake Superior

-226-
al, gae. Verh. int. Verein. theor, anj ew. Limnol. jj, 395 - 404.
Shilo, M. (1975) Factors involved in dynamics of algal blooms in nature. In Unifvinn. ConeexAn of Eoolorv,
Proc. First Internat. Conf. Lcol. pp. 127 - 132. The
Hague.
Shubert, L. E. (1974) Utility of dilute algal media. J.
Phvco_l. 10, (Suppl. ) p21 only (Abstr. ).
and Trainor, F. R. (1974) Scenedesmue morphogenesis:
control of the unicell stage with phosphorus. Bre
phyycol J. 2,1 - 7.
Sicko-Goad, L. 111., Crang, R. E. and Jensen, T. E. (1975)
Phosphate metabolism in blue-green algae IV. In situ
analysis of polyphosphate bodies by x-ray energy
dispersive analysis. Cytobiol. 11,430 - 437.
and Jensen, T. E. (1974) X-ray energy dispersive
analysis of polyphosphate bodies in P. ec; tonerarz borg*anur.
In 32nd Ann. Proe. rleotr. MicroE3oop. Soo. Am©r,. (ad:
Arceneaux, C. J. ). St Louis, 11issouri.
_ and (1976) Phosphate metabolism in blue-green
algae II. Change in phosphate distribution during
starvation and the "polyphosphate overplue" phenomenon in Plectonema bor anum. Am. J. Bot. , 183 - 188.
Sigma. (1974) Phosphatase: acid, alkaline and proatatio. Sigma Tech. Bull. No. 104.
Simon, R. D. (1977) Sporulation in the filamentoun
cyanobacterium Anabaena oylindricn: the course of spore formation. Arch. Miorobiol. 111,283 - 288.
Singh, H. 11. (1967) Genetic control of aporuletion in the
blue-green alga Anabaena doliolum Bharadwaja. Plantr,
ZU, 33 - 38.
and Srivastava, B. 3. (1968) Studies on morphogenesi© in a blue-green alga 1, Effect of inorganic nitrogen sources on developmental morphology of Anabaen doliolum. Can. J. Microbiol. j, 1341 - 1346.
and Sunita, K. 1.1. (1974) A biochemical study of spore germination in the blue-green alga Anebanna doliolum. J. exp. Bot. 837 - 845.
Singh, R. N. (1942a) A new species of the genus Ranhidop sie

-227-
Fritsch and Rich (Raphidonsis indica sp. nov. )
exhibiting morphological variants. J. Indian bot. Soo.
21,239 - 243.
(1942b) Wollea bharadwa s. ep. nov., and its autecology.
Ann. Bot. N. S. 6,593 - 606.
and Dikshit, R. F. (1976). Mutaeenesia of the blue-
green alga, Anabaena doliolur Bharadwaja. Mutation it ea. 65 - 78.
Singh, H. N. and Sinha, H. (1967) Genetic control of
cellular differentiation in blue-green algae. Bull.
Nat. Inst. Sei., India No. 34,131 - 145. (Pron. Symp.
Newer Trends in Taxonomy, New Delhi).
Sinha, B. D. and Kumar, H. D. (1973) A mutational study of
the nitrogen-fixing blue-green alga Anabanna doliolum.
Ann. Bot. 2Z, 673 - 679.
Sinker, C. A. (1962) The North Shropshire mores and mosses:
a background for ecologists. Fld Stud. 1,101 - 138.
Sirenko, L. A., Chernousova, V. M., Arendarchuk, V. V. and
Kozitskaya, V. N. (1969) Factors of mass development of
blue-green algae. Hydrobiol J. ! j, 1-8.
Smith, R. V. (1973) Evidence for the obligate dependanoe of
a Lough Neagh strain of Oscillatorin redekii Van Goor
on the presence of bacteria for growth. Symp.
Prokaryotic Photoeynthetio Or tniems, Freiburg.
pp. 89 - 90 (Abstr. ).
and Foy, R. H. (1974) Improved hydrogen ion buffering
of media for the culture of fresh-water algae. Br.
phycol J. , 2,239 - 245.
Spratt, E. R. (1911) Some observations on the life history
of Anabaena cycadaceae. Ann. Bot. 25,369 - 380.
Stewart, W. D. P. (1971) Physiological studies on nitrogen-
fixing blue-green algae. In Biolorioel Nitro, on Fixation
in Natural and Aitriculteral Habitat rý (ede: Lie, T. A.
and I": ulder, B. G. ) pp. 377 - 391. Pl. Soil, special
volume. (1972) Algal metabolism and water pollution in the
Tay Region. Proc. R. Soo. Bdinb. 71,209 - 224. and Alexander, G. (1971) Phosphorus availability and
nitrogenase activity in aquatic blue-greon algae.

-228-
Freshwater Biol. 1,389 - 4U4.
, Fitzgerald, G. P. and Burris, R. H. (1967) In situ
studies on N2 fixation using the acetylene reduction technique. Proc. natn. Acad. Sci. U. S. A. 58,2071 - 2078.
md (1968) Acetylene reduction by
nitrogen fi:: ing blue-green algae. Arch. Tlikrobiol. 629
336 - 348.
and (1970) Acetylene reduction assay for determining phosphorus availability in 'Iisconcin
lakes. Proc. natn. Acad. Sci., U. S. A. 66,1104 - 1111.
and Pearson, H. W. (1970) Effects of aerobic and
anaerobic conditions on growth and metabolism on blue-
green algae. Yroc. R. Soc. B 1l, 293 - 311.
Stoermer, E. F. and Ladewski, T. B. (1976) Apparent optimal
temperatures for the occurrence of some common
phytoplankton species in southern Lake Michigan. Great
Lakes Res. Div., Publication 18. Univ. of Michigan,
Ann Arbor.
Sud'ina, O. G., 5hnyukova, Ye. 1., Kostlan, N. V.,
Mushak, Y. 0. and Tupyk, 14. D. (1973) Biochemical
features of blue-green algae which cause water-bloom. Ukr. Bot. Zh. 30,497 - 504. [Exiglish trunalation].
Sverdrup, H. U. (1953) On conditions for the vernal blooming of phytoplankton. J. Cons. perm. int. Exnlor.
Iter. 18,287 - 295.
Telling, J. F. (1971) The underwater light climate no a
controlling factor in the production ecology of fresh-
water phytoplankton. I'iitt. int Verein. theor. anaew. Limnol. 19,214 - 243.
(1976) The depletion of carbon dioxide from lake
water by phytoplankton. J. Lcol. 64,79 - 121.
Taylor, B. F., lee, C. C. and Bunt, J. S. (1973) Nitrogen
fixation associated with the marine blue-Green alga, Trichodesmium, as measured by the acetylene reduction technique. Arch. Mikrobiol. 88,205 - 212.
Taylor, F. J. (1975) Contamination of water samples during
collection (Note). II. Z. J. Mar. Freshwat. Res. 119. Telitchenko, M. I";., Teytsarian, G. V. and Shirkova, Ye. L.

-229-
(1970) Trace eler. ýents and algal "bloon". Hydrobiol+J.
6,1 - 6.
Thompson, L. M. N. and Macleod, R. A. (1974) Biochemical
localisation of alkaline phosphatase in the cell wall
of a marine pseudorioz. ad. J. Bract. 117,819 - 826.
Tischer, J. (1957) Untersuchungen i1ber die granulgren
Einschlüsse und das Reductions - Oxidationavormogen der
Cyanophyceen. Arch. Mikrobiol. al, 400 - 428.
Topachevskiy, A. V., Braginskiy, L. P. and Sirenko, L. A.
(1969) Massive development of blue-green algae as a
product of the ecosystem of a reservoir. Hydrobiol. J.
5,1 - 10. [English tranalutioni.
Trainor, F. R. and Shubert, L. E. (1974) Scenedesmus,
morphogeneois: colony control in dilute media. J. rhveo_
10,28 - 30.
Tyagi, V. V. S. (1974) Some observations on the pattern of
sporulation in a blue-green alga, Anabaena doliol. um.
Ann. Bot. M, 1107 - 1111.
Vance, B. D. (1965) Composition and succession of
cyanophyceaii water blooms. J. Yh col. 1,81 - 86.
Vasil'eva, V. E. and Levitin, M. G. (1974) Effect of carbon
dioxide starvation on certain blue-green algae. Finiol.
Rast. Most. )_ 21,1207 - 1211.
Vladimirova, K. S. (1968) The correlation between the
phytoplankton and the phytomicrobenthos of reservoirs.
In "Tsvetnie" Vody ("Blooming, Water) (ad: Topuohuvokky,
A. V. ) pp. 67 - 81. Izdat "Naukova" Durnka, Kiev,
U. S. S. R. [English translationJ.
Volk, S. L. and Phinney, H. K. (1968) Mineral requirements
for the growth of Anabaena spiroides in vitro. Can. J.
Bot. 4-6,619 - 630.
Vollenweider, R. A. (1974) A Manual on Methods for
Measuring, Primary Production in Aquatic Environments.
International Biological Programme Handbook No. 12.
Blackwell ; scientific Publications, Oxford, London and Edinburgh.
Walsby, A. E. (1971) The pressure relationships of Can
vacuoles. Proc. R. Soc. B J, 301 - 326.
and Booker, N. J. (1976) The physiology of water

-23C-
bloom formation by planktonic blue-green algae. Br. -
phycol J. 11,200 only (Abstr. ).
and Bichelberger, 11.11. (1968) The fine structure of
gas vacuoles released from the cells of the blue-green
alga Anabaena flos-acguse. Arch. Ilikrobiol. 60,76 - 83.
and Klever, A. R. (1974) The role of gas vacuoles in
the aicrostratification of a population of Oscillatoria
aagardhii var. isothrix in Deming Lake, Ilinnesota. Arch.
H_ý! drobiol . 74,375 - 392.
Wardle, T. (1897) The breaking of Copmere. Trans. North
Staffs. Field Club 1897 volume, 110 - 123.
Watanabe, A. and Yamamoto, Y. (1967) Heterotrophic nitrogen fixation by the blue-green alga Anabaenopsis circularie.
" Nature, Lond. 21A, 738.
Mosre, 11. M. and Benenann, J. R. (1973) Nitrogen fixation by
Anabaena cylindrica II. Nitrogenase activity during
induction and ageing of batch cultures. Arch. Mikrobiol.
22,101 - 112.
Westlake, D. F. (1966) The light climate for plants in
rivers. In Light as an Ecological Factor (eds:
Bainbridge, R., Evans, G. C. and Rackham, 0. ) pp. 99 - 119. Blackwell Scientific Publishers, Oxford and Edinburgh.
White, 11. S. and Wetzel, R. G. (1975) Nitrogen, phosphorus,
particulate and colloidal carbon content of aedimenting
sexton of a hard-water lake. Verh. int. Verein. theor.
annew. L mnol. 12,330 - 339. Whitton, B. A., Kirkby, S. M., Peat, A. and Sinclair, C-
(1973) Environmental effects on the morphology of flivulariaceae trichomes. Srmn. Prokaryotic Photosynthetic Organisms, Preiburg. pp. 180 - 181 (Abstr. ).
Wieringa, K. T. (1968) A new method for obtaining bacteria- free cultures of blue-green algae. Antonie van Leeuwenhoek , 54 - 56.
Wildman, R. B., Loescher, J. H. and Winger, C. L. (1975)
Development and germination of akinetes of Anhanizomenon flos-aquae. J. Phtºcol. 11,96 - 104.
Woelkerling, W. J., Kowal, R. R. and Gough, S. B. (1976)
Sedgwick - Rafter cell counts: a procedural analysis.

-231-
ý11- drobiolotIia 48,95 - 107.
''loll;, C. P. (1965) Control of cporulation in blue-green
algae. Devl Biol. 12,15 - 35.
(1956) Evidence of a role of heterocysts in the
sporulation of a blue-green alga. Am. J. Bot. ,
260 - 262.
(1968) Movement of carbon from vegetative cello to
heterocysts in Anabaena cylindrica. J. Bact. 269
2138 - 2143.
(1975) Differentiation and pattern formation in
fL lamentous blue-green algae. In Spores, Vol. VI.
PP. 85 - 96. Am. Soc. Microbiol.
Wood, E. D., Armstrong, F. A. J. and Rioharde, F. A. (1967)
Determination of nitrate in sea water by cadmium-copper
4, reduction to nitrite. J. mar. biol. Aso. U. R..
23 - 31. Wyatt, J. T. and Silvey, J. X. G. (1969) Nitrogen fixation
by Gloeoc_ansa. Science. N. Y. 165,908 - 909.

Proc. R. Soc. Lond. B. 196,317-332 (1977)
Printed in Great Britain
Sporulation and the development of planktonic blue-green
algae in two Salopian meres
BY J. A. ROTHER AND P. FAY
Department of Botany and Biochemistry, Westfield College, University of London
(Communicated by G. E. Fogg, F. R. S. - Received 23 June 1976)
Factors affecting akineto formation and the survival of planktonic blue- green algal populations after the formation of `water blooms' were investi- gated in two eutrophic kataglacial lakes. It is considered that the induction of sporulation may be due to extreme conditions at the water surface during the summer to which the algal bloom is exposed, rather than to nutrient, especially orthophosphate, deficiency. It has not been shown conclusively that akinetes constitute an overwintering stage of the planktonic algal life cycle, or that akinetes germinate in the spring to produce the inoculum for subsequent vegetative growth. Comparisons of planktonic, sedimenting and benthic algal material indicate that germina- tion shortly after maturation may provide the greater part of the over- wintering planktonic vegetative populations. Population sizes and life cycles appear to be influenced by the physical factors which control the stability of the water column.
INTRODUCTION
`Water bloom' formation has been shown by Reynolds (1971,1972,1973a) to result from the redistribution of buoyant planktonic blue-green algae. Buoyancy is imparted by gas vacuoles, and the rate of surface accumulation is dependent on the amount of gas vacuoles within the algal cell, the degree of turbulence within the water body and the size of the alga. The larger colonial species have a greater potential for rapid movement through the water column, both in terms of buoyancy and sedimentation (Reynolds & Walsby 1975). The conditions at the surface are highly unfavourable to the survival of vegetative algal material, and algae frequently undergo photo-oxidative death after bloom formation (Abeliovich & Shilo 1972; Boyd, Prather & Parks 1975). Some colony forming genera (Micro-
cystis and Coelosphaerium) are more resistant to damage than the filamentous forms discussed in this paper (Reynolds 1967). Since much of the total algal bio-
mass may be concentrated within a few centimetres at the surface during a bloom, the survival of some of this material will be critical for the development of future
populations. An important adaptation of many heterocystous blue-green algae in conditions
unfavourable for vegetative growth appears to be the formation of akinetes, [ 317 1

318 J. A. Rother and P. Fay
resistant cells well protected by an elaborate envelope (Fogg, Stewart, Fay & Walsby 1973) each of which differentiates from a single vegetative cell. Earlier
studies indicate that akinetes but not vegetative filaments survive in the surface mud sediments during the winter and germinate in the spring (Rose 1934; Roelofs & Oglesby 1970; Wildman, Loescher & Winger 1975). Germination of akinetes at a time when the lakes are still unstratified, apparently occurred in response to more favourable conditions of light and temperature (Reynolds 1971,1972). The
significance of this process is doubtful as the number of akinetes is never very large
and hence germination of akinetes may not contribute significantly to the popula- tion maximum (Reynolds 1972,1975). Growth can mostly be accounted for by
vegetative reproduction of a small overwintering stock of planktonic material. Non-sporulating species, especially Microcystis, have been shown to remain viable during the winter in the mud sediments of the deeper lakes, giving rise to plank- tonic populations the following spring (Sirenko, Chernousova, Arendarchuk & Kozitskaya 1966; Reynolds & Rogers 1976).
In laboratory experiments on akinete formation, it has been shown that a reduction in concentration of a critical nutrient such as orthophosphate (Wolk 1965) or the increase of extracellular products (Fisher & Wolk 1976) induces akinete development, that is, the phenomenon is characteristic of stationary phase cultures. Field studies on factors affecting akinete production are relatively sparse. Roelofs & Oglesby (1970) conclude from field and laboratory observations on Gloeotrichia ec1 inulata that akinete formation is not a result of unfavourable conditions, as akinetes were seen during the logarithmic phase of growth.
An attempt was made in this investigation to assess the importance of akinetes in the formation of planktonic algal propulations of a number of species, in two contrasted lacustrine environments, and to determine which factors controlled the various stages of the algal life cycle. The early stages of development may shed light on the mechanism of the annual succession of the heterocystous blue-green algal types and give an indication of the possible size of the population maximum.
THE SITES
Two meres, Kettle Mere and White Mere, of the Ellesmere group were investi- gated (figures la-c). Kettle Mere (National Grid reference: SJ 418 341) one of the smallest of the Salopian (Shropshire) meres, has a surface area of 2 ha and a maximum depth of 7 m. It is well sheltered from the wind, being at the bottom of a deep conical depression and fringed with woodland. White Mere (National Grid reference: SJ 415 330) with a surface area of 26 ha and a maximum depth of 15 m (in average between 4 and 8 m) is more exposed. Neither of the meres have signifi- cant inflows or outflows and rely on phreatic and surface drainage for water supply. Both meres are eutrophic and support large populations of blue-green algae. More detailed descriptions of the Meres are available in articles by Griffiths 1925), Gorham (1957), Sinker (1962) and Reynolds (1973b).

Sporulation of planktonic blue-green algae 319
METHODS
Sampling
Plankton samples from the sites indicated on figures 1b and 1c were collected using a Ruttner sampler, were preserved in the field in Lugol's iodine and enume- rated using the iodine sedimentation and inverted microscope technique of Lund, Kipling & Le Cren (1958). More rapid sedimentation which is less prone to re- suspension, particularly of the larger colonial gas vacuolate species, was achieved by collapsing the gas vacuoles using the `hammer, cork and bottle' technique of Klebahn (1895). Estimates of heterocyst and akinete frequency were made by
counting a minimum of 1000 cells along randomly chosen filaments. Total sus- pended solids and algal dry mass were determined by weighing the amount of particulate material after filtration onto Whatman GF/C glass fibre filters and drying at 110 °C under atmospheric pressure for 24 h.
During 1975 a Jenkin surface mud sampler (Mortimer 1942) was used to collect mud samples at the sediment-water interface, at other times the Ruttner sampler was used. The top 5-10 cm section of the core was used to enumerate algal fila- ments, akinetes and empty akinete envelopes. Due to the loose and flocculent
FzoU I. The Ellesmere group of meres. (a) Location of meres. (b) Kettle Mere, indicat. ing position of sediment traps (0) and sampling point (0). (c) White Mere, symbols as in (b).

320 J. A. Rother and P. Fay
nature of the surface mud deposits it was not possible to section the cores and therefore the data are expressed in terms of number per unit volume. The mud samples were preserved in 4% formaldehyde. In order to facilitate identification of the algal material against the background of detritus, a Zeiss Standard 18 Micro- scope with Epi-fluorescence condenser IV/F was used, utilizing the natural fluorescence of chlorophyll a. This was supplemented by observations using normal transmission light microscopy.
Sedimentation of algal material through the water column was followed by using sediment traps. Each trap consisted of 14 or 16 plastic tubes 1.5 cm in dia- meter, held in a plastic container and suspended 2m above the surface mud sediments, below the summer thermocline. The amount of material collected in this type of trap has been shown by Kirchner (1975) not to vary on a unit area basis for traps of different sizes. Surface wave action was minimized by using submerged floats attached 2m above the trap itself. The trap was located by a small surface buoy attached to the submerged float by a loose line. No growth of epiphytes occurred on traps positioned 4m below the water surface, and any accumulation of bacterial material on the tubes was minimized by replacing them with a new set of clean tubes when samples were collected. These requirements prescribed that the traps were positioned in water which was not less than 6m deep. Total sedimentation rates were assessed in terms of dry mass of material sedimented per square metre per day. Quantitative and qualitative estimations of the sedimentation of algal material were also made.
Analysi8 Profiles of oxygen saturation (using an Electronic Instruments Ltd oxygen
probe), temperature, and light penetration (using a Griffin Environmental Comparator) were obtained.
Analyses of orthophosphate were carried out on sterile filtered samples using the molybdenum blue method as described by Golterman (1971) or on site using a Hach Direct Reading Engineers' Laboratory (model DR-EL/2). The analysis procedure used in the latter is derived from a method described in the American Public Health Association Standard Methods (1971) and is similar in principle to the method used in the laboratory. The results obtained are comparable.
Analyses of labile cellular orthophosphate were made on washed filtered algal material following the method of Fitzgerald & Nelson (1966), and assays of alkaline phosphatase activity were performed using the p-nitrophenol phosphate technique as described in Sigma Technical Bulletin No. 104 (1974). Algal material for these analyses was concentrated by allowing the buoyant material to float to the surface of the vessel, and then pipetting off a known amount of the dense suspension. Aliquots of the concentrate were used to determine dry mass. The results are expressed as milligrams of orthophosphate or enzyme activity per milligram of dry alga.

Sporulation of planktonic blue-green algae 321
TABLE 1. AMOUNT OF ALGAL MATERIAL AT THE WATER SURFACE : FILAMENTS
COLONIES OF MICROCYSTIS) PER MILLILITRE, KETTLE MERE
date A. circi- A. foa. A. apiro- A. aoli- Aph. foa- Micro. (1974) nalia aquae idea taria aquae cyatis 10/6 277 0 0 0 3363 0 16/6 500 0 0 0 700 0
2/7 0 0 40 0 100 0 9/7 25 0 40 0 100 2
18/7 31 0 8 0 35 0 30/7 0 0 1 0 21 4 13/8 0.2 0.4 4.4 0 0 49 29/8 1.6 0 9 0 0 98
9/9 0 0 7 0 0 80 27/9 10 29 5 0 0 61 20/11 0.1 0.5 0 0 0.2 2.5 16/12 0.03 0.01 0 0 0 1.9
(1975) 14/3 0 000 0 1.1
5/5 104 51 00 1 0.6 9/6 22 68 7 14 28 13
23/6 0 0 0.9 1.3 6.3 10 7/7 0 2.7 3.3 4.7 0 2.7
22/7 0 001.4 0 56 16/8 0 05 225 0 44 18/8 0.2 00 120 0 74 23/8 2 06 255 24 33 26/8 0 0 0.3 251 28 28 6/10 0 0 1.5 0 0 0
G. echinulata was not observed.
RESULTS
Among the most important factors which control the annual cycle of growth and reproduction of algae are light and temperature. The differing responses of the
various algal species result in the typical succession of the bloom forming species (tables 1 and 2) in the meres investigated. Generally the filamentous species are the first to bloom, especially Anabaena circinalis (Kütz. ) Hansgirg and Aplaani-
zomenon floc-aquae (L. ) Ralfs., which are followed by Anabaena flos-aquae (Lyng. ) Breb., Anabaena solitaria Kleb., Anabaena spiroides Kleb. and Gloeotrichia echinu- lata (Smith) Richt. Microcystis aeruginosa Kütz. typically blooms in the autumn. The differences between the two meres are probably the result of differences in the physical factors. White Mere is considerably more exposed to wind action than Kettle Mere and consequently does not show thermal stratification until July (figure 2c) whereas Kettle Mere is stably stratified from June to September (figure 2a). Changes of dissolved oxygen (figure 2b, d) also reflect the stability of the water column. Below the relatively shallow epilimnion of Kettle Mere the
water becomes almost totally deoxygenated. This is less marked in White Mere

322 J. A. Rother and P. Fay
TABLE 2. AMOUNT OF ALGAL MATERIAL AT THE WATER SURFACE: FILAMENTS
COLONIES OF MICROCYSTIS) PER MILLILITRE, WHITE MERE
date A. circi- A. fos- A. Spiro- A. soli- Aph. fos- Micro. (1974) nalis aquae ides taria aquae cystic
10/6 0 0 0 0 0 0 16/6 40 0 0 0 0 0 2/7 235 0 0 0 0 86 9/7 366 17 0 0 0 4
18/7 603 301 247 20 0 219 30/7 3189 2035 6 7 0 0.4 13/8 490 0 18 8 0 11 29/8 20 21 7 5 0 12
9/9 106 50 1 1 8 36 27/9 109 5 0.2 0.2 15 33 20/11 18 71 0 0.6 3.3 0.7 16/12 4.5 24 0 1.3 0.2 0.2
(1975) 24/3 181 177 0 0 0 0
5/5 0 22 0 0 1 0 9/6 0 6 0 0 0 0
23/6 7 8 0 0 0 0 7/7 557 391 7 6 2.2 0.7
22/7 17 29 1.4 55 14 0 16/8 0 1741 0.4 4 0 0 18/8 0 965 0.2 4 2 3.4 23/8 0 680 0 0.6 0 7 26/8 0 741 0 1.4 0 8 6/10 0 0.04 0 0 0 0.6
Colonies of 0. echinulata were observed in the plankton in July and August 1974 and 1975.
which is stable for a shorter period of time. In both meres there are periods of supersaturation of dissolved oxygen in the surface layers during calm weather. The earlier stratification of Kettle Mere produces an earlier and more intense blooming of algae. As a consequence of the earlier stratification, of the shallow epilimnion and of the considerable absorbance (table 3) of the peaty water, popula- tion levels are generally lower in Kettle Mere than in White Mere. Populations also undergo a more dramatic decline which could be as a result of the greater surface concentration of the algae in Kettle Mere.
During this time of calm weather and intense solar radiation, and with the greater part of the algal population floating within a few millimetres of the surface of the water, a sudden change in the physiological state of the algae takes place, which manifests itself in extensive akinete formation. This process is particularly rapid in Kettle Mere and this rapidity accounts for the lack of detailed records in table 4. The situation is similar in White Mere, though modified by physical factors (table 4). The onset of akinete formation coincides with the stabilization of the water column and the formation of surface scums of the species concerned.

Sporulation of planktonic blue-green algae 323
z
O b
db
4)
10 b
tl ö
q ti oý L4 äý v
1
äo
ao$ pD
ti
b
dý f7
GH
rü
(uz) ugdep

324 J. A. Rother and P. Fay
TABLE 3. DEPTH OF 10 % OF SURFACE RADIATION
depth (m) depth (m)
date Kettle White date Kettle White (1974) Mere Mere (1975) Mere Mere
10/0 1.5 4.5 24/3 1.6 2.0 18/8 1.0 3.0 5/5 2.0 4.1 2/7 1.0 2.5 2/6 2.0 3.7 9/7 1.0 1.7 2 310 1.4 4.7
18/7 1.0 1.7 7/7 1.4 3.0 30/7 1.0 1.0 22/7 0.9 1.9 13/8 1.5 1.0 18/8 1.0 1.5 29/8 1.0 1.7 18/8 1.0 1.0
9/9 1.5 1.6 23/8 1.0 1.3 20/10 0.9 2.0 26/8 1.0 1.0 20/11 1.0 - 6/10 2.5 1.0 16/12 1.0 1.5 27/10 1.0 5.0
18/11 1.0 2.0
TABLE 4. AKINETE PERCENTAGE OF ALGAE FROM THE MERES
Aphani.
A. circi- A. fl o8- A. epiro- A. Soli- zomenon nalis aquae ide8 taria
, los-aquae
date r--ýý----, ý-ý--, ý--ý-ý ý-ýý (1974) KM WM KM WM KM WM KM WM KM WM
18/7 t--000---- 30/7 - 3.1 - 0.2 - 3.0 - 1.1 -- 13/8 -02.0 1.0 - 4.1 - 1.5 -- 29/8 0 0.1 - 1.2 0.5 4.6 - 2.5 --
9/9 ---------- 27/9 ttttt-----
(1975)
24/3 -0-0-0-0-- 5/5 0-000----- 9/6 0$ -000-0-0-
23/6 -t-------- 7/7 -t--0.2 -0---
22/7 -0-0--00.3 -0 16/8 -0-0.2 --0.1 --- 18/8 ---0.1 --0-0- 23/8 ---1.0 --2.1 -0- 26/8 ---0------
6/10 ---0-----0
t Akinetes observed but insufficient for quantitative estimation. $ In Kettle Mere A. circinalis had undergone very extensive sporulation by early June;
on 9 June clumps of akinetes were found in the sheltered areas of the mere, but little remained in the plankton at the sampling site.
-, insufficient material of the species concerned. 0, no akinetes. KM, Kettle Mere. WM, White Mere.

Sporulation of planktonic blue-green algae 325 In contrast to the well-marked succession of species in Kettle Mere, many of the filamentous species in White Mere form surface blooms and produce akinetes more or less simultaneously.
The process of sporulation effectively removes the alga concerned from the plankton. The transformation of a gas vacuolate vegetative cell into an akinote lacking gas vesicles and accumulating large cyanophycin bodies leads to a con- siderable increase in the specific gravity of the cell. This could be the reason for the
rapid sedimentation of akinetes. Sedimentation is enhanced by the aggregation of akinetes into large clumps and by the breakdown of filamentous material. While in general clumps of detached akinetes sediment more rapidly than the surviving vegetative material, the greater turbulence in the surface layers of White More during July-August 1975, breaks up the clumps so that filaments and akinetes sediment at an equal pace. Rates of sedimentation of the total particulate matter are given in figure 3a. Maximum rates of sedimentation of particulate material generally correspond with high rates of sedimentation of algal material, particu- larly of Mierocystis in Kettle Mere during the autumn, and of Asterionella in White Mere in late spring. The smaller peaks of sedimentation in Kettle More are also reflections of the algal contribution (figure 3b). The pattern of sedimentation in White Mere (figure 3c) is disrupted by the earlier breakdown of stratification in August resulting in the resuspension of bottom deposits. Although the main algal component in the sedimented matter in Kettle Mere is A. circinalis, and the data in figure 3b are the total of all the Anabaena species, the clear succession of species results in distinct periods of sedimentation for each species. The minor increase during the second half of August 1975 in Kettle Mere is due to the sporulation and sedimentation of A. solitaria.
There is little survival during the winter of vegetative filaments in the mud of Kettle Mere. The only period during which material was observed in the mud was immediately after sedimentation. Filaments appeared to persist longer at the mud surface in White Mere (figure 4a) but much of this is due to an interchange
of material between the plankton and the benthos when stratification breaks down,
rather than indicating long survival of sedimented vegetative material. In both mares there is a considerable and rapid input of akinetes to the surface mud layers during the period immediately after a bloom (figure 4 b, c) but there is also an equally rapid decline in numbers in both meres. (Note that the data on akinete which are expressed in terms of numbers per millilitre cannot be quantitatively compared with data on rates of sedimentation expressed as numbers per square metre per day. )
Orthophosphate concentrations are given in table 5. In both mores the concentra- tion is high throughout the year with only a slight decrease in the surface waters during stratification. Analyses of intracellular labile orthophosphate, an indication of surplus phosphate within the cell, show relatively high values of 0.01-0.05 mg orthophosphate per milligramme algal dry mass, particularly when compared with stationary phase batch cultures. The high levels of available phosphate are also reflected by the low levels of alkaline phosphatase activity (Fitzgerald &

326 J. A. Rother and P. Fay
" o
1
I. N O N O N O
1-'ABP '-tu 2 (e-01 x99 9Ibo. capJt) T-'c'ep : -w 1_ &gp '-ui xaquinu x 9-01
aequznu x 9-01
z O_
öw
mmý A mýý
ý' ao
vi n
z mý o
o hm
n +ý

Sporulation of planktonic blue-green algae
tl. "O cý o eý o 0 0
N cV
Gldtuvs T_jui aegmnu x a-Ol
zm m
ob
ým
d° 'O ä¢ý
ti IH 0
q
os
aa. q
Ö
.r
zö
mý n
C7 rM1"1
327

328 J. A. Rother and P. Fay
Nelson 1966). Algal material collected from surface blooms had alkaline phosphat- ase activities in the range of 1.0 to 2.0 Sigma units per milligramme dry mass of alga. (1 Sigma unit is equivalent to the liberation of 1 µM p-nitrophenol per h. ) This is comparable with activities recorded in logarithmically growing laboratory
cultures of algal species isolated from the meres. Old cultures in the post-log- arithmic phase of growth had activities of 3-6 Sigma units with low levels of orthophosphate remaining in the medium at this stage of growth.
TABLE 5. ORTHOPHOSPHATE CONCENTRATIONS IN THE MERES
(mg P04 P PER LITRE), 1975
site Kettle Mere White Mere depth of
sample Om 3m 6m Om Mt B$
515 1.70 1.80 1.20 0.67 0.70 0.80 9/6 1.30 0.75 2.30 0.80 0.23 0.75
23/6 0.34 0.69 1.18 0.32 0.50 0.97 7/7 0.35 0.69 1.70 0.33 0.85 1.31
22/7 0.40 0.74 1.78 0.60 0.58 1.08 16/8 0.98 1.30 2.50 0.83 0.98 2.30 18/8 0.74 1.61 3.20 1.31 0.66 2.70 23/8 1.66 1.70 4.60 1.52 2.60 4.70 26/8 1.50 1.07 0.92 1.65 0.66 2.95 29/9 - - - 1.01 1.01 -
6/10 0.45 0.56 1.24 1.03 1.03 1.04 27/10 0.56 0.56 1.26 1.65 1.80 1.81 15/11 0.77 0.75 0.75 0.82 0.82 0.86
j Middle of water column (3-4 m). $1m above bottom (6-9 m).
DISCUSSION
The coincidence of akinete production with dense surface accumulations of algal material suggests that light may play a critical role, rather than nutrient deficiency,
especially of orthophosphate in the induction of sporulation. Despite the dense
scum of algae at the water surface, the individual cells do not appear to be de- ficient in phosphorus. Blue-green algae are capable of storing through luxury uptake, sufficient phosphorus reserves, in the form of polyphosphate bodies, to support the growth of two further generations when transferred to a medium deficient in phosphorus (Batterton & Van Baalen 1968). Thus, even if diffusion of phosphates through the stagnant surface layers of water were the only means of transport to the algal cell, there would be sufficient reserves for a limited period of active growth. The analyses of labile orthophosphate and also alkaline phosphat-
e activity of algae collected during a bloom indicate that either the supply from the water to the algal cells is sufficient to maintain a high level within the cell or that whatever reserves were accumulated previously were not utilized.
High light intensity together with supersaturated oxygen conditions results in the photo-oxidative death and bacterial breakdown of algal material (Abeliovich

Sporulation of planktonic blue-green algae 329
& Shilo 1972). The production and release of organic compounds may initiate
sporulation in the manner suggested by Fisher & Wolk (1976), the light intensity
and quality (particularly in the u. v. region) may have a direct photoinductive effect of the type described by Lazaroff (1966) and Lazaroff & Schiff (1962), or akinete production may be the result of a physiological imbalance between
nitrogen fixation and photosynthesis while the latter is inhibited by the high light intensity. Tyagi (i974) observed that akinete production in Anabaena doliolum $haradw. is inhibited by inorganic nitrogen compounds but is stimulated by
glucose. In this species, akinete differentiation begins in cells which are equidistant between two heterocysts, and sporulation proceeds centrifugally. However, in all the planktonic species examined in the meres, the first akinete is developed from the third vegetative cell from the heterocyst and sporulation proceeds centri- petally to produce a short chain of two or three akinetes. Any small variation in the number of cells between the first akinete and the heterocyst may be due to cell division subsequent to the formation of the akinete. Large variations may be due to filament breakage resulting in the separation of closely associated heterocysta
and akinetes. This seems to be a consistent feature of the planktonic Anabaena species, and a similar relationship has also been observed in Aph. floc-aquae. A considerable amount of variability in the number of cells between heterocyst and akinete was observed in Anabaena planktonica Brunth. by Burger (1974)
although 60 % of akinetes were found four or less cells away from the heterocyst. From the consistent relationship between heterocysta and akinetea in the species
examined, it would seem to be that heterocysts exert some degree of control over akinete production (Wolk 1966). However, it seems that some external factor is
required to trigger off the process. During the calm weather of early December 1975, a dense surface scum of A. circinalis accumulated at the lee shore of White Mere. The filaments appeared intact and healthy, and they incorporated hetero-
cysts but no akinetes. This supports the idea that a high level of solar radiation is
required for sporulation. The increase of cytoplasmic density results in the sedimentation of akinetes to
the mud surface. Although there is a rapid increase in the number of akinetes in the surface deposits, there is also an equally rapid decline in numbers soon after in both meres. This could be, but is probably not, due to lifting of material by turbulence, since Kettle Mere was still stratified in early October of both years 1974 and 1975, as was White Mere at the end of July. Very few akinetes were observed in the plankton after sedimentation had occurred, certainly not enough to account for the fall in numbers in the mud even if it is considered that the material was evenly distributed through the water column. The decline is probably due to akinete germination soon after maturation, which may account for the
small increase of the filamentous population observed about a month after the
occurrence of surface blooming. From the point of view of future development of the algae this would be more important in Kettle Mere, where algal numbers were reduced to undetectable levels, than in White Mere where a considerable proportion
12 Vol. 196. B.

330 J. A. Rother and P. Fay
of the population had survived blooming. Although only a few instances of akinete germination have been observed in plankton samples, up to 60 % of the akinetes examined were empty while still within the plankton. Germination shortly after formation has been observed in laboratory cultures of isolates of algae from the meres and also in cultures of Anabaena cylindrica Lemm. (Fay 1969). The generally large overwintering planktonic populations of vegetative filaments, especially in White Mere, indicate that akinete germination is not an important source of algal material during the spring as most of the growth can be accounted for by the material already within the plankton. There was no marked decrease of akinetes in the spring, or any signs of extensive germination. From the observation that akinetes germinate soon after their maturation, their function is seen more in terms of survival of algal populations during the period of blooming, rather than promoting their survival during the winter. In Kettle Mere, where the over- wintering planktonic populations are much lower, akinete germination during the spring may be more important to the early development of algal populations.
Griffiths (1925) observed that Aph. flos-aquae did not produce akinetes in Kettle Mere, but did so in a nearby mere, The Mere, Ellesmere. During the period of this investigation no akinetes were observed in the Kettle Mere strain, but the White Mere strain, although a minor component of the phytoplankton, produced akinetes. This is probably a phenotypic rather than a genotypic difference, as akinetes have been produced in the Kettle Mere strain which has been isolated and grown in the laboratory. However, any extrapolations to the field from experiments done in the laboratory using isolates grown from natural water bodies must be viewed with caution due to the genetic plasticity of blue-green algae. During 1974 a dense surface bloom of Apl&. flos-aquae was observed in Kettle Mere, which rapidly decreased in numbers. During the rest of the year, no material was found, either in the plankton or in the sediments. In 1975 the size of the population of the species was less than 1% of that in the previous year. This supports the idea that akinetes are important for the survival of a sufficient stock of algal material to produce a large population in future years.
We are indebted to the Trustees of the Grosvenor Estate for permission to work at White Mere, to Mr R. K. Mainwaring and Major K. W. Pettergree for permission to work at Kettle Mere, to Mr L. R. Jebb for allowing access to the shore of White Mere and for storage facilities, and to the Freshwater Biological Association for the use of the Meres Laboratory at Preston Montford Field Centre. We are grateful to Dr C. S. Reynolds for helpful comments on the manuscript of this paper. This work has been carried out with the support of a grant from the Natural Environ- ment Research Council.

Sporulation of planktonic blue-green algae 331
REFERENCES
Abeliovich, A. & Shilo, M. 1972 Photo-oxidative death in blue-green bacteria. J. Bact. 111, 682-689.
American Public Health Association. 1971 Standard methods for the examination of water and wastewater, 13th ed. New York: Am. Publ. Health Assoc., Inc.
Batterton, J. C. & Van Baalen, C. 1968 Phosphorus deficiency and phosphate uptake in the blue-green alga Anacystis nidulans. Can. J. Microbiol. 14,341-348.
Boyd, C. E., Prather, E. E. & Parks, R. W. 975 Sudden mortality of a massive phyto. plankton bloom. Weed Sei. 23,61-67.
Burger, J. 1974 A study of two populations of Anabaena planktonica (Cyanophyta) from Minnesota. Phycologia 13,125-129.
Fay, P. 1969 Metabolic activities of isolated spores of Anabaena cylindrica. J. exp. Bot. 20, 100-109.
Fisher, R. W. & Volk, C. P. 1976 Substance stimulating the differentiation of spores of the blue-green alga Cylindrospermum lich. eniforme. Nature, Lend. 259,394-395.
Fitzgerald, G. P. & Nelson, T. C. 1966 Extractive and enzymatic analyses for limiting or surplus phosphorus in algae. J. Phycol. 2,32-37.
Fogg, G. E., Stewart, W. D. P., Fay, P. & Walsby, A. E. 1973 The blue-green algae. London and New York: Academic Press.
Golterman, H. L. 1971 Methods for chemical analysis of fresh waters. I. B. P. Handbook no. 8. Oxford, London and Edinburgh: Blackwell Scientific Publications.
Gorham, E. 1957 The chemical composition of some waters from lowland lakes in Shropshire, England. Tellus 9,174-179.
Griffiths, B. Al. 1925 Studies in the phytoplankton of the lowland waters of Great Britain. III. The phytoplankton of Shropshire, Cheshire and Staffordshire. J. Linn. Soc. (Bot. ) 47,75-98.
Kirchner, W. B. 1975 An evaluation of sediment trap methodology. Limnol. Oceanogr. 20, 657-660.
Klebahn, H. 1895 Gasvakuolen, ein Bestandteil der Zellen der wasserblütebildenden Phyco- chromaceen. Flora, Jena 80,241-282.
Lazaroff, N. 1966 Photoinduction and photoreversal of the Nostocacean developmental cycle. J. Phycol. 2,7-17.
Lazaroff, N. & Schiff, J. 1962 Action spectrum for developmental photo-induction of the blue-green alga Nostoc muscorum. Science, N. Y. 137,603-604.
Lund, J. W. G., Kipling, C. & Le Cren, E. D. 1958 The inverted microscope method of estimating algal numbers and the statistical basis of estimating by counting. Hydro- biologia 11,143-170.
Mortimer, C. H. 1942 The exchange of dissolved substances between mud and water in lakes. J. Ecol. 30,147-201.
Reynolds, C. S. 1967 The breaking of the Shropshire Mares: Some recent investigations. Bull. Shrops. Conserv. Trust. 10,9-14.
Reynolds, C. S. 1971 The ecology of the planktonic blue-green algae in the North Shropshire mares, England. Fld Stud. 3,409-432.
Reynolds, C. S. 1972 Growth, gas vacuolation and buoyancy in a natural population of a blue-green alga. Freshwat. Biol. 2,87-106.
Reynolds, C. S. 1973 a Growth and buoyancy of Microcystis aeruginosa Kütz. emend. Elenkin in a shallow eutrophic lake. Proc. R. Soc. Lond. B 184,29-50.
Reynolds, C. S. 1973b Phytoplankton periodicity of some North Shropshire meres. Br.
phycol. J. 8,301-320. Reynolds, C. S. 1975 Interrelations of photosynthetic behaviour and buoyancy regulation
in a natural population of a blue-green alga. Freshwat. Biol. 5,323-338. Reynolds, C. S. & Rogers, D. A. 1976 Seasonal variations in the vertical distribution and
buoyancy of 1llicrocystis aeruginosa Kütz. emend. Elenkin in Rostherne Mere, England. Hydrobiologia 48,17-23.
Reynolds, C. S. & Walsby, A. E. 1975 Water-blooms. Biol. Rev. 50,437-481. I2-2




















