
Variation in Paleoindian Fauna Use on the Great Plains and Rocky Mountains of North America.
20
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Variation in Paleoindian Fauna Use on the Great Plains and Rocky Mountains of North America.

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Quaternary International 191 (2008) 34–52
Variation in Paleoindian fauna use on the Great Plains and RockyMountains of North America
Matthew E. Hill Jr.�
Department of Anthropology, University of Iowa, 114 MacBride Hall, Iowa City, IA 52242, USA
Available online 10 October 2007
Abstract
Drawing on data from 69 zooarchaeological assemblages from Paleoindian sites in the Great Plains and Rocky Mountains, this paper
explores factors underlying changes in prehistoric diet breadth. Results indicate that different site types provide different perspectives on
Paleoindian faunal use. Kill assemblages can only inform on the exploitation of large game, while the full variety of prey used by foragers
is represented at camps. In contrast to prior claims, however, the inclusion of kill site assemblages in zooarchaeological analyses does not
appear to bias our perspective of Paleoindian subsistence. Rather, there is strong indication that Paleoindian foragers targeted different
types of prey depending on which habitat they occupied. In more-diverse environments of alluvial valleys and foothill/mountains,
foragers exploited large game whenever possible, but they also targeted a suite of smaller prey items. In more uniform grassland
environments, subsistence strategies focused almost exclusively on hunting large prey, such as bison and mammoth. These results also
provide limited support for prior studies that argue for intensified resource use and increased diet breadth in response to early Holocene
environmental changes, at least in certain settings.
r 2007 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
The use of animal resources by Paleoindian hunter-gatherers has been a key research issue for archaeologistsfor nearly 80 years. In the last few decades the debate overthis issue has revolved around what I consider a falsedichotomy: specialized big game hunting versus broad-spectrum foraging (e.g., Amick, 1994; Waguespack andSurovell, 2003; Cannon and Meltzer, 2004). Hunting large-bodied animals is argued to play a primary role inPaleoindian subsistence (Todd, 1991; Bement, 2003;Waguespack, 2003, 2007; Waguespack and Surovell,2003), technology (Frison, 1991), mobility and land usestrategies (Kelly and Todd, 1988), and the success of theinitial settlement of the Americas (Surovell, 2000; Wague-spack and Surovell, 2003). Some even suggest that humanhunting was a primary cause for the extinction of 35 generaof Rancholabrean fauna (Alroy, 1998, 2001; Martin andSteadman, 1999; Haynes, 2002a, b; Martin, 2005; Surovellet al., 2005).
This study does not wade into the murky pool that is thespecialist-generalist debate. Instead, I draw attention toimportant epistemological issues related to Paleoindianfaunal use, as well as to the need to quantitatively analyzezooarchaeological patterning to obtain a more completeunderstanding of prehistoric life ways and land use.Drawing on data from 69 Paleoindian assemblages in the
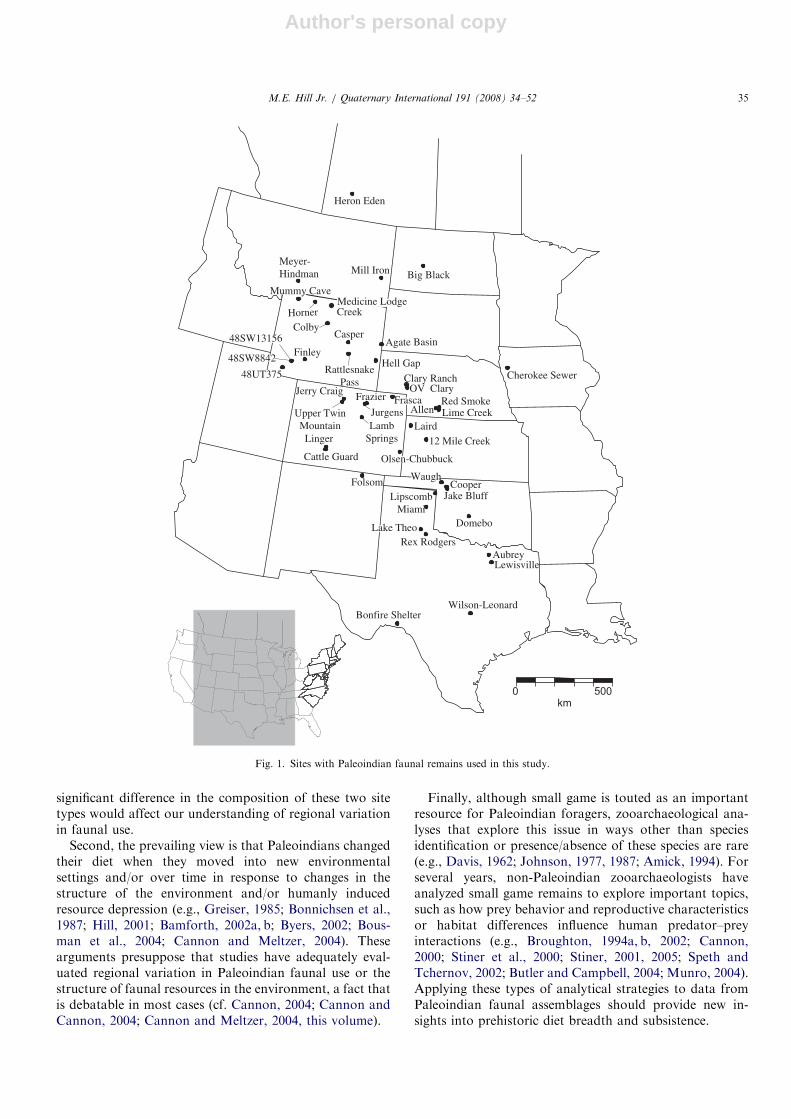
Great Plains and Rocky Mountains (Fig. 1 and Table 1),this paper explores several issues, including the biasedconsideration of kill sites, long-term changes in dietbreadth, and exploitation of small game. First, archae-ologists commonly claim that kill assemblages bias ourinferences about Paleoindian subsistence (Meltzer, 1988,1993, 1995; Gero, 1993; Cannon and Meltzer, 2004;Kornfeld and Larson, this volume). Specifically, it isclaimed that by including data from kill sites thecontribution of hunting of large-game taxa to thePaleoindian diet is over estimated. However, this claim israrely quantitatively evaluated so it is not often clear whateffect this ‘‘bias’’ actually has. For example, Paleoindianarchaeologists do not often systematically compare thecomposition of the zooarchaeological record from kill sitesto those from camps. Further it is not clear how even a
ARTICLE IN PRESS
1040-6182/$ - see front matter r 2007 Elsevier Ltd and INQUA. All rights reserved.
doi:10.1016/j.quaint.2007.10.004
�Tel.: +1 319 335 2890; fax: +1 319 335 0653.
E-mail address: [email protected]
Author's personal copy
significant difference in the composition of these two sitetypes would affect our understanding of regional variationin faunal use.
Second, the prevailing view is that Paleoindians changedtheir diet when they moved into new environmentalsettings and/or over time in response to changes in thestructure of the environment and/or humanly inducedresource depression (e.g., Greiser, 1985; Bonnichsen et al.,1987; Hill, 2001; Bamforth, 2002a, b; Byers, 2002; Bous-man et al., 2004; Cannon and Meltzer, 2004). Thesearguments presuppose that studies have adequately eval-uated regional variation in Paleoindian faunal use or thestructure of faunal resources in the environment, a fact thatis debatable in most cases (cf. Cannon, 2004; Cannon andCannon, 2004; Cannon and Meltzer, 2004, this volume).
Finally, although small game is touted as an importantresource for Paleoindian foragers, zooarchaeological ana-lyses that explore this issue in ways other than speciesidentification or presence/absence of these species are rare(e.g., Davis, 1962; Johnson, 1977, 1987; Amick, 1994). Forseveral years, non-Paleoindian zooarchaeologists haveanalyzed small game remains to explore important topics,such as how prey behavior and reproductive characteristicsor habitat differences influence human predator–preyinteractions (e.g., Broughton, 1994a, b, 2002; Cannon,2000; Stiner et al., 2000; Stiner, 2001, 2005; Speth andTchernov, 2002; Butler and Campbell, 2004; Munro, 2004).Applying these types of analytical strategies to data fromPaleoindian faunal assemblages should provide new in-sights into prehistoric diet breadth and subsistence.
ARTICLE IN PRESS
0 500
Big Black
Cattle Guard
48UT375
FinleyHell Gap
Mummy Cave
Red Smoke
Meyer-Hindman
Bonfire Shelter
Heron Eden
Olsen-Chubbuck
Frazier
Domebo
CooperLipscomb
Miami
Waugh
Clary Ranch
Medicine LodgeCreek
12 Mile Creek
Agate Basin
AubreyLewisville
Jerry Craig
Casper
Laird
Colby
Folsom
Horner
Lake Theo
LambSprings
Wilson-Leonard
Mill Iron
Frasca
RattlesnakePass
48SW13156
Allen Lime CreekUpper TwinMountain
Jurgens
Linger
Cherokee Sewer
Jake Bluff
OV Clary
48SW8842
Rex Rodgers
km
Fig. 1. Sites with Paleoindian faunal remains used in this study.
M.E. Hill Jr. / Quaternary International 191 (2008) 34–52 35

Author's personal copy
1.1. History of research
Almost immediately following the important discoveriesat the Folsom type site, archaeologists began askingquestions about what prey Paleoindian hunter-gatherersdid or did not use (e.g., Barbour and Schultz, 1932;Roberts, 1935). For most of the twentieth centuryPaleoindians were portrayed as specialized hunters ofbison and mammoth, based on the numerous occasionswhere remains of these species were recovered in associa-tion with well-made fluted or lanceolate projectile points
(Sellards, 1952; Wormington, 1957). When evidence for useof small game was recovered from Paleoindian sites, it wasoften assumed that these resources were simply supple-ments to a diet otherwise rich in big game (Holder andWilke, 1949; Davis, 1953, 1962).Recent excavations, utilizing careful excavation techni-
ques and fine-screening of back dirt, largely uphold thetraditional view of the primacy of bison and mammothremains in Paleoindian zooarchaeological assemblages.This is true for both kill sites (e.g., Hofman et al., 1995;Collins et al., 1997; Hill and Hofman, 1997; Hofman andBlackmar, 1997; Bement, 1999; Kornfeld et al., 1999;Hofman and Todd, 2001; Bement and Carter, 2003;Mandel and Hofman, 2003) and camp localities (e.g.,Jodry, 1999; Williams, 2000; Ferring, 2001; Hill et al., 2006,this volume; cf., Mallouf and Mandel, 1997; Baker, 1998;Smith et al., 2003; Byers et al., 2005).Despite these findings, there is a growing number of
archaeologists who argue that the contribution of large-game hunting to Paleoindian subsistence is exaggerated(e.g., Johnson, 1977; Bonnichsen et al., 1987; Amick, 1994;Cannon and Meltzer, 2004; Kornfeld and Larson, thisvolume). According to these critics, empirical evidence andtheoretical concepts raise questions about whether or not ahigh reliance on big game hunting was even possible forhunter-gatherers. For example, these researchers note thatseveral regions in North America, such as the northeastand Great Basin, have produced little direct evidence forthe exploitation of these types of fauna (Meltzer, 1988;Cannon and Meltzer, 2004; Cannon and Meltzer, thisvolume). Findings from behavioral ecology models andtheories concerning population density in large-bodiedmammals also question whether or not large game wouldhave been abundant enough to support even modesthunter-gatherer populations (Geist, 1998, 1999; Byers andUgan, 2005). Finally, some researchers suggest that thecurrent perspective of Paleoindians as specialized big gamehunters is the result of biases introduced through thepreferential preservation and archaeological discovery ofmegafauna bone beds over accumulations of small-bodiedanimals (Grayson and Meltzer, 2002).In summary, despite progress over the last decade in
excavation of newly discovered sites (e.g., Hill andHofman, 1997; Bement, 1999; Ferring, 2001; Bement andCarter, 2003; Hill et al., 2006) and the reanalysis ofimportant curated zooarchaeological assemblages (e.g.,Jones, 1999; Hill, 2001; Borresen, 2002; Byers, 2002; Hill,2002a, b, 2006; Hill and Hill, 2002; Byerly et al., 2005;Meltzer, 2006), Paleoindian faunal studies have come to animpasse. The research community has become deeplydivided over how the diet of Paleoindians should beportrayed. As a result, there has been little progress onfinding either a consensus on this issue or generating abetter understanding of the factors underlying the variationobserved in the Paleoindian zooarchaeological record.The current paper tries to move beyond this impasse. By
exploring geographic and diachronic variation in the
ARTICLE IN PRESS
Table 1
Site with Paleoindian faunal remains (see Fig. 1 for locations)
Site State Reference
12 Mile Creek KS Hill (2002b)
48SW8842 WY Byers et al. (2005)
48SW13156 WY Byers et al. (2005)
48UT375 WY Smith et al. (2003)
Agate Basin WY Hill (2001)
Allen NE Bamforth et al. (2005)
Aubrey TX Ferring (2001)
Big Black ND Williams (2000)
Bonfire Shelter TX Byerly et al. (2005)
Casper WY Frison (1974)
Cattle Guard CO Jodry (1999)
Cherokee Sewer IA Whittaker (1997)
Clary Ranch NE Hill et al. (2006)
Colby WY Frison and Todd (1986)
Cooper OK Bement (1999)
Domebo OK Leonhardy (1966)
Finley WY Hill (2007)
Folsom NM Meltzer (2006)
Frasca CO Fulgham and Stanford (1982)
Frazier CO Borresen (2002)
Hell Gap WY Byers (2002); Knell et al. (2002)
Heron Eden Saskatchewan Corbeil (1995)
Horner WY Frison and Todd (1987)
Jake Bluff OK Bement and Carter (2003)
Jerry Craig CO Kornfeld and Frison (2000)
Jurgens CO Wheat (1979)
Laird KS Hofman and Blackmar, 1997
Lake Theo TX Baxevanis (1997)
Lamb Spring CO McCartney (1983)
Lewisville TX Winkler (1982)
Lime Creek NE Jones (1999)
Linger CO Jodry (1999)
Lipscomb TX Todd et al. (1992)
Medicine Lodge Creek WY Walker (1975)
Miami TX Sellards (1938)
Mill Iron WY Frison (1996)
Mummy Cave WY Hughes (2003)
Myers–Hindman MT Cannon and Cannon (2004)
Olsen–Chubbuck CO Wheat (1972)
OV Clary NE Hill et al. (2006)
Rattlesnake Pass WY Smith and McNees (1990)
Red Smoke NE Hill (2007)
Rex Rodgers TX Hill (2006)
Upper Twin Mountain CO Kornfeld et al. (1999)
Waugh OK Hill and Hofman (1997)
Wilson–Leonard TX Baker (1998)
M.E. Hill Jr. / Quaternary International 191 (2008) 34–5236

Author's personal copy
human predation of different-sized game it may be possibleto understand how Paleoindian occupations in differenthabitats influenced diet breadth and resource exploitationpatterns. Specifically, I investigate the correspondencebetween the types of environmental settings occupied byPaleoindian foragers (grasslands, woodlands, riverinevalleys) and the predation strategies of those foragers(i.e., reliance on collecting small game versus hunting largegame) (see Hill, 2007).
2. Foraging theory and changes in diet breadth
This paper uses models developed from human beha-vioral ecology (e.g., Charnov, 1976; Orians and Pearson,1979; Winterhalder, 2001) to generate a series of a priori
testable hypotheses concerning how prehistoric foragersrespond to diachronic (temporal) and synchronic (regional)changes in prey availability.
Based on results from prey-choice models it is antici-pated that preferred food items will be pursued wheneverthey are encountered, and that dietary expansion isexpected when encounter rates with preferred prey typesdecline (Emlen, 1966; MacArthur and Pianka, 1966;Charnov, 1976; Heller, 1980). Therefore, when the supplyof high-ranked prey is abundant, a forager’s diet shouldnarrow to include preferred types in quantities dispropor-tionate to their environmental abundance. Conversely,when the supply of preferred prey declines, foragers’choices become less selective and more common andlower-quality foods will be added to the diet.
Previous studies, however, demonstrate that classifica-tion systems of prey rank can vary significantly. Tradi-tionally, optimal foraging theory assumes that foragers willprefer the prey with the highest post-encounter return rate
(Emlen, 1966; MacArthur and Pianka, 1966; Schoener,1971). As Kuhn and Stiner’s (2001) recent review of the netyields (kJ/h) for various food classes shows, body size,because of its strong correlation with post-encounter returnrate, is an appropriate proxy estimate of prey rank in mostcases (also Stephens and Krebs, 1986; Simms, 1987; Szuterand Bayham, 1989; Broughton and Grayson, 1993;Broughton, 1994a; Ugan, 2005). Based on this system itis assumed that foragers preferentially target the largestprey available and increase their use of smaller prey asencounters with preferred-sized prey decline.
However, other measures, such as protein content, fatvalue, nutrients, or desire for prestige, can also be used tocategorize the relative value of prey items (e.g., Hawkes etal., 1982; Belovsky, 1988; Kelly, 1995; Hockett and Haws,2003). Stiner and colleagues (Stiner et al., 2000; Stiner,2001, 2005; Stiner and Munro, 2002), for example, suggestthat prey classification systems relying solely on body sizepotentially overlook significant differences in handlingcosts for animals of comparable size. This is because aforager’s harvesting strategies must respond to differentlocomotor habits and predator avoidance tactics for preyof comparable value. Stiner (2001, 2005) suggests that
relatively slow moving or immobile prey that are easy toharvest, such as tortoises and shellfish will be of higherrank than comparably sized prey that are quick, agile, and/or below ground dwelling (e.g., hares, partridges, and fish).Here prey speed and predator avoidance strategy is used asa proxy of capture costs.Resource availability and quality is heterogeneous (i.e.,
patchy) both across space and through time and, as aresult, prehistoric foragers must decide which patch tooccupy, how long to use it, and which resources to exploitwhile in that patch (Charnov, 1976; Wien, 1976; McNa-mara, 1982). Models developed by Charnov (1976) andOrians and Pearson (1979) suggest foragers will preferen-tially utilize patches rich in high-return resources over onescontaining primarily low-return resources. A forager isanticipated to remain in a patch as long as the returnsexceed the costs associated with traveling and exploiting anew patch (Nagaoka, 2002, 2005).However, resource overexploitation by foragers, either
through continuous use of a patch or increased number offoragers using the same patch, will eventually result in areduction of the quality of that patch. Broughton (2002, p.63) suggests that in response to the depression of localresources quality, foragers will (1) more intensively uselower-ranked prey in those patches, (2) devote greatereffort to travel to distant, less-depleted patches, or (3) both(also Cannon, 2000, 2003). Regardless of which responseforagers choose, they invariably sacrifice a substantialdegree of foraging efficiency (Butler and Campbell, 2004).
2.1. Paleoenvironment, prey abundance, and hunting
pressure
As the discussion in Section 2 highlights, understandinghow the abundance of high-ranked prey change isimportant for making inferences about Paleoindian faunalexploitation patterns. As I have proposed elsewhere (Hill,2007), the reorganization of terrestrial biotic communitiesat the Pleistocene–Holocene transition and the concurrentchanges in human population demography likely had astrong influence on the abundance of fauna in the GreatPlains and Rocky Mountains (Graham and Lundelius,1984; FAUNMAP Working Group, 1994).At the end of the Pleistocene, much of the central and
western North America was a broad expanse of grasslandand parkland environments populated with a multitude ofmegafaunal species (Kurten and Anderson, 1980; Lunde-lius et al., 1983; Graham and Mead, 1987; Grayson, 1993).Due to significant overlap in feeding strategies and likelyhigh seasonal mortality, the absolute number of anyparticular species was fairly low (Guthrie, 1984; Geist,1998, 1999; Feranec and MacFadden, 2000; Martin andWilliams, 2007). At the same time, considering the sparsepopulations of human hunters on the landscape, theirimpact on game animals was likely limited (Wobst, 1974;Fiedel, 2004; cf. Martin, 2005). Therefore, prior to the endof the Pleistocene, while foragers could expect to encounter
ARTICLE IN PRESSM.E. Hill Jr. / Quaternary International 191 (2008) 34–52 37

Author's personal copy
a wide diversity of megafauna taxa across most habitats,the absolute abundance of this prey class was such that itwas unlikely that humans could exclude lower-ranked preyitems from their diet (Byers and Ugan, 2005).
The close of the Pleistocene and the extinction of mostmegafauna species wrought dramatic alterations to thelandscape (Lundelius et al., 1983; Owens-Smith, 1987;Grayson and Meltzer, 2002, 2003; Cannon, 2004; cf.Martin and Williams, 2007). However, these changes werenot necessarily detrimental for human hunters. While bisonwas one of the few very large land mammal species (e.g.,moose, and muskox) left in North America, released frommuch of their prior browsing competition and mostpredation pressure from large carnivores, populationsunderwent a rapid increase, albeit within a more limitedgeographic area, mostly within grasslands of the GreatPlains and adjacent areas (MacDonald, 1981; Guthrie,1984; FAUNMAP Working Group, 1994). This periodalso witnessed an expansion in populations of medium-sized artiodactyls such as deer, pronghorn, and elk as theyfilled niches left open by the extinction of megafauna(Martin and Williams, 2007).
Because human population density was still likely quitelow during this period and bison population density wasincreasing in certain regions, especially the Great Plains,Paleoindian foragers likely found hunting large prey quiterewarding (Amick, 1996; MacDonald, 1998; Hofman,1999). Certainly foragers would have found themselves inareas, such as the American Southwest and Great Basin,which were previously rich in bison, mammoths, andhorses, but were now lacking in large game. In addition, itwas likely that nowhere were bison so abundant that smalland medium game could be totally ignored as food sources.
As the Holocene progressed, increases in mean tempera-ture and the contribution of monsoonal precipitationacross the mid-continent of North America resulted inimportant shifts in sediment deposition patterns (Holliday,1997; Olson and Porter, 2002), the composition of bioticcommunities (Davis et al., 1986; Jodry, 1999; Baker et al.,2000), and stream and pond hydrology (Neck, 1995;Yansa, 1998; Leonard and Reasoner, 1999) that wouldhave had important affects on the abundance of largefauna. These environmental changes were, however, slowto appear and their expressions varied across the land-scape. By approximately 9500–9000 rcybp bison popula-tions appear to have decreased to the point where they hadan effect on human subsistence patterns at least in portionsof the Rocky Mountains (e.g., MacDonald, 1981; Hughes,2003; Byers et al., 2005). Environmental conditions withinthe Great Plains during the Paleoindian period do notappear to have degraded to the point to discourage at leastperiodic mass killing of bison, as evidenced at sites such asHeron Eden, Horner, and Finley (Frison and Todd, 1987;Corbeil, 1995).
It was likely only towards the end of the Paleoindianperiod that foragers, especially in mountains settings,turned to intensively exploiting lower ranked, lower risk
resources (e.g., plants and small game). During the latePaleoindian period there is evidence for increased occupa-tional intensity and possibly the beginnings of regionaliza-tion of foraging populations, especially in alluvial valleysand mountain settings (e.g., Husted, 1969; Frison, 1992;Bamforth et al., 2005; LaBelle, 2005). The combined effectsof increased human population density and decreasedmobility and environmental degradation likely would haveresulted in a depression to the abundance and structure ofthe faunal resources on which these foragers relied.
3. Methods for measuring diet breadth and prey rank
Diet breadth can be measured as total number of fooditems in the diet and/or greater proportional evennessamong high and low ranked prey items in the diet. Thisstudy utilizes two different richness and evenness measuresto analyze Paleoindian subsistence.Prey rank is defined using two different classification
systems. The first system groups prey according to packagesize following previous approaches (Bayham, 1979; Szuterand Bayham, 1989; Broughton, 1994a) and utilizespreviously established body size categories (i.e., Klein,1976; Brain, 1981): size class 1 (live body mass o22 kg),size class 2 (22–113 kg), size class 3 (113–340 kg), and sizeclass 4 (4340 kg). The number of identifiable specimens(NISP) from each body size class is summed to calculatethe proportional representation of three prey body sizeabundance indices (AI): large-fauna index (Class 4),medium-fauna index (Classes 3 and 2) and small faunaindex (Class 1).The AI values are calculated by combining NISP counts
for all taxa within each body size category and thencomparing the contribution of each specific body size classto the entire faunal assemblage. For example, the calcula-tion of large-fauna AI follows the equation: large-faunaAI ¼
Psize class 4 NISP/(
Psize class 1 NISP+
Psize
class 2 NISP+P
size class 3 NISP+P
size class 4 NISP).This resulting index generates values ranging between 0and 1.0, with high values indicating greater representationof fauna of this size category.The second classification system is a modified version of
a scheme developed by Stiner (2001, 2005) and Stiner et al.(2000) (see also Wolverton, 2005), and groups the mostabundant small- and medium-bodied prey species intothree categories according to body size, predator avoidancestrategy, and speed: (1) medium-sized ungulates, (2) smallslow game, and (3) small difficult-to-catch game (Table 2).The goal is to organize medium and small prey based onthe difficulty or risk foragers face in harvesting these preyitems. A few small- and medium-sized taxa utilized byPaleoindians were not included in this classification systembecause they did not fit neatly into just one prey typecategory; none of the ignored taxa were well represented,and excluding them does not influence the results presentedhere.
ARTICLE IN PRESSM.E. Hill Jr. / Quaternary International 191 (2008) 34–5238

Author's personal copy
The Reciprocal of Simpson’s Index (RSI) is used as ameasure of assemblage evenness. This index is calculatedusing the equation RSI ¼ 1/
P(pi)2, where p represents the
proportion of each prey type for array i (Stiner and Munro,2002; adapted from Simpson, 1949; Levins, 1968). As onlythree potential prey classes are used in this study, the RSIranges from 1 (least even) to 3 (most even).
3.1. The study sample
Zooarchaeological data from the 69 Paleoindian assem-blages forms the basis of this study (Table 3). These dataare drawn from both published reports and the author’sown analyses. Assemblages were selected from majorhabitat settings of the Great Plains and Rocky Mountains(alluvial valley n ¼ 16; foothill/mountain n ¼ 30; plains/rolling hills n ¼ 23) and together span the period fromapproximately 11,500 to 7200 rcybp.1 Multifunction short-term camps (n ¼ 40) are slightly better represented in thissample than special-use kill/carcass-processing locales(n ¼ 29).2
Preservation and recovery quality were generally highfor all assemblages. Systematic matrix screening waspracticed at more than 84% (n ¼ 58) of the assemblages,although the proportion of unscreened components wasbiased towards alluvial (n ¼ 6; 38%) and plains/rollinghills (n ¼ 4; 17%) settings. Only a single assemblage (3%)from foothill/mountain settings was not screened. Priorwork demonstrates that screening treatment does notsignificantly influence the analyses of these zooarchaeolo-gical assemblages (Hill, 2007).A total of 130,365 specimens (NISP) is present in this
study, of which more than 113,000 specimens (87%) wereidentified to genus or finer taxonomic distinction. Follow-ing the procedure outlined by Grayson and Meltzer (2002)and Cannon and Meltzer (2004, pp. 1959–1960) onlyfaunal remains showing unequivocal evidence for humanbutchery and consumption were included in this study.3
4. Results
As recognized previously (Amick, 1994; Waguespackand Surovell, 2003; Hemmings, 2004; Hill, 2007), althoughmore than 31 fauna taxa are represented in Paleoindianfaunal assemblages, only a few species account for the vastmajority of faunal remains recovered (Table 4). The sixmost abundant taxa (i.e., bison, pronghorn, hares/rabbits,turtle, mammoth, and bighorn sheep) account for over99% (n ¼ 112,571) of identifiable remains used in thisstudy. That a small suite of fauna dominates Paleoindianfaunal assemblages is partially explained by the abundanceof multi-animal bison kills and processing sites included inthe study sample. However, even if kill sites are notconsidered, Paleoindian assemblages are still dominated bysame set of taxa (Hill, 2007). Although a broad spectrum ofprey entered their diet, Paleoindian foragers primarily
ARTICLE IN PRESS
Table 2
Summary of life history, predator defense strategy, and capture requirements for common medium and small prey in Paleoindian assemblages
Prey classificationa Predator defense strategy Reproductive
productivityaCapture
techniques
Taxa
Medium ungulates Rapid running, freezing, charging Slow Hunt/trap Deer, bighorn sheep, pronghorn
Small slow Freezing, hiding, armor Slow Gather Turtle, mussels
Small difficult to
catch
Hiding, running, subterranean escape,
nocturnal activity,
Fast Hunt/trap Birds, lagomorph, rodents, prairie dog,
beaver, fish
aClassification and productivity system used in this study is adapted from Stiner (2001, 2005) and Stiner et al. (2000).
1The habitat settings used in this study were chosen because site
occupants in these areas would have differential access to a suite of
important resources, as well as unique constraints (Kay, 1998; Kornfeld
and Osborn, 2003). The foothill/mountain setting includes localities
situated in (1) true alpine setting, (2) high altitude intermountain grassland
valleys (e.g., Middle Park and San Luis Valley), or (3) foothill
environments immediately adjacent to true mountain slopes. Plains/
rolling hills settings include both the High Plains proper and adjacent
grassland environments (e.g., Edwards Plateau, Rolling hills of Oklaho-
ma). Alluvial valley habitats are restricted to very large, permanent river
systems, where sites were located directly within a major drainage valley
(or at least in a former floodplain terrace) in order to be included in this
group.2Site function definitions follow those of Sellet (1999) and Wheat (1978,
1979) (see Judge, 1973; LaBelle, 2005), and are based primarily on the
characteristics of the chipped stone assemblage at each locality and
secondarily on non-faunal characteristics concerning site organization and
structure. A component was defined as a kill/processing locale if it was
associated with a small chipped stone assemblage with a low diversity of
tool types. Weaponry, butchery, and hide working tools dominate kill/
processing assemblages, and chipped stone debitage, if present, consisted
mostly of tool resharpening flakes and utilized flakes. Multi-function
camps are characterized by large lithic assemblages with numerous non-
weaponry tools, a high quantity of debitage, and preforms in various
stages of preparation. A low proportion of projectile points characterize
the stone-tool assemblages at camps. When projectile points are present at
camps, they are commonly broken and often present only in the form
point bases. Kill/processing localities were sometimes associated with one
(footnote continued)
or more physical feature, such as a steep-sided arroyo, that presumably
was used to trap animals during a kill. Camping localities often lack such
features, but instead were associated with cooking/heating, storage, or
habitation features.3The criteria for determine whether a taxon was used and consumed at a
particular site include: (1) presence of unambiguous butchery marks (e.g.,
cut marks or impact cones); (2) evidence of intentional burning or
cooking; (3) unquestionable and direct spatial association of projectile
points and butchery tools with faunal remains; and (4) association of
faunal remains with constructed cultural features. This study excluded any
remains suggested to be of non-cultural origin by the original analyst.
M.E. Hill Jr. / Quaternary International 191 (2008) 34–52 39

Author's personal copyARTIC
LEIN
PRES
S
Table 3
Summary information for 69 Paleoindian zooarchaeological assemblages used in this study
Site Screena Site
functionb
14C Agec NISP Large AI
(Class 4)
Medium
AI (Class
3/2)
Small AI
(Class 1)
%Medium
ungulate
%Small
slow
%Small
difficult
Alluvial valley sites
Allen—Occupation 2 N C 8500 279 0.06 0.16 0.77 0.16 0.03 0.81
Allen—Intermediate zone N C 9500 473 0.15 0.24 0.60 0.28 0.02 0.70
Allen—Occupation 1 N C 10,493 1763 0.47 0.07 0.45 0.12 0.10 0.78
Aubrey—Camp A and B; pond C/E1; red sand S Y C 11,588 1523 0.34 0.01 0.64 0.03 0.74 0.24
Big Black Y C 10,500C 836 0.41 0.02 0.57 0.00 0.00 1.00
Cherokee sewer—Level IIIa Y K 7183 144 1.00 0.00 0.00 — — —
Frazier N K 9650 1188 0.99 0.00 0.01 0.25 0.00 0.75
Jurgens—Area 1 Y C 9070c 2527 0.97 0.01 0.02 0.37 0.06 0.58
Jurgens—Area 2 Y C 9070c 155 0.61 0.37 0.03 0.93 0.00 0.07
Jurgens—Area 3 Y K 9070 2993 0.99 0.01 0.00 0.74 0.00 0.26
Lewisville Y C 11,000c 466 0.05 0.04 0.92 0.04 0.04 0.92
Lime creek—Zone I N C 9120 564 0.26 0.22 0.52 0.14 0.03 0.83
Red smoke—Strata 80–90 N C 8810 6165 0.55 0.35 0.10 0.60 0.07 0.33
Wilson Leonard—Units Isi, Isi/Icl, Igl/Isi, Isi/Isi-
C, Isi/Icl/Isi-C
Y C 11,200 311 0.12 0.74 0.14 0.00 0.11 0.89
Wilson Leonard—Unit II Y C 9294 606 0.01 0.66 0.33 0.01 0.10 0.89
Wilson Leonard—Upper unit I Y C 9663 430 0.00 0.75 0.24 0.06 0.29 0.65
Foothills/mountain sites
48SW8842 Y C 8490 147 0.00 0.00 1.00 0.00 0.00 1.00
48SW13156 Y C 8725 172 0.00 0.02 0.98 0.00 0.00 1.00
48UT375—Level 1 Y C 8476 926 0.00 0.00 1.00 0.00 0.00 1.00
48UT375—Level 2 Y C 7890 3190 0.00 0.00 1.00 0.00 0.00 1.00
Cattle Guard Y C 10800 3517 1.00 0.00 0.00 — — —
Colby Y K 10961 463 1.00 0.00 0.00 — — —
Finley Y K 9009 8326 1.00 0.00 0.00 — — —
Folsom N K 10,450 3640 1.00 0.00 0.00 — — —
Hell Gap—Locality II (Agate Basin) Y C 8600 1318 0.99 0.01 0.00 1.00 0.00 0.00
Hell Gap—Locality V (Cody) Y C 8781 1374 0.99 0.00 0.01 0.75 0.00 0.25
Horner I Y C 8788 2157 0.99 0.01 0.00 0.81 0.19 0.00
Horner II Y K 9630 7097 1.00 0.00 0.00 1.00 0.00 0.00
Jerry Craig Y K 9390 326 1.00 0.00 0.00 — — —
Linger Y C 9000c 462 1.00 0.00 0.00 — — —
Medicine Lodge Creek—Deep Deer Faunule Y C 9700* 65 0.03 0.35 0.62 0.37 0.00 0.63
Medicine Lodge Creek—Rodent Level Faunule Y C 9590 3,880 0.00 0.01 0.99 0.01 0.00 0.99
Medicine Lodge Creek—Fire Pit Faunule Y C 8240 95 0.00 0.28 0.72 0.09 0.00 0.91
Medicine Lodge Creek—Pryor Stemmed Faunule Y C 8191 52 0.02 0.48 0.50 0.49 0.00 0.51
Mill Iron Y C 11,168 655 1.00 0.00 0.00 — — —
Mummy Cave—Layer 12 Y C 8100 179 0.00 0.91 0.09 0.86 0.00 0.14
Mummy Cave—Layer 11 Y C 8136 95 0.00 1.00 0.00 1.00 0.00 0.00
Mummy Cave—Layer 10 Y C 8200 145 0.00 0.94 0.06 0.93 0.00 0.07
Mummy Cave—Layer 9 Y C 8307 108 0.00 0.99 0.01 0.99 0.00 0.01
Mummy Cave—Layer 8 Y C 8455 85 0.00 0.98 0.02 0.96 0.00 0.04
Mummy Cave—Layer 6 Y C 8500 46 0.00 0.38 0.63 0.26 0.00 0.74
M.E
.H
illJ
r./
Qu
atern
ary
Intern
atio
na
l1
91
(2
00
8)
34
–5
240

Author's personal copyARTIC
LEIN
PRES
S
Mummy Cave—Layer 4 Y C 9244 33 0.00 0.52 0.48 0.36 0.00 0.64
Mummy Cave—Layer 1 Y C 9500 35 0.00 0.14 0.86 0.12 0.00 0.88
Myers–Hindman—Unit 1 Y C 9000 37 0.38 0.62 0.00 1.00 0.00 0.00
Rattlesnake Pass Y K 9860 424 0.99 0.01 0.00 0.80 0.00 0.20
Upper Twin mountain Y K 10,392 222 1.00 0.00 0.00 — — —
Plains/rolling hills sites
12 Mile Creek N K 10,503 871 1.00 0.00 0.00 — — —
Agate Basin—Area 2—AB Y K 10,430 3179 1.00 0.00 0.00 — — —
Agate Basin—Area 2—Folsom Y C 10,699 1330 0.78 0.22 0.00 1.00 0.00 0.00
Agate Basin—Area 3—Hell Gap Y K 10,445 986 1.00 0.00 0.00 — — —
Bonfire Shelter—Bone Bed 2 Y K 9871 2261 1.00 0.00 0.00 — — —
Casper Y K 10,025 7146 1.00 0.00 0.00 — — —
Clary Ranch Y K 9040 1920 0.96 0.00 0.04 0.00 0.22 0.78
Cooper—Lower Kill Y K 10,050 900 1.00 0.00 0.00 — — —
Cooper—Middle Kill Y K 10,040c 1200 1.00 0.00 0.00 — — —
Cooper—Upper Kill Y K 10,030c 1450 1.00 0.00 0.00 — — —
Domebo N K 11,091 75 1.00 0.00 0.00 — — —
Frasca—Area I Y K 8908 7526 1.00 0.00 0.00 — — —
Heron Eden Y K 9037 22,901 1.00 0.00 0.00 1.00 0.00 0.00
Jake Bluff Y K 10,762 167 1.00 0.00 0.00 — — —
Laird Y K 8495 322 1.00 0.00 0.00 — — —
Lake Theo-Folsom Y C 10,400c 6919 1.00 0.00 0.00 — — —
Lamb Spring Y K 9000c 579 1.00 0.00 0.00 — — —
Lipscomb Y K 10820 1202 1.00 0.00 0.00 — — —
Miami N K 11,000c 54 1.00 0.00 0.00 — — —
Olsen–Chubbuck N K 9395 5370 1.00 0.00 0.00 — — —
OV Clary—Middle Y C 9123 1033 0.97 0.00 0.03 0.00 0.39 0.61
Rex Rodgers Y K 9391 1886 1.00 0.00 0.00 — — —
Waugh Y C 10,393 1403 1.00 0.00 0.00 — — —
aY ¼ yes; N ¼ no.bC ¼ camp locality; K ¼ kill/processing locality.cEstimated age.
M.E
.H
illJ
r./
Qu
atern
ary
Intern
atio
na
l1
91
(2
00
8)
34
–5
241

Author's personal copy
relied on only a small number of prey for their subsistence(Cannon and Meltzer, this volume).
The following sections explore several dimensions of thePaleoindian zooarchaeological record. Section 4.1 consid-ers whether including kill assemblages in analyses influ-ences our perspective on the importance of megafauna
hunting to Paleoindian subsistence. Section 4.2 explorespatterning in the use of different-sized prey species overtime. Finally, Section 4.3 discusses evidence for resourceintensification during the Paleoindian period.
4.1. Large-game abundance and bias from kill sites
As noted in Section 4, bison remains from kill/processinglocalities represent a majority of the total fauna recoveredfrom Paleoindian assemblages. But does this fact mean thatkill site assemblages obscure the realities of Paleoindiansubsistence? Fig. 2 compares the distribution of large-faunaAI values for kill/processing localities to those from camps.A simple comparison of median AI values shows that biggame is much less abundant at camps (.1; n ¼ 40) than atkills (1.0; n ¼ 29), and non-parametric analysis indicatesthese differences to be statistically significant (Kolmogor-ov–Smirnov D ¼ 0.73; w2 ¼ 35.9; po0.0001), and unre-lated to sample size difference as measured by total NISPbetween the two site types (Kolmogorov–SmirnovD ¼ 0.29; w2 ¼ 5.51; po0.13).This result is not surprising. Mass-death assemblages,
such as those that make up the bulk of Paleoindian kills,are expected to contain primarily the remains from a singlespecies, whereas foragers are expected to transport avariety of prey types back to camp localities. However,this is not to suggest that bison remains are absent from allcamps or that camps from different habitats have the sameproportion of taxa represented. For example, the faunalassemblages from camps at Agate Basin-Area 2 (Folsom),Cattle Guard, Horner I-, Hell Gap-Localities II (AgateBasin) and V (Cody), Jurgen-Area 1, Linger, Lake Theo,Mill Iron, OV Clary, and Waugh consist almost exclusivelyof bison remains. In addition, large-fauna AI values for killassemblages are significantly higher than those from campsin alluvial (Kolmogorov–Smirnov D ¼ 1.0; w2 ¼ 9.80;p ¼ 0.02) and foothill/mountains settings (Kolmogorov–S-mirnov D ¼ 0.78; w2 ¼ 13.2; p ¼ 0.003) but not in plains/rolling hill settings (Kolmogorov–Smirnov D ¼ 0.15;w2 ¼ 0.28; p40.99) (see Table 3).
ARTICLE IN PRESS
Table 4
Assemblage ubiquity, total NISP, and NISP/site for identifiable fauna
reported from the sites listed in Table 1
Common name Number of
components (%)
NISP NISP/site
Bison 50 (83) 108,230 2164.6
Pronghorn 16 (27) 1323 82.7
Hares/rabbits 23 (38) 1310 57.0
Turtle 14 (23) 693 49.5
Mammoth 3 (5) 592 197.3
Bighorn sheep 12 (20) 423 35.3
Deer 23 (38) 284 12.3
Prairie dog 4 (7) 254 63.5
Bird 21 (35) 247 11.8
Beaver 11 (18) 189 17.2
Snakes 8 (13) 182 22.8
Fish 10 (17) 130 13
Canids 10 (17) 114 11.4
Mussels/mollusks 1 (2) 87 87
Amphibians 5 (8) 45 9
Badger/weasel 6 (10) 38 6.3
Elk 5 (8) 24 4.8
Bear 3 (5) 23 7.7
Horse 1 (2) 13 13
Marmot/woodchuck 4 (7) 11 2.8
Porcupine 3 (5) 7 2.3
Wolf 3 (5) 6 2
Muskrat 2 (3) 4 2
Raccoon 1 (2) 4 4
Fox 2 (3) 4 2
Lizards 2 (3) 3 1.5
Moose 1 (2) 2 2
Skunk 1 (2) 2 2
Coyote 1 (2) 2 2
Alligator 1 (2) 1 1
Lynx/bobcat 1 (2) 1 1
Total 114,248
0
0.2
0.4
0.6
0.8
1
Lar
ge F
auna
AI
Val
ue (
Size
Cla
ss 4
)
Campsn=40
Kill/processingn=29
Median
Outliers
75 percentile
25 Percentile
Outliers
Fig. 2. Box plot comparison of large fauna (size class 4) abundance index (AI) between specialized kill/processing localities and multifunction short-term
camps. Kolmogorov–Smirnow two-sample test (D ¼ 0.731; w2 ¼ 35.9; po.0001) reject null hypothesis that two samples have the same distributions.
M.E. Hill Jr. / Quaternary International 191 (2008) 34–5242

Author's personal copy
Based on these results, it might seem reasonableto assume that including assemblages from kills andprocessing sites into analyses distorts our view ofPaleoindian diet. However, as depicted in Fig. 3,non-parametric analyses indicate that the distributions oflarge-fauna AI values by habitat settings do not signifi-cantly differ between samples including both kills andcamps and samples restricted only to camps. Regardless ofwhether kills are included or not, the range of large-faunaAI values for foothill/mountain and alluvial valley sites iswide, but median large-fauna AI values remain consistentlylow. Plains/rolling hill assemblages, on the other hand,consist almost exclusively of remains of bison andmammoth.
These results suggest that kill sites are not ‘‘biasing’’our perspective of Paleoindian subsistence. Rather,foragers appear to be targeting different types ofprey depending on which habitat setting they occupy.In the more-diverse environments such as alluvialvalleys and foothill/mountains, foragers likely exploitedthe largest prey types whenever possible, but also targeteda wide range of smaller faunal resources as well. Incontrast, in more uniform grassland environmentsPaleoindians focused almost exclusive on hunting bisonand mammoth and appeared to make very little use ofsmall game.
4.2. Temporal trends
Based on the environmental history of the project areaoutlined in Section 2.2, I predict that the abundance ofbison (and mammoth) taken by Paleoindian huntersshould decrease through time as a result of increasingaridity and changes to the biotic communities during theearly Holocene, and possibly as a result of increasinghunting pressure. I previously discussed some importantregional differences in the extent to which foragersexploited prey of different body size through time (Hill,2007). This work showed that plains/rolling hills campsassemblages have very high proportion of large game forall periods. In alluvial assemblages large-fauna AI valuesdecrease through time, while the AI index for both mediumand small fauna increase through time, although none ofthese trends were statistically significant. In contrast,foothill/mountain assemblages show a long-term patternfor significant decreases in large-fauna AI values and acorresponding increase in the medium-fauna index values.Small-fauna index in foothill/mountain assemblagesshowed no significant temporal trend. As discussed in Hill(2007, p. 430), these results are not a function of the size ofthe faunal sample, as measured by NISP.While trends in AI values by body size are informative
for tracking how different-sized prey were used through
ARTICLE IN PRESS
Allu
vial
Val
ley
n=
13
Foot
hills
/Mou
ntai
n n
=23
Plai
ns/R
ollin
g H
ills
n=
4
n=16
n=30
n=23
0
0.2
0.4
0.6
0.8
1
0
0.2
0.4
0.6
0.8
1
Lar
ge F
auna
AI
Val
ue(S
ize
Cla
ss 4
)L
arge
Fau
na A
I V
alue
(Siz
e C
lass
4)
Median
Outliers
75 percentile
25 Percentile
Outliers
Fig. 3. Box plot comparison of large-fauna (size class 4) AI among alluvial valley, foothill/mountains, and plains/rolling hill (A) with both kill/processing
localities and multifunction short-term camps, and (B) multifunction short-term camps only. Kolmogorov–Smirnow two-sample tests do not reject the
null hypothesis that the distributions of large-fauna AI values for sites in alluvial valley (D ¼ 0.188; w2 ¼ 1.009; p40.99), foothill/mountain (D ¼ 0.183;
w2 ¼ 1.74; p ¼ 0.84), and plains/rolling hills (D ¼ 0.120; w2 ¼ 0.195; p40.99) differ between samples that included both kills and camps and samples that
included only camps.
M.E. Hill Jr. / Quaternary International 191 (2008) 34–52 43

Author's personal copy
time as shown in Hill (2007), it may ignore important shiftsin the overall composition of diets. As a result, I explorethe degree of evenness in the representation of different sizeprey through the use of the RSI calculated based on thethree body size categories (small, medium, and large). Onlycamp assemblages are considered here because, unlike kills,they provide data on a broad spectrum of prey items. Also,as the sample of plains/rolling hill camps is very small, onlyresults from alluvial valley and foothill/mountain campsare discussed.
Fig. 4 shows that foothill/mountain camp assemblagesgenerally yield low levels of evenness in diet breadth. Thetrend in the RSI was flat and not correlated with time(Spearman’s rho ¼ 0.02; p ¼ 0.94; n ¼ 23). Interestingly,there was a significant inverse correlation between the RSIand sample size (Spearman’s rho ¼ �0.69; po0.001;n ¼ 23). These results indicate that for most foothill/mountain assemblages a single body size class dominates.In early Paleoindian assemblages that prey is typicallybison, but later assemblages consist of either small ormedium fauna. In only a handful of foothill/mountainassemblages is there an even mixture of body sizes preyrepresented. Assemblages with a mixture of body sizesconsist of only a few specimens, so these results may notrepresentative of human predation practices because ofsampling biases.
These findings suggest that most foothill/mountain siteswere not residential camps to which a wide variety ofresources were transported. Instead, these assemblages arethe end result of collection focused on one or two preytypes. Assemblages dominated by medium-size ungulateslikely represent seasonal hunting camps and usually consist
of bighorn sheep and deer (e.g., Hughes, 2003). Assem-blages dominated by small game are likely the results ofshort-term residences where lagomorphs or various rodentswere important dietary resources (e.g., Walker, 1975).Evenness values from alluvial camp assemblages are
significantly higher than those from foothill/mountainenvironments (Kolmogorov–Smirnov two-sample testsD ¼ 0.609; w2 ¼ 12.309; p ¼ 0.004), and show a slight,although insignificant, trend for increasing values throughtime (Spearman’s rho ¼ 0.07, p ¼ 0.80; n ¼ 13). Foralluvial camps, the RSI values do not correlate withsample size (Spearman’s rho ¼ 0.31; p ¼ 0.29; n ¼ 13).Unlike in foothill/mountain settings, Paleoindian foragersoccupying alluvial habitats preyed on a relatively equalmixture of different-sized game animals. This implies thatalluvial camps were residential bases from which harvest ofa mixed-size fauna occurred. The poor temporal resolutionat most sites makes it difficult to determine whether or notthese different prey were collected concurrently or sequen-tially. The two stratified sites of Allen and Wilson–Leonardexhibit variability in evenness measurement through timewith no apparent temporal trend (Fig. 5).
4.3. Use of small- and medium-size game and evaluating
evidence for resource depression
Up to this point, the results of this study strongly suggestthat there are significant regional differences in Paleoindianprey exploitation patterns and, at least for alluvial valleysites, there is some evidence for dietary expansion throughtime. However, what factor(s) are driving these patterns(such as environmental differences or humanly-inducedresource depression) is not clear. Stiner (2001, p. 6993)suggests that shifts in foragers’ use of different body sizeclasses (which largely follow Linnean taxonomic cate-gories) may be explainable by climate-driven environmen-tal changes or geographic variations in biotic communities
ARTICLE IN PRESS
.75
1
1.25
1.5
1.75
2
2.25
2.5
2.75
Eve
ness
, Bod
y Si
ze
7000 8000 9000 10000 11000 12000
Radiocarbon (14C) Age
Foothills/MountainAlluvial Valley
Fig. 4. Evenness in the representation of three prey categories (large-,
medium-, and small-bodied fauna) using the Reciprocal of Simpson’s
Index (3 ¼ most even) versus radiocarbon age. Best-fit regression line
drawn through alluvial valley (r2 ¼ 0.02; F ¼ 0.189; df ¼ 1,11; p ¼ 0.67)
and foothills/mountains (r2 ¼ 0.001; F ¼ 0.017; df ¼ 1,21; p ¼ 0.89)
multifunction short-term camp assemblages separately. Reciprocal of
Simpson’s Index from alluvial camp assemblages are significantly higher
than those from foothill/mountains (Kolmogorov–Smirnow two-sample
D ¼ 0.609; w2 ¼ 12.309; p ¼ 0.004).
8000
8500
9000
9500
10000
10500
11000
11500
1.2 1.3 1.4 1.5 1.6 1.7 1.8 1.9 2
Wilson Leonard
Allen
Rad
ioca
rbon
(14
C)
Age
Evenness, Body Size
Fig. 5. Plot of evenness in the representation of three prey categories
based on body size (large-, medium-, and small-bodied fauna) using the
Reciprocal of Simpson’s Index (3 ¼ most even) by radiocarbon age for the
foothill/mountain multifunction short-term camps of Allen and Wilson
Leonard.
M.E. Hill Jr. / Quaternary International 191 (2008) 34–5244

Author's personal copy
(see also Wolverton, 2005). To track significant changes inthe intensity of human resource exploitation, she (Stiner etal., 1999, 2000; Stiner, 2003) suggests that the abundance ofsmall-bodied, low-turnover prey species can serve assymptoms of threshold effects in predator–prey systems.The greater representation of high- versus low-turnoverprey is suggestive of increasing harvesting pressure. It mustbe acknowledged that this approach does not preclude thepossibility that environmental factors are also influencingprey choice. To explore the trends are seen in the
Paleoindian record, the most common small- and med-ium-sized faunal species were regrouped based on body sizeand predator avoidance strategies (see Section 3).A triangle graph depicting the proportions of these
different prey categories reveals some interesting differ-ences in exploitation of small and medium prey betweenhabitat settings (Fig. 6). First, there are very few sites withevidence for use of small slow game. Except for the HornerI assemblage, foothill/mountain assemblages lack evidencefor predation of either turtles or mollusks. Alluvial
ARTICLE IN PRESS
Small Difficult
DominatedSmall Slow
Dominated
Alluvial Valley Sites
Foothill/Mountain Sites
100%Medium Ungulates
100%Small Difficult
100%Small Slow
(0% SmallSlow)
(0% SmallDifficult)
(0% MediumUngulates)
Big Black
Wilson Leonard-Early Paleo
Wilson Leonard-Unit II
Aubrey
LewisvilleWilson-Leonard-
Upper III
Allen-OC1
Lime CreekAllen-OC2Frazier
Allen- IZ
Jurgens-Area 1
Red Smoke-Strat. 80-90
Jurgens-Area 3
Jurgens-Area 2
48SW1315648SW884248UT375-L148UT375-L2
MLC-Rodent Level
MLC-Fire PitMummy Cave-L1
Mummy Cave-L6
Mummy Cave-L4
MLC-Deep Deer
MLC-Pryor Stemmed
Hell Gap-Loc.V
Rattlesnake Pass
Horner I
Mummy Cave-L12
Mummy Cave-L8Mummy Cave-L9
Hell Gap-Loc.IIHorner II
Mummy Cave-L11Myers-Hindman
Medium Ungulate
Dominated
Fig. 6. Triangular graph of proportional representation of three prey categories based on forager harvesting strategies and prey body size and predator
avoidance strategies (medium ungulate, small slow, small difficult) for alluvial valley and foothill/mountain multifunction short-term camps
(abbreviations: MLC ¼Medicine Lodge Creek). Data on large fauna abundance are not depicted on this graph.
M.E. Hill Jr. / Quaternary International 191 (2008) 34–52 45

Author's personal copy
assemblages, on the other hand, generally show at least aminimal presence of these prey types. Allen-OC1, RedSmoke, and several levels at Wilson–Leonard showmoderate use of these types of prey, but small, slow gameonly dominate the zooarchaeological assemblage at Au-brey. The paucity of these prey from foothill/mountainsites likely relates to the fact that both turtle and molluskspopulations preferentially inhabit lower altitude riparianenvironments.
Second, a paucity of small, slow fauna means that theprimary axis for faunal use is along the simpler continuumbetween sites dominated by large- and medium-sized preyand those dominated by small, difficult-to-catch prey. Sitesdistributed at the top of the graph (i.e., those with a highproportions of medium-size ungulate remains) all appear tobe assemblages resulting from fairly efficient collectionstrategies. These sites represent camps where either themedium-size game was the primary prey taken or asecondary resources acquired along with bison. The faunalassemblages from Myers–Hindman and a number of stratafrom Mummy Cave were dominated with bighorn sheepremains, which is not surprising given that both sites aresituated in areas of rugged terrain, the preferred habitat forbighorn sheep (Shackleton et al., 1999).
At the other sites scattered at the top of this graph (i.e.,Hell Gap—localities II and V; Horner I; Red Smoke, andJurgen-Area 2), the economic focus was apparently bison.The relatively modest number of pronghorn and deerremains at these sites suggests their use was of secondaryimportance. Perhaps medium-sized ungulates were eitheravailable in the same settings where the hunters searchedfor large game, or less likely, these taxa were mixed in withbison being targeted and were killed as part of the largerkill event.
On the other hand, sites showing evidence for a highlevel harvesting of small game (e.g., lagomorphs and/orrodents) likely represent a fundamentally different foragingapproach that required different strategies and equipmentthan those utilized in the procurement of either large ormedium-size game (Frison, 1991, 2004). For sites in thelower left corner of Fig. 6, large- and medium-sized prey,make up only a relative small proportion of the assem-blage, if they are present at all. The faunal collections fromthese sites appear to represent systematic and intentionalcollection of small-bodied game (Walker, 1975; Hughes,2003).
A focus on small, difficult-to-catch prey likely developedbecause sources of higher ranked resources in these settingswere not available or stores of large game remains (if theyexisted at all) were exhausted. These types of small gamewould have been available in the immediate vicinity of thesites, so collection likely occurred opportunistically bysmall foraging task groups that were gathering other foodand non-food resources (see Amick, 1994). The level ofsmall game use represented at these site, while substantial,was not at a comparable level to the systematic masscollection of rabbits, for example, as seen in Late
Prehistoric Great Basin sites (e.g., Madsen, 1993; Ugan,2005).Overall, the results are at least suggestive of some
humanly induced resource intensification during the laterportions of the Paleoindian period (Jones, 1999; Bamforth,2002a, b). Support for this hypothesis comes from the factthat the most common types of small prey taken are notthose that are slow reproducing (low turnover) or easilycaught types (e.g., turtle and shell fish), but instead are thequick reproducing (high turnover), difficult to catch types(e.g., birds, rabbits/hares, and subterranean rodents).If harvesting pressure on small-bodied resources was low
throughout the entire Paleoindian period, one wouldexpect relatively intensive use of the easier-to-harvestsmall-bodied prey through time. Fig. 7 shows that at leastfor alluvial valley sites this is not the case. Easily harvestedsmall prey types are better represented in early Paleoindianassemblages than in later assemblages, where they are rare.An opposite trend is seen for the abundance of difficult-to-harvest small prey, which generally increases in abundancethrough time. It is perhaps significant that the only localitywhere turtle remains are very abundant is the 11,500-year-old Aubrey site, which is the oldest site in the project areaand one likely occupied at a time when human population
ARTICLE IN PRESS
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
% S
mal
l Slo
w P
rey
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
% S
mal
l Dif
ficu
lt P
rey
7000 8000 9000 10000 11000 12000
Radiocarbon (14C) Age
Fig. 7. Plot of (A) %Slow small fauna and (B) %difficult small fauna
versus radiocarbon age for multifunction short-term camp assemblages in
alluvial valley habitats with best-fit regression lines.
M.E. Hill Jr. / Quaternary International 191 (2008) 34–5246

Author's personal copy
density and, therefore also harvesting pressure, was at itslowest level.
While these patterns are suggestive, it must be acknowl-edged that the strength and direction of these trends arestrongly influenced by a few sites. For example, if theAubrey site is dropped from consideration, the slope ofdecline in the use small, slow prey over time flattenssignificantly. Likewise, omissions of the Red Smoke andAubrey sites, which have a low frequency of small,difficult-to-catch prey, produces a graph with no obvioustrend in the use of this prey over time. The results of thisstudy cannot prove that human hunting caused resourcedepression, but only suggest this may have been the case. Itis clear that more quantitative studies of small animal useare needed to clarify this situation.
5. Discussion
This paper explores several aspects of the nature of thePaleoindian zooarchaeological record through a quantita-tive analysis of data from a number of sites spread acrossthe landscape. Specifically, it evaluated whether biasintroduced through collection strategies or site selectioninfluences our view of role big game hunting played inPaleoindian subsistence. It also explored how prehistoricdiets varied through time and across space in response toenvironmental differences or abundance of bison andmammoth populations.
My results generally support the trends proposed by thelarge prey abundance model outlined earlier in this paper.In addition, as I have previously suggested, site location isa strong determining factor in use of faunal resources (Hill,2007). As expected, early Paleoindian foragers focusedmostly on hunting bison and mammoths irrespective of thehabitat occupied. But later in time, when mammoth wereextinct and bison became regionally more scarce, large-game hunting became less common at least in foothill/mountain settings, although it continued at moderate levelsin alluvial valley settings and at very high levels in plains/rolling hills environments. With the decrease in abundanceof bison in foothill/mountain settings, foragers appeared toshift greater attention towards use of medium-sizedungulates and smaller game. While deer and pronghornare also exploited in plains/rolling hills or alluvial valleysettings, they appear to be supplemental resources to bison.
With the resolution of available information I cannotdetermine whether Paleoindian foragers settled in alluvialvalleys and foothill/mountain settings because they pro-vided a more-diverse choice of foodstuffs and otherresources than plains/rolling hills, or if the Paleoindiansdiet became diversified as occupation duration of thesehabitats extended. Nevertheless, it is possible to speculateabout the covariance between the Paleoindian diet and theeffects of regional differences in Paleoindians land use,resource distribution, and group mobility.
The high reliance on large game in plains/rolling hillhabitats likely is a response to the abundance of these taxa
in this habitat. Also, because hunters have to travel fardistances to find their prey in these settings and howfrequently they would have to move away from grasslandareas to acquire much needed wood and high-quality rawmaterial, there is also a relationship between search timeand prey choice. When search time increases or prey mustbe pursued over long-distance, foragers shift their selectioncriterion concerning which resource to exploit. The fartherforagers travel, the more likely they are to stop targetingsmall game and focus instead on finding large game(Schoener, 1971; Bettinger et al., 1997).Various researchers have suggested that Paleoindians
spent more time and used a greater diversity of resources inalluvial valleys and foothill/mountain environments thanthey did in plains/rolling hill environments (Surovell, 2003;Bamforth et al., 2005). It must be acknowledged thatarchaeologists do not really understand why groupsdecided to remain longer in these settings. Prehistoricpeople in these environments certainly spent time perform-ing a variety of activities related to tool production, foodpreparation and processing, in addition to habitationactivities. These settings would also encourage greaterduration of use because of the availability of protectedsettings, wood, and potable water.Regardless of our inability to understand exactly why
Paleoindian foragers were less mobile in these alluvialvalleys and foothill/mountains, this longer periods ofhabitat residence is associated with increased reliance onalternative game. This greater duration of time spent inthese habitats may have resulted in more resources beingcollected from these areas (Knell, 2007). This continuedexploitation in these resources patches would likelyeventually resulted in a decrease in the relative abundanceof key resources (e.g., Schoener, 1971; Charnov et al., 1976;Cannon, 2000). This explanation has been used by priorresearchers to explain other evidence for presumedintensification of resource exploitation during the Paleoin-dian period (Jones, 1999; Bamforth, 2002a; Borresen,2002).The results of this study show a general trend for Late
Paleoindian period sites in alluvial valleys, and to a lesserextent in foothill/mountain settings, with evidence for anexpansion of the diet in terms of number and type of preyitems. Part of this decrease in foraging efficiency was a shiftfrom one of nearly exclusive use of large prey items to oneof mixed use of large and small prey items, all of which arecostly to harvest. Unfortunately, the results of this studycannot show statistically meaningful trends on this issue.What is clear is that when Paleoindian foragers did notutilize large-bodied game, they primarily relied on faunawith high reproductive rates and that were generally harderto catch. This suggests that the easiest to catch small gamewas not readily available for exploitation due to environ-mental factors or previous hunting pressure, or both.Although not discussed here, strategies for expanding
diet breadth and decreased foraging efficiency would likelycorrelate with a pattern for increased processing of
ARTICLE IN PRESSM.E. Hill Jr. / Quaternary International 191 (2008) 34–52 47

Author's personal copy
carcasses, as suggested by Hill (2001, pp. 258–259) andByers (2002) (see also Cannon, 2003; Nagaoka, 2005,2006). This type of processing, although labor intensive,was designed to increase the caloric and fat yield from thesecarcasses. Perhaps significantly, during the Paleoindianperiod bone processing techniques appear to be limited tocold marrow extraction of major limb element (Jodry,1999; Hill, 2001). That Paleoindians do not appear to haveattempted ‘‘grease-rendering’’ (i.e., heat-in-liquid) or in-tensive processing of lower limb elements or crania suggeststhat while they were under some nutritional stress, it wassignificantly less than that faced by later hunters in NorthAmerica or Paleolithic peoples in the Old World (e.g.,Speth, 1983; Speth and Spielmann, 1983; Reeves, 1990;Stiner, 2002, 2003).
6. Conclusions
Three conclusions are drawn from the results of thisstudy, including:
� Site function has a significant effect on the compositionof Paleoindian faunal assemblages. Kill assemblages aregenerally dominated by single large-game taxa, whilecamp assemblages contain a larger diversity of preyrepresented. However, the abundance of large gameremains does not significantly differ between sampleswith both kills and camps and samples with just camps,when compared by different habitat settings.� Prey use changes based on habitat setting. Paleoindian
foragers targeted different prey types depending onwhich habitat setting they occupied. In the more-diverseenvironments (alluvial valleys and foothill/mountains)foragers exploited the largest prey whenever possible,but they also targeted a wide array of smaller faunalresources. In more uniform grassland environments,Paleoindians focused almost exclusively on huntingbison and mammoth.� There is limited evidence for increasing resource
intensification, as measured by increased exploitationof smaller game and decreased foraging efficiency,during the latter part of the Paleoindian period.Evidence for these changes is most apparent in alluvialvalley and foothill/mountain settings. The differenceslikely relate to prehistoric land use patterns andenvironmental conditions that affected abundance oflarge game.
Acknowledgements
I would like to acknowledge Margaret Beck, MatthewG. Hill, Bryan Hockett, Jack Hofman, Vance Holliday,Jason LaBelle, Ed Knell, Mary Stiner, Todd Surovell,Nicole Waguespack, and Chris Widga for helping shapemy thinking on the issues covered in this paper. Specialthanks also goes to Larry Martin of the University of
Kansas Museum of Natural History (12 Mile Creek), JackHofman of the University of Kansas Department ofAnthropology (Waugh), George Corner of the NebraskaState Museum (Red Smoke), Jeff Indeck and Rolla Shallerof the Panhandle-Plains Historical Museum (Rex Rod-gers), and George Frison, Marcel Kornfeld, MarthaRodgers, and Danny Walker of the University of Wyoming(Finley) for allowing me access to museum collections.Financial support for various stages of my research wasprovided by the Carroll D. Clark Fund of the Departmentof Anthropology at the University of Kansas, the HauryFund of the Department of Anthropology at the Universityof Arizona, and the George C. Frison Institute of theUniversity of Wyoming. An earlier version of this paperwas presented at the 2006 ICAZ conference, and was muchimproved as a result of comments received from MargaretBeck and Steve Wolverton.
References
Alroy, J., 1998. Putting North America’s End-Pleistocene megafaunal
extinctions in context: large-scale analyses of spatial patterns,
extinction rates, and size distributions. In: MacPhee, R. (Ed.),
Extinctions in Near Time. Kluwer Academic/Plenum Publishers,
New York, pp. 105–143.
Alroy, J., 2001. A multispecies overkill simulation of the End-Pleistocene
megafaunal mass extinction. Science 292, 1893–1896.
Amick, D.S., 1994. Folsom diet breadth and land use in the American
southwest. Unpublished Ph.D. Dissertation, Department of Anthro-
pology, University of New Mexico, Albuquerque.
Amick, D.S., 1996. Regional patterns of folsom mobility and land use in
the American southwest. World Archaeology 27, 411–426.
Baker, B.W., 1998. Vertebrate faunal remains from the 1/4- and 1/8-inch
screens. In: Collins, M.B., Wilson-Leonard (Eds.), An 11,000-year
Archaeological Record of Hunter-gatherers in Central Texas, 5.
Studies in Archaeology, vol. 31. Texas Archaeological Research
Laboratory and Archaeological Studies Program Report 10. Texas
Department of Transportation, Austin, pp. 1463–1509.
Baker, R.G., Fredlund, G.G., Mandel, R.D., Bettis III, E.A., 2000.
Holocene environments of the central Great Plains: multi-proxy
evidence from alluvial sequences, southeastern Nebraska. Quaternary
International 67, 75–88.
Bamforth, D.B., 2002a. High-tech foragers? Folsom and later Paleoindian
technology on the Great Plains. Journal of World Prehistory 16,
55–98.
Bamforth, D.B., 2002b. The Paleoindian occupation of the medicine creek
drainage, southwestern Nebraska. In: Roper, D.C. (Ed.), Medicine
Creek: Seventy Years of Archaeological Investigations. The University
of Alabama Press, Tuscaloosa, pp. 54–83.
Bamforth, D.B., Becker, M., Hudson, J., 2005. Intrasite spatial analysis,
ethnoarchaeology, and Paleoindian land-use on the Great Plains: The
Allen site. American Antiquity 70, 561–580.
Barbour, E.H., Schultz, C.B., 1932. The Scottsbluff bison quarry and its
artifacts. Bulletin of the Nebraska State Museum 34 (1), 283–286.
Baxevanis, S.E., 1997. Collection management and analysis of the Lake
Theo Folsom bone assemblage. Unpublished Master’s Thesis,
Museum Science, Texas Tech University, Lubbock.
Bayham, F.E., 1979. Factors influencing the archaic pattern of animal
utilization. Kiva 44, 219–235.
Belovsky, G.E., 1988. An optimal foraging-based model of hunter-
gatherer population dynamics. Journal of Anthropological Archae-
ology 7, 329–372.
Bement, L.C., 1999. Bison Hunting at Cooper Site: Where Lightning Bolts
Drew Thundering Herds. University of Oklahoma Press, Norman.
ARTICLE IN PRESSM.E. Hill Jr. / Quaternary International 191 (2008) 34–5248

Author's personal copy
Bement, L.C., 2003. Constructing the cooper model of Folsom bison kills
on the Southern Plains. Great Plains Research 13, 27–41.
Bement, L.C., Carter, B., 2003. Clovis bison hunting at the Jake bluff site,
NW Oklahoma. Current Research in the Pleistocene 20, 5–7.
Bettinger, R.L., Malhi, R., McCarthy, H., 1997. Central place models of
acorn and mussel processing. Journal of Archaeological Science 24,
887–899.
Bonnichsen, R., Stanford, D.J., Fastook, J.L., 1987. Environmental
change and developmental history of human adaptive patterns: the
Paleoindian case. In: Ruddiman, W.F., Wright, H.E. (Eds.), North
America and Adjacent Oceans During the Last Deglaciation.
Geological Society of America, Boulder, pp. 43–424.
Borresen, J.A., 2002. A faunal analysis of the Frazier site, an Agate Basin-
age Bison Kill-Butchery site in Northeastern Colorado. Unpublished
Master’s Thesis, Department of Anthropology, University of Tennes-
see, Knoxville.
Bousman, C.B., Baker, B.W., Kerr, A.C., 2004. Paleoindian archeology in
Texas. In: Pertula, T.K. (Ed.), The Prehistory of Texas. Texas A & M
University Press, College Station, pp. 15–97.
Brain, C.K., 1981. The Hunters or the Hunted? Introduction to African
Cave Taphonomy. The University of Chicago Press, Chicago.
Broughton, J.M., 1994a. Declines in mammalian foraging efficiency
during the Late Holocene, San Francisco bay. Journal of Anthro-
pological Archaeology 13, 371–401.
Broughton, J.M., 1994b. Late Holocene resource intensification in the
Sacamento valley, California: The Vertebrate Evidence. Journal of
Archaeological Science 12, 501–514.
Broughton, J.M., 2002. Prey spatial structure and behavior affect
archaeological tests of optimal foraging models: examples from
Emeryville Shellmound Vertbrate fauna. World Archaeology 34,
60–83.
Broughton, J.M., Grayson, D.K., 1993. Diet breadth, adaptive change,
and the White Mountain faunas. Journal of Archaeological Science 20,
331–336.
Butler, V.L., Campbell, S.K., 2004. Resource intensification and resource
depression in the Pacific Northwest of North America: a zooarchaeo-
logical review. Journal of World Prehistory 18, 327–405.
Byerly, R.M., Cooper, J.R., Meltzer, D.J., Hill, M.E., LaBelle, J.M., 2005.
On bonfire shelter (Texas) as a Paleoindian jump: an assessment using
GIS and zooarchaeology. American Antiquity 70, 595–629.
Byers, D.A., 2002. Paleoindian fat-seeking behavior: The Hell Gap Site
locality II Agate Basin faunal assemblage. Plains Anthropologist 47,
359–377.
Byers, D.A., Ugan, A., 2005. Should we expect large game specialization
in the Late Pleistocene? An optimal foraging perspective on Early
Paleoindian prey choice. Journal of Archaeological Science 32,
1624–1640.
Byers, D.A., Smith, C.S., Broughton, J.M., 2005. Holocene artiodactyl
populations histories and large game hunting in the Wyoming Basin,
USA. Journal of Archaeological Science 32, 125–142.
Cannon, M.D., 2000. Large mammals relative abundance in Pithouse and
Pueblo period Archaeofaunas from Southwestern New Mexico:
resource depression among the Mimbres–Mogollon. Journal of
Anthropological Archaeology 19, 317–347.
Cannon, M.D., 2003. A model of central place forager prey choice and an
application to faunal remains from the Mimbres Valley, New Mexico.
Journal of Anthropological Archaeology 22, 1–25.
Cannon, M.D., 2004. Geographic variability in North American Mammal
community richness during the terminal Pleistocene. Quaternary
Science Reviews 23, 1099–1123.
Cannon, K.P., Cannon, M.B., 2004. Zooarchaeological and wildlife
management in the Greater Yellowstone ecosystem. In: Lyman, R.L.,
Cannon, K.P. (Eds.), Zooarchaeology and Conservation Biology. The
University of Utah Press, Salt Lake City, pp. 45–60.
Cannon, M.D., Meltzer, D.J., 2004. Early Paleoindian foraging: examin-
ing the faunal evidence for large mammal specialization and
regional variability in prey choice. Quaternary Science Review 23,
1955–1987.
Cannon, M.D., Meltzer, D.B., this volume. Explaining variability in Early
Paleoindian foraging. Quaternary International.
Charnov, E.L., 1976. Optimal foraging, the marginal value theorem.
Theoretical Population Biology 9, 129–136.
Charnov, E.L., Orians, G.H., Hyatt, K., 1976. Ecological implications of
resource depression. The American Naturalist 110, 247–259.
Collins, M.B., Stanford, D.J., Hofman, J.L., Jodry, M.A., Rose, R.O.,
Todd, L.C., Kilber, K., Blackmar, J.M., 1997. Cody down south: the
Seminole-Rose Site in West Texas. Current Research in the Pleistocene
14, 15–18.
Corbeil, M.R., 1995. The Archaeology and Taphonomy of the Heron
Eden Site, Southwestern Saskatchewan. Unpublished M.A. Thesis,
Department of Anthropology and Archaeology, University of
Saskatchewan, Saskatoon.
Davis, E.M., 1953. Recent data from two Paleoindian sites on Medicine
Creek, Nebraska. American Antiquity 18, 380–386.
Davis, E.M., 1962. Archaeology of the Lime Creek Site in Southwestern
Nebraska. Special Publication No. 3. University of Nebraska State
Museum, Lincoln.
Davis, O.K., Sheppard, J.C., Robertson, S., 1986. Contrasting climatic
histories for the Snake River Plains, Idaho, resulting from multiple
thermal maximum. Quaternary Research 26, 321–339.
Emlen, J.M., 1966. The role of time and energy in food preference. The
American Naturalist 100, 611–617.
FAUNMAP Working Group, 1994. FAUNMAP: a database document-
ing Late Quaternary distribution of mammal species in the United
States. Illinois State Museum Scientific Papers 25, Illinois State
Museum, Springfield.
Feranec, R.S., MacFadden, B.J., 2000. Evolution of the grazing niche in
Pleistocene mammals from Florida: evidence from stable isotopes.
Palaeogeography, Palaeoclimatology, Palaeoecology 162, 155–169.
Ferring, C.R., 2001. The archaeology and paleoecology of the Aubrey
Clovis Site (41DN479), University of North Texas, Denton County,
Texas. Report submitted to US Army Corp of Engineers, Fort Worth
District, Fort Worth.
Fiedel, S.J., 2004. Rapid migrations by Arctic hunting peoples. In: Barton,
C.M., Clark, G.A., Yesner, D.R., Pearson, G.A. (Eds.), The
Settlement of the American Continents: a Multidisciplinary Approach
to Human Biogeography. The University of Arizona Press, Tucson,
pp. 79–84.
Frison, G.C., 1974. The Casper Site: a Hell Gap Bison Kill on the High
Plains. Academic Press, New York.
Frison, G.C., 1991. Prehistoric Hunters of the High Plains. Academic
Press, San Diego.
Frison, G.C., 1992. The foothills-mountain and the open plains: the
dichotomy in Paleoindian subsistence strategies between two ecosys-
tems. In: Stanford, D.J., Day, J.S. (Eds.), Ice Age Hunters of the
Rockies. Denver Museum of Natural History or University Press of
Colorado, Niwot, pp. 323–342.
Frison, G.C. (Ed.), 1996. The Mill Iron Site. University of New Mexico
Press, Albuquerque.
Frison, G.C., 2004. Survival by Hunting: Prehistoric Human Predators
and Animal Prey. University of California Press, Berkley, CA.
Frison, G.C., Todd, L.C. (Eds.), 1986. The Colby Mammoth Site:
Taphonomy and Archaeology of a Clovis Kill in Northern Wyoming.
University of New Mexico Press, Albuquerque.
Frison, G.C., Todd, L.C. (Eds.), 1987. The Horner Site: The Type Site of
the Cody Cultural Complex. Academic Press, Orlando.
Fulgham, T., Stanford, D.J., 1982. The Frasca site: a preliminary report.
Southwestern Lore 48, 1–9.
Geist, V., 1998. Deer of the World: their Evolution, Behavior, and
Ecology. Stackpole Books, Mechanicburg, PA.
Geist, V., 1999. Periglacial ecology, large mammals, and their significance
to human biology. In: Bonnichsen, R., Turnmire, K.L. (Eds.), Ice Age
Peoples of North America: Environments, Origins, and Adaptations of
the First Americans. Oregon State University, Corvallis, pp. 78–94.
Gero, J., 1993. The social world of prehistoric facts: gender and power in
Paleoindian research. In: du Cross, H., Smith, L. (Eds.), Women in
ARTICLE IN PRESSM.E. Hill Jr. / Quaternary International 191 (2008) 34–52 49

Author's personal copy
Archaeology, A Feminist Critique. Occasional Papers in Prehistory
No. 23, Department of Prehistory, Australian National University,
Canberra, pp. 31–40.
Graham, R.W., Lundelius, E.L., 1984. Coevolutionary disequilibrium and
Pleistocene extinctions. In: Martin, P.S., Klein, R.G. (Eds.), Quatern-
ary Extinction: a Prehistoric Revolution. University of Arizona Press,
Tucson, pp. 223–249.
Graham, R.W., Mead, J.I., 1987. Environmental fluctuations and
evolution of mammalian faunas during the last deglaciation in North
America. In: Ruddiman, W.F., Wright, H.E. (Eds.), North American
and Adjacent Oceans during the last Deglaciation. Geological Society
of America, Boulder, pp. 371–402.
Grayson, D.K., 1993. The Desert’s Past: A Natural History of the Great
Basin. Smithsonian Institution Press, Washington, DC.
Grayson, D.K., Meltzer, D.J., 2002. Clovis hunting and large mammal
extinction: a critical review of evidence. Journal of World Prehistory
16, 313–359.
Grayson, D.K., Meltzer, D.J., 2003. A requiem for North American
overkill. Journal of Archaeological Science 30, 585–593.
Greiser, S.T., 1985. Predictive models of hunter–gatherer subsistence and
settlement strategies on the Central High Plains. Plains Anthropologist
Memoir No. 20, Part 2, Lincoln.
Guthrie, R.D., 1984. Alaskan megabucks, megabulls, and megarams: the
issue of Pleistocene gigantism. In: Genoways, H.H., Dawson, M.R.
(Eds.), Contributions in Quaternary Vertebrate Paleontology: A
Volume in Memorial to John E. Guilday. Special publication in
Carnegie Museum of Natural History No. 8, Pittsburgh, pp. 482–510.
Haynes, G., 2002a. The catastrophic extinction of North American
mammoths and mastodons. World Archaeology 33, 391–416.
Haynes, G., 2002b. The Early Settlement of the Americas: the Clovis Era.
Cambridge University Press, Cambridge.
Hawkes, K., Hill, K., O’Connell, J.F., 1982. Why hunter gather: optimal
foraging and the ache of Eastern Paraguay. American Ethnologist 9,
379–398.
Heller, R., 1980. On optimal diet in a patchy environment. Theoretical
Population Biology 17, 201–214.
Hemmings, C.A., 2004. The organic Clovis: a single continent-wide
cultural adaptation. Unpublished Ph.D. Dissertation, Department of
Anthropology, University of Florida, Gainesville.
Hill, M.G., 2001. Paleoindian diet and subsistence behavior on the
Northwestern Great Plains of North America. Unpublished Ph.D.
Dissertation, Department of Anthropology, University of Wisconsin,
Madison.
Hill Jr., M.E., 2002a. The Milnesand Site: site formation study of a
Paleoindian bison bonebed in Eastern New Mexico. Plains Anthro-
pologist 47, 323–337.
Hill Jr., M.E., 2002b. The folsom-age 12 Mile Creek Bison bonebed in
Western Kansas. In: Dort, Jr., W. (Ed.), TER-QUA Symposium
Series, vol. 3. Institute for Tertiary-Quaternary Studies, University of
Kansas, Lawrence, pp. 53–70.
Hill Jr., M.E., 2006. Rex Rodgers bonebed: reevaluation of site formation.
Current Research in the Pleistocene 23, 108–110.
Hill Jr., M.E., 2007. A moveable feast: variations in faunal resource use
among Central and Western North American Paleoindian sites.
American Antiquity 72, 417–438.
Hill Jr., M.E., Hofman, J.L., 1997. The Waugh site: a Folsom-age bison
bonebed in Northwestern Oklahoma. Plains Anthropologist 42,
63–83.
Hill Jr., M.E., Hill, M.G., 2002. Season of bison mortality at the Jurgens
site, Weld County, Colorado. Current Research in the Pleistocene 19,
104–106.
Hill, M.G., May, D.W., Rapson, D.J., 2006. A new, stratified Late
Paleoindian locality on the Clary Ranch, Ash Hallow, Garden County,
Nebraska. Current Research in the Pleistocene 23, 110–112.
Hill, M.G., May, D.W., Rapson, D.J., Boehm, A.R., Otarola-Castillo, E.,
this volume. Faunal exploitation by early Holocene hunter/gatherers
on the Great Plains of North America: evidence from the Clary Ranch
Sites. Quaternary International.
Hockett, B., Haws, J., 2003. Nutritional ecology and diachronic trends in
Palaeolithic diet and health. Evolutionary Anthropology 12,
211–216.
Hofman, J.L., 1999. Unbounded hunters: Folsom bison hunting on the
Southern Plains ca 10,500BP, the lithic evidence. In: Jaubert, J.,
Brugal, J.-P., David, F., Enloe, J.G. (Eds.), Le Bison: Gibier et Moyen
de Subsistance des Hommes du Paleolithique aux Paleoindiens des
Grandes Plaines. APDCA, Antibes, pp. 383–415.
Hofman, J.L., Blackmar, J.M., 1997. The Paleoindian Laird Bison bone
bed in Northwestern Kansas. The Kansas Anthropologist 18, 45–58.
Hofman, J.L., Todd, L.C., 2001. Tyranny in the archaeological record of
specialized hunters. In: Gerlach, S.C., Murray, M.S. (Eds.), People and
Wildlife in Northern North America: Essays in Honor of R. Dale
Guthrie. BAR International Series 944. Archaeo Press, Oxford, pp.
200–215.
Hofman, J.L., Hill Jr., M.E., Johnson, W.C., Sather, D.T., 1995. Norton:
an Early-Holocene Bison bone bed in Western Kansas. Current
Research in the Pleistocene 12, 19–21.
Holder, P., Wilke, J., 1949. The frontier culture complex: a preliminary
report of the prehistoric hunters camp in Southwestern Nebraska.
American Antiquity 14, 260–266.
Holliday, V.T., 1997. Paleoindian Geoarchaeology of the Southern High
Plains. University of Texas Press, Austin.
Hughes, S.S., 2003. Beyond the Altithermal: the role of climate change in
the prehistoric adaptation of Northwestern Wyoming. Unpublished
Ph.D. Dissertation, Department of Anthropology, University of
Washington, Seattle.
Husted, W.M., 1969. Bighorn Canyon Archaeology, Smithsonian Institute
River Basin Surveys Publications in Salvage Archeology No. 12,
Lincoln.
Jodry, M.A., 1999. Folsom technological and socioeconomic strategies:
view from Stewart’s cattle guard and the upper Rio Grande Basin,
Colorado. Unpublished Ph.D. Dissertation, Department of Anthro-
pology, American University, Washington, DC.
Johnson, E., 1977. Animal food resources of Paleoindians. In: Johnson, E.
(Ed.), Paleoindian Lifeways. Museum Journal (West Texas Museum
Association) 15, 107–122.
Johnson, E. (Ed.), 1987. Lubbock Lake: Late Quaternary Studies on the
Southern High Plains. Texas A & M University Press, College Station.
Jones, D.C., 1999. Late Paleoindian/early archaic archaeology and
subsistence: a taphonomic analysis of the Lime Creek Site (25ST41),
Southwestern Nebraska. Unpublished Master’s Thesis, Department of
Anthropology, Colorado State University, Fort Collins.
Judge, W.J., 1973. Paleoindian Occupation of the Central Rio Grande
Valley in NewMexico. University of New Mexico Press, Albuquerque.
Kay, M., 1998. The great plains setting. In: Wood, W.R. (Ed.),
Archaeology of the Great Plains. University of Kansas Press,
Lawrence, pp. 16–47.
Kelly, R.L., 1995. The foraging spectrum: diversity in hunter–gatherer
lifeways. Smithsonian Institution Press, Washington.
Kelly, R.L., Todd, L.C., 1988. Coming into the country: Early
Paleoindian hunting and mobility. American Antiquity 53, 231–244.
Klein, R.G., 1976. The mammalian fauna of the Klasies River mouth sites,
Southern Cape province, South Africa. South African Archaeological
Bulletin 31, 75–98.
Knell, E.J., 2007. The organization of Late Paleoindian Cody Complex
land-use on the northern American great plains. Unpublished Ph.D.
Dissertation, Department of Anthropology, Washington State Uni-
versity, Pullman.
Knell, E.J., Hill, M.G., Izeta, A., Kornfeld, M., Haynes Jr., C.V., Frison,
G.C., 2002. The Cody-complex component at locality V of the Hell
Gap site, Wyoming. Current Research in the Pleistocene 19, 49–52.
Kornfeld, M., Frison, G.C., 2000. Paleoindian occupation of the high
country: the case of Middle Park Colorado. Plains Anthropologist 45,
129–153.
Kornfeld, M., Larson, M.L., this volume. Bonebeds and other Myths:
Paleoindian to Archaic transition on North American Great Plains
and Rocky Mountains. Quaternary International, this volume.
ARTICLE IN PRESSM.E. Hill Jr. / Quaternary International 191 (2008) 34–5250

Author's personal copy
Kornfeld, M., Osborn, A.J. (Eds.), 2003. Island on the Plains: Ecological
and Social, and Ritual Use of Landscapes. University of Utah Press,
Salt Lake City.
Kornfeld, M., Frison, G.C., Larson, M.L., Miller, J.C., Saysette, J., 1999.
Paleoindian bison procurement and Paleoenvironments in Middle
Park of Colorado. Geoarchaeology 14, 655–674.
Kuhn, S.L., Stiner, M.C., 2001. The antiquity of hunter–gatherers. In:
Panter-Brick, C., Layton, R.H., Rowley-Conwy, P. (Eds.), Hunter–-
gatherers: an Interdisciplinary Perspective. Cambridge University
Press, Cambridge, pp. 99–142.
Kurten, B., Anderson, E., 1980. Pleistocene Mammals of North America.
Columbia University Press, New York.
LaBelle, J.M., 2005. Hunter–gatherer foraging variability during the early
Holocene of the Central Plains of North America. Unpublished Ph.D.
Dissertation, Southern Methodist University, Dallas.
Leonard, E.M., Reasoner, M.A., 1999. A continuous Holocene glacial
record inferred from postglacial lake sediments in Banff National
Park, Alberta, Canada. Quaternary Research 51, 1–13.
Leonhardy, F., 1966. Domebo: a Paleoindian Mammoth Kill in the
Prairie-Plains. Contribution of the Museum of the Great Plains, No. 1.
The Great Plains Historical Association, Lawton, Oklahoma.
Levins, R., 1968. Evolution in Changing Environments: Some Theoretical
Explorations. Princeton University Press, Princeton.
Lundelius Jr., E.L., Graham, R.W., Anderson, E., Guilday, J., Holman,
J.A., Steadman, D.W., Webb, S.D., 1983. Terrestrial vertebrate
faunas. In: Porter, S.C. (Ed.), Late Quaternary Environments of the
United States. The Late Pleistocene, vol. 1. University of Minnesota
Press, Minneapolis, pp. 311–353.
MacArthur, R.H., Pianka, E.R., 1966. On optimal use of a patchy
environment. The American Naturalist 100, 603–609.
MacDonald, J.N., 1981. North American Bison: their Classification and
Evolution. University of California Press, Berkeley, CA.
MacDonald, D.H., 1998. Subsistence, sex, and cultural transmission in
Folsom culture. Journal of Anthropological Archaeology 17, 217–239.
Madsen, D.B., 1993. Testing diet breadth models: examining adaptive
change in the Late Prehistoric Great Basin. Journal of Archaeological
Science 20, 321–329.
Mallouf, R.J., Mandel, R.D., 1997. Horace rivers: a Late-Plainview
component in the Northeastern Texas panhandle. Current Research in
the Pleistocene 14, 50–52.
Mandel, R.D., Hofman, J.L., 2003. Geoarchaeological investigations at
the Winger Site: a Late Paleoindian bison bonebed in Southwestern
Kansas, USA. Geoarchaeology 18, 129–144.
Martin, P.S., 2005. Twilight of the Mammoths: Ice Age Extinctions and
the Rewilding of America. University of California Press, Berkeley,
CA.
Martin, P.S., Steadman, D.W., 1999. Prehistoric extinctions on Islands
and continents. In: MacPhee, R.D.E. (Ed.), Extinction in Near Time.
Kluwer Academic/Plenum Publishers, New York, pp. 17–52.
Martin, L.D., Williams, D.R., 2007. Late-Pleistocene biodiversity in the
Central Plains. Current Research in the Pleistocene 23, 172–173.
McCartney, P.H., 1983. An archaeological analysis of bison remains from
the Cody Paleo-Indian site of Lamb Spring. Unpublished Master’s
Thesis, Department of Anthropology, University of Arizona, Tucson.
McNamara, J., 1982. Optimal patch use in a stochastic environment.
Theoretical Population Biology 21, 269–288.
Meltzer, D.J., 1988. Late Pleistocene human adaptations in Eastern North
America. Journal of World Prehistory 2, 1–52.
Meltzer, D.J., 1993. Is there a Clovis adaptation. In: Soffer, O., Praslov,
N.D. (Eds.), From Kostenki to Clovis: Upper Paleolithic-Paleo-Indian
Adaptations. Plenum Press, New York, pp. 293–310.
Meltzer, D.J., 1995. Clocking the first Americans. Annual Review of
Anthropology 24, 21–45.
Meltzer, D.J., 2006. Folsom: New Archaeological Investigations of a
Classic Bison Kill. University of California Press, Berkeley, CA.
Munro, N.D., 2004. Zooarchaeological measures of hunting pressure and
occupation intensity in the Natufian. Current Anthropology 45
(Suppl.), S5–S33.
Nagaoka, L., 2002. The effects of resource depression on foraging
efficiency, diet breath, and patch use in Southern New Zealand.
Journal of Anthropological Archaeology 21, 419–442.
Nagaoka, L., 2005. Declining foraging efficiency and Moa Carcass
exploitation in Southern New Zealand. Journal of Archaeological
Science 32, 1328–1338.
Nagaoka, L., 2006. Prehistoric Seal Carcass exploitation at the Shag
Mouth Site, New Zealand. Journal of Archaeological Science 33,
242–258.
Neck, R.W., 1995. Molluscan remains. In: Holliday, V.T. (Ed.),
Stratigraphy and Paleoenvironments of Late Quaternary Valley Fills
on the Southern High Plains. Geological Society of America Memoir
186, Boulder, pp. 59–67.
Olson, C.G., Porter, D.A., 2002. Isotopic and geomorphic evidence for
Holocene climate, Southwestern Kansas. Quaternary International 87,
29–44.
Orians, G.H., Pearson, N.E., 1979. On the theory of central place
foraging. In: Horn, D.J., Stairs, G.R., Mitchell, R.D. (Eds.), Analysis
of Ecological Systems. Ohio State University Press, Columbus, pp.
155–177.
Owens-Smith, R.N., 1987. Pleistocene extinctions: the pivotal role of
megaherbivores. Paleobiology 13, 351–361.
Reeves, B.O.K., 1990. Communal Bison hunters of the Northern Plains.
In: Davis, L.B., Reeves, B.O.K. (Eds.), Hunters of the Recent Past.
Unwin Hyman, Boston, pp. 168–194.
Roberts, F.H.H., 1935. A Folsom complex: a preliminary report on
investigations at the Lindenmeier in Northern Colorado. Smithsonian
Miscellaneous Collections 94, 1–35.
Schoener, T.W., 1971. Theory of feeding strategies. Annual Review of
Ecology and Systematics 2, 369–404.
Sellards, E.H., 1938. Artifacts associated with fossil elephant. Bulletin of
the Geological Society of America 49, 999–1010.
Sellards, E.H., 1952. Early Man in America. University of Texas Press,
Austin, TX.
Sellet, F., 1999. A dynamic view of Paleoindian assemblages at the Hell
Gap Site, Wyoming: reconstructing lithic technological systems.
Unpublished Ph.D. Dissertation, Department of Anthropology,
Southern Methodist University, Dallas.
Shackleton, D.M., Shank, C.C., Wikeem, B.M., 1999. Natural history of
rocky mountain and California bighorn sheep. In: Valdez, R.,
Krausman, P.R. (Eds.), Mountain Sheep of North America. The
University of Arizona Press, Tucson, pp. 78–138.
Simms, C.R., 1987. Behavioral ecology and hunter–gatherer foraging:
an example from the Great Basin. BAR International Series 381,
1–157.
Simpson, E.H., 1949. Measurement of diversity. Nature 163, 688.
Smith, C.S., McNees, L.M., 1990. Rattlesnake pass site: a Folsom
occupation in South Central Wyoming. Plains Anthropologist 35,
273–289.
Smith, C.S., Reust, T.P., Russell, R.D., 2003. Site 48UT375: Late
Paleoindian period subsistence and land use patterns in the Green
River Basin, Wyoming. Plains Anthropologist 48, 133–151.
Speth, J., 1983. Bison Kills and Bone Counts. University of Chicago Press,
Chicago.
Speth, J.D., Spielmann, K., 1983. Energy source, protein metabolism, and
hunter–gatherer subsistence strategies. Journal of Anthropological
Archaeology 2, 1–31.
Speth, J.D., Tchernov, E., 2002. Middle Paleolithic tortoise use at Kebara
Cave (Israel). Journal of Archaeological Science 59, 471–483.
Stephens, D.W., Krebs, J.R., 1986. Foraging Theory. Princeton University
Press, Princeton, NJ.
Stiner, M.C., 2001. Thirty years on the ‘‘Broad Spectrum Revolution’’ and
Paleolithic demography. The Proceedings of the National Academy of
Sciences 98, 6993–6996.
Stiner, M.C., 2002. Carnivory, coevolution, and the geographic spread of
the genus Homo. Journal of Archaeological Research 10, 1–63.
Stiner, M.C., 2003. Zooarchaeological evidence for resource intensifica-
tion in Algarve, Southern Portugal. Promontoria 1, 27–58.
ARTICLE IN PRESSM.E. Hill Jr. / Quaternary International 191 (2008) 34–52 51

Author's personal copy
Stiner, M.C., 2005. The faunas of Hayonim Cave Israel: a 200,000-year
Record of Paleolithic Diet, Demography, and Society. American
School of Prehistoric Research Bulletin, vol. 48. Peabody Museum of
Archaeology, Harvard University Press, Cambridge, MA.
Stiner, M.C., Munro, N.D., 2002. Approaches to prehistoric diet breadth,
demography, and prey ranking systems in time and space. Journal of
Archaeological Method and Theory 9, 175–208.
Stiner, M.C., Munro, N.D., Surovell, T.A., Tchernov, E., Bar-Yosef, O.,
1999. Paleolithic population growth pulses evidenced by small animal
exploitation. Science 283, 190–194.
Stiner, M.C., Munro, N.D., Surovell, T.A., 2000. The tortoise and the
hare: small-game use, the broad-spectrum revolution and Paleolithic
demography. Current Anthropology 41, 39–73.
Surovell, T.A., 2000. Early Paleoindian women, children, mobility, and
fertility. American Antiquity 65, 493–509.
Surovell, T.A., 2003. The behavioral ecology of Folsom Lithic Technol-
ogy. Ph.D. Dissertation, University of Arizona, Tucson; University
Microfilms, Ann Arbor.
Surovell, T.A., Waguespack, N., Brantingham, P.J., 2005. Global
archaeological evidence for Proboscidean overkill. Proceeding of the
National Academy of Sciences 102, 6231–6236.
Szuter, C.R., Bayham, F.E., 1989. Sedentism and prehistoric animal
procurement among desert horticulturalists of the North American
Southwest. In: Kent, S. (Ed.), Farmers as Hunters: the Implications of
Sedentism. Cambridge University Press, Cambridge, pp. 80–95.
Todd, L.C., 1991. Seasonality studies and Paleoindian subsistence
strategies. In: Stiner, M.C. (Ed.), Human Predators and Prey
Mortality. Westview Press, Boulder, pp. 215–238.
Todd, L.C., Hofman, J.L., Schultz, C.B., 1992. Faunal analysis and
Paleoindian studies: a reexamination of the Lipscomb Bison bonebed.
Plains Anthropologist 37, 137–165.
Ugan, A., 2005. Climate, bone density, and resource depression: what is
driving variation in large and small game in Fremont Archaeofaunas?
Journal of Anthropological Archaeology 24, 227–251.
Waguespack, N.M., 2003. Clovis hunting and the organization of
subsistence labor. Ph.D. Dissertation, University of Arizona, Tucson;
University Microfilms, Ann Arbor.
Waguespack, N.M., 2007. Why we’re still arguing about the Pleistocene
occupation of the Americas. Evolutionary Anthropology 16, 63–74.
Waguespack, N.M., Surovell, T., 2003. Clovis hunting strategies, or how
to make out on plentiful resources. American Antiquity 68, 333–352.
Walker, D.N., 1975. A cultural and ecological analysis of the vertebrate
fauna from the medicine lodge creek site (48BH499). Unpublished
Masters Thesis, Department of Anthropology, University of Wyom-
ing, Laramie.
Wheat, J.B., 1972. The Olsen-Chubbuck Site: a Paleoindian Bison Kill.
Memoir No. 26. Society for American Archaeology, Washington, DC.
Wheat, J.B., 1978. Olsen-Chubbuck and Jurgens Sites: four aspects of
Paleo-Indian economy. In: Davis, L.B., Wilson, M. (Eds.), Bison
Procurement and Utilization: a Symposium. Memoir 14. Plains
Anthropologist, vol. 23, pp. 84–89.
Wheat, J.B., 1979. The Jurgens Site. Plains Anthropologists Memoir 15,
Lincoln.
Whittaker, W., 1997. The Cherokee excavation revisited: prehistoric
adaptation and change in the eastern great plains. Unpublished
Master’s Thesis, Department of Anthropology, University of Iowa,
Iowa City.
Williams, J.D. (Ed.), 2000. The Big Black Site (32DU955C): a Folsom
Complex Workshop in Knife River Flint Quarry Area, North Dakota.
Washington State University Press, Pullman.
Wien, J.A., 1976. Population responses to patchy environments. Annual
Review of Ecology and Systematics 7, 81–120.
Winkler, D.A., 1982. Re-evaluation of the vertebrate fauna from the
Lewisville archeological site, Denton County, TX. University of North
Texas. Report submitted to US Army Corp of Engineers, Dallas.
Winterhalder, B., 2001. The behavioral ecology of hunter–gatherers. In:
Panter-Brick, C., Layton, R.H., Rowley-Conwy, P. (Eds.), Hunter–-
Gatherers: an Interdisciplinary Perspective. Cambridge University
Press, Cambridge, pp. 12–38.
Wobst, H.M., 1974. Boundary conditions for Paleolithic social systems: a
simulation approach. American Antiquity 39, 147–178.
Wolverton, S., 2005. The effect of the Hypsithermal on prehistoric
foraging efficiency in Missouri. American Antiquity 70, 91–106.
Wormington, H.M., 1957. Ancient man in North America. Denver
Museum of Natural History, Popular Series No. 4, Denver.
Yansa, C.H., 1998. Holocene Paleoenvironment and Paleohydrology of a
Prairie Pothole in Southern Saskatchewan, Canada. Journal of
Paleolimnology 19, 429–441.
ARTICLE IN PRESSM.E. Hill Jr. / Quaternary International 191 (2008) 34–5252




















