
Transcription factor XBP-1 is expressed during osteoblast differentiation and is transcriptionally...
7
UNCORRECTED PROOF Transcription factor XBP-1 is expressed during osteoblast differentiation and is transcriptionally regulated by parathyroid hormone (PTH) A. Zambelli a,b, * , E. Mongiardini b , S.N. Villegas b , N.G. Carri b , R.P. Boot-Handford a , G.A. Wallis a a Wellcome Trust Centre for Cell-Matrix Research, 2.205 Stopford Building, School of Biological Sciences, University of Manchester, Oxford Road, Manchester M13 9PT, UK b Instituto Multidisciplinario de Biologı´a Celular (IMBICE), CC403 (1900) La Plata, Argentina Received 10 September 2004; revised 3 March 2005; accepted 25 March 2005 13 Abstract 14 X-box-binding protein 1 (XBP-1) is a basic-region leucine zipper protein in the cyclic AMP response element binding protein/ 15 activating transcription factor (CREB/ATF) family of transcription factors involved in different cell-differentiation processes. We 16 have investigated the expression of XBP-1 in differentiating MC3T3-E1 osteoblastic cells. Cultures were treated with ascorbic acid 17 (AA) and b-glycerophosphate (BGP) to induce differentiation. Under these conditions, the basal transcription of xbp-1 increases at 18 day 2 following induction, peaks at day 5 and decreases thereafter. This result showed that xbp-1 gene is differentially expressed 19 during MC3T3-E1 cell differentiation. Detection of XBP-1 by immunofluorescence at days 0 (control culture without AA and BGP), 20 8 and 21 showed that the protein has a major cytoplasmic perinuclear location. In addition, xbp-1 is transcriptionally upregulated by 21 parathyroid hormone within 2.5 h of treatment and decreases thereafter. 22 Ó 2005 Published by Elsevier Ltd on behalf of International Federation for Cell Biology. 23 Keywords: XBP-1; PTH; Osteoblast differentiation 25 1. Introduction 26 X-box-binding protein 1 (XBP-1) is a basic-region 27 leucine zipper protein of the cyclic AMP response 28 element binding protein/activating transcription factor 29 (CREB/ATF) family of transcription factors. In vitro 30 studies have demonstrated down-regulation of the xbp-1 31 gene by B cell-specific activator protein (BSAP), di- 32 merization of XBP-1 protein with c-Fos (which also 33 belongs to the CREB/ATF family), and a decrease in 34 MHC class II gene expression when antisense xbp-1 35 sequences were introduced into Raji cells (Ono et al., 36 1991; Reimold et al., 1996). Disruption of the xbp-1 37 gene in mice resulted in embryonic lethality prior to 38 skeletogenesis and revealed that this transcription factor 39 is essential for liver growth (Reimold et al., 2000). 40 Furthermore, xbp-1-null B-lymphocytes fail to undergo 41 terminal differentiation to plasma cells (Reimold et al., 42 2001). 43 Studies on the transcription of xbp-1 revealed that it 44 is induced by activating transcription factor 6 (ATF6) YCBIR1349_proof ĸ 13 May 2005 ĸ 1/7 Abbreviations: XBP-1, X-box-binding protein 1; PTH, parathyroid hormone; PTHrP, PTH-related protein; PTH1R, PTH/PTHrP re- ceptor; AA, ascorbic acid; BGP, b-glycerophosphate. * Corresponding author. Present address: Advanta Semillas, Ruta 226 Km 60.5, CC30 (7620) Balcarce, Argentina. Tel.: C54 2266 430001; fax: C54 2266 430002. E-mail address: [email protected] (A. Zambelli). 1065-6995/$ - see 1 front matter Ó 2005 Published by Elsevier Ltd on behalf of International Federation for Cell Biology. doi: 2 10.1016/j.cellbi.2005.03.018 ARTICLE IN PRESS DTD 5 www.elsevier.com/locate/cellbi Cell Biology International -- (2005) ---e---
Transcript of Transcription factor XBP-1 is expressed during osteoblast differentiation and is transcriptionally...

13
141516171819202122
23
25
26272829
1
2
ARTICLE IN PRESSDTD 5
www.elsevier.com/locate/cellbi
Cell Biology International -- (2005) ---e---
ECTEDPROOF
Transcription factor XBP-1 is expressed during osteoblastdifferentiation and is transcriptionally regulated by
parathyroid hormone (PTH)
A. Zambelli a,b,*, E. Mongiardini b, S.N. Villegas b, N.G. Carri b,R.P. Boot-Handford a, G.A. Wallis a
a Wellcome Trust Centre for Cell-Matrix Research, 2.205 Stopford Building, School of Biological Sciences, University of Manchester,
Oxford Road, Manchester M13 9PT, UKb Instituto Multidisciplinario de Biologı́a Celular (IMBICE), CC403 (1900) La Plata, Argentina
Received 10 September 2004; revised 3 March 2005; accepted 25 March 2005
Abstract
X-box-binding protein 1 (XBP-1) is a basic-region leucine zipper protein in the cyclic AMP response element binding protein/activating transcription factor (CREB/ATF) family of transcription factors involved in different cell-differentiation processes. We
have investigated the expression of XBP-1 in differentiating MC3T3-E1 osteoblastic cells. Cultures were treated with ascorbic acid(AA) and b-glycerophosphate (BGP) to induce differentiation. Under these conditions, the basal transcription of xbp-1 increases atday 2 following induction, peaks at day 5 and decreases thereafter. This result showed that xbp-1 gene is differentially expressedduring MC3T3-E1 cell differentiation. Detection of XBP-1 by immunofluorescence at days 0 (control culture without AA and BGP),
8 and 21 showed that the protein has a major cytoplasmic perinuclear location. In addition, xbp-1 is transcriptionally upregulated byparathyroid hormone within 2.5 h of treatment and decreases thereafter.� 2005 Published by Elsevier Ltd on behalf of International Federation for Cell Biology.
Keywords: XBP-1; PTH; Osteoblast differentiation
R
303132333435363738394041424344
UNCOR1. Introduction
X-box-binding protein 1 (XBP-1) is a basic-regionleucine zipper protein of the cyclic AMP responseelement binding protein/activating transcription factor(CREB/ATF) family of transcription factors. In vitro
Abbreviations: XBP-1, X-box-binding protein 1; PTH, parathyroid
hormone; PTHrP, PTH-related protein; PTH1R, PTH/PTHrP re-
ceptor; AA, ascorbic acid; BGP, b-glycerophosphate.
* Corresponding author. Present address: Advanta Semillas, Ruta
226 Km 60.5, CC30 (7620) Balcarce, Argentina. Tel.: C54 2266
430001; fax: C54 2266 430002.
E-mail address: [email protected] (A.
Zambelli).
YCBIR1349_proof �
1065-6995/$ - see front matter � 2005 Published by Elsevier Ltd on behalf
doi:10.1016/j.cellbi.2005.03.018
studies have demonstrated down-regulation of the xbp-1gene by B cell-specific activator protein (BSAP), di-merization of XBP-1 protein with c-Fos (which alsobelongs to the CREB/ATF family), and a decrease inMHC class II gene expression when antisense xbp-1sequences were introduced into Raji cells (Ono et al.,1991; Reimold et al., 1996). Disruption of the xbp-1gene in mice resulted in embryonic lethality prior toskeletogenesis and revealed that this transcription factoris essential for liver growth (Reimold et al., 2000).Furthermore, xbp-1-null B-lymphocytes fail to undergoterminal differentiation to plasma cells (Reimold et al.,2001).
Studies on the transcription of xbp-1 revealed that itis induced by activating transcription factor 6 (ATF6)
13 May 2005 � 1/7
of International Federation for Cell Biology.

T
45464748495051525354555657585960616263646566676869707172737475767778798081828384858687888990
91
92
93949596
979899100101
102
103104105106107108109110111112113114115116117118119120121122123124125126127128129130131132
133
134135136137138139140141142143144145146
ARTICLE IN PRESSDTD 5
2 A. Zambelli et al. / Cell Biology International -- (2005) ---e---
UNCORREC
and the mRNA is spliced by IRE1 endonuclease inresponse to stress on the endoplasmic reticulum. Hence,the spliced form of xbp-1 can be translated and theresultant protein activates the unfolded protein response(UPR) (Yoshida et al., 2001). In situ hybridization onsections of mouse embryos demonstrated that xbp-1 istranscribed at quite high levels in osteoblasts andchondroblasts (Clauss et al., 1993). However, there isno information about the role of XBP-1 during bonedevelopment.
Parathyroid hormone (PTH) and PTH-related pro-tein (PTHrP) play crucial roles in regulating bonedevelopment and homeostasis. The importance ofPTHrP and its receptor in skeletal development hasbeen emphasized by gene deletion experiments in mice(Amizuka et al., 1994). Animals in which the PTHrPgene was ablated died immediately after birth, andshowed a skeletal dysplasia characterized by short limbsdue to accelerated chondrocyte maturation (Amizukaet al., 1994). In addition to the knockout studies, furtherinformation regarding the effects of PTH/PTHrP hasbeen obtained using osteoblastic cell lines. PTH orPTHrP binds to the PTH/PTHrP receptor (PTH1R) onosteoblasts and activates protein kinase A and Cpathways (Fitzpatrick and Bilezikian, 1999), ultimatelyresulting in the regulation of genes encoding, alkalinephosphatase, type I collagen, metalloproteinase, osteo-pontin and bone sialoprotein, and dihydroxyvitamin D3expression (Fitzpatrick and Bilezikian, 1999; Yamaguchiet al., 2000).
The anabolic action of PTH has been reported to bedependent on cAMP (Fitzpatrick and Bilezikian, 1999).Interestingly, PTH and PTHrP rapidly increase thelevels of c-fos and fra-2 mRNA in osteoblastic cell lines(McCauley et al., 1997, 2001). Since XBP-1 is known todimerize with c-Fos (Ono et al., 1991) and c-Fos itselfhas been proposed to play a role in osteoblastdifferentiation and bone formation (McCabe et al.,1995; Machwate et al., 1995; Grigoriadis et al., 1995;Matsuo et al., 2000; Winchester et al., 2000), it ispossible that XBP-1 may also play an important role inbone development. Consequently, the aim of this studywas to examine the expression of the xbp-1 gene in vitroduring osteoblast differentiation, the effect of PTH (1e34) on its transcription and the intracellular distributionof the XBP-1 protein.
2. Materials and methods
2.1. MC3T3-E1 cultures
MC3T3-E1 cells, a mouse osteoblastic cell line, werecultured in D-MEM medium (Gibco BRL) supple-mented with 10% fetal calf serum (FCS), 100 U/mlpenicillin, and 100 mg/ml streptomycin. Cells were
YCBIR1349_proof � 13
EDPROOF
seeded at 2! 104 cells/cm2 and grown in 10% FCS/D-MEM containing 50 mg/ml of ascorbic acid (AA) and10 mM of b-glycerophosphate (BGP) (Green et al.,2000). PTH treatment was performed using the humanPTH (1e34) peptide (Peninsula Laboratories) at 0.1 mM.
2.2. Immunofluorescence analysis
MC3T3-E1 cells treated with AA and BGP werecultured for 0 (control), 8 and 21 days. As the cultureshad reached confluence at days 8 and 21, immunoflu-orescence detection of protein in single cells was notpossible at these time points. Thus, cells from the 0, 8and 21 day cultures were trypsinized one day before theassay, then seeded on cover slips at a density of2! 104 cells/cm2, put into a culture chamber, andcultured overnight in a humid chamber to allow cell-attachment to the surface. The following day the cellswere fixed with 4% paraformaldehyde for 20 min. Thecells were rinsed in PBS and permeabilized with TritonX-100, 0.25%, for 15 min. After rinsing in PBS, a set ofslides was either incubated overnight at 4 �C witha rabbit polyclonal antibody raised against rat XBP-1(Reimold et al., 2000) diluted 1:1000 or with a rabbitpolyclonal antibody against rat c-Fos (Santa CruzBiotechnology) diluted 1:1000. Slides were washed withPBS and incubated with secondary biotinylated goatIgG anti-rabbit IgG at 1:1000 for 1 h at roomtemperature and then washed with PBS. Thereafter,they were incubated with the fluorochrome Texas Redconjugated to avidin for 45 min at room temperature.After rinsing with PBS, cell nuclei were counterstainedwith DAPI solution, mounted with Fluoromount(Vector) and observed in an inverted fluorescencemicroscope (Zeiss). Controls with no primary antibodyand with normal rabbit serum were included in allimmunodetections. No signal was detected in any of thecontrols.
2.3. RNA isolation and northern-blot analysis
Total RNA was isolated from cells by the guanidinethiocyanate/acid phenol extraction method (Chomczyn-ski and Sacchi, 1987). RNA was electrophoresed ona 1% (w/v) denaturing agarose gel containing 2.2 M offormaldehyde and transferred onto nitrocellulose mem-brane (Hybond N). The mouse xbp-1 cDNA probe wasobtained by RT-PCR from total RNA isolated fromMC3T3-E1 cultures and using the primers 5#-AATGGTCCTTTATCAGTTTCTC-3# and 5#-GCATTGTACTTTTTAATTGCAC-3# based on the mouse xbp-1 genesequence (GenBank accession AF027963). The 272-bp-long amplification product was cloned and sequencedto confirm that it corresponded to the xbp-1 gene. In
May 2005 � 2/7

T
147148149150151152153154155156157158159160161162163164165
166
167168169170171172173174
175
176177178179180181182183184185186187188189190191192193194195
196197198199200201202
ARTICLE IN PRESSDTD 5
3A. Zambelli et al. / Cell Biology International -- (2005) ---e---
UNCORREC
the same way, a c-fos cDNA probe was obtained byRT-PCR using the primers 5#-GGTTCATTATTGGAATTAAC-3# and 5#-CAACTTAAATGCTTTTATTG-3#(GenBank accession X06769). The c-fos cDNA frag-ment (445 bp long) was cloned and sequenced. A bovinepth1r probe was previously isolated from a subtractedgrowth-plate library (Hillarby et al., 1996). The clonewas sequenced confirming identity to the PTH1R.Radiolabeled probes were made using the randompriming method (Random Primers DNA LabelingSystem, Gibco BRL) using (a32P)dATP (NEN). North-ern hybridization of the filters with the xbp-1, c-fos andpth1r probes was performed at 68 �C (Sambrook et al.,1989). Blots were washed in 0.2! SSC/0.1% SDS andexposed to X-ray film at �70 �C with intensifyingscreens. Blots were stripped and reprobed using a cDNAprobe for 18S ribosomal RNA (Manuelidis and Ward,1984). A Kodak Digital Imaging system was used toquantify the signal intensity.
2.4. Statistical analysis
To measure the basal expression of xbp-1 mRNApost-mineralization, experiments were performed intriplicate and transcriptional regulation by PTH wasassayed in five independent experiments. In eachexperiment, treatments were performed in triplicate.The results were analyzed using ANOVA followed byTukeyeKramer multiple comparison test to determinestatistical differences between groups.
3. Results
Northern-blot analysis of total RNA isolated fromMC3T3-E1 osteoblastic cell cultures identified xbp-1mRNA. To evaluate whether this expression wasmodulated during osteoblast differentiation, MC3T3-E1 cells were cultured with media supplemented withAA and BGP, to induce osteoblast differentiation. TotalRNA was isolated after 0, 2, 5, 7 and 9 days of inductionand mRNA levels of xbp-1, c-fos and pth1r wereinvestigated by northern blotting. The expressionpatterns of the latter two genes have been consideredas osteoblast differentiation markers (McCabe et al.,1995; McCauley et al., 1996; Lynch et al., 1998; Purpuraet al. 2003; von Stechow et al., 2004). The analysisrevealed that, after 2 days of culture, the level of xbp-1mRNA increased about eightfold above that of thecontrol cultures (day 0, p! 0.05). The maximum xbp-1transcription was observed at day 5, although xbp-1mRNA was still significantly increased at day 7 comparedto the day 0 cells. After 9 days of culture, the xbp-1 levelsdid not differ significantly from the control (Fig. 1).
YCBIR1349_proof �
EDPROOF
The pattern of expression of xbp-1 was similar to thatof pth1r and c-fos. The transcription of pth1r wasincreased about fourfold after 2 days of culture withrespect to the day 0 control ( p! 0.05), and remainedsignificantly higher than the control at days 5, 7 and 9(Fig. 1, p! 0.05). The c-fos gene exhibited an increasein transcription at day 5 with respect to the control
a 0 2 5 7 9
xbp-1
18S
18S
18S
pth1r
c-fos
Day
b
0 2 5 7 9
0
1
2
3
4
c-fos
xbp-1
pth1r
Days in culture
Ge
ne
/1
8S
ra
tio
Fig. 1. MC3T3-E1 differentiation and xbp-1, c-fos and pth1r mRNAs
expression. (a) MC3T3-E1 cells were treated with ascorbic acid (AA)
and b-glycerophosphate (BGP). After 0, 2, 5, 7 and 9 days of culture,
total RNA was isolated for the analysis of xbp-1, c-fos and pth1r
expression by northern blot. (b) The intensity of each band was
quantified as described in Section 2 and the mean valuesG SEM of the
ratio between xbp-1, c-fos or pth1r mRNAs and their respective 18S
RNA signal intensity were plotted for each treatment day. The
autoradiograms shown are representative of three independent experi-
ments performed on different days.
13 May 2005 � 3/7
203204205206207208209210211212213214215216217218219220221222223224225226227228
229230231232233234235236237238239240241242243244245
246
247248249250251
ARTICLE IN PRESSDTD 5
4 A. Zambelli et al. / Cell Biology International -- (2005) ---e---
(undetectable signal), decreasing at days 7 and 9 (Fig. 1,p! 0.05).
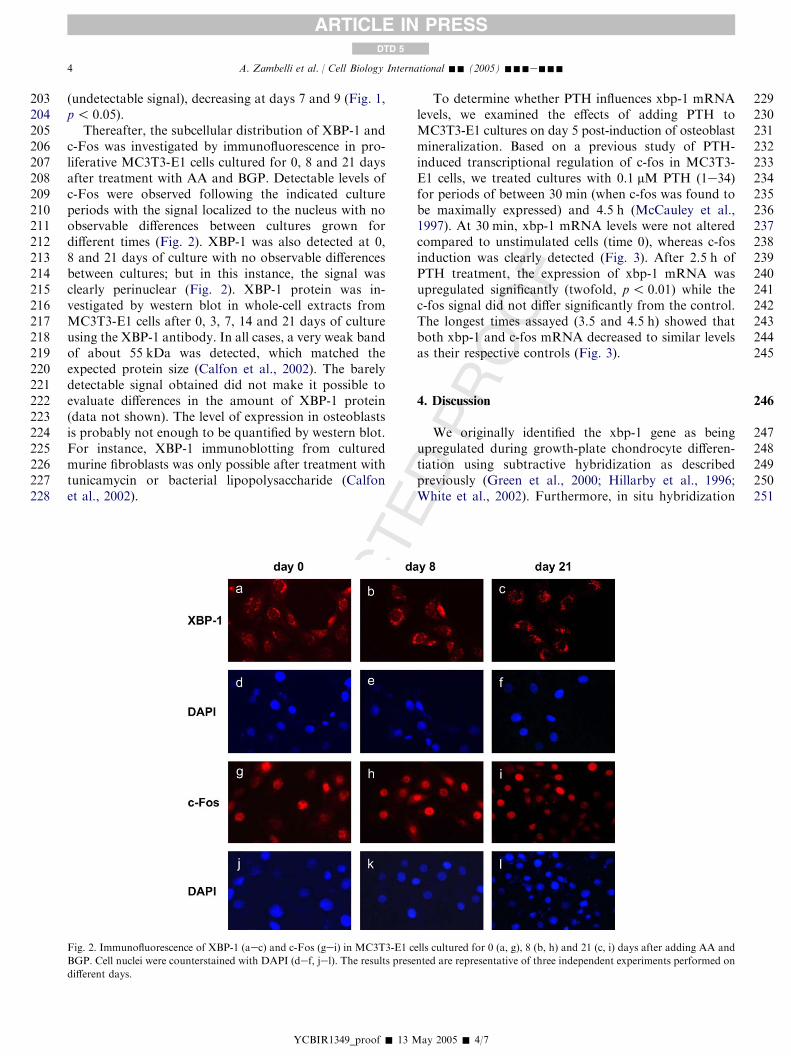
Thereafter, the subcellular distribution of XBP-1 andc-Fos was investigated by immunofluorescence in pro-liferative MC3T3-E1 cells cultured for 0, 8 and 21 daysafter treatment with AA and BGP. Detectable levels ofc-Fos were observed following the indicated cultureperiods with the signal localized to the nucleus with noobservable differences between cultures grown fordifferent times (Fig. 2). XBP-1 was also detected at 0,8 and 21 days of culture with no observable differencesbetween cultures; but in this instance, the signal wasclearly perinuclear (Fig. 2). XBP-1 protein was in-vestigated by western blot in whole-cell extracts fromMC3T3-E1 cells after 0, 3, 7, 14 and 21 days of cultureusing the XBP-1 antibody. In all cases, a very weak bandof about 55 kDa was detected, which matched theexpected protein size (Calfon et al., 2002). The barelydetectable signal obtained did not make it possible toevaluate differences in the amount of XBP-1 protein(data not shown). The level of expression in osteoblastsis probably not enough to be quantified by western blot.For instance, XBP-1 immunoblotting from culturedmurine fibroblasts was only possible after treatment withtunicamycin or bacterial lipopolysaccharide (Calfonet al., 2002).
YCBIR1349_proof � 13
DPROOF
To determine whether PTH influences xbp-1 mRNAlevels, we examined the effects of adding PTH toMC3T3-E1 cultures on day 5 post-induction of osteoblastmineralization. Based on a previous study of PTH-induced transcriptional regulation of c-fos in MC3T3-E1 cells, we treated cultures with 0.1 mM PTH (1e34)for periods of between 30 min (when c-fos was found tobe maximally expressed) and 4.5 h (McCauley et al.,1997). At 30 min, xbp-1 mRNA levels were not alteredcompared to unstimulated cells (time 0), whereas c-fosinduction was clearly detected (Fig. 3). After 2.5 h ofPTH treatment, the expression of xbp-1 mRNA wasupregulated significantly (twofold, p! 0.01) while thec-fos signal did not differ significantly from the control.The longest times assayed (3.5 and 4.5 h) showed thatboth xbp-1 and c-fos mRNA decreased to similar levelsas their respective controls (Fig. 3).
4. Discussion
We originally identified the xbp-1 gene as beingupregulated during growth-plate chondrocyte differen-tiation using subtractive hybridization as describedpreviously (Green et al., 2000; Hillarby et al., 1996;White et al., 2002). Furthermore, in situ hybridization
UNCORRECTE
Fig. 2. Immunofluorescence of XBP-1 (aec) and c-Fos (gei) in MC3T3-E1 cells cultured for 0 (a, g), 8 (b, h) and 21 (c, i) days after adding AA and
BGP. Cell nuclei were counterstained with DAPI (def, jel). The results presented are representative of three independent experiments performed on
different days.
May 2005 � 4/7

T
252253254255256257258259260261262263264
265266267268269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310311312313314315316317318319320
ARTICLE IN PRESSDTD 5
5A. Zambelli et al. / Cell Biology International -- (2005) ---e---
UNCORREC
studies have shown that xbp-1 is expressed in vivo byboth chondrocytes and osteoblasts during mouseskeletal development (Clauss et al., 1993). The patternof transcription of xbp-1 was found to be very similar tothat of the genes encoding the tissue inhibitor ofmetalloproteinase (TIMP), alkaline phosphatase (ALP)and osteonectin (SPARC) throughout development inbone and cartilage (Clauss et al., 1993). XBP-1, like c-Fos, is a basic-region leucine zipper protein of theCREB/ATF family of transcription factors; in fact,these transcription factors can form heterodimers (Onoet al., 1991; Vinson et al., 2002). Mice lacking the xbp-1gene die prior to skeletogenesis and thus give no insight
a 0 0.5 2.5 3.5 4.5Hrs
xbp-1
18S
c-fos
18S
b
Control 0.5 2.5 3.5 4.5
0
1
2
3
4
5
6
xbp-1
c-fos
*
*
Time of PTH stimulation (Hrs)
Gen
e/18S
ratio
Fig. 3. Effect of PTH (1e34) on xbp-1 and c-fos mRNA expression. (a)
MC3T3-E1 cells were treated for 0.5, 2.5, 3.5, and 4.5 h with 0.1 mM
PTH (1e34), total RNA isolated and northern blotted. (b) The
intensity of each xbp-1 and c-fos band was quantified and the mean
valuesG SEM of the ratio between xbp-1 or c-fos and their 18S RNA
signal intensity were plotted for the indicated PTH-stimulation times.
The autoradiograms shown are representative of three independent
experiments performed on different days.
YCBIR1349_proof �
EDPROOF
into the role of this transcription factor in boneformation (Reimold et al., 2000).
We therefore looked for expression of xbp-1 in theMC3T3-E1 cell line. This cell line is a clonal osteoblast-like line, which, when cultured in the presence ofAA and BGP, undergoes a specified pattern of dif-ferentiation, which comprises: proliferation (days 1e10);extracellular matrix maturation and mineralization(days 10e20); and apoptotic mediated tissue mainte-nance (beyond day 20) (McCabe et al., 1995; McCauleyet al., 1996; Franceschi and Iyer, 1992; Lynch et al.,1998). Also, this cell line expresses pth1r (McCauleyet al., 1996) and higher levels of pth1r and c-fos havebeen observed at days 1 and 8 of culture compared topost-proliferative differentiating stages (McCabe et al.,1995; McCauley et al., 1996; Lynch et al., 1998).MC3T3-E1 cells rapidly (30e45 min) upregulated c-fosmRNA levels upon exposure to 0.1 mM PTH (1e34).This increased c-fos expression is sustained for up to 1 hand decreases thereafter (McCauley et al., 1997).
We have found that MC3T3-E1 cells also differen-tially express xbp-1 during osteoblast differentiation andthat expression increases at 2, 5 and 7 days of cultureand decreases at day 9. The pattern of expression ofxbp-1 is similar to that of pth1r and c-fos (Fig. 1)indicating that xbp-1 is also differentially expressedduring osteoblast differentiation.
We next investigated the intracellular distribution ofthe XBP-1 and c-Fos proteins by immunofluorescence.Both proteins were identified in MC3T3-E1 osteoblastsduring the proliferative and maturation stages. Stainingfor c-Fos was clearly located in the nucleus. In contrast,XBP-1 staining was predominantly perinuclear and faintwithin the nucleus. This finding was unexpectedconsidering that XBP-1 has been classified as a tran-scription factor. Recently, the upregulation and over-expression of the hXBP-1 gene has been described inprimary breast cancers (Fujimoto et al., 2003). Immu-nohistochemistry demonstrated strong staining for thehXBP-1 protein in the cytoplasm of cancer cells that wasabsent in normal breast ductal epithelial and myoepi-thelial cells; however, no explanation for the predomi-nantly cytoplasmic location of XBP-1 in the cancer cellswas given (Fujimoto et al., 2003).
So far, c-Fos (a leucine zipper transcription factor)was the only transcription factor found to have directcytoplasmic function through a mechanism independentof its genomic AP-1 activity. In quiescent fibroblastcultures stimulated with FCS, c-Fos associates with theendoplasmic reticulum, regulating synthesis/replenish-ment of phospholipid molecules required for signaltransduction pathways, and later on regulates transcrip-tion of enzymes involved in cell-growth and morpho-logical differentiation (Bussolino et al., 2001; Borioliet al., 2002; Gil et al., 2004). Our present results showthat XBP-1, like c-Fos, is expressed during osteoblast
13 May 2005 � 5/7

321322323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372373374375376
377378379380381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
ARTICLE IN PRESSDTD 5
6 A. Zambelli et al. / Cell Biology International -- (2005) ---e---
UNCORREC
differentiation, which reinforces the previous finding ofFujimoto et al. (2003), constituting a new exampleof a transcription factor with a main cytoplasmiclocalization.
However, we cannot exclude that XBP-1 is alsopresent in the nucleus of osteoblasts, as the level may bebelow the detection limit of our immunofluorescentassay. It is difficult to explain the cytoplasmic functionof XBP-1 in osteoblasts. At the moment we can onlyspeculate that, as c-Fos does in growing fibroblasts,XBP-1 may modulate phospholipid degradation, affect-ing the formation of lipid second messenger productsinvolved in the signaling of osteoblast differentiation. Ithas recently been demonstrated that the phospholipidssphingosine-1-phosphate and lysophosphatidic acid pre-vent apoptosis in cultured osteoblasts (Grey et al.,2002). Thus, a possible function that remains to bedemonstrated is that XBP-1 could be associated with thesynthesis of phospholipid involved in the regulation ofosteoblast apoptosis.
PTH is known to play a critical role in osteoblastdifferentiation, in part through cAMP-dependant in-duction of c-fos. We therefore examined first whetherPTH could affect expression of xbp-1 and secondwhether the effects were similar to that seen for c-fos.We found that PTH does indeed increase xbp-1 mRNAlevels significantly. It is therefore possible that the effectsinduced by PTH are characterized by a shift in thepattern of expression of transcription factors capable offorming heterodimers that may modulate expression ofdifferent target genes in a temporal manner. However,since XBP-1 and c-Fos in osteoblasts seem to be locatedin different subcellular compartments, it would seemunlikely that XBP-1 is exerting its effect througha heterodimeric complex with c-fos. XBP-1 and c-Fosmay therefore be playing distinct and independent rolesin osteoblast differentiation.
The anabolic effect of PTH on bone and its use fortreatment of pathologies associated with bone-mass lossis well known (Neer et al., 2001; Finkelstein et al., 2003).It was proposed that the observed increased boneformation after treatment with PTH is, among othercauses, due to prevention of osteoblast apoptosis (Jilkaet al., 1999). The present results show that XBP-1 isupregulated by PTH and that the subcellular locationof the protein is mainly cytoplasmic. Based on thedocumented anti-apoptotic effects of both PTH andphospholipids on osteoblasts, the PTH/XBP-1 relation-ship shown here, and the observed cytoplasmic locationof XBP-1, we can propose that the anti-apoptotic effectof PTH could in part be produced through pathwaysinvolving the cytoplasmic activity of XBP-1.
The results reported here indicate that xbp-1 is a genedifferentially expressed during osteoblast differentiationand upregulated by PTH. Its cellular distribution mightindicate that XBP-1 acts not only as a transcription
YCBIR1349_proof � 1
TEDPROOF
factor in the nucleus, but may have additional extranu-clear functions. These data suggest that XBP-1 requiresfurther investigation to address its specific role, likeother leucine zipper transcription factors, during theprogression of osteoblast differentiation.
References
Amizuka N, Warshawsky H, Henderson JE, Goltzman D,
Karaplis AC. Parathyroid hormone-related peptide-depleted mice
show abnormal epiphyseal cartilage development and altered
endochondral bone formation. J Cell Biol 1994;126:1611e23.
Borioli GA, Fanani ML, Caputto BL, Maggio B. c-Fos is a surface
pressure-dependent diverter of phospholipase activity. Biochem
Biophys Res Commun 2002;295:964e9.Bussolino DF, Guido ME, Gil GA, Borioli GA, Renner ML,
Grabois VR, et al. c-Fos associates with the endoplasmic reticulum
and activates phospholipid metabolism. FASEB J 2001;15:556e8.
Calfon M, Zeng H, Urano F, Till JH, Hubbard SR, Harding HP, et al.
IRE1 couples endoplasmic reticulum load to secretory capacity by
processing the XBP-1 mRNA. Nature 2002;415:92e6.
Chomczynski P, Sacchi N. Single step method of RNA isolation by
acid guanidinium thiocyanateephenolechloroform extraction.
Anal Biochem 1987;162:156e9.
Clauss IM, Gravallese EM, Darling JM, Shapiro F, Glimcher MJ,
Glimcher LH. In situ hybridization studies suggest a role for the
basic region-leucine zipper protein hXBP-1 in exocrine glands and
skeletal development during mouse embryogenesis. Dev Dyn
1993;197:146e56.
Finkelstein JS, Hayes A, Hunzelman JL, Wyland JJ, Lee H, Neer RM.
The effects of parathyroid hormone, alendronate, or both in men
with osteoporosis. N Engl J Med 2003;349:1216e26.
Fitzpatrick LA, Bilezikian JP. Parathyroid hormone: structure,
function and dynamics. In: Seibel MJ, Robins SP, Bilezikian JP,
editors. Dynamics of bone and cartilage metabolism. San Diego:
Academic Press; 1999. p. 187.
Franceschi RT, Iyer BS. Relationship between collagen synthesis and
expression of the osteoblast phenotype in MC3T3-E1 cells. J Bone
Miner Res 1992;7:235e46.
Fujimoto T, Onda M, Nagai H, Nagahata T, Ogawa K, Emi M.
Upregulation and overexpression of human X-box binding protein
1 (hXBP-1) gene in primary breast cancers. Breast Cancer 2003;
10:301e6.
Gil GA, Bussolino DF, Portal MM, Pecchio AA, Renner ML,
Borioli GA, et al. c-Fos activated phospholipid synthesis is
required for neurite elongation in differentiating PC12 cells. Mol
Biol Cell 2004;15:1881e94.
Green H, Canfield AE, Hillarby MC, Grant ME, Boot-Handford RP,
Freemont AJ, et al. The ribosomal protein QM is expressed
differentially during vertebrate endochondral bone development.
J Bone Miner Res 2000;15:1066e75.
Grey A, Chen Q, Callon K, Xu X, Reid IR, Cornish J. The
phospholipids sphingosine-1-phosphate and lysophosphatidic acid
prevent apoptosis in osteoblastic cells via a signaling pathway
involving Gi proteins and phosphatidylinositol-3 kinase. Endocri-
nology 2002;143:4755e63.
Grigoriadis AE, Wang ZQ, Wagner EF. Fos and bone cell
development: lessons from a nuclear oncogene. Trends Genet 1995;
11:436e41.
Hillarby MC, King KE, Brady G, Grant ME, Wallis GA, Boot-
Handford RP. Localization of gene expression during endochon-
dral ossification. Ann N Y Acad Sci 1996;785:263e5.
Jilka RL, Weinstein RS, Bellido T, Roberson P, Parfitt AM,
Manolagas SC. Increased bone formation by prevention of
3 May 2005 � 6/7

T
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
ARTICLE IN PRESSDTD 5
7A. Zambelli et al. / Cell Biology International -- (2005) ---e---
C
osteoblast apoptosis with parathyroid hormone. J Clin Invest
1999;104:439e46.
Lynch MP, Capparelli C, Stein JL, Stein GS, Lian JB. Apoptosis
during bone-like tissue development in vitro. J Cell Biochem
1998;68:31e49.Machwate M, Jullienne A, Moukhtar M, Marie PJ. Temporal
variation of c-fos proto-oncogene expression during osteoblast
differentiation and osteogenesis in developing rat bone. J Cell
Biochem 1995;57:62e70.
Manuelidis L, Ward DC. Chromosomal and nuclear distribution of
the HindIII 1.9 kb human DNA repeat segment. Chromosoma
1984;91:28e38.Matsuo K, Owens JM, Tonko M, Elliot C, Chambers TJ, Wagner EF.
Fosl1 is a transcriptional target of c-Fos during osteoclast
differentiation. Nat Genet 2000;24:184e7.
McCabe LR, Kockz M, Lian J, Stein J, Stein G. Selective expression of
fos- and jun-related genes during osteoblast proliferation and
differentiation. Exp Cell Res 1995;218:255e62.
McCauley LK, Koh AJ, Beecher CA, Cui Y, Rosol TJ, Franceschi RT.
PTH/PTHrP receptor is temporally regulated during osteoblast
differentiation and is associated with collagen synthesis. J Cell
Biochem 1996;61:638e47.
McCauley LK, Koh AJ, Beecher CA, Rosol TJ. Proto-oncogene c-fos
is transcriptionally regulated by parathyroid hormone (PTH) and
PTH-related protein in a cyclic adenosine monophosphate-
dependent manner in osteoblastic cells. Endocrinology 1997;138:
5427e33.McCauley K, Koh-Paige AJ, Chen H, Chen C, Ontiveros C, Irwin R,
et al. Parathyroid hormone stimulates fra-2 expression in osteo-
blastic cells in vitro and in vivo. Endocrinology 2001;142:1975e81.Neer RM, Arnaud CD, Zanchetta JR, Prince R, Gaich GA,
Reginster JY, et al. Effect of parathyroid hormone (1e34) on
fractures and bone mineral density in postmenopausal women with
osteoporosis. N Engl J Med 2001;344:1434e41.Ono SJ, Liou HC, Davidon R, Strominger JL, Glimcher LH. Human
X-box-binding protein 1 is required for the transcription of a subset
of human class II major histocompatibility genes and forms
heterodimer with c-fos. Proc Natl Acad Sci USA 1991;88:4309e12.
UNCORRE
YCBIR1349_proof �
EDPROOF
Purpura KA, Zandstra PW, Aubin JE. Fluorescence activated cell
sorting reveals heterogeneous and cell non-autonomous osteopro-
genitor differentiation in fetal rat calvaria cell populations. J Cell
Biochem 2003;90:109e20.
Reimold AM, Ponath PD, Li YS, Hardy RR, David CS,
Strominger JL, et al. Transcription factor B cell lineage-specific
activator protein regulates the gene for human X-box binding
protein 1. J Exp Med 1996;183:393e401.Reimold AM, Etkin A, Clauss I, Perkins A, Friend DS, Zhang J, et al.
An essential role in liver development for transcription factor
XBP-1. Genes Dev 2000;14:152e7.
Reimold AM, Iwakoshi NN, Manis J, Vallabhajosyula P, Szomolanyi-
Tsuda E, Gravallese EM, et al. Plasma cell differentiation requires
the transcription factor XBP-1. Nature 2001;412:300e7.
Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory
manual. New York: Cold Spring Harbor; 1989.
Vinson C, Myakishev M, Acharya A, Mir AA, Moll JR, Bonovich M.
Classification of human B-ZIP proteins based on dimerization
properties. Mol Cell Biol 2002;22:6321e35.von Stechow D, Zurakowski D, Pettit AR, Muller R, Gronowicz G,
Chorev M, et al. Differential transcriptional effects of PTH and
estrogen during anabolic bone formation. J Cell Biochem 2004;
15:476e90.White AH, Watson RE, Newman B, Freemont AJ, Wallis GA.
Annexin VIII is differentially expressed by chondrocytes in the
mammalian growth plate during endochondral ossification and in
osteoarthritic cartilage. J Bone Miner Res 2002;17:1851e8.Winchester SK, Selvamurugan N, D’alonzo RC, Partridge NC.
Developmental regulation of collagenase-3 mRNA in normal,
differentiating osteoblasts through the activator protein-1 and the
runt domain binding sites. J Biol Chem 2000;275:23310e8.
Yamaguchi A, Komori T, Suda T. Regulation of osteoblast
differentiation mediated by bone morphogenic protein, hedgehogs,
and Cbfa1. Endocr Rev 2000;21:393e411.Yoshida H, Matsui T, Yamamoto A, Okada T, Mori K. XBP1 mRNA
is induced by ATF6 and spliced by IRE1 in response to ER stress
to produce a highly active transcription factor. Cell 2001;107:
881e891.
13 May 2005 � 7/7




















