
Treeholes as Calling, Breeding, and Developmental Sites for the Amazonian Canopy Frog, Phrynohyas...
11
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Treeholes as Calling, Breeding, and Developmental Sites for the Amazonian Canopy Frog, Phrynohyas resinifictrix (Hylidae) Author(s): Luis Schiesari, Marcelo Gordo, Walter Hödl Source: Copeia, 2003(2):263-272. 2003. Published By: The American Society of Ichthyologists and Herpetologists DOI: http://dx.doi.org/10.1643/0045-8511(2003)003[0263:TACBAD]2.0.CO;2 URL: http://www.bioone.org/doi/full/10.1643/0045-8511%282003%29003%5B0263%3ATACBAD %5D2.0.CO%3B2 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Treeholes as Calling, Breeding, and Developmental Sites for the Amazonian Canopy Frog, Phrynohyas...

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors nonprofit publishers academic institutions researchlibraries and research funders in the common goal of maximizing access to critical research
Treeholes as Calling Breeding and Developmental Sites for the AmazonianCanopy Frog Phrynohyas resinifictrix (Hylidae)Author(s) Luis Schiesari Marcelo Gordo Walter HoumldlSource Copeia 2003(2)263-272 2003Published By The American Society of Ichthyologists and HerpetologistsDOI httpdxdoiorg1016430045-8511(2003)003[0263TACBAD]20CO2URL httpwwwbiooneorgdoifull1016430045-8511282003290035B02633ATACBAD5D20CO3B2
BioOne (wwwbiooneorg) is a nonprofit online aggregation of core research in the biological ecological andenvironmental sciences BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies associations museums institutions and presses
Your use of this PDF the BioOne Web site and all posted and associated content indicates your acceptance ofBioOnersquos Terms of Use available at wwwbiooneorgpageterms_of_use
Usage of BioOne content is strictly limited to personal educational and non-commercial use Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder
q 2003 by the American Society of Ichthyologists and Herpetologists
Copeia 2003(2) pp 263ndash272
Treeholes as Calling Breeding and Developmental Sites for theAmazonian Canopy Frog Phrynohyas resinifictrix (Hylidae)
LUIS SCHIESARI MARCELO GORDO AND WALTER HODL
In this paper we present information on the natural history reproductive anddevelopmental strategies of the canopy treefrog Phrynohyas resinifictrix Phrynohyasresinifictrix is a widespread Amazonian species breeding exclusively in water-filledtreeholes in terra-firme rain forests Females laid relatively large floating egg massesin the treeholes Gut content analyses indicated that tadpoles are generalists andpredominantly macrophageous basing their diet on detritus and conspecific fertil-ized eggs of younger cohorts Tadpole guts were found to contain up to 62 intacteggs This cannibalistic interaction is presumably of major importance for the com-pletion of the larval stage and may result in depletion of entire egg complementsStrong site fidelity interference competition for treeholes by territorial males con-tinuous use of treeholes year after year repeated oviposition in treeholes and tem-poral constancy in density of calling males are consistent with the hypotheses thatadequate treeholes are a limiting resource for P resinifictrix populations or thatlocation of unoccupied treeholes in the rain-forest canopy involves substantial riskor energetic costs
Neste artigo apresentamos informacoes sobre a historia natural e sobre as estra-tegias reprodutivas e de desenvolvimento da perereca de dossel Phrynohyas resini-fictrix Phrynohyas resinifictrix e uma especie amplamente distribuıda em florestaspluviais tropicais de terra-firme da Amazonia que se reproduz exclusivamente emocos-de-arvore preenchidos com agua Nestes ocos as femeas depositaram desovasflutuantes relativamente grandes A analise de conteudos intestinais revelou que osgirinos possuem habitos alimentares generalistas e predominantemente macrofagi-cos sendo a dieta baseada em detrito e ovos fertilizados coespecıficos de coortesposteriores Encontramos nos intestinos um maximo de 62 ovos intactos Esta in-teracao canibalıstica e presumivelmente de grande importancia para a completacaodo desenvolvimento larval mas pode resultar na eliminacao de desovas inteiras Apresenca de forte fidelidade ao sıtio reprodutivo competicao de interferencia entremachos territoriais utilizacao contınua de ocos-de-arvore ano apos ano oviposturarepetida em ocos-de-arvore e a constancia temporal na densidade de machos vo-calizantes sao observacoes consistentes com a hipotese de que ocos-de-arvore ade-quados constituem recurso limitante para populacoes de P resinifictrix ou que alocalizacao de ocos-de-arvore desocupados no dossel da floresta envolve risco oucustos energeticos substanciais
PHYTOTELMS are plants or parts of plantsthat hold rainwater such as palm leaf
fronds and axils bracts inflorescences fruitsbromeliads and treeholes These water bodiesmay increase the habitat available for aquaticorganisms up to 50000 liters per hectare insome moist tropical locations (Fish 1983) How-ever animal development in phytotelms is con-strained by several physico-chemical as well asbiological properties Among these are thesmall water volumes low turnover rates of theaccumulated water and predominance of de-caying processes resulting in low levels of dis-solved oxygen and low availability and predict-ability of food Nevertheless the use of phyto-telms as breeding andor developmental sites is
found in four of the more than 30 reproductivemodes exhibited by anuran amphibians (Duell-man and Trueb 1986 Haddad and Hodl 1997Haddad and Sawaya 2000) and at least five tad-pole ecomorphotypes are associated with life inthese microhabitats (Lannoo et al 1987)
Herein we present data on the biology of theAmazonian hylid treefrog Phrynohyas resinifictrix(Goeldi 1907) known locally as canuaru cu-nuaru or canoeiro (Lutz 1973) Phrynohyas re-sinifictrix is known from undisturbed Amazonianterra-firme and occasionally flooded rain forestsin scattered localities in Peru Bolivia EcuadorColombia Venezuela Brazil Suriname andFrench Guiana (Frost 1985 pers obs) Malescall from widely spaced treeholes in primary for-
264 COPEIA 2003 NO 2
est Long-range communication is effected witha loud low-pitched call which is carried forlong distances through the use of the rain forestlow frequency window (Zimmerman and Hodl1983 Hodl 1991) When present the hollowportion of the treehole may act as a resonatingchamber increasing the broadcasting distancefor the calls (Hodl 1991)
Phrynohyas resinifictrix tadpoles collected fromtreeholes have been the subject of detailed eco-morphological studies (Grillitsch 1992 Schie-sari et al 1996) which predicted an ability toingest large food particles and a reliance on pul-monary as well as branchial respiration Withthe exception of the studies mentioned abovevery little information is available about theecology of this species
We show here that P resinifictrix is capable ofcompleting its entire life cycle in the forest can-opy by breeding exclusively in large water-filledtreeholes We present data on the physical andbiological aspects of the treeholes used as call-ing and breeding sites by the species as well asestimates of calling male population densitiesQuantitative dietary data are examined to clar-ify the strategies allowing completion of meta-morphosis in these unproductive microhabitatsWe discuss the importance of predation of eggsand larvae and low interspecific competition asmajor forces maintaining oviposition in phyto-telms Finally we discuss the hypothesis thattreeholes constitute a limiting resource for Presinifictrix populations and suggest that malesmonopolizing treeholes may obtain multiplemates
MATERIALS AND METHODS
Fieldwork was conducted in two Amazonianterra-firme rain-forest sites Panguana Peru(lower Rio Llullapichis 98379S 748569W) byWH from October to November 1988 and theReserva Florestal Adolfo Ducke (hereforth re-ferred to as lsquolsquoManausrsquorsquo) located about 25 kmnorth of Manaus Amazonas Brazil (038089S608029W) by LS and MG from November 1990to March 1991 and from January to April 1993Sporadic additional observations were conduct-ed from January to June 1994 and from Augustto October 1994 Annual rainfall is 2635 mm inPanguana (Aichinger 1987) and 2075 mm inManaus (mean values from 1931 to 1960 Brasil1978) The rainy season occurs between Sep-tember and May (Panguana) or December andMay (Manaus) with monthly rainfall of morethan 200 mm whereas the driest months Julyand August (plus September in Manaus) re-ceive less than 60 mm each It is noteworthy that
during the study period mean annual rainfall inManaus was 2660 mm 30 higher than the av-erage (data collected at an EMBRAPA meteo-rological station 5 km north of the study site)
We searched for calling males while walkingalong a single trail 1470 m long in Panguana(totaling at least 94 km walked in 32 nights) andalong a system of trails 8854 m long in Manaus(totaling at least 151 km walked in 36 nights in19901991 and 145 km walked in 57 nights in1993) We located the trees from which malefrogs were calling by approaching the source ofthe loud call during the night and subsequentlypressing an ear tightly against tree trunks Thetrunk hosting the calling male was the one thatbest conducted sound vibrations Wheneverpossible treeholes were reached either bymeans of ladders lsquolsquogriffesrsquorsquo (metal hooks withspikes used for tree climbing see Mori 1987for description and illustration) or mountain-eering equipment We located 10 trees in Pan-guana and 19 in Manaus and were able to in-spect three and seven treeholes respectively
We estimated the density of calling males bydividing the number of regularly calling males(the abundance) by the length of trails walked(individualskm linear density) and by the areaacoustically sampled (individualsha density byarea) To estimate the area acoustically sampledwe multiplied the length of trails walked bytwice the maximum distance inside the forest inwhich calls were above ambient noise levelsThese were determined with a CEL-493 Preci-sion Impulse Integrating Sound Level Meterand a CEL-296 Third Octave Autoscan FilterSet Although in some cases calls were withinhuman hearing range from distances up to 250m sound pressure levels were consistently abovethe ambient noise level at a maximal distanceof 160 m at the 800 Hz terz filter band At thisfrequency and distance sound levels were be-tween 24 dB and 32 dB (linear peak relative to20 mNm2) Therefore we employed the con-servative value of 160 m for our estimates ofarea acoustically sampled
We also measured diameter at breast height(DBH) of the host tree dimensions of the tree-hole opening water surface and depth of thetreehole and its height Treehole water vol-umes were either directly measured by suckingout water through a hose and pouring it into ameasuring cup or estimated by employing thecylinder volume formula to the water surfaceand treehole depth measured A cylindricalmodel appears to be a good estimator of tree-hole shape no significant differences werefound between the actual and estimated vol-umes of three treeholes for which both sets of
265SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
data are available (Student t-test t 5 068 P 5057)
Treeholes were inspected for 20ndash30 min onone to 30 occasions depending on accessibilityTreehole limnological parameters were mea-sured on 19 March 1993 and 31 May 1994 (Ma-naus) Values of pH dissolved oxygen concen-tration (DO) and algal chlorophyll concentra-tion (as a measure of phytoplankton standingcrop) of the surface water present in four tree-holes two bromeliads a small temporary pondand a dammed creek were analyzed We deter-mined pH with a Corning pH meter 130 andDO with an YSI Dissolved Oxygen meter eitherin situ or on the ground after 500 ml of waterwere collected with minimum disturbanceChlorophyll a was extracted from 250 mL of wa-ter and analyzed in spectrophotometer for ex-tinctions at 7500 A (Strickland and Parsons1968) Classification of water body trophic type(productivity) followed Wetzel (1983)
Eggs and tadpoles were collected with smallcircular nets or sucked with a hose and pre-served in 5 formalin Identity of larvae wasconfirmed by rearing until metamorphosis orby the aid of literature sources (Hero 1990 Gri-llitsch 1992 Schiesari et al 1996) Voucherspecimens were deposited in the Museum of Zo-ology of the University of Sao Paulo (MZUSP128321) Larvae were staged according to Gos-ner (1960) To quantify the dietary habits oftadpoles from Manaus we analyzed the gut con-tents of 32 tadpoles of several developmentalstages ranging from Gosner (1960) stage 25 to42 from a single treehole These tadpoles werecollected in four samples taken at roughlymonthly intervals for a period of five months inthe rainy season For a comparison among tree-holes we analyzed gut contents of three to fivetadpoles in the same developmental stage (stage25) collected at one time from four treeholesWe used the following procedure After remov-ing the entire intestinal mass a fragment of 1cm of the foregut was taken and its contentssuspended in 01 ml of distilled water on a mi-croscope slide and examined at a magnificationof 250 to 4003 We quantified the contents byrandomly selecting 10 microscopic fields perslide and noting the presence or absence ineach field of the following five food categoriesdetritus (nonidentifiable particulate organicmatter) plant fragments algae arthropod frag-ments and anuran eggs Dietary compositionwas expressed as mean 6 1 SE of the percent-ages of occurrence of each food item in the in-dividuals analyzed Because more than one foodcategory may be present per microscopic fieldthe sum of the frequencies of occurrence of all
categories may exceed 100 When guts werefilled with eggs the abundance of this item wasassumed to be 100 We then counted intacteggs for the length of the digestive tract Par-tially or totally digested eggs could be identifiedby the texture of residue and confirmed by theobservation of yolk platelets under microscopicexamination To determine whether ingestedeggs were fertilized embryonic developmentwas ascertained by external morphology (Gos-ner 1960) and by histological analysis of somehematoxylin-eosin stained eggs of at least onetadpole gut per sample The latter procedureprovided evidence of early embryonic develop-ment which might not be easily detected by ex-ternal morphology
RESULTS
TreeholesmdashTreeholes used by calling males of Presinifictrix occurred from 22ndash32 m high in treeswith DBH ranging from 13ndash92 cm (mean 6 SE449 6 62 cm n 5 15) No significant differ-ences in DBH were found between trees usedin Panguana and Manaus (Student t-test t 5022 P 5 083)
Treehole openings were circular to ellipticalwith areas ranging from 628ndash4514 cm2 (18046 396 n 5 10 Table 1) Treeholes invariablyfaced upward thus allowing the replenishmentof evaporating water via rainfall and the accu-mulation of allochthonous organic matter Bot-toms of treeholes were usually covered by oneor more layers of dead leaves and decaying or-ganic matter There was no sign of resin liningon the treehole as described by Goeldi (1907)Treehole depth ranged from 15ndash190 cm (8666 277 cm n 5 9 Table 1) Treehole water vol-umes varied from 06ndash900 liters (182 6 93 li-ters n 5 9) but tadpoles were found only intreeholes larger than 28 liters (Table 1) Con-sidering only treeholes sampled during therainy season maximum tadpole abundanceshowed no association to treehole height (P 5017) but was positively associated with treeholewater volume (Y 5 3868 1 362 3 Volume r2
5 077 F 5 1983 P 5 00043) as indicated bylinear regressions No associations were foundbetween maximum tadpole density and tree-hole height (P 5 077) or volume (P 5 078)
Water was less acidic in treeholes (pH 5 6816 029 range 590ndash690 n 5 4) than in pond(564) creek (409) or bromeliads (378 6 02range 357ndash398 n 5 2) Treeholes nearly brack-eted the variation in DO found in other waterbodies DO in treeholes varied from 02ndash39mgl (23 6 11 mgl) when compared to thepond (09 mgl) bromeliads (27 6 06 mgl
266 COPEIA 2003 NO 2
TABLE 1 CHARACTERISTICS OF TREEHOLES USED BY Phrynohyas resinifictrix AND MAXIMUM ABUNDANCE AND DEN-SITY OF TADPOLES OBSERVED PER TREEHOLE
Tree
NumberDBH(cm)
Treehole characteristics
Height(m)
Opening(cm)
Depth(cm)
Volume(L)
Tadpoles
Maxabundance(No inds)
Maxdensity
(IndsL)
Manaus123c
456
341549mdashmdash32
115
32128
145
13 3 1311 3 1123 3 2515 3 1520 3 1321 3 21
19059351932
260
253a
58a
158b
66b
81b
900b
237930
33128337
9371612
0502
1572374
7 27 13 13 3 13 75 89a 10 113
Panguana123
572733
5210422
14 3 0720 3 0810 3 08
70mdash15
28a
mdash06a
83d
0
286mdash
0
Treehole volumes were either a directly measured or b estimatedc Treehole examined at the end of the dry seasond Tadpoles of Dendrobates quinquevittatus species group
range 21ndash33) and creek (41 mgl) Phyto-plankton chlorophyll a concentration was low-est in the creek (031 ugl ultraoligotrophic)followed by treeholes (088 6 032 ugl range015ndash165 ultraoligotrophic to oligotrophic)and pond (449 ugl mesotrophic) See Mag-nusson and Hero (1991) for limnological pa-rameters of additional terrestrial water bodies atthe Manaus study site
Treeholes used by males of P resinifictrix wereformed in the main trunk after the decayingand breaking of larger branches some treesseemed to be in poor condition and at least twowere decaying In Manaus seven of the treeswere identified in the field as Eschweilera cf odo-ra Lecythidaceae (n 5 3 individuals) Aspido-sperma abscurinervea Apocynaceae Trichilia spMeliaceae unidentified Cariocaraceae and un-identified Mimosaceae In Panguana one of thetrees was an unidentified Moraceae
Adult behaviormdashMales called exclusively fromtreeholes On no occasion was this speciesfound in any of the intensively sampled terres-trial water bodies or in other smaller phyto-telms commonly found in the study areas suchas buriti palm leaf axils (Mauritia flexuosa) orground bromeliads (Guzmania spp Streptocalyxspp) Males often called from sundown (1830h) until sunup (0455 h) when they often leftthe treehole but remained in the canopy Call-ing season lasted from August to April althougha few called regularly during the dry seasonMales appeared to show strong site fidelity Callswere heard from the same trees night after
night In Panguana one male continuously re-turned to the same treehole for a period of 20nights In Manaus we toe-clipped and recap-tured a male in treehole 2 four months later(1990) In the following field season (1993) wetoe-clipped and recaptured three males aggres-sively interacting inside the same treehole onfour occasions over two months Aggressive in-teractions between males were common and in-cluded a particular calling repertoire (Zimmer-man and Hodl 1983) and physical attempts toimpede the opponent to enter or remain in thetreehole
Treeholes were often occupied in multipleyears For example five of the six surviving treesused in 1990 were still occupied in 1993 andfive of nine trees used in 1993 were still occu-pied in 1994
Abundance of calling malesmdashCalling males of Presinifictrix were widely spaced in the forest Wefound 48 calling maleskm (015ha) in Pan-guana and from 17ndash20 calling maleskm(005ndash006ha) in Manaus (Table 2) Abun-dances were 24ndash28 times higher in Panguanathan in Manaus Densities were remarkably con-stant in Manaus during the study period
Premetamorphic biology EggsmdashEgg clutches arelaid in water and consist of a gelatinous eggmass that primarily floats on or near the surfacebut often adheres to the inner wall of the tree-hole The darkly pigmented eggs are small(diam 158 6 006 mm n 5 6) and surroundedby gelatinous capsules (diam 428 6 021 mm
267SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
TABLE 2 SEASONAL ESTIMATES OF THE ABUNDANCE
AND DENSITY (LINEAR AND BY AREA) OF CALLING MALES
OF Phrynohyas resinifictrix IN PANGUANA AND MANAUS
Season
Abun-dance
ofcallingmales
Density of calling males
Linear(males
km)Area
(malesha)Area21
(hamale)
Panguana1988 7 48 015 68
Manaus1990199119931994
1095
191720
006005006
171188163
Fig 1 Number of intact eggs found in the guts oftadpoles of Phrynohyas resinifictrix through ontogeny(n 5 32 treehole 2) The dashed line represents anarbitrarily defined developmental threshold at stage32 after which eggs are efficiently used as food re-source Bubble size is proportional to sample size(varying from 1ndash6)
TABLE 3 DIET OF Phrynohyas resinifictrix LARVAE Occurrence expressed as Mean 6 SE
Stages n
Occurrence of food categories ()
AlgaePlant
fragmentsArthropodfragments Detritus Eggs
One treehole sampled over five months25ndash41 32 166 6 51 100 6 28 53 6 16 359 6 84 597 6 87
Four treeholes sampled at one time25252525
5443
60 6 25150 6 65300 6 41300 6 100
180 6 58275 6 103200 6 4133 6 33
00 6 0075 6 4875 6 7500 6 00
580 6 132975 6 25750 6 50730 6 22
600 6 18200 6 0000 6 00
300 6 100
Total44 175 6 39 114 6 23 46 6 13 446 6 67 523 6 72
n 5 6) At the moment of collection clutch sizevaried from 106ndash1540 eggs (436 6 234 eggs n5 6) with as many as six clutches laid in a singletreehole during a period of four months (De-cember 1990 to April 1991)
Larval dietmdashThe diet of P resinifictrix larvaeconsists largely of conspecific eggs and detritusless common items include algae plant frag-ments (mainly leaf fragments but also pollen)and arthropod fragments (insect mouthpartsheads and wing fragments lepidopteran scalesand mosquito larvae bristles Table 3)
Fifty-two percent of all guts examined (n 544) contained eggs in different phases of diges-tion (Table 3) Tadpoles from stages 32ndash42 wereable to swallow whole eggs with capsules andsometimes gelatin (Fig 1) whereas smaller tad-poles ingested egg fragments presumablyrasped with their keratinized mouthparts Wheneggs were available individuals ingested asmany as possible and their guts became greatlyextended The enlarged foregut contained upto 62 whole eggs (Fig 1) plus partially digested
eggs that could not be counted Most eggs ex-amined were fertilized as indicated by externalor histological observation of cell cleavage andembryo development (up to stage 16) Four ofsix clutches laid during a five-month period intreehole 2 at Manaus contained fertilized eggsthe two others could not be analyzed becauseno eggs remained in the gelatinous mass by thetime of observation
Detritus was the second most frequent fooditem consumed observed in 446 of micro-scopic fields when eggs were not available de-
268 COPEIA 2003 NO 2
tritus occurrence reached 975 (Table 3) Allother food items were found much less fre-quently Algae were never abundant althoughsporadically occurring in 175 of all micro-scopic fields or up to 30 in tadpoles in earlydevelopmental stages Plant fragments occurredin 114 of all microscopic fields or up to275 in tadpoles in early developmental stag-es Arthropod fragments were only present in46 of all microscopic fields examined (Table3)
DISCUSSION
Nonaquatic oviposition behavior in frogsevolved independently in many lineages andhas been interpreted as a strategy to avoid pre-dation competition or unfavorable abiotic con-ditions in the ancestral water body (Crump1982 Duellman and Trueb 1986) From anecological rather than evolutionary perspectivethese same processes might be currently oper-ating to maintain arboreal oviposition Magnus-son and Hero (1991) studied an anuran assem-blage at our Central Amazonian rain-forest fieldsite They found that the percentage of specieswith terrestrial eggs and aquatic larvae in a wa-ter body was positively correlated with an indexof egg predation pressure in that water body butonly weakly correlated with the number of spe-cies of potential competitors or abiotic condi-tions such as dissolved oxygen content per-centage of dry days and temperature These ob-servations led the authors to suggest that pre-dation is a major factor maintaining nonaquaticoviposition behavior in this contemporary an-uran assemblage
Oviposition in phytotelms might also bemaintained by the high predation pressure interrestrial water bodies Predation risk is pre-sumably lower in phytotelms because theheight small size concealment and often widespacing of these water bodies reduce predatoraccess to the eggs and tadpoles In addition un-favorable abiotic conditions in phytotelms suchas the potentially low dissolved oxygen levelsand short hydroperiod constrain the survival ofaquatic predators In this study few actual orpotential predators of eggs (wasps small dytis-cid beetles and heterospecific oophagic andcannibalistic tadpoles) tadpoles (spiders) andadults (a colubrid snake Philodryas viridissimus)have been observed inside treeholes inhabitedby P resinifictrix This contrasts strongly with theubiquity of insect and fish predators found inground water bodies at the study site (Magnus-son and Hero 1991)
In addition our observations suggest that
current interspecific competition among Ama-zonian frogs for phytotelms as breeding and de-velopmental sites is minimal and could be an-other important force maintaining ovipositionin these water bodies Only four of 130 lowlandAmazonian frog species breed in phytotelms(Hodl 1990) Moreover in Central Amazonia Presinifictrix is the only anuran to breed exclusive-ly in large water-filled treeholes whereas Osteo-cephalus oophagus breeds in a variety of phyto-telms such as bromeliads buriti (Mauritia fle-xuosa) palm leaf axils palm bracts and only oc-casionally in treeholes ( Jungfer and Schiesari1995 pers obs) Calling adults eggs or larvaeof P resinifictrix and O oophagus were neverfound in the extensively sampled ground waterbodies indicating specialization for a phytotel-mic breeding site Yet tadpoles of these specieswere never found coexisting in any of the phyto-telms sampled indicating strong habitat segre-gation Several species of Dendrobates have ter-restrial oviposition but transport tadpoles tophytotelms after hatching for instance we ob-served in a single occasion Dendrobates cf quin-quevittatus inside treeholes from which P resini-fictrix called More often however Dendrobatesspecies use small-sized ground phytotelms (egCaldwell and Araujo 1998) Overall this strongpattern of habitat segregation among phytotelmtypes in which more than one species are sel-dom found to coexist contrasts with ground wa-ter bodies where typically more than three andsometimes up to eight tadpole species coexist(Magnusson and Hero 1991)
Therefore several lines of evidence suggestthat both predation pressure and interspecificcompetition are lower in phytotelms than in ter-restrial water bodies and could contribute to themaintenance of the use of phytotelms as breed-ing and developmental sites for Amazonian an-urans in general and P resinifictrix in particularIn other systems however an unavailability ofterrestrial water bodies may act as a strong forcemaintaining phytotelm reproduction This ap-pears to be the case in Caribbean Islands suchas Jamaica where historical and geological fac-tors result in phytotelms being the predominantlentic water bodies (Thompson 1996)
Phytotelms are a harsh environment for thedevelopment of anuran embryos and larvaeConstrained volumes of noncirculating waterdominated by decaying processes sometimes re-sult in very low dissolved oxygen concentra-tions Assuming that treehole breeding is de-rived in the genus Phrynohyas early develop-ment of large inflated functional lungs in oth-er pond-dwelling species of the genus may beconsidered a useful morphophysiological trait
269SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
for the colonization of phytotelms by P resinific-trix Lungs are also used as hydrostatic organskeeping tadpoles close to the water surface andmay reduce the energy expenditure involved inrepeatedly swimming to the surface to bob air(Feder 1984 Feder and Moran 1985 Lannooet al 1987)
An important biotic characteristic of phyto-telms is the unpredictability and low availabilityof high quality food In most tadpole species forwhich dietary data are available algae of pe-riphyton and phytoplankton constitute majorfood resources in addition to low-quality detri-tus (eg Altig and Johnston 1989 Kupferberget al 1994) Narrow treehole openings oftencovered by overlying canopy may result in anearly aphotic habitat in which primary pro-ductivity is low and therefore likely insufficientto sustain tadpole growth
Detritus derived from the decomposition offallen leaf litter and treehole walls constitutedthe basis for the speciesrsquo diet when eggs werenot available The nutritive value of detritus islow (Kupferberg et al 1994) and several spe-cies of generalized pond tadpoles fail to reachmetamorphosis when restricted to this food type(Werner and Glennemeier 1999 LS unpubl)In this context oophagy may be of major im-portance for the success of individual larvae inreaching metamorphosis because eggs providea highly nutritious and energetic food source(Crump 1992) However detritivory could alsobe of crucial importance especially for smallertadpoles Prior to stage 32 tadpoles are gape-limited and therefore inefficient egg feedersAfter this stage tadpoles are capable of swallow-ing large quantities of whole eggs leading toaccelerated growth and development
Opportunistic oophagy is common in anuranlarvae (Magnusson and Hero 1991 Crump1992 Petranka and Kennedy 1999) In severalphytotelm-developing anurans however oop-hagy results from complex parental behavior inwhich the mother periodically returns to thephytotelm to lay fertilized or unfertilized eggsfor nutrition of its progeny (Lannoo et al 1987Weygoldt 1987 Crump 1995) For some ofthese species it was demonstrated that oophagyis obligate that is tadpoles fail to reach meta-morphosis if eggs are not provided by the moth-er (Weygoldt 1987 Thompson 1996 Jungferand Weygoldt 1999) In P resinifictrix howeverthere are several observations suggesting thatoophagy is opportunistic cannibalism ratherthan parental care in the form of periodic eggdeposition for obligate tadpole nutrition Firstas noted by Jungfer and Proy (1998) P resinific-trix egg clutches are large (maximum clutch size
in the field 1540 eggs this study in terraria4336 eggs Jungfer and Proy 1998) comparedto phytotelm-breeding species that intentionallyprovide eggs to their offspring For instanceconsidering only hylids on average 158 eggs arelaid by Anotheca spinosa ( Jungfer 1996) 258 byOsteocephalus oophagus ( Jungfer and Weygoldt1999) 276 per bromeliad by Osteopilus brunneus(Lannoo et al 1987) and 13 by Phyllodytes lu-teolus (Giaretta 1996) Second as a conse-quence of the large clutch size interclutch in-terval is long in P resinifictrix in a regularly sam-pled treehole six clutches were deposited overa period of four months (around one clutch ev-ery 20 days) which coincides with the inter-clutch interval of pairs in captivity (20 daysrange 7ndash37 n 5 29 Jungfer and Proy 1998)This long interclutch interval contrasts stronglywith the interclutch interval of species that areknown to be obligate oophagic For instanceeggs are deposited every 49 days in A spinosa( Jungfer 1996) 51 days in O oophagus ( Jung-fer and Weygoldt 1999) and 44 days in Obrunneus (Thompson 1996) Jungfer and Proy(1998) found that tadpoles of P resinifictrixstarve if dependent only on the motherrsquos infre-quent oviposition as the food source but thatmetamorphic success in an experimental con-tainer was high if in addition to leaf-litter eggswere added every 17 days Third an opportu-nistic oophagic nutrition is suggested by our ob-servation that eggs are fertilized unlike some ofthe more lsquolsquoadvancedrsquorsquo forms of anuran parentalcare through laying of unfertilized trophic eggs(Weygoldt 1987) although obligate oophagyhas been documented for species laying fertil-ized eggs (Thompson 1996) Fourth althoughextensive reduction of oral structures character-izes several of the obligate oophagic phytotelm-developing species (Lannoo et al 1987) P re-sinifictrix retains typical generalized pond-typelarval morphology with only moderate structur-al reduction when compared to its pond breed-ing congeners (Schiesari et al 1996) Experi-mental evidence is necessary to unambiguouslydemonstrate that oophagy is not obligate in Presinifictrix however the arguments above pro-vide indirect support for the hypothesis thatconsumption of eggs is important but may notbe obligate for metamorphic success
We suggest that among the premetamorphicstages of P resinifictrix life cycle cannibalism andintraspecific competition are the major forcesacting on the survival of eggs and larvae re-spectively Evidence for this hypothesis are thelow nutritive value of food other than eggs andor high unpredictability of food in the treeholethe voracious oophagic habits of the tadpole
270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo

q 2003 by the American Society of Ichthyologists and Herpetologists
Copeia 2003(2) pp 263ndash272
Treeholes as Calling Breeding and Developmental Sites for theAmazonian Canopy Frog Phrynohyas resinifictrix (Hylidae)
LUIS SCHIESARI MARCELO GORDO AND WALTER HODL
In this paper we present information on the natural history reproductive anddevelopmental strategies of the canopy treefrog Phrynohyas resinifictrix Phrynohyasresinifictrix is a widespread Amazonian species breeding exclusively in water-filledtreeholes in terra-firme rain forests Females laid relatively large floating egg massesin the treeholes Gut content analyses indicated that tadpoles are generalists andpredominantly macrophageous basing their diet on detritus and conspecific fertil-ized eggs of younger cohorts Tadpole guts were found to contain up to 62 intacteggs This cannibalistic interaction is presumably of major importance for the com-pletion of the larval stage and may result in depletion of entire egg complementsStrong site fidelity interference competition for treeholes by territorial males con-tinuous use of treeholes year after year repeated oviposition in treeholes and tem-poral constancy in density of calling males are consistent with the hypotheses thatadequate treeholes are a limiting resource for P resinifictrix populations or thatlocation of unoccupied treeholes in the rain-forest canopy involves substantial riskor energetic costs
Neste artigo apresentamos informacoes sobre a historia natural e sobre as estra-tegias reprodutivas e de desenvolvimento da perereca de dossel Phrynohyas resini-fictrix Phrynohyas resinifictrix e uma especie amplamente distribuıda em florestaspluviais tropicais de terra-firme da Amazonia que se reproduz exclusivamente emocos-de-arvore preenchidos com agua Nestes ocos as femeas depositaram desovasflutuantes relativamente grandes A analise de conteudos intestinais revelou que osgirinos possuem habitos alimentares generalistas e predominantemente macrofagi-cos sendo a dieta baseada em detrito e ovos fertilizados coespecıficos de coortesposteriores Encontramos nos intestinos um maximo de 62 ovos intactos Esta in-teracao canibalıstica e presumivelmente de grande importancia para a completacaodo desenvolvimento larval mas pode resultar na eliminacao de desovas inteiras Apresenca de forte fidelidade ao sıtio reprodutivo competicao de interferencia entremachos territoriais utilizacao contınua de ocos-de-arvore ano apos ano oviposturarepetida em ocos-de-arvore e a constancia temporal na densidade de machos vo-calizantes sao observacoes consistentes com a hipotese de que ocos-de-arvore ade-quados constituem recurso limitante para populacoes de P resinifictrix ou que alocalizacao de ocos-de-arvore desocupados no dossel da floresta envolve risco oucustos energeticos substanciais
PHYTOTELMS are plants or parts of plantsthat hold rainwater such as palm leaf
fronds and axils bracts inflorescences fruitsbromeliads and treeholes These water bodiesmay increase the habitat available for aquaticorganisms up to 50000 liters per hectare insome moist tropical locations (Fish 1983) How-ever animal development in phytotelms is con-strained by several physico-chemical as well asbiological properties Among these are thesmall water volumes low turnover rates of theaccumulated water and predominance of de-caying processes resulting in low levels of dis-solved oxygen and low availability and predict-ability of food Nevertheless the use of phyto-telms as breeding andor developmental sites is
found in four of the more than 30 reproductivemodes exhibited by anuran amphibians (Duell-man and Trueb 1986 Haddad and Hodl 1997Haddad and Sawaya 2000) and at least five tad-pole ecomorphotypes are associated with life inthese microhabitats (Lannoo et al 1987)
Herein we present data on the biology of theAmazonian hylid treefrog Phrynohyas resinifictrix(Goeldi 1907) known locally as canuaru cu-nuaru or canoeiro (Lutz 1973) Phrynohyas re-sinifictrix is known from undisturbed Amazonianterra-firme and occasionally flooded rain forestsin scattered localities in Peru Bolivia EcuadorColombia Venezuela Brazil Suriname andFrench Guiana (Frost 1985 pers obs) Malescall from widely spaced treeholes in primary for-
264 COPEIA 2003 NO 2
est Long-range communication is effected witha loud low-pitched call which is carried forlong distances through the use of the rain forestlow frequency window (Zimmerman and Hodl1983 Hodl 1991) When present the hollowportion of the treehole may act as a resonatingchamber increasing the broadcasting distancefor the calls (Hodl 1991)
Phrynohyas resinifictrix tadpoles collected fromtreeholes have been the subject of detailed eco-morphological studies (Grillitsch 1992 Schie-sari et al 1996) which predicted an ability toingest large food particles and a reliance on pul-monary as well as branchial respiration Withthe exception of the studies mentioned abovevery little information is available about theecology of this species
We show here that P resinifictrix is capable ofcompleting its entire life cycle in the forest can-opy by breeding exclusively in large water-filledtreeholes We present data on the physical andbiological aspects of the treeholes used as call-ing and breeding sites by the species as well asestimates of calling male population densitiesQuantitative dietary data are examined to clar-ify the strategies allowing completion of meta-morphosis in these unproductive microhabitatsWe discuss the importance of predation of eggsand larvae and low interspecific competition asmajor forces maintaining oviposition in phyto-telms Finally we discuss the hypothesis thattreeholes constitute a limiting resource for Presinifictrix populations and suggest that malesmonopolizing treeholes may obtain multiplemates
MATERIALS AND METHODS
Fieldwork was conducted in two Amazonianterra-firme rain-forest sites Panguana Peru(lower Rio Llullapichis 98379S 748569W) byWH from October to November 1988 and theReserva Florestal Adolfo Ducke (hereforth re-ferred to as lsquolsquoManausrsquorsquo) located about 25 kmnorth of Manaus Amazonas Brazil (038089S608029W) by LS and MG from November 1990to March 1991 and from January to April 1993Sporadic additional observations were conduct-ed from January to June 1994 and from Augustto October 1994 Annual rainfall is 2635 mm inPanguana (Aichinger 1987) and 2075 mm inManaus (mean values from 1931 to 1960 Brasil1978) The rainy season occurs between Sep-tember and May (Panguana) or December andMay (Manaus) with monthly rainfall of morethan 200 mm whereas the driest months Julyand August (plus September in Manaus) re-ceive less than 60 mm each It is noteworthy that
during the study period mean annual rainfall inManaus was 2660 mm 30 higher than the av-erage (data collected at an EMBRAPA meteo-rological station 5 km north of the study site)
We searched for calling males while walkingalong a single trail 1470 m long in Panguana(totaling at least 94 km walked in 32 nights) andalong a system of trails 8854 m long in Manaus(totaling at least 151 km walked in 36 nights in19901991 and 145 km walked in 57 nights in1993) We located the trees from which malefrogs were calling by approaching the source ofthe loud call during the night and subsequentlypressing an ear tightly against tree trunks Thetrunk hosting the calling male was the one thatbest conducted sound vibrations Wheneverpossible treeholes were reached either bymeans of ladders lsquolsquogriffesrsquorsquo (metal hooks withspikes used for tree climbing see Mori 1987for description and illustration) or mountain-eering equipment We located 10 trees in Pan-guana and 19 in Manaus and were able to in-spect three and seven treeholes respectively
We estimated the density of calling males bydividing the number of regularly calling males(the abundance) by the length of trails walked(individualskm linear density) and by the areaacoustically sampled (individualsha density byarea) To estimate the area acoustically sampledwe multiplied the length of trails walked bytwice the maximum distance inside the forest inwhich calls were above ambient noise levelsThese were determined with a CEL-493 Preci-sion Impulse Integrating Sound Level Meterand a CEL-296 Third Octave Autoscan FilterSet Although in some cases calls were withinhuman hearing range from distances up to 250m sound pressure levels were consistently abovethe ambient noise level at a maximal distanceof 160 m at the 800 Hz terz filter band At thisfrequency and distance sound levels were be-tween 24 dB and 32 dB (linear peak relative to20 mNm2) Therefore we employed the con-servative value of 160 m for our estimates ofarea acoustically sampled
We also measured diameter at breast height(DBH) of the host tree dimensions of the tree-hole opening water surface and depth of thetreehole and its height Treehole water vol-umes were either directly measured by suckingout water through a hose and pouring it into ameasuring cup or estimated by employing thecylinder volume formula to the water surfaceand treehole depth measured A cylindricalmodel appears to be a good estimator of tree-hole shape no significant differences werefound between the actual and estimated vol-umes of three treeholes for which both sets of
265SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
data are available (Student t-test t 5 068 P 5057)
Treeholes were inspected for 20ndash30 min onone to 30 occasions depending on accessibilityTreehole limnological parameters were mea-sured on 19 March 1993 and 31 May 1994 (Ma-naus) Values of pH dissolved oxygen concen-tration (DO) and algal chlorophyll concentra-tion (as a measure of phytoplankton standingcrop) of the surface water present in four tree-holes two bromeliads a small temporary pondand a dammed creek were analyzed We deter-mined pH with a Corning pH meter 130 andDO with an YSI Dissolved Oxygen meter eitherin situ or on the ground after 500 ml of waterwere collected with minimum disturbanceChlorophyll a was extracted from 250 mL of wa-ter and analyzed in spectrophotometer for ex-tinctions at 7500 A (Strickland and Parsons1968) Classification of water body trophic type(productivity) followed Wetzel (1983)
Eggs and tadpoles were collected with smallcircular nets or sucked with a hose and pre-served in 5 formalin Identity of larvae wasconfirmed by rearing until metamorphosis orby the aid of literature sources (Hero 1990 Gri-llitsch 1992 Schiesari et al 1996) Voucherspecimens were deposited in the Museum of Zo-ology of the University of Sao Paulo (MZUSP128321) Larvae were staged according to Gos-ner (1960) To quantify the dietary habits oftadpoles from Manaus we analyzed the gut con-tents of 32 tadpoles of several developmentalstages ranging from Gosner (1960) stage 25 to42 from a single treehole These tadpoles werecollected in four samples taken at roughlymonthly intervals for a period of five months inthe rainy season For a comparison among tree-holes we analyzed gut contents of three to fivetadpoles in the same developmental stage (stage25) collected at one time from four treeholesWe used the following procedure After remov-ing the entire intestinal mass a fragment of 1cm of the foregut was taken and its contentssuspended in 01 ml of distilled water on a mi-croscope slide and examined at a magnificationof 250 to 4003 We quantified the contents byrandomly selecting 10 microscopic fields perslide and noting the presence or absence ineach field of the following five food categoriesdetritus (nonidentifiable particulate organicmatter) plant fragments algae arthropod frag-ments and anuran eggs Dietary compositionwas expressed as mean 6 1 SE of the percent-ages of occurrence of each food item in the in-dividuals analyzed Because more than one foodcategory may be present per microscopic fieldthe sum of the frequencies of occurrence of all
categories may exceed 100 When guts werefilled with eggs the abundance of this item wasassumed to be 100 We then counted intacteggs for the length of the digestive tract Par-tially or totally digested eggs could be identifiedby the texture of residue and confirmed by theobservation of yolk platelets under microscopicexamination To determine whether ingestedeggs were fertilized embryonic developmentwas ascertained by external morphology (Gos-ner 1960) and by histological analysis of somehematoxylin-eosin stained eggs of at least onetadpole gut per sample The latter procedureprovided evidence of early embryonic develop-ment which might not be easily detected by ex-ternal morphology
RESULTS
TreeholesmdashTreeholes used by calling males of Presinifictrix occurred from 22ndash32 m high in treeswith DBH ranging from 13ndash92 cm (mean 6 SE449 6 62 cm n 5 15) No significant differ-ences in DBH were found between trees usedin Panguana and Manaus (Student t-test t 5022 P 5 083)
Treehole openings were circular to ellipticalwith areas ranging from 628ndash4514 cm2 (18046 396 n 5 10 Table 1) Treeholes invariablyfaced upward thus allowing the replenishmentof evaporating water via rainfall and the accu-mulation of allochthonous organic matter Bot-toms of treeholes were usually covered by oneor more layers of dead leaves and decaying or-ganic matter There was no sign of resin liningon the treehole as described by Goeldi (1907)Treehole depth ranged from 15ndash190 cm (8666 277 cm n 5 9 Table 1) Treehole water vol-umes varied from 06ndash900 liters (182 6 93 li-ters n 5 9) but tadpoles were found only intreeholes larger than 28 liters (Table 1) Con-sidering only treeholes sampled during therainy season maximum tadpole abundanceshowed no association to treehole height (P 5017) but was positively associated with treeholewater volume (Y 5 3868 1 362 3 Volume r2
5 077 F 5 1983 P 5 00043) as indicated bylinear regressions No associations were foundbetween maximum tadpole density and tree-hole height (P 5 077) or volume (P 5 078)
Water was less acidic in treeholes (pH 5 6816 029 range 590ndash690 n 5 4) than in pond(564) creek (409) or bromeliads (378 6 02range 357ndash398 n 5 2) Treeholes nearly brack-eted the variation in DO found in other waterbodies DO in treeholes varied from 02ndash39mgl (23 6 11 mgl) when compared to thepond (09 mgl) bromeliads (27 6 06 mgl
266 COPEIA 2003 NO 2
TABLE 1 CHARACTERISTICS OF TREEHOLES USED BY Phrynohyas resinifictrix AND MAXIMUM ABUNDANCE AND DEN-SITY OF TADPOLES OBSERVED PER TREEHOLE
Tree
NumberDBH(cm)
Treehole characteristics
Height(m)
Opening(cm)
Depth(cm)
Volume(L)
Tadpoles
Maxabundance(No inds)
Maxdensity
(IndsL)
Manaus123c
456
341549mdashmdash32
115
32128
145
13 3 1311 3 1123 3 2515 3 1520 3 1321 3 21
19059351932
260
253a
58a
158b
66b
81b
900b
237930
33128337
9371612
0502
1572374
7 27 13 13 3 13 75 89a 10 113
Panguana123
572733
5210422
14 3 0720 3 0810 3 08
70mdash15
28a
mdash06a
83d
0
286mdash
0
Treehole volumes were either a directly measured or b estimatedc Treehole examined at the end of the dry seasond Tadpoles of Dendrobates quinquevittatus species group
range 21ndash33) and creek (41 mgl) Phyto-plankton chlorophyll a concentration was low-est in the creek (031 ugl ultraoligotrophic)followed by treeholes (088 6 032 ugl range015ndash165 ultraoligotrophic to oligotrophic)and pond (449 ugl mesotrophic) See Mag-nusson and Hero (1991) for limnological pa-rameters of additional terrestrial water bodies atthe Manaus study site
Treeholes used by males of P resinifictrix wereformed in the main trunk after the decayingand breaking of larger branches some treesseemed to be in poor condition and at least twowere decaying In Manaus seven of the treeswere identified in the field as Eschweilera cf odo-ra Lecythidaceae (n 5 3 individuals) Aspido-sperma abscurinervea Apocynaceae Trichilia spMeliaceae unidentified Cariocaraceae and un-identified Mimosaceae In Panguana one of thetrees was an unidentified Moraceae
Adult behaviormdashMales called exclusively fromtreeholes On no occasion was this speciesfound in any of the intensively sampled terres-trial water bodies or in other smaller phyto-telms commonly found in the study areas suchas buriti palm leaf axils (Mauritia flexuosa) orground bromeliads (Guzmania spp Streptocalyxspp) Males often called from sundown (1830h) until sunup (0455 h) when they often leftthe treehole but remained in the canopy Call-ing season lasted from August to April althougha few called regularly during the dry seasonMales appeared to show strong site fidelity Callswere heard from the same trees night after
night In Panguana one male continuously re-turned to the same treehole for a period of 20nights In Manaus we toe-clipped and recap-tured a male in treehole 2 four months later(1990) In the following field season (1993) wetoe-clipped and recaptured three males aggres-sively interacting inside the same treehole onfour occasions over two months Aggressive in-teractions between males were common and in-cluded a particular calling repertoire (Zimmer-man and Hodl 1983) and physical attempts toimpede the opponent to enter or remain in thetreehole
Treeholes were often occupied in multipleyears For example five of the six surviving treesused in 1990 were still occupied in 1993 andfive of nine trees used in 1993 were still occu-pied in 1994
Abundance of calling malesmdashCalling males of Presinifictrix were widely spaced in the forest Wefound 48 calling maleskm (015ha) in Pan-guana and from 17ndash20 calling maleskm(005ndash006ha) in Manaus (Table 2) Abun-dances were 24ndash28 times higher in Panguanathan in Manaus Densities were remarkably con-stant in Manaus during the study period
Premetamorphic biology EggsmdashEgg clutches arelaid in water and consist of a gelatinous eggmass that primarily floats on or near the surfacebut often adheres to the inner wall of the tree-hole The darkly pigmented eggs are small(diam 158 6 006 mm n 5 6) and surroundedby gelatinous capsules (diam 428 6 021 mm
267SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
TABLE 2 SEASONAL ESTIMATES OF THE ABUNDANCE
AND DENSITY (LINEAR AND BY AREA) OF CALLING MALES
OF Phrynohyas resinifictrix IN PANGUANA AND MANAUS
Season
Abun-dance
ofcallingmales
Density of calling males
Linear(males
km)Area
(malesha)Area21
(hamale)
Panguana1988 7 48 015 68
Manaus1990199119931994
1095
191720
006005006
171188163
Fig 1 Number of intact eggs found in the guts oftadpoles of Phrynohyas resinifictrix through ontogeny(n 5 32 treehole 2) The dashed line represents anarbitrarily defined developmental threshold at stage32 after which eggs are efficiently used as food re-source Bubble size is proportional to sample size(varying from 1ndash6)
TABLE 3 DIET OF Phrynohyas resinifictrix LARVAE Occurrence expressed as Mean 6 SE
Stages n
Occurrence of food categories ()
AlgaePlant
fragmentsArthropodfragments Detritus Eggs
One treehole sampled over five months25ndash41 32 166 6 51 100 6 28 53 6 16 359 6 84 597 6 87
Four treeholes sampled at one time25252525
5443
60 6 25150 6 65300 6 41300 6 100
180 6 58275 6 103200 6 4133 6 33
00 6 0075 6 4875 6 7500 6 00
580 6 132975 6 25750 6 50730 6 22
600 6 18200 6 0000 6 00
300 6 100
Total44 175 6 39 114 6 23 46 6 13 446 6 67 523 6 72
n 5 6) At the moment of collection clutch sizevaried from 106ndash1540 eggs (436 6 234 eggs n5 6) with as many as six clutches laid in a singletreehole during a period of four months (De-cember 1990 to April 1991)
Larval dietmdashThe diet of P resinifictrix larvaeconsists largely of conspecific eggs and detritusless common items include algae plant frag-ments (mainly leaf fragments but also pollen)and arthropod fragments (insect mouthpartsheads and wing fragments lepidopteran scalesand mosquito larvae bristles Table 3)
Fifty-two percent of all guts examined (n 544) contained eggs in different phases of diges-tion (Table 3) Tadpoles from stages 32ndash42 wereable to swallow whole eggs with capsules andsometimes gelatin (Fig 1) whereas smaller tad-poles ingested egg fragments presumablyrasped with their keratinized mouthparts Wheneggs were available individuals ingested asmany as possible and their guts became greatlyextended The enlarged foregut contained upto 62 whole eggs (Fig 1) plus partially digested
eggs that could not be counted Most eggs ex-amined were fertilized as indicated by externalor histological observation of cell cleavage andembryo development (up to stage 16) Four ofsix clutches laid during a five-month period intreehole 2 at Manaus contained fertilized eggsthe two others could not be analyzed becauseno eggs remained in the gelatinous mass by thetime of observation
Detritus was the second most frequent fooditem consumed observed in 446 of micro-scopic fields when eggs were not available de-
268 COPEIA 2003 NO 2
tritus occurrence reached 975 (Table 3) Allother food items were found much less fre-quently Algae were never abundant althoughsporadically occurring in 175 of all micro-scopic fields or up to 30 in tadpoles in earlydevelopmental stages Plant fragments occurredin 114 of all microscopic fields or up to275 in tadpoles in early developmental stag-es Arthropod fragments were only present in46 of all microscopic fields examined (Table3)
DISCUSSION
Nonaquatic oviposition behavior in frogsevolved independently in many lineages andhas been interpreted as a strategy to avoid pre-dation competition or unfavorable abiotic con-ditions in the ancestral water body (Crump1982 Duellman and Trueb 1986) From anecological rather than evolutionary perspectivethese same processes might be currently oper-ating to maintain arboreal oviposition Magnus-son and Hero (1991) studied an anuran assem-blage at our Central Amazonian rain-forest fieldsite They found that the percentage of specieswith terrestrial eggs and aquatic larvae in a wa-ter body was positively correlated with an indexof egg predation pressure in that water body butonly weakly correlated with the number of spe-cies of potential competitors or abiotic condi-tions such as dissolved oxygen content per-centage of dry days and temperature These ob-servations led the authors to suggest that pre-dation is a major factor maintaining nonaquaticoviposition behavior in this contemporary an-uran assemblage
Oviposition in phytotelms might also bemaintained by the high predation pressure interrestrial water bodies Predation risk is pre-sumably lower in phytotelms because theheight small size concealment and often widespacing of these water bodies reduce predatoraccess to the eggs and tadpoles In addition un-favorable abiotic conditions in phytotelms suchas the potentially low dissolved oxygen levelsand short hydroperiod constrain the survival ofaquatic predators In this study few actual orpotential predators of eggs (wasps small dytis-cid beetles and heterospecific oophagic andcannibalistic tadpoles) tadpoles (spiders) andadults (a colubrid snake Philodryas viridissimus)have been observed inside treeholes inhabitedby P resinifictrix This contrasts strongly with theubiquity of insect and fish predators found inground water bodies at the study site (Magnus-son and Hero 1991)
In addition our observations suggest that
current interspecific competition among Ama-zonian frogs for phytotelms as breeding and de-velopmental sites is minimal and could be an-other important force maintaining ovipositionin these water bodies Only four of 130 lowlandAmazonian frog species breed in phytotelms(Hodl 1990) Moreover in Central Amazonia Presinifictrix is the only anuran to breed exclusive-ly in large water-filled treeholes whereas Osteo-cephalus oophagus breeds in a variety of phyto-telms such as bromeliads buriti (Mauritia fle-xuosa) palm leaf axils palm bracts and only oc-casionally in treeholes ( Jungfer and Schiesari1995 pers obs) Calling adults eggs or larvaeof P resinifictrix and O oophagus were neverfound in the extensively sampled ground waterbodies indicating specialization for a phytotel-mic breeding site Yet tadpoles of these specieswere never found coexisting in any of the phyto-telms sampled indicating strong habitat segre-gation Several species of Dendrobates have ter-restrial oviposition but transport tadpoles tophytotelms after hatching for instance we ob-served in a single occasion Dendrobates cf quin-quevittatus inside treeholes from which P resini-fictrix called More often however Dendrobatesspecies use small-sized ground phytotelms (egCaldwell and Araujo 1998) Overall this strongpattern of habitat segregation among phytotelmtypes in which more than one species are sel-dom found to coexist contrasts with ground wa-ter bodies where typically more than three andsometimes up to eight tadpole species coexist(Magnusson and Hero 1991)
Therefore several lines of evidence suggestthat both predation pressure and interspecificcompetition are lower in phytotelms than in ter-restrial water bodies and could contribute to themaintenance of the use of phytotelms as breed-ing and developmental sites for Amazonian an-urans in general and P resinifictrix in particularIn other systems however an unavailability ofterrestrial water bodies may act as a strong forcemaintaining phytotelm reproduction This ap-pears to be the case in Caribbean Islands suchas Jamaica where historical and geological fac-tors result in phytotelms being the predominantlentic water bodies (Thompson 1996)
Phytotelms are a harsh environment for thedevelopment of anuran embryos and larvaeConstrained volumes of noncirculating waterdominated by decaying processes sometimes re-sult in very low dissolved oxygen concentra-tions Assuming that treehole breeding is de-rived in the genus Phrynohyas early develop-ment of large inflated functional lungs in oth-er pond-dwelling species of the genus may beconsidered a useful morphophysiological trait
269SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
for the colonization of phytotelms by P resinific-trix Lungs are also used as hydrostatic organskeeping tadpoles close to the water surface andmay reduce the energy expenditure involved inrepeatedly swimming to the surface to bob air(Feder 1984 Feder and Moran 1985 Lannooet al 1987)
An important biotic characteristic of phyto-telms is the unpredictability and low availabilityof high quality food In most tadpole species forwhich dietary data are available algae of pe-riphyton and phytoplankton constitute majorfood resources in addition to low-quality detri-tus (eg Altig and Johnston 1989 Kupferberget al 1994) Narrow treehole openings oftencovered by overlying canopy may result in anearly aphotic habitat in which primary pro-ductivity is low and therefore likely insufficientto sustain tadpole growth
Detritus derived from the decomposition offallen leaf litter and treehole walls constitutedthe basis for the speciesrsquo diet when eggs werenot available The nutritive value of detritus islow (Kupferberg et al 1994) and several spe-cies of generalized pond tadpoles fail to reachmetamorphosis when restricted to this food type(Werner and Glennemeier 1999 LS unpubl)In this context oophagy may be of major im-portance for the success of individual larvae inreaching metamorphosis because eggs providea highly nutritious and energetic food source(Crump 1992) However detritivory could alsobe of crucial importance especially for smallertadpoles Prior to stage 32 tadpoles are gape-limited and therefore inefficient egg feedersAfter this stage tadpoles are capable of swallow-ing large quantities of whole eggs leading toaccelerated growth and development
Opportunistic oophagy is common in anuranlarvae (Magnusson and Hero 1991 Crump1992 Petranka and Kennedy 1999) In severalphytotelm-developing anurans however oop-hagy results from complex parental behavior inwhich the mother periodically returns to thephytotelm to lay fertilized or unfertilized eggsfor nutrition of its progeny (Lannoo et al 1987Weygoldt 1987 Crump 1995) For some ofthese species it was demonstrated that oophagyis obligate that is tadpoles fail to reach meta-morphosis if eggs are not provided by the moth-er (Weygoldt 1987 Thompson 1996 Jungferand Weygoldt 1999) In P resinifictrix howeverthere are several observations suggesting thatoophagy is opportunistic cannibalism ratherthan parental care in the form of periodic eggdeposition for obligate tadpole nutrition Firstas noted by Jungfer and Proy (1998) P resinific-trix egg clutches are large (maximum clutch size
in the field 1540 eggs this study in terraria4336 eggs Jungfer and Proy 1998) comparedto phytotelm-breeding species that intentionallyprovide eggs to their offspring For instanceconsidering only hylids on average 158 eggs arelaid by Anotheca spinosa ( Jungfer 1996) 258 byOsteocephalus oophagus ( Jungfer and Weygoldt1999) 276 per bromeliad by Osteopilus brunneus(Lannoo et al 1987) and 13 by Phyllodytes lu-teolus (Giaretta 1996) Second as a conse-quence of the large clutch size interclutch in-terval is long in P resinifictrix in a regularly sam-pled treehole six clutches were deposited overa period of four months (around one clutch ev-ery 20 days) which coincides with the inter-clutch interval of pairs in captivity (20 daysrange 7ndash37 n 5 29 Jungfer and Proy 1998)This long interclutch interval contrasts stronglywith the interclutch interval of species that areknown to be obligate oophagic For instanceeggs are deposited every 49 days in A spinosa( Jungfer 1996) 51 days in O oophagus ( Jung-fer and Weygoldt 1999) and 44 days in Obrunneus (Thompson 1996) Jungfer and Proy(1998) found that tadpoles of P resinifictrixstarve if dependent only on the motherrsquos infre-quent oviposition as the food source but thatmetamorphic success in an experimental con-tainer was high if in addition to leaf-litter eggswere added every 17 days Third an opportu-nistic oophagic nutrition is suggested by our ob-servation that eggs are fertilized unlike some ofthe more lsquolsquoadvancedrsquorsquo forms of anuran parentalcare through laying of unfertilized trophic eggs(Weygoldt 1987) although obligate oophagyhas been documented for species laying fertil-ized eggs (Thompson 1996) Fourth althoughextensive reduction of oral structures character-izes several of the obligate oophagic phytotelm-developing species (Lannoo et al 1987) P re-sinifictrix retains typical generalized pond-typelarval morphology with only moderate structur-al reduction when compared to its pond breed-ing congeners (Schiesari et al 1996) Experi-mental evidence is necessary to unambiguouslydemonstrate that oophagy is not obligate in Presinifictrix however the arguments above pro-vide indirect support for the hypothesis thatconsumption of eggs is important but may notbe obligate for metamorphic success
We suggest that among the premetamorphicstages of P resinifictrix life cycle cannibalism andintraspecific competition are the major forcesacting on the survival of eggs and larvae re-spectively Evidence for this hypothesis are thelow nutritive value of food other than eggs andor high unpredictability of food in the treeholethe voracious oophagic habits of the tadpole
270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo

264 COPEIA 2003 NO 2
est Long-range communication is effected witha loud low-pitched call which is carried forlong distances through the use of the rain forestlow frequency window (Zimmerman and Hodl1983 Hodl 1991) When present the hollowportion of the treehole may act as a resonatingchamber increasing the broadcasting distancefor the calls (Hodl 1991)
Phrynohyas resinifictrix tadpoles collected fromtreeholes have been the subject of detailed eco-morphological studies (Grillitsch 1992 Schie-sari et al 1996) which predicted an ability toingest large food particles and a reliance on pul-monary as well as branchial respiration Withthe exception of the studies mentioned abovevery little information is available about theecology of this species
We show here that P resinifictrix is capable ofcompleting its entire life cycle in the forest can-opy by breeding exclusively in large water-filledtreeholes We present data on the physical andbiological aspects of the treeholes used as call-ing and breeding sites by the species as well asestimates of calling male population densitiesQuantitative dietary data are examined to clar-ify the strategies allowing completion of meta-morphosis in these unproductive microhabitatsWe discuss the importance of predation of eggsand larvae and low interspecific competition asmajor forces maintaining oviposition in phyto-telms Finally we discuss the hypothesis thattreeholes constitute a limiting resource for Presinifictrix populations and suggest that malesmonopolizing treeholes may obtain multiplemates
MATERIALS AND METHODS
Fieldwork was conducted in two Amazonianterra-firme rain-forest sites Panguana Peru(lower Rio Llullapichis 98379S 748569W) byWH from October to November 1988 and theReserva Florestal Adolfo Ducke (hereforth re-ferred to as lsquolsquoManausrsquorsquo) located about 25 kmnorth of Manaus Amazonas Brazil (038089S608029W) by LS and MG from November 1990to March 1991 and from January to April 1993Sporadic additional observations were conduct-ed from January to June 1994 and from Augustto October 1994 Annual rainfall is 2635 mm inPanguana (Aichinger 1987) and 2075 mm inManaus (mean values from 1931 to 1960 Brasil1978) The rainy season occurs between Sep-tember and May (Panguana) or December andMay (Manaus) with monthly rainfall of morethan 200 mm whereas the driest months Julyand August (plus September in Manaus) re-ceive less than 60 mm each It is noteworthy that
during the study period mean annual rainfall inManaus was 2660 mm 30 higher than the av-erage (data collected at an EMBRAPA meteo-rological station 5 km north of the study site)
We searched for calling males while walkingalong a single trail 1470 m long in Panguana(totaling at least 94 km walked in 32 nights) andalong a system of trails 8854 m long in Manaus(totaling at least 151 km walked in 36 nights in19901991 and 145 km walked in 57 nights in1993) We located the trees from which malefrogs were calling by approaching the source ofthe loud call during the night and subsequentlypressing an ear tightly against tree trunks Thetrunk hosting the calling male was the one thatbest conducted sound vibrations Wheneverpossible treeholes were reached either bymeans of ladders lsquolsquogriffesrsquorsquo (metal hooks withspikes used for tree climbing see Mori 1987for description and illustration) or mountain-eering equipment We located 10 trees in Pan-guana and 19 in Manaus and were able to in-spect three and seven treeholes respectively
We estimated the density of calling males bydividing the number of regularly calling males(the abundance) by the length of trails walked(individualskm linear density) and by the areaacoustically sampled (individualsha density byarea) To estimate the area acoustically sampledwe multiplied the length of trails walked bytwice the maximum distance inside the forest inwhich calls were above ambient noise levelsThese were determined with a CEL-493 Preci-sion Impulse Integrating Sound Level Meterand a CEL-296 Third Octave Autoscan FilterSet Although in some cases calls were withinhuman hearing range from distances up to 250m sound pressure levels were consistently abovethe ambient noise level at a maximal distanceof 160 m at the 800 Hz terz filter band At thisfrequency and distance sound levels were be-tween 24 dB and 32 dB (linear peak relative to20 mNm2) Therefore we employed the con-servative value of 160 m for our estimates ofarea acoustically sampled
We also measured diameter at breast height(DBH) of the host tree dimensions of the tree-hole opening water surface and depth of thetreehole and its height Treehole water vol-umes were either directly measured by suckingout water through a hose and pouring it into ameasuring cup or estimated by employing thecylinder volume formula to the water surfaceand treehole depth measured A cylindricalmodel appears to be a good estimator of tree-hole shape no significant differences werefound between the actual and estimated vol-umes of three treeholes for which both sets of
265SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
data are available (Student t-test t 5 068 P 5057)
Treeholes were inspected for 20ndash30 min onone to 30 occasions depending on accessibilityTreehole limnological parameters were mea-sured on 19 March 1993 and 31 May 1994 (Ma-naus) Values of pH dissolved oxygen concen-tration (DO) and algal chlorophyll concentra-tion (as a measure of phytoplankton standingcrop) of the surface water present in four tree-holes two bromeliads a small temporary pondand a dammed creek were analyzed We deter-mined pH with a Corning pH meter 130 andDO with an YSI Dissolved Oxygen meter eitherin situ or on the ground after 500 ml of waterwere collected with minimum disturbanceChlorophyll a was extracted from 250 mL of wa-ter and analyzed in spectrophotometer for ex-tinctions at 7500 A (Strickland and Parsons1968) Classification of water body trophic type(productivity) followed Wetzel (1983)
Eggs and tadpoles were collected with smallcircular nets or sucked with a hose and pre-served in 5 formalin Identity of larvae wasconfirmed by rearing until metamorphosis orby the aid of literature sources (Hero 1990 Gri-llitsch 1992 Schiesari et al 1996) Voucherspecimens were deposited in the Museum of Zo-ology of the University of Sao Paulo (MZUSP128321) Larvae were staged according to Gos-ner (1960) To quantify the dietary habits oftadpoles from Manaus we analyzed the gut con-tents of 32 tadpoles of several developmentalstages ranging from Gosner (1960) stage 25 to42 from a single treehole These tadpoles werecollected in four samples taken at roughlymonthly intervals for a period of five months inthe rainy season For a comparison among tree-holes we analyzed gut contents of three to fivetadpoles in the same developmental stage (stage25) collected at one time from four treeholesWe used the following procedure After remov-ing the entire intestinal mass a fragment of 1cm of the foregut was taken and its contentssuspended in 01 ml of distilled water on a mi-croscope slide and examined at a magnificationof 250 to 4003 We quantified the contents byrandomly selecting 10 microscopic fields perslide and noting the presence or absence ineach field of the following five food categoriesdetritus (nonidentifiable particulate organicmatter) plant fragments algae arthropod frag-ments and anuran eggs Dietary compositionwas expressed as mean 6 1 SE of the percent-ages of occurrence of each food item in the in-dividuals analyzed Because more than one foodcategory may be present per microscopic fieldthe sum of the frequencies of occurrence of all
categories may exceed 100 When guts werefilled with eggs the abundance of this item wasassumed to be 100 We then counted intacteggs for the length of the digestive tract Par-tially or totally digested eggs could be identifiedby the texture of residue and confirmed by theobservation of yolk platelets under microscopicexamination To determine whether ingestedeggs were fertilized embryonic developmentwas ascertained by external morphology (Gos-ner 1960) and by histological analysis of somehematoxylin-eosin stained eggs of at least onetadpole gut per sample The latter procedureprovided evidence of early embryonic develop-ment which might not be easily detected by ex-ternal morphology
RESULTS
TreeholesmdashTreeholes used by calling males of Presinifictrix occurred from 22ndash32 m high in treeswith DBH ranging from 13ndash92 cm (mean 6 SE449 6 62 cm n 5 15) No significant differ-ences in DBH were found between trees usedin Panguana and Manaus (Student t-test t 5022 P 5 083)
Treehole openings were circular to ellipticalwith areas ranging from 628ndash4514 cm2 (18046 396 n 5 10 Table 1) Treeholes invariablyfaced upward thus allowing the replenishmentof evaporating water via rainfall and the accu-mulation of allochthonous organic matter Bot-toms of treeholes were usually covered by oneor more layers of dead leaves and decaying or-ganic matter There was no sign of resin liningon the treehole as described by Goeldi (1907)Treehole depth ranged from 15ndash190 cm (8666 277 cm n 5 9 Table 1) Treehole water vol-umes varied from 06ndash900 liters (182 6 93 li-ters n 5 9) but tadpoles were found only intreeholes larger than 28 liters (Table 1) Con-sidering only treeholes sampled during therainy season maximum tadpole abundanceshowed no association to treehole height (P 5017) but was positively associated with treeholewater volume (Y 5 3868 1 362 3 Volume r2
5 077 F 5 1983 P 5 00043) as indicated bylinear regressions No associations were foundbetween maximum tadpole density and tree-hole height (P 5 077) or volume (P 5 078)
Water was less acidic in treeholes (pH 5 6816 029 range 590ndash690 n 5 4) than in pond(564) creek (409) or bromeliads (378 6 02range 357ndash398 n 5 2) Treeholes nearly brack-eted the variation in DO found in other waterbodies DO in treeholes varied from 02ndash39mgl (23 6 11 mgl) when compared to thepond (09 mgl) bromeliads (27 6 06 mgl
266 COPEIA 2003 NO 2
TABLE 1 CHARACTERISTICS OF TREEHOLES USED BY Phrynohyas resinifictrix AND MAXIMUM ABUNDANCE AND DEN-SITY OF TADPOLES OBSERVED PER TREEHOLE
Tree
NumberDBH(cm)
Treehole characteristics
Height(m)
Opening(cm)
Depth(cm)
Volume(L)
Tadpoles
Maxabundance(No inds)
Maxdensity
(IndsL)
Manaus123c
456
341549mdashmdash32
115
32128
145
13 3 1311 3 1123 3 2515 3 1520 3 1321 3 21
19059351932
260
253a
58a
158b
66b
81b
900b
237930
33128337
9371612
0502
1572374
7 27 13 13 3 13 75 89a 10 113
Panguana123
572733
5210422
14 3 0720 3 0810 3 08
70mdash15
28a
mdash06a
83d
0
286mdash
0
Treehole volumes were either a directly measured or b estimatedc Treehole examined at the end of the dry seasond Tadpoles of Dendrobates quinquevittatus species group
range 21ndash33) and creek (41 mgl) Phyto-plankton chlorophyll a concentration was low-est in the creek (031 ugl ultraoligotrophic)followed by treeholes (088 6 032 ugl range015ndash165 ultraoligotrophic to oligotrophic)and pond (449 ugl mesotrophic) See Mag-nusson and Hero (1991) for limnological pa-rameters of additional terrestrial water bodies atthe Manaus study site
Treeholes used by males of P resinifictrix wereformed in the main trunk after the decayingand breaking of larger branches some treesseemed to be in poor condition and at least twowere decaying In Manaus seven of the treeswere identified in the field as Eschweilera cf odo-ra Lecythidaceae (n 5 3 individuals) Aspido-sperma abscurinervea Apocynaceae Trichilia spMeliaceae unidentified Cariocaraceae and un-identified Mimosaceae In Panguana one of thetrees was an unidentified Moraceae
Adult behaviormdashMales called exclusively fromtreeholes On no occasion was this speciesfound in any of the intensively sampled terres-trial water bodies or in other smaller phyto-telms commonly found in the study areas suchas buriti palm leaf axils (Mauritia flexuosa) orground bromeliads (Guzmania spp Streptocalyxspp) Males often called from sundown (1830h) until sunup (0455 h) when they often leftthe treehole but remained in the canopy Call-ing season lasted from August to April althougha few called regularly during the dry seasonMales appeared to show strong site fidelity Callswere heard from the same trees night after
night In Panguana one male continuously re-turned to the same treehole for a period of 20nights In Manaus we toe-clipped and recap-tured a male in treehole 2 four months later(1990) In the following field season (1993) wetoe-clipped and recaptured three males aggres-sively interacting inside the same treehole onfour occasions over two months Aggressive in-teractions between males were common and in-cluded a particular calling repertoire (Zimmer-man and Hodl 1983) and physical attempts toimpede the opponent to enter or remain in thetreehole
Treeholes were often occupied in multipleyears For example five of the six surviving treesused in 1990 were still occupied in 1993 andfive of nine trees used in 1993 were still occu-pied in 1994
Abundance of calling malesmdashCalling males of Presinifictrix were widely spaced in the forest Wefound 48 calling maleskm (015ha) in Pan-guana and from 17ndash20 calling maleskm(005ndash006ha) in Manaus (Table 2) Abun-dances were 24ndash28 times higher in Panguanathan in Manaus Densities were remarkably con-stant in Manaus during the study period
Premetamorphic biology EggsmdashEgg clutches arelaid in water and consist of a gelatinous eggmass that primarily floats on or near the surfacebut often adheres to the inner wall of the tree-hole The darkly pigmented eggs are small(diam 158 6 006 mm n 5 6) and surroundedby gelatinous capsules (diam 428 6 021 mm
267SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
TABLE 2 SEASONAL ESTIMATES OF THE ABUNDANCE
AND DENSITY (LINEAR AND BY AREA) OF CALLING MALES
OF Phrynohyas resinifictrix IN PANGUANA AND MANAUS
Season
Abun-dance
ofcallingmales
Density of calling males
Linear(males
km)Area
(malesha)Area21
(hamale)
Panguana1988 7 48 015 68
Manaus1990199119931994
1095
191720
006005006
171188163
Fig 1 Number of intact eggs found in the guts oftadpoles of Phrynohyas resinifictrix through ontogeny(n 5 32 treehole 2) The dashed line represents anarbitrarily defined developmental threshold at stage32 after which eggs are efficiently used as food re-source Bubble size is proportional to sample size(varying from 1ndash6)
TABLE 3 DIET OF Phrynohyas resinifictrix LARVAE Occurrence expressed as Mean 6 SE
Stages n
Occurrence of food categories ()
AlgaePlant
fragmentsArthropodfragments Detritus Eggs
One treehole sampled over five months25ndash41 32 166 6 51 100 6 28 53 6 16 359 6 84 597 6 87
Four treeholes sampled at one time25252525
5443
60 6 25150 6 65300 6 41300 6 100
180 6 58275 6 103200 6 4133 6 33
00 6 0075 6 4875 6 7500 6 00
580 6 132975 6 25750 6 50730 6 22
600 6 18200 6 0000 6 00
300 6 100
Total44 175 6 39 114 6 23 46 6 13 446 6 67 523 6 72
n 5 6) At the moment of collection clutch sizevaried from 106ndash1540 eggs (436 6 234 eggs n5 6) with as many as six clutches laid in a singletreehole during a period of four months (De-cember 1990 to April 1991)
Larval dietmdashThe diet of P resinifictrix larvaeconsists largely of conspecific eggs and detritusless common items include algae plant frag-ments (mainly leaf fragments but also pollen)and arthropod fragments (insect mouthpartsheads and wing fragments lepidopteran scalesand mosquito larvae bristles Table 3)
Fifty-two percent of all guts examined (n 544) contained eggs in different phases of diges-tion (Table 3) Tadpoles from stages 32ndash42 wereable to swallow whole eggs with capsules andsometimes gelatin (Fig 1) whereas smaller tad-poles ingested egg fragments presumablyrasped with their keratinized mouthparts Wheneggs were available individuals ingested asmany as possible and their guts became greatlyextended The enlarged foregut contained upto 62 whole eggs (Fig 1) plus partially digested
eggs that could not be counted Most eggs ex-amined were fertilized as indicated by externalor histological observation of cell cleavage andembryo development (up to stage 16) Four ofsix clutches laid during a five-month period intreehole 2 at Manaus contained fertilized eggsthe two others could not be analyzed becauseno eggs remained in the gelatinous mass by thetime of observation
Detritus was the second most frequent fooditem consumed observed in 446 of micro-scopic fields when eggs were not available de-
268 COPEIA 2003 NO 2
tritus occurrence reached 975 (Table 3) Allother food items were found much less fre-quently Algae were never abundant althoughsporadically occurring in 175 of all micro-scopic fields or up to 30 in tadpoles in earlydevelopmental stages Plant fragments occurredin 114 of all microscopic fields or up to275 in tadpoles in early developmental stag-es Arthropod fragments were only present in46 of all microscopic fields examined (Table3)
DISCUSSION
Nonaquatic oviposition behavior in frogsevolved independently in many lineages andhas been interpreted as a strategy to avoid pre-dation competition or unfavorable abiotic con-ditions in the ancestral water body (Crump1982 Duellman and Trueb 1986) From anecological rather than evolutionary perspectivethese same processes might be currently oper-ating to maintain arboreal oviposition Magnus-son and Hero (1991) studied an anuran assem-blage at our Central Amazonian rain-forest fieldsite They found that the percentage of specieswith terrestrial eggs and aquatic larvae in a wa-ter body was positively correlated with an indexof egg predation pressure in that water body butonly weakly correlated with the number of spe-cies of potential competitors or abiotic condi-tions such as dissolved oxygen content per-centage of dry days and temperature These ob-servations led the authors to suggest that pre-dation is a major factor maintaining nonaquaticoviposition behavior in this contemporary an-uran assemblage
Oviposition in phytotelms might also bemaintained by the high predation pressure interrestrial water bodies Predation risk is pre-sumably lower in phytotelms because theheight small size concealment and often widespacing of these water bodies reduce predatoraccess to the eggs and tadpoles In addition un-favorable abiotic conditions in phytotelms suchas the potentially low dissolved oxygen levelsand short hydroperiod constrain the survival ofaquatic predators In this study few actual orpotential predators of eggs (wasps small dytis-cid beetles and heterospecific oophagic andcannibalistic tadpoles) tadpoles (spiders) andadults (a colubrid snake Philodryas viridissimus)have been observed inside treeholes inhabitedby P resinifictrix This contrasts strongly with theubiquity of insect and fish predators found inground water bodies at the study site (Magnus-son and Hero 1991)
In addition our observations suggest that
current interspecific competition among Ama-zonian frogs for phytotelms as breeding and de-velopmental sites is minimal and could be an-other important force maintaining ovipositionin these water bodies Only four of 130 lowlandAmazonian frog species breed in phytotelms(Hodl 1990) Moreover in Central Amazonia Presinifictrix is the only anuran to breed exclusive-ly in large water-filled treeholes whereas Osteo-cephalus oophagus breeds in a variety of phyto-telms such as bromeliads buriti (Mauritia fle-xuosa) palm leaf axils palm bracts and only oc-casionally in treeholes ( Jungfer and Schiesari1995 pers obs) Calling adults eggs or larvaeof P resinifictrix and O oophagus were neverfound in the extensively sampled ground waterbodies indicating specialization for a phytotel-mic breeding site Yet tadpoles of these specieswere never found coexisting in any of the phyto-telms sampled indicating strong habitat segre-gation Several species of Dendrobates have ter-restrial oviposition but transport tadpoles tophytotelms after hatching for instance we ob-served in a single occasion Dendrobates cf quin-quevittatus inside treeholes from which P resini-fictrix called More often however Dendrobatesspecies use small-sized ground phytotelms (egCaldwell and Araujo 1998) Overall this strongpattern of habitat segregation among phytotelmtypes in which more than one species are sel-dom found to coexist contrasts with ground wa-ter bodies where typically more than three andsometimes up to eight tadpole species coexist(Magnusson and Hero 1991)
Therefore several lines of evidence suggestthat both predation pressure and interspecificcompetition are lower in phytotelms than in ter-restrial water bodies and could contribute to themaintenance of the use of phytotelms as breed-ing and developmental sites for Amazonian an-urans in general and P resinifictrix in particularIn other systems however an unavailability ofterrestrial water bodies may act as a strong forcemaintaining phytotelm reproduction This ap-pears to be the case in Caribbean Islands suchas Jamaica where historical and geological fac-tors result in phytotelms being the predominantlentic water bodies (Thompson 1996)
Phytotelms are a harsh environment for thedevelopment of anuran embryos and larvaeConstrained volumes of noncirculating waterdominated by decaying processes sometimes re-sult in very low dissolved oxygen concentra-tions Assuming that treehole breeding is de-rived in the genus Phrynohyas early develop-ment of large inflated functional lungs in oth-er pond-dwelling species of the genus may beconsidered a useful morphophysiological trait
269SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
for the colonization of phytotelms by P resinific-trix Lungs are also used as hydrostatic organskeeping tadpoles close to the water surface andmay reduce the energy expenditure involved inrepeatedly swimming to the surface to bob air(Feder 1984 Feder and Moran 1985 Lannooet al 1987)
An important biotic characteristic of phyto-telms is the unpredictability and low availabilityof high quality food In most tadpole species forwhich dietary data are available algae of pe-riphyton and phytoplankton constitute majorfood resources in addition to low-quality detri-tus (eg Altig and Johnston 1989 Kupferberget al 1994) Narrow treehole openings oftencovered by overlying canopy may result in anearly aphotic habitat in which primary pro-ductivity is low and therefore likely insufficientto sustain tadpole growth
Detritus derived from the decomposition offallen leaf litter and treehole walls constitutedthe basis for the speciesrsquo diet when eggs werenot available The nutritive value of detritus islow (Kupferberg et al 1994) and several spe-cies of generalized pond tadpoles fail to reachmetamorphosis when restricted to this food type(Werner and Glennemeier 1999 LS unpubl)In this context oophagy may be of major im-portance for the success of individual larvae inreaching metamorphosis because eggs providea highly nutritious and energetic food source(Crump 1992) However detritivory could alsobe of crucial importance especially for smallertadpoles Prior to stage 32 tadpoles are gape-limited and therefore inefficient egg feedersAfter this stage tadpoles are capable of swallow-ing large quantities of whole eggs leading toaccelerated growth and development
Opportunistic oophagy is common in anuranlarvae (Magnusson and Hero 1991 Crump1992 Petranka and Kennedy 1999) In severalphytotelm-developing anurans however oop-hagy results from complex parental behavior inwhich the mother periodically returns to thephytotelm to lay fertilized or unfertilized eggsfor nutrition of its progeny (Lannoo et al 1987Weygoldt 1987 Crump 1995) For some ofthese species it was demonstrated that oophagyis obligate that is tadpoles fail to reach meta-morphosis if eggs are not provided by the moth-er (Weygoldt 1987 Thompson 1996 Jungferand Weygoldt 1999) In P resinifictrix howeverthere are several observations suggesting thatoophagy is opportunistic cannibalism ratherthan parental care in the form of periodic eggdeposition for obligate tadpole nutrition Firstas noted by Jungfer and Proy (1998) P resinific-trix egg clutches are large (maximum clutch size
in the field 1540 eggs this study in terraria4336 eggs Jungfer and Proy 1998) comparedto phytotelm-breeding species that intentionallyprovide eggs to their offspring For instanceconsidering only hylids on average 158 eggs arelaid by Anotheca spinosa ( Jungfer 1996) 258 byOsteocephalus oophagus ( Jungfer and Weygoldt1999) 276 per bromeliad by Osteopilus brunneus(Lannoo et al 1987) and 13 by Phyllodytes lu-teolus (Giaretta 1996) Second as a conse-quence of the large clutch size interclutch in-terval is long in P resinifictrix in a regularly sam-pled treehole six clutches were deposited overa period of four months (around one clutch ev-ery 20 days) which coincides with the inter-clutch interval of pairs in captivity (20 daysrange 7ndash37 n 5 29 Jungfer and Proy 1998)This long interclutch interval contrasts stronglywith the interclutch interval of species that areknown to be obligate oophagic For instanceeggs are deposited every 49 days in A spinosa( Jungfer 1996) 51 days in O oophagus ( Jung-fer and Weygoldt 1999) and 44 days in Obrunneus (Thompson 1996) Jungfer and Proy(1998) found that tadpoles of P resinifictrixstarve if dependent only on the motherrsquos infre-quent oviposition as the food source but thatmetamorphic success in an experimental con-tainer was high if in addition to leaf-litter eggswere added every 17 days Third an opportu-nistic oophagic nutrition is suggested by our ob-servation that eggs are fertilized unlike some ofthe more lsquolsquoadvancedrsquorsquo forms of anuran parentalcare through laying of unfertilized trophic eggs(Weygoldt 1987) although obligate oophagyhas been documented for species laying fertil-ized eggs (Thompson 1996) Fourth althoughextensive reduction of oral structures character-izes several of the obligate oophagic phytotelm-developing species (Lannoo et al 1987) P re-sinifictrix retains typical generalized pond-typelarval morphology with only moderate structur-al reduction when compared to its pond breed-ing congeners (Schiesari et al 1996) Experi-mental evidence is necessary to unambiguouslydemonstrate that oophagy is not obligate in Presinifictrix however the arguments above pro-vide indirect support for the hypothesis thatconsumption of eggs is important but may notbe obligate for metamorphic success
We suggest that among the premetamorphicstages of P resinifictrix life cycle cannibalism andintraspecific competition are the major forcesacting on the survival of eggs and larvae re-spectively Evidence for this hypothesis are thelow nutritive value of food other than eggs andor high unpredictability of food in the treeholethe voracious oophagic habits of the tadpole
270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo

265SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
data are available (Student t-test t 5 068 P 5057)
Treeholes were inspected for 20ndash30 min onone to 30 occasions depending on accessibilityTreehole limnological parameters were mea-sured on 19 March 1993 and 31 May 1994 (Ma-naus) Values of pH dissolved oxygen concen-tration (DO) and algal chlorophyll concentra-tion (as a measure of phytoplankton standingcrop) of the surface water present in four tree-holes two bromeliads a small temporary pondand a dammed creek were analyzed We deter-mined pH with a Corning pH meter 130 andDO with an YSI Dissolved Oxygen meter eitherin situ or on the ground after 500 ml of waterwere collected with minimum disturbanceChlorophyll a was extracted from 250 mL of wa-ter and analyzed in spectrophotometer for ex-tinctions at 7500 A (Strickland and Parsons1968) Classification of water body trophic type(productivity) followed Wetzel (1983)
Eggs and tadpoles were collected with smallcircular nets or sucked with a hose and pre-served in 5 formalin Identity of larvae wasconfirmed by rearing until metamorphosis orby the aid of literature sources (Hero 1990 Gri-llitsch 1992 Schiesari et al 1996) Voucherspecimens were deposited in the Museum of Zo-ology of the University of Sao Paulo (MZUSP128321) Larvae were staged according to Gos-ner (1960) To quantify the dietary habits oftadpoles from Manaus we analyzed the gut con-tents of 32 tadpoles of several developmentalstages ranging from Gosner (1960) stage 25 to42 from a single treehole These tadpoles werecollected in four samples taken at roughlymonthly intervals for a period of five months inthe rainy season For a comparison among tree-holes we analyzed gut contents of three to fivetadpoles in the same developmental stage (stage25) collected at one time from four treeholesWe used the following procedure After remov-ing the entire intestinal mass a fragment of 1cm of the foregut was taken and its contentssuspended in 01 ml of distilled water on a mi-croscope slide and examined at a magnificationof 250 to 4003 We quantified the contents byrandomly selecting 10 microscopic fields perslide and noting the presence or absence ineach field of the following five food categoriesdetritus (nonidentifiable particulate organicmatter) plant fragments algae arthropod frag-ments and anuran eggs Dietary compositionwas expressed as mean 6 1 SE of the percent-ages of occurrence of each food item in the in-dividuals analyzed Because more than one foodcategory may be present per microscopic fieldthe sum of the frequencies of occurrence of all
categories may exceed 100 When guts werefilled with eggs the abundance of this item wasassumed to be 100 We then counted intacteggs for the length of the digestive tract Par-tially or totally digested eggs could be identifiedby the texture of residue and confirmed by theobservation of yolk platelets under microscopicexamination To determine whether ingestedeggs were fertilized embryonic developmentwas ascertained by external morphology (Gos-ner 1960) and by histological analysis of somehematoxylin-eosin stained eggs of at least onetadpole gut per sample The latter procedureprovided evidence of early embryonic develop-ment which might not be easily detected by ex-ternal morphology
RESULTS
TreeholesmdashTreeholes used by calling males of Presinifictrix occurred from 22ndash32 m high in treeswith DBH ranging from 13ndash92 cm (mean 6 SE449 6 62 cm n 5 15) No significant differ-ences in DBH were found between trees usedin Panguana and Manaus (Student t-test t 5022 P 5 083)
Treehole openings were circular to ellipticalwith areas ranging from 628ndash4514 cm2 (18046 396 n 5 10 Table 1) Treeholes invariablyfaced upward thus allowing the replenishmentof evaporating water via rainfall and the accu-mulation of allochthonous organic matter Bot-toms of treeholes were usually covered by oneor more layers of dead leaves and decaying or-ganic matter There was no sign of resin liningon the treehole as described by Goeldi (1907)Treehole depth ranged from 15ndash190 cm (8666 277 cm n 5 9 Table 1) Treehole water vol-umes varied from 06ndash900 liters (182 6 93 li-ters n 5 9) but tadpoles were found only intreeholes larger than 28 liters (Table 1) Con-sidering only treeholes sampled during therainy season maximum tadpole abundanceshowed no association to treehole height (P 5017) but was positively associated with treeholewater volume (Y 5 3868 1 362 3 Volume r2
5 077 F 5 1983 P 5 00043) as indicated bylinear regressions No associations were foundbetween maximum tadpole density and tree-hole height (P 5 077) or volume (P 5 078)
Water was less acidic in treeholes (pH 5 6816 029 range 590ndash690 n 5 4) than in pond(564) creek (409) or bromeliads (378 6 02range 357ndash398 n 5 2) Treeholes nearly brack-eted the variation in DO found in other waterbodies DO in treeholes varied from 02ndash39mgl (23 6 11 mgl) when compared to thepond (09 mgl) bromeliads (27 6 06 mgl
266 COPEIA 2003 NO 2
TABLE 1 CHARACTERISTICS OF TREEHOLES USED BY Phrynohyas resinifictrix AND MAXIMUM ABUNDANCE AND DEN-SITY OF TADPOLES OBSERVED PER TREEHOLE
Tree
NumberDBH(cm)
Treehole characteristics
Height(m)
Opening(cm)
Depth(cm)
Volume(L)
Tadpoles
Maxabundance(No inds)
Maxdensity
(IndsL)
Manaus123c
456
341549mdashmdash32
115
32128
145
13 3 1311 3 1123 3 2515 3 1520 3 1321 3 21
19059351932
260
253a
58a
158b
66b
81b
900b
237930
33128337
9371612
0502
1572374
7 27 13 13 3 13 75 89a 10 113
Panguana123
572733
5210422
14 3 0720 3 0810 3 08
70mdash15
28a
mdash06a
83d
0
286mdash
0
Treehole volumes were either a directly measured or b estimatedc Treehole examined at the end of the dry seasond Tadpoles of Dendrobates quinquevittatus species group
range 21ndash33) and creek (41 mgl) Phyto-plankton chlorophyll a concentration was low-est in the creek (031 ugl ultraoligotrophic)followed by treeholes (088 6 032 ugl range015ndash165 ultraoligotrophic to oligotrophic)and pond (449 ugl mesotrophic) See Mag-nusson and Hero (1991) for limnological pa-rameters of additional terrestrial water bodies atthe Manaus study site
Treeholes used by males of P resinifictrix wereformed in the main trunk after the decayingand breaking of larger branches some treesseemed to be in poor condition and at least twowere decaying In Manaus seven of the treeswere identified in the field as Eschweilera cf odo-ra Lecythidaceae (n 5 3 individuals) Aspido-sperma abscurinervea Apocynaceae Trichilia spMeliaceae unidentified Cariocaraceae and un-identified Mimosaceae In Panguana one of thetrees was an unidentified Moraceae
Adult behaviormdashMales called exclusively fromtreeholes On no occasion was this speciesfound in any of the intensively sampled terres-trial water bodies or in other smaller phyto-telms commonly found in the study areas suchas buriti palm leaf axils (Mauritia flexuosa) orground bromeliads (Guzmania spp Streptocalyxspp) Males often called from sundown (1830h) until sunup (0455 h) when they often leftthe treehole but remained in the canopy Call-ing season lasted from August to April althougha few called regularly during the dry seasonMales appeared to show strong site fidelity Callswere heard from the same trees night after
night In Panguana one male continuously re-turned to the same treehole for a period of 20nights In Manaus we toe-clipped and recap-tured a male in treehole 2 four months later(1990) In the following field season (1993) wetoe-clipped and recaptured three males aggres-sively interacting inside the same treehole onfour occasions over two months Aggressive in-teractions between males were common and in-cluded a particular calling repertoire (Zimmer-man and Hodl 1983) and physical attempts toimpede the opponent to enter or remain in thetreehole
Treeholes were often occupied in multipleyears For example five of the six surviving treesused in 1990 were still occupied in 1993 andfive of nine trees used in 1993 were still occu-pied in 1994
Abundance of calling malesmdashCalling males of Presinifictrix were widely spaced in the forest Wefound 48 calling maleskm (015ha) in Pan-guana and from 17ndash20 calling maleskm(005ndash006ha) in Manaus (Table 2) Abun-dances were 24ndash28 times higher in Panguanathan in Manaus Densities were remarkably con-stant in Manaus during the study period
Premetamorphic biology EggsmdashEgg clutches arelaid in water and consist of a gelatinous eggmass that primarily floats on or near the surfacebut often adheres to the inner wall of the tree-hole The darkly pigmented eggs are small(diam 158 6 006 mm n 5 6) and surroundedby gelatinous capsules (diam 428 6 021 mm
267SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
TABLE 2 SEASONAL ESTIMATES OF THE ABUNDANCE
AND DENSITY (LINEAR AND BY AREA) OF CALLING MALES
OF Phrynohyas resinifictrix IN PANGUANA AND MANAUS
Season
Abun-dance
ofcallingmales
Density of calling males
Linear(males
km)Area
(malesha)Area21
(hamale)
Panguana1988 7 48 015 68
Manaus1990199119931994
1095
191720
006005006
171188163
Fig 1 Number of intact eggs found in the guts oftadpoles of Phrynohyas resinifictrix through ontogeny(n 5 32 treehole 2) The dashed line represents anarbitrarily defined developmental threshold at stage32 after which eggs are efficiently used as food re-source Bubble size is proportional to sample size(varying from 1ndash6)
TABLE 3 DIET OF Phrynohyas resinifictrix LARVAE Occurrence expressed as Mean 6 SE
Stages n
Occurrence of food categories ()
AlgaePlant
fragmentsArthropodfragments Detritus Eggs
One treehole sampled over five months25ndash41 32 166 6 51 100 6 28 53 6 16 359 6 84 597 6 87
Four treeholes sampled at one time25252525
5443
60 6 25150 6 65300 6 41300 6 100
180 6 58275 6 103200 6 4133 6 33
00 6 0075 6 4875 6 7500 6 00
580 6 132975 6 25750 6 50730 6 22
600 6 18200 6 0000 6 00
300 6 100
Total44 175 6 39 114 6 23 46 6 13 446 6 67 523 6 72
n 5 6) At the moment of collection clutch sizevaried from 106ndash1540 eggs (436 6 234 eggs n5 6) with as many as six clutches laid in a singletreehole during a period of four months (De-cember 1990 to April 1991)
Larval dietmdashThe diet of P resinifictrix larvaeconsists largely of conspecific eggs and detritusless common items include algae plant frag-ments (mainly leaf fragments but also pollen)and arthropod fragments (insect mouthpartsheads and wing fragments lepidopteran scalesand mosquito larvae bristles Table 3)
Fifty-two percent of all guts examined (n 544) contained eggs in different phases of diges-tion (Table 3) Tadpoles from stages 32ndash42 wereable to swallow whole eggs with capsules andsometimes gelatin (Fig 1) whereas smaller tad-poles ingested egg fragments presumablyrasped with their keratinized mouthparts Wheneggs were available individuals ingested asmany as possible and their guts became greatlyextended The enlarged foregut contained upto 62 whole eggs (Fig 1) plus partially digested
eggs that could not be counted Most eggs ex-amined were fertilized as indicated by externalor histological observation of cell cleavage andembryo development (up to stage 16) Four ofsix clutches laid during a five-month period intreehole 2 at Manaus contained fertilized eggsthe two others could not be analyzed becauseno eggs remained in the gelatinous mass by thetime of observation
Detritus was the second most frequent fooditem consumed observed in 446 of micro-scopic fields when eggs were not available de-
268 COPEIA 2003 NO 2
tritus occurrence reached 975 (Table 3) Allother food items were found much less fre-quently Algae were never abundant althoughsporadically occurring in 175 of all micro-scopic fields or up to 30 in tadpoles in earlydevelopmental stages Plant fragments occurredin 114 of all microscopic fields or up to275 in tadpoles in early developmental stag-es Arthropod fragments were only present in46 of all microscopic fields examined (Table3)
DISCUSSION
Nonaquatic oviposition behavior in frogsevolved independently in many lineages andhas been interpreted as a strategy to avoid pre-dation competition or unfavorable abiotic con-ditions in the ancestral water body (Crump1982 Duellman and Trueb 1986) From anecological rather than evolutionary perspectivethese same processes might be currently oper-ating to maintain arboreal oviposition Magnus-son and Hero (1991) studied an anuran assem-blage at our Central Amazonian rain-forest fieldsite They found that the percentage of specieswith terrestrial eggs and aquatic larvae in a wa-ter body was positively correlated with an indexof egg predation pressure in that water body butonly weakly correlated with the number of spe-cies of potential competitors or abiotic condi-tions such as dissolved oxygen content per-centage of dry days and temperature These ob-servations led the authors to suggest that pre-dation is a major factor maintaining nonaquaticoviposition behavior in this contemporary an-uran assemblage
Oviposition in phytotelms might also bemaintained by the high predation pressure interrestrial water bodies Predation risk is pre-sumably lower in phytotelms because theheight small size concealment and often widespacing of these water bodies reduce predatoraccess to the eggs and tadpoles In addition un-favorable abiotic conditions in phytotelms suchas the potentially low dissolved oxygen levelsand short hydroperiod constrain the survival ofaquatic predators In this study few actual orpotential predators of eggs (wasps small dytis-cid beetles and heterospecific oophagic andcannibalistic tadpoles) tadpoles (spiders) andadults (a colubrid snake Philodryas viridissimus)have been observed inside treeholes inhabitedby P resinifictrix This contrasts strongly with theubiquity of insect and fish predators found inground water bodies at the study site (Magnus-son and Hero 1991)
In addition our observations suggest that
current interspecific competition among Ama-zonian frogs for phytotelms as breeding and de-velopmental sites is minimal and could be an-other important force maintaining ovipositionin these water bodies Only four of 130 lowlandAmazonian frog species breed in phytotelms(Hodl 1990) Moreover in Central Amazonia Presinifictrix is the only anuran to breed exclusive-ly in large water-filled treeholes whereas Osteo-cephalus oophagus breeds in a variety of phyto-telms such as bromeliads buriti (Mauritia fle-xuosa) palm leaf axils palm bracts and only oc-casionally in treeholes ( Jungfer and Schiesari1995 pers obs) Calling adults eggs or larvaeof P resinifictrix and O oophagus were neverfound in the extensively sampled ground waterbodies indicating specialization for a phytotel-mic breeding site Yet tadpoles of these specieswere never found coexisting in any of the phyto-telms sampled indicating strong habitat segre-gation Several species of Dendrobates have ter-restrial oviposition but transport tadpoles tophytotelms after hatching for instance we ob-served in a single occasion Dendrobates cf quin-quevittatus inside treeholes from which P resini-fictrix called More often however Dendrobatesspecies use small-sized ground phytotelms (egCaldwell and Araujo 1998) Overall this strongpattern of habitat segregation among phytotelmtypes in which more than one species are sel-dom found to coexist contrasts with ground wa-ter bodies where typically more than three andsometimes up to eight tadpole species coexist(Magnusson and Hero 1991)
Therefore several lines of evidence suggestthat both predation pressure and interspecificcompetition are lower in phytotelms than in ter-restrial water bodies and could contribute to themaintenance of the use of phytotelms as breed-ing and developmental sites for Amazonian an-urans in general and P resinifictrix in particularIn other systems however an unavailability ofterrestrial water bodies may act as a strong forcemaintaining phytotelm reproduction This ap-pears to be the case in Caribbean Islands suchas Jamaica where historical and geological fac-tors result in phytotelms being the predominantlentic water bodies (Thompson 1996)
Phytotelms are a harsh environment for thedevelopment of anuran embryos and larvaeConstrained volumes of noncirculating waterdominated by decaying processes sometimes re-sult in very low dissolved oxygen concentra-tions Assuming that treehole breeding is de-rived in the genus Phrynohyas early develop-ment of large inflated functional lungs in oth-er pond-dwelling species of the genus may beconsidered a useful morphophysiological trait
269SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
for the colonization of phytotelms by P resinific-trix Lungs are also used as hydrostatic organskeeping tadpoles close to the water surface andmay reduce the energy expenditure involved inrepeatedly swimming to the surface to bob air(Feder 1984 Feder and Moran 1985 Lannooet al 1987)
An important biotic characteristic of phyto-telms is the unpredictability and low availabilityof high quality food In most tadpole species forwhich dietary data are available algae of pe-riphyton and phytoplankton constitute majorfood resources in addition to low-quality detri-tus (eg Altig and Johnston 1989 Kupferberget al 1994) Narrow treehole openings oftencovered by overlying canopy may result in anearly aphotic habitat in which primary pro-ductivity is low and therefore likely insufficientto sustain tadpole growth
Detritus derived from the decomposition offallen leaf litter and treehole walls constitutedthe basis for the speciesrsquo diet when eggs werenot available The nutritive value of detritus islow (Kupferberg et al 1994) and several spe-cies of generalized pond tadpoles fail to reachmetamorphosis when restricted to this food type(Werner and Glennemeier 1999 LS unpubl)In this context oophagy may be of major im-portance for the success of individual larvae inreaching metamorphosis because eggs providea highly nutritious and energetic food source(Crump 1992) However detritivory could alsobe of crucial importance especially for smallertadpoles Prior to stage 32 tadpoles are gape-limited and therefore inefficient egg feedersAfter this stage tadpoles are capable of swallow-ing large quantities of whole eggs leading toaccelerated growth and development
Opportunistic oophagy is common in anuranlarvae (Magnusson and Hero 1991 Crump1992 Petranka and Kennedy 1999) In severalphytotelm-developing anurans however oop-hagy results from complex parental behavior inwhich the mother periodically returns to thephytotelm to lay fertilized or unfertilized eggsfor nutrition of its progeny (Lannoo et al 1987Weygoldt 1987 Crump 1995) For some ofthese species it was demonstrated that oophagyis obligate that is tadpoles fail to reach meta-morphosis if eggs are not provided by the moth-er (Weygoldt 1987 Thompson 1996 Jungferand Weygoldt 1999) In P resinifictrix howeverthere are several observations suggesting thatoophagy is opportunistic cannibalism ratherthan parental care in the form of periodic eggdeposition for obligate tadpole nutrition Firstas noted by Jungfer and Proy (1998) P resinific-trix egg clutches are large (maximum clutch size
in the field 1540 eggs this study in terraria4336 eggs Jungfer and Proy 1998) comparedto phytotelm-breeding species that intentionallyprovide eggs to their offspring For instanceconsidering only hylids on average 158 eggs arelaid by Anotheca spinosa ( Jungfer 1996) 258 byOsteocephalus oophagus ( Jungfer and Weygoldt1999) 276 per bromeliad by Osteopilus brunneus(Lannoo et al 1987) and 13 by Phyllodytes lu-teolus (Giaretta 1996) Second as a conse-quence of the large clutch size interclutch in-terval is long in P resinifictrix in a regularly sam-pled treehole six clutches were deposited overa period of four months (around one clutch ev-ery 20 days) which coincides with the inter-clutch interval of pairs in captivity (20 daysrange 7ndash37 n 5 29 Jungfer and Proy 1998)This long interclutch interval contrasts stronglywith the interclutch interval of species that areknown to be obligate oophagic For instanceeggs are deposited every 49 days in A spinosa( Jungfer 1996) 51 days in O oophagus ( Jung-fer and Weygoldt 1999) and 44 days in Obrunneus (Thompson 1996) Jungfer and Proy(1998) found that tadpoles of P resinifictrixstarve if dependent only on the motherrsquos infre-quent oviposition as the food source but thatmetamorphic success in an experimental con-tainer was high if in addition to leaf-litter eggswere added every 17 days Third an opportu-nistic oophagic nutrition is suggested by our ob-servation that eggs are fertilized unlike some ofthe more lsquolsquoadvancedrsquorsquo forms of anuran parentalcare through laying of unfertilized trophic eggs(Weygoldt 1987) although obligate oophagyhas been documented for species laying fertil-ized eggs (Thompson 1996) Fourth althoughextensive reduction of oral structures character-izes several of the obligate oophagic phytotelm-developing species (Lannoo et al 1987) P re-sinifictrix retains typical generalized pond-typelarval morphology with only moderate structur-al reduction when compared to its pond breed-ing congeners (Schiesari et al 1996) Experi-mental evidence is necessary to unambiguouslydemonstrate that oophagy is not obligate in Presinifictrix however the arguments above pro-vide indirect support for the hypothesis thatconsumption of eggs is important but may notbe obligate for metamorphic success
We suggest that among the premetamorphicstages of P resinifictrix life cycle cannibalism andintraspecific competition are the major forcesacting on the survival of eggs and larvae re-spectively Evidence for this hypothesis are thelow nutritive value of food other than eggs andor high unpredictability of food in the treeholethe voracious oophagic habits of the tadpole
270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo
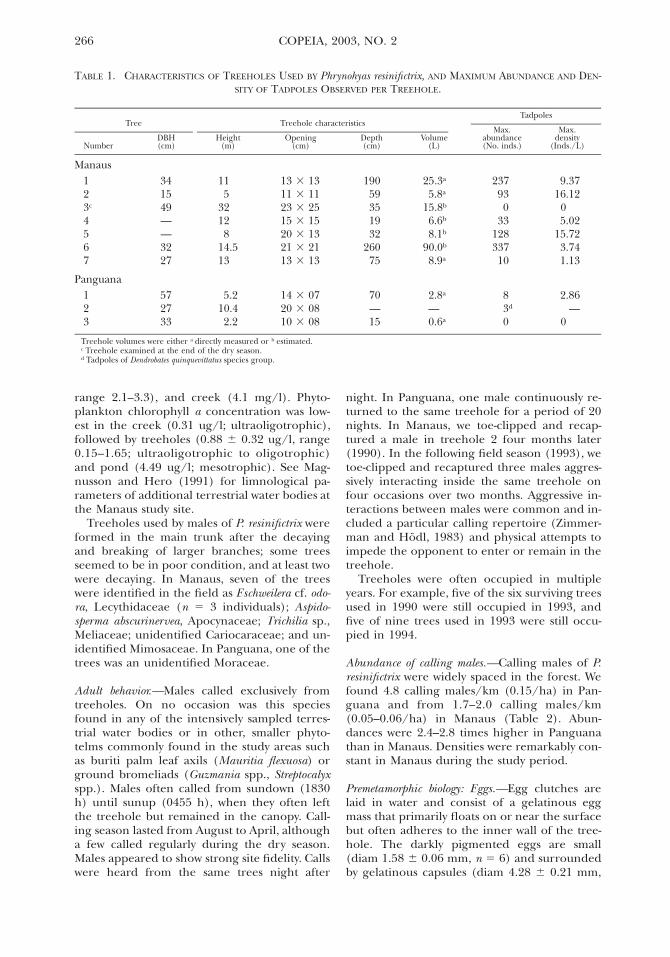
266 COPEIA 2003 NO 2
TABLE 1 CHARACTERISTICS OF TREEHOLES USED BY Phrynohyas resinifictrix AND MAXIMUM ABUNDANCE AND DEN-SITY OF TADPOLES OBSERVED PER TREEHOLE
Tree
NumberDBH(cm)
Treehole characteristics
Height(m)
Opening(cm)
Depth(cm)
Volume(L)
Tadpoles
Maxabundance(No inds)
Maxdensity
(IndsL)
Manaus123c
456
341549mdashmdash32
115
32128
145
13 3 1311 3 1123 3 2515 3 1520 3 1321 3 21
19059351932
260
253a
58a
158b
66b
81b
900b
237930
33128337
9371612
0502
1572374
7 27 13 13 3 13 75 89a 10 113
Panguana123
572733
5210422
14 3 0720 3 0810 3 08
70mdash15
28a
mdash06a
83d
0
286mdash
0
Treehole volumes were either a directly measured or b estimatedc Treehole examined at the end of the dry seasond Tadpoles of Dendrobates quinquevittatus species group
range 21ndash33) and creek (41 mgl) Phyto-plankton chlorophyll a concentration was low-est in the creek (031 ugl ultraoligotrophic)followed by treeholes (088 6 032 ugl range015ndash165 ultraoligotrophic to oligotrophic)and pond (449 ugl mesotrophic) See Mag-nusson and Hero (1991) for limnological pa-rameters of additional terrestrial water bodies atthe Manaus study site
Treeholes used by males of P resinifictrix wereformed in the main trunk after the decayingand breaking of larger branches some treesseemed to be in poor condition and at least twowere decaying In Manaus seven of the treeswere identified in the field as Eschweilera cf odo-ra Lecythidaceae (n 5 3 individuals) Aspido-sperma abscurinervea Apocynaceae Trichilia spMeliaceae unidentified Cariocaraceae and un-identified Mimosaceae In Panguana one of thetrees was an unidentified Moraceae
Adult behaviormdashMales called exclusively fromtreeholes On no occasion was this speciesfound in any of the intensively sampled terres-trial water bodies or in other smaller phyto-telms commonly found in the study areas suchas buriti palm leaf axils (Mauritia flexuosa) orground bromeliads (Guzmania spp Streptocalyxspp) Males often called from sundown (1830h) until sunup (0455 h) when they often leftthe treehole but remained in the canopy Call-ing season lasted from August to April althougha few called regularly during the dry seasonMales appeared to show strong site fidelity Callswere heard from the same trees night after
night In Panguana one male continuously re-turned to the same treehole for a period of 20nights In Manaus we toe-clipped and recap-tured a male in treehole 2 four months later(1990) In the following field season (1993) wetoe-clipped and recaptured three males aggres-sively interacting inside the same treehole onfour occasions over two months Aggressive in-teractions between males were common and in-cluded a particular calling repertoire (Zimmer-man and Hodl 1983) and physical attempts toimpede the opponent to enter or remain in thetreehole
Treeholes were often occupied in multipleyears For example five of the six surviving treesused in 1990 were still occupied in 1993 andfive of nine trees used in 1993 were still occu-pied in 1994
Abundance of calling malesmdashCalling males of Presinifictrix were widely spaced in the forest Wefound 48 calling maleskm (015ha) in Pan-guana and from 17ndash20 calling maleskm(005ndash006ha) in Manaus (Table 2) Abun-dances were 24ndash28 times higher in Panguanathan in Manaus Densities were remarkably con-stant in Manaus during the study period
Premetamorphic biology EggsmdashEgg clutches arelaid in water and consist of a gelatinous eggmass that primarily floats on or near the surfacebut often adheres to the inner wall of the tree-hole The darkly pigmented eggs are small(diam 158 6 006 mm n 5 6) and surroundedby gelatinous capsules (diam 428 6 021 mm
267SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
TABLE 2 SEASONAL ESTIMATES OF THE ABUNDANCE
AND DENSITY (LINEAR AND BY AREA) OF CALLING MALES
OF Phrynohyas resinifictrix IN PANGUANA AND MANAUS
Season
Abun-dance
ofcallingmales
Density of calling males
Linear(males
km)Area
(malesha)Area21
(hamale)
Panguana1988 7 48 015 68
Manaus1990199119931994
1095
191720
006005006
171188163
Fig 1 Number of intact eggs found in the guts oftadpoles of Phrynohyas resinifictrix through ontogeny(n 5 32 treehole 2) The dashed line represents anarbitrarily defined developmental threshold at stage32 after which eggs are efficiently used as food re-source Bubble size is proportional to sample size(varying from 1ndash6)
TABLE 3 DIET OF Phrynohyas resinifictrix LARVAE Occurrence expressed as Mean 6 SE
Stages n
Occurrence of food categories ()
AlgaePlant
fragmentsArthropodfragments Detritus Eggs
One treehole sampled over five months25ndash41 32 166 6 51 100 6 28 53 6 16 359 6 84 597 6 87
Four treeholes sampled at one time25252525
5443
60 6 25150 6 65300 6 41300 6 100
180 6 58275 6 103200 6 4133 6 33
00 6 0075 6 4875 6 7500 6 00
580 6 132975 6 25750 6 50730 6 22
600 6 18200 6 0000 6 00
300 6 100
Total44 175 6 39 114 6 23 46 6 13 446 6 67 523 6 72
n 5 6) At the moment of collection clutch sizevaried from 106ndash1540 eggs (436 6 234 eggs n5 6) with as many as six clutches laid in a singletreehole during a period of four months (De-cember 1990 to April 1991)
Larval dietmdashThe diet of P resinifictrix larvaeconsists largely of conspecific eggs and detritusless common items include algae plant frag-ments (mainly leaf fragments but also pollen)and arthropod fragments (insect mouthpartsheads and wing fragments lepidopteran scalesand mosquito larvae bristles Table 3)
Fifty-two percent of all guts examined (n 544) contained eggs in different phases of diges-tion (Table 3) Tadpoles from stages 32ndash42 wereable to swallow whole eggs with capsules andsometimes gelatin (Fig 1) whereas smaller tad-poles ingested egg fragments presumablyrasped with their keratinized mouthparts Wheneggs were available individuals ingested asmany as possible and their guts became greatlyextended The enlarged foregut contained upto 62 whole eggs (Fig 1) plus partially digested
eggs that could not be counted Most eggs ex-amined were fertilized as indicated by externalor histological observation of cell cleavage andembryo development (up to stage 16) Four ofsix clutches laid during a five-month period intreehole 2 at Manaus contained fertilized eggsthe two others could not be analyzed becauseno eggs remained in the gelatinous mass by thetime of observation
Detritus was the second most frequent fooditem consumed observed in 446 of micro-scopic fields when eggs were not available de-
268 COPEIA 2003 NO 2
tritus occurrence reached 975 (Table 3) Allother food items were found much less fre-quently Algae were never abundant althoughsporadically occurring in 175 of all micro-scopic fields or up to 30 in tadpoles in earlydevelopmental stages Plant fragments occurredin 114 of all microscopic fields or up to275 in tadpoles in early developmental stag-es Arthropod fragments were only present in46 of all microscopic fields examined (Table3)
DISCUSSION
Nonaquatic oviposition behavior in frogsevolved independently in many lineages andhas been interpreted as a strategy to avoid pre-dation competition or unfavorable abiotic con-ditions in the ancestral water body (Crump1982 Duellman and Trueb 1986) From anecological rather than evolutionary perspectivethese same processes might be currently oper-ating to maintain arboreal oviposition Magnus-son and Hero (1991) studied an anuran assem-blage at our Central Amazonian rain-forest fieldsite They found that the percentage of specieswith terrestrial eggs and aquatic larvae in a wa-ter body was positively correlated with an indexof egg predation pressure in that water body butonly weakly correlated with the number of spe-cies of potential competitors or abiotic condi-tions such as dissolved oxygen content per-centage of dry days and temperature These ob-servations led the authors to suggest that pre-dation is a major factor maintaining nonaquaticoviposition behavior in this contemporary an-uran assemblage
Oviposition in phytotelms might also bemaintained by the high predation pressure interrestrial water bodies Predation risk is pre-sumably lower in phytotelms because theheight small size concealment and often widespacing of these water bodies reduce predatoraccess to the eggs and tadpoles In addition un-favorable abiotic conditions in phytotelms suchas the potentially low dissolved oxygen levelsand short hydroperiod constrain the survival ofaquatic predators In this study few actual orpotential predators of eggs (wasps small dytis-cid beetles and heterospecific oophagic andcannibalistic tadpoles) tadpoles (spiders) andadults (a colubrid snake Philodryas viridissimus)have been observed inside treeholes inhabitedby P resinifictrix This contrasts strongly with theubiquity of insect and fish predators found inground water bodies at the study site (Magnus-son and Hero 1991)
In addition our observations suggest that
current interspecific competition among Ama-zonian frogs for phytotelms as breeding and de-velopmental sites is minimal and could be an-other important force maintaining ovipositionin these water bodies Only four of 130 lowlandAmazonian frog species breed in phytotelms(Hodl 1990) Moreover in Central Amazonia Presinifictrix is the only anuran to breed exclusive-ly in large water-filled treeholes whereas Osteo-cephalus oophagus breeds in a variety of phyto-telms such as bromeliads buriti (Mauritia fle-xuosa) palm leaf axils palm bracts and only oc-casionally in treeholes ( Jungfer and Schiesari1995 pers obs) Calling adults eggs or larvaeof P resinifictrix and O oophagus were neverfound in the extensively sampled ground waterbodies indicating specialization for a phytotel-mic breeding site Yet tadpoles of these specieswere never found coexisting in any of the phyto-telms sampled indicating strong habitat segre-gation Several species of Dendrobates have ter-restrial oviposition but transport tadpoles tophytotelms after hatching for instance we ob-served in a single occasion Dendrobates cf quin-quevittatus inside treeholes from which P resini-fictrix called More often however Dendrobatesspecies use small-sized ground phytotelms (egCaldwell and Araujo 1998) Overall this strongpattern of habitat segregation among phytotelmtypes in which more than one species are sel-dom found to coexist contrasts with ground wa-ter bodies where typically more than three andsometimes up to eight tadpole species coexist(Magnusson and Hero 1991)
Therefore several lines of evidence suggestthat both predation pressure and interspecificcompetition are lower in phytotelms than in ter-restrial water bodies and could contribute to themaintenance of the use of phytotelms as breed-ing and developmental sites for Amazonian an-urans in general and P resinifictrix in particularIn other systems however an unavailability ofterrestrial water bodies may act as a strong forcemaintaining phytotelm reproduction This ap-pears to be the case in Caribbean Islands suchas Jamaica where historical and geological fac-tors result in phytotelms being the predominantlentic water bodies (Thompson 1996)
Phytotelms are a harsh environment for thedevelopment of anuran embryos and larvaeConstrained volumes of noncirculating waterdominated by decaying processes sometimes re-sult in very low dissolved oxygen concentra-tions Assuming that treehole breeding is de-rived in the genus Phrynohyas early develop-ment of large inflated functional lungs in oth-er pond-dwelling species of the genus may beconsidered a useful morphophysiological trait
269SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
for the colonization of phytotelms by P resinific-trix Lungs are also used as hydrostatic organskeeping tadpoles close to the water surface andmay reduce the energy expenditure involved inrepeatedly swimming to the surface to bob air(Feder 1984 Feder and Moran 1985 Lannooet al 1987)
An important biotic characteristic of phyto-telms is the unpredictability and low availabilityof high quality food In most tadpole species forwhich dietary data are available algae of pe-riphyton and phytoplankton constitute majorfood resources in addition to low-quality detri-tus (eg Altig and Johnston 1989 Kupferberget al 1994) Narrow treehole openings oftencovered by overlying canopy may result in anearly aphotic habitat in which primary pro-ductivity is low and therefore likely insufficientto sustain tadpole growth
Detritus derived from the decomposition offallen leaf litter and treehole walls constitutedthe basis for the speciesrsquo diet when eggs werenot available The nutritive value of detritus islow (Kupferberg et al 1994) and several spe-cies of generalized pond tadpoles fail to reachmetamorphosis when restricted to this food type(Werner and Glennemeier 1999 LS unpubl)In this context oophagy may be of major im-portance for the success of individual larvae inreaching metamorphosis because eggs providea highly nutritious and energetic food source(Crump 1992) However detritivory could alsobe of crucial importance especially for smallertadpoles Prior to stage 32 tadpoles are gape-limited and therefore inefficient egg feedersAfter this stage tadpoles are capable of swallow-ing large quantities of whole eggs leading toaccelerated growth and development
Opportunistic oophagy is common in anuranlarvae (Magnusson and Hero 1991 Crump1992 Petranka and Kennedy 1999) In severalphytotelm-developing anurans however oop-hagy results from complex parental behavior inwhich the mother periodically returns to thephytotelm to lay fertilized or unfertilized eggsfor nutrition of its progeny (Lannoo et al 1987Weygoldt 1987 Crump 1995) For some ofthese species it was demonstrated that oophagyis obligate that is tadpoles fail to reach meta-morphosis if eggs are not provided by the moth-er (Weygoldt 1987 Thompson 1996 Jungferand Weygoldt 1999) In P resinifictrix howeverthere are several observations suggesting thatoophagy is opportunistic cannibalism ratherthan parental care in the form of periodic eggdeposition for obligate tadpole nutrition Firstas noted by Jungfer and Proy (1998) P resinific-trix egg clutches are large (maximum clutch size
in the field 1540 eggs this study in terraria4336 eggs Jungfer and Proy 1998) comparedto phytotelm-breeding species that intentionallyprovide eggs to their offspring For instanceconsidering only hylids on average 158 eggs arelaid by Anotheca spinosa ( Jungfer 1996) 258 byOsteocephalus oophagus ( Jungfer and Weygoldt1999) 276 per bromeliad by Osteopilus brunneus(Lannoo et al 1987) and 13 by Phyllodytes lu-teolus (Giaretta 1996) Second as a conse-quence of the large clutch size interclutch in-terval is long in P resinifictrix in a regularly sam-pled treehole six clutches were deposited overa period of four months (around one clutch ev-ery 20 days) which coincides with the inter-clutch interval of pairs in captivity (20 daysrange 7ndash37 n 5 29 Jungfer and Proy 1998)This long interclutch interval contrasts stronglywith the interclutch interval of species that areknown to be obligate oophagic For instanceeggs are deposited every 49 days in A spinosa( Jungfer 1996) 51 days in O oophagus ( Jung-fer and Weygoldt 1999) and 44 days in Obrunneus (Thompson 1996) Jungfer and Proy(1998) found that tadpoles of P resinifictrixstarve if dependent only on the motherrsquos infre-quent oviposition as the food source but thatmetamorphic success in an experimental con-tainer was high if in addition to leaf-litter eggswere added every 17 days Third an opportu-nistic oophagic nutrition is suggested by our ob-servation that eggs are fertilized unlike some ofthe more lsquolsquoadvancedrsquorsquo forms of anuran parentalcare through laying of unfertilized trophic eggs(Weygoldt 1987) although obligate oophagyhas been documented for species laying fertil-ized eggs (Thompson 1996) Fourth althoughextensive reduction of oral structures character-izes several of the obligate oophagic phytotelm-developing species (Lannoo et al 1987) P re-sinifictrix retains typical generalized pond-typelarval morphology with only moderate structur-al reduction when compared to its pond breed-ing congeners (Schiesari et al 1996) Experi-mental evidence is necessary to unambiguouslydemonstrate that oophagy is not obligate in Presinifictrix however the arguments above pro-vide indirect support for the hypothesis thatconsumption of eggs is important but may notbe obligate for metamorphic success
We suggest that among the premetamorphicstages of P resinifictrix life cycle cannibalism andintraspecific competition are the major forcesacting on the survival of eggs and larvae re-spectively Evidence for this hypothesis are thelow nutritive value of food other than eggs andor high unpredictability of food in the treeholethe voracious oophagic habits of the tadpole
270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo
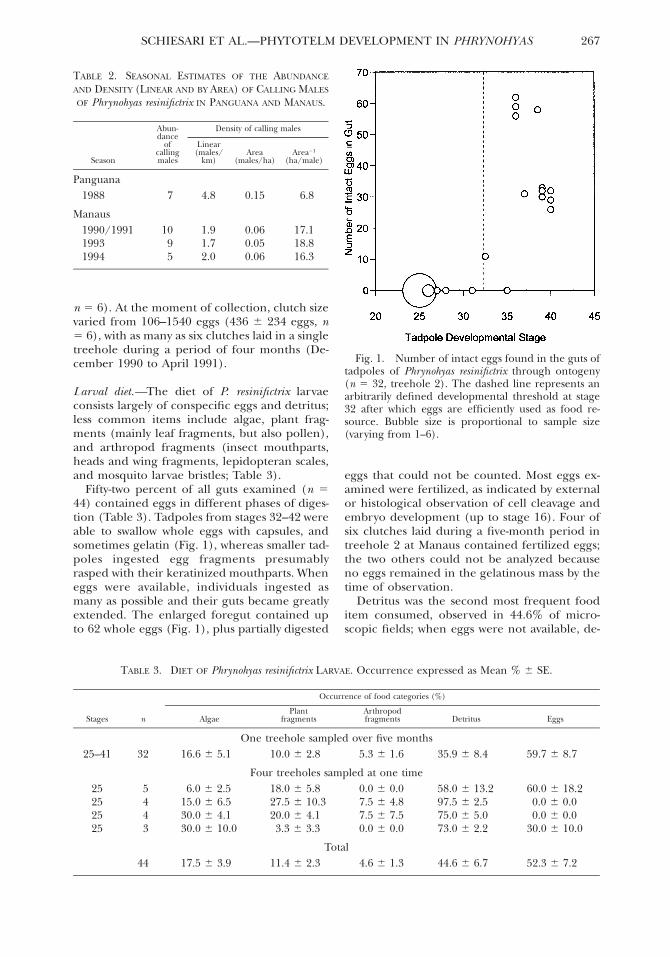
267SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
TABLE 2 SEASONAL ESTIMATES OF THE ABUNDANCE
AND DENSITY (LINEAR AND BY AREA) OF CALLING MALES
OF Phrynohyas resinifictrix IN PANGUANA AND MANAUS
Season
Abun-dance
ofcallingmales
Density of calling males
Linear(males
km)Area
(malesha)Area21
(hamale)
Panguana1988 7 48 015 68
Manaus1990199119931994
1095
191720
006005006
171188163
Fig 1 Number of intact eggs found in the guts oftadpoles of Phrynohyas resinifictrix through ontogeny(n 5 32 treehole 2) The dashed line represents anarbitrarily defined developmental threshold at stage32 after which eggs are efficiently used as food re-source Bubble size is proportional to sample size(varying from 1ndash6)
TABLE 3 DIET OF Phrynohyas resinifictrix LARVAE Occurrence expressed as Mean 6 SE
Stages n
Occurrence of food categories ()
AlgaePlant
fragmentsArthropodfragments Detritus Eggs
One treehole sampled over five months25ndash41 32 166 6 51 100 6 28 53 6 16 359 6 84 597 6 87
Four treeholes sampled at one time25252525
5443
60 6 25150 6 65300 6 41300 6 100
180 6 58275 6 103200 6 4133 6 33
00 6 0075 6 4875 6 7500 6 00
580 6 132975 6 25750 6 50730 6 22
600 6 18200 6 0000 6 00
300 6 100
Total44 175 6 39 114 6 23 46 6 13 446 6 67 523 6 72
n 5 6) At the moment of collection clutch sizevaried from 106ndash1540 eggs (436 6 234 eggs n5 6) with as many as six clutches laid in a singletreehole during a period of four months (De-cember 1990 to April 1991)
Larval dietmdashThe diet of P resinifictrix larvaeconsists largely of conspecific eggs and detritusless common items include algae plant frag-ments (mainly leaf fragments but also pollen)and arthropod fragments (insect mouthpartsheads and wing fragments lepidopteran scalesand mosquito larvae bristles Table 3)
Fifty-two percent of all guts examined (n 544) contained eggs in different phases of diges-tion (Table 3) Tadpoles from stages 32ndash42 wereable to swallow whole eggs with capsules andsometimes gelatin (Fig 1) whereas smaller tad-poles ingested egg fragments presumablyrasped with their keratinized mouthparts Wheneggs were available individuals ingested asmany as possible and their guts became greatlyextended The enlarged foregut contained upto 62 whole eggs (Fig 1) plus partially digested
eggs that could not be counted Most eggs ex-amined were fertilized as indicated by externalor histological observation of cell cleavage andembryo development (up to stage 16) Four ofsix clutches laid during a five-month period intreehole 2 at Manaus contained fertilized eggsthe two others could not be analyzed becauseno eggs remained in the gelatinous mass by thetime of observation
Detritus was the second most frequent fooditem consumed observed in 446 of micro-scopic fields when eggs were not available de-
268 COPEIA 2003 NO 2
tritus occurrence reached 975 (Table 3) Allother food items were found much less fre-quently Algae were never abundant althoughsporadically occurring in 175 of all micro-scopic fields or up to 30 in tadpoles in earlydevelopmental stages Plant fragments occurredin 114 of all microscopic fields or up to275 in tadpoles in early developmental stag-es Arthropod fragments were only present in46 of all microscopic fields examined (Table3)
DISCUSSION
Nonaquatic oviposition behavior in frogsevolved independently in many lineages andhas been interpreted as a strategy to avoid pre-dation competition or unfavorable abiotic con-ditions in the ancestral water body (Crump1982 Duellman and Trueb 1986) From anecological rather than evolutionary perspectivethese same processes might be currently oper-ating to maintain arboreal oviposition Magnus-son and Hero (1991) studied an anuran assem-blage at our Central Amazonian rain-forest fieldsite They found that the percentage of specieswith terrestrial eggs and aquatic larvae in a wa-ter body was positively correlated with an indexof egg predation pressure in that water body butonly weakly correlated with the number of spe-cies of potential competitors or abiotic condi-tions such as dissolved oxygen content per-centage of dry days and temperature These ob-servations led the authors to suggest that pre-dation is a major factor maintaining nonaquaticoviposition behavior in this contemporary an-uran assemblage
Oviposition in phytotelms might also bemaintained by the high predation pressure interrestrial water bodies Predation risk is pre-sumably lower in phytotelms because theheight small size concealment and often widespacing of these water bodies reduce predatoraccess to the eggs and tadpoles In addition un-favorable abiotic conditions in phytotelms suchas the potentially low dissolved oxygen levelsand short hydroperiod constrain the survival ofaquatic predators In this study few actual orpotential predators of eggs (wasps small dytis-cid beetles and heterospecific oophagic andcannibalistic tadpoles) tadpoles (spiders) andadults (a colubrid snake Philodryas viridissimus)have been observed inside treeholes inhabitedby P resinifictrix This contrasts strongly with theubiquity of insect and fish predators found inground water bodies at the study site (Magnus-son and Hero 1991)
In addition our observations suggest that
current interspecific competition among Ama-zonian frogs for phytotelms as breeding and de-velopmental sites is minimal and could be an-other important force maintaining ovipositionin these water bodies Only four of 130 lowlandAmazonian frog species breed in phytotelms(Hodl 1990) Moreover in Central Amazonia Presinifictrix is the only anuran to breed exclusive-ly in large water-filled treeholes whereas Osteo-cephalus oophagus breeds in a variety of phyto-telms such as bromeliads buriti (Mauritia fle-xuosa) palm leaf axils palm bracts and only oc-casionally in treeholes ( Jungfer and Schiesari1995 pers obs) Calling adults eggs or larvaeof P resinifictrix and O oophagus were neverfound in the extensively sampled ground waterbodies indicating specialization for a phytotel-mic breeding site Yet tadpoles of these specieswere never found coexisting in any of the phyto-telms sampled indicating strong habitat segre-gation Several species of Dendrobates have ter-restrial oviposition but transport tadpoles tophytotelms after hatching for instance we ob-served in a single occasion Dendrobates cf quin-quevittatus inside treeholes from which P resini-fictrix called More often however Dendrobatesspecies use small-sized ground phytotelms (egCaldwell and Araujo 1998) Overall this strongpattern of habitat segregation among phytotelmtypes in which more than one species are sel-dom found to coexist contrasts with ground wa-ter bodies where typically more than three andsometimes up to eight tadpole species coexist(Magnusson and Hero 1991)
Therefore several lines of evidence suggestthat both predation pressure and interspecificcompetition are lower in phytotelms than in ter-restrial water bodies and could contribute to themaintenance of the use of phytotelms as breed-ing and developmental sites for Amazonian an-urans in general and P resinifictrix in particularIn other systems however an unavailability ofterrestrial water bodies may act as a strong forcemaintaining phytotelm reproduction This ap-pears to be the case in Caribbean Islands suchas Jamaica where historical and geological fac-tors result in phytotelms being the predominantlentic water bodies (Thompson 1996)
Phytotelms are a harsh environment for thedevelopment of anuran embryos and larvaeConstrained volumes of noncirculating waterdominated by decaying processes sometimes re-sult in very low dissolved oxygen concentra-tions Assuming that treehole breeding is de-rived in the genus Phrynohyas early develop-ment of large inflated functional lungs in oth-er pond-dwelling species of the genus may beconsidered a useful morphophysiological trait
269SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
for the colonization of phytotelms by P resinific-trix Lungs are also used as hydrostatic organskeeping tadpoles close to the water surface andmay reduce the energy expenditure involved inrepeatedly swimming to the surface to bob air(Feder 1984 Feder and Moran 1985 Lannooet al 1987)
An important biotic characteristic of phyto-telms is the unpredictability and low availabilityof high quality food In most tadpole species forwhich dietary data are available algae of pe-riphyton and phytoplankton constitute majorfood resources in addition to low-quality detri-tus (eg Altig and Johnston 1989 Kupferberget al 1994) Narrow treehole openings oftencovered by overlying canopy may result in anearly aphotic habitat in which primary pro-ductivity is low and therefore likely insufficientto sustain tadpole growth
Detritus derived from the decomposition offallen leaf litter and treehole walls constitutedthe basis for the speciesrsquo diet when eggs werenot available The nutritive value of detritus islow (Kupferberg et al 1994) and several spe-cies of generalized pond tadpoles fail to reachmetamorphosis when restricted to this food type(Werner and Glennemeier 1999 LS unpubl)In this context oophagy may be of major im-portance for the success of individual larvae inreaching metamorphosis because eggs providea highly nutritious and energetic food source(Crump 1992) However detritivory could alsobe of crucial importance especially for smallertadpoles Prior to stage 32 tadpoles are gape-limited and therefore inefficient egg feedersAfter this stage tadpoles are capable of swallow-ing large quantities of whole eggs leading toaccelerated growth and development
Opportunistic oophagy is common in anuranlarvae (Magnusson and Hero 1991 Crump1992 Petranka and Kennedy 1999) In severalphytotelm-developing anurans however oop-hagy results from complex parental behavior inwhich the mother periodically returns to thephytotelm to lay fertilized or unfertilized eggsfor nutrition of its progeny (Lannoo et al 1987Weygoldt 1987 Crump 1995) For some ofthese species it was demonstrated that oophagyis obligate that is tadpoles fail to reach meta-morphosis if eggs are not provided by the moth-er (Weygoldt 1987 Thompson 1996 Jungferand Weygoldt 1999) In P resinifictrix howeverthere are several observations suggesting thatoophagy is opportunistic cannibalism ratherthan parental care in the form of periodic eggdeposition for obligate tadpole nutrition Firstas noted by Jungfer and Proy (1998) P resinific-trix egg clutches are large (maximum clutch size
in the field 1540 eggs this study in terraria4336 eggs Jungfer and Proy 1998) comparedto phytotelm-breeding species that intentionallyprovide eggs to their offspring For instanceconsidering only hylids on average 158 eggs arelaid by Anotheca spinosa ( Jungfer 1996) 258 byOsteocephalus oophagus ( Jungfer and Weygoldt1999) 276 per bromeliad by Osteopilus brunneus(Lannoo et al 1987) and 13 by Phyllodytes lu-teolus (Giaretta 1996) Second as a conse-quence of the large clutch size interclutch in-terval is long in P resinifictrix in a regularly sam-pled treehole six clutches were deposited overa period of four months (around one clutch ev-ery 20 days) which coincides with the inter-clutch interval of pairs in captivity (20 daysrange 7ndash37 n 5 29 Jungfer and Proy 1998)This long interclutch interval contrasts stronglywith the interclutch interval of species that areknown to be obligate oophagic For instanceeggs are deposited every 49 days in A spinosa( Jungfer 1996) 51 days in O oophagus ( Jung-fer and Weygoldt 1999) and 44 days in Obrunneus (Thompson 1996) Jungfer and Proy(1998) found that tadpoles of P resinifictrixstarve if dependent only on the motherrsquos infre-quent oviposition as the food source but thatmetamorphic success in an experimental con-tainer was high if in addition to leaf-litter eggswere added every 17 days Third an opportu-nistic oophagic nutrition is suggested by our ob-servation that eggs are fertilized unlike some ofthe more lsquolsquoadvancedrsquorsquo forms of anuran parentalcare through laying of unfertilized trophic eggs(Weygoldt 1987) although obligate oophagyhas been documented for species laying fertil-ized eggs (Thompson 1996) Fourth althoughextensive reduction of oral structures character-izes several of the obligate oophagic phytotelm-developing species (Lannoo et al 1987) P re-sinifictrix retains typical generalized pond-typelarval morphology with only moderate structur-al reduction when compared to its pond breed-ing congeners (Schiesari et al 1996) Experi-mental evidence is necessary to unambiguouslydemonstrate that oophagy is not obligate in Presinifictrix however the arguments above pro-vide indirect support for the hypothesis thatconsumption of eggs is important but may notbe obligate for metamorphic success
We suggest that among the premetamorphicstages of P resinifictrix life cycle cannibalism andintraspecific competition are the major forcesacting on the survival of eggs and larvae re-spectively Evidence for this hypothesis are thelow nutritive value of food other than eggs andor high unpredictability of food in the treeholethe voracious oophagic habits of the tadpole
270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo

268 COPEIA 2003 NO 2
tritus occurrence reached 975 (Table 3) Allother food items were found much less fre-quently Algae were never abundant althoughsporadically occurring in 175 of all micro-scopic fields or up to 30 in tadpoles in earlydevelopmental stages Plant fragments occurredin 114 of all microscopic fields or up to275 in tadpoles in early developmental stag-es Arthropod fragments were only present in46 of all microscopic fields examined (Table3)
DISCUSSION
Nonaquatic oviposition behavior in frogsevolved independently in many lineages andhas been interpreted as a strategy to avoid pre-dation competition or unfavorable abiotic con-ditions in the ancestral water body (Crump1982 Duellman and Trueb 1986) From anecological rather than evolutionary perspectivethese same processes might be currently oper-ating to maintain arboreal oviposition Magnus-son and Hero (1991) studied an anuran assem-blage at our Central Amazonian rain-forest fieldsite They found that the percentage of specieswith terrestrial eggs and aquatic larvae in a wa-ter body was positively correlated with an indexof egg predation pressure in that water body butonly weakly correlated with the number of spe-cies of potential competitors or abiotic condi-tions such as dissolved oxygen content per-centage of dry days and temperature These ob-servations led the authors to suggest that pre-dation is a major factor maintaining nonaquaticoviposition behavior in this contemporary an-uran assemblage
Oviposition in phytotelms might also bemaintained by the high predation pressure interrestrial water bodies Predation risk is pre-sumably lower in phytotelms because theheight small size concealment and often widespacing of these water bodies reduce predatoraccess to the eggs and tadpoles In addition un-favorable abiotic conditions in phytotelms suchas the potentially low dissolved oxygen levelsand short hydroperiod constrain the survival ofaquatic predators In this study few actual orpotential predators of eggs (wasps small dytis-cid beetles and heterospecific oophagic andcannibalistic tadpoles) tadpoles (spiders) andadults (a colubrid snake Philodryas viridissimus)have been observed inside treeholes inhabitedby P resinifictrix This contrasts strongly with theubiquity of insect and fish predators found inground water bodies at the study site (Magnus-son and Hero 1991)
In addition our observations suggest that
current interspecific competition among Ama-zonian frogs for phytotelms as breeding and de-velopmental sites is minimal and could be an-other important force maintaining ovipositionin these water bodies Only four of 130 lowlandAmazonian frog species breed in phytotelms(Hodl 1990) Moreover in Central Amazonia Presinifictrix is the only anuran to breed exclusive-ly in large water-filled treeholes whereas Osteo-cephalus oophagus breeds in a variety of phyto-telms such as bromeliads buriti (Mauritia fle-xuosa) palm leaf axils palm bracts and only oc-casionally in treeholes ( Jungfer and Schiesari1995 pers obs) Calling adults eggs or larvaeof P resinifictrix and O oophagus were neverfound in the extensively sampled ground waterbodies indicating specialization for a phytotel-mic breeding site Yet tadpoles of these specieswere never found coexisting in any of the phyto-telms sampled indicating strong habitat segre-gation Several species of Dendrobates have ter-restrial oviposition but transport tadpoles tophytotelms after hatching for instance we ob-served in a single occasion Dendrobates cf quin-quevittatus inside treeholes from which P resini-fictrix called More often however Dendrobatesspecies use small-sized ground phytotelms (egCaldwell and Araujo 1998) Overall this strongpattern of habitat segregation among phytotelmtypes in which more than one species are sel-dom found to coexist contrasts with ground wa-ter bodies where typically more than three andsometimes up to eight tadpole species coexist(Magnusson and Hero 1991)
Therefore several lines of evidence suggestthat both predation pressure and interspecificcompetition are lower in phytotelms than in ter-restrial water bodies and could contribute to themaintenance of the use of phytotelms as breed-ing and developmental sites for Amazonian an-urans in general and P resinifictrix in particularIn other systems however an unavailability ofterrestrial water bodies may act as a strong forcemaintaining phytotelm reproduction This ap-pears to be the case in Caribbean Islands suchas Jamaica where historical and geological fac-tors result in phytotelms being the predominantlentic water bodies (Thompson 1996)
Phytotelms are a harsh environment for thedevelopment of anuran embryos and larvaeConstrained volumes of noncirculating waterdominated by decaying processes sometimes re-sult in very low dissolved oxygen concentra-tions Assuming that treehole breeding is de-rived in the genus Phrynohyas early develop-ment of large inflated functional lungs in oth-er pond-dwelling species of the genus may beconsidered a useful morphophysiological trait
269SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
for the colonization of phytotelms by P resinific-trix Lungs are also used as hydrostatic organskeeping tadpoles close to the water surface andmay reduce the energy expenditure involved inrepeatedly swimming to the surface to bob air(Feder 1984 Feder and Moran 1985 Lannooet al 1987)
An important biotic characteristic of phyto-telms is the unpredictability and low availabilityof high quality food In most tadpole species forwhich dietary data are available algae of pe-riphyton and phytoplankton constitute majorfood resources in addition to low-quality detri-tus (eg Altig and Johnston 1989 Kupferberget al 1994) Narrow treehole openings oftencovered by overlying canopy may result in anearly aphotic habitat in which primary pro-ductivity is low and therefore likely insufficientto sustain tadpole growth
Detritus derived from the decomposition offallen leaf litter and treehole walls constitutedthe basis for the speciesrsquo diet when eggs werenot available The nutritive value of detritus islow (Kupferberg et al 1994) and several spe-cies of generalized pond tadpoles fail to reachmetamorphosis when restricted to this food type(Werner and Glennemeier 1999 LS unpubl)In this context oophagy may be of major im-portance for the success of individual larvae inreaching metamorphosis because eggs providea highly nutritious and energetic food source(Crump 1992) However detritivory could alsobe of crucial importance especially for smallertadpoles Prior to stage 32 tadpoles are gape-limited and therefore inefficient egg feedersAfter this stage tadpoles are capable of swallow-ing large quantities of whole eggs leading toaccelerated growth and development
Opportunistic oophagy is common in anuranlarvae (Magnusson and Hero 1991 Crump1992 Petranka and Kennedy 1999) In severalphytotelm-developing anurans however oop-hagy results from complex parental behavior inwhich the mother periodically returns to thephytotelm to lay fertilized or unfertilized eggsfor nutrition of its progeny (Lannoo et al 1987Weygoldt 1987 Crump 1995) For some ofthese species it was demonstrated that oophagyis obligate that is tadpoles fail to reach meta-morphosis if eggs are not provided by the moth-er (Weygoldt 1987 Thompson 1996 Jungferand Weygoldt 1999) In P resinifictrix howeverthere are several observations suggesting thatoophagy is opportunistic cannibalism ratherthan parental care in the form of periodic eggdeposition for obligate tadpole nutrition Firstas noted by Jungfer and Proy (1998) P resinific-trix egg clutches are large (maximum clutch size
in the field 1540 eggs this study in terraria4336 eggs Jungfer and Proy 1998) comparedto phytotelm-breeding species that intentionallyprovide eggs to their offspring For instanceconsidering only hylids on average 158 eggs arelaid by Anotheca spinosa ( Jungfer 1996) 258 byOsteocephalus oophagus ( Jungfer and Weygoldt1999) 276 per bromeliad by Osteopilus brunneus(Lannoo et al 1987) and 13 by Phyllodytes lu-teolus (Giaretta 1996) Second as a conse-quence of the large clutch size interclutch in-terval is long in P resinifictrix in a regularly sam-pled treehole six clutches were deposited overa period of four months (around one clutch ev-ery 20 days) which coincides with the inter-clutch interval of pairs in captivity (20 daysrange 7ndash37 n 5 29 Jungfer and Proy 1998)This long interclutch interval contrasts stronglywith the interclutch interval of species that areknown to be obligate oophagic For instanceeggs are deposited every 49 days in A spinosa( Jungfer 1996) 51 days in O oophagus ( Jung-fer and Weygoldt 1999) and 44 days in Obrunneus (Thompson 1996) Jungfer and Proy(1998) found that tadpoles of P resinifictrixstarve if dependent only on the motherrsquos infre-quent oviposition as the food source but thatmetamorphic success in an experimental con-tainer was high if in addition to leaf-litter eggswere added every 17 days Third an opportu-nistic oophagic nutrition is suggested by our ob-servation that eggs are fertilized unlike some ofthe more lsquolsquoadvancedrsquorsquo forms of anuran parentalcare through laying of unfertilized trophic eggs(Weygoldt 1987) although obligate oophagyhas been documented for species laying fertil-ized eggs (Thompson 1996) Fourth althoughextensive reduction of oral structures character-izes several of the obligate oophagic phytotelm-developing species (Lannoo et al 1987) P re-sinifictrix retains typical generalized pond-typelarval morphology with only moderate structur-al reduction when compared to its pond breed-ing congeners (Schiesari et al 1996) Experi-mental evidence is necessary to unambiguouslydemonstrate that oophagy is not obligate in Presinifictrix however the arguments above pro-vide indirect support for the hypothesis thatconsumption of eggs is important but may notbe obligate for metamorphic success
We suggest that among the premetamorphicstages of P resinifictrix life cycle cannibalism andintraspecific competition are the major forcesacting on the survival of eggs and larvae re-spectively Evidence for this hypothesis are thelow nutritive value of food other than eggs andor high unpredictability of food in the treeholethe voracious oophagic habits of the tadpole
270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo

269SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
for the colonization of phytotelms by P resinific-trix Lungs are also used as hydrostatic organskeeping tadpoles close to the water surface andmay reduce the energy expenditure involved inrepeatedly swimming to the surface to bob air(Feder 1984 Feder and Moran 1985 Lannooet al 1987)
An important biotic characteristic of phyto-telms is the unpredictability and low availabilityof high quality food In most tadpole species forwhich dietary data are available algae of pe-riphyton and phytoplankton constitute majorfood resources in addition to low-quality detri-tus (eg Altig and Johnston 1989 Kupferberget al 1994) Narrow treehole openings oftencovered by overlying canopy may result in anearly aphotic habitat in which primary pro-ductivity is low and therefore likely insufficientto sustain tadpole growth
Detritus derived from the decomposition offallen leaf litter and treehole walls constitutedthe basis for the speciesrsquo diet when eggs werenot available The nutritive value of detritus islow (Kupferberg et al 1994) and several spe-cies of generalized pond tadpoles fail to reachmetamorphosis when restricted to this food type(Werner and Glennemeier 1999 LS unpubl)In this context oophagy may be of major im-portance for the success of individual larvae inreaching metamorphosis because eggs providea highly nutritious and energetic food source(Crump 1992) However detritivory could alsobe of crucial importance especially for smallertadpoles Prior to stage 32 tadpoles are gape-limited and therefore inefficient egg feedersAfter this stage tadpoles are capable of swallow-ing large quantities of whole eggs leading toaccelerated growth and development
Opportunistic oophagy is common in anuranlarvae (Magnusson and Hero 1991 Crump1992 Petranka and Kennedy 1999) In severalphytotelm-developing anurans however oop-hagy results from complex parental behavior inwhich the mother periodically returns to thephytotelm to lay fertilized or unfertilized eggsfor nutrition of its progeny (Lannoo et al 1987Weygoldt 1987 Crump 1995) For some ofthese species it was demonstrated that oophagyis obligate that is tadpoles fail to reach meta-morphosis if eggs are not provided by the moth-er (Weygoldt 1987 Thompson 1996 Jungferand Weygoldt 1999) In P resinifictrix howeverthere are several observations suggesting thatoophagy is opportunistic cannibalism ratherthan parental care in the form of periodic eggdeposition for obligate tadpole nutrition Firstas noted by Jungfer and Proy (1998) P resinific-trix egg clutches are large (maximum clutch size
in the field 1540 eggs this study in terraria4336 eggs Jungfer and Proy 1998) comparedto phytotelm-breeding species that intentionallyprovide eggs to their offspring For instanceconsidering only hylids on average 158 eggs arelaid by Anotheca spinosa ( Jungfer 1996) 258 byOsteocephalus oophagus ( Jungfer and Weygoldt1999) 276 per bromeliad by Osteopilus brunneus(Lannoo et al 1987) and 13 by Phyllodytes lu-teolus (Giaretta 1996) Second as a conse-quence of the large clutch size interclutch in-terval is long in P resinifictrix in a regularly sam-pled treehole six clutches were deposited overa period of four months (around one clutch ev-ery 20 days) which coincides with the inter-clutch interval of pairs in captivity (20 daysrange 7ndash37 n 5 29 Jungfer and Proy 1998)This long interclutch interval contrasts stronglywith the interclutch interval of species that areknown to be obligate oophagic For instanceeggs are deposited every 49 days in A spinosa( Jungfer 1996) 51 days in O oophagus ( Jung-fer and Weygoldt 1999) and 44 days in Obrunneus (Thompson 1996) Jungfer and Proy(1998) found that tadpoles of P resinifictrixstarve if dependent only on the motherrsquos infre-quent oviposition as the food source but thatmetamorphic success in an experimental con-tainer was high if in addition to leaf-litter eggswere added every 17 days Third an opportu-nistic oophagic nutrition is suggested by our ob-servation that eggs are fertilized unlike some ofthe more lsquolsquoadvancedrsquorsquo forms of anuran parentalcare through laying of unfertilized trophic eggs(Weygoldt 1987) although obligate oophagyhas been documented for species laying fertil-ized eggs (Thompson 1996) Fourth althoughextensive reduction of oral structures character-izes several of the obligate oophagic phytotelm-developing species (Lannoo et al 1987) P re-sinifictrix retains typical generalized pond-typelarval morphology with only moderate structur-al reduction when compared to its pond breed-ing congeners (Schiesari et al 1996) Experi-mental evidence is necessary to unambiguouslydemonstrate that oophagy is not obligate in Presinifictrix however the arguments above pro-vide indirect support for the hypothesis thatconsumption of eggs is important but may notbe obligate for metamorphic success
We suggest that among the premetamorphicstages of P resinifictrix life cycle cannibalism andintraspecific competition are the major forcesacting on the survival of eggs and larvae re-spectively Evidence for this hypothesis are thelow nutritive value of food other than eggs andor high unpredictability of food in the treeholethe voracious oophagic habits of the tadpole
270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo

270 COPEIA 2003 NO 2
and the variable but potentially high larval den-sities found in the treeholes (up to 16 individ-ualsliter) which can deplete entire egg clutch-es
In the speciesrsquo original description Goeldi(1907) reported and illustrated in an apparentcombination of Indian legend and personal ob-servation that P resinifictrix constructs a basinmade of carefully chosen tree resins inside hol-low branches This nest accumulates fresh waterthat remains uncontaminated by decayingwood therefore providing lsquolsquoexcellent condi-tions for the hatching and development of theeggs and tadpolesrsquorsquo (Goeldi 1907) No sign ofresin lining was observed in any of the treeholessampled Lutz (1973) attributed this misconcep-tion to the occasional use by the species of cav-ities that had been lined with wax by wild beesknown to live in hollow trees
In this paper we demonstrated that P resini-fictrix is specialized for breeding in treeholesTherefore the availability of large water-hold-ing treeholes in the rain-forest canopy may havestrong impacts on both individual behavior andpopulation dynamics Although we did not mea-sure the availability of treeholes the followingevidence is consistent with the hypothesis oftreeholes constituting a limiting resource for Presinifictrix populations Calling males showstrong site fidelity and aggressive behavior in de-fense of the treehole multiple males were oftenobserved competing for a treehole Treeholeswere occupied most nights A regularly moni-tored treehole had at least six egg clutches in aperiod of five months indicating that males thatmonopolize a treehole may attain high repro-ductive success All treeholes inspected housedtadpoles during the entire rainy season Alter-natively if the cost or risk in locating a newunoccupied treehole is high males might cuethe presence of adequate treeholes by othermalesrsquo advertisement calls and selection mightas well favor strong male site fidelity and ag-gressiveness male satellite behavior and femaleoviposition in treeholes occupied by cannibal-istic tadpoles The alternative hypothesis how-ever that treeholes are not a limiting resourcebut are expensive in energy or risk to searchfor and find might not explain the constancy inthe density of calling males year after yearwhich is suggestive of resource limitation
The ecology and evolution of the use ofphytotelms as developmental sites for anuranamphibians has been increasingly identified asa promising area of research The present studycontributes to the understanding of phytotelmichabitats and of some of the most importanttraits involved in their colonization Future re-
search efforts should focus on experimental ap-proaches to the study of the larval feeding ecol-ogy and to the abiotic and biotic factors affect-ing the population dynamics of phytotelmic an-urans in general and P resinifictrix in particular
ACKNOWLEDGMENTS
M Henzl G de Mattos Accacio R Sousa PStumpe and B Wallnofer helped with field-work L Coelho identified in the field treeshosting P resinifictrix B Forsberg and T Pimen-tel helped with limnological analyses and DZago in the preparation and analysis of histo-logical sections This study was improved withthe criticism provided by M Trefaut RodriguesM Martins B Grillitsch R Lehtinen W Mag-nusson M Zimmerman and S McCauley Theauthors are grateful to W Magnusson and Ins-tituto Nacional de Pesquisas da Amazonia(INPA) for logistical support in Manaus and forH-W Koepcke and J Diller for use of their pri-vate field station lsquolsquoPanguanarsquorsquo near the villageof Llullapichis Peru LS is grateful to the su-pervision of M Trefaut Rodrigues to an under-graduate grant from Fundacao de Amparo aPesquisa do Estado de Sao Paulo (FAPESP pro-cess 911131-0) and by a doctoral grant of Con-selho Nacional de Desenvolvimento Cientıfico eTecnologico (CNPq process 20009397-5) dur-ing final preparation of the manuscript Speci-mens were collected under the IBAMA permits07592 and 03393 DEVIS-DIFAS Without thesupport of W Kerr the company of B Zimmer-man during many night walks in the ReservaDucke and the support of the Austrian ScienceFoundation (FWF projects 4346 5403 and6389) WH would never have had the chance toget involved with this frog species This study isdedicated to the memory of Werner C A Bo-kermann (1929ndash1995) whose knowledge of anddedication to the study of Brazilian vertebratesinfluenced a generation of naturalists
LITERATURE CITED
AICHINGER A N 1987 Annual activity patterns of an-urans in a seasonal Neotropical environment Oe-cologia 71583ndash592
ALTIG R AND G F JOHNSTON 1989 Guilds of anuranlarvae relationships among developmental modesmorphologies and habitats Herpetol Monogr 381ndash109
BRASIL 1978 Departamento Nacional de ProducaoMineral Projeto RADAMBRASIL Folha SA-20 Ma-naus Rio de Janeiro Brasil
CALDWELL J P AND M C DE ARAUJO 1998 Canni-balistic interactions resulting from indiscriminate
271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo

271SCHIESARI ET ALmdashPHYTOTELM DEVELOPMENT IN PHRYNOHYAS
predatory behavior in tadpoles of poison frogs(Anura Dendrobatidae) Biotropica 3092ndash103
CRUMP M L 1982 Amphibian reproductive ecologyon the community level p 21ndash36 In Herpetolog-ical communities N J Scott Jr (ed) Wildl ResRept No 13 US Fish and Wildlife Service Wash-ington DC
mdashmdashmdash 1992 Cannibalism in amphibians p 256ndash276In Cannibalism ecology and evolution among di-verse taxa M A Elga and B J Crespi (eds) Ox-ford Univ Press Oxford
mdashmdashmdash 1995 Parental care p 518ndash567 In Amphib-ian biologymdashsocial behaviour H Heatwole and BK Sullivan (eds) Surrey Beatty and Sons Chip-ping Norton New South Wales Australia
DUELLMAN W E AND L TRUEB 1986 Biology of am-phibians McGraw-Hill Book Co New York
FEDER M E 1984 Consequences of aerial respirationfor amphibian larvae p 71ndash86 In Respiration andmetabolism of embryonic vertebrates R S Sey-mour (ed) W Junk Publ Dordrecht Netherlands
mdashmdashmdash AND C M MORAN 1985 Effect of water depthon costs of aerial respiration and its alternatives intadpoles of Rana pipiens Can J Zool 63643ndash648
FISH D 1983 Phytotelmata fauna and flora p 1ndash27In Phytotelmata terrestrial plants as hosts foraquatic insect communities J H Frank and L PLounibos (eds) Plexus Publ Medford NJ
FROST D R 1985 Amphibian species of the world Ataxonomic and geographical reference Allen PressLawrence KS
GIARETTA A A 1996 Reproductive specializations ofthe bromeliad hylid frog Phyllodytes luteolus J Her-petol 3096ndash97
GOELDI E A 1907 Description of Hyla resinifictrixGoeldi a new Amazonian treefrog peculiar for itsbreeding habits Proc Zool Soc Lond 1907135ndash140
GOSNER K L 1960 A simplified table for staging an-uran embryos with notes on identification Herpe-tologica 16183ndash190
GRILLITSCH B 1992 Notes on the tadpole of Phrynoh-yas resinifictrix (GOELDI 1907) Buccopharyngealand external morphology of a tree hole dwellinglarva (Anura Hylidae) Herpetozoa 551ndash66
HADDAD C F B AND W HODL 1997 New reproduc-tive mode in anurans bubble nest in Chiasmocleisleucosticta (Microhylidae) Copeia 1997585ndash589
mdashmdashmdash AND R J SAWAYA 2000 Reproductive modesof Atlantic Forest hylid frogs a general overviewand the description of a new mode Biotropica 32862ndash871
HERO J-M 1990 An illustrated key to tadpoles oc-curring in the Central Amazonian rainforest Ma-naus Amazonas Brasil Amazoniana 11201ndash262
HODL W 1990 Reproductive diversity in Amazonianlowland frogs Fortschritte der Zoologie 3841ndash60
mdashmdashmdash 1991 Phrynohyas resinifictrix (Hylidae Anu-ramdashcalling behaviour Wiss Film 4263ndash70
JUNGFER K H 1996 Reproduction and parental careof the coronated treefrog Anotheca spinosa (Stein-dachner 1864) (Anura Hylidae) Herpetologica5225ndash32
mdashmdashmdash AND C PROY 1998 Phrynohyas resinifictrix
(Goeldi 1907) der Frosch der seine Frau bei Voll-mond ruft Geschichte und Fortpflanzungsverhal-ten im Terrarium Herpetofauna 2019ndash29
mdashmdashmdash AND L C SCHIESARI 1995 Description of aCentral Amazonian and Guianan treefrog genusOsteocephalus (Anura Hylidae) with oophagous tad-poles Alytes 131ndash13
mdashmdashmdash AND P WEYGOLDT 1999 Biparental care inthe tadpole-feeding Amazonian treefrog Osteocep-halus oophagus Amphib-Reptilia 20235ndash249
KUPFERBERG S J J C MARKS AND M E POWERS 1994Effects of variation in natural algal and detrital dietson larval anuran (Hyla regilla) life-history traits Co-peia 1994446ndash457
LANNOO M J D S TOWNSEND AND R J WASSERSUG1987 Larval life in the leaves arboreal tadpoletypes with special attention to the morphologyecology and behavior of the oophagous Osteopilusbrunneus (Hylidae) larva Fieldiana Zool New Ser381ndash31
LUTZ B 1973 Brazilian species of Hyla Univ of TexasPress Austin
MAGNUSSON W E AND J-M HERO 1991 Predationand the evolution of complex oviposition behaviourin Amazon rainforest frogs Oecologia 86310ndash318
MORI S A 1987 The Lecythidaceae of a lowlandNeotropical Forest La Fumee Mountain FrenchGuiana Mem New York Bot Gard 441ndash190
PETRANKA J W AND C A KENNEDY 1999 Pond tad-poles with generalist morphology is it time to re-consider their functional roles in aquatic commu-nities Oecologia 120621ndash631
SCHIESARI L C B GRILLITSCH AND C VOGEL 1996Comparative morphology of phytotelmonous andpond-dwelling larvae of four neotropical treefrogspecies (Anura Hylidae Osteocephalus oophagus Os-teocephalus taurinus Phrynohyas resinifictrix Phrynoh-yas venulosa) Alytes 13109ndash139 1 Erratum 14127ndash128
STRICKLAND J D H AND T R PARSONS 1968 A prac-tical handbook of seawater analysis Bull Fish ResBd Can 167
THOMPSON R L 1996 Larval habitat ecology andparental investment of Osteopilus brunneus (Hyli-dae) p 259ndash269 In Contributions to West Indianherpetology a tribute to Albert Schwartz R Powelland R W Henderson (eds) Society for the Studyof Amphibians and Reptiles Ithaca NY
WERNER E E AND K GLENNEMEIER 1999 The influ-ence of forest canopy cover on the breeding ponddistributions of several amphibian species Copeia19991ndash12
WETZEL R G 1983 Limnology Saunders CollegePublishing Orlando FL
WEYGOLDT P 1987 Evolution of parental care in dartpoison frogs (Amphibia Anura Dendrobatidae) Zzool Syst Evol-forsch 2551ndash67
ZIMMERMANN B AND W HODL 1983 Distinction ofPhrynohyas resinifictrix (Goeldi 1907) from P venu-losa (Laurenti 1768) based on acoustical and be-havioural parameters (Amphibia Anura Hylidae)Zool Anz Jena 211341ndash352
(LS) DEPARTAMENTO DE ZOOLOGIA INSTITUTO
272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo

272 COPEIA 2003 NO 2
DE BIOCIENCIAS UNIVERSIDADE DE SAO PAULOCP 11461 05422-970 SAO PAULO SAO PAULOBRAZIL (MG) DEPARTAMENTO DE BIOLOGIAUNIVERSIDADE DO AMAZONAS 69068 MANAUSAM BRAZIL AND (WH) ZOOLOGISCHES INSTI-TUT UNIVERSITAT WIEN ALTHANSTRASSE 14 A-1090 VIENNA AUSTRIA PRESENT ADDRESS
(LS) DEPARTMENT OF ECOLOGY AND EVOLU-TIONARY BIOLOGY UNIVERSITY OF MICHIGANANN ARBOR MICHIGAN 48109-1048 E-mail(LS) lschiesaumichedu Send reprint re-quests to LS Submitted 15 Aug 2002 Ac-cepted 14 Nov 2002 Section editor M JLannoo




















