
Transcription Termination and RNA Degradation Contribute to Silencing of RNA Polymerase II...
11
Molecular Cell Article Transcription Termination and RNA Degradation Contribute to Silencing of RNA Polymerase II Transcription within Heterochromatin Lidia Vasiljeva, 1 Minkyu Kim, 1 Nihal Terzi, 1 Luis M. Soares, 1 and Stephen Buratowski 1, * 1 Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 240 Longwood Avenue, Boston, MA 02115, USA *Correspondence: [email protected] DOI 10.1016/j.molcel.2008.01.011 SUMMARY Within the heterochromatin of budding yeast, RNA polymerase II (RNAPII) transcription is repressed by the Sir2 deacetylase. Although heterochromatic si- lencing is generally thought to be due to limited ac- cessibility of the underlying DNA, there are several reports of RNAPII and basal transcription factors within silenced regions. Analysis of the rDNA array re- vealed cryptic RNAPII transcription within the ‘‘non- transcribed’’ spacer region. These transcripts are terminated by the Nrd1/Sen1 complex and degraded by the exosome. Mutations in this pathway lead to de- creased silencing and dramatic chromatin changes in the rDNA locus. Interestingly, Nrd1 mutants also show higher levels of rDNA recombination, suggest- ing that the cryptic RNAPII transcription might have a physiological role in regulating rDNA copy number. The Nrd1/Sen1/exosome pathway also contributes to silencing at telomeric loci. These results suggest that silencing of heterochromatic genes in Saccharo- myces cerevisiae occurs at both transcriptional and posttranscriptional levels. INTRODUCTION In the budding yeast Saccharomyces cerevisiae, silent chromatin (heterochromatin) includes the rRNA-encoding DNA array (rDNA), telomeric regions, and silent mating-type loci. Within these regions, transcription of reporter genes by RNA polymer- ase II (RNAPII) is repressed. Histone deacetylation by Sir2 plays an important role in heterochromatin formation (Moazed, 2001a, 2001b). In addition, histone methylation by Set1 and chromatin remodeling by Swi/Snf complex also contribute to silencing (Briggs et al., 2001; Bryk et al., 2002; Dror and Winston, 2004). The mechanism by which heterochromatin inhibits transcription is still unclear. One model suggests that the compact structure of heterochromatin regulates DNA accessibility. However, other studies found that heterochromatin is accessible for RNAPII re- cruitment, suggesting inhibition occurs after RNAPII assembly into the initiation complex (Sekinger and Gross, 1999, 2001). Eukaryotic rRNA genes are separated by ‘‘nontranscribed’’ spacers, NTS1 and NTS2 (Figure 1A). Ribosomal RNA genes are arranged as tandem repeats of 100–200 copies, a subset of which are highly transcribed by RNA polymerase I (RNAPI) and RNA polymerase III (RNAPIII) within the nucleolus (Preuss and Pi- kaard, 2007; Venema and Tollervey, 1995, 1999). RNAPII-tran- scribed reporter genes inserted into rDNA array are silenced, but the primary function of heterochromatic rDNA is probably to prevent recombination and loss of rDNA repeats (reviewed in Moazed, 2001a). It is unclear how RNAPII silencing is maintained in rDNA while this region is highly transcribed by two other poly- merases. A recent analysis of RNAPII distribution throughout the S. cerevisiae genome revealed that RNAPII is present within het- erochromatin (Steinmetz et al., 2006). Similar analyses in human cells also detected RNAPII at promoters of genes that were not producing full-length transcripts (Barski et al., 2007; Guenther et al., 2007). Transcription is a complex process regulated at mul- tiple levels, and these data suggest that events downstream of RNAPII recruitment can contribute to silencing. We have shown that the RNA-binding protein Nrd1 recognizes the RNAPII C-terminal domain (CTD) phosphorylated at Ser5 (L.V., M.K., S.B., and A. Meinhart, unpublished data) and associ- ates with promoter-proximal RNAPII transcription complexes (Nedea et al., 2003). This allows Nrd1, in a complex with the RNA-binding protein Nab3 and RNA helicase Sen1, to control transcription early in elongation by promoting RNAPII termination (Arigo et al., 2006a; Steinmetz et al., 2006). The Nrd1 complex mediates termination of RNAPII transcripts such as short (%500 nt) mRNAs, snoRNAs, and recently described cryptic un- stable transcripts or CUTs (Arigo et al., 2006b; Kim et al., 2006; Steinmetz et al., 2001; Steinmetz et al., 2006; Thiebaut et al., 2006). In eukaryotes, RNA can be degraded in a 3 0 / 5 0 direction by the exosome complex (Vanacova and Stefl, 2007). We showed that the Nrd1/Nab3/Sen1 recruits the exosome to 3 0 ends of RNA substrates to promote RNA degradation or processing (Vasiljeva and Buratowski, 2006). Because the Nrd1/exosome complex can terminate and de- grade RNAPII transcripts, we tested whether it plays a role in re- pressing RNAPII transcription within heterochromatin. We report here that the Nrd1/Sen1/exosome pathway represses CUT RNAs produced by RNAPII within rDNA loci. Furthermore, loss of proper CUT termination leads to dramatic changes in the rDNA chromatin (most notably an increase in histone acetylation) Molecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier Inc. 313
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Transcription Termination and RNA Degradation Contribute to Silencing of RNA Polymerase II...

Molecular Cell
Article
Transcription Termination and RNA DegradationContribute to Silencing of RNA Polymerase IITranscription within HeterochromatinLidia Vasiljeva,1 Minkyu Kim,1 Nihal Terzi,1 Luis M. Soares,1 and Stephen Buratowski1,*1Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, 240 Longwood Avenue, Boston,
MA 02115, USA
*Correspondence: [email protected] 10.1016/j.molcel.2008.01.011
SUMMARY
Within the heterochromatin of budding yeast, RNApolymerase II (RNAPII) transcription is repressed bythe Sir2 deacetylase. Although heterochromatic si-lencing is generally thought to be due to limited ac-cessibility of the underlying DNA, there are severalreports of RNAPII and basal transcription factorswithin silenced regions. Analysis of the rDNA array re-vealed cryptic RNAPII transcription within the ‘‘non-transcribed’’ spacer region. These transcripts areterminated by the Nrd1/Sen1 complex and degradedby the exosome. Mutations in this pathway lead to de-creased silencing and dramatic chromatin changesin the rDNA locus. Interestingly, Nrd1 mutants alsoshow higher levels of rDNA recombination, suggest-ing that the cryptic RNAPII transcription might havea physiological role in regulating rDNA copy number.The Nrd1/Sen1/exosome pathway also contributesto silencing at telomeric loci. These results suggestthat silencing of heterochromatic genes in Saccharo-myces cerevisiae occurs at both transcriptional andposttranscriptional levels.
INTRODUCTION
In the budding yeast Saccharomyces cerevisiae, silent chromatin
(heterochromatin) includes the rRNA-encoding DNA array
(rDNA), telomeric regions, and silent mating-type loci. Within
these regions, transcription of reporter genes by RNA polymer-
ase II (RNAPII) is repressed. Histone deacetylation by Sir2 plays
an important role in heterochromatin formation (Moazed, 2001a,
2001b). In addition, histone methylation by Set1 and chromatin
remodeling by Swi/Snf complex also contribute to silencing
(Briggs et al., 2001; Bryk et al., 2002; Dror and Winston, 2004).
The mechanism by which heterochromatin inhibits transcription
is still unclear. One model suggests that the compact structure of
heterochromatin regulates DNA accessibility. However, other
studies found that heterochromatin is accessible for RNAPII re-
cruitment, suggesting inhibition occurs after RNAPII assembly
into the initiation complex (Sekinger and Gross, 1999, 2001).
Mo
Eukaryotic rRNA genes are separated by ‘‘nontranscribed’’
spacers, NTS1 and NTS2 (Figure 1A). Ribosomal RNA genes
are arranged as tandem repeats of 100–200 copies, a subset of
which are highly transcribed by RNA polymerase I (RNAPI) and
RNA polymerase III (RNAPIII) within the nucleolus (Preuss and Pi-
kaard, 2007; Venema and Tollervey, 1995, 1999). RNAPII-tran-
scribed reporter genes inserted into rDNA array are silenced,
but the primary function of heterochromatic rDNA is probably to
prevent recombination and loss of rDNA repeats (reviewed in
Moazed, 2001a). It is unclear how RNAPII silencing is maintained
in rDNA while this region is highly transcribed by two other poly-
merases. A recent analysis of RNAPII distribution throughout the
S. cerevisiae genome revealed that RNAPII is present within het-
erochromatin (Steinmetz et al., 2006). Similar analyses in human
cells also detected RNAPII at promoters of genes that were not
producing full-length transcripts (Barski et al., 2007; Guenther
et al., 2007). Transcription is a complex process regulated at mul-
tiple levels, and these data suggest that events downstream of
RNAPII recruitment can contribute to silencing.
We have shown that the RNA-binding protein Nrd1 recognizes
the RNAPII C-terminal domain (CTD) phosphorylated at Ser5
(L.V., M.K., S.B., and A. Meinhart, unpublished data) and associ-
ates with promoter-proximal RNAPII transcription complexes
(Nedea et al., 2003). This allows Nrd1, in a complex with the
RNA-binding protein Nab3 and RNA helicase Sen1, to control
transcription early in elongation by promoting RNAPII termination
(Arigo et al., 2006a; Steinmetz et al., 2006). The Nrd1 complex
mediates termination of RNAPII transcripts such as short
(%500 nt) mRNAs, snoRNAs, and recently described cryptic un-
stable transcripts or CUTs (Arigo et al., 2006b; Kim et al., 2006;
Steinmetz et al., 2001; Steinmetz et al., 2006; Thiebaut et al.,
2006). In eukaryotes, RNA can be degraded in a 30 / 50 direction
by the exosome complex (Vanacova and Stefl, 2007). We showed
that the Nrd1/Nab3/Sen1 recruits the exosome to 30 ends of RNA
substrates to promote RNA degradation or processing (Vasiljeva
and Buratowski, 2006).
Because the Nrd1/exosome complex can terminate and de-
grade RNAPII transcripts, we tested whether it plays a role in re-
pressing RNAPII transcription within heterochromatin. We report
here that the Nrd1/Sen1/exosome pathway represses CUT
RNAs produced by RNAPII within rDNA loci. Furthermore, loss
of proper CUT termination leads to dramatic changes in the
rDNA chromatin (most notably an increase in histone acetylation)
lecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier Inc. 313
Molecular Cell
Multiple Levels of Heterochromatic Silencing
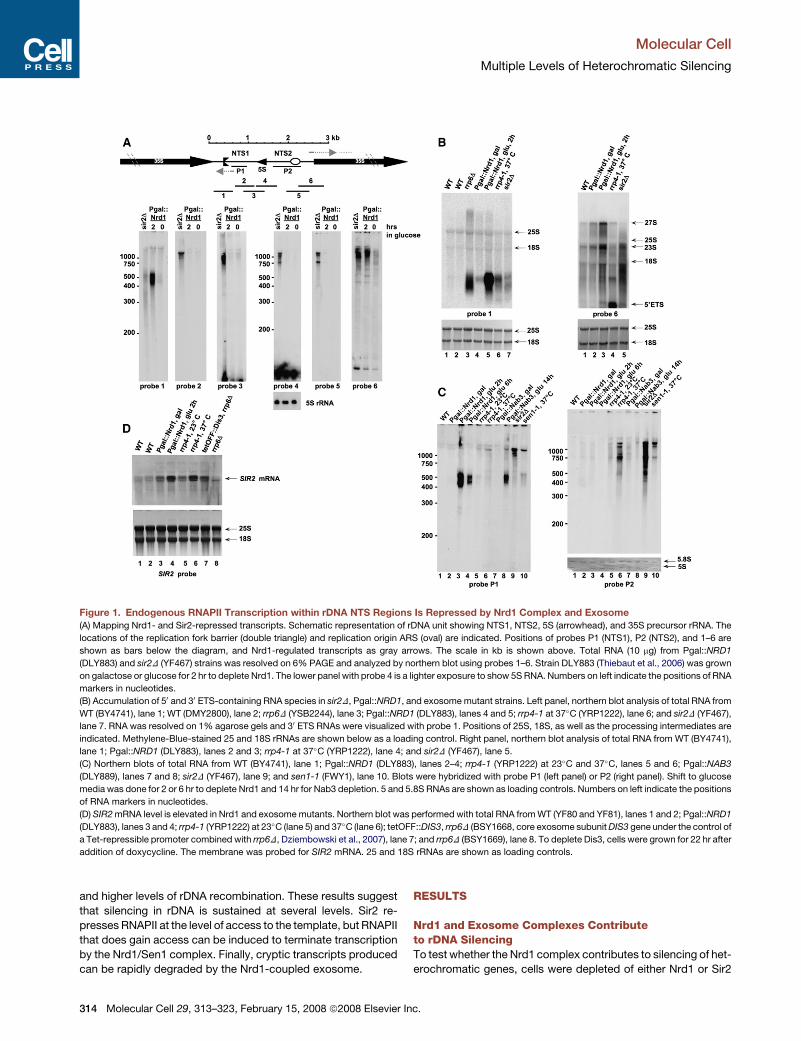
Figure 1. Endogenous RNAPII Transcription within rDNA NTS Regions Is Repressed by Nrd1 Complex and Exosome
(A) Mapping Nrd1- and Sir2-repressed transcripts. Schematic representation of rDNA unit showing NTS1, NTS2, 5S (arrowhead), and 35S precursor rRNA. The
locations of the replication fork barrier (double triangle) and replication origin ARS (oval) are indicated. Positions of probes P1 (NTS1), P2 (NTS2), and 1–6 are
shown as bars below the diagram, and Nrd1-regulated transcripts as gray arrows. The scale in kb is shown above. Total RNA (10 mg) from Pgal::NRD1
(DLY883) and sir2D (YF467) strains was resolved on 6% PAGE and analyzed by northern blot using probes 1–6. Strain DLY883 (Thiebaut et al., 2006) was grown
on galactose or glucose for 2 hr to deplete Nrd1. The lower panel with probe 4 is a lighter exposure to show 5S RNA. Numbers on left indicate the positions of RNA
markers in nucleotides.
(B) Accumulation of 50 and 30 ETS-containing RNA species in sir2D, Pgal::NRD1, and exosome mutant strains. Left panel, northern blot analysis of total RNA from
WT (BY4741), lane 1; WT (DMY2800), lane 2; rrp6D (YSB2244), lane 3; Pgal::NRD1 (DLY883), lanes 4 and 5; rrp4-1 at 37�C (YRP1222), lane 6; and sir2D (YF467),
lane 7. RNA was resolved on 1% agarose gels and 30 ETS RNAs were visualized with probe 1. Positions of 25S, 18S, as well as the processing intermediates are
indicated. Methylene-Blue-stained 25 and 18S rRNAs are shown below as a loading control. Right panel, northern blot analysis of total RNA from WT (BY4741),
lane 1; Pgal::NRD1 (DLY883), lanes 2 and 3; rrp4-1 at 37�C (YRP1222), lane 4; and sir2D (YF467), lane 5.
(C) Northern blots of total RNA from WT (BY4741), lane 1; Pgal::NRD1 (DLY883), lanes 2–4; rrp4-1 (YRP1222) at 23�C and 37�C, lanes 5 and 6; Pgal::NAB3
(DLY889), lanes 7 and 8; sir2D (YF467), lane 9; and sen1-1 (FWY1), lane 10. Blots were hybridized with probe P1 (left panel) or P2 (right panel). Shift to glucose
media was done for 2 or 6 hr to deplete Nrd1 and 14 hr for Nab3 depletion. 5 and 5.8S RNAs are shown as loading controls. Numbers on left indicate the positions
of RNA markers in nucleotides.
(D) SIR2 mRNA level is elevated in Nrd1 and exosome mutants. Northern blot was performed with total RNA from WT (YF80 and YF81), lanes 1 and 2; Pgal::NRD1
(DLY883), lanes 3 and 4; rrp4-1 (YRP1222) at 23�C (lane 5) and 37�C (lane 6); tetOFF::DIS3, rrp6D (BSY1668, core exosome subunit DIS3 gene under the control of
a Tet-repressible promoter combined with rrp6D, Dziembowski et al., 2007), lane 7; and rrp6D (BSY1669), lane 8. To deplete Dis3, cells were grown for 22 hr after
addition of doxycycline. The membrane was probed for SIR2 mRNA. 25 and 18S rRNAs are shown as loading controls.
and higher levels of rDNA recombination. These results suggest
that silencing in rDNA is sustained at several levels. Sir2 re-
presses RNAPII at the level of access to the template, but RNAPII
that does gain access can be induced to terminate transcription
by the Nrd1/Sen1 complex. Finally, cryptic transcripts produced
can be rapidly degraded by the Nrd1-coupled exosome.
314 Molecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier
RESULTS
Nrd1 and Exosome Complexes Contributeto rDNA SilencingTo test whether the Nrd1 complex contributes to silencing of het-
erochromatic genes, cells were depleted of either Nrd1 or Sir2
Inc.

Molecular Cell
Multiple Levels of Heterochromatic Silencing
and transcription was assayed within NTS1 and NTS2 of the
rDNA array. Deletion of SIR2 resulted in accumulation of several
RNA species of molecular weight from 750 to >1000 nt through-
out the entire region, except for an area just 30 to the 35S rRNA in
NTS1 (Figure 1A, probes 1–6, and Figure 1B, probe 6, lane 5).
The probe 1 region overlaps the 30 external spacer (30 ETS) of
rRNA (Figure S1A available online), and Sir2 has only a minor
effect here (Figure 1A, probe 1, and Figure 1B, probe 1, lane 7).
In contrast, in Nrd1-depleted cells, two new transcripts are
seen. The first is a heterogeneous CUT just downstream of the
35S 30 end (Figure 1A, probe 1, and Figure 1B, probe 1, lane
5). Strand-specific probes show that this RNA is transcribed in
a direction opposite to that of the ribosomal RNAs (Figure S2D).
Primer extension (Figure S2B) revealed two 50 ends roughly 300
bp upstream of the replication fork block. Mapping of 30 ends
(Figure S2C) indicates heterogeneity, accounting for the broad
band seen on the northern blot. This transcript is also seen
upon depletion of Nab3 and to a lesser extent in a sen1-1 mutant,
two other components of the Nrd1 complex (Figure 1C, probe
P1, lanes 8 and 10). In the SEN1 mutant, additional RNA species
were detected resembling those observed in sir2D cells (Fig-
ure 1C, probes P1 and P2, lanes 9 and 10), suggesting that
Sen1 may have additional functions compared to Nrd1.
A second transcript starting near the 50 end of the 35S RNA
(Figure 1A, probe 6), in the same orientation as the rDNA tran-
script (Figure S2D), was seen upon Nrd1 or Sir2 depletion. These
RNAs resemble a previously described RNAPII transcript initiat-
ing just upstream of the RNAPI promoter. This RNAPII promoter
can produce 35S rRNA when the RNAPI factor UAF (upstream
activation factor) is mutated (Oakes et al., 1999; Vu et al., 1999).
Analyzing the same RNA with probe 6 on an agarose gel, accumu-
lation of larger species containing 50 external transcribed spacer
(50 ETS) sequences was observed in both Nrd1- and Sir2-de-
pleted cells. For Nrd1 depletion, the mobilities of the new RNA
species correspond to the positions of 23S and 27S RNA precur-
sors not normally observed during 18S rRNA processing (Fig-
ure 1B, probe 6, lane 3). These species are the result of improper
processing (Venema and Tollervey, 1999, see Figure S1A). This
aberrant processing may be because RNAPII transcripts are
not efficiently coupled to the rRNA processing machinery. In con-
trast, the majority of new transcripts seen in sir2D cells are shorter
than 18S rRNA (Figure 1B, probe 6, lane 5), excluding the possi-
bility that these transcripts could be used as functional 18S RNA.
This may explain why a sir2D mutation was unable to allow cells to
use RNAPII instead of RNAPI to synthesize 35S rRNA unless
combined with a UAF mutation (Oakes et al., 1999).
The Nrd1 termination pathway is coupled to exosome-depen-
dent trimming or degradation of transcripts. Therefore, transcrip-
tion in the NTS was also assayed in mutants of two exosome sub-
units: Rrp4, an essential component of the exosome core, and
Rrp6, an auxiliary factor associated specifically with the nuclear
exosome. During normal rRNA processing (Figure S1A), the 50
ETS region of 35S rRNA is cleaved from the pre-rRNA and de-
graded by the exosome. As expected, a temperature-sensitive
allele of RRP4 (rrp4-1) showed stabilization of the �0.6 kb 50
ETS fragment (Figure 1B, probe 6, lane 4) (de la Cruz et al.,
1998; Mitchell et al., 2003). In the rrp4-1 strain, there is also accu-
mulation of the NTS1 CUT (Figure 1B, probe 1, lane 6) and
Mole
increased amounts of the NTS2 RNA (Figure 1C, probe P2, lanes
5 and 6). Deletion of the Rrp6 subunit also had an effect on the
stability of the NTS1 transcript (Figure 1B, probe 1, lane 3). These
data suggest that the exosome can degrade cryptic transcripts
that have escaped the earlier repressive actions of Nrd1 and Sir2.
To confirm that NTS1 and NTS2 transcription observed in the
mutants is due to RNAPII, the temperature-sensitive allele rpb1-1
was combined either with rrp6D, sen1-1, or sir2D. In the rpb1-1
background at nonpermissive temperature (37�C), cryptic NTS
transcripts were greatly reduced in all three mutant backgrounds
(Figures S1C–S1E). However, a stable fraction of 50 ETS RNA
was detected in the rpb1-1, rrp6D mutant at 37�C, presumably
representing the RNAPI transcript that is usually degraded by
the exosome (Figure S1E, lane 8). No effect of rpb1-1 was
seen for the RNAPIII-transcribed U6 gene (Figure S1B).
To verify that the silencing effects of Nrd1 and exosome com-
plex mutants are not indirectly due to lower levels of Sir2, SIR2
mRNA expression was assayed by northern blot (Figure 1D).
Surprisingly, SIR2 mRNA levels were elevated in the exosome
mutants or upon Nrd1 depletion. Nrd1 and exosome appear to
directly or indirectly downregulate SIR2 expression, so loss of si-
lencing upon Nrd1 depletion clearly cannot be due to the loss of
SIR2. Levels of Sir2 are carefully regulated; although SIR2 dele-
tion has no effect on growth rate, SIR2 overexpression is toxic
(Holmes et al., 1997; Matecic et al., 2002), perhaps due to over-
active silencing (Fritze et al., 1997). Overexpression of SIR2 in the
Nrd1 and exosome mutants may be an attempt to compensate
for incomplete heterochromatic silencing.
To determine whether repression of RNAPII transcription by
Sir2 and the Nrd1/Sen1/exosome pathway were working to-
gether or in parallel, double mutants were analyzed (Figure S3).
Deletion of SIR2 combined with Nrd1 or Rrp4 depletion led to in-
creased levels of the NTS1 CUT (Figure S3B) and appearance of
larger unprocessed species. At NTS2, loss of Sir2 and Rrp4
showed additive effects (Figure S3C), although this was less strik-
ing for the combination of Sir2 and Nrd1 depletion (Figure S3D).
These results are consistent with the hypothesis that Sir2 re-
presses transcription at initiation whereas Nrd1 and exosome
repress RNA production at later stages of gene expression.
Reporter Genes Inserted into the rDNA Arrayor Telomeric Regions Can Be Repressedby Nrd1 and the ExosomeIt was important to examine whether the Nrd1/exosome com-
plex specifically affected the endogenous NTS transcripts or
was a more general contributor to heterochromatic silencing.
Strains carrying a URA3 reporter gene integrated either in the
middle of NTS1, just upstream of the mapped CUT (DMY2804),
or near NTS2 in the 50 ETS area actively transcribed by RNAPI
(DMY2800) were tested (Figures 2B and 2C and Figure S4). As
previously observed (Huang et al., 2006), both rDNA reporters
were silenced, whereas a reporter inserted at a euchromatic lo-
cation was active (Figure 2D). As expected, derepression of both
rDNA reporters was seen in sir2D cells. Interestingly, a Nrd1 mu-
tant (D39-169) or deletion of an exosome subunit (rrp6D) dere-
pressed the NTS2 reporter (Figure 2B), and this result was con-
firmed by northern blot (Figure 2C). Southern blotting showed
that the increase of URA3 expression in the NRD1 and RRP6
cular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier Inc. 315

Molecular Cell
Multiple Levels of Heterochromatic Silencing
Figure 2. A URA3 Reporter Integrated in rDNA Can Be Derepressed by Nrd1 and Rrp6 Mutants
(A) Schematic of rDNA unit (see Figure 1A for features) with position of the DMY2800 URA3 reporter shown. Gray arrows show positions and orientation of RNAPII
transcripts.
(B) Derivatives of strain DMY2800 (YSB2243, YSB2246, YSB2245, YSB2244, and YSB2164; see Table S1) were assayed for growth with 10-fold serial dilutions on
media containing (+URA) or lacking uracil (�URA) or containing 5-FOA. Lack of growth on FOA indicates URA3 expression.
(C) Northern blot analysis of URA3 expression in Nrd1-depleted, rrp6D, or sir2D cells. All strains carry the NTS2::URA3 reporter except for DMY2798 (lane 6), in
which URA3 gene is inserted in a euchromatic region as a positive control for URA3 expression. The lower panel shows 5S rRNA as a loading control.
(D) NRD1, RRP6, and SIR2 mutations do not affect a euchromatic URA3 reporter (DMY2798). All strains carry the euchromatic URA3 reporter except for rows 3
and 4, which show NTS1::URA3 (DMY2804) and NTS2::URA3 (DMY2800) reporter strains as positive controls for repression.
mutant strains is not due to an increase in URA3 copy number
(Figure S5). Unexpectedly, the NTS1 reporter was not affected
by mutation of Nrd1 or exosome, but careful mapping of the re-
porter integration site revealed that URA3 inserted only 35 bp up-
stream of the NTS1 CUT initiation site, very likely disrupting the
CUT promoter (Figures S2 and S4). Finally, no effect of Nrd1 or
Rrp6 mutations was seen on the expression of the euchromatic
URA3 reporter (Figure 2D). These results suggest that Nrd1/exo-
some silencing can affect reporter genes within heterochroma-
tin, but location relative to native CUTs may lead to position-
dependent effects.
316 Molecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier I
To test whether Nrd1 and exosome play a role in silencing at
telomeres, we used strain UCC4602 containing a URA3 reporter
gene adjacent to the left telomere of chromosome VII and an
ADE2 reporter inserted near the right telomere of chromosome
V (Singer et al., 1998). Cells lacking Sir2 show strong derepres-
sion of both telomeric reporters, as indicated by the inability to
grow on 5-FOA (Figure 3A) and the lack of red colored sectors
(Figure 3C). Interestingly, the Nrd1 mutant strain showed re-
duced silencing of the URA3 telomeric reporter, and northern
blotting showed a 2-fold increase in URA3 RNA level (Figure 3B).
In addition, there was reduced sectoring, indicating partial
nc.

Molecular Cell
Multiple Levels of Heterochromatic Silencing
Figure 3. Nrd1 and Exosome Mutants Derepress Telomeric Reporters
Strain UCC4602 and its derivatives (YSB2146, YSB2147, YSB2165, YSB2166, YSB2261, and YSB2265) contain URA3 and ADE2 reporters at telomeres of
chromosomes VII and V, respectively.
(A) 10-fold serial dilutions of cells were plated on nonselective (+URA) or selective media (�URA and 5-FOA). URA3 repression is indicated by growth on 5-FOA.
(B) Northern blot analysis of URA3 expression in UCC4602 upon Nrd1 glucose depletion and sir2D cells. Nonrepressed control strain (DMY2798) has the URA3
gene at a euchromatic location. The lower panel shows 5S RNA as a loading control. Normalized URA3 levels were calculated relative to control DMY2798 and are
listed below each lane.
(C) Cells were plated on medium with a low concentration of adenine. ADE2 expression is indicated by the red sectors.
derepression of the ADE2 reporter. Deletion of RRP6 dere-
pressed the ADE2, but not URA3 reporter. The reason for this
discrepancy is unclear. Combining sir2D with nrd1 or rrp6 mu-
tants gave results similar to sir2D alone. In agreement with the re-
porter assays, accumulation of endogenous transcripts derived
from the chromosome V telomeric region was seen in Nrd1- or
Nab3-depleted cells as well as sen1-1 or rrp4-1 mutants (data
not shown).
To test if Nrd1 and exosome function in silencing at mating-
type loci, levels of the endogenous a1 transcript and expression
of a TRP1 reporter integrated at HMR were tested in the mutant
strains (Figure S6). Unlike rDNA and telomeric regions, silencing
at mating-type loci was unaffected in the mutants. Given that
rDNA and telomeric heterochromatin exhibit epigenetic instabil-
ity while silent mating loci are stable, this result could suggest
that the Nrd1/Sen1/exosome pathway affects switching be-
tween inactive and active states of chromatin.
Nrd1 Mediates Termination of RNAPII Transcriptionwithin Active rDNA RepeatsTo explore Nrd1 function at rDNA, its recruitment to NTS1 and
NTS2 was assayed by chromatin immunoprecipitation (ChIP)
Mole
(Figure 4A). Nrd1 crosslinks to a region overlapping the NTS1
CUT (PCR products 13–16). This profile resembles that of RNA-
PII (Figure 4B). This RNAPII peak was also observed in a ge-
nome-wide analysis (Steinmetz et al., 2006) and coincides with
the previously described putative ‘‘E-pro’’ promoter (Kobayashi
and Ganley, 2005).
Nrd1 functions in transcription termination, so we tested
whether it regulates RNAPII occupancy at the rDNA locus. Inter-
estingly, Nrd1 depletion causes RNAPII levels to increase slightly
throughout the entire NTS region but strongly over the NTS1 CUT
(Figures 4B and 4C). The ChIP pattern suggests that transcrip-
tion initiates near primer set 15 but extends into regions 12–14
in the absence of Nrd1-dependent termination. This increase is
different from that seen in a sir2D strain, where the overall RNAPII
pattern remains the same as wild-type (WT) but at much higher
levels (Figure S7). Interestingly, although the RNAPII ChIP signal
increased 6- to 10-fold in the absence of Sir2 (Figure S6) and
2-fold upon Nrd1 depletion (Figure 4C), the intensity of cryptic
transcripts is often equal or higher in case of Nrd1 depletion (Fig-
ures 1A–1C). This lack of correlation is consistent with the idea
that Nrd1/exosome contributes to silencing by acting at post-
initiation steps downstream of Sir2.
cular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier Inc. 317

Molecular Cell
Multiple Levels of Heterochromatic Silencing
Figure 4. Nrd1 Mediates Termination of RNAPII Transcription in NTS1
(A) Nrd1 crosslinks to the NTS1 CUT region. At the top is a schematic diagram of the rDNA repeat (as in Figure 1A) showing positions of the ChIP PCR products
(black bars below with corresponding numbers, from Huang et al., 2006). ChIP was performed with Nrd1-TAP (YF817) and untagged (BY4741) strains. Lower
panel shows PCR products, and middle panel shows data quantification as described in the Experimental Procedures.
(B) RNAPII crosslinks to the NTS1 CUT region and is stimulated by glucose. ChIP analysis with anti-Rpb3 antibody was carried out in Nrd1 WT (W303) cells grown
on galactose and glucose media.
(C) Nrd1 depletion leads to defective RNAPII termination at the NTS1 CUT. ChIP with anti-Rpb3 antibody in Pgal::NRD1 (DLY883) cells grown either on galactose
or on glucose for 2 hr is shown.
(D) RNAPII transcription occurs in transcriptionally active rDNA repeats. ChIP with anti-Rpb3 antibody in a strain containing reduced rDNA repeat number (25
copies all active, NOY1071) or a WT strain containing 190 copies of rDNA (NOY1064).
In all panels, error bars show standard deviation from three repetitions.
Within the 150–200 yeast rDNA repeats, at most half are ac-
tively transcribing at a given time (Dammann et al., 1995). RNAPII
transcription could be occurring in repeats that are transcription-
ally inactive for RNAPI and RNAPIII transcription, or else all three
polymerases could be active on the same repeats. To distinguish
between these possibilities, we tested for crosslinking of RNAPII
in the rDNA repeats of a strain that contains only 25 rDNA re-
peats, all of which are necessarily active to support viability
(Cioci et al., 2003). This strain showed a peak of RNAPII cross-
linking that was about half that seen in an isogenic control strain
with 190 repeats, of which roughly 70–90 should be active
(Figure 4D). This suggests that transcription of the NTS1 CUT
takes place in transcriptionally active rDNA repeats. Interest-
ingly, RNAPII crosslinking at NTS1 was stronger in glucose ver-
sus galactose, even in a WT background (Figure 4B). Because
the number of active rDNA repeats is proportional to cellular
318 Molecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier In
growth rate, we suspect that faster growth in glucose may lead
to more copies of the NTS1 CUT being transcribed.
Readthrough Transcription of the NTS1 CUT Leadsto Chromatin Changes and Increased Recombinationbetween rDNA RepeatsRNAPII transcription has been linked to increased recombination
(Huertas and Aguilera, 2003; Kobayashi and Ganley, 2005), and
Sir2 deacetylase represses crossovers between rDNA repeats
(Moazed, 2001a, 2001b). We therefore tested whether Nrd1
also affects histone acetylation and recombination within
rDNA. ChIP for histone H3 revealed that nucleosome density is
not uniform throughout the NTS region (Figure 5A). There is
reduced histone crosslinking over the NTS1 CUT region and
near the 50 end of the 35S RNA where both RNAPI and RNAPII
initiate. This pattern is consistent with the finding that yeast
c.

Molecular Cell
Multiple Levels of Heterochromatic Silencing
Figure 5. Nrd1 Regulates Histone Modifications and Unequal Crossovers in rDNA
(A) The CUT promoter regions have lower nucleosome density. ChIP analysis with anti-H3 antibody was carried out for Nrd1 WT (W303) and Pgal::NRD1 (DLY883)
cells grown on glucose media for 2 hr. Numbers at bottom refer to PCR products as diagrammed in Figure 4A.
(B) Loss of Nrd1 leads to higher levels of H3 K4 trimethylation. ChIP with anti-H3K4me3 antibody was carried out with the same chromatin used in (A). Signals
were quantitated by a PhosphorImager and normalized to total H3.
(C) Loss of Nrd1 leads to higher levels of histone H4 acetylation. ChIP with anti-H4 tetra-acetyl antibody was carried out as in (B).
(D) Frequency of unequal rDNA crossovers is increased in Nrd1 mutants. Percent of the ADE2 gene loss (% marker loss) was calculated for a wild-type strain (WT,
DMY3010), a sir2D strain (DMY3011), a Pgal::NRD1 strain containing plasmids with WT NRD1 (YSB2193), nrd1-102 (YSB2192), nrd1D2-165 (YSB2194), or
a nrd1D::KAN strain with plasmids encoding WT NRD1 (YSB2205), nrd1-102 (YSB2207), nrd1D2-165 (YSB2206), or nrd1D39-169 (YSB2211). The relevant
chromosomal genotype is shown above each bar while the plasmid encoded NRD1 allele is shown underneath. Error bars show standard deviation from three
repetitions.
RNAPII promoters are generally nucleosome free (Liu et al.,
2005).
Histone H3K4 trimethylation and H4 acetylation correlate with
RNAPII transcription (Guenther et al., 2007; Barski et al., 2007
and references therein). ChIP experiments showed peaks of
H3K4me3 near the mapped RNAPII transcription start sites
(Figure 5B, primers 15/16 and 22/23). Upon Nrd1 depletion,
a strong increase in H3K4me3 at the NTS1 CUT promoter region
is seen, correlating with the increased RNAPII. Histone methyla-
tion changes in the spacer region have also been reported in
cells lacking Sir2 (Li et al., 2006). In addition, H4 acetylation levels
are higher throughout the NTS regions when Nrd1 is depleted,
but there is a remarkably strong increase in the region from the
NTS1 CUT promoter continuing into the 30 end of the 35S
rRNA region (Figure 5C). Interestingly, the acetylation increase
is very strong at primer pairs 4 and 6, which are 2–3 kb beyond
readthrough transcription from the NTS1 CUT (Figure 4). The ob-
servation that CUT-related chromatin changes can extend be-
yond the actual site of transcription suggests a model for how
Nrd1 mutants can derepress reporter genes within heterochro-
matin. The passage of RNAPII disrupts the repressive chromatin
configuration locally, but the effect may be long range due to the
‘‘spreading’’ nature of Sir2 repression.
Mol
Because histone deacetylation in rDNA repeats suppresses
interrepeat recombination, we tested the effect of NRD1 muta-
tion on the loss of an ADE2 marker placed within rDNA. NRD1
alleles were expressed in the context of either a chromosomal
nrd1D deletion or metabolic depletion (Pgal::NRD1). Recombi-
nation was quantitated by counting frequency of marker loss,
indicated by red-sectored colonies (Huang et al., 2006). As
expected, cells lacking Sir2 showed a significant increase in
recombination compared to an isogenic wild-type strain (Fig-
ure 5D). Cells expressing a wild-type copy of NRD1 from a low
copy plasmid showed a slightly higher frequency of marker
loss compared to a strain carrying a genomic copy of NRD1,
possibly due to altered expression levels of Nrd1 produced
from a plasmid (Arigo et al., 2006a). However, cells expressing
mutant alleles of NRD1 showed significantly increased frequen-
cies of recombination (Figure 5D).
DISCUSSION
Recent findings challenge the view that heterochromatin is com-
pletely inaccessible to transcription factors. In S. cerevisiae,
RNAPII crosslinks to heterochromatic areas within rDNA, adja-
cent to telomeres, and at silent mating-type loci (Steinmetz
ecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier Inc. 319

Molecular Cell
Multiple Levels of Heterochromatic Silencing
et al., 2006). Based on ChIP experiments, Sekinger and Gross
(2001) proposed that heterochromatin blocks transcription
downstream of PIC formation, whereas Chen and Widom
(2005) suggested that only partial PICs formed. Results in other
organisms also indicate that silencing of heterochromatic genes
can occur downstream of RNAPII PIC assembly. Experiments in
Drosophila suggest that Polycomb silencing does not com-
pletely block PIC assembly (Dellino et al., 2004; Schwartz
et al., 2004), and maintenance of centromeric heterochromatin
in Schizosaccharomyces pombe is linked to ongoing RNAPII
transcription (Buhler et al., 2006; Motamedi et al., 2004).
Results presented here may help resolve this issue. Although
Sir2 reduces RNAPII crosslinking in rDNA, depletion or muta-
tions in the Nrd1 complex (Nrd1, Nab3, or Sen1) or exosome
(Rrp6 and Rrp4) reveal the presence of heterochromatic RNAPII
transcripts that are normally degraded by this pathway. These
mutations also derepress reporter genes integrated into the
rDNA array or telomeric regions, although no effect was seen
at silent mating-type loci. We note that, while our paper was be-
ing reviewed, another group also reported that exosome or Nrd1
mutants reveal rDNA CUTs (Houseley et al., 2007).
Our data suggest a three-level model for silencing of hetero-
chromatic genes in S. cerevisiae (Figure 6). At the first level,
Sir2 and associated proteins restrict DNA accessibility. However,
transcripts that escape this repression can be minimized at a sec-
ond level by the Nrd1/Sen1 termination complex, which prevents
RNAPII from extending too far within the heterochromatic region.
Finally, rapid RNA degradation by the exosome provides a third
level of repression. This multilevel process can also explain why
we and others see derepression of telomeric silencing in Nrd1
and exosome mutants (Houseley et al., 2007; Wang et al., 2008).
Although these three levels of repression could function inde-
pendently, our results suggest there are important connections.
The Nrd1/Sen1 termination complex is physically and function-
ally linked to the exosome (Vasiljeva and Buratowski, 2006), con-
necting levels 2 and 3. Here we show that failure to properly
terminate the NTS1 CUT via the Nrd1/Sen1 complex leads to
dramatic changes in chromatin modifications (connecting levels
2 and 1), with a particularly striking increase in histone acetylation
(Figure 5). These chromatin changes antagonize Sir2-dependent
repression, and the resulting increased accessibility of the chro-
matin leads to further activation of RNAPII transcription and
increased recombination between rDNA repeats.
Interestingly, increased acetylation is observed well into the
35S rDNA region, several kilobases beyond CUT transcription.
The long-range acetylation changes may occur via disruption
of the Sir2 complex RENT. RENT is localized to the replication
fork block within NTS1 by the Fob1 protein and to the RNAPI pro-
moter region, yet disruption of this complex increases histone
acetylation throughout the rDNA region (Huang and Moazed,
2003; Moazed, 2001a). Loss of Nrd1-mediated termination leads
to increased transcription through the replication fork block and
may disrupt RENT recruitment.
The proposed multilevel repression system in S. cerevisiae has
interesting parallels to heterochromatic repression in S. pombe.
In fission yeast, both heterochromatin formation and subsequent
degradation of nascent heterochromatic transcripts are carried
out by the RITS RNA interference (RNAi) machinery; paradoxi-
320 Molecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier In
cally, this mechanism actually requires ongoing RNAPII tran-
scription to generate siRNAs (Buhler et al., 2006; Motamedi
et al., 2004). Recent experiments in S. pombe revealed an addi-
tional RNAi-independent level of silencing mediated by the exo-
some (Buhler et al., 2007; Murakami et al., 2007; Wang et al.,
2008). Although two groups did not observe changes in histone
modification at heterochromatic reporter genes in S. pombe
exosome mutants (Buhler et al., 2007; Murakami et al., 2007), an-
other study found that H3K9 methylation was strongly reduced in
rrp6D cells (Nicolas et al., 2007). Although the components of the
exosome are conserved in all eukaryotes studied to date (Vana-
cova and Stefl, 2007), the RNAi machinery is absent from S. cer-
evisiae. Given the likelihood that the RNAi pathway existed early
in yeast evolution and was lost in the budding yeast lineage, it is
tempting to speculate that the exosome contribution to silencing
has been conserved from before the divergence.
Why is RNAPII transcription within rDNA repeats silenced, and
if silencing is important, why have at least two RNAPII promoters
persisted in this region? The most likely role for Sir2 deacetylation
in the ‘‘silenced’’ NTS is to regulate recombination and thereby
rDNA copy number, which fluctuates depending upon the cellular
growth rate (Kobayashi and Ganley, 2005; Kobayashi et al.,
Figure 6. Multiple Levels for Silencing of Heterochromatic Genes in
S. cerevisiae
1. Chromatin modifications (histone H4 and H3 deacetylation by Sir2) regulate
initial recruitment of RNAPII to DNA. Acetyl groups are shown as flags.
2. Nrd1/Sen1 complex triggers termination of cryptic RNAPII transcription that
does occur in heterochromatin.
3. The exosome contributes to repression by degrading RNA produced by
RNAPII despite Sir2 and Nrd1 actions.
c.

Molecular Cell
Multiple Levels of Heterochromatic Silencing
2004). Transcription can affect cohesin binding and thereby lead
to the rDNA repeat expansion and contraction (Kobayashi et al.,
2004; Huang et al., 2006), and transcription itself can stimulate
recombination (Huertas and Aguilera, 2003). Interestingly, the
Nrd1-regulated CUT RNAs coincide with the locations of two re-
combination hotspots: HOT1 E in NTS1 and HOT1 I in NTS2 (Ko-
bayashi and Ganley, 2005; Voelkel-Meiman et al., 1987). Our data
suggest that RNAPII transcription of the NTS1 CUT could directly
affect rDNA recombination. The NTS1 CUT region has lower nu-
cleosome density (Figure 5A), and the level of RNAPII at the CUT
is significantly higher when cells are grown in glucose versus ga-
lactose (Figure 4B). If favorable growth conditions increase CUT
transcription, the frequency of occasional readthrough events
would also increase. Transcripts detected in exosome mutants,
which do not have a termination defect (Kim et al., 2006), show
that RNAPII transcribes through the replication fork block and be-
yond. Transcription of the CUT correlates with increased histone
acetylation (Figure 5C), and increased template accessibility
could lead to a higher probability of recombination between
rDNA repeats.
In support of this model, elevated rDNA recombination fre-
quency is observed in NRD1 mutants (Figure 5D). Also, two re-
cent reports found that S. cerevisiae and S. pombe strains with
mutations in exosome or the exosome-associated TRAMP com-
plex exhibit variation in rDNA copy number (Houseley et al., 2007;
Wang et al., 2008). In contrast to our results with Nrd1 mutants,
Houseley et al. (2007) did not see changes in recombination fre-
quency in a TRAMP mutant. Based on genetic interactions be-
tween TRAMP and topoisomerase I, they proposed that NTS1
CUT expression might induce DNA damage at the replication
fork block, leading to changes in rDNA copy number. However,
another possibility is that exosome mutants have a small effect
on CUT expression or termination via interaction with the Nrd1/
Sen1 complex, leading to the changes in chromatin structure
we report here.
Another possible function for RNAPII transcription in the NTS
could be to regulate rRNA synthesis. Regulation is known to
occur at several levels. One mechanism controls the number of
active rRNA genes, and a second regulates the amount of
rRNA produced per gene (Russell and Zomerdijk, 2005). RNAPII
transcription occurs in active rDNA repeats (Figure 4D). The pas-
sage of RNAPII through the rDNA spacer region could affect ri-
bosome expression by changing nucleosome positions or his-
tone modifications at the RNAPI enhancer (within NTS1) or
promoter (within NTS2) (Dammann et al., 1995; Kobayashi and
Ganley, 2005; Voelkel-Meiman et al., 1987). It is also possible
that the intergenic RNA itself has a more direct role regulating
rRNA expression: intergenic transcripts produced by RNAPI in
mammalian cells were recently shown to recruit a histone mod-
ifying complex to rDNA promoter regions (Mayer et al., 2006).
The relationship between RNAPII and RNAPI transcription is
not well understood, but it is becoming evident that they are
not independent. RNAPI activity becomes dispensable if rRNA
synthesis is put under the control of a strong RNAPII promoter
(Lal et al., 2004). Furthermore, endogenous rDNA can be tran-
scribed by RNAPII when RNAPI initiation is impaired (Conrad-
Webb and Butow, 1995; Oakes et al., 1999; Vu et al., 1999).
Upon depletion of Nrd1, RNAPII-produced rRNA precursors
Mo
are inefficiently processed (Figure 1B), but under certain condi-
tions, these may be sufficient for viability.
The results presented here and in several other recent papers
(Camblong et al., 2007; Houseley et al., 2007; Martens et al.,
2004, 2005) show that transcription of noncoding regions in
yeast can have important regulatory functions. Although the
transcripts themselves may not have a function, the act of tran-
scription itself can affect protein interactions with nearby DNA to
influence important biological functions.
EXPERIMENTAL PROCEDURES
Yeast Strains and Plasmids
S. cerevisiae strains used in this study are listed in Table S1. Plasmids pJC580,
pJC643, and pJC720 were from J. Corden (Conrad et al., 2000) and
pRS314nrd1D39-169 from D. Brow (Steinmetz and Brow, 1998). A list of oligo-
nucleotides used is given in Table S2. The plasmid expressing Nrd1 with inter-
nal deletion from 151 to 214 aa was made by inverse PCR of pJC580 with
primers Nrd1_ 215aa_up and Nrd1_150aa_low to create yeast expression
plasmid pRS415-nrd1D151-214.
Silencing Assay
Silencing of the reporter genes inserted into the rDNA, telomeres, and the
mating-type loci (see Table S1 for strains) was monitored as described (van
Leeuwen and Gottschling, 2002). Cells were grown on rich medium to an
OD600 of 1.0, 107 cells were concentrated, and 10-fold serial dilutions were
spotted onto synthetic complete medium or medium containing 5-FOA. Plates
were incubated at 30�C for 2–3 days. The growth of the mutants strains was
compared to isogenic WT strains.
rDNA Recombination Assay
Assays were done as previously described (Huang et al., 2006). Cell cultures
from a white colony of strain UCC4602 or derivatives thereof (Table S1) were
grown to an OD600 of 1.0 and plated for single cells onto synthetic medium
containing low adenine concentration (27 mM). Plates were incubated for
3 days (5 days for mutants) at 30�C and then at 4�C for 2 days to enhance
development of red color. Sectors that lost the ADE2 reporter from the rDNA
array turn red due to accumulation of an intermediate in the adenine biosyn-
thetic pathway. The percentage of marker loss was calculated by dividing
the number of red-sectored colonies by the total number of colonies (at least
12,000). Completely red colonies indicate marker loss before plating and were
excluded. The experiment was repeated at least three times with different
isolates, and standard deviation was calculated for error bars.
Northern Blotting
Northern blot experiments were performed as previously described (Kim et al.,
2006; Vasiljeva and Buratowski, 2006). Primers used to generate probes are
described in Table S2. Strand-specific probes (probe 1 sense, probe 1 anti-
sense and probe 6 sense, probe 6 antisense) were made by single-step
PCR in the presence of [a-32P]dATP with only one primer (either sense or an-
tisense). For detection of URA3 mRNA, a 1:1 mixture of two probes was used.
One URA3-specific probe was made by digesting pRS316 with NdeI and ApaI.
A �500 bp restriction fragment complementary to the 50 part of URA3 was gel
purified and labeled (Megaprime DNA labeling system, Amersham Biosci-
ence). Another URA3-specific probe was made by PCR using URA3_89_up
and URA3_754_down primers. For SIR2 probe, the SIR2 ORF was PCR ampli-
fied from genomic DNA with oligos SIR2_50 and SIR2_30, the product was di-
gested by BamHI, and a�600 bp fragment was gel purified and labeled. RNAs
were resolved on acrylamide or agarose gels as indicated with Ambion Cen-
tury Plus markers used to determine sizes.
ChIP
ChIP experiments were done as previously described (Huang et al., 2006;
Keogh and Buratowski, 2004) with some modifications. Conditions were opti-
mized to be within a linear range of the exponential PCR curve. For rDNA loci,
lecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier Inc. 321

Molecular Cell
Multiple Levels of Heterochromatic Silencing
21 amplification cycles were carried out with 4 ml of decrosslinked template
DNA (1:4 dilution for immunoprecipitated, 1:200 dilution for INPUT DNA). For
the control noncoding ChrV region, 26 cycles were used for amplification
with 4 ml of DNA (no dilution for immunoprecipitated, 1:50 dilution for INPUT
DNA). Each chromatin sample was normalized first for the differences in
DNA concentration and those numbers were used to calculate fold enrichment
values for each reaction:
Relative enrichment = [rDNA(IP) / NoncodingChrV(IP)] / [rDNA(INPUT) / Non-
coding ChrV (INPUT)] 3 100.
The standard deviations were calculated from three independent experi-
ments. Primers used for PCR are listed in the Table S2. Histone antibodies
were from Upstate, anti-Rpb3 from Neoclone.
Supplemental Data
Supplemental Data include Supplemental Experimental Procedures, eight fig-
ures, and two tables and can be found with this article online at http://www.
molecule.org/cgi/content/full/29/3/313/DC1/.
ACKNOWLEDGMENTS
We thank D. Moazed, J. Corden, D. Brow, D. Libri, M. Nomura, and D. Gottsch-
ling for yeast strains and plasmids. We are grateful to K. Ahmad’s lab for help
with imaging and R. Buratowski for help with supplementary tables. We also
thank D. Moazed, M. Buhler, K. Mekhail, M. Oakes, M. Nomura, D. Libuda,
and F. Winston for helpful discussions. This research was supported by grants
GM46498 and GM56663 to S.B. from NIH. L.V. is a Special Fellow of the Leu-
kemia and Lymphoma Society. L.M.S. is a recipient of an EMBO Long Term
Postdoctoral Fellowship.
Received: October 5, 2007
Revised: December 12, 2007
Accepted: January 30, 2008
Published: February 14, 2008
REFERENCES
Arigo, J.T., Carroll, K.L., Ames, J.M., and Corden, J.L. (2006a). Regulation of
yeast NRD1 expression by premature transcription termination. Mol. Cell 21,
641–651.
Arigo, J.T., Eyler, D.E., Carroll, K.L., and Corden, J.L. (2006b). Termination of
cryptic unstable transcripts is directed by yeast RNA-binding proteins Nrd1
and Nab3. Mol. Cell 23, 841–851.
Barski, A., Cuddapah, S., Cui, K., Roh, T.Y., Schones, D.E., Wang, Z., Wei, G.,
Chepelev, I., and Zhao, K. (2007). High-resolution profiling of histone methyl-
ations in the human genome. Cell 129, 823–837.
Briggs, S.D., Bryk, M., Strahl, B.D., Cheung, W.L., Davie, J.K., Dent, S.Y.,
Winston, F., and Allis, C.D. (2001). Histone H3 lysine 4 methylation is mediated
by Set1 and required for cell growth and rDNA silencing in Saccharomyces
cerevisiae. Genes Dev. 15, 3286–3295.
Bryk, M., Briggs, S.D., Strahl, B.D., Curcio, M.J., Allis, C.D., and Winston, F.
(2002). Evidence that Set1, a factor required for methylation of histone H3, reg-
ulates rDNA silencing in S. cerevisiae by a Sir2-independent mechanism. Curr.
Biol. 12, 165–170.
Buhler, M., Verdel, A., and Moazed, D. (2006). Tethering RITS to a nascent
transcript initiates RNAi- and heterochromatin-dependent gene silencing.
Cell 125, 873–886.
Buhler, M., Haas, W., Gygi, S.P., and Moazed, D. (2007). RNAi-dependent and
-independent RNA turnover mechanisms contribute to heterochromatic gene
silencing. Cell 129, 707–721.
Camblong, J., Iglesias, N., Fickentscher, C., Dieppois, G., and Stutz, F. (2007).
Antisense RNA stabilization induces transcriptional gene silencing via histone
deacetylation in S. cerevisiae. Cell 131, 706–717.
Chen, L., and Widom, J. (2005). Mechanism of transcriptional silencing in
yeast. Cell 120, 37–48.
322 Molecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier I
Cioci, F., Vu, L., Eliason, K., Oakes, M., Siddiqi, I.N., and Nomura, M. (2003).
Silencing in yeast rDNA chromatin: reciprocal relationship in gene expression
between RNA polymerase I and II. Mol. Cell 12, 135–145.
Conrad, N.K., Wilson, S.M., Steinmetz, E.J., Patturajan, M., Brow, D.A.,
Swanson, M.S., and Corden, J.L. (2000). A yeast heterogeneous nuclear
ribonucleoprotein complex associated with RNA polymerase II. Genetics
154, 557–571.
Conrad-Webb, H., and Butow, R.A. (1995). A polymerase switch in the synthe-
sis of rRNA in Saccharomyces cerevisiae. Mol. Cell. Biol. 15, 2420–2428.
Dammann, R., Lucchini, R., Koller, T., and Sogo, J.M. (1995). Transcription in
the yeast rRNA gene locus: distribution of the active gene copies and chroma-
tin structure of their flanking regulatory sequences. Mol. Cell. Biol. 15, 5294–
5303.
de la Cruz, J., Kressler, D., Tollervey, D., and Linder, P. (1998). Dob1p (Mtr4p) is
a putative ATP-dependent RNA helicase required for the 30 end formation of
5.8S rRNA in Saccharomyces cerevisiae. EMBO J. 17, 1128–1140.
Dellino, G.I., Schwartz, Y.B., Farkas, G., McCabe, D., Elgin, S.C., and Pirrotta,
V. (2004). Polycomb silencing blocks transcription initiation. Mol. Cell 13, 887–
893.
Dror, V., and Winston, F. (2004). The Swi/Snf chromatin remodeling complex is
required for ribosomal DNA and telomeric silencing in Saccharomyces cerevi-
siae. Mol. Cell. Biol. 24, 8227–8235.
Dziembowski, A., Lorentzen, E., Conti, E., and Seraphin, B. (2007). A single
subunit, Dis3, is essentially responsible for yeast exosome core activity. Nat.
Struct. Mol. Biol. 14, 15–22.
Fritze, C.E., Verschueren, K., Strich, R., and Easton Esposito, R. (1997). Direct
evidence for SIR2 modulation of chromatin structure in yeast rDNA. EMBO J.
16, 6495–6509.
Guenther, M.G., Levine, S.S., Boyer, L.A., Jaenisch, R., and Young, R.A.
(2007). A chromatin landmark and transcription initiation at most promoters
in human cells. Cell 130, 77–88.
Holmes, S.G., Rose, A.B., Steuerle, K., Saez, E., Sayegh, S., Lee, Y.M., and
Broach, J.R. (1997). Hyperactivation of the silencing proteins, Sir2p and
Sir3p, causes chromosome loss. Genetics 145, 605–614.
Houseley, J., Kotovic, K., El Hage, A., and Tollervey, D. (2007). Trf4 targets
ncRNAs from telomeric and rDNA spacer regions and functions in rDNA
copy number control. EMBO J. 26, 4996–5006.
Huang, J., and Moazed, D. (2003). Association of the RENT complex with
nontranscribed and coding regions of rDNA and a regional requirement for
the replication fork block protein Fob1 in rDNA silencing. Genes Dev. 17,
2162–2176.
Huang, J., Brito, I.L., Villen, J., Gygi, S.P., Amon, A., and Moazed, D. (2006).
Inhibition of homologous recombination by a cohesin-associated clamp
complex recruited to the rDNA recombination enhancer. Genes Dev. 20,
2887–2901.
Huertas, P., and Aguilera, A. (2003). Cotranscriptionally formed DNA:RNA
hybrids mediate transcription elongation impairment and transcription-associ-
ated recombination. Mol. Cell 12, 711–721.
Keogh, M.C., and Buratowski, S. (2004). Using chromatin immunoprecipitation
to map cotranscriptional mRNA processing in Saccharomyces cerevisiae.
Methods Mol. Biol. 257, 1–16.
Kim, M., Vasiljeva, L., Rando, O.J., Zhelkovsky, A., Moore, C., and Buratowski,
S. (2006). Distinct pathways for snoRNA and mRNA termination. Mol. Cell 24,
723–734.
Kobayashi, T., and Ganley, A.R. (2005). Recombination regulation by
transcription-induced cohesin dissociation in rDNA repeats. Science 309,
1581–1584.
Kobayashi, T., Horiuchi, T., Tongaonkar, P., Vu, L., and Nomura, M. (2004).
SIR2 regulates recombination between different rDNA repeats, but not recom-
bination within individual rRNA genes in yeast. Cell 117, 441–453.
Lal, A., Mazan-Mamczarz, K., Kawai, T., Yang, X., Martindale, J.L., and
Gorospe, M. (2004). Concurrent versus individual binding of HuR and AUF1
to common labile target mRNAs. EMBO J. 23, 3092–3102.
nc.

Molecular Cell
Multiple Levels of Heterochromatic Silencing
Li, C., Mueller, J.E., and Bryk, M. (2006). Sir2 represses endogenous polymer-
ase II transcription units in the ribosomal DNA nontranscribed spacer. Mol.
Cell. Biol. 17, 3848–3859.
Liu, C.L., Kaplan, T., Kim, M., Buratowski, S., Schreiber, S.L., Friedman, N.,
and Rando, O.J. (2005). Single-nucleosome mapping of histone modifications
in S. cerevisiae. PLoS Biol. 3, e328. 10.1371/journal.pbio.0030328.
Martens, J.A., Laprade, L., and Winston, F. (2004). Intergenic transcription is
required to repress the Saccharomyces cerevisiae SER3 gene. Nature 429,
571–574.
Martens, J.A., Wu, P.Y., and Winston, F. (2005). Regulation of an intergenic
transcript controls adjacent gene transcription in Saccharomyces cerevisiae.
Genes Dev. 19, 2695–2704.
Matecic, M., Stuart, S., and Holmes, S.G. (2002). SIR2-induced inviability is
suppressed by histone H4 overexpression. Genetics 162, 973–976.
Mayer, C., Schmitz, K.M., Li, J., Grummt, I., and Santoro, R. (2006). Intergenic
transcripts regulate the epigenetic state of rRNA genes. Mol. Cell 22, 351–361.
Mitchell, P., Petfalski, E., Houalla, R., Podtelejnikov, A., Mann, M., and Toller-
vey, D. (2003). Rrp47p is an exosome-associated protein required for the 30
processing of stable RNAs. Mol. Cell. Biol. 23, 6982–6992.
Moazed, D. (2001a). Common themes in mechanisms of gene silencing. Mol.
Cell 8, 489–498.
Moazed, D. (2001b). Enzymatic activities of Sir2 and chromatin silencing. Curr.
Opin. Cell Biol. 13, 232–238.
Motamedi, M.R., Verdel, A., Colmenares, S.U., Gerber, S.A., Gygi, S.P., and
Moazed, D. (2004). Two RNAi complexes, RITS and RDRC, physically interact
and localize to noncoding centromeric RNAs. Cell 119, 789–802.
Murakami, H., Goto, D.B., Toda, T., Chen, E.S., Grewal, S.I., Martienssen,
R.A., and Yanagida, M. (2007). Ribonuclease activity of Dis3 is required for
mitotic progression and provides a possible link between heterochromatin
and kinetochore function. PLoS ONE 2, e317. 10.1371/journal.pone.0000317.
Nedea, E., He, X., Kim, M., Pootoolal, J., Zhong, G., Canadien, V., Hughes, T.,
Buratowski, S., Moore, C.L., and Greenblatt, J. (2003). Organization and
function of APT, a subcomplex of the yeast cleavage and polyadenylation
factor involved in the formation of mRNA and small nucleolar RNA 30-ends.
J. Biol. Chem. 278, 33000–33010.
Nicolas, E., Yamada, T., Cam, H.P., Fitzgerald, P.C., Kobayashi, R., and
Grewal, S.I. (2007). Distinct roles of HDAC complexes in promoter silencing,
antisense suppression and DNA damage protection. Nat. Struct. Mol. Biol.
14, 372–380.
Oakes, M., Siddiqi, I., Vu, L., Aris, J., and Nomura, M. (1999). Transcription
factor UAF, expansion and contraction of ribosomal DNA (rDNA) repeats,
and RNA polymerase switch in transcription of yeast rDNA. Mol. Cell. Biol.
19, 8559–8569.
Preuss, S., and Pikaard, C.S. (2007). rRNA gene silencing and nucleolar dom-
inance: insights into a chromosome-scale epigenetic on/off switch. Biochim.
Biophys. Acta 1769, 383–392.
Russell, J., and Zomerdijk, J.C. (2005). RNA-polymerase-I-directed rDNA
transcription, life and works. Trends Biochem. Sci. 30, 87–96.
Mo
Schwartz, Y.B., Kahn, T.G., Dellino, G.I., and Pirrotta, V. (2004). Polycomb
silencing mechanisms in Drosophila. Cold Spring Harb. Symp. Quant. Biol.
69, 301–308.
Sekinger, E.A., and Gross, D.S. (1999). SIR repression of a yeast heat shock
gene: UAS and TATA footprints persist within heterochromatin. EMBO J. 18,
7041–7055.
Sekinger, E.A., and Gross, D.S. (2001). Silenced chromatin is permissive to
activator binding and PIC recruitment. Cell 105, 403–414.
Singer, M.S., Kahana, A., Wolf, A.J., Meisinger, L.L., Peterson, S.E., Goggin,
C., Mahowald, M., and Gottschling, D.E. (1998). Identification of high-copy
disruptors of telomeric silencing in Saccharomyces cerevisiae. Genetics
150, 613–632.
Steinmetz, E.J., and Brow, D.A. (1998). Control of pre-mRNA accumulation by
the essential yeast protein Nrd1 requires high-affinity transcript binding and
a domain implicated in RNA polymerase II association. Proc. Natl. Acad. Sci.
USA 95, 6699–6704.
Steinmetz, E.J., Conrad, N.K., Brow, D.A., and Corden, J.L. (2001). RNA-
binding protein Nrd1 directs poly(A)-independent 30-end formation of RNA
polymerase II transcripts. Nature 413, 327–331.
Steinmetz, E.J., Warren, C.L., Kuehner, J.N., Panbehi, B., Ansari, A.Z., and
Brow, D.A. (2006). Genome-wide distribution of yeast RNA polymerase II
and its control by Sen1 helicase. Mol. Cell 24, 735–746.
Thiebaut, M., Kisseleva-Romanova, E., Rougemaille, M., Boulay, J., and Libri,
D. (2006). Transcription termination and nuclear degradation of cryptic unsta-
ble transcripts: a role for the nrd1-nab3 pathway in genome surveillance. Mol.
Cell 23, 853–864.
van Leeuwen, F., and Gottschling, D.E. (2002). Assays for gene silencing in
yeast. Methods Enzymol. 350, 165–186.
Vanacova, S., and Stefl, R. (2007). The exosome and RNA quality control in the
nucleus. EMBO Rep. 8, 651–657.
Vasiljeva, L., and Buratowski, S. (2006). Nrd1 interacts with the nuclear
exosome for 30 processing of RNA polymerase II transcripts. Mol. Cell 21,
239–248.
Venema, J., and Tollervey, D. (1995). Processing of pre-ribosomal RNA in
Saccharomyces cerevisiae. Yeast 11, 1629–1650.
Venema, J., and Tollervey, D. (1999). Ribosome synthesis in Saccharomyces
cerevisiae. Annu. Rev. Genet. 33, 261–311.
Voelkel-Meiman, K., Keil, R.L., and Roeder, G.S. (1987). Recombination-stim-
ulating sequences in yeast ribosomal DNA correspond to sequences regulat-
ing transcription by RNA polymerase I. Cell 48, 1071–1079.
Vu, L., Siddiqi, I., Lee, B.S., Josaitis, C.A., and Nomura, M. (1999). RNA
polymerase switch in transcription of yeast rDNA: role of transcription factor
UAF (upstream activation factor) in silencing rDNA transcription by RNA
polymerase II. Proc. Natl. Acad. Sci. USA 96, 4390–4395.
Wang, S.W., Stevenson, A.L., Kearsey, S.E., Watt, S., and Bahler, J. (2008).
Global role for polyadenylation-assisted nuclear RNA degradation in
post-transcriptional gene silencing. Mol. Cell. Biol. 28, 656–665.
lecular Cell 29, 313–323, February 15, 2008 ª2008 Elsevier Inc. 323




















