
Trace and soft body fossils from the Pedroche Formation (Ovetian, Lower Cambrian of the Sierra de...
26
Trace and soft body fossils from the Pedroche Formation (Ovetian, Lower Cambrian of the Sierra de Córdoba, S Spain) and their relation to the Pedroche event Ichnofossiles et fossiles à corps mous de la Formation Pedroche (Ovétien, Cambrien Inférieur de la Sierra de Córdoba, sud de l’Espagne) et leur relation avec l’événement Pedroche José Antonio Gámez Vintaned a, * , Eladio Liñán a , Eduardo Mayoral b , María Eugenia Dies a,d , Rodolfo Gozalo c , Fernando Muñiz b a Departamento de ciencias de la tierra, Área y museo de paleontología, facultad de ciencias, universidad de Zaragoza, E-50009 Zaragoza, Spain b Departamento de geodinámica y paleontología, facultad de ciencias experimentales, campus de El Carmen, universidad de Huelva, avenida de las Fuerzas Armadas, s/n, E-21071 Huelva, Spain c Departamento de geología, facultad de Ciencias biológicas, universitat de València, E-46100 Burjassot, Spain d Department of geology, Lund university, Sölvegatan 12, SE-22362 Lund, Sweden Received 13 October 2004; accepted 22 April 2005 Available online 22 June 2006 Abstract The low Lower Cambrian rocks from the Sierra de Córdoba contain one of the best successions in Europe, which consists of well exposed mixed facies with abundant fossil assemblages showing long stratigraphical ranges throughout the Pedroche Formation. These assemblages in- clude diverse Ovetian archaeocyaths, trilobites, small shelly fossils, calcimicrobes, trace fossils and stromatolites. Trace fossils are still poorly known, and thus they are the main objective of this work. Ichnological data are obtained from the Arroyo de Pedroche 1, Arroyo de Pedroche 2 and Puente de Hierro sections. Trace fossils include the ichnogenera Bergaueria, aff. Bilinichnus, Cochlichnus, aff. Cosmorhaphe?, Cylindrich- nus, Dactyloidites, Dimorphichnus, Diplichnites, Monocraterion, Palaeophycus, aff. Phycodes, Planolites, Psammichnites, Rusophycus, Sko- lithos, Torrowangea and Treptichnus, as well as faecal pellets, meniscate trace fossils and others. They are abundant in shales and sandstones, and indicate important changes in the benthic conditions with respect to the underlying Torreárboles Formation. Changes in fossil assemblages within Member I of the Pedroche Formation indicate palaeoecological disruptions, which led to the disappearance of numerous archaeocyath species and the decrease of stromatolite biodiversity. This was followed by dominance of trilobite and brachiopod assemblages, accompanied by trace fossils of the Psammichnites ichnosp. A ichnoassociation. This biotic turnover (Pedroche event) occurred at the lower part of the archae- ocyath Zone III, within the Bigotina bivallata biozone. The diagnoses of the ichnospecies Cochlichnus anguineus and Dactyloidites cabanasi are emended. © 2006 Elsevier SAS. All rights reserved. Résumé Les roches du Cambrien inférieur basal dans la Sierra de Córdoba contiennent une des meilleures successions en Europe. Elle se compose de faciès mixtes bien exposées et d’abondants assemblages de fossiles qui montrent les longues distributions stratigraphiques dans la Formation Pedroche. Ces assemblages sont composés d’archaeocyathes, de trilobites, de small shelly fossils, de calcimicrobes, de traces fossiles et de stromatolites de l’Ovetian. Les traces fossiles sont encore très peu connues et, par conséquent, elles sont ici notre principal objectif. Les traces fossiles de trois coupes sont étudiées : coupes de l’Arroyo de Pedroche 1, de Arroyo de Pedroche 2 et de Puente de Hierro. Des traces fossiles (lesquelles incluent les ichnogenres Bergaueria, aff. Bilinichnus, Cochlichnus, aff. Cosmorhaphe?, Cylindrichnus, Dactyloidites, Dimorphichnus, http://france.elsevier.com/direct/GEOBIO/ Geobios 39 (2006) 443–468 * Corresponding author. E-mail address: [email protected] (J.A. Gámez Vintaned). 0016-6995/$ - see front matter © 2006 Elsevier SAS. All rights reserved. doi:10.1016/j.geobios.2005.04.004
Transcript of Trace and soft body fossils from the Pedroche Formation (Ovetian, Lower Cambrian of the Sierra de...

http://france.elsevier.com/direct/GEOBIO/
Geobios 39 (2006) 443–468
* CorrespE-mail a
0016-6995/doi:10.1016
Trace and soft body fossils from the Pedroche Formation
onding author.ddress: game
$ - see front m/j.geobios.200
(Ovetian, Lower Cambrian of the Sierra de Córdoba,
S Spain) and their relation to the Pedroche eventIchnofossiles et fossiles à corps mous de la Formation Pedroche(Ovétien, Cambrien Inférieur de la Sierra de Córdoba,
sud de l’Espagne) et leur relation avec l’événement Pedroche
José Antonio Gámez Vintaned a,*, Eladio Liñán a, Eduardo Mayoral b, María Eugenia Dies a,d,Rodolfo Gozalo c, Fernando Muñiz b
aDepartamento de ciencias de la tierra, Área y museo de paleontología, facultad de ciencias, universidad de Zaragoza, E-50009 Zaragoza, Spain
z@uniza
atter ©5.04.004
bDepartamento de geodinámica y paleontología, facultad de ciencias experimentales,
campus de El Carmen, universidad de Huelva, avenida de las Fuerzas Armadas, s/n, E-21071 Huelva, SpaincDepartamento de geología, facultad de Ciencias biológicas, universitat de València, E-46100 Burjassot, SpaindDepartment of geology, Lund university, Sölvegatan 12, SE-22362 Lund, Sweden
Received 13 October 2004; accepted 22 April 2005Available online 22 June 2006
Abstract
The low Lower Cambrian rocks from the Sierra de Córdoba contain one of the best successions in Europe, which consists of well exposedmixed facies with abundant fossil assemblages showing long stratigraphical ranges throughout the Pedroche Formation. These assemblages in-clude diverse Ovetian archaeocyaths, trilobites, small shelly fossils, calcimicrobes, trace fossils and stromatolites. Trace fossils are still poorlyknown, and thus they are the main objective of this work. Ichnological data are obtained from the Arroyo de Pedroche 1, Arroyo de Pedroche 2and Puente de Hierro sections. Trace fossils include the ichnogenera Bergaueria, aff. Bilinichnus, Cochlichnus, aff. Cosmorhaphe?, Cylindrich-nus, Dactyloidites, Dimorphichnus, Diplichnites, Monocraterion, Palaeophycus, aff. Phycodes, Planolites, Psammichnites, Rusophycus, Sko-lithos, Torrowangea and Treptichnus, as well as faecal pellets, meniscate trace fossils and others. They are abundant in shales and sandstones,and indicate important changes in the benthic conditions with respect to the underlying Torreárboles Formation. Changes in fossil assemblageswithin Member I of the Pedroche Formation indicate palaeoecological disruptions, which led to the disappearance of numerous archaeocyathspecies and the decrease of stromatolite biodiversity. This was followed by dominance of trilobite and brachiopod assemblages, accompanied bytrace fossils of the Psammichnites ichnosp. A ichnoassociation. This biotic turnover (Pedroche event) occurred at the lower part of the archae-ocyath Zone III, within the Bigotina bivallata biozone. The diagnoses of the ichnospecies Cochlichnus anguineus and Dactyloidites cabanasi areemended.© 2006 Elsevier SAS. All rights reserved.
Résumé
Les roches du Cambrien inférieur basal dans la Sierra de Córdoba contiennent une des meilleures successions en Europe. Elle se compose defaciès mixtes bien exposées et d’abondants assemblages de fossiles qui montrent les longues distributions stratigraphiques dans la FormationPedroche. Ces assemblages sont composés d’archaeocyathes, de trilobites, de small shelly fossils, de calcimicrobes, de traces fossiles et destromatolites de l’Ovetian. Les traces fossiles sont encore très peu connues et, par conséquent, elles sont ici notre principal objectif. Les tracesfossiles de trois coupes sont étudiées : coupes de l’Arroyo de Pedroche 1, de Arroyo de Pedroche 2 et de Puente de Hierro. Des traces fossiles(lesquelles incluent les ichnogenres Bergaueria, aff. Bilinichnus, Cochlichnus, aff. Cosmorhaphe?, Cylindrichnus, Dactyloidites, Dimorphichnus,
r.es (J.A. Gámez Vintaned).
2006 Elsevier SAS. All rights reserved.

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468444
Diplichnites, Monocraterion, Palaeophycus, aff. Phycodes, Planolites, Psammichnites, Rusophycus, Skolithos, Torrowangea et Treptichnus, ainsique des pelotes fécales, traces fossiles ménisquées et autres) sont abondantes dans les argilites et grès, et indiquent des changements importantsdans les conditions du benthos par rapport à la Formation Torreárboles subjacente. Les assemblages fossiles dans le Membre I de la FormationPedroche indiquent des changements paléoécologiques qui conduisent à la disparition de nombreuses espèces d’archaeocyathes et à la diminutionde la biodiversité des stromatolites. Ensuite, les associations de trilobites et de brachiopodes sont dominantes, ainsi que les traces fossiles del’ichnoassociation à Psammichnites ichnosp. A. Cette crise écologique (événement Pedroche) a eu lieu à la limite entre les zones II et III d’ar-chaeocyathes, dans la biozone à trilobites de Bigotina bivallata. On émende les diagnoses de l’ichnoespèces Cochlichnus anguineus and Dacty-loidites cabanasi.© 2006 Elsevier SAS. All rights reserved.
Keywords: Spain; Lower Cambrian; Palaeoecology; Trace fossils; Pedroche event
Mots clés : Espagne ; Cambrien inférieur ; Paléoécologie ; Traces fossiles ; Événement Pedroche
1. Introduction
The Sierra de Córdoba is among the most important areasfor understanding the Lower Cambrian palaeontology of theIberian Peninsula. Since Hernández Pacheco (1907) found ar-chaeocyaths in limestones near the city of Córdoba, a numberof authors described additional archaeocyaths, crustaceans,trace fossils, trilobites, stromatolites, small shelly fossils, bra-chiopods, and soft-bodied fossils (for references see Liñán etal., 2005). In the Pedroche Formation (Liñán Guijarro, 1978),calcareous and siliciclastic beds alternate; numerous horizonsbear archaeocyaths and trilobites; they are well exposed alongthe Arroyo de Pedroche (N of Córdoba), in one of the bestLower Cambrian successions of western Europe. Moreover,the Sierra de Córdoba is the type area for the Corduban andOvetian stages. The biostratigraphic and biochronological unitsestablished allow international correlation with northernFrance, Sardinia, Morocco, Siberia, Antarctica and SW China(Perejón, 1986; Moreno-Eiris, 1987d; Pillola, 1991; Liñán etal., 2005).
The aim of this paper is to describe the ichnology of Mem-ber I of the Pedroche Formation in three selected sections(Fig. 1) where trilobites associated with archaeocyaths havebeen also studied, and to describe the palaeoecology of thelower Ovetian substage.
2. Geological setting
The geology of the Sierra de Córdoba was described in theclassic works of Liñán Guijarro (1974, 1978) and synthesizedby Perejón et al. (1996). The Neoproterozoic Volcanic–Sedi-mentary Complex unconformably underlies the Lower Cam-brian sequence, which consists (from bottom to top) of the Tor-reárboles, Pedroche and Santo Domingo formations.
The Torreárboles Formation is composed of conglomerate,sandstone, arkose and shale. Three Corduban ichnoassem-blages were described by Fedonkin et al. (1985). The firstone contains Cochlichnus ichnosp., Planolites ichnosp. andSkolithos ichnosp., and the second Phycodes pedum Seilacher,1955, Phycodes palmatum (Hall, 1852), Planolites cf. bever-leyensis (Billings, 1862) and Treptichnus ichnosp. The ichno-logical contents of both assemblages suggest a Lower Cordu-ban age. The third ichnoassemblage is characterised by the first
record of Rusophycus ichnosp. (indicative of an Upper Cordu-ban age; Liñán et al., 1993: Fig. 3), accompanied by Bilinich-nus ichnosp. and the soft-bodied metazoan Tiernavia tiernaeFedonkin in Fedonkin et al., 1985.
The Pedroche Formation conformably overlies the Torreár-boles Formation. It is composed of limestone and shale, withscarce sandstone and dolostone, and contains archaeocyaths,trilobites (aff. Bigotinella, Bigotina, Lemdadella, Serrania,Eoredlichia and Neoredlichiidae), brachiopods (Paterina), al-gae, calcimicrobes (Renalcis and Epiphyton), small shelly fos-sils, bradoriids, hyolithids, columnar stromatolites (Vetella cf.sarfatiae Schmitt, 1979, Charaulachia cordubensis Schmitt,1983 and Vetella nodosa Schmitt, 1983, all defined at the Ar-royo de Pedroche 1 section), trace fossils (Bergaueria, aff. Bi-linichnus, Cochlichnus, aff. Cosmorhaphe?, Cylindrichnus,Dactyloidites, Dimorphichnus, Diplichnites, Monocraterion,Palaeophycus, aff. Phycodes, Planolites, Psammichnites, Ru-sophycus, Skolithos, Torrowangea, Treptichnus, faecal pellets,meniscate trace fossils and others), and the probable soft-bod-ied coelenterates Tiernavia and aff. Tiernavia. Fossil assem-blages suggest an early Ovetian age for the Pedroche Forma-tion.
The Santo Domingo Formation conformably overlies thePedroche Formation and is composed of red shale, dolostoneand chert-bearing limestone. Scarce small shelly fossils, stro-matolites (Stratifera), calcimicrobes (Renalcis) and brachio-pods have been reported from the Member Orive (García-Her-nández and Liñán, 1983).
The Castellar Formation lies conformably above the SantoDomingo Formation and is composed of sandstone and con-glomerate with trace fossils. It records the uppermost LowerCambrian.
Basal sandstones and siltstones of the conformably overly-ing Los Villares Formation contain Middle Cambrian trilobitesand trace fossils (Liñán Guijarro, 1978; Liñán et al., 1995; Li-ñán Guijarro et al., 2004).
3. Stratigraphy and biochronology
The ichnofossils studied herein were collected from Mem-ber I of the Pedroche Formation at three sections (Figs. 3–5).
The Puente de Hierro section (PH) is the classic localitywhere early Lower Cambrian trilobites of Sierra Morena were
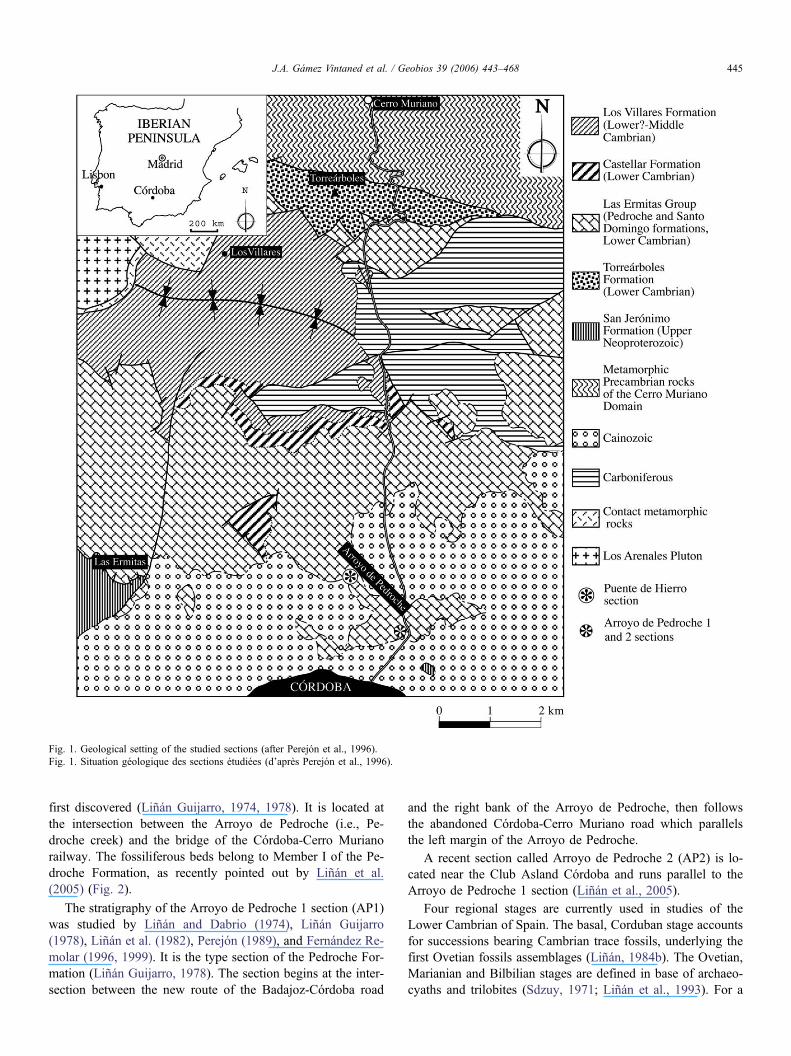
Fig. 1. Geological setting of the studied sections (after Perejón et al., 1996).Fig. 1. Situation géologique des sections étudiées (d’après Perejón et al., 1996).
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 445
first discovered (Liñán Guijarro, 1974, 1978). It is located atthe intersection between the Arroyo de Pedroche (i.e., Pe-droche creek) and the bridge of the Córdoba-Cerro Murianorailway. The fossiliferous beds belong to Member I of the Pe-droche Formation, as recently pointed out by Liñán et al.(2005) (Fig. 2).
The stratigraphy of the Arroyo de Pedroche 1 section (AP1)was studied by Liñán and Dabrio (1974), Liñán Guijarro(1978), Liñán et al. (1982), Perejón (1989), and Fernández Re-molar (1996, 1999). It is the type section of the Pedroche For-mation (Liñán Guijarro, 1978). The section begins at the inter-section between the new route of the Badajoz-Córdoba road
and the right bank of the Arroyo de Pedroche, then followsthe abandoned Córdoba-Cerro Muriano road which parallelsthe left margin of the Arroyo de Pedroche.
A recent section called Arroyo de Pedroche 2 (AP2) is lo-cated near the Club Asland Córdoba and runs parallel to theArroyo de Pedroche 1 section (Liñán et al., 2005).
Four regional stages are currently used in studies of theLower Cambrian of Spain. The basal, Corduban stage accountsfor successions bearing Cambrian trace fossils, underlying thefirst Ovetian fossils assemblages (Liñán, 1984b). The Ovetian,Marianian and Bilbilian stages are defined in base of archaeo-cyaths and trilobites (Sdzuy, 1971; Liñán et al., 1993). For a

Fig. 2. Cambrian stratigraphy of the Sierra de Córdoba (modified from Perejón et al., 1996).Fig. 2. Stratigraphie du Cambrien dans la Sierra de Córdoba (modifié d’après Perejón et al., 1996).
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468446
recent synthesis on the biochronology of the Ovetian of theSierra de Córdoba see Liñán et al. (2005). These authors haveestablished three trilobite interval biozones for the lower Ove-tian of Sierra Morena: Bigotina bivallata Biozone, Lemdadellalinarease Biozone and Lemdadella perejoni Biozone. They are
limited by the first appearance datum (FAD) of the index spe-cies of each biozone, following an evolutionary lineage (i.e.interval filozones). The upper boundary of the Lemdadellaperejoni Biozone is placed at the first occurrence of Eoredli-chia cf. ovetensis (Sdzuy in Liñán and Sdzuy, 1978) which is a

Fig. 3. Stratigraphy of the Puente de Hierro section (modified from Liñán Guijarro, 1978).Fig. 3. Stratigraphie de la section Puente de Hierro (modifié d’après Liñán Guijarro, 1978).
J.A.Gám
ezVintaned
etal.
/Geobios
39(2006)
443–468
447

Fig. 4. Stratigraphy of the Arroyo de Pedroche 1 section (modified from Liñán and Dabrio, 1974).Fig. 4. Stratigraphie de la section Arroyo de Pedroche 1 (modifié d’après Liñán et Dabrio, 1974).
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468448
young descendant of this lineage (Liñán et al., 2005). Accord-ing to these authors, the B. bivallata and L. linaresae Biozonescorrelate to Zone 0 of Morocco, and the L. perejoni Biozone to
Zone I (F. tazemmourtensis Zone). The three Spanish biozonesprobably correspond to the Lower Atdabanian Stage of Siberiaand correlative beds on the East European Platform and in Ant-

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 449
arctica. Finally, Pillola (1991) correlated Lower Cambrian bedswith the trilobite genus Eoredlichia from SW China, SW Sar-dinia, S France, Spain and Morocco.
4. Systematic palaeontology
4.1. Coelenterata
The material studied herein is housed in the Museo Paleon-tológico de la Universidad de Zaragoza-Gobierno de Aragón atZaragoza, Spain, under references MPZ 2002/844 and MPZ2002/923.
Class INCERTAE SEDISGenus Tiernavia Fedonkin in Fedonkin Liñán and Perejón,
1985.Type species: Tiernavia tiernae Fedonkin in Fedonkin Li-
ñán and Perejón, 1985.
Tiernavia tiernae Fedonkin in Fedonkin et al., 1985.Fig. 6(9).Material: One specimen preserved as a convex mould in
claystone (MPZ 2002/923).Description: Fossil subcircular in outline, cup-shaped, with
a truncated apical (lower) end surrounded by two narrow an-nular depressions. Diameter is ca. 15 mm and height is6.3 mm.
Interpretation and discussion: Tiernavia is a soft-bodiedfossil described from Lower Cambrian lutites of the upper JuliaMember of the Torreárboles Formation in Sierra de Córdoba.Fedonkin (in Fedonkin et al., 1985) interpreted Tiernavia asmoulds of imprints left by sessile, soft-bottom dwelling,polyps. Our specimen is incomplete, but the determination isstill possible.
Stratigraphical distribution and biochronology: Arroyode Pedroche 1 section, level 9 (upper part, above the Pedrocheevent). Lower Ovetian.
aff. TiernaviaFig. 6(5, 6).Material: One specimen preserved as a convex mould with-
in a siltstone bed (MPZ 2002/844).Description: Bag-shaped fossil with a lower, somewhat
flattened hemispherical expansion (25 mm in diameter) orna-mented with small tubercles (1 mm in width), which is sepa-rated from an upper part by a sharp narrowing (15 mm in dia-meter). The upper part of the fossil (27 mm in width) showsthree lobes on one side (the other one is not visible) separatedby longitudinal furrows (arrow in Fig. 6(5)). The total length ofthe fossil (measured almost horizontally) is 38 mm; the presentvertical dimension is 11.2 mm.
Interpretation and discussion: The interpretation of thisbody fossil as a trace fossil is not plausible due to its sharpnarrowing, the ornamentation and the type of preservation. Inturn, these features point to the mould of the impression left bya sessile, endosedimentary soft-bodied animal (probably of cni-
darian affinities) preserved in situ. The original orientationcould be more or less vertical, appearing today near the hori-zontal due to compaction flattening. Although this fossil showsa quite distinctive morphology, the scarcity of material (onespecimen) prevents against defining a new taxon, and thus itis placed in open nomenclature, tentatively close to Tiernavia,which is the closest form known.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
4.2. Trace fossils
The English terminology used in this work follows the mul-tilingual glossary of Gámez Vintaned and Liñán (1996). Thematerial studied herein is housed in the Museo Paleontológicode la Universidad de Zaragoza-Gobierno de Aragón at Zarago-za, Spain, under references from MPZ 2002/712 to MPZ 2002/843, MPZ 2002/845 to MPZ 2002/922, MPZ 2002/924, MPZ2002/925, MPZ 2002/928, MPZ 2002/931, MPZ 2002/932,MPZ 2002/951 and MPZ 2002/952.
Ichnogenus Bergaueria Prantl, 1946.Type ichnospecies: Bergaueria perata Prantl, 1946.Diagnosis: Bag forming or cylindrical protuberances on the
lower surface of the bedding plane. The surface of these pro-tuberances is smooth (from Prantl, 1946).
Bergaueria perata Prantl, 1946.Fig. 6(1–4).Material: Two specimens preserved as full relief in silt-
stone (MPZ 2002/742 and 845).Description: Bergaueria wider than deep, vertical to obli-
que, with thinly lined, parallel burrow walls and a lower termi-nation which is either rounded or show a central knob (Fig. 6(1)). Transverse section is near circular; both fine and strongtransverse constrictions are present on the wall. The largestspecimen is 22.7 mm in diameter and 17.3 mm deep. Theuppermost part of the cylinder of specimen MPZ 2002/742(18 mm in diameter) is surrounded by an outer circular bandwith concentric structure (28.5 mm in diameter) that extendsdownwards forming a flat funnel 6.3 mm in height (Fig. 6(2)).
Interpretation and discussion: Despite the presence oftransverse constrictions (a feature related to the burrowingtechnique) and the absence of radial depressions (a non-diag-nostic character, yet common), we find advisable to classifythis form as B. perata. This also applies to the central cylinderof specimen MPZ 2002/742, but the upper, flat funnel is a fea-ture not described until now within this ichnotaxon. The con-centric structure of the funnel (somewhat similar to Conosti-chus) indicates that the burrowing process encompassed twophases, finishing with the penetration of an axial stalk downinto the soft sediment. Other ichnospecies of Bergaueria (andConostichus) have very distinctive features which are lackingin our material.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468450

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 451
Ichnogenus Bilinichnus Fedonkin and Palij in Palij, Postiand Fedonkin, 1979.
Type ichnospecies: Bilinichnus simplex Fedonkin and Palijin Palij et al., 1979.
aff. Bilinichnus.Fig. 7(9).Material: Two specimens preserved as convex hyporelief
in siltstone (MPZ 2002/863 and 896).Description: A pair of two blunt, parallel ridges of uneven
longitudinal profile (i.e. the height varies) separated by an irre-gular to slightly concave area. Each ridge seems to be com-posed of segments aligned in a row or slightly imbricated,which renders a pelletoidal appearance. The width is equalfor each ridge of the pair (0.5 mm) and is uniform along thepath, which is gently sinuous or curved. Spacing between thetwo ridges of specimen MPZ 2002/863 varies along its pathfrom 1.7 to 2.6 mm and then to 1.7 mm. The area in betweenis slightly concave and of anastomosed texture (Fig. 7(9)). Spe-cimen MPZ 2002/896 shows a constant spacing between thetwo ridges (1.5 mm) and the area in between is irregular. Thisspecimen is actually composed of two paths aligned into anarch but overlapping at the junction. The two specimens are28 and 34 mm long, respectively.
Interpretation and discussion: These ichnofossils stronglyresemble Bilinichnus by their general morphology, dimensionsand mode of preservation. However, Bilinichnus displayssmooth ridges of even longitudinal profile and is interpretedas an exogenous trail (Fedonkin, 1981). We compare our twopoorly preserved specimens with Bilinichnus because it is pre-sent in early Lower Cambrian rocks from Sierra de Córdoba,but they could also be compared with Diplopodichnus becauseof their pelletoidal appearance (Buatois, pers. comm.). More-over, Buatois et al. (1998) noted that imprints are not necessa-rily preserved in Diplopodichnus, and Bilinichnus could there-fore be considered as a potential junior synonym, pending re-examination of the holotype. On the other hand, the particulartexture of the area between the ridges in specimen MPZ 2002/863 recalls somehow the impressions left by some soft-bodiedworms, hence its interpretation as an ichnofossil is not conclu-sive.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Cochlichnus Hitchcock, 1858.Type ichnospecies: Cochlichnus anguineus Hitchcock,
1858.Diagnosis: Regular, sinusoidal, horizontal trails and bur-
rows resembling a compressed and stretched corkscrew. Theoverall width of an individual trace may change progressively(from Gluszek, 1995).
Fig. 5. Stratigraphy of the Arroyo de Pedroche 2 section (modified from Liñán et aFig. 5. Stratigraphie de la section Arroyo de Pedroche 2 (modifié d’après Liñán et
Cochlichnus anguineus Hitchcock, 1858.Fig. 8(9d) and Fig. 9(9b).Material: Ten studied specimens preserved as full relief in
siltstone and very fine-grained sandstone (MPZ 2002/761, 765,775, 825, 833, 870, 872, 873, 880 and 903).
Emended diagnosis: Simple, smooth, horizontally undulat-ing trails and burrows. Slight vertical undulations can be pre-sent as well. The first-order path is either straight or slightlycurved. Horizontal wave length may change along the pathand is markedly larger than the wave amplitude.
Gluszek (1995) emended the diagnosis of the ichnogenus,and applied the same diagnosis to the type ichnospecies C. an-guineus. Given that several distinct ichnospecies compose thisichnogenus, it is necessary a new diagnosis.
Description: Simple, horizontal or vertically undulatingburrows, most of them showing a straight first-order path anda few arched. The second-order path is a lax sinuous line, withindex R being nearly 4. Unrectified length ranges from 10 to65 mm, and width from 0.5 to 1.8 mm. Wave length rangefrom 0.6 to 12.0 mm. Cochlichnus anguineus often cuts acrossprevious traces, such as Dactyloidites cabanasi (Fig. 8(9d))and Treptichnus ichnosp. A (Fig. 9(9b)).
Interpretation and discussion: Cochlichnus anguineus dif-fers from Cochlichnus ichnosp. A in showing a straight first-order path, and from this point of view it is similar to Coch-lichnus ichnosp. B described by Gámez-Vintaned and Liñán(in Van Amerom et al., 1973) from Spanish Lower Permianfine-grained volcanic ashes deposited in small ponds of freshwater, which nevertheless shows a very irregular second-ordersinuous path.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section; Arroyo de Pedroche 1section, level 9 (upper part, above the Pedroche event). LowerOvetian.
Cochlichnus ichnosp. AFig. 10(1a).Material: Five specimens, four of them preserved as full
relief in siltstone and very fine-grained sandstone (MPZ2002/727, 731, 740 and 871), and one as concave semireliefin siltstone (MPZ 2002/884).
Description: Simple, smooth, horizontal burrows of sinu-ous general (i.e. first order) path. The second-order path is alax sinuous line with index R (λ/2A; Gámez-Vintaned and Li-ñán in Van Amerom et al., 1993) being nearly 4. The ampli-tude of the first-order sinuous path may vary within a singlespecimen. Unrectified length ranges from 31 to 100 mm, andwidth from 0.5 to 1.7 mm. The first-order wavelength rangesfrom 31 to 88 mm; the first-order 2A parameter (twice thewave amplitude) ranges from 4.7 to 10.0 mm, which result inindex R values from 6.5 to 22.0. The second-order wavelengthranges from 1.5 to 6.0 mm, with index R being nearly 4. Coch-
l., 2005).al., 2005).

Fig. 6. Soft-bodied fossils and ichnofossils from the Pedroche Formation. Material coated with ammonium chloride (except 1–6, and 11). Scale bars = 1 cm. 1–4.Bergaueria perata preserved as full relief in siltstone. 1, 2. Specimen MPZ 2002/742; 1: bottom view (note the central knob); 2: upper view (the concentric structureis visible). 3, 4. Specimen MPZ 2002/845; 3: bottom view; 4: lateral view. 5, 6. aff. Tiernavia (MPZ 2002/844). The contour of the fossil is paralleled by white dots.5: bottom view, the arrow points to longitudinal furrow separating two lobes in the upper part of the fossil; 6: lateral view. 7. Latex mould of aff. Cosmorhaphe?preserved as concave semirelief in siltstone (MPZ 2002/877). White arrow points to third-order undulation. Black arrow indicates the trilobite Lemdadella linaresae
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468452

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 453
lichnus ichnosp. A cuts across previous specimens of Dactyloi-dites cabanasi.
Interpretation and discussion: As stated by Gámez-Vin-taned and Liñán (in Van Amerom et al., 1993), the genusCochlichnus needs a diagnostic revision in the light of geome-trical patterns and morphometry. Stanley and Pickerill (1998)included all previous ichnospecies in C. anguineus, which isthe type ichnospecies by monotypy. Fillion and Pickerill(1990) characterised C. anguineus by the absence of ornamen-tation. We agree with these authors in that Cochlichnus in-cludes trails and burrows. Cochlichnus ichnosp. A describedhere is a smooth form which differs from both C. anguineusHitchcock, 1858 and C. kochi (Ludwig, 1869) in having a sin-uous first-order path in addition to the “regular”, smaller undu-lations. Moreover, our material differs from C. kochi in havingan index R markedly larger (nearly 4) for the second-orderwaves. Nevertheless, the number of studied specimens is notappropriate for defining a new ichnospecies.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Cosmorhaphe Fuchs, 1895.Type ichnospecies: Cosmorhaphe sinuosa (Azpeitia Moros,
1933).
aff. Cosmorhaphe?Fig. 6(7).Material: One specimen preserved as concave semirelief in
siltstone (MPZ 2002/877).Description: Simple, horizontal trace fossil showing three
orders of undulations. The first-order path is a loosely guidedmeander with a wavelength of 35 mm and angular turns. Thesecond-order path is a sinuous to angulate line with wavelengthranging from 12 to 34 mm. The third-order path is sinuouswith a wavelength of 2 mm and very low amplitude. The rec-tified length of the trace is 120 mm; width equals 1.2 mm.
Interpretation and discussion: Cosmorhaphe has only twoorders of undulations, whilst our specimen shows three. Thesecond-order undulation (which is added to the Cosmorhaphepattern in our case) emphasises the guide of the otherwiseloose first-order meanders, indicating thigmotaxis, which is akind of behaviour also present in Cosmorhaphe but not inCochlichnus. Following ichnotaxonomic pattern analyses from
Liñán and Sdzuy, 1978. 8. Undetermined burrow preserved as sandy full relief withiin claystone (MPZ 2002/923). 10. Faecal pellets preserved in full relief, in a circular a(a: MPZ 2002/850). Cylindrichnus concentricus (b: MPZ 2002/924). Both preservarrangements (MPZ 2002/883).Fig. 6. Fossiles à corps mous et ichnofossiles de la Formation Pedroche. Matériel cou1–4. Bergaueria perata préservé comme relief complet dans la siltite. 1, 2. Spécisupérieure (on peut observer la structure concentrique). 3, 4. Spécimen MPZ 2002/8contour du fossile est marqué par des points blancs. 5 : vue inférieure, la flèche indifossile ; 6 : vue latérale. 7. Moule à latex de aff. Cosmorhaphe? préservé comme seondulations de troisième ordre. La flèche noire indique le trilobite Lemdadella linaresablonneux dans l’argilite (MPZ 2002/898). 9. Tiernavia tiernae préservé commecomme relief complet avec un arrangement subcirculaire (MPZ 2002/885). 11. aff. PMPZ 2002/924). Tous les deux préservés comme relief complet. 12. Pelotes fécales2002/883).
Seilacher (1977), the undulations pattern described here couldbe used for erecting a new ichnotaxon, which is beyond thisarticle because of the scarce material. On the other hand, thisform may also resemble physical structures (Buatois and Uch-man, pers. comm.), and hence a question mark is included afterthe ichnogenus.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Cylindrichnus Toots in Howard, 1966.Type ichnospecies: Cylindrichnus concentricus Toots in
Howard, 1966.
Cylindrichnus concentricus Toots in Howard, 1966.Fig. 6(11b).Material: Two specimens preserved as full relief in very
fine-grained sandstone (MPZ 2002/924 and 925).Description: Transverse sections of short, vertical burrows,
from 2.4 to 3.5 mm in diameter. Undistinct zoned fill com-posed of a narrow core of sandstone and a wide mantle formedby multiple concentric layers of fine-grained lutite and alternat-ing thicknesses.
Interpretation and discussion: Liñán et al. (1995) inter-preted Cylindrichnus concentricus preserved in Middle Cam-brian lutites from the Los Villares Formation as endichnial,feeding and dwelling burrows produced by a semi sessile en-dobenthic animal within a soft substratum rich in nutrients.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Dactyloidites Hall, 1886.Type ichnospecies: Dactyloidites bulbosus Hall, 1886.
Dactyloidites cabanasi (Meléndez in Cabanás, 1966).Figs. 8,7(1–5, 8d) and Fig. 10(1b)*1966. Anthoichnites cabanasi, nov. gen. (parataxon), n. sp.
- Meléndez in Cabanás, pp.82–83, Figs. 1–3.v1986. Haentzschelinia sp. ? - Díez Balda, pp. 53, Pl. VI,
Fig. c.?v1999a. Dactyloidites ichnosp. - Palacios, Gámez, Fernán-
dez-Remolar and Liñán, pp. 265.?v1999b. Dactyloidites ichnosp. - Palacios, Gámez Vin-
taned, Fernández-Remolar and Liñán, pp. 1017.
n claystone (MPZ 2002/898). 9. Tiernavia tiernae preserved as a convex mouldrrangement (MPZ 2002/885). 11. aff. Phycodes in a very fine-grained sandstoneed as full relief. 12. Full relief faecal pellets of unequal size under different
vert avec du chlorure d’ammonium (sauf de 1–6 et 11). Barre d’échelle = 1 cm.men MPZ 2002/742; 1 : vue inférieure (remarquer le nœud central); 2 : vue45; 3 : vue inférieure; 4 : vue latérale. 5, 6. aff. Tiernavia (MPZ 2002/844). Leque le sillon longitudinal qui sépare les deux lobes dans la partie supérieure dumirelief concave dans la siltite (MPZ 2002/877). La flèche blanche indique lessae Liñán et Sdzuy, 1978. 8. Terrier indéterminé préservé comme relief completmoule convexe dans l’argilite (MPZ 2002/923). 10. Pelotes fécales préservéshycodes dans grès très fin (a : MPZ 2002/850). Cylindrichnus concentricus (b :comme relief complet de dimensions inégales et arrangements différents (MPZ

Fig. 7. Ichnofossils from the Pedroche Formation. All photographs are bottom views, except items 2, 3 and 4. Material coated with ammonium chloride (exceptthe sections). Scale bars = 1 cm. 1. Dactyloidites cabanasi with four radial burrows (MPZ 2002/734). Top view. 2. Vertical section across bioturbated siltstone(under water). Two retrusive spreite burrows belonging to different specimens of D. cabanasi are shown in transverse section. The one above (b) preserves the mainburrow on its final stage (arrow; MPZ 2002/798), while the one below does not (MPZ 2002/797). 3. Same sample of bioturbated siltstone, showing the shaft (arrow)
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468454

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 455
Material: Seventy-five specimens preserved as full relief insiltstone and fine to very fine-grained sandstone, visible eitherfrom the bottom or the top of the strata as well as in cross-section (MPZ 2002/712-717, 719, 720, 725, 726, 729, 734-737, 741, 743-746, 748-758, 760, 763, 764, 767-770, 772,774, 776-779, 781, 783-788, 795-798, 800, 801, 804, 805,813, 819, 820, 824, 826, 827, 830-832, 836-841 and 928).
Emended diagnosis: Petaloid to rosetted trace fossils com-posed of four to six radial, horizontal, retrusive spreiten bur-rows connected to a narrow central shaft.
Description: Radial burrows are predominantly petaloid, innumber from four to six but most frequently five. Basic bio-metric parameters are summarised on Table 1 and Fig. 11.Spreiten burrows are plane-convex to biconvex. The retrusivespreite is conspicuous in cross-sections and also in bottom andtop views; laminae are arranged concave up both across andalong the burrows (Fig. 8(2, 3, 10)). The main burrow (centralshaft) may be present (Fig. 7(3) and Fig. 8(1a, 8a)) or absent(Fig. 7(1, 5) and Fig. 8(1b)). Unweathered specimens mayshow faint concave striae, which trace the spreite inside; con-cavity points mostly toward the central shaft (Fig. 8(4)).Weathering reveals the three-dimensional architecture of thespreite (Fig. 7(1, 5) and Fig. 8(3, 10)). A narrow, longitudinalfurrow may run on the lower surface of some radial burrows(Fig. 8(1a) and Fig. 10(1b)). Burrow infill may differ from thesurrounding matrix. On some occasions tiny, scarce pellets arelocated on the proximal region of the petal (Fig. 8(4)).
Good radial symmetry is absent only in a few specimens,like the six-armed MPZ 2002/836 (Fig. 8(2)) and MPZ 2002/781 (Fig. 8(7)).
Interpretation and discussion: Dactyloidites is very abun-dant in the heterolithic facies of the Puente de Hierro section,composed of fine to very fine-grained sandstone and siltstone.It is absent from the prevailing fine-grained mudstones of theArroyo de Pedroche 1 section.
The radial geometry and the spreite structure lead to inter-pret our specimens of Dactyloidites cabanasi as feeding bur-row systems (fodinichnia) produced by soft-bodied mud eaterswithin nutrient-rich, soft substrata. Simultaneous occupation of
of D. cabanasi in longitudinal section (MPZ 2002/841). 4. Different spreiten of sev2002/797). 5. Retrusive spreite in a specimen of D. cabanasi with four radial bur6. Dimorphichnus ichnosp. A (a: MPZ 2002/789). Dimorphichnus cf. obliquus (b:preserved as convex hyporelief in fine-grained sandstone. 7. Diplichnites ichnosp. indpreserved as convex hyporelief on a very fine-grained sandstone (from a–c: MPDactyloidites cabanasi (d: MPZ 2002/719). 9. aff. Bilinichnus preserved as convexFig. 7. Ichnofossiles de la Formation Pedroche. Toutes les vues sont prises dès la bas(sauf les coupes). Barre d’échelle = 1 cm. 1. Dactyloidites cabanasi avec quatre terrisiltite bioturbée (submergé dans l’eau). On observe la coupe transversale de deux terrCelui supérieur (b) préserve le terrier principal dans son stade final (flèche ; MPZantérieur de la siltite bioturbée montre le tuyau (flèche) de D. cabanasi en coupe lonD. cabanasi (a : MPZ 2002/795, b : MPZ 2002/796, c : MPZ 2002/797). 5. Traversetraverse sont concaves vers le haut le long des terriers (MPZ 2002/838). 6. Dimorph2002/790). Diplichnites ichnosp. indét. (c : MPZ 2002/791). Tous sont préservés com2002/889). Moule artificiel d’un épirelief concave. 8. Rusophycus didymus préservé2002/722 et MPZ 2002/723). Dans la partie haute de la figure : Dactyloidites cabandans la siltite (MPZ 2002/863).
a given area of substratum by a number of producers probablyoccurred. Specimens of Dactyloidites cabanasi may touch butdo not overprint each other, thus suggesting phobotaxis. Theexclusivity of retrusive forms is highly compatible with softsubstrata undergoing moderate but rather continuous sedimen-tary agradation in a shallow sublittoral setting of moderate en-ergy. Differences in spreite convexity and ornamentation mayreflect slight differences in the water contents of the mud dur-ing the process of production and early stages of burial. Thehigher or lower relief of the hypichnial bulge depends partiallyon the burrowing technique and, to a bigger extent, on the tex-tural contrast between the exploited sediment and the underly-ing substratum. The maximum bulge in Dactyloidites cabanasiappears when this is produced within sediment of coarse silt tofine-grained sandstone and the bottom of the radial burrowsreaches a level of underlying clay.
Our material shows a constructional pattern very close tothe interpretation given by Fürsich and Bromley (1985) forthe so-called Dactyloidites canyonensis (Bassler, 1941). As-suming their interpretation of Dactyloidites as a valid ichno-genus for rosetted spreite structures, we propose to transferthe ichnospecies Anthoichnites cabanasi Meléndez in Cabanás,1966 to the ichnogenus Dactyloidites because of priority. Butcontrary to the opinion of Fürsich and Bromley (1985), we findnot advisable to include Brooksella canyonensis Bassler, 1941within Dactyloidites, given that the Neoproterozoic material ofBassler (1941) is still to be well understood. This is in agree-ment with the statement by Agirrezabala and De Gibert (2004:204) that Brooksella canyonensis and Dactyloidites asteroides“are in need of a new systematic revision, including an assess-ment of their possible inclusion within the ichnogenus Dacty-loidites”. Dactyloidites cabanasi differs from Gyrophyllites inlacking the helicoidal, stacked pattern of leaf-shaped feedingburrows.
Hence we find convenient to maintain the ichnospecific des-ignation given by Meléndez in Cabanás (1966).
Stratigraphical distribution and biochronology: Lowerand middle part of the Puente de Hierro section (type-locality
eral specimens of D. cabanasi (a: MPZ 2002/795, b: MPZ 2002/796, c: MPZrows. Laminae are arranged concave up along the burrows (MPZ 2002/838).MPZ 2002/790). Diplichnites ichnosp. indet. (c: MPZ 2002/791). All of themet. (MPZ 2002/889). Artificial cast of concave epirelief. 8. Rusophycus didymusZ 2002/721, MPZ 2002/722 and MPZ 2002/723). At the top of the figure:hyporelief in siltstone (MPZ 2002/863).e, avec l’exception de 2, 3 et 4. Matériel couvert avec du chlorure d’ammoniumers radiaux (MPZ 2002/734). Vue supérieure. 2. Coupe verticale en travers de laiers avec traverse rétrusive qui appartient à différents spécimens de D. cabanasi.2002/798), au contraire de celui inférieur (MPZ 2002/797). 3. L’échantillongitudinale (MPZ 2002/841). 4. Différentes traverses de plusieurs spécimens derétrusive d’un spécimen de D. cabanasi à quatre terriers radiaux. Les lames deichnus ichnosp. A (a : MPZ 2002/789). Dimorphichnus cf. obliquus (b : MPZme hyporelief convexe dans des grès fins. 7. Diplichnites ichnosp. indét. (MPZcomme hyporelief convexe dans des grès très fins (a–c : MPZ 2002/721, MPZasi (d : MPZ 2002/719). 9. aff. Bilinichnus préservé comme hyporelief convexe

Fig. 8. Ichnofossils from the Pedroche Formation. Material coated with ammonium chloride. Scale bars = 1 cm. 1. Dactyloidites cabanasi (a, b: MPZ 2002/800, 801).Meniscate burrow (c: MPZ 2002/802). Faecal pellets (d: MPZ 2002/803). Psammichnites ichnosp. A (e: MPZ 2002/931). 2. Dactyloidites cabanasi (MPZ 2002/836).3. Dactyloidites cabanasi (MPZ 2002/752). 4. Dactyloidites cabanasi (MPZ 2002/748). The arrow show faint, concave striae reflecting the spreiten infill.5. Dactyloidites cabanasi (a–e: MPZ 2002/712-716). Psammichnites ichnosp. A (f: MPZ 2002/718). 6. Dactyloidites cabanasi (a: MPZ 2002/729). Psammichnites
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468456

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 457
of the ichnospecies); Arroyo de Pedroche 2 section, level 12.Lower Ovetian.
Ichnogenus Dimorphichnus Seilacher, 1955.Type ichnospecies: Dimorphichnus obliquus Seilacher,
1955.
Dimorphichnus cf. obliquus Seilacher, 1955.Fig. 7(6b).Material: One specimen preserved as convex hyporelief in
fine-grained sandstone (MPZ 2002/790).Description: Trace fossil composed of two sets, each with
five or four slightly sigmoidal, raker impressions and a similarnumber of obscure and blunt, pusher impressions arranged par-allel to those. The trace is overprinted (and to some extent ob-scured) by specimen MPZ 2002/791 (Diplichnites ichnosp. in-det.). Total length of the trace is 25 mm, total width is 19 mm.Length of each raker set is 10.5 mm; width of raker sets are11.4 and 10.0 mm.
Interpretation and discussion: According to Seilacher(1955), Dimorphichnus is a grazing trace formed by trilobiteswhen scratching the sea bottom with the appendages in searchfor food. Our specimen is interrupted on its middle part by thelater production of Diplichnites.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Dimorphichnus ichnosp. A.Fig. 7(6a).Material: One specimen preserved as convex hyporelief in
fine-grained sandstone (MPZ 2002/789).Description: Dimorphichnus composed of a set of two
arched, convergent, 30-mm long (unrectified) raker impres-sions and two blunt pusher impressions. Separation betweenraker impressions diminishes gradually from 4.4 to 2.0 mm.
Discussion: This specimen differs from D. obliquus in thearched path of the raker impressions, instead of sigmoidal.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Diplichnites Dawson, 1873.Type ichnospecies: Diplichnites aenigma Dawson, 1873.
Diplichnites ichnosp. indet.Fig. 7(6c, 7).
ichnosp. A (b: MPZ 2002/730). Cochlichnus ichnosp. A (c: MPZ 2002/731). Torr2002/781). 8. Dactyloidites cabanasi (a, b: MPZ 2002/819, 820). Psammichnites ic823). 9. Dactyloidites cabanasi (a–c: MPZ 2002/830-832). Cochlichnus anguineusFig. 8. Ichnofossiles du membre inférieur de la Formation Pedroche. Matériel coucabanasi (a, b : MPZ 2002/800, 801). Terrier ménisqué (c : MPZ 2002/802). Pelote2. Dactyloidites cabanasi (MPZ 2002/836). 3. Dactyloidites cabanasi (MPZ 2002/75MPZ 2002/712-716). Psammichnites ichnosp. A (f : MPZ 2002/718). 6. DactyloiditeCochlichnus ichnosp. A (c : MPZ 2002/731). Torrowangea ichnosp. indét. (d : Mcabanasi (a, b : MPZ 2002/819, 820). Psammichnites ichnosp. A (c, d : MPZ 20cabanasi (a–c : MPZ 2002/830-832). Cochlichnus anguineus (d : MPZ 2002/833).
Material: Three specimens, two of them preserved as con-vex hyporelief (MPZ 2002/791 and 922) and one as concaveepirelief (MPZ 2002/889), all in fine-grained sandstone.
Description: Only one out of the three specimens is pre-served complete, showing two convergent rows of tracks. Theother two preserve only one row. Length of the complete speci-men is 22.0 mm and width varies progressively from 12.0 to6.7 mm; individual imprints are less than 1 mm wide. Lengthof each incomplete specimen is 12.5 mm, with individual im-prints from 1.5 to 3.5 mm in width.
Interpretation and discussion: Trackways produced by tri-lobites walking on the sea floor. In the two incomplete speci-mens, appendage imprints left by one side of the body may notbe preserved due to undertrace deficit preservation (cleavagerelief).
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Monocraterion Torell, 1870.Type ichnospecies: Monocraterion tentaculatum Hall,
1847.
Monocraterion ichnosp. indet.Fig. 9(4, 6).Material: Three studied specimens, one preserved as full
relief within very fine-grained sandstone (MPZ 2002/810),and two as convex hyporeliefs in fine-grained quartzite (MPZ2002/846 and 848).
Description: Conical structures from 3.0 to 11.6 mm in dia-meter and from 2.2 to 5.2 mm in height. The two largest speci-mens, preserved in quartzite, show a truncated apical (lower)termination.
Interpretation and discussion: Monocraterion is inter-preted as domichnia left by a variety of producers. Our speci-mens appear in beds deposited at moments of increasing waterenergy. It is noteworthy the co-existence of Monocraterion ich-nosp. indet. with Psammichnites gigas together in the samesample (7 cm thick) of fine-grained quartzite (Fig. 9(6)); theformer predates Psammichnites within a clean, sandy substrate.These specimens reflect two different feeding and living beha-viours, and probably record two stages of the ecological suc-cession posterior to the high energy, event bed.
Although Schlirf and Uchman (2004) suggest restriction onthe use of Monocraterion to the type-material, we follow thegeneralised criterion of including vertical, cylindrical burrows,
owangea ichnosp. indet. (d: MPZ 2002/732). 7. Dactyloidites cabanasi (MPZhnosp. A (c, d: MPZ 2002/821, 822). Skolithos ichnosp. indet. (e: MPZ 2002/(d: MPZ 2002/833). 10. Dactyloidites cabanasi (MPZ 2002/839). Top view.vert avec du chlorure d’ammonium. Barre d’échelle = 1 cm. 1. Dactyloiditess fécales (d : MPZ 2002/803). Psammichnites ichnosp. A (e : MPZ 2002/931).2). 4. Dactyloidites cabanasi (MPZ 2002/748). 5. Dactyloidites cabanasi (a–e :s cabanasi (a : MPZ 2002/729). Psammichnites ichnosp. A (b : MPZ 2002/730).PZ 2002/732). 7. Dactyloidites cabanasi (MPZ 2002/781). 8. Dactyloidites02/821, 822). Skolithos ichnosp. indét. (e : MPZ 2002/823). 9. Dactyloidites10. Dactyloidites cabanasi (MPZ 2002/839). Vue supérieure.

Fig. 9. Ichnofossils from the Pedroche Formation. Material coated with ammonium chloride (except the section in 2). Scale bars = 1 cm. 1. Top view of twoPsammichnites gigas. One specimen (b: MPZ 2002/817), coming from below, intersects another one (a: MPZ 2002/816). 2. Same as previous, in cross-section (underwater). White, dotted contour: incomplete, elliptical burrow of MPZ 2002/817. Black arrows: path of the ciliated “snorkel”, inclined to the right, which produced asinuous track (not visible on Fig. 9(1)) on top of MPZ 2002/817. White arrow: the same for MPZ 2002/816, but inclined to the left side and only partially preserved
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468458

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 459
which are funnel-shaped at the top, in this ichnogenus, until acomprehensive revision is accomplished.
Stratigraphical distribution and biochronology: Arroyode Pedroche 1 section, level 2; middle part of the Puente deHierro section. Lower Ovetian.
Ichnogenus Palaeophycus Hall, 1847.Type ichnospecies: Palaeophycus tubularis Hall, 1847.
Palaeophycus cf. alternatus Pemberton and Frey, 1982.Fig. 10(5b).Material: One specimen preserved as full relief in very
fine-grained sandstone (MPZ 2002/862).Description: Simple, cylindrical, weakly lined and straight,
horizontal burrow with faint, parallel longitudinal striae andover-imposed faint constrictions. Infilling stuff is the same ashost rock. Diameter varies from 0.5 to 1.5 mm; length, 60 mm.
Interpretation and discussion: Striae and constrictions donot alternate (as in P. alternatus Pemberton and Frey, 1982) butrather coincides in place all along the trace fossil.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Palaeophycus striatus Hall, 1852.Fig. 9(7d) and Fig. 10(5a, 6b).Material: Five specimens preserved as full relief in fine to
very fine-grained sandstone and siltstone (MPZ 2002/812, 851,852, 855 and 861). Additional studied material remains in thefield.
Description: Simple, cylindrical, weakly lined and mostlystraight, horizontal burrows with faint, parallel longitudinalstriae. Same infilling stuff as host rock. Cross-sections are cir-cular and range from 0.5 to 8.0 mm; length from 21 to 60 mm.
Interpretation and discussion: Domichnia whose reinfor-cements (lining), although thin, permitted the open burrows tobe maintained. Building behaviour simple, with straight first-
due to intersection of MPZ 2002/817 from below. 3. Psammichnites gigas preservedof Monocraterion ichnosp. indet. (a: MPZ 2002/846) and cross-section of Psammichin 6. See palaeoecological interpretation in text. 5. Psammichnites ichnosp. A prepathway of the trace. (MPZ 2002/ 814). 6. Bottom view ofMonocraterion ichnosp. in4. The latter is partially eroded but the straight, central cord is still visible. Arrows indepirelief on fine-grained sandstone (a: MPZ 2002/806). Treptichnus ichnosp. B (b:ichnosp. indet. (c: MPZ 2002/811) and Palaeophycus striatus (d: MPZ 2002/812), bpreserved as concave epirelief on fine-grained sandstone (MPZ 2002/807); same samrelief in siltstone (a: MPZ 2002/902), and Cochlichnus anguineus (b: MPZ 2002/9Fig. 9. Ichnofossiles de la Formation Pedroche. Matériel couvert avec du chlorure ddeux Psammichnites gigas. Un des spécimens (b : MPZ 2002/817) en travers de l’audans l’eau). Contour blanc et pointillé : terrier incomplet et elliptique du MPZ 2002/une trace sinueuse (pas visible dans la Fig. 9(1)) dans le haut de MPZ 2002/817. Flèpréservé partiellement dû au croisement au-dessous par MPZ 2002/817. 3. Psammicremplissage ménisqué. 4. Vue latérale de Monocraterion ichnosp. indét. (a : MPZphotographiés à l’air. La vue inférieure est montrée à 6. Voyez l’interprétation phyporelief convexe dans des grès très fins. Les flèches montrent la trajectoire de la tMPZ 2002/846) et Psammichnites gigas (b : MPZ 2002/847) montrés à 4. Le dernieLes flèches indiquent les marges du terrier. 7. Treptichnus ichnosp. A préservé comichnosp. B (b : MPZ 2002/808) en connexion ontogénétique avec MPZ 2002/806.MPZ 2002/812), les deux en relief complet. 8. Treptichnus ichnosp. A (flèches) com(MPZ 2002/807) ; même échantillon qu’à la figure 7. 9. Vue dès la base de Treptichnet Cochlichnus anguineus (b : MPZ 2002/903) en relief complet dans la siltite et d
order path, but sinuosity along this is observed in one specimen(Fig. 10(5a)).
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section; Arroyo de Pedroche 1section, levels 2 and 4. Lower Ovetian.
Palaeophycus ichnosp. A.
Fig. 10(2, 6a).
Material: Five specimens preserved as full relief in veryfine-grained sandstone and (mostly) in siltstone (MPZ 2002/853, 854, 856, 857 and 865).
Description: Simple, bulbous, weakly lined, straight (hori-zontal) or gently curved (either in the horizontal or verticalplane) burrows with distinct, regularly spaced, transverse con-strictions. Infilling stuff is the same as host rock. Cross-sec-tions are circular and diameter may change abruptly over shortdistances: it ranges from 2.0 to 5.0 mm; length from 9.5 to20.0 mm.
Interpretation and discussion: The presence of transverseconstrictions as the only burrow sculpture and the bulbousmorphology prevent identifying this ichnofossil with any ofthe ichnospecies of Palaeophycus previously recognised, in-cluding Palaeophycus serratus McCann, 1993 which show an-nulae faintly marked and a constant diameter. The distinct mor-phological characteristics would make this form suitable for anew ichnospecies. Nevertheless, we consider the material col-lected as insufficient.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Palaeophycus ichnosp. indet.
Material: Two specimens preserved as concave semi-reliefin siltstone and as full relief in very fine-grained sandstone(MPZ 2002/866 and 867).
as concave epirelief (MPZ 2002/849) showing meniscate infill. 4. Lateral viewnites gigas (b: MPZ 2002/847) photographed in air. The bottom view is shownserved as convex hyporelief in very fine-grained sandstone. Arrows show thedet. (a: MPZ 2002/846) and Psammichnites gigas (b: MPZ 2002/847) shown inicate the margins of the burrow. 7. Treptichnus ichnosp. A preserved as concaveMPZ 2002/808) in ontogenetic connection with MPZ 2002/806. Torrowangeaoth in full relief. 8. Treptichnus ichnosp. A (arrows) composed of four burrowsple as shown in 7. 9. Bottom view of Treptichnus ichnosp. A preserved as full
03) in full relief in siltstone and very fine-grained sandstone.’ammonium (sauf le coupe à 2). Barre d’échelle = 1 cm. 1. Vue supérieure detre (a : MPZ 2002/816). 2. Le même spécimen en coupe transversale (submergé817. Flèches noires : trajectoire du « snorkel » cilié, incliné à droite, qui produitche blanche : le même pour MPZ 2002/816 mais incliné à gauche et seulementhnites gigas préservé comme épirelief concave (MPZ 2002/849) qui montre un2002/846) et coupe transversale de Psammichnites gigas (b : MPZ 2002/847),aléoécologique dans le texte. 5. Psammichnites ichnosp. A préservé commerace. (MPZ 2002/814). 6. Vue inférieure des Monocraterion ichnosp. indét. (a :r est partiellement érodé, mais on peut encore observer le cordon central, droit.me épirelief concave dans des grès très fins (a : MPZ 2002/806). TreptichnusTorrowangea ichnosp. indét. (c : MPZ 2002/811) et Palaeophycus striatus (d :posé par quatre terriers préservés comme épirelief concave dans grès très finsus ichnosp. A préservé comme relief complet dans la siltite (a : MPZ 2002/902),es grès très fins.

Fig. 10. Ichnofossils from the Pedroche Formation. Material coated with ammonium chloride. Scale bars = 1 cm. 1. Bottom view of Cochlichnus ichnosp. A (a: MPZ2002/727), Dactyloidites cabanasi (b: MPZ 2002/726) and Planolites annularis (c: MPZ 2002/728). 2. Palaeophycus ichnosp. A preserved as full relief in very fine-grained sandstone (MPZ 2002/853). 3. Top view of Planolites montanus preserved as full relief (a: MPZ 2002/904, b: MPZ 2002/905). 4. Top view ofPsammichnites ichnosp. A. White arrows show the axial, sinuous track of the trace (a: MPZ 2002/911). Torrowangea rosei preserved as full relief (b: MPZ 2002/951;
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468460
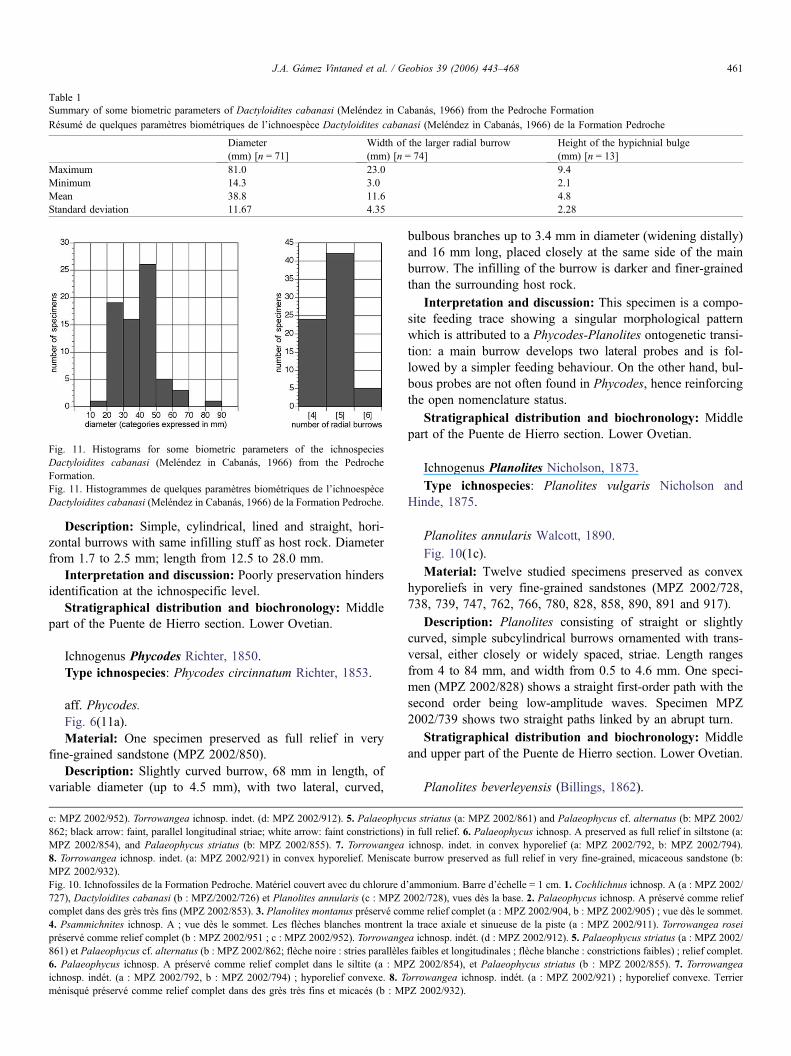
Fig. 11. Histograms for some biometric parameters of the ichnospeciesDactyloidites cabanasi (Meléndez in Cabanás, 1966) from the PedrocheFormation.Fig. 11. Histogrammes de quelques paramètres biométriques de l’ichnoespèceDactyloidites cabanasi (Meléndez in Cabanás, 1966) de la Formation Pedroche.
Table 1Summary of some biometric parameters of Dactyloidites cabanasi (Meléndez in Cabanás, 1966) from the Pedroche Formation
Résumé de quelques paramètres biométriques de l’ichnoespèce Dactyloidites cabanasi (Meléndez in Cabanás, 1966) de la Formation Pedroche
Diameter(mm) [n = 71]
Width of the larger radial burrow(mm) [n = 74]
Height of the hypichnial bulge(mm) [n = 13]
Maximum 81.0 23.0 9.4Minimum 14.3 3.0 2.1Mean 38.8 11.6 4.8Standard deviation 11.67 4.35 2.28
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 461
Description: Simple, cylindrical, lined and straight, hori-zontal burrows with same infilling stuff as host rock. Diameterfrom 1.7 to 2.5 mm; length from 12.5 to 28.0 mm.
Interpretation and discussion: Poorly preservation hindersidentification at the ichnospecific level.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Phycodes Richter, 1850.Type ichnospecies: Phycodes circinnatum Richter, 1853.
aff. Phycodes.Fig. 6(11a).Material: One specimen preserved as full relief in very
fine-grained sandstone (MPZ 2002/850).Description: Slightly curved burrow, 68 mm in length, of
variable diameter (up to 4.5 mm), with two lateral, curved,
c: MPZ 2002/952). Torrowangea ichnosp. indet. (d: MPZ 2002/912). 5. Palaeophyc862; black arrow: faint, parallel longitudinal striae; white arrow: faint constrictions)MPZ 2002/854), and Palaeophycus striatus (b: MPZ 2002/855). 7. Torrowangea8. Torrowangea ichnosp. indet. (a: MPZ 2002/921) in convex hyporelief. MeniscatMPZ 2002/932).Fig. 10. Ichnofossiles de la Formation Pedroche. Matériel couvert avec du chlorure d727), Dactyloidites cabanasi (b : MPZ/2002/726) et Planolites annularis (c : MPZ 2complet dans des grès très fins (MPZ 2002/853). 3. Planolites montanus préservé com4. Psammichnites ichnosp. A ; vue dès le sommet. Les flèches blanches montrentpréservé comme relief complet (b : MPZ 2002/951 ; c : MPZ 2002/952). Torrowange861) et Palaeophycus cf. alternatus (b : MPZ 2002/862; flèche noire : stries parallèles6. Palaeophycus ichnosp. A préservé comme relief complet dans le siltite (a : MPichnosp. indét. (a : MPZ 2002/792, b : MPZ 2002/794) ; hyporelief convexe. 8. Tménisqué préservé comme relief complet dans des grès très fins et micacés (b : MP
bulbous branches up to 3.4 mm in diameter (widening distally)and 16 mm long, placed closely at the same side of the mainburrow. The infilling of the burrow is darker and finer-grainedthan the surrounding host rock.
Interpretation and discussion: This specimen is a compo-site feeding trace showing a singular morphological patternwhich is attributed to a Phycodes-Planolites ontogenetic transi-tion: a main burrow develops two lateral probes and is fol-lowed by a simpler feeding behaviour. On the other hand, bul-bous probes are not often found in Phycodes, hence reinforcingthe open nomenclature status.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Planolites Nicholson, 1873.Type ichnospecies: Planolites vulgaris Nicholson and
Hinde, 1875.
Planolites annularis Walcott, 1890.Fig. 10(1c).Material: Twelve studied specimens preserved as convex
hyporeliefs in very fine-grained sandstones (MPZ 2002/728,738, 739, 747, 762, 766, 780, 828, 858, 890, 891 and 917).
Description: Planolites consisting of straight or slightlycurved, simple subcylindrical burrows ornamented with trans-versal, either closely or widely spaced, striae. Length rangesfrom 4 to 84 mm, and width from 0.5 to 4.6 mm. One speci-men (MPZ 2002/828) shows a straight first-order path with thesecond order being low-amplitude waves. Specimen MPZ2002/739 shows two straight paths linked by an abrupt turn.
Stratigraphical distribution and biochronology: Middleand upper part of the Puente de Hierro section. Lower Ovetian.
Planolites beverleyensis (Billings, 1862).
us striatus (a: MPZ 2002/861) and Palaeophycus cf. alternatus (b: MPZ 2002/in full relief. 6. Palaeophycus ichnosp. A preserved as full relief in siltstone (a:ichnosp. indet. in convex hyporelief (a: MPZ 2002/792, b: MPZ 2002/794).e burrow preserved as full relief in very fine-grained, micaceous sandstone (b:
’ammonium. Barre d’échelle = 1 cm. 1. Cochlichnus ichnosp. A (a : MPZ 2002/002/728), vues dès la base. 2. Palaeophycus ichnosp. A préservé comme reliefme relief complet (a : MPZ 2002/904, b : MPZ 2002/905) ; vue dès le sommet.
la trace axiale et sinueuse de la piste (a : MPZ 2002/911). Torrowangea roseia ichnosp. indét. (d : MPZ 2002/912). 5. Palaeophycus striatus (a : MPZ 2002/faibles et longitudinales ; flèche blanche : constrictions faibles) ; relief complet.Z 2002/854), et Palaeophycus striatus (b : MPZ 2002/855). 7. Torrowangeaorrowangea ichnosp. indét. (a : MPZ 2002/921) ; hyporelief convexe. TerrierZ 2002/932).

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468462
Material: One specimen preserved as full relief in fine-grained quartzite (MPZ 2002/818).
Description: Almost straight, smooth, simple subcylindricalburrow, 40 mm long and 2.5 mm wide. Burrow fill is slightlydifferent to the host rock.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Planolites montanus Richter, 1937.
Fig. 10(3a, b).
Material: Sixteen studied specimens preserved as full reliefin claystone, siltstone and very fine-grained sandstone (MPZ2002/860, 868, 879, 888, 897, 899, 900, 904, 905, 906, 909,913–916 and 918).
Description: Variably curved to undulating, simple subcy-lindrical burrows of length ranging from 3.4 to 20.0 mm; widthranges from 0.5 to 2.9 mm and may change along the path.Surface is usually smooth, but rounded longitudinal ridgesmay be present (MPZ 2002/904).
Stratigraphical distribution and biochronology: Arroyode Pedroche 1 section, levels 2, 9 (upper part, above the Pe-droche event) and 11; middle part of the Puente de Hierro sec-tion. Lower Ovetian.
Ichnogenus Psammichnites Torell, 1870.
Type ichnospecies: Arenicolites gigas Torell, 1868.
Psammichnites gigas (Torell, 1868).
Fig. 9(1–3, 4b, 6b).
Material: Four specimens collected, two of them preservedas full relief at the top of a bed (MPZ 2002/816 and 817), onepreserved as concave epirelief (MPZ 2002/849), and one pre-served as full relief at the bottom of a bed (MPZ 2002/847).All were found in fine-grained quartzite. Additional material isin one large block of rippled sandstone which remains in thefield (level AP1/9).
Description: Horizontal burrows, elliptical in cross-section,showing a complex structure which includes an axial, sinuoustrack on the upper levels and an axial depression on the middlelevels of the trace, both preserved as cleavage relief (Fig. 9(1,2). The bottom of the burrow show meniscate structures (Fig. 9(3)) and a central, straight to slightly sinuous cord (arrowed inFig. 9(1a)). In Fig. 9(2) the cross-section shows the specimenMPZ 2002/816 disrupted and shifted upwards by the cross-over of specimen MPZ 2002/817 slightly from below (white,dotted contour on the photograph). Length of the burrowsamong collected material range from 60 to 175 mm, widthfrom 12 to 21.6 mm, and preserved thickness from 2.0 to13 mm. Specimens remaining in the field show larger dimen-sions (ca. 35–40 mm in width) and display lasso patterns.
Interpretation and discussion: The complex morphologyof Psammichnites gigas resulted from the bulldozing of a rela-tively large soft-bodied marine animal (probably a flat worm)parallel to the bedding plane a few centimetres below the sur-face, which collected organic detritus from the sediment sur-
face with a ciliated “snorkel” and backfilled actively the bur-row (Seilacher and Gámez-Vintaned, 1995; Seilacher Drexlerand Seilacher, 1999). The pendular movement of this organcombined with body motion to produce the sinuous track ob-served in upper levels of the trace, as the sediment was sharplycut (arrows on Fig. 9(2)). As pointed out by the cited authors,the animal possibly employed a two-level mode of feeding,inhaling oxygenated water and detrital food from the surfaceand pumping H2S for bacterial chemosymbionts from the porewater.
Stratigraphical distribution and biochronology: Arroyode Pedroche 1 section, levels 2 and 9 (lower part, below thePedroche event); middle part of the Puente de Hierro section.Lower Ovetian.
Psammichnites ichnosp. A.Fig. 8(1e, 5f, 6b, 8c, d) and Fig. 9(5).Material: Sixteen specimens preserved as full relief and
convex hyporelief in very fine-grained sandstone (MPZ 2002/718, 730, 814, 815, 821, 822, 834, 835, 842, 859, 864, 892,907, 908, 911 and 931).
Description: Basic morphology is similar to that of P. gigasbut differs in that burrow lining is present and size is smaller.Burrows range from 13 to 112 mm in length and from 1.2 to11.3 mm in width. Also, the higher number of specimens per-mits one to reconstruct the complete set of features of the trace,which further includes fine transverse striae on the lowermostsurface of the burrow, meniscate backfilling above this and anaxial, straight cord (MPZ 2002/718 in Fig. 8(5f)). The axial,sinuous track is also visible from the bottom part of the trace.Specimen MPZ 2002/821 shows a sinuous path (wavelength,62 mm) and cross over one specimen of Dactyloidites cabanasi(Fig. 8(8c)); the latter feature is observed also elsewhere.
Interpretation and discussion: Morphological similaritieswith P. gigas point to the same basic behaviour, but differencesin size and burrow lining may give support to a separate ich-nospecies.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section; Arroyo de Pedroche 1section, levels 9 (upper part, above the Pedroche event) and11. Lower Ovetian.
Ichnogenus Rusophycus Hall, 1852.Type ichnospecies: Fucoides biloba Vanuxem, 1842.
Rusophycus didymus (Salter, 1856).Fig. 7(8a–c).Material: Three specimens preserved as convex hyporelief
on a single slab of very fine-grained sandstone (MPZ 2002/721to 723).
Description: Shallow, coffee-bean bilobate traces with fewscratch marks which are not organised into sets. The midlinefurrow widens to the rear (as the total width of the trace does)in one opisthocline specimen (MPZ 2002/721) and is homoge-neously narrow (as is the total width) in the other two. TheV-angle of marks is ca. 45° in MPZ 2002/721 and near 180°

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 463
in the others. The traces are 7.4–8.5 mm wide and 10–14 mmlong. All the three appear parallel to four long, cylindrical bur-rows, and near two specimens of Dactyloidites cabanasi.
Interpretation and discussion: The general shape of thetraces and their tiny sizes allow the identification with R. didy-mus. Usually too faint scratch mark patterns make this ichnos-pecies rather poorly defined. In our case, variations within theendopodite scratch marks pattern are due to differences in thedigging behaviour of the arthropod producer. Some authorsconsidered that the holotype of Rusophycus didymus is prob-ably of inorganic origin (Pickerill, personal communication inMángano et al., 1996). Further studies on the holotype are thusneeded.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Ichnogenus Skolithos Haldemann, 1840.
Type ichnospecies: Skolithos linearis Haldemann, 1840.
Skolithos ichnosp. indet.
Fig. 8(8e).
Material: Four studied specimens preserved as full relief infine-grained and very fine-grained sandstone (MPZ 2002/823,829, 869 and 895). Additional material remains in the field.
Description: Vertical to oblique, cylindrical burrows ofshort length (most frequently less than 1 cm) and a few milli-metres in diameter (from 3.6 to 5.3 mm).
Stratigraphical distribution and biochronology: Arroyode Pedroche 1 section, levels 2 and 4; middle part of the Puentede Hierro section. Lower Ovetian.
Ichnogenus Torrowangea Webby, 1970.
Type ichnospecies: Torrowangea rosei Webby, 1970.
Torrowangea rosei Webby, 1970.
Fig. 10(4b, c).
Material: Two specimens preserved as full relief in silt-stone (MPZ 2002/951 and 952).
Description: Simple, meandering horizontal burrows inwhich thickenings and constrictions alternate. Diameter from1.2 to 1.5 mm; rectified length up to 23 mm.
Interpretation and discussion: These traces are interpretedas shallow feeding burrows actively infilled with pellets.Although the two specimens are incompletely preserved,meandering allows identifying them as T. rosei Webby, 1970.
Stratigraphical distribution and biochronology: Arroyode Pedroche 1 section, level 11. Lower Ovetian.
Torrowangea ichnosp. indet.
Fig. 8(6d), 9(7c) and Fig. 10(4d, 7, 8a).
Material: Fourteen specimens preserved as full relief andconcave epirelief in fine to very fine-grained sandstone andsiltstone (MPZ 2002/732, 792-794, 811, 874-876, 878, 893,894, 912, 919 and 921).
Description: Isolated, simple, thin, straight to slightly wind-ing, horizontal burrows in which thickenings and constrictionsalternate. Diameter from a few tenths of millimetre up to1.6 mm; length up to 55 mm.
Interpretation and discussion: These traces are interpretedas shallow feeding burrows actively infilled with pellets. Theisolation and the lack of intense windings or loops prevents theidentification with T. rosei Webby, 1970. In the absence ofquick turns producing angles on otherwise straight paths, theichnospecies T. angulata Gámez-Vintaned and Liñán, 1993 canalso be discarded.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section; Arroyo de Pedroche 1section, levels 11 and 12. Lower Ovetian.
Ichnogenus Treptichnus Miller, 1889.
Type ichnospecies: Treptichnus bifurcus Miller, 1889.
Treptichnus ichnosp. A.
Fig. 9(7a, 8, 9a).
Material: Six specimens, two of them preserved as full re-lief at the bottom of very fine-grained sandstone (MPZ 2002/733 and 799), three preserved as concave epirelief on fine-grained sandstone (MPZ 2002/806, 807 and 809) and one asfull relief in siltstone (MPZ 2002/902).
Description: Straight sets of individual burrows curved hor-izontal to vertically of similar length arranged into a zigzagpattern; one is connected to another at their lower parts. Indi-vidual burrows are from 11.4 to 20.7 mm in length, and from1.0 to 2.1 in width. Sets are from 22.4 to 74.0 mm, and from5.5 to 16.0 mm in width.
Interpretation and discussion: The appearance of Trep-tichnus varies according to the level at which the trace is ob-served (a basal level is shown in MPZ 2002/806; Fig. 9(7a)),and also if viewed either from the bottom (Fig. 9(9a)) or thetop of the stratum (Fig. 9(7a, 8)).
Stratigraphical distribution and biochronology: Arroyode Pedroche 1 section, level 9 (upper part, above the Pedrocheevent); middle part of the Puente de Hierro section. LowerOvetian.
Treptichnus ichnosp. B.
Fig. 9(7b).
Material: One specimen preserved as concave epirelief onfine-grained sandstone (MPZ 2002/808).
Description: A set of four straight individual burrows ar-ranged into a square pattern. The set is in ontogenetic connec-tion with a set of burrows organised into a Treptichnus ich-nosp. A pattern (Fig. 9(7b, a)). Individual burrows are from6.8 to 14.2 mm in length, and 0.7 mm in width; the total lengthof the set is 27 mm, and its total width 12.5 mm.
Interpretation and discussion: Although this morphologywould not fit strictly with Treptichnus, the ontogenetic conti-nuity with Treptichnus ichnosp. A and the same type of bifur-cation (each successive burrow branching from the distal part

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468464
of the previous one) support the inclusion within this ichno-genus.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
Undetermined burrow.Fig. 6(8).Material: One specimen preserved as sandy full relief with-
in claystone (MPZ 2002/898).Description: Trace fossil characterised by a very simple
pattern composed of horizontal burrows of elliptical section,3 mm in width and a total length of 55 mm, branching at acuteangles. Ornamentation consists of fine, faint longitudinalridges.
Interpretation and discussion: This ichnofossil stronglyresembles Phycodes ichnosp. A described by Gámez Vintanedand Mayoral (1995) from lutites of the Middle CambrianMurero Formation at Murero (Cadenas Ibéricas, Spain).
Stratigraphical distribution and biochronology: Arroyode Pedroche 1 section, level 9 (upper part, above the Pedrocheevent). Lower Ovetian.
Faecal pellets.Fig. 6(10, 12) and Fig. 8(1d).Material: Numerous specimens preserved as full relief, and
as concave semireliefs, in siltstone and fine-grained, micaceoussandstone. Among them, nine items are given the referencesMPZ 2002/773, 803, 881-883, 885-887 and 901.
Description: Pellets can be found either grouped or scat-tered on the rock. Scattered pellets range from 0.2 to 4.7 mmin diameter. MPZ 2002/881, 883 and 901.
Groupings are either linear or in circles. Linear groups areeither straight and simple or curved and double (MPZ 2002/886; two parallel lines are 5 mm apart), accounting for a fewpellets from 0.8 to 1.8 mm in diameter each. Total lengthreaches up to 30 mm.
Circular groupings show 5–15 pellets each, reaching up to14.7 mm in diameter. Diameter of pellets range from 0.3 to1.4 mm. MPZ 2002/773, 803, 882, 885 and 887.
Interpretation and discussion: The material is interpretedas faecal pellets after its morphology, arrangement patterns andmode of preservation. Prefossilization processes (Seilacher,2002) should be invoked for preservation of primary morphol-ogies.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section; Arroyo de Pedroche 1section, levels 9 (upper part, above the Pedroche event) and11. Lower Ovetian.
Meniscate trace fossils.Fig. 8(1c) and Fig. 10(8b).Material: Six studied specimens preserved as full relief in
siltstone (MPZ 2002/724, 759, 771, 782 and 802) and in veryfine-grained, micaceous sandstone (MPZ 2002/932).
Description: Straight to slightly curved, simple subcylind-rical to vertically flattened burrows with meniscate infilling of
the same type as the host rock. Burrows with closely spacedmenisci (Fig. 10(8b)) seem to lack lining, while those withwidely spaced menisci (Fig. 8(1c)) have a thin lining. Lengthranges from a few millimetres to 50 mm, and width from 1.4 to9.0 mm.
Interpretation and discussion: The material probably in-cludes two ichnotaxa related to Taenidium.
Stratigraphical distribution and biochronology: Middlepart of the Puente de Hierro section. Lower Ovetian.
5. Palaeoenvironments and palaeoecological turnover atthe Pedroche event
Sedimentological studies on the Member I of the PedrocheFormation at the Arroyo de Pedroche 1 and other sections inSierra de Córdoba were made by Zamarreño and Debrenne(1977) and by Moreno-Eiris (1987a, 1987b, 1987c).
The base of the Arroyo de Pedroche 1 (AP1) section is cov-ered by quaternary deposits. According to Perejón et al. (1996),the characteristics of the Cambrian sediments suggest a shallowcarbonate platform with intermittent clastic influence, as it isrecorded by the silts and sandstones, interbedded with lime-stones and sandy limestones (Fig. 4).
This alternation of terrigenous and carbonates is repeatedthroughout the section and is interpreted as a shallowing se-quence. This sequence begins with terrigenous deposits, siltsand sandstones, which are coarse bedded and show parallellamination, cross-bedding and ripples. Some thin lenticularlimestone beds are interbedded with terrigenous sediments.
The carbonates at top of the section are thicker and showdifferent facies types, such as oolitic, oncolitic, calcimicrobe,stromatolitic, bioclastic and mound limestones.
Oolitic limestones are often present and these grainstonesconstitute ooid shoals. In this section calcimicrobe limestonesand mounds are present on top of the oolitic bars.
The biogenic limestones are made of several componentswhich change according to environmental conditions. The cal-cimicrobe limestones are made of boundstones of Renalcis andEpiphyton, and peloidal wackestones-packstones, formed in aprotected environment. Stromatolitic limestones have beenfound in several levels of the section, sometimes associatedwith scattered archaeocyaths (Schmitt, 1983).
The small mounds are formed by archaeocyaths and calci-microbes. The cups of Protopharetra bigoti Debrenne, 1964and Agastrocyathus gregarius Debrenne, 1961 constitutebranching colonies which are related to Nochoroicyathus, Cos-cinocyathus, and they are surrounded by calcimicrobes such asEpiphyton, Renalcis and Girvanella. These boundstones consti-tute a framework, which has been previously described inLower Cambrian of the Poleta Formation, Nevada (Rowland,1984) and the Nebida Formation, Sardinia (Bechstädt et al.,1985).
During the algal activity and the development of themounds on this carbonate-terrigenous platform, ooid shoalswere formed, which were related to algal limestones in a pro-tected area where terrigenous material was deposited. Small

J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 465
reef mounds were developed on top of the oolitic bars. Theterrigenous deposit is more important during certain episodes,until it prevails over carbonate sedimentation.
In summary, the sedimentary evolution of the Member I ofthe Pedroche Formation is continuous, without importantbreaks in sedimentation.
From a palaeoecological point of view, the upper part of thelowermost Cambrian Torreárboles Formation records the onsetof widespread, open sublittoral ecosystems at the beginning ofthe Cambrian transgression in Sierra Morena (Liñán and Fer-nández Carrasco, 1984; Liñán and Quesada, 1990). Arthro-pods, annelids and coelenterates colonized the sea bottom, pro-ducing distinctive trace fossils (Fedonkin et al., 1985; Liñán,1984a). Subsequently, biothermal organisms colonized themarine platform, and carbonate constructions with archaeo-cyaths and calcimicrobes developed during the deposition ofthe Pedroche Formation (Member I) (Perejón, 1975a, 1975b,1975c, 1976a, 1976b, 1977, 1989; Moreno-Eiris, 1987d). Perireefal communities of small shelly fossils, sponges, bradoriidsand trilobites also developed in carbonate substrates, whileclastic substrates accommodated soft-bodied communities pro-ducing ichnocoenoses.
Archaeocyatha are well represented in the AP1 section,where four zones were recognised by Perejón (1986, 1994).The boundary between Zones II and III represent the extinctionof characteristic archaeocyath genera, followed by the radiationof new forms (Perejón, 1989).
Stromatolites spread at the lower-middle part of the Pe-droche Formation (Member I) and decreased at the base ofthe Bigotina bivallata biozone (Fig. 4). The decrease in stro-matolite diversity during Early Cambrian times is a generaltrend which is also recorded in other regions of the world. Inthe Anti-Atlas of Morocco, it took place within the Lemdadellahorizon of the Lemdad section (Sdzuy, 1978; Schmitt, 1979).In Russia, stromatolites diversified from Vendian to Lower At-dabanian in Uchur-Maya and Anabar regions (Krylov, 1963;Komar, 1966, 1973; Semikhatov et al., 1970; Keller, 1973).The stromatolite taxa distinguished in the Arroyo de Pedrochesection are Vetella cf. sarfatiae, Charaulachia cordobensis andVetella nodosa, all of them from the archaeocyath Zone II.Charaulachia and Vetella are two Neoproterozoic generawhich disappeared from Morocco (High Atlas), the SiberianPlatform (Charaulach and Olenek) and the Sierra de Córdobain Early Cambrian times (Schmitt, 1979,1983).
Some ichnoassemblages from the lower part of the ArroyoPedroche 1 section (levels 1 to 4) are dominated by both hor-izontal and short, vertical domichnia, while others show fodi-nichnia and pascichnia, all preserved in substrata of grain sizefrom siltstone to medium sandstone, suggesting a shallow mar-ine environment with rather clean substrata under relativelyhigh energy conditions. Ichnotaxa of the Cruziana ichnofacies(including Psammichnites gigas) alternate here with ichnogen-era typical of the Skolithos ichnofacies (Fig. 4), reflectingshort-term, alternating energy conditions which produced dif-ferent grain-sizes in substrata.
Ichnological data are very scarce from the middle part of thesection. In the upper part (archaeocyath Zone III), ichnoassem-blages of a different composition are recorded in siltstone andfine-grained sandstone and show a change in the Psammich-nites ethology. They can be grouped into a well-characterisedPsammichnites ichnosp. A ichnoassociation (this term, sensuGámez Vintaned and Mayoral Alfaro, 1992, is defined in baseof the ichnotaxon or ichnotaxa which are recurrently present ina number of ichnocoenoses) and show a variety of fodinichnia,pascichnia and horizontal domichnia indicative of enriched, in-fralittoral substrata under moderate energy conditions (Cru-ziana ichnofacies). The Psammichnites ichnosp. A ichnoasso-ciation occurs immediately above the last occurrence ofstromatolites and coincides with the trilobite and brachiopodexpansion (Fig. 4). A very similar set of ichnoassemblages isrecorded in fine-grained sandstone and siltstone at the Puentede Hierro section (Lemdadella linaresae biozone, archaeocyathZone III), with the only difference of the great abundance ofDactyloidites cabanasi. This points to substrata even richerthan for the upper part of the Arroyo Pedroche 1 section, underotherwise very similar palaeoecological conditions (Fig. 3).The ubiquity of Dactyloidites cabanasi in the lower and middleparts of the Puente de Hierro section permit those ichnoassem-blages to be grouped into a Dactyloidites ichnoassociation.
Trilobites of the basal levels of the Arroyo de Pedroche 1section occur together with other perireefal groups, preservedin calcareous levels. The species Bigotina bivallata appearsabove the disappearance of characteristic archaeocyaths ofZone II, preserved in limestones and coinciding with the lastrecord of columnar stromatolites. Trilobites start to abound co-inciding with the onset of Psammichnites ichnosp. A ichnoas-sociation, being preserved mainly in siliciclastic rocks, out ofthe reefal influence. Soon after, the brachiopod facies (repre-sented here by the genus Paterina) are recorded for the firsttime in the section.
In summary, low Lower Cambrian (lower Ovetian) rocksfrom Sierra de Córdoba record a biotic turnover (Pedrocheevent; Liñán, 2003) which took place at the lower part of theBigotina bivallata biozone, a few meters above the disappear-ance of characteristic archaeocyaths of Zone II of Perejón(1986, 1989) and immediately above the last record of colum-nar stromatolites. At that time, palaeoecological conditionsshifted from a shallow, somewhat restricted infralittoral envir-onment (recording archeocyath, stromatolite and scarce trilo-bite facies) to a less energetic, probably deeper, infralittoralenvironment (with archeocyath, brachiopod and abundant trilo-bite facies). Thus, trace and body fossils provide evidencessupporting a remarkable palaeoecological disturbance, whichwas likely to cause the seeming decrease of stromatolite-build-ing communities and the expansion of trilobite, brachiopod andsoft-bodied metazoan biotas, in sublittoral (infralittoral) envir-onments. A remarkable palaeoecological shift is also recordedin horizons of similar age at sections in Morocco and Russiahence pointing to a global event, which confirmation will re-quire further studies.

Fu
Fü
Gá
Gá
Gá
Gá
Ga
Glu
Ha
Ha
Ha
Ha
He
Hit
Ho
Ke
Ko
Ko
Kry
Liñ
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468466
Acknowledgements
Financial support was given by the Dirección General deInvestigación, Spanish Ministerio de Educación y Cultura (Pro-jects BTE2000-1145-C02 C1 and C2, BTE2003-04997) and bythe Diputación General de Aragón (Grupo Consolidado E-17,“Patrimonio y Museo Paleontológico”). M.E. Dies benefitsfrom a postdoctoral grant of the Spanish Ministerio de Educa-ción y Ciencia in Lund University. Comments from two re-viewers, doctors L. Buatois and A. Uchman, are acknowl-edged.
References
Agirrezabala, L.M., De Gibert, J.M., 2004. Paleodepth and paleoenvironmentof Dactyloidites ottoi (Genitz, 1849) from Lower Cretaceous deltaic depos-its (Basque-Cantabrian basin, West Pyrenees). Palaios 19, 276–291.
Azpeitia Moros, F., 1933. Datos para el estudio paleontológico del Flysch dela Costa Cantábrica y de algunos otros puntos de España. Boletín del Ins-tituto Geológico y Minero de España 53 (1a serie), 1–65.
Bassler, R.S., 1941. A supposed jellyfish from the pre-Cambrian of the GrandCanyon. Proceedings of the United States National Museum 89 (3104),519–522.
Bechstädt, T., Boni, M., Selg, M., 1985. The Lower Cambrian of SW Sardi-nia: from a clastic tidal shelf to an isolated carbonate platform. Facies 12,113–140.
Billings, E., 1862. New species of fossils from different parts of the Lower,Middle and Upper Silurian rocks of Canada. In: Billings, E. (Ed.), Palaeo-zoic fossils, Vol. 1: containing descriptions and figures of new or littleknown species of organic remains from the Silurian rocks. (1861–1865).Geological Survey of Canada, Ottawa, pp. 96–168.
Buatois, L.A., Mángano, M.G., Maples, C.G., Lanier, W.P., 1998. Taxonomicreassessment of the ichnogenus Beaconichnus and additional examplesfrom the Carboniferous of Kansas, USA. Ichnos 5, 287–302.
Cabanás, G., 1966. Notas estratigráficas de la provincia de Córdoba. Con unanota sobre un nuevo fósil del Cambriano por Bermudo Meléndez. Notas yComunicaciones del Instituto Geológico y Minero de España 90, 77–84.
Dawson, J.W., 1873. Impressions and footprints of aquatic animals and imita-tive markings, on Carboniferous rocks. American Journal of Science 5 (3),16–24.
Debrenne, F., 1961. Nouvelles données sur la faune d’Archaeocyatha du JbelTaïssa (Anti-Atlas occidental). Notes et Mémoires du Service géologiquedu Maroc 20, 7–37.
Debrenne, F., 1964. Archaeocyatha. Contribution à l’étude des faunes cam-briennes du Maroc, de Sardaigne et de France. Notes et Mémoires du Ser-vice géologique du Maroc 179, 5–265.
Díez Balda, M.A., 1986. El Complejo esquisto-grauváquico, las series paleo-zoicas y la estructura hercínica al Sur de Salamanca. Ediciones Universi-dad de Salamanca, Salamanca (162 pp.).
Fedonkin, M.A., 1981. Belomorskaja biota venda (dokembriiskaja besskelet-naja fauna severa Russkoi platformi). [The Vendian White Sea biota (Pre-cambrian non-skeletal fauna of the North Russian Platform)]. Trudi Geolo-gicheskogo Instituta, Akademija Nauk SSSR 342, 1–100 (in Russian).
Fedonkin, M., Liñán, E., Perejón, A., 1985. Icnofósiles de las rocas precám-brico-cámbricas de la Sierra de Córdoba, España. Boletín de la Real Socie-dad Española de Historia Natural (Geología) 81 (1983), 5–14.
Fernández Remolar, D.C., 1996. Los microfósiles problemáticos del CámbricoInferior de la Sierra de Córdoba: datos preliminares. Coloquios de Paleon-tología 48, 161–173.
Fernández Remolar, D.C., 1999. Las calizas fosforíticas del Ovetiense Inferiorde la Sierra de Córdoba, España. Boletín de la Real Sociedad Española deHistoria Natural (Geológica) 95, 15–45.
Fillion, D., Pickerill, R.K., 1990. Ichnology of the Upper Cambrian? to LowerOrdovician Bell Island and Wabana groups of eastern Newfoundland, Ca-nada. Palaeontographica Canadiana 7, 1–119.
chs, T., 1895. Studien über Fucoiden und Hieroglyphen. Kaiserlichen Aka-demie der Wissenschaften zu Wien, mathematisch-naturwissenschaftlicheClasse. Denkschriften 62, 369–448.rsich, F.T., Bromley, R.G., 1985. Behavioural interpretation of a rosettedspreite trace fossil: Dactyloidites ottoi (Geinitz). Lethaia 18, 199–207.mez-Vintaned, J.A., Liñán, E., 1993. 5 Paléoichnologie. In: Van Amerom,H.W.J., Broutin, J., Ferrer, J., Gámez-Vintaned, J.A., Gisbert, J., Liñán, E.(Eds.), Les flores du Permien basal et la paléoichnologie de la fosse deFombuena (province de Zaragoza, Espagne). Mededelingen Rijks Geolo-gische Dienst 48, pp. 19–26.mez Vintaned, J.A., Liñán, E., 1996. Revisión de la terminología icnológicaen español. Revista Española de Paleontología 11, 155–176.mez Vintaned, J.A., Mayoral Alfaro, E., 1992. Primeras aportaciones a laPaleoicnología del Grupo Murero (Cámbrico inferior-medio) en Murero.(Provincia de Zaragoza. Cadena Ibérica Occidental). Geogaceta 12, 100–102.mez Vintaned, J.A., Mayoral, E., 1995. Paleoicnología del Grupo Mesones(Cámbrico Inferior-Medio) en Murero (Cadena Ibérica Occidental, NE deEspaña). In: Gámez Vintaned, J.A., Liñán, E. (Eds.), Memorias de las IVJornadas Aragonesas de Paleontología: “La expansión de la vida en elCámbrico”. Libro homenaje al Prof. Klaus Sdzuy. Institución “Fernandoel Católico”, Zaragoza, pp. 219–252.rcía Hernández, M., Liñán, E., 1983. Estromatolitos y facies asociadas en laFormación Santo Domingo (Cámbrico inferior de la Sierra de Córdoba).In: Comba, J.A. (Ed.), Libro Jubilar J.M. Ríos, 3. Instituto Geológico yMinero de España, Madrid, pp. 125–132.szek, A., 1995. Invertebrate trace fossils in the continental deposits of anUpper Carboniferous coal-bearing succession, Upper Silesia, Poland. Stu-dia Geologica Polonica 108, 171–202.ldemann, S.S., 1840. Supplement to number one of “A monograph of theLimniades, and other fresh-water univalve shells of North America”, con-taining descriptions of apparently new animals in different classes, and thenames and characters of the subgenera in Paludina and Anculosa. J. Dob-son, Philadelphia (pp. 1–3).ll, J., 1847. Palaeontology of New York. Volume 1, C. Van Benthuysen,Albany (338 pp.).ll, J., 1852. Palaeontology of New York. Volume 2, C. Van Benthuysen,Albany (362 pp.).ll, J., 1886. Note on some obscure organisms in the roofing slate of Wa-shington County, New York. In: Trustees New York State Museum of Nat-ural History, 39th annual Report. New York State Museum of Natural His-tory, New York, p. 160.rnández Pacheco, E., 1907. Los martillos de piedra y las piedras con cazo-letas de las antiguas minas de cobre de la Sierra de Córdoba. Boletín de laReal Sociedad Española de Historia Natural 7, 279–292.chcock, E., 1858. Ichnology of New England. A report on the sandstone ofthe Connecticut Valley, especially its fossil footmarks. W. White, Boston(220 pp.).ward, J.D., 1966. Characteristic trace fossils in Upper Cretaceous sand-stones of the Book Cliffs and Wasatch Plateau. Utah Geological andMineralogical Survey, Bulletin 80, 35–53.ller, B.M., 1973. Vendian, Yudomian, and terminal Riphean (Vendomian).Izvestija Akademii nauk SSSR, Seriya Geologicheskaya 1973, 86–92 (inRussian).mar, V.A., 1966. Upper Precambrian stromatolites in the northern part ofthe Siberian Platform and their stratigraphic significance. Trudi Geologi-cheskogo Instituta, Akademija Nauk SSSR 154, 1–122 (in Russian).mar, V.A., 1973. Novye dannye o korrelyatsii rifeiskikh otlozhenii Yuzh-nogo Urala i Sibiri. Izvestija Akademii nauk SSSR, Seriya Geologiches-kaya 1973, 30–36 (in Russian).lov, I.N., 1963. Columnar branching stromatolites of the Riphean depositsof the Southern Ural and their significance for the stratigraphy of theUpper Precambrian. Trudi Geologicheskogo Instituta, Akademija NaukSSSR 69, 1–133 (in Russian).án, E., 1984a. Los icnofósiles de la Formación Torreárboles (¿Precámbri-co?-Cámbrico Inferior) en los alrededores de Fuente de Cantos, Badajoz.Cuadernos do Laboratorio Xeolóxico de Laxe 8, 47–74.

Liñ
Liñ
Liñ
Liñ
Liñ
Liñ
Liñ
Liñ
Liñ
Liñ
Liñ
Liñ
Liñ
Lu
Má
Mc
Mi
Mo
Mo
Mo
Mo
Nic
Nic
Pal
Pal
Pal
Pem
Per
Per
Per
Per
Per
Per
Per
Per
Per
Per
Pil
Pra
Ric
RicRic
Ro
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468 467
án, E., 1984b. Introducción al problema de la Paleogeografía del Cámbricode Ossa Morena. Cuadernos do Laboratorio Xeolóxico de Laxe 8, 283–314.án, E., 2003. Bioeventos y bioestratigrafía en el Cámbrico de España. In:Lamolda, M.A. (Ed.), Bioevents: their stratigraphical records, patterns andcauses. Caravaca de la Cruz, June 3rd-8th 2003 (pp. 32–34).án, E., Álvaro, J., Gozalo, R., Gámez-Vintaned, J.A., Palacios, T., 1995. ElCámbrico Medio de la Sierra de Córdoba (Ossa-Morena, S de España): tri-lobites y paleoicnología. Implicaciones bioestratigráficas y paleoambien-tales. Revista Española de Paleontología 10, 219–238.án, E., Dabrio, C., 1974. Litoestratigrafía del tramo inferior de la Forma-ción de Pedroche (Cámbrico Inferior) (Córdoba). Acta Geológica Hispáni-ca 9, 21–26.án, E., Dies, M.E., Gámez Vintaned, J.A., Gozalo, R., Mayoral, E., Muñiz,F., 2005. Lower Ovetian (Lower Cambrian) trilobites and biostratigraphyof the Pedroche Formation (Sierra de Córdoba, southern Spain). Geobios38, 365–381.án, E., Fernández-Carrasco, J., 1984. La Formación Torreárboles y la pa-leogeografía del límite Precámbrico-Cámbrico en Ossa Morena (FlancoNorte de la alineación Olivenza-Monesterio). Cuadernos do LaboratorioXeolóxico de Laxe 8, 315–328.án Guijarro, E., 1974. Las formaciones cámbricas del norte de Córdoba.Acta Geológica Hispánica 9, 15–20.án Guijarro, E., 1978. Bioestratigrafía de la Sierra de Córdoba. Tesis doc-torales de la Universidad de Granada, 191. Secretariado de Publicacionesde la Universidad de Granada, Granada (212 pp.).án Guijarro, E., Dies Álvarez, M.E., Gámez Vintaned, J.A., Gozalo Gutié-rrez, R., Mayoral Alfaro, E., Muñiz Guinea, F., Chirivella Martorell, J.B.,2004. Hallazgo del trilobites Alueva hastata (Sdzuy, 1958) y de pistas fó-siles en el Cámbrico Medio temprano de Sierra Morena. Consecuencia re-gionales y paleoambientales. In: Baquedano, E., Rubio Jara, S. (Eds.), Mis-celánea en Homenaje a Emiliano Aguirre. Volumen II. Paleontología. ZonaArqueológica, 4, Museo Arqueológico Regional de la Comunidad de Ma-drid, Alcalá de Henares, pp. 256–263.án, E., Moreno-Eiris, E., Perejón, A., Schmitt, M., 1982. Fossils from thebasal levels of the Pedroche Formation, Lower Cambrian (Sierra Morena,Córdoba, Spain). Boletín de la Real Sociedad Española de Historia Natural(Geología) 79 (1981), 277–286.án, E., Perejón, A., Sdzuy, K., 1993. The Lower-Middle Cambrian stagesand stratotypes from the Iberian Peninsula: a revision. Geological Maga-zine 130, 817–833.án, E., Quesada, C., 1990. Part V Ossa-Morena Zone. 2. Stratigraphy. 2.2.Rift Phase (Cambrian). In: Dallmeyer, R.D., Martínez García, E. (Eds.),Pre-Mesozoic Geology of Iberia. Springer-Verlag, Berlin, Heidelberg, pp.259–266.án, E., Sdzuy, K., 1978. A trilobite from the Lower Cambrian of Córdoba(Spain) and its stratigraphical significance. Senckenbergiana lethaea 59,387–399.dwig, R., 1869. Fossile Pflanzenreste aus den palaeolithischen Formationender Umgebung von Dillenburg, Biedenkopf und Friedberg und aus demSaalfeldischen. Palaeontographica 17, 105–128.ngano, M.G., Buatois, L.A., Aceñolaza, G.F., 1996. Trace fossils and sedi-mentary facies from a Late Cambrian-Early Ordovician tide-dominatedshelf (Santa Rosita Formation, northwest Argentina): implications for ich-nofacies models of shallow marine successions. Ichnos 5, 53–88.Cann, T., 1993. A Nereites ichnofacies from the Ordovician-Silurian WelshBasin. Ichnos 3, 39–56.ller, S.A., 1889. North American geology and palaeontology for the use ofamateurs, studentes and scientists. Western Methodist Book Concern, Cin-cinnati (664 pp.).reno-Eiris, E., 1987a. Los montículos arrecifales de Algas y Arqueociatosdel Cámbrico Inferior de Sierra Morena. I: Estratigrafía y Facies. BoletínGeológico y Minero 98, 295–317.reno-Eiris, E., 1987b. Los montículos arrecifales de Algas y Arqueociatosdel Cámbrico Inferior de Sierra Morena. II: Las algas calcáreas. BoletínGeológico y Minero 98, 449–459.
reno-Eiris, E., 1987c. Los montículos arrecifales de Algas y Arqueociatosdel Cámbrico Inferior de Sierra Morena. III: Microfacies y Diagénesis. Bo-letín Geológico y Minero 98, 591–621.reno-Eiris, E., 1987d. Los montículos arrecifales de Algas y Arqueociatosdel Cámbrico Inferior de Sierra Morena. IV: Bioestratigrafía y Sistemáticade los Arqueociatos. Boletín Geológico y Minero 98, 729–779.holson, H.A., 1873. Contributions to the study of the errant annelides ofthe older Palaeozoic rocks. Proceedings of the Royal Society of London21, 288–290.holson, H.A., Hinde, G.J., 1875. Notes on the fossils of the Clinton, Nia-gara, and Guelph formations of Ontario, with descriptions of new species.Canadian Journal of Science, Literature, and History, new series 14, 137–140.acios, T., Gámez, J.A., Fernández-Remolar, D., Liñán, E., 1999a. New pa-laeontological data at the Vendian-Cambrian transition in central Spain.Journal of Conference Abstracts 4 (1), 265 (EUG 10, Abstract Volume).acios, T., Gámez Vintaned, J.A., Fernández-Remolar, D., Liñán, E., 1999b.The lowermost Cambrian in the Valdelacasa Anticline (central Spain):some new palaeontological data. Journal of Conference Abstracts 4 (3),1017 (XV RGOP Abstract Volume).ij, V.M., Posti, E., Fedonkin, M.A., 1979. Soft-bodied metazoa and tracefossils of the Vendian and Lower Cambrian. In: Keller, B.M., Rozanov,A. Yu (Eds.), Upper Precambrian and Cambrian paleontology of East-Eur-opean Platform. Akademija Nauk SSSR, Moskva, pp. 49–82 (in Russian).[English translation in: Urbanek, A., Rozanov, A. Yu. (Eds.), PublishingHouse Wydawnictwa Geologiczne, Warszawa, pp. 56–93. Year 1983].berton, S.G., Frey, R.W., 1982. Trace fossil nomenclature and the Plano-lites-Palaeophycus dilemma. Journal of Paleontology 56, 843–881.ejón, A., 1975a. Nuevas faunas de Arqueociatos del Cámbrico inferior deSierra Morena. Tecniterrae 8, 8–29.ejón, A., 1975b. Arqueociatos de lo subórdenes Monocyathina y Dokido-cyathina. Boletín de la Real Sociedad Española de Historia Natural (Geo-logía) 73, 125–145.ejón, A., 1975c. Arqueociatos regulares del Cámbrico inferior de SierraMorena (SW de España). Boletín de la Real Sociedad Española de HistoriaNatural (Geología) 73, 147–193.ejón, A., 1976a. Nuevas faunas de Arqueociatos del Cámbrico Inferior deSierra Morena (II). Tecniterrae 9, 7–24.ejón, A., 1976b. Nuevos datos sobre los Arqueociatos de Sierra Morena.Estudios Geológicos 32, 5–33.ejón, A., 1977. Arqueociatos con túmulos en el Cámbrico inferior de Cór-doba (Sierra Morena Oriental). Estudios Geológicos 33, 545–555.ejón, A., 1986. Bioestratigrafía de los Arqueociatos en España. Cuadernosde Geología Ibérica 9 (1984), 213–316.ejón, A., 1989. Arqueociatos del Ovetiense en la sección del ArroyoPedroche, Sierra de Córdoba, España. Boletín de la Real Sociedad Españo-la de Historia Natural (Geología) 84, 143–247.ejón, A., 1994. Palaeogeographic and biostratigraphic distribution of Ar-chaeocyatha in Spain. Courier Forschungs-Institut Senckenberg 172, 341–354.ejón, A., Moreno-Eiris, E., Liñán, E., Sequeiros, L., 1996. II.3. September16. The Cambrian of the Sierra de Córdoba (Ossa-Morena Zone. SierraMorena). In: Liñán, E., Gámez Vintaned, J.A., Gozalo, R. (Eds.), II FieldConference of the Cambrian Stage Subdivision Working Groups. Interna-tional Subcommission on Cambrian Stratigraphy. Spain, 13-21 September1996. Field Trip Guide and Abstracts. Universidad de Zaragoza, Zaragoza,pp. 1–129.lola, G.L., 1991. Trilobites du Cambrien inférieur du SW de la Sardaigne,Italie. Palaeontographia Italica 78, 1–173.ntl, F., 1946. Two new problematic trails from the Ordovician of Bohemia.Académie Tchèque des Sciences, Bulletin International, Classe desSciences Mathématiques, Naturelles et de la Médecine 46 (1945), 49–59.hter, R., 1850. Aus der thüringischen Grauwacke. Deutsche GeologischeGesellschaft, Zeitschrift 2, 198–206.hter, R., 1853. Gaea von Salfeld. Programm d. Realsch. Saalfeld, 3–32.hter, R., 1937. Marken und Spuren aus allen Zeiten. I-II. SenckenbergianaBiologica 13, 299–342.wland, S.M., 1984. Were there framework reefs in the Cambrian? Geology12, 181–183.

Sal
Sch
Sch
Sch
Sd
Sd
Sei
Sei
Sei
Sei
Sei
Sem
Sta
To
To
Va
Va
Wa
We
Zam
J.A. Gámez Vintaned et al. / Geobios 39 (2006) 443–468468
ter, J.W., 1856. On fossil remains in the Cambrian Rocks of the Longmyndand North Wales. The Quarterly Journal of the Geological Society of Lon-don 12, 246–251.lirf, M., Uchman, A., 2004. Revision of the ichnogenus Sabellarifex Rich-ter and the value of ichnotaxobases in simple vertical trace fossils. In: Bua-tois, L.A., Mángano, M.G. (Eds.), Ichnia 2004, First International Con-gress on Ichnology, April 19-23, 2004. Museo Paleontológico EgidioFeruglio, Trelew, Patagonia, Argentina, Abstract Book. Museo Paleontoló-gico Egidio Feruglio, Trelew, p. 76.mitt, M., 1979. The section of Tiout (Precambrian/Cambrian boundarybeds, Anti-Atlas, Morocco): Stromatolites and their Biostratigraphy. Arbei-ten der Paläontologische Institut Würzburg 2, 1–188.mitt, M., 1983. Columnar stromatolites from the Lower Cambrian Pe-droche Formation, Sierra Morena, S-Spain. Boletín de la Real SociedadEspañola de Historia Natural (Geología) 80 (1982), 5–23.zuy, K., 1971. Acerca de la correlación del Cámbrico inferior de la Penín-sula Ibérica. I Congreso Hispano-Luso-Americano de Geología Econó-mica, Sección 1, Geología 2, 753–768.zuy, K., 1978. The Precambrian-Cambrian boundary beds in Morocco (Pre-liminary Report). Geological Magazine 1158394.lacher, A., 1977. Pattern analysis of Paleodictyon and related trace fossils.In: Crimes, T.P., Harper, J.C. (Eds.), Trace fossils 2. Geological JournalSpecial Issue 9, pp. 289–334.lacher, A, 1955. IV. Spuren und Lebensweise der Trilobiten. In: Schinde-wolf, O.H., Seilacher, A (Eds.), Beiträge zur Kenntnis des Kambriums inder Salt Range (Pakistan). Akademie der Wissenschaften und der Literaturin Mainz, mathematisch-naturwissenschaftliche Klasse, Abhandlungen, pp.342–372.lacher, A., 2002. Non olet: the strange taphonomy of coprolites and colo-lites. In: De Renzi, M., Pardo Alonso, M.V., Belinchón, M., Peñalver, E.,Montoya, P., Márquez-Aliaga, A. (Eds.), Current Topics on Taphonomyand Fossilization. Col-lecció Encontres, 5. Ajuntament de Valencia, Valen-cia, pp. 233–240.
lacher, A., Gamez-Vintaned, J.A., 1995. Psammichnites gigas: Ichnologicalexpression of the Cambrian explosion. In: Cherchi, A. (Ed.), Proceedings.Sixth Paleobenthos International Symposium. Alghero, 28-30 october1995. Dipartimento di Scienze della Terra, Università di Cagliari, Cagliari,pp. 151–152.lacher Drexler, E., Seilacher, A., 1999. Undertraces of Sea Pens and MoonSnails and possible fossil counterparts. Neues Jahrbuch für Geologie undPaläontologie, Abhandlungen 214, 195–210.ikhatov, M.A., Komar, V.A., Serebryakov, S.N., 1970. The Yudomiancomplex of stratotypical area. Trudi Geologicheskogo Instituta, AkademijaNauk SSSR 210, 1–207 (in Russian).nley, D.C.A., Pickerill, R.K., 1998. Systematic ichnology of the Late Ordo-vician Georgian Bay Formation of southern Ontario, eastern Canada. Roy-al Ontario Museum Life Sciences Contributions 162, 1–55.rell, O.M., 1868. Bidrag till Sparagmitetagens geognosi och paleontologi.Acta Universitatis Lundensis, Lunds Universitets Årsskrift 4, 1–40.rell, O.M., 1870. Petrificata Suecana Formationis Cambricae. Acta Univer-sitatis Lundensis, Lunds Universitets Årsskrift 6, 1–14.n Amerom, H.W.J., Broutin, J., Ferrer, J., Gámez-Vintaned, J.A., Gisbert,J., Liñán, E., 1993. Les flores du Permien basal et la paléoichnologie de lafosse de Fombuena (province de Zaragoza, Espagne). Mededelingen RijksGeologische Dienst 48, 1–63.nuxem, L., 1842. Geology of New York, pt. III, comprising the survey ofthe 3d geological district. W. and A. White and J. Visscher, Albany.lcott, C.D., 1890. Descriptive notes of new genera and species from theLower Cambrian or Olenellus Zone of North America. US National Mu-seum Proceedings 12 (1889), 33–46.bby, B.D., 1970. Late Precambrian trace fossils from New South Wales.Lethaia 3, 79–109.arreño, I., Debrenne, F., 1977. Sédimentologie et biologie des construc-tions organogènes du Cambrien inférieur du Sud de l’Espagne. Mémoiresdu Bureau de Recherches Géologiques et Minières 89, 49–61.




















