
A solution to modelling vegetation condition for whole-of-landscape conservation planning

- the role of local landscape structure in the vegetation composition of field boundaries - 375Applied Vegetation Science 11: 375-386, 2008doi: 10.3170/2008-7-18486, published online 6 May 2008© IAVS; Opulus Press Uppsala.
What is the role of local landscape structure in the vegetation composition of field boundaries?
Aavik, Tsipe1*; Augenstein, Isabel2,3; Bailey, Debra4,5; Herzog, Felix4,6; Zobel, Martin1,7 & Liira, Jaan1,8
1Institute of Ecology and Earth Sciences, Department of Botany, University of Tartu, 40 Lai St., Tartu EE-51005, Estonia;2UFZ Helmholz Centre for Environmental Research, Theodor-Lieser-Str. 4; DE-06120 Halle, Germany; Current address:
Chair for Strategies of Landscape Management, TU Munich, Germany; 3E-mail [email protected];4Agroscope Reckenholz Tänikon Research Station ART, Reckenholzstrasse 191, CH-8046 Zurich, Switzerland;
5E-mail [email protected]; 6E-mail [email protected]; 7E-mail [email protected]; 8E-mail [email protected]; *Corresponding author; Fax +37 27376222; E-mail [email protected]
AbstractQuestion: How distinct is the flora of field boundaries? How does the structure of field boundaries determine the composi-tion of vegetation?Location: Estonia, six 4 km × 4 km agricultural areas. Methods: We studied the vegetation of fields and field boundaries using 2 m × 2 m sample plots. We estimated the frequency of species in both habitat types, applied an MRPP test to analyse the vegetation composition of field boundaries with various combinations of landscape features (ditches, roads, tree and bush layers) illustrating this by DCA ordination, and used indicator species analysis to determine the characteristic species of each boundary type.Results: Ca. 45% of the flora of field boundaries comprised species found on agricultural land. Most typical species in fields – agrotolerants – were also the most common in field boundaries. The vegetation of road verges and grassy boundaries consisted mainly of disturbance-tolerant species. Woody boundaries were characterised by shade-tolerant and nitrophilous species. Ditch banks included species typical of moist habitats and semi-natural grasslands. Few threatened or protected species were observed.Conclusions: The vegetation composition of field boundaries varied due to the complex effects of landscape structure around and in these boundaries. Plant species in agricultural landscapes can be classified into two broad emergent groups on the basis of their different responses to agricultural disturbances – agrotoler-ant species and nature-value species. Agrotolerant species are promoted by agriculture, nature-value species include rare weeds and habitat specialists. We suggest that high-nature-value species should prevail in monitoring the effects of land-use intensifica-tion on biodiversity rather than total species richness.
Keywords: Agricultural landscape; Agrotolerant species; Ditch bank; Forest edge; Grassy field boundary; Hedgerow; Road verge; Tree-line.
Nomenclature: Flora Europaea (Tutin et al. 2001).
Abbreviation: DCA = Detrended correspondence analysis; MRPP = Multi response permutation procedure.
Introduction
The transition from traditional to intensive agriculture during the last century has resulted in drastic changes in the environmental conditions of agricultural landscapes (Matson et al. 1997; Tilman et al. 2001; Green et al. 2005) and is associated with a steep decline in the bio-diversity of agroecosystems (Andrén 1997; Stoate et al. 2001; Robinson & Sutherland 2002; Foley et al. 2005). The vegetation of agricultural landscapes is affected mostly by large-scale changes in landscape structure, such as the declining area and heterogeneity of natural and semi-natural habitats (Bruun 2000; Fahrig 2003; Kivinen et al. 2006; Liira et al. 2008). Small patches of natural or semi-natural vegetation and semi-natural linear elements (e.g. field boundaries), referred to as green veining (Grashof-Bokdam & Langevelde 2005), are expected to support the maintenance of biodiversity in agroecosystems.
In sensu lato, green veining includes zones of field boundaries and field margin strips (sensu Marshall & Moonen 2002). The field margin strip is defined as the buffering edge zone in a field between the crop and boundary of permanent vegetation, including sown grass or flower strips and farm tracks. The field boundary is a permanent vegetation zone outside agriculturally man-aged areas, e.g. hedgerows, tree-lines, forest edges, ditch verges and road verges (Marshall & Moonen 2002). In the current study we concentrate on field boundaries as the permanent source of biodiversity in the green vein-ing system.
A number of factors affect the vegetation composi-tion in field boundaries. Agricultural land-use intensity seriously threatens the environmental quality of green veining habitats via seepage of fertilisers and herbicides from adjacent fields (Kleijn & Snoeijing 1997; Snoo & van der Poll 1999; Aude et al. 2003). Enhanced nitrogen

376 aavik, t. et al.
and phosphorus concentrations facilitate the growth of competitive perennials and fast-growing ruderals and may cause a decrease in species richness and changes in species and functional composition (Boatman et al. 1994; Kleijn & Snoeijing 1997; Marshall & Moonen 2002; Liira et al. 2008). Due to agricultural disturbance, we could expect compositional overlap between the floras of fields and boundaries. Despite the large number of studies on the compositional pattern and diversity of field bound-ary vegetation (for a review, see Grashof-Bokdam & Langevelde 2005), few studies have attempted to specify the specific support function of field boundaries on plant diversity in comparison with agricultural land (Marshall & Arnold 1995; Kiss et al. 1997; Jobin et al. 1997).
In addition to land-use effects, environmental condi-tions in field boundaries are influenced by small-scale structural features in the landscape near field boundaries. For instance, the presence of trees and shrub layer may provide suitable conditions for shade-tolerant species (Le Coeur et al. 2002; Petit et al. 2004). Regularly mown road verges and field boundaries may compensate the habitat loss for species characteristic of semi-natural grasslands (Hovd & Skogen 2005; Cousins 2006). Drainage ditches enhance the diversity of moisture-demanding species (Blomqvist et al. 2003; Milsom et al. 2004). Studies on field boundary vegetation have usually addressed only on one structural aspect, e.g. the vegetation of hedgerows (McCollin et al. 2000; Boutin et al. 2002; Aude et al. 2003; Deckers et al. 2004), ditches (Blomqvist et al. 2003; Milsom et al. 2004; Manhoudt et al. 2005) or road verges (Tikka et al. 2001; Hovd & Skogen 2005; Cousins 2006). Comparative studies on different types of field bounda-ries are still rare (Freemark et al. 2002). Furthermore, the fact that field boundaries usually encompass more than one structural aspect has mostly been overlooked. For example, the vegetation composition of ditch verges have been shown to depend on presence or absence of trees or shrub layers (Le Coeur et al. 1997).
The importance of field boundaries as the last resort of biodiversity in agricultural landscapes has yet to be evaluated. Particularly, the interactions between the vegetation and the structure of field boundaries are not completely understood. The aim of this study was to analyse the importance of surrounding landscape struc-ture on the field boundary plant species composition. In particular, we asked: how distinct is the flora of field boundaries compared to agricultural fields, and how does the landscape structure affect the species composition of boundaries?
Material and Methods
Study sites
The methodological framework of the study is based on the pan-European project ‘Greenveins’, the aim of which was to detect the relationship between biodiversity, the structure of green veining elements and land-use intensity (Schweiger et al. 2005; Herzog et al. 2006; Dormann et al. 2007; Billeter et al. 2008; Liira et al. 2008). Fieldwork was carried out in six Estonian agricultural landscapes each of which had an area of 4 km × 4 km. Landscape study sites were distributed over the Estonian mainland with the aim to represent various combinations of the gradients of agricultural land-use intensity and landscape structure (Fig. 1). Two sites, Are and Vihtra, feature extensive cattle breeding and most of the agricultural land is used as rotational grassland. Ar-able fields dominate in two landscapes, Väike-Maarja and Ilmatsalu. The last two sites, Viiratsi and Abja-Paluoja, comprise of a mixture of different management regimes, such as extensive cattle breeding, pig farming and in-tensive crop cultivation (Table 1). A group of land users from each landscape was interviewed to assess the land-use intensity (for detailed description of the assessment method, see Herzog et al. 2006). The interview included questions on the area of utilised agricultural land, main crops, the use of fertilizers and pesticides and grazing intensity. The mean area-weighted annual nitrogen use varied from 34 kg/ha in Vihtra to 319 kg/ha in Viiratsi. There were no intensively managed fields (> 150 kg-N/ha) in Vihtra, while in Viiratsi intensive fields formed 67% of agricultural area. The mean number of crops in rotation ranged from 1.8 crops in Are to 7.7 in Ilmatsalu. The mean annual use frequency of pesticides varied from 0.04 in Are to 2.72 applications in Abja.
The maps of landscape structure were digitised on the basis of aerial photos, Estonian Basic Map and cadastral maps provided by the Estonian Land Board. Landscape elements were classified into broad classes, such as forest, grassland, non-greenvein habitats and their boundaries. The total percentage area of green veining in the land-scape ranged from 17% in Ilmatsalu to 33% in Vihtra. Forest was the dominant natural habitat in all study sites. In four landscapes (Vihtra, Are, Abja, Ilmatsalu), open ground ditches or small streams were common, whereas the landscape in Viiratsi was characterised by a number of small lakes. The edge density of natural and semi-natural communities varied from 13 km/km² in Väike-Maarja to 36.9 km/km² in Vihtra. The highest density of hedgerows and tree-lines was noted in Are and Vihtra (0.63 and 0.32 km/km², respectively). Stone piles and small woodlots were present in all landscapes, most notably in Väike-Maarja and Are.

- the role of local landscape structure in the vegetation composition of field boundaries - 377
Vegetation data
Vegetation was sampled in fields and field bounda-ries, typically one plot per landscape element. Fields were classified into rotational grasslands and arable land (including grain, rape or potato fields). Field boundaries were defined as homogeneous semi-natural linear fea-tures with predominantly perennial vegetation on the outer border of fields, being 0.5 m - 5 m wide – grassy field boundaries, ditch banks, road verges etc. From each study site, from 160 (Abja-Paluoja) to 194 (Are) sample plots of 2 m × 2 m were described with a 1:5
ratio of agricultural land to field boundaries. A plot size of 1 m × 4 m was used in narrow boundaries. The total number of sample plots in the six sites was 1019 – 123 from fields and 896 from field boundaries.
Vascular plant species were identified in each sample plot. The abundance of all vascular plant species was estimated on the basis of the Braun-Blanquet scale (van der Maarel 1979). In addition, the small-scale landscape structural features within a 10 m radius from the centre of the plot (the presence-absence of trees or shrubs, ditch and road) were determined during fieldwork and from maps.
Fig. 1. The location of 4 km × 4 km landscape study sites in Estonia.
Table 1. Land-use intensity, landscape structure and species richness of landscape study sites.
Landscape study site Are Vihtra Viiratsi Väike-Maarja Ilmatsalu Abja-Paluoja
Utilised agricultural area (ha) 1046 952 971 1182 1218 1084Intensively managed land (%) 13 0 67 48 43 0Nitrogen fertilization (kg/ha/y) 38 34 319 168 150 102Livestock density (livestock units/ha) 1.6 0.2 0.3 0.9 0.7 0.1Pesticides (applications/year) 0.04 0.57 1.11 0.93 1.96 2.72Edge density (m/km²) 23124 36945 23165 12974 14527 22896Cover of green veining elements (% of study site) 1) Woodland 26.7 27.7 24 20.6 12.3 19.9 2) Semi-natural 3.8 5.8 5.5 2.7 4.7 5.7 3) Total 30.5 33.6 29.5 23.5 17 25.7
Number of plant species 1) Field boundaries 252 213 190 192 183 173 2) Fields 51 60 73 68 57 83 3) Total 254 213 200 196 184 187

378 aavik, t. et al.
Plant species were assigned Ellenberg ecological indicator values of light, soil fertility and moisture. These values estimate the optimal position of a species along particular environmental gradients, such as light, soil nitrogen content, moisture or acidity (Ellenberg et al. 1991). Mean abundance-weighted Ellenberg scores were calculated for each sample plot.
We set a 10% frequency threshold to define species to be frequent in fields or boundary areas. The data on habitat preferences of species are based on Paal (1997). Species frequency data and conservation status were retrieved from Kukk & Kull (2005). Species invasion status was evaluated according to Kukk (2001). Kukk (1999) was used to assess plant species tolerance to hu-man activities and Estonian Red Data Book (Lilleleht 1998) to verify species rarity.
Data analyses
MRPP (Multi-Response Permutation Procedures) based on the Sørensen distance measure (Mielke & Berry 2001) was used to determine the differences in the species composition (based on presence/absence data) among the different combinations of the presence/absence of small-scale landscape structural features at the field boundary (Table 2). Detrended Correspondence Analysis (DCA) was performed to illustrate the variation in plant spe-cies composition in different field boundary types. The presence/absence data was used, and the species that occurred in less than 1% of sample plots were excluded from the ordination analysis. We used an after-the-fact evaluation of the effectiveness of ordination based on relative Euclidean distance (McCune & Grace 2002). We calculated Spearman rank correlation coefficients between ordination axes and the following variables: Ellenberg ecological values of light, soil fertility and moisture; the percentage of natural and semi-natural habitats per landscape; mean nitrogen fertilization rate per landscape; the presence/absence indicator variable of a ditch within 10 m radius, the presence/absence indicator of a road, and the presence/absence indicator of a tree or bush layer.
Indicator species analysis (Dufrene & Legendre 1997) was used to detect species typical of field boundaries characterised by different combinations of small-scale landscape structural features. The boundary types to be compared for characteristic species were derived from the results of MRPP test. The randomness of indicator value distribution within species was tested using Monte Carlo simulation tests (4999 runs). DCA, MRPP and indicator species analysis were carried out using PC-Ord ver. 5.10 (McCune & Grace 2002).
Results
General characteristics of field boundaries
The analysis of the landscape structure within a 10 m radius of the sampling plots showed that most of the boundaries encompassed more than one landscape structural feature. Nearly half (46.8%) of field bounda-ries were ‘pure types’ (consisting of only one landscape structural aspect), while the other half of boundaries were complexes of several landscape features (Table 2). The most common field boundary types were the combina-tion of ditch verge and woody vegetation (27.3%), fol-lowed by road verge (22.4%) and tree-line or hedgerow (19.2%). The least represented field boundary type was the complex of ditch and road verge (4.6%), ditch verge (3%) and grassy field boundary (2.2%).
General vegetation characteristics
A total of 384 vascular plant species were recorded in six landscapes: 170 species were found on arable land and rotational grasslands, and 371 species in field boundaries. Nearly all species (99%) recorded in fields were also present in field boundaries, whereas these spe-cies constituted 45% of field boundary flora.
The most frequent species on agricultural land were Taraxacum officinale (in 56% of fields), Elymus repens (50%) and Cirsium arvense (37%). The main cultivar spe-cies of rotational grasslands – Phleum pratense – was also
Table 2. The complexes of structural features of field boundaries and the sample size (number of plots).
Boundary type Structural characteristic of field boundary (10 m radius) Road Ditch Tree/shrub Sample size %
Grassy field boundary (G) - - - 20 2.2Road verge (R) 1 - - 201 22.4Ditch verge (D) - 1 - 27 3.0Woody boundary, forest edge, tree-line, hedgerow (W) - - 1 172 19.2Woody road verge (WR) 1 - 1 77 8.6Ditch with road (DR) 1 1 - 41 4.6Woody ditch verge (WD) - 1 1 244 27.3Woody ditch verge with road (WDR) 1 1 1 114 12.7
- the role of local landscape structure in the vegetation composition of field boundaries - 379
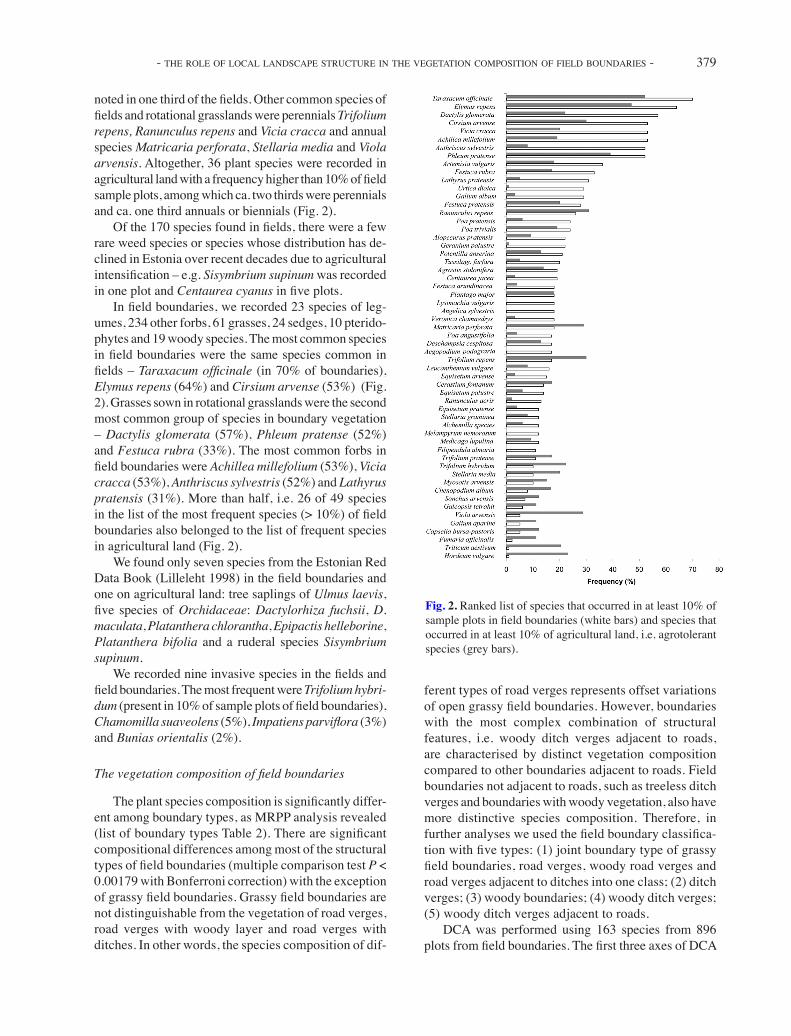
noted in one third of the fields. Other common species of fields and rotational grasslands were perennials Trifolium repens, Ranunculus repens and Vicia cracca and annual species Matricaria perforata, Stellaria media and Viola arvensis. Altogether, 36 plant species were recorded in agricultural land with a frequency higher than 10% of field sample plots, among which ca. two thirds were perennials and ca. one third annuals or biennials (Fig. 2).
Of the 170 species found in fields, there were a few rare weed species or species whose distribution has de-clined in Estonia over recent decades due to agricultural intensification – e.g. Sisymbrium supinum was recorded in one plot and Centaurea cyanus in five plots.
In field boundaries, we recorded 23 species of leg-umes, 234 other forbs, 61 grasses, 24 sedges, 10 pterido-phytes and 19 woody species. The most common species in field boundaries were the same species common in fields – Taraxacum officinale (in 70% of boundaries), Elymus repens (64%) and Cirsium arvense (53%) (Fig. 2). Grasses sown in rotational grasslands were the second most common group of species in boundary vegetation – Dactylis glomerata (57%), Phleum pratense (52%) and Festuca rubra (33%). The most common forbs in field boundaries were Achillea millefolium (53%), Vicia cracca (53%), Anthriscus sylvestris (52%) and Lathyrus pratensis (31%). More than half, i.e. 26 of 49 species in the list of the most frequent species (> 10%) of field boundaries also belonged to the list of frequent species in agricultural land (Fig. 2).
We found only seven species from the Estonian Red Data Book (Lilleleht 1998) in the field boundaries and one on agricultural land: tree saplings of Ulmus laevis, five species of Orchidaceae: Dactylorhiza fuchsii, D. maculata, Platanthera chlorantha, Epipactis helleborine, Platanthera bifolia and a ruderal species Sisymbrium supinum.
We recorded nine invasive species in the fields and field boundaries. The most frequent were Trifolium hybri-dum (present in 10% of sample plots of field boundaries), Chamomilla suaveolens (5%), Impatiens parviflora (3%) and Bunias orientalis (2%).
The vegetation composition of field boundaries
The plant species composition is significantly differ-ent among boundary types, as MRPP analysis revealed (list of boundary types Table 2). There are significant compositional differences among most of the structural types of field boundaries (multiple comparison test P < 0.00179 with Bonferroni correction) with the exception of grassy field boundaries. Grassy field boundaries are not distinguishable from the vegetation of road verges, road verges with woody layer and road verges with ditches. In other words, the species composition of dif-
ferent types of road verges represents offset variations of open grassy field boundaries. However, boundaries with the most complex combination of structural features, i.e. woody ditch verges adjacent to roads, are characterised by distinct vegetation composition compared to other boundaries adjacent to roads. Field boundaries not adjacent to roads, such as treeless ditch verges and boundaries with woody vegetation, also have more distinctive species composition. Therefore, in further analyses we used the field boundary classifica-tion with five types: (1) joint boundary type of grassy field boundaries, road verges, woody road verges and road verges adjacent to ditches into one class; (2) ditch verges; (3) woody boundaries; (4) woody ditch verges; (5) woody ditch verges adjacent to roads.
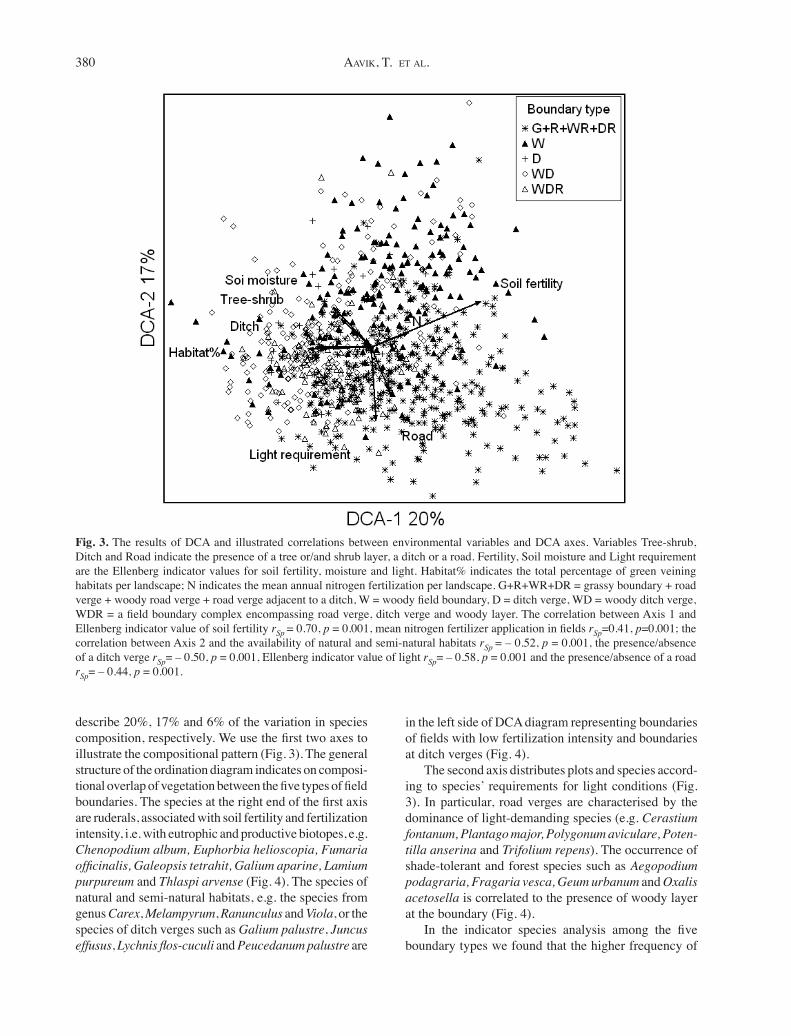
DCA was performed using 163 species from 896 plots from field boundaries. The first three axes of DCA
Fig. 2. Ranked list of species that occurred in at least 10% of sample plots in field boundaries (white bars) and species that occurred in at least 10% of agricultural land, i.e. agrotolerant species (grey bars).
380 aavik, t. et al.
describe 20%, 17% and 6% of the variation in species composition, respectively. We use the first two axes to illustrate the compositional pattern (Fig. 3). The general structure of the ordination diagram indicates on composi-tional overlap of vegetation between the five types of field boundaries. The species at the right end of the first axis are ruderals, associated with soil fertility and fertilization intensity, i.e. with eutrophic and productive biotopes, e.g. Chenopodium album, Euphorbia helioscopia, Fumaria officinalis, Galeopsis tetrahit, Galium aparine, Lamium purpureum and Thlaspi arvense (Fig. 4). The species of natural and semi-natural habitats, e.g. the species from genus Carex, Melampyrum, Ranunculus and Viola, or the species of ditch verges such as Galium palustre, Juncus effusus, Lychnis flos-cuculi and Peucedanum palustre are
in the left side of DCA diagram representing boundaries of fields with low fertilization intensity and boundaries at ditch verges (Fig. 4).
The second axis distributes plots and species accord-ing to species’ requirements for light conditions (Fig. 3). In particular, road verges are characterised by the dominance of light-demanding species (e.g. Cerastium fontanum, Plantago major, Polygonum aviculare, Poten-tilla anserina and Trifolium repens). The occurrence of shade-tolerant and forest species such as Aegopodium podagraria, Fragaria vesca, Geum urbanum and Oxalis acetosella is correlated to the presence of woody layer at the boundary (Fig. 4).
In the indicator species analysis among the five boundary types we found that the higher frequency of
Fig. 3. The results of DCA and illustrated correlations between environmental variables and DCA axes. Variables Tree-shrub, Ditch and Road indicate the presence of a tree or/and shrub layer, a ditch or a road. Fertility, Soil moisture and Light requirement are the Ellenberg indicator values for soil fertility, moisture and light. Habitat% indicates the total percentage of green veining habitats per landscape; N indicates the mean annual nitrogen fertilization per landscape. G+R+WR+DR = grassy boundary + road verge + woody road verge + road verge adjacent to a ditch, W = woody field boundary, D = ditch verge, WD = woody ditch verge, WDR = a field boundary complex encompassing road verge, ditch verge and woody layer. The correlation between Axis 1 and Ellenberg indicator value of soil fertility rSp = 0.70, p = 0.001, mean nitrogen fertilizer application in fields rSp=0.41, p=0.001; the correlation between Axis 2 and the availability of natural and semi-natural habitats rSp = – 0.52, p = 0.001, the presence/absence of a ditch verge rSp= – 0.50, p = 0.001, Ellenberg indicator value of light rSp= – 0.58, p = 0.001 and the presence/absence of a road rSp= – 0.44, p = 0.001.

- the role of local landscape structure in the vegetation composition of field boundaries - 381
several species is associated with particular structural features of field boundaries. The pooled type of bounda-ries that includes grassy boundaries, road verges and the combinations of road verges with ditch and with woody layer is characterised by species typical of open and disturbed habitats: particularly Taraxacum officinale (86% of plots), but also Achillea millefolium, Artemisia vulgaris, Plantago major and Poa annua; or weed spe-cies Capsella bursa-pastoris and Matricaria perforata (Table 3). Most of these species are also common in arable fields and rotational grasslands (Table 3).
The herb layer of woody boundaries (forest edges, tree-lines, hedgerows) is characterised by shade-tolerant and nitrophilous species Elymus repens, Urtica dioica and Rubus idaeus (Table 3).
The list of characteristic species of ditch verges is the longest, including typical species of semi-natural grasslands such as Centaurea jacea, Galium boreale and Ranunculus acris (Table 3), and species characteristic of moist and nutrient-rich soils such as Filipendula ulmaria, Geranium palustre, Aegopodium podagraria
and Angelica sylvestris. None of these species is wide-spread in agricultural land. The only species with higher requirements for moisture and characteristic of woody ditch verges is Deschampsia cespitosa.
Common species in the boundary complex of ditch, road, and woody layer encompass several grassland species – Dactylis glomerata, Leucanthemum vulgare and Medicago lupulina, and species typical of habitats with higher nutrient availability – Cirsium arvense and Lysimachia vulgaris (Table 3).
Several species were revealed to be generalists oc-curring frequently in almost all types of field boundaries (Table 3), such as Phleum pratense and Vicia cracca that occurred in more than 50% of plots. Common graminoids including Festuca pratensis, Poa pratensis and P. trivialis were also often present in all types of field boundaries.
Fig. 4. DCA ordination diagram of field boundary plant species that occurred in at least 1% of sample plots in field boundaries (i.e. 163 species). For correlations see Fig. 3.

382 aavik, t. et al.
Discussion
Agricultural land constitutes ca. 45% of the European Union and 20% of Estonian land area. Even if we focused only on utilised agricultural areas and field boundaries we still recorded nearly 20% of the regional flora of Estonia (1441 vascular plant species; Kukk 1999). Hence, the habitats in agricultural landscapes seem to support a con-siderable proportion of biodiversity. We found, however, that the flora of field boundaries has a large overlap with the flora of agricultural land (ca. 45%), which is consistent with the findings of Marshall (1989) and Jobin et al. (1997)
who also concluded that ca. 40%-50% of the vegetation in hedgerows and forest edges can be found in the adjacent crops. Field boundaries provide thus habitats for weeds, generalists and disturbance-tolerant species, many of which can also be found on adjacent arable land (Marshall 1989; Kleijn & Verbeek 2000; Manhoudt et al. 2005). The frequent occurrence of common, disturbance-tolerant generalist species can be related to the general character-istics of agricultural landscapes, such as relatively higher disturbance frequency and mean fertilization rate and the loss of natural habitats. Those species perform success-fully in agricultural landscapes even when the amount
Table 3. List of indicator species that have significant (P < 0.05) indicator values in five field boundary types, created according to the MRPP test: G+R = pooled type consisting grassy boundary + road verge + woody road verge + road verge adjacent to ditch, W = woody field boundary, D = ditch verge, WD = woody ditch verge, WDR = woody ditch and road verge. Species that were frequent in all types of field boundaries are listed at the end of the Table (Generalist). Agri% = the frequency of a particular species on agricultural land; I.v. = Indicator value; P = significance of randomization test. Species names in bold denote agrotolerant spe-cies, i.e. the species was present in at least 10 % of sample plots on agricultural land.
Species Frequency (%) Agri% Group I.v. P G+R+... W D WD WDR
Taraxacum officinale 56 G+R+... 22 0.004 86 49 63 61 73Artemisia vulgaris 18 G+R+... 21 0.001 56 30 15 14 39Plantago major 16 G+R+... 19 0.006 35 3 0 7 18Achillea millefolium 20 G+R+... 18 0.020 69 20 59 47 65Matricaria perforata 29 G+R+... 13 0.005 31 10 7 10 14Capsella bursa-pastoris 11 G+R+... 11 0.002 13 1 0 0 1Poa annua 2 G+R+... 10 0.006 17 1 4 1 5Elymus repens 50 W 21 0.013 68 79 44 54 55Urtica dioica 1 W 15 0.010 21 51 44 26 25Rubus idaeus - W 13 0.001 3 23 0 8 7Lathyrus pratensis 5 D 23 0.008 23 14 67 39 54Geranium palustre 1 D 21 0.004 10 18 56 36 29Centaurea jacea 2 D 19 0.004 15 8 48 21 32Anthriscus sylvestris 10 D 19 0.014 50 52 74 44 68Ranunculus acris 2 D 17 0.006 11 9 37 17 7Alopecurus pratensis 9 D 16 0.002 14 15 48 30 33Festuca rubra 17 D 16 0.013 39 14 56 27 54Galium boreale 1 D 14 0.004 3 3 26 9 6Aegopodium podagraria - D 13 0.006 12 27 37 21 6Leontodon hispidus - D 12 0.002 1 1 15 0 1Angelica sylvestris - D 12 0.016 3 9 37 36 33Ranunculus auricomus - D 11 0.004 2 2 22 11 9Filipendula ulmaria - D 11 0.007 4 9 30 18 17Potentilla erecta 1 D 10 0.003 1 3 19 7 5Deschampsia cespitosa 13 WD 11 0.026 8 11 19 31 21Melilotus alba 0 WDR 25 0.002 10 0 7 1 38Galium album 2 WDR 20 0.001 24 13 48 33 59Dactylis glomerata 24 WDR 19 0.023 68 48 56 41 75Cirsium arvense 37 WDR 17 0.047 53 51 48 48 68Potentilla anserina 13 WDR 14 0.006 24 9 11 20 38Calamagrostis epigeios 1 WDR 12 0.003 6 4 7 13 26Equisetum palustre 6 WDR 12 0.007 6 3 26 20 33Medicago lupulina 10 WDR 11 0.011 19 2 7 3 25Lysimachia vulgaris - WDR 11 0.017 7 13 19 30 33Leucanthemum vulgare 8 WDR 10 0.019 11 6 19 23 30Phleum pratense 39 Generalist 0.386 52 41 44 57 55Vicia cracca 23 Generalist 0.540 50 51 59 55 59Festuca pratensis 21 Generalist 0.052 31 13 41 27 42Poa pratensis 7 Generalist 0.229 31 19 30 14 32Poa trivialis 19 Generalist 0.683 23 27 15 26 22Ranunculus repens 31 Generalist 0.070 23 20 19 35 24Tussilago farfara 5 Generalist 0.229 20 12 19 24 26

- the role of local landscape structure in the vegetation composition of field boundaries - 383
of natural and semi-natural habitats is negligible. We therefore suggest defining them as ‘agrotolerant species‘ – species that are enhanced by anthropogenic activities and intensive land-use. In contrast, some other studies have shown the distinctiveness of field boundary flora from arable fields (Marshall & Arnold 1995; Kiss et al. 1997). Depending on the land-use intensity and habitat properties, green veining elements might act as refugia for habitat specialists (Cousins & Eriksson 2001; Smart et al. 2002). However, we found that besides relatively large overlap of the vegetation in field boundaries and fields, the boundaries hosted few species of high conservation value and several invasive species.
The present study confirms the importance of small-scale landscape structure (within a 10-m scale) in affect-ing the vegetation of field boundaries (Le Coeur et al. 1997; Baudry et al. 2000; Cherrill et al. 2001). Widely used EUNIS classification (Davies & Moss 2002) is relatively robust and may be well applicable in some landscapes. However, concerning ecological responses of species, it seems that EUNIS system does not carry sufficient information about the environmental vari-ation within field boundaries. A classification of field boundaries encompassing several landscape features (e.g. ditch verge with shrubs and road) into the EUNIS types may be very observer influenced. In studied land-scapes, nearly half of field boundaries had a complex structure, i.e. encompassed more than one landscape feature which means that more sophisticated methods in the boundary classification are required. The results of MRPP analysis indicate that field boundary habitats can be divided broadly into (1) open drier or fresh habitats (boundaries related to road verges and grassy vegetation strips between the fields), (2) open moist habitats (ditch verges), (3) moist habitats with woody vegetation (woody ditch verges), (4) other woody boundaries such as forest edges, tree-lines and hedgerows and (5) complex field boundaries encompassing ditch verge and road verge with woody cover. The reason for applying such a complex habitat classification is the need to take into account the heterogeneity of habitat conditions which enables the co-existence of species with different ecological requirements.
The gradients of soil moisture and light availability seem to be the most critical in the formation of species composition as this can be seen in the distinction among drier boundaries (road verges, grassy boundaries), ditch verges and woody boundaries (i.e. forest edges, tree-lines and hedgerows). The pooled group of road verges and grassy boundaries was characterised by species with higher requirements for light and nitrogen. Most of these are agrotolerant species. Road verges are subject to intensive anthropogenic disturbances – they are mown, bush or tree saplings are cut and the parts closest to the
road are mechanically disturbed by vehicles. They are also characterised by specific ecological conditions such as ample light, frequent drought, and leaching of salts from roads or mineral nutrients from adjacent fields. Therefore, disturbance-tolerant or ruderal species tend to dominate in these habitats (Ullmann & Heindl 1989; Forman & Alexander 1998; Truscott et al. 2005). The function of such road verges as alternative habitats and dispersal corridors for semi-natural grassland species seems very questionable. However, wider road verges can encompass gradual decrease in disturbance and increase in moisture, and may thus still be important habitats for a number species of semi-natural grasslands (Cherrill et al. 2001; Hovd & Skogen 2005; Bratli et al. 2006; Cousins 2006).
Ditch verges are not subject to such intensive me-chanical disturbances, but they are mown at least once every few years and shrubs are cleared. The lower disturbance rate, combined with the removal of more competitive fast-growing species and reduced inflow of nutrients, has enabled the development of diverse plant association composed mainly of perennials (including orchids), otherwise common in permanent grasslands (Paal 1997; Blomqvist et al. 2003; Milsom et al. 2004). The lower parts of ditches are critical for the survival of phreatophytes and moisture-demanding plants (Deckers et al. 2004).
It has been shown that woody linear habitats may contribute to the diversity of forest plant species in agri-cultural landscapes (Baudry et al. 2000; McCollin et al. 2000; Boutin et al. 2002; Aude et al. 2003; Roy & de Blois 2006). Indeed, we recorded several forest species in forest edges, tree-lines or hedgerows. Ditch verges with woody layer provided habitat for species characteristic of paludified forests. However, woody boundaries were also suitable for ruderal species preferring high soil nutrient content, which is an indication of agricultural disturbances and chemical pollution from neighbouring fields (De Blois et al. 2002; Deckers et al. 2004). In ad-dition, the light and soil conditions of hedgerows differ from the conditions in woodland interiors (McCollin et al. 2000) and the survival of woodland species is likely to be limited. The significance of woody field boundaries as habitats or dispersal corridors for forest species may, therefore, be frequently over-estimated.
In EU member states, the biodiversity restoration and monitoring programs in agricultural landscapes started a few years ago or are in the process of preparation. Biodiversity monitoring demands optimal methodologies and indicators to assess the current status of biodiversity and to detect temporal trends (Büchs 2003; Brooks & Kennedy 2004). It has been suggested that species rich-ness estimate per se does not adequately demonstrate the status of biodiversity in agricultural landscapes (Büchs

384 aavik, t. et al.
et al. 2003; Clergue et al. 2005; Waldhardt et al. 2003), because it does not reflect the compositional aspects and the status of species more prone to agricultural distur-bances or habitat loss. Our results confirm the concern that the species of natural and semi-natural communities could be replaced by generalist species, nitrophilous weeds and invasive species, while the total number of species may remain constant or show only small vari-ations. The main effort of biodiversity conservation in agricultural landscapes should evidently be focused on habitat specialists (Waldhardt et al. 2003) – species characteristic of natural and semi-natural habitats as well as of declining weeds. Both these species groups could be joined into the group of ‘high-nature-value species’or ‘conservation-demanding species‘.
The presence of rare or Red Data Book species has been suggested as an alternative indicator for the evalu-ation of diversity in agricultural landscapes (Weibull & Östman 2003; Clergue et al. 2005). However, the results of our study indicate that field boundaries host very few rare, endangered or protected species and consequently, the approach based on rare species might be impractical in the assessment of the biodiversity in agroecosystems.
Therefore, studies that aim to evaluate the impact of agricultural activities on the plant species diversity of green veining habitats have to distinguish between differ-ent species response groups and ecologically reasonable habitat types. Particularly, we suggest the use of a very simple, empirically derived and theoretically motivated species classification based on the response of species to agricultural land-use – agrotolerant species and nature-value species. The last group can be more indicative of detecting the changes in agricultural landscapes.
Acknowledgements. We owe special thanks to the farmers and all the numerous helpers. Particularly we thank Oliver Parrest, Silvia Lotman, Ave Anniste, Viivi Aavik and Piret Annus for participating in the fieldwork. We are grateful to Dr. Robert Szava-Kovac who kindly commented the draft of the paper. Funding was received from the GREENVEINS project within the Energy, Environment and Sustainable development Pro-gramme (FP5) of the European Commission (contract number EVK2-CT-2000-00082), the project AGRIPOPES within EUROCORES program of the European Scientific Foundation and University of Tartu (SF0180098s08).
References
Andrén, H. 1997. Habitat fragmentation and changes in bio-diversity. Ecological Bulletins 46: 171-181.
Aude, E., Tybirk, K. & Bruus Pedersen, M. 2003. Vegetation di-versity of conventional and organic hedgerows in Denmark. Agriculture, Ecosystems & Environment 99: 135-147.
Baudry, J., Burel, F., Thenail, C. & Le Coeur, D. 2000. A holistic landscape ecological study of the interactions between far-ming activities and ecological patterns in Brittany, France. Landscape and Urban Planning 50: 119-128.
Billeter, R., Liira, J., Bailey, D., Bugter, R., Arens, P., Au-genstein, I., Aviron, S., Baudry, J., Bukáček, R., Burel, F., Cerny, M., De Blust, G., De Cock, R., Diekötter, T., Dietz, H., Dirksen, J., Dormann, C., Durka, W., Frenzel, M., Hamersky, R., Hendrickx, F., Herzog, F., Klotz, S., Koolstra, B., Lausch, A., Le Coeur, D., Maelfait, J.P., Op-dam, P., Roubalova, M., Schermann, A., Schermann, N., Schmidt, T., Schweiger, O., Smulders, M.J.M., Speelmans, M., Simova, P., Verboom, J., van Wingerden, W.K.R.E., Zobel, M. & Edwards, P.J. 2008. Indicators for biodiversity in agricultural landscapes: a pan-European study. Journal of Applied Ecology 45: 141-150.
Blomqvist, M.M., Vos, P., Klinkhamer, P.G.L. & ter Keurs, W. J. 2003. Declining plant species richness of grassland ditch banks – a problem of colonisation or extinction? Biological Conservation 109: 391-406.
Boatman, N.D., Rew L.J., Theaker, A.J. & Froud-Williams, R.J. 1994. The impact of nitrogen fertilisers on field margin flora. In: Boatman, N. (ed.) Field margins: Integrating agriculture and conservation. BCPC Monograph no. 58, pp. 209-214. British Crop Protection Council, Alton, UK.
Boutin, C., Jobin, B., Bélanger, L. & Choinière, L. 2002. Plant diversity in three types of hedgerows adjacent to cropfields. Biodiversity and Conservation 11: 1-25.
Bratli, H., Økland, T., Økland, R.H., Dramstad, W.E., Elven, R., Engan, G., Fjellstad, W., Heegaard, E., Pedersen, O. & Solstad, H. 2006. Patterns of variation in vascular plant species richness and composition in SE Norwegian agricul-tural landscapes. Agriculture, Ecosystems & Environment 114: 270-286.
Brooks, T. & Kennedy, E. 2004. Biodiversity barometers. Nature 431: 1046-1047.
Bruun, H.H. 2000. Patterns of species richness in dry grass-land patches in an agricultural landscape. Ecography 23: 641-650.
Büchs, W. 2003. Biotic indicators for biodiversity and sustaina-ble agriculture – introduction and background. Agriculture, Ecosystems & Environment 98: 1-16.
Büchs, W., Harenberg, A., Zimmermann, J. & Weiss, B. 2003. Biodiversity, the ultimate agri-environmental indicator? Potential and limits for the application of faunistic elements as gradual indicators in agroecosystems. Agriculture, Eco-systems & Environment 98: 99-123.
Cherrill, A., Mercer, C., McClean, C. & Tudor, G. 2001. Asses-sing the floristic diversity of hedge networks: a landscape perspective. Landscape Research 26: 55-64.
Clergue, B., Amiaud, B., Pervanchon, F., Lasserre-Joulin, F. & Plantureux, S. 2005. Biodiversity: function and assessment

- the role of local landscape structure in the vegetation composition of field boundaries - 385
in agricultural areas. A review. Agronomy for Sustainable Development 25: 1-15.
Cousins, S.A.O. 2006. Plant species richness in midfield islets and road verges – the effect of landscape fragmentation. Biological Conservation 127: 500-509.
Cousins, S.A.O. & Eriksson, O. 2001. Plant species occur-rences in a rural hemiboreal landscape: effects of remnant habitats, site history, topography and soil. Ecography 24: 461-469.
Davies, C. E. & Moss, D. 2002. EUNIS Habitat Classification. Final Report to the European Topic Centre on Nature Pro-tection and Biodiversity. European Environment Agency, Copenhagen, DK.
De Blois, S., Domon, G. & Bouchard, A. 2002. Factors affecting plant species distribution in hedgerows of southern Quebec. Biological Conservation 105: 355-367.
Deckers, B., Hermy, M. & Muys, B. 2004. Factors affecting plant species composition of hedgerows: relative im-portance and hierarchy. Acta Oecologia 26: 23-37.
complete reference in the reference list would then be:Dormann, C.F., Schweiger, O., Augenstein, I., Bailey, D., Bille-
ter, R., De Blust, G., DeFilippi, R., Frenzel, M., Hendrickx, F., Herzog, F., Klotz, S., Liira, J., Maelfait, J.P., Schmidt, T., Speelmans, M., van Wingerden, W.K.R.E. & Zobel, M. 2007. Effects of landscape structure and land-use intensity on similarity of plant and animal communities. Global Ecology and Biogeography 16: 774-787.
Dufrêne, M. & Legendre, P. 1997. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecological Monographs 67: 345-366.
Ellenberg, H., Weber, H.E., Düll, R., Wirth, V., Werner, W. & Paulißen, D. 1991. Zeigerwerte von Pflanzen in Mittel-europa. Scripta Geobotanica 18.
Fahrig, L. 2003. Effects of habitat fragmentation on Biodi-versity. Annual Review of Ecology and Systematics 34: 487-515.
Foley, J.A., De Fries, R., Asner, G.P., Barford, C., Bonan, G., Carpenter, S.R., Chapin, F.S., Coe, M.T., Daily, G.C., Gibbs, H.K., Helkowski, J.H., Holloway, T., Howard, E.A., Kucharik, C.J., Monferda, C., Patz, J.A., Prentice, I.C., Ramankutty, N. & Snyder, P.K. 2005. Global consequences of land use. Science 309: 570-574.
Forman, R.T.T. & Alexander, L.E. 1998. Roads and their major ecological effects. Annual Review of Ecology and Systematics 29: 207-31.
Freemark, KE., Boutin, C. & Keddy, C. J. 2002. Importance of farmland habitats for conservation of plant species. Conservation Biology 16: 399-412.
Grashof-Bokdam, C.J. & Langevelde, F. 2005. Green veining: landscape determinants of biodiversity in European agricul-tural landscapes. Landscape Ecology 20: 417-439.
Green, R.E., Cornell, S.J., Scharlemann, J.P.W. & Balmford, A. 2005. Farming and the fate of wild nature. Science 307: 550-555.
Herzog, F., Steiner, B., Bailey, D., Baudry, J., Billeter, R., Bukáček, R., De Blust, G., De Cock, R., Dirksen, J., Dor-mann, C., De Filippi, R., Frossard, E., Liira, J., Stöckli, R., Schmidt, T., Thenail, C., Van Wingerden, W. & Bugter, R. 2006. Assessing the intensity of temperate European
agriculture at the landscape scale. European Journal of Agronomy 24: 165-181.
Hovd, H. & Skogen, A. 2005. Plant species in arable field margins and road verges of central Norway. Agriculture, Ecosystems & Environment 110: 257-265.
Jobin, B., Boutin, C. & DesGrandes, J.-L. 1997. Effect of agri-cultural practices on the flora of hedgerows and woodland edges in southern Quebec. Canadian Journal of Plant Science 77: 293-299.
Kiss, J., Penksza, K., Tóth, F. & Kadar, F. 1997. Evaluation of fields and field margins in nature production capacity with special regard to plant protection. Agriculture, Ecosystems & Environment 63: 227-232.
Kivinen, S., Luoto, M., Kuussaari, M. & Helenius, J. 2006. Multi-species richness of boreal agricultural landscapes: effects of climate, biotope, soil and geographical location. Journal of Biogeography 33: 862-875.
Kleijn, D. & Snoeijing, G. I. J. 1997. Field boundary vegeta-tion and the effects of agrochemical drift: botanical change caused by low levels of herbicide and fertilizer. Journal of Applied Ecology 34: 1413-1425.
Kleijn, D. & Verbeek, M. 2000. Factors affecting the species composition of arable field boundary vegetation. Journal of Applied Ecology 37: 256-266.
Kukk, T. 1999. Eesti taimestik (Estonian flora). Estonian Acad-emy of Sciences, Tartu-Tallinn, EE. (In Estonian.)
Kukk, T. 2001. Taimed. Tulnukaid sama palju kui Kesk-Euroopas (Plants. As much invasive species as in Central Europe). In: Eek-Piirsoo, L. (ed.) Võõrliigid Eestis. Kes on tulnuktaimed ja –loomad ning mida nendega peale hakata, pp. 15-20. Estonian Ministry of Environment, Tallinn, EE. (In Estonian.)
Kukk, T. & Kull, T. 2005. Atlas of the Estonian Flora. Institute of Agricultural and Environmental Sciences of the Estonian University of Life Sciences, Tartu, EE.
Le Coeur, D., Baudry, J. & Burel, B. 1997. Field margins plant assemblages: variation partitioning between local and landscape factors. Landscape and Urban Planning 37: 57-71.
Le Coeur, D., Baudry, J., Burel, B. & Thenail, C. 2002. Why and how we should study field boundary biodiversity in an agrarian landscape context. Agriculture, Ecosystems & Environment 89: 23-40.
Liira, J., Schmidt, T., Aavik, T., Arens, P., Augenstein, I., Bailey, D., Billeter, R., Bukáček, R., Burel, F., De Blust, G., De Cock, R., Dirksen, J., Edwards, P.J., Hamerský, R., Herzog, F., Klotz, S., Kühn, I., Le Coeur, D., Miklová, P., Roubalova, M., Schweiger, O., Smulders, M J.M., van Wingerden, W.K.R.E., Bugters, R. & Zobel, M. 2008. Plant functional group composition and large-scale species richness in European agricultural landscapes. Journal of Vegetation Science 19: 3-14.
Lilleleht, V. (ed.) 1998. Eesti Punane Raamat (Estonian Red Data Book). Estonian Academy of Sciences, EE.
Manhoudt, A.G.E., Udo de Haes, H.A. & de Snoo, G.R. 2005. An indicator of plant species richness of semi-natural habi-tats and crops on arable farms. Agriculture, Ecosystems & Environment 109: 166-174.
Marshall, E.J.P. 1989. Distribution patterns of plants associated

386 aavik, t. et al.
with arable field edges. Journal of Applied Ecology 26: 247-257.
Marshall, E.J.P. & Arnold, G.M. 1995. Factors affecting field weed and field margin flora on a farm in Essex, UK. Landscape and Urban Planning 31: 205-216.
Marshall, E.J.P. & Moonen, A.C. 2002. Field margins in northern Europe: their functions and interactions with agriculture. Landscape and Urban Planning 89: 5-21.
Matson, P.A., Parton, W.J., Power, A.G. & Swift, M.J. 1997. Agricultural Intensification and Ecosystem Properties. Science 277: 504-509.
McCollin, D., Jackson, J.I., Bunce, R.G.H., Barr, C.J. & Stuart, R. 2000. Hedgerows as habitat for woodland plants. Journal of Environmental Management 60: 77-90.
McCune, B. & Grace, J.B. 2002. Indicator species analysis. In: Analysis of ecological communities, pp. 198-204. OR, US.
Mielke, P.V., Jr. & Berry, K.J. 2001. Permutation methods: A distance function approach. Springer Series in Statistics. Springer Verlag, New York, NY, US.
Milsom, T.P., Sherwood, A.J., Rose, S.C., Town, S.J. & Runham, S.R. 2004. Dynamics and management of plant communities in ditches bordering arable fenland in eastern England. Agriculture, Ecosystems & Environment 103: 85-99.
Paal, J. 1997. Eesti taimkatte kasvukohatüüpide klassifikatsioon (Classification of Estonian habitat types). Estonian Ministry of the Environment, Tallinn, EE. (In Estonian.)
Petit, S., Griffiths, L., Smart, S.S., Smith, G.M., Stuart, R.C. & Wright, S.M. 2004. Effects of area and isolation of wood-land patches on herbaceous plant species richness across Great Britain. Landscape Ecology 19: 463-471.
Robinson, R. A. & Sutherland, W. J. 2002. Post-war changes in arable farming and biodiversity in Great Britain. Journal of Applied Ecology 39: 157-176.
Roy V. & de Blois, S. 2006. Using functional traits to assess the role of hedgerow corridors as environmental filters for forest herbs. Biological Conservation 130: 592-603.
Schweiger, O., Maelfait, J.P., Van Wingerden, W., Hendrickx, F., Billeter, R., Speelmans, M., Augenstein, I., Aukema, B., Aviron, S., Bailey, D., Bukáček, R., Burel, F., Diekötter, T., Dirksen, J., Frenzel, M., Herzog, F., Liira, J., Roubalova, M. & Bugter, R. 2005. Quantifying the impact of environ-mental factors on arthropod communities in agricultural landscapes across organizational levels and spatial scales. Journal of Applied Ecology 42: 1129-1139.
Smart, S.M., Bunce, R.G.H., Firbank, L.G. & Coward, P. 2002. Do field boundaries act as refugia for grassland plant species diversity in intensively managed agricultural landscapes in Britain? Agriculture, Ecosystems & Environment 91: 73-87.
Snoo, G.R. & van der Poll, R.J. 1999. Effect of herbicide drift on adjacent boundary vegetation. Agriculture, Ecosystems & Environment 73: 1-6.
Stoate, C., Boatman, N.D., Borralho, R.J., Rio Carvalho, C., de Snoo, G.R. & Eden, P. 2001. Ecological impacts of arable intensification in Europe. Journal of Environmental Management 63: 337-365.
Tikka, P. M., Högmander, H. & Koski, P. S. 2001. Road and
railway verges serve as dispersal corridors for grassland plants. Landscape Ecology 16: 659-666.
Tilman, D., Fargione, J., Wolff, B., D’Antonio, C., Dobson, A., Howarth, R., Schindler, D., Schlesinger, W. H., Simberloff, D. & Swackhamer, D. 2001. Forecasting agriculturally dri-ven global environmental change. Science 292: 281-284.
Truscott, A.M., Palmer, S.C.F., McGowan, G.M., Cape, J.N. & Smart, S. 2005. Vegetation composition of roadside verges in Scotland: the effects of nitrogen deposition, disturbance and management. Environmental Pollution 136: 109-118.
Tutin, T.G., Heywood, V.H., Burges, N.A., Valentine, D.H., Walters, S.M. & Webb, D.A. 2001. Flora Europaea. 5 Vols. and CD-ROM Pack. 2nd.ed. Cambridge University Press, Cambridge, UK.
Ullmann, I. & Heindl, B. 1989. Geographical and ecological differentiation of roadside vegetation in temperate Europe. Botanica Acta 102: 261-269.
van der Maarel, E. 1979. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 29: 97-114.
Waldhardt, R., Simmering, D. & Albrecht, H. 2003. Floristic diversity at the habitat scale in agricultural landscapes of Central Europe – summary, conclusions and perspectives. Agriculture, Ecosystems & Environment 98: 79-85.
Weibull, A.-C. & Östman, Ö. 2003. Species composition in agroecosystems: The effect of landscape, habitat, and farm management. Basic and Applied Ecology 4: 349-361.
Received 29 January 2007;Accepted 12 February 2008;
Co-ordinating Editor: A. Moody.
Copyright © 2022 FDOKUMEN




















