
Pharmacokinetic basis for nonadditivity of intraocular pressure lowering in timolol combinations
Upload
independentCategory
view
0download
0
Animal Physiology
Triglyceride-lowering Effect of RespiratoryUncoupling in White Adipose TissueMartin Rossmeisl,* Jan Kovar,† Ivo Syrovy,* Pavel Flachs,* Dagmar Bobkova,† Frantisek Kolar,‡Rudolf Poledne,† and Jan Kopecky*
AbstractROSSMEISL, MARTIN, JAN KOVAR, IVO SYROVY,PAVEL FLACHS, DAGMAR BOBKOVA, FRANTISEKKOLAR, RUDOLF POLEDNE, AND JAN KOPECKY.Triglyceride-lowering effect of respiratory uncoupling inwhite adipose tissue. Obes Res. 2005;13:835–844.Objective: Hypolipidemic drugs such as bezafibrate andthiazolidinediones are known to induce the expression ofmitochondrial uncoupling proteins (UCPs) in white adiposetissue. To analyze the potential triglyceride (TG)-loweringeffect of respiratory uncoupling in white fat, we evaluatedsystemic lipid metabolism in aP2-Ucp1 transgenic micewith ectopic expression of UCP1 in adipose tissue.Research Methods and Procedures: Hemizygous and ho-mozygous transgenic mice and their nontransgenic litter-mates were fed chow or a high-fat diet for up to 3 months.Total TGs, nonesterified fatty acids, and the composition ofplasma lipoproteins were analyzed. Hepatic TG productionwas measured in mice injected with Triton WR1339. Up-take and the use of fatty acids were estimated by measuringadipose tissue lipoprotein lipase activity and fatty acid ox-idation, respectively. Adipose tissue gene expression wasassessed by quantitative reverse transcriptase-polymerasechain reaction.Results: Transgene dosage and the high-fat diet interactedto markedly reduce plasma TGs. This was reflected bydecreased concentrations of very-low-density lipoproteinparticles in the transgenic mice. Despite normal hepatic TG
secretion, the activity of lipoprotein lipase in epididymal fatwas enhanced by the high-fat diet in the transgenic mice ina setting of decreased re-esterification and increased in situfatty acid oxidation.Discussion: Respiratory uncoupling in white fat may lowerplasma lipids by enhancing their in situ clearance and ca-tabolism.
Key words: epididymal fat, lipoprotein lipase, fatty acidoxidation, transgenic mice, uncoupling protein
IntroductionA growing body of evidence suggests that hypertriacyl-
glycerolemia is a risk factor for cardiovascular disease,namely because of associated atherogenic factors, i.e., in-creased concentrations of triglyceride (TG)1-rich lipopro-teins, small low-density lipoprotein (LDL) particles, andlow high-density lipoprotein (HDL)-cholesterol (1,2). Var-ious treatment strategies aimed at decreasing plasma TGconcentration are known to affect the synthesis of lipopro-teins in the liver and/or to increase clearance of TGs byperipheral tissues (3). For instance, the lipid-lowering ef-fects of the widely used hypolipidemic drugs, fibrates, canbe attributed to changes in the hepatic gene expression (4,5).As a result, both reduced secretion of very-low-densitylipoprotein (VLDL) particles and enhanced catabolism ofTG-rich particles contribute to the hypolipidemic effect offibrates (6). In contrast, the antidiabetic agents thiazo-lidinediones (TZD) lower serum TG concentrations in rats,primarily by enhancing serum TG removal through theactivation of lipoprotein lipase (LPL) in adipose tissue,without affecting TG production in vivo (3).
Treatments with TZD and some fibrates such as bezafi-brate also induce gene expression of mitochondrial uncou-
Received for review July 27, 2004.Accepted in final form February 22, 2005.The costs of publication of this article were defrayed, in part, by the payment of pagecharges. This article must, therefore, be hereby marked “advertisement” in accordance with18 U.S.C. Section 1734 solely to indicate this fact.*Department of Adipose Tissue Biology and Center for Applied Genomics, Academy ofSciences of the Czech Republic; ‡Department of Developmental Cardiology, Institute ofPhysiology, Academy of Sciences of the Czech Republic; and †Institute for Clinical andExperimental Medicine and Center for Experimental Cardiovascular Research, Prague,Czech Republic.Address correspondence to Jan Kopecky, Department of Adipose Tissue Biology, Instituteof Physiology, Academy of Sciences of the Czech Republic, Videnska 1083, 142 20 Prague4, Czech Republic.E-mail: [email protected] © 2005 NAASO
1 Nonstandard abbreviations: TG, triglyceride; LDL, low-density lipoprotein; HDL, high-density lipoprotein; VLDL, very-low-density lipoprotein; TZD, insulin-sensitizing drugsthiazolidinediones; LPL, lipoprotein lipase; UCP, mitochondrial uncoupling protein; FA,fatty acid; PCR, polymerase chain reaction; NEFA, non-esterified fatty acid; IDL, interme-diate-density lipoprotein; apo, apolipoprotein; PEPCK, phosphoenolpyruvate carboxyki-nase; AMP, adenosine 5�-monophosphate; AMPK, AMP-activated protein kinase; PPAR,peroxisome proliferator-activated receptor.
OBESITY RESEARCH Vol. 13 No. 5 May 2005 835

pling proteins (UCPs), including UCP1, UCP2, and UCP3,in adipose tissue of rats and mice (7–13). Because all UCPsmay lower mitochondrial ATP production through respira-tory uncoupling (14–16), thus stimulating substrate oxida-tion in mitochondria, it is possible to speculate that theeffect of these lipid-lowering compounds may be caused, inpart, by an increased degradation of fatty acids (FAs) withinadipose tissue depots. However, both fibrates and TZD havea rather complex pattern of action in vivo, and the role ofrespiratory uncoupling per se in adipose tissue in the regu-lation of plasma TGs could not be precisely defined.
Transgenic aP2-Ucp1 mice (17), expressing the UCP1gene from a highly fat-specific (18) aP2 gene promoter,represent an established model of respiratory uncoupling inadipose tissue (19–21). These animals resist the develop-ment of obesity induced by genetic or dietary factors, re-flecting a lower accumulation of TG in all fat depots exceptgonadal fat (17,22). Resistance to obesity results from re-spiratory uncoupling (20) in white fat but not in brown fat(19,23) and is conveyed by the stimulation of endogenoussubstrate oxidation (19) and the depression of in situ FAsynthesis (24) in white fat. In our previous study (22), amild but significant depression of plasma TG in hemizygousaP2-Ucp1 transgenic mice suggested an effect of adiposetissue respiratory uncoupling on lipid metabolism.
The aim of this study was to analyze the mechanism ofthe lipid-lowering effect of respiratory uncoupling in whitefat. Our results show that decreased efficiency of mitochon-drial energy conversion in white fat may lower plasma TGsby enhancing their in situ clearance and catabolism.
Research Methods and ProceduresAnimals
Transgenic aP2-Ucp1 mice (17) were originally obtainedfrom the Jackson Laboratory (Bar Harbor, ME), and themouse colony was established in the animal facility of theInstitute of Physiology. Home-produced hemizygous andhomozygous transgenic male mice, carrying low and highdosages of the aP2-Ucp1 transgene, respectively, togetherwith their nontransgenic littermate controls, were identifiedby real-time quantitative polymerase chain reaction (PCR;LightCycler; Roche, Penzberg, Germany) using primers P1(CCGCAGACGACAGGAAGGTGAAG) and P2 (CTG-ACGGAGCTGGGTTAGGTATGG) specific for the endog-enous aP2 gene promoter (reaction A) and primers P3(GGCCCCCATTGGTCACTCC) and P4 (CTGATGCGG-GCACGAAACC) specific for the aP2-Ucp1 transgene con-struct (reaction B). Transgene dosage was assessed as a ratiobetween the products of reactions A and B. The mice weremaintained on a 12:12-hour light-dark cycle (light startingat 6:00 AM) at 20 °C. After weaning at 4 weeks of age, theanimals were single-caged and allowed unrestricted accessto water and standard chow. At 3 months of age, some mice
were randomly assigned to a high-fat diet, whereas the restwere kept on regular chow and served as controls for theeffect of the high-fat diet (22). The animals were fed ahigh-fat diet for variable periods of time. (See Figure 1 forschematic illustration of the experimental protocol withrespect to age and dietary treatment of animals.) The high-fat diet induced obesity in the control mice, whereas bodyweight was reduced in the aP2-Ucp1 mice proportionally tothe transgene dosage (see Tables 1 and 2 in ref. 23). Ani-mals were killed by cervical dislocation under diethyl etheranesthesia, and epididymal and subcutaneous dorsolumbarwhite fat depots (19) were dissected and stored at –70 °C forthe estimation of LPL activity and RNA analysis. Theexperiments were conducted under the guidelines for theuse and care of laboratory animals of the Institute of Phys-iology.
Plasma Nonesterified Fatty Acids, Total TG,Cholesterol, and Lipoprotein Concentrations
Plasma was obtained from the truncal blood of fastedanimals (16 hours; 6:00 PM to 10:00 AM) and used for theestimation of non-esterified fatty acids (NEFAs; kit 994–75409 NEFA C; Wako Chemicals, Richmond, VA), totalTG, and total cholesterol (Roche Diagnostics, Basel, Swit-zerland). For lipoprotein analysis, plasma was pooled fromsubgroups of five to eight nontransgenic and hemizygoustransgenic mice. Aliquots of fresh plasma (0.75 to 3.1 mL)containing EDTA (1 mg/mL) and benzamidine (0.3 mg/mL)were subjected to sequential ultracentrifugation at densitiesof 1.006 and 1.063 g/mL (100,000 g, 18 hours, 12 °C) in a40.3 rotor (Beckman Instruments, Palo Alto, CA) to sepa-rate VLDL (d � 1.006 g/mL), intermediate-density lipopro-tein (IDL) and LDL fraction (IDL � LDL; d � 1.006 to1.063 g/mL), and HDL (d � 1.063 to 1.21 g/mL). The IDL� LDL fraction was concentrated and desalted on Ultra-free-15 centrifugal filtration devices (Millipore, Bedford,
Figure 1: Schematic illustration of the experimental protocol withrespect to age and dietary treatment of animals at the time whenparticular experiments were performed. Age-matched mice of ap-propriate genotypes fed standard chow served as controls for theeffect of a high-fat diet. 3-HB, 3-hydroxybutyrate.
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
836 OBESITY RESEARCH Vol. 13 No. 5 May 2005

MA). Total TG and total cholesterol in lipoprotein fractionswere measured using enzymatic tests (Roche Diagnostics).The VLDL and IDL � LDL fractions were delipidated, andconcentrations of apolipoprotein (apo) B-100 and apo B-48were determined using a 3% to 20% gradient sodium do-decyl sulfate-polyacrylamide gel electrophoresis (25). Thecorresponding bands on gels were quantified using GS-800Calibrated Densitometer and Quantity One software.Prestained sodium dodecyl sulfate-polyacrylamide gel elec-trophoresis standards (Broad Range) were from BioRadLaboratories (161-0317; Hercules, CA).
Hepatic TG Secretion and KetogenesisA modification of the previously published procedures
(3,26) was used. One day before the experiment, at 6:00 PM,the mouse diet was removed, and the animals were allowedfree access to water and fat-free food (dry cereal). Theexperiment was performed between 9:00 AM and 1:00 PM inthe control and hemizygous transgenic mice anesthetized byintraperitoneal injection of pentobarbital (50 �g/g bodyweight). The animals were kept at 37 °C. The jugular veinwas surgically exposed, and mice were injected intrave-nously with 10% Tyloxapol (Triton WR-1339; T-8761;Sigma-Aldrich, St. Louis, MO; 500 mg/kg body weight).Under these conditions, the nonionic detergent blocks thelipolytic degradation of TG-rich lipoproteins in the periph-eral tissues, and the rise in plasma TG is proportional to theproduction of VLDL in the liver (27). Blood was collectedfrom the jugular vein before and 2 hours after the injection,and plasma TG levels were subsequently assessed to deter-mine their absolute rise during the experiment. The valueswere also expressed relative to liver mass (26). Hepaticketogenesis was assessed by measuring plasma levels of3-hydroxybutyrate in fed animals using a kit Autokit 3-HBfrom Wako Chemicals.
LPL ActivityThe enzyme activity was estimated in detergent extracts
of adipose tissues dissected from fasted mice as describedbefore (28).
FA OxidationOxidation of oleic acid was measured by 3H2O release
from fragments (�35 mg) of adipose tissue as describedbefore (29). Because fasting itself increases FA oxidation inadipose tissue (30), the effect of aP2-Ucp1 transgene on FAoxidation was assessed in adipose tissue of mice in the fedstate. The results are expressed either per milligram ofprotein or per milligram of tissue. Protein concentration wasmeasured using a bicinchoninic acid procedure.
Gene Expression in Adipose TissueTotal RNA was isolated using TRIzol Reagent (Invitro-
gen, Carlsbad, CA). Gene expression was analyzed by re-
verse transcription followed by the real-time quantitativePCR (LightCycler Instrument; F. Hoffman-La Roche, Basel,Switzerland) with primers specific for mouse phosphoenol-pyruvate carboxykinase (PEPCK; forward, CCAGCCAGT-GCCCCATTATTGAC; reverse, TTTGCCGAAGTTGTAG-CCGAAGAA). Expression levels of cyclophilin-� (forward,ACTACGGGCCTGGCTGGGTGAG; reverse, TGCCGGA-GTCGACAATGATGA) mRNA were used to correct for in-ter-sample variations. The detailed protocol has been describedbefore (31).
Statistical AnalysisData were evaluated by ANOVA:single factor (Excel;
Microsoft, Redmond, WA) and by ANOVA with Newman-Keuls posthoc multiple comparison method as describedbefore (22), when indicated. All differences were judged tobe significant at p � 0.05.
ResultsPlasma Lipids
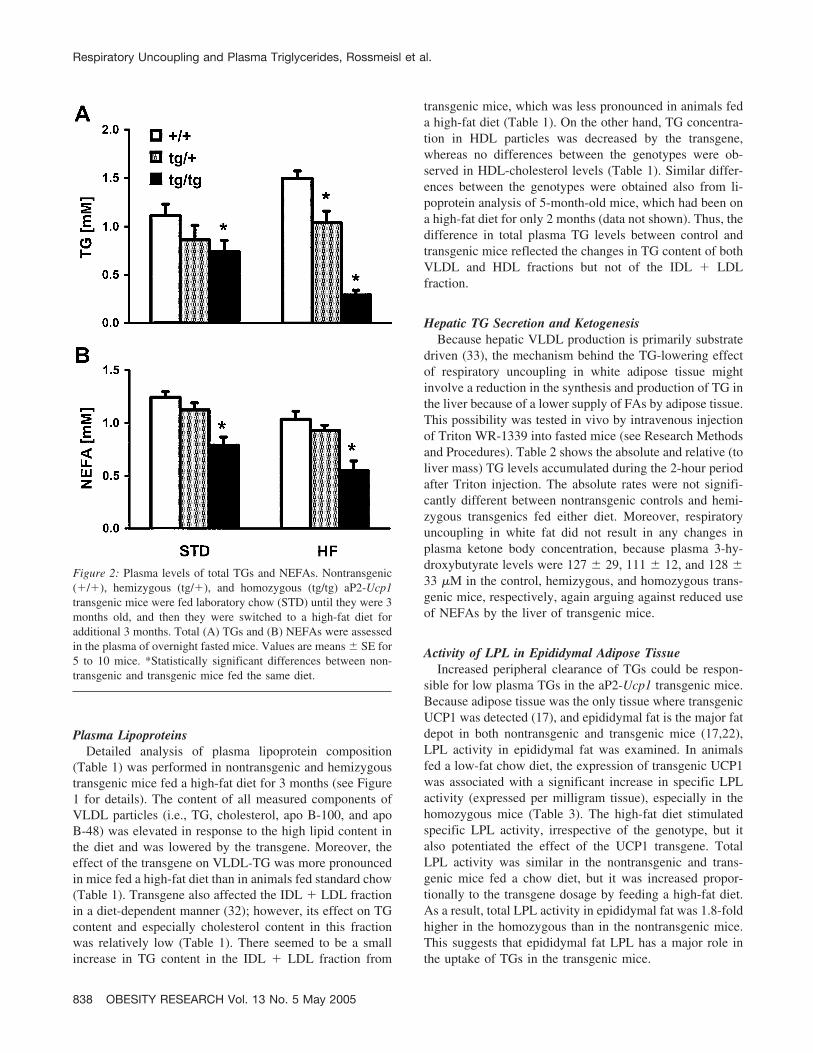
It was shown previously (22) that a mild but significantdepression of plasma TG occurred in hemizygous aP2-Ucp1transgenic mice, irrespective of the fat content in their diet.In this study, we used both hemizygous and homozygoustransgenic mice, i.e., mice that carry various transgenedosages, to analyze in detail the effect of respiratory uncou-pling in white fat on systemic lipid metabolism and itsinteraction with a high-fat diet. (See Figure 1 for detailswith regard to age and dietary treatment of animals.) Figure2 shows plasma TGs in nontransgenic and transgenic mice.Plasma TG levels were reduced by the transgene in adose-dependent manner (Figure 2). The high-fat diet ele-vated plasma TGs in the control nontransgenic mice com-pared with their chow diet-fed counterparts (p � 0.05);however TG levels did not change in the hemizygous trans-genic mice and even decreased (p � 0.01) in the homozy-gotes in response to a high-fat diet. As a result, in thehomozygous transgenics fed a high-fat diet, plasma TGlevels were 2.5-fold lower compared with their counterpartsfed a chow diet (Figure 2). This difference was observed inseveral independent experiments. Plasma NEFAs in thetransgenic mice were also relatively low (Figure 2), al-though significant differences were found only betweencontrol nontransgenic and homozygous transgenic mice. Onthe other hand, cholesterol levels were much less affectedby the transgene in mice that were on a high-fat diet for 2months (see Figure 1 for experimental details), becauseplasma cholesterol concentrations were 2.89 � 0.08, 2.51 �0.05, and 2.47 � 0.18 mM in the nontransgenic, hemizy-gous, and homozygous transgenics, respectively (p � 0.05,control versus transgene). Thus, the overexpression of trans-genic UCP1 in white fat leads to profound effects at thelevel of systemic lipid metabolism.
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
OBESITY RESEARCH Vol. 13 No. 5 May 2005 837
Plasma LipoproteinsDetailed analysis of plasma lipoprotein composition
(Table 1) was performed in nontransgenic and hemizygoustransgenic mice fed a high-fat diet for 3 months (see Figure1 for details). The content of all measured components ofVLDL particles (i.e., TG, cholesterol, apo B-100, and apoB-48) was elevated in response to the high lipid content inthe diet and was lowered by the transgene. Moreover, theeffect of the transgene on VLDL-TG was more pronouncedin mice fed a high-fat diet than in animals fed standard chow(Table 1). Transgene also affected the IDL � LDL fractionin a diet-dependent manner (32); however, its effect on TGcontent and especially cholesterol content in this fractionwas relatively low (Table 1). There seemed to be a smallincrease in TG content in the IDL � LDL fraction from
transgenic mice, which was less pronounced in animals feda high-fat diet (Table 1). On the other hand, TG concentra-tion in HDL particles was decreased by the transgene,whereas no differences between the genotypes were ob-served in HDL-cholesterol levels (Table 1). Similar differ-ences between the genotypes were obtained also from li-poprotein analysis of 5-month-old mice, which had been ona high-fat diet for only 2 months (data not shown). Thus, thedifference in total plasma TG levels between control andtransgenic mice reflected the changes in TG content of bothVLDL and HDL fractions but not of the IDL � LDLfraction.
Hepatic TG Secretion and KetogenesisBecause hepatic VLDL production is primarily substrate
driven (33), the mechanism behind the TG-lowering effectof respiratory uncoupling in white adipose tissue mightinvolve a reduction in the synthesis and production of TG inthe liver because of a lower supply of FAs by adipose tissue.This possibility was tested in vivo by intravenous injectionof Triton WR-1339 into fasted mice (see Research Methodsand Procedures). Table 2 shows the absolute and relative (toliver mass) TG levels accumulated during the 2-hour periodafter Triton injection. The absolute rates were not signifi-cantly different between nontransgenic controls and hemi-zygous transgenics fed either diet. Moreover, respiratoryuncoupling in white fat did not result in any changes inplasma ketone body concentration, because plasma 3-hy-droxybutyrate levels were 127 � 29, 111 � 12, and 128 �33 �M in the control, hemizygous, and homozygous trans-genic mice, respectively, again arguing against reduced useof NEFAs by the liver of transgenic mice.
Activity of LPL in Epididymal Adipose TissueIncreased peripheral clearance of TGs could be respon-
sible for low plasma TGs in the aP2-Ucp1 transgenic mice.Because adipose tissue was the only tissue where transgenicUCP1 was detected (17), and epididymal fat is the major fatdepot in both nontransgenic and transgenic mice (17,22),LPL activity in epididymal fat was examined. In animalsfed a low-fat chow diet, the expression of transgenic UCP1was associated with a significant increase in specific LPLactivity (expressed per milligram tissue), especially in thehomozygous mice (Table 3). The high-fat diet stimulatedspecific LPL activity, irrespective of the genotype, but italso potentiated the effect of the UCP1 transgene. TotalLPL activity was similar in the nontransgenic and trans-genic mice fed a chow diet, but it was increased propor-tionally to the transgene dosage by feeding a high-fat diet.As a result, total LPL activity in epididymal fat was 1.8-foldhigher in the homozygous than in the nontransgenic mice.This suggests that epididymal fat LPL has a major role inthe uptake of TGs in the transgenic mice.
Figure 2: Plasma levels of total TGs and NEFAs. Nontransgenic(�/�), hemizygous (tg/�), and homozygous (tg/tg) aP2-Ucp1transgenic mice were fed laboratory chow (STD) until they were 3months old, and then they were switched to a high-fat diet foradditional 3 months. Total (A) TGs and (B) NEFAs were assessedin the plasma of overnight fasted mice. Values are means � SE for5 to 10 mice. *Statistically significant differences between non-transgenic and transgenic mice fed the same diet.
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
838 OBESITY RESEARCH Vol. 13 No. 5 May 2005

FA Oxidation in Epididymal Fat of Mice Fed a High-fatDiet
Increased delivery of TGs to epididymal adipose tissue oftransgenic mice fed a high-fat diet would also imply in-creased in situ oxidation of FA because of respiratory un-coupling induced by the transgenic UCP1. Therefore, oleateoxidation by epididymal fat was measured in the control,hemizygous, and homozygous transgenic mice fed a high-fat diet for 1 month (Figure 3). Oleate oxidation, expressedeither per milligram of protein (Figure 3A) or per milligramof tissue (Figure 3B), was elevated by the transgene in adose-dependent manner. As a result, it was �2-fold higherin the homozygous transgenic than in nontransgenic mice
when expressed per milligram of protein (Figure 3A), andthe difference between the genotypes was even higher whenFA oxidation was calculated per milligram of tissue (Figure3B).
Effect of Transgene and Diet on the Expression ofPEPCK in Epididymal Fat
Re-esterification of FA within adipocytes is an importantmechanism participating in the regulation of adipose tissuemass (34–36) and could be affected by the expression of theaP2-Ucp1 transgene. As a marker of glyceroneogenesis andFA re-esterification (36), the expression of the PEPCK genewas quantified in epididymal fat of overnight fasted mice
Table 1. Effect of diet on lipoprotein composition in nontransgenic and aP2-Ucp1 transgenic mice
STD HF
�/� tg/� �/� tg/�
VLDL (d � 1.006)TG (mM) 0.22 0.22 0.69 0.37Cholesterol (mM) 0.06 0.06 0.11 0.05Apo B-100 (mg/liter) 3.3 0.9 7.5 3.9Apo B-48 (mg/liter) 1.3 0.7 5.7 2.8
IDL � LDL (1.006 � d � 1.063)TG (mM) 0.10 0.23 0.13 0.16Cholesterol (mM) 0.14 0.13 0.59 0.51Apo B-100 (mg/liter) 81.4 79.4 74.3 120.0Apo B-48 (mg/liter) 3.7 3.1 3.4 4.4
HDL (d � 1.063)TG (mM) 0.57 0.25 0.76 0.52Cholesterol (mM) 1.80 1.94 3.16 2.96
Lipoprotein composition was characterized in pooled plasma samples (five to eight mice per group; see Research Methods and Proceduresfor details) from the control nontransgenic (�/�) and hemizygous (tg/�) aP2-Ucp1 transgenic mice fed either standard chow (STD) or ahigh-fat (HF) diet.
Table 2. Hepatic TG secretion in nontransgenic and aP2-Ucp1 transgenic mice
STD HF
Absolute Relative Liver (g) Absolute Relative Liver (g)
�/� 12.76 � 1.46 9.11 � 0.99 1.50 � 0.56 11.28 � 2.29 7.66 � 1.89 1.66 � 0.21tg/� 13.23 � 1.40 9.19 � 0.78 1.46 � 0.59 10.94 � 1.33 5.83 � 0.60 1.87 � 0.11
Mice (8 to 11 per group) were injected intravenously with Triton WR-1339, and TG levels were assessed 2 hours after injection. Data aregiven as means � SE. Plasma TG levels are expressed either as absolute values (mM) or relative to liver mass (mM/g tissue). Differencesbetween genotypes within each type of diet were not statistically significant.STD, standard chow; HF, high fat.
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
OBESITY RESEARCH Vol. 13 No. 5 May 2005 839

fed either standard chow or a high-fat diet for 2 months(Figure 4). Transgene markedly down-regulated PEPCKgene expression, irrespective of the diet or the dosage of thetransgene (p � 0.01; Figure 4).
DiscussionA major finding of this study was that respiratory uncou-
pling in white adipose tissue is capable of lowering plasmaTGs while enhancing FA oxidation and the activity of LPLin the tissue. This effect was particularly enhanced by ahigh-fat diet. Thus, changes in energy conversion in whiteadipocytes affected the whole-body lipid metabolism andmight contribute to lipid-lowering effects of some fibratesand TZD that increase the expression of UCP homologuesin white adipose tissue (see the Introduction).
Uptake of FA by white adipose tissue, mediated by LPL,is the main mechanism of plasma TG clearance during thepostprandial period (37,38). Results of this study indicatesignificantly higher specific, as well as total, LPL activity inthe epididymal fat depot of transgenic compared with wild-type mice, namely in the homozygous animals, and furtherstimulation of the LPL activity by a high-fat diet. These datasuggest a major role for epididymal fat in FA catabolismand a TG-lowering effect in the aP2-Ucp1 mice fed ahigh-fat diet. It is unlikely that the subcutaneous adiposetissue, which was greatly reduced already in the hemizygoustransgenic mice (17,22) and was 10-fold less in the homozy-
gous transgenics compared with the nontransgenic mice feda high-fat diet (data not shown), contributes significantly toplasma TG clearance in these animals. Interscapular brownadipose tissue could not be involved either, because it isvirtually absent in the homozygous and strongly reduced inthe hemizygous transgenic mice (23). However, the stimu-
Table 3. Effect of diet and genotype on weight andLPL activity in the epididymal fat depot
Tissue weight(mg)
LPL activity
Specific Total
STD�/� 843 � 217 0.20 � 0.01 190 � 79tg/� 897 � 160 0.29 � 0.04* 278 � 74tg/tg 477 � 40†‡ 0.48 � 0.08†‡ 224 � 29
HF�/� 1884 � 134 0.45 � 0.13 820 � 194tg/� 1875 � 220 0.81 � 0.25 1319 � 300tg/tg 1075 � 133†‡ 1.49 � 0.39†‡ 1487 � 339†
Specific LPL activity (nmol FA/min per milligram of tissue) andtotal LPL activity (nmol FA/min per fat depot) were assessed indetergent extracts of adipose tissue. Values are means � SE forfour to six mice.Symbols indicate significant differences (by ANOVA with New-man-Keuls posthoc multiple comparison method): *�/� vs. tg/�;†�/� vs. tg/tg; ‡tg/� vs. tg/tg.STD, standard chow; HF, high fat.
Figure 3: Oxidation of FA in epididymal adipose tissue. Four-month-old nontransgenic (�/�), hemizygous (tg/�), and homozy-gous (tg/tg) aP2-Ucp1 transgenic mice were fed a high-fat diet for1 month. Oleate oxidation was measured by 3H2O release infragments of adipose tissue and expressed either (A) per milligramof protein or (B) per milligram of tissue. Protein concentrations inthe nontransgenic, hemizygous, and homozygous transgenic micewere 7.33 � 1.16, 8.41 � 0.49, and 9.63 � 0.85 mg protein/gtissue, respectively. Data are means � SE (n � 6 for nontransgenicand hemizygous transgenic mice; n � 4 for homozygous trans-genics). *Statistically significant differences between nontrans-genic and transgenic mice.
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
840 OBESITY RESEARCH Vol. 13 No. 5 May 2005

lation of LPL activity in other tissues, triggered by the lowplasma levels of NEFAs, cannot be excluded (39,40).
In the transgenic mice, LPL activity in adipose tissue isapparently stimulated in response to the activation ofFA oxidation in adipocytes of animals fed a high-fat diet(Figure 3). The mechanistic link between intracellular FAoxidation and LPL activity is not clear. However, the aden-osine 5�-monophosphate (AMP)-activated protein kinase(AMPK) that acts as a metabolic master switch activated bya depression in the intracellular energy charge (41) could beinvolved in the stimulation of FA oxidation by respiratoryuncoupling. This kinase is activated and its content in-creased in white fat depots of the aP2-Ucp1 mice, includingepididymal fat (29), where the transgenic UCP1 decreasesmitochondrial membrane potential (20), lowers the ATP/AMP ratio (29), increases mitochondrial biogenesis (21),and stimulates endogenous substrate oxidation (19). Similarto the changes seen in adipose tissue of the aP2-Ucp1transgenic mice, TZD stimulates AMPK (42) and increasesfat use in adipocytes by increasing mitochondrial biogene-sis, FA oxidation, and oxygen consumption (8), while in-ducing the expression of various UCPs (9–12), includingUCP1 (7,8,13).
In contrast to transgenic UCP1 that down-regulatesPEPCK, TZD stimulate the expression of PEPCK (36), therate-limiting enzyme in glyceroneogenesis and marker ofFA re-esterification in adipocytes (36). By this mechanism,and by up-regulation of glycerol kinase (43), TZD stimu-
lates futile TG/FA cycling. Re-esterification of FAs in adi-pocytes plays an important role in intracellular fuel parti-tioning. For instance, in normal fed rats, 49.7% ofendogenous FAs are reesterified, 50.1% are released, andonly 0.2% are oxidized (30). Moreover, increased expres-sion of PEPCK in adipose tissue of mice has been associ-ated with obesity (34), and conversely, its down-regulationthrough a mutation in the promoter binding site for itsregulatory factor peroxisome proliferator-activated receptor(PPAR)-� reduces adipose tissue size (35). Thus, the in-crease in re-esterification of FAs might be a part of themechanism by which TZD support adipogenesis (8). On theother hand, the mechanism responsible for the reduction ofadiposity and the low plasma TG in the aP2-Ucp1 trans-genic mice most likely relies on the induction of fat use andinhibition of in situ lipogenesis (24) in adipocytes ratherthan on futile cycling of FA re-esterification. This conclu-sion is further supported by other findings, where increasedAMPK activity in adipose tissue caused by exercise led to adown-regulation of lipogenesis and FA re-esterificationthrough the inhibition of mitochondrial glycerol-3-phos-phate acyltransferase (44). Reduced expression of PEPCKin epididymal adipose tissue of aP2-Ucp1 transgenic mice ismost likely caused by a down-regulation of its upstreamregulatory factor PPAR-� (29,35). However, reduced FAre-esterification in adipose tissue of transgenic mice couldalso be the result of metabolic adaptations that counteractincreased FA use and reduce NEFA production by theUCP1-expressing adipocytes, similarly to a model of hor-mone-sensitive lipase-deficient mice (45).
Because NEFAs are believed to be the main driving forcefor the production of VLDLs in the liver (33), it can bespeculated that lower plasma NEFA concentrations in trans-genic mice would result in lower VLDL production and,therefore, lower VLDL and TG concentrations. However,we did not observe any significant changes in TG secretionfrom the liver (Table 2). If there are no differences in VLDLproduction, the livers of transgenic mice must mobilize TGfor VLDL-TG production from their intracellular sources tocompensate for their lower circulating NEFA levels. Indeed,it was observed previously (see Figure 3 in ref. 22) that, inthe hemizygous aP2-Ucp1 transgenic mice fed a high-fatdiet, the relative TG content in the liver was �2-fold higherthan in control animals, suggesting increased hepatic denovo lipogenesis as a compensatory mechanism. The factthat there are no differences between control and transgenicmice in the use of NEFAs by the liver is further supportedby the finding that plasma 3-hydroxybutyrate levels as amarker of hepatic ketogenesis were not different betweenthe groups.
Both transgenic mice overexpressing human LPL in mul-tiple tissues including adipose (46) and mice deficient inhormone-sensitive lipase (39) have shown elevated LPL
Figure 4: Gene expression of PEPCK in epididymal adipose tissueof 5-month-old nontransgenic (�/�), hemizygous (tg/�), andhomozygous (tg/tg) aP2-Ucp1 transgenic mice that were fed ahigh-fat (HF) diet for 2 months. Nontransgenic and hemizygoustransgenic mice of the same age fed standard chow (STD) servedas a control for the effect of the HF diet. Overnight fasted micewere analyzed. Bars are means � SE (n � 6 except for hemizy-gous and homozygous transgenic mice fed HF diet with n � 4).*Statistically significant differences between nontransgenic andtransgenic mice fed the same diet.
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
OBESITY RESEARCH Vol. 13 No. 5 May 2005 841

activity in white fat and improved lipoprotein profiles withreduced plasma TG and VLDL-TG in the fasted state,similar to our transgenic model. Increased activity of LPLassociated with UCP1-mediated respiratory uncoupling iswell documented during the activation of brown fat ther-mogenic function by cold exposure (47). Moreover, hyper-leptinemia induced by adenovirus transfer in rats up-regu-lates LPL as well as UCP1, UCP2, and enzymes of FAoxidation in white fat (48,49), while lowering plasma TGs(50). Transgenic overexpression of both transcription factorPPAR-� (51) and winged helix/forkhead transcription factorFOXC2 (52) in adipose tissue of mice induces a markedexpression of the UCP1 gene and a significant reduction inplasma TGs. In the first type of transgenic animals, simi-larly to aP2-Ucp1 mice, the hypotriglyceridemic effect ofthe transgene was augmented by the high-fat diet, whereasplasma cholesterol levels were normal (51). Importantly,compared with the other models (51,52), only in the aP2-Ucp1 mice is it primarily the UCP1-induced respiratoryuncoupling in white but not brown adipocytes (19,21,23)that exerts the hypolipidemic effect. A recent study (53)showing that human PPAR-� coactivator-1� is able to in-crease the expression of human UCP1 in human whiteadipocytes strengthens the view that induction of respiratoryuncoupling in white adipose tissue could be a useful strat-egy for the treatment of obesity and disturbed lipid metab-olism in metabolic disorders such as obesity and type 2diabetes.
In conclusion, the results of this study show that respira-tory uncoupling in adipocytes of white fat leads to profoundchanges of systemic lipid metabolism. Overexpression ofUCP1 in white fat was capable of protecting mice not onlyfrom dietary obesity but also from hyperlipidemia associ-ated with a high-fat feeding. Thus, respiratory uncoupling inwhite fat could play a role in the way some fibrates andTZD exert their hypolipidemic effects.
AcknowledgmentsWe thank Dr. Jaroslav Vorlicek for statistical analysis,
Jaroslava Bemova and Sona Hornova for technical assis-tance, and A. Kotyk for critical reading of the manuscript.This work was supported by grants 303/02/1220 (to J.K.)and 303/03/P127 (to M.R.) from the Grant Agency of theCzech Republic and research project AVOZ 5011922.
References1. Hokanson JE, Austin MA. Plasma triglyceride level is a risk
factor for cardiovascular disease independent of high-densitylipoprotein cholesterol level: a meta-analysis of population-based prospective studies. J Cardiovasc Risk. 1996;3:213–9.
2. Grundy SM. Hypertriglyceridemia, atherogenic dyslipide-mia, and the metabolic syndrome. Am J Cardiol. 1998;81:18B–25B.
3. Lefebvre AM, Peinado-Onsurbe J, Leitersdorf I, et al.Regulation of lipoprotein metabolism by thiazolidinediones
occurs through a distinct but complementary mechanism rel-ative to fibrates. Arterioscler Thromb Vasc Biol. 1997;17:1756–64.
4. Staels B, Vu-Dac N, Kosykh VA, et al. Fibrates downregu-late apolipoprotein C-III expression independent of inductionof peroxisomal acyl coenzyme A oxidase. A potential mech-anism for the hypolipidemic action of fibrates. J Clin Invest.1995;95:705–12.
5. Andersson Y, Majd Z, Lefebvre AM, et al. Developmentaland pharmacological regulation of apolipoprotein C-II geneexpression. Comparison with apo C-I and apo C-III generegulation. Arterioscler Thromb Vasc Biol. 1999;19:115–21.
6. Schoonjans K, Staels B, Auwerx J. Role of the peroxisomeproliferator-activated receptor (PPAR) in mediating the effectsof fibrates and fatty acids on gene expression. J Lipid Res.1996;37:907–25.
7. Digby JE, Montague CT, Sewter CP, et al. Thiazolidinedi-one exposure increases the expression of uncoupling protein 1in cultured human preadipocytes. Diabetes. 1998;47:138–41.
8. Wilson-Fritch L, Nicoloro S, Chouinard M, et al. Mito-chondrial remodeling in adipose tissue associated with obesityand treatment with rosiglitazone. J Clin Invest. 2004;114:1281–9.
9. Cabrero A, Llaverias G, Roglans N, et al. Uncouplingprotein-3 mRNA levels are increased in white adipose tissueand skeletal muscle of bezafibrate-treated rats. Biochem Bio-phys Res Commun. 1999;260:547–56.
10. Cabrero A, Alegret M, Sanchez RM, Adzet T, Laguna JC,Vazquez M. Bezafibrate reduces mRNA levels of adipocytemarkers and increases fatty acid oxidation in primary cultureof adipocytes. Diabetes. 2001;50:1883–90.
11. Matsuda J, Hosoda K, Itoh H, et al. Increased adiposeexpression of the uncoupling protein-3 gene by thiazo-lidinediones in Wistar fatty rats and in cultured adipocytes.Diabetes. 1998;47:1809–14.
12. Emilsson V, O’Dowd J, Wang S, et al. The effects ofrexinoids and rosiglitazone on body weight and uncouplingprotein isoform expression in the Zucker fa/fa rat. Metabolism.2000;49:1610–5.
13. Fukui Y, Masui S, Osada S, Umesono K, Motojima K. Anew thiazolidinedione, NC-2100, which is a weak PPAR-gamma activator, exhibits potent antidiabetic effects and in-duces uncoupling protein 1 in white adipose tissue of KKAyobese mice. Diabetes. 2000;49:759–67.
14. Ricquier D, Bouillaud F. The uncoupling protein homo-logues: UCP1, UCP2, UCP3, StUCP and AtUCP. Biochem J.2000;345:161–79.
15. Echtay KS, Winkler E, Frischmuth K, Klingenberg M.Uncoupling proteins 2 and 3 are highly active H� transportersand highly nucleotide sensitive when activated by coenzymeQ (ubiquinone). Proc Natl Acad Sci U S A. 2001;98:1416–21.
16. Fink BD, Hong YS, Mathahs MM, Scholz TD, Dillon JS,Sivitz WI. UCP2-dependent proton leak in isolated mamma-lian mitochondria. J Biol Chem. 2002;277:3918–25.
17. Kopecky J, Clarke G, Enerback S, Spiegelman B, KozakLP. Expression of the mitochondrial uncoupling protein genefrom the aP2 gene promoter prevents genetic obesity. J ClinInvest. 1995;96:2914–23.
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
842 OBESITY RESEARCH Vol. 13 No. 5 May 2005

18. Pelton PD, Zhou L, Demarest KT, Burris TP. PPARgammaactivation induces the expression of the adipocyte fatty acidbinding protein gene in human monocytes. Biochem BiophysRes Commun. 1999;261:456–8.
19. Kopecky J, Rossmeisl M, Hodny Z, Syrovy I, Horakova M,Kolarova P. Reduction of dietary obesity in the aP2-Ucptransgenic mice: mechanism and adipose tissue morphology.Am J Physiol. 1996;270:E776–86.
20. Baumruk F, Flachs P, Horakova M, Floryk D, Kopecky J.Transgenic UCP1 in white adipocytes modulates mitochon-drial membrane potential. FEBS Lett. 1999;444:206–10.
21. Rossmeisl M, Barbatelli G, Flachs P, et al. Expression of theuncoupling protein 1 from the aP2 gene promoter stimulatesmitochondrial biogenesis in unilocular adipocytes in vivo.Eur J Biochem. 2002;269:1–10.
22. Kopecky J, Hodny Z, Rossmeisl M, Syrovy I, Kozak LP.Reduction of dietary obesity in the aP2-Ucp transgenic mice:physiology and adipose tissue distribution. Am J Physiol.1996;270:E768–75.
23. Stefl B, Janovska A, Hodny Z, et al. Brown fat is essentialfor cold-induced thermogenesis but not for obesity resistancein aP2-Ucp mice. Am J Physiol. 1998;274:E527–33.
24. Rossmeisl M, Syrovy I, Baumruk F, Flachs P, Janovska P,Kopecky J. Decreased fatty acid synthesis due to mitochon-drial uncoupling in adipose tissue. FASEB J. 2000;14:1793–800.
25. Kotite L, Bergeron N, Havel RJ. Quantification of apoli-poproteins B-100, B-48, and E in human triglyceride-richlipoproteins. J Lipid Res. 1995;36:890–900.
26. Li X, Catalina F, Grundy SM, Patel S. Method to measureapolipoprotein B-48 and B-100 secretion rates in an individualmouse: evidence for a very rapid turnover of VLDL andpreferential removal of B-48- relative to B-100-containinglipoproteins. J Lipid Res. 1996;37:210–9.
27. Otway S, Robinson AM. The use of a non-ionic detergent(triton WR1339) to determine rates of triglyceride entry intothe circulation of the rat under different physiological condi-tions. J Physiol. 1967;190:321–32.
28. Iverius P-H, Ostlund-Lindqvist A-M. Preparation, charac-terization, and measurement of lipoprotein lipase. In: AlbersJJ, Segrest JP, eds. Methods in Enzymology, Plasma Lipopro-teins, Part B. New York: Academic Press; 1986, pp. 691–704.
29. Matejkova O, Mustard KJ, Sponarova J, et al. Possibleinvolvement of AMP-activated protein kinase in obesity re-sistance induced by respiratory uncoupling in white fat. FEBSLett. 2004;569:245–8.
30. Wang T, Zang Y, Ling W, Corkey BE, Guo W. Metabolicpartitioning of endogenous fatty acid in adipocytes. Obes Res.2003;11:880–7.
31. Flachs P, Novotny J, Baumruk F, et al. Impaired nor-adrenaline-induced lipolysis in white fat of aP2-Ucp1 trans-genic mice is associated with changes in G-protein levels.Biochem J. 2002;364:369–76.
32. Nishina PM, Verstuyft J, Paigen B. Synthetic low and highfat diets for study of atherosclerosis in the mouse. J Lipid Res.1990;31:859–69.
33. Lewis GF. Fatty acid regulation of very low density lipopro-tein production. Curr Opin Lipidol. 1997;8:146–53.
34. Franckhauser S, Munoz S, Pujol A, et al. Increased fattyacid re-esterification by PEPCK overexpression in adiposetissue leads to obesity without insulin resistance. Diabetes.2002;51:624–30.
35. Olswang Y, Cohen H, Papo O, et al. A mutation in theperoxisome proliferator-activated receptor gamma-bindingsite in the gene for the cytosolic form of phosphoenolpyruvatecarboxykinase reduces adipose tissue size and fat content inmice. Proc Natl Acad Sci U S A. 2002;99:625–30.
36. Tordjman J, Chauvet G, Quette J, Beale EG, Forest C,Antoine B. Thiazolidinediones block fatty acid release byinducing glyceroneogenesis in fat cells. J Biol Chem. 2003;278:18785–90.
37. Frayn KN, Summers LKM. Substrate fluxes in skeletalmuscle and white adipose tissue and their importance in thedevelopment of obesity. In: Kopelman PG, Stock MJ, eds.Clinical Obesity. Oxford: Blackwell Science; 1998, pp. 129–157.
38. Fielding PE, Fielding CJ. Dynamics of lipoprotein transportin the human circulatory system. In: Vance DE, Vance J, eds.Biochemistry of Lipids, Lipoproteins and Membranes. Am-sterdam: Elsevier; 1996, pp. 495–516.
39. Haemmerle G, Zimmermann R, Strauss JG, et al.Hormone-sensitive lipase deficiency in mice changes theplasma lipid profile by affecting the tissue-specific expressionpattern of lipoprotein lipase in adipose tissue and muscle.J Biol Chem. 2002;277:12946–52.
40. Levak-Frank S, Hofmann W, Weinstock PH, et al. Inducedmutant mouse lines that express lipoprotein lipase in cardiacmuscle, but not in skeletal muscle and adipose tissue, havenormal plasma triglyceride and high-density lipoprotein-cholesterol levels. Proc Natl Acad Sci U S A. 1999;96:3165–70.
41. Winder WW, Hardie DG. AMP-activated protein kinase, ametabolic master switch: possible roles in type 2 diabetes.Am J Physiol. 1999;277:E1–10.
42. Saha AK, Avilucea PR, Ye JM, Assifi MM, Kraegen EW,Ruderman NB. Pioglitazone treatment activates AMP-activated protein kinase in rat liver and adipose tissue in vivo.Biochem Biophys Res Commun. 2004;314:580–5.
43. Guan HP, Li Y, Jensen MV, Newgard CB, Steppan CM,Lazar MA. A futile metabolic cycle activated in adipocytesby antidiabetic agents. Nat Med. 2002;8:1122–8.
44. Ruderman NB, Park H, Kaushik VK, et al. AMPK as ametabolic switch in rat muscle, liver and adipose tissue afterexercise. Acta Physiol Scand. 2003;178:435–42.
45. Zimmermann R, Haemmerle G, Wagner EM, Strauss JG,Kratky D, Zechner R. Decreased fatty acid esterificationcompensates for the reduced lipolytic activity in hormone-sensitive lipase-deficient white adipose tissue. J Lipid Res.2003;44:2089–99.
46. Shimada M, Shimano H, Gotoda T, et al. Overexpression ofhuman lipoprotein lipase in transgenic mice. Resistence todiet-induced hypertriglyceridemia and hypercholesterolemia.J Biol Chem. 1993;268:17924–9.
47. Carneheim C, Nedergaard J, Cannon B. Beta-adrenergicstimulation of lipoprotein lipase in rat brown adipose tissue
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
OBESITY RESEARCH Vol. 13 No. 5 May 2005 843

during acclimation to cold. Am J Physiol. 1984;246:E327–33.48. Zhou Y-T, Shimabukuro M, Koyama K, et al. Induction by
leptin of uncoupling protein-2 and enzymes of fatty acidoxidation. Proc Natl Acad Sci U S A. 1997;94:6386–90.
49. Zhou Y-T, Wang Z-W, Higa M, Newgard CB, Unger RH.Reversing adipocyte differentiation: Implications for treat-ment of obesity. Proc Natl Acad Sci U S A. 1999;96:2391–5.
50. Chen G, Koyama K, Yuan X, et al. Disappearance of bodyfat in normal rats induced by adenovirus-mediated leptin genetherapy. Proc Natl Acad Sci U S A. 1996;93:14795–9.
51. Wang YX, Lee CH, Tiep S, et al. Peroxisome-proliferator-activated receptor delta activates fat metabolism to preventobesity. Cell. 2003;113:159–70.
52. Cederberg A, Gronning LM, Ahren B, Tasken K, CarlssonP, Enerback S. FOXC2 is a winged helix gene that counter-acts obesity, hypertriglyceridemia, and diet-induced insulinresistance. Cell. 2001;106:563–73.
53. Tiraby C, Tavernier G, Lefort C, et al. Acquirement ofbrown fat cell features by human white adipocytes. J BiolChem. 2003;278:33370–6.
Respiratory Uncoupling and Plasma Triglycerides, Rossmeisl et al.
844 OBESITY RESEARCH Vol. 13 No. 5 May 2005
Copyright © 2022 FDOKUMEN




















