
Transfer of leaf rust and stripe rust resistance from Aegilops umbellulata Zhuk. to bread wheat (...
11
RESEARCH ARTICLE Transfer of leaf rust and stripe rust resistance from Aegilops umbellulata Zhuk. to bread wheat (Triticum aestivum L.) Parveen Chhuneja Satinder Kaur R. K. Goel M. Aghaee-Sarbarzeh M. Prashar H. S. Dhaliwal Received: 29 June 2007 / Accepted: 8 October 2007 / Published online: 31 October 2007 Ó Springer Science+Business Media B.V. 2007 Abstract Aegilops umbellulata acc. 3732, an excel- lent source of resistance to major wheat diseases, was used for transferring leaf rust and stripe rust resistance to cultivated wheat. An amphiploid between Ae. umbellulata acc. 3732 and Triticum durum cv. WH890 was crossed with cv. Chinese Spring Ph I to induce homoeologous pairing between Ae. umbellulata and wheat chromosomes. The F 1 was crossed to the susceptible Triticum aestivum cv. ‘WL711’ and leaf rust and stripe rust resistant plants were selected among the backcross progenies. Homozygous lines were selected and screened against six Puccinia triticina and four Puccinia striiformis f. sp. tritici pathotypes at the seedling stage and a mixture of prevalent pathotypes of both rust pathogens at the adult plant stage. Genomic in situ hybridization in some of the selected introgression lines detected two lines with complete Ae. umbellulata chromosomes. Depending on the rust reactions and allelism tests, the introgres- sion lines could be classified into two groups, comprising of lines with seedling leaf rust resistance gene Lr9 and with new seedling leaf rust and stripe rust resistance genes. Inheritance studies detected an additional adult plant leaf rust resistance gene in one of the introgression lines. A minimum of three putatively new genes—two for leaf rust resistance (LrU1 and LrU2) and one for stripe rust resistance (YrU1) have been introgressed into wheat from Ae. umbellulata. Two lines with no apparent linkage drag have been identified. These lines could serve as sources of resistance to leaf rust and stripe rust in breeding programs. Keywords Aegilops umbellulata Á Homoeologous pairing Á Leaf rust Á Puccinia striiformis Á Puccinia triticina Á Stripe rust Á Triticum aestivum Á Wheat Á Wide hybridization Introduction Leaf rust caused by Puccinia triticina Eriks. (Pt) and stripe rust caused by Puccinia striiformis Westend f. sp. tritici (Pst) are the most important foliar diseases P. Chhuneja (&) Á S. Kaur Á R. K. Goel Á M. Aghaee-Sarbarzeh Á H. S. Dhaliwal Department of Plant Breeding, Genetics and Biotechnology, Punjab Agricultural University, Ludhiana 141 004, India e-mail: [email protected] M. Prashar Directorate of Wheat Research, Regional Research Station, Shimla, India Present Address: M. Aghaee-Sarbarzeh Agricultural Organization of Kermanshah, Kermanshah, Iran Present Address: H. S. Dhaliwal Indian Institute of Technology, Roorkee, India 123 Genet Resour Crop Evol (2008) 55:849–859 DOI 10.1007/s10722-007-9289-3
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Transfer of leaf rust and stripe rust resistance from Aegilops umbellulata Zhuk. to bread wheat (...

RESEARCH ARTICLE
Transfer of leaf rust and stripe rust resistance from Aegilopsumbellulata Zhuk. to bread wheat (Triticum aestivum L.)
Parveen Chhuneja Æ Satinder Kaur Æ R. K. Goel Æ M. Aghaee-Sarbarzeh ÆM. Prashar Æ H. S. Dhaliwal
Received: 29 June 2007 / Accepted: 8 October 2007 / Published online: 31 October 2007
� Springer Science+Business Media B.V. 2007
Abstract Aegilops umbellulata acc. 3732, an excel-
lent source of resistance to major wheat diseases, was
used for transferring leaf rust and stripe rust resistance
to cultivated wheat. An amphiploid between Ae.
umbellulata acc. 3732 and Triticum durum cv.
WH890 was crossed with cv. Chinese Spring PhI to
induce homoeologous pairing between Ae. umbellulata
and wheat chromosomes. The F1 was crossed to the
susceptible Triticum aestivum cv. ‘WL711’ and leaf
rust and stripe rust resistant plants were selected among
the backcross progenies. Homozygous lines were
selected and screened against six Puccinia triticina
and four Puccinia striiformis f. sp. tritici pathotypes at
the seedling stage and a mixture of prevalent
pathotypes of both rust pathogens at the adult plant
stage. Genomic in situ hybridization in some of the
selected introgression lines detected two lines with
complete Ae. umbellulata chromosomes. Depending
on the rust reactions and allelism tests, the introgres-
sion lines could be classified into two groups,
comprising of lines with seedling leaf rust resistance
gene Lr9 and with new seedling leaf rust and stripe rust
resistance genes. Inheritance studies detected an
additional adult plant leaf rust resistance gene in one
of the introgression lines. A minimum of three
putatively new genes—two for leaf rust resistance
(LrU1 and LrU2) and one for stripe rust resistance
(YrU1) have been introgressed into wheat from Ae.
umbellulata. Two lines with no apparent linkage drag
have been identified. These lines could serve as sources
of resistance to leaf rust and stripe rust in breeding
programs.
Keywords Aegilops umbellulata �Homoeologous pairing � Leaf rust �Puccinia striiformis � Puccinia triticina �Stripe rust � Triticum aestivum � Wheat �Wide hybridization
Introduction
Leaf rust caused by Puccinia triticina Eriks. (Pt) and
stripe rust caused by Puccinia striiformis Westend f.
sp. tritici (Pst) are the most important foliar diseases
P. Chhuneja (&) � S. Kaur � R. K. Goel �M. Aghaee-Sarbarzeh � H. S. Dhaliwal
Department of Plant Breeding, Genetics and
Biotechnology, Punjab Agricultural University,
Ludhiana 141 004, India
e-mail: [email protected]
M. Prashar
Directorate of Wheat Research, Regional Research
Station, Shimla, India
Present Address:M. Aghaee-Sarbarzeh
Agricultural Organization of Kermanshah, Kermanshah,
Iran
Present Address:H. S. Dhaliwal
Indian Institute of Technology, Roorkee, India
123
Genet Resour Crop Evol (2008) 55:849–859
DOI 10.1007/s10722-007-9289-3

of wheat the worldwide. Genetic resistance is the
most effective, economical and environmentally
friendly means to reduce losses due to the rust
diseases. More than 58 leaf rust and 40 stripe rust
resistance genes have been designated so far
(McIntosh et al. 2005; Kuraparthy et al. 2007a, b),
the most of which condition hypersensitive reactions
and interact with respective pathogens in a gene-
for-gene fashion (Flor 1942). As the resistance
conferred by single major resistance genes is not
durable, hence a constant search for new sources of
rust resistance is necessary to combat the highly
variable rust pathogens. In addition, the replacement
of highly variable land races by high yielding pure-
line varieties in many parts of the world has narrowed
down the genetic base of modern wheats. Wild
progenitor and non-progenitor species of wheat are
highly valuable source of additional resistance genes
(Dvorak 1977; Sharma and Gill 1983; Hammer 1985;
Gale and Miller 1987; Jiang et al. 1994; Singh et al.
1998, 2007). Many genes conferring resistance to rust
diseases, powdery mildew, cereal cyst nematode and
insect-pests were transferred from Aegilops species
into wheat (Jiang et al. 1994; Friebe et al. 1996;
Romero et al. 1998; Dhaliwal et al. 2003; Marais
et al. 2005a). Some of these genes transferred from
distantly related species have been exploited com-
mercially but others seem to be associated with yield
penalty due to the linkage drag (Young and Tanksley
1989).
Several strategies have been used for transferring
the alien segments that are smaller than complete
chromosome arms from non-progenitor wild species
into wheat. Sears (1956) transferred a leaf rust
resistance gene (Lr9) from Ae. umbellulata Zhuk. to
wheat through irradiation. Masoudi-Nejad et al.
(2002) used gametocidal genes for the transfer of
alien chromosome segments to wheat. Alien genes
from non-progenitor species have also been trans-
ferred to the wheat through induced homoeologous
chromosome pairing between the wheat and the
alien chromosomes (Jiang et al. 1994; Friebe et al.
1996) by using mutant or null alleles of Ph1 gene
(Sears 1972, 1981) or by using PhI, an epistatic
inhibitor of Ph1 gene from Aegilops speltoides
Tausch (Riley et al. 1968; Chen et al. 1994; Aghaee
et al. 2000). The transfers made by homoeologous
recombination are genetically compensating and the
size of the transferred alien segment is often
proportional to the linkage drag (Jiang et al. 1994;
Friebe et al. 1996). The identification and charac-
terization of cytologically undetectable primary
recombinants in wheat-Aegilops geniculata Roth
(syn. Aegilops ovata auct. non L.) introgression
carrying Lr57 and Yr40 (Kuraparthy et al. 2007a)
and wheat-Aegilops triuncialis L. introgression car-
rying Lr58 (Kuraparthy et al. 2007b), demonstrated
the feasibility of transfer of small alien desirable
segments without linkage drag. Resistance for biotic
and abiotic stresses and some quality traits have
been introgressed from progenitor and non-progen-
itor species into T. durum Desf. also using various
chromosome engineering and molecular techniques
(review by Ceoloni and Jauhar 2006). T. durum has
been reported to be equally amenable to alien
introgressions but relatively less buffered for toler-
ating gene transfers as compared to T. aestivum.
Ae. umbellulata, a non-progenitor diploid species
with the UU genome, was earlier found to be an
excellent source of resistance for various diseases
(Valkoun et al. 1985; Dhaliwal et al. 1991; Singh and
Dhaliwal 2000). This paper reports the introgression
of resistance to leaf rust and stripe rust from Ae.
umbellulata acc. 3732 to the hexaploid wheat.
Materials and methods
Plant material
A leaf rust and stripe rust resistant Ae. umbellulata
acc. 3732 from USSR, supplied by Dr J. P. Gustafson,
University of Missouri, Columbia, USA, was used for
developing a synthetic amphiploid with a susceptible
T. durum cv. WH890. The amphiploid T. durum
WH890-Ae. umbellulata acc. 3732 was crossed with
cv. Chinese Spring {CS (PhI)} (Chen et al. 1994) to
induce homoeologous pairing (Fig. 1). The resultant
F1 plants were crossed with a leaf rust and stripe rust
susceptible genotype, WL711(NN), which has kr
(crossability) alleles and is a near-isogenic line of
T. aestivum L. cv. WL711. The resistant F1 plants
from the cross T. durum WH890-Ae. umbellulata//
CSPh1/3/WL711(NN) were selfed and backcrossed to
WL711(NN) to recover the recurrent genotype. Some
of the selected BC1F1 plants were selfed to generate
BC1F6–7 and others were backcrossed to generate
BC2F5. In the segregating generations, selection for
850 Genet Resour Crop Evol (2008) 55:849–859
123

leaf rust resistance was carried out by screening the
genetically enhanced material at the seedling stage
against two most virulent and prevalent Pt pathotypes
(121R63-1 and 21R5). The same plants were
screened for terminal disease severity for stripe rust
and leaf rust at adult plant stage in the field at the
Punjab Agricultural University (PAU), Ludhiana,
India.
Rust screening
Rust tests on homozygous F8, BC1F6–7 and BC2F5
wheat-Ae. umbellulata introgression lines (ILs) along
with the parental lines were performed using five Pt
pathotypes viz. IR5, 109R31-1, 121R63-1, 21R5 and
253R31 (Lr19 virulent) and four Pst pathotypes viz.
46S102, 47S103, 46S119 (Yr9 virulent) and 78S84
(Yr27 virulent) at the two-leaf seedling stage
(Table 1). A leaf rust resistance gene Lr9 was
transferred from Ae. umbellulata by Sears (1956).
To determine whether the leaf rust resistance gene(s)
in the wheat-Ae. umbellulata ILs were different from
Lr9, these ILs were also tested with a Lr9 virulent Pt
pathotype, 121R127 at the Regional Research
Station, Shimla, India. The seedlings were inoculated
Fig. 1 Schematic representation of the crossing strategy
adopted for transferring leaf and stripe rust resistance genes
from Ae. umbellulata to hexaploid wheat T. aestivum cv.
WL711 through induced homoeologous pairing using Chinese
Spring stock with homoeologous pairing inducer gene, PhIgene (Ph inhibitor). T. aestivum cv. WL711 (NN) with kr(crossability) alleles was used as recipient parent
Ta
ble
1A
vir
ule
nce
/vir
ule
nce
afo
rmu
lae
of
leaf
rust
(P.
trit
icin
a)
and
stri
pe
rust
(P.
stri
ifo
rmis
)p
ath
oty
pes
use
dfo
rth
esc
reen
ing
of
wh
eat-
Ae.
um
bel
lula
tain
tro
gre
ssio
nli
nes
Pat
ho
typ
eA
vir
ule
nce
Vir
ule
nce
P.
trit
icin
a
IR5
Lr1
,L
r9,
Lr1
9,
Lr2
0,
Lr2
4,
Lr2
5L
r2,
Lr3
,L
r10,
Lr1
1,
Lr1
4,
Lr1
5,
Lr1
6,
Lr1
7,
Lr1
8,
Lr2
1,
Lr2
3,
Lr2
6
10
9R
31
-1L
r9,
Lr1
9,
Lr2
4,
Lr2
5,
Lr2
6L
r1,
Lr2
,L
r3,
Lr1
0,
Lr1
1,
Lr1
4,
Lr1
5,
Lr1
6,
Lr1
7,
Lr1
8,
Lr2
0,
Lr2
1,
Lr2
3
12
1R
63
-1L
r9,
Lr1
9,
Lr2
4,
Lr2
5L
r1,
Lr2
,L
r3,
Lr1
0,
Lr1
1,
Lr1
4,
Lr1
5,
Lr1
6,
Lr1
7,
Lr1
8,
Lr2
0,
Lr2
1,
Lr2
3,
Lr2
6
12
1R
12
7L
r19,
Lr2
4,
Lr2
5L
r1,
Lr2
,L
r3,
Lr9
,L
r10,
Lr1
1,
Lr1
4,
Lr1
5,
Lr1
6,
Lr1
7,
Lr1
8,
Lr2
0,
Lr2
1,
Lr2
3,
Lr2
6
21
R5
Lr9
,L
r15,
Lr1
9,
Lr2
4,
Lr2
5L
r1,
Lr2
,L
r3,
Lr1
0,
Lr1
1,
Lr1
4,
Lr1
6,
Lr1
7,
Lr1
8,
Lr2
0,
Lr2
1,
Lr2
3,
Lr2
6
25
3R
31
Lr9
,L
r24,
Lr2
5L
r1,
Lr2
,L
r3,
Lr1
0,
Lr1
1,
Lr1
6,
Lr1
7,
Lr1
9,
Lr2
0,
Lr2
1,
Lr2
3,
Lr2
6
P.
stri
ifo
rmis
46
S1
19
Yr1
,Y
r5,
Yr1
0,
Yr1
5,
Yr2
4,
Yr2
5,
Yr2
6,
Yr2
7Y
r2,
Yr3
,Y
r4,
Yr6
,Y
r7,
Yr8
,Y
r9,
Yr1
7,
Yr1
8
46
S1
02
Yr1
,Y
r5,
Yr9
,Y
r10,
Yr1
5,
Yr1
7,
Yr2
4,
Yr2
5,
Yr2
6,
Yr2
7Y
r2,
Yr3
,Y
r4,
Yr6
,Y
r7,
Yr8
,Y
r18
47
S1
03
Yr5
,Y
r9,
Yr1
0,
Yr1
5,
Yr1
7,
Yr2
4,
Yr2
5,
Yr2
6,
Yr2
7Y
r1,
Yr2
,Y
r3,
Yr4
,Y
r6,
Yr7
,Y
r8
78
S8
4Y
r1,
Yr5
,Y
r10,
Yr1
5,
Yr1
7,
Yr2
4,
Yr2
5,
Yr2
6Y
r2,
Yr3
,Y
r4,
Yr6
,Y
r7,
Yr8
,Y
r9,
Yr2
7
aA
tth
ese
edli
ng
stag
ein
the
gla
ssh
ou
seu
nd
erst
and
ard
con
dit
ion
s(N
ayar
etal
.1
99
7)
Genet Resour Crop Evol (2008) 55:849–859 851
123

with a mixture of rust urediospores and talc. The
inoculated seedlings were incubated in a dark cham-
ber at 20 ± 1�C at 100% RH for 16 h (Nayar et al.
1997). After the incubation, the trays were shifted to
a green house maintained at 20 ± 2�C. Infection
types (ITs) were recorded 14 days after the inocula-
tion using a 0–4 scale (Stakman et al. 1962) where IT
0; = no uredinia or other macroscopic sign of
infection, ; = no uredinia but small hypersensitive
necrotic or chlorotic flecks present, 1 = small uredi-
nia surrounded by necrosis, 3 = medium uredinia
with or without chlorosis, 3+ = large uredinia with
or without chlorosis and 33+ = large uredinia with-
out chlorosis. For adult plant screening, the
introgression lines were planted in 1.5 m rows with
row-to-row distance of 20 cm and plant-to-plant
distance of 10 cm. Infector rows of the susceptible
cultivar Agra Local were planted all around the
experimental plot and sprayed with a mixture of
urediniospores of Pt pathotypes (109R31-1, 121R63-
1 and 21R5) and Pst pathotypes (46S102, 47S103 and
46S119). At the adult plant stage, data were recorded
according to the modified Cobb’s scale (Peterson
et al. 1948) which included disease severity (percent
leaf area affected) and infection type viz. 0 =
immune; R = resistant, MR = moderately resistant;
MS = moderately susceptible and S = susceptible.
Cytological studies
Spikes were fixed at the pre-booting stage in Cornoy’s
solution II (6 ethanol:3 chloroform:1 glacial acetic
acid) and transferred to ethanol (70%) after 48 h.
Squash preparations of pollen mother cells (PMCs) at
the metaphase I were made in acetocarmine (2%) to
study the chromosome number and pairing behavior
in the selected wheat-Ae. umbellulata introgression
lines. Genomic in situ hybridization (GISH) was used
to detect alien introgression in some of the selected
rust resistant wheat-Ae. umbellulata ILs. GISH was
done as described in Zhang et al. (2001) using Ae.
umbellulata (2n = 14, UU) genomic DNA as the
probe and Chinese Spring (CS) genomic DNA as
blocker. Total genomic DNA was extracted from CS
wheat and Ae. umbellulata using the CTAB method as
modified by Saghai-Maroof et al. (1984). The DNA
from Ae. umbellulata was sheared using a needle to an
average size of 200 bp and CS DNA was autoclaved
for 12 min. The genomic Ae. umbellulata DNA was
labeled with fluorescein (FITC) dUTP using nick
translation. A ratio of 35–40:1 of wheat competitor
DNA to the labeled probe (Ae. umbellulata DNA) was
used in the hybridization mixture. Chromosomes were
counterstained with Propidium Iodide (PI) and
mounted in Vectashield (Vector Laboratories). The
slides were analyzed with an epifluorescence Zeiss
Axioplan 2 microscope and images were captured
using a SPOT CCD (charge-coupled-device) camera
operated with SPOT 2.1 software (Diagnostic Instru-
ments) and processed with Photoshop 5.5 (Adobe
Systems).
Results
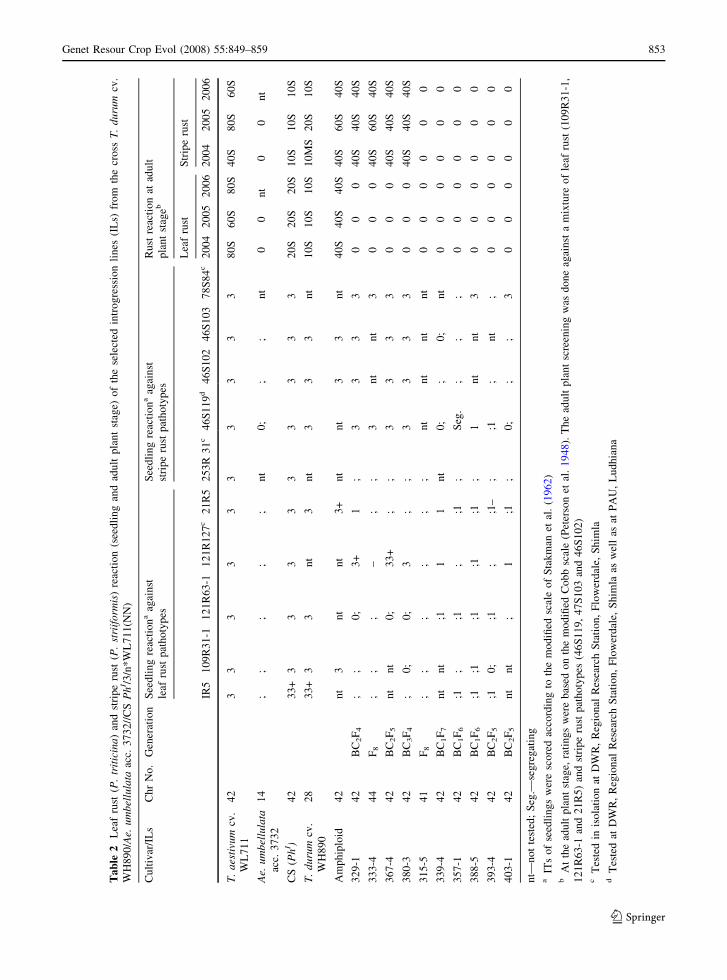
Triticum durum cv. WH890, CS (PhI) and the
recurrent parent WL711(NN) were susceptible against
all the P. triticina and P. striiformis pathotypes used at
the seedling stage whereas at the adult plant stage,
both ‘WH890’ and CS (PhI) exhibited low severities
with susceptible responses to leaf rust as well as stripe
rust in different years (Table 2). The recurrent parent
WL711(NN) showed 60S–80S leaf rust and 40S–80S
stripe rust reactions at the adult plant stage. Ae.
umbellulata accession 3732 was resistant to all the
tested Pt and Pst pathotypes at the seedling stage and
showed hypersensitive reactions to both the rusts at
the adult plant stage. The amphiploid was susceptible
to Pt and Pst pathotypes at the seedling stage and
showed a terminal disease severity of 40S for leaf rust
as well as stripe rust at the adult plant stage (Table 2).
However, plants with rust resistance were recovered
from the selfed and the backcross progenies.
Homozygous wheat-Ae. umbellulata ILs viz.
PAU#315-5, 329-1, 333-4, 339-4, 357-1, 367-4,
380-3, 388-5, 393-4 and 403-1 with leaf and/or stripe
rust resistance were selected and analyzed for the
chromosome number, chromosome pairing and the
effectiveness of resistance against the most virulent
and prevalent Pt and Pst pathotypes. The cytogenetic
characterization of the introgression lines using
meiotic analysis detected normal chromosome num-
ber and pairing in most of the ILs. The ILs 329-1,
357-1, 367-4, 380-3, 388-5, 393-4 and 403-1 had
2n = 42 and showed normal chromosome pairing
with 17–19 ring and 2–4 rod bivalents (Fig. 2a–c) at
the metaphase I. IL315-5 had 2n = 41 and IL333-4
852 Genet Resour Crop Evol (2008) 55:849–859
123
Ta
ble
2L
eaf
rust
(P.
trit
icin
a)
and
stri
pe
rust
(P.
stri
ifo
rmis
)re
acti
on
(see
dli
ng
and
adu
ltp
lan
tst
age)
of
the
sele
cted
intr
og
ress
ion
lin
es(I
Ls)
fro
mth
ecr
oss
T.
du
rum
cv.
WH
89
0/A
e.u
mb
ellu
lata
acc.
37
32
//C
SP
hI /3
/n*
WL
71
1(N
N)
Cu
ltiv
ar/I
Ls
Ch
rN
o.
Gen
erat
ion
See
dli
ng
reac
tio
na
agai
nst
leaf
rust
pat
ho
typ
es
See
dli
ng
reac
tio
na
agai
nst
stri
pe
rust
pat
ho
typ
es
Ru
stre
acti
on
atad
ult
pla
nt
stag
eb
Lea
fru
stS
trip
eru
st
IR5
10
9R
31
-11
21
R6
3-1
12
1R
12
7c
21
R5
25
3R
31
c4
6S
11
9d
46
S1
02
46
S1
03
78
S8
4c
20
04
20
05
20
06
20
04
20
05
20
06
T.
aes
tivu
mcv
.
WL
71
1
42
33
33
33
33
33
80
S6
0S
80
S4
0S
80
S6
0S
Ae.
um
bel
lula
taac
c.3
73
2
14
;;
;;
;n
t0
;;
;n
t0
0n
t0
0n
t
CS
(Ph
I )4
23
3+
33
33
33
33
32
0S
20
S2
0S
10
S1
0S
10
S
T.
du
rum
cv.
WH
89
0
28
33
+3
3n
t3
nt
33
3n
t1
0S
10
S1
0S
10
MS
20
S1
0S
Am
ph
iplo
id4
2n
t3
nt
nt
3+
nt
nt
33
nt
40
S4
0S
40
S4
0S
60
S4
0S
32
9-1
42
BC
2F
4;
;0
;3
+1
;3
33
30
00
40
S4
0S
40
S
33
3-4
44
F8
;;
;–
;;
3n
tn
t3
00
04
0S
60
S4
0S
36
7-4
42
BC
2F
5n
tn
t0
;3
3+
;;
33
33
00
04
0S
40
S4
0S
38
0-3
42
BC
3F
4;
0;
0;
3;
;3
33
30
00
40
S4
0S
40
S
31
5-5
41
F8
;;
;;
;;
nt
nt
nt
nt
00
00
00
33
9-4
42
BC
1F
7n
tn
t;1
11
nt
0;
;0
;n
t0
00
00
0
35
7-1
42
BC
1F
6;1
;;1
;;1
;S
eg.
;;
;0
00
00
0
38
8-5
42
BC
1F
6;1
;1;1
;1;1
;1
nt
nt
30
00
00
0
39
3-4
42
BC
2F
5;1
0;
;1;
;1–
;;1
;n
t;
00
00
00
40
3-1
42
BC
2F
5n
tn
t;
1;1
;0
;;
;3
00
00
00
nt—
no
tte
sted
;S
eg.—
seg
reg
atin
ga
ITs
of
seed
lin
gs
wer
esc
ore
dac
cord
ing
toth
em
od
ified
scal
eo
fS
tak
man
etal
.(1
96
2)
bA
tth
ead
ult
pla
nt
stag
e,ra
tin
gs
wer
eb
ased
on
the
mo
difi
edC
ob
bsc
ale
(Pet
erso
net
al.
19
48
).T
he
adu
ltp
lan
tsc
reen
ing
was
do
ne
agai
nst
am
ixtu
reo
fle
afru
st(1
09
R3
1-1
,
12
1R
63
-1an
d2
1R
5)
and
stri
pe
rust
pat
ho
typ
es(4
6S
11
9,
47
S1
03
and
46
S1
02
)c
Tes
ted
inis
ola
tio
nat
DW
R,
Reg
ion
alR
esea
rch
Sta
tio
n,
Flo
wer
dal
e,S
him
lad
Tes
ted
atD
WR
,R
egio
nal
Res
earc
hS
tati
on
,F
low
erd
ale,
Sh
imla
asw
ell
asat
PA
U,
Lu
dh
ian
a
Genet Resour Crop Evol (2008) 55:849–859 853
123

was observed to be an addition line with 2n = 44.
The meiotic analysis of the F1 plants from the cross
of IL393-4 with cv. PBW343, a widely adapted wheat
cultivar, detected 21II consisting of 19 ring and 2 rod
bivalents (Fig. 2d) suggesting the presence of a small
alien introgression. The GISH analysis of IL 315-5
detected four Ae. umbellulata chromosomes, one
submetacentric and three with subterminal centro-
meres (Fig. 3). GISH analysis of IL333-4 detected a
pair of Ae. umbellulata chromosomes with subtermi-
nal centromeres (Fig. 3). GISH analysis was also
conducted for IL339-4 with 2n = 42, however, no
alien chromatin was detected (results not shown).
Rust screening of wheat-Ae. umbellulata
introgression lines
To test the effectiveness of putatively new rust
resistance genes, 10 selected homozygous wheat-Ae.
umbellulata ILs were screened as seedlings and adult
plants against the most virulent pathotypes of Pt and
Pst prevalent in the Indian subcontinent. All the ILs
were resistant to Pt pathotypes IR5, 109R31-1,
121R63-1, 21R5 and 253R31 at the seedling stage
(Table 2). Three ILs 329-1, 367-4 and 380-3 showed
susceptible seedling responses against pathotype
121R127, indicating the presence of Lr9. All the
other ILs (IL333-4 could not be tested for 121R127)
were resistant to 121R127, suggesting the introgres-
sion of leaf rust resistance genes, other than Lr9, from
Ae. umbellulata. All the selected ILs showed com-
plete leaf rust resistance at the adult plant stage
(Table 2) for 3 years (Fig. 4).
These ILs were also tested at the seedling stage
against Pst pathotypes 46S102, 47S103, 46S119 and
78S84. Genotypes including ILs 339-4, 357-1, 388-5,
393-4 and 403-1 were resistant (Table 2) at the
seedling stage indicating the presence of a seedling
resistance gene for stripe rust in these ILs. In contrast,
Fig. 2 Pollen mother cells
in wheat-Ae. umbellulataintrogression lines (a) 380-3
(2n = 42) with 19 ring and
2 rod bivalents, (b) 393-4
with 19 ring and 2 rod
bivalents, (c) 403-1
(2n = 42) with 17 ring and
4 rod bivalents and (d) F1
plant from the cross of
IL393-4 with bread wheat
cv. PBW343 (2n = 42)
with 19 ring and 2 rod
bivalents
Fig. 3 Genomic in situhybridization (GISH) of the
mitotic metaphase
chromosomes of wheat-Ae.umbellulata interspecific
lines T315-5 (a) and T333-4
(b) using total genomic
DNA of Ae. umbellulata as
a probe, visualized by
yellow-green FITC
fluorescence; chromosomes
were counterstained with
propidium iodide and
fluoresce red
854 Genet Resour Crop Evol (2008) 55:849–859
123

ILs 388-5 and 403-1 showed susceptible infection
types against the Pst pathotype 78S84 and need to be
retested. Remaining ILs viz. 329-1, 333-4, 367-4 and
380-3 were susceptible at the seedling stage. IL 315-5
was not tested at the seedling stage. Introgression
lines 329-1, 333-4, 367-4 and 380-3 showed moder-
ate to high adult plant stripe rust responses (40S–
60S). On the contrary, ILs 315-5, 339-4, 357-1,
388-5, 393-4 and 403-1 exhibited complete stripe rust
resistance at the adult plant stage (Fig. 4).
Inheritance studies for leaf rust resistance
Introgression lines 367-4 (leaf rust resistant and stripe
rust susceptible, proposed to carry Lr9) and 403-1
(leaf rust and stripe rust resistant, proposed to carry
putatively new genes) were crossed with the recurrent
parent WL711 and segregation for resistance was
studied in the F2 population. F1 derived from the
IL367-4/WL711 cross was resistant to leaf rust at the
seedling and adult plant stage. The F2 population
showed monogenic segregation at both the seedling
and the adult plant stage (Table 3). In F3, out of 79
progenies screened at the seedling and adult plant
stage, 17 progenies were homozygous resistant, 43
segregating and 19 homozygous susceptible for leaf
rust (v2(1:2:1) = 0.66) confirming the segregation of a
single leaf rust resistance gene. The F1 derived from
IL 403-1/WL711 cross was resistant to both leaf rust
and stripe rust indicating that the resistance was
dominant. When tested for leaf rust seedling resis-
tance, F2 population segregated into 104 resistant (R)
and 48 susceptible (S) plants with v2(3:1) = 3.5
indicating segregation of a single dominant gene.
All the 152 plants (both resistant and susceptible)
were transplanted in the field and screened for
terminal leaf rust severity at the adult plant stage.
Out of the 48 plants that were susceptible at the
seedling stage, 28 were resistant at the adult plant
stage, indicating the presence of an adult plant
resistance (APR) gene for leaf rust resistance. At
the adult plant stage, this F2 population segregated
into 132R: 20S plants with v2(15:1) = 12.4 (Table 3).
Only selected F3 progenies with seedling and/or APR
genes were evaluated for leaf rust resistance. The
Table 3 Segregation of leaf rust (P. triticina) resistance in the F2 from the crosses of the introgression lines derived from the cross T.durum cv. WH890/Ae. umbellulata acc. 3732//CS PhI/3/3*WL711(NN) with the recurrent parent WL711 (NN)
IL Stage No. of plants in F2 Expected ratio Calculated v2 valuec
Resistant Susceptible Total
367-4 Seedling a 68 27 95 3:1 0.59
Adult plantb 67 24 91 3:1 0.09
403-1 Seedlinga 104 48 152 3:1 3.5
Adult plantb 132 20 152 15:1 12.4
a Seedling screening with leaf rust pathotype 121R63-1b Adult plant screening in the field using a mixture of leaf rust and stripe rust pathotypesc v2 value for significance at P = 0.05 is 3.84 (df = 1)
Fig. 4 Leaf and stripe
reaction of the parents and
introgression lines
developed from the cross: T.durum cv. WH890/Ae.umbellulata acc. 3732//CS
PhI/3/*nWL711 (NN) at the
adult plant stage against a
mixture of leaf rust and
stripe rust pathotypes
Genet Resour Crop Evol (2008) 55:849–859 855
123

progeny testing of the 30 selected leaf rust resistant
F2 plants confirmed the transfer of a seedling
resistance gene and an APR gene for leaf rust in
IL403-1 from Ae. umbellulata.
Allelic tests
Evaluation of wheat-Ae. umbellulata ILs at the
seedling stage with different Pt isolates led to the
identification of two groups of lines, one group with
Lr9 and other with a putatively new leaf rust
resistance gene effective at the seedling stage. To
further test that the leaf rust seedling resistance genes
in the two groups were different, crosses were made
between IL 393-4, resistant to all leaf rust and stripe
rust pathotypes and IL 380-3, susceptible to leaf rust
pathotype 121R127 and all stripe rust pathotypes. All
the F1 plants were resistant to both the rusts. In the F2,
however, out of a total of 85 plants, 82 were resistant
to leaf rust at the seedling as well as at the adult plant
stage (Table 4) and three were susceptible, indicating
segregation of two dominant genes (v2(15:1) = 1.07).
The F2 population was also scored for stripe rust
reaction at the adult plant stage in the field, which
segregated into 69R and 16S plants (v2(3:1) = 1.73)
indicating a single gene for stripe rust resistance in
the IL393-4. F2 population from the cross of two ILs
(T393-4 and T357-1) with putatively new leaf rust
and stripe rust resistance genes did not show any
segregation for leaf rust and stripe rust indicating that
the same genes were present in both the ILs.
Discussion
In the present study, we are reporting the transfer of
putatively new leaf rust and stripe rust resistance
genes from a non-progenitor species Ae. umbellulata
to bread wheat through induction of homoelogous
pairing. An accession of Ae. umbellulata with
multiple disease resistance was hybridized with a
susceptible durum cultivar to produce a synthetic
amphiploid. The amphiploid was, however, suscep-
tible, indicating the presence of suppressors in the A
and/or B genomes of durum wheat (Aghaee et al.
2001). Ae. umbellulata leaf rust and stripe rust
resistance genes were recovered following the crosses
of the amphiploid with the bread wheat cv. WL711
due to the segregation of the suppressors of the durum
genomes. Based on the leaf and stripe rust reaction of
the seedling and adult plant stages and the tests of
allelism, the introgression lines could be grouped into
two broad categories. Introgression lines 329-1, 333-
4, 367-4 and 380-3 carried seedling effective leaf rust
resistance gene only, similar to Lr9, transferred
previously from Ae. umbellulata into the wheat as a
homoeologous chromosome transfer 6BS.6BL-6UL
(Sears 1961; Friebe et al. 1995). The inheritance
studies in the IL 367-4 also confirmed the transfer of
a single leaf rust resistance gene only. All the other
six ILs viz. 315-5, 339-4, 357-1, 388-5, 393-4 and
403-1 carried putatively new seedling leaf rust and
stripe rust resistance gene(s). Inheritance studies in
the IL 403-1 showing monogenic segregation at the
seedling stage and digenic segregation at the adult
Table 4 Allelic tests of wheat-Ae. umbellulata introgression lines T393-4 with T380-3 (leaf rust resistant and stripe rust susceptible)
and T357-1 (leaf and stripe rust resistant)
No. of plants in F2 Expected ratio Calculated v2 value
Resistant Susceptible Total
F2: T393-4/T380-3
Seedlinga 82 3 85 15:1 1.07
Adult plant leaf rustb 82 3 85 15:1 1.07
Adult plant stripe rustb 69 16 85 3:1 1.73
F2: T393-4/T 357-1
Seedling leaf rust 111 0 111 – –
Adult plant leaf rust and stripe rust 111 0 111 – –
a Seedling screening with leaf rust pathotype 121R63-1b Adult plant screening in the field using a mixture of leaf and stripe rust pathotypesc v2 value for significance at P = 0.05 is 3.84 (df = 1)
856 Genet Resour Crop Evol (2008) 55:849–859
123

plant stage indicated that this introgression line
carried a gene each for seedling and adult plant leaf
rust resistance. The segregation for the seedling leaf
rust resistance gene fitted well to the monogenic ratio.
However, the two gene segregation observed at the
adult plant stage was distorted suggesting that the
APR gene for leaf rust might be carried on a larger
alien segment. Such distortions in the transmission of
alien genes have often been observed (Endo 1990;
Prins and Marais 1999; Marais et al. 2005b). APR
gene for leaf rust in IL403-1 appeared to have been
masked due to the presence of the seedling resistance
gene indicating that some other ILs with seedling
resistance gene for leaf rust might also carry the APR
gene. The cv. Chinese Spring parent used for the
induction of homoeologous pairing carried an APR
gene for leaf rust, Lr34 (McIntosh 1992; Singh and
Gupta 1992). Chinese Spring showed terminal leaf
rust severity of 20S in the field while the F2 plants
and one of the ILs homozygous for APR gene (data
not given) developed 5MR leaf rust indicating that
the APR gene identified in the IL403-1 was different
from Lr34. The standard Ae. umbellulata genome has
three chromosomes with submedian and four chro-
mosomes with subterminal centromeres (Friebe et al.
1995). The Ae. umbellulata chromosome, with sub-
terminal centromere, present in IL333-4 might be 6U
which is known to carry Lr9 (Sears 1956). Hence the
putatively new rust resistance genes might be located
on other subterminal/submetacentic chromosome(s)
observed in 315-5. The other introgression lines,
derived from the same cross amphiploid/CS Ph1//
WL711(NN), might be carrying different sizes of the
introgressed alien segments. Linkage drag was
apparent from the variation in the plant type of these
leaf and stripe rust resistant interspecific lines. The
plant type of the IL315-5 was the most affected, with
very poor, grass-like phenotype with narrow leaves,
reduced height, small spikes and delayed flowering.
The GISH analysis confirmed that this IL had very
high amount of the alien chromatin. IL393-4
appeared to carry the smallest alien segment with
leaf rust and stripe rust resistance genes since it did not
have any apparent linked, deleterious phenotypic
effects. Presence of only bivalents in the F1 derived
from 393-4/PBW343 cross also indicated that 393-4 did
not carry complete Ae. umbellulata chromosome.
Another IL339-4 carried cytologically undetectable
introgression. The IL393-4 has been selected for further
studies on the molecular mapping of putative novel leaf
rust and stripe rust resistance genes and for transferring
these genes to other elite wheat backgrounds.
Except for 1BS-1RL wheat-rye translocation in
Veery series of CIMMYT wheat lines, most of the alien
introgressions had limited use in practical breeding
because of cytological instability of alien chromosome
segments incorporated in non-homoeologous regions or
because of the linkage of the undesirable genes on the
large alien segments (Friebe et al. 1996). However, by
screening a large number of recombinants, we were
successful in recovering the IL 393-4 with apparently
least linkage drag and IL 339-4 with cytologically
undetectable alien segment. Identification of the wheat-
Ae. umbellulata introgression lines with no apparent
linkage drag clearly suggests that it is possible to
transfer novel and useful genetic variability from wild
species with little undesirable genetic information.
During the present investigation, three putatively
new rust resistance genes, one seedling and one APR
gene for leaf rust temporarily designated as LrU1 and
LrU2 and one seedling gene for stripe rust, temporarily
designated as YrU1 have been transferred from a single
accession of Ae. umbellulata to T. aestivum cv. WL711.
In addition to these new genes, a leaf rust resistance
gene that may be similar to Lr9 has been transferred.
Transfer of three novel rust resistance genes including
one APR gene from single Ae. umbellulata accession
clearly demonstrates the presence of a high level of
useful variability among related species and the
importance of these sources for germplasm enhance-
ment. Mapping of these resistance genes using
molecular markers is in progress and will facilitate
their cataloguing and utilization for wheat germplasm
enhancement, especially in the Indian subcontinent.
Acknowledgements Financial assistance provided by the
USDA-ARS under the Project IN-ARS-842 and Grant Number
FG-In-792 to carry out this research work is gratefully
acknowledged. We thank Dr. B S Gill, Distinguished
Professor and Dr. Bernd Friebe, WGGRC, KSU, USA, for
providing laboratory facilities for GISH analysis. We are also
grateful to Dr. H S Bariana, Australian Cereal Rust Control
Program, University of Sydney, Australia for a critical review
of the manuscript and valuable suggestions.
References
Aghaee-Sarbarzeh M, Singh H, Dhaliwal HS (2000) Ph1 gene
derived from Ae. speltoides induces homoeologous
Genet Resour Crop Evol (2008) 55:849–859 857
123

pairing in wide crosses of Triticum aestivum. J Heredity
91:417–421
Aghaee-Sarbarzeh M, Dhaliwal HS, Chhuneja P, Singh H
(2001) Suppression of rust resistance genes from distantly
related species in Triticum durum-Aegilops amphiploids.
Wheat Inf Ser 92:12–16
Ceoloni C, Jauhar PP (2006) Chromosome engineering of the
durum wheat genome: strategies and applications of
potential breeding value. In: Singh RJ, Jauhar PP (eds)
Genetic resources, chromosome engineering and crop
improvement: Cereals. CRC Press, Taylor & Francis
Group, London, pp 27–59
Chen PD, Tsujimoto H, Gill BS (1994) Transfer of PhI genes
promoting homoeologous pairing from Triticum spelto-ides to common wheat. Theor Appl Genet 88:97–101
Dhaliwal HS, Singh H, Gupta S, Bagga PS, Gill KS (1991)
Evaluation of Aegilops and wild Triticum species for
resistance to leaf rust (Puccinia recondita f.sp. tritici) of
wheat. Intern J Trop Agric 9:118–121
Dhaliwal HS, Chhuneja P, Gill RK, Goel RK, Singh H (2003)
Introgression of disease resistance genes from related
species into cultivated wheats through interspecific
hybridization. Crop Improv 29:1–18
Dvorak J (1977) Transfer of leaf rust resistance from Aegilopsspeltoides to Triticum aestivum. Can J Genet Cytol
19:133–141
Endo TR (1990) Gametocidal chromosomes and their induc-
tion of chromosome mutations in wheat. Jpn J Genet
65:135–152
Flor HH (1942) Inheritance of pathogenicity in Melampsoralini. Phytopathology 32:653–669
Friebe B, Jiang J, Tuleen N, Gill BS (1995) Standard karyotype
of Aegilops umbellulatum and the characterization of the
derived chromosome addition and translocation lines in
common wheat. Theor Appl Genet 90:150–156
Friebe B, Jiang J, Raupp WJ, McIntosh RA, Gill BS (1996)
Characterization of wheat-alien translocations conferring
resistance to diseases and pests: current status. Euphytica
91:59–87
Gale MD, Miller TE (1987) The introduction of alien genetic
variation into wheat. In: Lupton FGH (ed) Wheat
breeding: its scientific basis. Chapman and Hall, UK,
pp 173–210
Hammer K (1985) Studies towards a monographic treatment of
wild plant collections: Aegilops L. – resistance tests.
Kulturpflanze 33:123–131
Jiang J, Friebe B, Gill BS (1994) Recent advances in alien gene
transfer in wheat. Euphytica 73:199–212
Kuraparthy V, Chhuneja P, Dhaliwal HS, Kaur S, Bowden RL,
Gill BS (2007a) Characterization and mapping of cryptic
alien introgression from Aegilops geniculata with novel
leaf rust and stripe rust resistance genes Lr57 and Yr40 in
wheat. Theor Appl Genet 114:1379–1389
Kuraparthy V, Sood S, Chhuneja P, Dhaliwal HS, Kaur S,
Bowden RL, Gill BS (2007b) A cryptic wheat-Aegilopstriuncialis translocation with leaf rust resistance gene
Lr58. Crop Sci 47:1995–2003
Marais GF, McCallum B, Snyman JE, Pretorius ZA, Marais AS
(2005a) Leaf rust and stripe rust resistance genes Lr54 and
Yr37 transferred to wheat from Aegilops kotschyi. Plant
Breed 124:538–541
Marais GF, Pretorius ZA, Wellings CR, McCallum B, Marais
AS (2005b) Leaf rust and stripe rust resistance genes
transferred to common wheat from Triticum dicoccoides.
Euphytica 143:115–123
Masoudi-Nejad A, Nasuda S, McIntosh RA, Endo TR (2002)
Transfer of rye chromosome segments to wheat by a ga-
metocidal system. Chromosome Res 10:349–357
McIntosh RA (1992) Close genetic linkage of genes conferring
adult plant resistance to leaf rust and stripe rust in wheat.
Plant Pathol 41:523–527
McIntosh RA, Devos KM, Dubcovsky J, Rogers WJ, Morris
CF, Appels R, Anderson OD (2005) Catalogue of gene
symbols: 2005 supplement. In: KOMUGI—Integrated
Wheat Science Database. http://www.grs.nig.ac.jp/wheat/
komugi/genes/macgene/supplement2005.pdf
Nayar SK, Prashar M, Bhardwaj SC (1997) Manual of current
techniques in wheat rusts. Research Bull No. 2, 32 pp,
Regional Station. Flowerdale, Shimla 171002, India
Peterson RF, Campbell AB, Hannah AE (1948) A diagram-
matic scale for rust intensity on leaves and stems of
cereals. Can J Res 26:496–500
Prins R, Marais GF (1999) A genetic study of the gametocidal
effect of the Lr19 translocation of common wheat. S Afr J
Plant Soil 16:10–14
Riley R, Chapman V, Johnson R (1968) The incorporation of
alien disease resistance in wheat by genetic interference
with the regulation of meiotic chromosome synapsis.
Genet Res Camb 12:198–219
Romero M, Montes MJ, Sin E, Lopez-Brana I, Duce A, Martin-
Sanchez JA, Andres MF, Delibes A (1998) A cereal cyst
nematode (Heterodera avenae) resistance gene transferred
from Aegilops triuncialis to hexaploid wheat. Theor Appl
Genet 96:1135–1140
Saghai-Maroof MA, Soliman KM, Jorgensen RA, Allard RW
(1984) Ribosomal DNA spacer length polymorphisms in
barley: Mendelian inheritance, chromosomal location, and
population dynamics. Proc Natl Acad Sci USA 81:8014–
8018
Sears ER (1956) The transfer of leaf rust resistance from
Aegilops umbellulata to wheat. Brookhaven Symp in Biol.
No. 9, Genetics in Plant Breeding, pp 1–22
Sears ER (1961) Identification of the wheat chromosome car-
rying leaf rust resistance from Aegilops umbellulata.
Wheat Inf Serv 12:12–13
Sears ER (1972) Chromosome engineering in wheat. In:
Stadler Genetics Symp. 4. Univ. of Missouri, Columbia,
USA, pp 23–38
Sears ER (1981) Transfer of alien genetic material to wheat. In:
Evans LT, Peacock WJ (eds) Wheat science today and
tomorrow, pp 75–89
Sharma HC, Gill BS (1983) Current status of wide hybridiza-
tion in wheat. Euphytica 32:17–31
Singh H, Dhaliwal HS (2000) Intraspecific genetic diversity for
resistance to wheat rusts in wild Triticum and Aegilopsspecies. Wheat Inf Serv 90:21–30
Singh RP, Gupta AK (1992) Expression of wheat leaf rust
resistance gene Lr34 in seedlings and adult plants. Plant
Dis 76:489–491
Singh H, Grewal TS, Dhaliwal HS, Pannu PPS, Bagga PS
(1998) Sources of leaf rust and stripe rust resistance in
wild relatives of wheat. Crop Improv 25:26–33
858 Genet Resour Crop Evol (2008) 55:849–859
123

Singh K, Chhuneja P, Ghai M, Kaur S, Goel RK, Bains NS,
Keller B, Dhaliwal HS (2007) Molecular mapping of leaf
and stripe rust resistance genes in Triticum monococcumand their transfer to hexaploid wheat. In: Buck H, Nisi JE,
Solomon N (eds) Wheat production in stressed environ-
ments. Developments in Plant Breeding, vol 12. Springer,
Netherlands, pp 779–786
Stakman EC, Stewart DH, Loegering WQ (1962) Identification
of physiologic races of Puccinia graminis var. tritici.USDA Agri Res Serv No E617 (Rev.), p 53
Valkoun J, Hammer K, Kucerova D, Bartos P (1985) Disease
resistance in the genus Aegilops L. – stem rust, leaf rust,
stripe rust and powdery mildew. Kulturpflanze 33:
133–153
Young ND, Tanksley SD (1989) RFLP analysis of the size of
chromosomal segment retained around TM-2 locus of
tomato during backcross breeding. Theor Appl Genet
77:353–359
Zhang P, Friebe B, Lukaszewski AJ, Gill BS (2001) The
centromere structure in robertsonian wheat-rye translo-
cation chromosomes indicates that centric breakage-
fusion can occur at different positions within the primary
constriction. Chromosoma 110:335–344
Genet Resour Crop Evol (2008) 55:849–859 859
123




















