
Water entry into detached root systems saturates with increasing externally applied pressure; a...
12
PHYSIO8911 P891 14-05-02 10:14:00 PDF mh a203891 PPL1O342 UNCORRECTED PROOF PHYSIOLOGIA PLANTARUM 114: 000. 2002 Copyright C Physiologia Plantarum 2002 Printed in Denmark – all rights reserved ISSN 0031-9317 Water entry into detached root systems saturates with increasing externally applied pressure; a result inconsistent with models of simple passive diffusion R.J.Neil Emery a, * and Christophe Salon a a Institut National de la Recherche Agronomique, Unite ´ de Malherbologie et Agronomie 17 Rue Sully, BP 86510, 21065 Dijon CEDEX, France b Biology Department, Trent University, Peterborough, Ontario, K9J 7B8, Canada *Corresponding author, e-mail: nemery/trentu.ca Received 14 August 2001; revised 20 August 2001; in final form 18 February 2002 The most widely accepted model of radial water entry from the soil into the xylem of roots is based on principles of ordi- nary passive diffusion. However, long-standing problems with this model remain unresolved, which concern variable intrinsic properties of conductivity, L p , changing reflection coefficients, s, and inaccurate resolution of osmotic differentials between the soil and xylem. Our study re-examined pressure flow re- lationships in isolated roots of tomato (Lycopersicon escu- lentum Mill. cv. Montfavet), pea (Pisum sativum cv. Baccara) and soybean (Glycine max L. Merryl cv. Essor) manipulated in a pressure chamber. In addition to problems previously rec- Introduction There has been substantial progress concerning the na- ture and location(s) of the greatest resistance(s) to the uptake and transport of water through roots (Steudle and Peterson 1998, Tyerman et al. 1999). However, some fundamental details remain uncharacterized. This may be due, at least in part, to problems with the application of current quantitative models to data of water flow through roots. The ordinary passive diffusion model Quantitative data is often acquired from experiments de- scribing the transport of water through whole root sys- tems by analysing flow of xylem from root systems sub- jected to artificially raised pressure in chambers designed to accommodate roots still in potted soil (Fiscus 1977, Miller 1985, Passioura 1988, Markhart and Smit 1990, Maggio and Joly 1995, Jackson et al. 1996). The most widely accepted physical model applied to the flow data Physiol. Plant. 114, 2002 1 ognized with the simple passive diffusion model, a new con- flict, flow saturation, was observed at high pressures. Experi- ments revealed that the plateau in flow, J max seen at high pressures followed natural rhythms diurnally and develop- mentally, and was not due to root damage or unnatural flow restriction. Near the end of the photoperiod, J max closely cor- related with root dry mass. The above inconsistencies between observations in pressure-flow kinetics and ordinary passive diffusion indicate that either the current model should be ad- justed or a new model should be proposed. generated from these experiments is analogous to models of ordinary passive diffusion across a membrane (Nobel 1991). J J Ω P j (C o j ª c i j ) [1] Where J j is flux of species j across the membrane of a given area (quantity/area/time, i.e. m 3 /m 2 /s), P j is the permeability coefficient of species j (length/time, i.e. m/ s) and (c j æ – cji) expresses the driving force across the membrane (referring here to the concentration of j out- side and inside). The analogous root model is suitable for a system of membranes (Fiscus 1977, Passioura 1988, Jackson et al. 1996): J Ω L p (DP ª s Dp) [2] Where L p is the collective membrane conductivity (l/ time, m s ª1 ) and (DP– s Dp) is the driving force sum- ming up components of water potential. DP and Dp are Q1 Q2
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Water entry into detached root systems saturates with increasing externally applied pressure; a...

PHYSIO8911 P891 14-05-02 10:14:00 PDF mh a203891 PPL1O342
UN
CO
RR
ECTE
DPR
OO
F
PHYSIOLOGIA PLANTARUM 114: 000. 2002 Copyright C Physiologia Plantarum 2002
Printed in Denmark – all rights reserved ISSN 0031-9317
Water entry into detached root systems saturates with increasingexternally applied pressure; a result inconsistent with models of simplepassive diffusion
R.J.Neil Emerya,* and Christophe Salona
aInstitut National de la Recherche Agronomique, Unite de Malherbologie et Agronomie 17 Rue Sully, BP 86510, 21065 Dijon CEDEX,FrancebBiology Department, Trent University, Peterborough, Ontario, K9J 7B8, Canada*Corresponding author, e-mail: nemery/trentu.ca
Received 14 August 2001; revised 20 August 2001; in final form 18 February 2002
The most widely accepted model of radial water entry fromthe soil into the xylem of roots is based on principles of ordi-nary passive diffusion. However, long-standing problems withthis model remain unresolved, which concern variable intrinsicproperties of conductivity, Lp, changing reflection coefficients,s, and inaccurate resolution of osmotic differentials betweenthe soil and xylem. Our study re-examined pressure flow re-lationships in isolated roots of tomato (Lycopersicon escu-lentum Mill. cv. Montfavet), pea (Pisum sativum cv. Baccara)and soybean (Glycine max L. Merryl cv. Essor) manipulatedin a pressure chamber. In addition to problems previously rec-
Introduction
There has been substantial progress concerning the na-ture and location(s) of the greatest resistance(s) to theuptake and transport of water through roots (Steudleand Peterson 1998, Tyerman et al. 1999). However, somefundamental details remain uncharacterized. This maybe due, at least in part, to problems with the applicationof current quantitative models to data of water flowthrough roots.
The ordinary passive diffusion model
Quantitative data is often acquired from experiments de-scribing the transport of water through whole root sys-tems by analysing flow of xylem from root systems sub-jected to artificially raised pressure in chambers designedto accommodate roots still in potted soil (Fiscus 1977,Miller 1985, Passioura 1988, Markhart and Smit 1990,Maggio and Joly 1995, Jackson et al. 1996). The mostwidely accepted physical model applied to the flow data
Physiol. Plant. 114, 2002 1
ognized with the simple passive diffusion model, a new con-flict, flow saturation, was observed at high pressures. Experi-ments revealed that the plateau in flow, Jmax seen at highpressures followed natural rhythms diurnally and develop-mentally, and was not due to root damage or unnatural flowrestriction. Near the end of the photoperiod, Jmax closely cor-related with root dry mass. The above inconsistencies betweenobservations in pressure-flow kinetics and ordinary passivediffusion indicate that either the current model should be ad-justed or a new model should be proposed.
generated from these experiments is analogous tomodels of ordinary passive diffusion across a membrane(Nobel 1991).
JJ Ω Pj(Coj ª ci
j) [1]
Where Jj is flux of species j across the membrane of agiven area (quantity/area/time, i.e. m3/m2/s), Pj is thepermeability coefficient of species j (length/time, i.e. m/s) and (cjæ – cji) expresses the driving force across themembrane (referring here to the concentration of j out-side and inside).
The analogous root model is suitable for a system ofmembranes (Fiscus 1977, Passioura 1988, Jackson et al.1996):
J Ω Lp(DP ª s Dp) [2]
Where Lp is the collective membrane conductivity (l/time, m sª1) and (DP – s Dp) is the driving force sum-ming up components of water potential. DP and Dp are
Q1
Q2

UN
CO
RR
ECTE
DPR
OO
F
the differences in hydrostatic pressure and osmotic press-ure, respectively, from outside the root to inside the xy-lem. s is called the reflection coefficient, which measuresinteraction between water and solutes passing the mem-brane through common paths and gives an indication ofthe selectivity of the membrane for water, or leakinessfor other solutes such as ions. The appeal of this modelis its simplicity and its apparent ability to qualitativelyexplain the relationship between hydrostatic pressureand xylem flow. However, this model cannot resolve cer-tain aspects of experimental data including diurnal fluc-tuations of Lp and an unexplained offset on the pressureaxis of pressure-flow curves (Passioura 1988). Althoughthe intercept extrapolated from the linear portion of thecurve theoretically should indicate the osmotic pressureof the soil solution, it rarely does, and a term to describethe pressure intercept offset, Poff, has been introduced tothe equation (Passioura 1988):
J Ω Lp(DP ª Poff ª s Dp) [3]
Fiscus (1986) mathematically described diurnal changesin components of Equation 3, but the explanation wascomplex and the underlying physiological mechanism re-mained a black box. Murphy (2000) explained Poff in afour-component model incorporating asymmetric solutedistribution across the endodermis as a result of phloemdelivery of photoassimilates. However, testable predic-tions arising from the putative role of the phloem haveyet to be investigated. Despite these problems, use of themodel has perpetuated in many recent studies and re-views (Carvajal et al. 1996, 2000, Jackson et al. 1996,Fennel and Markhart 1998, Henzler et al. 1999,Quintero et al. 1999, Seel and Jeschke 1999, Tyermanet al. 1999, Wan and Zwiazek 1999).
Its apparent phenomenological fit but lack of physio-logical resolution for some of its parameters hasprompted a description of the simple passive diffusionmodel and its ability to explain flow data as ‘simple yetbaffling’ (Passioura and Munns 1984). Our study re-examines pressure flow relationships in isolated rootsmanipulated in a pressure chamber. Pressure flow re-lationships was investigated at different times of day anddevelopmental stages for three species, including twothat have been used extensively before (tomato andsoybean) and one that was yet unstudied (pea), by using(1) external pressure range which was increased to higherlevels than normally reported in previous studies and (2)techniques such as pressure cycling and reverse-pressuregradients.
Materials and methods
Plant material and growing conditions
All plants were grown in a glasshouse in Dijon underconditions that were designed to optimise homogeneityof both shoot and root growth as described below. Thesoil mixture that was developed specifically to the studyof root systems and avoids artefactual problems associ-
Physiol. Plant. 114, 20022
ated with pressure application to more sandy soils (Pas-sioura 1988): a bottom layer of expanded fireclay balls(6–10 mm diameter) facilitated effective pot drainage,while an unpacked mixture of high absorption Americanaluminium oxide (Al2SiO2, 0.5–4.0 mm, OIL DRIA.US-Special Typ IIIR, Lobbe, Germany) and clay par-ticles (2–5 mm) (50 : 50 v/v) allowed easy and homogene-ous diffusion of air and water and efficient water reten-tion. Roots were easily separated from this soil mixture.
An automatic watering system ensured plants wereregularly supplied with nutrient solution (P, K and oligo-elements; N was supplied to tomato and soybean; os-motic potential was ª0.065 MPa) adjusted to give aneutral pH after interaction with the aluminium oxide.The nutrient solution was delivered automatically ac-cording to the weights of the pots to maintain a volu-metric soil water content that was close to that achievedafter pots were freshly watered and allowed to drain(about 1.03 l per 4.90 l of soil mixture).
Seeds of tomato (Lycopersicon esculentum Mill. cv.Montfavet), pea (Pisum sativum cv. Baccara inoculatedwith Rhizobium leguminosarum) and soybean (Glycinemax L. Merryl cv. Essor, indeterminate group 00, non-inoculated) were sown in 5 l pots at a density of 8 seedsper pot. After seedling establishment the plants werethinned to the 4 most homogeneous per pot. Side shootswere removed regularly. Three different series of plantswere used; all of which were grown from May to Novem-ber under natural light. Developmental stages of theplants used for the studies were as follows: tomato plantswere between the 7 and 10 leaf stage, peas between theonset of flowering to the end of seed filling and soybeanswere nearing the end of seed filling. The roof of theglasshouse was normally left open, but it closed auto-matically during rain. Temperatures were maintained be-tween 5 and 24æC through the use of heating and thesliding feature of the roof.
Pressure chamber and collection of xylem sap
The experiments were not water-limited as pots were wellwatered with nutrient solution and allowed to drain nor-mally just before the plants were de-topped just belowthe cotyledonary node. For pea this required a small ex-cavation of soil. The pot complete with soil and rootsystem was placed into a pressure chamber similar tothat previously described by Else et al. (1994) with somemodifications. The lid and cylindrical body were madeof welded iron (240 mm i.d., 260 mm high; ComptoirMetallurgique Dijonnais, Dijon, France). The chamberhad a flat rubber joint (4 mm thick and 30 mm wide)which sealed the connection between the iron lid andbody. Twenty bolts fixed the lid firmly to the bodyallowing the application of pressure of up to 1.00 MParead from a 0–1.00 MPa manometer. All joints fixed intothe lid including the pressure inlet and four double-end-ed couplings that form quick joints with rigid tubingwere rated leak resistant up to 2.0 MPa (Prevost Ltd,Annecy-le-Vieux, France). After rinsing and blotting the

UN
CO
RR
ECTE
DPR
OO
F
cut surface thoroughly with distilled water, short sleevesof supple silicon tubing were fitted over the cut plantstumps. The internal diameter of the silicon tubingvaried according to the species and maturity of theplants and was selected to give a snug airtight seal. Theother end of the sleeve was fit around a longer length ofTygon tubing, which in turn was connected to a rigidtube inserted into the bottom end of chamber lid coup-ling. All tubing-tubing connections were made usingshort silicon sleeves like those connecting the hypocotylstump to the system. On the outside of the lid rigid tub-ing fit into the other end of the coupling was joined toa short length of supple tubing which directed exudateinto pre-weighed Eppendorf tubes. Thus the pressuresealing occurred between the tubing and coupler ratherthan against the hypocotyl stump. Four couplingsallowed simultaneous collection of exudate from up tofour plants in the same pot. Mounting and sealing ofthe plant in the pressure chamber took less than 10 minfrom when the stem was cut.
Chamber pressure was increased by using compressedair injected through an inlet in the lid connected to avalve. A manual valve allowed fine regulation for chang-ing or holding pressures constant over long periods oftime. The initial 20 min of exudate flow was discarded,since it was comprised of the time for mounting the rootsystem into the pressure chamber and the commence-ment of flow at 0.05 MPa and could not be reliablyquantified. Thereafter, at a given pressure, or series ofpressures, exudate was collected at regular intervals(usually of 10 min), and weighed to determine its volumeand, thus, xylem flow rate. Estimates of Lp and Pi weredetermined as the slope of the regression of Jv on ap-plied pressure (SAS Proc REG), including only pointssubjectively considered as comprising the linear phase ofthe curve.
After pressure manipulations were finished the rootswere carefully recovered by washing. They were subse-quently dried in an oven at 70æC for 48 h, and weighed.
Once a plant was subjected to pressure-flow manipula-tion it was not used in further experiments. In experi-ments involving pressure-flow relationships at differenthours throughout an entire day, separate homogeneouslygrown plants were used at each hour.
Transpiration
Transpiration was measured in parallel with the press-ure-flow manipulations from intact plants that were de-velopmentally homologous to those in the chamber. Therate of transpiration was measured gravimetrically fromat least 6 intact pots of plants. The pots were wateredand a weight was taken after drainage through the bot-tom of the pot. At intervals of one or more hours thepots were re-weighed to give an indication of water loss.From this amount water loss from control pots (whichcontained the same soil mixture but no plants) was sub-tracted and the remaining water loss was attributed totranspiration.
Physiol. Plant. 114, 2002 3
Results
Pressure-flow relationships
Using procedures similar to those of Jackson et al.(1996) an ascending pressure gradient was applied toroot systems of tomato in an effort to recreate flows thatmatched transpiration rates in intact plants. Fig. 1Ashows a curve typical of those obtained. Flows per entire
Fig. 1. Effect of an ascending pressure gradient on the rates of exu-date flow from roots of tomato (A), soybean (B) and pea (C). Theinitial 20 min of exudate flow were discarded and collections of 10min from 0.05 to 0.60 MPa were made thereafter. The single plantcurves were chosen as qualitatively representative of others observedfor the same species. For tomato, the methods for deriving termsof simple passive diffusion and facilitated passive diffusion modelsare displayed. For the simple passive diffusion a straight line was fitto the linear portion of the curve at medium pressures to obtain aslope (Lp) and pressure axis intercept (Pi). Maximum flow, Fmax (orJmax when expressed per g DW) was estimated from the flow plateauat high pressure and P0.5 was determined as the pressure at whichhalf Fmax (or Jmax) was observed. Transpiration rates of intact con-trol plants are indicated (arrow and value) for each species.

UN
CO
RR
ECTE
DPR
OO
F
root systems, F, were 30–50% lower than those reportedpreviously (Jackson et al. 1996), but this probably re-flects different cultivar and culture conditions. At lowpressures the curves were qualitatively similar since, asDP was increased in small steps, F from de-topped rootsincreased curvilinearly at first, and then linearly athigher pressures. Rates of transpiration were reachedusing less than 0.40 MPa and well within the linear rangeof flows (50–80% of the maximum flow per the entireroot system, Fmax) generated with the pressure gradient.However, at 0.45 MPa exudate flow began to saturate,and further increase in pressure distinguished a plateauin the pressure-flow curve giving it an overall sigmoidalshape, a feature not reported previously in studies thathave measured pressure induced xylem flow. Fig. 1A alsoindicates how the parameters of the curves were derivedaccording to interpretations previously proposed frommodels of simple passive diffusion. A straight line wasfitted by regression to the linear phase of the pressure-flow curve, which was extrapolated back to the pressureaxis to give an intercept value, Pi, that should theoretic-ally equal the osmotic potential of the soil psoil. Theslope of the fitted straight line, Lp, is considered ameasure of root water conductivity. Other parametersthat are not derived from simple passive diffusion arealso marked on the figure. The maximum flow per rootsystem, Fmax, was estimated from the highest flow ratesachieved. P0.5 was determined as the pressure that re-sulted in half the estimated Fmax. For comparisons in-volving transpiration per plant Fmax was used, but inother cases Jmax was calculated by dividing Fmax by theroot DW. Used this way, root DW was taken as an indi-cator of the layers of surface area water passes throughwhen entering the xylem from the soil. Comparisons influx could then be made among root systems of differentsizes, states of development and treatments. The meansto directly determine surface area were not available, butit was assumed that root DW varies linearly with surfacearea. This should be true since roots were relatively ho-mogeneous with respect to diameter within species andamong experiments.
Similar experiments were done with soybean, since itwas used previously to study pressure induced xylemflow (Fiscus 1977, Markhart and Smit 1990), to checkwhether saturation of flow was common across species(Fig. 1B). Like tomato, the curve was sigmoidal, andsaturation occurred at about 0.40 MPa. Unlike tomato,Fmax appeared to be barely adequate or slightly undertranspirational loss, since rates of transpiration of intactplants did not always occur within the linear portion ofthe curve, but instead usually happened as flow satu-rated.
Finally, pressure-flow curves were tested in pea (Fig.1C), since availability of a detailed model of its phenolo-gical progression would allow precise comparisonsamong experiments (e.g. Ney and Turc 1993, Rocheet al. 1998, Lhuiller-Soundele et al. 1999). Accordingly,it represents an excellent model species to study waterflow through roots according to development. The gen-
Physiol. Plant. 114, 20024
eral curve shape for pea was different from tomato andsoybean since flow from pea roots was linear from theinitial low pressure of 0.05 MPa onwards. Once againsaturation of flow was observed with increasing pressuregradients, but flow from pea saturated at lower pressuresthan either tomato or soybean. These differences re-sulted in a curve that was more hyperbolic than sig-moidal. With the exception of a few rare cases underunusually humid conditions, transpirational loss ofwater from intact peas in the glasshouse far exceededexudate flow from roots under pressure. For a set of 17experiments encompassing measurements across thephotoperiod and covering reproductive development(from flowering to the end of seed filling) Fmax ac-counted for only 3.9–40% of transpirational demands.The shortfall between isolated root supply and transpi-ration loss was considerable since measurements of po-tential water stocks in the shoots were very small (fromabout 1–3 ml during the reproductive cycle of peas,which would supply from 5 to 60 min of water if it alonesupplied transpiration) and made little impact on waterbudgets.
Results in Fig. 1 are curves obtained from single plantschosen because they were qualitatively representative oftheir species as inferred from over 65 pressure-flow ma-nipulations. It is important to note however, that par-ameters such as Lp, Pi, Fmax (or Jmax Ω Fmax/root DW)and P0.5 varied among different series of plants, amongdifferent aged plants within a series (developmentally)and among the same-aged plants diurnally (discussedbelow). Interestingly, although the water potential of thenutrient solution used to water the plants was constantamong experiments at ª0.065 MPa, Pi rarely occurredclose to this value.
Tests of root system integrity
Since flow saturation has not been reported before, itwas important to determine if it was a result of rootdamage caused by the application of high pressure. Ifthe integrity of the root were not compromised, onewould expect that cycling between saturating and subsa-turating pressures would result in predictable and con-stant flows characteristic of each pressure. On the otherhand, damaged roots might be expected to have morevariable flow, especially if pressure cycling caused furtherdegradation in root integrity.
In tomato, cycling of external pressure applicationfrom saturating (0.55 MPa) back to subsaturating (0.20MPa) pressures gave flows, at low pressures, slightlygreater than those obtained before saturation was ini-tially achieved (Fig. 2). This effect has been observedpreviously when pressure drops were imposed onsoybean roots, even when saturating pressures werenever imposed on the root system (Fiscus 1977). Fur-thermore, during the pressure cycling, the maximal flowsreached, Jmax, were the same each time a saturatingpressure was applied. Likewise, each drop to 0.20 MPaproduced a constant flow for that pressure.

UN
CO
RR
ECTE
DPR
OO
F
Fig. 2. Effect of cycling pressure between a saturating (0.55 MPa;?) and subsaturating (0.20 MPa; ?) pressure on exudate flow per gDW, J, of tomato roots following an ascending pressure gradient (?).The curve displays flow measurements in the chronological order inwhich they were made. Characteristics used to describe the ascend-ing pressure-flow curve are indicated including the saturation offlow (Jmax), the pressure at which half-maximal flow is observed(P0.5), the slope of the linear phase of the curve (Lp), and the press-ure-axis intercept derived from extrapolation from the linear por-tion of the curve (Pi).
Despite utmost care, occasionally root systems wereobviously injured, probably due to tissue rupture duringeither mounting in the pressure chamber or after increas-ing the application of external pressure. Two types ofinjuries were noted: the most common was likely a rup-ture of tissue that allowed easy passage of air throughthe system consisting of the root and connected tubing.This most likely occurred close to where the root stumpswas sealed by silicon tubing due to mechanical manipu-lation causing a cracking in the outer epidermal layersand cortex and creating a longitudinal fissure throughwhich air could travel and break the seal formed by thesilicon tubing. It was characterized by decreased xylemflow and a strongly increasing flow of air; such replicateswere obvious and were discarded. The other type of in-jury was also characterized by decreased flow, but waslikely due to a constriction by pinching. Problems of thisnature were also recognized by a deviation from theusual pressure-flow relationship (Fig. 3). Such deviationsconsisted of premature and unstable flow saturation andtheir occurrence could be confirmed by pressure cycling.Figure 3 shows flows from one healthy vs. three dam-aged, pinched roots from plants manipulated simul-taneously in the pressure chamber. If pressure wasdropped from saturating to subsaturating levels, the ef-fect was the opposite to that of undamaged roots (de-scribed above for Fig. 2) since flow would increase at thelower pressure, presumably due to an alleviation of thepinching effect. Any replicates that showed such devi-ations were not used further in pressure flow analyses.
Physiol. Plant. 114, 2002 5
Generally, pea had a more fragile root system, as indi-cated by a higher frequency of root injury due to asmaller and more irregularly shaped stem or root cross-section as compared to the thicker, rounder more robustones of tomato or soybean. Otherwise the integrity ofthe pressure-flow relationships could be impressivelystrong. One mature pea root system was observed towithstand a large amplitude in pressure cycling between0.1 MPa (16.4 ∫ 1.9 ml minª1) to 0.7 MPa (28.8 ∫ 1.9 mlminª1).
Exudate flow in descending pressure gradients
It has been observed previously that a diminishing flowrate may not follow the same kinetics as an incrementingflow rate (Fiscus 1977, Passioura and Munns 1984). Fur-thermore, the analysis of any differences between in-creasing- and decreasing-pressure gradients could givemore insight to the nature of the resistance towardswater entering roots. Descending pressure gradientswere imposed on tomato root systems to see if character-istics of the pressure-flow curves such as Jmax, P0.5, Lp
and Pi were constant, or a consequence of the directionin pressure gradient application.
Pressure flow relationships were investigated in two re-gimes involving declining-pressure gradients (Fig. 4A,B).In one case, an initial increasing pressure gradient (using0.05 MPa increments) was applied up to flow saturation(0.05–0.55 MPa) and was followed by a mirror-imagedeclining pressure gradient (Fig. 4A). The curve of theincreasing pressure gradient was typical of those ob-
Fig. 3. The response in exudate flow per g DW, J, of four individualtomato plants exposed simultaneously to an ascending pressuregradient, followed by cycling between saturating (0.45 MPa) andsubsaturating (0.20 MPa) pressures. The curves display flow meas-urements in the chronological order in which they were made. Onereplicate (?) shows flows typical for those not impeded by damagedor restriction caused by pressure manipulations. Three other repli-cates (?, k and ô) are typical of flows physically constricted around0.20–0.30 MPa of pressure. Note the opposite response (slowing offlow versus alleviation) of flow between the undamaged and flowrestricted replicates after pressure drops to 0.20 MPa.
UN
CO
RR
ECTE
DPR
OO
F
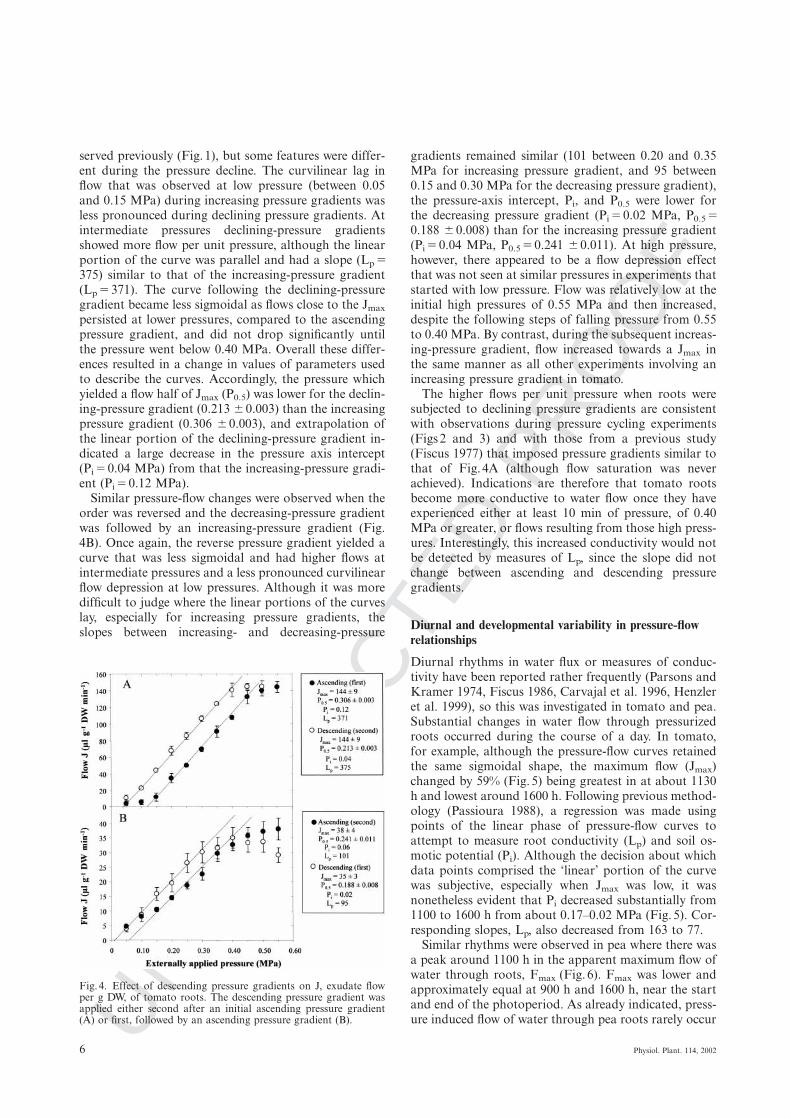
served previously (Fig. 1), but some features were differ-ent during the pressure decline. The curvilinear lag inflow that was observed at low pressure (between 0.05and 0.15 MPa) during increasing pressure gradients wasless pronounced during declining pressure gradients. Atintermediate pressures declining-pressure gradientsshowed more flow per unit pressure, although the linearportion of the curve was parallel and had a slope (Lp Ω375) similar to that of the increasing-pressure gradient(Lp Ω 371). The curve following the declining-pressuregradient became less sigmoidal as flows close to the Jmax
persisted at lower pressures, compared to the ascendingpressure gradient, and did not drop significantly untilthe pressure went below 0.40 MPa. Overall these differ-ences resulted in a change in values of parameters usedto describe the curves. Accordingly, the pressure whichyielded a flow half of Jmax (P0.5) was lower for the declin-ing-pressure gradient (0.213 ∫ 0.003) than the increasingpressure gradient (0.306 ∫ 0.003), and extrapolation ofthe linear portion of the declining-pressure gradient in-dicated a large decrease in the pressure axis intercept(Pi Ω 0.04 MPa) from that the increasing-pressure gradi-ent (Pi Ω 0.12 MPa).
Similar pressure-flow changes were observed when theorder was reversed and the decreasing-pressure gradientwas followed by an increasing-pressure gradient (Fig.4B). Once again, the reverse pressure gradient yielded acurve that was less sigmoidal and had higher flows atintermediate pressures and a less pronounced curvilinearflow depression at low pressures. Although it was moredifficult to judge where the linear portions of the curveslay, especially for increasing pressure gradients, theslopes between increasing- and decreasing-pressure
Fig. 4. Effect of descending pressure gradients on J, exudate flowper g DW, of tomato roots. The descending pressure gradient wasapplied either second after an initial ascending pressure gradient(A) or first, followed by an ascending pressure gradient (B).
Physiol. Plant. 114, 20026
gradients remained similar (101 between 0.20 and 0.35MPa for increasing pressure gradient, and 95 between0.15 and 0.30 MPa for the decreasing pressure gradient),the pressure-axis intercept, Pi, and P0.5 were lower forthe decreasing pressure gradient (Pi Ω 0.02 MPa, P0.5 Ω0.188 ∫ 0.008) than for the increasing pressure gradient(Pi Ω 0.04 MPa, P0.5 Ω 0.241 ∫ 0.011). At high pressure,however, there appeared to be a flow depression effectthat was not seen at similar pressures in experiments thatstarted with low pressure. Flow was relatively low at theinitial high pressures of 0.55 MPa and then increased,despite the following steps of falling pressure from 0.55to 0.40 MPa. By contrast, during the subsequent increas-ing-pressure gradient, flow increased towards a Jmax inthe same manner as all other experiments involving anincreasing pressure gradient in tomato.
The higher flows per unit pressure when roots weresubjected to declining pressure gradients are consistentwith observations during pressure cycling experiments(Figs 2 and 3) and with those from a previous study(Fiscus 1977) that imposed pressure gradients similar tothat of Fig. 4A (although flow saturation was neverachieved). Indications are therefore that tomato rootsbecome more conductive to water flow once they haveexperienced either at least 10 min of pressure, of 0.40MPa or greater, or flows resulting from those high press-ures. Interestingly, this increased conductivity would notbe detected by measures of Lp, since the slope did notchange between ascending and descending pressuregradients.
Diurnal and developmental variability in pressure-flowrelationships
Diurnal rhythms in water flux or measures of conduc-tivity have been reported rather frequently (Parsons andKramer 1974, Fiscus 1986, Carvajal et al. 1996, Henzleret al. 1999), so this was investigated in tomato and pea.Substantial changes in water flow through pressurizedroots occurred during the course of a day. In tomato,for example, although the pressure-flow curves retainedthe same sigmoidal shape, the maximum flow (Jmax)changed by 59% (Fig. 5) being greatest in at about 1130h and lowest around 1600 h. Following previous method-ology (Passioura 1988), a regression was made usingpoints of the linear phase of pressure-flow curves toattempt to measure root conductivity (Lp) and soil os-motic potential (Pi). Although the decision about whichdata points comprised the ‘linear’ portion of the curvewas subjective, especially when Jmax was low, it wasnonetheless evident that Pi decreased substantially from1100 to 1600 h from about 0.17–0.02 MPa (Fig. 5). Cor-responding slopes, Lp, also decreased from 163 to 77.
Similar rhythms were observed in pea where there wasa peak around 1100 h in the apparent maximum flow ofwater through roots, Fmax (Fig. 6). Fmax was lower andapproximately equal at 900 h and 1600 h, near the startand end of the photoperiod. As already indicated, press-ure induced flow of water through pea roots rarely occur

UN
CO
RR
ECTE
DPR
OO
F
Fig. 5. Changes in effect of ascending pressure gradients on exudateflow from tomato roots sampled from mid-day to late-afternoon.Three experiments were conducted, which started at 1130 h (g),1400 h (?) and 16 : 00 h (?). Comparisons are made on level of flowat saturation (Jmax) and the pressure at which half-maximal flow isobserved (P0.5). In addition a straight line was fit to the linear por-tion of the curves and extrapolated backward to determine thepressure axis intercept (Pi) and apparent conductivity as indicatedby the slope (Lp).
Fig. 6. Patterns in gravimetrically determined transpiration rates (?)of glasshouse grown peas compared to the pattern of Fmax (?), thelevel of maximum exudate flow from isolated root systems deter-mined from pressure-flow curves.
at rates great enough to account for transpirationallosses (Fig. 1). The two-component model of water entryinto roots predicts that water flow should increase dur-ing the periods of greatest transpiration (Steudle andPeterson 1998), so the timing of rhythms in Fmax andwater supply shortfalls were investigated according tochanges in transpiration. As indicated by Fmax, thegreatest potential water flow occurred in the mid-morn-ing, whereas transpiration of peas in the glasshouse wasusually low in the morning and gradually increased fromthe morning at 1100 h until peaking in mid-afternoon(Fig. 6). As indicated by Fmax compared to transpi-rational loss, there would be large shortfalls in watersupply and the most serious apparent deficits would oc-cur during the maximum rates of transpiration in themid-afternoon.
The rhythms in maximal water flow through pea rootsper root biomass, Jmax, were analysed in detail at 6 dif-ferent stages from early flowering to the end of seed fill-
Physiol. Plant. 114, 2002 7
ing (Fig. 7A). The amplitude of the diurnal rhythms pro-gressed in a logical sequence with development. Theysteadily increased from early flowering and reached amaximum from the middle to end of flowering. Fromthat point the amplitude of rhythm dropped consider-ably, coinciding with the middle to end of seed filling.Jmax declined from midday to late afternoon at eachstage of development, except for the end of grain fillingwhen Jmax was constant throughout the photoperiod. By1600 h Jmax converged to a similar rate among develop-mental stages. The constancy of Jmax measured at 1600h was analysed further by plotting maximal water flowper root system, Fmax, versus biomass using the series ofplants shown in Fig. 7A and an additional developmen-tal series of measurements that spanned the reproductivecycle of pea. The plot shows that Fmax in the late after-noon is strongly associated with root biomass (r2 Ω 0.96;Fig. 7B). This association held across different series of
Fig. 7. Patterns in apparent root conductivity determined frompressurized isolated roots of glasshouse grown peas: the diurnalpatterns in the maximum exudate flow per g DW of root (Jmax ΩFmax gª1 DW) from midday maximums to late afternoon minimumsfor 6 different stages of development from flowering at the firstreproductive node (F1) to the end of seed filling at the 6th repro-ductive node (ESF 6) SSF Ω start of seed filling (A). Relationshipbetween maximum exudate flow per root system, Fmax, samplednear the end of the day at 1600 h, to root DW (B).

UN
CO
RR
ECTE
DPR
OO
FQ3
experiments, and spanned almost a 3-fold change in rootbiomass.
Discussion
Problems with ordinary passive diffusion models
Ordinary passive diffusion models (Equation 2) kin-etically describe a J shaped curve (Newman 1976, Fiscus1977, Passioura 1988, Jackson et al. 1996). The initialslow increase of flow with pressure is perceived to be aresult of a total driving force during which the osmoticcomponent plays a significant role. Afterwards, a linearphase commences once osmotic potential effects becomenegligible at faster flows. Problems with ordinary passivediffusion models of water entry and flow through rootshave been known for quite some time and include vari-able intrinsic properties of conductivity, Lp, changing re-flection coefficients, s, and the inability to accuratelyresolve osmotic differentials between the soil and the xy-lem (Passioura 1988).
Our results reveal similar phenomenon and problemswith their interpretation. According to the model, theslope of the linear portion of the curve should be ameasure of membrane conductivity, Lp. Deciding whichpoints comprised the linear portion of the curve for re-gressions was subjective, especially for plants with lowerflows. In fact, current results indicated that the pressureflow curves in tomato and soybean are sigmoidal and,accordingly, have an inflection point about which theslope changes; therefore, the practice of deriving con-stants from intercepts and slopes of a regressed line isunstable. Even if one ignores this, systematic use of line-fitting showed that Lp was subject to large variationswith changing external pressure, both diurnally and de-velopmentally. These changes cannot be rationalizedeasily with simple passive diffusion.
The model predicts that the pressure axis interceptfrom an extrapolation of the linear portion of the press-ure-flow curve should correspond directly with the os-motic potential of the soil (psoil), but it rarely does (Pas-sioura 1988). The model has been amended with the in-clusion of an intercept-offset term, Poff, for which thereis no confirmed physiological explanation. The serious-ness of this was highlighted in one study of chilled rootsystems of spinach for which the pressure axis interceptwas as high as 0.20 MPa despite the fact that the rootsystems were in a nutrient solution with an osmotic po-tential of no less than ª0.05 MPa (Fennel and Markh-art 1998). Present data underline the same problem intomato and pea. In our experiments with tomato, theosmotic potential of the nutrient solution was constant,yet large differences in pressure axis intercepts were ob-served both among experiments (Figs 2,3,4 and 5) andwithin experiments (Figs 4A,B and 5). Moreover, inpeas, it was not uncommon to have a negative pressureaxis intercept despite the fact that the root systems weresurrounded by nutrient solution with an osmotic poten-tial of ª0.065 MPa, which therefore should have re-
Physiol. Plant. 114, 20028
sulted in a positive pressure axis intercept at 0.065 MPa.Present results also highlight a conflict previously not
revealed or discussed by earlier studies. Once the linearphase of the pressure-flow curve begins, Equation 2 pre-dicts that it should increase linearly in proportion to theapplied pressure ad infinitum. In practice this is not thecase. The three species tested each reached a saturatingmaximum flow (Fig. 1), that persisted even when all theindications were that root systems were undamaged bypressure application. As discussed below, the maximumflow appeared to be related to root biomass and integ-rally regulated according to developmental stage of theplant and time of day (Figs 5,6 and 7A,B). Can satu-ration be explained by adapting the simple passive dif-fusion model to incorporate some sort of inherent mem-brane ‘resistance’ that changes at the arrival of highwater potentials? This study does not completely dis-prove that water crosses the membrane by non-Stokesiandiffusion from the random thermal motion of the lipidtails. Using the Newman’s tangent test (Newman 1976),our data reveal a rather large probable accumulation ofsolutes at the principle barrier (limiting resistance) in theroot, which might be created from solvent drag. Thiscould tend to decrease the driving force at the principlebarrier and explain saturation. However, we did not ob-serve changes in saturation between plants that receivedhalf or full-strength nutrient solution prior to pressurechamber manipulations. Furthermore, plants flushed forseveral hours and supplied with distilled water through-out experiments neither prevented saturation nor alteredthe diurnal characteristics in saturated flow rates (Jmax)in tomato or pea roots. Finally, like current determi-nations of Lp and Pi, Newman’s tangent test relies onthe assignment of a straight line through the linear partof pressure-flow plots and is open to the same criticismoutlined above.
Is water conductivity by isolated root systemsrepresentative of intact plants?
The composite transport model of roots (Steudle andPeterson 1998) has been used to provide explanationsfor some of the problems associated with pressure flowcurves and the application of ordinary passive diffusionformalism. For example, the apoplastic entry of water isperceived to have a changing role in water absorptionbased on the demands of transpiration, which may ex-plain why Lp changes with pressure (Steudle et al. 1993).Accordingly, parallel apoplastic and transcellular path-ways would play an important role during the passageof water across the different root tissues. The apoplasticpaths are envisaged to lessen hydraulic resistance byallowing increased flow during periods of high transpi-ration, whereas water channels in the transcellular pathwould finely adjust water uptake in older, suberizedparts of roots lacking a substantial apoplastic compo-nent.
While this may be true, relationships observed for peabetween transpiration and pressure-flow data occurred

UN
CO
RR
ECTE
DPR
OO
F
inversely to the predictions of the two component hy-pothesis. In the glasshouse, peak transpiration for peaoccurred at about 5 h after the mid-day peak in maxi-mum water flow through pressurized roots, Fmax. In fact,peak transpiration occurred when Fmax was apparentlyat its lowest. Moreover, the work of Henzler et al. (1999)showed that transpiration rate can be manipulated inde-pendently of the diurnal rhythm of root conductivity,which would be inconsistent with the hypothesis thatroot conductivity was a limiting factor for transpiration.Considering these discrepancies between transpirationand conductivity, there is serious doubt whether waterflow through pressurized root systems is representativeof what occurs in an intact transpiring plant. There werelarge deficits between maximum potential supply ofwater from an individual root system (Fmax) and transpi-rational losses. The greatest deficits occurred betweenmid- to late-seed filling, when plants were transpiringtheir greatest amounts of water, but not visibly sufferingfrom drought.
Other studies that applied pressure to isolated rootsystems have not reported differences in between maxi-mum rates of flow from isolated roots and intact plants.Early on, Kramer (1959) warned that vacuum extractedsap flowed well below rates of transpiration, but surpris-ingly few studies have tried to compare the delivery ofwater from decapitated pressurized roots with measuredtranspiration rates. An exception is the work of Elseet al. (1995) that showed tomato roots pressurized above0.30 MPa could furnish an amount of water sufficientfor transpiration. Present results with tomato agreed,since a pressure between 0.35 and 0.40 MPa was satisfac-tory. Tomato in this respect could be an exceptional spe-cies. Our results with soybean indicate maximum flow is,at best, barely adequate and usually deficient for transpi-rational demands. Perhaps most researchers workingwith decapitated root systems have not concerned them-selves with accounting for transpiration because theyhave never observed or reported flow saturation at highpressures. Since ordinary passive diffusion models pre-dict an ever-increasing flow with pressure, one couldmerely extrapolate flow curves to calculate the kind ofpressure gradient required for transpirational require-ments. However, if pressure-flow saturation occurs be-low transpiration rates this possibility is precluded.
By contrast, in studies of intact plants of several differ-ent species (Hordeum vulgare, Lupinus albus, tomato andZea mays) discrepancies between rates of transpirationand pressure induced xylem flow rates have not been re-ported (Passioura and Munns 1984, Else et al. 1995,Stirzaker and Passioura 1996, Tiekstra et al. 2000).Those studies used a split-top pressure chamber design(Passioura 1980) that allowed pressurization of the rootsystem without de-topping the plant and xylem collec-tion from a cut leaf or petiole tip.
There is a strong possibility that at least one path ofwater flow is lost when shoots are excised. If true, thisappears to be the major route of water flow to the shootin the case of pea. It is difficult to speculate among the
Physiol. Plant. 114, 2002 9
several possible causes that might explain the loss of awater entry route brought about by the lack of a shootor loss of continuity in the water column (Passioura1988, Markhart and Smit 1990). Our pressure gradientswere generated by ‘pushing’ from outside the root byraising external pressure, but transpiration operates bytension or ‘pulling’ from the inside by lowering internalpressure through cohesion and flow. Although the DPwould be the same, physically this could have differenteffects on the conformation of membranes, water chan-nels or apoplasm (i.e. Shone and Clarkson 1988). How-ever, conductance of pressurized intact plants changeslinearly, even at pressures close to 2 MPa (Munns andPassioura 1984), so this explanation seems unlikely.
There may be a lost apoplastic route. Parallel apo-plastic and transcellular pathways are proposed to playdifferent roles during the passage of water across roottissues (Steudle et al. 1993). Increased apoplastic flowhas been implicated during periods of high flow requiredby midday transpiration. Perhaps what is left after re-moving the shoot is the water channel component in thetranscellular path that is normally implicated in onlyfine adjustments of water uptake in older, suberizedparts of roots lacking a substantial apoplastic compo-nent tissues (Steudle et al. 1993). Intact plants may notsuffer the same fate if they act like a manifold distri-bution system releasing the pressure drop more gradu-ally over a larger distance and area (i.e. the leaves ofmany nodes) rather than suddenly at the cut root stumpas in a shootless root system. The latter situation couldlead to an apoplastic plug if a sudden pressure dropcaused a form of intercellular compression.
Why has flow saturation not been reported earlier?
Although many examples of studies using isolated rootsystems have not reported flow saturation, they have notpresented pressures adequately high according to presentresults (i.e. greater than 0.40 MPa for tomato). Perhapsthere has been concern that the plateau was an artefactof damaged roots. However, the pressures used here wereless than 0.70 MPa, and far lower than those known toaffect cell physiology in plants (Passioura 1988). Obser-vations of the present study indicate that Jmax is acharacteristic of de-topped but otherwise well-func-tioning roots. For example, once flow saturation hadbeen reached and the pressure decreased, flow rates re-turned consistently to values equal to or greater thanthose measured at pre-saturation rates during the initialpressure increase. Subsequent return to saturating press-ures resulted in the same value of Jmax. This sort ofpressure cycling could be repeated several times with thesame root system without changing flow rates at specificsubsaturating or saturating pressures, and could be donewith pressure changes as large as 0.50 MPa.
During pressure cycling experiments, undamaged oruninhibited root systems were easily discerned fromdamaged or flow-inhibited root systems, for which flowappeared to be pinched off at higher pressures. For

UN
CO
RR
ECTE
DPR
OO
F
Q4
pinched roots, a drop in pressure actually increased flow,presumably by relieving the pinch, and subsequent re-turn to the higher pressure decreased flow once again.Pressure-inhibited and well-functioning roots were oc-casionally observed in the same experiment revealing adeparture of the former from the normally smooth sig-moidal kinetics (Fig. 3). Maximum flows were typicallyreached earlier than those of unobstructed roots and thetransition from linear flow to plateau was sudden ratherthan smooth and gradual. Moreover, the maximum flowdeclined with further pressure increase in pinched roots.
Another commonly observed type of root damagelikely resulted from lesions in the root during the fittingof the root system to the pressure chamber. This oc-curred most commonly with peas, which did not havethe woody strength of tomato or soybean, especially inthe region of tubing attachment. In those cases the flowof air increased and that of sap decreased as pressurewas increased. Root damage caused by lesions or pinch-ing might be expected to occur in a relatively randomfashion, whereas our observations of Jmax followed pre-dictable diurnal patterns, similar to those determinedfor root conductivity observed in several other plant sys-tems (as based on measurements of Lp or other anal-ogous parameters; Parsons and Kramer 1974, Passioura1988, Henzler et al. 1999).
Finally, careful scrutinization of previous studiessometimes reveals the tendency for saturating flow athigh pressures. For example, in studies of tomato, onecan see a point of saturation between 0.32 and 0.38 MPa(Maggio and Joly 1995) and a tendency for flow to leveloff at 0.40 MPa (Jackson et al. 1996). In the case ofMaggio and Joly (1995), despite the apparent flow satu-ration in some treatments, the curves were deemed‘highly linear’ and a straight line was fit to the data.
An hypothesis based on facilitated passive diffusion
Present results and problems with the current mathemat-ical modelling of water uptake in pressurized roots couldbe reconciled mechanistically with facilitated passive dif-fusion and the existence of water channels, aquaporins,for transmembrane water entry into roots. Considerablephysiological and molecular studies indicate flow islargely symplastic and that aquaporins may provide thegreatest resistance to water flow through root systems(Markhart and Smit 1990, Maggio and Joly 1995, Car-vajal et al. 1996, 2000, Henzler et al. 1999, Kjellbomet al. 1999, Quintero et al. 1999, Wan and Zwiazek1999).
Facilitated passive diffusion differs from ordinarypassive diffusion on criteria (Nobel 1991) that are strik-ingly similar to attributes of aquaporin-mediated waterflow. Facilitated diffusion is mediated by passive, selec-tive transporters that allow certain solutes to cross amembrane to the exclusion of other solutes and at fluxeslarger than those expected for ordinary passive diffusion(except at very high changes in water potential).
Likewise, aquaporins are transporters comprised of
Physiol. Plant. 114, 200210
proteins that form channels to allow the passage ofwater. The driving forces of these channels are hydraulicor osmotic in nature and are not active; while they per-mit or facilitate the movement of water through mem-branes they do not act as pumps (Chrispeels and Maurel1994). Aquaporins are highly selective and allow waterto pass freely while excluding ions or other small solutes.Accordingly, they are functionally defined as having ahigher level of permeability than would be expectedbased on a model of ordinary passive diffusion (Maurel1997, Tyerman et al. 1999). In fact, the usual tests forthe presence of functional aquaporins are the same asthose for the detection of facilitated passive diffusion.This includes membrane crossing at lowered activationenergy (Ea) and ratio of hydraulic permeability to dif-fusional permeability (Pf/Pd) greater than 1 (Kleinhans1998, Coury et al. 1999). Aquaporin Ea in structuralmodels indicates a low value of 16 kJ molª1 that is simi-lar to movement in bulk water and the corresponding Pf/Pd ratio is greater than unity in systems where diffusionfacilitated by aquaporins is known (Schäffner 1998).
Selectivity of facilitated passive diffusion is generallypoorly understood but the presence of transporters isassumed. Flux would saturate as all transporters becameoccupied at higher solute concentrations.
We are unaware of any studies reporting a saturationof aquaporin mediated water flow. However, in principle,saturation should occur once the finite number of aqua-porins in a given membrane are occupied at high outsidewater potential. It should be noted that binding is notneeded to explain how the process saturates. Any trans-port system that has a finite capacity will become satu-rated as that capacity is approached. Protein-mediatedprocesses saturate because the number of protein mol-ecules per cell is finite. Aquaporins somehow catalysewater movement through membranes. Regardless of themechanism, which is poorly understood, one can thinkof Jmax as the product of protein abundance and turn-over number (the number of ‘catalysed’ events per aqua-porin molecule per unit time). Since aquaporin abun-dance is finite, saturation is certainly possible, since thenumber of water molecules that can pass through theaquaporin channel per second is certainly finite. More-over, according to our observations, water entry intoisolated roots seems to saturate at physiologically rel-evant water flows.
Further evidence that aquaporins may be involved in-cludes our observations of short-term regulation (diurn-ally) and long-term regulation (with changes in growthphase). Such flow regulation has recently been inten-sively documented for water crossing membranes viaaquaporins (Maurel 1997, Johansson et al. 1998). Evi-dence already exists showing that water flow throughisolated roots is limited by the conductivity of aquapor-ins in the transcellular pathway in tomato (Maggio andJoly 1995) and a wide range of other plants includingmonocots, dicots, trees and legumes (Caravajal et al.1996, 2000, Henzler et al. 1999, Kjellbom et al. 1999,Quintero et al. 1999, Wan and Zwiazek 1999). Therefore,

UN
CO
RR
ECTE
DPR
OO
F
the water flow kinetics we observed in isolated roots oftomato, pea, and soybean may be more amenable to fa-cilitated passive diffusion and models based on enzymekinetics (Emery and Salon 2001).
Acknowledgements – We thank M. J. Jackson and R. S. Cole forsharing the detailed plans of their pressure chamber (University ofBristol, Lancaster, UK). Technical expertise was provided by G.Sanchez, F. Marmont and C. Jeudy in the design, building andmaintenance of the pressure chamber and by V. Durey for plantcultivation (INRA, Dijon, France). We are grateful to F. Marty,John Passioura and four anonymous reviewers for their criticism.
ReferencesCarvajal M, Cerda A, Martınez V (2000) Does calcium ameliorate
the negative effect of NaCl on melon root water transport byregulating aquaporin activity. New Phytol 145: 439–447
Carvajal M, Cooke DT, Clarkson DT (1996) Responses of wheatplants to nutrient deprivation may involve the regulation ofwater channel function. Planta 199: 372–381
Chrispeels MJ, Maurel C (1994) Aquaporins: the molecular basisof facilitated water movement through living plant cells? PlantPhysiol 105: 9–13
Coury LA, Hiller M, Mathal JC, Jones EW, Zeidel ML, BrodskyJL (1999) Water transport across yeast vacuolar and plasmamembrane-targeted secretory vesicles occurs by passive dif-fusion. J Bacteriol 181: 4437–4440
Else MA, Davies WJ, Whitford PN, Hall KC, Jackson MJ (1994)Concentrations of abscisic acid and other solutes in xylem sapfrom root systems of tomato and castor-oil plants are distortedby wounding and variable sap flow. J Exp Bot 45: 317–232
Else MA, Hall KC, Arnold GM, Davies WJ, Jackson MJ (1995)Export of abscisic acid, 1-aminocyclopropane-1-carboxylic acid,phosphate, and nitrate from roots to shoots of flooded tomatoplants. Plant Physiol 107: 377–384
Emery RJN, Salon C (2001) Apparent aquaporin mediated waterentry into pressurised detached root systems of pea, soybeanand tomato follows saturating facilitated diffusion kinetics. Pro-ceedings of the 4th European Conference in Grain Legumes.European Association for Grain Legume Research, Paris, pp308–309
Fennel A, Markhart AH III (1998) Rapid acclimation of root hy-draulic conductance to low temperature. J Exp Bot 49: 879–884
Fiscus EL (1977) Determination of hydraulic and osmotic prop-erties of soybean root systems. Plant Physiol 59: 1013–1020
Fiscus EL (1986) Diurnal changes in volume and solute transportcoefficients of Phaseolus roots. Plant Physiol 80: 752–759
Henzler T, Waterhouse RN, Smyth AJ, Carvajal M, Cooke DT,Schäffner AR, Steudle E, Clarkson DT (1999) Diurnal vari-ations in hydraulic conductivity and root pressure can be corre-lated with the expression of putative aquaporins in the roots ofLotus japonicus. Planta 210: 50–60
Jackson MB, Davies WJ, Else MA (1996) Pressure-flow relation-ships, xylem solutes and root hydraulic conductance in floodedtomato plants. Ann Bot 77: 17–24
Johansson I, Karlsson M, Shukla VK, Chrispeels MJ, Larsson C,Kjellbom P (1998) Water transport activity of the plasma mem-brane aquaporin PM28A is regulated by phosphorylation. PlantCell 10: 451–459
Kjellbom P, Larsson C, Johansson I, Karlsson M, Johanson U(1999) Aquaporins and water homeostasis in plants. TrendsPlant Sci 4: 308–314
Kleinhans FW (1998) Membrane permeability modelling: Kedem-
Edited by R. Munns
Physiol. Plant. 114, 2002 11
Katchalsky vs. a two-parameter formalism. Cryobiology 37:271–289
Kramer PJ (1959) Transpiration and the water economy of plants.In: Steward FC (ed) Plant Physiology: a Treatise, Vol. I: Plantsin Relation to Water and Solutes. Academic Press, New York,NY, pp 607–726
Lhuiller-Soundele A, Munier-Jolain NG, Ney B (1999) Dependenceof seed nitrogen concentration on plant nitrogen availabilityduring the seed filling pea. Eur J Agron 11: 157–166
Maggio A, Joly RJ (1995) Effects of mercuric chloride on the hy-draulic conductivity of tomato root systems. Plant Physiol 109:331–335
Markhart AH, IIISmit B (1990) Measurement of root hydraulicconductance. HortScience 25: 282–287
Maurel C (1997) Aquaporins and water permeability of plant mem-branes. Annu Rev Plant Physiol Plant Mol Biol 48: 399–429
Miller DM (1985) Studies of root function in Zea mays. IV. Effectsof applied pressure on the hydraulic conductivity and volumeflow through the excised root. Plant Physiol 77: 168–174
Munns R, Passioura JB (1984) Hydraulic resistance of plants. III.Effects of NaCl in barley and lupin. Aust J Plant Physiol 11:351–359
Murphy R (2000) Some compartmental models of the root: steady-state behaviour. J Theor Biol 207: 557–576
Newman EI (1976) Interaction between osmotic- and pressure-in-duced water flow in plant roots. Plant Physiol 57: 738–739
Ney B, Turc O (1993) Heat-unit-based description of the repro-ductive development of pea. Crop Sci 34: 141–146
Nobel PS (1991) Physicochemical and Environmental Plant Physi-ology. Academic Press, San Diego
Parsons LR, Kramer PJ (1974) Diurnal cycling in root resistanceto water movement. Physiol Plant 30: 19–23
Passioura JB (1980) The transport of water from soil to the shootin wheat seedlings. J Exp Bot 31: 333–345
Passioura JB (1988) Water transport in and to roots. Annu RevPlant Physiol Plant Mol Biol 39: 245–265
Passioura JB, Munns R (1984) Hydraulic resistance of plants. II.Effects of rooting medium, and time of day, in barley and lupin.Aust J Plant Physiol 11: 341–350
Quintero JM, Fournier JM, Benlloch M (1999) Water transport insunflower root systems: effects of ABA, Ca2π status and HgCl2.J Exp Bot 50: 1607–1612
Roche R, Jeuffroy M, Ney B (1998) A model to simulate the finalnumber of reproductive nodes in pea (Pisum sativum L.). AnnBot 81: 545–555
Schäffner AR (1998) Aquaporin function, structure, and expres-sion: are there more surprises to surface in water relations? Plan-ta 204: 131–139.
Seel WE, Jeschke WD (1999) Simultaneous collection of xylem sapfrom Rhinanthus minor and the hosts Hordeum and Trifolium:hydraulic properties, xylem sap composition and effects ofattachment. New Phytol 143: 281–298
Shone MGT, Clarkson DT (1988) Rectification of radial water flowin the hypodermis of nodal roots of Zea mays. Plant Soil 111:223–229
Steudle E, Murrmann M, Peterson CA (1993) Transport of waterand solutes across maize roots modified by puncturing the endo-dermis. Plant Physiol 103: 335–349
Steudle E, Peterson CA (1998) How does water get through roots?J Exp Bot 49: 775–788
Stirzaker RJ, Passioura JB (1996) The water relations of the root–soil interface. Plant Cell Environ 19: 201–208
Tiekstra AE, Else MA, Jackson MB (2000) External pressuresbased on leaf water potentials do not induce xylem sap to flowat rates of whole plant transpiration from roots of flooded orwell-drained tomato and maize plants. Impact of shoot hydrau-lic resistances. Ann Bot 86: 665–674
Tyerman SD, Bohnert HJ, Maurel C, Steudle E, Smith JAC (1999)Plant aquaporins: their molecular biology, biophysics and sig-nificance for plant water relations. J Exp Bot 50: 1055–1071
Wan X, Zwiazek JJ (1999) Mercuric chloride effects on root watertransport in aspen seedlings. Plant Physiol 121: 939–946

UN
CO
RR
ECTE
DPR
OO
F
Journal: Physiologia PlantarumArticle: PPL1O342
AUTHOR QUERY FORMDear Author,
Queries from the Production Editor are listed on the last page of the proof. The text to which the queries refer is indicated onthe proof by numbers (e.g. Q1) in the margin.Many thanks for your assistance.
Q1 Au: m3/m2/s. Should this be m s ª1?
Remarks:
Q2 Au: m/s. Should this be m s ª1?
Remarks:
Q3 Au: [2]. Please insert Equation 2 from the hard copy
Remarks:
Q4 Caravajal et al. 1996 has been changed to Carvajal et al. 1996 so that this citation matches the list
Remarks:
Physiol. Plant. 114, 200212




















