
Transport of particulate organic carbon in the Amazon River: insights from river sediment...
23
Source, transport and fluxes of Amazon River particulate organic carbon: Insights from river sediment depth-profiles Julien Bouchez a,b,⇑ , Valier Galy c , Robert G. Hilton d,a , Je ´ro ˆ me Gaillardet a,e , Patricia Moreira-Turcq f , Marcela Andrea Pe ´rez g , Christian France-Lanord h , Laurence Maurice i a Institut de Physique du Globe de Paris, Equipe de Ge ´ochimie-Cosmochimie – Universite ´ Paris Diderot – Sorbonne Paris Cite ´, 1 rue Jussieu, 75238 Paris cedex 05, France b German Research Centre for Geosciences, Helmholtz Centre Potsdam, Telegrafenberg, 14473 Potsdam, Germany c Woods Hole Oceanographic Institution, 266 Woods Hole Rd., Woods Hole, MA 02543-1050, USA d Department of Geography, Durham University, South Road, Durham DH1 3LE, UK e Institut Universitaire de France, France f Institut de Recherche pour le De ´veloppement, 32 avenue Henri Varagnat, 93143 Bondy, France g Departamento de Geoquı ´mica, Instituto de Quı ´mica, Universidade Federal Fluminense, Outeiro de Sa ˜o Joa ˜o Batista, s/n, 24020-007 Niteroı ´-RJ, Brazil h Centre de Recherches Pe ´trographiques et Ge ´ochimiques – Centre National pour la Recherche Scientifique, 15 rue Notre-Dame-des-Pauvres, 54501 Vandoeuvre-le `s Nancy, France i Ge ´osciences Environnement Toulouse – Institut de Recherche pour le De ´veloppement, 14 avenue Edouard Belin, 31400 Toulouse, France Received 5 March 2013; accepted in revised form 24 February 2014; Available online 11 March 2014 Abstract In order to reveal particulate organic carbon (POC) source and mode of transport in the largest river basin on Earth, we sampled the main sediment-laden tributaries of the Amazon system (Solimo ˜ es, Madeira and Amazon) during two sampling campaigns, following vertical depth-profiles. This sampling technique takes advantage of hydrodynamic sorting to access the full range of solid erosion products transported by the river. Using the Al/Si ratio of the river sediments as a proxy for grain size, we find a general increase in POC content with Al/Si, as sediments become finer. However, the sample set shows marked variability in the POC content for a given Al/Si ratio, with the Madeira River having lower POC content across the measured range in Al/Si. The POC content is not strongly related to the specific surface area (SSA) of the suspended load, and bed sediments have a much lower POC/SSA ratio. These data suggest that SSA exerts a significant, yet partial, control on POC transport in Amazon River suspended sediment. We suggest that the role of clay mineralogy, discrete POC particles and rock-derived POC warrant further attention in order to fully understand POC transport in large rivers. To examine the source of POC in the Amazon Basin in more detail, we use radiocarbon ( 14 C) content and the stable isotope composition (d 13 C) of POC. 14 C activity demonstrates that rock-derived POC is a significant component of river bed sediments and contributes to POC across the river depth-profiles of suspended sediments. We estimate that the flux of rock-derived POC may reach 10% of the total POC export by the Amazon River. After correcting for rock-derived POC input, we find that POC from the terrestrial biosphere (biospheric POC) is mostly sourced from C3-plants. Higher biospheric POC d 13 C values in the Madeira River (by 0.5–1&) are best explained by a small (<5%) contribution of C4-grasses from Bolivian savannas. Finally, we use 14 C to estimate the mean age of biospheric POC exported from the Amazon Basin. The data show that biospheric POC is younger in the Solimo ˜ es River (1120 years) than in the Madeira River (2850 years). These http://dx.doi.org/10.1016/j.gca.2014.02.032 0016-7037/Ó 2014 Elsevier Ltd. All rights reserved. ⇑ Corresponding author at: Institut de Physique du Globe de Paris, Equipe de Ge ´ochimie-Cosmochimie – Universite ´ Paris Diderot – Sorbonne Paris Cite ´, 1 rue Jussieu, 75238 Paris cedex 05, France. Tel.: +33 (0)1 8395 74 37. E-mail address: [email protected] (J. Bouchez). www.elsevier.com/locate/gca Available online at www.sciencedirect.com ScienceDirect Geochimica et Cosmochimica Acta 133 (2014) 280–298
Transcript of Transport of particulate organic carbon in the Amazon River: insights from river sediment...

Available online at www.sciencedirect.com
www.elsevier.com/locate/gca
ScienceDirect
Geochimica et Cosmochimica Acta 133 (2014) 280–298
Source, transport and fluxes of Amazon River particulateorganic carbon: Insights from river sediment depth-profiles
Julien Bouchez a,b,⇑, Valier Galy c, Robert G. Hilton d,a, Jerome Gaillardet a,e,Patricia Moreira-Turcq f, Marcela Andrea Perez g, Christian France-Lanord h,
Laurence Maurice i
a Institut de Physique du Globe de Paris, Equipe de Geochimie-Cosmochimie – Universite Paris Diderot – Sorbonne Paris Cite, 1 rue Jussieu,
75238 Paris cedex 05, Franceb German Research Centre for Geosciences, Helmholtz Centre Potsdam, Telegrafenberg, 14473 Potsdam, Germany
c Woods Hole Oceanographic Institution, 266 Woods Hole Rd., Woods Hole, MA 02543-1050, USAd Department of Geography, Durham University, South Road, Durham DH1 3LE, UK
e Institut Universitaire de France, Francef Institut de Recherche pour le Developpement, 32 avenue Henri Varagnat, 93143 Bondy, France
g Departamento de Geoquımica, Instituto de Quımica, Universidade Federal Fluminense, Outeiro de Sao Joao Batista, s/n, 24020-007
Niteroı-RJ, Brazilh Centre de Recherches Petrographiques et Geochimiques – Centre National pour la Recherche Scientifique, 15 rue
Notre-Dame-des-Pauvres, 54501 Vandoeuvre-les Nancy, Francei Geosciences Environnement Toulouse – Institut de Recherche pour le Developpement, 14 avenue Edouard Belin, 31400 Toulouse, France
Received 5 March 2013; accepted in revised form 24 February 2014; Available online 11 March 2014
Abstract
In order to reveal particulate organic carbon (POC) source and mode of transport in the largest river basin on Earth, wesampled the main sediment-laden tributaries of the Amazon system (Solimoes, Madeira and Amazon) during two samplingcampaigns, following vertical depth-profiles. This sampling technique takes advantage of hydrodynamic sorting to access thefull range of solid erosion products transported by the river. Using the Al/Si ratio of the river sediments as a proxy for grainsize, we find a general increase in POC content with Al/Si, as sediments become finer. However, the sample set shows markedvariability in the POC content for a given Al/Si ratio, with the Madeira River having lower POC content across the measuredrange in Al/Si. The POC content is not strongly related to the specific surface area (SSA) of the suspended load, and bedsediments have a much lower POC/SSA ratio. These data suggest that SSA exerts a significant, yet partial, control onPOC transport in Amazon River suspended sediment. We suggest that the role of clay mineralogy, discrete POC particlesand rock-derived POC warrant further attention in order to fully understand POC transport in large rivers.
To examine the source of POC in the Amazon Basin in more detail, we use radiocarbon (14C) content and the stableisotope composition (d13C) of POC. 14C activity demonstrates that rock-derived POC is a significant component of riverbed sediments and contributes to POC across the river depth-profiles of suspended sediments. We estimate that the flux ofrock-derived POC may reach �10% of the total POC export by the Amazon River. After correcting for rock-derived POCinput, we find that POC from the terrestrial biosphere (biospheric POC) is mostly sourced from C3-plants. Higher biosphericPOC d13C values in the Madeira River (by �0.5–1&) are best explained by a small (<5%) contribution of C4-grasses fromBolivian savannas. Finally, we use 14C to estimate the mean age of biospheric POC exported from the Amazon Basin. Thedata show that biospheric POC is younger in the Solimoes River (1120 years) than in the Madeira River (2850 years). These
http://dx.doi.org/10.1016/j.gca.2014.02.032
0016-7037/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author at: Institut de Physique du Globe de Paris, Equipe de Geochimie-Cosmochimie – Universite Paris Diderot –Sorbonne Paris Cite, 1 rue Jussieu, 75238 Paris cedex 05, France. Tel.: +33 (0)1 8395 74 37.
E-mail address: [email protected] (J. Bouchez).

J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 281
ages are much younger than the corresponding estimates of sediment residence time in the basin, suggesting that lowlandareas and/or young POC from above-ground biomass may contribute disproportionately to the biomarker signals in terrig-enous sediments of the Amazon delta.� 2014 Elsevier Ltd. All rights reserved.
1. INTRODUCTION
Over geological time-scales, the main long-term sinks foratmospheric CO2 are carbon consumption by silicateweathering coupled to carbonate precipitation (Garrelsand Mackenzie, 1971; Gaillardet et al., 1999) and organiccarbon (OC) burial in marine and terrestrial sediments(Burdige, 2005; Galy et al., 2007a). These CO2 sinks actto negate the main sources of CO2 to the atmosphere,through volcanic and metamorphic degassing (Beckeret al., 2008), and oxidation of OC contained in sedimentaryrocks (Galy et al., 2008a; Bouchez et al., 2010; Hilton et al.,2011). Rivers play a central role in these C transfers (Sarmi-ento and Sundquist, 1992; Ludwig et al., 1996) by trans-porting solid products of erosion containing OC exportedfrom the terrestrial biosphere and mobilized from sedimen-tary rocks (Blair et al., 2004; Aufdenkampe et al., 2011). Inmany river basins these particles travel through extensiveriver networks and floodplains, which act as reactors whereOC can be processed, buried or destroyed (Richey et al.,2002; Battin et al., 2009). Constraining the fate of OC trans-ported through large rivers is essential to better understandhow erosion and weathering impact the global C cycle.
Particulate organic carbon (POC, i.e. present in particlescoarser than 0.22, 0.45, or 0.7 lm) has received consider-able focus because of its close relationship with clastic sed-iment. If POC survives transport, it may be buried insedimentary deposits and result in a long-term sink ofatmospheric CO2 (Galy et al., 2007a). However, thisstrongly depends upon the source of riverine POC, whichcan simply be considered in terms of two components:“rock-derived” OC (sometimes termed “fossil” OC or“petrogenic” OC), and “biospheric” OC derived fromabove-ground biomass (C3- and C4-plants), soils and river-ine autotrophic living organisms (Blair et al., 2003; Galyet al., 2008a; Hilton et al., 2008; Hilton et al., 2010). Bio-spheric POC can be reactive and has been shown to be par-tially oxidized during transport through large river systems,which results in a CO2 source over short timescales (Richeyet al., 2002; Mayorga et al., 2005). Variations in the resi-dence time of biospheric OC can also influence the atmo-spheric inventory of CO2 (Galy and Eglinton, 2011). Overgeological timescales (>105 years) the imbalance betweenthe oxidation of rock-derived POC and burial of biosphericPOC governs the net sink of atmospheric CO2 (Galy et al.,2007a; Hilton et al., 2012), whereas the burial of rock-de-rived OC has no net effect (Galy et al., 2008a; Hiltonet al., 2011). For these reasons, it is essential to quantifythe contribution of these POC sources in large rivers.
In addition to considering POC source, the mode oftransport of POC during fluvial transport must also beconstrained. River POC is transported either as OC closely
associated with inorganic clasts (Mayer, 1999) or as discreteOC particles. The former is thought to be relatively refrac-tory during riverine transport and after deposition (e.g.Hedges and Keil, 1995), while discrete OC particles maybe more prone to oxidation because of (1) their higherbuoyancy, hindering their ability to sink and reach suboxicand anoxic deep waters and sediments after delivery tolakes and oceans and (2) facilitated access to organic matterfor microbial organisms. Globally, POC is thought to belargely associated with mineral surfaces, on the basis of abroad correlation between POC content and specific surfacearea (SSA) in river sediments, estuaries and ocean margins(e.g. Keil et al., 1997). The observed OC content per unit ofmineral surface area (“OC-loading”) of river sediments typ-ically falls between 0.4 and 1.0 mg POC m�2 (Keil et al.,1994; Mayer, 1994a; Blair and Aller, 2012), correspondingto a monolayer of organic matter coating all mineral sur-faces (Mayer, 1994b). However, this first-order model ischallenged by the observation that OC is present at mineralsurfaces as discrete patches rather than as a continuouslayer (Ransom et al., 1997; Mayer, 1999). Furthermore,OC may have a high affinity for clay surfaces rather thanfor any mineral surface (Ransom et al., 1998; Bock andMayer, 2000; Curry et al., 2007; Galy et al., 2008b). Finally,discrete OC particles can comprise a significant componentof total POC from mountainous catchments to downstreamareas (West et al., 2011). Despite the global significance oflarge rivers POC delivery to the ocean, the source and modeof transport of POC in these settings are still poorlyconstrained.
Large tropical rivers drain continental areas of high pri-mary productivity (Nebel et al., 2001) and contribute signif-icantly to OC transport to the ocean (Hedges et al., 1986a;Spencer et al., 2012). The largest of these tropical river sys-tems is the Amazon Basin. Most of the significant body ofwork on the source and fate of riverine POC in the AmazonBasin has been based on suspended sediment collected atthe channel surface, mid-depth, or using depth-integratingsampling methods, then sieved into two size fractions: finePOC (FPOC, <63 lm) and coarse POC (CPOC, >63 lm).While informative, these techniques mask the hydrody-namic sorting processes at play in large river channels,which can contain novel information on the dominant pro-cesses controlling river sediment chemistry (e.g. Galy et al.,2008b). In contrast, we sampled river sediment and POCthroughout the water column, following depth-profilesfrom the channel surface to the river bed (Bouchez et al.,2011a). The samples capture the full range of sedimentgrain sizes transported by the river, extending from claysto coarse sands (Bouchez et al., 2011b). We use riverdepth-profiles of POC content, stable isotope compositionand radiocarbon activity to provide new insight on the

282 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298
mode of transport of POC in large rivers, make the firstassessment of the contribution of rock-derived POC toAmazon POC flux, and estimate the average age of bio-spheric POC transported in the Amazon Basin.
2. SETTING, SAMPLES, AND METHODS
2.1. The Amazon Basin
2.1.1. Geological and hydrological setting
The Amazon is the largest river system in the world interms of basin area and water discharge (around 6400 km3
year�1; Callede et al., 2000), and exports ca. 600 Mt year�1
of sediment (Filizola and Guyot, 2009). The headwaters ofthe Amazon Basin drain the Andes, while most of the basincomprises the vast lowland area of tropical South America.The lowland area is underlain by the Guyana and BrazilianPrecambrian shields to the west, and by several kilometersof mostly terrestrial sediment to the east (Fig. 1). The mainAmazon tributaries in terms of sediment supply are theSolimoes River draining the Northern Peruvian andEcuadorian Andes, and the Madeira River draining thesouthern Peruvian and Bolivian Andes (Gibbs, 1967). TheMadeira Basin mostly drains (meta)sedimentary Paleozoicand Mesozoic rocks, with large outcrops of organic-richshales and minor areas of intrusive volcanic rocks (Carlottoet al., 1996). In the Andean part of the Solimoes, (meta)sedimentary rocks also dominate alongside outcrops of acidto intermediate igneous rocks (Putzer, 1984). The other largetributaries are mostly sediment-starved, exhibit low dis-solved loads (Gaillardet et al., 1997), drain almost exclusivelythe lowland area, and are either characterized by high dis-solved organic matter concentrations (e.g. Negro River) orby a high in situ primary productivity (e.g. Tapajos River).The lowland Amazon Basin rivers is characterized by ana-branching channels (Latrubesse, 2008) and large floodplainsconsisting of a complex assembly of lakes, secondarychannels, and seasonally inundated forests. These wetlands,totaling more than 300,000 km2 in the Central Amazon
-85˚ - 80˚ -75˚ -70˚ -65˚-20˚
-15˚
-10˚
-5˚
0˚
5˚
500 km
acific ceanaAndes
ShieldAmazonbasin limit
-62 ˚ -60 ˚ -58 ˚ -56˚-4˚
-2˚
-1˚
Rio Negro
Solim›es
Mad
eira
T
Trombetas
ManacapuruFoz Madeira
Iracema
ParintinsObidos
Amazon
Solim
500 km
Fig. 1. Map of the Amazon Basin, with locations at which depth profiles(high water stage of the Solimoes and Amazon rivers, low water stageSolimoes and Amazon River, high water stage of the Madeira River).
(Hess et al., 2003) and through which 30% of the Amazonwater discharge transits (Richey et al., 1989; Bonnet et al.,2007), represent sites of important sediment andOC exchange and processing (Dunne et al., 1998;Maurice-Bourgoin et al., 2007; Moreira et al., 2012).
2.1.2. Previous work on particulate organic carbon in the
Amazon River
The Amazon River drains large tropical ecosystemswhich contain between 40 and 100 PgC in the lowlands(Houghton et al., 2001). The total (dissolved plus particu-late) estimated OC export to the ocean is 32–36 Mt year�1,30–40% of which is POC (Richey et al., 1990; Moreira-Turcq et al., 2003). During riverine transport in the low-lands, 0.5 Gt year�1 of CO2 are degassed to the atmosphere(Richey et al., 2002). The dominant source of respired OC isthought to be rapidly cycling, recently photosynthesizedOC (Hedges et al., 1986b; Richey et al., 1990), with 14C agesaround 5 years (Mayorga et al., 2005). In addition to bio-spheric OC, rock-derived POC is oxidized during transportthrough the lowland plains of the Amazon, resulting in asignificant long-term flux of CO2 to the atmosphere(0.25 MtC year�1 for the Madeira floodplain; Bouchezet al., 2010). Only 6–10% of the POC flux delivered to theAtlantic Ocean is deposited onto the shelf (Schlunz et al.,1999). Furthermore, 65–70% of terrestrial POC depositedon the Amazon shelf is thought to be oxidized over centen-nial to millennial timescales (Kuehl et al., 1986; Aller et al.,1996).
Most of the POC transported as suspended sediment bythe rivers in the lowland is present in the grain size fraction<63 lm, (FPOC; Richey et al., 1990), with <5% of the OCtransported as CPOC (>63 lm). Previous work hasestablished compositional differences between FPOC andCPOC, in terms of d13C, 14C-activity, C:N ratios, and lig-nin, aldose and amino acids yields and compositions(Hedges et al., 1986a,b, 1994; Richey et al., 1990; Quayet al., 1992; Mayorga et al., 2005). Most of these differencesare larger than (i) temporal variability at a given site, (ii)
-60˚ -55˚ -50˚ -45˚
-54˚
apaj
os
›es
Mad
eira
Amazon
of suspended sediment and bed sediment were sampled in June 2005of the Madeira River) and March 2006 (rising water stage of the

J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 283
longitudinal variations along the Amazon mainstem, and(iii) differences between tributaries. These differencesbetween grain size fractions were thus interpreted as reflect-ing differences in OC source and extent of OC degradation.The CPOC is thought to be mostly composed of well-preserved, contemporaneous woody debris and tree leavesfrom floodplain C3-metabolism plants, along with a minorcontribution (0–10%) of debris from C4-floodplain grasses(Hedges et al., 1986a,b, 1994). The FPOC, although ulti-mately deriving from similar plant sources, exhibits a morealtered state and older 14C-ages, respectively attributed tomicrobial degradation and longer residence times in soilsbefore export (Hedges et al., 1986a, 1994). FPOC is re-ported to be physically associated with inorganic materialas a result of sorption of OC onto mineral surfaces in soils(Hedges and Keil, 1995; Keil et al., 1997; Perez et al., 2011).The role of phytoplankton in OC composition in sediment-laden tributaries of the Amazon Basin is thought to besmall (Wissmar et al., 1981), although this figure has re-cently been disputed (Ellis et al., 2012). A longitudinaltrend in d13C values of FPOC along the mainstem led Quayet al. (1992) and Hedges et al. (2000) to estimate that be-tween 35% and 60% of the POC exported at the mouthwas derived from the lowland areas rather than from theAndes. This was attributed to progressive incorporationof OC photosynthesized in the lowlands, possibly combinedwith oxidation of upland OC.
2.2. Sampling
The Solimoes, Madeira, and Amazon rivers weresampled along channel depth-profiles during two cruises(Bouchez et al., 2011a,b), in June 2005 and March 2006,within the framework of the HyBAm program (Fig. 1,http://www.ore-hybam.org). The June 2005 sampling cam-paign corresponds to the high water stage of the SolimoesRiver (discharge normalized to the mean dischargeQ=Qmean = 1.22) and the low water stage of the MadeiraRiver ðQ=Qmean = 0.78), while the March 2006 samplingcampaigns corresponds to the high water stage of theMadeira River ðQ=Qmean = 1.69) and rising water stage ofthe Solimoes River ðQ=Qmean = 1.06). The Solimoes andMadeira rivers were sampled near their mouth at Manaca-puru and Foz Madeira, respectively. The Amazon main-stem was sampled at Iracema (downstream from theSolimoes-Negro confluence), Parintins (downstream fromthe confluence with the Madeira) and Obidos (the lastgauging station upstream from the Amazon mouth).
Sampling at discrete depths in large rivers grants accessto a range of the river solid load in terms of grain size, min-eralogy and chemical composition. Sampling procedureand sample treatment are described in detail in Bouchezet al. (2011a). In summary here, at each location, river bot-tom depth and water velocity were first recorded alongtransects using Acoustic Doppler Current Profiling (ADCP;Filizola and Guyot, 2003). One to three depth-profiles ofriver water samples were then collected. Each river depth-profile was composed of three to five samples collected witha point-sampler. River water samples were then filtered at0.22 lm-porosity, using PSE (Millipore) filter membranes.
Suspended sediments were retrieved from the filter sheetsand dried at 60 �C in the lab and aliquoted for the differentanalyses. At each site, one to six bed sediment samples weredredged from the bottom of the river. One bed sedimentsample was also taken near the mouth of the Negro River.The full results for sediment concentration, grain size, andinorganic chemistry are reported and discussed in detail inBouchez et al. (2011a,b,c).
2.3. Analytical methods
POC content (wt% of total sediment) and d13C (in &ofrelative deviation of the 13C/12C ratio as compared toVPDB) were determined on 5–20 mg-sample aliquots atthe Centre de Recherches Petrographiques et Geochimiques(CRPG), Vandoeuvre-les-Nancy, France, using a modifiedEurovector EuroEA3028-HT elemental analyzer coupledto a GV Instruments IsoPrime continuous-flow isotopemass spectrometer (Tables 1 and 2; Galy et al., 2007b;Bouchez et al., 2010). As the contribution of carbonates tototal C is small in sediments from the lowland Amazon(Mayorga et al., 2005), the aliquots were not decarbonatedprior to analyses in order to minimize bias on POC contentand d13C values (Galy et al., 2007b). Trace amounts of car-bonate would only slightly alter POC and d13C measure-ments (Appendix A). Although our results are internallyconsistent, our data might not be directly comparable toacid-insoluble (decarbonated) data reported elsewhere.The contribution of the blank (measured on a pure quartzpowder processed in the same way as the samples) was<0.01%. This blank contribution was not corrected for.Accuracy was checked and external reproducibility was esti-mated through repeated analyses of the in-house standardsoil “M2” (Galy et al., 2007b). External reproducibilities(2r) for standards were estimated to be 0.02% for POCcontent and 0.05& for d13C. Duplicate analysis for POCand d13C were carried out on 16 sediment samples by mea-suring separate powdered sample aliquots, and suggestrepeatability of 0.05% for POC content and 0.17& ford13C. These values are higher than the instrumental preci-sion and comparable to the bias introduced by the presenceof trace carbonate in our samples (Appendix A). We attri-bute such differences mostly to sample heterogeneity andconsider them as the best estimates of the overall data uncer-tainty. When duplicates were measured, the average POCcontent and d13C values were calculated and used in plotsand statistical analyses.
Activity of 14C was measured by accelerator mass spec-trometry at the LMC14 National Facility, Saclay, France,after off-line OC combustion and cryogenic C purification.Aliquots were decarbonated prior to analysis, as even traceamounts of 14C-extinct carbonate can significantly alter 14Cmeasurements. The amount of CO2 released during com-bustion was measured by barometric methods, allowingfor the calculation of the POC content after decarbonation,POCdecarb (or “acid-insoluble”, Table 3). These POCdecarb
values are typically 20% lower than POC content measuredon non-decarbonated aliquots because of solubilization oforganic matter during decarbonation (Galy et al., 2007b).14C values were corrected for instrumental mass dependent

284 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298
fractionation (normalization to a d13C value of �25&) andexpressed as fraction modern carbon (Fmod), relative to 95%of the 14C activity of the oxalic acid standard (Stuiver andPolach, 1977). An analytical 2r-uncertainty of 0.003 wasestimated for Fmod values.
Specific surface area (SSA) was measured on 0.3 to1.0 g-aliquots using the N2 adsorption isotherms method(Brunauer et al., 1938), at the Laboratoire d’Environne-ment et Mineralurgie (LEM), Nancy, France, to a precisionof 0.5 m2 g�1 (Table 4). Some of these measurements weredone on amalgamated suspended load samples, due toinsufficient amount of material in single samples. Grain sizedistribution was determined on 50 mg-aliquots dispersed indistilled water using a laser granulometer Malvern Master-sizer at the Laboratoire d’Environnement et de Mineralur-gie, Nancy, France (Bouchez et al., 2011a). Repeatability ofthe grain size distribution parameters (such as mean,median, or mode) was estimated to be around 10%. Thechemical composition of the inorganic phase was measuredat the SARM (Service d’Analyses des Roches et desMineraux, http://helium.crpgnancy.fr/SARM) NationalFacility, Vandoeuvre-les-Nancy, France, using ICP-OESfor major elements and ICP-MS for minor and traceelements, after dry ashing and alkali fusion (Bouchezet al., 2011b). Typical uncertainties are 3% for majorelements and better than 10% for minor and traceelements.
3. RESULTS
3.1. POC content
POC content in the main tributaries of the lowlandAmazon Basin (Tables 1 and 2) varies from 0.05% to1.02% in bedload samples (average value 0.17 � 0.27% 1standard deviation, n = 17; 13 samples have POC contentbelow 0.10%), and from 0.60% to 2.22% in suspended loadsamples (1.17 � 0.34%, n = 83). For both sampling cam-paigns, the highest average POC contents in the suspendedload are measured for the Amazon at Iracema(1.49 � 0.28%, n = 4 for the high water stage and1.22 � 0.33%, n = 7 for the rising water stage). The lowestPOC contents in the suspended load are obtained for theMadeira (1.23 � 0.15%, n = 7 for the low water stage and0.76 � 0.07%, n = 11 for the high water stage). These num-bers are within the range previously reported for Amazonrivers lowland sites (Hedges et al., 1986a, 1994; Richeyet al., 1990; Quay et al., 1992; Moreira-Turcq et al., 2003).
The largest variability in suspended load POC contentwith depth for a given sampling site is observed for theAmazon River at Parintins (1.47 � 0.48%, n = 8) and Obi-dos (1.24 � 0.41%, n = 9) during the high water stage (witha relative standard deviation, RSD of 33% for both sites)and for the Amazon River at Iracema during the risingwater stage (RSD = 27%). The lowest variability is ob-tained for the Madeira River (RSD = 12% for both lowand high water stage). The Madeira River also exhibitsthe largest and most significant difference (Student’s t-test:p < 0.0001) between the two sampling campaigns, whilethe Solimoes and the Amazon rivers display distributions
of POC content that are not significantly different fromone sampling campaign to the other.
Trends of decreasing POC content with increasing sam-pling depth are observed for several vertical profiles of theSolimoes and the Amazon at Iracema, for both samplingcampaigns (Fig. 2). POC content is constant with depthin the other profiles (e.g. profile #2 of the Solimoes duringthe high water stage, Table 1). Differences exist between dif-ferent depth-profiles within a given sampling campaign; forexample for the high water stage of the Solimoes, profile #1yields systematically higher POC contents than profile #2(Table 2). In the Madeira, POC contents are invariant withdepth (Fig. 2B). The Amazon at Parintins and Obidos dis-play increasing POC contents towards the surface, but withsignificant lateral heterogeneity for Obidos during highwater stage (Fig. 2C and D).
3.2. Stable isotope composition
Values of POC d13C range from �25.8 to �27.5& in bedsediment samples (�26.5 � 0.5&, n = 17), and from �26.5to �29.0&in suspended load samples (�27.9 � 0.6&,n = 83, Tables 1 and 2, Fig. 3). Suspended load is thusdepleted in 13C when compared to bed sediment, but iswithin the range previously reported for the Amazon River(Hedges et al., 1986a, 1994; Quay et al., 1992; Mayorgaet al., 2005; Mortillaro et al., 2011; Moreira-Turcq et al.,2013). The most depleted d13C values in the suspended loadare observed for the Solimoes (�28.3 � 0.4&, n = 13during the rising water stage and �28.2 � 0.3&, n = 8during high water stage) and the Amazon at Iracema(�28.2 � 0.2&, n = 7 during the rising water stage and�28.4 � 0.3&, n = 4 during the high water stage). TheNegro River bed sediment has the most depleted d13C value(�29.6&, n = 1). By contrast, the Madeira suspended loadexhibits the most enriched d13C values (�27.6 � 0.3&,n = 7 during the low water stage and �26.8 � 0.3&,n = 11 during the high water stage). The Amazon mainstemsuspended load shows intermediate values (for example atObidos: �27.7 � 0.3&, n = 10 during the rising water stageand �28.1 � 0.2&, n = 9 during the high water stage).
For a given sampling campaign, d13C value distributionsare statistically similar for the Solimoes and the Amazon atIracema. Similarly, d13C value distributions are statisticallysimilar for the Amazon at Parintins and at Obidos. Distri-bution of d13C values (Fig. 3) are significantly different be-tween the two sampling campaigns for the Madeira(p < 0.0004) for the Amazon at Parintins and Obidos(p < 0.005), but not for the Solimoes dataset (includingthose from the Amazon at Iracema). No systematic trendof d13C with sampling depth was observed in the depth-profiles.
3.3. Activity of 14C, rock-derived POC content and age of
biospheric POC
In depth-profile suspended sediments, 14C activityranges between 0.37 and 0.86 Fmod (Table 3; Bouchezet al., 2010). At each location, bed sediment is moredepleted in 14C than suspended load. The Fmod values
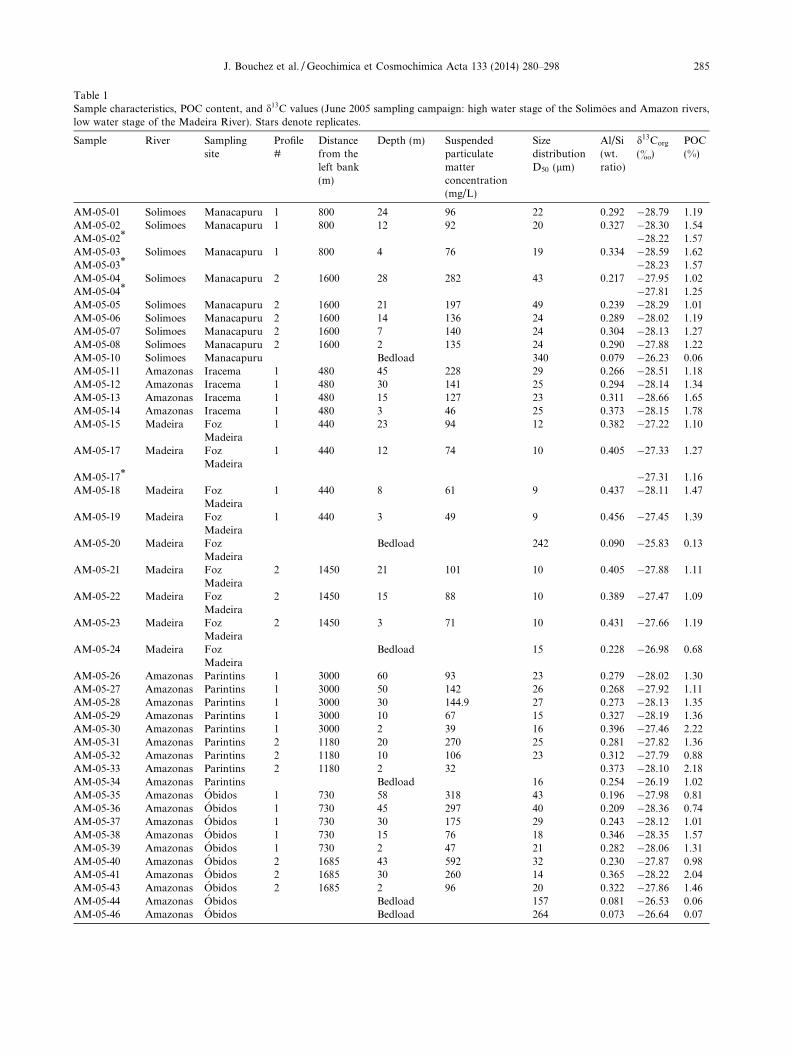
Table 1Sample characteristics, POC content, and d13C values (June 2005 sampling campaign: high water stage of the Solimoes and Amazon rivers,low water stage of the Madeira River). Stars denote replicates.
Sample River Samplingsite
Profile#
Distancefrom theleft bank(m)
Depth (m) Suspendedparticulatematterconcentration(mg/L)
SizedistributionD50 (lm)
Al/Si(wt.ratio)
d13Corg
(&)POC(%)
AM-05-01 Solimoes Manacapuru 1 800 24 96 22 0.292 �28.79 1.19AM-05-02 Solimoes Manacapuru 1 800 12 92 20 0.327 �28.30 1.54AM-05-02* �28.22 1.57AM-05-03 Solimoes Manacapuru 1 800 4 76 19 0.334 �28.59 1.62AM-05-03* �28.23 1.57AM-05-04 Solimoes Manacapuru 2 1600 28 282 43 0.217 �27.95 1.02AM-05-04* �27.81 1.25AM-05-05 Solimoes Manacapuru 2 1600 21 197 49 0.239 �28.29 1.01AM-05-06 Solimoes Manacapuru 2 1600 14 136 24 0.289 �28.02 1.19AM-05-07 Solimoes Manacapuru 2 1600 7 140 24 0.304 �28.13 1.27AM-05-08 Solimoes Manacapuru 2 1600 2 135 24 0.290 �27.88 1.22AM-05-10 Solimoes Manacapuru Bedload 340 0.079 �26.23 0.06AM-05-11 Amazonas Iracema 1 480 45 228 29 0.266 �28.51 1.18AM-05-12 Amazonas Iracema 1 480 30 141 25 0.294 �28.14 1.34AM-05-13 Amazonas Iracema 1 480 15 127 23 0.311 �28.66 1.65AM-05-14 Amazonas Iracema 1 480 3 46 25 0.373 �28.15 1.78AM-05-15 Madeira Foz
Madeira1 440 23 94 12 0.382 �27.22 1.10
AM-05-17 Madeira FozMadeira
1 440 12 74 10 0.405 �27.33 1.27
AM-05-17* �27.31 1.16AM-05-18 Madeira Foz
Madeira1 440 8 61 9 0.437 �28.11 1.47
AM-05-19 Madeira FozMadeira
1 440 3 49 9 0.456 �27.45 1.39
AM-05-20 Madeira FozMadeira
Bedload 242 0.090 �25.83 0.13
AM-05-21 Madeira FozMadeira
2 1450 21 101 10 0.405 �27.88 1.11
AM-05-22 Madeira FozMadeira
2 1450 15 88 10 0.389 �27.47 1.09
AM-05-23 Madeira FozMadeira
2 1450 3 71 10 0.431 �27.66 1.19
AM-05-24 Madeira FozMadeira
Bedload 15 0.228 �26.98 0.68
AM-05-26 Amazonas Parintins 1 3000 60 93 23 0.279 �28.02 1.30AM-05-27 Amazonas Parintins 1 3000 50 142 26 0.268 �27.92 1.11AM-05-28 Amazonas Parintins 1 3000 30 144.9 27 0.273 �28.13 1.35AM-05-29 Amazonas Parintins 1 3000 10 67 15 0.327 �28.19 1.36AM-05-30 Amazonas Parintins 1 3000 2 39 16 0.396 �27.46 2.22AM-05-31 Amazonas Parintins 2 1180 20 270 25 0.281 �27.82 1.36AM-05-32 Amazonas Parintins 2 1180 10 106 23 0.312 �27.79 0.88AM-05-33 Amazonas Parintins 2 1180 2 32 0.373 �28.10 2.18AM-05-34 Amazonas Parintins Bedload 16 0.254 �26.19 1.02AM-05-35 Amazonas Obidos 1 730 58 318 43 0.196 �27.98 0.81AM-05-36 Amazonas Obidos 1 730 45 297 40 0.209 �28.36 0.74AM-05-37 Amazonas Obidos 1 730 30 175 29 0.243 �28.12 1.01AM-05-38 Amazonas Obidos 1 730 15 76 18 0.346 �28.35 1.57AM-05-39 Amazonas Obidos 1 730 2 47 21 0.282 �28.06 1.31AM-05-40 Amazonas Obidos 2 1685 43 592 32 0.230 �27.87 0.98AM-05-41 Amazonas Obidos 2 1685 30 260 14 0.365 �28.22 2.04AM-05-43 Amazonas Obidos 2 1685 2 96 20 0.322 �27.86 1.46AM-05-44 Amazonas Obidos Bedload 157 0.081 �26.53 0.06AM-05-46 Amazonas Obidos Bedload 264 0.073 �26.64 0.07
J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 285
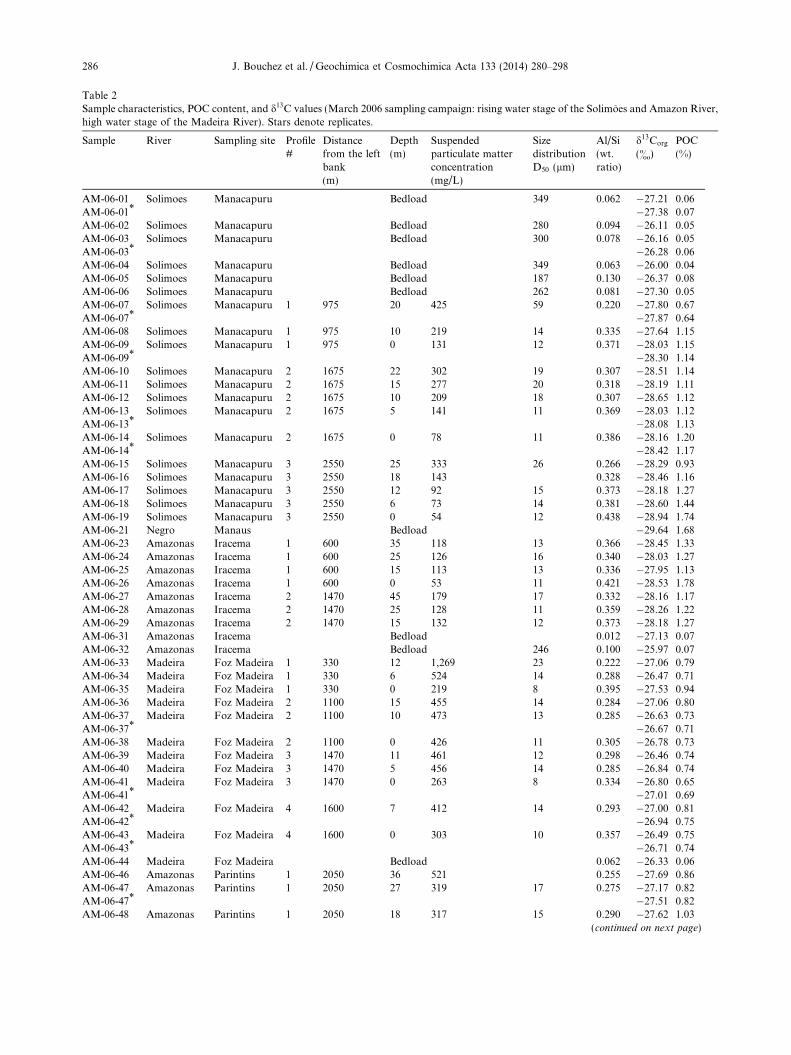
Table 2Sample characteristics, POC content, and d13C values (March 2006 sampling campaign: rising water stage of the Solimoes and Amazon River,high water stage of the Madeira River). Stars denote replicates.
Sample River Sampling site Profile#
Distancefrom the leftbank(m)
Depth(m)
Suspendedparticulate matterconcentration(mg/L)
SizedistributionD50 (lm)
Al/Si(wt.ratio)
d13Corg
(&)POC(%)
AM-06-01 Solimoes Manacapuru Bedload 349 0.062 �27.21 0.06AM-06-01* �27.38 0.07AM-06-02 Solimoes Manacapuru Bedload 280 0.094 �26.11 0.05AM-06-03 Solimoes Manacapuru Bedload 300 0.078 �26.16 0.05AM-06-03* �26.28 0.06AM-06-04 Solimoes Manacapuru Bedload 349 0.063 �26.00 0.04AM-06-05 Solimoes Manacapuru Bedload 187 0.130 �26.37 0.08AM-06-06 Solimoes Manacapuru Bedload 262 0.081 �27.30 0.05AM-06-07 Solimoes Manacapuru 1 975 20 425 59 0.220 �27.80 0.67AM-06-07* �27.87 0.64AM-06-08 Solimoes Manacapuru 1 975 10 219 14 0.335 �27.64 1.15AM-06-09 Solimoes Manacapuru 1 975 0 131 12 0.371 �28.03 1.15AM-06-09* �28.30 1.14AM-06-10 Solimoes Manacapuru 2 1675 22 302 19 0.307 �28.51 1.14AM-06-11 Solimoes Manacapuru 2 1675 15 277 20 0.318 �28.19 1.11AM-06-12 Solimoes Manacapuru 2 1675 10 209 18 0.307 �28.65 1.12AM-06-13 Solimoes Manacapuru 2 1675 5 141 11 0.369 �28.03 1.12AM-06-13* �28.08 1.13AM-06-14 Solimoes Manacapuru 2 1675 0 78 11 0.386 �28.16 1.20AM-06-14* �28.42 1.17AM-06-15 Solimoes Manacapuru 3 2550 25 333 26 0.266 �28.29 0.93AM-06-16 Solimoes Manacapuru 3 2550 18 143 0.328 �28.46 1.16AM-06-17 Solimoes Manacapuru 3 2550 12 92 15 0.373 �28.18 1.27AM-06-18 Solimoes Manacapuru 3 2550 6 73 14 0.381 �28.60 1.44AM-06-19 Solimoes Manacapuru 3 2550 0 54 12 0.438 �28.94 1.74AM-06-21 Negro Manaus Bedload �29.64 1.68AM-06-23 Amazonas Iracema 1 600 35 118 13 0.366 �28.45 1.33AM-06-24 Amazonas Iracema 1 600 25 126 16 0.340 �28.03 1.27AM-06-25 Amazonas Iracema 1 600 15 113 13 0.336 �27.95 1.13AM-06-26 Amazonas Iracema 1 600 0 53 11 0.421 �28.53 1.78AM-06-27 Amazonas Iracema 2 1470 45 179 17 0.332 �28.16 1.17AM-06-28 Amazonas Iracema 2 1470 25 128 11 0.359 �28.26 1.22AM-06-29 Amazonas Iracema 2 1470 15 132 12 0.373 �28.18 1.27AM-06-31 Amazonas Iracema Bedload 0.012 �27.13 0.07AM-06-32 Amazonas Iracema Bedload 246 0.100 �25.97 0.07AM-06-33 Madeira Foz Madeira 1 330 12 1,269 23 0.222 �27.06 0.79AM-06-34 Madeira Foz Madeira 1 330 6 524 14 0.288 �26.47 0.71AM-06-35 Madeira Foz Madeira 1 330 0 219 8 0.395 �27.53 0.94AM-06-36 Madeira Foz Madeira 2 1100 15 455 14 0.284 �27.06 0.80AM-06-37 Madeira Foz Madeira 2 1100 10 473 13 0.285 �26.63 0.73AM-06-37* �26.67 0.71AM-06-38 Madeira Foz Madeira 2 1100 0 426 11 0.305 �26.78 0.73AM-06-39 Madeira Foz Madeira 3 1470 11 461 12 0.298 �26.46 0.74AM-06-40 Madeira Foz Madeira 3 1470 5 456 14 0.285 �26.84 0.74AM-06-41 Madeira Foz Madeira 3 1470 0 263 8 0.334 �26.80 0.65AM-06-41* �27.01 0.69AM-06-42 Madeira Foz Madeira 4 1600 7 412 14 0.293 �27.00 0.81AM-06-42* �26.94 0.75AM-06-43 Madeira Foz Madeira 4 1600 0 303 10 0.357 �26.49 0.75AM-06-43* �26.71 0.74AM-06-44 Madeira Foz Madeira Bedload 0.062 �26.33 0.06AM-06-46 Amazonas Parintins 1 2050 36 521 0.255 �27.69 0.86AM-06-47 Amazonas Parintins 1 2050 27 319 17 0.275 �27.17 0.82AM-06-47* �27.51 0.82AM-06-48 Amazonas Parintins 1 2050 18 317 15 0.290 �27.62 1.03
(continued on next page)
286 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298

0.5 1.0 1.5 2.0POC (%)
0
5
10
15
20
25
30
Dep
th (
m)
Solimões
risinghigh
highlow
Madeira
POC (%)
0
10
20
30
40
50
60
Dep
th (
m)
Amazon (Parintins)
risinghigh
Amazon (Iracema)
risinghigh
POC (%)
0
10
20
30
40
50
60
Dep
th (
m)
2.5
Amazon (Óbidos)
risinghigh
0.5 1.0 1.5 2.0 2.5 0.5 1.0 1.5 2.0
0.5 1.0 1.5 2.0POC (%)
0
5
10
15
20
25
Dep
th (
m)
A B
C D
Fig. 2. POC content vs. depth in Amazon river suspended sediment. The uncertainty on sampling depth is ca. 1 m (Bouchez et al., 2011b).Labels “high”, “low”, and “rising” refer to the water stage.
Table 2 (continued)
Sample River Sampling site Profile#
Distancefrom the leftbank(m)
Depth(m)
Suspendedparticulate matterconcentration(mg/L)
SizedistributionD50 (lm)
Al/Si(wt.ratio)
d13Corg
(&)POC(%)
AM-06-51 Amazonas Parintins 1 2050 8 106 10 0.390 �28.14 1.19AM-06-52 Amazonas Parintins 1 2050 0 88 8 0.430 �28.17 1.22AM-06-53 Amazonas Parintins Bedload 237 0.087 �26.39 0.10AM-06-54 Amazonas Parintins Bedload 117 0.134 �27.52 0.18AM-06-55 Amazonas Obidos 1 630 55 341 24 0.214 �27.74 0.60AM-06-56 Amazonas Obidos 1 630 40 193 14 0.308 �27.96 1.04AM-06-57 Amazonas Obidos 1 630 25 198 13 0.325 �27.51 1.07AM-06-58 Amazonas Obidos 1 630 10 124 11 0.331 �27.55 1.11AM-06-59 Amazonas Obidos 1 630 0 101 9 0.398 �28.02 1.20AM-06-60 Amazonas Obidos 2 1900 35 482 17 0.292 �27.14 0.90AM-06-61 Amazonas Obidos 2 1900 20 159 11 0.352 �27.39 1.08AM-06-62 Amazonas Obidos 2 1900 0 84 8 0.430 �27.98 1.22AM-06-63 Amazonas Obidos 3 1200 45 249 13 0.315 �27.10 0.76AM-06-63* �27.47 0.91AM-06-65 Amazonas Obidos 3 1200 0 81 11 0.423 �28.03 1.23
J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 287

10
20
30
40
50
% s
ampl
e
Madeirahigh water stage
Madeiralow water stage
Amazon@ Parintinsand îbidoshigh water stage rising water stage
Solim›es + Amazon @ Iracema
10
20
30
40
% s
ampl
e
-30 -29 -28 -270
10
20
30
40
% s
ampl
e
δ13C (ä)-29 -28 -27 -26
δ13C (ä)-26
A
B C
D E
50
60
Bedload samples
Solim›es high water stageSolim›es rising water stageMadeira low water stage
Madeira high water stage
Amazon rising water stage
Amazon high water stage
Amazon@ Parintinsand îbidos
Fig. 3. Distribution of d13C values in suspended sediments of Amazon Basin main tributaries. The distributions yielded by the two samplingcampaigns were not statistically different for the Solimoes (see text). Therefore, they are plotted as a single distribution in panel (A). For theMadeira (B and C) and the Amazon at Obidos (D and E), distinct distributions were obtained. In each diagram, the number of bins has beenadapted to the distribution for the sake of clarity. Bed sediment d13C values are plotted for comparison (see legend).
288 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298
measured here extend the range previously reported for theAmazon ðFmod = 0.55–1.0; Mayorga et al., 2005) towardlower Fmod values, and are consistent with the value re-ported by Moreira-Turcq et al. (2013) for the Amazon nearObidos (Fmod ’ 0.8). In river sediments, POC can be inter-preted as a binary mixture of rock-derived OC and bio-spheric OC with distinct 14C content (e.g. Blair et al.,2003; Galy et al., 2008a; Blair and Aller, 2012). Althoughit is known that the 14C activity of rock-derived POC is 0,both the 14C activity of the biospheric pool (Fmodbio
) andthe contribution of rock-derived OC to total POC(POCrock ) are unknown. In order to determine the two un-knowns, we use an end member mixing analysis as Galyet al. (2008a). Details are given in Appendix B and inBouchez et al. (2010).
The Solimoes, Madeira and Amazon (at Obidos) riversall display relatively low POCrock between 0.02 � 0.03% forthe Madeira and 0.06 � 0.05% for the Amazon at Obidos(Fig. 4, Table 3). While this corresponds to only 3–10%
of total POC in suspended load samples, rock-derivedOC contributes significantly to POC in most bed sedimentsamples (up to ca. 50% in the Solimoes, for example).These numbers are lower than those reported for the Gan-ges–Brahmaputra system, where the proportion of rock-de-rived POC to total POC can reach 20% in suspended loadand 100% in bedload (Galy et al., 2007a), and for the An-dean headwaters of the Madre de Dios, where this contri-bution can reach 80% in suspended sediment (Clarket al., 2013).
The mean activity of 14C in the biospheric POC poolusing the end member mixing approach (Galy et al.,2008a; Galy and Eglinton, 2011) is the lowest for the Ma-deira (Fmodbio
= 0.71 � 0.04) and significantly higher forthe Solimoes and the Amazon at Obidos (respectively0.87 � 0.03 and 0.84 � 0.04; Fig. 4, Table 3). These num-bers correspond to 14C conventional ages of 2750 yearsfor the Madeira River, 1120 years for the Solimoes and1400 years for the Amazon at Obidos.

0.4 0.8 1.20.0
1.2
0.8
0.4
0.0
POC (%)
Mod
ern
C (
%)
2σ
Solim›es
risinghigh
Madeira
Amazon
susp. bed
high
risinghigh
Fig. 4. Modern C (POC � Fmod, with Fmod = fraction moderncarbon, which is the sample 14C activity relative to 95% of theactivity of the oxalic acid standard) as a function of POC content(Table 3). Samples from a given river site (including bed andsuspended sediments and the two sampling campaigns) definedistinct straight lines (stippled lines), showing that the concentra-tion of rock-derived POC (POCrock, for which the 14C activity is 0)and the Fmod of the biospheric POC (Fmodbio
) are both, at first-order,invariant with time and grain size for a given river (Galy et al.,2008b; Bouchez et al., 2010; Galy and Eglinton, 2011). Thecorresponding values can be determined from the slope and theintercept of each lines (Appendix B).
J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 289
3.4. Specific surface area
Specific Surface Area (SSA) is lower in Amazon Basinbed sediment samples (3.9 m2 g�1 in the Madeira and5.4 m2 g�1 in the Solimoes) than in suspended load, whereit ranges from 10.5 to 22.8 m2 g�1 in the Solimoes and from10.5 to 19.6 m2 g�1 for the Madeira (Table 4).
4. DISCUSSION
Important variations in POC content, isotope composi-tion, and SSA are observed along depth-profiles of Amazon
Table 314C activity (expressed as fraction modern carbon, Fmod) and POC contenaliquots analyzed for 14C activity (Bouchez et al., 2010). Absolute conbiospheric pool (Fmodbio
) are also reported for each river (Appendix B, S
Sample River Sampling site Depth (m)
AM-05-04 Solimoes Manacapuru 28AM-05-08 Solimoes Manacapuru 2AM-05-10 Solimoes Manacapuru BedloadAM-06-10 Solimoes Manacapuru 22AM-06-36 Madeira Foz Madeira 15AM-06-38 Madeira Foz Madeira 0AM-06-44 Madeira Foz Madeira BedloadAM-05-35 Amazon Obidos 58AM-05-37 Amazon Obidos 30AM-05-39 Amazon Obidos 2AM-06-64 Amazon Obidos 20AM-06-66 Amazon Obidos Bedload
River sediments. Herein we interpret these variations interms of hydrodynamic sorting of inorganic particles andorganic matter within the channel, attempting to decipherthe modes of POC transport. We then re-assess the contri-bution of different sources (rock-derived, biospheric fromC3 and C4 plants) to the Amazon POC load. Finally, weassess the variability in the mean age of biospheric POCtransported in the Amazon River.
4.1. Transport of POC in the Amazon Basin
4.1.1. Control of POC content by sediment grain size and
specific surface area
Our results demonstrate the variability of POC contentin Amazon river sediments across a given cross-section(Fig. 2). While lateral variability can be attributed toshort-term variations in sediment transport capacity inthe channel, variations with depth are most likely relatedto hydrodynamic sorting of particles following their grainsize (Bouchez et al., 2011a). It is widely recognized thatPOC content in sediments carried by large rivers varies withgrain size, and is commonly enriched in fine size fractionswhich can offer larger mineral surface area (e.g. Hedgesand Keil, 1995; Keil et al., 1997; Galy et al., 2008b). To fur-ther examine this link between POC and inorganic sedimentload, here we use measurements of SSA and Al/Si ratios.Al/Si ratios can serve as a proxy for sediment grain size dis-tribution (Galy et al., 2008b; Bouchez et al., 2011b). Firstwe note that based on previous work, sediment samplesfrom depth-profiles can be interpreted as a mixture between(1) a coarse-grained, quartz-rich, low-Al/Si end-memberbest represented by bed sediment and (2) a fine-grained,clay-rich, high Al/Si end-member best represented by sus-pended sediment sampled near the channel surface. In theAmazon, the correlation between Al/Si and SSA(Fig. 5A), mostly set by the low Al/Si bed sediment sam-ples, shows that high-Al/Si, fine particles transported nearthe channel surface have higher SSA than low-Al/Si, coarseparticles typically present near or at the river bottom.
In the Solimoes and Madeira sample set, POC increaseswith SSA (Fig. 5B). The slope in the POC–SSA diagram isclassically referred to as “OC-loading” in mg POC m�2
(Keil et al., 1997; Galy et al., 2008b). For suspended
t after decarbonation (POCdecarb, “acid-insoluble”, Appendix A) forcentration of rock-derived OC (POCrock) and 14C activity of the
upplementary Fig. 1).
POCdecarb (%) Fmod POCrock (%) Fmodbio
0.79 0.86 0.03 0.871.13 0.83 �0.02 �0.030.06 0.370.95 0.830.62 0.71 0.02 0.710.65 0.68 �0.03 �0.040.05 0.450.65 0.79 0.06 0.840.92 0.78 �0.05 �0.041.22 0.810.93 0.770.65 0.77

0.0 0.1 0.2 0.3Al/Si
0
5
10
15
20
25
SS
A (
m /g
)2
),%(COP0.0 0.5 1.0 1.5
Solim›esMadeira
OC
-load
ing
= 0.
4 m
g OC
/m2
OC-loading = 1.0 mg OC/m
2
A B
Gan
ges
Brahmaputra
Suspended load
Solim›esMadeira
Bed sediments
Fig. 5. (A) Specific surface area (SSA) vs. Al/Si in Amazon river sediment (Tables 1–3). Al/Si is used here as a proxy for grain size distribution(Bouchez et al., 2011a). The trends displayed by the Ganges and Brahmaputra rivers (Galy et al., 2008a) are shown as grey stippled lines forcomparison. The linear best fit over the whole dataset is shown as a continuous black line (SSA [m2 g�1] = 44.2 � Al/Si + 0.9, R2 = 0.58),although different residual variances are obtained when rivers are considered separately (R2 = 0.33 for the Solimoes, R2 = 0.89 for theMadeira). (B) SSA vs. POC content (Tables 1 and 2). The range of trends of OC-loading commonly observed in river sediment (Blair andAller, 2012) is shown. All analyzed samples are from the March 2006 sampling campaign (rising water stage of the Solimoes and Amazonrivers, high water stage of the Madeira River).
Table 4Specific surface area (SSA) measurements. For composite samples, Al/Si, POC and d13C were calculated from the properties of the individualsamples used to produce the composite mixture, along with the weight proportions. AM-06-S1 is a mixture between samples AM-06-07, AM-06-10 and AM-06-15; AM-06-S2 a mixture between samples AM-06-08, AM-06-11 and AM-06-16, and AM-06-S3 a mixture between samplesAM-06-09, AM-06-13, AM-06-14 and AM-06-18. AM-06-M1 is a mixture between samples AM-06-35 and AM-06-43.
Sample River Sampling site Sample type Al/Si POC (%) d13C (&) SSA (m2 g�1)
AM-06-05 Solimoes Manacapuru Bedload 0.130 0.08 �26.4 5.4AM-06-S1 Solimoes Manacapuru Composite 0.241 0.79 �28.1 12.6AM-06-S2 Solimoes Manacapuru Composite 0.325 1.14 �28.1 22.8AM-06-S3 Solimoes Manacapuru Composite 0.374 1.18 �28.3 10.5AM-06-33 Madeira Foz Madeira Suspended load 0.222 0.79 �27.1 10.5AM-06-34 Madeira Foz Madeira Suspended load 0.288 0.71 �26.5 10.9AM-06-44 Madeira Foz Madeira Bedload 0.062 0.06 �26.3 3.9AM-06-M1 Madeira Foz Madeira Composite 0.365 0.79 �26.8 19.6
290 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298
sediments, the OC-loadings defined by the depth-profiles ofSolimoes and Madeira sediments are consistent with thosemeasured on grain size separates of depth-integrated Ama-zon suspended sediment samples (between 0.4 and 1.0 mgPOC m�2) and from other rivers systems (Fig. 5B; Keilet al., 1997; Blair and Aller, 2012). However, they do notdefine a single value of OC-loading, with bed sediments fall-ing outside of the range of relationships reported by Blairand Aller (2012).
When we use Al/Si as a proxy for grain size to expandthe sample set, we find a broad positive relationship be-tween POC content and the Al/Si ratio throughout sus-pended and bed sediments collected over the Amazonlowland basin (Fig. 6). To the first order, this shows thatfine particles contain more POC than coarse particles.However, there are significant spatial and temporal varia-tions between rivers. For a given grain size (Al/Si ratio),the data suggests more POC is present in sediments of theSolimoes River during high-water stage than during risingwater (this is also true for the Amazon at Iracema). Regard-less of the water stage, Madeira river sediments of a given
grain size bear less POC than sediments from the othersampling locations.
Therefore, over the Amazon lowland basin, in generalPOC content increases with decreasing grain size of themineral load. However, the nature of the link betweenPOC and inorganic matter appears to differ between (1)bed and suspended sediments and (2) different rivers. Inthe following sub-sections we discuss the potential controlson OC-loading in the Amazon before considering the vari-ability in the sources of POC (Section 4.2).
4.1.2. Controls on OC-loading in bed sediment
River bed sediments lie off the “OC-loading” trends de-fined by Keil et al. (1997) and have a relatively low POCcontent for their SSA (Fig. 5B). This observation is similarto that made by Arnarson and Keil (2001) on sedimentsfrom the Pacific margin, where the POC/SSA ratio was low-est in coarse sediment. The 14C-activity of the bed sedimentsuggests that rock-derived POC contributes significantly tototal POC (ca. 50%, Table 3). Rock-derived POC is oftenencountered within clasts or as large discrete particles and

0.20 0.25 0.30 0.35 0.40 0.50
Al/Si
-29
-28
-27
-26
δ13C
(‰
)
0.45
Solimõesrisinghigh
Madeirahighlow
Fig. 7. d13C vs. Al/Si in Amazon river suspended sediments.Dotted lines represent linear best-fits for each of the rivers (using allprofiles from both sampling date).
0.0 0.1 0.2 0.3 0.4 0.5Al/Si
0.0
0.5
1.0
1.5
2.0
2.5P
OC
(%
)
GangesBrahm.
Solim›es
risinghigh
Madeira
Amazon
susp. bed
highlow
risinghigh
Fig. 6. POC content vs. Al/Si in Amazon river sediment. Therelationships obtained for the Ganges and Brahmaputra rivers(Galy et al., 2008a) are indicated as grey bands for comparison. Al/Si is a proxy for grain size distribution (Bouchez et al., 2011a).Fine, clay-rich sediment are characterized by a high Al/Si ratio.Coarse, quartz-rich sediment have a low Al/Si ratio. In the text, werefer to the “Solimoes” relationship, including the data from theAmazon at Iracema, to the “Madeira” relationship and to the“Amazon” relationship including the data from both Parintins andObidos sampling sites. These trends include both suspended andbed sediments.
J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 291
so is not bound to clastic grain surfaces (Galy et al., 2008a;Bouchez et al., 2010). We therefore suggest that at the lowSSA prevailing in coarse bed sediments (ca. 5 m2 g�1), POCis largely influenced by the presence of discrete particles ofrock derived POC, rather than by mineral-bound OC.Importantly, this suggests that a weak control of POC con-tent by SSA is also likely for low Al/Si-suspended sedimentstransported near the channel bed (Tables 1 and 2; Bouchezet al., 2011a).
4.1.3. Controls on OC-loading in suspended sediment
Amazon suspended sediment samples plot within the do-main of OC-loading relationships derived by Keil et al.(1997) for river sediments (0.4–1.0 mg POC m�2; Fig. 5B),suggesting a control of POC content by SSA. However,three observations suggest that important additional fac-tors may control POC content in Amazon suspended sedi-ments: (a) the correlation between SSA and POC content isrelatively poor compared to that between SSA and Al/Si(Fig. 5); (b) the two sampling campaigns yield differentPOC-Al/Si relationships for the Solimoes River (Fig. 6);(c) the Solimoes River sediments transport more POC thanthe Madeira River sediments at a given Al/Si (Fig. 6).Importantly, rock-derived POC is a relatively small contrib-utor to total POC in these suspended sediment samples(with POCrock of ca 0.05% for a total POC content higherthan 0.7%, Tables 1–3), hence the amount of rock-derivedPOC should not be a major factor controlling OC-loadingof the suspended load. In addition, the influence of in situ
primary productivity at high water stage is likely to be min-
or, as high turbidity hinders phytoplankton growth (Wiss-mar et al., 1981). We therefore invoke two factors whichcan control POC content, in addition to SSA.
First, although SSA allows for a crude quantification ofthe amount of clay minerals present in a sample, as shownby the correlation between SSA and Al/Si ratio (Fig. 5A), itdoes not account for mineral surface charge and chemistry,particle geometry or crystalline structure (Mayer et al.,2004; Curry et al., 2007). It is likely that these parameters,and hence type of clay mineral assemblage, play an impor-tant role on OC-loading, as suggested by Galy et al. (2008b)for the Ganges–Brahmaputra River. In fact, Guyot et al.(2007) report that the clay fraction of the Solimoes Rivercontains more smectite and less chlorite than that of theMadeira River. Given the high specific surface area ofsmectite, a modest contribution from smectite may affectsediment SSA but not OC content. Therefore these miner-alogical differences may explain the different POC-Al/Sirelationships of these two rivers (Fig. 6).
A second possible explanation is related to the presenceof discrete organic debris in the POC load. The ability ofrivers to transport organic debris harvested from soils andvegetation in upland areas has been demonstrated to be en-hanced at high water stage (West et al., 2011; Clark et al.,2013). By analogy, discrete organic debris could be inputto the Solimoes River during flooding of adjacent banksand floodplains at high water stage, thereby increasingOC-loading (Fig. 6). The abundance of discrete POC isnot controlled by SSA, but is likely to be enhanced inhigh-Al/Si sediment samples sampled near the channel sur-face, if discrete OC particles are more buoyant than the restof the riverine sedimentary material.
If significant, this enrichment may alter the d13C compo-sition of POC as a function of grain size, provided that dis-crete and mineral-bound POC have significantly differentd13C values. There is some indication from previous workthat this may be the case, as suggested by (a) the average

-29 -28 -27 -26 -25 -24
δ13C
0.0
0.2
0.4
0.6
0.8
1.0
Fm
od
1.2
rock OC
Solim›es
risinghigh
Madeira
Amazon
susp. bed
high
high
Fig. 8. Activity of 14C (expressed as fraction modern carbon, Fmod)vs. d13C of organic carbon in Amazon River depth-profiles. Thesamples having the lowest Fmod and least negative d13C result froma binary mixture between biospheric POC (which is in turn amixture between different sources) and rock-derived POC. Thelinear best-fit (thick black line) through bed and deep suspendedsediments (n = 4 combined) is the mixing line corresponding to thisbinary mixture. Extrapolation of this line to Fmod = 0 yields therock-derived d13C POC value. If bed sediment contained enoughtrace carbonates such that their d13C is biassed (Appendix A), therange of d13C would extend to more negative values.
292 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298
d13C values of �30.1& reported by Hedges et al. (1986a)for leaves (forming a large fraction of discrete POC), and(b) the d13C values of �26.8& reported by Quay et al.(1992) for FPOC (thought to reflect mineral-bound OC)of the Solimoes and Madeira. Therefore, an increasing con-tribution of discrete POC to total POC toward the channelsurface may result in decreasing d13C values with decreasingsampling depth, decreasing sediment grain size, or increas-ing Al/Si.
Although no systematic trend between sampling depthand d13C was observed, we note that suspended sedimentd13C is negatively correlated with Al/Si in the Solimoesand Madeira rivers (R2 = 0.10, p < 0.07 and R2 = 0.56,p < 0.001 for the Solimoes and the Madeira, respectively;Fig. 7). The broad negative trends may be consistent withan enrichment of discrete POC toward the channel surfaceand suggest a partial control of POC content by the discretePOC particles. However, the samples vary considerably ind13C for a given Al/Si. In addition, discrete POC particlesand mineral-bound POC from soils may have d13C valueswhich partially overlap (Clark et al., 2013). Nevertheless,together our observations warrant further research intothe role of clay mineral assemblage and discrete POC trans-port in large river systems.
4.2. Source of POC in the Amazon Basin
4.2.1. Contribution of rock-derived OC
Our measurements of 14C content of POC in the Ama-zon Basin demonstrate that rock-derived POC is presentin river sediments of the major tributaries (Table 3). Thiswarrants an assessment of its influence on POC fluxes inthe Amazon Basin. In Amazon River sediment, absolutecontent of rock-derived POC (POCrock ) has been shownto be relatively invariant throughout depth-profiles(Fig. 4). Therefore, POCrock does not depend on grain sizeand is present in a range of clastic particles or as discreteparticles (Galy et al., 2008a; Bouchez et al., 2010). Valuesof POCrock (Table 3, content in weight %) thus can be mul-tiplied by depth-integrated sediment fluxes to calculaterock-derived OC fluxes (Supplementary material). We usethe upper limit of the 95% confidence interval (mean + 2r)to estimate the maximum flux of rock-derived POC ex-ported by the rivers of the Amazon Basin. Considering bothsampling campaigns, the calculated POCrock fluxes are�10 kg s�1 for the Solimoes and range between 45 and70 kg s�1 for the Amazon at Obidos. For the Madeira Riv-er, the maximum estimates for the POCrock flux are<1 kg s�1 during the low water stage and �10 kg s�1 duringthe high water stage. We estimate that for our samplingdates the maximum rock-derived POC contribution is 5–8% of the total, depth-integrated POC flux of the Solimoesand the Madeira rivers and �10% for the Amazon at Obi-dos (Supplementary material). These fluxes would representC sequestered from the atmosphere in the geological pastand their potential contribution to the modern river loadcannot be ignored.
At these lowland locations of the Amazon River, previ-ous work has found that most of the rock-derived POC is ofgraphitic structure (Bouchez et al., 2010). Graphitic carbon
is thought to be resistant to oxidation during riverine trans-port and is therefore locked into the geological C cycle(Galy et al., 2008a). This oxidation-resistant fraction maytherefore be transferred quantitatively to the Amazon delta.Combining the rock-derived POC concentration found forthe Amazon at Obidos (Table 3; Bouchez et al., 2010) withdecadal estimates of the Amazon sediment flux from rivergauging (600 Mt year�1; Filizola and Guyot, 2009), we esti-mate that the long-term flux of rock-derived, refractoryPOC exported at Obidos is between 0.06 and0.6 MtC year�1. These estimates are similar to the flux ofC to the atmosphere derived from rock-derived OC oxida-tion in the Madeira floodplain (0.25 MtC year�1; Bouchezet al., 2010), suggesting that a significant fraction ofrock-derived POC escapes oxidation during transit in theAmazon Basin. The flux of rock-derived POC transferredto the oceans can also be compared to OC burial on theAmazon shelf. Richey et al. (1990) report a flux of totalPOC exported by the Amazon of 14 MtC year�1. If we as-sume that the burial efficiency reported for POC depositedon the shelf (35–40%; Aller et al., 1996) applies to the wholeAmazon depositional system, the flux of Amazon POC bur-ial is ca. 5 MtC year�1. Therefore, the flux of refractoryrock-derived POC exported by the Amazon system esti-mated here represents 1–10% of this long-term POC burialflux. Despite the significant uncertainties, these calculationsdemonstrate that an important component of the POCwhich escapes oxidation during transport and isexported to the Amazon shelf may be refractory, graphitic,
rock

J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 293
rock-derived POC, likely to be sourced from erosion inmountain catchments of the Andes (Clark et al., 2013).
4.2.2. Downstream evolution of biospheric POC
Previous work has estimated that between 35% and 60%of the POC exported at the Amazon mouth is derived fromthe lowland areas (Quay et al., 1992; Hedges et al., 2000).However, this was based on a downstream decrease in bulkFPOC d13C values along the mainstem of the Amazon. Asour work demonstrates, FPOC is likely to comprise a mix-ture of biospheric POC and rock-derived POC, rather thansolely biospheric POC. The rock-derived POC content ofthe Beni River at the outlet of the Andes is 0.26 � 0.11%(Bouchez et al., 2010), for total POC contents around1.5% (Hedges et al., 2000). Therefore, ca. 15% of FPOCmay be derived from rocks in the Andean part of the Ma-deira Basin. In order to infer the isotopic composition ofbiospheric POC leaving the Andes, the d13C compositionof rock-derived POC has to be identified. We use the factthat the activity of 14C in Amazon river depth-profiles isnegatively related to d13C composition, with bed sedimentdriving this relationship because of their higher relativerock-derived POC content (Fig. 8). Suspended load sam-pled near the channel surface displays the most depletedd13C values and the highest Fmod. Extrapolating this trendto Fmod = 0, the estimated d13Crock is between �24.3&
and �25.7&, consistent with the values reported by Clarket al. (2013). When we take rock-derived POC contributionand d13C composition into account, we can correct the d13Cvalue of bulk POC reported by Hedges et al. (2000) for theBeni (�28.2& and �26.9& for coarse and fine POC,respectively). We calculate the d13C of the biospheric POCexported from the Bolivian Andes to be �28.9& (CPOC)and �27.4& (FPOC). The relatively enriched POC d13Cvalues in the Peruvian headwaters of the Solimoes River(Aufdenkampe et al., 2007; Townsend-Small et al., 2007)might also partially stem from a significant contributionof rock-derived OC, as shown for the headwaters of theMadre de Dios by Clark et al. (2013).
Downstream in the suspended load of the Solimoes (orMadeira) River, the concentrations of POCrock found here(Table 3) are equivalent to only a few % of total POC. Gi-ven the typical d13C value of rock-derived POC (Clarket al., 2013), such contribution does not affect significantlyd13C values of bulk suspended load sampled near the sur-face or around mid-depth. This means that, in the centralAmazon, bulk d13C values reported by Quay et al. (1992)(�26.8 � 0.5&and �27.4 � 0.4&for CPOC and FPOC,respectively) are likely to reflect primarily biospheric POCd13C values.
Therefore, no large decrease in d13C values is apparentalong the course of the Madeira when only biospheric OCis considered. The POC replacement in floodplains inferredby Hedges et al. (2000) might occur, but our analysis showsthat biospheric d13C values are not sensitive to this process.As riverine POC is a complex mixture of a vast composi-tional range of terrestrial organic molecules, compound-specific measurements (d13C, 14C activity) might betterreveal POC fate, processing, and replacement during trans-fer through the Amazon lowland.
4.2.3. Source of biospheric POC
As discussed by Hedges et al. (1986a), the d13C valuesexhibited by biospheric POC in the Solimoes (in the range�28.5 to �28.0&, Fig. 3) are compatible with a dominantlowland C3-plant source. However, the Madeira biosphericPOC displays enriched d13C values across the range of sed-iment grain sizes carried by the river (Fig. 7), rangingbroadly from �28.0& to �26.0&. At least one additionalsource is required to explain the Madeira POC d13C values,characterized by a less depleted d13C. Phytoplankton islikely not a significant component of POC in our samplesgiven the high turbidity of water at the time of sampling(Wissmar et al., 1981). Plants growing at high altitudes(Bird et al., 1994) in the Andean upland of the Madeira Ba-sin may contribute to the Madeira mouth POC as an en-riched d13C-source. However we show above that thebiospheric POC d13C value at the Andean outlet is relativelyclose to that measured at the mouth of the Madeira byQuay et al. (1992) and in this study (�26.8 � 0.5&; Ta-ble 1). Alternatively, a low-d13C component in the MadeiraRiver could be derived from the floodplain C4-grassessourced from the Beni floodplain where vast savanna eco-systems are present (e.g. Llanos de Moxos; Hamiltonet al., 2004). Using the isotopic composition of C4-lowlandgrasses (d13C = �12.2&; Hedges et al., 1986b), a contribu-tion of only 3–5% from these grasses is sufficient to explainthe d13C values measured in POC at the mouth of the Ma-deira. This calculated contribution of C4-grasses is compa-rable to that inferred by Hedges et al. (1986a) for theAmazon mainstem CPOC (0–10%). The enriched d13C val-ues of POC during the high water stage of the Madeira(Fig. 3) may therefore reflect a higher input of POC fromthe savannas during this period, consistent with rapid trans-port of OC from upstream areas (i.e. Beni floodplain) underdynamic hydrological conditions.
Therefore, using the end-member mixing approach ofGaly et al. (2008a), we are able to quantify the contributionof rock-derived POC to Amazon POC, allowing for aninterpretation of biospheric POC stable isotope composi-tion. In the following section, we examine the 14C age ofbiospheric POC transported in the Amazon Basin.
4.3. POC age and timescale of sediment transport in the
Amazon Basin
Using an established technique, we use the 14C-activityand POC content of the river sediment samples to estimatethe mean age of biospheric POC transported in the AmazonBasin (Fig. 4; Galy et al., 2008a). The 14C ages of 2750, 1120,and 1400 years obtained for POC transported in the Ma-deira, Solimoes, and Amazon at Obidos, respectively, showthat on average biospheric POC resides for a significant per-iod of time in the Amazon Basin before being exported tothe ocean. This is clear when we consider the contemporary14C age of biospheric POC exported from a small mountainwatershed in the Andean headwaters of the Amazon (Clarket al., 2013). The ages are similar to those reported for theGanges River system by Galy and Eglinton (2011). Theseobservations hint that POC aged in soils and floodplainsmakes an important contribution transported in large rivers.

294 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298
The contrast in biospheric POC age between the Ma-deira (2750 years) and the Solimoes (1120 years) is in thesame direction as observed for the timescale of clastic sed-iment transport in the Madeira (22,000 years) and Solimoes(5000 years), as inferred from U-series disequilibria(Dosseto et al., 2006a,b). This suggests that sedimentdynamics may play an important role on biospheric OCresidence time in large river floodplains. Floodplaindestruction by erosional processes (such as bank erosionor sediment scouring during floods) is known to return alarge amount of inorganic sediment to the river channel(Dunne et al., 1998). This sediment is likely to be associatedwith POC from soils and overlying vegetation. However, atour three sampling locations, the age of biospheric POC issignificantly younger than the timescale of sediment trans-port reported by Dosseto et al. (2006a,b). Therefore, de-spite the close link between clastic sediment characteristicsand POC transport (e.g. Fig. 6), at the basin scale, theresidence time of sedimentary material is longer than theresidence time of C in the ecosystems.
This finding potentially points to heterotrophic process-ing and turnover of organic matter, which will limit the ageof POC contained within soils compared to the mineral sub-strate (Trumbore, 1993; Torn et al., 1997). In addition, recy-cling of POC may occur during downstream fluvialtransport, deposition and re-suspension. These processeswould act to lower the net age of biospheric POC relativeto the timescale of sediment transport. If biospheric POC ex-ported by large rivers inherits its younger age from process-ing during downstream transport, its biochemicalfingerprints (e.g. d13C or C:N ratios) are likely to preferen-tially reflect the characteristics of lowland ecosystems ratherthan those from the whole basin (e.g. Eckard et al., 2007).This would have important implications for understandingthe impact of biospheric POC transport on C-budgets incatchments (Galy and Eglinton, 2011) and for the interpre-tation of biomarker signals in the sedimentary record (e.g.Kastner and Goni, 2003; Bendle et al., 2010).
Alternatively, the mean biospheric POC ages might re-flect a more complex age structure (Rosenheim and Galy,2013) with POC of an age comparable to inorganic sedi-ment residence time (e.g. deep/old soil OC) mixing withyounger OC, potentially of contemporaneous age (e.g. livestanding biomass and recently deposited litter). If the age oflarge river biospheric POC results from a complex mixtureof different OC pools, it is essential to extract compound-specific signals from the sedimentary record to faithfullyreconstruct paleo-environmental shifts in large river basins.
5. CONCLUSIONS
The present study examines, for the first time, the POCcontent and isotopic composition of Amazon River sedi-ments collected along river depth-profiles. This samplingtechnique grants access to the full range of sediment grainsizes transported by a large river, and provides new insighton the source, mode of transport and fluxes of POC in thelargest river system in the world.
The amount of POC for a given river sediment size frac-tion, or OC-loading, appears to be controlled in the
suspended load by (1) the quantity of surface area; (2) thetype of surface area; (3) the presence of discrete organicparticles. The contribution of the latter to total POC de-serves further scrutiny, as if buoyant, discrete POC maybe oxidized in deltas or in the oceans over a relatively shorttimescale, in contrast to POC more closely associated withclastic sediment, which may deposited and protected fromoxidation.
Using the 14C content of the river POC, we show thatrock-derived POC is a major component of POC in riverbed sediment and contributes to POC at all depths in thesampled rivers. We estimate that rock-derived POC con-tributes a maximum of 10% of the POC exported to theocean by the Amazon River. The exported rock-derivedOC may be an important contributor to offshore OC burialbecause it is mostly comprised of graphitic carbon whichmay escape remineralization following deposition.
Having accounted for rock-derived POC, we are able toexamine the isotopic composition of biospheric POC. In theupland tributaries (e.g. Beni River), rock-derived POC canbe significant and its d13C-enriched isotope compositionmust be accounted for before interpreting downstreamchanges in biospheric d13C values. Having done this, wesuggest minimal change in the d13C values of biosphericPOC in the major Amazon tributaries (e.g. the Madeira),which calls into question previous estimates of significantreprocessing of biospheric POC during transit in the upperAmazon Basin, based on the bulk stable isotopecomposition.
The 14C activities of POC are used to estimate the meanage of biospheric POC transported by rivers in the AmazonBasin. Biospheric POC is, on average, younger in the Soli-moes (1120 years) than the Madeira (2750 years), which isconsistent with the longer residence time of clastic sedimentin the Madeira basin estimated from U-series disequilibria.However, the average biospheric POC ages are much youn-ger than estimated sediment residence times (5000–22,000 years). This may be due to a significant contributionof POC from young above-ground biomass, which mixeswith an older component of biospheric POC. In this case,14C ages would allow for a faithful reconstruction of pastchanges in environmental conditions only if extracted fromspecific compounds. Future work is needed to characterizethe age distribution of POC exported by the Amazon andother large rivers in order to test this hypothesis.
ACKNOWLEDGEMENTS
This work was funded by the “Reliefs de la Terre” INSU-CNRS program. The field campaigns, supported by the FrenchInstitute for Research and Development (IRD), were realized inthe frame of the HyBAm (Hydrology and Geochemistry of theAmazonian Basin, http://www.mpl.ird/hybam/, cooperation agree-ment with the Brazilian Research Centre – CNPq-no 492685/2004-5). We sincerely acknowledge the Brazilian Institutions andUniversities (ANA, UnB, UFF, CPRM) which collaborate in thisproject. V. Galy was supported by the National Science Founda-tion (Grants OCE-0851015 and OCE-0928582). C. Guilmette andA. Razafitianamaharavo are thanked for their help in the lab. Thismanuscript benefited from thorough reviews by Brad Rosenheimand two anonymouns reviewers, and from insightful comments

J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 295
by Associate Editor Peter Hernes. This is IPGP contribution no.3509.
APPENDIX A. ABSENCE OF DECARBONATION
PRIOR TO POC AND 13C MEASUREMENTS
No decarbonation was carried out before POC contentand d13C measurements. River sediments from the centralAmazon Basin are known to be carbonate-poor (Mayorgaet al., 2005; Galy et al., 2007b) such that the presence oftrace carbonates is unlikely to influence the measuredPOC content. Furthermore, avoiding decarbonation pre-vents partial dissolution of labile OC, which would resultin POC contents artificially lowered by up to 20%, andpotentially biassed POC d13C values (Galy et al., 2007b).However, the presence of trace carbonates could bias themeasured d13C compositions towards less negative values.This is not the case for the present sample set. First, sequen-tial extractions carried out on Amazon River bed sedimentsamples using phosphoric acid yielded carbonate contentsof 0.03% and 0.08% (combined calcite and dolomite, inwt % CaCO3) for the Madeira and Amazon rivers, andfor the Solimoes River, respectively (Galy et al., 2007b).Note that these levels, which correspond to 0.004% and0.010% C, respectively, are close to the detection limit ofthe method and are negligible in terms of bias on thePOC content measured on non-decarbonated samples.Moreover, assuming a d13C value of ca. 0&for the carbon-ate component, the presence of such trace amounts of car-bonates in the sediment would induce a shift of only ca.
�0.15& on the bulk d13C value measured on suspendedload, which is still lower than the uncertainty stemmingfrom sample heterogeneity. For some bed sediment sam-ples, this shift could actually be of a few &.
APPENDIX B. ROCK-DERIVED POC CONTENT AND
AGE OF BIOSPHERIC POC FROM 14C-ACTIVITY
In river sediments, POC can be interpreted as a binarymixture of rock-derived OC and biospheric OC with dis-tinct 14C-activities (e.g. Blair et al., 2003; Galy et al.,2008a; Blair and Aller, 2012). The mixing equation forPOC is then:
Fmod ¼POCrock
POC� Fmodrock
þ POCbio
POC� Fmodbio
ð1Þ
Although it is known that the 14C activity of rock-derivedPOC is 0, both the 14C activity of the biospheric pool(Fmodbio
) and the contribution of rock-derived OC to totalPOC (POCrock ) are unknown. In order to determine thetwo unknowns, we use the method developed byGaly et al. (2008a). As Fmodrock
= 0 and POC =POCrock + POCbio:
ModernC ¼ POC � Fmodbio� POCrock � Fmodbio
ð2Þ
with ModernC ¼ POC � Fmod. Assuming that a set ofsamples has invariant Fmodbio
and POCrock and that theseend-members are well-mixed, these samples should definea linear trend in a diagram representing Modern C vs.
POC. These conditions are met by Amazon sediment sam-ples from a given location, regardless of the depth-profile orof the sampling date (Bouchez et al., 2010). The values ofPOCrock and Fmodbio
can both be determined from the slopeand intercept of each line in Fig. 4 (Galy et al., 2008a; Bou-chez et al., 2010; Galy and Eglinton, 2011). Results areshown in Table 3. To carry out this analysis, the POC con-tent measured after decarbonation, POCdecarb (“acid-insolu-ble”), was used.
APPENDIX C. SUPPLEMENTARY DATA
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.gca.2014.02.032.
REFERENCES
Aller R. C., Blair N. E., Xia Q. and Rude P. D. (1996)Remineralization rates, recycling, and storage of carbon inAmazon shelf sediments. Cont. Shelf Res. 16, 753–786.
Arnarson T. S. and Keil R. G. (2001) Organic–mineral interactionsin marine sediments studied using density fractionation and X-ray photoelectron spectroscopy. Org. Geochem. 32, 1401–1415.
Aufdenkampe A. K., Mayorga E., Hedges J. I., Llerena C., QuayP. D., Gudeman J., Krusche A. V. and Richey J. E. (2007)Organic matter in the Peruvian headwaters of the Amazon:compositional evolution from the Andes to the lowlandAmazon mainstem. Org. Geochem. 38, 337–364.
Aufdenkampe A. K., Mayorga E., Raymond P. A., Melack J. M.,Doney S. C., Alin S. R., Aalto R. A. and Yoo K. (2011)Riverine coupling of biogeochemical cycles between land,oceans, and atmosphere. Front. Ecol. Environ. 9, 53–60.
Battin T. J., Kaplan L. A., Findlay S., Hopkinson C. S., Marti E.,Packman A. I., Newbold J. D. and Sabater F. (2009) Biophys-ical controls on organic carbon fluxes in fluvial networks. Nat.
Geosci. 1, 95–100.Becker J. A., Bickle M. J., Galy A. and Holland T. J. B. (2008)
Himalayan metamorphic CO2 fluxes: quantitative constraintsfrom hydrothermal springs. Earth Planet. Sci. Lett. 265, 616–629.
Bendle J. A., Weijers J. W. H., Maslin M. A., Damste J. S. S.,Schouten S., Hopmans E. C., Boot C. S. and Pancost R. D.(2010) Major changes in glacial and Holocene terrestrialtemperatures and sources of organic carbon recorded in theAmazon fan by tetraether lipids. Geochem. Geophys. Geosyst.
11, 03308.Bird M. I., Haberle S. G. and Chivas A. R. (1994) Effect of altitude
on the carbon?isotope composition of forest and grassland soilsfrom Papua New Guinea. Global Biogeochem. Cycles 8(1), 13–22.
Blair N. E., Leithold E. L., Ford S. T., Peeler K. A., Holmes J. C.and Perkey D. W. (2003) The persistence of memory: the fate ofancient sedimentary organic carbon in a modern sedimentarysystem. Geochim. Cosmochim. Acta 67, 63–73.
Blair N. E., Leithold E. L. and Aller R. C. (2004) From bedrock toburial: the evolution of particulate organic carbon acrosscoupled watershed-continental margin systems. Marine Geol.
92, 141–156.Blair N. E. and Aller R. C. (2012) The fate of terrestrial organic
carbon in the marine environment. Annu. Rev. Marine Sci. 4,401–423.
Bock M. J. and Mayer L. M. (2000) Mesodensity organo-clayassociations in a near-shore sediment. Mar. Geol. 163, 65–75.

296 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298
Bonnet M., Barroux G., Martinez J.-M., Seyler F., Moreira-TurcqP., Cochonneau G., Melack J., Boaventura G. R., Maurice-Bourgoin L., Leon J., Roux E., Calmant S., Kosuth P., GuyotJ.-L. and Seyler P. (2007) Floodplain hydrology in an Amazonfloodplain lake (Lago Grande de Curuaı). J. Hydrol. 349, 18–30.
Bouchez J., Beyssac O., Galy V., Gaillardet J., France-Lanord C.,Maurice L. and Moreira-Turcq P. (2010) Oxidation of petro-genic organic carbon in the Amazon floodplain as a source ofatmospheric CO2. Geology 38, 255–258.
Bouchez J., Metivier F., Lupker M., Maurice L., Perez M.,Gaillardet J. and France-Lanord C. (2011a) Prediction ofdepth-integrated fluxes of suspended sediment in the AmazonRiver: particle aggregation as a complicating factor. Hydrol.
Processes 25, 778–794.Bouchez J., Gaillardet J., France-Lanord C., Maurice L. and
Dutra-Maia P. (2011b) Grain-size controls of river suspendedsediment geochemistry: insights from Amazon River depth-profiles. Geophys. Geochem. Geosyst. 12, GC00380.
Bouchez J., Lupker M., Gaillardet J., France-Lanord C. andMaurice L. (2011c) How important is it to integrated riverinesuspended sediment chemical composition with depth? Cluesfrom Amazon River depth-profiles. Geochim. Cosmochim. Acta
75, 6955–6970.Brunauer S., Emmet P. H. and Teller E. J. (1938) Adsorption of
gases in multimolecular layers. Am. Chem. Soc. 60.Burdige D. J. (2005) Burial of terrestrial organic matter in marine
sediments: a re-assessment. Global Biogeochem. Cycles 19,GB4011.
Callede J., Kosuth P., Guyot J.-L. and Guimaraes V. (2000)Discharge determination by acoustic Doppler current profilers(ADCP): a moving bottom error correction method and itsapplication on the river Amazon at Obidos. Hydrol. Sci. J. 45,911–924.
Carlotto V., Gil W., Cardenas J. and Chavez R. (1996) Geologıa delos cuadrangulos de Urubamba y Calca. Instituto GeologicoMinero y Metalurgico Boletin, 65 Serie A, maps 27-r y 27-s,scale 1:100,000.
Clark K. E., Hilton R. G., West A. J., Malhi Y., Grocke D. R.,Bryant C. L., Ascough P. L., Robles Caceres A. and New M.(2013) New views on old carbon in the Amazon River: insightfrom the source of organic carbon eroded from the PeruvianAndes. Geochem. Geophys. Geosyst. 14, 1644–1659.
Curry K. J., Bennett R. H., Mayer L. M., Curry A., Abril M.,Biesiot P. M. and Hulbert M. H. (2007) Direct visualization ofclay microfabric signatures driving organic matter preservationin fine-grained sediment. Geochim. Cosmochim. Acta 71, 1709–1720.
Dosseto A., Bourdon B., Gaillardet J., Allegre C. J. and Filizola N.(2006a) Time-scale and conditions of weathering under tropicalclimate: study of the Amazon basin with U-series. Geochim.
Cosmochim. Acta 70, 71–89.Dosseto A., Bourdon B., Gaillardet J., Maurice-Bourgoin L. and
Allegre C. J. (2006b) Weathering and transport of sediments inthe Bolivian Andes: time constraints from uranium-seriesisotopes. Earth Planet. Sci. Lett. 248, 759–771.
Dunne T., Mertes L. A. K., Meade R. H., Richey J. E. andForsberg B. R. (1998) Exchanges of sediment between the floodplain and channel of the Amazon River in Brazil. Geol. Soc.
Am. Bull. 110, 450–467.Eckard R. S., Hernes P. J., Bergamaschi B. A., Stepanauskas R.
and Kendall C. (2007) Landscape scale controls on the vascularplant component of dissolved organic carbon across a fresh-water delta. Geochim. Cosmochim. Acta 71, 5968–5984.
Ellis E. E., Richey J. E., Aufdenkampe A. K., Krusche A. V., QuayP. D., Salimon C. and Brandao da Cunha H. (2012) Factors
controlling water-column respiration in rivers of the central andsouthwestern Amazon Basin. Limnol. Oceanogr. 57(2), 527–540.
Filizola N. and Guyot J.-L. (2003) The use of Doppler technologyfor suspended sediment discharge determinations on the RiverAmazon at Obidos. Hydrol. Sci. J. 49, 143–153.
Filizola N. and Guyot J.-L. (2009) Suspended sediment yield in theAmazon basin: an assessment using the Brazilian national dataset. Hydrol. Proc. 23, 3207–3215.
Gaillardet J., Dupre B., Allegre C. J. and Negrel P. (1997)Chemical and physical denudation in the Amazon River Basin.Chem. Geol. 142, 141–173.
Gaillardet J., Dupre B., Louvat P. and Allegre C. J. (1999) Globalsilicate weathering and CO2 consumption rates deduced fromthe chemistry of large rivers. Chem. Geol. 159, 3–30.
Galy V., France-Lanord C., Beyssac O., Faure P., Kudrass H. andPalhol F. (2007a) Efficient carbon burial in the Bengal fansustained by the Himalayan erosional system. Nature 450, 407–410.
Galy V., Bouchez J. and France-Lanord C. (2007b) Determinationof Total Organic Carbon content and d13C in carbonate-richdetrital sediments. Geostand. Geoanal. Res. 31, 199–207.
Galy V., Beyssac O., France-Lanord C. and Eglinton T. (2008a)Recycling of graphite during Himalayan erosion: a geologicalstabilisation of carbon in the crust. Science 322, 943–945.
Galy V., France-Lanord C. and Lartiges B. (2008b) Loading andfate of particulate organic carbon from the Himalaya to theGanga–Brahmaputra delta. Geochim. Cosmochim. Acta 72,1767–1787.
Galy V. and Eglinton T. (2011) Protracted storage of biosphericcarbon in the Ganges–Brahmaputra basin. Nat. Geosci. 4, 843–847.
Garrels R. M. and Mackenzie F. T. (1971) Evolution of Sedimen-
tary Rocks. Norton, New York.Gibbs R. J. (1967) Amazon River system: environmental factors
that control its dissolved and suspended load. Science 156,1734–1737.
Guyot J.-L., Jouanneau J.-M., Suares L., Boaventura G. R.,Maillet N. and Lagane C. (2007) Clay mineral composition ofriver sediments in the Amazon Basin. Catena 71, 340–356.
Hamilton S. K., Sippel S. J. and Melack J. M. (2004) Seasonalinundation patterns in two large savanna floodplains of SouthAmerica: the Llanos de Moxos (Bolivia) and the Llanos delOrinoco (Venezuela and Colombia). Hydrol. Processes 18,2103–2116.
Hedges J. I., Clarke W. A., Quay P. D., Richey J. E., Devol A. H.and Santos U. de M. (1986a) Compositions and fluxes ofparticulate organic material in the Amazon River. Limnol.
Oceanogr. 31, 717–738.Hedges J. I., Ertel J. R., Quay P. D., Grootes P. M., Richey J. E.,
Devol A. H., Farwell G. W., Schmidt F. W. and Salati E.(1986b) Organic carbon-14 in the Amazon River System.Science 231, 1129–1131.
Hedges J. I., Cowie G. L., Richey J. E., Quay P., Benner R., StromM. and Forsberg B. R. (1994) Origins and processing of organicmatter in the Amazon River as indicated by carbohydrates andamino acids. Limnol. Oceanogr. 39, 743–761.
Hedges J. I. and Keil R. G. (1995) Sedimentary organic matterpreservation: an assessment and speculative synthesis. Mar.
Chem. 49, 81–115.Hedges J. I., Mayorga E., Tsamakis E., McClain M. E., Aufdenk-
ampe A., Quay P., Richey J. E., Benner R., Opsahl S., Black B.,Pimentel T., Quintanilla J. and Maurice L. (2000) Organicmatter in Bolivian tributaries of the Amazon River: a compar-ison to the lower mainstream. Limnol. Oceanogr. 45, 1449–1466.
Hess L. L., Melack J. M., Novo E. M. L. M., Barbosa C. C. F. andGastil M. (2003) Dual-season mapping of wetland inundation

J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298 297
and vegetation for the central Amazon basin. Remote Sens.
Environ. 87, 404–428.Hilton R. G., Galy A. and Hovius N. (2008) Riverine particulate
organic carbon from an active mountain belt: importance oflandslides. Global Biogeochem. Cycles 22, GB1017.
Hilton R. G., Galy A., Hovius N., Horng M.-J. and Chen H.(2010) The isotopic composition of particulate organic carbonin mountain rivers of Taiwan. Geochim. Cosmochim. Acta 74,3164–3181.
Hilton R. G., Galy A., Hovius N., Horng M.-J. and Chen H.(2011) Efficient transport of fossil organic carbon to the oceanby steep mountain rivers: an orogenic carbon sequestrationmechanism. Geology 39(1), 71–74.
Hilton R. G., Galy A., Hovius N., Kao S.-J., Horng M.-J. andChen H. (2012) Climatic and geomorphic controls on theerosion of terrestrial biomass from subtropical mountain forest.Global Biogeochem. Cycles 26, GB3014.
Houghton R. A., Lawrence K. T., Hackler J. L. and Brown S.(2001) The spatial distribution of forest biomass in the BrazilianAmazon: a comparison of estimates. Global Change Biol. 7,731–746.
Kastner T. P. and Goni M. A. (2003) Constancy in the vegetationof the Amazon Basin during the late Pleistocene: evidence fromthe organic matter composition of Amazon deep sea fansediments. Geology 31(4), 291–294.
Keil R. G., Tsamakis E., BorFuh C., Giddings J. C. and Hedges J.I. (1994) Mineralogical and textural controls on the organiccomposition of coastal marine sediments: hydrodynamic sepa-ration using SPLITT fractionation. Geochim. Cosmochim. Acta
58, 879–893.Keil R. G., Mayer L. M., Quay P. D., Richey J. E. and Hedges J. I.
(1997) Loss of organic matter from riverine particles in deltas.Geochim. Cosmochim. Acta 61, 1507–1511.
Kuehl S. A., DeMaster D. J. and Nittrouer C. A. (1986) Nature ofsediment accumulation on the Amazon continental shelf. Cont.
Shelf Res. 6(1-2), 209–225.Latrubesse E. M. (2008) Patterns of anabranching channels: the
ultimate end-member adjustment of mega rivers. Geomorphol-
ogy 101, 130–145.Ludwig W., Probst J.-L. and Kempe S. (1996) Predicting the
oceanic input of organic carbon by continental erosion. Global
Biogeochem. Cycles 10(1), 23–41.Maurice-Bourgoin L., Bonnet M.-P., Martinez J.-M., Kosuth P.,
Cochonneau G., Moreira-Turcq P., Guyot J.-L., Vauchel P.,Filizola N. and Seyler P. (2007) Temporal dynamics of waterand sediments exchanges between the Curuaı floodplain and theAmazon River, Brazil. J. Hydrol. 335, 140–156.
Mayer L. M. (1994a) Surface area control of organic carbonaccumulation in continental shelf sediments. Geochim. Cosmo-
chim. Acta 58, 1271–1284.Mayer L. M. (1994b) Relationships between mineral surfaces and
organic carbon concentrations in soils and sediments. Chem.
Geol. 114, 347–363.Mayer L. M. (1999) Extent of coverage of mineral surfaces by
organic matter in marine sediments. Geochim. Cosmochim. Acta
63, 207–215.Mayer L. M., Schick L. L., Hardy K. R., Wagai R. and McCarthy
J. (2004) Organic matter in small mesopores in sediments andsoils. Geochim. Cosmochim. Acta 68, 3863–3872.
Mayorga E., Aufdenkampe A. K., Masiello C. A., Krusche A. V.,Hedges J. I., Quay P. D., Richey J. E. and Brown T. A. (2005)Young organic matter as a source of carbon dioxide outgassingfrom Amazonian rivers. Nature 436, 538–541.
Moreira-Turcq P., Seyler P., Guyot J.-L. and Etcheber H. (2003)Exportation of organic carbon from the Amazon River and itsmain tributaries. Hydrol. Processes 17, 1329–1344.
Moreira L. S., Moreira-Turcq P., Turcq B., Caquineau S. andCordeiro R. C. (2012) Paleoclimatic changes and carbonaccumulation in the Santa Ninha, and Amazonian floodplainlake. J. Paleolimnol. 48, 339–350.
Moreira-Turcq P., Bonnet M.-P., Amorim M., Bernardes M.,Lagane C., Maurice L., Perez M. and Seyler P. (2013) Seasonalvariability in concentration, composition, age, and fluxes ofparticulate organic carbon exchanged between the floodplainand Amazon River. Global Biogeochem. Cycles 27, GBC20022.
Mortillaro J. M., Abril G., Moreira-Turcq P., Sobrinho R. L.,Perez M. and Meziane T. (2011) Fatty acid and stable isotope(d13C, d15N) signatures of particulate organic matter in thelower Amazon River: seasonal contrasts and connectivitybetween floodplain lakes and the mainstem. Org. Geochem.
42, 1159–1168.Nebel G., Dragsted J. and Salazar Vega A. (2001) Litter fall,
biomass and net primary production in flood plain forests in thePeruvian Amazon. Forest Ecol. Manag. 150, 93–102.
Perez M. A. P., Moreira-Turcq P., Gallard H., Allard T. andBenedetti M. (2011) Dissolved organic matter dynamic in theAmazon basin: sorption by mineral surfaces. Chem. Geol. 286,158–168.
Putzer, H. (1984) The geological evolution of the Amazon Basinand its mineral resources. In: The Amazon: Limnology and
Landscape Ecology of a Mighty Tropical River and its Basin (ed.H. Sioli). Dordrecht, The Netherlands, pp. 15–46.
Quay P. D., Wilbur D. O., Richey J. E., Hedges J. I., Devol A. H.and Victoria R. (1992) Carbon cycling in the Amazon River:implications from the 13C compositions of particles and solutes.Limnol. Oceanogr. 37, 857–871.
Ransom B., Bennett R. H., Baerwald R. and Shea K. (1997) TEMstudy of in situ organic matter on continental margins:occurrence and the monolayer hypothesis. Mar. Geol. 138, 1–9.
Ransom B., Kim D., Kastner M. and Wainwright S. (1998)Organic matter preservation on continental slopes: importanceof mineralogy and surface area. Geochim. Cosmochim. Acta 62,1329–1345.
Richey J. E., Mertes L., Dunne T., Victoria R., Forsberg B. R.,Tancredi A. and Oliveira E. (1989) Source and routing of theAmazon River flood wave. Global Biogeochem. Cycles 3, 191–204.
Richey J. E., Hedges J. I., Devol A. H., Quay P. D., Victoria R.,Martinelli L. and Forsberg B. R. (1990) Biogeochemistry ofcarbon in the Amazon River. Limnol. Oceanogr. 35, 352–371.
Richey J. E., Melack J. M., Aufdenkampe A. K., Ballester V. M.and Hess L. L. (2002) Outgassing from Amazonian rivers anwetlands as a large tropical source of atmospheric CO2. Nature
416, 617–620.Rosenheim B. E. and Galy V. (2013) Direct measurement of
riverine particulate organic carbon age structure. Geophys. Res.
Lett. 39, L19703.Sarmiento J. L. and Sundquist E. T. (1992) Revised budget for the
oceanic uptake of anthropogenic carbon dioxide. Nature 356,589–592.
Schlunz B., Schneider R. R., Muller P. J., Showers W. J. and WeferG. (1999) Terrestrial organic carbon accumulation on theAmazon deep sea fan during the last glacial sea level low stand.Chem. Geol. 159, 263–281.
Spencer R. G. M., Hernes P. J., Aufdenkampe A. K., Baker A.,Gulliver P., Stubbins A., Aiken G. R., Dyda R. Y., Butler K.D., Mwamba V. L., Mangangu A. M., Wabakanghanzi J. N.and Six J. (2012) An initial investigation into the organic matterbiogeochemistry of the Congo River. Geochim. Cosmochim.
Acta 84, 614–627.Stuiver M. and Polach H. A. (1977) Reporting of 14C data.
Radiocarbon 19(3), 355–363.

298 J. Bouchez et al. / Geochimica et Cosmochimica Acta 133 (2014) 280–298
Torn M. S., Trumbore S. E., Chadwick O. A., Vitousek P. M. andHendricks D. M. (1997) Mineral control of soil organic carbonstorage and turnover. Nature 389, 170–173.
Townsend-Small A., Noguera J. L., McClain M. E. and Brandes J.A. (2007) Radiocarbon and stable isotope geochemistry oforganic matter in the Amazon headwaters, Peruvian Andes.Global Biogeochem. Cycles 21, GB2029.
Trumbore S. E. (1993) Comparison of carbon dynamics in tropicaland temperate soils using radiocarbon measurements. Global
Biogeochem. Cycles 7(2), 275–290.
West A. J., Lin C.-W., Lin T.-C., Hilton R. G., Tanaka M., ChangC.-T., Lin K.-C., Galy A., Sparkes R. and Hovius N. (2011)Mobilization and transport of coarse woody debris by largestorms. Limnol. Oceanogr. 56, 77–85.
Wissmar R. C., Richey J. E., Stallard R. F. and Edmond J. M.(1981) Plankton metabolism and carbon processes in theAmazon River, its tributaries, and floodplain waters, Peru–Brazil, May–June 1977. Ecology 62, 1622–1633.
Associate editor: Peter Hernes

Supplementary Material to Bouchez et al. (2014): depth-integrated suspended POCfluxes and concentrations
POC content in Amazon River suspended load varies within single depth-profiles(Fig. 2). This variability has to be accounted for when estimating POC fluxes and con-centrations. In order to calculate the instantaneous depth-integrated POC fluxes fromdepth-profiles in the Amazon River, the method developed by Bouchez et al. (2011a,b,c)is applied here. The depth-integration method is based on a fit of the depth profilesof suspended particulate matter (SPM) concentration by the Rouse model (e.g. Rouse,1950). The Rouse model is applied to each grain size fraction separately, and the result-ing modeled SPM profiles are extrapolated laterally to the entire channel cross-section,such that SPM is predicted for each grain size at any point in the cross section (Bouchezet al., 2011a). The sediment grain size distribution and the associated parameters (suchas the median, D10, D90) can then be calculated at any point in the channel cross-section.Using the linear correlation between the grain size distribution D90 and the Al/Si ra-tio (Bouchez et al., 2011b), the Al/Si ratio can also be calculated over the cross-section(Bouchez et al., 2011c). We use the linear POC-Al/Si relationships (specific to each riverand to each sampling campaign in the case of the Solimões and the Amazon River, Fig.6) to model the POC content of suspended sediment at any point in the channel cross-section. Combining water velocity data retrieved in the field from ADCP with modelledSPM concentration and POC content over the whole cross-section, the local flux of OCcan be predicted at any point in the cross-section. Finally, spatial integration of the POCflux along the vertical and lateral directions allows estimation of the POC flux integratedover the whole channel (Tab. S1). The uncertainty on these depth-integrated POC fluxesis ca. 40% (Bouchez et al., 2011c). These estimates do not take into account the POC fluxcarried by bed sediments, which is thought to be only ca. 1% of the total sediment flux inthe Amazon (Dunne et al., 1998). Importantly, they only reflect instantaneous fluxes atthe times of sampling, since the depth-profiles characteristics (particularly changes wa-ter velocity profiles, SPM concentration and grain size, and OC-loading) vary greatlyduring the hydrological cycle of the river.
The largest instantaneous suspended POC fluxes are obtained for the Amazon atÓbidos (397 and 604 kg.s−1 in June 2005 and March 2006, respectively), which is thesite at which the largest sediment flux is also obtained. The smallest were estimatedto be carried by the Madeira (14 and 132 kg.s−1 in June 2005 and March 2006, respec-tively). Altogether, a reasonable agreement between our estimates and those of Richeyet al. (1990) demonstrates the accuracy of the method for estimating depth-integratedfluxes of POC.
Dividing depth-integrated POC fluxes by depth-integrated sediment fluxes, depth-integrated POC concentrations can be estimated (Tab. S1). Rather constant depth-integratedPOC concentrations are obtained for the Amazon at Iracema and Óbidos, while signif-icant temporal variations are observed for the Solimões and the Madeira. These valuesare compared in Fig. S1 with the average of POC contents measured at the channel sur-face, for each sampling site and date. The relative uncertainty on these depth-integratedPOC concentrations is estimated at ca. 15% (Bouchez et al., 2011c). In most cases, depth-integrated POC contents are ca. up to twice as low as those measured at the channelsurface.
1

River Sampling site Sampling date Depth-integrated (Al/Si)int Depth-integrated POCint (%) Depth-integrated
SPM flux (kg/s) POC flux (kg/s) rock-derived POC flux (kg/s)
Solimões Manacapuru June 2005 17400 0.212 168 0.97 8
Solimões Manacapuru March 2006 22800 0.209 155 0.68 12
Amazon Iracema June 2005 17000 0.268 217 1.28
Amazon Iracema March 2006 13100 0.332 153 1.17
Madeira Foz Madeira June 2005 1300 0.392 14 1.08 0.7
Madeira Foz Madeira March 2006 19400 0.248 132 0.68 10
Amazon Óbidos June 2005 61300 0.220 604 0.99 68
Amazon Óbidos March 2006 39700 0.327 397 1.00 44Table 1Table S1: Depth-integrated instantaneous fluxes of suspended sediments and POC for the Solimões, Madeiraand Amazon at Iracema and Óbidos, calculated using Rouse models (Bouchez et al., 2011c) and the correlationbetween POC content and Al/Si (Fig. 6). The method was not applied to the Amazon at Parintins becauseof the lack of sufficiently reliable ADCP water velocity data. Depth-integrated Al/Si (Al/Siint) were calcu-lated from depth-integrated Al and Si fluxes reported by Bouchez et al. (2011c). Depth-integrated POC con-centration (POCint) were obtained by dividing depth-integrated POC fluxes by depth-integrated suspendedsediment fluxes. The overall relative uncertainty on depth-integrated fluxes is ca. 40% and that on depth-integrated concentration is ca. 15% (Bouchez et al., 2011c).
2

0.0
0.5
1.0
1.5
2.0
0.0 0.5 1.0 1.5 2.0Surface POC content (%)
Dep
th-in
tegr
ated
PO
C c
onte
nt (%
)
1:1
1:2
SolimõesMadeira
IracemaObidosMarch 2006June 2005
Figure S1: Comparison between depth-integrated POC content and POC content esti-mated as an average of measurements on channel surface suspended sediment samples,for the Solimões, Madeira and Amazon rivers.
3

References
Bouchez J., Métivier F., Lupker M., Maurice L., Perez M., Gaillardet J. and France-LanordC. (2011a) Prediction of depth-integrated fluxes of suspended sediment in the Ama-zon River: particle aggregation as a complicating factor. Hydrol. Proc., 25:778-794.
Bouchez J., Gaillardet J., France-Lanord C., Maurice L. and Dutra-Maia P. (2011b) Grain-size controls of river suspended sediment geochemistry: insights from Amazon Riverdepth-profiles. Geophys. Geochem. Geosystems, 12:GC00380.
Bouchez J., Lupker M., Gaillardet J., France-Lanord C. and Maurice L. (2011c) Howimportant is it to integrated riverine suspended sediment chemical composition withdepth? Clues from Amazon River depth-profiles. Gochim. Cosmochim. Acta, 75:6955-6970.
Dunne T., Mertes L.A.K., Meade R.H., Richey J.E. and Forsberg B.R. (1998) Exchangesof sediment between the flood plain and channel of the Amazon River in Brazil. Geol.Soc. Am. Bull., 110:450-467.
Richey J.E., Hedges J.I., Devol A.H., Quay P.D., Victoria R., Martinelli L. and ForsbergB.R. (1990) Biogeochemistry of carbon in the Amazon River. Limnol. Oceanogr., 35:352-371.
Rouse H. (1950) Engineering Hydraulics. Wiley, New York.
4




















