
VIGS VECTORS FOR GENE SILENCING: Many Targets, Many Tools
27
Annu. Rev. Plant Biol. 2004. 55:495–519 doi: 10.1146/annurev.arplant.55.031903.141803 Copyright c 2004 by Annual Reviews. All rights reserved First published online as a Review in Advance on February 25, 2004 VIGS VECTORS FOR GENE SILENCING: Many Targets, Many Tools Dominique Robertson Departments of Botany and Genetics, North Carolina State University, Raleigh, North Carolina 27695–7612; email: Niki [email protected] Key Words RNAi, hpRNA, functional genomics, geminiviruses ■ Abstract The discovery that plants recognize and degrade invading viral RNA caused a paradigm shift in our understanding of viral/host interactions. Combined with the discovery that plants cosuppress their own genes if they are transformed with ho- mologous transgenes, new models for both plant intercellular communication and viral defense have emerged. Plant biologists adapted homology-based defense mechanisms triggered by incoming viruses to target individual genes for silencing in a process called virus-induced gene silencing (VIGS). Both VIGS- and dsRNA-containing transforma- tion cassettes are increasingly being used for reverse genetics as part of an integrated approach to determining gene function. Virus-derived vectors silence gene expression without transformation and selection. However, because viruses also alter gene expres- sion in their host, the process of VIGS must be understood. This review examines how DNA and RNA viruses have been modified to silence plant gene expression. I discuss advantages and disadvantages of VIGS in determining gene function and guidelines for the safe use of viral vectors. CONTENTS INTRODUCTION ..................................................... 496 Viruses as Vectors ................................................... 497 Biology of VIGS .................................................... 498 When the Silencer is the Target ......................................... 501 Inoculation of Viral Vectors and Safety ................................... 501 Optimizing Silencing ................................................. 505 Pathways for Diffusible Silencing Signals ................................ 506 Gene Function Studies ................................................ 507 FUNCTIONAL GENOMICS ............................................ 508 Future Directions .................................................... 509 APPENDIX .......................................................... 510 1543-5008/04/0602-0495$14.00 495 Annu. Rev. Plant Biol. 2004.55:495-519. Downloaded from www.annualreviews.org by Universidad Autonoma de Queretaro on 01/15/14. For personal use only.
Transcript of VIGS VECTORS FOR GENE SILENCING: Many Targets, Many Tools

27 Apr 2004 15:11 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18) P1: GDL10.1146/annurev.arplant.55.031903.141803
Annu. Rev. Plant Biol. 2004. 55:495–519doi: 10.1146/annurev.arplant.55.031903.141803
Copyright c© 2004 by Annual Reviews. All rights reservedFirst published online as a Review in Advance on February 25, 2004
VIGS VECTORS FOR GENE SILENCING: ManyTargets, Many Tools
Dominique RobertsonDepartments of Botany and Genetics, North Carolina State University,Raleigh, North Carolina 27695–7612; email: Niki [email protected]
Key Words RNAi, hpRNA, functional genomics, geminiviruses
■ Abstract The discovery that plants recognize and degrade invading viral RNAcaused a paradigm shift in our understanding of viral/host interactions. Combined withthe discovery that plants cosuppress their own genes if they are transformed with ho-mologous transgenes, new models for both plant intercellular communication and viraldefense have emerged. Plant biologists adapted homology-based defense mechanismstriggered by incoming viruses to target individual genes for silencing in a process calledvirus-induced gene silencing (VIGS). Both VIGS- and dsRNA-containing transforma-tion cassettes are increasingly being used for reverse genetics as part of an integratedapproach to determining gene function. Virus-derived vectors silence gene expressionwithout transformation and selection. However, because viruses also alter gene expres-sion in their host, the process of VIGS must be understood. This review examines howDNA and RNA viruses have been modified to silence plant gene expression. I discussadvantages and disadvantages of VIGS in determining gene function and guidelinesfor the safe use of viral vectors.
CONTENTS
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 496Viruses as Vectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 497Biology of VIGS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 498When the Silencer is the Target . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 501Inoculation of Viral Vectors and Safety . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 501Optimizing Silencing . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 505Pathways for Diffusible Silencing Signals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 506Gene Function Studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 507
FUNCTIONAL GENOMICS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 508Future Directions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 509
APPENDIX . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 510
1543-5008/04/0602-0495$14.00 495
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
496 ROBERTSON
INTRODUCTION
The first indication that plants can be transformed with engineered genes broke theground for plant biotechnology and promised a new era of crop plant protectionand yield enhancement (8, 32). However, transgene expression is not entirelypredictable. RNA levels from the same gene transformed into different plants canvary and in some cases transgene expression is lost during development or insubsequent generations (4, 7). This was dramatically illustrated by the discoverythat not only transgene but also endogenous plant gene expression is subject tosilencing by ectopic expression of homologous transgenes (104, 145). Despiteactive transcription, mRNA accumulation for both genes decreased, a processcalled post-transcriptional gene silencing (PTGS).
The parallel discovery that transgene silencing can also impact RNA virusinfections if the transgene and virus share significant homology led to a model ofplant-mediated RNA degradation as a defense mechanism (24, 84, 119; reviewedin 83). Subsequent studies demonstrated that the transgene was not required totrigger this plant defense pathway. Nontransgenic plants that displayed recoveryphenotypes from wild-type virus infection also showed characteristics similar togene silencing in that viral RNA was eliminated in tissue where recovery wasevident (2, 20).
The nature of the recovery mechanism was not understood until PTGS1 wasassociated with a diffusible silencing signal. This was first demonstrated by graft-ing transgenic stocks of tobacco showing PTGS with the scions from a plant thatexpressed the same transgene (108). Efficient transmission of a silencing signaloccurred even when the stock and scion were separated by a 30-cm nontransgenicstem (double graft). In a second report, spread of transgene silencing was initi-ated by syringe-mediated agroinfiltration of a reporter gene expression cassetteinto a single leaf ofN. benthamianaactively expressing a chromosomal copy ofthe reporter gene (147). Expression of the reporter gene, encoding jellyfish greenfluorescent protein (GFP) could be visualized using UV illumination. Movementof an unidentified silencing factor into upper leaves and new growth could bechronicled in real time as UV-illuminated silenced tissues lost all green fluores-cence. Remarkably, even promoterless dsDNA with homology to GFP introducedby biolistics triggered systemic GFP silencing (149).
Antisense- and sense-mediated inhibition of gene expression was commonlyused to downregulate gene expression in plants and inC. elegans, but its efficiencyvaried in different transformants. A breakthrough that started the use of RNAias a general silencing tool occurred when it was found that only small amounts
1PTGS is similar to RNAi, which is here referred to as interfering RNA from dsRNA.siRNA refers to 22-nt oligonucleotide dimers used to initiate silencing. hpRNA refers toconstructs for transcribing inverted repeats separated by nonhomologous or intron sequences(sometimes referred to as panhandles). smRNA refers to small species of RNA (less than100 nt) associated with PTGS.
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 497
(a few molecules) of dsRNA injected intoC. eleganswere needed for widespreadsilencing similar to PTGS in plants (19a, 29, 70, 100). The low efficiency of bothsense- and antisense-mediated silencing in plants was then explained by proposingthat dsRNA was produced from complex integration patterns in different transfor-mants. Subsequently, transgene constructs containing an expression cassette ofinverted repeats (IR-PTGS or hpRNA) were found to be much more efficient thaneither antisense- or sense-mediated silencing alone (157). The similarities betweenPTGS-related mechanisms in a variety of different organisms have been reviewed(16, 19).
Another breakthrough occurred when smRNAs (21–28 nt) homologous togenes silenced by PTGS were found in silenced tissue (43, 44). In mammals andDrosophila, 22-nt siRNA molecules are sufficient to induce silencing spread (12,46). There are reports of 22-nt siRNA-mediated silencing in plants (74, 146), buttransitive RNA, dsRNA that flanks the region of homology, was also found. Transi-tive RNA fractionates with smRNA but lacks homology to the inducing RNA, andis likely produced by the host enzyme, RNA-dependent RNA polymerase (RdRP).In C. elegans, which also shows transitive RNA, siRNAs can initiate silencing butsubstantially less dsRNA is needed if longer RNAs are used (A. Fire, personalcommunication). In other organisms that encode RdRP, such as plants and fungi,dsRNA larger than 22 nt can initiate silencing, but the specific nature of the mobileRNA species in plants and fungi remains to be determined (98, 116).
This review covers the silencing of endogenous plant genes initiated from re-combinant viral vectors, coined virus-induced gene silencing (VIGS) (126). RNAand DNA virus vectors are described and compared with respect to host range,host interactions, and method of use. Examples from the literature are highlightedto show relative effectiveness of silencing in different classes of genes, and the ad-vantages and disadvantages of using viral vectors compared to methods requiringtransformation. An excellent review covering genomic initiatives for using bothVIGS (primarily RNA virus-based VIGS) and hpRNA chromosomal cassettes forsilencing should also be consulted to provide a comprehensive picture of RNAi inplants (158).
Viruses as Vectors
Many different RNA and DNA viruses have been modified to serve as vectors forgene expression (reviewed in 118, 138). Some viruses, such as tobacco mosaicvirus (TMV), potato virus X (PVX), and tobacco rattle virus (TRV), can be usedfor both protein expression and gene silencing (6, 77, 92, 93). Not all RNA virus-derived expression vectors will be useful as silencing vectors because many, suchas TEV, have potent anti-silencing proteins that directly interfere with host silenc-ing machinery (5, 68). DNA viruses have not been used extensively as expressionvectors due to their size constraints for movement (109). However, a nonmobilemaize streak virus-derived vector (MSV, in the Mastrevirus genus ofGeminiviri-dae) has been successfully used for long-term production of protein in maize cellcultures (110).
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
498 ROBERTSON
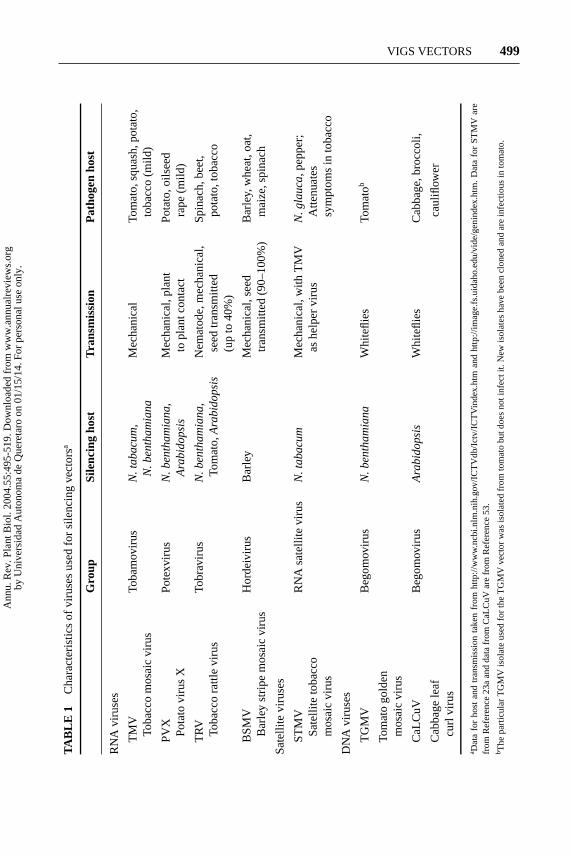
Using viral vectors to silence endogenous plant genes requires cloning homol-ogous gene fragments into the virus without compromising viral replication andmovement. This was first demonstrated in RNA viruses by inserting sequences intoTMV (77), and then for DNA viruses by replacing the coat protein gene with ahomologous sequence (72). These reports used visible markers for gene silencingPDSandChlI, providing a measure of the tissue specificity of silencing.
Table 1 shows some general characteristics for currently available virus-derivedgene silencing vectors. Most viruses are plus-strand RNA viruses or satellites,whereas tomato golden mosaic virus (TGMV) and cabbage leaf curl virus (CaL-CuV) are DNA viruses. Although RNA viruses replicate in the cytoplasm, DNAviruses replicate in plant nuclei using host DNA replication machinery. Both typesof viruses induce diffusible, homology-dependent systemic silencing of endoge-nous genes. However, the extent of silencing spread and the severity of viral symp-toms can vary significantly in different host plants and host/virus combinations.With the variety of viruses and the diversity of infection patterns, transmissionvectors, and plant defenses it is not surprising that viruses differ with respect tosilencing (137). Because the continuing development of virus-based silencing vec-tors can extend VIGS to economically important plants, it is useful to considersome of the characteristics of successful VIGS vectors.
Biology of VIGS
RNA viruses replicate cytoplasmically using their own polymerase and host cy-toplasmic membranes, ribosomes, and proteins (22, 60, 106). Detailed protocolsfor making and using virus-derived silencing vectors have been published andshould be consulted for more information (23, 89). The RNA viruses shown inTable 1 can be mechanically inoculated using in vitro transcribed RNA copies ofthe plus-strand genome. An easier method uses agroinfection of cloned vectorsfor TRV and PVX (86, 120). Following agroinfiltration of an expression cassettecontaining a cDNA copy of the genome, a single-stranded mRNA resembling thevirus is transcribed by the host RNA polymerase and exported to the cytoplasm.The first sequence on the mRNA is then translated to produce the viral replicase(an RdRP). Replication of similar mRNA molecules containing a 3′-virus-specificrecognition sequence is initiated and the viral vector moves into new cells. RNAviruses with multiple genome components, such as TRV, can be cloned and coinoc-ulated with high efficiency (120). Most RNA virus–derived vectors are insertionvectors (118) and contain a duplicated subgenomic promoter preceding the inser-tion site. There are two versions of the TRV vector, one with an insertion site andone with a duplicate subgenomic promoter and insertion site (86, 89). Becauseeach RNA virus-encoded protein is required for efficient movement and repli-cation, gene replacement vectors have not been successfully used for silencing(118).
Satellite viruses are small RNAs (800–1300 nt) that usually encode their owncoat protein but rely on a second virus for replication (22). A silencing system
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.
23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 499TA
BLE
1C
hara
cter
istic
sof
viru
ses
used
for
sile
ncin
gve
ctor
sa
Gro
upS
ilenc
ing
host
Tran
smis
sion
Pat
hoge
nho
st
RN
Avi
ruse
s
TM
VTo
bam
oviru
sN
.ta
ba
cum
,M
echa
nica
lTo
mat
o,sq
uash
,pot
ato,
Toba
cco
mos
aic
viru
sN
.be
nth
am
ian
ato
bacc
o(m
ild)
PV
XP
otex
viru
sN
.be
nth
am
ian
a,M
echa
nica
l,pl
ant
Pot
ato,
oils
eed
Pot
ato
viru
sX
Ara
bid
op
sis
topl
antc
onta
ctra
pe(m
ild)
TR
VTo
brav
irus
N.b
en
tha
mia
na,
Nem
atod
e,m
echa
nica
l,S
pina
ch,b
eet,
Toba
cco
rattl
evi
rus
Tom
ato,A
rab
ido
psi
sse
edtr
ansm
itted
pota
to,t
obac
co(u
pto
40%
)
BS
MV
Hor
deiv
irus
Bar
ley
Mec
hani
cal,
seed
Bar
ley,
whe
at,o
at,
Bar
ley
strip
em
osai
cvi
rus
tran
smitt
ed(9
0–10
0%)
mai
ze,s
pina
ch
Sat
ellit
evi
ruse
s
ST
MV
RN
Asa
telli
tevi
rus
N.t
ab
acu
mM
echa
nica
l,w
ithT
MV
N.g
lau
ca,p
eppe
r;S
atel
lite
toba
cco
ashe
lper
viru
sA
ttenu
ates
mos
aic
viru
ssy
mpt
oms
into
bacc
o
DN
Avi
ruse
s
TG
MV
Beg
omov
irus
N.b
en
tha
mia
na
Whi
tefli
esTo
mat
ob
Tom
ato
gold
enm
osai
cvi
rus
CaL
CuV
Beg
omov
irus
Ara
bid
op
sis
Whi
tefli
esC
abba
ge,b
rocc
oli,
Cab
bage
leaf
caul
iflow
ercu
rlvi
rus
a Dat
afo
rho
stan
dtr
ansm
issi
onta
ken
from
http
://w
ww
.ncb
i.nlm
.nih
.gov
/ICT
Vdb
/Ictv
/ICT
Vin
dex.
htm
and
http
://im
age.
fs.u
idah
o.ed
u/vi
de/g
enin
dex.
htm
.D
ata
for
ST
MV
are
from
Ref
eren
ce23
aan
dda
tafr
omC
aLC
uVar
efr
omR
efer
ence
53.
b The
part
icul
arT
GM
Vis
olat
eus
edfo
rth
eT
GM
Vve
ctor
was
isol
ated
from
tom
ato
butd
oes
noti
nfec
tit.
New
isol
ates
have
been
clon
edan
dar
ein
fect
ious
into
mat
o.
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
500 ROBERTSON
[a satellite virus–induced silencing system (SVISS)] was developed that uses satel-lite TMV (STMV) as a vector for spreading silencing fragments and TMV as ahelper virus in tobacco (38). In this system, 100–300-nt fragments are inserted intothe satellite genome and the resulting vector is coinoculated with a helper virus.PVX, TMV, and the TMV satellite virus do not have strong anti-silencing proteins,whereas the cucumber mosaic virus, tobacco etch virus, and cymbidium ringspotvirus have proteins that can reverse transgene silencing (10, 40, 49, 91; reviewedin 144).
Two geminiviruses, TGMV and CaLCuV, have been used to generate silenc-ing vectors. As with other members of the Begomovirus genus ofGeminiviridae,the TGMV and CaLCuV genomes consist of two single-stranded circular DNAmolecules called the A and B components, each is about 2.5 kb in size (45, 53).DNA viruses replicate through dsDNA intermediates in plant nuclei. They dependon host transcription and translation to produce proteins that interact with cellcycle regulators to induce host DNA replication machinery (25, 76). Infectiousvectors have been cloned that contain direct repeats of the viral-replication origin–containing common region flanking the viral genome (124). Replicating unit lengthvectors depends on the AC1 gene and results in infectious DNA genomes (26).DNA replication occurs using host enzymes and double-stranded viral DNA in-termediates (42, 47, 47a).
Geminivirus-derived vectors move systemically without the 800-bp coat pro-tein gene and can carry foreign sequence instead (14, 50, 133), but only in certainhost/virus combinations. Although the A component can replicate in the absenceof the B component (124), movement proteins encoded in the B component arerequired for systemic infection (81). The TGMV genome is stably propagated withinserts ranging from 100–800 bp (112), but African cassava mosaic virus (ACMV)vectors acquire additional sequence from other parts of its genome by recombiningto restore its original size (75). Results for both TGMV and ACMV vectors usedN. benthamiana, suggesting that the viruses themselves exhibited different proper-ties with respect to movement. The molecular size of geminivirus-derived vectorsis easily determined by DNA gel blot hybridizations of restriction enzyme digestedviral DNA because the virus replicates to high numbers in plant nuclei (138). Thus,DNA gel blots can be used to determine the stability of the inserted DNA silencingfragment.
Because geminiviruses induce host DNA replication and alter plant gene ex-pression (9, 103; reviewed in 47), their vectors may be considered unsuitable fortesting gene function. However, two results argue against this conclusion: Bothviral DNA replication and induction of PCNA are cell autonomous (103), and thereplication of geminivirus-derived silencing vectors is greatly reduced comparedto replication of the wild-type virus (73, 112). Both in situ hybridization of viralDNA in silenced tissues and DNA gel blots show that viral DNA levels are re-duced in silenced tissue. SilencingChlI from a mutant, phloem-limited TGMVfurther demonstrates that extensive silencing can occur in uninfected cells (112).Vectors carrying similarly sized unrelated DNA can be used to control the impact
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 501
of virus-associated changes in gene expression. Similar controls have been used toseparate target gene silencing phenotypes and virus effects in RNA vectors (159).
When the Silencer is the Target
RNA viruses are targets for silencing because their genome is directly affectedby host-mediated dsRNA degradation. Inoculation of movement-competent PVX-GFP into GFP-expressing plants results in eliminating the inducing vector (126).However, the same vector carrying sequence homologous to the endogenousgeneRbcS(small subunit of ribulose-bisphosphate carboxylase/oxygenase) wasnot eliminated from PVX:RbcS-silenced plants. DNA methylation is associatedwith PVX-GFP-silenced GFP transgenes, but the PVX:RbcS-silenced endogenousgenes lacked extensive methylation (64). RNA-directed DNA methylation waspreviously reported (114, 155) and may be a general epigenetic mechanism (65,154, 156). It is not understood why transgenes are methylated by VIGS whereasendogenous genes are not.
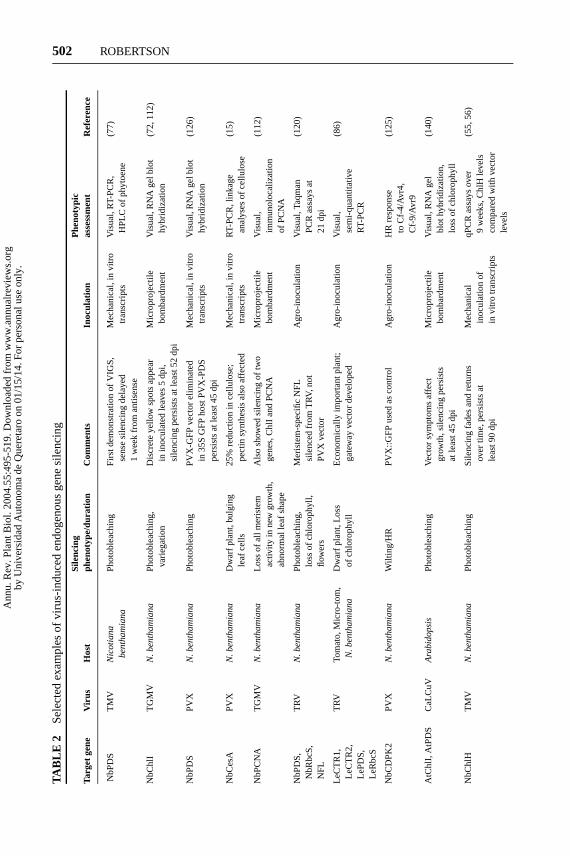
Levels of recombinant TMV vectors with homology to an endogenous genefluctuate, andChlH silencing also fades and reappears (55). Table 2 shows thatsome RNA vectors produce strong silencing but then it decreases. Although elim-inating a recombinant PVX silencing vector did not occur when an endogenousgene was targeted, it was not clear how long silencing was maintained in newgrowth (126). In contrast to RNA viruses, there is no evidence for eliminationof recombinant geminivirus genomes, even when they contain sequences withhomology to a transgene. For example, systemic spread of GFP silencing is notfollowed by recovery from the virus when a CaLCuV vector is used to infectArabidopsiscontaining a 35S-GFP transgene (140). In contrast to ssDNA virusesstudied so far, the wild-type retrovirus-like CaMV can be eliminated by silencing-related recovery (2), perhaps because the RNA form of its genome is a target(112).
Wild-type geminivirus infections can show attenuation of symptoms similar toRNA viruses, but viral DNA accumulation is reduced, not eliminated (59, 123).Replicating DNA viruses do not tolerate methylation (28) and it is enigmatic thatDNA viruses can elicit silencing of endogenous plant genes while maintainingenough expression of their own genes to sustain infection. The requirement ofRdRP for DNA VIGS, but not RNA VIGS, inArabidopsis(160) provides furtherevidence that silencing does not eliminate DNA viruses. Note that even thoughwild-type tomato yellow leaf curl geminivirus infections are associated with siR-NAs, the symptoms of this pathogen are severe (90).
Inoculation of Viral Vectors and Safety
Recombinant viruses require special treatment and approval from plant healthregulatory authorities must be obtained before the viruses are received and used.Because RNA viruses, especially barley stripe mosaic virus (BSMV), are usedin their entirety for silencing vectors, unintentional inoculation by mechanical
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.
23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
502 ROBERTSONTA
BLE
2S
elec
ted
exam
ples
ofvi
rus-
indu
ced
endo
geno
usge
nesi
lenc
ing
Sile
ncin
gP
heno
typi
cTa
rget
gene
Viru
sH
ost
phen
otyp
e/du
ratio
nC
omm
ents
Inoc
ulat
ion
asse
ssm
ent
Ref
eren
ce
NbP
DS
TM
VN
ico
tian
aP
hoto
blea
chin
gF
irstd
emon
stra
tion
ofV
IGS
,M
echa
nica
l,in
vitr
oV
isua
l,R
T-P
CR
,(7
7)b
en
tha
mia
na
sens
esi
lenc
ing
dela
yed
tran
scrip
tsH
PLC
ofph
ytoe
ne1
wee
kfr
oman
tisen
se
NbC
hlI
TG
MV
N.b
en
tha
mia
na
Pho
tobl
each
ing,
Dis
cret
eye
llow
spot
sap
pear
Mic
ropr
ojec
tile
Vis
ual,
RN
Age
lblo
t(7
2,11
2)va
riega
tion
inin
ocul
ated
leav
es5
dpi,
bom
bard
men
thy
brid
izat
ion
sile
ncin
gpe
rsis
tsat
leas
t52
dpi
NbP
DS
PV
XN
.be
nth
am
ian
aP
hoto
blea
chin
gP
VX
-GF
Pve
ctor
elim
inat
edM
echa
nica
l,in
vitr
oV
isua
l,R
NA
gelb
lot
(126
)in
35S
GF
Pho
stP
VX
-PD
Str
ansc
ripts
hybr
idiz
atio
npe
rsis
tsat
leas
t45
dpi
NbC
esA
PV
XN
.be
nth
am
ian
aD
war
fpla
nt,b
ulgi
ng25
%re
duct
ion
ince
llulo
se;
Mec
hani
cal,
invi
tro
RT-
PC
R,l
inka
ge(1
5)le
afce
llspe
ctin
synt
hesi
sal
soaf
fect
edtr
ansc
ripts
anal
yses
ofce
llulo
se
NbP
CN
AT
GM
VN
.be
nth
am
ian
aLo
ssof
allm
eris
tem
Als
osh
owed
sile
ncin
gof
two
Mic
ropr
ojec
tile
Vis
ual,
(112
)ac
tivity
inne
wgr
owth
,ge
nes,
Chl
Iand
PC
NA
bom
bard
men
tim
mun
oloc
aliz
atio
nab
norm
alle
afsh
ape
ofP
CN
A
NbP
DS
,T
RV
N.b
en
tha
mia
na
Pho
tobl
each
ing,
Mer
iste
m-s
peci
ficN
FL
Agr
o-in
ocul
atio
nV
isua
l,Ta
qman
(120
)N
bRbc
S,
loss
ofch
loro
phyl
l,si
lenc
edfr
omT
RV,
not
PC
Ras
says
atN
FL
flow
ers
PV
Xve
ctor
21dp
i
LeC
TR
1,T
RV
Tom
ato,
Mic
ro-t
om,
Dw
arfp
lant
,Los
sE
cono
mic
ally
impo
rtan
tpla
nt;
Agr
o-in
ocul
atio
nV
isua
l,(8
6)Le
CT
R2,
N.b
en
tha
mia
na
ofch
loro
phyl
lga
tew
ayve
ctor
deve
lope
dse
mi-q
uant
itativ
eLe
PD
S,
RT-
PC
RLe
Rbc
S
NbC
DP
K2
PV
XN
.be
nth
am
ian
aW
iltin
g/H
RP
VX
::GF
Pus
edas
cont
rol
Agr
o-in
ocul
atio
nH
Rre
spon
se(1
25)
toC
f-4/
Avr
4,C
f-9/
Avr
9
AtC
hlI,
AtP
DS
CaL
CuV
Ara
bid
op
sis
Pho
tobl
each
ing
Vect
orsy
mpt
oms
affe
ctM
icro
proj
ectil
eV
isua
l,R
NA
gel
(140
)gr
owth
,sile
ncin
gpe
rsis
tsbo
mba
rdm
ent
blot
hybr
idiz
atio
n,at
leas
t45
dpi
loss
ofch
loro
phyl
l
NbC
hlH
TM
VN
.be
nth
am
ian
aP
hoto
blea
chin
gS
ilenc
ing
fade
san
dre
turn
sM
echa
nica
lqP
CR
assa
ysov
er(5
5,56
)ov
ertim
e,pe
rsis
tsat
inoc
ulat
ion
of9
wee
ks,C
hlH
leve
lsle
ast9
0dp
iin
vitr
otr
ansc
ripts
com
pare
dw
ithve
ctor
leve
ls
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 503Ta
rget
gene
Viru
sH
ost
Sile
ncin
gph
enot
ype
Com
men
tsIn
ocul
atio
nP
heno
typi
cas
sess
men
tR
efer
ence
HvP
DS
BS
MV
Bar
ley
Pho
tobl
each
ing
Qua
ntita
tive
PC
Ras
says
show
Mec
hani
cal,
thre
eH
PLC
used
to(5
8)si
lenc
ing
incr
ease
sth
enin
vitr
otr
ansc
ribed
dete
ctph
ytoe
nede
crea
ses
betw
een
5–25
dpi
RN
As
AtT
auA
TR
V35
SA
tTau
A-G
FP
Loss
of35
S-T
uA-G
FP
Sho
wed
that
TM
V-d
sRed
Mec
hani
cali
nocu
latio
nC
onfo
calm
icro
scop
y,(3
6)tr
ansg
enic
expr
essi
on,
still
mov
esin
35S
-AtT
au-G
FP
ofin
vitr
otr
ansc
ripts
tran
sgen
eex
pres
sion
N.b
en
tha
mia
na
sile
nced
tissu
e,bu
tsile
ncin
gin
guar
dce
llsre
mai
nsof
endo
geno
usN
bTau
Aho
mol
ogis
notd
emon
stra
ted
NbE
DS
TR
V35
S-N
tNge
neH
yper
sens
itive
Com
plic
ated
syst
embu
tA
gro-
inoc
ulat
ion
ofV
isua
l,R
NA
gelb
lot
(111
)tr
ansf
orm
edre
spon
seto
TM
V,cl
ear
phen
otyp
eT
RV
::ED
Sth
enhy
brid
izat
ion
N.b
en
tha
mia
na
syst
emic
necr
osis
TM
V::G
FP
;or
TR
V::E
DS
then
TR
V::P
toor
TR
V::R
xth
enT
MV
::GF
P
PS
IIox
ygen
TR
VN
.be
nth
an
ian
aLi
ghtg
reen
leav
es10
Xen
hanc
edT
MV
repl
icat
ion
Agr
o-in
ocul
atio
nV
isua
l,R
NA
gelb
lot
(1)
evol
ving
com
plex
,re
port
ed,v
erifi
edin
PS
IIhy
brid
izat
ion
33K
subu
nit
herb
icid
etr
eate
dpl
ants
NtP
DS
,NtC
hsA
,S
TM
VN
.ta
ba
cum
Vario
us,s
trik
ing
100–
300
ntsi
lenc
ing
Co-
inoc
ulat
ion
ofVa
rious
phen
otyp
es(3
8)N
tRbc
S,N
tCes
A,
vect
orvi
sual
phen
otyp
esfr
agm
ents
optim
al,s
ilenc
ing
invi
tro
tran
scrib
eddo
cum
ente
dby
NtT
K,N
tALS
,w
ithT
MV
pers
ists
atle
asts
ixm
onth
sR
NA
and
TM
Vle
afim
ages
NtP
PX
,NtG
ln,
help
ervi
rus
extr
act
NtR
pII,
NtA
ct,
NtN
PK
1N
tPA
RP
NtP
DS
,NtE
DS
1,T
RV
35S
-Nge
neLo
ssof
hype
rsen
sitiv
eC
ompl
icat
edsy
stem
but
Agr
o-in
ocul
atio
nV
isua
l,(8
7)N
tRar
1,tr
ansf
orm
edre
spon
seto
TM
V::G
FP
clea
rph
enot
ype
ofT
RV
::ED
S,T
RV
::Rar
1,se
mi-q
uant
itativ
eN
tNpr
1/N
im1
N.b
en
tha
mia
na
TR
V::P
DS
sile
ncin
gin
uppe
rT
RV
::NP
R1/
NIM
1th
enR
T-P
CR
leav
esse
enat
5dp
i,pe
rsis
tant
TM
V::G
FP
NbS
KP
1,N
bSG
T1,
TR
V35
S-N
gene
Pre
senc
eof
TM
V-G
FP
TR
Vus
edfo
rsi
lenc
ing,
Agr
o-in
ocul
atio
nS
emi-q
uant
itativ
eP
CR
(88)
NbC
SN
3,N
bCS
N8
N.b
en
tha
mia
na
infe
ctio
nT
MV
and
PV
Xve
ctor
sus
edfo
rex
pres
sion
ofF
LAG
-or
HA
-epi
tope
tagg
edge
nes
toco
nfirm
inte
ract
ion
(Co
ntin
ue
d)
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
504 ROBERTSON
TAB
LE2
(Co
ntin
ue
d)
Targ
etge
neV
irus
Hos
tS
ilenc
ing
phen
otyp
eC
omm
ents
Inoc
ulat
ion
Phe
noty
pic
asse
ssm
ent
Ref
eren
ce
NbF
tsH
PV
XN
.be
nth
am
ian
aP
hoto
blea
chin
gP
heno
copi
esAra
bid
op
sisv
ar1
Mec
hani
cali
nocu
latio
nR
T-P
CR
,im
mun
oblo
t(1
29)
mut
atio
n(F
tsH
hom
olog
)of
invi
tro
tran
scrip
ts
NbP
AF,
NbR
PN
9T
RV
N.b
en
tha
mia
na
Ass
ayed
20dp
iE
xcel
lent
cont
rols
,N,C
,and
Agr
o-in
ocul
atio
nS
emi-q
uant
itativ
e(7
1)fo
rpr
ogra
mm
edfu
ll-le
ngth
PA
Fsi
lenc
ing
RT
PC
Rof
mul
tiple
cell
deat
hco
ntru
cts
assa
yed
gene
ste
sted
NbP
DS
,HvP
DS
TM
VN
.be
nth
am
ian
a,P
hoto
blea
chin
gIR
inse
rten
hanc
edsp
read
ofM
echa
nica
lV
isua
l,Q
PC
R(7
9)B
SM
Vba
rley
begi
ns12
dpiT
MV,
sile
ncin
g,40
–60
ntIR
used
,14
dpib
arle
y20
0nt
IRno
teffe
ctiv
e
NtC
DP
K1,
NtR
PN
3P
VX
N.b
en
tha
mia
na
Cel
ldea
th,a
bnor
mal
RT
PC
Rof
mul
tiple
gene
sA
gro-
inoc
ulat
ion
RT-
PC
R(8
2)st
omat
alde
velo
pmen
tto
test
com
plex
resp
onse
,as
says
at20
dpi
NbW
IPK
,NbS
IPK
PV
XN
.be
nth
am
ian
aH
yper
sens
itive
Tim
eco
urse
RT-
PC
Rov
er4
hM
echa
nica
lR
T-P
CR
(131
)re
spon
sefo
llow
ing
chal
leng
ew
ithin
ocul
atio
nof
Imm
unob
lots
,kin
ase
fung
alel
icito
rin
vitr
o-m
ade
assa
ysfo
llow
ing
tran
scrip
tsw
ound
ing
orel
icito
r
WIP
K,S
IPK
PV
X35
S-N
gene
Loss
ofT
MV
-indu
ced
Bot
hW
IPK
and
SIP
KA
gro-
inoc
ulat
ion
Imm
unob
lots
(85)
N.b
en
tha
mia
na
kina
seex
pres
sion
prot
eins
asse
ssed
inre
cipr
ocal
sile
ncin
gex
pts
P58
(IP
K)
PV
XN
.be
nth
am
ian
aP
lant
deat
hfo
llow
ing
VIG
Sre
sults
confi
rmed
inA
gro-
inoc
ulat
ion
Vis
ual,
RT-
PC
R(1
3)vi
rus
infe
ctio
nA
rab
ido
psi
skno
ckou
tmut
ant
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 505
transmission must be considered and appropriate measures must be taken toprevent it.
Viruses from DNA vectors such as CaLCuV lack a coat protein and are notinfectious, but the same precautions should be taken, especially when the coatprotein is present. Geminiviruses are transmitted by whiteflies (Begomoviruses)or leaf hoppers (Mastreviruses and Curtoviruses). The TGMV vector comes froma virus that can no longer be transmitted by whiteflies and is no longer infectious intomato. Other isolates of TGMV were recently reported in Brazil and are infectiousin tomato (35). CaLCuV was identified in Florida as a pathogen of cabbage (53).The CaLCuV silencing vector developed forArabidopsislacks the coat proteinand therefore cannot be transmitted by whiteflies (139). Without the coat protein,CaLCuV is not infectious in cabbage, broccoli, and cauliflower by microprojectilebombardment (M. Flores & D. Robertson, unpublished data). Geminiviruses canevolve rapidly under field conditions (117), and even noninfectious vectors derivedfrom viral pathogens should be contained at all times (135, 136). The presenceof whiteflies should be monitored carefully and, in certain agricultural regions,geminivirus vectors should be used only if they lack a coat protein.
Several methods are commonly employed to deliver viral silencing vectorsto plants, including: mechanical inoculation using in vitro transcribed RNA orextracts from infected leaves, agroinoculation, and microprojectile bombardment.Mechanical inoculation is time consuming but can increase the efficiency of silenc-ing in certain hosts such asArabidopsis(120). Agroinoculation or agroinfiltrationhave been developed for both DNA and RNA viruses, as well as for transient si-lencing in the absence of virus (27, 130, 141). Tobacco, tomato, and barley vectorshave been developed that show extensive silencing with attenuated symptoms, andagroinfiltration of TRV vectors is becoming the vector of choice for many investiga-tors (Table 2). A web-based movie demonstrating syringe inoculation of Agrobac-terium into leaves can be found at http://www.sainsbury-laboratory.ac.uk/david-baulcombe/Services/agroInfil1.mpg, courtesy of D. Baulcombe. Agroinfiltrationof N. benthamianaleaves is routine and a similarly robust procedure is being devel-oped forArabidopsis(S. Kjemtrup, personal communication). The use of syringeswith needles can be used reliably (J. Ascensio & L. Hanley-Bowdoin, unpublisheddata), but the procedure is not suitable for high throughput.
Microprojectile bombardment of plasmid DNA-coated tungsten or gold micron-sized particles have been extremely useful for DNA viruses (102). Alternativemethods for virus inoculation are also available (34, 122).
Optimizing Silencing
The effectiveness of VIGS depends on environmental variables. For example,plants grown at higher temperatures show a stronger silencing response (97, 134).For TGMV, N. benthamianagrown in a 4′′ pot shows better silencing than whengrown in a 2′′ pot (C. Jordan & D. Robertson, unpublished work). Silencing inArabidopsisis more effective under short days, and inoculation at the 4–6-leaf
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
506 ROBERTSON
stage produces silencing in rosette leaves whereas inoculation at 10–12 leavestargets inflorescence tissue (140). These variables are easier to sort out when usinga target gene with a predicted visual or quantifiable phenotype.
Having a single gene as a silencing reference can help to sort out these variables.Choosing a target gene for a positive silencing control should be done carefullybecause different targets may impact VIGS vector replication, movement, andextent or timing of silencing, especially if unknown targets are characterized.Because transgene silencing can be propagated independently of the vector, its useas a silencing control for endogenous gene silencing is limited (54, 64, 72, 112,126).
GSA, PDS, ChlI, ChlH, andPAIgenes have visual phenotypes following silenc-ing (11, 57, 61, 77). The sequence from the tobaccoChlI ortholog (also calledsu)was originally identified by transposon tagging thesulfur locus of tobacco (31, 48,105).Chl1-silencedN. benthamianashow a uniform progression of silencing andflower normally and set seed, but they have reduced leaf size, stature, and numberof flowers (112). Silencing ofChlI from TGMV has been maintained for morethan 52 days by pruning plants to prevent them from producing seed (112).
Choosing only one gene for a silencing control is important for standardiz-ing epigenitic variation due to environmental conditions. For example, silencingdifferent subunits (ChlH versus ChlI) of the magnesium chelatase complex is ex-pected to produce different results. Expression levels of the nuclear-encodedLHCPmRNA in ChlH andChlI Arabidopsismutants showed that the nuclear-encodedLHCP mRNA decreased inchlH but not inchlI. TheChlH subunit participatesin a plastid-to-nucleus signaling pathway that regulates photosynthesis-associatedgene expression (99). Photobleaching could also impact silencing results; however,when chloramphenicol was added tochlH mutants to inhibit chloroplast transla-tion, LHCP expression was restored. Therefore, repression depended on activechloroplast translation in the albino plants. Although silencingChlH may causeunintended downregulation of other photosynthesis-related genes, bothChlH andChlI are reliable markers and are useful for studying the silencing dynamics ofendogenous genes (55, 56).
Pathways for Diffusible Silencing Signals
To use VIGS to target specific genes, it is useful to understand the dynamic natureof silencing. Homology-dependent gene silencing was previously classified asTGS or PTGS depending on whether transcripts for the silenced gene were foundin the nucleus (94). Silencing machinery is not necessarily active in TGS, whereasPTGS degrades incoming viral RNA and causes a dominant silencing phenotype(37, 80). There is now abundant evidence that RNA causes changes in chromatinstructure that prevent transcription leading to TGS (63, 113, 114, 150, 151, 155).In plants, a picture is emerging where transgenes are methylated in regions thatcorrespond to RNA homology, whereas endogenous genes appear to escape suchmethylation (95, 96).
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 507
The idea that there are similar pathways for virus movement and silencingsignals is supported by the identification of nontoxic levels of cadmium as aninhibitor of PTGS spread and tobamovirus movement in plants (142) and thecorrelation of anti-silencing and viral RNA accumulation in different tissues andplants (69, 128, 148). Other pathways for silencing signals cannot be ruled out andevidence for two kinds of movement has been presented (54). The dynamic natureof plasmodesmatal trafficking of nucleic acids was shown using a TMV vectorengineered to carry an additional copy of its movement protein translationallyfused to GFP (107). Epidermal spread of GFP from an inoculated area was transientand occurred only in cells at the leading edge of the TMV infection.
The dynamic nature of early events in endogenous gene silencing by TGMV::ChlI in Nicotiana is unusual in that discrete yellow spots 0.5–3 mm form ap-proximately five days post-bombardment. Spots are also seen whenN. tabacum,a TGMV host that requires the coat protein gene for movement, is bombarded(C. Peele & D. Robertson, unpublished work). Similar spots are not seen inCaLCuV::ChlI-infectedArabidopsisand it takes 12–14 days to see evidence of si-lencing in new growth (140). The rapid appearance and uniformity of TGMV::ChlI-induced silencing spots inN. benthamianacould be evidence for an apoplasticcomponent of spread (112). Noncell-autonomous movement of nucleic acids andprotein is an exciting area of research (51, 127, 159).
VIGS of transcribed sequences is confined to the inoculated plant and is lostalong with the viral inducer in subsequent generations. VIGS of pretranscribedsequences, such as promoters, can cause methylation, presumably due to RNA-DNA interactions (64). Methylated promoters are not transcribed efficiently, andshow reduced expression of their genes. Methylation is associated with chromatinchanges and can be inherited independently from the VIGS vector. Paradoxically,TRV carrying 35S sequence can cause methylation of 35S promoter sequencesthat extend into subsequent generations, but the TRV vector itself is not seedtransmitted, although the wild-type TRV is (65; Table 1).
Gene Function Studies
VIGS has been used to silence a wide variety of genes in plants (Table 2). Therehave been elegant studies combining VIGS with biochemical and genetic methodsto determine gene function, and they are producing a coherent picture of genefunction (13, 15, 62, 71, 82). Loss of function mutations of some of these geneswould be embryo lethals (such as PCNA, which is required for DNA replication,and mutations affecting chlorophyll stability). VIGS has been especially power-ful for dissecting signaling components involved in disease resistance (87, 111,125). Dramatic phenotypic effects are seen in pathogen-infected host plants si-lenced for the corresponding defense genes. Note that most of these studies usedN. benthamianaas a host (Table 2).
N. benthamianahas been extensively used for silencing studies because it is asuitable host for a wide range of viruses (Table 2). It can be readily transformed by
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
508 ROBERTSON
Agrobacterium cocultivation of leaf discs, self-pollinates, flowers rapidly, and is ofsmaller stature than tobacco.N. benthamianahas advantages over Arabidopsis thatinclude limited symptoms and abundant leaf material for biochemical analysis.N.tabacum, tomato, and barley also have well-developed VIGS vectors. VIGS vectorsfor Arabidopsis are currently limited by the need to passage vectors throughN.benthamianato get virions (TRV) or severe symptoms (CaLCuV). Modificationsin these vectors, or host plant mutations, may increase the effectiveness of TRVand CaLCuV in Arabidopsis.
The power of VIGS is its rapid initiation of silencing in intact wild-type ortransgenic plants. The ability to reliably silence 1–2 genes can provide materialfor biochemical analysis, metabolic profiling, and transcript profiling, if suitablecontrols are included. The use of RNA-based silencing to modulate mRNA lev-els can be an advantage and some RNA vectors show predictable increases anddecreases of silencing over time (58). Networks of genetic and protein interac-tions change when mRNA levels for individual genes are altered and informationcan be obtained for both medium and high levels of silencing, as demonstrated inArabidopsis (18). Keep in mind that silencing is a method for modulating geneexpression, not eliminating it. Quantitative measures of target mRNA levels shouldbe measured for each silencing event to be useful for genomic studies.
In-depth characterization of gene function must be accompanied by other meth-ods to verify VIGS-related analyses. Reverse transcriptase PCR (RT-PCR) of genenetworks, testing overexpression lines, and using transgenic lines carrying hairpinsilencing cassettes can help to insure that effects are gene specific rather than VIGSspecific. For plants with sequenced genomes, homology searches can be done toavoid unintended silencing. Potential effects of transitive silencing, which refer tosilencing initiated from mRNA sequences adjacent to the area of target homology,should also be considered. In plants, transitive RNAi has only been demonstratedfor transgenes (121, 143, 146).
FUNCTIONAL GENOMICS
Integration of RNAi data in combination with transcript profiling and protein-protein interaction studies is yielding computer-intensive ways to explore develop-ment and gene regulation (39). RNAi inC. eleganshas been used for genome-widefunctional genomics (33, 66, 67). Because different kinds of information can begained from large-scale genomics approaches, it would be useful to develop similarsystems in plants. hpRNA has been used on plants in functional genomics (152).VIGS can supplant hpRNA studies by targeting genes that are hard to silence byhpRNA and by providing a bridge for testing conservation of gene function fromArabidopsisto crop plants.
Large-scale silencing projects using transformation with constructs that con-tain hpRNA are in progress and high-throughput vectors are available (41, 52).
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 509
There is an extensive collection of RNA silencing–based transformants, PCRprimers, and vectors forArabidopsischromatin-associated genes (for informa-tion, see http://www.chromdb.org/). The project also shows quantitative data foreach transformant using RNA gel blots or RT-PCR. This is especially importantbecause RNA-based silencing does not eliminate gene expression and silencinglevels vary in different transformants (18).
There are caveats to using hpRNA to determine gene function. Although thehpRNA-induced silencing is heritable, epigenetic effects may result in unexpectedsilencing in subsequent generations, and transcriptional silencing of the inducinghpRNA locus may restore target gene expression (7, 78). Archiving and maintain-ing seed stocks are now being done for T-DNA insertion lines, but they requirean enormous investment. Archiving vectors is easier, and the Chromatin databaseproject has made such constructs available through TheArabidopsisInforma-tion Resource (TAIR). Additionally, unless transformation technologies improve,it will be hard to generate similar resources for plants that are recalcitrant totransformation.
Summaries of high-throughput screens using VIGS were reported inNicotianausing TMV and in barley using BSMV (30). In a second report, a screen of 5000genes for disease resistance resulted in approximately 100 clones whose silencinginterfered with a cell death assay inN. benthamiana(89). Of these, about 10 geneswere directly involved in disease resistance and 90 had unrelated loss of cell deathphenotypes. Although only a small number of genes were identified, this studydemonstrates that VIGS can be used to rapidly identify candidates for furthertesting.
Although there has been a lack of a suitable high-throughput VIGS vectorfor Arabidopsis, there are many public resources available for determining genefunction (one is TAIR at http://arabidopsis.org/). VIGS could be a useful additionfor verifying mutant phenotypes from plants that may have more than one mutationand for silencing combination of genes. The geminivirus vector CaLCuV causesextensive silencing inArabidopsisbut the associated symptoms limit the usefulnessof this vector, especially for developmental studies (140). Because geminivirussymptoms and silencing can be uncoupled (101a), refinements to the CaLCuVVIGS system will improve its usefulness in the near future.
Future Directions
Libraries of VIGS vectors with sequenced inserts would be useful resources forfunctional genomics studies. Unlike transgenes, which are subject to epigeneticmodifications, VIGS vectors can be used for reliable silencing and can be used indifferent genetic backgrounds and for various screens. This versatility is especiallyuseful because many phenotypes are not evident until proper environmental con-ditions are reached. Environment by genotype variations (for example, in stresstolerance assays) will likely require large numbers of plants and it is difficult topredict what genes will be useful for further testing before these kinds of studies are
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
510 ROBERTSON
performed. Because seed storage and archiving can be labor-intensive processes,developing community resources for RNA silencing–based vectors would be use-ful. Known combinations of PCR primers for silencing sequences could be archivedalong with descriptions of genotypes used and resulting phenotypes. For example,the internationalC. elegansdatabase incorporates a searchable index for RNAiphenotypes at http://www.wormbase.org/db/searches/rnaisearch. The databasearchives information for PCR primers and includes time-lapse studies of develop-ment (115).
An advantage of using viruses for reverse genetics is that most hard-to-transformcrop plants are susceptible to viruses, which could in turn be potential silencingvectors. Viruses with broad host range, such as BSMV and TRV, will be usefulfor extending functional data from model systems to crop plants. Geminivirusesare also attractive for gene silencing vectors because their genome structure isconserved and they infect a wide range of crop plants including soybean, cotton,and vegetable crops. Once a VIGS system is identified and optimized, one can testorthologous genes from model systems for conservation of function because it ispossible that similar genes will produce proteins that acquire different functionsin different plants (21, 101).
Identifying a plant that is supersensitive to silencing may be possible whenthe mechanism of silencing in plants is better understood. Such a mutant inC. eleganshas been identified and maps (counterintuitively) to a RdRP-like gene(132). However, until a method for inducing PTGS of endogenous genes in plantsis developed that is comparable to RNAi inC. elegans, combinations of VIGSand hpRNA stable transformation methods will likely be the method of choice forfunctional genomics in most plant species.
ACKNOWLEDGMENTS
I am indebted to George Allen, Chad Jordan, and Miguel Flores for many stimu-lating discussions, and to Susanne Kjemtrup, Anton Calloway, and Linda Hanley-Bowdoin for suggestions and help.
APPENDIX
PDS, phytoene desaturase;ChsA, chalcone synthase;RbcS, small subunit ofribulose-bisphosphate carboxylase/oxygenase;TK, plastid transketolase;CesA1,cellulose synthase subunit A;AlS, acetolactate synthase.Ppx, protoporphyrin IXoxidase;Gln1, glutamine synthetase;RPII, RNA polymerase II;Cat1, catalase1 PK1, kinasePARP, poly(ADP-ribose) polymerase;N gene, Toll Interleukin1/nucleotide-binding site/leucine-rich repeat resistance gene; TuA, alpha tubu-lin, WIPK, wound-induced protein kinase,SIPK, salicylic acid–induced proteinkinase;ChlH (magnesium chelatase subunit H),p58(IPK) ortholog of dsRNA-dependent PKR inhibitor;PAF, alpha subunit of 26S proteosome,RPN1, secondcomponent of 26S proteosome.
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 511
The Annual Review of Plant Biologyis online at http://plant.annualreviews.org
LITERATURE CITED
1. Abbink TE, Peart JR, Mos TN,Baulcombe DC, Bol JF, Linthorst HJ.2002. Silencing of a gene encoding a pro-tein component of the oxygen-evolvingcomplex of photosystem II enhances virusreplication in plants.Virology 295:307–19
2. Al-Kaff NS, Covey SN,Kreike MM, PageAM, Pinder R, Dale PJ. 1998. Transcrip-tional and posttranscriptional plant genesilencing in response to a pathogen.Sci-ence279:2113–15
3. Al-Kaff NS, Kreike MM, Covey SN,Pitcher R, Page AM, Dale PJ. 2000.Plants rendered herbicide-susceptible bycauliflower mosaic virus-elicited suppres-sion of a 35S promoter-regulated trans-gene.Nat. Biotechnol.18:995–99
4. Allen GC, Spiker S, Thompson WF. 2000.Use of matrix attachment regions (MARs)to minimize transgene silencing.PlantMol. Biol. 43:361–76
5. Anandalakshmi R, Pruss GJ, Ge X,Marathe R, Mallory AC, et al. 1998. A vi-ral suppressor of gene silencing in plants.Proc. Natl. Acad. Sci. USA95:13079–84
6. Angell SM, Baulcombe DC. 1999. Tech-nical advance: potato virus X amplicon-mediated silencing of nuclear genes.PlantJ. 20:357–62
7. Ascenzi R, Ulker B, Todd JJ, SowinskiDA, Schimeneck CR, et al. 2003. Analysisof trans-silencing interactions using tran-scriptional silencers of varying strengthand targets with and without flanking nu-clear matrix attachment regions.Trans-genic Res.12:305–18
8. Barton KA, Binns AN, Matzke AJ,Chilton MD. 1983. Regeneration of in-tact tobacco plants containing full lengthcopies of genetically engineered T-DNA,and transmission of T-DNA to R1progeny.Cell 32:1033–43
9. Bass HW, Nagar S, Hanley-Bowdoin L,
Robertson D. 2000. Chromosome con-densation induced by geminivirus infec-tion of mature plant cells.J. Cell Sci.113:1149–60
10. Beclin C, Berthome R, Palauqui JC,Tepfer M, Vaucheret H. 1998. Infectionof tobacco or Arabidopsis plants by CMVcounteracts systemic post-transcriptionalsilencing of nonviral (trans)genes.Virol-ogy252:313–17
11. Bender J, Fink GR. 1995. Epigenetic con-trol of an endogenous gene family is re-vealed by a novel blue fluorescent mutantof Arabidopsis. Cell83:725–34
12. Bernstein E, Caudy AA, Hammond SM,Hannon GJ. 2001. Role for a bidentate ri-bonuclease in the initiation step of RNAinterference.Nature409:363–66
13. Bilgin DD, Liu Y, Schiff M, Dinesh-Kumar SP. 2003. P58(IPK), a plantortholog of double-stranded RNA-dependent protein kinase PKR inhibitor,functions in viral pathogenesis.Dev. Cell4:651–61
14. Brough CL, Hayes RJ, Morgan AJ, CouttsRHA, Buck KW. 1988. Effects of muta-genesis in vitro on the ability of clonedtomato golden mosaic virus DNA to infectNicotiana benthamianaplants.J. Gen. Vi-rol. 69:503–14
15. Burton RA, Gibeaut DM, Bacic A, Find-lay K, Roberts K, et al. 2000. Virus-induced silencing of a plant cellulose syn-thase gene.Plant Cell12:691–706
16. Cerutti H. 2003. RNA interference: trav-eling in the cell and gaining functions?Trends Genet.19:39–46
17. Chi JT, Chang HY, Wang NN, ChangDS, Dunphy N, Brown PO. 2003.Genomewide view of gene silencing bysmall interfering RNAs.Proc. Natl. Acad.Sci. USA100:6343–46
18. Chuang CF, Meyerowitz EM. 2000. Spe-cific and heritable genetic interference
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
512 ROBERTSON
by double-stranded RNA inArabidop-sis thaliana. Proc. Natl. Acad. Sci. USA97:4985–90
19. Cogoni C, Macino G. 2000. Post-transcriptional gene silencing across king-doms.Curr. Opin. Genet. Dev.10:638–43
19a. Couzin J. 2002. Breakthrough of the year.Small RNAs make big splash.Science298:2296–97
20. Covey SN,Al-Kaff NS. 2000. Plant DNAviruses and gene silencing.Plant Mol.Biol. 43:307–22
21. Cremer F, Coupland G. 2003. Distinctphotoperiodic responses are conferred bythe same genetic pathway inArabidopsisand in rice.Trends Plant Sci.8:405–7
22. Dallwitz MJ, Zurcher EJ. 1996.PlantViruses Online: Descriptions and Listsfrom the VIDE Database, ed. AA Brunt, KCrabtree, MJ Dallwitz, AJ Gibbs, L Wat-son, EJ Zurcher. UK: C.A.B. Intl. 1484 pp.http://image.fs.uidaho.edu/vide/refs.htm
23. Dinesh-Kumar SP, Anandalakshmi R,Marathe R, Schiff M, Liu Y. 2003. Virusinduced gene silencing.Methods in Mol.Biol. 236:287–94
23a. Dodds JA. 1999. Satellite tobacco mo-saic virus.Curr. Top. Microbiol. Immunol.239:145–57
24. Dougherty WG, Lindbo JA, Smith HA,Park TD, Swaney S, Proebsting WL.1994. RNA-mediated virus resistance intransgenic plants: exploitation of a cel-lular pathway possibly involved in RNAdegradation.Mol. Plant Microbe. Inter-act.7:544–52
25. Egelkrout EM, Robertson D, Hanley-Bowdoin L. 2001. Proliferating cell nu-clear antigen transcription is repressedthrough an E2F consensus element and ac-tivated by geminivirus infection in matureleaves.Plant Cell13:1437–52
26. Elmer JS, Brand L, Sunter G, GardinerWE, Bisaro DM, Rogers SG. 1988. Ge-netic analysis of the tomato golden mosaicvirus. II. The product of the AL1 codingsequence is required for replication.Nu-cleic Acids Res.16:7043–60
27. Elmer JS, Sunter G, Gardiner WE,Brand L, Browning CK, et al. 1988.Agrobacterium-mediated inoculation ofplants with tomato golden mosaic virusDNAs. Plant Mol. Biol.10:225–34
28. Ermak G, Paszkowski U, Wohlmuth M,Scheid OM, Paszkowski J. 1993. Cyto-sine methylation inhibits replication ofAfrican cassava mosaic virus by 2 dis-tinct mechanisms.Nucleic Acids Res.21:3445–50
29. Fire A, Xu S, Montgomery MK, KostasSA, Driver SE, Mello CC. 1998. Po-tent and specific genetic interference bydouble-stranded RNA in Caenorhabditiselegans.Nature391:806–11
30. Fitzmaurice WP, Holzberg S, Lindbo JA,Padgett HS, Palmer KE, et al. 2002. Epi-genetic modification of plants with sys-temic RNA viruses.Omics6:137–51
31. Fitzmaurice WP, Nguyen LV, WernsmanEA, Thompson WF, Conkling MA. 1999.Transposon tagging of the sulfur gene oftobacco using engineered maize Ac/Ds el-ements.Genetics153:1919–28
32. Fraley RT, Rogers SG, Horsch RB,Sanders PR, Flick JS, et al. 1983. Expres-sion of bacterial genes in plant cells.Proc.Natl. Acad. Sci. USA80:4803–7
33. Fraser AG, Kamath RS, ZipperlenP, Martinez-Campos M, Sohrmann M,Ahringer J. 2000. Functional genomicanalysis of C. elegans chromosome Iby systematic RNA interference.Nature408:325–30
34. Gal-On A, Meiri E, Elman C, Gray DJ,Gaba V. 1997. Simple hand-held de-vices for the efficient infection of plantswith viral-encoding constructs by particlebombardment.J. Virol. Methods64:103–10
35. Galvao RM, Mariano AC, Luz DF, Alfe-nas PF, Andrade EC, et al. 2003. A natu-rally occurring recombinant DNA-A of atypical bipartite begomovirus does not re-quire the cognate DNA-B to infect Nico-tiana benthamiana systemically.J. Gen.Virol. 84:715–26
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 513
36. Gillespie T, Boevink P, Haupt S, RobertsAG, Toth R, et al. 2002. Functional analy-sis of a DNA-shuffled movement proteinreveals that microtubules are dispensablefor the cell-to-cell movement of tobaccomosaic virus.Plant Cell14:1207–22
37. Goodwin J, Chapman K, Swaney S,Parks TD, Wernsman EA, DoughertyWG. 1996. Genetic and biochemical dis-section of transgenic RNA-mediated virusresistance.Plant Cell8:85–105
38. Gossele V, Fache I, Meulewaeter F, Cor-nelissen M, Metzlaff M. 2002. SVISS-anovel transient gene silencing system forgene function discovery and validation intobacco plants.Plant J.32:859–66
39. Grant BD, Wilkinson HA. 2003. Func-tional genomic maps in Caenorhabditis el-egans.Curr. Opin. Cell Biol.15:206–12
40. Guo HS, Ding SW. 2002. A viral pro-tein inhibits the long range signaling ac-tivity of the gene silencing signal.EMBOJ. 21:398–407
41. Guo HS, Fei JF, Xie Q, Chua NH. 2003. Achemical-regulated inducible RNAi sys-tem in plants.Plant J.34:383–92
42. Gutierrez C. 2000. Geminiviruses and theplant cell cycle.Plant Mol. Biol.43:763–72
43. Hamilton A, Voinnet O, Chappell L,Baulcombe D. 2002. Two classes ofshort interfering RNA in RNA silencing.EMBO J.21:4671–79
44. Hamilton AJ, Baulcombe DC. 1999. Aspecies of small antisense RNA in post-transcriptional gene silencing in plants.Science286:950–52
45. Hamilton WDO, Stein VE, Coutts RHA,Buck KW. 1984. Complete nucleotidesequence of the infectious cloned DNAcomponents of tomato golden mosaicvirus: Potential coding regions and reg-ulatory sequences.EMBO J.3:2197–205
46. Hammond SM, Bernstein E, Beach D,Hannon GJ. 2000. An RNA-directed nu-clease mediates post-transcriptional genesilencing in Drosophila cells.Nature404:293–96
47. Hanley-Bowdoin L, Settlage SB, OrozcoBM, Nagar S, Robertson D. 2000. Gem-iniviruses: models for plant DNA repli-cation, transcription, and cell cycle reg-ulation. Crit. Rev. Biochem. Mol. Biol.35:105–40
47a. Hanley-Bowdoin L, Settlage SB, Robert-son D. 2004. Reprogramming plant geneexpression—a prerequisite to geminivirusDNA replication. Mol. Plant Path. Int.In press
48. Hansson A, Kannangara CG, vonWettstein D, Hansson M. 1999. Molecu-lar basis for semidominance of missensemutations in the XANTHA-H (42-kDa)subunit of magnesium chelatase.Proc.Natl. Acad. Sci. USA96:1744–49
49. Havelda Z, Hornyik C, Crescenzi A, Bur-gyan J. 2003. In situ characterization ofCymbidium Ringspot Tombusvirus infec-tion-induced posttranscriptional genesilencing in Nicotiana benthamiana.J.Virol. 77:6082–86
50. Hayes RJ, Petty IDT, Coutts RHA, BuckKW. 1988. Gene amplification and ex-pression in plants by a replicating gem-inivirus vector.Nature334:179–82
51. Haywood V, Kragler F, Lucas WJ. 2002.Plasmodesmata: pathways for protein andribonucleoprotein signaling.Plant Cell14(Suppl.):S303–25
52. Helliwell C, Waterhouse P. 2003. Con-structs and methods for high-throughputgene silencing in plants.Methods30:289–95
53. Hill JE, Strandberg JO, Hiebert E,Lazarowitz SG. 1998. Asymmetric in-fectivity of pseudorecombinants of cab-bage leaf curl virus and squash leaf curlvirus: implications for bipartite gemi-nivirus evolution and movement.Virology250:283–92
54. Himber C, Dunoyer P, MoissiardG, Ritzenthaler C, Voinnet O. 2003.Transitivity-dependent and -independentcell-to-cell movement of RNA silencing.EMBO J.22:4523–33
55. Hiriart JB, Aro EM, Lehto K. 2003.
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
514 ROBERTSON
Dynamics of the VIGS-mediatedchimeric silencing of the Nicotiana ben-thamiana ChlH gene and of the tobaccomosaic virus vector.Mol. Plant MicrobeInteract.16:99–106
56. Hiriart JB, Lehto K, Tyystjarvi E, JunttilaT, Aro EM. 2002. Suppression of a keygene involved in chlorophyll biosynthesisby means of virus-inducing gene silenc-ing. Plant Mol. Biol.50:213–24
57. Hofgen R, Axelsen KB, KannangaraCG, Schuttke I, Pohlenz HD, et al.1994. A visible marker for antisensemRNA expression in plants: inhibition ofchlorophyll synthesis with a glutamate-1-semialdehyde aminotransferase anti-sense gene.Proc. Natl. Acad. Sci. USA91:1726–30
58. Holzberg S, Brosio P, Gross C, Pogue GP.2002. Barley stripe mosaic virus-inducedgene silencing in a monocot plant.PlantJ. 30:315–27
59. Hormuzdi SG, Bisaro DM. 1995. Geneticanalysis of beet curly top virus: Exami-nation of the roles of L2 and L3 genesin viral pathogenesis.Virology206:1044–54
60. Hull R. 2001.Matthew’s Plant Virology.San Diego: Academic. 340 pp.
61. Jeddeloh JA, Bender J, Richards EJ. 1998.The DNA methylation locus DDM1 is re-quired for maintenance of gene silencingin Arabidopsis.Genes Dev.12:1714–25
62. Jin H, Axtell MJ, Dahlbeck D, EkwennaO, Zhang S, et al. 2002. NPK1, anMEKK1-like mitogen-activated proteinkinase kinase kinase, regulates innate im-munity and development in plants.Dev.Cell 3:291–97
63. Jones AL, Thomas CL, Maule AJ. 1998.De novo methylation and co-suppressioninduced by a cytoplasmically replicatingplant RNA virus.EMBO J.17:6385–93
64. Jones L, Hamilton AJ, Voinnet O, ThomasCL, Maule AJ, Baulcombe DC. 1999.RNA-DNA interactions and DNA methy-lation in post-transcriptional gene silenc-ing. Plant Cell11:2291–301
65. Jones L, Ratcliff F, Baulcombe DC. 2001.RNA-directed transcriptional gene silenc-ing in plants can be inherited inde-pendently of the RNA trigger and re-quires Met1 for maintenance.Curr. Biol.11:747–57
66. Kamath RS, Ahringer J. 2003. Genome-wide RNAi screening in Caenorhabditiselegans.Methods30:313–21
67. Kamath RS, Fraser AG, Dong Y, PoulinG, Durbin R, et al. 2003. Systematicfunctional analysis of the Caenorhabdi-tis elegans genome using RNAi.Nature421:231–37
68. Kasschau KD, Carrington JC. 1998. Acounterdefensive strategy of plant viruses:suppression of posttranscriptional gene si-lencing.Cell 95:461–70
69. Kasschau KD, Carrington JC. 2001.Long-distance movement and replicationmaintenance functions correlate with si-lencing suppression activity of potyviralHC-Pro.Virology285:71–81
70. Ketting RF, Plasterk RH. 2000. A geneticlink between co-suppression and RNA in-terference in C. elegans.Nature404:296–98
71. Kim M, Yang KS, Kim YK, Paek KH, PaiHS. 2003. Molecular characterization ofNbPAF encoding the alpha6 subunit of the20S proteasome in Nicotiana benthami-ana.Mol. Cells15:127–32
72. Kjemtrup S, Sampson KS, Peele CG,Nguyen LV, Conkling MA, et al. 1998.Gene silencing from plant DNA carriedby a Geminivirus.Plant J.14:91–100
73. Deleted in proof74. Klahre U, Crete P, Leuenberger SA, Igle-
sias VA, Meins F Jr. 2002. High molecularweight RNAs and small interfering RNAsinduce systemic posttranscriptional genesilencing in plants.Proc. Natl. Acad. Sci.USA99:11981–86
75. Klinkenberg FA, Ellwood S, Stanley J.1989. Fate of African cassava mosaicvirus coat protein deletion mutants afteragroinoculation.J. Gen. Virol.70:1837–44
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 515
76. Kong LJ, Orozco BM, Roe JL, Nagar S,Ou S, et al. 2000. A geminivirus replica-tion protein interacts with the retinoblas-toma protein through a novel domain todetermine symptoms and tissue speci-ficity of infection in plants.EMBO J.19:3485–95
77. Kumagai MH, Donson J, della-Cioppa G,Harvey D, Hanley K, Grill LK. 1995. Cy-toplasmic inhibition of carotenoid biosyn-thesis with virus-derived RNA.Proc.Natl. Acad. Sci. USA92:1679–83
78. Kumpatla SP, Hall TC. 1998. Recurrentonset of epigenetic silencing in rice har-boring a multi-copy transgene.Plant J.14:129–35
79. Lacomme C, Hrubikova K, Hein I. 2003.Enhancement of virus-induced gene si-lencing through viral-based production ofinverted-repeats.Plant J.34:543–53
80. Lawrence RJ, Pikaard CS. 2003.Transgene-induced RNA interference: astrategy for overcoming gene redundancyin polyploids to generate loss-of-functionmutations.Plant J.36:114–21
81. Lazarowitz SG, Beachy RN. 1999. Viralmovement proteins as probes for intracel-lular and intercellular trafficking in plants.Plant Cell11:535–48
82. Lee SS, Cho HS, Yoon GM, Ahn JW,Kim HH, Pai HS. 2003. Interaction ofNtCDPK1 calcium-dependent protein ki-nase with NtRpn3 regulatory subunit ofthe 26S proteasome in Nicotiana tabacum.Plant J.33:825–40
83. Lindbo JA, Fitzmaurice WP, Della-Cioppa G. 2001. Virus-mediated repro-gramming of gene expression in plants.Curr. Opin. Plant Biol.4:181–85
84. Lindbo JA, Silva-Rosales L, ProebstingWL, Dougherty WG. 1993. Induction ofa highly specific anti-viral state in trans-genic plants: implications for gene reg-ulation and virus resistance.Plant Cell5:1749–59
85. Liu Y, Jin H, Yang KY, Kim CY, Baker B,Zhang S. 2003. Interaction between twomitogen-activated protein kinases dur-
ing tobacco defense signaling.Plant J.34:149–60
86. Liu Y, Schiff M, Dinesh-Kumar SP. 2002.Virus-induced gene silencing in tomato.Plant J.31:777–86
87. Liu Y, Schiff M, Marathe R, Dinesh-Kumar SP. 2002. Tobacco Rar1, EDS1and NPR1/NIM1 like genes are requiredfor N-mediated resistance to tobacco mo-saic virus.Plant J.30:415–29
88. Liu Y, Schiff M, Serino G, Deng XW,Dinesh-Kumar SP. 2002. Role of SCFubiquitin-ligase and the COP9 signalo-some in the N gene-mediated resistanceresponse to tobacco mosaic virus.PlantCell 14:1483–96
89. Lu R, Martin-Hernandez AM, Peart JR,Malcuit I, Baulcombe DC. 2003. Virus-induced gene silencing in plants.Methods30:296–303
90. Lucioli A, Noris E, Brunetti A, TavazzaR, Ruzza V, et al. 2003. Tomato yellowleaf curl Sardinia virus rep-derived re-sistance to homologous and heterologousgeminiviruses occurs by different mecha-nisms and is overcome if virus-mediatedtransgene silencing is activated.J. Virol.77:6785–98
91. Lucy AP, Guo HS, Li WX, Ding SW.2000. Suppression of post-transcriptionalgene silencing by a plant viral protein lo-calized in the nucleus.EMBO J.19:1672–80
92. MacFarlane SA, Popovich AH. 2000. Ef-ficient expression of foreign proteins inroots from tobravirus vectors.Virology267:29–35
93. Mallory AC, Parks G, Endres MW,Baulcombe D, Bowman LH, et al. 2002.The amplicon-plus system for high-levelexpression of transgenes in plants.Nat.Biotechnol.20:622–25
94. Matzke MA, Matzke AJM. 1995.Homology-dependent gene silencing intransgenic plants: what does it really tellus?Trends Genet.11:1–3
95. Matzke MA, Matzke AJM, Pruss GJ,Vance VB. 2001. RNA-based silencing
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
516 ROBERTSON
strategies in plants.Curr. Opin. Genet.Dev.11:221–27
96. Mette MF, Aufsatz W, van der WindenJ, Matzke MA, Matzke AJM. 2000. Tran-scriptional silencing and promoter methy-lation triggered by double-stranded RNA.EMBO J.19:5194–201
97. Meza TJ, Kamfjord D, Hakelien AM,Evans I, Godager LH, et al. 2001. Thefrequency of silencing in Arabidopsisthaliana varies highly between progeny ofsiblings and can be influenced by envi-ronmental factors.Transgenic Res.10:53–67
98. Mlotshwa S, Voinnet O, Mette MF,Matzke M, Vaucheret H, et al. 2002. RNAsilencing and the mobile silencing signal.Plant Cell14:S289–301
99. Mochizuki N, Brusslan JA, Larkin R,Nagatani A, Chory J. 2001. Arabidop-sis genomes uncoupled 5 (GUN5) mutantreveals the involvement of Mg-chelataseH subunit in plastid-to-nucleus signaltransduction.Proc. Natl. Acad. Sci. USA98:2053–58
100. Montgomery MK, Fire A. 1998.Double-stranded RNA as a mediator insequence-specific genetic silencing andco-suppression.Trends Genet.14:255–58
101. Mouradov A, Cremer F, Coupland G.2002. Control of flowering time: interact-ing pathways as a basis for diversity.PlantCell 14(Suppl.):S111–30
102. Muangsan N, Robertson D. 2004. Gemi-nivirus vectors for transient gene silencingin plants.Methods Molec. Biol.236:101–15
103. Nagar S, Hanley-Bowdoin L, RobertsonD. 2002. Host DNA replication is inducedby geminivirus infection of differentiatedplant cells.Plant Cell14:2995–3007
104. Napoli C, Lemieux C, Jorgensen R. 1990.Introduction of a chimeric chalcone syn-thase gene into petunia results in re-versible co-suppression of homologousgenes in trans.Plant Cell2:279–89
105. Nguyen LV. 1995.Transposon taggingand isolation of the sulfur gene in to-
bacco (Nicotiana tabacum).PhD thesis.N.C. State Univ.
106. Noueiry AO, Ahlquist P. 2003. Bromemosaic virus RNA replication: revealingthe role of the host in RNA virus replica-tion. Annu. Rev. Phytopathol.41:77–98
107. Oparka KJ, Prior DA, Santa Cruz S, Pad-gett HS, Beachy RN. 1997. Gating of epi-dermal plasmodesmata is restricted to theleading edge of expanding infection sitesof tobacco mosaic virus (TMV).Plant J.12:781–89
108. Palauqui JC, Elmayan T, Pollien JM,Vaucheret H. 1997. Systemic acquiredsilencing: transgene-specific post-transcriptional silencing is transmittedby grafting from silenced stocks to non-silenced scions.EMBO J.16:4738–45
109. Palmer KE, Rybicki EP. 2001. Investiga-tion of the potential of maize streak virusto act as an infectious gene vector in maizeplants.Arch. Virol.146:1089–104
110. Palmer KE, Thomson JA, Rybicki EP.1999. Generation of maize cell lines con-taining autonomously replicating maizestreak virus-based gene vectors.Arch. Vi-rol. 144:1345–60
111. Peart JR, Cook G, Feys BJ, Parker JE,Baulcombe DC. 2002. An EDS1 ortho-logue is required for N-mediated resis-tance against tobacco mosaic virus.PlantJ. 29:569–79
112. Peele C, Jordan CV, Muangsan N, Tur-nage M, Egelkrout E, et al. 2001.Silencing of a meristematic gene us-ing geminivirus-derived vectors.Plant J.27:357–66
113. Pelissier T, Thalmeir S, Kempe D, SangerHL, Wassenegger M. 1999. Heavy denovo methylation at symmetrical and non-symmetrical sites is a hallmark of RNA-directed DNA methylation.Nucleic AcidsRes.27:1625–34
114. Pelissier T, Wassenegger M. 2000. ADNA target of 30 bp is sufficient for RNA-directed DNA methylation.RNA6:55–65
115. Piano F, Schetter AJ, Mangone M, SteinL, Kemphues KJ. 2000. RNAi analysis
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 517
of genes expressed in the ovary ofCaenorhabditis elegans.Curr. Biol. 10:1619–22
116. Pickford AS, Cogoni C. 2003. RNA-mediated gene silencing.Cell Mol. LifeSci.60:871–82
117. Pita JS, Fondong VN, Sangare A, Otim-Nape GW, Ogwal S, Fauquet CM.2001. Recombination, pseudorecombina-tion and synergism of geminiviruses aredeterminant keys to the epidemic of se-vere cassava mosaic disease in Uganda.J.Gen. Virol.82:655–65
118. Pogue GP, Lindbo JA, Garger SJ, Fitz-maurice WP. 2002. Making an ally froman enemy: plant virology and the new agri-culture.Annu. Rev. Phytopathol.40:45–74
119. Ratcliff F, Harrison BD, Baulcombe DC.1997. A similarity between viral defenseand gene silencing in plants.Science276:1558–60
120. Ratcliff F, Martin-Hernandez AM,Baulcombe DC. 2001. Technical ad-vance. Tobacco rattle virus as a vector foranalysis of gene function by silencing.Plant J.25:237–45
121. Ratcliff FG, MacFarlane SA, BaulcombeDC. 1999. Gene silencing without DNA.RNA-mediated cross-protection betweenviruses.Plant Cell11:1207–16
122. Redinbaugh MG, Louie R, Ngwira P,Edema R, Gordon DT, Bisaro DM. 2001.Transmission of viral RNA and DNAto maize kernels by vascular punctureinoculation. J. Virol. Methods98:135–43
123. Rigden JE, Krake LR, Rezaian MA, DryIB. 1994. ORF C4 of tomato leaf curlgeminivirus is a determinant of symptomseverity.Virology204:847–50
124. Rogers SG, Bisaro DM, Horsch RB,Fraley RT, Hoffmann NL, et al. 1986.Tomato golden mosaic virus A componentDNA replicates autonomously in trans-genic plants.Cell 45:593–600
125. Romeis T, Ludwig AA, Martin R, JonesJD. 2001. Calcium-dependent protein ki-
nases play an essential role in a plant de-fence response.EMBO J.20:5556–67
126. Ruiz MT, Voinnet O, Baulcombe DC.1998. Initiation and maintenance ofvirus-induced gene silencing.Plant Cell10:937–46
127. Ruiz-Medrano R, Xoconostle-Cazares B,Lucas WJ. 2001. The phloem as a con-duit for inter-organ communication.Curr.Opin. Plant Biol.4:202–9
128. Saenz P, Salvador B, Simon-Mateo C,Kasschau KD, Carrington JC, Garcia JA.2002. Host-specific involvement of theHC protein in the long-distance move-ment of potyviruses.J. Virol. 76:1922–31
129. Saitoh H, Terauchi R. 2002. Virus-induced silencing of FtsH gene in Nico-tiana benthamiana causes a strikingbleached leaf phenotype.Genes Genet.Syst.77:335–40
130. Schob H, Kunz C, Meins F Jr. 1997.Silencing of transgenes introduced intoleaves by agroinfiltration: a simple, rapidmethod for investigating sequence re-quirements for gene silencing.Mol. Gen.Genet.256:581–85
131. Sharma PC, Ito A, Shimizu T, Terauchi R,Kamoun S, Saitoh H. 2003. Virus-inducedsilencing of WIPK and SIPK genes re-duces resistance to a bacterial pathogen,but has no effect on the INF1-inducedhypersensitive response (HR) in Nico-tiana benthamiana.Mol. Genet. Genomics269:583–91
132. Simmer F, Tijsterman M, Parrish S,Koushika SP, Nonet ML, et al. 2002. Lossof the putative RNA-directed RNA poly-merase RRF-3 makes C. elegans hyper-sensitive to RNAi.Curr. Biol.12:1317–19
133. Sudarshana MR, Wang HL, Lucas WJ,Gilbertson RL. 1998. Dynamics of beandwarf mosaic geminivirus cell-to-cell andlong-distance movement in Phaseolusvulgaris revealed, using the green fluores-cent protein.Mol. Plant-Microbe Interact.11:277–91
134. Szittya G, Silhavy D, Molnar A, HaveldaZ, Lovas A, et al. 2003. Low temperature
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
518 ROBERTSON
inhibits RNA silencing-mediated defenceby the control of siRNA generation.EMBO J.22:633–40
135. Tepfer M. 2002. Risk assessment of virus-resistant transgenic plants.Annu. Rev.Phytopathol.40:467–91
136. Tepfer M, Jacquemond M. 1996. Sleepingsatellites: a risky prospect.Nat. Biotech-nol. 14:1226
137. Teycheney PY, Tepfer M. 2001. Virus-specific spatial differences in the inter-ference with silencing of the chs-A genein non-transgenic petunia.J. Gen. Virol.82:1239–43
138. Timmermans M, Das O, Messing J. 1994.Geminivirus and their uses as extrachro-mosomeal replicons.Annu. Rev. PlantPhysiol.45:79–112
139. Deleted in proof140. Turnage MA, Muangsan N, Peele CG,
Robertson D. 2002. Geminivirus-basedvectors for gene silencing in Arabidopsis.Plant J.30:107–14
141. Turpen TH, Turpen AM, Weinzettl N, Ku-magai MH, Dawson WO. 1993. Trans-fection of whole plants from woundsinoculated with Agrobacterium tumefa-ciens containing cDNA of tobacco mosaicvirus.J. Virol. Methods42:227–39
142. Ueki S, Citovsky V. 2001. RNA commutesto work: regulation of plant gene expres-sion by systemically transported RNAmolecules.BioEssays23:1087–90
143. Vaistij FE, Jones L, Baulcombe DC. 2002.Spreading of RNA targeting and DNAmethylation in RNA silencing requirestranscription of the target gene and a pu-tative RNA-dependent RNA polymerase.Plant Cell14:857–67
144. Vance V, Vaucheret H. 2001. RNA silenc-ing in plants–defense and counterdefense.Science292:2277–80
145. van der Krol AR, Mur LA, Beld M, MolJN, Stuitje AR. 1990. Flavonoid genes inpetunia: addition of a limited number ofgene copies may lead to a suppression ofgene expression.Plant Cell2:291–99
146. Vanitharani R, Chellappan P, Fauquet
CM. 2003. Short interfering RNA-mediated interference of gene expressionand viral DNA accumulation in culturedplant cells.Proc. Natl. Acad. Sci. USA100:9632–36
147. Voinnet O, Baulcombe DC. 1997. Sys-temic signalling in gene silencing.Nature389:553
148. Voinnet O, Lederer C, Baulcombe DC.2000. A viral movement protein preventsspread of the gene silencing signal inNicotiana benthamiana.Cell 103:157–67
149. Voinnet O, Vain P, Angell S, BaulcombeDC. 1998. Systemic spread of sequence-specific transgene RNA degradation inplants is initiated by localized introduc-tion of ectopic promoterless DNA.Cell95:177–87
150. Volpe T, Schramke V, Hamilton GL,White SA, Teng G, et al. 2003. RNAinterference is required for normal cen-tromere function in fission yeast.Chro-mosome Res.11:137–46
151. Volpe TA, Kidner C, Hall IM, Teng G,Grewal SI, Martienssen RA. 2002. Reg-ulation of heterochromatic silencing andhistone H3 lysine-9 methylation by RNAi.Science297:1833–37
152. Wang MB, Waterhouse PM. 2002. Appli-cation of gene silencing in plants.Curr.Opin. Plant Biol.5:146–50
153. Ward BM, Medville R, Lazarowitz SG,Turgeon R. 1997. The geminivirus BL1movement protein is associated withendoplasmic reticulum-derived tubulesin developing phloem cells.J. Virol.71:3726–33
154. Wassenegger M. 2000. RNA-directedDNA methylation. Plant Mol. Biol.43:203–20
155. Wassenegger M, Heimes S, Riedel L,Sanger HL. 1994. RNA-directed de novomethylation of genomic sequences inplants.Cell 76:567–76
156. Wassenegger M, Pelissier T. 1998. Amodel for RNA-mediated gene silencingin higher plants.Plant Mol. Biol.37:349–62
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

23 Mar 2004 17:1 AR AR213-PP55-19.tex AR213-PP55-19.sgm LaTeX2e(2002/01/18)P1: GDL
VIGS VECTORS 519
157. Waterhouse PM, Graham MW, Wang MB.1998. Virus resistance and gene silencingin plants can be induced by simultaneousexpression of sense and antisense RNA.Proc. Natl. Acad. Sci. USA95:13959–64
158. Waterhouse PM, Helliwell CA. 2003. Ex-ploring plant genomes by RNA-inducedgene silencing.Nat. Rev. Genet.4:29–38
159. Wu X, Dinneny JR, Crawford KM, Rhee
Y, Citovsky V, et al. 2003. Modes of in-tercellular transcription factor movementin the Arabidopsis apex.Development130:3735–45
160. Muangsan N, Beclin C, Vaucheret H,Robertson D. 2004. Genetic requirementsfor geminivirus-mediated VIGS of en-dogenous genes defines a new branch forthe silencing pathway in plants.Plant J.In press
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

P1: FRK
March 24, 2004 1:1 Annual Reviews AR213-FM
Annual Review of Plant BiologyVolume 55, 2004
CONTENTS
AN UNFORESEEN VOYAGE TO THE WORLD OF PHYTOCHROMES,Masaki Furuya 1
ALTERNATIVE NAD(P)H DEHYDROGENASES OF PLANTMITOCHONDRIA, Allan G. Rasmusson, Kathleen L. Soole,and Thomas E. Elthon 23
DNA METHYLATION AND EPIGENETICS, Judith Bender 41
PHOSPHOENOLPYRUVATE CARBOXYLASE: A NEW ERA OFSTRUCTURAL BIOLOGY, Katsura Izui, Hiroyoshi Matsumura,Tsuyoshi Furumoto, and Yasushi Kai 69
METABOLIC CHANNELING IN PLANTS, Brenda S.J. Winkel 85
RHAMNOGALACTURONAN II: STRUCTURE AND FUNCTION OF ABORATE CROSS-LINKED CELL WALL PECTIC POLYSACCHARIDE,Malcolm A. O’Neill, Tadashi Ishii, Peter Albersheim, and Alan G. Darvill 109
NATURALLY OCCURRING GENETIC VARIATION IN ARABIDOPSISTHALIANA, Maarten Koornneef, Carlos Alonso-Blanco, andDick Vreugdenhil 141
SINGLE-CELL C4 PHOTOSYNTHESIS VERSUS THE DUAL-CELL (KRANZ)PARADIGM, Gerald E. Edwards, Vincent R. Franceschi,and Elena V. Voznesenskaya 173
MOLECULAR MECHANISM OF GIBBERELLIN SIGNALING IN PLANTS,Tai-ping Sun and Frank Gubler 197
PHYTOESTROGENS, Richard A. Dixon 225
DECODING Ca2+ SIGNALS THROUGH PLANT PROTEIN KINASES,Jeffrey F. Harper, Ghislain Breton, and Alice Harmon 263
PLASTID TRANSFORMATION IN HIGHER PLANTS, Pal Maliga 289
SYMBIOSES OF GRASSES WITH SEEDBORNE FUNGAL ENDOPHYTES,Christopher L. Schardl, Adrian Leuchtmann, Martin J. Spiering 315
TRANSPORT MECHANISMS FOR ORGANIC FORMS OF CARBON ANDNITROGEN BETWEEN SOURCE AND SINK, Sylvie Lalonde,Daniel Wipf, and Wolf B. Frommer 341
vii
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.

P1: FRK
March 24, 2004 1:1 Annual Reviews AR213-FM
viii CONTENTS
REACTIVE OXYGEN SPECIES: METABOLISM, OXIDATIVE STRESS,AND SIGNAL TRANSDUCTION, Klaus Apel and Heribert Hirt 373
THE GENERATION OF Ca2+ SIGNALS IN PLANTS,Alistair M. Hetherington and Colin Brownlee 401
BIOSYNTHESIS AND ACCUMULATION OF STEROLS, Pierre Benveniste 429
HOW DO CROP PLANTS TOLERATE ACID SOILS? MECHANISMS OFALUMINUM TOLERANCE AND PHOSPHOROUS EFFICIENCY,Leon V. Kochian, Owen A. Hoekenga, and Miguel A. Pineros 459
VIGS VECTORS FOR GENE SLIENCING: MANY TARGETS,MANY TOOLS, Dominique Robertson 495
GENETIC REGULATION OF TIME TO FLOWER IN ARABIDOPSIS THALIANA,Yoshibumi Komeda 521
VISUALIZING CHROMOSOME STRUCTURE/ORGANIZATION,Eric Lam, Naohiro Kato, and Koichi Watanabe 537
THE UBIQUITIN 26S PROTEASOME PROTEOLYTIC PATHWAY,Jan Smalle and Richard D. Vierstra 555
RISING ATMOSPHERIC CARBON DIOXIDE: PLANTS FACE THE FUTURE,Stephen P. Long, Elizabeth A. Ainsworth, Alistair Rogers,and Donald R. Ort 591
INDEXESSubject Index 629Cumulative Index of Contributing Authors, Volumes 45–55 661Cumulative Index of Chapter Titles, Volumes 45–55 666
ERRATAAn online log of corrections to Annual Review of Plant Biologychapters may be found at http://plant.annualreviews.org/
Ann
u. R
ev. P
lant
Bio
l. 20
04.5
5:49
5-51
9. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsid
ad A
uton
oma
de Q
uere
taro
on
01/1
5/14
. For
per
sona
l use
onl
y.




















