
Wnt2 signaling is necessary and sufficient to activate the airway smooth muscle program in the lung...
21
Wnt2 signaling is necessary and sufficient to activate the airway smooth muscle program in the lung by regulating myocardin/ Mrtf-B and Fgf10 expression Ashley M. Goss 1 , Ying Tian 1 , Lan Cheng 1 , Jifu Yang, Diane Zhou 1 , Ethan David Cohen 1 , and Edward E. Morrisey 1,2,3,4,* 1 Department of Medicine University of Pennsylvania Philadelphia, PA 19104 2 Department of Cell and Developmental Biology University of Pennsylvania Philadelphia, PA 19104 3 Cardiovascular Institute University of Pennsylvania Philadelphia, PA 19104 4 Institute for Regenerative Medicine University of Pennsylvania Philadelphia, PA 19104 Abstract Smooth muscle in the lung is thought to derive from the developing lung mesenchyme. Smooth muscle formation relies upon coordination of both autocrine and paracrine signaling between the budding epithelium and adjacent mesenchyme to govern its proliferation and differentiation. However, the pathways initiating the earliest aspects of smooth muscle specification and differentiation in the lung are poorly understood. Here, we identify the Wnt2 ligand as a critical regulator of the earliest aspects of lung airway smooth muscle development. Using Wnt2 loss and gain of function models, we show that Wnt2 signaling is necessary and sufficient for activation of a transcriptional and signaling network critical for smooth muscle specification and differentiation including myocardin/Mrtf-B and the signaling factor Fgf10. These studies place Wnt2 high in a hierarchy of signaling molecules that promote the earliest aspects of lung airway smooth muscle development. INTRODUCTION Disruption to lung smooth muscle development and function in humans can lead to chronic health complications including asthma and pulmonary hypertension (Camoretti-Mercado et al., 2003; Halayko et al., 1998; Halayko and Stephens, 1994; Humbert et al., 2004; Solway et al., 1998). Understanding the signaling pathways coordinating the various specification and differentiation processes occurring during smooth muscle development are critical to a complete understanding of lung development as well as pathogenesis in the postnatal lung. In the early lung bud, the condensing lung mesenchyme is thought to generate several cell lineages including smooth muscle cells, endothelial cells, and resident fibroblasts such as myofibroblasts, lipofibroblasts, and stromal fibroblasts (reviewed in (Morrisey and Hogan, 2010). At the level of the mainstem bronchi, smooth muscle develops partially around the © 2011 Elsevier Inc. All rights reserved * To whom correspondence should be addressed: Edward E. Morrisey, Ph.D. University of Pennsylvania 956 BRB II/III 421 Curie Blvd. Philadelphia, PA 19104 Phone: 215-573-3010 FAX: 215-573-2094 Email: [email protected]. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Dev Biol. Author manuscript; available in PMC 2012 August 15. Published in final edited form as: Dev Biol. 2011 August 15; 356(2): 541–552. doi:10.1016/j.ydbio.2011.06.011. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Transcript of Wnt2 signaling is necessary and sufficient to activate the airway smooth muscle program in the lung...

Wnt2 signaling is necessary and sufficient to activate the airwaysmooth muscle program in the lung by regulating myocardin/Mrtf-B and Fgf10 expression
Ashley M. Goss1, Ying Tian1, Lan Cheng1, Jifu Yang, Diane Zhou1, Ethan David Cohen1,and Edward E. Morrisey1,2,3,4,*
1Department of Medicine University of Pennsylvania Philadelphia, PA 191042Department of Cell and Developmental Biology University of Pennsylvania Philadelphia, PA191043Cardiovascular Institute University of Pennsylvania Philadelphia, PA 191044Institute for Regenerative Medicine University of Pennsylvania Philadelphia, PA 19104
AbstractSmooth muscle in the lung is thought to derive from the developing lung mesenchyme. Smoothmuscle formation relies upon coordination of both autocrine and paracrine signaling between thebudding epithelium and adjacent mesenchyme to govern its proliferation and differentiation.However, the pathways initiating the earliest aspects of smooth muscle specification anddifferentiation in the lung are poorly understood. Here, we identify the Wnt2 ligand as a criticalregulator of the earliest aspects of lung airway smooth muscle development. Using Wnt2 loss andgain of function models, we show that Wnt2 signaling is necessary and sufficient for activation ofa transcriptional and signaling network critical for smooth muscle specification and differentiationincluding myocardin/Mrtf-B and the signaling factor Fgf10. These studies place Wnt2 high in ahierarchy of signaling molecules that promote the earliest aspects of lung airway smooth muscledevelopment.
INTRODUCTIONDisruption to lung smooth muscle development and function in humans can lead to chronichealth complications including asthma and pulmonary hypertension (Camoretti-Mercado etal., 2003; Halayko et al., 1998; Halayko and Stephens, 1994; Humbert et al., 2004; Solwayet al., 1998). Understanding the signaling pathways coordinating the various specificationand differentiation processes occurring during smooth muscle development are critical to acomplete understanding of lung development as well as pathogenesis in the postnatal lung.In the early lung bud, the condensing lung mesenchyme is thought to generate several celllineages including smooth muscle cells, endothelial cells, and resident fibroblasts such asmyofibroblasts, lipofibroblasts, and stromal fibroblasts (reviewed in (Morrisey and Hogan,2010). At the level of the mainstem bronchi, smooth muscle develops partially around the
© 2011 Elsevier Inc. All rights reserved*To whom correspondence should be addressed: Edward E. Morrisey, Ph.D. University of Pennsylvania 956 BRB II/III 421 CurieBlvd. Philadelphia, PA 19104 Phone: 215-573-3010 FAX: 215-573-2094 Email: [email protected]'s Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to ourcustomers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review ofthe resulting proof before it is published in its final citable form. Please note that during the production process errors may bediscovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptDev Biol. Author manuscript; available in PMC 2012 August 15.
Published in final edited form as:Dev Biol. 2011 August 15; 356(2): 541–552. doi:10.1016/j.ydbio.2011.06.011.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

airway on the dorsal surface with cartilaginous rings forming on the ventral surface. Slightlymore posterior in the proximal airways, smooth muscle forms around the entire epitheliumwhile the most distal portion of the respiratory tree lacks smooth muscle (Tollet et al., 2001,2002). Airway smooth muscle development (ASM) closely follows the growth anddifferentiation of the airway epithelium temporally and spatially. At E11.5 in the mouse, asmooth muscle layer starts to envelop the trachea and proximal bronchi and continues todevelop around the proximal airway tubules as lung development progresses (Badri et al.,2008). Current theories suggest that ASM originates from a distal pool of Fgf10-expressingmesenchymal cells that surrounds the tip of the branching airway epithelium (Mailleux etal., 2005; Ramasamy et al., 2007). These distal cells proliferate and actively or passivelytranslocate to surround the proximal airways undergoing epithelial tube outgrowth andbranching.
Several signaling pathways are known to be critical in lung smooth muscle developmentincluding FGF, BMP, Shh, and Wnt signaling. Fgf10 signaling is important for ASMdevelopment and mediates this process in part, through signaling interactions with theepithelium. Fgf10 signaling from the mesenchyme activates a gene expression program inthe adjacent developing epithelium, which signals in a reciprocal fashion back to themesenchyme to direct smooth muscle differentiation (Mailleux et al., 2005). The pro-myogenic signals expressed in the developing airway epithelium include Shh and Bmp4 andthese factors direct smooth muscle differentiation along the branching epithelial tubule asFgf10-expressing cells condense around the primitive airway (Mailleux et al., 2005). Fgf9signaling from the mesothelium is also reported to mediate the development of immaturesmooth muscle cells in the distal mesenchyme by limiting the differentiation response of themesenchyme to Shh signaling (Colvin et al., 2001).
Published data indicate that Wnt/β-catenin signaling is active and plays a role during smoothmuscle development in the lung. From E14.5 to E18.5, Wnt/β-catenin signaling activity isobserved in the developing ASM (Shu et al., 2005). Wnt7b is a canonical Wnt ligandexpressed in the developing epithelium, and Wnt7b knockout mice display perinatalpulmonary hemorrhaging due to disrupted vascular smooth muscle (VSM) development(Shu et al., 2002). Further investigation into the mechanisms of Wnt7b signaling revealedthat Wnt7b broadly regulates the development of smooth muscle cells in the early lung budmesenchyme (Cohen et al., 2009). Additionally, treating lung explants with the canonicalWnt signaling inhibitor Dickkopf-1 (Dkk-1) impairs smooth muscle gene expression (DeLanghe et al., 2005).
Wnt2 is a ligand robustly expressed throughout the lung mesenchyme during the course oflung development. Wnt2 is first expressed in the mesoderm surrounding the anterior foregutendoderm from E9.0–E9.5 during the period when the lung is specified, and Wnt2 continuesto be expressed in the mesoderm condensing around the early lung bud to form the primitivelung mesenchyme with expression persisting up to perinatal timepoints (Goss et al., 2009;Monkley et al., 1996). The expression pattern of Wnt2 suggests that it may play a role in thedevelopment of mesenchymal-derived lineages in the lung. Previous work has shown thatWnt2, in cooperation with Wnt2b, is essential for specification of the respiratory lineage inthe anterior foregut endoderm (Goss et al., 2009). However, whether Wnt2 plays a role inregulating mesenchymal lineage differentiation in the lung is unclear. In this report, weshow that a loss of Wnt2 leads to a severe deficiency in ASM development in the lung. Weshow that Wnt2 acts upstream of Fgf10 and the critical transcription factors myocardin andMrtf-B to regulate early ASM differentiation in the multipotent lung mesenchyme.Moreover, Wnt2 acts in a paracrine manner to promote Wnt7b expression in the endodermwhich in turn promotes lung smooth muscle development. Thus, Wnt2 plays an important
Goss et al. Page 2
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

role in the lung mesenchyme by promoting the earliest stages of ASM development inaddition to its role in specifying the respiratory endoderm fate.
RESULTSWnt2−/− null lungs exhibit airway smooth muscle defects
Wnt2 is robustly expressed in the developing lung mesenchyme throughout lungdevelopment (Goss et al., 2009). To investigate whether Wnt2 regulates mesenchymaldevelopment, E18.5 wild type control and Wnt2−/− null mutant embryos were examined forsmooth muscle defects. Immunostaining for SM22α expression, a marker of smooth musclecells, revealed a reduction in smooth muscle surrounding the proximal and distal bronchiolarairways (Fig. 1A and B and data not shown). Higher magnification of Wnt2−/− null mutantlung cross-sections showed sporadic SM22α-expressing cells around the airways, resultingin significant gaps in the sub-epithelial ASM layer (Fig. 1C and D). Interestingly, SM22αexpression in the smooth muscle layer of adjacent pulmonary arteries appeared unchanged(Fig. 1A and B, arrows).
Wnt2 signaling through β-catenin regulates airway smooth muscle development in thelung mesenchyme
The Wnt/β-catenin transgenic reporter mice BAT-GAL and TOP-GAL reveal Wnt/β-cateninsignaling activity in the developing lung smooth muscle (Okubo and Hogan, 2004; Shu etal., 2005), and Wnt2 is reported to signal canonically through the Wnt/β-catenin pathway inthe lung (Goss et al., 2009). To determine if Wnt2 is required for Wnt/β-catenin signalingactivity in the developing ASM, Wnt2+/− mutants were crossed into the transgenic BAT-GALreporter background to yield Wnt2−/−:BAT-GAL embryos (Fig. 1E and F). In E14.5 controlBAT-GAL embryos, lacZ-positive cells were labeled in the developing epithelium and in thepresumptive ASM layer (Fig. 1E). In Wnt2−/−:BAT-GAL mutants, there was a reduction inthe number of lacZ-positive cells in the developing ASM layer, indicating a loss of Wnt/β-catenin activity in these cells (Fig. 1F). BAT-GAL activity was also reduced inWnt2−/−:BAT-GAL mutant lung epithelium as previously reported (Fig. 1F and (Goss et al.,2009). Additionally, immunostaining for Axin2, a Wnt/β-catenin transcriptional target (Jhoet al., 2002), showed reduced expression in the presumptive ASM layer of E14.5 Wnt2−/−
null lungs (Fig. 1G and H). Quantification of Axin2 gene expression in E14.5 control andWnt2−/− null lungs confirmed a significant reduction in the level of Axin2 expression in theabsence of Wnt2 signaling (Fig. 1I). Together, these data suggest that Wnt2 signals throughthe β-catenin dependent canonical pathway in the lung mesenchyme to mediate proper ASMdevelopment.
Wnt2 regulates development of immature smooth muscle from the lung mesenchymeWe hypothesized that the loss of mature ASM in Wnt2−/− null mutants was due to an earlyloss of SMC development in the primitive mesenchyme. SM22α is a marker of bothprimitive and mature SMCs (Solway et al., 1995). In E11.5 wild type embryos, SM22αexpression was observed in a sub-epithelial population of cells forming around thedeveloping proximal airway (Fig. 2A). In Wnt2−/− null embryos, this expression wasreduced indicating a loss of SMC differentiation in the absence of Wnt2 signaling (Fig. 2B).Quantitative PCR (Q-PCR) on E11.5 control and Wnt2−/− null lung buds confirmed anapproximate 50% reduction of SM22α expression in Wnt2−/− null lung buds (Fig. 2I). AtE14.5, a thick and contiguous layer of SM22α–expressing cells was observed in thedeveloping ASM of wild type embryos (Fig. 2C). In contrast, fewer SM22α expressing cellswere labeled in Wnt2−/− null lungs and the loss results in a thinned layer of ASM withpronounced gaps along the epithelium-mesenchyme border (Fig. 2D).
Goss et al. Page 3
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Pdgfrα and Pdgfrβ are receptor tyrosine kinases through which members of the platelet-derived growth factor family (Pdgf) of ligands signal to regulate proliferation, migration,and differentiation of embryonic mesenchyme (reviewed in (Betsholtz, 2004). Pdgfrα andPdgfrβ are broadly expressed in the multipotent mesenchymal cells of the early lung bud andare important in the development of smooth muscle cells from the lung mesenchyme(Bostrom et al., 1996; Hellstrom et al., 1999). Both receptors continue to be expressed in theembryonic lung in both ASM and VSM (Cohen et al., 2009).
In E11.5 wild type embryos, expression of Pdgfrβ was observed in mesenchymal cells thatare coalescing around the developing airways and vasculature (Fig. 2E). However, inWnt2−/− null embryos, Pdgfrβ expression was reduced throughout the mesenchyme (Fig.2F). Q-PCR on E11.5 control and Wnt2−/− null lung buds demonstrated a significantreduction in Pdgfrβ expression (Fig. 2I). In E14.5 wild type embryos, Pdgfrβ expression wasobserved in the sub-epithelial mesenchyme as well as the developing smooth muscleencompassing the vasculature (Fig. 2G, arrowhead). In Wnt2−/− null embryos, Pdgfrβexpression in the mesenchyme surrounding the airways was reduced whereas Pdgfrβexpression in the VSM did not appear to be effected (Fig. 2H, arrowhead). At E14.5 Pdgfrαwas also found to be robustly expressed in the presumptive ASM of wild type embryos andloss of Wnt2 signaling led to a reduction in Pdgfrα expression in the mesenchymesurrounding the airways (Supplemental Fig. 1). Together, this data suggests that loss ofWnt2 function disrupts the development of a SM22α+;Pdgfrα/β+ population of immatureSMCs in the early lung mesenchyme, which leads to a loss of mature ASM.
Wnt2 signaling is necessary for activation of the smooth muscle transcription factorsmyocardin and Mrtf-B in the lung
To assess whether the earliest stages of smooth muscle cell differentiation are activated inWnt2−/− null lungs, expression levels of the transcription factors myocardin and Mrtf-B wereexamined. Myocardin and Mrtf-B are members of the myocardin related transcription factor(Mrtf) family of serum response factor (SRF) co-factors and expressed in the developing andmature smooth muscle of several tissues including the lung (reviewed in (Wang and Olson,2004). Myocardin is a critical upstream regulator of myogenesis and in primitive smoothmuscle cells, myocardin transactivates numerous smooth muscle differentiation genesincluding SM22α and smooth muscle α-actin (SMA) (Wang et al., 2001; Wang et al., 2003).At E11.5, in situ hybridizations on wild type embryos showed myocardin expression aroundthe proximal airway of the primary lung bud (Figure 3A, arrowheads). In Wnt2−/− nullmutant embryos at both E11.5 and E14.5, there was a reduction in myocardin expressionsurrounding the developing airway (Fig. 3A–D). Quantification of myocardin expressionlevels in E11.5 control and Wnt2−/− null lung buds demonstrated an approximate 80%reduction in myocardin expression in Wnt2−/− null lung buds (Fig. 3E). Additionally,quantification of Mrtf-B expression was also significantly reduced in E11.5 Wnt2−/− nulllung buds as compared to control lung buds (Fig. 3E). The loss of myocardin and Mrtf-Bexpression in Wnt2−/− null lung buds indicates the down-regulation of an early myogenictranscriptional program in the lung, suggesting that Wnt2 is required for initiation of thesmooth muscle gene program in the developing lung.
Differentiation of other mesenchymal-derived lineages in Wnt2−/− null lungsMultiple cell types are hypothesized to derive from the developing lung mesenchymeincluding endothelial and lymphatic cell lineages (Chinoy, 2003). Moreover, lungmesenchyme could provide a necessary source of signaling factors, including Wnt2,required for proper investment of vascular endothelium from outside the lung. Thus, wesought to determine whether differentiation of other mesenchymal lineages includingvascular and lymphatic endothelial cell lineages were affected in Wnt2−/− mutants.
Goss et al. Page 4
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Q-PCR was performed on E12.5 control and Wnt2−/− null lung buds to assess endothelialmarker gene expression. Pecam1 expression was not significantly reduced in Wnt2−/− nulllung buds whereas Flk1 was slightly reduced to 85% of wild-type expression (Fig. 4B).Immunostaining confirmed that PECAM1 (CD31) expression was unaltered in Wnt2−/−
mutants (Supplemental Fig. 2). Members of the Vegf family of molecules are important forregulating vascular and lymphatic development in multiple tissue types (Lohela et al., 2009).Vegf-A, Vegf-C, and Vegf-D are expressed in the lung during development and Vegf-A andVegf-D are critical for proper vascular endothelial development while Vegf-C is necessaryfor lymphatic endothelial development (Karkkainen et al., 2004; Thebaud, 2007). Q-PCRrevealed comparable expression levels of Vegf-A and Vegf-D between control and Wnt2−/−
null lung buds (Fig. 4C and D). Interestingly, Vegf-C expression was significantly reducedin Wnt2−/− null lung buds (Fig. 4E). Vegf-C signals predominantly through Vegfr-3 and iscritical for early development of the lymphatic vasculature (Veikkola et al., 2001).Expression of Vegfr-3 was also analyzed in E12.5 control and Wnt2−/− null lung buds andQ-PCR showed a slight but not significant reduction in Vegfr-3 expression in the absence ofWnt2 function (Fig. 4F). Expression of Lyve-1, another marker of lymphatic endothelium(Banerji et al., 1999), was unchanged in Wnt2−/− null lungs at E12.5 (Fig. 4G). Together,these data indicate that the loss of Wnt2 signaling in the embryonic lung does notappreciably effect expression of most early endothelial marker genes with the exception ofminor decreases in Flk-1 and Vegf-C expression.
Expression of signaling pathways important for smooth muscle development in Wnt2−/−null lungs
Several signaling pathways are critical for smooth muscle development in the mouse lungincluding the Shh, Bmp, Wnt7b, and Fgf10 signaling pathways (reviewed in (Morrisey andHogan, 2010). Wnt2 is expressed in the primitive mesenchyme that is condensing around thetracheal bifurcation and presumptive lung bud (Goss et al., 2009), and Wnt2 signaling mayregulate smooth muscle development up or down-stream of any of these pathways. In situhybridization and Q-PCR on wild type and Wnt2−/− null mutant embryos showed thatWnt7b expression is reduced in Wnt2−/− null mutant foreguts (Fig. 5A–C). Fgf10 is a ligandthat is expressed in the developing lung mesenchyme and mediates lung bud outgrowth andASM development and has been shown to be a direct downstream target of canonical Wnt/β-catenin signaling (Cohen et al., 2007). Fgf10 expression in the mesenchyme of Wnt2−/−
null mutant lungs is reduced (Fig. 5D–F). Shh and Bmp4 are well-established epithelial-mesenchymal regulators of smooth muscle development in the lung during branchingmorphogenesis (Bellusci et al., 1997a; Bellusci et al., 1997b; Weaver et al., 2000; Weaver etal., 1999). In Shh−/− null mutant lungs there is a dramatic loss of smooth muscledevelopment (Bellusci et al., 1997a; Pepicelli et al., 1998), and activation of Bmp4 signalingbeen shown to be downstream of Fgf10 signaling (Mailleux et al., 2005). In situhybridization and Q-PCR show that Shh expression is not changed in Wnt2−/− null mutantlungs (Fig. 5G–I). Expression of Bmp4 is also unaltered in Wnt2−/− lungs (Fig. 5J–L).Taken together, these data suggests that Wnt2 signaling acts upstream of Wnt7b and Fgf10expression, but not Shh or Bmp4, in the primitive lung mesenchyme.
Wnt2 signaling is sufficient to promote smooth muscle gene expression and developmentin the lung
Although Wnt2−/− mutants display severe defects in ASM development, it is unclearwhether Wnt2 signaling is sufficient to promote smooth muscle differentiation anddevelopment in the lung mesenchyme. To test whether Wnt2 signaling was sufficient toactivate the smooth muscle differentiation program in mesenchymal cells, we treated themesenchymal cell line 10T1/2, a cell line known to differentiate into smooth muscle cellsunder certain culture conditions (Darland and D'Amore, 2001; Lockman et al., 2007; Wang
Goss et al. Page 5
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

et al., 2007b), with recombinant Wnt2 (rWnt2). Culturing 10T1/2 cells in the presence ofrWnt2 for 48 hours led to increased expression of several smooth muscle related genesincluding SM22α, SMA, Pdgfrα, and Pdgfrβ, as assessed by Q-PCR (Fig. 6A). Expression ofthe canonical Wnt target gene Axin2 was also significantly upregulated in the presence ofrWnt2, indicating activation of Wnt/β-catenin signaling in 10T1/2 cells by this recombinantligand (Fig. 6A).
To further investigate the effects of rWnt2 on smooth muscle development, E11.5 wild typelung buds were dissected and cultured for 48 hours with either control PBS or rWnt2 in alung explant assay. Explants were collected and examined for changes in smooth muscledevelopment by histology and Q-PCR. In lung bud explants cultured in the presence ofrWnt2, SM22α, myocardin, MRTFB, Fgf10, Pdgfrβ, and Pdgfrα expression were allsignificantly increased (Fig. 6B). This increase in smooth muscle gene expression wasaccompanied by increased smooth muscle cell development around the developing airwaysin the rWnt2 treated explants compared to control explants (Fig. 6C and D). Bmp4 andWnt7b expression were also examined by Q-PCR to determine whether rWnt2 activatedexpression of these pro-myogenic epithelial expressed factors. Interestingly in the presenceof rWnt2, expression of Wnt7b was significantly upregulated whereas Bmp4 expression wasnot significantly increased when compared to control wild type untreated lung bud explants(Fig. 6E). This data in conjunction with the decreased Wnt7b expression in Wnt2−/− nullmutants suggests that Wnt2 signaling coordinates smooth muscle development in partthrough activated Wnt7b signaling.
The whole lung bud explant assay does not permit the assessment of Wnt2 autocrinesignaling on activation of smooth muscle gene expression. To assess the role of autocrineWnt2 signaling in the lung mesenchyme in the absence of endoderm, primary cultures ofisolated E11.5 wild type lung mesenchyme were cultured in the absence or presence ofrWnt2 for 48 hours. In the presence of rWnt2, the expression of several smooth musclegenes were significantly upregulated including myocardin, MRTFB, Fgf10, SM22α, Pdgfrα,and SMA (Figure 6F). Surprisingly Fgf10 expression was increased 5-fold in the presence ofrWnt2, suggesting that Fgf10 signaling can be activated via autocrine Wnt2 signaling in theprimitive lung mesenchyme. Although activation of smooth muscle gene expressionoccurred in primary lung mesenchyme cultures treated with rWnt2, the overall magnitudewas less compared to whole lung bud explant cultures. These data suggest that Wnt2mediated paracrine signaling also contributes to smooth muscle gene expression anddevelopment in the lung.
Fgf10 signaling does not activate Wnt2 expression in the lungGenetic models have shown that Fgf10 signaling is necessary for ASM development in thelung (Mailleux et al., 2005; Ramasamy et al., 2007). Our data show that loss of Wnt2expression leads to decreased Fgf10 expression (Fig. 5D–F). However it is unclear whetherWnt2 simply acts upstream of Fgf10 signaling to promote smooth muscle development orwhether Fgf10 also cooperates to increase Wnt2 expression in a feed-forward loop. To testwhether Fgf10 signaling modulates Wnt2 expression, E11.5 wild type lung bud explantswere treated with either PBS or recombinant Fgf10 (rFgf10) and expression of Wnt2 wasanalyzed by Q-PCR. Wnt2 gene expression levels were unaffected by exogenous rFgf10 inwild type lung bud explants, suggesting that Fgf10 signaling neither activates nor inhibitsWnt2 expression (Fig. 7A). Treatment of lung bud explants with rFgf10 did result in asignificant upregulation of several smooth muscle marker genes including SM22α, SMA, andPdgfrα but not myocardin or Mrtf-B (Fig. 7A). This upregulation in smooth muscle geneexpression was confirmed by SM22α immunostaining showing increased ASM developmentin rFgf10 treated wild type explants (Fig. 7B and C). Previous reports examining Fgf10hypomorphic models suggested that Fgf10 regulates smooth muscle development through
Goss et al. Page 6
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

epithelial intermediates including Bmp4 (Mailleux et al. 2005). However, we did notobserve a significant increase in Bmp4 and Wnt7b expression in the presence of rFgf10(Supplemental Fig. 3).
Wnt2 regulates smooth muscle development through activation of Fgf10 signalingThe data thus far suggests that Wnt2 signaling mediates ASM development in part, throughactivation of Fgf10 signaling. To determine whether exogenous Fgf10 could rescue the lossof smooth muscle development in Wnt2−/− null mutant lungs, E11.5 lung buds weredissected from Wnt2+/− heterozygous mutant crosses for lung explant rescue assays withrFgf10. Lung bud explants were individually cultured in the presence of either PBS orrFgf10 for 48 hours, and subsequently harvested for histological sectioning and Q-PCR.rFgf10 treatment of Wnt2−/− null lung bud explants resulted in increased levels of SM22α,Pdgfrβ, and SMA (Fig. 7D). Expression of myocardin, Mrtf-B, and Pdgfrα was notstatistically changed (Fig. 7D). However, there was a less than significant rescue in Wnt7bexpression with addition of Fgf10 (Fig. 7D). Immunostaining for SM22α expression alsoshowed increased smooth muscle development in rFgf10 treated Wnt2−/− null lung budexplants supporting the Q-PCR findings (Fig. 7E–G). Overall, this data suggests that rFgf10acts downstream of Wnt2 to promote the expansion and differentiation of smooth musclecells in the lung that have already initiated expression of myocardin and Mrtf-B (Fig. 8).
DISCUSSIONOur data indicate that Wnt2 signaling is necessary and sufficient to promote thedifferentiation of ASM from the multipotent lung mesenchyme through direct or indirectactivation of a myogenic regulatory network including myocardin and Mrtf-B. These datashow that Wnt2 signaling activates Fgf10 signaling in the mesenchyme, which further drivesproliferation and smooth muscle differentiation into mature ASM. Our results also suggestthat Wnt2 signaling activates Wnt7b signaling in the adjacent epithelium, which is known tosignal in a paracrine manner to the sup-epithelial mesenchyme to promote smooth muscledifferentiation.
Smooth muscle development in the lung is poorly understood in part because the origins ofthe various smooth muscle lineages are still unclear and the pathways regulatingproliferation, migration, and differentiation of primitive smooth muscle cells are complexand unresolved. Recent genetic evidence demonstrates that Wnt/β-catenin signaling liesupstream of signals governing smooth muscle development in the mouse lung. Loss of thecanonical ligand Wnt7b in the embryonic lung epithelium leads to severe VSM defects,demonstrating a critical role for paracrine Wnt/β-catenin signaling between the developingepithelium and mesenchyme (Cohen et al., 2009; Shu et al., 2002). Additional studiesdemonstrate that genetic deletion of β-catenin in the primitive lung mesenchyme alsodisrupts ASM development (De Langhe et al., 2008), implicating a potential function forother canonical Wnt ligands expressed in the lung for proper ASM development. In thisstudy, we identify Wnt2 as a critical Wnt ligand required in the lung mesenchyme tomediate ASM development. We show that Wnt2 is required for BAT-GAL activity in ASMindicating that it acts through the β-catenin dependent canonical pathway to regulate ASMdevelopment.
While defects in ASM are observed Wnt2−/− null mutants, the VSM appears grossly normal.Wnt7b signaling from the lung epithelium has been shown to regulate VSM development(Cohen et al., 2009; Shu et al., 2002), and analysis of Wnt2−/− null mutant lungsdemonstrates that Wnt7b is expressed at about 50% of normal levels in Wnt2−/− nullmutants. Additionally, Wnt2 expression is reduced in Wnt7b−/− null lungs (unpublishedobservations), suggesting there is signaling interplay between epithelial and mesenchymal
Goss et al. Page 7
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

expressed Wnt ligands in the lung. If Wnt2 signaling is important for VSM development, thepotential effects from loss of Wnt2 signaling may be compensated by the continuedexpression of Wnt7b or Wnt2b, which is also expressed in the lung mesenchyme and couldfunction redundantly with Wnt2 signaling in VSM development (Goss et al., 2009). Thedistinct molecular pathways that regulate VSM versus ASM development in the lung arepoorly understood. Moreover, it is unclear whether ASM and VSM have a common ordistinct developmental origins. Disruption of other pathways including Shh result in defectsin both ASM and VSM (Miller et al., 2004). Thus, additional fate-mapping as well asdevelopmental studies will be required to determine the similarities and differences in VSMand ASM origins and cellular phenotypes.
A recent study describing conditional mutants in β-catenin signaling in the developing lungmesenchyme suggested a role for Wnt/β-catenin in the differentiation of the endotheliallineage (De Langhe et al., 2008). Wnt2−/− null embryoid bodies are also unable toeffectively differentiate into endothelial cells (Wang et al., 2007a). Our data suggests thateither Wnt2 is not required for lung endothelial development or that it acts redundantly withother Wnt ligands to regulate endothelial development in the lung.
Wnt2 is expressed during the earliest stages of lung morphogenesis suggesting the perinatalASM deficiency in Wnt2−/− null mutants could be the result of an early defect in thedevelopment of smooth muscle cells in the distal mesenchyme. Analysis of Wnt2−/− nullmutants during the early stages of myogenesis indicated reduced expression of the genesregulating the earliest aspects of smooth muscle cell differentiation from the primitivemesenchyme including myocardin, Mrtf-B, and Pdgfrβ These data indicate that Wnt2signaling regulates the development of the multipotent mesenchymal into immature smoothmuscle cells in the mouse lung.
A Wnt-Fgf regulatory network is thought to govern development of the lung mesenchyme(Yin et al., 2008), and mice carrying hypomorphic or null alleles of Fgf10 and Fgfr2, thecognate receptor of Fgf10, exhibit ASM deficiencies (De Langhe et al., 2006; Ramasamy etal., 2007). In Wnt2−/− null mutant lungs, both Fgf10 expression and Fgfr2 expression arereduced, implicating a Wnt2-Fgf10 signaling network in the coordination of ASMdevelopment. We show that Fgf10 signaling promotes ASM development throughupregulation of several mature smooth muscle markers but not the early smooth muscledifferentiation factors myocardin or Mrtf-B. These findings suggest that Fgf10 signalingmediates the expansion and differentiation of immature smooth muscle cell alreadycommitted to the ASM lineage.
In summary, these studies demonstrate that Wnt2 signaling is necessary for ASMdevelopment in the mouse lung. Wnt2 expression during early lung morphogenesis isrequired for activation of the smooth muscle gene expression program including myocardin,Mrtf-B, and Fgf10. Exogenous Fgf10 can partly rescue the loss of Wnt2 by promotingdifferentiation of cells already committed to the ASM lineage. Thus, these results define aWnt2-Fgf10 signaling axis governing early ASM development in the lung.
MATERIALS AND METHODSMice
Generation and genotyping information on the Wnt2−/− mutant mice has been described(Goss et al., 2009). Genotyping of the BAT-GAL mice has been described (Maretto et al.,2003). The University of Pennsylvania Institutional Animal Care and Use Committeeapproved all animal protocols.
Goss et al. Page 8
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

HistologyEmbryos were fixed in 4% paraformaldehyde for 24 hours, dehydrated in a series of ethanolwashes, and then embedded in paraffin for tissue sectioning. Dissected embryonic lung budsand foreguts were fixed in 4% paraformaldehyde overnight, dehydrated in a series ofmethanol washes and blocked a 10% normal goat serum/PBS solution before wholemountimmunostaining. Radioactive in situ hybridization and immunohistochemistry wereperformed as previously described (Shu et al., 2001). Tissue sections were stained with thefollowing antibodies and dilutions: anti-Axin2 (Abcam; 1:100), anti-Pdgfrβ (Cell SignalingTechnology Inc.; 1:50), anti-PECAM-1 (PharMingen; 1:500), and anti-SM22α (Abcam;1:500). β-galactosidase histochemical staining of embryos was performed as previouslydescribed by our laboratory (Shu et al., 2002).
Quantitative RT-PCRTotal RNA was isolated from lung tissue at the indicated time points using Trizol reagent,reverse transcribed using SuperScript First Strand Synthesis System (Invitrogen), and usedin quantitative real time PCR (Q-PCR) analysis using the oligonucleotides listed inSupplemental Table 1. RNA was isolated from at least 5 lung bud tissue samplescorresponding to each genotype.
Lung explant and primary mesenchyme culturesLung buds were dissected from E11.5 embryos and cultured in BGjb media (Invitrogen)supplemented with 0.1mg/ml ascorbic acid for 48 hours as previously described (Zhang etal., 2008). Explants were cultured on a 0.4μm membrane filter (BD Falcon). Formesenchymal cultures, whole lung buds were dissected at E11.5 and subjected tocollagenase digestion to give rise to single cells and mesenchymal cells were isolated viadifferential adhesion. For rescue experiments, embryos were collected from wild-typefemales or generated from Wnt2+/− heterozygous mutant crosses. Exogenous recombinanthuman Wnt2 protein (Novus Biologicals) was added to the lung explant media at aconcentration of 0.15ug/ml. Exogenous recombinant human Fgf10 (R & D Systems, Inc.)was added to the lung explant media at a concentration of 0.2ug/ml. At least 8 explants wereused for each experimental condition and treatment.
Cell Culture10T1/2 cells were cultured in Eagle's Basal medium with Earle's BSS, 2 mM L-glutamine,1.5 g/L sodium bicarbonate, and 10% heat-inactivated fetal bovine serum as suggested byATCC. Recombinant human Wnt2 protein was added to the 10T1/2 media at a concentrationof 0.15ug/ml, and cultured for 48 hours before harvesting for Q-PCR analysis.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsThese studies were supported by funding from the NIH (HL087825 and HL100405 to E.E.M.) and a PredoctoralFellowship from the American Heart Association to A.M.G..
REFERENCESBadri KR, Zhou Y, Schuger L. Embryological origin of airway smooth muscle. Proc Am Thorac Soc.
2008; 5:4–10. [PubMed: 18094078]
Goss et al. Page 9
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Banerji S, Ni J, Wang SX, Clasper S, Su J, Tammi R, Jones M, Jackson DG. LYVE-1, a newhomologue of the CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J Cell Biol.1999; 144:789–801. [PubMed: 10037799]
Bellusci S, Furuta Y, Rush MG, Henderson R, Winnier G, Hogan BL. Involvement of Sonic hedgehog(Shh) in mouse embryonic lung growth and morphogenesis. Development. 1997a; 124:53–63.[PubMed: 9006067]
Bellusci S, Grindley J, Emoto H, Itoh N, Hogan BL. Fibroblast growth factor 10 (FGF10) andbranching morphogenesis in the embryonic mouse lung. Development. 1997b; 124:4867–4878.[PubMed: 9428423]
Betsholtz C. Insight into the physiological functions of PDGF through genetic studies in mice.Cytokine Growth Factor Rev. 2004; 15:215–228. [PubMed: 15207813]
Bostrom H, Willetts K, Pekny M, Leveen P, Lindahl P, Hedstrand H, Pekna M, Hellstrom M, Gebre-Medhin S, Schalling M, Nilsson M, Kurland S, Tornell J, Heath JK, Betsholtz C. PDGF-A signalingis a critical event in lung alveolar myofibroblast development and alveogenesis. Cell. 1996; 85:863–873. [PubMed: 8681381]
Camoretti-Mercado B, Dulin NO, Solway J. Serum response factor function and dysfunction in smoothmuscle. Respir Physiol Neurobiol. 2003; 137:223–235. [PubMed: 14516728]
Chinoy MR. Lung growth and development. Front Biosci. 2003; 8:d392–415. [PubMed: 12456356]Cohen ED, Ihida-Stansbury K, Lu MM, Panettieri RA, Jones PL, Morrisey EE. Wnt signaling
regulates smooth muscle precursor development in the mouse lung via a tenascin C/PDGFRpathway. J Clin Invest. 2009; 119:2538–2549. [PubMed: 19690384]
Cohen ED, Wang Z, Lepore JJ, Lu MM, Taketo MM, Epstein DJ, Morrisey EE. Wnt/beta-cateninsignaling promotes expansion of Isl-1-positive cardiac progenitor cells through regulation of FGFsignaling. J Clin Invest. 2007; 117:1794–1804. [PubMed: 17607356]
Colvin JS, White AC, Pratt SJ, Ornitz DM. Lung hypoplasia and neonatal death in Fgf9-null miceidentify this gene as an essential regulator of lung mesenchyme. Development. 2001; 128:2095–2106. [PubMed: 11493531]
Darland DC, D'Amore PA. TGF beta is required for the formation of capillary-like structures in three-dimensional cocultures of 10T1/2 and endothelial cells. Angiogenesis. 2001; 4:11–20. [PubMed:11824373]
De Langhe SP, Carraro G, Tefft D, Li C, Xu X, Chai Y, Minoo P, Hajihosseini MK, Drouin J,Kaartinen V, Bellusci S. Formation and differentiation of multiple mesenchymal lineages duringlung development is regulated by beta-catenin signaling. PLoS One. 2008; 3:e1516. [PubMed:18231602]
De Langhe SP, Carraro G, Warburton D, Hajihosseini MK, Bellusci S. Levels of mesenchymal FGFR2signaling modulate smooth muscle progenitor cell commitment in the lung. Dev Biol. 2006;299:52–62. [PubMed: 16989802]
De Langhe SP, Sala FG, Del Moral PM, Fairbanks TJ, Yamada KM, Warburton D, Burns RC, BellusciS. Dickkopf-1 (DKK1) reveals that fibronectin is a major target of Wnt signaling in branchingmorphogenesis of the mouse embryonic lung. Dev Biol. 2005; 277:316–331. [PubMed: 15617677]
Goss AM, Tian Y, Tsukiyama T, Cohen ED, Zhou D, Lu MM, Yamaguchi TP, Morrisey EE. Wnt2/2band beta-catenin signaling are necessary and sufficient to specify lung progenitors in the foregut.Dev Cell. 2009; 17:290–298. [PubMed: 19686689]
Halayko AJ, Rector E, Stephens NL. Airway smooth muscle cell proliferation: characterization ofsubpopulations by sensitivity to heparin inhibition. Am J Physiol. 1998; 274:L17–25. [PubMed:9458796]
Halayko AJ, Stephens NL. Potential role for phenotypic modulation of bronchial smooth muscle cellsin chronic asthma. Can J Physiol Pharmacol. 1994; 72:1448–1457. [PubMed: 7767892]
Hellstrom M, Kalen M, Lindahl P, Abramsson A, Betsholtz C. Role of PDGF-B and PDGFR-beta inrecruitment of vascular smooth muscle cells and pericytes during embryonic blood vesselformation in the mouse. Development. 1999; 126:3047–3055. [PubMed: 10375497]
Humbert M, Morrell NW, Archer SL, Stenmark KR, MacLean MR, Lang IM, Christman BW, WeirEK, Eickelberg O, Voelkel NF, Rabinovitch M. Cellular and molecular pathobiology ofpulmonary arterial hypertension. J Am Coll Cardiol. 2004; 43:13S–24S. [PubMed: 15194174]
Goss et al. Page 10
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Jho EH, Zhang T, Domon C, Joo CK, Freund JN, Costantini F. Wnt/beta-catenin/Tcf signaling inducesthe transcription of Axin2, a negative regulator of the signaling pathway. Mol Cell Biol. 2002;22:1172–1183. [PubMed: 11809808]
Karkkainen MJ, Haiko P, Sainio K, Partanen J, Taipale J, Petrova TV, Jeltsch M, Jackson DG, TalikkaM, Rauvala H, Betsholtz C, Alitalo K. Vascular endothelial growth factor C is required forsprouting of the first lymphatic vessels from embryonic veins. Nat Immunol. 2004; 5:74–80.[PubMed: 14634646]
Lockman K, Taylor JM, Mack CP. The histone demethylase, Jmjd1a, interacts with the myocardinfactors to regulate SMC differentiation marker gene expression. Circ Res. 2007; 101:e115–123.[PubMed: 17991879]
Lohela M, Bry M, Tammela T, Alitalo K. VEGFs and receptors involved in angiogenesis versuslymphangiogenesis. Curr Opin Cell Biol. 2009; 21:154–165. [PubMed: 19230644]
Mailleux AA, Kelly R, Veltmaat JM, De Langhe SP, Zaffran S, Thiery JP, Bellusci S. Fgf10expression identifies parabronchial smooth muscle cell progenitors and is required for their entryinto the smooth muscle cell lineage. Development. 2005; 132:2157–2166. [PubMed: 15800000]
Maretto S, Cordenonsi M, Dupont S, Braghetta P, Broccoli V, Hassan AB, Volpin D, Bressan GM,Piccolo S. Mapping Wnt/beta-catenin signaling during mouse development and in colorectaltumors. Proc Natl Acad Sci U S A. 2003; 100:3299–3304. [PubMed: 12626757]
Miller LA, Wert SE, Clark JC, Xu Y, Perl AK, Whitsett JA. Role of Sonic hedgehog in patterning oftracheal-bronchial cartilage and the peripheral lung. Dev Dyn. 2004; 231:57–71. [PubMed:15305287]
Monkley SJ, Delaney SJ, Pennisi DJ, Christiansen JH, Wainwright BJ. Targeted disruption of theWnt2 gene results in placentation defects. Development. 1996; 122:3343–3353. [PubMed:8951051]
Morrisey EE, Hogan BL. Preparing for the first breath: genetic and cellular mechanisms in lungdevelopment. Dev Cell. 2010; 18:8–23. [PubMed: 20152174]
Okubo T, Hogan BL. Hyperactive Wnt signaling changes the developmental potential of embryoniclung endoderm. J Biol. 2004; 3:11. [PubMed: 15186480]
Pepicelli CV, Lewis PM, McMahon AP. Sonic hedgehog regulates branching morphogenesis in themammalian lung. Curr Biol. 1998; 8:1083–1086. [PubMed: 9768363]
Ramasamy SK, Mailleux AA, Gupte VV, Mata F, Sala FG, Veltmaat JM, Del Moral PM, De LangheS, Parsa S, Kelly LK, Kelly R, Shia W, Keshet E, Minoo P, Warburton D, Bellusci S. Fgf10dosage is critical for the amplification of epithelial cell progenitors and for the formation ofmultiple mesenchymal lineages during lung development. Dev Biol. 2007; 307:237–247.[PubMed: 17560563]
Shu W, Guttentag S, Wang Z, Andl T, Ballard P, Lu MM, Piccolo S, Birchmeier W, Whitsett JA,Millar SE, Morrisey EE. Wnt/beta-catenin signaling acts upstream of N-myc, BMP4, and FGFsignaling to regulate proximal-distal patterning in the lung. Dev Biol. 2005; 283:226–239.[PubMed: 15907834]
Shu W, Jiang YQ, Lu MM, Morrisey EE. Wnt7b regulates mesenchymal proliferation and vasculardevelopment in the lung. Development. 2002; 129:4831–4842. [PubMed: 12361974]
Shu W, Yang H, Zhang L, Lu MM, Morrisey EE. Characterization of a new subfamily of winged-helix/forkhead (Fox) genes that are expressed in the lung and act as transcriptional repressors. JBiol Chem. 2001; 276:27488–27497. [PubMed: 11358962]
Solway J, Forsythe SM, Halayko AJ, Vieira JE, Hershenson MB, Camoretti-Mercado B.Transcriptional regulation of smooth muscle contractile apparatus expression. Am J Respir CritCare Med. 1998; 158:S100–108. [PubMed: 9817732]
Solway J, Seltzer J, Samaha FF, Kim S, Alger LE, Niu Q, Morrisey EE, Ip HS, Parmacek MS.Structure and expression of a smooth muscle cell-specific gene, SM22 alpha. J Biol Chem. 1995;270:13460–13469. [PubMed: 7768949]
Thebaud B. Angiogenesis in lung development, injury and repair: implications for chronic lung diseaseof prematurity. Neonatology. 2007; 91:291–297. [PubMed: 17575472]
Goss et al. Page 11
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Tollet J, Everett AW, Sparrow MP. Spatial and temporal distribution of nerves, ganglia, and smoothmuscle during the early pseudoglandular stage of fetal mouse lung development. Dev Dyn. 2001;221:48–60. [PubMed: 11357193]
Tollet J, Everett AW, Sparrow MP. Development of neural tissue and airway smooth muscle in fetalmouse lung explants: a role for glial-derived neurotrophic factor in lung innervation. Am J RespirCell Mol Biol. 2002; 26:420–429. [PubMed: 11919078]
Veikkola T, Jussila L, Makinen T, Karpanen T, Jeltsch M, Petrova TV, Kubo H, Thurston G,McDonald DM, Achen MG, Stacker SA, Alitalo K. Signalling via vascular endothelial growthfactor receptor-3 is sufficient for lymphangiogenesis in transgenic mice. EMBO J. 2001; 20:1223–1231. [PubMed: 11250889]
Wang D, Chang PS, Wang Z, Sutherland L, Richardson JA, Small E, Krieg PA, Olson EN. Activationof cardiac gene expression by myocardin, a transcriptional cofactor for serum response factor.Cell. 2001; 105:851–862. [PubMed: 11439182]
Wang DZ, Olson EN. Control of smooth muscle development by the myocardin family oftranscriptional coactivators. Curr Opin Genet Dev. 2004; 14:558–566. [PubMed: 15380248]
Wang H, Gilner JB, Bautch VL, Wang DZ, Wainwright BJ, Kirby SL, Patterson C. Wnt2 coordinatesthe commitment of mesoderm to hematopoietic, endothelial, and cardiac lineages in embryoidbodies. J Biol Chem. 2007a; 282:782–791. [PubMed: 17098737]
Wang J, Li A, Wang Z, Feng X, Olson EN, Schwartz RJ. Myocardin sumoylation transactivatescardiogenic genes in pluripotent 10T1/2 fibroblasts. Mol Cell Biol. 2007b; 27:622–632. [PubMed:17101795]
Wang Z, Wang DZ, Pipes GC, Olson EN. Myocardin is a master regulator of smooth muscle geneexpression. Proc Natl Acad Sci U S A. 2003; 100:7129–7134. [PubMed: 12756293]
Weaver M, Dunn NR, Hogan BL. Bmp4 and Fgf10 play opposing roles during lung budmorphogenesis. Development. 2000; 127:2695–2704. [PubMed: 10821767]
Weaver M, Yingling JM, Dunn NR, Bellusci S, Hogan BL. Bmp signaling regulates proximal-distaldifferentiation of endoderm in mouse lung development. Development. 1999; 126:4005–4015.[PubMed: 10457010]
Zhang Y, Goss AM, Cohen ED, Kadzik R, Lepore JJ, Muthukumaraswamy K, Yang J, DeMayo FJ,Whitsett JA, Parmacek MS, Morrisey EE. A Gata6-Wnt pathway required for epithelial stem celldevelopment and airway regeneration. Nat Genet. 2008; 40:862–870. [PubMed: 18536717]
Goss et al. Page 12
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Highlights
- We show that Wnt2 signaling is required for lung smooth muscledevelopment
- Wnt2 acts by direct regulation of Fgf10 and acts to promote myocardin/Mrtf-B expression
- Wnt2 activates Wnt7b expression in the lung endoderm
Goss et al. Page 13
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
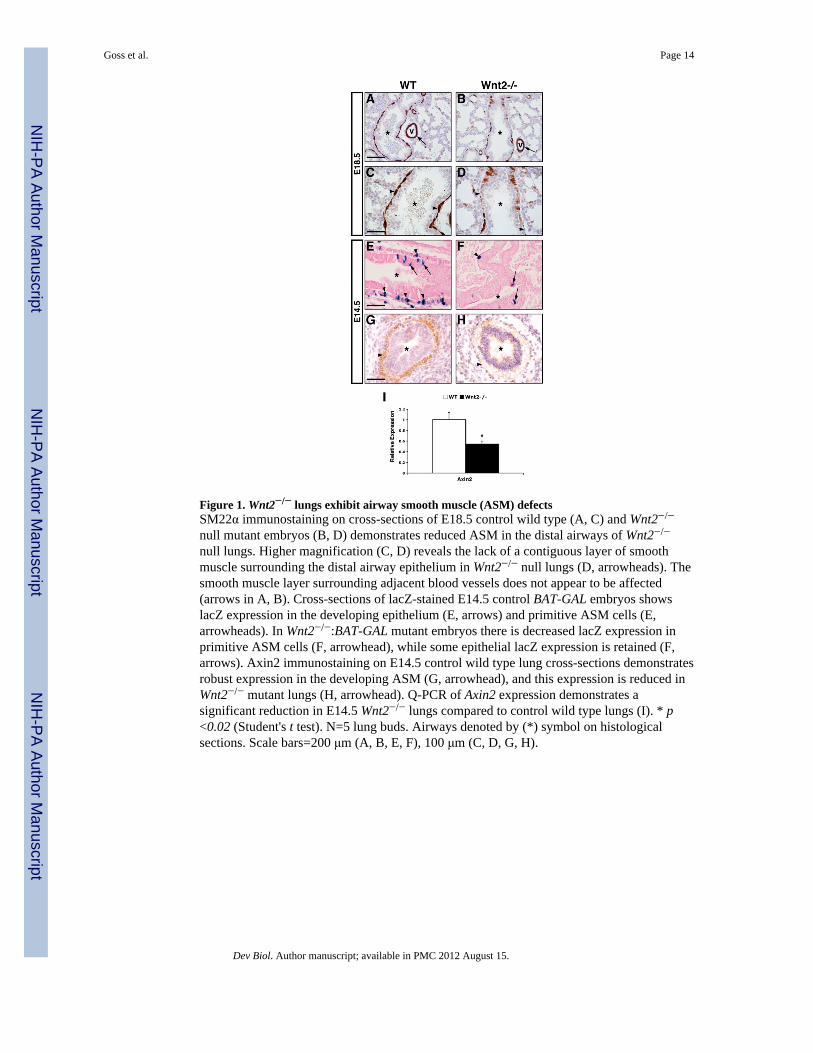
Figure 1. Wnt2−/− lungs exhibit airway smooth muscle (ASM) defectsSM22α immunostaining on cross-sections of E18.5 control wild type (A, C) and Wnt2−/−
null mutant embryos (B, D) demonstrates reduced ASM in the distal airways of Wnt2−/−
null lungs. Higher magnification (C, D) reveals the lack of a contiguous layer of smoothmuscle surrounding the distal airway epithelium in Wnt2−/− null lungs (D, arrowheads). Thesmooth muscle layer surrounding adjacent blood vessels does not appear to be affected(arrows in A, B). Cross-sections of lacZ-stained E14.5 control BAT-GAL embryos showslacZ expression in the developing epithelium (E, arrows) and primitive ASM cells (E,arrowheads). In Wnt2−/−:BAT-GAL mutant embryos there is decreased lacZ expression inprimitive ASM cells (F, arrowhead), while some epithelial lacZ expression is retained (F,arrows). Axin2 immunostaining on E14.5 control wild type lung cross-sections demonstratesrobust expression in the developing ASM (G, arrowhead), and this expression is reduced inWnt2−/− mutant lungs (H, arrowhead). Q-PCR of Axin2 expression demonstrates asignificant reduction in E14.5 Wnt2−/− lungs compared to control wild type lungs (I). * p<0.02 (Student's t test). N=5 lung buds. Airways denoted by (*) symbol on histologicalsections. Scale bars=200 μm (A, B, E, F), 100 μm (C, D, G, H).
Goss et al. Page 14
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
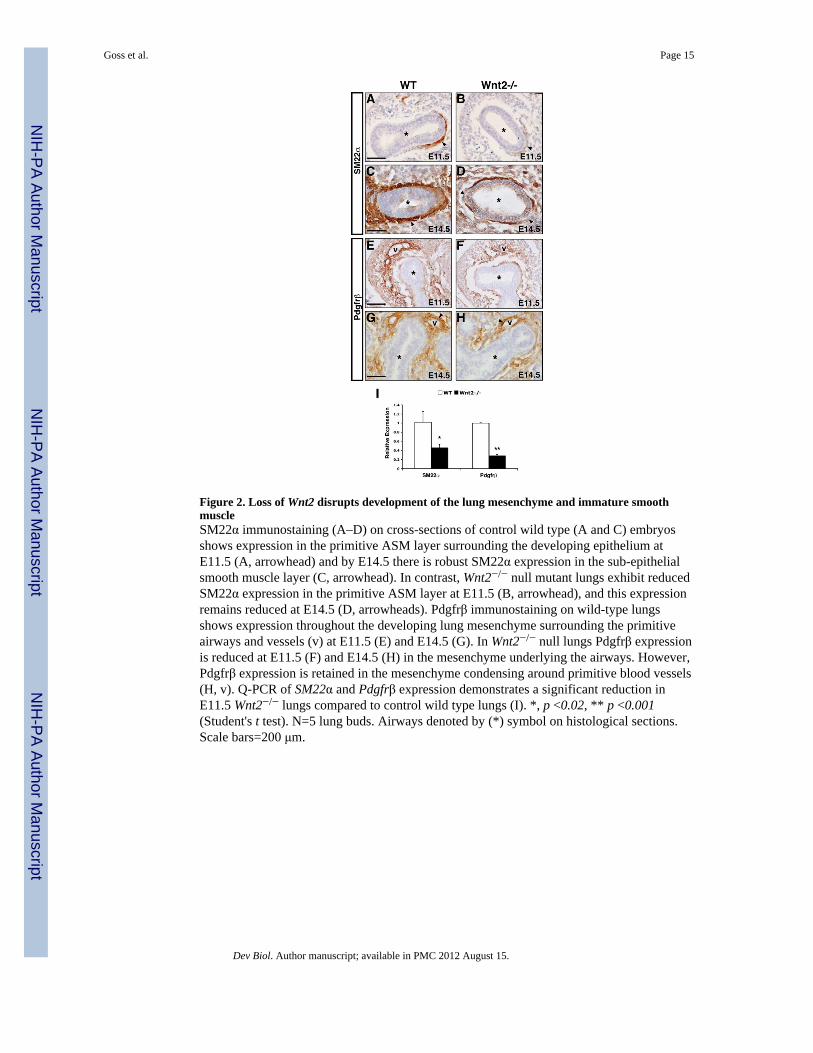
Figure 2. Loss of Wnt2 disrupts development of the lung mesenchyme and immature smoothmuscleSM22α immunostaining (A–D) on cross-sections of control wild type (A and C) embryosshows expression in the primitive ASM layer surrounding the developing epithelium atE11.5 (A, arrowhead) and by E14.5 there is robust SM22α expression in the sub-epithelialsmooth muscle layer (C, arrowhead). In contrast, Wnt2−/− null mutant lungs exhibit reducedSM22α expression in the primitive ASM layer at E11.5 (B, arrowhead), and this expressionremains reduced at E14.5 (D, arrowheads). Pdgfrβ immunostaining on wild-type lungsshows expression throughout the developing lung mesenchyme surrounding the primitiveairways and vessels (v) at E11.5 (E) and E14.5 (G). In Wnt2−/− null lungs Pdgfrβ expressionis reduced at E11.5 (F) and E14.5 (H) in the mesenchyme underlying the airways. However,Pdgfrβ expression is retained in the mesenchyme condensing around primitive blood vessels(H, v). Q-PCR of SM22α and Pdgfrβ expression demonstrates a significant reduction inE11.5 Wnt2−/− lungs compared to control wild type lungs (I). *, p <0.02, ** p <0.001(Student's t test). N=5 lung buds. Airways denoted by (*) symbol on histological sections.Scale bars=200 μm.
Goss et al. Page 15
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
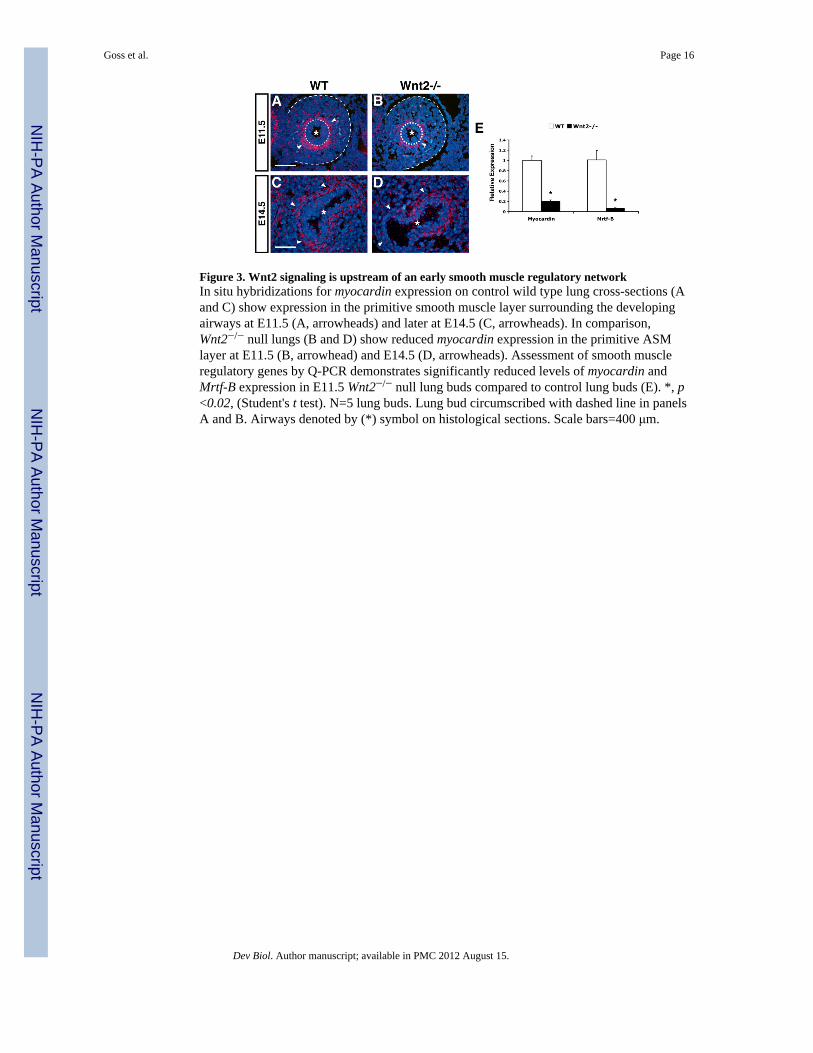
Figure 3. Wnt2 signaling is upstream of an early smooth muscle regulatory networkIn situ hybridizations for myocardin expression on control wild type lung cross-sections (Aand C) show expression in the primitive smooth muscle layer surrounding the developingairways at E11.5 (A, arrowheads) and later at E14.5 (C, arrowheads). In comparison,Wnt2−/− null lungs (B and D) show reduced myocardin expression in the primitive ASMlayer at E11.5 (B, arrowhead) and E14.5 (D, arrowheads). Assessment of smooth muscleregulatory genes by Q-PCR demonstrates significantly reduced levels of myocardin andMrtf-B expression in E11.5 Wnt2−/− null lung buds compared to control lung buds (E). *, p<0.02, (Student's t test). N=5 lung buds. Lung bud circumscribed with dashed line in panelsA and B. Airways denoted by (*) symbol on histological sections. Scale bars=400 μm.
Goss et al. Page 16
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
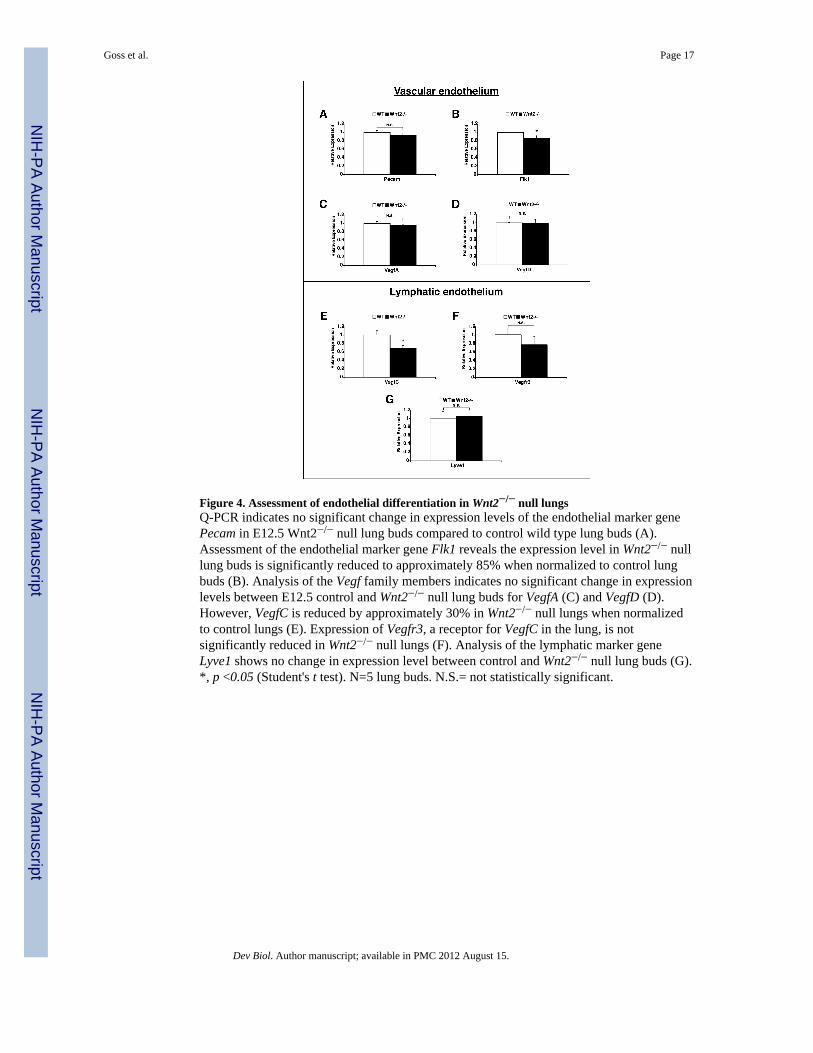
Figure 4. Assessment of endothelial differentiation in Wnt2−/− null lungsQ-PCR indicates no significant change in expression levels of the endothelial marker genePecam in E12.5 Wnt2−/− null lung buds compared to control wild type lung buds (A).Assessment of the endothelial marker gene Flk1 reveals the expression level in Wnt2−/− nulllung buds is significantly reduced to approximately 85% when normalized to control lungbuds (B). Analysis of the Vegf family members indicates no significant change in expressionlevels between E12.5 control and Wnt2−/− null lung buds for VegfA (C) and VegfD (D).However, VegfC is reduced by approximately 30% in Wnt2−/− null lungs when normalizedto control lungs (E). Expression of Vegfr3, a receptor for VegfC in the lung, is notsignificantly reduced in Wnt2−/− null lungs (F). Analysis of the lymphatic marker geneLyve1 shows no change in expression level between control and Wnt2−/− null lung buds (G).*, p <0.05 (Student's t test). N=5 lung buds. N.S.= not statistically significant.
Goss et al. Page 17
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. Evaluation of signaling factors important for smooth muscle development in Wnt2−/−
null lungsIn situ hybridization on wild-type (A, D, G, J) and Wnt2−/− null mutant embryos (B, E, H,K) to detect expression of Wnt7b (A and B), Fgf10 (D and E), Shh (G and H), and Bmp4 (Jand K) at E10.5 (A and B) and E11.5 (D,E,G,H,J,K). Q-PCR to detect expression of Wnt7b(C), Fgf10 (F), Shh (I), and Bmp4 (L) in wild type and Wnt2−/− null mutant lung buds atE11.5. *, p < 0.02, ** p <0.001 (Student's t test). N=5 lung buds. N.S.= not statisticallysignificant. Lung bud circumscribed with dashed line in panels D, E, J, and K. Primitiveairway denoted by (*) symbol on histological sections. Scale bars=100 μm (A–K).
Goss et al. Page 18
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
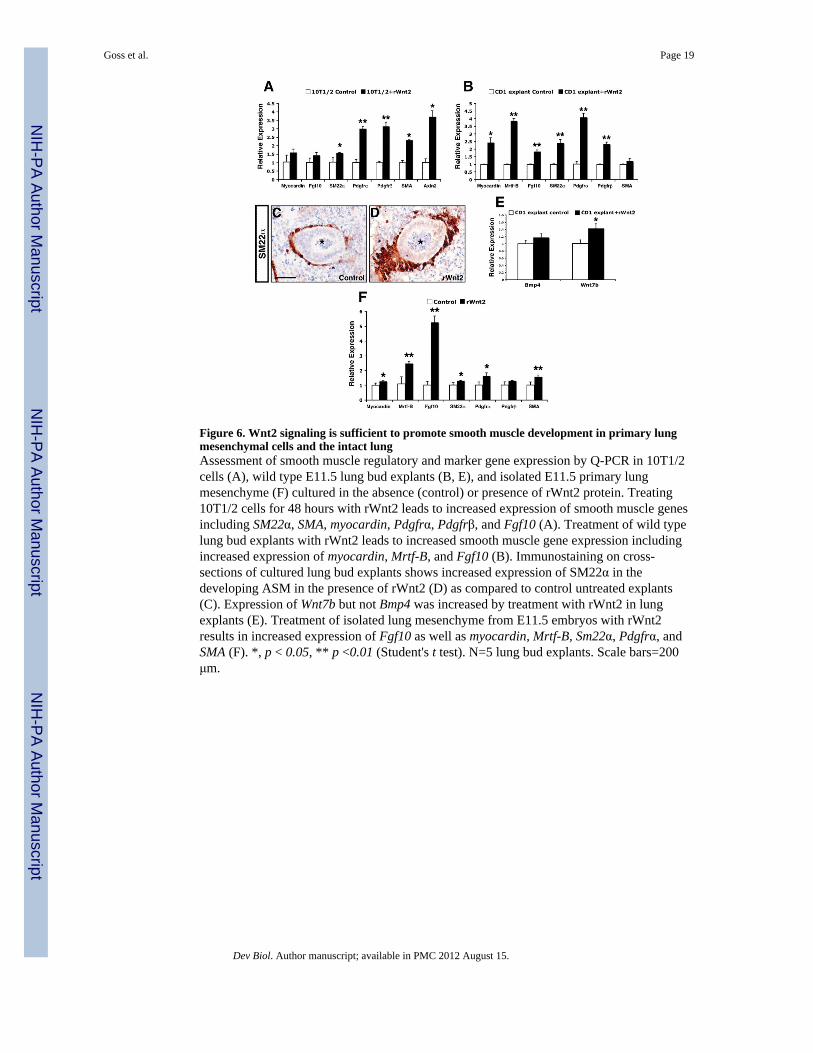
Figure 6. Wnt2 signaling is sufficient to promote smooth muscle development in primary lungmesenchymal cells and the intact lungAssessment of smooth muscle regulatory and marker gene expression by Q-PCR in 10T1/2cells (A), wild type E11.5 lung bud explants (B, E), and isolated E11.5 primary lungmesenchyme (F) cultured in the absence (control) or presence of rWnt2 protein. Treating10T1/2 cells for 48 hours with rWnt2 leads to increased expression of smooth muscle genesincluding SM22α, SMA, myocardin, Pdgfrα, Pdgfrβ, and Fgf10 (A). Treatment of wild typelung bud explants with rWnt2 leads to increased smooth muscle gene expression includingincreased expression of myocardin, Mrtf-B, and Fgf10 (B). Immunostaining on cross-sections of cultured lung bud explants shows increased expression of SM22α in thedeveloping ASM in the presence of rWnt2 (D) as compared to control untreated explants(C). Expression of Wnt7b but not Bmp4 was increased by treatment with rWnt2 in lungexplants (E). Treatment of isolated lung mesenchyme from E11.5 embryos with rWnt2results in increased expression of Fgf10 as well as myocardin, Mrtf-B, Sm22α, Pdgfrα, andSMA (F). *, p < 0.05, ** p <0.01 (Student's t test). N=5 lung bud explants. Scale bars=200μm.
Goss et al. Page 19
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 7. Wnt2 signaling regulates smooth muscle development through activation ofdownstream Fgf10 signalingAssessment of smooth muscle gene expression by Q-PCR in wild type lung bud explantstreated with either PBS (control) or rFgf10 (A). Treating wild type lung bud explants withrFgf10 leads to increased expression of smooth muscle genes including SM22α, SMA, andPdgfrα (A). SM22α immunostaining on cross-sections of lung bud explants treated withrFgf10 shows increased expression in the developing ASM as compared to control untreatedlung bud explants (B and C). Assessment of smooth muscle gene expression by quantitativePCR in cultured E11.5 Wnt2+/+ wild type and Wnt2−/− null lung bud explants (D).Expression levels of SM22α, Pdgfrβ, and SMA expression levels were significantlyupregulated in rFgf10-treated Wnt2−/− null lung bud explants as compared to untreatedWnt2−/− null lung bud explants while Wnt7b was increased to a non-significant level (D).Wnt2 expression levels were also measured to confirm the genotyping of lung bud explantpopulations (D). SM22α immunostaining on cross-sections of control Wnt2−/− null lung budexplants shows reduced expression in the primitive ASM as compared to Wnt2+/+ wild typeexplants (E and F). In the presence of rFgf10, there is increased SM22α expression in thedeveloping ASM, similar to that in Wnt2+/+ wild type lung bud explants (compare E and G).*, p < 0.05 (Student's t test). N=8 lung bud explants. n.s.=not significant. Scale bars=100μm.
Goss et al. Page 20
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
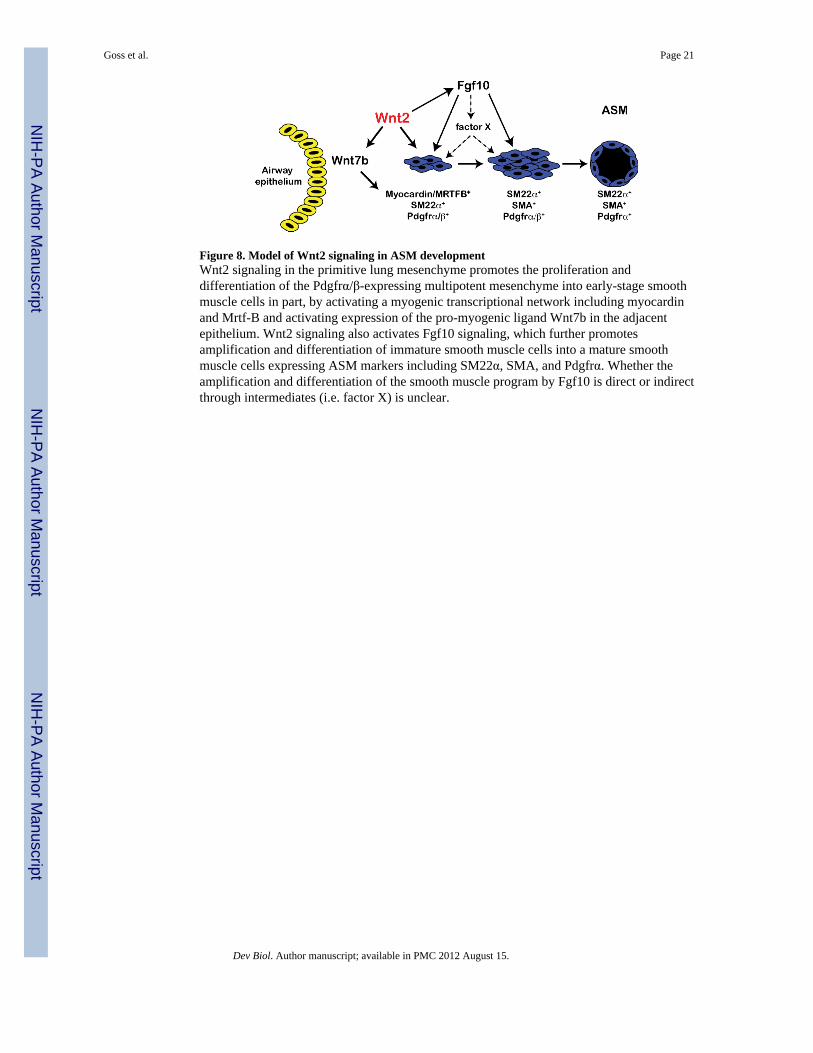
Figure 8. Model of Wnt2 signaling in ASM developmentWnt2 signaling in the primitive lung mesenchyme promotes the proliferation anddifferentiation of the Pdgfrα/β-expressing multipotent mesenchyme into early-stage smoothmuscle cells in part, by activating a myogenic transcriptional network including myocardinand Mrtf-B and activating expression of the pro-myogenic ligand Wnt7b in the adjacentepithelium. Wnt2 signaling also activates Fgf10 signaling, which further promotesamplification and differentiation of immature smooth muscle cells into a mature smoothmuscle cells expressing ASM markers including SM22α, SMA, and Pdgfrα. Whether theamplification and differentiation of the smooth muscle program by Fgf10 is direct or indirectthrough intermediates (i.e. factor X) is unclear.
Goss et al. Page 21
Dev Biol. Author manuscript; available in PMC 2012 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript












![The Necessary and Sufficient Condition of Military Defection for the Occurrence of Revolution, as Evident in the Arab Spring Uprisings[2012]](https://static.fdokumen.com/doc/165x107/631e9af1dc32ad07f307a996/the-necessary-and-sufficient-condition-of-military-defection-for-the-occurrence.jpg)






