
Environmental enrichment promotes improved spatial abilities and enhanced dendritic growth in the...
13
Behavioural Brain Research 163 (2005) 78–90 Research report Environmental enrichment promotes improved spatial abilities and enhanced dendritic growth in the rat Maria Giuseppa Leggio a,b,∗ , Laura Mandolesi b,c , Francesca Federico a,d , Francesca Spirito a,b , Benedetta Ricci b , Francesca Gelfo a , Laura Petrosini a,b a Department of Psychology, University of Rome “La Sapienza”, Via dei Marsi 78, 00185 Rome, Italy b IRCCS Santa Lucia Foundation, Rome c University of Naples “Parthenope”, Italy d University of Siena, Italy Received 14 February 2005; received in revised form 12 April 2005; accepted 14 April 2005 Available online 23 May 2005 Abstract An enriched environment consists of a combination of enhanced social relations, physical exercise and interactions with non-social stimuli that leads to behavioral and neuronal modifications. In the present study, we analyzed the behavioral effects of environmental complexity on different facets of spatial function, and we assessed dendritic arborisation and spine density in a cortical area mainly involved in the spatial learning, as the parietal cortex. Wistar rat pups (21 days old) were housed in enriched conditions (10 animals in a large cage with toys and a running wheel), or standard condition (two animals in a standard cage, without objects). At the age of 3 months, both groups were tested in the radial maze task and Morris water maze (MWM). Morphological analyses on layer-III pyramidal neurons of parietal cortex were performed in selected animals belonging to both experimental groups. In the radial maze task, enriched animals exhibited high performance levels, by exploiting procedural competencies and working memory abilities. Furthermore, when the requirements of the context changed, they promptly reorganized their strategies by shifting from prevalently using spatial procedures to applying mnesic competencies. In the Morris water maze, enriched animals more quickly acquired tuned navigational strategies. Environmental enrichment provoked increased dendritic arborisation as well as increased density of dendritic spines in layer-III parietal pyramidal neurons. © 2005 Elsevier B.V. All rights reserved. Keywords: Development; Spatial procedural learning; Spatial working memory; Neuronal changes; Dendritic spines; Parietal cortex 1. Introduction An enriched environment consists of a combination of enhanced social relations, physical exercise and interactions with non-social stimuli. Enriched conditions are known to have a significant impact on memory and learning abili- ties [8,30,46] as well as on neurogenesis and synaptoge- nesis [41,74,84]. In the 1960s, Rosenzweig, Diamond and co-workers pioneered studies on the mechanisms by which complex environment and experience influence brain func- tion, demonstrating significant changes in brain anatomy ∗ Corresponding author. Tel.: +39 0649917522; fax: +39 0649917522. E-mail address: [email protected] (M.G. Leggio). [19,20,45,46,65]. More recent studies have established that these structural and behavioral changes are associated with changes in brain neurochemistry and physiology. Several as- pects of hippocampal functioning, such as LTP, neurogenesis, dendritic spine growth and neurotrophin mRNA expression are reported to be enhanced by environmental enrichment [22,25,27,40,41,74,80]. Also, in the neocortical regions, en- richment induces clear neural alterations such as increase in thickness, dendritic branching, presynaptic vesicle number and synaptophysin levels [18,28,33,51,78]. However, in spite of the copious literature on environ- mental complexity effects, whether the beneficial effects of enrichment influence all aspects of learning, or are linked to particular components of learning abilities has not been 0166-4328/$ – see front matter © 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.bbr.2005.04.009
Transcript of Environmental enrichment promotes improved spatial abilities and enhanced dendritic growth in the...

Behavioural Brain Research 163 (2005) 78–90
Research report
Environmental enrichment promotes improved spatial abilities andenhanced dendritic growth in the rat
Maria Giuseppa Leggioa,b,∗, Laura Mandolesib,c, Francesca Federicoa,d, Francesca Spiritoa,b,Benedetta Riccib, Francesca Gelfoa, Laura Petrosinia,b
a Department of Psychology, University of Rome “La Sapienza”, Via dei Marsi 78, 00185 Rome, Italyb IRCCS Santa Lucia Foundation, Rome
c University of Naples “Parthenope”, Italyd University of Siena, Italy
Received 14 February 2005; received in revised form 12 April 2005; accepted 14 April 2005Available online 23 May 2005
Abstract
cial stimulit mplexity ond the spatiall toys and ar ested in ther erformedi e levels, bye y promptlyr ater maze,e rborisationa©
K
1
ewhtncct
thatd withal as-esis,
sionmentn-se inber
on-ts ofkedeen
0d
An enriched environment consists of a combination of enhanced social relations, physical exercise and interactions with non-sohat leads to behavioral and neuronal modifications. In the present study, we analyzed the behavioral effects of environmental coifferent facets of spatial function, and we assessed dendritic arborisation and spine density in a cortical area mainly involved in
earning, as the parietal cortex. Wistar rat pups (21 days old) were housed in enriched conditions (10 animals in a large cage withunning wheel), or standard condition (two animals in a standard cage, without objects). At the age of 3 months, both groups were tadial maze task and Morris water maze (MWM). Morphological analyses on layer-III pyramidal neurons of parietal cortex were pn selected animals belonging to both experimental groups. In the radial maze task, enriched animals exhibited high performancxploiting procedural competencies and working memory abilities. Furthermore, when the requirements of the context changed, theeorganized their strategies by shifting from prevalently using spatial procedures to applying mnesic competencies. In the Morris wnriched animals more quickly acquired tuned navigational strategies. Environmental enrichment provoked increased dendritic as well as increased density of dendritic spines in layer-III parietal pyramidal neurons.2005 Elsevier B.V. All rights reserved.
eywords:Development; Spatial procedural learning; Spatial working memory; Neuronal changes; Dendritic spines; Parietal cortex
. Introduction
An enriched environment consists of a combination ofnhanced social relations, physical exercise and interactionsith non-social stimuli. Enriched conditions are known toave a significant impact on memory and learning abili-
ies [8,30,46] as well as on neurogenesis and synaptoge-esis[41,74,84]. In the 1960s, Rosenzweig, Diamond ando-workers pioneered studies on the mechanisms by whichomplex environment and experience influence brain func-ion, demonstrating significant changes in brain anatomy
∗ Corresponding author. Tel.: +39 0649917522; fax: +39 0649917522.E-mail address:[email protected] (M.G. Leggio).
[19,20,45,46,65]. More recent studies have establishedthese structural and behavioral changes are associatechanges in brain neurochemistry and physiology. Severpects of hippocampal functioning, such as LTP, neurogendendritic spine growth and neurotrophin mRNA expresare reported to be enhanced by environmental enrich[22,25,27,40,41,74,80]. Also, in the neocortical regions, erichment induces clear neural alterations such as increathickness, dendritic branching, presynaptic vesicle numand synaptophysin levels[18,28,33,51,78].
However, in spite of the copious literature on envirmental complexity effects, whether the beneficial effecenrichment influence all aspects of learning, or are linto particular components of learning abilities has not b
166-4328/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2005.04.009

M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90 79
addressed. In adult rodents, environmental enrichment ini-tiated at almost any point in the lifespan seems to im-prove performances on all kinds of tasks, i.e. spatial ornon-spatial, associative or discriminative, mnesic or proce-dural [9,52,64,72,76,78]. This non-specific pattern of be-havioral improvement probably emerges because analysesfail to split complex functions into single components.Thus, it seemed interesting to analyze the effect of envi-ronmental enrichment on specific facets of cognitive perfor-mance.
In memory and learning abilities, spatial functions are offundamental importance to mobile organisms whose spatialmemories form and are organized in distinct but coordinatedframes, thus allowing well structured adaptive capacities.Since enriched-reared animals display superior performanceson some tasks tapping spatial functioning[28,73,75,80], itseems particularly important to identify the components ofspatial behavior that are enhanced most by enriched rearing.To measure spatial working memory abilities in rats, themost suitable task appears to be the radial maze. In fact, theuse of different radial maze protocols allows distinguishingspatial working memory abilities from procedural strategies.Furthermore, Morris water maze (MWM) allows separatelyassessing (localizatory) declarative memory from (explo-rative) procedural strategies[7,49]. These different facetsof spatial functioning are related to the activity of variousn h aspt -c[ ionsc icatet n ofs ayb tion,s ntialp e ofe ures[ etala ly in-t sity int dritica dentm ed pseso erredf sity[
ectso riousc logyo ouldi rilya use-f ngesw
2. Materials and methods
2.1. Animals
Initially, 40 Wistar rats were used in the experiment. Useful datawere collected on 36 rats; the remaining four animals were discardedbecause their behavior froze in the very first sessions of the radialmaze task. On the 21st postnatal day, an even number of littermatesof the same dam were randomly assigned to one of the two ex-perimental groups. The first group (N= 17) was reared in enrichedconditions (EC), and the other (N= 19) in standard conditions (SC).Animals were kept in the respective conditions until behavioral ex-periments were carried out 2.5/3 months later and also throughoutthe whole behavioral testing. Both groups of animals received thesame type of food. Food and water were provided ad libitum untilbehavioral testing.
2.2. Rearing conditions
The EC group rats were housed in groups of 10 in a large cage(100 cm× 50 cm× 70 cm), with one extra level constructed of gal-vanized wire mesh and connected by ramps of the same materialto create two interconnected levels. The cage contained wood shav-ings, a running wheel, a shelter (a house-shaped toy with a concaveopening), plastic coloured toys and small constructions. Through-out the enrichment period, the shelter and running wheel were keptin the cage, while the toys and constructions were changed once aw werem xplo-r ndledd , i.e.t
cage( cts.F sitiont l careb ma-n ringa ntact.
2
form(l falls,e d ofe mazew ll-litr cuesh
2con-
d Priort e theirw fied.I in ad tting.
bit-u atform
euronal structures, namely, neocortical regions, sucrefrontal[16,26,62,81]and parietal[4,12,21,43,61,68]cor-
ices, hippocampal circuits[10,48,49,54,55,58,67]and subortical areas, such as basal ganglia[56,83]and cerebellum15,29,59,66]. Despite clear instances where these regan operate independently of each other, other data indhat they can interact during new learning and retentiopatial information[42]. Thus, more than one structure me involved in processing single attributes of spatial funcuggesting the possibility for both parallel and sequerocessing. Taking into account the marked influencnvironmental enrichment on the neocortical struct
33,35,74]and the heavy involvement of associative parireas in different spatial aspects, it seemed particular
eresting to assess dendritic arborisation and spine denhe parietal cortex. Earlier studies have proven that denlterations are a sensitive index of experience-depenodifications of brain structures[44,74]. Because thendritic surface receives more than 95% of the synan a neuron, changes in synapse activity can be inf
rom analysis of dendritic arborisation and spine den35].
In conclusion, our specific aim was to compare the efff enriched versus standard rearing conditions on vaomponents of spatial functioning and on the morphof the parietal neurons. Differences between groups w
ndicate which components of spatial learning are primaffected by developmental manipulations, and provide a
ul behavioral and anatomical model to study plastic chaithin normal or injured brain.
eek. Also once a week, the feeding boxes and water bottlesoved to different cage points to encourage foraging and e
ative behaviors. Furthermore, each enriched animal was haaily for at least 10 min and allowed to forage in a large space
he whole lab room.The SC group rats were housed by twos in a standard
40 cm× 26 cm× 18 cm) containing wood shavings but no objeeeding boxes and water bottles were kept in the same po
hroughout the animals’ life. These animals received the usuay the stabulary staff, even if without particular and prolongedipulation. This procedure did not result in an impoverished reand also standard animals were accustomed to the “human” co
.3. Radial maze
The eight-arm radial maze consisted of a round central plat30 cm in diameter) from which eight arms (12.5 cm wide× 60 cmong) radiated like the spokes of a wheel. To avoid accidentalach arm was bordered by a clear rail (3 cm high). At the enach arm, there was a food well (5 cm diameter). The entireas white and stood 50 cm off the floor. It was located in a we
oom with one door and was surrounded by some extra-mazeeld in constant spatial relations throughout the experiment.
.3.1. Behavioral procedureFollowing the period of exposure to their respective housing
itions, both groups of animals were tested in the radial maze.o the habituation phase, rats were food-restricted to decreaseight 20–25%. However, housing conditions were not modi
n this phase, both groups of rats were gently handled for 10 may to habituate them to the researchers and experimental se
Following the initial food restriction and handling, a 2-day haation phase started. The rat was placed on the central maze pl

80 M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90
and allowed to explore the maze freely for 10 min and to eat the feeduniformly scattered throughout the maze. According to the proce-dure chosen, the arms were totally or partially baited with a piece ofpurina chow. The rat was placed on the central platform and allowedto make the runs or to explore the maze for 15 min.
2.3.2. Behavioral testingFull-baited maze procedure. All maze arms were baited with a
piece of purina chow at each session. After the habituation phase, therat was placed on the central platform and allowed to make 16 runsor to explore the maze for 15 min. The animal made an error when itcompletely entered an arm it had already visited in the same session.The animals underwent two sessions a day for five consecutive days.The inter-session interval was at least 3 h.
2.3.2.1. Forced-choice procedure.On the first day of testing, fourarms (for example, arms 1, 3, 4 and 7) were baited and the remainingarms were closed by a little door at their proximal end. The baitedarms were separated by different angles to prevent the animal fromreaching the solution by adopting a stereotyped pattern. The rat wasplaced on the central platform and allowed to explore all the openarms. Then, it spent 60 s in its cage before it was put back in themaze. In the second phase, free access to all arms was allowed, butonly the four previously closed arms were baited. This paradigm wasrepeated for five consecutive days with a different configuration ofarms closed each day. In this way, no fixed search pattern could beused.
2e full-
b pectst tial,o
timea isite nt-a isiteda s then pan,cd were:4 e ratsm
ces int rmsv aiteda sub-t at hadb ed asr
2
ular,p in an onl wasm whichw ents.T byt in
diameter) was placed in the middle of one cardinal quadrant (north-west, northeast, southwest, southeast), 30 cm from the side walls; itwas either submerged 2 cm below or elevated 2 cm above the waterlevel. Each rat was gently released into the water from pseudoran-domly varied starting points, so that it faced the center of the pool.The rat was allowed to swim around to find the platform. Blocks offour trials were presented to each rat, two blocks of trials per day.On reaching the platform, each rat was allowed to remain there for30 s before it was placed in the water again for the next trial. If arat failed to locate the platform within 120 s, it was guided there bythe experimenter and allowed to stay there for 30 s. In the first foursessions (Trials 1–16), the platform was hidden in the northwestpool quadrant (Place 1); in the next two sessions (Trials 17–24), theplatform was kept visible in the northeast quadrant (cue phase); inthe final four sessions (Trials 25–40), the platform was hidden inthe northeast quadrant (Place 2)[49]. The rats’ trajectories in thepool were monitored by a video camera mounted on the ceiling. Theresulting video signal was relayed to a monitor, allowing both on-and off-line analyses, and to an image analyzer (Ethovision, Noldus,Wageningen, The Netherlands). Thexandy coordinates of the rat’sposition were sampled and stored on disk.
2.4.1. Behavioral parametersParameters taken into account in analysing the MWM perfor-
mances were successes in finding the platform, latencies to findthe platform and swimming trajectories. Spatial and temporal dis-tribution of swimming trajectories, path length, swimming speed,percentage of time spent in inner or outer annuli, headings (devi-a ge oft pointc be-h g att ad-r rching( reasa nts,n di-r ol).
2
peri-m animalt imals( gi-c oft lem liza-t malli s to-p ctiona mitst . Thea g/kg)a pari-e thes Wat-s neB ma,S esr
.3.3. Behavioral parametersParameters used to assess the animals’ performances in th
aited maze procedure permitted emphasizing different task ashat were more closely, but not exclusively, linked to motor, spar procedural components.
Themotorparameters were: number of arms visited in thellowed (15 min), either right or wrong; total time needed to vight arms, either right or wrong.Spatialparameters were: percege of total errors, calculated as the percentage of wrongly vrms divided by the number of entries; first error, calculated aumber of arms visited including the first wrong visit; spatial salculated as the longest sequence of correctly visited arms.Proce-ural parameters measuring the structure of choice behavior5◦ angles, calculated as the percentage of that angle that thade in each session divided by the number of angles made.Parameters taken into account in analysing the performan
he forced-choice procedure were the following: number of aisited; working memory errors, considered as entries into unbrms. This parameter was further broken down into two error
ypes: across-phase errors, defined as entries into an arm theen entered during the first phase; within-phase errors, define-entries into an arm visited earlier in the same session.
.4. Morris water maze
The rats in both experimental groups were placed in a circlastic pool (diameter 140 cm) with white inside walls, locatedormally equipped laboratory room and uniformly lit by four ne
amps (40 W each) suspended 3 m from the ceiling. No effortade to enhance (or, vice versa, to diminish) extra-maze cues,ere held in constant spatial relations throughout the experimhe pool was filled with water (24◦C), 50 cm deep, made opaque
he addition of 2 l of milk. A white steel escape platform (10 cm
,
tion between the rat’s actual direction when leaving the edhe tank and a straight line from the start location to the tankontaining the platform) were considered to divide explorationavior into five main categories: circling (peripheral swimmin
he tank wall); extended searching (swimming in all pool quants, visiting the same areas more than once); restricted seaswimming in some pool quadrants but not visiting other tank at all); restricted circling (swimming in some peripheral quadraot visiting central tank areas at all); direct finding (swimmingectly toward the platform without any exploration around the po
.5. Analysis of neuronal morphology
Since behavioral outcomes of single specimens of both exental groups were rather homogeneous and there was no
hat performed at odds with the others, randomly selected anEC: N= 3; SC:N= 3) were chosen for the neuronal morpholoal analysis. To this aim, we used the in vivo Golgi-like fillinghe neurons described by Jiang et al.[36]. This simple and reliabethod allows high efficiency of labeling and a complete visua
ion of dendritic arborizations. Furthermore, by requiring very snjection sites and affording a good spatial resolution, it allowographic analyses of the neuronal circuits related to the injerea. Unlike the classical Golgi method, this in vivo method per
he characterization of all components of a selected projectionnimals were anesthetized with a solution of ketamine (90 mnd xylazine (15 mg/kg) i.p. administered. They received atal cortical injection 1 mm below the pial surface, according totereotaxic co-ordinates derived from the atlas of Paxinos andon [57] (ML −5, AP −1.5) of 10% biotinylated dextran amiDA (Molecular Probes, Eugene, OR) and 10 mM NMDA (Sigt. Louis, MO) in 0.01 M phosphate buffer (total injected volum
anging from 0.1 to 0.2�l).

M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90 81
Fig. 1. Golgi-like labeling of rat primary somatosensory cortex showing tracer injection. (A) Dark column marks BDA + NMDA injection site in area SI; (B)a higher magnification of the (A) insert. Scale bars: 1 mm.
After a survival time of 72 h, the animals were anesthetized andtranscardially perfused with phosphate buffered saline followed by4% paraformaldehyde. After removal, the brains were cryoprotectedin 30% buffered sucrose and cut on a freezing microtome into coro-nal sections of 50�m as proposed in the original report by Jianget al.[36]. Most of the BDA labeled neurons were evidenced usingthe avidin-biotin complex. Some sections were counterstained withthionin to visualize the cytoarchitectonic features of the cortex. Theinjection area centered in primary somatosensory cortex showed ev-idence of necrosis, probably due to the excitotoxic action of NMDA(Fig. 1). Most labeled cells were found in the cortex surroundingthe experimental lesion where no signs of excitotoxicity were noted.The radial distribution of labelled neurons was comparable in con-trol and enriched animals. In both groups the labelled parietal neu-rons chosen for the morphological analysis were located in layer IIIof parietal cortex from AP−2.8 to−3.8, areas corresponding to thesecondary somatosensory and posterior parietal cortices.
A researcher unaware of specimen identity made the morpholog-ical analysis with the aid of the Neurolucida software (Microbright-field, Colchester, VT). This system allows computer-assisted recon-struction of the dendritic arbor, thus providing accurate measure-ment of dendritic arborisation. Neurons were selected when theirlabeling was uniform and extending into the most distal branchesof apical and basal dendrites, where spines were clearly marked.Furthermore, the predominant plan of the dendritic arbors of theseneurons should be parallel to the plan of the section[36].
According to these criteria, in layer III of parietal cortex, 10 neu-rons were selected from the enriched animals and 10 neurons fromt neu-r ndardg C15,t d sep-an ntrics ascs ell. A
shell was the volume contained out to a given radius. Total numberof branches was calculated by summing the dendritic nodes startingfrom the cell body.
Spine number was calculated by summing all the spines fromthe initial protrusion of apical or basal dendrites. Spine density wascounted as the number of spines per segments of 10�m length alongthe entire dendritic extension. All protrusions of the dendritic mem-brane were considered in our study, regardless of their shape oractual function.
2.6. Statistical analysis
Metric unit results of animals belonging to the two experimentalgroups were first tested for homoscedasticity of variance and thencompared using one-way or two-way “p× q” analyses of variance(ANOVAs) with repeated measures, followed by multiple compar-isons using Duncan’s tests.
For morphological analyses, to take into account the correlationof data within the same animal, a nested-design ANOVA was ap-plied. Accordingly, the main factor was Group (between-subjectswith two levels), while Animal was a nested factor within Group.Since not-equal number of neurons (n) was examined in each animaland the degrees of freedom of error for each animal aren− 1, thetotal degrees of freedom for the mean square error (denominator forthe computation ofF-statistic) resulted 14.
3. Results
3
otaltp mostl d inF r of
he standard housed animals (enriched group: rat EC1, threeons; rat EC6, four neurons; rat EC12, three neurons and staroup: rat SC4, four neurons; rat SC11, three neurons; rat S
hree neurons). Apical and basal dendritic trees were examinerately by means of Sholl Analysis of sphere intersection[71]. Theumber of intersections of dendrites with a series of concepheres at 10�m intervals from the center of the cell body wounted for each neuron. Total dendritic length (in�m) was theum of the lengths of all processes passing through each sh
.1. Radial maze: full-baited procedure
SC group performed worse than the EC group for time and number of arms visited (Fig. 2A). The EC grouperformed better than SC group even on the parameters
inked to the computation of spatial factors, as displayeig. 2B. In particular, the EC group made a lower numbe
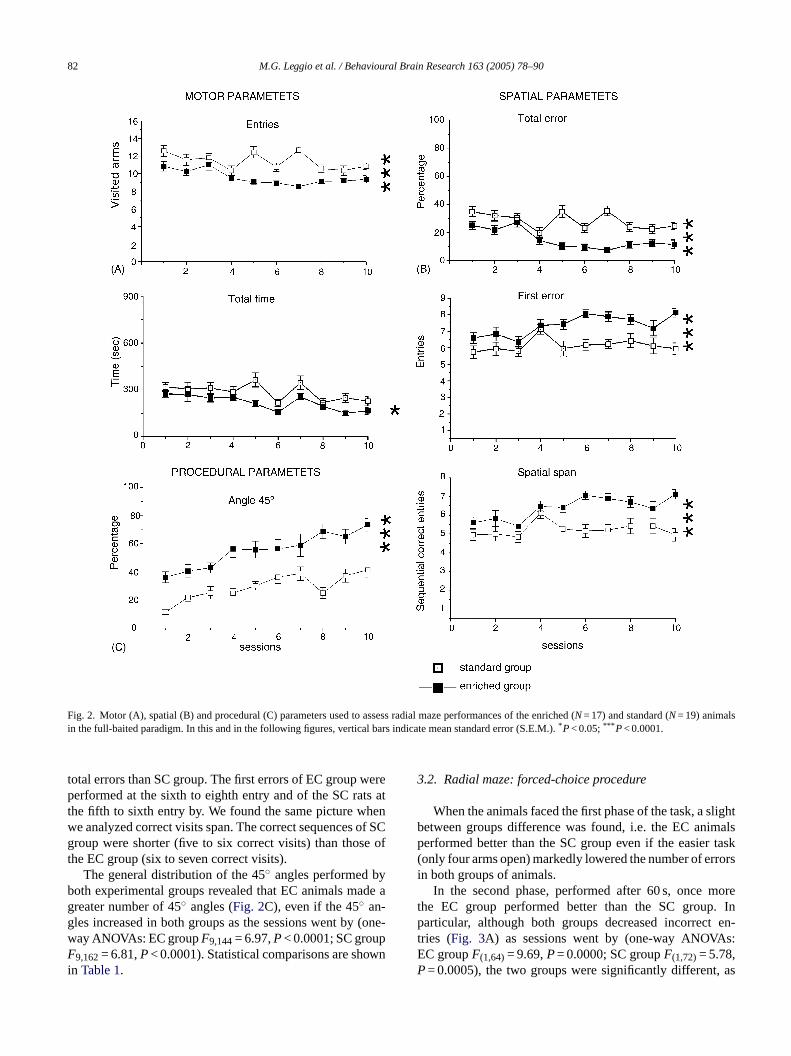
82 M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90
Fig. 2. Motor (A), spatial (B) and procedural (C) parameters used to assess radial maze performances of the enriched (N= 17) and standard (N= 19) animalsin the full-baited paradigm. In this and in the following figures, vertical bars indicate mean standard error (S.E.M.).*P< 0.05;*** P< 0.0001.
total errors than SC group. The first errors of EC group wereperformed at the sixth to eighth entry and of the SC rats atthe fifth to sixth entry by. We found the same picture whenwe analyzed correct visits span. The correct sequences of SCgroup were shorter (five to six correct visits) than those ofthe EC group (six to seven correct visits).
The general distribution of the 45◦ angles performed byboth experimental groups revealed that EC animals made agreater number of 45◦ angles (Fig. 2C), even if the 45◦ an-gles increased in both groups as the sessions went by (one-way ANOVAs: EC groupF9,144= 6.97,P< 0.0001; SC groupF9,162= 6.81,P< 0.0001). Statistical comparisons are shownin Table 1.
3.2. Radial maze: forced-choice procedure
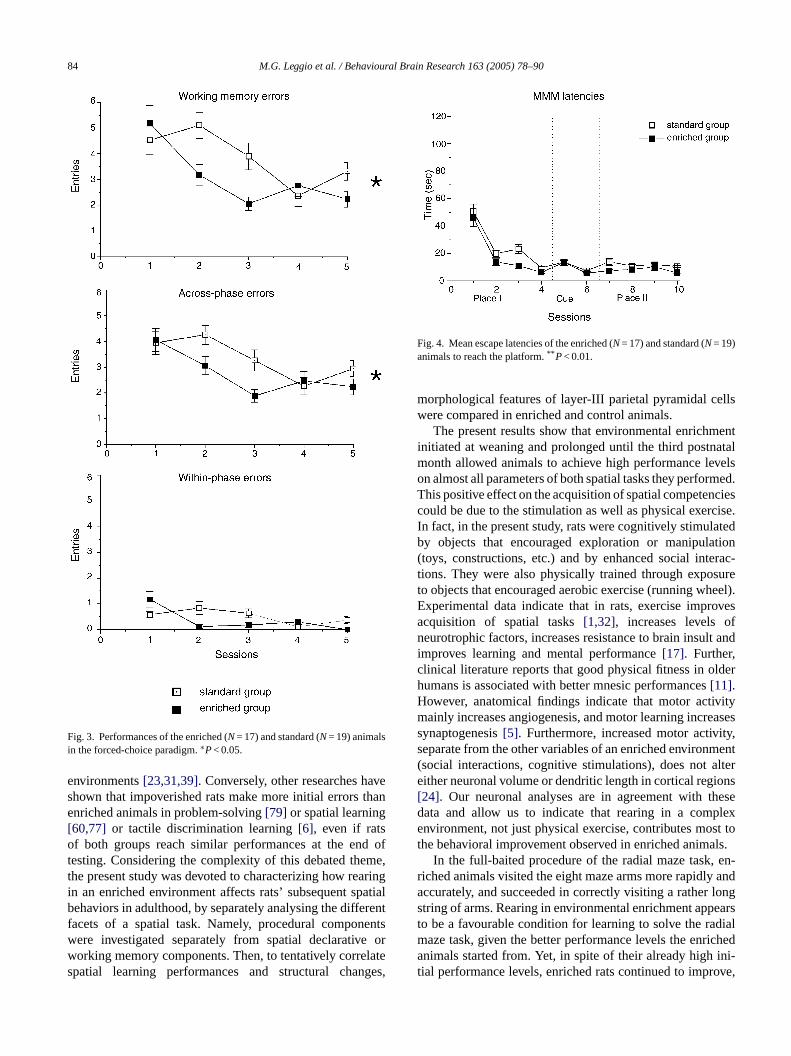
When the animals faced the first phase of the task, a slightbetween groups difference was found, i.e. the EC animalsperformed better than the SC group even if the easier task(only four arms open) markedly lowered the number of errorsin both groups of animals.
In the second phase, performed after 60 s, once morethe EC group performed better than the SC group. Inparticular, although both groups decreased incorrect en-tries (Fig. 3A) as sessions went by (one-way ANOVAs:EC groupF(1,64)= 9.69,P= 0.0000; SC groupF(1,72)= 5.78,P= 0.0005), the two groups were significantly different, as
M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90 83
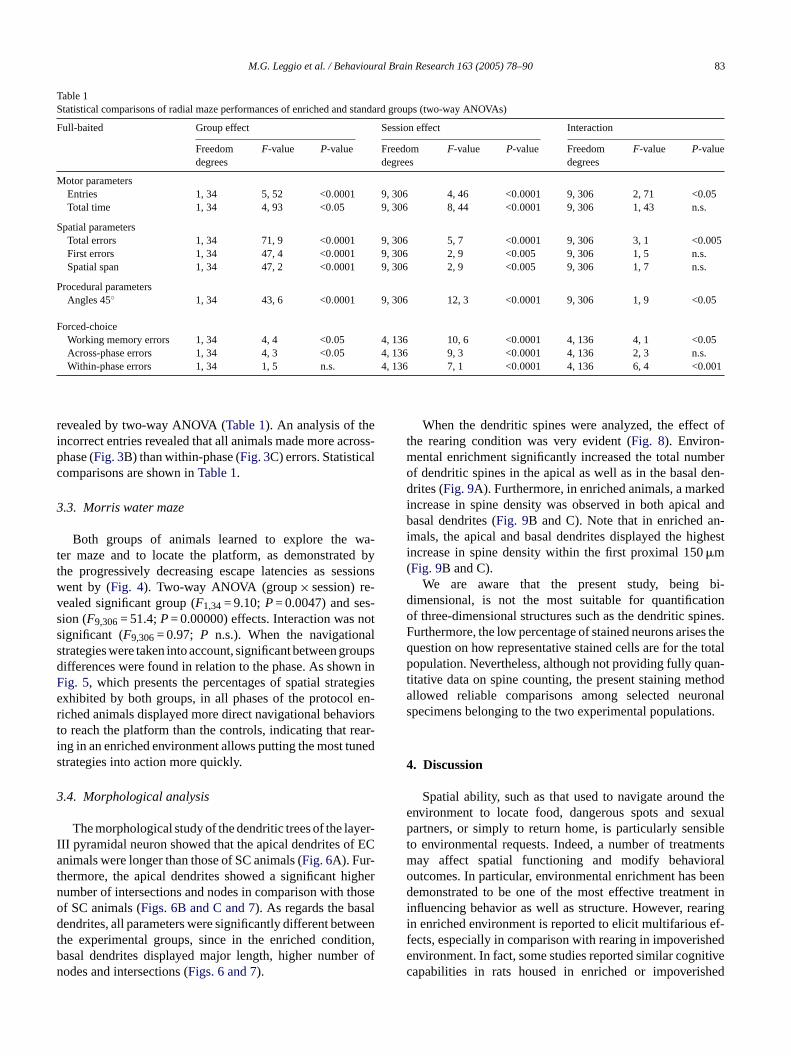
Table 1Statistical comparisons of radial maze performances of enriched and standard groups (two-way ANOVAs)
Full-baited Group effect Session effect Interaction
Freedomdegrees
F-value P-value Freedomdegrees
F-value P-value Freedomdegrees
F-value P-value
Motor parametersEntries 1, 34 5, 52 <0.0001 9, 306 4, 46 <0.0001 9, 306 2, 71 <0.05Total time 1, 34 4, 93 <0.05 9, 306 8, 44 <0.0001 9, 306 1, 43 n.s.
Spatial parametersTotal errors 1, 34 71, 9 <0.0001 9, 306 5, 7 <0.0001 9, 306 3, 1 <0.005First errors 1, 34 47, 4 <0.0001 9, 306 2, 9 <0.005 9, 306 1, 5 n.s.Spatial span 1, 34 47, 2 <0.0001 9, 306 2, 9 <0.005 9, 306 1, 7 n.s.
Procedural parametersAngles 45◦ 1, 34 43, 6 <0.0001 9, 306 12, 3 <0.0001 9, 306 1, 9 <0.05
Forced-choiceWorking memory errors 1, 34 4, 4 <0.05 4, 136 10, 6 <0.0001 4, 136 4, 1 <0.05Across-phase errors 1, 34 4, 3 <0.05 4, 136 9, 3 <0.0001 4, 136 2, 3 n.s.Within-phase errors 1, 34 1, 5 n.s. 4, 136 7, 1 <0.0001 4, 136 6, 4 <0.001
revealed by two-way ANOVA (Table 1). An analysis of theincorrect entries revealed that all animals made more across-phase (Fig. 3B) than within-phase (Fig. 3C) errors. Statisticalcomparisons are shown inTable 1.
3.3. Morris water maze
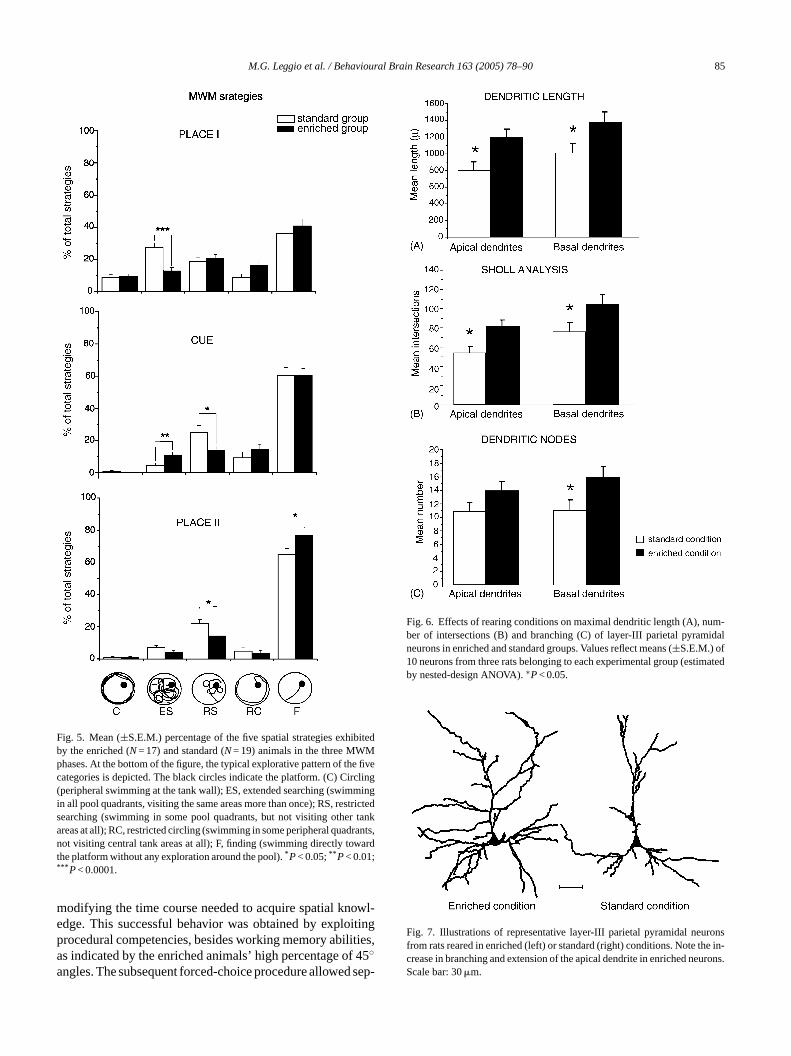
Both groups of animals learned to explore the wa-ter maze and to locate the platform, as demonstrated bythe progressively decreasing escape latencies as sessionswent by (Fig. 4). Two-way ANOVA (group× session) re-vealed significant group (F1,34= 9.10;P= 0.0047) and ses-sion (F9,306= 51.4;P= 0.00000) effects. Interaction was notsignificant (F9,306= 0.97; P n.s.). When the navigationalstrategies were taken into account, significant between groupsdifferences were found in relation to the phase. As shown inFig. 5, which presents the percentages of spatial strategiesexhibited by both groups, in all phases of the protocol en-riched animals displayed more direct navigational behaviorsto reach the platform than the controls, indicating that rear-ing in an enriched environment allows putting the most tunedstrategies into action more quickly.
3.4. Morphological analysis
yer-I f ECat ighern hoseo ald eent tion,b r ofn
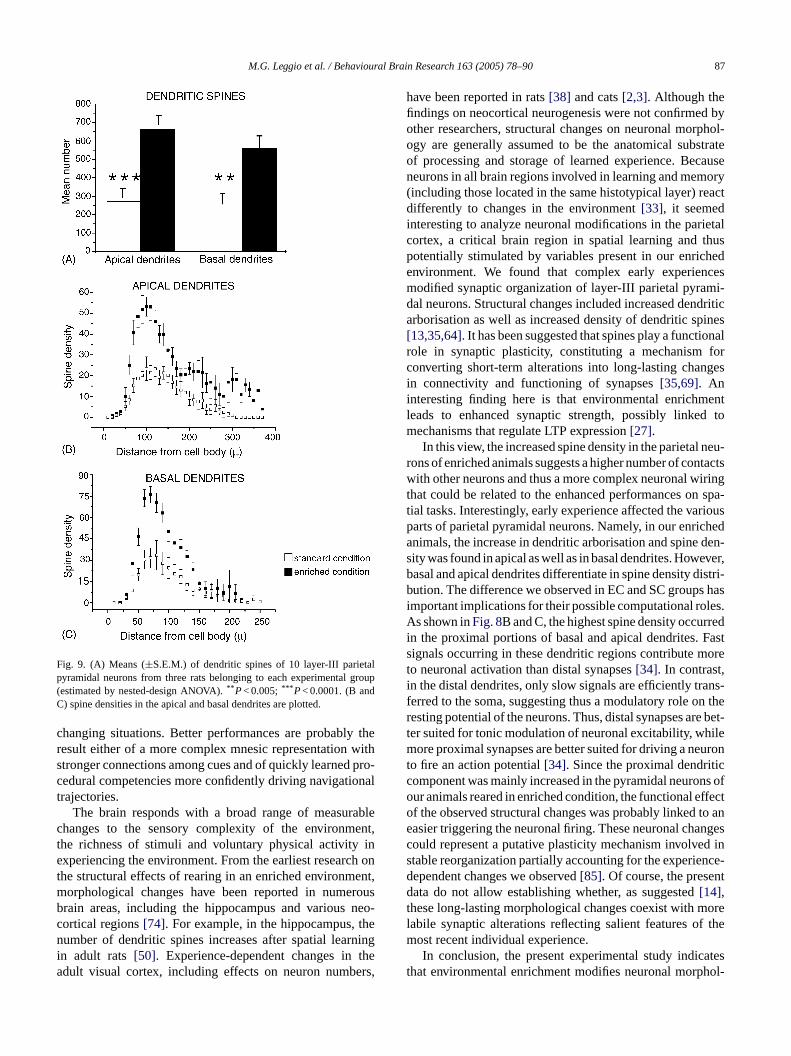
When the dendritic spines were analyzed, the effect ofthe rearing condition was very evident (Fig. 8). Environ-mental enrichment significantly increased the total numberof dendritic spines in the apical as well as in the basal den-drites (Fig. 9A). Furthermore, in enriched animals, a markedincrease in spine density was observed in both apical andbasal dendrites (Fig. 9B and C). Note that in enriched an-imals, the apical and basal dendrites displayed the highestincrease in spine density within the first proximal 150�m(Fig. 9B and C).
We are aware that the present study, being bi-dimensional, is not the most suitable for quantificationof three-dimensional structures such as the dendritic spines.Furthermore, the low percentage of stained neurons arises thequestion on how representative stained cells are for the totalpopulation. Nevertheless, although not providing fully quan-titative data on spine counting, the present staining methodallowed reliable comparisons among selected neuronalspecimens belonging to the two experimental populations.
4. Discussion
Spatial ability, such as that used to navigate around theenvironment to locate food, dangerous spots and sexualp iblet entsm ralo beend nt ini ringi ef-f hede itivec hed
The morphological study of the dendritic trees of the laII pyramidal neuron showed that the apical dendrites onimals were longer than those of SC animals (Fig. 6A). Fur-
hermore, the apical dendrites showed a significant humber of intersections and nodes in comparison with tf SC animals (Figs. 6B and C and 7). As regards the basendrites, all parameters were significantly different betw
he experimental groups, since in the enriched condiasal dendrites displayed major length, higher numbeodes and intersections (Figs. 6 and 7).
artners, or simply to return home, is particularly senso environmental requests. Indeed, a number of treatmay affect spatial functioning and modify behavioutcomes. In particular, environmental enrichment hasemonstrated to be one of the most effective treatme
nfluencing behavior as well as structure. However, rean enriched environment is reported to elicit multifariousects, especially in comparison with rearing in impoverisnvironment. In fact, some studies reported similar cognapabilities in rats housed in enriched or impoveris
84 M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90
Fig. 3. Performances of the enriched (N= 17) and standard (N= 19) animalsin the forced-choice paradigm.∗P< 0.05.
environments[23,31,39]. Conversely, other researches haveshown that impoverished rats make more initial errors thanenriched animals in problem-solving[79] or spatial learning[60,77] or tactile discrimination learning[6], even if ratsof both groups reach similar performances at the end oftesting. Considering the complexity of this debated theme,the present study was devoted to characterizing how rearingin an enriched environment affects rats’ subsequent spatialbehaviors in adulthood, by separately analysing the differentfacets of a spatial task. Namely, procedural componentswere investigated separately from spatial declarative orworking memory components. Then, to tentatively correlatespatial learning performances and structural changes,
Fig. 4. Mean escape latencies of the enriched (N= 17) and standard (N= 19)animals to reach the platform.** P< 0.01.
morphological features of layer-III parietal pyramidal cellswere compared in enriched and control animals.
The present results show that environmental enrichmentinitiated at weaning and prolonged until the third postnatalmonth allowed animals to achieve high performance levelson almost all parameters of both spatial tasks they performed.This positive effect on the acquisition of spatial competenciescould be due to the stimulation as well as physical exercise.In fact, in the present study, rats were cognitively stimulatedby objects that encouraged exploration or manipulation(toys, constructions, etc.) and by enhanced social interac-tions. They were also physically trained through exposureto objects that encouraged aerobic exercise (running wheel).Experimental data indicate that in rats, exercise improvesacquisition of spatial tasks[1,32], increases levels ofneurotrophic factors, increases resistance to brain insult andimproves learning and mental performance[17]. Further,clinical literature reports that good physical fitness in olderhumans is associated with better mnesic performances[11].However, anatomical findings indicate that motor activitymainly increases angiogenesis, and motor learning increasessynaptogenesis[5]. Furthermore, increased motor activity,separate from the other variables of an enriched environment(social interactions, cognitive stimulations), does not altereither neuronal volume or dendritic length in cortical regions[24]. Our neuronal analyses are in agreement with thesed plexe st tot als.
en-r anda longs earst dialm icheda ini-t ove,
ata and allow us to indicate that rearing in a comnvironment, not just physical exercise, contributes mo
he behavioral improvement observed in enriched animIn the full-baited procedure of the radial maze task,
iched animals visited the eight maze arms more rapidlyccurately, and succeeded in correctly visiting a rathertring of arms. Rearing in environmental enrichment appo be a favourable condition for learning to solve the raaze task, given the better performance levels the enrnimals started from. Yet, in spite of their already high
ial performance levels, enriched rats continued to impr
M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90 85
Fig. 5. Mean (±S.E.M.) percentage of the five spatial strategies exhibitedby the enriched (N= 17) and standard (N= 19) animals in the three MWMphases. At the bottom of the figure, the typical explorative pattern of the fivecategories is depicted. The black circles indicate the platform. (C) Circling(peripheral swimming at the tank wall); ES, extended searching (swimmingin all pool quadrants, visiting the same areas more than once); RS, restrictedsearching (swimming in some pool quadrants, but not visiting other tankareas at all); RC, restricted circling (swimming in some peripheral quadrants,not visiting central tank areas at all); F, finding (swimming directly towardthe platform without any exploration around the pool).*P< 0.05;** P< 0.01;*** P< 0.0001.
modifying the time course needed to acquire spatial knowl-edge. This successful behavior was obtained by exploitingprocedural competencies, besides working memory abilities,as indicated by the enriched animals’ high percentage of 45◦angles. The subsequent forced-choice procedure allowed sep-
Fig. 6. Effects of rearing conditions on maximal dendritic length (A), num-ber of intersections (B) and branching (C) of layer-III parietal pyramidalneurons in enriched and standard groups. Values reflect means (±S.E.M.) of10 neurons from three rats belonging to each experimental group (estimatedby nested-design ANOVA).∗P< 0.05.
Fig. 7. Illustrations of representative layer-III parietal pyramidal neuronsfrom rats reared in enriched (left) or standard (right) conditions. Note the in-crease in branching and extension of the apical dendrite in enriched neurons.Scale bar: 30�m.
86 M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90
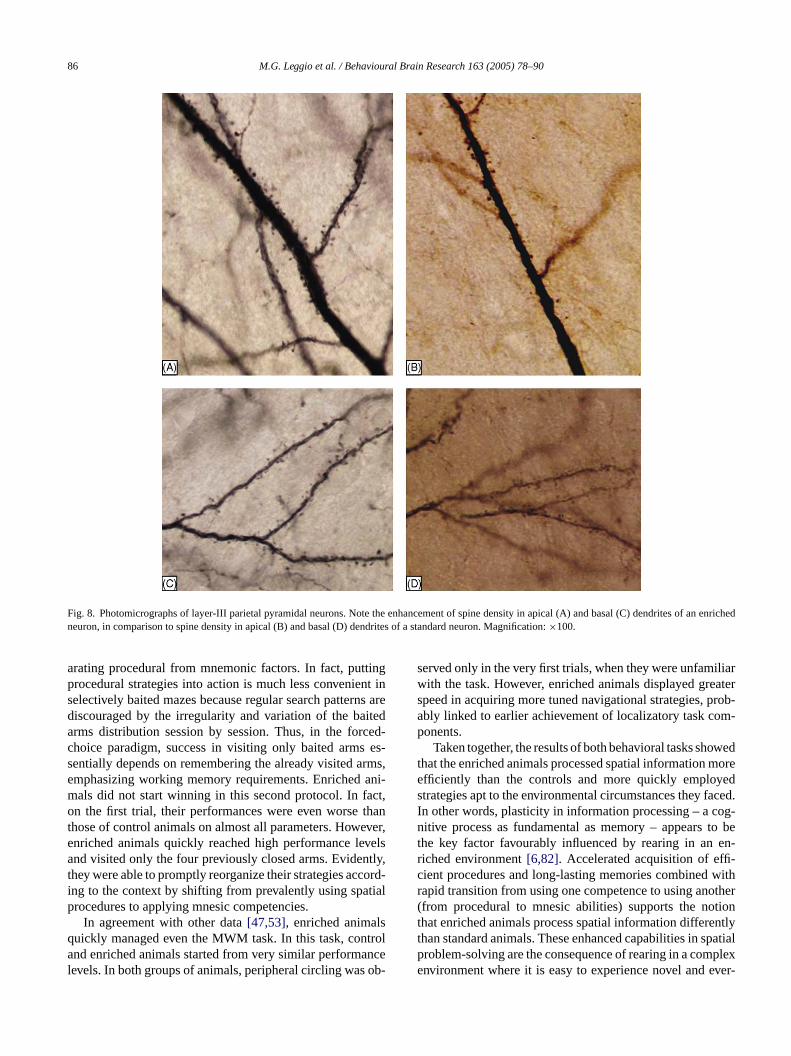
Fig. 8. Photomicrographs of layer-III parietal pyramidal neurons. Note the enhancement of spine density in apical (A) and basal (C) dendrites of an enrichedneuron, in comparison to spine density in apical (B) and basal (D) dendrites of a standard neuron. Magnification:×100.
arating procedural from mnemonic factors. In fact, puttingprocedural strategies into action is much less convenient inselectively baited mazes because regular search patterns arediscouraged by the irregularity and variation of the baitedarms distribution session by session. Thus, in the forced-choice paradigm, success in visiting only baited arms es-sentially depends on remembering the already visited arms,emphasizing working memory requirements. Enriched ani-mals did not start winning in this second protocol. In fact,on the first trial, their performances were even worse thanthose of control animals on almost all parameters. However,enriched animals quickly reached high performance levelsand visited only the four previously closed arms. Evidently,they were able to promptly reorganize their strategies accord-ing to the context by shifting from prevalently using spatialprocedures to applying mnesic competencies.
In agreement with other data[47,53], enriched animalsquickly managed even the MWM task. In this task, controland enriched animals started from very similar performancelevels. In both groups of animals, peripheral circling was ob-
served only in the very first trials, when they were unfamiliarwith the task. However, enriched animals displayed greaterspeed in acquiring more tuned navigational strategies, prob-ably linked to earlier achievement of localizatory task com-ponents.
Taken together, the results of both behavioral tasks showedthat the enriched animals processed spatial information moreefficiently than the controls and more quickly employedstrategies apt to the environmental circumstances they faced.In other words, plasticity in information processing – a cog-nitive process as fundamental as memory – appears to bethe key factor favourably influenced by rearing in an en-riched environment[6,82]. Accelerated acquisition of effi-cient procedures and long-lasting memories combined withrapid transition from using one competence to using another(from procedural to mnesic abilities) supports the notionthat enriched animals process spatial information differentlythan standard animals. These enhanced capabilities in spatialproblem-solving are the consequence of rearing in a complexenvironment where it is easy to experience novel and ever-
M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90 87
Fig. 9. (A) Means (±S.E.M.) of dendritic spines of 10 layer-III parietalpyramidal neurons from three rats belonging to each experimental group(estimated by nested-design ANOVA).** P< 0.005;*** P< 0.0001. (B andC) spine densities in the apical and basal dendrites are plotted.
changing situations. Better performances are probably theresult either of a more complex mnesic representation withstronger connections among cues and of quickly learned pro-cedural competencies more confidently driving navigationaltrajectories.
The brain responds with a broad range of measurablechanges to the sensory complexity of the environment,the richness of stimuli and voluntary physical activity inexperiencing the environment. From the earliest research onthe structural effects of rearing in an enriched environment,morphological changes have been reported in numerousbrain areas, including the hippocampus and various neo-cortical regions[74]. For example, in the hippocampus, thenumber of dendritic spines increases after spatial learningin adult rats [50]. Experience-dependent changes in theadult visual cortex, including effects on neuron numbers,
have been reported in rats[38] and cats[2,3]. Although thefindings on neocortical neurogenesis were not confirmed byother researchers, structural changes on neuronal morphol-ogy are generally assumed to be the anatomical substrateof processing and storage of learned experience. Becauseneurons in all brain regions involved in learning and memory(including those located in the same histotypical layer) reactdifferently to changes in the environment[33], it seemedinteresting to analyze neuronal modifications in the parietalcortex, a critical brain region in spatial learning and thuspotentially stimulated by variables present in our enrichedenvironment. We found that complex early experiencesmodified synaptic organization of layer-III parietal pyrami-dal neurons. Structural changes included increased dendriticarborisation as well as increased density of dendritic spines[13,35,64]. It has been suggested that spines play a functionalrole in synaptic plasticity, constituting a mechanism forconverting short-term alterations into long-lasting changesin connectivity and functioning of synapses[35,69]. Aninteresting finding here is that environmental enrichmentleads to enhanced synaptic strength, possibly linked tomechanisms that regulate LTP expression[27].
In this view, the increased spine density in the parietal neu-rons of enriched animals suggests a higher number of contactswith other neurons and thus a more complex neuronal wiringthat could be related to the enhanced performances on spa-t riousp heda den-s ever,b istri-b s hasi les.A redi Fasts oret ,i ns-f n ther e bet-t ilem uront icc ns ofo ffecto to ane ngesc d ins nce-d ntdt orel them
atest hol-
ial tasks. Interestingly, early experience affected the vaarts of parietal pyramidal neurons. Namely, in our enricnimals, the increase in dendritic arborisation and spineity was found in apical as well as in basal dendrites. Howasal and apical dendrites differentiate in spine density dution. The difference we observed in EC and SC group
mportant implications for their possible computational ros shown inFig. 8B and C, the highest spine density occur
n the proximal portions of basal and apical dendrites.ignals occurring in these dendritic regions contribute mo neuronal activation than distal synapses[34]. In contrastn the distal dendrites, only slow signals are efficiently traerred to the soma, suggesting thus a modulatory role oesting potential of the neurons. Thus, distal synapses arer suited for tonic modulation of neuronal excitability, whore proximal synapses are better suited for driving a ne
o fire an action potential[34]. Since the proximal dendritomponent was mainly increased in the pyramidal neurour animals reared in enriched condition, the functional ef the observed structural changes was probably linkedasier triggering the neuronal firing. These neuronal chaould represent a putative plasticity mechanism involvetable reorganization partially accounting for the experieependent changes we observed[85]. Of course, the preseata do not allow establishing whether, as suggested[14],
hese long-lasting morphological changes coexist with mabile synaptic alterations reflecting salient features of
ost recent individual experience.In conclusion, the present experimental study indic
hat environmental enrichment modifies neuronal morp

88 M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90
ogy and improves cognitive performances, consistently withstudies in humans that indicate neuroprotective effects ofthe intellectual stimulation[37,70]. Recently, it has beensuggested that “early environmental enrichment is associatedwith lower levels of antisocial behavior and schizotypalpersonality in adulthood”[63]. Environmental enrichment,by stimulating multiple sensory systems, exposing subjectsto complex spatial environments and providing numerousproblem-solving opportunities, appears be a useful tool toact on behavioral impairment characterized by cognitivedeficits. However, it has been not yet determined which envi-ronmental components (physical exercise, social interaction,cognitive stimulation) had beneficial effects on preventingdebilitating mental disorders. The experimental approachmay provide a useful support in separately analyzing any as-pect, allowing the application of more focused environmentaltherapy.
Acknowledgements
We gratefully acknowledge Dr. P. Pasqualetti for his kindstatistical assistance. This work was supported by MIURgrants to L.P.
R
vakadial
the
apsesom-
andtors.Ox-
WT.es an-Sci
ofviron-
maze.mp
and
[ ry: ampus
[ Theory
[ rietal
[13] Comery TA, Stamoudis CX, Irwin SA, Greenough WT. Increaseddensity of multiple-head dendritic spines on medium-sized spinyneurons of the striatum in rats reared in a complex environment.Neurobiol Learn Mem 1996;66:93–6.
[14] Coq JO, Xerri C. Environmental enrichment alters organizationalfeatures of the forepaw representation in the primary somatosensorycortex of adult rats. Exp Brain Res 1998;121:191–204.
[15] Colombel C, Lalonde R, Caston J. The effects of unilateral removalof the cerebellar hemispheres on spatial learning and memory in rats.Brain Res 2004;1004:108–15.
[16] Constantinidis C, Franowicz MN, Goldman-Rakic PS. The sensorynature of mnemonic representation in the primate prefrontal cortex.Nat Neurosci 2001;4:311–6.
[17] Cotman CW, Berchtold NC. Exercise: a behavioral interven-tion to enhance brain health and plasticity. Trends Neurosci2002;25:295–301.
[18] Diamond MC, Ingham CC, Johnson RE, Bennett EL, RosenzweigMR. Effects of environment on morphology of rat cerebral cortexand hippocampus. J Neurobiol 1976;7:75–85.
[19] Diamond MC, Law F, Rhodes H, Lindner B, Rosenzweig MR, KrechD, et al. Increases in cortical depth and glia numbers in rats subjectedto enriched environment. J Comp Neurol 1966;128:117–26.
[20] Diamond MC, Lindner B, Raymond A. Extensive cortical depth mea-surements and neuron size increases in the cortex of environmentallyenriched rats. J Comp Neurol 1967;131:357–64.
[21] DiMattia BD, Kesner RP. Spatial cognitive maps: differential roleof parietal cortex and hippocampal formation. Behav Neurosci1988;102:471–80.
[22] Duffy SN, Craddock KJ, Nguyen PV. Environmental enrichmentmodifies the PKA-dependence of hippocampal LTP and improveshippocampus-dependent memory. Learn Mem 2001;8:26–34.
[ nvi-plo-ehav
[ icalment.
[ bladphicatial3–6.
[ cam-aze
[ apticrain
[ andging
[ therningem
[ ntaltinc-
[ hley
[ l ac-raleuro-
[ exityrat.
eferences
[1] Anderson BJ, Rapp DN, Back DH, Mc Closkcy DP, Coburn-LitPS, Robinson JK. Exercise influences spatial learning in the rarm maze. Physiol Behav 2000;70:425–9.
[2] Beaulieu C, Colonnier M. Effect of richness of environment oncat visual cortex. J Comp Neurol 1987;266:478–91.
[3] Beaulieu C, Colonnier M. Number and size of neurons and synin the motor cortex of cats raised in different environmental cplexities. J Comp Neurol 1989;289:178–81.
[4] Berthoz A. Hippocampal and parietal contribution to topokinetictopographic memory. In: Burgess N, Jeffery KJ, O’Keefe J, ediThe hippocampal and parietal foundations of spatial cognition.ford: Oxford University Press; 1999. p. 1–490.
[5] Black JE, Isaacs KR, Anderson BJ, Alcantara AA, GreenoughLearning causes synaptogenesis, whereas motor activity causgiogenesis, in cerebellar cortex of adult rats. Proc Natl AcadUSA 1990;87:5568–72.
[6] Bourgeon S, Xerri C, Coq JO. Abilities in tactile discriminationtextures in adult rats exposed to enriched or impoverished enments. Behav Brain Res 2004;153:217–31.
[7] Brandeis R, Brandys J, Yehuda S. The use of the Morris waterin the study of memory learning. Int J Neurosci 1989;48:29–69
[8] Brown RT. Early experience and problem-solving ability. J CoPhysiol Psychol 1968;65:433–40.
[9] Bucci DJ, Chiba AA, Gallagher M. Spatial learning in malefemale long-Evans rats. Behav Neurosci 1995;109:180–3.
10] Chiba AA, Kesner RP, Jackson PA. Two forms of spatial memodouble dissociation between the parietal cortex and the hippocain the rat. Behav Neurosci 2002;116:874–83.
11] Chodzko-Zajko WJ, Schuler P, Solomon J, Heinl B, Ellis NR.influence of physical fitness on automatic and effortful memchanges in aging. Int J Aging Hum Dev 1992;35:265–85.
12] Colby CL, Duhamel JR. Spatial representations for action in pacortex. Cogn Brain Res 1996;5:105–15.
23] Engellenner WJ, Goodlett CR, Burright RG, Donovick PJ. Eronmental enrichment and restriction: effects on reactivity, exration and maze learning in mice with septal lesions. Physiol B1982;29:885–93.
24] Faherty CJ, Kerley D, Smeyne RJ. A Golgi-Cox morphologanalysis of neuronal changes induced by environmental enrichBrain Res Dev 2003;141:55–61.
25] Falkenberg T, Mohammed HK, Henriksson B, Persson H, WinB, Lindefors N. Increased expression of brain-derived neurotrofactor mRNA in rat hippocampus is associated with improved spmemory and enriched environment. Neurosci Lett 1992;138:15
26] Floresco SB, Seamans JK, Phillips AG. Selective roles for hippopal, prefrontal cortical, and ventral striatal circuits in radial-arm mtasks with or without a delay. J Neurosci 1997;17:1880–90.
27] Foster TC, Gagne J, Massicotte G. Mechanism of altered synstrength due to experience: relation to long-term potentiation. BRes 1996;736:243–50.
28] Frick KM, Fernadez SM. Enrichment enhances spatial memoryincreases synaptophysin levels in aged female mice. Neurobiol A2003;24:615–26.
29] Gaytan-Tocaven L, Olvera-Cortes ME. Bilateral lesion ofcerebellar-dentate nucleus impairs egocentric sequential leabut not egocentric navigation in the rat. Neurobiol Learn M2004;82:120–7.
30] Gardner EB, Boitano JJ, Mancino NS, D’Amico DP. Environmeenrichment and deprivation: effects on learning, memory and extion. Physiol Behav 1975;14:321–7.
31] Gill JR, Reid LD, Porter PB. Effects of restricted rearing on Lasstand performance. Psychol Rep 1966;19:239–42.
32] Gomez-Pinilla F, So V, Kesslak JP. Spatial learning and physicativity contribute to the induction of fibroblast growth factor: neusubstrates for increased cognition associated with exercise. Nscience 1998;85:53–61.
33] Greenough WT, Volkmar FR, Juraska J. Effects of rearing complon dendritic branching in frontolateral and temporal cortex of theExp Neurol 1973;41:371–8.

M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90 89
[34] Henze DA, Cameron W, Barrionuevo G. Dendritic morphology andits effects on the amplitude and rise-time of synaptic signals inhippocampal CA3 pyramidal cells. J Comp Neurol 1996;369:331–44.
[35] Hering H, Sheng M. Dendritic spines: structure, dynamics and reg-ulation. Nat Rev Neurosci 2001;2:880–8.
[36] Jiang X, Johnson RR, Burkhalter A. Visualization of dendritic mor-phology of cortical projection neurons by retrograde axonal tracing.J Neurosci Methods 1993;50:45–60.
[37] Kalmijn S, Feskens EJ, Launer LJ, Kromhout D. Longitudinal studyof the effect of apolipoprotein E4 allele on the association be-tween education and cognitive decline in elderly men. Br Med J1997;314:34–5.
[38] Kaplan MS. Neurogenesis in the 3-month-old rat visual cortex. JComp Neurol 1981;195:323–8.
[39] Kelche CR, Will BE. Effects of environment on functional re-covery after hippocampal lesions in adult rats. Physiol Behav1978;21:935–41.
[40] Kempermann G. Why new neurons? Possible functions for adulthippocampal neurogenesis. J Neurosci 2002;22:635–8.
[41] Kempermann G, Kuhn HG, Gage FH. More hippocampal neu-rons in adult mice living in an enriched environment. Nature1997;386:493–5.
[42] Kesner RP, Rogers J. An analysis of independence and interactionsof brain substrates that subserve multiple attributes, memory sys-tems, and underlying processes. Neurobiol Learn Mem 2004;82:199–215.
[43] Kesner RP, Farnsworth G, Kametani H. Role of parietal cortexand hippocampus in representing spatial information. Cereb Cortex1991;1:367–73.
[44] Kolb B, Gibb R, Gorny G. Experience-dependent changes in den-dritic arbor and spine density in neocortex vary qualitatively with
[ ntalchol
[ rainand
01–7.[ and
used
[ par-ntialpsy-
[ tion–3.
[ pinearn-
Natl
[ brainviron-
[ en-entate78.
[ nalationcent
[ ford:
[ itiveUn-brain
[56] Packard MG, Knowlton BJ. Learning and memory functions of thebasal ganglia. Annu Rev Neurosci 2002;25:563–93.
[57] Paxinos G, Watson C. The rat brain in stereotaxic coordinates. SanDiego: Academic Press; 1998.
[58] Pearce JM, Robert AD, Good M. Hippocampal lesions disrupt nav-igation based on cognitive maps but not heading vectors. Nature1998;396:75–7.
[59] Petrosini L, Molinari M, Dell’Anna ME. Cerebellar contribution tospatial event processsing: Morris water maze and T-maze. Eur JNeurosci 1996;8:1882–96.
[60] Pham TM, Ickes B, Albeck D, Soderstrom S, Granholm AC, Mo-hammed AH. Changes in brain nerve growth factor levels and nervegrowth factor receptors in rats exposed to environmental enrichmentfor 1 year. Neuroscience 1999;94:279–86.
[61] Pinto-Hamuy T, Montero VM, Torrealba F. Neurotoxic lesion ofanteromedial/posterior parietal cortex disrupts spatial maze memoryin blind rats. Behav Brain Res 2004;153:465–70.
[62] Porter MC, Burk JA, Mair RG. A comparison of the effects of hip-pocampal or prefrontal cortical lesions on three versions of delayednon-matching-to-sample based on positional or spatial cues. BehavBrain Res 2000;109:69–81.
[63] Raine A, Mellingen K, Liu J, Venables P, Mednick SA. Effects ofenvironmental enrichment at ages 3–5 years on schizotypal person-ality and antisocial behaviour at ages 17 and 23 years. Am J Psychol2003;160:1627–35.
[64] Rampon C, Tang YP, Goodhouse J, Shimizu E, Kyin M, Tsien JZ.Enrichment induces structural changes and recovery from nonspatialmemory deficits in CA1 NMDAR1-knockout mice. Nat Neurosci2000;3:238–44.
[65] Renner MJ, Rosenzweig MR. Enriched and impoverished environ-ments, vol. xi. New York: Springer; 1987, 134 p.
[ ingehav
[ adelwith
00;3:
[ rac-place
[ unit.
[ em-suc-
[ uen;
[ ivitychol
[ onatalrning
[ es of
[ onand
[ ch-avior
[ aidseural
[ H.ning
age and sex. Neurobiol Learn Mem 2003;79:1–10.45] Krech D, Rosenzweig MR, Bennett EL. Effects of environme
complexity and training on brain chemistry. J Comp Physiol Psy1960;53:509–19.
46] Krech D, Rosenzweig MR, Bennett EL. Relations between bchemistry and problem-solving among rats raised in enrichedimpoverished environments. J Comp Physiol Psychol 1962;55:8
47] Larsson F, Winblad B, Mohammed AH. Psychological stressenvironmental adaptation in enriched versus impoverished horats. Pharmacol Biochem Behav 2002;73:193–207.
48] Martin SJ, de Hoz L, Morris RGM. Retrograde amnesia neithertial nor complete hippocampal lesions in rats result in preferesparing of remote spatial memory, even after reminding. Neurochologia 2005;43:609–24.
49] Morris RGM, Garrud P, Rawlings JNP, O’Keefe J. Place navigaimpaired in rats with hippocampal lesion. Nature 1982;297:681
50] Moser MB, Trommald M, Andersen P. An increase in dendritic sdensity on hippocampal CA1 pyramidal cells following spatial leing in adult rats suggests the formation of new synapses. ProcAcad Sci USA 1994;91:12673–5.
51] Nakamura H, Kobayashi S, Ohashi Y, Ando S. Age-changes ofsynapses and synaptic plasticity in response to an enriched enment. J Neurosci Res 1999;56:307–15.
52] Nilsson M, Perfilieva E, Johansson U, Orwar O. Eriksson PSriched environment increases neurogenesis in the adult rat dgyrus and improves spatial memory. J Neurobiol 1999;39:569–
53] Oitzl MS, Workel JO, Fluttert M, Frosch F, De Kloet ER. Materdeprivation affects behaviour from youth to senescence: amplificof individual differences in spatial learning and memory in senesBrown Norway rats. Eur J Neurosci 2000;12:3771–80.
54] O’Keefe J, Nadel L. The hippocampus as a cognitive map. OxOxford University Press; 1978. p. 1–570.
55] O’Keefe J. A computational theory of the hippocampal cognmap. In: In: Storm-Mathisen J, Zimmer J, Ottersen OP, editors.derstanding the brain through the hippocampus, progress inresearch, vol. 83. Amsterdam: Elsevier; 1990. p. 301–12.
66] Rondi-Reig L, Le Marec N, Caston J, Mariani J. The role of climband parallel fibers inputs to cerebellar cortex in navigation. BBrain Res 2002;132:11–8.
67] Rosenbaum RS, Priselac S, Kohler S, Black SE, Gao F, NL, et al. Remote spatial memory in an amnesic personextensive bilateral hippocampal lesions. Nat Neurosci 201044–8.
68] Save E, Paz-Villagran V, Alexinsky T, Poucet B. Functional intetion between the associative parietal cortex and hippocampalcell firing in the rat. Eur J Neurosci 2005;21:522–30.
69] Shepherd GM. The dendritic spine: a multifunctional integrativeJ Neurophysiol 1996;75:2197–210.
70] Shimamura AP, Berry JM, Mangels JA, Rusting CL, Jurica PJ. Mory and cognitive abilities in university professors: evidence forcessful aging. Psychol Sci 1995;6:271–7.
71] Sholl DA. The organization of the cerebral cortex. London: Meth1956. p. 1–125.
72] Smith HV. Effects of environmental enrichment on open-field actand Hebb–Williams problem solving in rats. J Comp Physiol Psy1972;80:163–8.
73] Tees RC. The influences of sex, rearing environment, and necholine dietary supplementation on spatial and nonspatial leaand memory in adult rats. Dev Psychobiol 1999;35:328–42.
74] Van Praag H, Kempermann G, Gage FH. Neural consequencenvironmental enrichment. Nat Rev Neurosci 2000;1:191–8.
75] Van Waas M, Soffie M. Differential environmental modulationslocomotor activity, exploration and spatial behaviour in youngold rats. Physiol Behav 1996;59:265–71.
76] Varty GB, Paulus MP, Braff DL. Geyer MA environmental enriment and isolation rearing in the rat: effects on locomotor behand startle response plasticity. Biol Psych 2000;47:864–73.
77] Whishaw IQ, Zaborowski JA, Kolb B. Postsurgical enrichmentadult hemidecorticate rats on a spatial navigation task. Behav NBiol 1984;42:183–90.
78] Will BE, Rosenzweig MR, Bennett EL, Hebert M, MorimotoRelatively brief environmental enrichment aids recovery of lear

90 M.G. Leggio et al. / Behavioural Brain Research 163 (2005) 78–90
capacity and alters brain measures after postweaning brain lesionsin rats. J Comp Physiol Psychol 1977;91:33–50.
[79] Will BE, Rosenzweig MR, Bennett EL. Effects of differentialenvironments on recovery from neonatal brain lesions, measuredby problem-solving scores and brain dimensions. Physiol Behav1976;16:603–11.
[80] Williams BM, Luo Y, Ward C, Redd K, Gibson R, Kuczaj SA, etal. Environmental enrichment: effects on spatial memory and hip-pocampal CREB immunoreactivity. Physiol Behav 2001;73:649–58.
[81] Winocur G, Moscovitch M. Hippocampal and prefrontal cortexcontributions to learning and memory: analysis of lesion and ag-ing effects on maze learning in rats. Behav Neurosci 1990;104:544–51.
[82] Woodcock EA, Richardson R. Effects of environmental enrichmenton rate of contextual processing and discriminative ability in adultrats. Neurobiol Learn Mem 2000;73:1–10.
[83] Yeshenko O, Guazzelli A, Mizumori SJ. Context-dependent reorga-nization of spatial and movement representations by simultaneouslyrecorded hippocampal and striatal neurons during performance ofallocentric and egocentric tasks. Behav Neurosci 2004;118:751–69.
[84] Young D, Lawlor PA, Leone P, Dragunow M, During MJ. Environ-mental enrichment inhibits spontaneous apoptosis, prevents seizuresand is neuroprotective. Nat Med 1999;5:448–53.
[85] Yuste R, Bonhoeffer T. Morphological changes in dendritic spinesassociated with long-term synaptic plasticity. Annu Rev Neurosci2001;24:1071–89.




















