
Working Memory Training Promotes General Cognitive Abilities in Genetically Heterogeneous Mice
6
Current Biology 20, 777–782, April 27, 2010 ª2010 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2010.02.034 Report Working Memory Training Promotes General Cognitive Abilities in Genetically Heterogeneous Mice Kenneth R. Light, 1 Stefan Kolata, 1 Christopher Wass, 1 Alexander Denman-Brice, 1 Ryan Zagalsky, 1 and Louis D. Matzel 1, * 1 Program in Behavioral Neuroscience, Department of Psychology, Rutgers University, Piscataway, NJ 08854, USA Summary In both humans and mice, the efficacy of working memory capacity and its related process, selective attention, are each strongly predictive of individuals’ aggregate perfor- mance in cognitive test batteries [1–9]. Because working memory is taxed during most cognitive tasks, the efficacy of working memory may have a causal influence on individ- uals’ performance on tests of ‘‘intelligence’’ [10, 11]. Despite the attention this has received, supporting evidence has been largely correlational in nature (but see [12]). Here, genetically heterogeneous mice were assessed on a bat- tery of five learning tasks. Animals’ aggregate performance across the tasks was used to estimate their general cognitive abilities, a trait that is in some respects analogous to intelli- gence [13, 14]. Working memory training promoted an increase in animals’ selective attention and their aggregate performance on these tasks. This enhancement of general cognitive performance by working memory training was attenuated if its selective attention demands were reduced. These results provide evidence that the efficacy of working memory capacity and selective attention may be causally related to an animal’s general cognitive performance and provide a framework for behavioral strategies to promote those abilities. Furthermore, the pattern of behavior reported here reflects a conservation of the processes that regulate general cognitive performance in humans and infrahuman animals. Results Working Memory Exercise Promotes Selective Attention and General Cognitive Performance Procedures and results from individual tests are presented in the Supplemental Information available online. One group of animals received 12 days of training wherein they performed concurrently in two eight-choice radial arm mazes. This task was designed to promote a heavy reliance on all aspects of working memory (including aspects of short-term mainte- nance, capacity, and selective attention) and has been described previously [8, 9]. A second group received compa- rable handling absent any working memory training. When both groups of mice were subsequently tested on a mouse analog [9] of the human Stroop test, mice that received this working memory training performed significantly better than animals that had not undergone training. That is, task-relevant interference provoked fewer errors in animals that had previously undergone working memory training [Figure 1B; F(1,25) = 8.20, p < 0.01], suggesting that this training promoted an increase in the efficacy of selective attention. All animals were then tested on five independent learning tasks (passive avoidance, odor discrimination, associative fear conditioning, egocentric navigation in a Lashley maze, and spatial navigation in a water maze). Results from individual tasks in the learning battery are presented in the Supplemental Information. To obtain general learning scores for all mice, we subjected acquisition data from the five learning tasks to a principal components analysis. A primary factor was extracted with an eigenvalue of 1.51, which accounted for 30% of the variance across animals. From this primary factor, factor scores were extracted to represent each animal’s general learning ability. Factor scores, which are analogous to an average z score of an animal’s performance across all tasks, provide a sensitive index of an individual’s aggregate performance in the learning battery. However, the number of subjects used here is somewhat small by factor analytic stan- dards. Therefore, to verify the accuracy of these factor scores as descriptors of general learning performance, the factor scores were compared to each animal’s average rank perfor- mance (relative to others in the sample) across all five learning tasks (a method that has been previously reported [13]). Factor scores and average ranks were significantly correlated [r(25) = 0.61, p < 0.001]. Because factor scores are presumed to be a more precise measure of an animal’s aggregate performance across all tasks, these scores were used in all subsequent analyses to compare the performance of groups of animals. Mice trained on the concurrent radial arm maze task had significantly higher general learning scores than did animals that were not subjected to this working memory training [Fig- ure 1C; F(1,25) = 4.48, p < 0.05; Cohen’s d effect size = 0.94]. It was possible that working memory training differentially improved general learning scores in a subset of mice (e.g., mice from different regions of the distribution of general learning abilities). To assess this, we compared factor scores for the top and bottom performers on the learning battery across animals that had received or had not received working memory training (Figure 1D). The top half of mice that had undergone working memory training had significantly higher general learning abilities than the top half of animals that had not received working memory training [F(1,11) = 6.88, p < 0.05]. A similar difference was observed in those animals from the bottom halves of the distribution of learning abilities [F(1,11) = 13.92, p < 0.005], suggesting that working memory training promotes an improvement in general learning perfor- mance regardless of an animal’s position in the distribution of learning abilities. In this experiment, animals that underwent working memory training exhibited a commensurate increase in their aggregate performance in a battery of diverse learning tasks. Two alter- native explanations for these results are possible. First, it is conceivable that any manipulation outside of the home cage would have resulted in an increase in general learning abilities simply because it rescues the animals from the environmental deprivation of the home cage. However, this result stands in contrast to previous attempts to manipulate general learning abilities of mice through extensive and repeated exposure to *Correspondence: [email protected]
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Working Memory Training Promotes General Cognitive Abilities in Genetically Heterogeneous Mice

Working Memory Training Pr
Current Biology 20, 777–782, April 27, 2010 ª2010 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2010.02.034
Reportomotes
General Cognitive Abilitiesin Genetically Heterogeneous Mice
Kenneth R. Light,1 Stefan Kolata,1 Christopher Wass,1
Alexander Denman-Brice,1 Ryan Zagalsky,1
and Louis D. Matzel1,*1Program in Behavioral Neuroscience, Department ofPsychology, Rutgers University, Piscataway, NJ 08854, USA
Summary
In both humans and mice, the efficacy of working memorycapacity and its related process, selective attention, are
each strongly predictive of individuals’ aggregate perfor-mance in cognitive test batteries [1–9]. Because working
memory is taxed during most cognitive tasks, the efficacyof working memory may have a causal influence on individ-
uals’ performance on tests of ‘‘intelligence’’ [10, 11]. Despitethe attention this has received, supporting evidence has
been largely correlational in nature (but see [12]). Here,genetically heterogeneous mice were assessed on a bat-
tery of five learning tasks. Animals’ aggregate performanceacross the tasks was used to estimate their general cognitive
abilities, a trait that is in some respects analogous to intelli-gence [13, 14]. Working memory training promoted an
increase in animals’ selective attention and their aggregateperformance on these tasks. This enhancement of general
cognitive performance by working memory training wasattenuated if its selective attention demands were reduced.
These results provide evidence that the efficacy of working
memory capacity and selective attention may be causallyrelated to an animal’s general cognitive performance and
provide a framework for behavioral strategies to promotethose abilities. Furthermore, the pattern of behavior reported
here reflects a conservation of the processes that regulategeneral cognitive performance in humans and infrahuman
animals.
Results
Working Memory Exercise Promotes Selective Attentionand General Cognitive Performance
Procedures and results from individual tests are presented inthe Supplemental Information available online. One group ofanimals received 12 days of training wherein they performedconcurrently in two eight-choice radial arm mazes. This taskwas designed to promote a heavy reliance on all aspects ofworking memory (including aspects of short-term mainte-nance, capacity, and selective attention) and has beendescribed previously [8, 9]. A second group received compa-rable handling absent any working memory training. Whenboth groups of mice were subsequently tested on a mouseanalog [9] of the human Stroop test, mice that received thisworking memory training performed significantly better thananimals that had not undergone training. That is, task-relevantinterference provoked fewer errors in animals that hadpreviously undergone working memory training [Figure 1B;
*Correspondence: [email protected]
F(1,25) = 8.20, p < 0.01], suggesting that this training promotedan increase in the efficacy of selective attention.
All animals were then tested on five independent learningtasks (passive avoidance, odor discrimination, associativefear conditioning, egocentric navigation in a Lashley maze,and spatial navigation in a water maze). Results from individualtasks in the learning battery are presented in the SupplementalInformation. To obtain general learning scores for all mice, wesubjected acquisition data from the five learning tasks toa principal components analysis. A primary factor wasextracted with an eigenvalue of 1.51, which accounted for30% of the variance across animals. From this primary factor,factor scores were extracted to represent each animal’sgeneral learning ability. Factor scores, which are analogousto an average z score of an animal’s performance across alltasks, provide a sensitive index of an individual’s aggregateperformance in the learning battery. However, the number ofsubjects used here is somewhat small by factor analytic stan-dards. Therefore, to verify the accuracy of these factor scoresas descriptors of general learning performance, the factorscores were compared to each animal’s average rank perfor-mance (relative to others in the sample) across all five learningtasks (a method that has been previously reported [13]). Factorscores and average ranks were significantly correlated [r(25) =0.61, p < 0.001]. Because factor scores are presumed to bea more precise measure of an animal’s aggregate performanceacross all tasks, these scores were used in all subsequentanalyses to compare the performance of groups of animals.
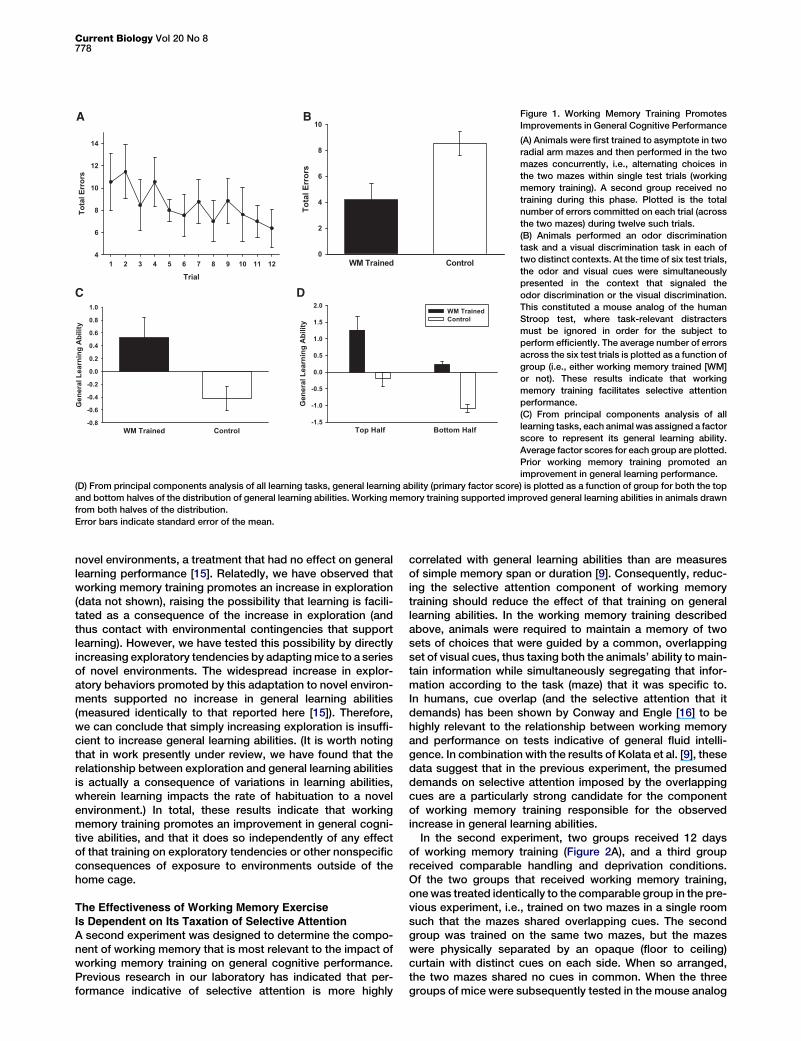
Mice trained on the concurrent radial arm maze task hadsignificantly higher general learning scores than did animalsthat were not subjected to this working memory training [Fig-ure 1C; F(1,25) = 4.48, p < 0.05; Cohen’s d effect size = 0.94].It was possible that working memory training differentiallyimproved general learning scores in a subset of mice (e.g.,mice from different regions of the distribution of generallearning abilities). To assess this, we compared factor scoresfor the top and bottom performers on the learning batteryacross animals that had received or had not received workingmemory training (Figure 1D). The top half of mice that hadundergone working memory training had significantly highergeneral learning abilities than the top half of animals that hadnot received working memory training [F(1,11) = 6.88, p <0.05]. A similar difference was observed in those animalsfrom the bottom halves of the distribution of learning abilities[F(1,11) = 13.92, p < 0.005], suggesting that working memorytraining promotes an improvement in general learning perfor-mance regardless of an animal’s position in the distributionof learning abilities.
In this experiment, animals that underwent working memorytraining exhibited a commensurate increase in their aggregateperformance in a battery of diverse learning tasks. Two alter-native explanations for these results are possible. First, it isconceivable that any manipulation outside of the home cagewould have resulted in an increase in general learning abilitiessimply because it rescues the animals from the environmentaldeprivation of the home cage. However, this result stands incontrast to previous attempts to manipulate general learningabilities of mice through extensive and repeated exposure to
A B
Trial
1 2 3 4 5 6 7 8 9 10 11 12
To
tal E
rro
rs
4
6
8
10
12
14
WM Trained Control
To
tal E
rro
rs
0
2
4
6
8
10
C D
WM Trained Control
Ge
neral L
ea
rn
in
g A
bility
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
Top Half Bottom Half
Gen
era
l L
ea
rn
in
g A
bility
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
WM Trained
Control
Figure 1. Working Memory Training Promotes
Improvements in General Cognitive Performance
(A) Animals were first trained to asymptote in two
radial arm mazes and then performed in the two
mazes concurrently, i.e., alternating choices in
the two mazes within single test trials (working
memory training). A second group received no
training during this phase. Plotted is the total
number of errors committed on each trial (across
the two mazes) during twelve such trials.
(B) Animals performed an odor discrimination
task and a visual discrimination task in each of
two distinct contexts. At the time of six test trials,
the odor and visual cues were simultaneously
presented in the context that signaled the
odor discrimination or the visual discrimination.
This constituted a mouse analog of the human
Stroop test, where task-relevant distracters
must be ignored in order for the subject to
perform efficiently. The average number of errors
across the six test trials is plotted as a function of
group (i.e., either working memory trained [WM]
or not). These results indicate that working
memory training facilitates selective attention
performance.
(C) From principal components analysis of all
learning tasks, each animal was assigned a factor
score to represent its general learning ability.
Average factor scores for each group are plotted.
Prior working memory training promoted an
improvement in general learning performance.
(D) From principal components analysis of all learning tasks, general learning ability (primary factor score) is plotted as a function of group for both the top
and bottom halves of the distribution of general learning abilities. Working memory training supported improved general learning abilities in animals drawn
from both halves of the distribution.
Error bars indicate standard error of the mean.
Current Biology Vol 20 No 8778
novel environments, a treatment that had no effect on generallearning performance [15]. Relatedly, we have observed thatworking memory training promotes an increase in exploration(data not shown), raising the possibility that learning is facili-tated as a consequence of the increase in exploration (andthus contact with environmental contingencies that supportlearning). However, we have tested this possibility by directlyincreasing exploratory tendencies by adapting mice to a seriesof novel environments. The widespread increase in explor-atory behaviors promoted by this adaptation to novel environ-ments supported no increase in general learning abilities(measured identically to that reported here [15]). Therefore,we can conclude that simply increasing exploration is insuffi-cient to increase general learning abilities. (It is worth notingthat in work presently under review, we have found that therelationship between exploration and general learning abilitiesis actually a consequence of variations in learning abilities,wherein learning impacts the rate of habituation to a novelenvironment.) In total, these results indicate that workingmemory training promotes an improvement in general cogni-tive abilities, and that it does so independently of any effectof that training on exploratory tendencies or other nonspecificconsequences of exposure to environments outside of thehome cage.
The Effectiveness of Working Memory Exercise
Is Dependent on Its Taxation of Selective AttentionA second experiment was designed to determine the compo-nent of working memory that is most relevant to the impact ofworking memory training on general cognitive performance.Previous research in our laboratory has indicated that per-formance indicative of selective attention is more highly
correlated with general learning abilities than are measuresof simple memory span or duration [9]. Consequently, reduc-ing the selective attention component of working memorytraining should reduce the effect of that training on generallearning abilities. In the working memory training describedabove, animals were required to maintain a memory of twosets of choices that were guided by a common, overlappingset of visual cues, thus taxing both the animals’ ability to main-tain information while simultaneously segregating that infor-mation according to the task (maze) that it was specific to.In humans, cue overlap (and the selective attention that itdemands) has been shown by Conway and Engle [16] to behighly relevant to the relationship between working memoryand performance on tests indicative of general fluid intelli-gence. In combination with the results of Kolata et al. [9], thesedata suggest that in the previous experiment, the presumeddemands on selective attention imposed by the overlappingcues are a particularly strong candidate for the componentof working memory training responsible for the observedincrease in general learning abilities.
In the second experiment, two groups received 12 daysof working memory training (Figure 2A), and a third groupreceived comparable handling and deprivation conditions.Of the two groups that received working memory training,one was treated identically to the comparable group in the pre-vious experiment, i.e., trained on two mazes in a single roomsuch that the mazes shared overlapping cues. The secondgroup was trained on the same two mazes, but the mazeswere physically separated by an opaque (floor to ceiling)curtain with distinct cues on each side. When so arranged,the two mazes shared no cues in common. When the threegroups of mice were subsequently tested in the mouse analog

A B
Trial
1 2 3 4 5 6 7 8 9 10 11 12
To
tal E
rro
rs
0
5
10
15
20
25
30Overlap
No Overlap
Overlap No Overlap Control
To
ta
l E
rro
rs
0
2
4
6
8
10
12
C D
Overlap No Overlap Control
Ge
ne
ra
l L
ea
rn
in
g A
bility
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
Top Half Bottom Half
Ge
ne
ra
l L
ea
rn
in
g A
bility
-2
-1
0
1
2
3Overlap
No Overlap
Control
Figure 2. Selective Attention Underlies the
Effects of Working Memory Training
(A) Animals were first trained to asymptote in two
radial arm mazes and then performed in the two
mazes concurrently, i.e., alternating choices in
the two mazes within single test trials (working
memory training). One group of animals received
training wherein the two mazes shared overlap-
ping visual cues (which guide the animals’
search), and one group received training on
mazes that were explicitly separated, sharing no
overlapping visual cues. The total number of
errors committed on each trial (across the two
mazes) during twelve such trials is plotted.
A third group received only equivalent handling
but did not perform in the mazes.
(B) Animals performed an odor discrimination
task and a visual discrimination task in each of
two distinct contexts. At the time of six test trials,
the odor and visual cues were simultaneously
presented in the context that signaled the
odor discrimination or the visual discrimination.
This constituted a mouse analog of the human
Stroop test, where task-relevant distracters
must be ignored in order for the animal to perform
efficiently. The total errors across the six test
trials are plotted as a function of group (i.e.,
working memory training with overlapping or no
overlapping visual cues, or no working memory
training). Prior working memory training facilitated selective attention, and that effect was accentuated if that training required animals to segregate infor-
mation related to shared (overlapping) cues.
(C) From principal components analysis of all learning tasks, general learning ability (primary factor score) is plotted as a function of group. Prior working
memory training promoted an improvement in general learning performance, and that effect was accentuated if that training required animals to segregate
information related to shared (overlapping) cues.
(D) From principal components analysis of all learning tasks, general learning ability (primary factor score) is plotted as a function of group for both the top
and bottom halves of the distribution of general learning abilities. Working memory training supported improved general learning abilities in animals drawn
from both halves of the distribution, and that effect was accentuated if that training required animals to segregate information related to shared (overlapping)
cues.
Error bars indicate standard error of the mean.
Working Memory Training and Intelligence779
of the human Stroop test, there was a strong trend towarda main effect of training history [Figure 2B; F(2,24) = 3.31, p =0.054]. Planned comparisons revealed a significant differencein the performance on the Stroop test between mice thatreceived working memory training with overlapping cues andeach of the remaining two groups (p < 0.05) but no differencebetween the animals trained with no overlapping cues andanimals not subject to any working memory training. Theseresults indicate that the added selective attention loadrequired to perform the working memory task when cues over-lapped promoted superior performance on the subsequenttest of selective attention. It is noted that the two forms ofradial arm maze training (with overlapping or nonoverlappingcues) made very different demands on the animals, and it ispossible that the difficulty performing these tasks (ratherthan selective attention demands per se) may promote differ-ences in performance in tests of selective attention. Toaddress whether the two radial arm maze tasks varied in diffi-culty, we performed a factorial analysis of variance on theacquisition data in the black maze for groups trained with over-lapping or nonoverlapping visual cues, and no differencebetween groups was observed [F(1,19) = 0.79]. Moreover, at test was performed to compare the performance of the twogroups on the last trial of this training, and again, no differencewas observed [t(19) = 1.43]. These results suggest that the twotasks were similarly difficult and thus that task difficulty wasnot a principal determinant of the observed improvement inselective attention.
Results from individual tasks in the learning battery are pre-sented in the Supplemental Information. In this experiment,a principal components analysis of learning performanceextracted a single factor with an eigenvalue of 1.73, whichaccounted for 35% of the variance in performance of individualanimals across all tasks. From this analysis, factor scores wereextracted that represented the general learning performance ofindividual animals. As in the previous experiment, we verifiedthe accuracy of these factor scores as descriptors of animals’aggregate performance in the learning battery by assessingthe correlation of these scores with animals’ average rankperformance across all five learning tasks. The two measureswere significantly correlated [r(25) = 0.84, p < 0.0001].
When factor scores from the three treatment conditionswere compared, a main effect of treatment was observed [Fig-ure 2C; F(2,24) = 10.57, p < 0.001, u2 effect size = 0.54]. Posthoc comparisons of factor scores revealed significant differ-ences between the group that received overlapping cuesduring working memory training and the group that receivedno overlapping cues during this training (p < 0.05) and animalsthat received no working memory training (p < 0.001). Lastly,a difference was observed between the group that receivedworking memory training with no overlapping cues and thegroup that received no working memory training (p < 0.05).
Again we assessed whether working memory training differ-entially affected the general learning performance of animalsdrawn from the top or bottom half of the distribution of learningabilities (Figure 2D). In the top half of the distribution, there was

Current Biology Vol 20 No 8780
a main effect of group [F(2,10) = 8.47, p < 0.01]. Plannedcomparisons revealed a significant difference between thegroup that received working memory training with overlappingcues and control animals that received no working memorytraining (p < 0.005). However, trends were seen toward differ-ences between mice that received working memory trainingwith overlapping cues relative to those that received thistraining with no overlapping cues (p = 0.09), as well as betweenmice that received working memory training with no overlap-ping cues and animals that had received no working memorytraining (p = 0.06). In the bottom half of the distribution oflearning abilities, the same pattern of results was observed.There was a significant main effect of group [F(2,10) = 13.97,p < 0.005]. Planned comparisons revealed significant differ-ences in general learning performance between mice thatreceived working memory training with overlapping cues andanimals that received no working memory training (p <0.001), mice that received working memory training with nooverlapping cues and those that did not receive workingmemory training (p < 0.05), and mice that received workingmemory training with overlapping or no overlapping cues(p < 0.05). Thus, working memory training again promoted animprovement in general learning abilities in animals in boththe top and bottom halves of the distribution of abilities.Importantly, working memory training without overlappingcues produced performances intermediate to training withoverlapping cues and no training in both the top and bottomhalves of the distribution.
This experiment provides a second demonstration thatworking memory training facilitates the implementation ofselective attention and promotes a commensurate increasein general learning abilities. When the selective attention loadof the complex radial arm maze task was reduced, the effectof working memory training on general learning abilities wassimilarly attenuated (although not eliminated). This diminishedyet persistent influence of working memory training with a lowselective attention load on general learning abilities can beaccounted for in two ways. First, it is possible that workingmemory training by itself, regardless of selective attentionload, is sufficient to promote an increase in general learningabilities. Consistent with this possibility, variations in workingmemory span have been determined to predict generallearning abilities in mice as well as general intelligence inhumans [10, 17], although they do so to a lesser degree thantests of working memory that more heavily tax the selectiveattention system. Another possibility is that overlapping cueswere simply reduced rather than eliminated, i.e., the animalsmay have used uncontrolled (i.e., undetected) aspects of thetraining context to guide their behavior. From the presentdata, we cannot determine which of these two, if either,accounts for the residual impact of working memory trainingwith a low selective attention load on the aggregate generallearning abilities of mice.
Discussion
The present set of experiments provides evidence that trainingregimens that ‘‘exercise’’ working memory promote anincrease in the efficacy of selective attention and a commensu-rate increase in general learning abilities. We have extendedprevious results obtained with human subjects [12, 18–22] bycomparing a working memory training regimen that specifi-cally taxes the selective attention system to one with a nomi-nally reduced selective attention load and find that the
selective attention load of this training is a critical determinantof the consequent improvement in general cognitive perfor-mance. This provides evidence that training regimens thatheavily tax the selective attention system have a particularlypotent capacity to increase general learning abilities in mice.
In humans, a diverse set of training methods have beenestablished that promote aspects of attention and self-regula-tion [22]. In combination with the parameters established herethat promote selective attention and general learning abilities,the capacity to modulate attention in humans has importantimplications for understanding intelligence and the modulationof this trait in humans. For instance, training programs thatpromote attention have been determined to have modesteffects on children’s performance on tests considered to beindicative of fluid intelligence [20, 21, 23], and this effect isaccentuated in children that express attention deficit disor-ders [18]. Qualitatively similar results have been observed inaged humans [24]. In combination, these results have begunto converge on the conclusion that the capacity for attentioncan be modified through structured training and that thistraining may commensurately impact performance thatreflects variations in intelligence.
One might ask whether the beneficial effects of workingmemory training reflect a consequence of that training orsimply reflect the experientially deprived state of the controlmice that did not receive training outside of the home cage—i.e., would any experience outside of the home cage havesimilar beneficial effects on general learning abilities? Relat-edly, it is possible that the manipulation of any correlate ofgeneral learning ability of mice might similarly impact thoseabilities. To assess these considerations, we have previouslydemonstrated that repeatedly exposing mice to novel environ-ments (for an amount of time comparable to that used here forthe implementation of working memory training) promotes anincrease in the animals’ subsequent exploration of new envi-ronments. However, this exposure to novel environmentsand the resultant increase in exploratory behaviors had noimpact on the animals’ general learning performance [15],despite the fact that native exploratory propensity has beenrepeatedly determined to strongly predict general learningabilities [13, 25, 26].
It is worth noting that in a manuscript currently under review,we report that the expression of a cluster of genes related todopamine signaling in the prefrontal cortex is directly relatedto the aggregate performance of mice in the cognitive testbattery described here, and the dopaminergic processes reg-ulated by these genes have been demonstrated to contributeto the efficacy of working memory [27–29]. It is thus temptingto speculate that working memory training, like that imposedhere, might upregulate the expression of these same genes,providing a potential epigenetic mechanism for the impact ofworking memory training on general cognitive abilities.
It has long been recognized that the processing compo-nents of the working memory system such as working memorycapacity and/or selective attention are strongly correlated withhumans’ performance on tests indicative of intelligence [30].‘‘Higher cognitive functions’’ (such as reasoning, comprehen-sion, and learning) are the hallmark of contemporary intelli-gence test batteries and form common colloquial descriptionsof ‘‘intelligence.’’ Thus, it is not surprising that workingmemory (or at least some of its subcomponents) have cometo be viewed by some as the potential latent factor thatunderlies general (fluid) cognitive abilities, i.e., intelligence[17, 31]. Accordingly, variations in working memory efficacy

Working Memory Training and Intelligence781
have been proposed to regulate individual differences inintelligence. Although theoretically compelling, evidence forthis causal relationship has been difficult to attain [10, 11].The results presented here, wherein we have demonstratedthat working memory training has a direct commensurateinfluence on general learning abilities, provide tentative sup-port for a causal relationship between the process (workingmemory) and the trait (general cognitive ability).
Experimental Procedures
Experiment 1 Subjects
Twenty-nine outbred CD-1 mice were obtained at 55 days of age, an age
typical of previous experiments in our laboratory, and were housed individ-
ually and maintained on ad libitum food and water unless noted otherwise in
a temperature-controlled vivarium on a 12 hr light/dark cycle. The mice were
acclimated to the vivarium for 1 week and were subsequently handled
(removed from the home cage and held by the experimenter for 60 s/day)
for 1 week prior to the start of training. Owing to a scheduling error, three
animals could not complete the experiment, resulting in final group
sizes of 13.
Experiment 1 Design
This experiment was a two-group, between-subjects design. Native explor-
atory tendencies are a good predictor of general learning abilities [13, 26],
and so animals were assigned to one of two groups based on their perfor-
mance on a pretest for propensity for exploration in an elevated plus
maze (see Supplemental Information). Working memory training was admin-
istered to one group, while the other was equivalently handled. Training and
testing were identical to that used previously in our laboratory [15, 25] and
are described in detail in the Supplemental Information. In brief, mice
were trained to asymptote on two distinct radial arm mazes, where the
animals collected food at the end of each of eight arms. The mazes were
located in the same room such that they shared common extramaze
visual cues (patterns of lights, pictures, and architectural details that are
used by the animal to guide its search). After performance in each of the
two mazes stabilized, animals then were tested on each of the two mazes
each day (3 hr intertrial interval) for 3 days (the order of testing in the two
mazes alternated across days). Subsequent to this initial training, the
animals then performed concurrently on both mazes once a day for 12 days
(constituting working memory training). During this training, mice alternated
choices in the two mazes and consequently were required to maintain
a memory of the choices in each maze and to segregate those memories
despite the overlapping extramaze visual cues. Thus, this training taxed
both the maintenance of information as well as working memory capacity
and selective attention.
Four days after working memory training (at 112 days of age), mice began
testing on a battery of five learning tasks (Lashley maze, passive avoidance,
spatial water maze, odor discrimination, and associative fear conditioning),
which has been described previously [13, 15, 26]. One week after comple-
tion of the learning battery, mice were tested for selective attention abilities
in a mouse-adapted version of the Stroop test [14]. In brief, mice were
trained to find food by choosing a correct odor (from three possible) in
one context and a correct visual stimulus (from three possible) in a second
context. In the critical test, mice were tested separately in the odor and
visual contexts; during the test trial, both sets of cues were presented simul-
taneously. To perform efficiently during this test trial, mice must ignore
those cues that are relevant to the context other than the current test
context and, in doing so, discriminate between cues relevant only to perfor-
mance in the current test context (e.g., visual cues should be ignored when
tested in the odor discrimination context). As we describe in detail else-
where [9], although no test is process pure, this test makes no nominal
demands on working memory span or duration but instead requires animals
to focus on task-specific cues while ignoring task-relevant distracters and is
thus conceptually consistent with contemporary descriptions of selective
attention [32, 33].
Experiment 2 Subjects
Thirty mice arrived in our laboratory at 45 days of age and were maintained
and tested as in experiment 1. Owing to a training error, three animals could
not complete the experiment, resulting in final group sizes of nine (working
memory training with overlapping cues), nine (working memory training with
no overlapping cues), and nine (no working memory training).
Experiment 2 Design
As in experiment 1, mice were all assessed in the elevated plus maze at
approximately 80 days of age. They were then assigned to one of three
groups, balanced for exploration in the elevated plus maze. One group
received radial arm maze training as described in experiment 1. A second
group received radial arm maze training as described in experiment 1,
although a floor-to-ceiling opaque curtain with a different set of visual
cues on each side was placed between the two radial arm mazes. This
curtain effectively split the room in half, leaving each maze in what was
essentially a distinct room with distinct extramaze cues. The relevant effect
of this alteration on the training of the second group was to have mice
trained on the same two radial arm mazes used for training the first group,
although in this case, the mazes shared no common cues. Because the
animals trained under these latter conditions need not segregate common
information according to the relevant maze, this training imposed reduced
demands on selective attention. A third group received no working memory
exercise. Mice in this group received equivalent food deprivation and food
reward (delivered in their home cages), as well as handling equivalent to
that administered to the working memory training groups.
Supplemental Information
Supplemental Information includes Supplemental Results and Supple-
mental Experimental Procedures and can be found with this article online
at doi:10.1016/j.cub.2010.02.034.
Acknowledgments
This work was supported by the National Institute on Aging (AG022698) and
the Busch Foundation (to L.D.M.).
Received: November 2, 2009
Revised: February 8, 2010
Accepted: February 9, 2010
Published online: March 25, 2010
References
1. Colom, R., Rebollo, I., Palacios, A., Jaun-Espinosa, M., and Kyllonen,
P.C. (2004). Working memory is (almost) perfectly predicted by g. Intel-
ligence 32, 277–296.
2. Conway, A.R., Cowan, N., Bunting, M., Therriault, D.J., and Scott, R.B.
(2002). A latent variable analysis of working memory capacity, short-
term memory capacity, processing speed, and general fluid intelli-
gence. Intelligence 30, 163–183.
3. Engle, R.W., Tuholski, S.W., Laughlin, W., and Conway, A.R. (1999).
Working memory, short-term memory, and general fluid intelligence:
A latent variable approach. J. Exp. Psychol. Gen. 128, 309–331.
4. Jarrold, C., and Towse, J.N. (2006). Individual differences in working
memory. Neuroscience 139, 39–50.
5. Sub, H.-M., Oberauer, K., Wittman, W.W., Wilhelm, O., and Schulze, R.
(2002). Working memory capacity explains reasoning ability—and
a bit more. Intelligence 30, 261–288.
6. Conway, A.R., Kane, M.J., and Engle, R.W. (2003). Working memory
capacity and its relation to general intelligence. Trends Cogn. Sci. 7,
547–552.
7. Kyllonen, P.C., and Christal, R.E. (1990). Reasoning ability is (little more
than) working memory capacity?! Intelligence 14, 389–433.
8. Kolata, S., Light, K., Townsend, D.A., Hale, G., Grossman, H., and Mat-
zel, L.D. (2005). Variations in working memory capacity predict indi-
vidual differences in general learning abilities among genetically diverse
mice. Neurobiol. Learn. Mem. 84, 241–246.
9. Kolata, S., Matzel, L.D., and Light, K. (2007). Selective attention is the
primary determinant of the relationship between working memory and
general learning abilities. Learn. Mem. 14, 22–28.
10. Matzel, L.D., and Kolata, S. (2010). Selective attention, working memory,
and animal intelligence. Neurosci. Biobehav. Rev. 34, 23–30.
11. Unsworth, N., and Engle, R.W. (2007). On the division of short-term and
working memory: An examination of simple and complex span and their
relation to higher order abilities. Psychol. Bull. 133, 1038–1066.
12. Jaeggi, S.M., Buschkuehl, M., Jonides, J., and Perrig, W.J. (2008).
Improving fluid intelligence with training on working memory. Proc.
Natl. Acad. Sci. USA 105, 6829–6833.

Current Biology Vol 20 No 8782
13. Matzel, L.D., Han, Y.R., Grossman, H., Karnik, M.S., Patel, D., Scott, N.,
Specht, S.M., and Gandhi, C.C. (2003). Individual differences in the
expression of a ‘‘general’’ learning ability in mice. J. Neurosci. 23,
6423–6433.
14. Kolata, S., Light, K., and Matzel, L.D. (2008). Domain-specific and
domain-general learning factors are expressed in genetically heteroge-
neous CD-1 mice. Intelligence 36, 619–629.
15. Light, K.R., Kolata, S., Hale, G., Grossman, H., and Matzel, L.D. (2008).
Up-regulation of exploratory tendencies does not enhance general
learning abilities in juvenile or young-adult outbred mice. Neurobiol.
Learn. Mem. 90, 317–329.
16. Conway, A.R., and Engle, R.W. (1994). Working memory and retrieval:
A resource-dependent inhibition model. J. Exp. Psychol. Gen. 123,
354–373.
17. Unsworth, N., and Engle, R.W. (2006). Simple and complex memory
spans and their relation to fluid abilities: Evidence from list-length
effects. J. Mem. Lang. 54, 68–80.
18. Klingberg, T., Forssberg, H., and Westerberg, H. (2002). Training of
working memory in children with ADHD. J. Clin. Exp. Neuropsychol.
24, 781–791.
19. Olesen, P.J., Westerberg, H., and Klingberg, T. (2004). Increased
prefrontal and parietal activity after training of working memory. Nat.
Neurosci. 7, 75–79.
20. Rueda, M.R., Rothbart, M.K., McCandliss, B.D., Saccomanno, L., and
Posner, M.I. (2005). Training, maturation, and genetic influences on
the development of executive attention. Proc. Natl. Acad. Sci. USA
102, 14931–14936.
21. Diamond, A., Barnett, W.S., Thomas, J., and Munro, S. (2007). Preschool
program improves cognitive control. Science 318, 1387–1388.
22. Tang, Y.Y., and Posner, M.I. (2009). Attention training and attention state
training. Trends Cogn. Sci. 13, 222–227.
23. Karbach, J., and Kray, J. (2009). How useful is executive control
training? Age differences in near and far transfer of task-switching
training. Dev. Sci. 12, 978–990.
24. Basak, C., Boot, W.R., Voss, M.W., and Kramer, A.F. (2008). Can training
in a real-time strategy video game attenuate cognitive decline in older
adults? Psychol. Aging 23, 765–777.
25. Grossman, H., Hale, G., Light, K., Kolata, S., and Matzel, L.D. (2007).
Pharmacological modulation of stress reactivity dissociates its role in
the determination of the relationship of exploration and general cogni-
tive abilities. Behav. Neurosci. 121, 949–964.
26. Matzel, L.D., Townsend, D.A., Grossman, H., Han, Y.R., Hale, G.,
Zappulla, M., Light, K., and Kolata, S. (2006). Exploration in outbred
mice covaries with general learning abilities irrespective of stress reac-
tivity, emotionality, and physical attributes. Neurobiol. Learn. Mem. 86,
228–240.
27. Sawaguchi, T., and Goldman-Rakic, P.S. (1991). D1 dopamine receptors
in prefrontal cortex: Involvement in working memory. Science 251,
947–950.
28. Schultz, W., Apicella, P., Ljungberg, T., Romo, R., and Scarnati, E.
(1993). Reward-related activity in the monkey striatum and substantia
nigra. Prog. Brain Res. 99, 227–235.
29. Thurley, K., Senn, W., and Luscher, H.R. (2008). Dopamine increases the
gain of the input-output response of rat prefrontal pyramidal neurons.
J. Neurophysiol. 99, 2985–2997.
30. Daneman, M., and Carpenter, P.A. (1980). Individual differences in
working memory and reading. J. Verbal Learn. Verbal Behav. 19,
450–466.
31. Mackintosh, N.J. (1998). IQ and Human Intelligence (Oxford: Oxford
University Press).
32. Baddeley, A. (2003). Working memory: Looking back and looking
forward. Nat. Rev. Neurosci. 4, 829–839.
33. Baddeley, A.D., and Logie, R.H. (1999). Working memory: The multiple
component model. In Models of Working Memory: Mechanisms of
Active Maintenance and Executive Control, A. Miyake and S. Priti,
eds. (Cambridge: Cambridge University Press).




















