
Toxicity assessment of sediments associated with various land-uses in coastal South Carolina, USA,...
10
Toxicity assessment of sediments associated with various land-uses in coastal South Carolina, USA, using a meiobenthic copepod bioassay Adriana C. Bejarano a, * , Keith A. Maruya b , G. Thomas Chandler a a Department of Environmental Health Sciences, Arnold School of Public Health, University of South Carolina, Columbia, SC 29208, USA b Skidaway Institute of Oceanography, Savannah, GA 31411, USA Abstract Coastal urbanization supplies surrounding estuarine environments with urban-related contaminants such as polycyclic aromatic hydrocarbons (PAHs), metals and pesticide mixtures. In our study, adult female and male copepods (Amphiascus tenuiremis) were chronically exposed to 18 sediment samples collected from sites influenced by different land-uses. Sediment samples were collected from three major suburban areas (Hilton Head, Kiawah Island and the Okatee River watershed) and a pristine site (North Inlet) in coastal South Carolina. Three-sediment bioassays (six sites per bioassay) were conducted by culturing copepods for 14 days in quadruplicate test sediments under flow-through conditions at 20 °C and 12:12 LD cycles. Adult survival and copepod reproductive output were quantified. Sediment samples were also analyzed by GC–MS for low and high molecular weight PAHs. Minimal adult mortality was observed in most sediment samples. However, sediments from Hilton Head Island and the Okatee River showed significant effects on copepod reproductive output (i.e., nauplii, copepodites and clutch size). Thus, we determined that reproductive endpoints rather than adult copepod survivorship were more sensitive to effects of contaminated sediments on A. tenuiremis. Furthermore, six (33%) of the 18 sites had a >25% reduction in copepod bioassay endpoints relative to controls, suggesting a high risk to long term A. tenuiremis population maintenance. Ó 2004 Elsevier Ltd. All rights reserved. Keywords: Sediments; Copepod Amphiascus tenuiremis; PAHs; Urbanization; ERMQ; Chronic exposure 1. Introduction The southeastern US seaboard has experienced tre- mendous urban expansion and population growth over the past 40 years. In South Carolina, urban growth, coupled with development of residential neighborhoods near major transportation routes and along estuarine marshes, is 6.2 times faster than population growth (Berkeley–Charleston–Dorchester Council of Govern- ments, (BCDCOG), pers. comm.). The proximity of various land-uses (i.e., shopping centers, parking lots, bridges, roads and golf course communities) to estuarine salt marshes and tidal creeks places enormous pressure on these coastal ecosystems. These areas provide nurs- eries for economically valuable fisheries as well as nest- ing and migratory resting sites for birds, while serving as filters for human-related contaminants. With urbaniza- tion comes an increase in impervious surface coverage resulting in increased storm water runoff, a major source of non-point source pollution to coastal and estuarine environments (Fulton et al., 1993). Urban runoff carries a wide variety of toxicants including polycyclic aromatic hydrocarbons (PAHs), metals and mixtures of pesticides that could cause deleterious effects on benthic fauna. Many studies have shown the usefulness of acute and chronic toxicity bioassays to evaluate the effects of estuarine contaminated sediment on a variety of organisms (McGee et al., 1999; Long, 2000). Toxicity bioassays using marine/estuarine organisms such as the sea urchin (Paracentrotus lividus), heart urchin (Echino- cardium cordatum), oyster (Crassostrea gigas), amphi- pod (Corophium volutator), and bacterium Vibrio fischeri (Geffard et al., 2001; Stronkhorst et al., 2003) * Corresponding author. Tel.: +1-803-777-6452; fax: +1-803-777- 3391. E-mail address: [email protected] (A.C. Bejarano). 0025-326X/$ - see front matter Ó 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2004.01.004 www.elsevier.com/locate/marpolbul Marine Pollution Bulletin 49 (2004) 23–32
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Toxicity assessment of sediments associated with various land-uses in coastal South Carolina, USA,...

www.elsevier.com/locate/marpolbul
Marine Pollution Bulletin 49 (2004) 23–32
Toxicity assessment of sediments associated with variousland-uses in coastal South Carolina, USA,using a meiobenthic copepod bioassay
Adriana C. Bejarano a,*, Keith A. Maruya b, G. Thomas Chandler a
a Department of Environmental Health Sciences, Arnold School of Public Health, University of South Carolina,
Columbia, SC 29208, USAb Skidaway Institute of Oceanography, Savannah, GA 31411, USA
Abstract
Coastal urbanization supplies surrounding estuarine environments with urban-related contaminants such as polycyclic aromatic
hydrocarbons (PAHs), metals and pesticide mixtures. In our study, adult female and male copepods (Amphiascus tenuiremis) were
chronically exposed to 18 sediment samples collected from sites influenced by different land-uses. Sediment samples were collected
from three major suburban areas (Hilton Head, Kiawah Island and the Okatee River watershed) and a pristine site (North Inlet) in
coastal South Carolina. Three-sediment bioassays (six sites per bioassay) were conducted by culturing copepods for 14 days in
quadruplicate test sediments under flow-through conditions at 20 �C and 12:12 LD cycles. Adult survival and copepod reproductive
output were quantified. Sediment samples were also analyzed by GC–MS for low and high molecular weight PAHs. Minimal adult
mortality was observed in most sediment samples. However, sediments from Hilton Head Island and the Okatee River showed
significant effects on copepod reproductive output (i.e., nauplii, copepodites and clutch size). Thus, we determined that reproductive
endpoints rather than adult copepod survivorship were more sensitive to effects of contaminated sediments on A. tenuiremis.
Furthermore, six (33%) of the 18 sites had a >25% reduction in copepod bioassay endpoints relative to controls, suggesting a high
risk to long term A. tenuiremis population maintenance.
� 2004 Elsevier Ltd. All rights reserved.
Keywords: Sediments; Copepod Amphiascus tenuiremis; PAHs; Urbanization; ERMQ; Chronic exposure
1. Introduction
The southeastern US seaboard has experienced tre-
mendous urban expansion and population growth over
the past 40 years. In South Carolina, urban growth,
coupled with development of residential neighborhoods
near major transportation routes and along estuarinemarshes, is 6.2 times faster than population growth
(Berkeley–Charleston–Dorchester Council of Govern-
ments, (BCDCOG), pers. comm.). The proximity of
various land-uses (i.e., shopping centers, parking lots,
bridges, roads and golf course communities) to estuarine
salt marshes and tidal creeks places enormous pressure
on these coastal ecosystems. These areas provide nurs-
*Corresponding author. Tel.: +1-803-777-6452; fax: +1-803-777-
3391.
E-mail address: [email protected] (A.C. Bejarano).
0025-326X/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.marpolbul.2004.01.004
eries for economically valuable fisheries as well as nest-
ing and migratory resting sites for birds, while serving as
filters for human-related contaminants. With urbaniza-
tion comes an increase in impervious surface coverage
resulting in increased storm water runoff, a major source
of non-point source pollution to coastal and estuarine
environments (Fulton et al., 1993). Urban runoff carriesa wide variety of toxicants including polycyclic aromatic
hydrocarbons (PAHs), metals and mixtures of pesticides
that could cause deleterious effects on benthic fauna.
Many studies have shown the usefulness of acute
and chronic toxicity bioassays to evaluate the effects
of estuarine contaminated sediment on a variety of
organisms (McGee et al., 1999; Long, 2000). Toxicity
bioassays using marine/estuarine organisms such as thesea urchin (Paracentrotus lividus), heart urchin (Echino-
cardium cordatum), oyster (Crassostrea gigas), amphi-
pod (Corophium volutator), and bacterium Vibrio
fischeri (Geffard et al., 2001; Stronkhorst et al., 2003)

24 A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32
have been successful in identifying contaminated sed-
iments with potential toxicological effects. This study
was done to assess risks posed by estuarine sediments
influenced by various land-uses to estuarine meioben-thic copepod survival, development and reproduc-
tion.
2. Materials and methods
2.1. Test organisms
The diosaccid harpacticoid copepod Amphiascus
tenuiremis inhabits the top 0.5 cm of muddy inter- and
subtidal estuarine sediments. This cosmopolitan cope-
pod, distributed from the Baltic Sea to the southern
Gulf of Mexico (Lang, 1948), has been mono-specifically
cultured (Chandler, 1986) in contaminant-free flow-
through sediment microcosms at 30 ppt, 23 �C, and a
12 h:12 h Light:Dark cycle. With a rapid generationtime of 21 days at 20 �C (Chandler and Green, 1996),
discrete life-stages, sexually dimorphic adults, and easily
quantifiable reproductive outputs, A. tenuiremis is an
ideal model organism for the assessment of sex specific
survival and reproductive effects (Chandler and Green,
1996).
2.2. Sediment collection and processing
Three 14-day partial life cycle bioassays with A. ten-
uiremis were conducted using salt marsh sediments col-
lected from sites adjacent to several land-uses within
three major suburban areas in coastal South Carolina,
USA. These areas included the relatively new (<10 yr)
suburban developments of the Okatee River watershed,
the somewhat older (<20 yr), less densely populatedKiawah Island resort, and the older (>40 yr) residential
Hilton Head Island area. The collection sites included
retention ponds and creeks within golf course commu-
nities and adjacent to impervious surfaces (i.e., paved
roadways and parking lots). Bioassay 1 was conducted
using sediments from Hilton Head Island (sites HTA,
PD2, PD3; Table 2) and Kiawah Island (sites TCE,
TCW, OR; Table 2), SC, USA. Bioassays 2 and 3 usedsediments from Hilton Head Island (sites HT1, HT2,
HT3, OM T1-P T2-H; Table 2) and the Okatee River
watershed (sites OCB, OCF, T3TG, T3C, HTB, BP;
Table 2). Within each site, 2-cm surface scrapes were
collected from �1 m2 of sediments and composited into
a single homogeneous sample. Composite samples were
placed in clean acetone-rinsed glass jars and stored at 4
�C for a maximum of 30 days until assayed. Within 5days prior to test initiation, ‘‘clean’’ salt marsh control
sediments were collected from a pristine, salt marsh
estuarine preserve little influenced by urbanization in
North Inlet, SC, USA (Sanger et al., 1999).
2.3. 14-day chronic copepod bioassay
Chronic sediment toxicity testing was performed
following procedures described by Chandler and Green(1996). Exposure chambers consisted of 50 ml Teflon
flasks with two 1-cm2 windows on opposite sides cov-
ered with a 63 lm Nitex mesh netting, allowing dripping
water flow through the flask at a rate of 5 ml/h.
Chambers were loaded with 20 ml-filtered (0.45 lm)
aerated artificial seawater (30 ppt, >90% DO satura-
tion), and 10 ml well homogenized and press-sieved
(6 250 lm fraction) sediments (four chambers per site).Chambers were loaded with laboratory cultured
non-gravid female and male copepods (25/each) and
placed in a flow-through system held in an incubator at
20 �C and 12:12 LD cycles. Every third day, a 1:1:1
algae mixture (Dunaliella terticolecta:Isochrysis gal-
bana:Phaeodactylum tricornutum) was added to a con-
centration of �107 cells/chamber (Coulter� Multisizer,
Coulter Electronics Inc., Hialeah, FL, USA). Waterquality (pH, dissolved oxygen, salinity) was monitored
every third day in the flow-through system, and
ammonia (NH3) measured in the exposure chambers at
the end of the bioassay.
After 14 days of culturing, parent copepods and off-
spring were collected by gently rinsing test sediments
through a 63 lm sieve. After checking for live/dead
individuals, samples were stained and fixed with 0.5%Rose-Bengal in 10% buffered formaldehyde, and indi-
viduals counted/staged manually using a microscope.
A total of 10 bioassay endpoints including sex spe-
cific survival, reproductive output (i.e., % gravid
females, clutch size per female, nauplius and copepo-
dites), indices of reproductive success (total offspring
(nauplii + copepodites), and realized production ((nau-
plii + copepodites)/surviving female)) as well as devel-opmental rate (copepodite-to-naupliar ratio) were
obtained for each site.
To assess effects of contaminated sediments on
copepod survival and reproduction, and to allow for
comparisons of among all sites, data from individual
replicates per site were referenced to mean endpoint
values from within bioassay control sediments. Values
are presented as percent change (%) relative to controls.Using this approach, bioassay endpoints were catego-
rized based on the number of test sediments (sites) in
which values of individual bioassay endpoints were re-
duced by >10% or >25% relative to controls. Similarly,
sites were categorized based on the number of bioassay
endpoints in which values were reduced by >10% or
>25% relative to controls. Based on our knowledge of
the natural history of A. tenuiremis, a >25% reduction inadult survival or reproductive output could potentially
influence long-term population growth and mainte-
nance, resulting in rapid population declines (Bejarano
and Chandler, 2003).

A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32 25
2.4. Sediment PAH analysis
Freeze-dried composite sediment samples (10 g) per
site (15 sites total) were homogenized with Hydro-matixTM and extracted with dichloromethane using an
Accelerated Solvent Extraction System (ASE 200, Dio-
nex Corp., Salt Lake City, UT, USA) under elevated
temperature and pressure. After exchanging the solvent
to hexane, each extract was treated with acid-activated
copper to remove elemental sulfur. Extracts were sepa-
rated into fractions of increasing solvent polarity (F1
and F2) using silica gel (2.0 g activated at 130 �C for >4h) packed into a glass column. All fractions were ex-
changed to hexane and reduced to 1.0 ml using a
TurboVap II (Zymark, Hopkinton, MA, USA).
The extracts containing polycyclic aromatic hydro-
carbons (PAH F1 and F2) were analyzed using a HP
6890 digital flow control GC coupled to a 5973 mass
selective detector (GC–MS), operating in the full scan
(50–350 amu at 1 cycles/s) electron ionization mode.Source and transfer line temperatures were 230 and 280
�C, respectively. The injector was programmed to track
the column oven temperature. The GC column oven was
programmed as follows: 60 �C (1 min hold); ramp to 150
�C at 10 �C/min; ramp to 300 �C at 4 �C/min (11 min
hold). Total run time was 60 min. A fused silica column
(30 m (L) · 0.25 mm (i.d.)) coated with 0.25 lm DB-
XLB (Agilent Technologies, Folsom, CA, USA) wasused to separate target PAH analytes.
Twenty-four PAH analytes (2–6 ring structures) were
quantified in all samples, except for the Kiawah Island
sites, based on calibration solutions of an authentic
standard mixture (SRM2260; National Institute of
Standards and Technology (NIST), Gaithersburg, MD,
USA). Target analytes included low molecular weight
(LMW; naphthalene [Naph], 2-methylnaphthalene,1-methylnaphthalene [1mN], biphenyl, 2,6-dimethyl-
naphthalene, acenaphthylene, acenaphthene [Ace-Nap], 2,
3,5-trimethylnaphthalene, fluorine [Fluor], phenanthrene
[Phen], anthracene [Anth] and 1-methylphenanthrene
[1mP]) and high molecular weight (HMW; fluoranthene
[Flu], pyrene [Pyr], benz[a]anthracene [B(a)A], chrysene
[Chry], benzo[b+k]fluoranthene [B(bk)F], benzo[e]py-
rene, benzo[a]pyrene, perylene [Per], indeno[1, 2,3-cd]py-rene, dibenz[a,h]anthracene and benzo[g,h,i]perylene)
PAHs. Total sediment PAH concentrations at the refer-
ence site (North Inlet, NI) have been measured previously
and are consistently low (<116 ng/g dw; Sanger et al.,
1999).
2.5. Sediment effects assessment
Based on the PAH levels at each of the sites, the
general risk of multiple, co-existing PAH exposures was
assessed following suggestions by Long et al., 1995) and
Hyland et al. (1999). For individual PAHs, Long et al.
(1995) published effects guidelines for sediment con-
centrations resulting in adverse effects for 10% (effects
range-low, ERL) and 50% (effects range median, ERM)
of the exposed organisms. For each site PAH-ERL andERM ratios were calculated by dividing PAH concen-
trations by the respective guideline. The ERM quotient
(ERM-Q) was then calculated by dividing the sum of all
ERM ratios per site by the number of analytes.
According to Hyland et al. (1999), the predictive risk of
observing degraded benthic invertebrate communities
is assumed low for ERMQ<0.02, moderate for
ERMQ >0.02)<0.077 and high for ERMQ>0.077.
2.6. Statistical analysis
All endpoints were tested for normality and homo-
geneity of variances using the Shapiro-Wilk’s ‘‘goodness-
of-fit test’’, and the Levene test, respectively. Variables
failing normality were transformed via logarithmic (i.e.,
Log10ðxþ 1Þ, square root ðpðxþ 0:5ÞÞ or angular (i.e.,Arcsine ðpxÞ) transformations where appropriate. Dif-
ferences in adult survival and reproductive output within
bioassays were determined by Analysis of Variance
(ANOVA; PROC GLM, SAS Institute Inc., Cary, NC,
USA). Within bioassays, multiple comparisons between
contaminated sediment samples and control sediments
were done by Dunnett’s procedure. All tests for sig-
nificance were performed using at a ¼ 0:05.The strength of association between bioassay end-
points and low and high molecular weight sediment PAH
(LMW and HMW) concentrations, and ERMQ values
was evaluated using the non-parametric Spearman Rank
correlation analysis (SPEARMAN; PROC FREQ, SAS
Institute Inc., Cary, NC, USA). The p-value of the cor-
relation tests the hypothesis of no association between
variables (correlation¼ 0), with rho ðqÞ as the measure ofcorrelation. This statistical analysis does not demon-
strate cause–effect relationships.
3. Results
3.1. 14-day chronic copepod bioassay
Water quality during the 14-day exposure and at
bioassay termination was consistent across bioassays,
and fell within acceptable limits for chronic sediment
bioassays (ASTM, 1992; Table 1). Dissolved oxygen
saturation (DO%) in the system was >95% during the
14-day exposure for all bioassays, and 70–85% in test
chambers at the end of the exposure.
In sediment samples from sites on Hilton Head andKiawah Island (Bioassay 1), adult survival and gravid
female abundances were not statistically different from
control sediments (ANOVA, p > 0:05; Table 2). How-
ever, reproductive outputs were severely reduced relative

Table 1
Water quality parameters (mean± 1 SD) in bioassay flow-through system during each 14-day chronic bioassay, and in exposure chambers at the end
of each bioassay
Bioassay Flow-through system Experimental chamber
Salinity (ppt) pH DO (%) Salinity pH DO (%) NH3 (lg/l)
1 30.0± 0.7 8.02± 0.08 96.1± 0.3 30.7± 0.5 7.99± 0.05 79.6± 5.0 62.9± 63.4
2 30.0± 0.1 8.09± 0.03 97.8± 0.8 30.6± 0.5 8.05± 0.02 74.7± 2.6 15.7± 19.6
3 30.0± 0.1 8.05± 0.09 95.7± 1.7 30.0± 0.1 7.93± 0.04 75.2± 5.4 20.9± 13.6
(1) Hilton Head and Kiawah Island sediments; (2,3) Hilton Head Island and Okatee River watershed sediments.
Table 2
Bioassay endpoints (mean± 1 SD) (4 replicates per site) from copepod chronic exposures to sediments collected adjacent to various land-uses
Bio-
assay
Site Land-use type Bioassay endpoints
Female
survival (%)
Male survival
(%)
Copepodites Nauplii Realized
offspring
production
Clutch size
1 NI Non-urban creek
(control)
79± 16 79± 15 319± 46 603± 65 48± 9 17± 6
HHI HTA Golf course––creek 82± 7 70± 10 89± 31� 375± 75� 23± 5� 13± 4�
HHI PD2 Golf course––Ret.
pond
82± 9 84± 13 102± 7� 349± 28� 22± 4� 11± 4�
HHI PD3 Suburban––Ret.
pond
80± 12 67± 8 90± 32� 343± 11� 22± 5� 10± 4�
KI TCE Golf course––Ret.
pond
87± 7 74± 3 396± 84 703± 91 50± 2 17± 5
KI TCW Suburban––creek 71± 8 84± 3 226± 47 431± 87� 37± 7 14± 4
KI OR Suburban––creek 73± 13 68± 14 308± 51 646± 59 54± 9 11± 4�
2 NI Non-urban creek
(control)
56± 14 31± 10 85± 27 216± 57 22± 5 13± 4
HHI HT1 Golf course––creek 52± 19 49± 13 72± 7 129± 46� 19± 9 12± 4
HHI HT2 Golf course––creek 40± 7 28± 12 85± 44 165± 24 25± 6 13± 4
HHI HT3 Golf course––creek 60± 9 48± 26 9± 6� 89± 34� 6± 2� 10± 5
ORW OM Outlet Mall––Ret.
pond
6± 2� 10± 7� 3± 2� 21± 12� 15± 6 5± 6�
ORW T1-P Suburban––creek 51± 7 50± 13 36± 18� 148± 40 15± 4 11± 4
ORW T3-H Hospital––Ret.
pond
2± 2� 5± 9� 0± 0� 0± 0� 0± 0� 0± 0�
3 NI Non-urban creek
(control)
81± 7 78± 7 3± 3 151± 23 8±1 8± 2
ORW OCB Highway––creek 78± 10 65± 14 3± 4 9± 6� 1± 0� 7± 1�
ORW OCF Suburban––creek 81± 4 72± 12 2± 1 15± 6� 1± 0� 7± 1�
ORW T3GC Golf course––Ret.
pond
68± 18 58± 14 2± 1 53± 3� 4± 1� 7± 1
ORW T3C Golf course––creek 83± 4 81± 8 6± 3 47± 13� 2± 1� 8± 3
HHI HTB Golf course––creek 78± 8 67± 19 12± 5� 327± 31� 17± 2� 11± 3�
B BP Golf course––creek 73± 11 72± 19 21± 7� 205± 7� 13± 3� 7± 2
(�) Represents significant difference from North Inlet sediments (NI, control sediments; a ¼ 0:05). HHI: Hilton Head Island, KI: Kiawah Island,
ORW: Okatee River watershed and B: City of Beaufort. Starting male and non-gravid female copepod abundances in each test chamber were 25
each.
26 A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32
to controls. For example, copepodite production was
reduced by 55–85% for all Hilton Head sites (HTA,
PD2, PD3) (176–271 copepodites; ANOVA, p <0:0001), while nauplii production was reduced by 27–56% (163–338 nauplii; ANOVA, p < 0:001) relative to
controls (including the Kiawah Island (TCW) site).
Except for sites TCE and TCW on Kiawah Island,
clutch size of surviving females was reduced by 12–54%
(2–9 embryos; ANOVA, p < 0:002). Consequently, real-ized offspring production in all Hilton Head samples
was 38–66% (ANOVA, p < 0:0002) lower than controls.
Moreover, copepodite-to-naupliar ratios were 21–72%(ANOVA, p < 0:008) lower than controls, indicating
either a significant reduction in naupliar production or
a reduction in numbers of nauplii successfully develop-
ing into copepodites.

Bioassay endpoints
%GF %F %M F/M C N C/N TO RO CS
Num
ber o
f tes
t sed
imen
ts
0
3
6
9
12
15
18 >10%>25%
50% test sediments
Fig. 1. Number of test sediments ðn ¼ 18Þ, in which bioassay endpoints
were reduced by >10% or >25% relative to controls. %GF¼% Gravid
female; %F¼% surviving females; %M¼% surviving males;
F/M¼ surviving female-to-male ratio; C¼ copepodites; N¼ nauplii;
C/N¼Copepodite-to-naupliar ratio; TO¼ total offspring; RO¼ rea-
lize offspring; and CS¼ clutch size.
A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32 27
In Bioassay 2 we tested sediments from Hilton Head
Island and the upper Okatee River watershed near
Bluffton. Adult copepod survival was consistently <30%
in OM and T3-H sediments (<15 adult copepods; AN-OVA, p < 0:0001) relative to controls. Adult copepods
exposed to T3-H sediments showed 100% reproduc-
tive failure. Compared to controls, sites OM, T1-P and
T3-H showed a reduction in copepodite production
>40% (>26 copepodites; ANOVA, p < 0:0001). Simi-
larly, most sites (except for HT1 and T1-P) showed a
reduction in naupliar production >40% (>87 nauplii;
ANOVA, p < 0:001). Despite the depressed naupliarproduction, realized offspring production was reduced
by >70% only for two sites (HT3 and T3-H), while co-
pepodite-to-naupliar ratios were reduced by >40% in
three sites (HT3, OM and T3-H; ANOVA, p < 0:0004).Bioassay 3 evaluated sediments from the Okatee
River watershed (OCB, OCF, T3GC, T3C), Hilton
Head (HTB) and the City of Beaufort retention pond
(BP). As with Bioassay 1, adult survival and incidence ofgravid females were not statistically different from con-
trols for any sample (p > 0:05; Table 2). Although
copepod production was unusually low in control sedi-
ments, in all Okatee River sites (OCB, OCF, T3TG and
T3C) nauplii and realized production were reduced by
>60% (>90 nauplii) and >68%, respectively. HTB and
BP showed significantly higher reproductive output than
those of controls (ANOVA, p ¼ 0:001). Due to lowcopepodite counts in controls, copepodite-to-naupliar
ratios in contaminated sediments were not statistically
different from controls (ANOVA, p > 0:05).In order to determine (1) how bioassay endpoints were
influenced by sediments from sites adjacent to several
land-uses; and (2) which sites posed the greatest risk to
A. tenuiremis, we pooled all of the bioassay results by
calculating changes in endpoints relative to controlsediment responses within bioassays. Specifically, the
number sites in which values of individual bioassay
endpoints were reduced by >10% or >25% relative to
control sediments (Fig. 1), were tallied, as were the
number of bioassay endpoints per site reduced by >10%
or >25% relative to within bioassay controls (Fig. 2).
Sites with >10% or >25% reduction in bioassay end-
points were categorized as of moderate or high risk to A.
tenuiremis, respectively.
Using this approach, male survival was reduced by
>10% relative to controls in 50% (9 of 18) of test sedi-
ments (Fig. 1). Copepodite and naupliar production,
and clutch size were reduced by >10% in 75% (12 of 18),
88% (14 of 18) and 75% (12 of 18), respectively; while
copepodite and naupliar production were reduced by
>25% in 50% (9 of 18) and 75% (12 of 18) of the sedi-ment tested, respectively. Total and realized production
were reduced by >10% in 88% (14 of 18) and 81% (13 of
18) of test sediments, and >25% in 75% (12 of 18) and
69% (11 of 18) of test sediments, respectively. Thus it is
apparent that reproductive endpoints to A. tenuiremis
(clutch size, naupliar and copepodite production), rather
than adult survivorship, are more sensitive indicatorsof chronic effects associated with exposure to these
sediments.
Similarly, 11 of the 18 sediments (HTA, PD2, PD3,
TCW, HT3, OM, T1-P, T3-H, OCB, OCF, T3GC)
showed a >10% reduction in more than half of the
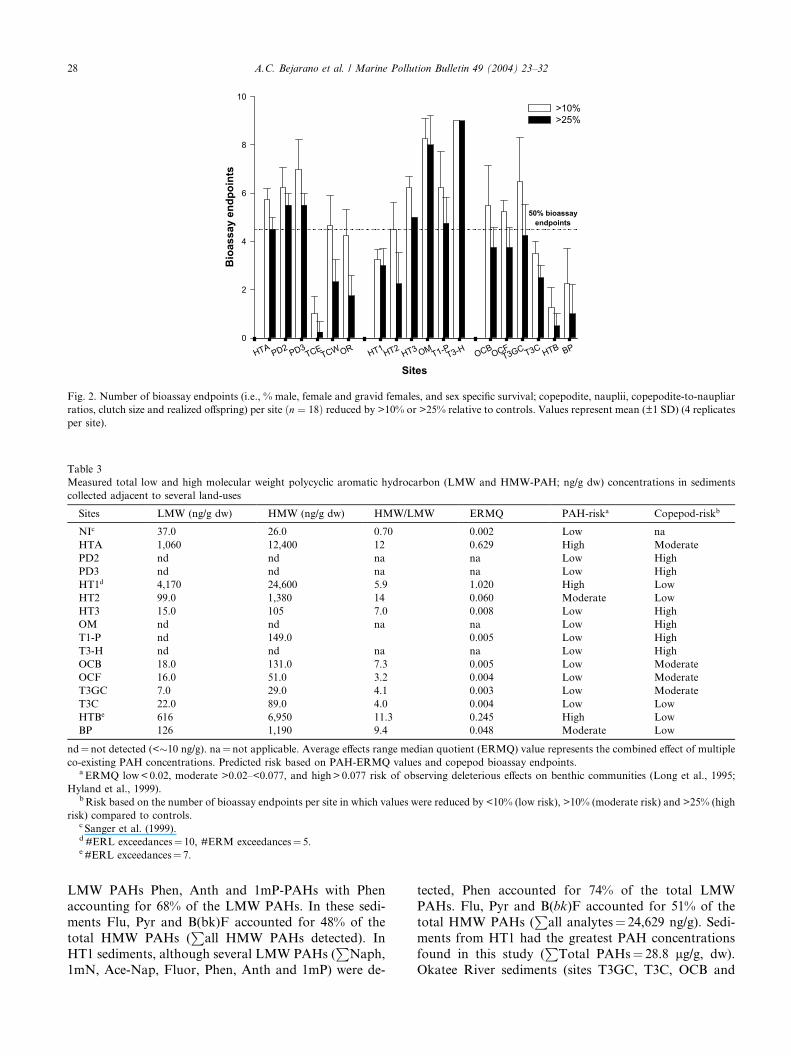
bioassay endpoints (Fig. 2) relative to controls. Six sites
(PD2, PD3, HT3, OM, T1-P, T3-H) showed a >25%
reduction in the same bioassay endpoints (Fig. 2). Theseresults indicate that 39%, 28% and 33% of the sites
respectively may pose a low, moderate or high risk,
to A. tenuiremis-like meiobenthic copepod populations
(Table 3).
3.2. Sediment chemistry
Sanger et al. (1999) reported a total PAH concen-tration (
P23 PAHs) of 116 ng/g (dw) for North Inlet
sediments (NI, control site). Normalization of the data
from Sanger et al. (1999) results in an estimated 37 ng/g
(dw) for low molecular weight PAHs (LMW; 2–3 rings)
and 26 ng/g (dw) for high molecular weight PAHs
(HMW; 4–6 rings; Table 3).
Sediment samples from PD2, PD3, OM and T3-H
had non-detectable levels of either LMW or HMWPAHs. In site HTA (Bioassay 1) Phen accounted for
100% of the LMW PAHs, while Flu, Pyr, and B(bk)F
accounted for �50% of the total HMW PAHs. Hilton
Head sites HT3, HT2, HT1 (Bioassay 2) had detectable
LMW and HMW PAHs. In HT3 sediments Phen ac-
counted for 100% of the LMW PAHs, while the only
HMW PAH detected wereP
Flu, Pyr, B(a)A, Chry,
B(bk)F. HT2 sediments had detectable levels of the
SitesHTA PD2 PD3
TCETCW OR HT1HT2 HT3 OMT1-PT3-H OCBOCFT3GC T3C HTB BP
Bio
assa
y en
dpoi
nts
0
2
4
6
8
10>10%>25%
50% bioassayendpoints
Fig. 2. Number of bioassay endpoints (i.e., % male, female and gravid females, and sex specific survival; copepodite, nauplii, copepodite-to-naupliar
ratios, clutch size and realized offspring) per site ðn ¼ 18Þ reduced by >10% or >25% relative to controls. Values represent mean (±1 SD) (4 replicates
per site).
Table 3
Measured total low and high molecular weight polycyclic aromatic hydrocarbon (LMW and HMW-PAH; ng/g dw) concentrations in sediments
collected adjacent to several land-uses
Sites LMW (ng/g dw) HMW (ng/g dw) HMW/LMW ERMQ PAH-riska Copepod-riskb
NIc 37.0 26.0 0.70 0.002 Low na
HTA 1,060 12,400 12 0.629 High Moderate
PD2 nd nd na na Low High
PD3 nd nd na na Low High
HT1d 4,170 24,600 5.9 1.020 High Low
HT2 99.0 1,380 14 0.060 Moderate Low
HT3 15.0 105 7.0 0.008 Low High
OM nd nd na na Low High
T1-P nd 149.0 0.005 Low High
T3-H nd nd na na Low High
OCB 18.0 131.0 7.3 0.005 Low Moderate
OCF 16.0 51.0 3.2 0.004 Low Moderate
T3GC 7.0 29.0 4.1 0.003 Low Moderate
T3C 22.0 89.0 4.0 0.004 Low Low
HTBe 616 6,950 11.3 0.245 High Low
BP 126 1,190 9.4 0.048 Moderate Low
nd¼ not detected (<�10 ng/g). na¼not applicable. Average effects range median quotient (ERMQ) value represents the combined effect of multiple
co-existing PAH concentrations. Predicted risk based on PAH-ERMQ values and copepod bioassay endpoints.a ERMQ low<0.02, moderate >0.02–<0.077, and high> 0.077 risk of observing deleterious effects on benthic communities (Long et al., 1995;
Hyland et al., 1999).bRisk based on the number of bioassay endpoints per site in which values were reduced by <10% (low risk), >10% (moderate risk) and >25% (high
risk) compared to controls.c Sanger et al. (1999).d #ERL exceedances¼ 10, #ERM exceedances¼ 5.e #ERL exceedances¼ 7.
28 A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32
LMW PAHs Phen, Anth and 1mP-PAHs with Phen
accounting for 68% of the LMW PAHs. In these sedi-
ments Flu, Pyr and B(bk)F accounted for 48% of the
total HMW PAHs (P
all HMW PAHs detected). In
HT1 sediments, although several LMW PAHs (P
Naph,
1mN, Ace-Nap, Fluor, Phen, Anth and 1mP) were de-
tected, Phen accounted for 74% of the total LMW
PAHs. Flu, Pyr and B(bk)F accounted for 51% of the
total HMW PAHs (P
all analytes¼ 24,629 ng/g). Sedi-
ments from HT1 had the greatest PAH concentrations
found in this study (P
Total PAHs¼ 28.8 lg/g, dw).
Okatee River sediments (sites T3GC, T3C, OCB and

Table 4
Spearman rank correlation ðqÞ of the bioassay endpoints versus total low and high molecular weight polycyclic aromatic hydrocarbons (LMW and
HMW-PAHs, ng/g dw), and average effects range median quotient (ERMQ) values ðn ¼ 72ÞBioassay endpoints LMW HMW ERMQ
% gravid females 0.36 (0.002) 0.18 (>0.05) 0.21 (>0.05)
% female survival 0.001 (>0.05) 0.03 (>0.05) 0.06 (>0.05)
% male survival )0.02 (>0.05) 0.02 (>0.05) 0.06 (>0.05)
Female-to-male ratio )0.01 (>0.05) )0.1 (>0.05) 0.13 (>0.05)
Copepodites 0.43 (0.0002) 0.1 (>0.05) 0.14 (>0.05)
Nauplii 0.48 (<0.0001) 0.08 (>0.05) 0.13 (>0.05)
Copepodite-to-nauplii ratios 0.3 (0.01) 0.15 (>0.05) 0.18 (>0.05)
Clutch size 0.37 (0.002) 0.27 (0.02) 0.35 (0.003)
Parenthesis represents the p-value of the correlation.
A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32 29
OCF; Bioassay 3) contained <22 ng/g LMW PAHs with
Phen accounting for 37–100%. HMW PAH ranged from
as low as 29 ng/g (T3GC) to 131 ng/g (OCB), with Flu,
Pyr, B(it bk)F and Per accounting for 63–80%. In con-
trast, BP and HTB had elevated LMW PAHs with Phen
accounting for 56% and 74% of the total LMW PAHs,
respectively. Although all HMW PAHs were detectedin these two sites, Flu, Pyr, Chry and B(bk)F accounted
for 54% and 57% of the HMW PAHs, respectively.
The distribution of PAH in environmental samples
may affect the toxic potential and thus pose risk to
benthic estuarine infauna. For example, the mutage-
nicity/carcinogenicity of HMW compounds such as
benzo[a]pyrene are well documented. Conversely, the
increased bioavailability via the aqueous phase of LMWPAH such as naphthalene and phenanthrene may pose a
greater risk due to narcosis. In contrast to the control
(North Inlet) sediment, all test sediments were enriched
with HMW PAH (Table 3). This distribution is consis-
tent with sediments from other coastal urban estuaries
(Kucklick et al., 1997; Kim et al., 1999) containing
weathered combustion or petroleum source PAH (Prahl
and Carpenter, 1983).Based on measured PAH concentrations and fol-
lowing the criteria developed by Hyland et al. (1999), the
estimated ERMQ (Table 3) indicates that sediments
from HTA, HT1 and HTB, may pose a high risk
(ERMQ >0.077) to associated sediment communities.
Similarly, sediments from HT2 and BP with ERMQ
values ranging >0.02 to <0.077, indicates that these
sediments may pose a moderate risk to associated sedi-ment communities. Sediments from the remaining 11
sites have ERMQ levels <0.02, indicating a low risk to
benthic assemblages. No apparent relationship between
HMW/LMW PAH and copepod rise were observed.
The results from the Spearman’s rank correlation
analysis for all reproductive output variables versus
both total LMW and HMW PAHs, and ERMQ values
are presented in Table 4. None of the endpoints (exceptclutch size) show a correlation with either HMW PAHs
or ERMQ values ðp > 0:05Þ. Surprisingly, percent gra-vid females, copepodites, nauplii, and copepodite-to-
naupliar ratios were all positively correlated with LMW
PAHs ðp < 0:05Þ. Clutch size was the only endpoint
positively correlated with both LMW and HMW PAHs,
and ERMQ values ðp < 0:05Þ.
4. Discussion
Concentrations and composition of PAHs in coastal
environments are reflective of land-use. Highly alkylated
and low molecular weight PAHs (LMW, naphthalenes)
dominate suburban and forested watersheds (Kucklick
et al., 1997; Sanger et al., 1999), while pyrogenic high
molecular weight PAHs (HMW, 4–6 rings) dominate
urban runoff (Kucklick et al., 1997; Menzie et al., 2002).
In our sediments, phenanthrene [Phen] was the mostcommonly detected LMW PAH, while fluoranthene
[Flu], pyrene [Pyr], chrysene [Chry], benzo [b+ k], fluo-
ranthene [B(bk)F] and perylene [Per] were the most
abundant among HMW PAHs.
Comparing measured total PAHs concentrations of
the current study with those reported by Sanger et al.
(1999) for South Carolina tidal creeks and salt marshes,
the majority of sites fell within PAH concentrationsreported for forested/reference sites. Sites with moderate
(HT2, BP) to high (HTA, HT1, HTB) total PAH con-
centrations were similar to coastal South Carolina’s
suburban and industrial/urban areas, respectively. Only
three of these sites (HTA, HT1, HTB) had total PAH
concentrations exceeding the highest concentration
(12,300 ng/g dw) found in estuarine sediments along the
Carolinian Province (Hyland et al., 1998). The pre-dominance of Phen, Flu, Pyr, Chry and B(bk)F is con-
sistent with the PAH combustion product signature
found in runoff water from Murrells Inlet, SC, USA
(Ngabe et al., 2000) and in sediment samples from
Charleston Harbor, SC (Kucklick et al., 1997).
The partial life cycle bioassay with the copepod A.
tenuiremis indicated that reproductive endpoints were
the most sensitive indicators of chronic effects followingexposure to sediments associated with coastal land-uses.
Reproductive outputs were depressed in 11 sediment
sample sites, with the most dramatic effects observed in
sediments from Hilton Head (HTA, PD2, PD3 and

30 A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32
HT3) and the Okatee River watershed (OM, T3-H, OCB
and OCF). Sites from Kiawah Island (TCE, TCW, OR)
did not cause any reproductive effects to A. tenuiremis.
Furthermore, based on all the bioassay endpoints, 33%of the sites (PD2, PD3, HT3, OM, T1-P and T3-H) may
pose a high risk to long term A. tenuiremis population
maintenance. In this study, we found a general lack of
agreement between the chronic toxicity of PAH con-
taminated sediments to A. tenuiremis and the cumulative
risk estimate based on effects range median quotient
(ERMQ) values for PAHs (Long et al., 1995 and Hyland
et al., 1999). Sediments from PD2, PD3, OM, HT3 andT3-H were found to pose a high risk to A. tenuiremis, yet
ERMQ for these sites place them in a low risk category.
Likewise, the ERMQ categorized HTA, HT1, and HTB
sediments as high potential risks to benthic fauna; yet
copepod bioassay results placed two of these sediments
(HT1, HTB) in low, and one (HTA) in the moderate risk
category. Furthermore a few sediment samples, with
PAH concentrations near or below instrument detectionlimits (PD2, PD3, OM and T3-H), and with ERMQ
values indicating low risk (the above plus HT3, T1-P,
T3-H, OCB and OCF), in some cases produced sig-
nificant effects on reproductive output. These results
suggest that observed deleterious effects on copepod
reproduction could not predicted by the ERMQ ap-
proach.
LMW PAHs are indicative of non-anthropogenicsources (Kucklick et al., 1997) and/or relatively fresh
petroleum contamination (Prahl and Carpenter, 1983).
LMW PAHs are also generally considered less toxic
than HMW PAHs (Eisler, 1987). Interestingly, a strong
positive correlation was seen between LMW PAHs and
percent gravid, copepodites, nauplii, and the develop-
mental rate (i.e., copepodite-to-naupliar ratios). Clutch
size was also positively correlated with LMW as well asHMW PAHs, which is consistent with a previous study
that found larger clutch sizes in female A. tenuiremis
chronically exposed to contaminated sediments (Fulton
et al., 1996). An increase in clutch size might be a
compensatory mechanism by which females reduce
PAH bioavailability. Another study with A. tenuiremis
found that female copepods chronically exposed to
chrysene had an increased yolk (lipovitellin) depositionto eggs relative to control females (Chandler and Volz,
in press). Also, female grass shrimp Paleomonetes pugio
exposed to pyrene (63 ppb) showed elevated vitellin
production, as well as increased embryo mortality
(Oberd€orster et al., 2000a). Thus, enhanced yolk pro-
duction, and potentially increased clutch size, may serve
as a mechanism for depuration of highly lipophilic
compounds such as PAHs, rendering these compoundsless biologically available to females (Oberd€orster et al.,2000b; Chandler and Volz, in press), but more available
to offspring. An alternative explanation for this positive
association is a mild bio-stimulatory effect due to low
level petroleum contamination, which in turn may
nourish higher trophic levels of the tidal creek benthic
food web.
The absence of a strong statistical correlation be-tween sediment PAHs (LMW, HMW, ERMQ) and
toxicity may be related to the presence of other
unmeasured anthropogenic contaminants (i.e., pesti-
cides, heavy metals) or unaccounted for physical (i.e.,
grain size) and chemical (i.e., organic matter content and
quality) sediment properties, which may have influenced
the nutritional status of test copepods as well as the
bioavailability of PAHs. A similar study (McGee et al.,1999) using PAH-laden field sediments found no sig-
nificant Spearman correlation between PAH concen-
trations and amphipod survival (10-day sediment
bioassay) and field densities. In this study, however,
amphipod survival was correlated with PAH ERMQ
values.
Despite a lack of inverse correlation between copepod
and amphipod bioassay endpoints and PAH concen-trations, PAHs can cause adverse effects on sediment
infauna. Although most PAHs exert narcotic toxicity
at environmental concentrations, the toxicity of PAHs
such as fluoranthene, pyrene, benzo[a]anthracene, anth-
racene and benzo(a)pyrene (all of which were detected
in some our sediment samples) can be exacerbated by
ultraviolet (UV)-photoactivation (Kosian et al., 1998;
Newsted and Giesy, 1987; Ankley et al., 1997). Bioas-says in this study were conducted under cool-fluorescent
lighting, so UV impacts would not be expected.
Other studies have shown the potential interaction of
PAHs with the arthropod ecdysteroid hormone system
(Oberd€orster et al., 1999, 2000b). Oberd€orster et al.
(1999) showed that PAHs enhance the in-vitro effects of
the hormone 20-Hydroxyecdysone (20HE; responsible
for crustacean molting, development and reproduction)on ecdysteroid-responsive imaginal disc cell lines by
reducing cell proliferation. Furthermore, PAHs could
interfere with the metabolism of ecysteroids in molting
crustaceans (Oberd€orster et al., 2000a,b). In the present
study, A. tenuiremis cultured in field contaminated sed-
iments from Hilton Head (HT1, HT3) and the Okatee
River (OCB, OCF) showed reduced titers of 20HE rel-
ative to controls (Bejarano, unpublished). For example,in late stage embryos microdissected from females ex-
posed to sediments from HT1 and HT3, 20HE titers
were 65% lower (11.7 ± 2.5 and 16.3 ± 8.4 fmol/embryo,
respectively, n ¼ 7) than 20HE in embryos from con-
trols (35.2 ± 7.8 fmol/embryo; n ¼ 7). Similarly, 20HE
titers in males exposed to sediments from OCB and
OCF were �60% lower (9 ± 2.2 and 9.1 ± 1.8 fmol/male,
respectively, n ¼ 10) compared to males exposed tocontrol sediments (24.9 ± 7.5 fmol/male, n ¼ 10). Re-
duced 20HE titers in HT1 and HT3 embryos might be
related to depressed naupliar production and develop-
mental rates in these sediments, since in this species late

A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32 31
stage embryos have elevated 20HE titers prior to
hatching into the naupliar stage (Block et al., 2003).
Conversely, the relationship between reduced 20HE in
males exposed to OCB and OCF sediments and repro-ductive output is less straightforward. As suggested in a
previous study, elevated 20HE in male A. tenuiremis
may be associated with spermatogenesis (Block et al.,
2003). In other arthropods (i.e., ticks) ecdysteroids have
been found to stimulate spermatocyte-DNA synthesis
during early spermatogenesis (Oliver and Dotson, 1993).
However, the link between 20HE titers and offspring
production under exposure to contaminants such asPAHs is currently not well understood for any crusta-
cean.
This study did not elucidate relationships among
sediment PAH concentrations and distributions, and
reproductive and developmental responses in A. tenui-
remis. Thus, determining the potential risk to indigenous
copepod populations inhabiting areas adjacent to vari-
ous land-use coastal environments is more complexthan considering PAH alone as a major pollutant.
Further research should focus on the analysis of multi-
ple anthropogenic-related contaminants (PAHs, heavy
metals, legacy and newly registered pesticides, PCBs),
and the addition of endocrine/hormonal endpoints in
toxicological models to better assess risks of contami-
nated sediments to estuarine meiobenthos.
Acknowledgements
The authors would like to thank T.L. Cary, S. Klo-
sterhaus and J.J. Rafalowski for technical assistance
with bioassay setup and sediment analyses, and two
anonymous reviewers for their useful comments on the
manuscript. This research was funded by the SouthAtlantic Bight Land Use-Coastal Ecosystem Study (LU-
CES) under an award from the Coastal Ocean Pro-
gram (COP), National Ocean Service (NOS), National
Oceanic and Atmospheric Administration (NOAA),
through the South Carolina Sea Grant Consortium,
Grant number NA960PO113. This publication does not
constitute an endorsement of any commercial product
or intend to be an opinion beyond scientific or otherresults obtained by NOAA. This research has not been
subject to either agencies’ peer or policy review, and
no endorsement should be inferred.
References
Ankley, G.T., Erickson, R.J., Sheedy, B.R., Kosian, P.A., Mattson,
V.R., Cox, J.S., 1997. Evaluation of models for predicting the
phototoxic potency of polycyclic aromatic hydrocarbons. Aquatic
Toxicology 37, 37–50.
ASTM, 1992. Standard guide for conducting 10-day static sediment
toxicity tests with marine and estuarine amphipods. In: ASTM,
1993, Annual Book of Standards, 11.04, E1367-92. American
Society for Testing and Materials, Philadelphia, PA, USA.
Bejarano, A.C., Chandler, G.T., 2003. Reproductive and developmen-
tal effects of atrazine on the estuarine meiobenthic copepod
Amphiascus tenuiremis. Environmental Toxicology and Chemistry
22, 3009–3016.
Block, D.S., Bejarano, A.C., Chandler, G.T., 2003. Ecdysteroid
concentrations through various life-stages of the meiobenthic
harpacticoid copepod, Amphiascus tenuiremis and the benthic
estuarine amphipod, Leptocheirus plumulosus. General and Com-
parative Endocrinology 132, 151–160.
Chandler, G.T., 1986. High density culture of meiobenthic herpacti-
coid copepods within a muddy sediment substrate. Canadian
Journal of Fisheries and Aquatic Sciences 43, 53–59.
Chandler, G.T., Green, A.S., 1996. A 14-day harpacticoid copepod
reproduction bioassay for laboratory and field contaminated
muddy sediments. In: Ostrander, G.K. (Ed.), Techniques in
Aquatic Toxicology. pp. 23–39.
Chandler, G.T., Volz, D.C. Semi-quantitative confocal laser scanning
microscopy applied to marine invertebrate ecotoxicology. Marine
Biotechnology, in press.
Eisler, R., 1987. Polycyclic aromatic hydrocarbon hazards to fish,
wildlife, and invertebrates: a synoptic review. US Fish Wild Life
Service. Biological Report, 85(1.11), 81 p.
Fulton, M.H., Scott, G.I., Fortner, A., Bidleman, T.F., Ngabe, B.,
1993. The effects of urbanization on small high salinity estuaries
of the Southeastern United states. Archives of Environmental
Contamination and Toxicology 25, 476–484.
Fulton, M.H., Chandler, G.T., Scott, G.I., 1996. Urbanization effects
on the fauna of a Southeastern USA. Bar-Built Estuary. In:
Vernberg, F.J., Vernberg, W.B. (Eds.), Sustainable Development in
the Southeastern Coastal Zone, vol. 20. Belle W. Baruch Library in
Marine Science, University of South Carolina Press, pp. 477–504.
Geffard, O., Budzinski, H., Augagneur, S., Seaman, M., His, E., 2001.
Assessment of sediment contamination by spermiotoxicity and
embryotoxicity bioassay with sea urchins (Paracentrotus lividus)
and oysters (Crassostrea gigas). Environmental Toxicology and
Chemistry 20 (7), 1605–1611.
Hyland, J.L., Balthis, L., Hackney, C.T., McRae, G., Ringwood,
A.H., Snoots, T.R., Van Dolah, R.F., Wade, T.L., 1998.
Environmental quality of estuaries of the Carolinian Province:
1995 Annual Statistical Summary for the 1995 EMAP-Estuaries
Demonstration Project in the Carolinian Province. NOAA Tech-
nical Memorandum NOS ORCA 123, National Ocean Service
Office of Ocean Resources Conservation and Assessment Coastal
Monitoring and Bio-effects Assessment Division. Charleston, SC,
p. 164.
Hyland, J.L., Van Dolah, R.F., Snoots, T.R., 1999. Predicting stress in
benthic communities of southeastern US estuaries in relation to
chemical contamination of sediments. Environmental Toxicology
and Chemistry 18, 2557–2564.
Kim, G.B., Maruya, K.A., Lee, R.F., Kang, S.G., Koh, C.H., Tanabe,
S., 1999. Distribution and sources of polycyclic aromatic hydro-
carbons from Kyeonggi Bay, Korea. Marine Pollution Bulletin 38,
7–15.
Kosian, P.A., Makynen, E.A., Monson, P.D., Mount, D.R., Spacie,
A., Mekenyan, O.G., Ankley, G.T., 1998. Application of toxicity-
based fractionation techniques and structure–activity relationship
models for the identification of phototoxic polycyclic aromatic
hydrocarbons in sediment pore water. Environmental Toxicology
and Chemistry 17, 1021–1033.
Kucklick, J.R., Sivertsen, S.K., Sanders, M., Scott, G.I., 1997. Factors
influencing polycyclic aromatic hydrocarbon distributions in South
Carolina estuarine sediments. Journal of Experimental Marine
Biology and Ecology 213, 13–29.
Lang, K., 1948. Monographie der Harpacticiden. Nordiska-Bokhen-
deln, Stockholm, Sweden.

32 A.C. Bejarano et al. / Marine Pollution Bulletin 49 (2004) 23–32
Long, E.R., 2000. Degraded sediment quality in US estuaries: a review
of magnitude and ecological implications. Ecological applications
10 (2), 338–349.
Long, E.R., MacDonald, D.D., Smith, S.L., Calder, F.D., 1995.
Incidence of adverse biological effects within ranges of chemical
concentrations in marine and estuarine sediments. Environmental
Management 19, 81–97.
McGee, B.L., Fisher, D.J., Yonkos, L.T., Ziegler, G.P., Turley, S.,
1999. Assessment of sediment contamination, acute toxicity, and
population viability of the estuarine amphipod Leptocheirus
plumulosus in Baltimore Harbor, Maryland, USA. Environmental
Toxicology and Chemistry 18, 2151–2160.
Menzie, C.A., Hoeppner, S.S., Cura, J.J., Freshman, J.S., LaFrey,
E.N., 2002. Urban and suburban storm water runoff as a source of
polycyclic aromatic hydrocarbons (PAHs) to Massachusetts estu-
arine and coastal environments. Estuaries 25, 165–176.
Newsted, J.L., Giesy, J.P., 1987. Predictive models for photoinduced
acute toxicity of polycyclic aromatic. Environmental Toxicology
and Chemistry 6, 445–461.
Ngabe, B., Bidleman, T.F., Scott, G.I., 2000. Polycyclic aromatic
hydrocarbons in storm runoff from urban and coastal South
Carolina. The Science of the Total Environment 255, 1–9.
Oberd€orster, E., Cottam, D.M., Wilmot, F.A., Milner, M.J., McLach-
lan, J.A., 1999. Interaction of PAHs and PCBs with ecdysone-
dependent gene expression and cell proliferation. Toxicology and
Applied Pharmacology 160, 101–108.
Oberd€orster, E., Rice, C.D., Irwin, L.K., 2000a. Purification of vitellin
from grass shrimp Palaemonetes pugio, generation of monoclonal
antibodies, and validation for the detection of lipovitellin in Crusta-
cea. Comparative Biochemistry and Physiology Part C 127, 199–207.
Oberd€orster, E., Brouwer, M., Hoexum-Brouwer, T., Manning, S.,
McLachlan, J.A., 2000b. Long-term pyrene exposure of grass
shrimp, Palaemonetes pugio, affects molting and reproduction of
males and offspring of exposed females. Environmental Health
Perspectives 108, 641–646.
Oliver, J.H., Dotson, E.M., 1993. Hormonal-control of molting and
reproduction in ticks. American Zoologist 33, 384–396.
Prahl, F.G., Carpenter, R., 1983. Polycyclic aromatic hydrocarbon
(PAH) phase associations in Washington coastal sediment. Geo-
chimica Cosmochimica Acta 47, 1013–1023.
Sanger, D.M., Holland, A.F., Scott, G.I., 1999. Tidal creek and salt
marsh sediments in South Carolina coastal estuaries. II. Distribu-
tion of organic contaminants. Archives of Environmental Con-
tamination and Toxicology 37, 458–471.
Stronkhorst, J., Schipper, C., Brils, J., Dubbeldam, M., Postma, J.,
van de Hoeven, N., 2003. Using marine bioassays to classify the
toxicity of Dutch Harbor sediments. Environmental Toxicology
and Chemistry 22, 1535–1547.




















