TRPV1: a therapeutic target for novel analgesic drugs?
10
TRPV1: a therapeutic target for novel analgesic drugs? Arpad Szallasi 1 , Francisco Cruz 2 and Pierangelo Geppetti 3 1 Departments of Pathology, Monmouth Medical Center, Long Branch, NJ 07740 and Drexel University College of Medicine, Philadelphia, PA 19102, USA 2 Department of Urology, Hospital S. Joa ˜ o and Faculty of Medicine, University of Porto, 4200-076 Porto, Portugal 3 Department of Critical Care Medicine and Surgery, University of Firenze, 50139 Firenze, Italy The vanilloid receptor TRPV1 is now recognized as a molecular integrator of painful stimuli ranging from noxious heat to endovanilloids in inflammation. Pharmacological blockade of TRPV1 represents a new strategy in pain relief. TRPV1 antagonists are expected to prevent pain by silencing receptors where pain is generated rather than stopping the propagation of pain, as most-traditional pain killers do. This hypothesis has already being tested in the clinic by administering small molecule TRPV1 antagonists (e.g. GlaxoSmithKline SB- 705498) for migraine and dental pain. Paradoxically, in some murine models of chronic pain, TRPV1-deficient mice exhibit more pain-related behavior than their wild- type littermates, indicating that the understanding of TRPV1 in pain is still incomplete. Moreover, there is mounting evidence to suggest the existence of func- tional TRPV1 both in the brain and in various non-neu- ronal tissues. The biological role of these receptors remains elusive, but their tissue distribution clearly indicates that they are involved in many more functions than just pain perception. Here, we review the potential therapeutic indications and adverse effects of TRPV1 antagonists. The transient receptor potential vanilloid subfamily member 1, TRPV1 It is an exciting time in pain research. The past ten years have seen the emergence of specific small molecule antagonists targeting receptors on sensory neurons that detect painful stimuli. Among these new therapeutic targets, TRPV1 has attracted the most attention [1]. TRPV1 (formerly known as the vanilloid receptor VR1) is probably still best recognized as the receptor for capsai- cin, which is responsible for the piquancy of hot chili peppers [2]. Capsaicin is unique among naturally occurring irritant compounds because the initial excitation that it evokes is followed by a durable refractory state during which previously excited neurons are unresponsive to a broad range of unrelated stimuli [2]. This effect, referred to as desensitization, has a clear therapeutic potential. As a matter of fact, capsaicin-containing creams have been in clinical use for decades for painful conditions such as diabetic neuropathy [3]. After its cloning in 1997 [4], the vanilloid receptor VR1 was re-named as the transient receptor potential vanilloid subfamily member 1 (TRPV1) to emphasize that this recep- tor belongs to the extended family of ‘thermo TRP’ channels [5]. Subsequently, TRPV1 knock-out mice, which lent further experimental proof that TRPV1 has a central role in the development of post-inflammatory thermal hyperal- gesia, were generated [6,7]. Importantly, it was also recog- nized that TRPV1 is upregulated both in some animal models of chronic neuropathic pain and in various human diseases. These findings have launched a major initiative of some pharmaceutical industries to identify small molecule TRPV1 antagonists with clinical potential to act as novel analgesic–antiphlogistic agents (Box 1 and Box 2). At pre- sent, several small molecule TRPV1 antagonists are either in preclinical studies or already undergoing Phase 1 (e.g. SB- 705498 for dental pain; www.clinicaltrials.gov/ct/show/ NCT00281684) or Phase 2 (e.g. SB-705498 for migraine; www.clinicaltrials.gov/ct/show/NCT00269022) clinical tri- als. Although the outcome of these trials is being awaited, questions regarding the clinical value of TRPV1 antagonists can be subdivided into two broad categories. The first cate- gory relates to the contribution of TRPV1 to inflammatory and neuropathic pain that determines the efficacy of antago- nists. The second category deals with the physiological roles of TRPV1 that might indicate potential adverse effects for antagonists. Here, we attempt to answer these questions based on the literature and our experience with TRPV1 agonists both in animal experiments and patients with migraine and bladder disorders. TRPV1 and its family Ironically, the founding member of the TRPV vanilloid receptor subfamily TRPV1 remains the only vanilloid receptor that is a target for capsaicin and other vanilloids. Up to date, the TRPV subfamily has six members, TRPV1– TRPV6. TRPV5 and TRPV6 seem to be constitutively active and are thought to have a role in vitamin D-depen- dent Ca 2+ uptake in the kidney and intestine, respectively [8,9]. TRPV4 was originally identified as an osmotic recep- tor detecting decreased osmolarity [10]. Interestingly, TRPV4 is co-expressed with TRPV1 in a subset of nocicep- tive neurons where it is sensitized by prostaglandin PGE 2 and has a role in inflammatory thermal hyperalgesia [11]. TRPV3 is co-expressed with TRPV1 and TRPV4 in keratinocytes where it might act as a heat sensor [12]. Review TRENDS in Molecular Medicine Vol.12 No.11 Corresponding author: Szallasi, A. ([email protected]). Available online 25 September 2006. www.sciencedirect.com 1471-4914/$ – see front matter ß 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.molmed.2006.09.001
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of TRPV1: a therapeutic target for novel analgesic drugs?

TRPV1: a therapeutic target for novelanalgesic drugs?Arpad Szallasi1, Francisco Cruz2 and Pierangelo Geppetti3
1 Departments of Pathology, Monmouth Medical Center, Long Branch, NJ 07740 and Drexel University College of Medicine,
Philadelphia, PA 19102, USA2 Department of Urology, Hospital S. Joao and Faculty of Medicine, University of Porto, 4200-076 Porto, Portugal3 Department of Critical Care Medicine and Surgery, University of Firenze, 50139 Firenze, Italy
Review TRENDS in Molecular Medicine Vol.12 No.11
The vanilloid receptor TRPV1 is now recognized as amolecular integrator of painful stimuli ranging fromnoxious heat to endovanilloids in inflammation.Pharmacological blockade of TRPV1 represents a newstrategy in pain relief. TRPV1 antagonists are expectedto prevent pain by silencing receptors where pain isgenerated rather than stopping the propagation of pain,as most-traditional pain killers do. This hypothesis hasalready being tested in the clinic by administering smallmolecule TRPV1 antagonists (e.g. GlaxoSmithKline SB-705498) for migraine and dental pain. Paradoxically, insome murine models of chronic pain, TRPV1-deficientmice exhibit more pain-related behavior than their wild-type littermates, indicating that the understanding ofTRPV1 in pain is still incomplete. Moreover, there ismounting evidence to suggest the existence of func-tional TRPV1 both in the brain and in various non-neu-ronal tissues. The biological role of these receptorsremains elusive, but their tissue distribution clearlyindicates that they are involved in many more functionsthan just pain perception. Here, we review the potentialtherapeutic indications and adverse effects of TRPV1antagonists.
The transient receptor potential vanilloid subfamilymember 1, TRPV1It is an exciting time in pain research. The past ten yearshave seen the emergence of specific small moleculeantagonists targeting receptors on sensory neurons thatdetect painful stimuli. Among these new therapeutictargets, TRPV1 has attracted the most attention [1].TRPV1 (formerly known as the vanilloid receptor VR1)is probably still best recognized as the receptor for capsai-cin, which is responsible for the piquancy of hot chilipeppers [2]. Capsaicin is unique among naturally occurringirritant compounds because the initial excitation that itevokes is followed by a durable refractory state duringwhich previously excited neurons are unresponsive to abroad range of unrelated stimuli [2]. This effect, referred toas desensitization, has a clear therapeutic potential. As amatter of fact, capsaicin-containing creams have been inclinical use for decades for painful conditions such asdiabetic neuropathy [3].
Corresponding author: Szallasi, A. ([email protected]).Available online 25 September 2006.
www.sciencedirect.com 1471-4914/$ – see front matter � 2006 Elsevier Ltd. All rights reserve
After its cloning in 1997 [4], the vanilloid receptor VR1was re-named as the transient receptor potential vanilloidsubfamily member 1 (TRPV1) to emphasize that this recep-tor belongs to the extended family of ‘thermo TRP’ channels[5]. Subsequently, TRPV1 knock-out mice, which lentfurther experimental proof that TRPV1 has a central rolein the development of post-inflammatory thermal hyperal-gesia, were generated [6,7]. Importantly, it was also recog-nized that TRPV1 is upregulated both in some animalmodels of chronic neuropathic pain and in various humandiseases. These findings have launched amajor initiative ofsome pharmaceutical industries to identify small moleculeTRPV1 antagonists with clinical potential to act as novelanalgesic–antiphlogistic agents (Box 1 and Box 2). At pre-sent, several small molecule TRPV1 antagonists are eitherinpreclinical studiesoralreadyundergoingPhase1 (e.g. SB-705498 for dental pain; www.clinicaltrials.gov/ct/show/NCT00281684) or Phase 2 (e.g. SB-705498 for migraine;www.clinicaltrials.gov/ct/show/NCT00269022) clinical tri-als. Although the outcome of these trials is being awaited,questions regarding the clinical value of TRPV1antagonistscan be subdivided into two broad categories. The first cate-gory relates to the contribution of TRPV1 to inflammatoryandneuropathic pain thatdetermines the efficacy of antago-nists. The second category dealswith the physiological rolesof TRPV1 that might indicate potential adverse effects forantagonists. Here, we attempt to answer these questionsbased on the literature and our experience with TRPV1agonists both in animal experiments and patients withmigraine and bladder disorders.
TRPV1 and its familyIronically, the founding member of the TRPV vanilloidreceptor subfamily TRPV1 remains the only vanilloidreceptor that is a target for capsaicin and other vanilloids.Up to date, the TRPV subfamily has six members, TRPV1–TRPV6. TRPV5 and TRPV6 seem to be constitutivelyactive and are thought to have a role in vitamin D-depen-dent Ca2+ uptake in the kidney and intestine, respectively[8,9]. TRPV4 was originally identified as an osmotic recep-tor detecting decreased osmolarity [10]. Interestingly,TRPV4 is co-expressed with TRPV1 in a subset of nocicep-tive neurons where it is sensitized by prostaglandin PGE2
and has a role in inflammatory thermal hyperalgesia [11].TRPV3 is co-expressed with TRPV1 and TRPV4 inkeratinocytes where it might act as a heat sensor [12].
d. doi:10.1016/j.molmed.2006.09.001

Box 1. Salient features of TRPV1 and their implication for
developing TRPV1 antagonists
� TRPV1 is activated by several stimuli, both exogenous and
endogenous, that have no recognizable relationship to each other.
Capsaicin: specific binding site is identified intracellularly.
Resiniferatoxin: recognition domain overlaps with capsaicin site.
Camphor: intracellular site, distinct from capsaicin-binding
domain.
Protons: interact at extracellular glutamate residues.
Heat: no specific response domain is identified.
� Group A antagonists inhibit both capsaicin and proton activation,
whereas Group B antagonists prevent TRPV1 activation only by
capsaicin. Channel-pore blockers (e.g. Ruthenium red) do not
interfere with binding but prevent activation by all stimuli.
� TRPV1 has several phosphorylation sites. Phosphorylation state
might be disease-dependent; phosphorylation influences profile
(agonist versus antagonist) of mixed agonist and antagonists.
546 Review TRENDS in Molecular Medicine Vol.12 No.11
Furthermore, TRPV3 might be a molecular target forplant-derived skin sensitizers [13]. TRPV2, initiallyisolated from the brain as a VR1-like protein (VRL1) withhigh-temperature threshold (>53 8C), forms heteromulti-mers with TRPV1 [14] and is upregulated during inflam-matory hyperalgesia [15]. In the CNS, the role of TRPV2remains enigmatic. Notably, TRPV2 is also expressed inaortic myocytes where (like TRPV4) it might be involved inosmosensing and cardiac-muscle degeneration [16].
The broad tissue distribution of TRPV1 and itsimplications for therapy and side effectsFunctional TRPV1 is expressed both in the brain (from theolfactory bulb to the cortex and from the basal ganglia tothe cerebellum) [17] and in various non-neuronal tissues.From a pharmacological point of view, this phenomenonmight be considered as a double-edged sword: TRPV1 inthese tissues might represent a novel therapeutic target
Box 2. Selected TRPV1 antagonists and their
pharmacological profiles
� SB-705498 (GlaxoSmithKline): successfully completed Phase I
clinical trials. At present, it is in Phase II for the acute treatment
of migraine (recruiting patients since December 2005; Clinical-
Trials.gov identifier: NCT00269022).
� A-425619 (Abbott) [1-isoquinolin-5-yl-3-(4-trifluoromethyl-benzyl)-
urea]: it inhibited mechanical allodynia induced by skin incision
and reversed mechanical hyperalgesia in established rat models of
persistent nociceptive and neuropathic pain [101].
� AMG-9810 (Amgen) [(E)-3-(4-t-Butylphenyl)-N-(2,3-dihydroben-
zo[b] [1,4] dioxin-6-yl)acrylamide]: it reverted thermal and
mechanical hyperalgesia induced by complete Freund’s adjuvant
(CFA) [102].
� BCTC (Neurogen) [N-(4-tertiarybutylphenyl)-4-(3-cholorphyridin-2-
yl)tetrahydropryazine-1(2H)-carbox-amide]: it significantly attenu-
ated thermal and mechanical hyperalgesia in the chronic constric-
tion injury model of neuropathic pain and CFA model of
inflammatory pain in rats [103].
� Nrgn-3 (Neurogen): it has better pharmacokinetics than BCTC.
� GRC 6127 (Glenmark): it is orally active; it reversed both FCA-
induced and partial sciatic-nerve legation-induced mechanical
hyperalgesia in the rat.
� JNJ-17203212 (JohnsonandJohnson): it relieved osteolytic sar-
coma-related bone pain in the mouse; it blocked nebulized citric-
acid-induced cough in guinea pigs [104].
www.sciencedirect.com
and/or mediate unforeseen adverse effects. Either way,these findings place TRPV1 in a much broader contextthan simply a polymodal pain sensor on primary afferentneurons.
Despite extensive research, the physiological function ofTRPV1 in the brain remains elusive. The results areexciting, but many are unconfirmed and/or contradictory.Upfront, it needs to be emphasized that: (i) TRPV1-defi-cient (knock-out) mice show no obvious deficits in theirmotor or cognitive functions; and (ii) the majority of pub-lished studies is based on the use of high doses of capsa-zepine, a first-generation TRPV1 antagonist, which isneither very potent nor selective [2]. In the mouse brain,an extensive neuronal co-localization was demonstratedbetween TRPV1-like and cannabinoid CB1 receptor-likeimmunoreactivities [18]. This is interesting because ana-ndamide and related endogenous cannabimimetic lipidsactivate both TRPV1 and CB1 receptors, in some casesevoking opposite biological actions [19,20]. In the rat sub-stantia nigra, TRPV1 is present on a subset of tyrosinehydroxylase-positive (dopaminergic) neurons [17]. Thisneuronal localization is consistent with the strong anti-hyperkinetic activity of capsaicin in a rat model of Hun-tington’s disease [21]. Furthermore, the TRPV1 antagonistcapsazepine, but not the CB1 blocker SR141716, wasshown to prevent the hypolocomotor effect of AM404, aninhibitor of anandamide uptake that potentiates theactions of endogenous cannabinoids in a mouse model(dopamine-transporter knockouts) of schizophrenia, imply-ing that the beneficial effects of endocannabinoids in thismodel might be mediated by TRPV1 [22]. Similarly, cap-sazepine abolished the beneficial action of AM404 inexperimental autoimmune encephalomyelitis, a rat modelof multiple sclerosis (MS) [23]. If these findings hold truealso in humans, targeting brain TRPV1 with agonistsmight constitute a novel therapeutic strategy for disordersthat are associated with hyperdopaminergia. Conversely,TRPV1 antagonists might be detrimental in patients withMS or schizophrenia. Up to now, these speculationshave no experimental support in humans. Nonetheless,the widespread distribution of TRPV1 in the brain posesthe question of whether CNS penetration is a desirabletrait for TRPV1 antagonists (Box 2).
In the bladder, the presence of TRPV1 wasdemonstrated in various cell types, including the urothe-lium, detrusor muscle and fibroblasts [24]. This ubiquitousexpression pattern is at variance with the phenotype ofTRPV1 knockout mice that exhibit some urine spotting butno other obvious deficits in genitourinary functions [25].TRPV1-like immunoreactivity was observed in humanskin biopsies, and capsaicin was shown to evoke Ca2+
uptake in keratinocytes in culture [26]. The biological roleof TRPV1 in the epithelium is not fully understood. Anattractive hypothesis is that, as in neurons, TRPV1 sub-serves a sensory role both in keratinocytes and in theurothelium [24]. Thus, noxious environmental stimuli thatact on TRPV1might generate pro-inflammatory mediatorsin epithelial cells that, in turn, can evoke a neurogenicinflammatory response, partially by sensitizing and acti-vating TRPV1 on sensory nerves (Figures 1 and Figure 2).If this is the case, TRPV1 is in the center of a positive

Figure 1. TRPV1 regulation on primary sensory neurons. TRPV1 can be activated either directly by agonists such as high temperature and low pH, or indirectly by liberating
the receptor from the inhibitory control of PIP2. PIP2 is cleaved by PLC, which, in turn, renders TRPV1 a downstream target for both NGF and the bradykinin receptor B2R.
Generally, TRPV1 sensitization involves receptor phosphorylation by PKA, PKC and CaMKII, whereas desensitization involves dephosphorylation by phosphatases such as
calcineurin (CN). Under resting conditions, TRPV1 is sequestered in intracellular compartments. When neurons are activated, TRPV1 is transported to the membrane in a
(SNARE)-dependent fashion. Membrane-bound TRPV1 interacts with cytoplasmic proteins as exemplified by b-tubulin (b-TUB). Abbreviations: AC, adenylyl cyclase; DAG,
diacylglycerol; EP1R, prostaglandin E2 receptor 1; EP2R, prostaglandin E2 receptor 2; Gs, stimulatory G protein; IP3, inositol triphosphate; SNARE, soluble
N-ethylmaleimide-sensitive factor.
Review TRENDS in Molecular Medicine Vol.12 No.11 547
pro-inflammatory feedback loop. A role for TRPV1 in hairgrowth has also been implied [27], linking this receptor tocell growth and differentiation. Interestingly, ablation byneonatal capsaicin treatment of TRPV1-expressing nervescan lead to focal alopecia, and capsaicin showed promise inthe treatment of psoriasis [28]. Originally, these actionswere attributed to depletion by capsaicin of substance P(SP) and calcitonin gene-related peptide (CGRP) – sensoryneuropeptides that stimulate keratinocyte proliferation invitro and markedly dilate cutaneous vessels, respectively[2]. Although sensory neuropeptides might regulate cuta-neous blood flow and skin homeostasis, these findingsdeserve a re-evaluation in light of the presence of TRPV1in keratinocytes [26,27].
A related phenomenon is the change in TRPV1expression during carcinogenesis. Up to now, the findingsare contradictory and no clear pattern is emerging. On theone hand, TRPV1 shows a gradual loss in immunoreactiv-ity in transitional cell carcinoma, with low-grade papillarycarcinoma being strongly TRPV1 positive and invasivecarcinoma being TRPV1 negative [29]. On the other hand,TRPV1 is apparently absent in normal uterine cervicalepithelium, whereas it is not only detectable but alsomediates apoptosis in response to agonists such as ana-ndamide in cervical carcinoma cells [30].
The biological function of TRPV1 in other tissuesremains elusive and questionable. An incomplete list ofsuch tissues includes gastric epithelial cells and entero-cytes [31], airway epithelial cells [32], liver [33], vascularendothelium [34], mast cells [2] and soft tissue (smoothmuscle and fibroblasts) [24]. The interrelation, if any,observed in different tissues between TRPV1 expressedin sensory nerve terminals and that expressed in adjacentcells of various types might result in either a synergistic
www.sciencedirect.com
action or an opposing effect, sometimes of difficultinterpretation. This dual (neural and non-neural) expres-sion of TRPV1 in diverse peripheral tissues might explainthe frequent conflicting results that have been obtainedwith TRPV1 agonists and antagonists and TRPV1 knock-out animals in the stomach, airways, and immune andvascular systems.
TRPV1 is regulated at multiple levels providingopportunities for pharmacological interventionTRPV1 is now recognized as a molecular integrator ofvarious noxious stimuli, both physical (noxious heat) andchemical (e.g. capsaicin analogs, protons and lipid deriva-tive endovanilloids) (Figure 1). This last group of moleculesincludes diverse agents such as cannabimimetic lipids (e.g.anandamide [19,20,35],N-arachidonoyl dopamine [36] andother endogenous unsaturated C18 N-acetylethanola-mines [37]), eicosanoid precursors (e.g. 12- and 5-hydro-peroxy-eicosatetraeonic acids) [38] and the satiety factorN-oleoylethanolamide [39]. Some agents directly open thechannel, whereas others act allosterically to reducethe heat activation threshold of TRPV1 (Figure 1). Amongthe endovanilloids (endogenous TRPV1 ligands), N-oleoy-lethanolamide has attracted recent attention as ananorexic lipid mediator [40] that reduces food intake inwild-type, but not in TRPV1-deficient, mice [41]. Thesefindings imply a novel role for TRPV1 in appetiteregulation.
Inflammatory mediators that act together can enhancethe potency of TRPV1 agonists (Figure 1). At the molecularlevel, TRPV1 is under the inhibitory control of phosphati-dylinositol bisphosphate (PIP2); hence, it can be liberatedfrom this control by phospholipase C (PLC), an enzymethat cleaves PIP2 [42] (Figure 1). This is important because
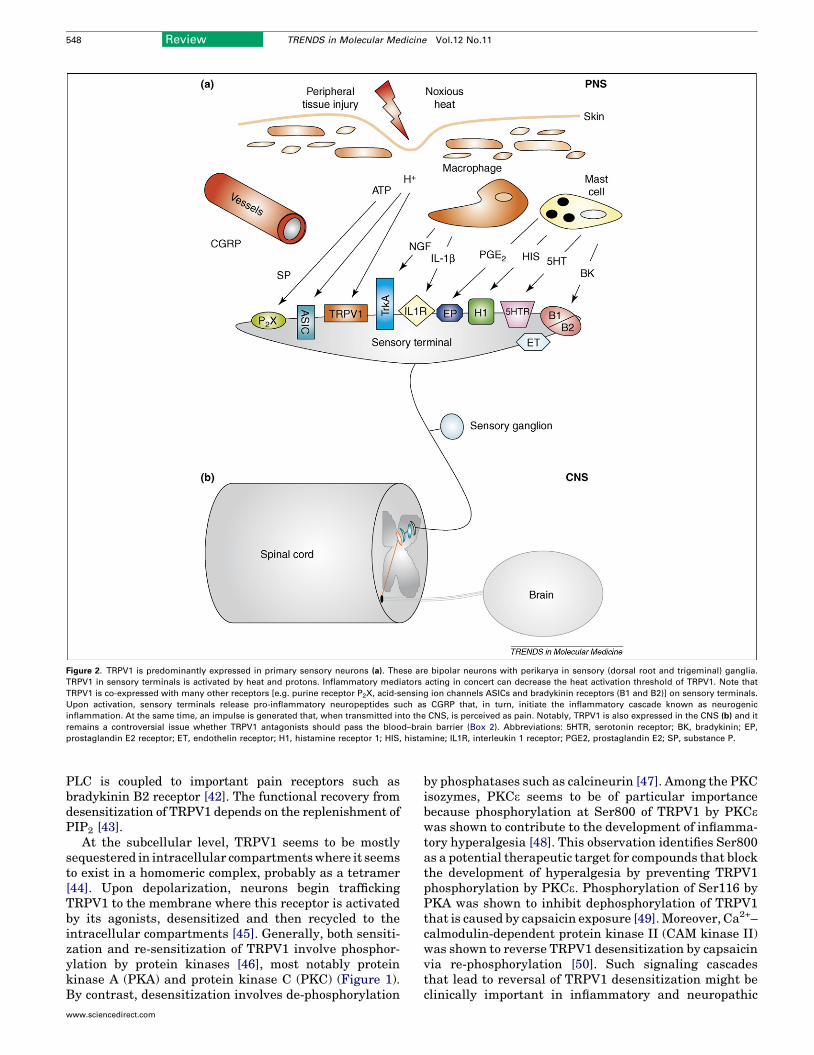
Figure 2. TRPV1 is predominantly expressed in primary sensory neurons (a). These are bipolar neurons with perikarya in sensory (dorsal root and trigeminal) ganglia.
TRPV1 in sensory terminals is activated by heat and protons. Inflammatory mediators acting in concert can decrease the heat activation threshold of TRPV1. Note that
TRPV1 is co-expressed with many other receptors [e.g. purine receptor P2X, acid-sensing ion channels ASICs and bradykinin receptors (B1 and B2)] on sensory terminals.
Upon activation, sensory terminals release pro-inflammatory neuropeptides such as CGRP that, in turn, initiate the inflammatory cascade known as neurogenic
inflammation. At the same time, an impulse is generated that, when transmitted into the CNS, is perceived as pain. Notably, TRPV1 is also expressed in the CNS (b) and it
remains a controversial issue whether TRPV1 antagonists should pass the blood–brain barrier (Box 2). Abbreviations: 5HTR, serotonin receptor; BK, bradykinin; EP,
prostaglandin E2 receptor; ET, endothelin receptor; H1, histamine receptor 1; HIS, histamine; IL1R, interleukin 1 receptor; PGE2, prostaglandin E2; SP, substance P.
548 Review TRENDS in Molecular Medicine Vol.12 No.11
PLC is coupled to important pain receptors such asbradykinin B2 receptor [42]. The functional recovery fromdesensitization of TRPV1 depends on the replenishment ofPIP2 [43].
At the subcellular level, TRPV1 seems to be mostlysequestered in intracellular compartmentswhere it seemsto exist in a homomeric complex, probably as a tetramer[44]. Upon depolarization, neurons begin traffickingTRPV1 to the membrane where this receptor is activatedby its agonists, desensitized and then recycled to theintracellular compartments [45]. Generally, both sensiti-zation and re-sensitization of TRPV1 involve phosphor-ylation by protein kinases [46], most notably proteinkinase A (PKA) and protein kinase C (PKC) (Figure 1).By contrast, desensitization involves de-phosphorylation
www.sciencedirect.com
by phosphatases such as calcineurin [47]. Among the PKCisozymes, PKCe seems to be of particular importancebecause phosphorylation at Ser800 of TRPV1 by PKCewas shown to contribute to the development of inflamma-tory hyperalgesia [48]. This observation identifies Ser800as a potential therapeutic target for compounds that blockthe development of hyperalgesia by preventing TRPV1phosphorylation by PKCe. Phosphorylation of Ser116 byPKA was shown to inhibit dephosphorylation of TRPV1that is caused by capsaicin exposure [49].Moreover, Ca2+–calmodulin-dependent protein kinase II (CAM kinase II)was shown to reverse TRPV1 desensitization by capsaicinvia re-phosphorylation [50]. Such signaling cascadesthat lead to reversal of TRPV1 desensitization might beclinically important in inflammatory and neuropathic

Review TRENDS in Molecular Medicine Vol.12 No.11 549
pain conditions. Of note, the phosphorylation state ofTRPV1 might be disease-specific. This is particularlyinteresting because phosphorylation by PKC of TRPV1can turn some antagonists into agonists [51].
At the cellular level, both upregulation anddownregulation of the gene encoding TRPV1 has beendemonstrated. In the rat, reduced levels of TRPV1mRNA were found following vanilloid desensitization[52], which is consistent with the reversible loss ofTRPV1 protein in these animals [2]. By contrast, ele-vated levels of TRPV1 mRNA and protein were shown inanimal models of inflammatory hyperalgesia [53]. Thelatter finding is in agreement with the increase inTRPV1-like immunoreactivity detected in painful humanconditions such as inflammatory bowel disease [54],vulvodynia [55] and mastalgia [56].
Disease-specific regulation of TRPV1Disease-related changes in TRPV1 expression arebi-directional. As discussed earlier, increased TRPV1-immunoreactive fiber innervation was demonstrated ininflamed human skin [56], vulva [55] and gastrointest-inal tract [54], in addition to tooth pulp with caries [57].This increase in TRPV1 expression was suggested tocontribute to the pathogenesis of various disease states(e.g. reflux esophagitis, inflammatory bowel disease,irritable bowel syndrome, vulvar allodynia and prurigonodularis), and implies a therapeutic value for TRPV1antagonists in their management. Animal experimentsalso showed elevated TRPV1 expression in sensory neu-rons in osteoarthritis [58]. This finding is in agreementwith the attenuation of experimental arthritis in TRPV1-deficient mice [59].
In contrast, downregulation by axotomy of TRPV1 insmall sensory neurons was demonstrated in the rat [60].Unexpectedly, the loss of TRPV1 in small (C-type) neuronswith unmyelinated fibers was accompanied by a concomi-tant fivefold expansion of TRPV1-positive large (A-type)neurons with myelinated fibers [61]. It can be speculatedthat this upregulated expression of TRPV1 in myelinatedfibers (A-neurons) is part of the ‘neuropathic painphenotypic switch’ and contributes to hyperalgesia. Asimilar phenomenon might also have an essential role inthe development and maintenance of diabetic neuropathy.In diabetic rats, the increased TRPV1 expression in A-neurons results in marked increases in both capsaicin- andproton-activated inward currents [62]. The role of TRPV1in diabetic neuropathy is, however, controversial; hyper-algesia was shown to develop earlier and be more severe inTRPV1-deficient mice compared with wild-type animals inthe streptozoicin model of diabetes [63]. It was proposedthat activation of TRPV1 by endovanilloids might releaseendogenous analgesic substances such as somatostatin[63]. In turn, somatostatin might negatively regulateTRPV1.
The mechanisms that can upregulate or downregulateTRPV1 expression are poorly understood. If elevatedTRPV1 has a role in the pathogenesis of certain diseases,then the pharmacological manipulation of the pathwaysthat upregulate TRPV1 has an untapped therapeuticpotential. In the focus of current interest are neurotrophic
www.sciencedirect.com
factors, most notably nerve growth factor (NGF). NGF isoverproduced in peripheral tissues during inflammationfrom where it is retrogradely transported to the cell bodiesof vanilloid-sensitive neurons [2]. NGF increases TRPV1protein [64], but not mRNA, levels in dorsal root ganglia(DRG) neurons in a p38 mitogen-activated protein kinase(MAPK)-dependent fashion, and via the RAC1–nicotina-mide adenine dinucleotide phosphate oxidase (NADPHoxidase) pathway [65]. In agreement with these results,neurotrophin-3, a negative modulator of NGF-receptor(trkA) expression, was shown to prevent upregulation ofTRPV1 mRNA and protein levels following chronic con-striction injury [66]. NGF might also directly sensitizeTRPV1 [42], and elevated NGF levels were found in nasalsecretions of patients with sensory hyper-reactivity of theairway mucosa after capsaicin challenge [67]. Takentogether, these findings imply a central and complex rolefor NGF in the regulation of TRPV1.
TRPV1 in bladder disordersFrom a theoretical point of view, TRPV1 antagonists are ofspecial interest in bladder disorders because TRPV1,whether overexpressed or not, might be activated by endo-genous ligands that can accumulate in the diseased tissue(Figure 2). Therefore, the most-obvious clinical applicationfor TRPV1 antagonists is in the treatment of pain andbladder hyperactivity that accompanies interstitial cysti-tis, a chronic inflammatory condition of the bladder ofunknown etiology. This notion has gained experimentalsupport by the demonstration of abnormal TRPV1responses to capsaicin in a feline model of interstitialcystitis that, similar to diabetes, are secondary to TRPV1phosphorylation by PKC [68]. In addition, TRPV1-likeimmunoreactivity is elevated in the neurogenic humanbladder [69]. Of relevance is the demonstration of elevatedNGF levels that accompany enhanced TRPV1 mRNAlevels in experimental detrusor instability [70].
Experimental evidence suggests that in models ofbladder inflammation the number of bladder nociceptivefibers that usually express TRPV1 is increased [71].Inflammation was shown to increase TRPV1 translationand to remove the receptor from the inhibitory action ofPIP2 [42]. Chronically inflamed bladders accumulateanandamide, a lipid that increases bladder reflex activityof intact bladders and enhances spinal c-fos expressionthrough a TRPV1-dependent mechanism [72]. Protons,another TRPV1 agonist [2,4], also increase in inflamedbladders. Finally, capsazepine decreases, albeit par-tially, the bladder reflex activity of chronic inflamedbladders [72].
Despite the mounting evidence that suggests atherapeutic value for TRPV1 antagonists in the sympto-matic treatment of interstitial cystitis (IC), a word ofcaution seems reasonable at the moment. Resiniferatoxin(RTX) has been assayed intravesically in IC patients withthe expectation that TRPV1 desensitization should be ableto decrease both pain and urinary frequency. The results ofthe two available placebo-controlled clinical trials areconflicting: one suggests clinical utility [73], whereas theother does not demonstrate any advantage from TRPV1desensitization [74].

550 Review TRENDS in Molecular Medicine Vol.12 No.11
Patients with neurogenic and non-neurogenic forms ofdetrusor overactivity, in contrast with IC patients,responded positively to intravesical RTX. The rationalewas based on the experimental evidence that a C-fiber-driven micturition reflex, usually inactive in the adult life,becomes enhanced and takes control of micturition in theoveractive bladder. A pooled analysis of the availableclinical trials in neurogenic and non-neurogenic casesindicates that RTX might increase bladder capacity by>40% and decrease the number of daily incontinenceepisodes by 60% [75]. Furthermore, in addition to provid-ing symptomatic relief, RTX given intravesically restoresnormal TRPV1 levels in the bladder [76]. It is, however,unclear if the clinical effect is due to the decrease levels ofTRPV1 or of other receptors, namely P2X3. In addition, noendovanilloid was identified in high concentration in thebladder of patients with detrusor overactivity. Therefore,the effect of a strong TRPV1 antagonist on urinaryfrequency and incontinence of patients with detrusoroveractivity is still unpredictable.
Migraine and other types of pain or inflammatoryconditionsThe ability to reduce thermal, but more importantlymechanical, hyperalgesia is a paramount feature ofanalgesic compounds. Studies in TRPV1-deficient miceclearly showed the role of TRPV1 in thermal, but notmechanical, hyperalgesia [6,7]. The old-generation antago-nist capsazepine confirmed this anti-hyperalgesic profile inthe rat but, surprisingly, it reduced also mechanical hyper-algesia in the guinea pig [77]. Recently developed antago-nists associated the ability to inhibit thermal andmechanical hyperalgesia in diverse pain models (Box 1).
At the recent American Pain Society meeting (2006) forthe first time a study in humans that reports the pharma-codynamic activity and anti-hyperalgesic properties of theTRPV1 antagonist SB-705498 was presented [78]. Thiscompound at a dose of 400 mg was safe and well tolerated,and reduced the areas of capsaicin- and ultraviolet B(UVB)-evoked flare. More importantly, SB-705498 reducedthe sensitivity to noxious heat at the site of capsaicin andUVB-evoked inflammation compared with placebo. Thus,it was concluded that SB-705498 has target-specific phar-macodynamic activity in humans, and the magnitude ofthe effect in inflammatory pain produced by UVB orcapsaicin was comparable to that of non steroidalanti-inflammatory drugs (NSAIDs).
The peculiar desensitizing property associated toTRPV1 activation has been largely used in different formsof neuropathic and inflammatory pain. Desensitization tocapsaicin has been used also to treat successfully cluster-headache patients [79] and, although in a limited numberof cases, patients with chronic migraine [80]. Capsaicintreatment, however, has important drawbacks includingthe acute pain response that limits its use to topical andtemporary applications [2]. By contrast, TRPV1 antago-nists should maintain the beneficial effect with a betterpharmacodynamic profile and enable systemic routes ofadministration. Menstruation and consumption of alco-holic beverages are common triggers of migraine headacheattacks. Estrogens increase TRPV1 sensitivity and ethanol
www.sciencedirect.com
lowers the threshold temperature for TRPV1 activation onsensory nerve terminals [81]. Finally, release of CGRP,possibly promoted by TRPV1 activation from trigeminalperivascular nerve fibers, seems to have a major role inmigraine [82]. These and additional findings offer a ratio-nale for testing TRPV1 antagonists for the acute treatmentof the migraine attack. After >100 years of studies onprimary afferent neurons and neurogenic inflammation,almost 50 years of basic and clinical investigation oncapsaicin and 10 year from the cloning of the TRPV1,the study of the promising anti-hyperalgesic property ofits first antagonist in humans opens new avenues for thestudy of highly needed new classes of analgesics.
TRPV1-deficient mice predict adverse effects forantagonistGenerally, TRPV1-deficientmice have a benign phenotype,including altered bladder function (high-frequencynon-voiding contractions) [25]. Recent findings, however,indicate that such animals might fare much worse thantheir wild-type littermates under pathological conditionssuch as post-ischemic recovery of the heart [83]. The well-known French Paradox – that is the protective action ofalcoholic beverages on coronary artery disease – might bein part explained with the recent finding that ethanol, bystimulating TRPV1 [81], releases CGRP and, by thismechanism, dilates coronary arteries and increases cor-onary blood flow [84] This is worrisome because somepatients on TRPV1 antagonists for pain might be moresusceptible to a heart attack during therapy. Confoundingthe problem is the possible masking of ischemic pain ofcardiac origin by TRPV1 antagonists, blocking C-fibersinnervating the heart. Of note, neuronal TRPV1 is down-regulated in rats with heart failure [85], indicating thatTRPV1-ligand effects can be unpredictable in patients withcomplex cardiovascular problems.
Mechanical hypersensitivity has an important role inthe abdominal pain experienced by patients with irritablebowel syndrome (IBS). In IBS patients, increased TRPV1levels were demonstrated in bowel biopsies [86]. It ispuzzling that increased susceptibility to experimentalcolitis was described in mice that lack TRPV1 [87], despiteprevious reports that TRPV1 antagonists attenuatedisease severity in such animals [88]. These findings sug-gest a protective role for TRPV1 against some diseases andindicate that TRPV1 antagonistsmight causemore diverseand severe side effects than initially believed. At present, itis unclear to what degree these findings apply to humans.
Future directionsThe TRPV1 literature is vast and suffers from manyunconfirmed and/or conflicting results. For example,recently both the presence [89] and absence [90] of TRPV1in mouse dendritic cells have been reported. Such discre-pant results need to be resolved. The function of TRPV1 inphysiological functions remains poorly understood. Thereis a clear gap between the unremarkable phenotype ofTRPV1-deficientmice and the broad interference of TRPV1antagonists with physiological functions, ranging frombody temperature regulation through motor functions toblood-pressure regulation. For instance, systemic TRPV1

Box 3. Outstanding questions
� Is it a desirable trait for a TRPV1 antagonist to pass the blood–brain
barrier?
Pro: Abbott compound with poor CNS penetration (A-795614)
exhibits much weaker activity in the osteoarthritis model than its
sister compound, A-784168, which freely passes the blood–brain
barrier. This finding implies a key role for central TRPV1 in the
broad-spectrum analgesia by TRPV1 antagonists (of note, it is
unclear if the target is spinal or supraspinal TRPV1).
Con: TRPV1 is expressed throughout the whole neuroaxis where its
physiological role is unknown.
� Is it possible to develop TRPV1 antagonists that prevent activation
by endovanilloids but spare heat sensitivity?
� Can TRPV1 antagonists be developed so that they selectively target
disease-specific phosphorylated form of TRPV1 but do not target
TRPV1 in normal tissues?
� What is the physiological role of TRPV1 in brain nuclei and non-
neuronal tissues? What adverse effects might TRPV1 antagonists
cause by interacting with TRPV1 in cells other than sensory
neurons?
� Can TRPV1 antagonism (associated with the inhibition of CGRP
release) be detrimental at the cardiovascular level?
Review TRENDS in Molecular Medicine Vol.12 No.11 551
antagonist treatment results in an increased bodytemperature [91]. In contrast, no difference in bodytemperature was noted between TRPV1-deficient miceand their wild-type littermates [92]. A potential caveatin the literature is its reliance on first-generation TRPV1antagonists such as capsazepine and iodo-RTX. Whengiven at high doses, these compounds might not only actas partial agonists for TRPV1 [93,94] but also target otherreceptors [2]. The use of indirect means (e.g. the endocan-nabinoid reuptake inhibitor AM404 [95] and the fatty acidamide hydrolase inhibitor URB597 [96]) to increase brainendovanilloid levels and, thereby, study the function ofbrain TRPV1 is an elegant but problematic approach.There is good evidence that anandamide might be gener-ated intracellularly in TRPV1-expressing neurons [97],bypassing the pathway that is sensitive to AM404 andURB597. AM404 was shown to directly interact withTRPV1 [98]. Clearly, the roles of brain TRPV1 should bedelineated by new-generation, selective TRPV1 antago-nists. Given the species-related differences in both theneurochemistry of capsaicin-sensitive neurons [2] andthe pharmacological properties of TRPV1 [99], one shouldexercise utmost caution when extrapolating resultsobtained in rodents to humans. Finally, there is prelimin-ary evidence that TRPV1 pharmacology might also be afunction of the disease state. For example, changes in thephosphorylation state of TRPV1 (that can be disease spe-cific [68]) might have a great influence of the potency ofagonists [100], or turn an agonist into an antagonist or viceversa [51]. In summary, TRPV1-deficient mice andfirst-generation TRPV1 antagonist results need to beconfirmed using new-generation, selective and potentTRPV1 antagonist in in vivo models that are relevant tohumans.
Concluding remarksEndovanilloids that act on TRPV1, which is a molecularintegrator of noxious stimuli, are believed to have a pivotalrole in certain pain conditions. Indeed, desensitization ofTRPV1 to the agonist capsaicin and RTX is a powerfulapproach to relieve inflammatory, post-operative andchronic neuropathic pain [2]. TRPV1 agonists, however,are doing much more than simply desensitizing TRPV1:they render the whole neuron unresponsive to a largeseries of stimuli unrelated to TRPV1 [2]. Mechanisms thatunderlie hyperalgesia and neuropathic pain are complexand incompletely understood (Figure 2). Clearly, manyneuronal receptors other than TRPV1 are involved inthe development and maintenance of hyperalgesia(Figure 2). A list of these receptors include other‘thermo-TRP’ channels, voltage-gated Ca2+, Na+, and K+
channels, NMDA receptors, and bradykinin, purinergicand prostaglandin receptors. Thus, selective TRPV1antagonists should provide some, but not complete, relieffrom pain. This is in line with the clinical experience thatthe GlaxoSmithKline compound SB-705498 [78] has aneffect against inflammatory pain comparable to that ofNSAIDs. In other words, TRPV1 antagonists are likelyto be used in conjunction with other analgesics. But do wereally need anotherNSAID-like drug? The answer dependson the therapeutic window of TRPV1 antagonists as
www.sciencedirect.com
compared with that of other NSAID-like drugs, and theirefficacy relative to existing analgesics against certain painconditions (e.g. neuropathic pain and migraine). For exam-ple, a proportion of migraine patients does not obtain painrelief when they are on triptan-class medications. ATRPV1 antagonist that could eliminate this residual painwould have a clear clinical value. As a matter of fact,GlaxoSmithKline is already actively recruiting patientsfor these trials (http://www.clinicaltrials.gov/ct/giu/show/NCT00269022).
As yet, the only known TRPV1-antagonist-related sideeffect in humans is a transient hyperthermic response. Theclinical significance of this hyperthermic response isunclear but TRPV1 antagonists probably should not beadministered to febrile patients. Nonetheless, there is aclear and unresolved discrepancy between the broad tissuedistribution of TRPV1 in the brain and various non-neu-ronal tissues and apparent lack of side effects of TRPV1antagonists other than elevated body temperature. Nodoubt, additional long-term studies are needed to assessthe efficacy and safety profile of TRPV1 antagonists(Box 3).
In conclusion, TRPV1antagonists alone or in conjunctionwith other analgesics will hopefully significantly improvethe quality of life of people with migraine and chronicintractable pain conditions.
AcknowledgementsWe thank Marcello Trevisani for preparing the figures. Furthermore, weapologize to those authors whose work could not be cited owing to lengthrestrictions.
References1 Krause, J.E. et al. (2005) Transient receptor potential ion channels as
targets for the discovery of pain therapeutics. Curr. Opin. Investig.Drugs 6, 48–57
2 Szallasi, A. and Blumberg, P.M. (1999) Vanilloid (capsaicin) receptorsand mechanisms. Pharmacol. Rev. 51, 159–212
3 Forst, T. et al. (2002) The influence of local capsaicin treatment onsmall nerve fiber function and neurovascular control in symptomaticdiabetic neuropathy. Acta Diabetol. 39, 1–6

552 Review TRENDS in Molecular Medicine Vol.12 No.11
4 Caterina, M.J. et al. (1997) The capsaicin receptor: a heat-activatedion channel in the pain pathway. Nature 389, 816–824
5 Moran, M.M. et al. (2004) TRP ion channels in the nervous system.Curr. Opin. Neurobiol. 14, 362–369
6 Caterina,M.J. et al. (2000) Impaired nociception and pain sensation inmice lacking the capsaicin receptor. Science 288, 306–313
7 Davis, J.B. et al. (2000) Vanilloid receptor-1 is essential forinflammatory thermal hyperalgesia. Nature 405, 183–187
8 Van Abel, M. et al. (2005) The epithelial calcium channels TRPV5 andTRPV6: regulation and implications for disease. NaunynSchmiedebergs Arch. Pharmacol. 371, 295–306
9 Meyer, M.B. et al. (2006) The human transient receptor potentialvanilloid type 6 distal promoter contains multiple vitamin D receptorbinding sites that mediate activation by 1,25-dihydroxyvitamin D3 inintestinal cells. Mol. Endocrinol. 20, 1447–1461
10 Liedtke, W. et al. (2003) Mammalian TRPV4 (VR-OAC) directsbehavioral responses to osmotic and mechanical stimuli inCaenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A. 100 (Suppl2), 14531–14536
11 Alessandri-Haber, N. et al. (2006) A transient receptor potentialvanilloid 4-dependent mechanism of hyperalgesia is engaged byconcerted action of inflammatory mediators. J. Neurosci. 26,3864–3874
12 Peier, A.M. et al. (2002) A heat-sensitive TRP channel expressed inkeratinocytes. Science 296, 2046–2049
13 Xu, H. et al. (2006) Oregano, thyme, and clove-derived flavors and skinsensitizers activate specific TRP channels. Nat. Neurosci. 9, 628–635
14 Liapi, A. and Wood, J.N. (2005) Extensive co-localization andheteromultimer formation of the vanilloid receptor-like proteinTRPV2 and the capsaicin receptor TRPV1 in the adult rat cerebralcortex. Eur. J. Neurosci. 22, 825–834
15 Shimosato, G. et al. (2005) Peripheral inflammation inducedup-regulation of TRPV2 expression in rat DRG. Pain 119, 225–232
16 Yang, X.R. et al. (2006) Functional expression of transient receptorpotential melastatin- and vanilloid-related channels in pulmonaryaterial and aortic smooth muscle. Am. J. Physiol. Lung Cell. Mol.Physiol. 290, L1267–L1276
17 Mezey, E. et al. (2000) Distribution of mRNA for vanilloid receptorsubtype 1 (VR1), and VR1-like immunoreactivity, in the centralnervous system of the rat and human. Proc. Natl. Acad. Sci. U. S. A.97, 3655–3660
18 Cristino, L. et al. (2006) Immunohistochemical localization ofcannabinoid type 1 and vanilloid transient receptor potentialvanilloid type 1 receptors in mouse brain. Neuroscience 139, 1405–1415
19 Pertwee, R.G. (2005) Pharmacological actions of cannabinoids.Handb. Exp. Pharmacol. 168, 1–51
20 De Petrocellis, L. and Di Marzo, V. (2005) Lipids as regulators of theactivity of transient receptor potential type 1 (TRPV1) channels. LifeSci. 77, 1651–1666
21 Lastres-Becker, I. et al. (2003) Compounds acting at theendocannabinoid and/or endovanilloid system reduce hyperkinesiasin a rat model of Huntington’s disease. J. Neurochem. 84, 1097–1109
22 Tzavara,E.T.etal. (2005)Endocannabinoidsactivatetransientreceptorpotential vanilloid 1 receptors to reduce hyperdopaminergia-relatedhyperactivity: therapeutic implications. Biol. Psychiatry 59, 508–515
23 Cabranes, A. et al. (2005) Decreased endocannabinoid levels in thebrain and beneficial effects of agents activating cannabinoid and/orvanilloid receptors in a rat model of multiple sclerosis.Neurobiol. Dis.20, 207–217
24 Birder, L.A. et al. (2001) Vanilloid receptor expression suggests asensory role for urinary bladder epithelial cells. Proc. Natl. Acad. Sci.U. S. A. 98, 13396–13401
25 Birder, L.A. et al. (2002) Altered urinary bladder function in micelacking the vanilloid receptor TRPV1. Nat. Neurosci. 5, 856–860
26 Inoue, K. et al. (2002) Functional vanilloid receptors in culturednormal human epidermal keratinocytes. Biochem. Biophys. Res.Commun. 291, 124–129
27 Bodo, E. et al. (2005) A hot new twist to hair biology: involvement ofvanilloid receptor-1 (VR1/TRPV1) signaling in human hair growthcontrol. Am. J. Pathol. 166, 985–998
28 Arnold, W.P. and van de Kerkhof, P.C. (1994) Topical capsaicin inpruritic psoriasis. J. Am. Acad. Dermatol. 31, 135
www.sciencedirect.com
29 Lazzeri, M. et al. (2005) Transient receptor potential vanilloid type 1(TRPV1) expression changes from normal urothelium to transitionalcell carcinoma of human bladder. Eur. Urol. 48, 691–698
30 Contassot, E. et al. (2004) Arachidonoyl ethanolamide inducedapoptosis of uterine cervix cancer cells via aberrantly expressedvanilloid receptor-1. Gynecol. Oncol. 93, 182–188
31 Kechagias, S. et al. (2005) Expression of vanilloid receptor-1 inepithelial cells of human antral gastric mucosa. Scand. J.Gastroenterol. 40, 775–782
32 Agopyan, N. et al. (2003) Vanilloid receptor activation by 2- and 10micrometer particles induces responses leading to apoptosis in humanairway epithelial cells. Toxicol. Appl. Pharmacol. 192, 21–35
33 Vriens, J. et al. (2004) TRP channels and medulation by hepatocytegrowth factor/scatter factor in human hepatoblastoma (HepG2) cells.Cell Calcium 36, 19–28
34 Golech, S.A. et al. (2004) Human brain endothelium: coexpression andfunction of vanilloid and endocannabinoid receptors. Brain Res. Mol.Brain Res. 132, 87–92
35 Singh Tahim, A. et al. (2005) Inflammatory mediators convertanandamide into a potent activator of the vanilloid type 1transient receptor potential receptor in nociceptive primarysensory neurons. Neuroscience 136, 539–548
36 Huang, S.M. et al. (2002) An endogenous capsaicin-like substancewith high potency at recombinant and native vanilloid VR1 receptors.Proc. Natl. Acad. Sci. U. S. A. 99, 8400–8405
37 Movahed, P. et al. (2005) Endogenous unsaturated C18 N-acylethanolamines are vanilloid receptor (TRPV1) agonists. J. Biol.Chem. 280, 38496–38504
38 Hwang, S.W. et al. (2000) Direct activation of capsaicin receptors byproducts of lipoxygenases: endogenous capsaicin-like substances.Proc. Natl. Acad. Sci. U. S. A. 97, 6155–6160
39 Ahern, G.P. (2003) Activation of TRPV1 by the satiety factoroleoylethanolamide. J. Biol. Chem. 278, 30429–30434
40 Proulx, K. et al. (2005) Mechanisms of oleoylethanolamide-inducedchanges in feeding behavior andmotor activity. Am. J. Physiol. Regul.Integr. Comp. Physiol. 289, R729–R737
41 Wang, X. et al. (2005) Oleoylethanolamide excites vagal sensoryneurons, induces visceral pain and reduces short-term food intakein mice via capsaicin receptor TRPV1. J. Physiol. 564, 541–547
42 Chuang, H.H. et al. (2001) Bradykinin and nerve growth factor releasethe capsaicin receptor from PI(4,5)P2-mediated inhibition. Nature411, 957–962
43 Liu,B. et al. (2005)Functional recovery fromdesensitizationofvanilloidreceptor TRPV1 requires resynthesis of phosphatidylinositol 4,5-biphosphate. J. Neurosci. 25, 4835–4843
44 Garcia-Sanz, N. et al. (2004) Identification of a tetramerizationdomain in the C terminus of the vanilloid receptor. J. Neurosci. 24,5307– 5314
45 Morenilla-Palao, C. et al. (2004) Regulated exocytosis contributes toprotein kinase C potentiation of vanilloid receptor activity. J. Biol.Chem. 279, 25665–25672
46 Suh, Y.G. and Oh, U. (2005) Activation and activators of TRPV1 andtheir pharmaceutical implication. Curr. Pharm. Des. 11, 2687–2698
47 Mohapatra, D.P. and Nau, C. (2005) Regulation of Ca2+-dependentdesensitization in the vanilloid receptor TRPV1 by calcineurin andcAMP-dependent protein kinase. J. Biol. Chem. 280, 13424–13432
48 Mandadi, S. et al. (2006) Increased sensitivity of desensitized TRPV1by PMA occurs through PKCe-mediated phosphorylation at S800.Pain 123, 106–116
49 Bhave, G. et al. (2002) cAMP-dependent protein kinase regulatesdesensitization of the capsaicin receptor (VR1) by directphosphorylation. Neuron 35, 721–731
50 Jung, J. et al. (2004) Phosphorylation of vanilloid receptor 1 by Ca2+/calmodulin-dependent kinase II regulates its vanilloid binding.J. Biol. Chem. 279, 7048–7054
51 Wang, Y. et al. (2003) High-affinity partial agonists of the vanilloidreceptor. Mol. Pharmacol. 64, 325–333
52 Donnerer, J. et al. (2005) Differential regulation of 3-b-hydroxysteroiddehydrogenase and vanilloid receptor TRPV1 mRNA in sensoryneurons by capsaicin and NGF. Pharmacology 73, 97–101
53 Amaya, F. et al. (2004) NGF and GDNF differentially regulate TRPV1expression that contributes to development of inflammatory thermalhyperalgesia. Eur. J. Neurosci. 20, 2303–2310

Review TRENDS in Molecular Medicine Vol.12 No.11 553
54 Yiangou, Y. et al. (2001) Vanilloid receptor 1 immunoreactivity ininflamed human bowel. Lancet 357, 1338–1339
55 Tympanidis, P. et al. (2004) increased vanilloid receptor VR1innervation of vulvodynia. Eur. J. Pain 8, 129–133
56 Gopinath, P. et al. (2005) Increased capsaicin receptor TRPV1 inskin nerve fibers and related vanilloid receptors TRPV3 andTRPV4 in keratinocytes in human breast pain. BMC WomensHealth 5, 2–7
57 Morgan, C.R. et al. (2005) Vanilloid receptor 1 expression in humantooth pulp in relation to caries and pain. J. Orofac. Pain 19, 248–260
58 Fernihough, J. et al. (2005) Regulation of calcitonin gene-relatedpeptide and TRPV1 in a rat model of osteoarthritis. Neurosci. Lett.388, 75–80
59 Barton, N.J. et al. (2006) Attenuation of experimental arthritis inTRPV1 knockout mice. Exp. Mol. Pathol DOI: 10.1016/j.yexmp.2006.04.007
60 Michael, G.J. and Priestley, J.V. (1999) Differential expression of themRNA for the vanilloid receptor subtype 1 in cells of the adultrat dorsal root and nodose ganglia and its downregulation byaxotomy. J. Neurosci. 19, 1844–1854
61 Ma, W. et al. (2005) Medium and large injured dorsal root ganglioncells increase TRPV1, accompanied by increased a2C-adrenoceptorco-expression and functional inhibition by clonidine. Pain 113,386–394
62 Hong, S. and Wiley, J.W. (2005) Early painful diabetic neuropathy isassociated with differential changes in the expression and function ofvanilloid receptor 1. J. Biol. Chem. 280, 618–627
63 Bolcskei, K. et al. (2005) Investigation of the role of TRPV1 receptorsin acute and chronic nociceptive processes using gene-deficient mice.Pain 117, 368–376
64 Zhang, X. et al. (2005) NGF rapidly increases membrane expression ofTRPV1 heat-gated ion channels. EMBO J. 24, 4211–4223
65 Puntambekar, P. et al. (2005) Essential role of Rac1/NADPH oxidasein nerve growth factor induction of TRPV1 expression. J. Neurochem.95, 1689–1703
66 Wilson-Gerwing, T.D. et al. (2005) Neurotrophin-3 suppressesthermal hyperalgesia associated with neuropathic pain andattenuates transient receptor potential vanilloid receptor-1expression in adult sensory neurons. J. Neurosci. 25, 758–767
67 Millqvist, E. et al. (2005) Changes in levels of nerve growth factor innasal secretions after capsaicin inhalation in patients with airwaysymptoms from scents and chemicals. Environ. Health Perspect. 113,849–852
68 Sculptoreanu, A. et al. (2005) Protein kinase C contributes toabnormal capsaicin responses in DRG neurons from cats withfeline interstitial cystitis. Neurosci. Lett. 381, 42–46
69 Brady, C.M. et al. (2004) Parallel changes in bladder suburothelialvanilloid receptor TRPV1 and pan-neuronal marker PGP9.5immunoreactivity in patients with neurogenic detrusoroveractivity after intravesical resiniferatoxin treatment. BJU Int.93, 770–776
70 Kim, J.C. et al. (2004) Nerve growth factor and vanilloid receptorexpression, and detrusor instability, after relieving bladder outletobstruction in rats. BJU Int. 94, 915–918
71 Dickson, A. et al. (2006) Peptidergic sensory and parasympatheticfiber sprouting in the mucosa of the rat urinary bladder in a chronicmodel of cyclophosphamide-induced cystitis. Neuroscience 139,671–685
72 Dinis, P. et al. (2004) Anandamide-evoked activation of vanilloidreceptor 1 contributes to the development of bladder hyperreflexiaand nociceptive transmission to spinal dorsal horn neurons in cystitis.J. Neurosci. 24, 11253–11263
73 Lazzeri, M. et al. (2004) Intravesical infusion of resiniferatoxin by atemporary in situ drug delivery system to treat interstitial cystitis: apilot study. Eur. Urol. 45, 98–102
74 Payne, C.K. et al. (2005) Intravesical resiniferatoxin for the treatmentof interstitial cystitis : a randomized, double-blind, placebo controlledtrial. J. Urol. 173, 1590–1594
75 Cruz, F. (2004) Mechanism involved in new therapies for overactivebladder. Urology 63 (Suppl 1), 65–73
76 Brady, C.M. et al. (2004) P2X3-immunoreactive nerve fibers inneurogenic dtrusor overactivity and the effect of intravesicalresiniferatoxin. Eur. Urol. 46, 247–253
www.sciencedirect.com
77 Walker, K.M. et al. (2003) The VR antagonist capsazepine reversesmechanical hyperalgesia in models of inflammatory and neuropathicpain. J. Pharmacol. Exp. Ther. 304, 56–62
78 Chizh, B et al. (2006) The TRPV1 antagonist SB705498 attenuatesTRPV1 receptor-mediated activity and inhibits inflammatoryhyperalgesia in humans: results from a phase I study. Am. PainSoc. Meeting, TX, USA, Abstract 6231
79 Sicuteri, F. et al. (1990) Substance P theory: a unique focus on thepainful and painless phenomena of cluster headache. Headache 30,69–79
80 Fusco, B.M. et al. (2003) Repeated intranasal capsaicin applications totreat chronic migraine. Br. J. Anaesth. 90, 812
81 Trevisani, M. et al. (2002) Ethanol elicits and potentiates noiceptorresponses via the vanilloid receptor-1. Nat. Neurosci. 5, 546–551
82 Geppetti, P. et al. (2005) CGRP and migraine: neurogenicinflammation revisited. J. Headache Pain 6, 61–70
83 Wang, L. and Wang, D.H. (2005) TRPV1 gene knockout impairspostischemic recovery in isolated perfused heart in mice.Circulation 112, 3617–3623
84 Gazzieri, D. et al. (2006) Ethanol dilates coronary arteries andincreases coronary flow via transient receptor potential 1 andcalcitonin gene-related peptide. Cardiovasc. Res. 70, 589–599
85 Smith, S.A. et al. (2005) The capsaicin-sensitive afferent neuron inskeletal muscle is abnormal in heart failure. Circulation 111,2056–2065
86 Chan, C.L. et al. (2003) Sensory fibres expressing capsaicin receptorTRPV1 in patients with rectal hypersensitivity and faecal urgency.Lancet 361, 385–391
87 Massa, F. et al. (2005) Vanilloid receptor (TRPV1)-deficient mice showincreased susceptibility to dinitrobenzene sulfonic acid inducedcolitis. J. Mol. Med. 83, 944–954
88 Kimball, E.S. et al. (2004) Vanilloid receptor 1 antagonists attenuatedisease severity in dextran sulphate sodium-induced colitis in mice.Neurogastroenterol. Motil. 16, 811–818
89 Basu, S. and Srivastava, P. (2005) Immunological role of neuronalvanilloid receptor 1 expressed on dendritic cells. Proc. Natl. Acad. Sci.U. S. A. 102, 5120–5125
90 O’Connell, P.J. et al. (2005) Dendritic cells do not transduceinflammatory stimuli via the capsaicin receptor TRPV1. FEBSLett. 579, 5135–5139
91 Swanson, D.M. et al. Identification and biological evaluation of 4-(3-trifluoromethylpyridin-2-yl)piperazine-1-carboxylic acid (5-trifluoromethylpyridin-2-yl)amide, a high affinity TRPV1 (VR1)vanilloid receptor antagonist. J. Med. Chem. 48, 1857–1872
92 Iida, T. et al. (2005) Attenuated fever response inmice lacking TRPV1.Neurosci. Lett. 378, 28–33
93 Shimizu, I. et al. (2005) 5-Iodoresiniferatoxin evokes hypothermia inmice and is a partial transient receptor potential vanilloid 1 agonist invitro. J. Pharmacol. Exp. Ther. 314, 1378–1385
94 Petho, G. et al. (2004) Effects of TRPV1 receptor antagonists onstimulated iCGRP release from isolated skin of rats and TRPV1mutant mice. Pain 109, 284–290
95 Maione, S. et al. (2006) Elevation of endocannabinoid levels in theventrolateral periaqueductal grey matter through inhibition of fattyacid amide hydrolase affects descending nociceptive pathways viaboth cannabinoid receptor type 1 and transient receptor potentialvanilloid type 1 receptors. J. Pharmacol. Exp. Ther. 316, 969–982
96 Lee, J. et al. (2006) A role for vanilloid receptor 1 (TRPV1) andendocannabinoid signaling in the regulation of spontaneous and L-DOPA induced locomotion in normal and reserpine-treated rats.Neuropharmacol DOI: 10.1016/j.neuropharm.2006.04.016
97 Van der Stelt, M. et al. (2005) Anandamide acts as an intracellularmessenger amplifying Ca2+ influx via TRPV1 channels. EMBO J. 24,3026–3037
98 Zygmunt, P.M. et al. (2000) The anandamide transport inhibitorAM404 activates vanilloid receptors. Eur. J. Pharmacol. 396, 39–42
99 McIntyre, P. et al. (2001) Pharmacological differences between thehuman and rat vanilloid receptor 1 (VR1). Br. J. Pharmacol. 132,1084–1094
100 Lizanecz, E. et al. (2006) Phophorylation-dependent desensitizationby anadamide of vanilloid receptor-1 (TRPV1) function in rat skeletalmuscle arterioles and in Chinese hamster ovary cells expressingTRPV1. Mol. Pharmacol. 69, 1015–1023

554 Review TRENDS in Molecular Medicine Vol.12 No.11
101 Honore, P. et al. (2005) A-425619 [1-isoquinolin-5-yl-3-(4-trifluoromethyl-benzyl)-urea], a novel transient receptor potentialtype V1 receptor antagonist, relieves pathophysiological painassociated with inflammation and tissue injury in rats.J. Pharmacol. Exp. Ther. 314, 410–421
102 Gavva, N.R. et al. (2005) AMG 9810 [(E)-3-(4-t-butylphenyl)-N-(2,3-dihydrobenzo[b][1,4]dioxin-6-yl_acrylamide], a novel vanilloidreceptor 1 (TRPV1) antagonist with antihyperalgesic properties.J. Pharmacol. Exp. Ther. 313, 474–484
Endeav
The quarterly magazine
philosophy o
You can access Ende
ScienceDirect, where you
editorial comment and a c
illustrated articles on th
Featuri
Death by hypnosis: an 189its European reverberat
NASA and the search for life iLinnaeus’ herbarium cabinet: a piece of furn
Civilising missions, natural history and British indFreudian snaps
Coming
Making sense of modernity’s maladies: economies of healEngineering fame: Isambard K
Disputed discovery: vivisection and experim"A finer and fairer future": commodifying wage earn
‘But man can do his duty’: Charles Darwin
And much, mu
Endeavour is available on Science
www.sciencedirect.com
103 Pomonis, J.D. et al. (2003) N-(4-Tertiarybutylphenyl)-4-(3-chorolphyridin-2-yl)tetrahydropyrazine-1(2H)-carbox-amide(BCTC), a novel, orally effective vanilloid receptor 1 antagonist withanalgesic properties: II. in vivo characterization in rat models ofinflammatory and neuropathic pain. J. Pharmacol. Exp. Ther. 306,387–393
104 Dax, S. et al. (2002) Vanilloid receptor antagonists: structure activityrelationships via parallel and targeted synthesis. Int. Symp. Med.Chem. 27, 93
our
for the history and
f science.
avour online on
’ll find book reviews,
ollection of beautifully
e history of science.
ng:
4 Hungarian case andions by E. Lafferton
n the universe by S.J. Dick
iture and its function by S. Muller-Wille
ustry: Livingstone in the Zambezi by L. Dritsas
by P. Fara
soon:
th and disease in the industrial revolution by M. Brown
ingdom Brunel by P. Fara
ent in the 19th century by C. Berkowitz
ers in American pulp science fiction by E. Drown
’s Christian belief by J. van der Heide
ch more. . .
Direct, www.sciencedirect.com