An unusual crinoid-barnacle association in the type area of the Danian (Paleocene), Denmark
Upload
independentCategory
view
4download
0
ISSN 0031�0301, Paleontological Journal, 2010, Vol. 44, No. 5, pp. 584–594. © Pleiades Publishing, Ltd., 2010Original Russian Text © L.B. Golovneva, 2010, published in Paleontologicheskii Zhurnal, 2010, No. 5, pp. 99–107.
584
INTRODUCTION
The genus Ginkgo L. of the modern flora includesthe only species G. biloba L., being the last survivedmember of the order Ginkgoales, which was commonin the Mesozoic and Cenozoic and comprised manygenera and species.
Leaf remains attributed to the genus Ginkgo (or theformal genus Ginkgoites Seward) appear in fossil florasas early as the Early Jurassic (Zhou, 1997). The heydayof the genus took place in the Late Jurassic–EarlyCretaceous (Samylina, 1967). During the Jurassic,Cretaceous, and Paleogene, Ginkgo had a circumpolarrange in both hemispheres (Tralau, 1968; Hill andCarpenter, 1999; Pole and Douglas, 1999). The rangeof the genus restricted in the Miocene up to Eurasia,and Ginkgo disappeared from Europe till the end of thePliocene (Samylina, 1967; Tralau, 1968).
Leaves of most Jurassic and Early Cretaceous spe�cies of Ginkgo are dissected into narrow lobes. Since themid�Cretaceous, entire�margined leaves that resembleleaves of the modern species gradually became domi�nants. The taxonomy of fossil species was originallybased on such morphological characters as degree ofdissection of the leaf blade, dimensions, petiole length,vein density, etc. However, the differentiation betweenspecies on a sole basis of morphological characters wasalways troublesome, since most species of Ginkgo havehighly variable leaf blades. Therefore, since Harris(1935) and Florin (1936, 1937), fossil species of Ginkgo
as well as members of many other groups of fossil gym�nosperms have been mostly diagnosed on epidermalcharacters, which are more diverse and less variablethan morphological characters.
Entire�margined leaves of Ginkgo were firstdescribed from a Late Miocene flora of Italy as Salis�buria adiantoides Unger (Unger, 1845). Heer (1878),who studied the Cretaceous flora of the SakhalinIsland, transferred this species into the genus Ginkgo.Nowadays, Ginkgo adiantoides (Unger) Heer is con�ventionally used as a formal name for entire�marginedleaf imprints of Ginkgo without preserved cuticle.
Florin (1937) was the first who studied the epider�mal morphology of fossil entire�margined species ofGinkgo, on the material from the Paleocene of Isle ofMull and the Pliocene of Germany. He describedleaves from the former locality as a new species,G. gardneri Florin, and assigned Pliocene leaves toGinkgo adiantoides. Later, Samylina (1967) proposed anew name, G. florinii Samylina, for the Germanremains, since the type specimen of G. adiantoidesfrom Italy lacks a phytoleim and its epidermal mor�phology is unknown. Several more Tertiary specieswere later described on the basis of epidermal mor�phology: G. spitsbergensis Manum from the Paleoceneof Spitsbergen and G. wyomingensis Manum from thePaleocene of North America (Manum, 1966), G. tza�gajanica Samylina from the Paleocene of the AmurRegion, G. occidentalis Samylina from the Miocene of
Variability in Epidermal Characters of Ginkgo tzagajanica Samylina (Ginkgoales) from the Paleocene of the Tsagayan Formation
(Amur Region) and the Taxonomy of Tertiary Species of GinkgoL. B. Golovneva
Komarov Botanical Institute, Russian Academy of Sciences, ul. Professora Popova 2, St. Petersburg, 197376 Russiae�mail: [email protected]
Received December 23, 2008
Abstract—The study of new material on Ginkgo tzagajanica Samylina and earlier published data has shownthat the epidermal morphology of the species in highly variable; several new characters were found. Occur�rence of different states of characters was analyzed, their frequency distribution was revealed, and typical andrare states of characters within variation rows were determined. The specific diagnosis of G. tzagajanica wasemended. The most variable characters are degree of undulation of the anticlinal walls of cells of abaxial andadaxial epidermises (the latter is more variable) and degree of development and number of papillae on ordi�nary epidermal cells. Less variable characters are degree of development of papillae on subsidiary cells of sto�mata and how the papillae cover the stomatal apertures. These are characters that bear maximal diagnosticsignificance for Cenozoic Ginkgo.
Key words: Ginkgo, cuticle, morphological variability, Tertiary.
DOI: 10.1134/S003103011005014X

PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
VARIABILITY IN EPIDERMAL CHARACTERS OF GINKGO TZAGAJANICA SAMYLINA 585
northern Caucasus Mountains and Trans�CarpathianRegion (Samylina, 1967), G. tatjanae Samylina etChelebaeva from the Late Eocene of the KamchatkaPeninsula and G. oishii Samylina from the Paleoceneof Japan (Samylina and Chelebaeva, 1986), andG. kamtschatica Budantsev from the Paleocene–EarlyEocene of the Kamchatka Peninsula (Budantsev,1983).
Unlike Cretaceous and Jurassic species of Ginkgo,Cenozoic members of the genus are characterized bymuch lesser developed epidermal and cuticular struc�tures, such as hairs, papillae, and cuticular thickenings(Samylina, 1967). This means that the amount ofqualitative characters and their combination is quitelimited. Cenozoic species of Ginkgo differ by bothqualitative and quantitative characters. As the numberof studied specimens of different species increased,many epidermal characters were shown to vary bothwithin a particular leaf and among different specimensfrom different localities. Some scientists concludedthat the diagnostic value of epidermal charactersshould be reevaluated and the taxonomic position ofsome species should be changed.
Thus, Boulter and Kva ek (1989) showed that thesizes of papillae on the adaxial epidermis vary inG. gardneri and united this species with G. spitsbergen�sis. Denk and Velitzelos (2002) united G. spitsbergensis,G. wyomingensis, G. tzagajanica, G. occidentalis, G. orien�talis, and G. florinii into the single species G. adian�toides mostly basing on their study of the epidermis inmodern Ginkgo biloba.
As a result, the diagnostic significance of nearly allmain epidermal characters of the Ginkgoales wasdoubted, such as the presence of papillae on the adaxialepidermis, degree of undulation of the anticlinal wallsof cells, and development of papillae on ordinary cellsof the adaxial epidermis and on subsidiary cells of sto�mata. The range of variability of G. adiantoides in termsof Denk and Velitzelos (2002) is so high that overlapsconsiderably with many Mesozoic species, also ques�tioning their taxonomic independence. By this, specieswith huge stratigraphic and geographic ranges and avery wide variability spectrum are introduced into pale�obotany. Epidermal analysis is practically discreditedfor the taxonomy of fossil Ginkgoales.
To evaluate the significance of the variability of epi�dermal characters for the taxonomy of Ginkgo withentire�margined leaves, a detailed study of the epider�mis in G. tzagajanica has been undertaken. G. tzaga�janica is one of the key species in the taxonomy of Ter�tiary members of Ginkgo. The assemblage of its char�acters is transitional between that of G. gardneri, whichis closer to Cretaceous species, and Miocene–Pliocene species, which are closer to modernG. biloba.
1 c
^
MATERIAL
The variability of epidermal characters of G. tzaga�janica was analyzed on the basis of newly collectedmaterials as well as published data. The type materialof G. tzagajanica comes from the deposits of the Tsa�gayan Formation, in the south of the Amur Region. Aspecimen that was described as the holotype(Samylina, 1967) was taken from a collection amassedby Konstantov, who published the first monograph onthe Tsagayan flora (Konstantov, 1914). The collectioncomes from the classic locality Belaya Gora on theBureya River and is currently kept at Central Scien�tific�Research Geological Exploration Museumnamed after academician F. N. Chernyshev (CNIGRMuseum, no. 370). The Tsagayan flora was character�ized in more detail by Kryshtofovich and Baikovskaya(1966). The latest monograph study of the flora wasaccomplished by Krassilov (1976). These authorsdated the flora to the Early Paleocene (Danian).
Remains of Ginkgo studied by Krassilov come fromthe same locality on the Bureya River as specimenscollected by Konstantov and are kept at the Institute ofBiology and Soil Sciences of the Russian Academy ofSciences, in Vladivostok. Krassilov (1976) believedthat the Tsagayan leaves were identical to leaves ofG. spitsbergensis by their epidermal characteristics andconsidered G. tzagajanica as a synonym of G. spitsber�gensis.
The material under present study comes from twolocalities in the Amur Region. Impressions of leaves ofGinkgo with preserved cuticle were collected byS. Manchester and were handed over to the presentauthor for this study. Currently, they are kept at themuseum of the University of Florida (FSU). In addi�tion, several specimens of Ginkgo were found in thecoal quarry “Pioner”, near the town of Raichikhinsk.These finds come from the lower part of Kivda beds,which are considered as the upper member of the Tsa�gayan Formation by some authors or the Kivda For�mation by others (Krassilov, 1976). This material iskept at the Botanical Institute of the Russian Academyof Sciences, in St. Petersburg (BIN, no. 1537). ThePaleocene flora of the Kivda beds from the suburb ofthe town of Raichikhinsk was studied by Kamaeva(1990) and Akhmetiev et al. (2002).
VARIABILITY OF EPIDERMAL CHARACTERS IN GINKGO TZAGAJANICA
FROM THE TSAGAYAN FORMATION
The variability of epidermal characters in G. tzaga�janica was mostly studied on cuticle specimens takenfrom central areas of leaf blades. The epidermal mor�phology of the leaf margin and near petiole was studiedby Samylina (1967).
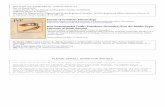
The leaves are hypostomous. The adaxial epidermisis indistinctly differentiated into costal and intercostalzones, with a gradual transition between them (Pl. 13,

586
PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
GOLOVNEVA
figs. 1, 2, 8). As a rule, bands of four to nine rows ofelongated tetra� or polygonal cells are situated overveins. The width of cells in the costal zone is 13–18 µm,and the length varies from 60 to 100 µm. The width ofthe costal zone is 100–170 µm. The intercostal zonesare wider, up to 200–350 µm. They include up to 10–19 rows of isodiametric or slightly elongated, irregu�larly tetragonal or irregularly pentagonal cells. Thewidth is up to 20–30 µm, and the length is 25–60 µm.The anticlinal walls of cells in the costal zones are usu�ally straight or, more rarely, weakly undulated, or, evenmore rarely, undulated (Pl. 13, figs. 1, 2, 6–8). Therange of the undulations is about 2–3 µm. The anticli�nal walls in intercostal zones are usually weakly undu�lated (Pl. 13, fig. 4); however, areas with nearly straightor undulated walls occur (Pl. 13, fig. 5). Although dis�tinct undulations characterize marginal areas of theleaf, undulations also occur in the central part of theleaf. In general, undulations are more distinct in inter�costal zones and near the upper margin of the leaf. Thepericlinal walls of cells are slightly convex. Many cellsof intercostal zones bear small dome�shaped centralcuticular thickenings, 12–20 µm in diameter and 5–10 µm in height (Pl. 13, figs. 7–10). Cells of the costalzones may bear central dome�shaped thickenings orless distinct longitudinal thickening stretched alongthe entire cell (Pl. 13, figs. 8, 9). The amount anddevelopment of thickenings vary considerably from anarea to an area of the leaf; thickenings are non�uni�form even within the same area (Pl. 13, fig. 10). Somespecimens are virtually devoid of thickenings and havenearly smooth or slightly undulating cuticle surface(Pl. 13, figs. 5, 6).
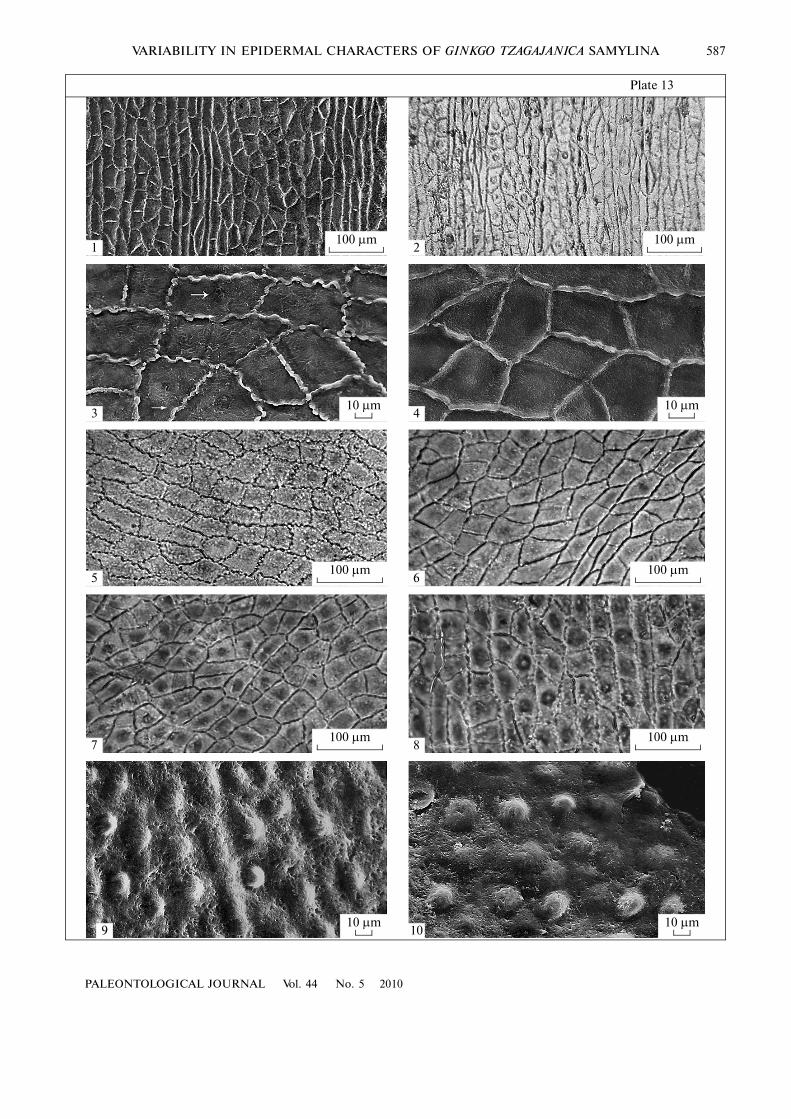
The costal/intercostal transition is sharp on theabaxial epidermis (Pl. 14, figs. 1, 2). The anticlinalwalls of cells are less distinct as well as boundariesbetween the cells. Where visible, the anticlinal wallsare usually straight (Pl. 14, figs. 5, 6, 10). Undulatedanticlinal walls were observed very rarely (Pl. 14,fig. 9). The periclinal walls of cells are usually convexor, more rarely, smooth. Cells of the costal zone havepericlinal walls that are slightly thickened along theentire length (Pl. 14, fig. 10); they also may bear asmall dome�shaped rounded thickening in the center
or a central hollow thin�walled papilla. The periclinalwalls of cells of the intercostal zone usually bear a dis�tinct papilla up to 10–15 µm high (Pl. 14, figs. 4, 8).More rarely, they bear a dome�shaped thickening(Pl. 14, fig. 3; Fig. 1f) and, even more rarely, hairlikepapillae or hairs up to 30–60 µm high (Pl. 14, fig. 9;Figs. 1a–1d). Some cells are completely devoid ofpapillae or thickenings (Pl. 14, fig. 10). In most speci�mens, papillae prevail and rare cuticular thickeningsare distributed more or less regularly. By the contrary,leaves with hairlike papillae have irregularly developedcuticular structures. All types of structures, fromthickenings to hairlike papillae and hairs, may occur ina given area (Pl. 13, fig. 9; Fig. 1b).
Stomata are situated and orientated irregularly(Pl. 14, fig. 4), without formation of regular rows.There are approximately up to five to seven stomataper a width of the intercostal zone (Pl. 14, figs. 1, 2).Contiguous stomata are occasionally present (Pl. 14,figs. 5, 6). The stomata are surrounded by five to sevensubsidiary cells. Occasionally, encircling cells sur�round the stoma with a ring, which can be incomplete(Pl. 14, fig. 5). Each subsidiary cell bears a proximalpapilla up to 15–25 µm long (Figs. 1g, 1h). Usually,these papillae completely cover the stoma (Pl. 14,fig. 8; Fig. 1e). More rarely, the papillae do not coverthe stoma completely and the latter is partially visible(Pl. 14, fig. 3; Fig. 1f). Open stomata surrounded bysmall papillae about 20–30 µm long are solitary(Pl. 14, fig. 7).
The following variant of the epidermal morphologyis most typical and common. The adaxial epidermis isdevoid of stomata; cells are with weakly undulatedanticlinal walls and weakly thickened periclinal walls.Cells of the abaxial epidermis are with smooth anticli�nal walls and a distinct papilla or cuticular thickeningson the periclinal wall. The stomata are surrounded bya ring of subsidiary cells with distinct papillae coveringthe stoma.
The most variable characters are the degree ofundulations of the anticlinal walls of cells of the adax�ial epidermis and degree of development of cuticularstructures on their periclinal walls. This variability isnearly always observed within a particular leaf, on its
E x p l a n a t i o n o f P l a t e 1 3
Figs. 1–10. Morphology of the adaxial epidermis of Ginkgo tzagajanica Samylina, Tsagayan Formation, Amur Region, Paleocene:(1) anticlinal walls of cells are straight and weakly undulating in costal zones and undulating and weakly undulating in intercostalzones, FSU, no. 49442, LM, inside view; (2) anticlinal walls of cells are straight in costal zones and undulating and weakly undu�lating in intercostal zones, some cells of intercostal zones have rounded cuticular thickenings on the periclinal walls, FSU,no. 49442, LM; (3) undulating anticlinal walls of cells of intercostal zone and scars corresponding to cuticular thickenings(arrows), BIN, no. 1537�39, SEM, inside view; (4) weakly undulating anticlinal walls of cells of intercostal zone, BIN, no. 1537�43,SEM, inside view; (5) undulating anticlinal walls of cells in costal and intercostal zones and periclinal walls devoid of cuticularthickenings, BIN, no. 1537�39, LM; (6) weakly undulating anticlinal walls of cells in costal and intercostal zones and minorcuticular thickenings of periclinal walls in some cells, BIN, no. 1537�39, LM; (7) weakly undulated and straight anticlinal wallsin the intercostal zone and periclinal walls with moderately developed rounded cuticular thickenings, BIN, no. 1537�39; (8) cellsof the intercostal zone showing undulated anticlinal walls, periclinal walls with distinct rounded cuticular thickenings, anticlinalwalls of cells in the costal zones are weakly undulated or straight, periclinal walls with cuticular rims, BIN, no. 1537�39, LM;(9) distinct rounded cuticular thickenings in the intercostal zone and cuticular rims in the costal zone, FSU, no. 49442, SEM,outside view; (10) variously developed cuticular thickenings, BIN, no. 1537�43, SEM, outside view.
PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
VARIABILITY IN EPIDERMAL CHARACTERS OF GINKGO TZAGAJANICA SAMYLINA 587
Plate 13
100 µm1
100 µm
10 µm10 µm3 4
2
5 6100 µm 100 µm
100 µm100 µm
10 µm 10 µm
7 8
9 10
588
PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
GOLOVNEVA
Plate 14
1100 µm 100 µm
10 µm10 µm(а) (а)
(b)
(b)
10 µm 10 µm
10 µm10 µm
100 µm 100 µm
2
43
5 6
7 8
9 10

PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
VARIABILITY IN EPIDERMAL CHARACTERS OF GINKGO TZAGAJANICA SAMYLINA 589
E x p l a n a t i o n o f P l a t e 1 4
Figs. 1–10. Abaxial epidermis of Ginkgo tzagajanica Samylina, Tsagayan Formation, Amur Region, Paleocene: (1, 2) general viewof the cuticle from outside (1) and inside (2), note costal zones with elongated cells and intercostal zones with stomata and papil�lae, FSU, no. 49443, SEM; (3) stomata completely (3a) or incompletely (3b) covered with papillae, ordinary epidermal cells withpapillae and cuticular thickenings, FSU, no. 49443, SEM, outside view; (4) stomata orientated under different angles and papil�lae on subsidiary cells of stomata and ordinary cells of the intercostal zones, BIN, no. 1537�43, SEM, inside view; (5) contiguousstomata, FSU, no. 49443, SEM, inside view; (6) contiguous stomata, note papillae on ordinary cells of the intercostal zone andstraight anticlinal walls of cells in the costal zone, FSU, no. 49443, SEM, inside view; (7) open stoma with weakly developedpapillae on the subsidiary cells and papillae on ordinary cells of the intercostal zone, BIN, no. 1537�39, LM; (8) stoma coveredwith densely contacting papillae, note papillae on ordinary cells of the intercostal zone, BIN, no. 1537�39, LM; (9) undulatinganticlinal walls of cells and variously developed papillae on ordinary epidermal cells, varying from normal to elongated and tohairlike, BIN, no. 1537�39, LM; (10) costal zone with elongated cells with cuticular rims and intercostal zone with stomata cov�ered with papillae, not ordinary cells with straight anticlinal walls, most of them without papillae or cuticular rims, FSU,no. 49442, LM.
different areas, but may vary from a specimen to aspecimen. The most variable character of the abaxialepidermis is the development of papillae on ordinaryepidermal cells of the intercostal zone. Usually, differ�ent combinations of papillae and cuticular thickeningsare observed. Hairlike papillae were not detected in allspecimens and occur only in some areas.
COMPARISON BETWEEN GINKGO TZAGAJANICA FROM THE TSAGAYAN FORMATION
AND OTHER LOCALITIES BY EPIDERMAL MORPHOLOGY
Outside the Amur Region, Ginkgo tzagajanica wasreported from the Minato Formation of the NodaGroup, Japan (Horiuchi and Kimura, 1986). The for�mation is dated to the Late Paleocene–Early Eoceneby palynological data. The authors studied about60 leaves focusing on the variability of epidermal char�acters in different areas of the leaf blade. By mostcharacters, the Japan leaves are very similar to Tsa�gayan leaves. However, their adaxial epidermis hasgreater undulations of the anticlinal walls of ordinarycells. Straight or weakly undulated walls only occur incostal zones. Cells of intercostal zones are character�ized by distinctly undulated walls. Such an importantcharacter as cuticular thickenings on the periclinalwalls of cells of the adaxial epidermis was not illus�trated, although rare occurrence of such thickeningswas mentioned in the text. In return, solitary stomatawere detected at the leaf blade base, near the petiole.In general, the anticlinal walls of cells at the leaf bladebase and near the margin are undulated in a greaterdegree.
The morphology of the abaxial epidermis in theJapan leaves is identical to that in the Tsagayan leaves.The authors noticed that papillae are irregularly dis�tributed in different areas of the leaf: they may bepresent on nearly each ordinary cell or may be virtuallyabsent; cuticular structures also vary from cuticularthickenings to ordinary and hairlike papillae. Thepresence of peglike hairs with thickened apices is alsoobserved (Horiuchi and Kimura, 1986, pl. 67, fig. 5).I believe that these structures are lower parts of ordi�nary broken hairs or hairlike papillae.
SYSTEMATIC PALEOBOTANY
The study of the new material of G. tzagajanica aswell as previously published descriptions and depic�tions has shown that the species is highly variable by itsepidermal morphology; several characters wererevealed for the first time. Occurrence of differentstates of characters was analyzed, their frequency dis�tribution was revealed, and typical and rare states ofcharacters within variation rows were determined. Thespecific diagnosis of G. tzagajanica was emended.
Family Ginkgoaceae Engler in Engler et Prantl, 1897
Genus Ginkgo L., 1771Ginkgo tzagajanica Samylina, 1967
Ginkgo tzagajanica: Samylina: Samylina, 1967, p. 314, pl. 4,figs. 3–9; Horiuchi and Kimura, 1986, p. 344, pl. 65, pl. 66, pl. 67,figs. 1, 3–6, pl. 68, text�figs. 2a–2d, 3c–3j.
G. oishii Samylina: Samylina and Chelebaeva, 1986, p. 100.G. adiantoides auct. non (Unger) Heer: Konstantov, 1914,
p. 15, pl. 4, figs. 2, 5, 6; Kryshtofovich and Baikovskaya, 1966,p. 227, pl. 1, fig. 1.
G. spitsbergensis auct. non Manum: Krassilov, 1976, p. 42,pl. 2, figs. 1–7.
Ginkgoites cf. adiantoides auct. non (Unger) Seward: Oishi,1938, p. 104, pls. 1–6, figs. A, B, C.
H o l o t y p e. CNIGR Museum, no. 370/18,Amur Region, Bureya River, Belaya Gora locality,Tsagayan Formation, Paleocene (specimen collectedby Konstantov, 1914, pl. 4, fig. 6).
E m e n d e d d i a g n o s i s. Leaves usually 6–9 cmlong and 4–10 cm wide, broadly fan�shaped, nearlyrounded or, more rarely, cuneate, entire or bilobed,with median incision never reaching halfway�downlamina. Base cuneate with basal angle 70° to 330°.Apical margin of lamina rounded, undulated, oftenwith several shallow incisions. Venation dichotomous;vein density 12–24 per 1 cm.
Leaves hypostomous, occasionally with rare sto�mata in marginal areas of adaxial epidermis. Adaxially,transition between costal and intercostal zones is grad�ual. Costal zones consist of four to nine rows of elon�gated tetra� or polygonal cells. Intercostal zones widerand contain up to 10–19 rows of isodiametric orslightly elongated irregularly tetragonal or polygonalcells. Anticlinal cell walls from straight to undulated
590
PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
GOLOVNEVA
100 µm 100 µm
10 µm10 µm
10 µm 10 µm
10 µm10 µm
(а) (b)
(d)(c)
(e) (f)
(g) (h)
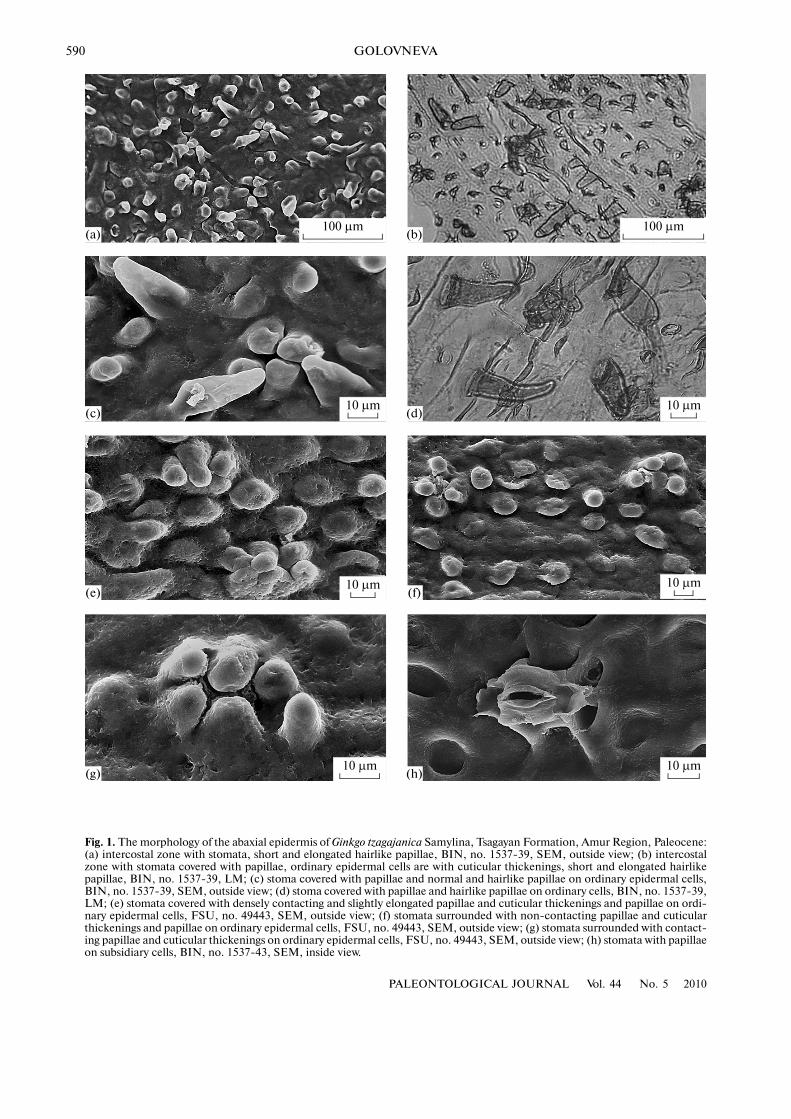
Fig. 1. The morphology of the abaxial epidermis of Ginkgo tzagajanica Samylina, Tsagayan Formation, Amur Region, Paleocene:(a) intercostal zone with stomata, short and elongated hairlike papillae, BIN, no. 1537�39, SEM, outside view; (b) intercostalzone with stomata covered with papillae, ordinary epidermal cells are with cuticular thickenings, short and elongated hairlikepapillae, BIN, no. 1537�39, LM; (c) stoma covered with papillae and normal and hairlike papillae on ordinary epidermal cells,BIN, no. 1537�39, SEM, outside view; (d) stoma covered with papillae and hairlike papillae on ordinary cells, BIN, no. 1537�39,LM; (e) stomata covered with densely contacting and slightly elongated papillae and cuticular thickenings and papillae on ordi�nary epidermal cells, FSU, no. 49443, SEM, outside view; (f) stomata surrounded with non�contacting papillae and cuticularthickenings and papillae on ordinary epidermal cells, FSU, no. 49443, SEM, outside view; (g) stomata surrounded with contact�ing papillae and cuticular thickenings on ordinary epidermal cells, FSU, no. 49443, SEM, outside view; (h) stomata with papillaeon subsidiary cells, BIN, no. 1537�43, SEM, inside view.

PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
VARIABILITY IN EPIDERMAL CHARACTERS OF GINKGO TZAGAJANICA SAMYLINA 591
with amplitude up to 2–3 µm; usually straight orslightly undulated in costal zones and slightly undu�lated in intercostal zones. Sinuosity is more distinct inintercostal zones and near upper margin of lamina. Per�iclinal walls slightly convex, thin, or with cuticular thick�enings. Cuticular thickenings in intercostal zones dome�like, 12–20 µm in diameter and 5–10 µm in height;cuticular thickenings in costal zones dome� or rimlike.Number and development of cuticular thickenings varyconsiderably in different areas of leaf lamina.
Transition between costal and intercostal zones issharp in abaxial epidermis. Anticlinal cell wallsstraight or, very rarely, undulated. Periclinal walls incostal zones thin or with dome� or rimlike cuticularthickenings. Periclinal walls in intercostal zones usu�ally with papilla up to 10–15 µm high, or with dome�shaped cuticular thickening, or, more rarely, smooth.Occasionally, thin�walled hairlike papillae or hairs upto 30–60 µm high occur, most often near margin orpetiole.
D e s c r i p t i o n. Leaf blade is broadly fan�shaped,nearly rounded or, more rarely bilobed, with roundedundulating upper margin, usually 6–9 cm long and 4–10 cm. Lobes of the leaf blade form the basal anglefrom 70° to 330°. The medial incision is no deeperthan a one half of the leaf blade length; several shallowincisions may be additionally present. The venation isdichotomous. The veins are thin and densely situated:12–24 veins occur per 1 cm.
Leaves are hypostomous, occasionally with rarestomata in marginal areas of the adaxial epidermis.Adaxially, transition between costal and intercostalzones is gradual. The costal zones consist of four tonine rows of elongated tetra� or polygonal cells. Theintercostal zones are wider and contain up to 10–19 rows of isodiametric or slightly elongated, irregu�larly tetragonal or polygonal cells. The anticlinal cellwalls vary from straight to undulated with amplitudeup to 2–3 µm. They are usually straight or slightlyundulated in the costal zones and slightly undulated inthe intercostal zones. Sinuosity is more distinct in theintercostal zones and near the upper margin of thelamina. The periclinal walls are slightly convex, thin,or with cuticular thickenings. The cuticular thicken�ings in intercostal zones are domelike, 12–20 µm indiameter and 5–10 µm in height; cuticular thicken�ings in the costal zones dome� or rimlike. Number anddevelopment of cuticular thickenings vary consider�ably in different areas of leaf lamina.
The transition between costal and intercostal zonesis sharp in the abaxial epidermis. The anticlinal cellwalls are straight or, very rarely, undulated. The peri�clinal walls in the costal zones are thin or with dome�or rimlike cuticular thickenings. The periclinal walls inthe intercostal zones are usually with a papilla up to10–15 µm high, or with a dome�shaped cuticularthickening, or, more rarely, smooth. Occasionally,
thin�walled hairlike papillae or hairs up to 30–60 µmhigh occur, most often near the leaf margin or petiole.
C o m p a r i s o n. Among species of Ginkgo, G. tza�gajanica show the maximal similarity to G. spitsbergen�sis, being considered as a synonym of the latter bysome authors (Krassilov, 1976). To date, G. tzagajanicais the most thoroughly studied Cenozoic species of thegenus: several dozens of specimens have been studiedwith help of scanning electron microscope and lightmicroscope. G. spitsbergensis has been studied in lessdetail, the description of its epidermal morphology isbased on few specimens studied in transmitted light(Manum, 1966).
Leaves of G. spitsbergensis are hypostomous. Cellsof the adaxial epidermis of G. spitsbergensis are withstraight or weakly undulating anticlinal walls and,more rarely, undulated walls. No cuticular thickeningswere detected on the periclinal walls. The abaxial epi�dermis is similar to that of G. tzagajanica, but papillosecells are usually less abundant, although there areareas with numerous papillae. Hairlike papillae orhairs were not found.
The known epidermal characters of G. spitsbergen�sis are within the range of variability of G. tzagajanica.However, cuticular structures are developed on bothepidermises of G. spitsbergensis in a lesser degree, andthe range of variability of all characters is much nar�rower. Some characteristic features of G. tzagajanicawere not observed in G. spitsbergensis, such as rare sto�mata and cuticular thickenings on the adaxial cuticleand hairlike papillae on the abaxial cuticle. There is apossibility that further study of G. spitsbergensis withapplication of a scanning electron microscope willreveal additional characters which will increase ordiminish the closeness between the two species. Onemore close species is G. wyomingensis. According toManum (1966, fig. 2B), it differs by strongly undulatedanticlinal walls of the adaxial epidermis.
By the morphology of the abaxial epidermis, G. tza�gajanica also resembles G. gardneri, although thenumber of papillae in G. gardneri is higher. However,the adaxial epidermis of G. gardneri differs from that ofG. tzagajanica by straight anticlinal walls of cells anddistinct papillae on the periclinal walls or, more rarely,cuticular thickenings (Boulter and Kva ek, 1989). Thepresence of distinct cuticular thickenings on the adax�ial epidermis is a character of many Mesozoic species.Among Cenozoic species of Ginkgo, they were onlyknown in G. gardneri, thus differentiating this speciesfrom other species. The find of cuticular thickenings inG. tzagajanica shows its similarity to G. gardneri andapparently testifies to the antiquity of the species.
Several trends were revealed in the epidermal mor�phology of species of Ginkgo during the Cenozoic(Samylina, 1967; Tralau, 1968; Horiuchi and Kimura,1986). These are less and less developed cuticularthickenings first on the adaxial epidermis and than on
1c
^

592
PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
GOLOVNEVA
the abaxial epidermis; diminishing papillae on ordi�nary cells of stomata covering the stomatal aperture ina lesser and lesser degree. In this row, G. tzagajanica isa transitional chain between G. gardneri, which haspapillae on both adaxial and abaxial epidermises, andG. spitsbergensis and G. wyomingensis, which havepapillae only on the abaxial epidermis. G. tzagajanicaoccasionally has irregular and variously developedcuticular thickenings on the adaxial epidermis. If suchthickenings are lacking, the epidermis of G. tzagajan�ica can be virtually identical to the epidermis ofG. spitsbergensis. However, the epidermis of G. spits�bergensis is currently insufficiently studied to be thor�oughly compared with G. tzagajanica; therefore, thesetwo species are so far considered as close but indepen�dent species.
Apart from G. tzagajanica, deposits of the MinatoFormation also contain compressions of Ginkgoites cf.adiantoides (Unger) Seward (Oishi, 1938). The adaxialsurface of these specimens bears cells with undulateanticlinal walls in both costal and intercostal zones,the periclinal walls lack any cuticular structures, andthe abaxial surface is characterized by straight anticli�nal walls of cells, distinct papillae in intercostal zones,papillae on subsidiary cells completely covering theaperture. Solitary stomata were found on the adaxialepidermis near the leaf margin. Samylina believed thatthese leaves were amphistomous and described on thisbasis a new species, G. oishii Samylina (Samylina andChelebaeva, 1986). However, the epidermal charac�ters of these leaves fit the characteristics of Ginkgo tza�gajanica, and I consider G. oishii as a synonym ofG. tzagajanica.
All listed�above species come from Paleocene orEocene deposits. They are characterized by a similar
morphology of the abaxial epidermis and differ indetails of the adaxial epidermis. Other, younger, spe�cies differ from G. tzagajanica by both adaxial andabaxial morphology. They have papillae developedaround guard cells of stomata in a much lesser degree,which results in an open stomatal aperture, and asmaller number of papillae on ordinary cells or eventhe absence of papillae.
In particular, leaves of Ginkgo from the Miocene ofGreece, which were identified by Denk and Velitzelos(2002) as G. tzagajanica, are characterized by stronglyundulated (with a range up to 5–6 µm in comparisonto 2–3 µm in G. tzagajanica) anticlinal walls of theadaxial epidermis and the complete absence of anystructures on the periclinal walls. The abaxial epider�mis of the Greek leaves bears less numerous papillaethan G. tzagajanica. Papillae on ordinary cells and nearstomata are shorter. Papillae never contact over sto�mata; completely open stomata are common. The epi�dermal characters of the leaves from the Miocene ofGreece answer the diagnosis of G. occidentalis from theMiocene of Trans�Carpathian Region and northernCaucasus Mountains.
O c c u r r e n c e. Tsagayan Formation of the AmurRegion, Russia; Minato Formation, Honshu Island,Japan. Paleocene–Eocene.
M a t e r i a l. BIN, no. 1537/39, 1537/43, FSU,no. 49442, FSU, no. 49443.
CONCLUSIONS
The epidermal characters of G. tzagajanica are veryvariable. The most variable character is the degree ofsinuosity of the anticlinal walls on the abaxial epider�mis and, particularly, on the adaxial epidermis. The
Anticlinal walls of cellsCharacters
straight weakly undulated undulated
absent
cuticular thickenings
papillae
Str
uctu
res
G. tzagajanica G. spitsbergensis G. wyomingensis G. gardneri
on p
eric
lin
al w
alls
Fig. 2. Variability spectra in characters of the adaxial epidermis overlapping in Paleocene–Eocene members of Ginkgo.

PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
VARIABILITY IN EPIDERMAL CHARACTERS OF GINKGO TZAGAJANICA SAMYLINA 593
degree of development and number of papillae onordinary cells of the upper and lower epidermis is alsosignificantly diverse. The most constant character isthe degree of development of papillae on subsidiarycells of stomata and how they cover the stomatal aper�ture. This is the character that bears the maximal diag�nostic significance for Tertiary species of Ginkgo. Mostcharacters vary not only from a specimen to a speci�men, but also within a particular specimen, from onearea to another.
Among all studied Cenozoic species of Ginkgo,G. tzagajanica has the most variable epidermis. It is sofar unclear if this reflects the fact that this species hasbeen so far studied in maximal detail or this variabilityrepresents its specific character. In spite of its greatvariability range, variation rows of all characters ofG. tzagajanica have distinct frequency extrema, thatenables one to provide the morphological characteris�tics of the species on the basis of the most typical statesof its characters.
I believe that the revealed variability does notdevaluate the diagnostic significance of epidermalcharacters, since their variation is not a randomassemblage of different states. The variability range ofdifferent states and the frequency of a given state of acharacter are important additional characters of thespecies; and even partial overlapping between varia�tion rows of different species should not be always con�sidered as a reason to unite these species.
Keeping in mind the limited number of availablediagnostic epidermal characters in Cenozoic speciesof Ginkgo, I incline that fusion of species is justified ifnot only a particular combination of characters coin�cides in the species under comparison, but also theentire spectra of their variability coincide. The consid�erable variability in some species does not necessarilymean that a similar variability exists in other species.Thus, Denk and Velitzelos (2002) included into onespecies six Cenozoic species of Ginkgo on the basis oftheir study of the epidermal morphology in G. biloba.However, such broad variability was not revealed inany of these fossil species. To revise the taxonomicposition of already described species, detailed study isneeded of the epidermis and variability spectra ofcharacters in all species under comparison.
ACKNOWLEDGMENTS
The study was financially supported by the RussianFoundation for Basic Research, project no. 07�04�01227, and Program of the Presidium of the RussianAcademy of Sciences “Origin and Evolution of theBiosphere.”
REFERENCES
1. M. A. Akhmetiev, T. V. Kezina, T. M. Kodrul, andS. R. Manchester, “Stratigraphy and Flora of the Cre�taceous–Paleogene Boundary Beds in the SoutheasternPart of the Zeya–Bureya Sedimentary Basin,” in Col�lection of Papers in Memory of Corresponding Member ofthe Russian Academy of Sciences Prof. V.A. Vakhrameevon Occasion of His 90th Birthday (GEOS, Moscow,2002), pp. 275–315 [in Russian].
2. M. C. Boulter and Z. Kva ek, “The Paleocene Flora ofthe Isle of Mull,” Spec. Pap. Palaeontol. 42, 1–149(1989).
3. L. Yu. Budantsev, History of the Arctic Flora of the EarlyCenophytic Epoch (Nauka, Leningrad, 1983) [in Rus�sian].
4. T. Denk and D. Velitzelos, “First Evidence of Epider�mal Structures of Ginkgo from the Mediterranean Ter�tiary,” Rev. Palaeobot. Palynol. (120), 1–15 (2002).
5. R. Florin, “Die fossilen Ginkgophyten von Franz�Joseph�Land nebst Erörterung über vermeintlich Cor�daitales mesozoischen Alters. I. Spezieller Teil,” Palae�ontogr. B 81 (3–6), 71–173 (1936).
6. R. Florin, “Die fossilen Ginkgophyten von Franz�Joseph�Land nebst Erörterung über vermeintlich Cor�daitales mesozoischen Alters. II. Algemeiner Teil,”Palaeontogr. B 82 (1–4), 1–72 (1937).
7. T. M. Harris, “The Fossil Flora of Scoresby Sound,Greenland. Part IV,” Medd. om Grønland 112 (1), 1–176 (1935).
8. O. Heer, “Miocene Flora der Insel Sachalin,” Mém.Acad. Imp. Sci. Saint�Pétersb., Ser. 7, 25, 1–61 (1878).
9. R. S. Hill and R. J. Carpenter, “Ginkgo Leaves fromPalaeogene Sediments in Tasmania,” Aust. J. Bot. 47,717–724 (1999).
10. Horiuchi J. and Kimura T., “Ginkgo tzagajanicaSamylina from the Palaeogene Noda Group, NortheastJapan, with Special Reference to Its External Morphol�ogy and Cuticular Features,” Trans. Proc. Palaeontol.Soc. Japan, N. S., No. 142, 341–353 (1986).
11. A. M. Kamaeva, Stratigraphy and Flora of the Creta�ceous–Paleogene Boundary Beds of the Zeya–BureyaDepression (Dal’nevost. Otd. Akad. Nauk SSSR, Kha�barovsk, 1990) [in Russian].
12. S. V. Konstantov, “Tertiary Flora of the Belogorsk Out�crop in the Lower Reaches of the Bureya River,”Tr. Geol. Kom., Nov. Ser., No. 113, 1–27 (1914).
13. V. A. Krassilov, Tsagayan Flora of the Amur Region(Nauka, Moscow, 1976) [in Russian].
14. A. N. Kryshtofovich and T. N. Baikovskaya, “UpperCretaceous Tsagayan Flora in the Amur Region,” inA.N. Kryshtofovich: Selected Papers (Akad. Nauk SSSR,Moscow�Leningrad, 1966), Vol. 3, pp. 184–320[in Russian].
15. S. Manum, “Ginkgo spitsbergensis n. sp. from thePaleocene of Spitsbergen and a Discussion of CertainTertiary Species of Ginkgo from Europe and NorthAmerica,” Norsk Polarinst. Årb. 1965, 49–58 (1966).
1c
^

594
PALEONTOLOGICAL JOURNAL Vol. 44 No. 5 2010
GOLOVNEVA
16. Oishi S., “On the Cuticles of Tertiary GinkgoitesLeaves from Kuzi, Iwate Prefecture,” J. Fac. Sci. Hokk.Univ. Ser. IV 4 (1–2), 103–106 (1938).
17. M. S. Pole and J. G. Douglas, “Bennettitales, Cycadales,and Ginkgoales from the Mid�Cretaceous of the Ero�manga Basin, Queensland, Australia,” Cret. Res. 20,523–538 (1999).
18. V. A. Samylina, “On the Final Stages of the History ofthe Genus Ginkgo L. in Eurasia,” Bot. Zh. 52 (3), 303–316 (1967).
19. V. A. Samylina and A. I. Chelebaeva, “New Data on theTertiary Species of Ginkgo in Soviet Eastern Asia,”Paleontol. Zh., No. 2, 97–102 (1986).
20. H. Tralau, “Evolutionary Trends in the Genus Ginkgo,”Lethaia 1, 63–101 (1968).
21. F. Unger, Synopsis Plantarum Fossilium (Leopold Voss,Leipzig, 1845).
22. Zhou Z., “Mesozoic Ginkgoalean Megafossils: A Sys�tematic Review,” in Ginkgo biloba—A Global Treasure:From Biology to Medicine, Ed. by Hori T. et al.(Springer�Verlag, Tokyo, 1997), pp. 183–206.
SPELL: 1. ok
Copyright © 2022 FDOKUMEN