
Where on earth can animals use a geomagnetic bi‐coordinate map for navigation?
9
1039 A bi-coordinate map sense based on magnetic information for homing animals was first suggested almost 130 yr ago (Viguier 1882). e idea is that animals may be able to use two different magnetic field parameters to determine their position in relation to a known goal. It has been pointed out that such a map would not necessarily function as our view of a printed map (Lohmann and Lohmann 2006). Instead animals familiar with the magnetic conditions in their goal area may be able to use the isolines of two different mag- netic parameters alternately to approach the goal as long as the two parameters are not parallel (Åkesson et al. 2003, Lohmann and Lohmann 2006). Map-based navigation, or true navigation, has been defined as the ability to determine spatial position in relation to a known goal, based only on information available at a distant release location (Griffin 1952, Able 2001). An animal that has learnt how two gradients vary in a familiar area, can extrapolate the gradi- ents to determine its position in relation to home or a known goal when displaced away from its familiar area, as long as the gradients vary in a consistent and predictable manner (Wallraff 1974, Wiltschko and Wiltschko 2003). Several papers have investigated the possibility of magnetic map use for navigation in different animals, e.g. newts (Fischer et al. 2001), alligators (Rodda 1984a), sea turtles (Lohmann and Lohmann 1994, 1996, Lohmann et al. 2001, 2004, Putman et al. 2011) and birds (Beck and Wiltschko 1988, Munro et al. 1997, Åkesson and Alerstam 1998, Fischer et al. 2003). In many of these studies animals were displaced in geomagnetic (but not in geographic) space and they showed a response in orientation or fuel deposi- tion behaviour depending on the geomagnetic coordinates in a way that suggested the use of magnetic cues in a map sense (Fischer et al. 2001, Fransson et al. 2001, Lohmann et al. 2001, 2004, Kullberg et al. 2007, Putman et al. 2011). For a long time true navigation was believed to be an ability unique for vertebrates (Walcott and Schmidt-Koenig 1973, Rodda 1984a, b, 1985, Phillips et al. 1995), but it has later been demonstrated to occur also in Caribbean spiny lobsters Panulirus argus (Boles and Lohmann 2003). Geomagnetic map use is however still a controversial subject and the literature presents conflicting evidence on this issue (Alerstam 2006, Freake et al. 2006). Several experimental attempts to disrupt homing success by attaching magnets to the animals have failed, and e.g. seabirds equipped with disturbance magnets showed a navigation performance that was similar to that of control individuals, suggesting that magnetic cues were not essential for their successful homing (Mouritsen et al. 2003, Bonadonna et al. 2005). ere are conflicting evidence also concerning the impor- tance of magnetic versus olfactory cues as a basis for the navigation abilities of homing pigeons (Mora et al. 2004, Gagliardo et al. 2009). A commonly addressed problem regarding a magnetic map sense in animals is the resolution necessary especially Ecography 35: 1039–1047, 2012 doi: 10.1111/j.1600-0587.2012.07507.x © 2012 e Authors. Ecography © 2012 Nordic Society Oikos Subject Editor: Erik Matthysen. Accepted 29 May 2012 Where on earth can animals use a geomagnetic bi-coordinate map for navigation? Jannika E. Boström, Susanne Åkesson and Thomas Alerstam J. E. Boström ([email protected]), S. Åkesson and T. Alerstam, Dept of Biology, Lund Univ., Ecology Building, SE-223 62 Lund, Sweden. Many animal taxa have been shown to possess the ability of true navigation. In this study we investigated the possibilities for geomagnetic bi-coordinate map navigation in different regions of the earth by analysing angular differences between isolines of geomagnetic total intensity and inclination. In ‘no-grid’ zones where isolines were running almost parallel, efficient geomagnetic bi-coordinate navigation would probably not be feasible. ese zones formed four distinct areas with a north-south extension in the northern hemisphere, whereas the pattern in the southern hemisphere was more diffuse. On each side of these zones there was often a mirror effect where identical combinations of the geomag- netic parameters appeared. is may potentially cause problems for species migrating long distances east-west across longitudes, since they may pass areas with identical geomagnetic coordinates. Migration routes assumed for four popula- tions of migratory passerine birds were used to illustrate the possibilities of geomagnetic bi-coordinate map navigation along different routes. We conclude that it is unlikely that animal navigation is universally based on a geomagnetic bi-coordinate map mechanism only, and we predict that the relative importance of geomagnetic coordinate information differs between animals, areas and routes, depending on the different conditions for bi-coordinate geomagnetic naviga- tion in different regions of the earth.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Where on earth can animals use a geomagnetic bi‐coordinate map for navigation?

1039
A bi-coordinate map sense based on magnetic information for homing animals was fi rst suggested almost 130 yr ago (Viguier 1882). Th e idea is that animals may be able to use two diff erent magnetic fi eld parameters to determine their position in relation to a known goal. It has been pointed out that such a map would not necessarily function as our view of a printed map (Lohmann and Lohmann 2006). Instead animals familiar with the magnetic conditions in their goal area may be able to use the isolines of two diff erent mag-netic parameters alternately to approach the goal as long as the two parameters are not parallel ( Å kesson et al. 2003, Lohmann and Lohmann 2006). Map-based navigation, or true navigation, has been defi ned as the ability to determine spatial position in relation to a known goal, based only on information available at a distant release location (Griffi n 1952, Able 2001). An animal that has learnt how two gradients vary in a familiar area, can extrapolate the gradi-ents to determine its position in relation to home or a known goal when displaced away from its familiar area, as long as the gradients vary in a consistent and predictable manner (Wallraff 1974, Wiltschko and Wiltschko 2003).
Several papers have investigated the possibility of magnetic map use for navigation in diff erent animals, e.g. newts (Fischer et al. 2001), alligators (Rodda 1984a), sea turtles (Lohmann and Lohmann 1994, 1996, Lohmann et al. 2001, 2004, Putman et al. 2011) and birds (Beck and Wiltschko 1988, Munro et al. 1997, Å kesson and Alerstam
1998, Fischer et al. 2003). In many of these studies animals were displaced in geomagnetic (but not in geographic) space and they showed a response in orientation or fuel deposi-tion behaviour depending on the geomagnetic coordinates in a way that suggested the use of magnetic cues in a map sense (Fischer et al. 2001, Fransson et al. 2001, Lohmann et al. 2001, 2004, Kullberg et al. 2007, Putman et al. 2011). For a long time true navigation was believed to be an ability unique for vertebrates (Walcott and Schmidt-Koenig 1973, Rodda 1984a, b, 1985, Phillips et al. 1995), but it has later been demonstrated to occur also in Caribbean spiny lobsters Panulirus argus (Boles and Lohmann 2003). Geomagnetic map use is however still a controversial subject and the literature presents confl icting evidence on this issue (Alerstam 2006, Freake et al. 2006). Several experimental attempts to disrupt homing success by attaching magnets to the animals have failed, and e.g. seabirds equipped with disturbance magnets showed a navigation performance that was similar to that of control individuals, suggesting that magnetic cues were not essential for their successful homing (Mouritsen et al. 2003, Bonadonna et al. 2005). Th ere are confl icting evidence also concerning the impor-tance of magnetic versus olfactory cues as a basis for the navigation abilities of homing pigeons (Mora et al. 2004, Gagliardo et al. 2009).
A commonly addressed problem regarding a magnetic map sense in animals is the resolution necessary especially
Ecography 35: 1039–1047, 2012 doi: 10.1111/j.1600-0587.2012.07507.x
© 2012 Th e Authors. Ecography © 2012 Nordic Society Oikos Subject Editor: Erik Matthysen. Accepted 29 May 2012
Where on earth can animals use a geomagnetic bi-coordinate map for navigation?
Jannika E. Boström, Susanne Åkesson and Thomas Alerstam
J. E. Boström ([email protected]), S. Åkesson and T. Alerstam, Dept of Biology, Lund Univ., Ecology Building, SE - 223 62 Lund, Sweden.
Many animal taxa have been shown to possess the ability of true navigation. In this study we investigated the possibilities for geomagnetic bi-coordinate map navigation in diff erent regions of the earth by analysing angular diff erences between isolines of geomagnetic total intensity and inclination. In ‘no-grid’ zones where isolines were running almost parallel, effi cient geomagnetic bi-coordinate navigation would probably not be feasible. Th ese zones formed four distinct areas with a north-south extension in the northern hemisphere, whereas the pattern in the southern hemisphere was more diff use. On each side of these zones there was often a mirror eff ect where identical combinations of the geomag-netic parameters appeared. Th is may potentially cause problems for species migrating long distances east-west across longitudes, since they may pass areas with identical geomagnetic coordinates. Migration routes assumed for four popula-tions of migratory passerine birds were used to illustrate the possibilities of geomagnetic bi-coordinate map navigation along diff erent routes. We conclude that it is unlikely that animal navigation is universally based on a geomagnetic bi-coordinate map mechanism only, and we predict that the relative importance of geomagnetic coordinate information diff ers between animals, areas and routes, depending on the diff erent conditions for bi-coordinate geomagnetic naviga-tion in diff erent regions of the earth.

1040
for short distance movements (Phillips 1996, Lohmann and Lohmann 2006). Newts typically move distances about 1 – 2 km (Fischer et al. 2001) and even though magnetic variations over such short distances are small, newts have been shown to be able to make use of magnetic informa-tion over these short ranges (Phillips et al. 1995, 2002, Fischer et al. 2001). Th e magnetic prerequisites for the successful use of a magnetic map are not often discussed and few studies aim to examine this. Å kesson and Alerstam (1998) used satellite tracks from wandering albatrosses from two diff erent breeding areas and examined the varia-tion in fi ve magnetic parameters (total fi eld intensity, hori-zontal fi eld intensity, vertical fi eld intensity, inclination and declination) within each track area to determine whether any two parameters formed a grid that could be used as a basis for a bi-coordinate map in the area. Th ey did not fi nd a general combination of any two magnetic parameters that could be used in all areas of the Southern Ocean and concluded that the irregular and weak gradi-ents in parts of the region makes the use of a bi-coordinate magnetic map unlikely ( Å kesson and Alerstam 1998).
In this study we wanted to give a global perspective of the prerequisites for magnetic bi-coordinate maps with focus primarily on migrating birds. Th e analysis was divided into two main parts: 1) the construction of a global map showing the angular diff erence between the isolines of total fi eld intensity and inclination, and 2) an illustration of the possibility to use a magnetic map based on total fi eld intensity and inclination along a few exemplifi ed migration routes.
We chose not to include magnetic declination in our analysis since we wanted to evaluate the use of a bi-coordinate map based purely on geomagnetic para-meters. Declination requires additional celestial/geographic information for the determination of the angular diff er-ence between geomagnetic and geographic north/south directions. Most studies reporting evidence for the use of magnetic map information in animal navigation refer to purely geomagnetic parameters (Freake et al. 2006), and declination as a combined magnetic/celestial cue has only rarely been considered (but see Å kesson and Alerstam 1998, Å kesson et al. 2005).
It should be noted that there may be regional diff erences in the usefulness not only of geomagnetic bi-coordinate navigation as analysed in this paper, but also of magnetic compass orientation, because magnetic loxodromes are shorter than geographic loxodromes in some regions of the earth (thus making constant magnetic compass ori-entation more favourable than constant geographic com-pass orientation) while the reverse situation holds in other regions (Alerstam 2001). In this analysis we will not deal with orientation but instead focus on true navigation and demonstrate and discuss the diff erent conditions for geomagnetic bi-coordinate navigation that exist in diff er-ent regions of the earth.
Magnetic anomalies may also pose problems for animals raised in these areas or passing them on migra-tion (Walcott 1978, Alerstam 1987, Schiff ner et al. 2011). Our aim with this analysis is to investigate geomagnetic bi-coordinate navigation on a global scale, and since magnetic anomalies only have a signifi cant impact on a regional or
local scale, they are not incorporated into our analysis. We will, however, return to the subject in the discussion.
Material and methods
Map analysis
For the map analysis we used two maps based on the WMM 2005 model (McLean et al. 2004) showing the variations in total fi eld intensity ( μ T) and inclination ( ° ) throughout the earth. Th e maps were shown in Mercator projection and included latitudes between 70 ° N and 70 ° S. Th ey were put together so that both magnetic parameters were shown in one map (Fig. 1). We divided the map into 3 ° lat. � 3 ° long. squares and the angular diff erence between the twoisolines within each square was estimated. Th ree diff erent intervals were used for the estimates: � 2 ° , 2 – 30 ° and � 30 ° . Th ese intervals were chosen to illustrate grids with diff erent levels of diffi culty for geomagnetic bi-coordinate navigation: � 2 ° : very hard or impossible, 2 – 30 ° : possibly presenting diffi culties and � 30 ° : useful and relatively well defi ned (Fig. 2, 3).
Th e precision of bi-coordinate magnetic navigation will depend on the animals ’ degree of sensitivity to the two geomagnetic gradients, the steepness of these gradients and the angular diff erence between the isolines of the two gradients. Th e animals will be able to locate an ‘ uncertainty area ’ around their goal that is defi ned by the limitations of magnetic sensitivity to the two gradients. Th is uncertainty area will be minimal (allowing the most precise naviga-tion) if the two gradients form a perpendicular grid and it will increase when the angular diff erence between isolines decreases. Geometric considerations show that, for small angles between isolines ( α ), the relative size of the uncer-tainty area (relative to that in a perpendicular grid) increases in direct proportion to 1/ α ( α in radians). Th is means that with an angular diff erence of e.g. 2 ° between gridlines, the uncertainty area has increased approximately 30-fold ( � 180/2 π ) in relation to the situation with a perpendi-cular grid, and the uncertainty area has of course become enlarged along the gridlines on either side of the goal (cf. Fig. 2). At what angle this increase and elongation of the uncertainty area will cause a problem for bi-coordinate navigation among diff erent animals is unknown, and our choice of 2 ° and 30 ° for the diff erent grid situations (Fig. 3) is arbitrary. However, the qualitative overall global pattern in Fig. 3 would remain the same if we had chosen other intervals of angles for the diff erent groups of geomag-netic grid, with e.g. the red zones becoming more narrow (but not disappearing) if we had chosen an even smaller angle than 2 ° to indicate almost parallel gridlines.
Th ere exists only limited information about the pos-sible sensitivity to magnetic inclination or intensity among diff erent animals (e.g. direct evidence that eastern red-spotted newts Notophthalmus viridescens can detect inclina-tion changes as small as 2 ° and 0.5 ° ; Fischer et al. 2001, Phillips et al. 2002, indirect indications that American alligators Alligator mississippiensis may detect changes in inclination as small as 0.01 – 0.02 ° ; Rodda 1984a; and neu-rophysiological responses by birds to changes in intensity

1041
Figure 1. Map showing the isolines for total fi eld intensity ( μ T, in blue) and inclination ( ° , in red). Th e distance between the isolines is 5 μ T for the total fi eld intensity and 20 ° for the inclination. Th e map is in Mercator projection and shows latitudes between 70 ° N and 70 ° S.
60°
????
15°
(a) (c)
(b)
Figure 2. Schematic drawing illustrating how bi-coordinate navigation from a distant location to a known goal (true navigation) is aff ected by the angular diff erence between isolines. Th e isolines of the two coordinates (geomagnetic parameters) are indicated by solid and dashed lines, respectively. Th ree examples illustrate situations with diff erent angles between isolines. Th e diagram shows the condi-tions experienced by an animal at four diff erent positions in relation to the goal. Note that even though the physical distances to the goal diff er between the positions, the gradient diff erences remain the same. Th e angular diff erences for the grids are (a) 60 ° , (b) 15 ° and (c) 0 ° . Arrows show the gradient directions towards the goal for each of the two coordinates (perpendicular to the isolines). Navigation towards the goal may be achieved by the animals reducing the gradient diff erences for both coordinates simultaneously or by alternately reducing the gradient diff erence for one coordinate (while moving along the isoline of the other coordinate) at a time. For navigation in a grid where isolines intersect at a small angle (case b) even small gradient diff erences from the goal may be associated with long physical dis-tances, and a very high accuracy of measuring gradients is required to establish a course towards the goal. In these situations, correct navigation will be very sensitive to small local or regional disturbances in the geomagnetic fi eld. When isolines are parallel (case c) the animal may only perform uni-coordinate navigation, which will allow it to localise the correct coordinate latitude but not longitude. Shaded areas in (a) and (b) illustrate the increase in uncertainty area around the goal with decreasing angle between gridlines, as explained in the text (Methods).
as small as 0.05 – 0.2 μ T; Beason and Semm 1987, Semm and Beason 1990). Even if the sensitivity to magnetic gra-dients diff er between diff erent animals (and perhaps even between populations that live and move within zones of a more or less favourable grid situation), making them more
or less well-equipped to navigate in areas with small angles between gridlines, the relative diffi culties for bi-coordinate navigation (as refl ected by the relative size of the uncer-tainty area; cf. above) will always increase with decreasing angle between gridlines. Our analysis serves to explore and

1042
We used the diff erent migration routes to plot the total fi eld intensities and the inclination angles against every 3 ° longitude along the route. Th e geomagnetic informa-tion was calculated using the WMM 2005 model (McLean et al. 2004).
Results
Map analysis
We produced a map showing the three angular diff erence intervals in diff erent colours (Fig. 3). Th e map analy-sis revealed a pattern of four quite distinct north/south zones in the northern hemisphere where the angular diff er-ence between the isolines for total fi eld intensity and inclina-tion diff ered by only 2 ° or less. Th ese zones are associated with a mirror-image eff ect, so that on either side of the zones there are places with identical geomagnetic fi eld intensity/inclination coordinates if the zones extend north-south (compare with Fig. 1). Th ere were also areas in the south-ern hemisphere where the angular diff erence between the two magnetic fi eld parameters diff ered by 2 ° or less (Fig. 3). It should be noted that, because of the secular variation of the geomagnetic fi eld (McLean et al. 2004), the spatial pattern of geomagnetic bi-coordinate conditions as shown in Fig. 3 is not static but will change with time.
Migration routes
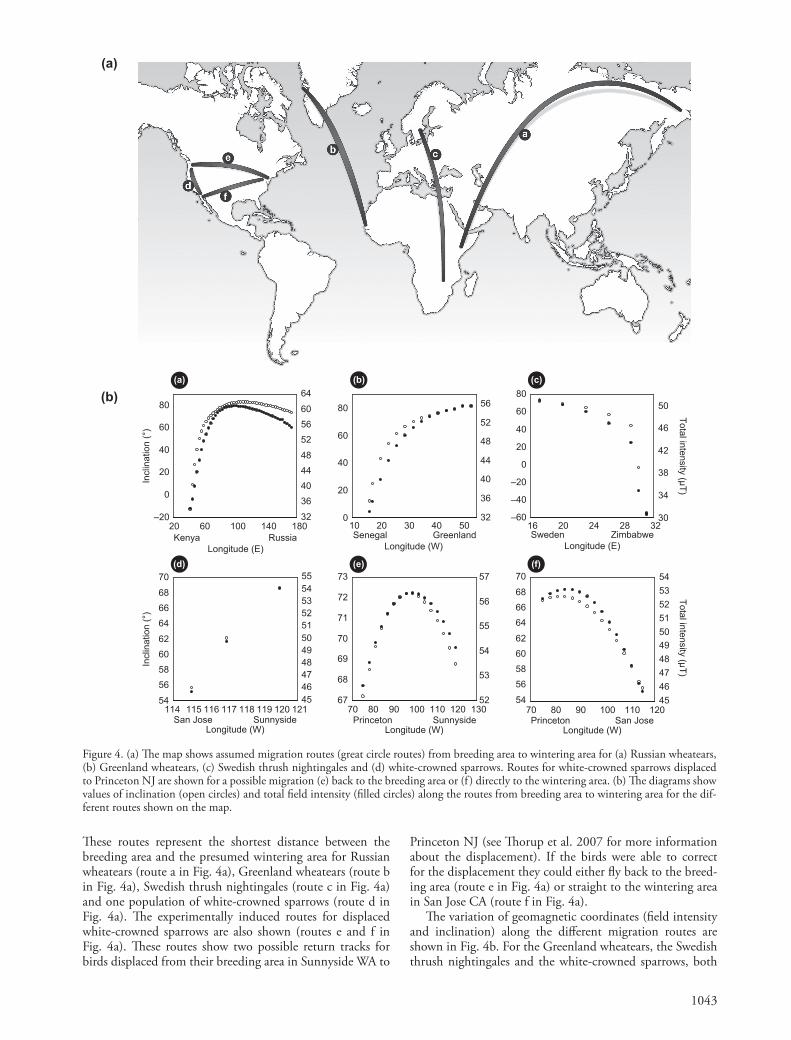
Th e assumed migration routes (great circle routes) for the populations included in the study are presented in Fig. 4a.
illustrate this relative diffi culty for bi-coordinate geomag-netic navigation on a global scale.
Migration routes
We assumed great circle routes for four examples of natural migrations and two experimentally induced migra-tions according to the equations given in Imboden and Imboden (1972) using Matlab 7.9.0 (MathWorks). Th ese routes do not necessarily depict the exact natural migra-tion routes followed by each population, since exact routes are unknown. Great circle routes were assumed for conve-nience, but principal patterns of geomagnetic coordinate changes along routes were the same for loxodromes and for routes intermediate between loxodromes and great circles.
Th e natural migrations included two populations of wheatear Oenanthe oenanthe , one population of thrush nightingale Luscinia luscinia and one population of white-crowned sparrow Zonotrichia leucophrys gambelii . Th ese routes are also representative for several other passerine spe-cies migrating in the diff erent areas (Newton 2008). Th e two experimentally induced routes were calculated using information from a translocation study on white-crowned sparrows in North America (Th orup et al. 2007). Th e thrush nightingale route was calculated in two steps, fi rst from Sweden to Egypt and then from Egypt to Zimbabwe (Fransson et al. 2001, 2005). We used these schematic routes to illustrate in a general way how geomagnetic coor-dinates may change along natural routes as well as along potential homing routes after displacement in ways that have consequences for true navigation based on a geomag-netic bi-coordinate map.
Figure 3. Illustration of the possibility for animals to use a bi-coordinate map based on geomagnetic inclination and total fi eld intensity on a global scale. Th e map was divided into 3 ° lat. � 3 ° long. squares and the angular diff erence between isolines of total fi eld intensity and inclination was estimated within each square. Areas with an angular diff erence of � 2 ° are shown in red, 2 – 30° in yellow and � 30 ° are shown in green.
1043
80 6064
80 5660
8050
70 80 90 100 110 120 13067
68
69
70
71
72
73
52
53
54
55
56
57
70 80 90 100 110 120545658606264666870
45464748495051525354
114 115 116 117 118 119 120 121545658606264666870
4546474849505152535455
San JosePrincetonSunnysideSunnysideSan Jose Princeton
Longitude (E) Longitude (W)
Longitude (W) Longitude (W) Longitude (W)
Longitude (E)
20 60 100 140 180–20
0
20
40
60
32364044485256
10 20 30 40 500
20
40
60
32
36
40
44
48
52
16 20 24 28 32–60
–40
–20
0
20
40
30
34
38
42
46
Kenya Russia Senegal Greenland Sweden Zimbabwe
Incl
inat
ion
(°)
Total intensity (µT)
Incl
inat
ion
(°)
Total intensity (µT)
(b) (c)(a)
(a)
(b)
(d) (f)(e)
Figure 4. (a) Th e map shows assumed migration routes (great circle routes) from breeding area to wintering area for (a) Russian wheatears, (b) Greenland wheatears, (c) Swedish thrush nightingales and (d) white-crowned sparrows. Routes for white-crowned sparrows displaced to Princeton NJ are shown for a possible migration (e) back to the breeding area or (f ) directly to the wintering area. (b) Th e diagrams show values of inclination (open circles) and total fi eld intensity (fi lled circles) along the routes from breeding area to wintering area for the dif-ferent routes shown on the map.
Th ese routes represent the shortest distance between the breeding area and the presumed wintering area for Russian wheatears (route a in Fig. 4a), Greenland wheatears (route b in Fig. 4a), Swedish thrush nightingales (route c in Fig. 4a) and one population of white-crowned sparrows (route d in Fig. 4a). Th e experimentally induced routes for displaced white-crowned sparrows are also shown (routes e and f in Fig. 4a). Th ese routes show two possible return tracks for birds displaced from their breeding area in Sunnyside WA to
Princeton NJ (see Th orup et al. 2007 for more information about the displacement). If the birds were able to correct for the displacement they could either fl y back to the breed-ing area (route e in Fig. 4a) or straight to the wintering area in San Jose CA (route f in Fig. 4a).
Th e variation of geomagnetic coordinates (fi eld intensity and inclination) along the diff erent migration routes are shown in Fig. 4b. For the Greenland wheatears, the Swedish thrush nightingales and the white-crowned sparrows, both

1044
navigation problematic or impossible. True navigation based on extrapolated geomagnetic bi-coordinate map gradients will become locally impossible in many anomaly areas where the gradients are greatly distorted and irregular and where the magnetic coordinates may vary out of range (navigationpossibilities will depend on the scale of movement in relation to the scale of irregularities in the geomagnetic fi eld, cf. Phillips 1996). However, this disturbance is only tem-porary for passing long-distance migrants since magnetic anomalies often are highly localised (and the distortion of the normal geomagnetic fi eld becomes reduced with increas-ing altitude) and the migrants can resume correct naviga-tion as soon as they have left the local anomaly areas. Studies of homing pigeons as well as migrating birds fl ying across magnetic anomalies (Walcott 1978, Alerstam 1987) or over an antenna system causing anomalous magnetic condi-tions (Larkin and Sutherland 1977) showed that the birds changed altitude or direction signifi cantly more often at the anomaly than over areas with a normal undisturbed geomagnetic fi eld, but the observed birds had soon passed the anomaly and restored stable fl ight paths. Th is indicates that local magnetic anomalies are not a major problem for long-distance bird migrants with only transient and small possible eff ects on the birds ’ magnetic map and compass senses (Alerstam 1987). Th e navigational performances by homing pigeons released in magnetic anomalies, however, are variable and seem to be dependent on conditions at the loft as well (Walcott 1992). A recent study using GPS devices to track homing pigeons released in a magnetic anomaly found that these birds needed longer time to establish their homeward direction but they managed to do so within 5 km from the release site, while still in the mag-netic anomaly area (Schiff ner et al. 2011). Even if local magnetic anomaly areas are numerous and widespread in all regions of the earth (Th é bault et al. 2010) they cover only a very small fraction of the earth ’ s surface, e.g. at most 1/1000 of the area of northern Scandinavia where there are several prominent magnetic anomalies (Alerstam 1987).
Animals moving locally in a magnetic anomaly area may use the anomaly to their own advantage by learning and incorporating the anomaly features as magnetic land-marks in a local mosaic map (Wiltschko et al. 2009). However, long-distance migrants raised within magnetic anomalies may learn an incorrect geomagnetic map by extrapolating the magnetic conditions within their home range and hence ending up migrating in the wrong direc-tion. Th ey may also learn an erroneous magnetic compass and misorient for that reason (Alerstam 1991). If such misnavigation or misorientation of long-distance migrants from magnetic anomalies actually occurs remains specula-tive and will anyway only involve a minor proportion of all migrants (cf. above about the small proportional coverage of magnetic anomaly areas).
Th e fact that animals moving within a ‘ no-grid ’ zone will probably not be able to use a bi-coordinate geomag-netic map for true navigation does of course not preclude that they use uni-coordinate magnetic navigation (or other navigational responses to the geomagnetic fi eld) in combi-nation with one or more non-magnetic map cues in these areas. Many animals, including several short-distance and partial migrants among birds as well as homing pigeons,
total fi eld intensity and inclination angle drop along the great circle route from the breeding area to the wintering area (diagrams b, c and d in Fig. 4b). For the Russian wheatears, however, both parameters fi rst increase before starting to drop at about 90 ° E (diagram a in Fig. 4b). Th e displaced white-crowned sparrows would also have to fl y initially towards higher total fi eld intensities and inclina-tions in order to reach the lower coordinate values of the breeding or wintering area (diagrams e and f in Fig. 4b).
Discussion
Th e magnetic map analysis reveals an interesting pattern in the northern hemisphere with four areas with a north/south extension, forming ‘ no-grid ’ zones (small angular diff erence, red markings in Fig. 3) where bi-coordinate navigation based on total fi eld intensity and inclination can be expected to be impossible or at least extremely diffi cult. It should be noted that other geomagnetic coordinates, like the horizontal or vertical components of the geomagnetic fi eld, are directly derived from the total fi eld intensity and angle of inclination, thus off ering no usefulness for bi-coordinate positioning when the two latter basic coordinates run virtually parallel. On a global scale the angular diff er-ence between the isolines is most commonly in the range 2 – 30 ° (moderate angular diff erence, yellow markings in Fig. 3), but there are also large areas with an angular dif-ference of 30 ° or more, typically above 50 ° N and below the equator (large angular diff erence, green markings in Fig. 3). Some of these areas coincide with extensive areas across ocean basins.
An interesting mirror eff ect appears in association with the ‘ no-grid ’ zones, where a particular magnetic bi-coordinate combination occurs on both sides of a ‘ no-grid’ area, if this area extends in a north-south direction. Th is could potentially cause large problems for animals migrat-ing east-west across the longitudes and across these areas, since they may encounter the same magnetic fi eld para-meters twice along the migration route. Many bird species migrating in east-west directions at northerly latitudes, not least shorebirds (Alerstam et al. 2007) but also seabirds, gulls, geese and terrestrial songbirds are crossing the ‘ no-grid ’ zones in Beringia and North America as well as in the Atlantic and in Asia (Alerstam 1990). Th is passage may expose the migrants to similar or identical magnetic fi eld combinations on either side of the ‘ no-grid ’ zone. Under these conditions an innate bi-coordinate map of the migra-tion route would be hard to attain and even a learned bi-coordinate map may be a challenge for experienced migrants. In order to overcome this problem animals probably have to rely on additional cues, independent of the geomagnetic fi eld.
However, Fig. 3 also shows that there are large regions in e.g. the South Atlantic Ocean, Indian Ocean, South Pacifi c Ocean and in NE North America/Greenland and N Europe/W Siberia, where the geomagnetic fi eld off ers very suitable conditions for bi-coordinate navigation among animals that keep their movements within these regions.
In many areas of the earth magnetic anomalies disturb the natural geomagnetic fi eld and make magnetic bi-coordinate

1045
In South America there is a ‘ no-grid ’ zone extending across the Caribbean southwards through the central part of northern South America. Th is could potentially cause problems for inter-continental migrants breeding in North America and wintering in Central or South America.
Th roughout Europe the angular diff erences between fi eld intensity and inclination isolines are mostly moderate, but with a zone of small angles in the west and an area with large isoline angles in the northeast (Fig. 3). Great Britain, Ireland and Portugal are more or less completely within a ‘ no-grid ’ area, suggesting that birds migrating or homing within these regions may not use the geomagnetic fi eld for bi-coordinate navigation.
Birds migrating across longitudes from eastern Asia or Alaska towards Europe or Africa will have to pass through ‘ no-grid ’ areas where coordinate changes will shift from increasing to decreasing (cf. Fig. 3, 4a).
A ‘ no-grid ’ zone runs through Australia from northwest to the central southern part (Fig. 3). Th ere are no areas where the isolines for total fi eld intensity and inclination form a very well defi ned grid that would be optimal for a magnetic bi-coordinate map for Australian migrants.
Oceanic migrations
In the northern hemisphere, a ‘ no-grid ’ zone stretches through the Bering Sea southwards to the Pacifi c Ocean. Th ere is also an area southwest of California and Mexico with little or no useful geomagnetic grid. An area with larger variation forming a well-defi ned grid is found west of South America, and a similar area stretches across the Philippines and to the east (Fig. 3).
A ‘ no-grid ’ zone extending from western Europe to the Caribbean splits the North Atlantic Ocean. On both sides of this divide the angular diff erences are moderate (Fig. 3), probably making parts of the North Atlantic Ocean a problematic area for magnetic bi-coordinate navigation.
In most parts of the South Atlantic Ocean, on the other hand, there is a nice grid that may form a basis for magnetic bi-coordinate navigation (Fig. 3; see also Å kesson and Alerstam 1998). Th ere are, however, areas with irregu-lar and weak gradients where bi-coordinate navigation can be expected to be diffi cult or even impossible (Fig. 1; Å kesson and Alerstam 1998).
Th ere are also irregular or weak gradients south of Australia (Fig. 1). In a large part of the Indian Ocean, south of India and east of Africa (Fig. 3), the isolines form a grid that may enable successful bi-coordinate navigation.
Migration routes
Both total fi eld intensity and inclination angle decrease along the presumed migration routes for Greenland wheatears and Swedish thrush nightingales fl ying from their European breeding areas to their African wintering areas (Fig. 4b). Th e thrush nightingales move within an area with a quite well defi ned bi-coordinate grid and would not be expected to encounter any navigational problems if using a geomagnetic bi-coordinate map. Th e Greenland wheatears will have to cross a ‘ no-grid ’ area in the Atlantic Ocean,
have their movements completely within the ‘ no-grid ’ zones in e.g. southern North America, Mexico and Great Britain. For these animals we expect that their homing/navigation cannot be based on geomagnetic bi-coordinate navigation. However, for migrants that pass these ‘ no-grid’ zones on migration, these areas do not necessarily act as barriers to navigation. Th ere are many conceivable special behaviours that may have evolved to allow the migrants to pass such zones without the need to rely on continuous geomagnetic bi-coordinate navigation.
If control of migration is primarily based on an innate clock-and-compass program during the fi rst outward (autumn) journeys (Berthold 1996, 2001), when juveniles are assumed to learn cues (e.g. geomagnetic coordinates) that will be used for navigation during future journeys in the adult life, then passing a ‘ no-grid ’ zone may not be a serious hurdle among either juveniles or adults. If adult migrants follow a succession of coordinate positions they will be able to fi nd the way across ‘ no-grid ’ zones and even pass widely separated areas with identical geomagnetic coordinates on either side of these zones without compli-cations. In contrast, the east-west passage of such ‘ no-grid ’ areas will be diffi cult for animals using true navigation based on extrapolated geomagnetic gradients in combination with one or a few geomagnetic goal positions. Mirror sites with identical geomagnetic coordinates on either side of no-grid zones will also cause diffi culties in a system where area-specifi c geomagnetic coordinates (sometimes referred to as magnetic signposts or beacons) trigger diff erent orientation responses, which are adaptive in the respec-tive areas. Th e response that is triggered depends solely on the actual geomagnetic conditions experienced by the ani-mal and not on preceding magnetic conditions, and this is the type of geomagnetic control system suggested for the orientation of young loggerhead sea turtles in the North Atlantic gyre (Lohmann et al. 2001, reviewed by Collett and Collett 2011). In regions where there are mirror sites with identical geomagnetic coordinates such a system would trigger the same response at the widely separated mirror sites, which may make this kind of system maladaptive in such regions.
Th ere is increasing evidence suggesting that more sophisticated control mechanisms than merely clock-and-compass orientation may guide also the fi rst outward jour-neys of juveniles, and that these mechanisms may include geomagnetic map information (Fransson et al. 2001, 2005, Freake et al. 2006). Th e pre-programmed bi-coordinate geomagnetic map information required to guide an animal east-west across a ‘ no-grid ’ zone, where magnetic gradients will become reversed along the route, must be much more elaborate than for journeys contained within regions of a well-defi ned geomagnetic grid (Fig. 3).
Continental migrations
North American migrants will face the challenge of a large ‘ no-grid ’ zone in the southern USA, and the rest of the con-tinent mostly exhibits moderate diff erences in isoline varia-tion (Fig. 3). Th e use of a geomagnetic bi-coordinate map on this continent thus seems unlikely for long-distance migrants and for birds migrating within the southern USA.

1046
possible on the basis of novel advanced tracking techniques. Furthermore, also for the planning and interpretation of displacement experiments (both geographically and in geo-magnetic space) it will be important to consider the geo-magnetic map coordinate patterns as illustrated in this paper. Th is will help to determine the possible importance and role of geomagnetic map cues in animal navigation.
Th e widely diff erent conditions for geomagnetic bi-coordinate navigation that exist in diff erent regions of the earth and the associated complications for migratory ani-mals lead us to two main conclusions: 1) animal navigation is probably not based universally on a geomagnetic bi-coordinate map mechanism alone, and 2) the relative impor-tance of geomagnetic map cues in animal navigation may well diff er between diff erent organisms, species, areas androutes, depending on the very diff erent conditions for geo-magnetic navigation in diff erent regions of the earth.
Acknowledgements – We are grateful to Gunilla Andersson and Emil Assarsson for help with the fi gures. Financial support was given by the Swedish Research Council (to TA and S Å ). Th e authors are associated with the Centre for Animal Movement Research (CAnMove) at Lund Univ.
References
Able, K. P. 2001. Th e concepts and terminology of bird navigation. – J. Avian Biol. 32: 174 – 182.
Å kesson, S. and Alerstam, T. 1998. Oceanic navigation: are there any feasible geomagnetic bi-coordinate combinations for albatrosses? – J. Avian Biol. 29: 618 – 625.
Å kesson, S. et al. 2003. Navigation by green turtles: which strategy do displaced adults use to fi nd Ascension Island? – Oikos 103: 363 – 372.
Å kesson, S. et al. 2005. Dramatic orientation shift of displaced birds in response to the geomagnetic fi eld. – Curr. Biol. 15: 1591 – 1597.
Alerstam, T. 1987. Bird migration across a stong magnetic anomaly. – J. Exp. Biol. 130: 63 – 86.
Alerstam, T. 1990. Bird migration. – Cambridge Univ. Press. Alerstam, T. 1991. Ecological causes and consequences of bird
orientation. – In: Berthold, P. (ed.), Orientation in birds. Birkh ä user, pp. 202 – 225.
Alerstam, T. 2001. Evaluation of long-distance orientation in birds on the basis of migration routes recorded by radar and satellite tracking. – J. Navig. 54: 393 – 403.
Alerstam, T. 2006. Confl icting evidence about long-distance animal navigation. – Science 313: 791 – 794.
Alerstam, T. et al. 2007. A polar system of intercontinental bird migration. – Proc. R. Soc. B 274: 2523 – 2530.
Beason, R. C. and Semm, P. 1987. Magnetic responses of the trigeminal nerve system of the bobolink ( Dolichonyx oryzivorus ). – Neurosci. Lett. 80: 229 – 234.
Beck, W. and Wiltschko, W. 1988. Magnetic factors control the migratory direction of pied fl ycatchers. – In: Ouellet, H. (ed.), Acta XIX Congress of International Ornithology. Univ. of Ottawa Press, pp. 1955 – 1962.
Berthold, P. 1996. Control of bird migration. – Chapman and Hall.
Berthold, P. 2001. Bird migration: a novel theory for the evolution, the control and the adaptability of bird migration. – J. Ornithol. 142: 148 – 159.
Boles, L. C. and Lohmann, K. J. 2003. True navigation and magnetic maps in spiny lobsters. – Nature 421: 60 – 63.
but since they cross it in a north-south direction it will prob-ably not pose any navigational problems (Fig. 3, 4).
Russian wheatears have a long migration route and in its initial phase they have to move a long distance across lon-gitudes from east to west. During this fl ight they will cross a ‘ no-grid ’ zone and they will also have to move towards increasing geomagnetic values before the ‘ no-grid ’ zone and then towards decreasing values (Fig. 4b). Independent of exact route this population will encounter the same prob-lem when crossing the ‘ no-grid ’ zone as long as the course across the ‘ no-grid ’ zone is not too diff erent from east-west. Th e wheatears may also encounter identical geomagnetic combinations on both sides of the ‘ no-grid ’ zone, making geomagnetic bi-coordinate navigation more problematic for this population.
Th e natural migration of the white-crowned sparrow breeding in Sunnyside WA does not pose any navigational diffi culties. However, after the cross-continental displace-ment the birds would encounter very diff erent geomagnetic situations during their return movements across the conti-nent (Fig. 4b). Along their natural migration route the white-crowned sparrows fl y almost due south towards a decreasing gradient (Th orup et al. 2007). After the eastward displace-ment to Princeton NJ the hypothetical migration routes to the breeding area and the wintering area both cross a ‘ no-grid ’ area and the birds would have to fl y towards increasing gradients east of the ‘ no-grid ’ area and towards decreasing gradients west of the ‘ no-grid ’ area, a very diff er-ent situation compared to their natural migration. A learned navigational map extrapolated to extend across the conti-nental US for adult white-crowned sparrows as suggested by Th orup et al. (2007) must be considered highly unlikely if based exclusively on geomagnetic parameters. A simi-lar displacement study on catbirds Dumetella carolinensis found no eff ect of magnetic manipulations, suggesting that these birds may rely on other, possibly olfactory, navigational cues (Holland et al. 2009).
Conclusions
Th e map analysis revealed several ‘ no-grid ’ areas where magnetic bi-coordinate navigation seems very diffi cult if at all possible, whereas other areas seem to off er almost ideal conditions for magnetic bi-coordinate navigation. Our analysis also showed that migration east-west across lon-gitudes may pose a problem for animals using a magnetic bi-coordinate map for navigation when crossing a ‘ no-grid ’ zone. Th ese individuals would have to move initially towards increasing values of fi eld intensity and inclination and then switch to moving towards decreasing values. On each side of the ‘ no-grid ’ zone the animal may also be exposed to identi-cal coordinates since the grid system is not unique, but the same magnetic combinations may be mirrored on each side of the ‘ no-grid ’ area. In order to overcome these problems animals navigating in and across ‘ no-grid ’ zones probably have to rely on additional cues.
We think that it will be useful and important to consider how geomagnetic coordinate conditions change along the exact migration routes of individual animals as demonstrations of such individual routes are now becoming

1047
Mora, C. V. et al. 2004. Magnetoreception and its trigeminal mediation in the homing pigeon. – Nature 432: 508 – 511.
Mouritsen, H. et al. 2003. Waved albatrosses can navigate with strong magnets attached to their head. – J. Exp. Biol. 206: 4155 – 4166.
Munro, U. et al. 1997. Evidence for a magnetite-based navigational “ map ” in birds. – Naturwissenschaften 84: 26 – 28.
Newton, I. 2008. Ecology of bird migration. – Academic Press. Phillips, J. B. 1996. Magnetic navigation. – J. Th eor. Biol. 180:
309 – 319. Phillips, J. B. et al. 1995. True navigation by an amphibian.
– Anim. Behav. 50: 855 – 858. Phillips, J. B. et al. 2002. Behavioral titration of a magnetic map
coordinate. – J. Comp. Physiol. A 188: 157 – 160. Putman, N. F. et al. 2011. Longitude perception and bicoordinate
magnetic maps in sea turtles. – Curr. Biol. 21: 1 – 4. Rodda, G. H. 1984a. Th e orientation and navigation of juvenile
alligators: evidence of magnetic sensitivity. – J. Comp. Physiol. 154: 649 – 658.
Rodda, G. H. 1984b. Homeward paths of displaced juvenile alliga-tors as determined by radiotelemetry. – Behav. Ecol. Sociobiol. 14: 241 – 246.
Rodda, G. H. 1985. Navigation in juvenile alligators. – Z. Tier-psychol. 68: 65 – 77.
Schiff ner, I. et al. 2011. Tracking pigeons in a magnetic anomaly and in magnetically “quiet” terrain. – Naturwissenschaften 98: 575 – 581.
Semm, P. and Beason, R. C. 1990. Responses to small magnetic variations by the trigeminal system of the bobolink. – R. Brain Res. Bull. 25: 735 – 740.
Th é bault, E. et al. 2010. Th e magnetic fi eld of the Earth ’ s lithosphere. – Space Sci. Rev. 155: 95 – 127.
Th orup, K. et al. 2007. Evidence for a navigational map stretching across the continental U.S. in a migratory songbird. – Proc. Natl Acad. Sci. USA 104: 18115 – 18119.
Viguier, C. 1882. Le sens de l ’ orientation et ses organes chez les ani-maux et chez l ’ homme. – Rev. Phil. France Etanger 14: 1 – 36.
Walcott, C. 1978. Anomalies in the earth ’ s magnetic fi eld increase the scatter of pigeons ’ vanishing bearings. – In: Schmidt-Koenig, K. and Keeton, W. T. (eds), Animal migra-tion, navigation and homing. Springer, pp. 141 – 151.
Walcott, C. 1992. Pigeons at magnetic anomalies – the eff ects of loft location. – J. Exp. Biol. 170: 127 – 141.
Walcott, C. and Schmidt-Koenig, K. 1973. Th e eff ect on pigeon homing of anesthesia during displacement. – Auk 90: 281 – 286.
Wallraff , H. G. 1974. Das Navigationssystem der V ö gel. – R. Oldenbourg Verlag.
Wiltschko, R. and Wiltschko, W. 2003. Avian navigation: from historical to modern concepts. – Anim. Behav. 65: 257 – 272.
Wiltschko, R. et al. 2009. A strong magnetic anomaly aff ects pigeon navigation. – J. Exp. Biol. 212: 2983 – 2990.
Bonadonna, F. et al. 2005. Orientation in the wandering albatross: interfering with magnetic perception does not aff ect orientation performance. – Proc. R. Soc. B 272: 489 – 495.
Collett, T. S. and Collett, M. 2011. Animal navigation: following signposts in the sea. – Curr. Biol. 21: R843 – R846.
Fischer, J. H. et al. 2001. Evidence for the use of magnetic map information by an amphibian. – Anim. Behav. 62: 1 – 10.
Fischer, J. H. et al. 2003. Magnetic navigation by an avian migrant. – In: Berthold, P. et al. (eds), Avian migration. Springer, pp. 423 – 432.
Fransson, T. et al. 2001. Magnetic cues trigger extensive refuelling. – Nature 414: 35 – 36.
Fransson, T. et al. 2005. Non-random distribution of ring recoveries from trans-Saharan migrants indicates species-specifi c stopover areas. – J. Avian Biol. 36: 6 – 11.
Freake, M. J. et al. 2006. Magnetic maps in animals: a theory comes of age? – Q. Rev. Biol. 81: 327 – 347.
Gagliardo, A. et al. 2009. Navigational abilities of adult and experienced homing pigeons deprived of olfactory or trigemi-nally mediated magnetic information. – J. Exp. Biol. 212: 3119 – 3124.
Griffi n, D. R. 1952. Bird navigation. – Biol. Rev. 27: 359 – 393. Holland, R. A. et al. 2009. Testing the role of sensory systems in
the migratory heading of a songbird. – J. Exp. Biol. 212: 4065 – 4071.
Imboden, C. and Imboden, D. 1972. Formel f ü r Orthodrome und Loxodrome bei der Berechnung von Richtung und Distanz zwischen Beringungs- und Wiederfundort. – Vogelwarte 26: 336 – 346.
Kullberg, C. et al. 2007. Fuelling decisions in migratory birds: geomagnetic cues override the seasonal eff ect. – Proc. R. Soc. B 274: 2145 – 2151.
Larkin, R. P. and Sutherland, P. J. 1977. Migrating birds respond to Project Seafarer ’ s electromagnetic fi eld. – Sience 195: 777 – 779.
Lohmann, K. J. and Lohmann, C. M. 1994. Detection of magnetic inclination angle by sea turtles: a possible mechanism for determining latitude. – J. Exp. Biol. 194: 23 – 32.
Lohmann, K. J. and Lohmann, C. M. 1996. Detection of magnetic fi eld intensity by sea turtles. – Nature 380: 59 – 61.
Lohmann, K. J. and Lohmann, C. M. F. 2006. Sea turtles, lobsters, and oceanic magnetic maps. – Mar. Freshw. Behav. Physiol. 39: 49 – 64.
Lohmann, K. J. et al. 2001. Regional magnetic fi elds as naviga-tional markers for sea turtles. – Science 294: 364 – 366.
Lohmann, K. J. et al. 2004. Animal behaviour: geomagnetic map used in sea-turtle navigation. – Nature 428: 909 – 910.
McLean, S. et al. 2004. Th e US/UK World Magnetic Model for 2005 – 2010. – NOAA Technical Report NESDIS/NGDC-1.




















