
Water dew-pointing with subsea gas dehydration to improve ...

Variability and correlations in bi-manual pointingmovements
J. Jasper Boessenkool *, Evert-Jan Nijhof, Casper J. Erkelens
Perceptual-Motor Integration Group, Helmholtz Institute, Utrecht University, P.O. Box 80.000, 3508 TA,
Utrecht, The Netherlands
Abstract
Bi-manual movement tasks present us with an intriguing paradox: on one hand it is clear
that we are capable of controlling our hands independently, yet at the same time we often
notice that one hand in¯uences the other. We investigated bi-manual pointing movement
towards a common target and hypothesised that if (parts of ) bi-manual movements are
planned as one functional unit, then there should be signi®cant correlations between the
movement parameters of each hand. Surprisingly, we found only consistently high correlation
coe�cients with regard to the reaction time. Our results demonstrate that trajectory formation
and adjustments of bi-manual movements are largely independent and hand-speci®c, con-
tradicting the notion that the left and right hand are controlled as one functional unit. Because
we ®nd that average, rather than individual, movements are correlated, we conclude that
coupling between the two hands occurs only at `task level'. In symmetrical tasks, movements
are initiated simultaneously. In asymmetrical tasks, however, there are small but very con-
sistent timing di�erences between the left and right hands. These di�erences are small com-
pared to the variability in reaction times and, therefore, we conclude that they result from an
intrinsic control property of the system of bi-manual movement generation. Ó 1999 Elsevier
Science B.V. All rights reserved.
Human Movement Science 18 (1999) 525±552
www.elsevier.com/locate/humov
* Corresponding author. Tel.: +31-30-253 2273; fax: +31-30-252 2664; e-mail: j.j.boessenkool@phy-
s.uu.nl
0167-9457/99/$ ± see front matter Ó 1999 Elsevier Science B.V. All rights reserved.
PII: S 0 1 6 7 - 9 4 5 7 ( 9 9 ) 0 0 0 2 1 - 4

PsycINFO classi®cation: 2330; 2520
Keywords: Multi-joint arm movements; Trajectory planning; Bi-manual; Motor control;
Human
1. Introduction
Bi-manual movement tasks present us with an intriguing paradox. On theone hand it is clear that we are capable of controlling our hands indepen-dently in sports, music and many daily-life tasks. On the other hand it hasbeen shown that movements of one hand are in¯uenced by movements of theother (Kelso, Southard & Goodman, 1979; Marteniuk & MacKenzie, 1980;Kelso, 1984). The later ®nding is often construed in terms of a commonorganisation principle that involves the control of both hands simulta-neously.
One way of investigating bi-manual behaviour is derived from rhythmicaltasks. Coupling between the movements of the left and right hand can clearlybe observed in a task introduced by Cohen (1971) and elaborated by Kelso(1984). When subjects tried to rhythmically ¯ex and extend their index ®ngersin anti-phase (i.e. one ®nger extends while the other ¯exes), they were able tomaintain this out-of-phase mode for low frequencies only. With increasingfrequencies, subjects switched, abruptly and involuntarily, to an in-phasemode in which homologous muscles were activated simultaneously. At higherfrequencies the symmetrical (in-phase) mode was found to be a more stableco-ordination pattern.
At the behavioural level, synergetic models have been used to assess thedegree of coupling between the hands. If movements of the left and righthand are described in terms of two oscillators, rhythmical behaviour can becharacterised by their relative phase relationship. Next, coupling betweenthe hands can be modelled with a non-linear interaction between these os-cillators and preferred co-ordination patterns in terms of minima of a po-tential function of the relative phase (Haken, Kelso & Bunz, 1985; Peper,Beek & van Wieringen, 1995). Although these models capture several in-teresting features of rhythmical behaviour, their interpretation in terms ofneurological structures is not clear. Moreover, it is di�cult to extend thesemodels to discrete movements (see Sch�oner (1990) for an attempt in onedimension) because the coupling between the hands is modelled as a po-
526 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

tential function of the relative phase between the movements of the twohands.
Another way of identifying the functional units of bi-manual movementorganisation is to focus on the interference in discrete bi-manual tasks. Thisapproach was inspired by the ®nding that in asymmetrical tasks, movementsof the two hands assimilate with the result that the characteristics of move-ments in bi-manual tasks di�er from those of equivalent movements in uni-manual conditions. In bi-manual pointing tasks with prescribed accuracy(target width), for instance, Kelso et al. (1979) found that subjects had astrong tendency to move their hands simultaneously. Even in conditions withunequal target widths, subjects were inclined to synchronise hand move-ments, thereby violating the well-known relationship between movementtime, amplitude and target width of (uni-manual) hand movements, knownas Fitts' Law (Fitts, 1954).
Notwithstanding the fact that both hands can move independently, it hasbeen suggested that control is simpli®ed by a task-dependent ``functionalgrouping of muscles'' (Kelso et al., 1979). Following the rationale putforward by Bernstein (1967), one can hypothesise that in order to controlthe redundancy of the movement system, co-ordination is achieved bycontrol of task-dependent synergies (Kugler, Kelso & Turvey, 1980). Withrespect to bi-manual movements on obvious assumption is that both handsare constrained ``to function as a single, synergistic unit within whichcomponent elements vary in a related manner (Kelso, Putnam & Goodman,1983)''. Others, however, have emphasised that di�erences between themovements of each hand can be observed when intrinsic movement times ofthe two hands become too dissimilar (Marteniuk & MacKenzie, 1980;Marteniuk, MacKenzie & Baba, 1984; Fowler, Duck, Mosher & Mathieson,1991).
It is di�cult to capture bi-manual interference, as found in discrete tasks,in a formal model but one is tempted to assume ``a unitary pattern of acti-vation (or program) governing these multi-limb actions'' (Swinnen, Young,Walter & Serrien, 1991). Strictly speaking, however, such a unitary pattern ofactivation can apply only to identical (i.e. symmetrical) movements of the leftand right hand. Some authors have suggested that bi-manual interference isdue to neural cross-talk between the trajectory formation of each hand(Marteniuk & MacKenzie 1980; Marteniuk et al., 1984; Swinnen et al., 1991;Spijkers, Heuer, Kleinsorge & van der Loo, 1997). In general, the concept ofcross-talk refers merely to qualitative phenomena of mutual assimilation ofmovement parameters in bi-manual tasks.
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 527

Another, not necessarily exclusive, interpretation of bi-manual interferencein discrete tasks is based on the assumption that some global aspects of bi-manual movements are controlled jointly, while other characteristics may begoverned separately for each hand (Schmidt, Zelaznik, Hawkins, Frank &Quinn Jr, 1979; Heuer, 1986; Franz, Zelaznik & McCabe, 1991). This in-terpretation derives from the idea that a certain class of actions is controlledin a more or less invariant way by means of a generalised motor program(structural features), whereas other parameters may vary from trial to trial(metrical features, Heuer, Schmidt & Ghodsian, 1995). In the case of bi-manual movements this implies that one can distinguish between ``. . .com-mon parameters [that] apply to movements of both hands, while speci®cparameters are speci®ed independently for the left and right hand'' (Heuer,1986). In fact, the concept of global aspects hints at integrated control similarto synergistic constraints, whereas hand-speci®c parameters allow both handsto make distinct movements simultaneously. Recently, Heuer et al. (1995)successfully elaborated on this idea when he investigated whether there wasan invariant relative timing strategy to control both unimanual and bi-manual movements.
Continuing in this vein we can hypothesise that if bi-manual movementsare partly coupled, trajectory generation can be classi®ed in two generalstages: a common stage in which part of the movement is planned for bothhands together (i.e. without reference to the left or right hand) and hand-speci®c structures in which movements are speci®ed independently of eachother. The contribution of these structures might vary with the speci®c taskand conditions. In situations in which the hands make essentially di�erentmovements, the contribution of common neural structures will necessarily bemodest, whereas in identical, simultaneous bi-manual movements commonstructures will be potentially more important. Ultimately, one can even as-sume that symmetrical bi-manual movements are controlled entirely bycommon neural pathways and that one activation pattern is sent to botharms, whereas largely di�erent, simultaneous movements of both hands arecontrolled entirely by hand-speci®c neural structures. In fact, this concept ofcommon and hand-speci®c structures allows a gradual transition from twoextreme positions, namely from completely independent to truly integratedcontrol of both arms.
Literature on discrete bi-manual tasks has scrutinised primarily temporalmovement parameters and has studied mainly integrated control of timing ofthe left and right hand. Despite well-known spatial characteristics of uni-manual hand paths, coupling between spatial parameters has received much
528 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

less attention. Some indications have been found that amplitudes are con-trolled independently (Schmidt et al., 1979; Heuer, 1986), whereas othershave reported assimilation e�ects in spatial parameters related to curvature(Kelso et al., 1983; Franz et al., 1991). Furthermore, most studies have fo-cused largely on assimilation e�ects that occur when movement tasks for theleft and right hand di�er (Kelso et al., 1979; Swinnen et al., 1991; Steen-bergen, Hulstijn, de Vries & Berger, 1996; Fowler et al., 1991). Integratedcontrol of bi-manual movements however, is most likely in identical, i.e.symmetrical, movements.
Characteristics of uni-manual pointing movements have been studiedfrequently, again following the idea of Bernstein (1967) that invariant fea-tures re¯ect control strategies of the trajectory formation system. For mostuni-manual reaching movements, the system is again redundant in severalways: in principle there is an in®nite set of joint rotations, velocity pro®les,muscle activations, etc. that can take the hand from one point to another.Therefore the system frequently has to select one particular neural activationpattern (out of an in®nite set of possible ones) that ful®ls a given task. De-spite this redundancy, pointing movements show several characteristics thatare invariant across subjects and tasks: generally, hand movements are gentlycurved, smooth and have a bell-shaped velocity pro®le (Morasso, 1981;Abend, Bizzi & Morasso, 1982). On the basis of these results, it has beenhypothesised that an extrinsic (hand) co-ordinate system is used internally togenerate arm movements (Morasso, 1981). Recently, we investigated theorigin of curvature in uni-manual pointing movements and concluded thatplanning and control mechanisms are the same for both hands becausecharacteristics of movements made by the left and right arm were found to bethe same (Boessenkool, Nijhof & Erkelens, 1998). It is therefore interesting toinvestigate which (if any) spatial characteristics are controlled jointly in bi-manual tasks.
Despite these invariant average characteristics of uni-manual pointingmovements, there is quite an amount of variability between individual handtrajectories in series of repeated trials. This variability can be explained by thepresence of variability in the system for trajectory formation. The notion of a``functional grouping of muscles'' raises the interesting question as to wherethe variability enters the system of hand movements generation: does it entervia common pathways or via hand speci®c structures of the trajectory for-mation? As an example, variability in the directions of binocular eye move-ments (saccades), made towards a single target, have been found to be tightlycoupled between the two eyes, which suggests that variability in saccades of
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 529

the left and right eyes originates from a common movement organisation(Erkelens & Sloot, 1995).
We hypothesise that if (parts of ) bi-manual movements are controlledjointly for both hands, there should be signi®cant correlations between themovement parameters of each hand. If, on the other hand, bi-manualmovements are controlled by independent, hand-speci®c structures, vari-ability in movements of one hand should be independent of the variability inmovements of the other hand. The objective of this study is to analyse thecorrelation of bi-manual movement parameters in common tasks in order toestimate which parameters of these movements are planned jointly for bothhands. We will investigate the characteristics of bi-manual hand movementswhich have previously been studied in uni-manual tasks. We will focus onboth temporal and spatial parameters.
Firstly, we tested possible coupling of the movement control of the twohands by analysing symmetrical bi-manual pointing movements towards acommon target. Secondly, we investigated the system's ability to adjust themotor programs of the two hands independently. In this second experi-ment we analysed bi-manual movements towards a common target andspeci®ed an intermediate point (via-point) for the preferred hand whichwas presented during the subject's reaction time. Via-points were chosensuch that an adjustment of the hand path was required. Thirdly, we an-alysed asymmetrical movements to test whether the movement parametersthat were found to be coupled for both hands in symmetrical movementsremained correlated when essentially di�erent movements were made byboth hands. For that purpose we presented subjects common targets towhich they had to make movements with a di�erent amplitude for eachhand.
2. Materials and methods
2.1. Subjects
Four subjects (two males and two females), aged 24±27, participated in theexperiments. Subjects did not have any known neuromuscular disorders. Allsubjects had normal or corrected-to-normal vision. Subject JB was left-handed, the other subjects were predominantly right-handed. All subjectswere naive relative to the purpose of the experiment, except for JB (one of theauthors).
530 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

2.2. Experimental set-up
Subjects were seated in front of a horizontal table that was positionedjust below shoulder level. Body movements were reduced by the use of achin rest. In order to ensure that equivalent hand paths originated fromequivalent motor programs, we restricted degrees of freedom of move-ments by immobilising wrist and ®nger joints with a wrist cu�. Otherwisesubjects could move freely and naturally; there was no support for theelbow. Targets, starting points and via-points consisted of small greenLEDLEDs mounted on a 50 ´ 50 cm2 board. Both the targets and the datarecording (see below) were controlled by an ATARIATARI Mega 1 microcom-puter. All movements could be made naturally and comfortably by allsubjects and no movements were made near the extremities of the work-space. During the experiments the room was completely darkened in orderto prevent subjects from adjusting their movements on the basis of visualinformation about their arm positions. Subjects were not given knowledgeof result.
2.3. Movement registration
Movements were recorded by measuring the positions of the subject's leftand right stretched index ®nger tip. A SELSPOTSELSPOT IIII infrared camera systemsampled at 500 Hz the positions of two small infrared LEDLEDs attached to thesubject's ®nger tips. The camera was attached to the ceiling about 2 m abovethe stimulus table. In order to reduce re¯ections, which introduce errors intothe sampled position of the infrared LEDLEDs, a black paper cone (length about1.5 m) was mounted around the camera.
The resolution of the system was 0.2 mm. Before each experiment a grid ofnine pre-de®ned positions was accurately measured for o�-line calibration.The calibration points were used to ®nd a linear transformation to convertthe (x, y) sample points to (x, y) hand positions using a linear, least-squaresmethod (Burden & Faires, 1988). The mean calibration error was 0.8 mm.E�ectively this step-up measured the projection of a three-dimensionalmovement on the horizontal (x, y)-plane. The vertical component of themovement was not measured but is known to be of the order of 5 cm(Rossetti, Stelmach, Desmurget, Prablane & Jeannerod, 1994). Because the(x, y)-signal is relatively una�ected by the vertical component, the error in-troduced by ignoring the vertical movement is small (<1 mm). A tiny metalpin (; 2 mm, negligible length) was mounted on the ®nger tip. This pin ®tted
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 531

a hole in the LEDLED board at the starting positions, in order to minimisevariations in starting position.
2.4. Experimental procedure
Subjects started each trial by releasing a foot-controlled switch whenevertheir hands were at the appropriate starting points and they were ready tomake the hand movements. After a randomised time of between 1 and 3 s thetarget was presented. Subjects were instructed to respond as quickly aspossible to the appearance of the target by making a ballistic movementtowards the target with both hands simultaneously. There were no accuracyconstraints and no instructions were given about the path the hands shouldtake.
In the ®rst experiment, movements were made towards the central targetLEDLED T1 (see Fig. 1). The target was located 25 cm in front of the subject onthe mid-sagittal axis so that movements were symmetrical for the left andright hand. Three di�erent starting positions were used and each movementwas repeated 30 times. Movements were mixed and presented in pseudo-random order. The starting positions were chosen such that movements re-quired primarily either elbow ¯exion (starting position 1), shoulder ¯exion
Fig. 1. Top view of the experimental set-up. Starting positions are depicted by open circles, targets and via-
points by ®lled symbols, arrows indicate movement directions. In Experiment 1 three starting positions
were used (1±3 for the left 10±30 for the right hand), whereas starting positions in Experiments 2 and 3 were
®xed. In Experiments 1 and 2 movements were made towards a ®xed target (T1 and T2, respectively),
whereas in the third experiment three targets were used.
532 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

(starting position 3) or a combination of both shoulder ¯exion and elbowextension (starting position 2).
The second experiment consisted of movements towards target LEDLED T2
that was located on the mid-sagittal axis 46 cm in front of the subjects (seeFig. 1). These movements required shoulder ¯exion and elbow extension andstarted from positions 2 and 20. In this experiment an intermediate LEDLED waspresented 100, 150 or 200 ms after the appearance of the target LEDLED. Subjectswere asked to respond as quickly as possible after the appearance of thetarget by making a ballistic movement to the target via the intermediatepoint. Three intermediate points were used which required movements thatwere either straight (v1) or curved inwards (v2) or outwards (v3). All via-points were presented 20 times for each delay time, resulting in 180 move-ments presented in pseudo-random order. In order to limit the visual anglebetween the target and via-points and to restrict the area of attention, in-termediate points were presented for the preferred hand only.
In the third experiment, both hands started from symmetrical positions (3and 30). The common targets (T3) however, were presented either to the leftor right of the subject's mid-sagittal axis, or as control condition, on this axis(thereby replicating one condition of Experiment 1). The target position wasvaried in pseudo-random order and movements were always made with bothhands. Again, subjects were asked to minimise reaction and movement timeand no speci®c instructions were given about the synchronisation of the twohands. Note that minimising both reaction and movement time implied thatif the target appeared to the left or right of the subject, subjects were asked tosynchronise the start but to desynchronise the end of the movements, sincemovement amplitude, and consequently movement time, was di�erent forboth hands in these asymmetrical conditions.
2.5. Data analysis
The data were numerically di�erentiated o�-line by taking the analyticalderivative of the second-order polynomial ®tted through a symmetrical,moving time window of 5 points (10 ms) (Press, Vetterling, Teukolsky &Flannery, 1992). Then the di�erentiated signal was digitally low-pass ®lteredto 10 Hz. This procedure was repeated to calculate the second derivativeof the position signal. In the ®rst two experiments, a velocity threshold of10 cm sÿ1 was used to detect the start and the end of the movement. Thisprocedure, however, was unsuitable for comparing movements with di�erentamplitudes, as studied in the third experiment. Because if the velocity pro®les
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 533

of movements with di�erent amplitudes can be considered as scaled versionsof each other, any method with a ®xed velocity threshold would introduceinappropriate timing di�erences. To avoid such errors, we used a relativevelocity threshold of 10% of the maximum velocity in the third experiment.
Hand paths were analysed by calculating four spatial parameters: initialdirection, e�ective direction, amplitude and curvature of the movements. Thetime course of movements was characterised by ®ve temporal parameters:reaction time, movement time, maximum velocity, maximum accelerationand skewness of the velocity pro®le. The initial direction was de®ned as thedirection from the initial position of the hand to its position after the ®rst3 cm of a movement (see Fig. 2). The e�ective direction was de®ned as thedirection from initial to ®nal position of the hand. It is important to dis-tinguish this e�ective direction from the movement direction towards the®nal position of the hand. Whereas the latter measures the direction in whichthe hand approaches the ®nal position, the e�ective direction measures theresulting direction of the whole movement. As there were no path con-straints, the e�ective direction measured the task ful®lment at the end of themovement irrespective of the hand path. The amplitude was de®ned as thedistance between initial and ®nal position of the hand and the curvature wasde®ned as the mean deviation from the straight line between these points(Boessenkool et al., 1998). Skewness of the velocity pro®le was characterisedby the relative acceleration phase (time from movement onset to maximumvelocity divided by movement time). Movements were described with refer-ence to a Cartesian co-ordinate system with the central target LEDLED (T1) asorigin, the positive x-axis to the right of the subject, parallel to the mid-
Fig. 2. De®nitions of movement parameters. Circles denote hand path, arrow indicates movement di-
rection.
534 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

frontal plane and the positive y-axis away from the subject in the mid-sagittalplane. Directions were described with reference to the positive x-axis (0°);angles increased counter clock-wise.
2.6. Correlations
To assess the strength of the coupling between the trajectory generation ofboth hands, we calculated the linear correlation coe�cients between pa-rameters of the left and right hand movements. In the extreme cases, wheremovements of both hands are either truly independent or completely cou-pled, linear correlation coe�cients of any parameter of the trajectory will bezero or unity, respectively. However, in intermediate cases, correlation co-e�cients also provide a useful interpretation in terms of variability added tocommon or hand-speci®c parts of the trajectory generation. The (squared)correlation coe�cient gives a direct estimate of the contribution of thecommon variability, relative to the total amount of variability observed in themovement parameters. In this way, correlation coe�cients provide an ade-quate assessment of the amount of coupling between the trajectory genera-tion of bi-manual movements. Since correlation coe�cients are extraordinarysensitive to statistical `outliers', all data were checked for the occurrence ofextreme values, but no exceptional movements were found.
3. Results
3.1. Experiment 1, symmetrical movements
3.1.1. Average movement parametersAll movements had a smooth, bell-shaped velocity pro®le that was roughly
symmetrical. Occasionally, a small secondary velocity peak was observedtowards the end of the movement. A factorial design, ANOVAANOVA, with subjectsas random factor and starting position and hand as ®xed e�ects, was used totest whether the main e�ects of starting position and hand used were signi-®cant. Di�erences between subjects could not be statistically analysed withthis ANOVAANOVA.
Reaction times varied somewhat with subject but we found no signi®canthand di�erences or any e�ect of starting position �F2;6 < 1; P � 0:05�. If re-action times re¯ect the time needed for information processing and planningof a forthcoming motor response, the duration of these processes does not
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 535

depend on the starting position. By contrast, other temporal parametersanalysed in this experiment: movement time, maximum velocity, maximumacceleration and skewness, all strongly depended on the starting position�F2;6 > 16:4; P < 0:004�. None of these parameters showed a signi®cant e�ectof the hand used �F1;3 < 3:9; P > 0:14�. Typically, movements that requiredmainly elbow ¯exion (starting position (1)) showed the highest maximumaccelerations, whereas movements which involved both elbow and shoulder¯exion (starting position (2)) showed lowest values for this parameter.Variations in mean maximum accelerations were quite extensive: sometimes,maximum accelerations of movements from starting position (1) were morethan twice as high as maximum accelerations of movements from startingposition (2). Maximum velocities showed a similar but less pronouncedpattern of variation: again maximum velocities were highest for movementsfrom starting position (1) and lowest for movements from starting position(2). The variations in maximum velocities and accelerations with startingposition were consistent for all subjects. Similarly, movement times wereinversely related to maximum accelerations and velocities: high maximumaccelerations led to low movement times. However, variations in movementtimes as a function of starting position were less distinct. Qualitatively, thedependence of maximum acceleration on the kind of movement corroboratesthe results obtained by Gordon, Ghilardi, Cooper and Ghez (1994). Thetypical dependences of mean maximum velocities and accelerations onstarting position can be explained by assuming that acceleration pro®les of allstarting positions are scaled versions of each other.
Average movement end-points were quite accurate, although small sys-tematic errors were found for most subjects. Typically, movement amplitudeswere slightly smaller than needed, possibly re¯ecting subjects' strategy not tocross their arms. In general, hand paths were slightly curved, the amount andsize of the curvature varying strongly between subjects and starting positions�F2;6 � 6:6; P � 0:03�. A similar dependence of curvature on subject andstarting position was reported in a previous study (Boessenkool et al., 1998).Movement amplitudes depended on starting position (F2;6 � 11:3; P � 0:009),whereas none of the spatial parameters depended on the hand used(F1;3 < 3:3; P > 0:17).
3.1.2. Variability in movement parametersThere was substantial variability in all movement parameters which de-
pended on subject and starting position. Typical coe�cients of variation(standard deviation divided by mean values) in temporal parameters ranged
536 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

from 5% to 25%. Generally, maximum velocities showed the lowest coe�-cients of variation (5±14%).
Hand paths between the various starting points and the target also showeda great amount of variability. To illustrate this point, we show in Fig. 3 handpaths from each starting position with the most extreme initial directionsmade by the right hand of a representative subject (EB). The variability inhand paths was not the same for all types of movements: movements thatrequired primarily rotation about the elbow joint (starting position (1), seeFig. 1) typically showed the least variability, whereas movements about theshoulder joint (3) and combined movements (2) showed much more vari-ability (Fig. 3). Standard deviations in the initial direction pooled oversubjects and hands ranged from 2.9° for movements from starting position(1) to 6.7° from (2) and 8.5° from position (3). Variability in e�ective direc-tions was much smaller; standard deviations ranged from 2.0° to 2.4° and2.8° for movements from starting positions (1), (2) and (3). This reduction indirectional variability was observed in all subjects and all starting positions.The ratio between standard deviations in initial and e�ective directions variedbetween 1.0 and 4.6 according to subject and starting position. A similarreduction in directional variability has been reported for eye movements(Erkelens & Sloot, 1995). The consequence of the considerable variability ininitial directions and more precise e�ective directions is that movementcurvature also showed a fair amount of variation. Variability in amplitudeswas modest, varying slightly with starting position. Highest variability wasfound for starting position (1), coe�cients of variation of amplitude weremaximally a mere 6% (about 1.5 cm).
Fig. 3. Example of hand paths of movements made in Experiment 1. Movements with the most extreme
initial direction of the right hand are shown.
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 537
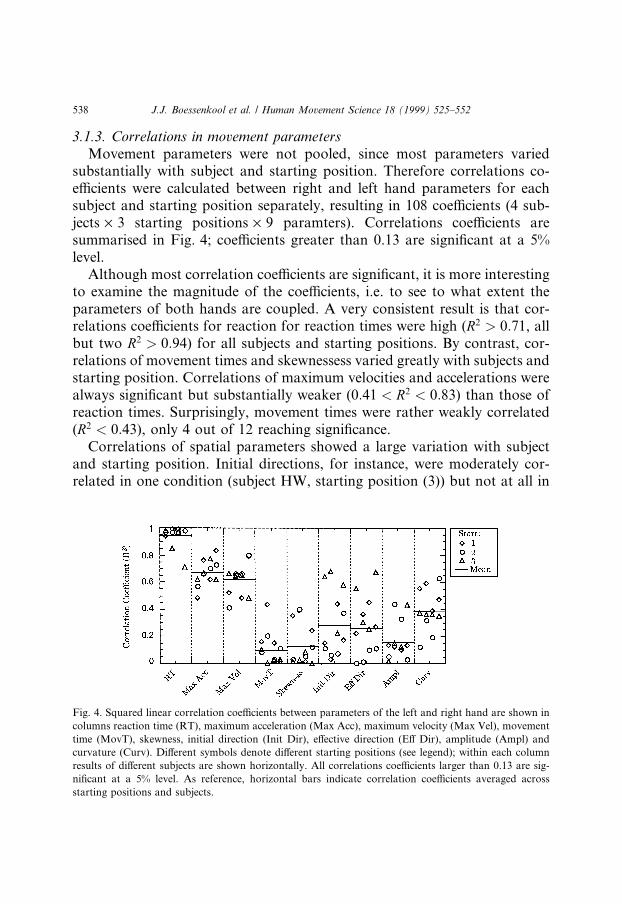
3.1.3. Correlations in movement parametersMovement parameters were not pooled, since most parameters varied
substantially with subject and starting position. Therefore correlations co-e�cients were calculated between right and left hand parameters for eachsubject and starting position separately, resulting in 108 coe�cients (4 sub-jects ´ 3 starting positions ´ 9 paramters). Correlations coe�cients aresummarised in Fig. 4; coe�cients greater than 0.13 are signi®cant at a 5%level.
Although most correlation coe�cients are signi®cant, it is more interestingto examine the magnitude of the coe�cients, i.e. to see to what extent theparameters of both hands are coupled. A very consistent result is that cor-relations coe�cients for reaction for reaction times were high (R2 > 0:71, allbut two R2 > 0:94) for all subjects and starting positions. By contrast, cor-relations of movement times and skewnessess varied greatly with subjects andstarting position. Correlations of maximum velocities and accelerations werealways signi®cant but substantially weaker (0:41 < R2 < 0:83) than those ofreaction times. Surprisingly, movement times were rather weakly correlated(R2 < 0:43), only 4 out of 12 reaching signi®cance.
Correlations of spatial parameters showed a large variation with subjectand starting position. Initial directions, for instance, were moderately cor-related in one condition (subject HW, starting position (3)) but not at all in
Fig. 4. Squared linear correlation coe�cients between parameters of the left and right hand are shown in
columns reaction time (RT), maximum acceleration (Max Acc), maximum velocity (Max Vel), movement
time (MovT), skewness, initial direction (Init Dir), e�ective direction (E� Dir), amplitude (Ampl) and
curvature (Curv). Di�erent symbols denote di�erent starting positions (see legend); within each column
results of di�erent subjects are shown horizontally. All correlations coe�cients larger than 0.13 are sig-
ni®cant at a 5% level. As reference, horizontal bars indicate correlation coe�cients averaged across
starting positions and subjects.
538 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

other conditions (starting positions (1) and (2)). Correlations of e�ectivedirections showed a typical variation with starting position: (1) and (3)yielded signi®cant correlations while correlations of starting position (2) werenot signi®cant. Correlations of initial directions exhibited the same variationwith starting position, but with two exceptions (out of 12).
Mean values of the correlation coe�cients demonstrate that the reactiontimes of ballistic bi-manual movements are very tightly coupled. Movementtimes are hardly coupled at all, which is in sharp contrast to the notion of``functional grouping of muscles'' (i.e. one activation pattern controls botharms). In other words, the timing of fast hand movements shows a fairamount of desynchronisation after almost perfectly simultaneous initiations.Furthermore, temporal movement parameters (other than movement time)seem to be more correlated than spatial parameters, although all parametersvary abundantly with subject and starting position.
3.2. Experiment 2, adjustments
In the second experiment, we presented an intermediate point shortlyafter the appearance of the target in order to investigate the subject's abilityto adjust a forthcoming hand movement. As the reaction times weresomewhat variable, the time available to the subject for adjusting theforthcoming movement varied and was to a certain extent beyond thecontrol of the experimenter. Therefore, time between the presentation of theintermediate point and movement onset was calculated o�-line; we will referto this time as ``adjustment time''. The intermediate points speci®edmovements that were either curved inwards, outwards or straight. Conse-quently, movement curvature is an appropriate measure of the adjustment.We expected the data to form two distinct sub-categories: a ®rst class ofshorter adjustment times in which subjects did not have su�cient time toprocess new information and showed movements symmetrical to those ofthe non-preferred hand, and a second class of longer adjustment times, inwhich subjects were able to account for the intermediate point and adjustedtheir movements accordingly. Fig. 5 shows curvature of the preferred handas a function of adjustment time (subject EB). Surprisingly, instead of twosub-categories, Fig. 5 clearly shows that movement curvature changedcontinuously with adjustment time. Movements via the intermediate point v2
were more negatively curved for increasing adjustment times, while move-ments via v3 showed an increasing positive curvature. Correlations betweencurvatures and adjustment times of the preferred hand were signi®cant for
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 539

all subjects and ranged between 0.43 and 0.74 for movements curved in-wards (via v2) and between ÿ0.47 and ÿ0.56 for movements curved out-ward (via v3). A similar gradual adjustment of movements has beenreported in double-step experiments for eye movements (Becker & J�urgens,1979) and hand movements (van Sonderen, Denier an der Gon & Gielen,1988).
Adjustments of motor programs of the preferred hand in response to thepresentation of intermediate points had virtually no e�ect on the other hand(Fig. 6). A modest, but signi®cant and consistent, correlation between ad-justment times and curvatures was found for movements by the preferredhand for all subjects. However, no signi®cant change in curvature was foundfor the other hand. The 60 movements (3 delays ´ 20 repetitions) with in-termediate point v3 of one representative subject have been sorted by ad-justment times. Fig. 6 shows average hand paths for the 5 movements havingthe shortest, median and longest adjustment times. It is clear that movementswith long adjustment times were made correctly via the intermediate point,whereas hand paths with short adjustment times were roughly symmetrical tothose of the other hand. Median adjustment times yielded paths that adaptedonly partially towards the intermediate point, again illustrating that move-ment modi®cation is a continuous function of adjustment time. However, insharp contrast to the modi®cations of the right-hand movements, paths bythe other hand are not a�ect at all, as is illustrated by the superposition of thethree average hand paths in Fig. 6.
Fig. 5. Curvature as a function of adjustment time (time between presentation of the intermediate point
and movement onset) of Experiment 2 subject EB. Filled (open) circles denote movements in the presence
of intermediate point v2 (v3) which requires positively (negatively) curved movements. Mean movement
curvature of the non-preferred hand is indicated by a dashed horizontal line.
540 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

In order to statistically analyse movement adaptation and possible inter-action between the hands, we calculated the mean movement parameters ofthe ®ve movements with the shortest and longest adjustment times. Allsubjects tended to make movements that were curved slightly inwards (pos-itive curvature). Therefore, movements with short adjustment times as well asmovements with intermediate point v1 had a positive curvature that wassigni®cant for most subjects. As mentioned before, all subjects showed asigni®cant correlation between adjustment times and curvatures for thepreferred hand. Consequently, curvatures of the ®ve movements with thelongest adjustment times were larger than those with the shortest adjustmentstimes. This di�erence was always signi®cant at a 5% con®dence level except inone subject in one condition. Subject HW showed a strong bias towardspositively (inwardly) curved movements. Therefore, the increase in meancurvature for movements with intermediate point v2 did not reach signi®-cance (two-tailed t-test8� 1.88, P� 0.096). Movements of the non-preferredhand were hardly in¯uenced by the increasing curvature of movement of thepreferred hand. Only in one case was there a signi®cant di�erence betweenmean curvatures of the ®ve movements with shortest and longest adjustmenttimes. Subject JB showed a signi®cant increase in the mean movement cur-vature of the non-preferred hand; this increase was approximately 12% of the
Fig. 6. Hand paths of movements of the left and right hand with intermediate point v3 of Experiment 2.
Average paths of 5 movements having longest (l), median (m) and shortest (s) adjustment times are shown
(subject EB).
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 541
increase found for the preferred hand in response to intermediate point v3
(two-tailed t-test8� 3.42, P� 0.009).
3.3. Experiment 3, movements of unequal amplitude
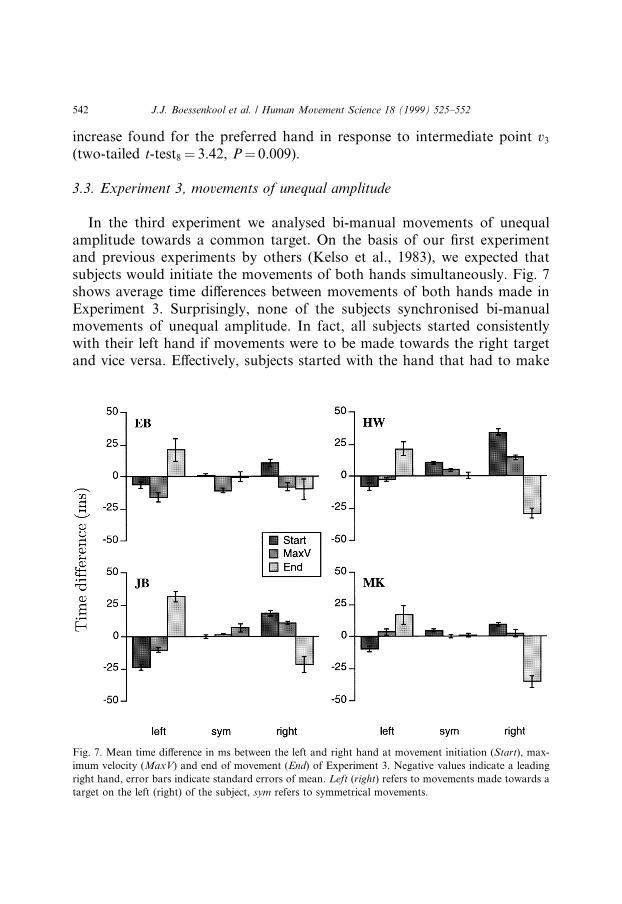
In the third experiment we analysed bi-manual movements of unequalamplitude towards a common target. On the basis of our ®rst experimentand previous experiments by others (Kelso et al., 1983), we expected thatsubjects would initiate the movements of both hands simultaneously. Fig. 7shows average time di�erences between movements of both hands made inExperiment 3. Surprisingly, none of the subjects synchronised bi-manualmovements of unequal amplitude. In fact, all subjects started consistentlywith their left hand if movements were to be made towards the right targetand vice versa. E�ectively, subjects started with the hand that had to make
Fig. 7. Mean time di�erence in ms between the left and right hand at movement initiation (Start), max-
imum velocity (MaxV) and end of movement (End) of Experiment 3. Negative values indicate a leading
right hand, error bars indicate standard errors of mean. Left (right) refers to movements made towards a
target on the left (right) of the subject, sym refers to symmetrical movements.
542 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

the larger movement, thereby desynchronising movement initiation. An-other consistent result was that subjects did not synchronise the end oftheir movements either. Fig. 7 clearly shows that the hand making thesmaller movement ended ®rst. By contrast, symmetrical (control) move-ments are very well synchronised, mean timing di�erences being less than10 ms.
Although timing di�erences were small, between 6 to 34 ms only, they wereconsistent in all subjects for all movements of unequal amplitude. All dif-ferences in movement initiation were signi®cant (two-tailed t-test29 P < 0.05);desynchronisation of the end of movements was signi®cant in all cases exceptone out of eight (4 subjects ´ 2 target positions). It is important to note thattiming di�erences at movement initiation were much smaller than the vari-ability in reaction times (31 to 68 ms). This means that not only averagemovements of the left and right hand exhibit subtle time di�erences, but thatin each individual trial movement initiation of one hand is delayed relative tothat of the other hand. In fact, the reaction times of both hands ± thoughslightly di�erent ± remained very well correlated, correlation coe�cientsranging between 0.92 and 0.99.
4. Discussion
4.1. Two hands, one functional unit?
We investigated bi-manual movement co-ordination in symmetrical taskswhich required identical movements for both hands. In such movements, thecontribution of common control structures will potentially be the most im-portant. In fact, we investigated bi-manual movements during optimal con-ditions of common control, since in theory these movements could becontrolled entirely as one functional unit, i.e. by a unitary activation patternsent to both hands. In literature, evidence for the existence of coupling be-tween the hands has been deduced mainly from assimilation e�ects that occurwhen the movements of each hand di�er from each other. Generally, subjectswere inclined to synchronise bi-manual movements even in asymmetricalconditions. We did indeed ®nd that in symmetrical movements, the meantemporal movement parameters for each hand did not di�er signi®cantly.This means that, on average, subjects tend to move both hands synchro-nously, as was predicted on the basis of earlier studies (Kelso et al., 1979;1983). However, as there was a fair amount of variability in both reaction
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 543

and movement times, in individual trials these times can di�er substantiallyfrom the mean values.
We argue that if bi-manual movements are constrained to act as a singlesynergy, movement parameters of each hand should vary jointly for bothhands together and should give rise to signi®cant correlation between theparameters of each hand. We try to explain the variability in movementparameters in terms of variability in the system of trajectory generation. If bi-manual movements are controlled partly in common for both hands, vari-ability will potentially stem from both common and independent sources.Firstly, variability in common neural structures of movement planning leadsto variations in the movements of both hands together. As an example, apossible common source of variability stems from the visual coding of theposition of the (common) target. Secondly, variability in hand-speci®cstructures of movement generation leads to independent variations ofmovements of each hand. As an example, a potential source of hand-speci®cvariability descends from the transformation of neural activation signals intomuscular activity. If the only source of variability in trajectory generationwas located in common neural pathways, both hands would truly be coupledin the sense that variations in movement parameters would be the same inboth hands. Consequently, correlation coe�cients of any movement pa-rameter would be equal to unity. On the other hand, if variability in hand-speci®c structures accounted for the observed variability, from an experi-mental point of view, both hands would be truly independent, variations inone hand being unrelated to variations in the other hand. In this case therewould only be correlations between tasks, e.g. covariation of average move-ment times in response to di�erent amplitudes, while within a certain bi-manual task (e.g. one particular distance) movements of each hand would beindependent.
From the result of the ®rst experiment it is clear that bi-manual move-ments are neither completely coupled nor fully independent. Correlationcoe�cients of reaction times are consistently high, from which we concludethat movement initiation is controlled together for both hands. Surprisingly,however, movement times show much lower correlation coe�cients, oftennot even reaching signi®cance. These results show that, although synchron-isation is present in average movements, individual movements may show afair amount of desynchronisation after simultaneous initiation. These resultspartially corroborate the ®ndings of Kelso et al. (1983), who also reportedhighly correlated reaction times in symmetrical (and asymmetrical) tasks. Bycontrast, however, they found that correlation coe�cients of movement times
544 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

were always signi®cant and ± although somewhat lower than those of reac-tion times ± varied between moderate and high (0.25 < R2 < 0.96 in sym-metrical conditions).
Correlations between spatial movement parameters are to some extentvariable, but generally quite modest. Therefore it is di�cult to maintain theview that both hands are essentially controlled as one functional unit. In fact,with respect to spatial parameters the hands are to a large extent independentof each other. This ®nding has important consequences for the interpretationof uni-manual control structures in bi-manual tasks. In the motor cortexneurons have been identi®ed which encode for the directions of handmovements (Georgopoulos, Kalaska, Caminiti & Massey, 1982). In addition,there have been numerous suggestions that direction and extent of pointingmovements are controlled by independent sensorimotor channels (Bock &Eckmiller, 1986; Soechting & Flanders, 1991; Gordon et al., 1994; Messier &Kalaska, 1997). Furthermore, errors of pointing movements have been as-cribed to the sensorimotor transformations during movement preparation(Soechting & Flanders, 1991). With respect to bi-manual co-ordination, theindependence of spatial parameters of each hand suggests that these pro-cesses operate at hand-speci®c levels.
The fact that movements of the hands are to a large extent independent ofeach other implies that the contribution of common structures of bi-manualmovement control is quite modest. It is only movement initiation that ap-pears to be controlled truly in common for both hands. In addition, however,moderate correlation coe�cients of maximum velocities and maximum ac-celerations (0.41 < R2 < 0.83) may re¯ect a partially common timing of the®rst phase of bi-manual movements. Interestingly, common control of theinitial phase of the movement does not hold for spatial parameters sincecorrelations between initial directions are low. The ®nding that averagemovements are synchronised rather than movements in individual trialssuggests that trajectory generation consists of two largely independent, hand-speci®c systems.
Results of the ®rst experiment show that bi-manual movement control isbasically independent for each hand. In the second experiment we investi-gated whether adjustments of the movements could also be made indepen-dently for each hand. As in the ®rst experiment, subjects preparedsymmetrical movements towards a common target, but an intermediate pointfor the preferred hand was presented during the subject's reaction time.Results show a continuously increasing curvature of movements of the pre-ferred hand with ``adjustment time'' (time between presentation of the in-
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 545

termediate point and movement onset). With large adjustment times subjectsmake correct via-point movements, whereas with short adjustment timessubjects make movements that are symmetrical to those of the non-preferredhand. Intermediate adjustment times yield a partial adjustment in response tothe presentation of the intermediate point. Despite this distinct e�ect onmovement curvature of the preferred hand, movements of the other handwere hardly a�ected. Comparison of movements with the shortest andlongest adjustment times show that movements of the other hand are hardlyin¯uenced by the adjustment made to movements of the preferred hand.Therefore, we conclude that not only planning but also adjustments ofmovements are made by two independent, hand-speci®c systems.
Interestingly, the absence of change in curvature of the movements of thenon-preferred hand is in contrast to the assimilation e�ects which have beenreported in other discrete movements and in a drawing task. Partial assimi-lation of curvature was observed when subjects were asked to draw lines withone hand and circles with the other hand simultaneously (Franz et al., 1991).Similarly, in discrete bi-manual movements both trajectories were found tochange if a hurdle was placed mid-way between starting point and target ofone hand (Kelso et al., 1983). Although the latter authors did not quantifymovement curvature, it was clear that the e�ect of the barrier on curvaturewas much stronger in the hand that had to move over it than in the otherunconstrained hand.
A number of noteworthy di�erences in the experimental paradigms maypossibly account for discrepancies between our present ®ndings and earlierstudies on bi-manual interference. Firstly, there were no accuracy constraintsand subjects did not have to compromise between speed and accuracy as isrequired in Fitts' task. Secondly, all movements were made in the dark, sothat no visual corrections could be made. We hypothesise that previouslyfound interactions between asymmetrical movements of both hands (Mar-teniuk & MacKenzie, 1980; Kelso et al., 1983; Swinnen et al., 1991) re¯ect thedi�culties of combining two di�erent motor tasks rather than a commoncontrol of homologous muscles. This concept also provides a possible ex-planation for the strong correlations in movement times found by Kelso et al.(1983). These authors studied symmetrical movements, but targets weredi�erent for each hand and there was a prescribed accuracy (target width).Symmetrical conditions were mixed with bi-manual movements of unequaldi�culty, i.e. di�erent targets widths for each hand. This experimental con-dition may be regarded as two di�erent tasks for each hand and consequentlymay cause an interaction between the hands at ``tasks level''.
546 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

4.2. Timing during movements of unequal amplitude
In the third experiment we analysed bi-manual movements of unequalamplitude to test whether reaction times remained correlated if one hand hadto make a substantially larger movement than the other hand. On the basis ofour ®rst experiment and previous experiments by others (Kelso et al., 1983),we expected that subjects would initiate both hands simultaneously. In orderto comply with the instruction to move as fast as possible, subjects wouldthen have to desynchronise the end of the movements. On the other hand, ifthe timing of both hands is governed by a single pattern, one would expectone hand to slow down so that the movements would start and ®nish si-multaneously. Results of Experiment 3 clearly show that subjects do notsynchronise either the start or the end of movements. On the contrary, thereare consistent timing di�erences between the movements of each hand.Subjects started with the hand making the larger movement and delayed theinitiation of the movement of the other hand which nevertheless ended ear-lier. In fact, di�erences in movement time were more or less distributedsymmetrically between the start and the end of movements, which impliesthat both hands reached their peak velocity at approximately the same time.These timing di�erences between paired left and right hand movements weresmall in comparison to the variability in reaction times. Therefore, we con-clude that these subtle timing di�erences are an essential property of thesystem of bi-manual movement generation.
Again our results partially con®rm earlier results on bi-manual interfer-ence. Literature on timing di�erences between the hands is inconclusive.Di�erences in movement initiation have been reported in some asymmetricaltasks (Marteniuk et al., 1984; Fowler et al., 1991), whereas other authorshave stressed the simultaneity of movement initiation (Kelso et al., 1983;Swinnen, Young, Walter & Serrien, 1995). Similarly, di�erences at the end ofthe movements have been reported when movements become too dissimilar(Marteniuk et al., 1984; Fowler et al., 1991), whereas other authors haveemphasised that timing di�erences between the hands were non-signi®cant,although there was ``a clear tendency for the easy hand to reach its target®rst'' (Kelso et al., 1983).
Note that, despite the timing di�erences between the hands, movementinitiation of both hands remained tightly coupled because correlation coef-®cients were invariably high. It is unclear why and how one hand is delayedwith respect to the other. One possibility is that movement initiation isplanned in common neural structures that inhibit trajectory generation of
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 547

one hand in order to retain the simultaneity of moments of peak velocity inasymmetrical conditions. Another possibility is that the activation patterns ofboth hands are planned independently but are afterwards synchronised inorder to minimise di�erences in the timing of peak velocities. In fact, there isneurophysiological evidence that bi-manual co-ordination involves activityof speci®c cells (Donchin, Gribova, Steinberg, Bergman & Vaadia, 1998) andregions (Viviani, Perani, Grassi, Bettinardi & Fazio, 1998) in the motorcortex. The reason for this synchronisation, however, remains obscure.
4.3. Limitations of our approach
We investigated bi-manual movements during optimal conditions ofcommon control, since in theory, identical movements can be controlledentirely by a unitary activation pattern sent to both hands. In order to elu-cidate the role of common neural structures, we analysed correlation coe�-cients of both temporal and spatial movement parameters. However, somecaution should be observed, in interpreting correlations between movementparameters in terms of a coupling between the control mechanisms of eachhand.
Note that in the absence of variability, correlations cannot be de®ned andhence our approach would be inapplicable. Despite the unambiguous taskand uniform initial conditions however, there was a substantial amount ofvariability in all movement parameters; this must result from variations in themechanisms of movement control. Furthermore, the fact that reaction timesare found to be highly correlated whereas initial directions are not, indicatesthat both common and hand-speci®c structures contribute to variability inmovement planning.
Our approach rests on the premise that variations in bi-manual move-ments are related to the process of trajectory generation. Correlation coef-®cients assess the contribution of common variability relative to the totalamount of observed variability. However, if variability is not related to theprocess of movement control, it is impossible to infer the role of commonneural pathways from correlations between movement parameters. In fact,irrespective of the value of the correlation coe�cients, one can always assumethat symmetrical bi-manual movements are controlled by one activationpattern that is sent to both hands. One is then forced to assume that, in thecase of low correlation coe�cients, the common activation patterns do notcontribute to the observed variability. Then, in order to explain variability inresponses, one has to assume that variability is introduced only at a hand-
548 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

speci®c phase of the process. Conversely, one can maintain the view that bi-manual movements are controlled by two hand-speci®c systems that shareonly a common source of variability. If variability is unrelated to the controlprocess, correlation coe�cients exclude an interpretation in terms of com-mon neural structures of movement control. However, irrespective of theneurological interpretation, high (low) correlation of movement parametersbetween the hands implies a common (independent) control at the behaviourlevel.
We tacitly assume that, given a speci®c task and condition, the meanvalues of movement parameters re¯ect the subject's intention or the ``ideal''outcome of the planning process. The relationship between movementplanning and measured movement parameters, however, may be obscuredby feedback mechanisms. As there was no visual feedback, measured pa-rameters can result only from feed-forward planning and possible prop-rioceptive (a�erent signals) and internal (e�erent signals) feedback. E�ectsof feedback were probably small, because all movements were ballistic.Nevertheless, consistent reductions in directional variability suggest thatfast feedback mechanisms were involved in the generation of the move-ments.
The set of measured parameters is to some extent arbitrary in the sensethat the movement generation mechanism may use a di�erent set of internalparameters. However, despite the in®nite number of possible trajectories thatcomply with the task requirements, the subject's responses form a morelimited class of actual trajectories. In fact, the consistent variations of meanmaximum velocity and accelerations found in Experiment 1 suggest thatmaximum velocity and acceleration are not varied independently by thesubjects, but that acceleration pro®les are more or less scaled versions of eachother. In other words, despite the limited set of, theoretically independent,measured parameters, our results indicate that maximum velocity and max-imum acceleration are mutually dependent. Moreover, observed variability inmeasured (external) parameters must originate from variability in parametersused in the system of trajectory generation. As the observed parametersdescribe the gross features of bi-manual movements relevant to the task, weinfer from the lack of correlation in these parameters that bi-manualmovement control consists of two largely independent systems. More subtlefeatures of hand trajectories however, may originate from coupling betweenthe movements of each hand. Therefore, we argue that correlations betweenobserved parameters are an adequate assessment of the degree of functionalcoupling (i.e. at a behaviour level) between the two hands.
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 549

5. Conclusions
We conclude that bi-manual trajectory generation consists of two largelyindependent, hand-speci®c systems. Therefore it is di�cult to maintain theview that movements of both hands are controlled by a single activationpattern. Common control of bi-manual movements is limited to reactiontimes. We suggest that the coupling between the hands exists mainly at ``tasklevel'', synchronising the tasks of both hands. We conclude that interactionsbetween movements of the two hands, as observed in asymmetrical tasks,result from the di�culties of performing two unrelated (motor) tasks si-multaneously rather than from non-optimal properties of common control(i.e. co-ordinative structure).
Acknowledgements
The work was supported by a grant from the Life Sciences Foundation(SLW) of the Netherlands Organisation for Scienti®c Research (NWO). Wethank Hans Kolijn for help with the experimental set-up, Pieter Schiphorstfor developing software and Sheila McNab for linguistic advice.
References
Abend, W., Bizzi, E., & Morasso, P. (1982). Human arm trajectory formation. Brain, 105, 331±348.
Becker, W., & J�urgens, R. (1979). An analysis of the saccadic system by means of double step stimuli.
Vision Research, 19, 967±983.
Bernstein, N. (1967). The co-ordination and regulation of movements. Oxford: Pergamon Press.
Bock, O., & Eckmiller, R. (1986). Goal-directed arm movements in absence of visual guidance: evidence
for amplitude rather than position control. Experimental Brain Research, 62, 451±458.
Boessenkool, J. J., Nijhof, E. J., & Erkelens, C. J. (1998). A comparison of curvatures of left and right
hand movements in a simple pointing task. Experimental Brain Research, 120, 369±376.
Burden, R. L., & Faires, J. D. (1988). Numerical Analysis (4th ed.), Boston: PWS-Kent.
Cohen, L. (1971). Synchronous bimanual movements performed by homologous and non-homologous
muscles. Perceptual and Motor Skills, 32, 639±644.
Donchin, O., Gribova, A., Steinberg, O., Bergman, H., & Vaadia, E. (1998). Primary motor cortex is
involved in bimanual coordination. Nature, 395, 274±278.
Erkelens, C. J., & Sloot, O. B. (1995). Initial directions and landing positions of binocular saccades. Vision
Research, 35, 3297±3303.
Fitts, P. M. (1954). The information capacity of the human motor system in controlling the amplitude
movement. Journal of Experimental Psychology, 47, 381±391.
Fowler, B., Duck, T. Mosher, M., & Mathieson, B. (1991). The coordination of bimanual aiming
movements: Evidence for progressive desynchronization. Quarterly Journal of Experimental Psychol-
ogy [A], 43(2), 205±221.
550 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552

Franz, E. A., Zelaznik, H. N., & McCabe, G. (1991). Spatial topological constraints in a bimanual task.
Acta Psychologica, 77, 137±151.
Georgopoulos, A. P., Kalaska, J. F., Caminiti, R., & Massey, J. T. (1982). On the relations between the
direction of two-dimensional arm movements and cell discharge in primate motor cortex. The Journal
of Neuroscience, 2(11), 1527±1537.
Gordon, J., Ghilardi, M. F., Cooper, S. E., & Ghez, C. (1994). Accuracy of planar reaching movements.
II. Systematic extent errors resulting from inertial anisotropy. Experimental Brain Research, 99, 112±
130.
Haken, H., Kelso, J. A. S., & Bunz, H. (1985). A theoretical model of phase transitions in human hand
movements. Biological Cybernetics, 51, 347±356.
Heuer, H. (1986). Intermanual interactions during programming of aimed movements: Converging
evidence on common and speci®c parameters of control. Psychological Research, 48, 37±46.
Heuer, H., Schmidt, R. A., & Ghodsian, D. (1995). Generalised motor programs for rapid bimanual tasks:
A two-level multiplicative-rate model. Biological Cybernetics, 73, 343±356.
Kelso, J. A. S., Southard, D. L., & Goodman, D. (1979). On the nature of human interlimb coordination.
Science, 203, 1029±1031.
Kelso, J. A. S., Putnam, C. A., & Goodman, D. (1983). On the space-time structure of human interlimb
co-ordination. Quarterly Journal of Experimental Psychology, 35A, 347±375.
Kelso, J. A. S. (1984). Phase transitions and critical behavior in human bimanual coordination. American
Journal of Physiology: Regulatory, Integrative and Comparative Physiology, 15, R1000-R1004.
Kugler, P. N., Kelso, J. A. S., & Turvey, M. T. (1980). On the concept of coordinative structures as
dissimipative structures: I. Theoretical lines of convergence. In G. E. Stelmach & J. Requin, Tutorials
in motor behavior (pp. 3±47). Amsterdam: North-Holland Publishing Co.
Marteniuk, R. G., & MacKenzie, C. L. (1980). A preliminary theory of two-hand co-ordinated control. In
G. E. Stelmach & J. Requin, Tutorials in motor behavior (pp. 185±197). Amsterdam: North-Holland.
Marteniuk, R. G., MacKenzie, C. L., & Baba, D. M. (1984). Bimanual movement control: Information
processing and interaction e�ects. The Quarterly Journal of Experimental Psychology, 36A, 335±365.
Messier, J., & Kalaska, J. F. (1997). Di�erential e�ect of task conditions on errors of direction and extent
of reaching movements. Experimental Brain Research, 115, 469±478.
Morasso, P. (1981). Spatial control of arm movements. Experimental Brain Research, 42, 223±227.
Peper, C. E., Beek, P. J., & van Wieringen P. C. W. (1995). Frequency induced transitions in bimanual
tapping. Biological Cybernetics, 73, 301±309.
Press, W. H., Vetterling, W. T., Teukolsky, S. A., & Flannery, B. P., (1992). Numerical recipes in C: The
art of scienti®c computing (2nd ed.), Cambridge: Cambridge University Press.
Rossetti, Y., Stelmach, G., Desmurget, M., Prablane, C., & Jeannerod, M. (1994). The e�ect of viewing the
static hand prior to movement onset on pointing kinematics and variability. Experimental Brain
Research, 101, 323±330.
Sch�oner, G. (1990). A dynamic theory of coordination of discrete movement. Biological Cybernetics, 63,
257±270.
Schmidt, R. A., Zelaznik, H., Hawkins, B., Frank, J. S., & Quinn Jr. J. T. (1979). Motor-output
variability: A theory for the accuracy of rapid motor acts. Psychological Review, 86, 415±451.
Soechting, J. F., & Flanders, M. (1991). Deducing central algorithms of arm movement control from
kinematics. In D. R. Humphrey & H.-J. Freund, Motor Control: Concepts and Issues. New York:
Wiley.
van Sonderen, J. F., Denier van der Gon, J. J., & Gielen, C. C. A. M. (1988). Conditions determining early
modi®cation of motor programmes in response to changes in target location. Experimental Brain
Research, 71, 320±328.
Spijkers, W., Heuer, H., Kleinsorge, T., & van der Loo, H. (1997). Preparation of bimanual movements
with same and di�erent amplitudes: Speci®cation interference as revealed by reaction time. Acta
Psychologica, 96, 207±227.
J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552 551

Steenbergen, B., Hulstijn, W., de Vries A., & Berger, M. (1996). Bimanual movement coordination in
spastic hemiparesis. Experimental Brain Research, 110, 91±98.
Swinnen, S. P., Young, D. E., Walter, C. B., & Serrien, D. J. (1991). Control of asymmetrical bimanual
movements. Experimental Brain Research, 85, 163±173.
Swinnen, S. P., Young, D. E., Walter, C. B., & Serrien, D. J. (1995). The organization of patterns of
multilimb coordination as revealed through reaction time measures. Experimental Brain Research, 104,
153±162.
Viviani, P., Perani, D., Grassi, F., Bettinardi V., & Fazio, F. (1998). Hemispheric asymmetries and
bimanual asynchrony in left- and right-handers. Experimental Brain Research, 120, 531±536.
552 J.J. Boessenkool et al. / Human Movement Science 18 (1999) 525±552
Copyright © 2022 FDOKUMEN



















