
What are the reasons for the Prussian carp expansion in the upper Elbe River, Czech Republic?
14
What are the reasons for the Prussian carp expansion in the upper Elbe River, Czech Republic? O. S LAVI ´ K *† AND L. B ARTOS ˇ ‡ *T G M – Water Research Institute, Podbabska ´ 30, 160 62 Prague 6, Czech Republic and ‡Institute of Animal Production, Ethology Group, 104 00 Prague 10 – Uhrˇı´neˇves, Czech Republic Fifty five Prussian carp Carassius auratus gibelio were radio-tracked weekly along the course of the upper Elbe River, for 15 months. In addition, diurnal movements were observed in three different habitat types (main channel, navigation canal and floodplain areas). Fish species composition from 10 fish ladders was regularly analysed in 1996, 2000 and 2001, and patterns of juvenile (0þ year) fish distribution and abundance were collected in 2000 and 2001, to support telemetric data on longitudinal migrations, and habitat preference within the studied stretch. Downstream migrations dominated in all seasons, being most intensive in spring and reaching a maximum distance of 85 km. Migration stopped when specimens found a floodplain area, where they occupied larger home ranges compared to the main channel. Migrations upstream of up to 2 km were rare, and specimens did not enter fish ladders. Spontaneous reproduction only occurred on the floodplain and its success was very low. The results suggested that neither upstream migrations, nor spontaneous reproduction represented the key mechanism for the Prussian carp invasion into the Elbe catchment area of the Czech Republic. It can be assumed that escapes from aquaculture activities are primarily responsible for its expansion. # 2004 The Fisheries Society of the British Isles Key words: downstream migration; expansion; floodplain area; reproduction; river slope. INTRODUCTION Prussian carp Carassius auratus gibelio (Bloch) is an introduced species in central Europe. The rapid increase in its populations in many European coun- tries has been recorded since the late 1960s and early 1970s. The primary motive for its translocation was for aquaculture (Holcˇı´k, 1991). Holcˇı´k & Z ˇ itnˇan (1978) described its probable transfer from East Europe and Asia into the European part of Russia. In 1954 the Prussian carp was introduced into Szarvas, Hungary for aquaculture, and escaped into the Ko¨ro¨s and the Tisza Rivers. Its subsequent expansion included the whole Danube catchment area in Bulgaria, Romania and Slovakia. Balon (1963) recorded the species in Slovakia in the early 1960s. By the 1970s, Prussian carp had expanded westward through the Dyje and Morava Rivers (Czech Republic; Barusˇ & Lusk, 1978). Recently, †Author to whom correspondence should be addressed. Tel.: þ420 2 20197224; fax: þ420 2 24310759; email: [email protected] Journal of Fish Biology (2004) 65 (Supplement A), 240–253 doi:10.1111/j.1095-8649.2004.00560.x, available online at http://www.blackwell-synergy.com 240 # 2004 The Fisheries Society of the British Isles
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of What are the reasons for the Prussian carp expansion in the upper Elbe River, Czech Republic?

What are the reasons for the Prussian carp expansion in
the upper Elbe River, Czech Republic?
O. SLAVIK*† AND L. BARTOS‡
*T G M – Water Research Institute, Podbabska 30, 160 62 Prague 6, CzechRepublic and ‡Institute of Animal Production, Ethology Group, 104 00
Prague 10 – Uhrıneves, Czech Republic
Fifty five Prussian carp Carassius auratus gibelio were radio-tracked weekly along the course of
the upper Elbe River, for 15 months. In addition, diurnal movements were observed in three
different habitat types (main channel, navigation canal and floodplain areas). Fish species
composition from 10 fish ladders was regularly analysed in 1996, 2000 and 2001, and patterns
of juvenile (0þ year) fish distribution and abundance were collected in 2000 and 2001, to
support telemetric data on longitudinal migrations, and habitat preference within the studied
stretch. Downstream migrations dominated in all seasons, being most intensive in spring and
reaching a maximum distance of 85 km. Migration stopped when specimens found a floodplain
area, where they occupied larger home ranges compared to the main channel. Migrations
upstream of up to 2 km were rare, and specimens did not enter fish ladders. Spontaneous
reproduction only occurred on the floodplain and its success was very low. The results
suggested that neither upstream migrations, nor spontaneous reproduction represented the
key mechanism for the Prussian carp invasion into the Elbe catchment area of the Czech
Republic. It can be assumed that escapes from aquaculture activities are primarily responsible
for its expansion. # 2004 The Fisheries Society of the British Isles
Key words: downstream migration; expansion; floodplain area; reproduction; river slope.
INTRODUCTION
Prussian carp Carassius auratus gibelio (Bloch) is an introduced species incentral Europe. The rapid increase in its populations in many European coun-tries has been recorded since the late 1960s and early 1970s. The primary motivefor its translocation was for aquaculture (Holcık, 1991). Holcık & Zitnan (1978)described its probable transfer from East Europe and Asia into the Europeanpart of Russia. In 1954 the Prussian carp was introduced into Szarvas, Hungaryfor aquaculture, and escaped into the Koros and the Tisza Rivers. Itssubsequent expansion included the whole Danube catchment area in Bulgaria,Romania and Slovakia. Balon (1963) recorded the species in Slovakia inthe early 1960s. By the 1970s, Prussian carp had expanded westward throughthe Dyje and Morava Rivers (Czech Republic; Barus & Lusk, 1978). Recently,
†Author to whom correspondence should be addressed. Tel.: þ420 2 20197224; fax: þ420 2 24310759;
email: [email protected]
Journal of Fish Biology (2004) 65 (SupplementA), 240–253
doi:10.1111/j.1095-8649.2004.00560.x,availableonlineathttp://www.blackwell-synergy.com
240# 2004TheFisheries Society of theBritish Isles

the species represent 4% of the total number of fish caught in the Elbe River inthe Czech Republic (O. Slavık, unpubl. data).It can be assumed that the increasing population density of the Prussian carp
results from its successful reproductive strategy and high ability to colonize newenvironments. The main cause of its reproductive success is usually attributed togynogenesis, where females are able to use sperm of particularly phytophylouscyprinids but even also unspecific salmonids (Penaz et al., 1979). Further factorsin the species successful reproductive strategy include multiple spawning, lowmortality of eggs, long spawning period, high fecundity and early maturation(Holcık, 1980a). Reduction of predator densities due to river channelization andexcessive fishing could also contribute to a rapid increase in Prussian carppopulations (Holcık, 1980b; Holcık & Kmet’, 1986).Reasons for the spatial expansion of Prussian carp are not so clear
(Holcık, 1980a). Its expansion through aquaculture, together with thecommon carp Cyprinus carpio L., is historically known, and has continuedrecently (Lelek & Buhse, 1992; Barus & Oliva, 1995). Intensive migrations ofthe species in riverine systems are another possible explanation. Someauthors have argued that the Prussian carp has high migration ability onthe basis of its frequent occurrence in running waters and high concentra-tions below weirs (Lusk et al., 1977; Barus & Lusk, 1978), however, accuratedata describing migrations of the species are missing. According to Holcık(1980a) Prussian carp penetrate onto floodplain areas during high flows. Inthe Danube River, specimens preferred floodplain areas and slow flowingarms, and were only rarely caught in the main channel (Holcık, 1980a).Available data, however, indicates that the only presently documentedcause of the spatial expansion of Prussian carp is escapes from aquaculturefacilities.This present study addressed the question as to whether high migratory
ability and reproductive success are key factors in the spatial expansion of thePrussian carp in the Elbe River in the Czech Republic. If high migratory activitywere responsible then it should be possible to observe both upstream anddownstream migrations along the main channel of the Elbe River. If reproduct-ive success plays a key role, then the behavioural strategy of phytophylousPrussian carp would be to reach the floodplain areas and high flows would beconsidered as the main factor (Holcık, 1980a).Migrations of 55 individuals of Prussian carp in the channel of the Czech
portion of the Elbe River were observed during 15 months using telemetry.Specimens from three localities (Elbe and Dyje Rivers and the Musov Dam)were tagged, and released into the three types of habitat (main channel,floodplain area and navigation canal) of the Elbe River, in order to comparethe migratory ability of separate populations whose origins were assumed tobe different. The telemetric data were supported by analysis of fish speciescomposition from 10 fish ladders situated on the Elbe River. The data wereregularly collected in 1996, 2000 and 2001. Furthermore, the diurnal behav-iour of Prussian carp was monitored over 15 months. The analysis of pat-terns of distribution and abundance of young-of-the-year (YOY) fish alongthe channel of the Elbe River provided additional information on habitatpreferences.
MIGRATION OF PRUSSIAN CARP 241
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

MATERIAL AND METHODS
STUDY AREA
The study was carried out on the Elbe River, in the Czech Republic. The river rises at1383m above sea level. It has a total length of 1091�47 km with a catchment area of148 000 km2. The Czech portion of the river is 367�95 km and has a catchment area of51 394 km2. The river has channelized beds and the relics of the original floodplain area(e.g. blind arms and oxbows). The river stretch studied was c. 160 km long limited by thefrontier with Germany [Fig. 1(a)], and included also the navigation canal (110 river km)for shipment connecting the Elbe and Vltava Rivers. The canal is 10�1 km long with thedepth between 2�5 and 3m, and width between 18 and 36m. Its banks are reinforced withrocks and concrete with the slope of 1 : 1�5. There are 10 fish ladders, consisting of pools(2� 2m) with surface notches (0�3� 0�3m) and submerged orifices (0�3� 0�3m) in thestudy area (Larinier et al., 2002). Eight locations for YOY collection were chosen[Fig. 1(a)]. Migrations of radio-tagged specimens were observed within an c. 90 km longriver stretch (Fig. 1; 40–130 river km). The average flow throughout the study period was341�1m3 with a winter maximum (1150m3) and a minimum in summer (132m3); riverslope of the observed stretch is shown in Fig. 1(b).
FISH CAPTURE AND TAGGING
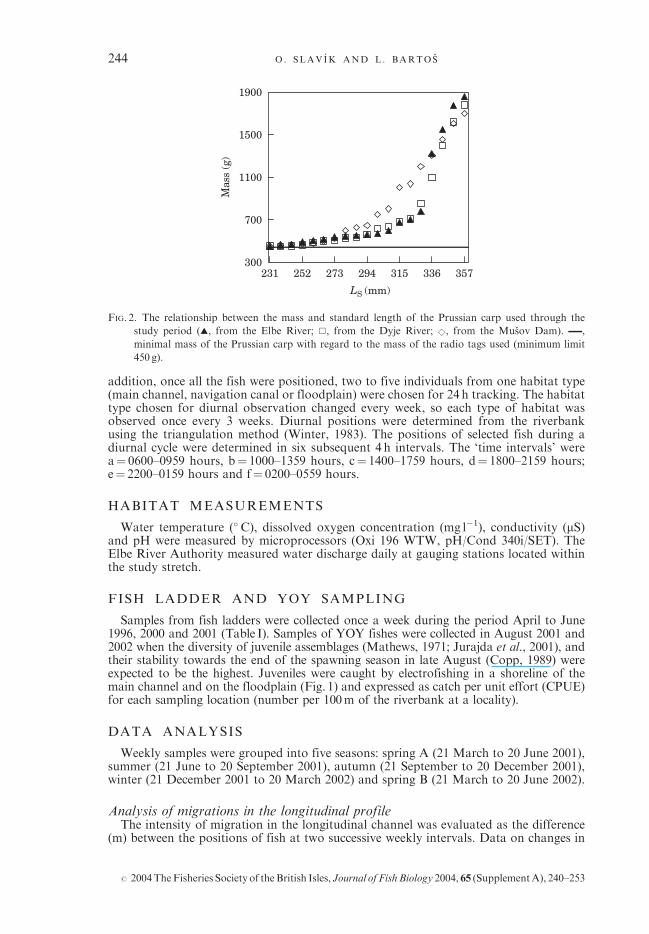
A total of 57 Prussian carp were caught on 18 March 2001 by electrofishing (650V,4A, pulsed D.C.). Specimens from three localities of the Elbe and Dyje Rivers weretagged to compare migrating ability in populations from different environments. Nine-teen fish from the local population were caught in the Elbe River: nine individuals in themain channel (Aa; 50�210 N; 14�280 E), six in the floodplain area (Ab; 50�170 N; 14�310 E)and four in the navigation canal (Ac; 50�190 N; 14�210 E) [Fig. 1(a)]. Nineteen specimenswere caught in the Dyje River, the origin of which was assumed to be the Danube River(Lusk et al., 1977; Barus & Lusk, 1978). The Dyje River (Danube catchment area) is305 km long with the catchment area of 13 418 km2. Fish were caught within the 3 kmlong river stretch (B) [Fig. 1(a)] situated 2–5 km upstream of the confluence with theMorava River. Nineteen specimens were caught in the Musov dam (450 ha) situated onthe Dyje River (C) [Fig. 1(a)] where the species was not observed before filling in 1975(Lusk, 1980) and the origin of which is, therefore, assumed to be from aquaculturesources. Fish from the Morava and Dyje Rivers were transported in tanks with oxygenadded, and 2 days before tag implantation were acclimatized in a cage in the Elbe River.The average mass of fish was 793 g (range 450 to 1855 g), and the average standard length(LS) was 313mm (range 231 to 357mm). All tagged individuals were females, the sexbeing determined during transmitter implantation. Fish were anaesthetized with 2-phenoxy-ethanol (0�2ml l�1). Radio transmitters (MCFT 3EM, 8�9 g in air, 11� 49mm,with operational life of c. 399 days; Lotek Engineering, Inc., Canada) were implantedinto the body cavity through a midventral incision that was closed by three separatestitches, using sterile braided, absorbable suture (Ethicon Coated Vicryl). The massof the transmitter never exceeded 2% of fish body mass in the air (Winter, 1983;Fig. 2). Fish were released at three sites representing different habitat types of theElbe River: Aa (n¼ 27), Ab (n¼ 18) and Ac (n¼ 12) [Fig. 1(a)] after they recoveredtheir equilibrium, and showed spontaneous swimming activity (c. 5min after surgery).Two specimens were caught by anglers and were not replaced. A total of 55 Prussiancarp were used for further analyses.
SAMPLING PROCEDURES
All the fish were tracked weekly during the period 21 March 2001 to 20 June 2002 inthe 90 km long, studied stretch of the Elbe River channel [40–130 river km, Fig. 1(a)].Two radio receivers (SRX_400 receiver firmware versions W5, W31) and a three-elementYagi antenna were used for the telemetry. Fish positions were recorded on a map. In
242 O. SLAV IK AND L. BARTOS
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

MC, 6 km
1
(a)
(b)
2
3
4
5Ac
Aa
Ab
6
7
8
FL, I, 40 km
N
FL, II, 82 kmFL, III, 92 kmFL, IV, 103 kmFL, V, 116 kmFL, VI, 123 kmFL, VII, 130 kmFL, VIII 138 kmFL, IX, 145 kmFL, X, 160 km
GERMANY
CZECH REPUBLIC
VLTAVARIVER
Ab Aa Ac(km 110)(km 114)
(km 110·0)Altitude
Baltic Sea
170,00
160,00
150,00
140,00
130,00
120,00110,00
150 100 50 0·00
STA
TE
BO
UN
DA
RY
CZE
CH
RE
PU
BL
IC -
GE
RM
AN
Y
River kilometre [rkm]
FL, X, 1
60 km
FL, IX, 1
45 km
FL, VIII
, 138
km
FL, VII,
130
km
FL, VI,
123 km
FL, V, 1
16 km
FL, IV, 1
03 km
FL, III,
92 km
FL, II,
82 km
FL, I, 4
0 km
D.P.
(km 116)
MC, 15 kmMC, 58 kmFA, 59 km
MC, 114 kmFA, 117 kmFA, 158 kmMC, 159 km
Prague
PRAGUE
A
B
C
Vlt
ava
Riv
er
VL
TA
VA
RIV
ER
ElbeRiver
ELBE
RIVER
ELBE RIVER
0 10 20 30 40 50 km
1234
56
78
FIG. 1. (a) Map showing the locations where the Prussian carps were caught (A, the Elbe River; B, Dyje
River; C, Musov Dam) and, in detail, the studied stretch of the upper Elbe River, where fish were
released [Aa, main channel (114 river km); Ab, flood plain area (116 river km); Ac, navigation canal
(110 river km)]. Fish ladders (FL) are numbered and river km indicated (e.g. FL I, 40 river km);
YOY were collected in the main channel (MC) or flood plain area (FA) (e.g. MC, ¶, 6 river km).
(b) Upper Elbe River slope.
MIGRATION OF PRUSSIAN CARP 243
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253
addition, once all the fish were positioned, two to five individuals from one habitat type(main channel, navigation canal or floodplain) were chosen for 24 h tracking. The habitattype chosen for diurnal observation changed every week, so each type of habitat wasobserved once every 3 weeks. Diurnal positions were determined from the riverbankusing the triangulation method (Winter, 1983). The positions of selected fish during adiurnal cycle were determined in six subsequent 4 h intervals. The ‘time intervals’ werea¼ 0600–0959 hours, b¼ 1000–1359 hours, c¼ 1400–1759 hours, d¼ 1800–2159 hours;e¼ 2200–0159 hours and f¼ 0200–0559 hours.
HABITAT MEASUREMENTS
Water temperature (� C), dissolved oxygen concentration (mg l�1), conductivity (mS)and pH were measured by microprocessors (Oxi 196 WTW, pH/Cond 340i/SET). TheElbe River Authority measured water discharge daily at gauging stations located withinthe study stretch.
FISH LADDER AND YOY SAMPLING
Samples from fish ladders were collected once a week during the period April to June1996, 2000 and 2001 (Table I). Samples of YOY fishes were collected in August 2001 and2002 when the diversity of juvenile assemblages (Mathews, 1971; Jurajda et al., 2001), andtheir stability towards the end of the spawning season in late August (Copp, 1989) wereexpected to be the highest. Juveniles were caught by electrofishing in a shoreline of themain channel and on the floodplain (Fig. 1) and expressed as catch per unit effort (CPUE)for each sampling location (number per 100m of the riverbank at a locality).
DATA ANALYSIS
Weekly samples were grouped into five seasons: spring A (21 March to 20 June 2001),summer (21 June to 20 September 2001), autumn (21 September to 20 December 2001),winter (21 December 2001 to 20 March 2002) and spring B (21 March to 20 June 2002).
Analysis of migrations in the longitudinal profileThe intensity of migration in the longitudinal channel was evaluated as the difference
(m) between the positions of fish at two successive weekly intervals. Data on changes in
1900
1500
1100
700
300231 252 273 294 315 336 357
Mas
s (g
)
LS (mm)
FIG. 2. The relationship between the mass and standard length of the Prussian carp used through the
study period (~, from the Elbe River; &, from the Dyje River; &, from the Musov Dam). ,
minimal mass of the Prussian carp with regard to the mass of the radio tags used (minimum limit
450 g).
244 O. SLAV IK AND L. BARTOS
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

TABLEI.
Totalnumber
offishes
andthepercentageofPrussiancarp
andother
speciescollectedfrom
fish
laddersin
1996,2001and2002
andyoung-of-the-yearcollectedfrom
thefloodplain
areas(FA)andthemain
channel
(MC)oftheupper
ElbeRiver
Number
of
fishes
(n)
Year
Prussiancarp
(%)
Phytophilous
cyprinids(%
)Other
cyprinids(%
)Other
species(%
)
Fishladders
2121
1996
00� 30
34� 80
64� 90
Adultsonly
5537
2001
0� 02
5� 20
93� 10
1� 68
2724
2002
03� 80
91� 20
5� 00
Environments
FA;1674
2001
0� 58
36� 62
50� 10
12� 70
YOY
FA;1700
2002
0� 57
55� 00
34� 70
9� 73
MC;1281
2001
08� 10
72� 30
19� 60
MC;1017
2002
04� 90
90� 40
4� 70
M IGRATION OF PRUSSIAN CARP 245
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

fish positions within the channel were used to analyse downstream or upstream migra-tions. All tagged specimens were observed weekly.
Analysis of fish movements during 24 hThe distance (m) between fish positions at two subsequent 4 h intervals during a
diurnal cycle were used to determine the intensity of movement during a diurnal cycle.Only data from the diurnal observations were used in an analysis describing a homerange and its size. Two to five tagged individuals were selected weekly for this observa-tion. Individuals that migrated in one direction only during the whole 24 h were excludedfrom further analysis. Instead of estimating the home range size in the traditional way,e.g. as a fixed kernel estimation using least squares cross-validation (Seaman et al., 1998),the size of the Prussian carp home range was determined as the set of all specimen move-ments during the 24 h; home range¼ (downstream distanceþupstream distance)� (leftside distanceþ right side distance).
STATISTICAL ANALYSIS
Statistical analyses were performed with the aid of the Statistical Analysis System (SAS2 V. 8.2) package. ANOVA cannot be used for designs that are unbalanced, i.e. where thenumber of observations for the different combinations of class variables is unequal.Hence, two general linear models (GLM) were used instead (Cody & Smith, 1997).Data were square root transformed for normality before GLM analysis. Class andmodel definitions as well as result characteristics, such as d.f. and F values, are identicalto those used with ANOVA (SAS System).
The factors used in the GLM I analysis of migrations of fish within the longitudinalchannel were classes: ‘season’ (spring A, B; summer; autumn; winter), ‘habitat’ (mainchannel; navigation canal; floodplain area) and ‘fish origin’ (Elbe River; Dyje River;Musov Dam). Migratory intensity, river slope, flow, water temperature and dissolvedoxygen were the dependent variables.
The factors in the GLM II analysis of fish movements during 24 h were classes: ‘season’(spring A, B; summer; autumn; winter), ‘habitat’ (main channel; navigation canal; flood-plain area) and ‘time interval’ (a; b; c; d; e; f). ‘Interval’ was nested to ‘season’ and‘habitat’. All interaction terms were tested, but are not reported unless statisticallysignificant. Intensity of movement, the home range size, water temperature and dissolvedoxygen were dependent variables.
Variables entered the analysis as a sum of observations per class as defined in theGLM. The arithmetic mean for a group may not accurately reflect response for thatgroup in unbalanced designs with more than one effect, since it does not take othereffects into account. Least-square means are, in effect, within-group means appropriatelyadjusted for the other effect in the model (further referred as ‘adjusted means’). There-fore, adjusted means were used in the model output. Adjusted means were computed foreach class, and differences between classes were tested by a t-test. Relationships betweenvariables were studied by computing residual Spearman correlation coefficients (i.e. rankcorrelation between the adjusted data, Table II).
RESULTS
SEASONAL ASPECTS OF MIGRATIONS IN THELONGITUDINAL PROFILE
In the longitudinal profile, downstream migration of the Prussian carp in theriver channel was significantly dominant and reached a maximum distance of85 km. By contrast upstream migration was only up to 2 km. Differencesin intensity of downstream (F65,1218, P< 0�0001) and upstream (F65,1218,
246 O. SLAV IK AND L. BARTOS
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

P< 0�0001) migration were supported by GLM I significance. Furthermore,migratory intensity in both directions varied according to season [Fig. 3(a)].The intensity of downstream (F4,1218, P< 0�0001) and upstream (F4,1218,
TABLE II. Residual Spearman rank correlation between environmental variables andintensity of the Prussian carp migrations across seasons
Environmental variables
Prussian carp migration River slope (S) River flow (Q)
Spring A Downstream 0�58*** 0�05 NSn¼ 356 Upstream �0�06 NS �0�001 NSSummer Downstream 0�5*** 0�06 NSn¼ 315 Upstream �0�06 NS �0�02 NSAutumn Downstream 0�45** 0�04 NSn¼ 212 Upstream �0�1 NS �0�03 NSWinter Downstream 0�4* 0�03 NSn¼ 233 Upstream �0�01 NS �0�02 NSSpring B Downstream 0�7*** 0�16 NSn¼ 103 Upstream �0�15 NS �0�03 NS
*P< 0�05, **P< 0�001, ***P< 0�0001. NS, non-significant.
(a) (b)
(c)
0
0
2
4
9
6
8
14
12
10
0
2
4
6
8
14
12
10
Inte
nsi
ty o
f m
igra
tion
s
Spring A Summer Autumn Winter Spring B Main channel Navigation canal
Spring A Summer Autumn Winter Spring B
45
36
27
18
Fis
h oc
curr
ence
in d
iffe
ren
t ha
bita
t ty
pes
FIG. 3. Intensity (mean� S.E. of transformed square root data) of Prussian carp migrations ( , downstream
and &, upstream) between (a) seasons and (b) habitat types and (c) changes in habitat preference
across seasons (~, flood plain area, &, main channel and ^, navigation canal).
MIGRATION OF PRUSSIAN CARP 247
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

P< 0�0001) migrations was high during both springs (A, B). No statisticaldifferences, either between upstream or downstream migrations were recordedin summer, autumn and winter, nor was there any difference among seasons.Similarly, neither downstream (F2,1218, P¼ 0�1) nor upstream (F2,1218, P¼ 0�1)migrations showed dependence on the origin of tagged fish (Elbe and DyjeRivers and Musov Dam).Where the main channel and navigation canal were compared, a higher
intensity of migrations occurred in the main channel, where the downstreamorientation was clearly dominant [F1,1218, P< 0�0093, Fig. 3(b)]. In the naviga-tion canal, migrations in both directions were similar suggesting that fish in thislentic environment were not oriented in a particular direction. In any season,downstream migration of fish mostly stopped when they found a floodplainarea, where they stayed for the remaining part of the study period. This isshown by a gradual increase in the number of tagged specimens recorded asoccupying the floodplain area from spring A to spring B [Fig. 3(c)].
MIGRATIONS IN THE LONGITUDINAL PROFILE V . RIVERSLOPE AND FLOW
The intensity of seasonal migrations, in relation to river slope, reached amaximum in spring B (F65,1218, P< 0�0004; Fig. 4 and Table II). By contrast, thevariability in river slope passed was not significant during weak upstreammigrations (F65,1218, P¼ 0�1). No relationship was recorded between the inten-sity of migration in either direction and flow (Table II).
ANALYSES OF FISH PATTERNS FROM FISH LADDERS
Within the 120 km long river stretch of the Elbe River, 10 382 individuals offish from 10 fish ladders were caught and analysed. Although cyprinids werenumerous in the fish ladders, only one Prussian carp was found in 2001 (Fig. 1,FL V, 116 river km), and other phytophylous species only rarely occurred in theladders (Table I). Low occurrence of individuals in the ladders indicated theirvery weak effort to migrate upstream.
0·10
0·08
0·06
0·04
0·02
0·00
Riv
er s
lope
Spring A Summer Autumn Winter Spring B
FIG. 4. Intensity (mean� S.E. of transformed square root data) of Prussian carp downstream migrations
in relation to river slope by season.
248 O. SLAV IK AND L. BARTOS
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

ANALYSIS OF YOY ASSEMBLAGES
Only four and three, YOY Prussian carp were caught in 2001 and 2002respectively, in the river stretch where migrations of adults were observed(Table I). All individuals were found in the oxbows of the floodplain thatwere connected with the main channel. This reinforces the assumption thatsuccessful reproduction only takes place in the floodplain.
THE HOME RANGE SIZE, HABITAT PREFERENCE ANDDIURNAL BEHAVIOUR
The size of the home range differed according to habitat type, being largest infloodplains, smaller in the main channel and minimal in the navigation canal(F2,720, P< 0�0001; Fig. 5). The intensity of movement by habitat type, seasonand time interval was tested. Interactions between these factors were significant(F60,720, P< 0�05), and therefore could not be analysed separately. In the flood-plain area, intensity of movements significantly differed from the main channeland navigation canal, whilst in the channel and canal, intensity of movementswas very similar (Fig. 6). During the two springs, differences in intensity ofmovement among all three habitats were relatively small. The exceptionoccurred in the floodplain area during morning intervals, when intensity ofmovement increased [Fig. 6(a), (e)]. In summer, the intensity of Prussian carpmovements in the floodplain areas increased during the night, and culminatedearly in the morning. In the main channel and navigation canal these move-ments occurred mainly during the afternoon and at dusk [Fig. 6(b)]. In autumnand winter, the trend in movements was similar in all three habitats, although itwas significantly more intensive in the floodplain areas [Fig. 6(c), (d)].
DISCUSSION
The results of these investigations suggest that upstream migration ofPrussian carp is not the key factor influencing its occurrence in the ElbeRiver. Tagged fish neither entered the fish ladders, nor showed the effort needed
4·0
3·6
3·2
2·8
2·4
2·0Navigation canal Main channel Flood plain area
Hom
e ra
nge
siz
e
FIG. 5. The size of the Prussian carp home range (mean� S.E. of transformed square root data) in
different habitat types.
MIGRATION OF PRUSSIAN CARP 249
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

0
2
4
6(a)
(b)
(c)
(d)
(e)
0
2
4
6
0
2
4
6
0
2
4
6
0
Time (hours)
2
4
6
Inte
nsi
ty o
f m
ovem
ents
0600
–095
910
00–1
359
1400
–175
918
00–2
159
2200
–015
902
00–0
559
FIG. 6. Intensity of the Prussian carp movements during diurnal cycle by seasons (a) spring A,
(b) summer, (c) autumn, (d) winter and (e) spring B (&, main channel; ~, navigation canal;
&, flood plain area).
250 O. SLAV IK AND L. BARTOS
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

to pass through reaches of higher river slope. Comparison of migratory activityin the main channel and navigation canal showed that increasing river slopeacted as a barrier to upstream migrations. In the canal, where the river slopeapproached zero, fish migrated in both directions, whilst in the main channel,downstream migration clearly dominated [Fig. 3(b)]. Upstream migration,which was originally thought to be the main mechanism of the Prussian carpexpansion in the Morava and Dyje Rivers, was supported by the high con-centration of specimens below weirs, and by their occurrence in fast flowingstretches (Lusk et al., 1977; Barus & Lusk, 1978). Later authors did not confirmthese findings; in the Morava River, the species represented only 1�2% of thetotal number of fishes caught below the weirs during the spring spawningseason (Jurajda et al., 1998). It can be assumed that short distance, upstreammovements presumably reflected increasing activity by Prussian carp searchingfor suitable spawning areas in spring, and local changes in spatial distributionduring other seasons. It would also correspond with the data showing thepreference for slow flowing habitats by this species (Nikolskij, 1953; Holcık,1980a, b).Weakening of migratory instinct may be considered as another reason
responsible for the weak upstream migrations recorded in the Elbe River. Forexample, Harcup et al. (1984) discussed an attenuation and extinction ofmigratory instinct in salmonids that occupied environments where free migra-tion along the river channel was permanently restricted. A similar process inPrussian carp populations may arise through a different mechanism. Based ondata compiled on differences in behaviour, Barus & Oliva (1995) distinguishedmonosexual populations as migratory and bisexual populations as stationaryforms in the original area of occurrence of the species. This assumption wouldalso correspond with the theory that the Prussian carp was autochthonous incentral Europe (Marsili, 1726; Mahen, 1931; Berg, 1949; Barus & Oliva, 1995)and in the Danube catchment area (Balon, 1966), and that its invasion pre-sumably involves only introduced monosexual forms. In addition, in 1995, thefirst recordings in the Czech Republic were of males from the Rivers Moravaand Dyje (S. Lusk, pers. comm.). Based on available data contemporary popu-lations of the Prussian carp in this area probably slowly transformed to thebisexual stationary form. In such a case, it would be possible that during the1960s and 1970s, the species undertook intensive migrations in the Danubecatchment area.Downstream migration of the Prussian carp was a highly significant factor in
its behaviour, and no differences in migratory behaviour were recorded amongindividuals from different original localities (Elbe and Morava Rivers and theMusov Dam). Individuals stopped downstream migration when they found afloodplain area, and had not left this habitat by the end of the study period[Fig. 3(c)]. It appears probable that the main channel of the Elbe River, in theCzech Republic, with its high slope, does not represent suitable habitat forPrussian carp, whilst the floodplain areas do. Significant divergences inbehaviour of individuals in different types of habitats were recorded. IndividualPrussian carp showed larger home ranges and higher movement activity inshallow floodplain areas with submerged vegetation and low velocities, ascompared with those in the main channel and navigation canal with channelized
MIGRATION OF PRUSSIAN CARP 251
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

riverbanks without vegetation. On the contrary, no differences in the size of thehome range and movement activity were recorded between the channel andcanal. The unifying effect of the channelized riverbed on the behaviour ofPrussian carp could also be a reason why no differences in behaviour of speci-mens from different original localities were observed. A similar effect of riverchannelization on behaviour of fishes has been described (Annett, 1998; Slavık& Bartos, 2001).No relationship between variability in flow and migrations of Prussian carp was
found. The relationship between flow ascent and penetration of Prussian carp intothe floodplain area described by Holcık (1980a, b) probably reflected the usualreproductive strategy of phytophilous species (Balon, 1975) in lowland rivers.Very low numbers of juvenile Prussian carp were caught in the studied stretch
of the Elbe River, and these only occurred in the floodplain areas. This agreeswith data from the Morava River, where YOY assemblages from the mainchannel and the floodplain area were compared: in the floodplain area, Prussiancarp juveniles were present, whilst in the main channel they were absent(Jurajda et al., 2001). In the Dyje River floodplain, Prussian carp YOY evenrepresented the second most numerous species (Bartosova & Jurajda, 2001). It isnot clear, why such a small number of juveniles were recorded during thepresent study. Phytophilous cyprinids, however, were the least numerous repro-ductive group in the Elbe River, (O. Slavık, unpubl. data), and the resultingabsence of suitable spawning partners for gynogenetic females could be a reasonfor the low spawning success.Aquaculture appeared to be responsible for the recruitment of the Prussian
carp in the Elbe River. Although records of the production of this species inCzech aquaculture are not available, values are estimated in hundreds of tonnes(J. Vostradovsky, pers. comm.). Direct escape of the Prussian carp during ponddraw down, and its introduction by anglers together with juveniles of thecommon carp into rivers, are repeated every year. In the upper part of theCzech portion of the Elbe River, spontaneous expansion of the Prussian carpinto the tributaries would not be expected because of the higher river slopes thatlimit its upstream migration. Similarly, there is no reason to expect a rapid,spontaneous increase in population density of the species due to the limitedfloodplain areas connected with the main channel that represents the onlysuitable habitat for its reproduction.
We thank J. Holcık for kind submission of the literature, E. Thorstad and an anonymousreferee for their constructive comments, P. Horky and the technical staff at the WaterResearch Institute in Prague for valuable assistance during the experimental period,P. Jurajda for capture of fish in the Dyje River and S. Lusk and J. Vostradovsky for valuableassistance during the project planning. The study was financially supported by Ministryof Environment of the Czech Republic within the frame of the VaV/510/1/99 project.
References
Annett, A. C. (1998). Hunting behaviour of Florida large mouth bass, Micropterussalmoides floridanus, in a channelized river. Environmental Biology of Fishes 53,75–87.
Balon, K. E. (1963). K nalezu karase strıbriteho eurasijskeho. Ziva 11, 205–206 (in Czech).
252 O. SLAV IK AND L. BARTOS
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253

Balon, K. E. (1966). Ryby Slovenska. Bratislava: Obzor (in Slovak).Balon, K. E. (1975). Reproductive guilds of fishes: a proposal and definitions. Journal of
the Fisheries and Research Board of Canada 32, 821–864.Bartosova, S. & Jurajda, P. (2001). A comparison of 0þ fish communities in borrow pits
under different flooding regime. Folia Zoologica 4, 305–315.Barus, V. & Lusk, S. (1978). Karas strıbrity – novy druh v nasich vodach. Rybarstvı 4,
80–81 (in Czech).Barus, V. & Oliva, O. (1995). Mihulovci a ryby – Petromyzontes a Osteichthyes II. Praha:
Academia (in Czech, with English summary).Berg, L. S. (1949). Ryby presnych vod SSSR i sopredelnych stran 2. Moskva, Leningrad:
Izdanie AN SSSR (in Russian).Cody, R. P. & Smith, K. J. (1997). Applied Statistics and the SAS Programming Language.
Upper Saddle River, NJ: Prentice-Hall.Copp, G. H. (1989). The habitat diversity and fish reproduction function of floodplain
ecosystems. Environmental Biology of Fishes 26, 1–27.Harcup, M. F., Williams, R. & Ellis, M. D. (1984). Movements of brown trout, Salmo
trutta L., in the River Gwyddon, South Wales. Journal of Fish Biology 24, 415–426.Holcık, J. (1980a). Carassius auratus (Pisces) in the Danube River. Acta Scientiarum
Naturalum Academiae Scientarum Bohemoslovacae – Brno 14, 1–43.Holcık, J. (1980b). Possible reason for expansion of Carassius auratus (Linnaeus, 1758)
(Teleostei, Cyprinidae) in the Danube River Basin. Internationale Revue dergesamten Hydrobiologie 65, 673–679.
Holcık, J. (1991). Fish introduction in Europe with particular reference to its central andeastern part. Canadian Journal of Fisheries and Aquatic Sciences 48 (Suppl. 1), 13–23.
Holcık, J. & Kmet’, T. (1986). Simple models of the population dynamics of some fishspecies from the lower reaches of the Danube. Folia Zoologica 35, 183–191.
Holcık, J. & Zitnan, R. (1978). On the expansion and origin of Carassius auratus inCzechoslovakia. Folia Zoologica 27, 279–288.
Jurajda, P., Hohausova, E. &Gelnar,M. (1998). Seasonal dynamics of fish abundance belowa migration barrier in the lower regulated Morava River. Folia Zoologica 3, 215–223.
Jurajda, P., Reichard, M., Hohausova, E. & Cerny, J. (2001). Comparison of 0þ fishcommunities between regulated-channelized and floodplain stretches of the RiverMorava. Archivum fur Hydrobiology, Large Rivers 12 (Suppl. 135), 187–202.
Larinier, M., Travade, F. & Porcher, J. P. (2002). Fish ways: biological basis, design criteriaand monitoring. Bulletin Francais de la Peche et de la Pisciculture 364 (Suppl.), 54–82.
Lelek, A. & Buhse, G. (1992). Fische des Rheins. Berlin: Springer-Verlag.Lusk, S. (1980). Development of the fish population in the Musov reservoir in the first
year after filling. Folia Zoologica 30, 249–261.Lusk, S., Barus, V. & Vesely, V. (1977). On the occurrence of Carassius auratus in the
Morava River drainage area. Folia Zoologica 26, 377–381.Mahen, J. (1931). Prıspevek k systematice ryb kaprovitych. Sbornık klubu prırodovedcu
Brno 12, 33–47 (in Czech).Marsili, A. F. (1726). Danubius pannonico-myscus obsevationibus. . . Tomus IV. Amsterdam:
Hagae Comitum.Mathews, C. P. (1971). Contribution of young fish to total production of fish in the River
Thames near Reading. Journal of Fish Biology 3, 157–180.Nikolskij, G. V. (1953). Reka Amur i jeje ryby. Chabarovsk: Knizneje izdanije.Penaz, M., Rab, P. & Prokes, M. (1979). Cytological analysis, gynogenesis and early
development of Carassius auratus gibelio. Acta Scientiarum Naturalum AcademiaeScientiarum Bohemoslovacae – Brno 13, 1–33.
Seaman, D. E., Griffith, B. & Powel, R. A. (1998). KERNELHR: a program forestimating animal home ranges. Wildlife Society Bulletin 26, 95–100.
Slavık, O. & Bartos, L. (2001). Spatial distribution and temporal variance of fishcommunities in the channelized and regulated Vltava River (Central Europe).Environmental Biology of Fishes 61, 47–55.
Winter, J. D. (1983). Underwater biotelemetry. In Fisheries Techniques (Nielsen, L. A. &Johnsen, D., eds), pp. 371–395. Bethesda, MD: American Fisheries Society.
MIGRATION OF PRUSSIAN CARP 253
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 65 (SupplementA), 240–253




















