
Towards a revision of Candoninae (Crustacea: Ostracoda): Australian representatives of the...
18
Subterranean Biology 2: 91-108, 2004 INTRODUCTION The genus Candonopsis was established at the end of the 19th century by Vávra (1891) to include the spe- cies Candona kingsleii, described earlier from south- east England (Brady and Robertson 1870). At that time Candonopsis and Candona Baird, 1845 were the only two genera in the subfamily Candoninae. The original generic diagnosis of the genus Candona was so broad (see Baird 1845) that it continued for many years to ac- cumulate many phylogenetically very distinct species. On the other hand, the diagnosis of the genus Cando- nopsis given by Vávra (1891) was much narrower and it allowed inclusion of only very closely related species. However, due to the further enrichment of our knowl- edge of ostracods and the discovery of new species, the generic diagnosis of the genus Candonopsis has been revised twice. Danielopol (1980) added characters of the cleaning leg, hemipenis and Zenkerʼs organ, while recently Karanovic & Marmonier (2002) gave more details of the soft parts morphology, and broadened the diagnosis substantially. Klie (1932) divided the genus Candonopsis into three groups of species based upon the presence of spines on the furcal claws. However, this division was shown to be unsustainable after the discov- ery of many transitional forms (Karanovic and Marmo- nier 2002). At present 25 recent species are recognized in this genus. A key to their identification was provided by Karanovic and Marmonier (2002). The majority of the Candonopsis species (19) are distributed in the Southern Hemisphere, with 12 sur- face and seven subterranean species. Of the six species known from Europe and North America, two are surface and four are subterranean inhabitants. Continents where the Candonopsis fauna is the most diverse are Africa with eight species (Vávra 1895; Daday 1910, 1913; Klie 1944; Rome 1962; Löffler 1968), and Australia with six species (Brady 1886; Karanovic and Marmonier 2002). Only one species is known so far from the Indian sub- continent (Gupta 1988) and three others from Sumatra, Java and the Solomon Islands, respectively (Klie 1932; Harding 1962). Until recently, four species were known from Central and South America (Vávra 1898; Daday 1905; Méhes 1913; Broodbakker 1983), but Karanovic and Marmonier (2002) proposed the exclusion of three species, retaining only Candonopsis hummelincki Broodbakker, 1983 as a valid representative of the ge- nus. The three excluded species are more closely related to the genera Cubacandona Broodbakker 1983 and Car- ibecandona Broodbakker, 1983, both known only from Central and South America (Broodbakker, 1983). The search for the type material of these species has been unsuccessful and a proper redescription cannot be given; I therefore only tentatively assign them to the genus Cu- bacandona. However, one should not exclude the pos- sibility that they may represent a new genus, or at least some of them. The similarity of the soft parts of Cando- nopsis, Cubacandona and Caribecandona is prominent Towards a revision of Candoninae (Crustacea, Ostracoda): on the genus Candonopsis Vávra, with descriptions of new taxa Ivana KARANOVIC Western Australian Museum, Francis Street, Perth 6000, W. A. Australia ABSTRACT Candonopsini trib. nov. is described in the present paper to encompass Candonopsis Vávra, 1891 Cubacandona Broodbakker, 1983 and Caribecandona Broodbakker, 1983. The new tribe stands apart from the rest of Candoninae by the appearance of Zenk- erʼs organ, hemipenis, prehensile palps, and chaetotaxies of the furca, and the second and third thoracopods. Abcandonopsis sub- gen. nov. of the genus Candonopsis is described in this paper. It contains the following three species: C. (A.) williami Karanovic and Marmonier, 2002, Candonopsis (Abcandonopsis) aula sp. nov., and C. (A.) indoles sp. nov. The first species is designated as the type species of the subgenus. Both new species are known only from the subterranean waters of the Kimberley Region of Western Australia, while C. (A.) williami is endemic to the subterranean waters of the Murchison Region of Western Australia. Abcandonop- sis differs from the nominal subgenus by characters of the carapace and the chaetotaxy of the first, second and third thoracopods. In the present paper three species of the subgenus Candonopsis are synonymized, all following examination of the relevant type mate- rial. Candonopsis calva Harding, 1962 is synonymized with C. (C.) sumatrana Klie, 1932, while C. dorsorecta Rome, 1962 and C. depressa Rome, 1962 are both synonymized with C. (C.) navicula Daday, 1910. An additional description of the latter species is provided. The record of Candonopsis tenuis (Brady) from Western Samoa is added to the synonymy of C. sumatrana which is also redescribed in this paper. The following new combinations are proposed: Cubacandona anisitsi (Daday, 1905) new combination; Cubacandona columbiensis (Méhes, 1913) new combination; and Cubacandona falclandica (Vávra, 1898) new combination. After this revision the genus Candonopsis includes 24 recent species. Key words: Candoninae, Candonopsis, revision, subterranean waters, Australia
Transcript of Towards a revision of Candoninae (Crustacea: Ostracoda): Australian representatives of the...

90 T.R. Sawicki, J.R. Holsinger, T.M. Iliffe Subterranean Biology 2: 91-108, 2004
INTRODUCTION
The genus Candonopsis was established at the end of the 19th century by Vávra (1891) to include the spe-cies Candona kingsleii, described earlier from south-east England (Brady and Robertson 1870). At that time Candonopsis and Candona Baird, 1845 were the only two genera in the subfamily Candoninae. The original generic diagnosis of the genus Candona was so broad (see Baird 1845) that it continued for many years to ac-cumulate many phylogenetically very distinct species. On the other hand, the diagnosis of the genus Cando-nopsis given by Vávra (1891) was much narrower and it allowed inclusion of only very closely related species. However, due to the further enrichment of our knowl-edge of ostracods and the discovery of new species, the generic diagnosis of the genus Candonopsis has been revised twice. Danielopol (1980) added characters of the cleaning leg, hemipenis and Zenkerʼs organ, while recently Karanovic & Marmonier (2002) gave more details of the soft parts morphology, and broadened the diagnosis substantially. Klie (1932) divided the genus Candonopsis into three groups of species based upon the presence of spines on the furcal claws. However, this division was shown to be unsustainable after the discov-ery of many transitional forms (Karanovic and Marmo-nier 2002). At present 25 recent species are recognized in this genus. A key to their identification was provided by Karanovic and Marmonier (2002).
The majority of the Candonopsis species (19) are distributed in the Southern Hemisphere, with 12 sur-face and seven subterranean species. Of the six species known from Europe and North America, two are surface and four are subterranean inhabitants. Continents where the Candonopsis fauna is the most diverse are Africa with eight species (Vávra 1895; Daday 1910, 1913; Klie 1944; Rome 1962; Löffler 1968), and Australia with six species (Brady 1886; Karanovic and Marmonier 2002). Only one species is known so far from the Indian sub-continent (Gupta 1988) and three others from Sumatra, Java and the Solomon Islands, respectively (Klie 1932; Harding 1962). Until recently, four species were known from Central and South America (Vávra 1898; Daday 1905; Méhes 1913; Broodbakker 1983), but Karanovic and Marmonier (2002) proposed the exclusion of three species, retaining only Candonopsis hummelincki Broodbakker, 1983 as a valid representative of the ge-nus. The three excluded species are more closely related to the genera Cubacandona Broodbakker 1983 and Car-ibecandona Broodbakker, 1983, both known only from Central and South America (Broodbakker, 1983). The search for the type material of these species has been unsuccessful and a proper redescription cannot be given; I therefore only tentatively assign them to the genus Cu-bacandona. However, one should not exclude the pos-sibility that they may represent a new genus, or at least some of them. The similarity of the soft parts of Cando-nopsis, Cubacandona and Caribecandona is prominent
Towards a revision of Candoninae (Crustacea, Ostracoda): on the genus Candonopsis Vávra, with descriptions of new taxa
Ivana KARANOVIC
Western Australian Museum, Francis Street, Perth 6000, W. A. Australia
ABSTRACT Candonopsini trib. nov. is described in the present paper to encompass Candonopsis Vávra, 1891 Cubacandona Broodbakker,
1983 and Caribecandona Broodbakker, 1983. The new tribe stands apart from the rest of Candoninae by the appearance of Zenk-erʼs organ, hemipenis, prehensile palps, and chaetotaxies of the furca, and the second and third thoracopods. Abcandonopsis sub-gen. nov. of the genus Candonopsis is described in this paper. It contains the following three species: C. (A.) williami Karanovic and Marmonier, 2002, Candonopsis (Abcandonopsis) aula sp. nov., and C. (A.) indoles sp. nov. The first species is designated as the type species of the subgenus. Both new species are known only from the subterranean waters of the Kimberley Region of Western Australia, while C. (A.) williami is endemic to the subterranean waters of the Murchison Region of Western Australia. Abcandonop-sis differs from the nominal subgenus by characters of the carapace and the chaetotaxy of the first, second and third thoracopods. In the present paper three species of the subgenus Candonopsis are synonymized, all following examination of the relevant type mate-rial. Candonopsis calva Harding, 1962 is synonymized with C. (C.) sumatrana Klie, 1932, while C. dorsorecta Rome, 1962 and C. depressa Rome, 1962 are both synonymized with C. (C.) navicula Daday, 1910. An additional description of the latter species is provided. The record of Candonopsis tenuis (Brady) from Western Samoa is added to the synonymy of C. sumatrana which is also redescribed in this paper. The following new combinations are proposed: Cubacandona anisitsi (Daday, 1905) new combination; Cubacandona columbiensis (Méhes, 1913) new combination; and Cubacandona falclandica (Vávra, 1898) new combination. After this revision the genus Candonopsis includes 24 recent species.
Key words: Candoninae, Candonopsis, revision, subterranean waters, Australia

92 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 93
and their separate position within the subfamily Can-doninae is so distinct that a new tribe is described in the present paper to include the three genera.
Almost all Candonopsis species are known to be restricted to the continent or island from which they have been described. There are only two exceptions: Candonopsis tenuis (Brady, 1886) and C. putealis Klie, 1932. Described and redescribed from New South Wales (Brady 1886; Sars 1896), the first species is also found in the Perth Basin (Karanovic and Marmonier 2002), Sumatra (Klie 1932) and Western Samoa (Victor and Fernando 1978). After rechecking the material of the last record, deposited in the Canadian Museum of Nature, it became clear that Victor and Fernando (1978) were not dealing with C. tenuis at all, but with C. sumatrana Klie, 1932. Also, after rechecking the type material of C. cal-va Harding, 1962, deposited in the British Museum, this species is added as a junior synonym of C. sumatrana. In the present paper C. sumatrana is redescribed after the specimens both from Western Samoa and the Solomon Islands. The material of C. tenuis from Sumatra could not be checked, and, unfortunately, Klie (1932) gave just a short note about it. C. putealis, described from Su-matra (Klie 1932), was recorded from south-west India (Victor and Fernando 1979). However, the descriptions and drawings of C. putealis given by Victor and Fern-ando (1979) more closely resemble C. navicula Daday, 1910 described from Kenya. Because the Indian mate-rial of C. putealis could not be checked, this similarity is only further discussed in this paper. Two other species are synonymized with C. navicula after examination of their type specimens: C. dorsorecta Rome, 1962 and C. depressa Rome, 1962, both described from Lake Tan-ganyika by Rome (1962).
Two new species of the genus Candonopsis are described in the present paper based upon the speci-mens collected in Australian subterranean waters. They display some peculiarities in the characters of both the carapace and soft parts which they share with another Australian species, Candonopsis williami Karanovic and Marmonier, 2002. A new subgenus is established to encompass these three species.
MATERIALS AND METHODS
The samples from the Kimberley Region were col-lected with haul-nets (mesh size 250 or 350 microme-ters) from the water pumping and monitoring bores used by the Argyle Diamond Mine for industrial purposes. Haul-nets are simple plankton nets of different sizes suitable for the bore, which range from 30 to 180 mm in diameter. Weighed nets were lowered down into the bore with one bottle screwed on its distal part then hauled through the water column, usually a number of times. All samples were sorted while alive under the stereo-microscope and the ostracods were then fixed in 70% ethanol and assigned a field number (prefix BES).
Ostracods were dissected in an equal mixture of distilled water and glycerol with fine entomological needles (mark 000). Dissected appendages and valves were mounted in Faureʼs medium. All non-dissected material is preserved in 70% ethyl-alcohol in glass test-tubes. Drawings were prepared using a drawing tube at-tachment on a Leica-DMLS microscope, with C-PLAN achromatic objectives. All material of the new species is deposited in the Western Australian Museum. Mate-rial of Candonopsis tenuis (Brady, 1886) from Western Samoa is deposited in the Canadian Museum of Nature. The type material of Candonopsis calva Harding, 1962 is deposited in the British Museum, while the type mate-rial of Candonopsis dorsorecta Rome, 1962 and C. de-pressa Rome, 1962 are deposited in the Royal Belgium Institute of Natural Sciences.
In the systematic part of this paper the length of all segments was measured in the middle of the segments, and length ratios are presented beginning with the proximal segment. The terminology of the appendages follows Martens (1998), while chaetotaxy of all limbs follows the model proposed by Broodbakker & Danielo-pol (1982), revised for the antenna by Martens (1987), and the third thoracopod by Meisch (1996). Lobes on the hemipenis are designated according to Danielopol (1969). In front of the abbreviations for the setae on the endopodal segments of the third thoracopod and setae on the first thoracopod letter “T” is added, not to be con-fused with the abbreviations for hemipenis gibven in the abbreviation list bellow.
Abbreviations used in text and figure legends: a - lateral lobe on hemipenis; A1 - antennula; A2 - an-tenna; b - medial lobe on hemipenis; B-R - Bou-Rouche sample; BES - prefix for the field number; BM - British Museum; CMNC - Canadian Museum of Nature Crusta-cea; d1, d2, dp - setae on basal segment T3; Fu - furca; G1, G2, G3, GM, Gm - antennal claws; H - height; L - length; LV - left valve; Md - mandible; Mxl -maxillula; RBINS - Royal Belgium Institute of Naturale Sciences; RV - right valve; Ta, Taʼ, Tb, Td - setae on T1; Te, Tf, Tg, Th1, Th2, Th3 - setae on endopodal segments T3; T1, T2, T3 - first, second and third thoracopods; t1, t2 - medial setae on the second endopodal segment A2; W - width; WAM - Western Australian Museum; Y, ya, y1, y2, y3 - aesthetascs; z1, z2, z3 - apical setae on the sec-ond endopodal segment of A2.
RESULTS
Family Candonidae Kaufmann, 1900Subfamily Candoninae Kaufmann, 1900
Tribe Candonopsini trib. nov.
Type genus: Candonopsis Vavra, 1891.Other genera: Caribecandona Broodbakker, 1983;
Cubacandona Broodbakker, 1983.

92 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 93
Diagnosis: Carapace shape various (reniform, elongated, trapezoidal), ornamentation possible, hinge adont. Central muscle scars conforming to general pat-tern of Candoninae. A1 7- or 6-segmented. Male sexual bristles on A2 present or absent. Terminal claw of Md palp broadly fused with the apical segment, externally accompanied with one distinctly weaker claw; exopodite with eight filaments. Penultimate segment of Mxl palps dilated. T1 with one or two “Ta” setae. Exopodite with two or three rays. Prehensile palps with only one subter-minal sclerotized structure being well developed (other one small or completely absent). T2 5-segmented, T3 5-segmented. Terminal segment T3 with two long and one short seta. Fu without posterior seta, posterior claw well developed, reduced or missing. Hemipenis with triangular and extended lobe “a”, lobe “b” flat, lobe “h” very small or absent. Sometimes two “a” lobes present. Female Genital field without processes. Zenkerʼs organ well developed and stocky in appearance, with seven whorls of spines.
Genus Candonopsis Vávra, 1891Subgenus Abcandonopsis subgen. nov.
Type species, here designated: Candonopsis (Abcan-donopsis) williami Karanovic and Marmonier, 2002
Other species: Candonopsis (Abcandonopsis) aula sp. nov.; C (A.) indoles sp. nov.
Diagnosis: Carapace reniform, smooth. In dorsal view, female carapace strongly asymmetrical. Marginal pore canals short, straight and dense. A1 7-segmented. A2 with 3-segmented endopodite in females, and 4-segmented in males (male sexual bristles developed). Terminal segment of Md-palp three times longer than wide, apical claw broadly fused with segment. Penul-timate segment of Mxl palp dilated. T1 with only one Ta seta. Male prehensile palps assymetrical, right one more stocky. Both palps with only one sclerotized seta. T2 5-segmented; basal segment without any seta. T3 5-segmented; basal segment with only d1 and dp setae present, endopodal setae Te and Tf missing. Terminal segment of same appendage with two long and one small seta. Posterior furcal seta absent, posterior claw absent or well developed. Hemipenis: lobe “a” more or less triangular, but always distinctly higher than the other two lobes. Female genital field without any ap-pendages.
Etymology: The subgenus name is composed of the Latin preposition “Ab” meaning from, and the genus name Candonopsis. Gender feminine.
Candonopsis (Abcandonopsis) aula, new speciesFigs 1-18
Material: 1) Holotype ™ (dissected on one slide - WAM C28442), two paratype ™ (one dissected - WAM
C28443, other in alcohol - WAM C28444), from the bore 13, Argyle Diamond Mine, Kimberley, Western Australia, 16043 ̓ S 128024 ̓ E; 10 October 2002; col-lectors W. F. Humphreys and R. Webb (BES: 9696). 2) One ¢ (decomposed and dissected on one slide - WAM C28445) from the bore MB13S, Argyle Diamond Mine, Kimberley, Western Australia, Australia; 16043ʼS 128024”E; 13 October 2002; collectors W. F. Humphreys and R. Webb (BES: 9754).
Female (holotype)L of LV 0.519 mm, RV 0.49 mm. Greatest H lies ar-
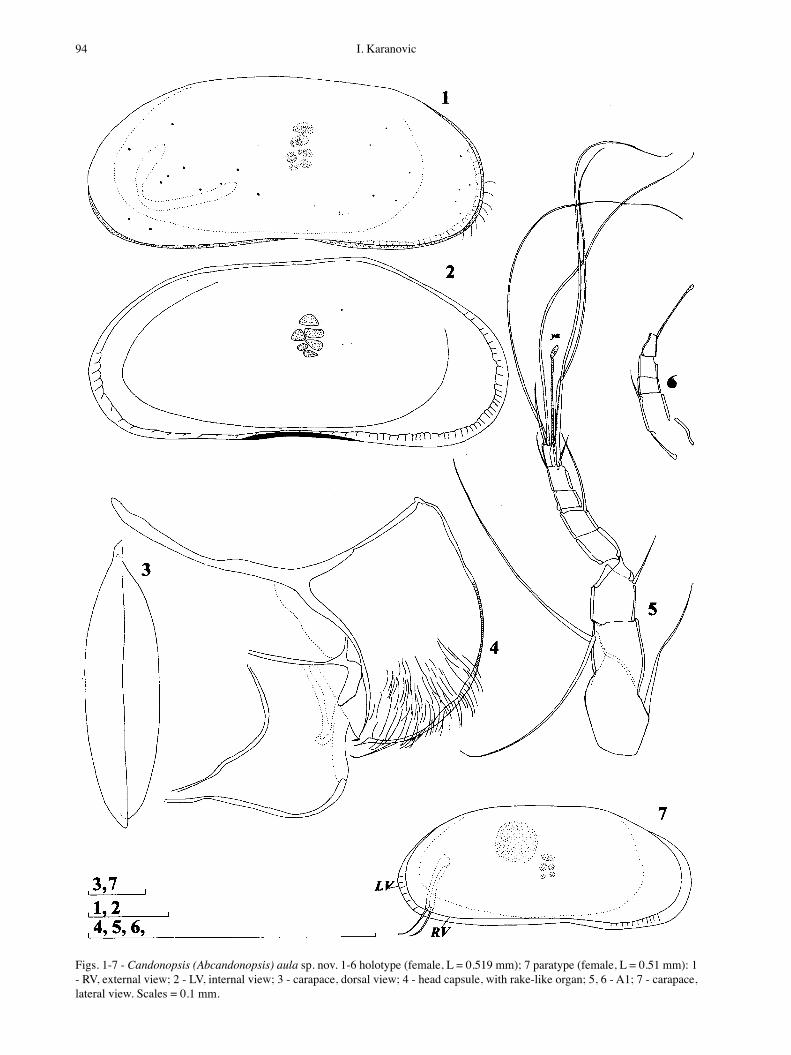
round mid-L and equals 41% of L. Greatest W lies also around middle, equals 25% of L. In lateral view (Fig. 1, 2) valves reniform. Dorsal margin straight in middle, rounded towards posterior and sloping anteriorly. Both posterior and anterior margins rounded and equally wide. Ventral margin concave arround mid-L. Calci-fied inner lamella not wide: anteriorly 16%, posteriorly 8% of total L. Marginal pore canals short, straight and dense. Selvage peripheral. In dorsal view (Fig. 3) LV distinctly overlaps RV on both ends. Valve surface smooth and covered with fine, short hairs. Colour pale white.
A1 (Fig. 5) 7-segmented. First segment with one seta antero-proximally and without any setae antero-distally, same segment with two long setae postero-distally. Next segment with one seta almost reaching distal end of third segment. Third segment without any seta, fourth with one anterior seta reaching distal end of terminal seg-ment, fifth segment without setae. Penultimate segment with one posterior seta (exceeds distal end of terminal segment), two long anterior setae, and very tiny alpha seta. Terminal segment distally with one short seta, two long setae and aesthetasc (ya) which only two times longer than terminal segment. L ratios of five endopodal segments 1.8 : 1.3 : 1 : 1.3 : 1.16.
A2 (Fig. 8) with 3-segmented endopodite. Exopodite consists of plate with one long and two short setae. First endopodal segment interno-distally with one long and one short seta. Penultimate segment with one seta ex-terno-medially, while interno-medially only one “t” seta observed. Only seta z1 visible. Claws G1, G2 and G3 of subequal L, and 1.7 times longer than first endopodal seg-ment. Claw GM 1.4 times longer than first endopodal seg-ment, while Gm about as long as same segment. All claws distally serrated. Only aesthetascs Y and y3 observed. Aesthetasc y1 fallen of, but its position clearly visible. Aesthetasc Y about as long as first endopodal segment, y3 slightly smaller than corresponding segment. L ratios of three endopodal segments equal 2.6 : 1.8 : 1.
Md (Fig. 10) with coxa, 4-segmented palpus, and exopodite with eight rays. Coxa with seven teeth. First segment of Md palp on outer edge without any seta, while internally with one smooth and short seta (alpha), two long plumed setae (more distal one S1), and one plumed small seta (S2). Second segment externally with two setae. Internally same segment with 3+2 setae in
94 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 95
Figs. 1-7 - Candonopsis (Abcandonopsis) aula sp. nov. 1-6 holotype (female, L = 0.519 mm); 7 paratype (female, L = 0.51 mm): 1 - RV, external view; 2 - LV, internal view; 3 - carapace, dorsal view; 4 - head capsule, with rake-like organ; 5, 6 - A1; 7 - carapace, lateral view. Scales = 0.1 mm.

94 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 95
Figs. 8-12 - Candonopsis (Abcandonopsis) aula sp. nov. Holotype (female, L = 0.519 mm): 8 - A2; 9 - Fu with genutal field; 10 - Md; 11 - T3; 12 - Fu attachment. Scale = 0.1 mm.

96 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 97
Figs. 13-18 - Candonopsis (Abcandonopsis) aula sp. nov. 13, 17, 18 male (WAM C28445); 14, 15, 16 holotype (female, L = 0.519 mm): 13 - Zenkerʼs organ; 14 - Mxl; 15 - T2; 16 - T1; 17 - Fu; 18 - A2. Scale = 0.1 mm.

96 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 97
bunch, all plumed. Penultimate segment externo-medi-ally with one seta (exceeds penultimate segment), one gamma seta (smooth), and two setae intero-distally. Terminal segment with broadly fused terminal claw (distally plumed), one stronger seta externally, and one weak seta internally.
Mxl (Fig. 14). Exopodite with about 19 filaments. Palp 2-segmented. First segment dilated and with four plumed setae, second segment with two claws and three setae. Terminal segment rectangular.
Rake-like organ (Fig. 4) with numerous small teeth. T1 (Fig. 16) with one Ta seta, and setae Tb and Td
setae present. Two setae observed on exopodite. Length ratios of three setae on endopodite equal 1 : 2 : 2.2.
T2 (Fig. 15) 5-segmented. Basal segment without any seta. First and second endopodal segments with one seta each. Penultimate segment distally with two setae. Terminal segment with two setae and one long, distally serrated claw, 1.3 times as long as three distal segments combined.
T3 (Fig. 11) 5-segmented. Second and third endopo-dal segments completely separated. Basal segment with only d1 and dp seta present, first one unusually long. First two endopodal segments lack any seta, while pe-nultimate segment carries Tg seta. L ratios of Th1, Th2, and Th3 setae equal 1 : 2 : 2.
Fu (Fig. 9) with posterior claw missing. Anterior claw well developed, serrated, and almost as long as anterior furcal margin. Claw without spine. Anterior and posterior setae absent as well. Genital field simple, and without any processes.
MaleA2 (Fig. 18) with 4-segmented endopodite. Only one
male bristle observed (t2). Claws G1 and G3 reduced into small, weak claws, G1 3.5 times longer than termi-nal segment, G3 as long as corresponding segment. Seta z1 not observed, z2 claw like z3 seta-like. G2 as long as z2. GM and Gm claws on terminal segment present.
Fu (Fig. 17) with less rounded claw than in female.Zenkerʼs organ (Fig. 13) with seven whorls of
spines.The only male collected has completely decalcified
valves and decomposed soft parts, so the shape of cara-pace, and morphology of other soft parts could not be observed.
Etymology: The species name is the Latin noun aula (feminine), meaning courtyard of a Greek house, hall of a Roman house.
VariabilityThe only variability noticed is on the A1 in the
holotype. Namely, one A1 has a seta anteriorly on the fourth segment (Fig. 5) while on the other A1 the same segment carries that seta posteriorly (Fig. 6). In the para-type female (Fig. 7) both A1 have the same seta situated anteriorly.
Candonopsis (Abcandonopsis) indoles new speciesFigs 19-33
Material: 1) Holotype, subadult ™ (dissected on slide WAM C28446), from the bore MB 30S, Argyle Dia-mond Mine, Kimberley, Western Australia, Australia, 16041 ̓S 128027 ̓E; 10 October 2002; collectors W. F. Humphreys and R. Webb (BES: 9700); 2) One ¢ (dis-integrated and dissected on one slide - WAM C28447), one juvenile ™ (dissected on one slide WAM C28448); from the bore MB 29D, Argyle Diamond Mine, Kimber-ley, Western Australia, Australia, 16041ʼS 128027 ̓E; 10 October 2002; collectors W. F. Humphreys and R. Webb (BES: 9692).
Female (holotype)L of LV 0.52 mm, RV 0.49 mm. Greatest H lies ar-
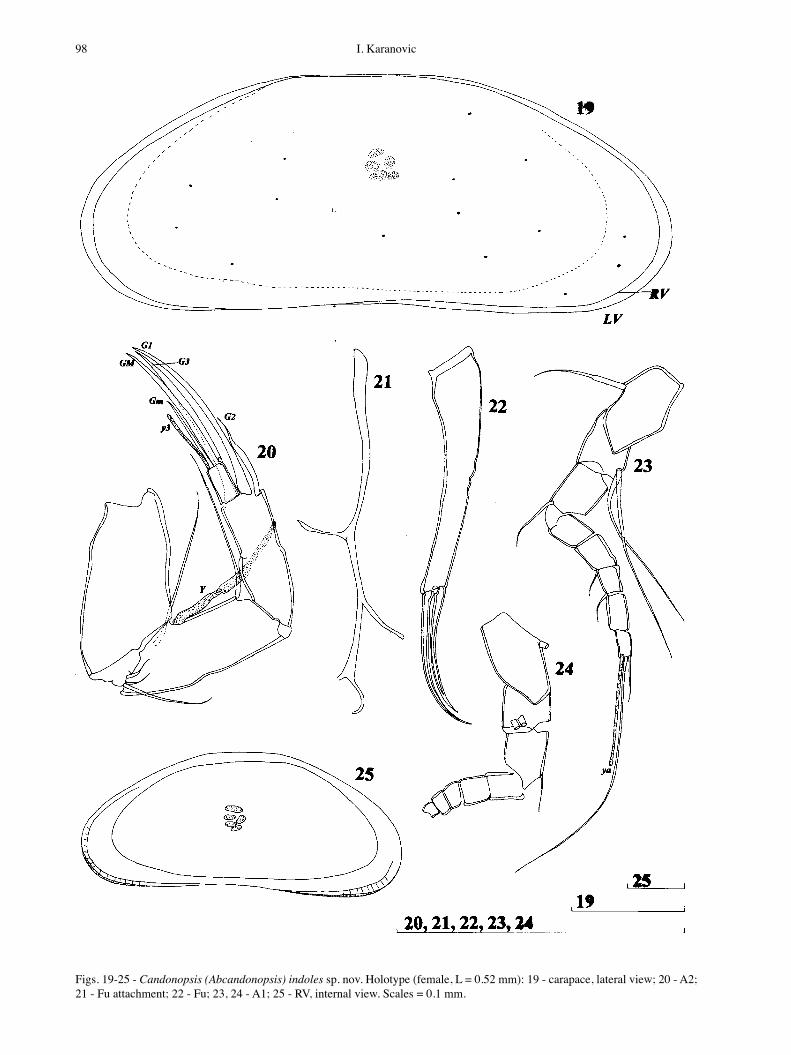
round mid-L and equals 38% of L. Greatest W lies also around middle and equals about 25% of L. In lateral view (Fig. 19, 25) valves reniform to slightly trapezoidal Dorsal margin straight in middle, equally slopping down towards both anterior and posterior ends Both posterior and anterior margins rounded and equally wide. Ventral margin concave arround middle. Calcified inner lamella not wide: anteriorly 14%, posteriorly 8% of total L Marginal pore canals short straight and dense. Selvage peripheral. In dorsal view LV distinctly overlaps RV on both ends. Valve surface smooth and covered with fine, short hairs. Colour pale white.
A1 (Fig. 23) 7-segmented. First segment with one seta antero-proximally and without any setae antero-distally, same segment with two long setae postero-dis-tally. Next segment with one seta almost reaching distal end of third segment. Third segment without any seta, fourth and fifth segments with one anterior seta each. Setae on penultimate segment not observed. Terminal segment distally with one short seta, one long setae and aesthetasc (ya) which four times longer than terminal segment. L ratios of five endopodal segments as 1.25 : 1.25 : 1 : 1.4 : 1.
A2 (Fig. 20) with 3-segmented endopodite. Ex-opodite consists of plate with one long and two short setae. First endopodal segment interno-distally with one long seta. Medio-lateral setae on penultimate segment not observed, but their position clearly visible. No “z” setae observed. Claws G1 and G3 of subequal L, and as long as first endopodal segment. G2 less than half as long as G1 and G3 (probably subadult state of char-acter). Claw GM 0.85 times as long as first endopodal segment, Gm about two times longer than terminal seg-ment. Only aesthetascs Y and y3 observed. Aesthetasc Y slightly shorter than first endopodal segment, y3 1.8 times longer than terminal segment. L ratios of three endopodal segments equal 4.5 : 3.4 : 1.
Md palp (Fig. 31) with 4-segmented palpus. First seg-ment of Md palp on outer edge without any seta, while internally with only two plumed setae observed (more dis-tal one S1). Second segment externally with two setae. In-
98 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 99
Figs. 19-25 - Candonopsis (Abcandonopsis) indoles sp. nov. Holotype (female, L = 0.52 mm): 19 - carapace, lateral view; 20 - A2; 21 - Fu attachment; 22 - Fu; 23, 24 - A1; 25 - RV, internal view. Scales = 0.1 mm.

98 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 99
Figs. 26-33 - Candonopsis (Abcandonopsis) indoles sp. nov. 26, 27, 28, 33 male (WAM C 28447, L = 0.52 mm); 29, 30, 31, 32 holotype (female, L = 0.52 mm): 26 - RV, external view; 27 - hemipenis; 28 - segments of A1; 29 - head capsule with rake-like organ; 30 - T3; 31 - Md palp; 32 - T2; 33 - Fu trunk. Scales = 0.1 mm.

100 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 101
ternally same segment with 3+2 setae in bunch, long ones plumed. Penultimate segment externo-medially with one seta (not exceeds penultimate segment), one gamma seta (smooth), and two setae interno-distally. Terminal segment with broadly fused terminal claw (distally plumed), one stronger seta externally, and one weak seta internally.
Mxl similar to that of previous species.Rake-like organ (Fig. 29) with numerous small
teeth. T1 with one Ta seta, setae Tb and Td present. Two
setae observed on exopodite. Length ratios of three setae on endopodite similar to those of previous species.
T2 (Fig. 32) 5-segmented. Basal segment without any seta. First endopodal segment with one seta, follow-ing one without any seta observed. Penultimate segment distally with one seta. Terminal segment with two setae and one long, distally serrated claw, 1.2 times as long as the three distal segments combined.
T3 (Fig. 30) 5-segmented. Penultimate segment incompletely divided. Basal segment with only d1 and dp present. First two endopodal segments lack any seta, while penultimate segment carries Tg seta. L ratios of Th1, Th2, and Th3 setae equal 1 : 2.7 : 2.7.
Fu (Fig. 22) with both claws well developed. Ante-rior and posterior setae absent. L ratios of anterior Fu margin, anterior and posterior claws equal 1.75 : 1.1 : 1.
MaleCarapace reniform in lateral view (Fig. 26). L 0.52
mm. Greatest H equals 40% of L. L ratios of five distal endopodal segments on A1 (Fig. 28) equal 1.8 : 2 : 1.4 : 1.7 : 1. Setae on A1 not observed, all fallen off. A2 com-pletely decomposed and not observed, as well as Md, Mxl, prehensile palps, T2 and T3.
Fu (Fig. 33) with longer Fu trunk than in female. Both claws fallen off, however their position clearly visible.
Hemipenis (Fig. 27) with rounded lobe “a”. Lobe “b” flat, “h” not observed. Internal parts decomposed, only two loops on duct visible.
Zenkerʼs organ with seven whorls of spines.
VariabilityThe only noticed variability is that on the A1 in
holotype. One A1 in the holotype female has deformed segments (Fig. 24).
Etymology: The species name is the Latin noun in-doles (feminine) meaning character.
Subgenus Candonopsis Vávra, 1891
Candonopsis (Candonopsis) sumatrana Klie, 1932Figs 34-46
SynonymyCandonopsis sumatrana n. sp. Klie 1932: 454-455,
Figs 16-23.
Candonopsis calva n. sp. Harding 1962: 58-59, Figs 36-44. New synonymy.
Candonopsis tenuis (Brady, 1886). Victor and Fern-ando 1978: 420-421. Misidentification.
Material: : 1) Six females (one dissected on a slide, others in alcohol - CMNC 2002-0090, specimens la-belled as Candonopsis tenuis), from the crab holes, Western Samoa, Pacific Ocean; 1975; collector J. S. Pillai; 2) Three females (one dissected on slide, oth-ers in alcohol - BM 1962.12.10.2, specimens labelled as paratypes of Candonopsis calva), from the stomach of Eleotris fusca, Lake Te-Nggano, Rennell Island; 18 October 1951; coll. T. Wolff.
Redescription of femaleL from 0.77 mm to 0.88 mm. Greatest H lies slightly
behind middle and equals 51% of L. Greatest W lies around middle and equals 32% of L. In lateral view (Fig. 24, 36, 45) valves reniform. Dorsal margin evenly rounded towards posterior end and slopping down to-wards anterior end. Both posterior and anterior margins rounded, posterior one slightly narrower. Ventral margin concave around middle. Calcified inner lamella not wide: anteriorly 9%, posteriorly 6% of total L Marginal pore canals short, straight and dense. Selvage peripheral. In dorsal view (Fig. 35) LV very slightly overlaps RV only on posterior end. Valve surface smooth and densely covered with long hairs.
A1 (Fig. 39) 7-segmented. First segment with one seta antero-proximally and one antero-distally, same segment with two long setae postero-distally. Follow-ing segment with one seta almost reaching distal end of fourth segment. Third segment posteriorly with one short seta, fourth and fifth segments posteriorly with one short seta each, anteriorly with two long setae each. Pe-nultimate segment with one seta almost reaching distal end of terminal segment, one seta three times as long, two long setae, and very small (alpha) seta. Terminal segment distally with one short seta, two long setae and aesthetasc (ya) which only 1.36 times longer than terminal segment. L ratios of five endopodal segments 1.4 :1 : 1.25 : 1.25 : 1.25. Posterior margins of first three endopodal segments covered with fine and short hairs.
A2 (Fig. 43) with 3-segmented endopodite. Ex-opodite consists of plate with one long and two short setae. First endopodal segment interno-distally with one long and one short seta. Penultimate segment with two seta externo-medially, and two “t” seta interno-medially. Seta z1 transformed into claw which exceeds distal end of terminal segment; seate z2 and z3 slightly shorter. Claws G1, G2 and G3 of subequal L, 1.35 times longer than first endopodal segment. Claw GM as long as first endopodal segment, Gm three times longer than termi-nal segment. All claws medially serrated. All aesthetascs observed. Aesthetasc Y about half as long as first endop-odal segment, y1 not reaching t setae, y2 not reaching distal end of terminal segment, y3 1.7 times longer than

100 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 101
Figs. 34-39 - Candonopsis (Candonopsis) sumatrana Klie, 1932. Female (CMNC 2002-0090, L = 0.78 mm): 34 - carapace, lateral view; 35 - carapace, dorsal view; 36 - RV, external view; 37 - Md palp; 38 - rake-like organ; 39 - A1. Scales = 0.1 mm.
102 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 103
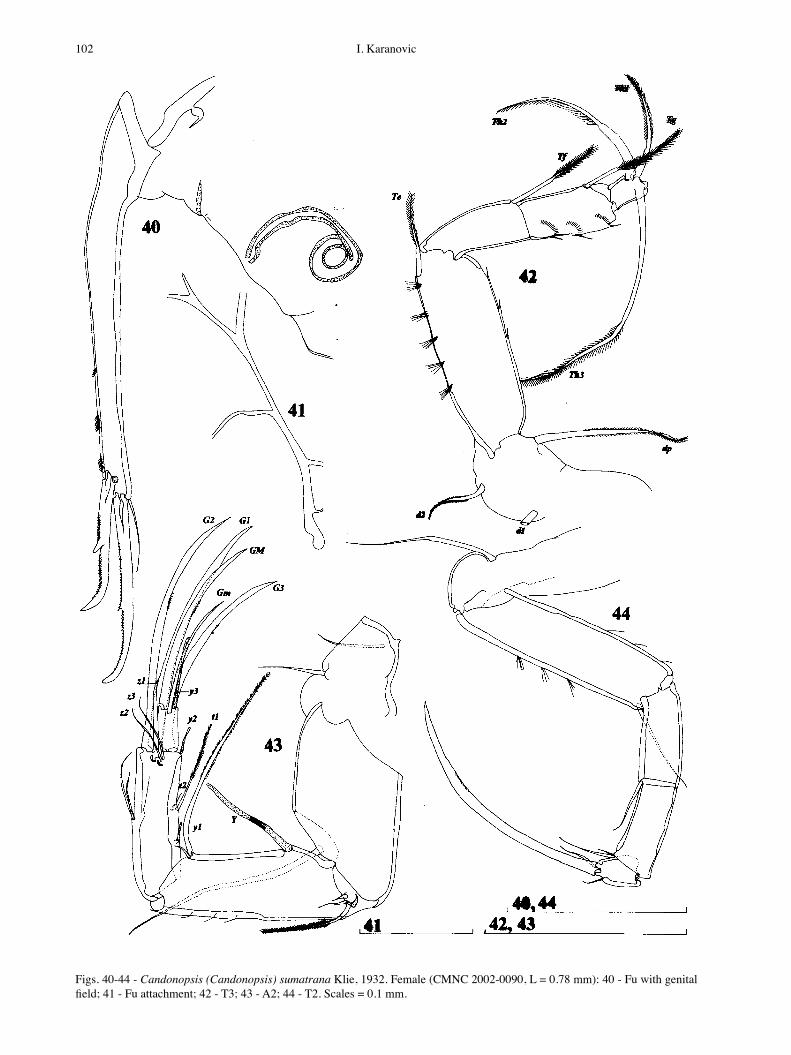
Figs. 40-44 - Candonopsis (Candonopsis) sumatrana Klie, 1932. Female (CMNC 2002-0090, L = 0.78 mm): 40 - Fu with genital field; 41 - Fu attachment; 42 - T3; 43 - A2; 44 - T2. Scales = 0.1 mm.

102 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 103
Figs. 45-54. 45, 46 - Candonopsis (Candonopsis) sumatrana Klie, 1932, female (BM 1962.12.10.2, L = 0.76 mm); 47-54 Can-donopsis navicula Daday, 1910; 47, 48, 49, 50, 51 female (RBINS OC 511, L = 1.00 mm); 52, 53, 54 female (RBINS OC 512b): 45 - carapace, lateral view; 46- Fu; 47 - RV, internal view; 48 - terminal segment of Md palp; 49 - Fu; 50 - T3; 51 - five terminal segments A1; 52 - Fu; 53 - terminal segment of Md palp; 54 - six terminal segments A1. Scales = 0.1 mm.

104 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 105
terminal segment. L ratios of three endopodal segments equal 4.3 : 3.4 : 1.
Md palp (Fig. 37) with two setae externally on second segment, 3+2 setae internally. Penultimate seg-ment with two setae externally and four setae distally. Terminal segment with broadly fused claw, one seta externally, and one interanally. Same segment six times longer than wide.
Rake-like organ (Fig.38) with approximately 20 small teeth.
T1 with two Ta seta, and both Tb and Td setae present. Three seta observed on exopodite.
T2 (Fig. 44) 5-segmented. Basal segment with one long seta. First and second endopodal segments with one seta each. Penultimate segment distally with two setae. Terminal segment with two setae and one long, medi-ally serrated claw, about as long as three distal segments combined.
T3 (Fig. 42) 5-segmented. Second and third endopo-dal segments completely separated. Basal segment with d1, d2 and dp setae present. First three endopodal seg-ments with one seta each. L ratios of Th1, Th2, and Th3 setae equal 1 : 1.4 : 2. All T3 setae plubed.
Fu (Figs 40, 46) with both claws well developed. Anterior claw with prominent spine, posterior claw with small one. Anterior seta well developed and almost reaching middle of anterior claw. L ratios of anterior margin, anterior and posterior claws equal 2.2 : 1.4 : 1. Posterior furcal margin with groups of small spines. Genital field simple, and without any processes.
Males reported by Klie (1932). The populations from Western Samoa and Solomon Islands are both partheno-genetic.
VariabilityKlieʼs (1932) specimens seem to have a slightly more
protruded anterior end of carapace and a wider calcified inner lamella. Specimens from the Solomon Islands have asymmetrical valves (RV overlapping LV, Fig. 45), but this is an artefact due to the carapace being completely decalcified. No other variability was noticed.
Candonopsis (Candonopsis) navicula Daday, 1910Figs 47-54
SynonymyCandonopsis navicula n. sp. Daday 1910: 226-227,
Plate 16, Figs 30-35.Candonopsis depressa n. sp. Rome 1962: 9-12, Fig.
2. New synonymy.Candonopsis dorsorecta n. sp. Rome 1962: 13-16,
Fig. 3. New synonymy.
Material: 1) One female (dissected on a slide - RBINS OC 511, specimen labelled as the type of Can-donopsis depressa), from the Station 8, Kungwe Bay, Lake Tanganyika, Tanzania; 12-13 December 1946; 2) One female (dissected on a slide - RBINS OC 512b,
specimen labelled as the type of Candonopsis dor-sorecta), station 260, between Kigoma and Ujiji, Lake Tanganyika, Tanzania; 9 December 1946.
Additional description of femaleL around 1.00 mm. Greatest H lies on posterior third
and equals about 50% of L. Dorsal margin evenly round-ed towards posterior end, and slopping down towards anterior end, while flat medially (Fig. 47). Posterior margin slightly wider than anterior one. Calcified inner lamella narrow: anteriorly 16%, posteriorly 11% of L. Marginal pore canals short, straight and dense. Selvage peripheral.
A1 (Fig. 51, 54) 7-segmented. First segment with one seta antero-proximally and one antero-distally, same segment with two long setae postero-distally. Fol-lowing segment with one seta exceeding distal end of fourth segment. Third segment posteriorly with one seta. Fourth and fifth segments posteriorly with one short seta and anteriorly with two long setae each. Penultimate segment with one seta almost reaching distal end of terminal segment, one long seta, very small (alpha) seta and two long setae anteriorly. Terminal segment distally with one claw-like seta, one long seta plus one distinctly shorter seta, and aesthetasc (ya) which as long as termi-nal segment. L ratios of five endopodal segments 1.6 :1 : 1 : 1 : 1.25.
A2 with 3-segmented endopodite. Exopodite con-sists of plate with one long and two short setae. First endopodal segment intero-distally with one long and one short seta. Penultimate segment with two seta ex-tero-medially, and two “t” seta interno-medially. Seta z1 transformed into claw which slightly exceeds distal end of terminal segment; seate z2 and z3 slightly shorter. Claws G1, G2 and G3 of subequal L, and as long as first endopodal segment. Claw GM slightly shorter than first endopodal segment, while Gm three times longer than terminal segment. All claws medially serrated. L ratios of three endopodal segments equal 5.3 : 4.1 : 1.
Md (Fig. 48, 53), and Mxl similar to that of the previ-ous species.
Rake-like organ with numerous small teeth. T1 with two Ta setae, setae Tb and Td setae present.
Two setae observed on exopodite.T2 5-segmented. Basal segment with one long seta.
First and second endopodal segments with one seta each. Penultimate segment distally with two setae. Terminal segment with two setae and one long, medially serrated claw, about as long as three distal segments combined.
T3 (Fig. 50) 5-segmented. Second and third endopo-dal segments completely separated. Basal segment with d1, d2 and dp setae present. All endopodal segments with one seta each, and all very small. L ratios of Th1, Th2, and Th3 setae equal 1 : 2.3 : 4. All T3 setae plumed.
Fu (Figs 49, 50) with both claws well developed, each carrying prominent spine. Anterior seta very short, and more claw like, often hardly visible. L ratios of ante-

104 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 105
rior margin, anterior and posterior claws equal 2 : 1.4 : 1. Posterior furcal margin with small spines. Genital field simple, and without any processes.
Only juvenile males reported (Daday 1910), while Rome (1962) found only females.
VariabilityRome (1962) reported two setae on the T1 exopodite
in Candonopsis dorsorecta and three in C. depressa. However, I have found two setae in both examined specimens, while Daday (1910) reported three setae on exopodite. Rome (1962) drew quite well-developed an-terior setae on Fu, but actually both observed specimens from Lake Tanganyika have very small, even claw-like setae. However, there are some differences between the two populations and they are as follows: appearance of the furcal trunks (Figs 49, 52), length of external seta on terminal segment of Md palp (Figs 48, 53), absence/presence of posterior seta on the third segment on A1, L of claws on terminal segment A1, absence/presence of a long seta on same segment (Figs 51, 54).
DISCUSSION
Candonopsis (Abcandonopsis) aula subgen. nov., sp. nov. and C. (A.) indoles subgen. nov., sp. nov. are very similar and, because of that, probably very closely re-lated. The main difference between them is the morphol-ogy of Fu. Namely, the first species completely lacks a posterior furcal claw, while the latter one has both claws well-developed. Unfortunately, the males were already disintegrated when collected, and it is therefore impos-sible to compare the morphology of their soft parts and carapace. Also, two females of C. (A.) indoles are im-mature and some morphological differences between this species and C. (A.) aula may be characters that will change in the last moult. One example is the short G2 claw in C. (A.) indoles (Fig. 20) which will probably, after the last moult, become as long as the claws G1 and G3, what is the case with all the other Candonopsis species, including C. (A.) aula (Fig. 8). It is also pos-sible that, until reaching the adult stage C. indoles will undergone some changes in the carapace shape, but the number of setae on all the appendages will most prob-ably stay the same. Unfortunately, the developmental pattern of Candoninae is poorly known. The third rep-resentative of the new subgenus, and its type species, is Candonopsis (Abcandonopsis) williami Karanovic and Marmonier, 2002, which is endemic to the subterranean waters of the Murchison Region, Western Australia (Ka-ranovic and Marmonier 2002). This species has both fur-cal claws well developed and, because of that, it is more similar to C. (A.) indoles than to C. (A.) aula. However, C. williami can be separated from C. indoles by the shape of the carapace (dorsal margin is more rounded), and the appearance of the hemipenis (“a” lobe is more triangular). The Murchison species can be separated
from the both new species also by having antero-distal seta on the first segment of A1, slightly more elongated terminal segment of Md palp, two setae externally (in-stead of one in both new species), and five setae distally (instead of three in both new species) on the penultimate segment of the same appendage.
The following character set separates Abcandonop-sis from the nominal subgenus: 1. valves of females are strongly asymmetrical; 2. only one Ta seta is present; 3. there is no basal seta on the T2 protopodite; 4. only two setae are present on the basal segment of T3; 5. Tf seta is missing. On the other hand, Abcandonopsis is very similar to all Candonopsis species in the general appearance of the hemipenis, prehensile palps and Md. A single Ta seta on the protopodite T1 is recorded in an-other two Australian subterranean species, Candonopsis (C.) kimberleyi Karanovic and Marmonier, 2002 and C. (C.) westaustraliensis Karanovic and Marmonier, 2002. They both have an isolated position in the subgenus because the first species has a strongly ornamented cara-pace, while the latter has a reduced posterior furcal claw. The reduction of furcal parts, in many different ways, seems to be common in the genus Candonopsis. First of all, all species lack the posterior furcal seta. Cando-nopsis (C.) thienemanni Schäfer, 1945, described from the subterranean waters of Greece (Schäfer 1945), has transformed the posterior furcal claw into a long seta, while C. (C.) urmilae Gupta, 1988, known from the subterranean waters of India (Gupta 1988) has a very short furcal ramus, compared with the furcal claws. And, finally, the two Australian species are the most extreme examples with the posterior furcal claw reduced into a short seta (C. westaustraliensis), or posterior furcal claw completely missing (C. aula). Obviously, all fur-cal reductions (except the absence of the posterior furcal seta) are autapomorphic in the genus Candonopsis and cannot be features at the generic level, like in some other Candoninae. Also, these reductions could be explained as stygomorphies, because all the above mentioned spe-cies are subterranean dwellers.
While each subterranean species in the genus Can-donopsis has at least one prominent character by which it is easily distinguished from its congeners, surface water species are more difficult from a taxonomic point of view. They all have more or less reniform and smooth carapaces, all setae on the appendages (typical for the genus) are present, only the posterior furcal seta being absent. The features most often used for their discrimi-nation are details of the hemipenis morphology, and presence of spines on the furcal claws. The first one is of no help when the species reproduce parthenogenetically, which is often the case in this genus. The latter charac-ter was the basis for Klieʼs (1932) division of the genus Candonopsis into the following three groups of species: 1. both furcal claws carry one prominent spine; 2. both claws without a spine; 3. only the posterior furcal claw has a prominent spine. In Klieʼs time groups even had separate geographic distribution, i.e. species from the

106 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 107
first and the last groups were known only from Africa and Australia, while species from the second group were restricted to Europe and the Americas. Later on Brood-bakker (1983) described Candonopsis (C.) hummelincki from Central and South America, which has prominent spines on both furcal claws. Also, Karanovic and Mar-monier (2002) found in Australia two species without any furcal spines, and three with variable degree and sexual dimorphism in the development of spines on one or both furcal claws. This was the main reason why the same authors synonymized Candonopsis fessleri Löf-fler, 1968 with C. nama Daday, 1913, and questioned the validity of several other Candonopsis species, one of which is Candonopsis (C.) calva Harding, 1962. This species was described only from females, from a fish gut in the Solomon Islands (Harding 1962). Candonopsis calva is characterized by the reniform carapace and a spine prominent only on the posterior furcal claw. After examining its type material, this species is here synonymized with Candonopsis (C.) sumatrana Klie, 1932, because there is actually not a single feature that could separate C. calva from C. sumatrana. Another species, very similar to C. sumatrana is Candonopsis (C.) tenuis (Brady, 1886). However, these two species can be separated by the shape of the valves (females of C. sumatrana have dorsal margin evenly rounded, while females of C. tenuis have more elongated valves, and one flat part on the dorsal margin), and the structure of the furcal claws (both claws have one prominent spine in C. tenuis). Except from Australia, where it reproduces sexually, C. tenuis is also known from two parthenoge-netic populations from Sumatra (Klie, 1832) and West-ern Samoa (Victor and Fernando 1978). Material of the last record is deposited in the Canadian Museum of Nature and it was recently kindly loaned to me for examination. The specimens from Western Samoa have a more reniform carapace, and a spine prominent only on the posterior furcal claw. These characteristics add Victor and Fernandoʼs (1978) record to the synonymy of C. sumatrana. Unfortunately, I was unable to check Klieʼs (1932) material from Sumatra. Variability in the appearance of carapace and furcal claws has not been recorded in the Australian populations of C. tenuis, nor in the populations of C. sumatrana and, because of that, there is no reason to treat these geografically separated populations other than as separate species. The follow-ing three species also have a spine only on the posterior furcal claw: C. (C.) nama, C. (C.) bujukensis Löffler, 1968, and C. (C.) putealis Klie, 1932. They can be sepa-rated from C. sumatrana by the structure of hemipenis (all three species) and details of the carapace (the second and the last one). Candonopsis nama and C. bujukensis are only known from Africa (Daday 1913; Löffler 1968), while C. putealis was described from Sumatra (Klie 1932) and later recorded from south-west India (Victor and Fernando 1979). The last record is most probably erroneous, because this species has a reniform carapace, while C. putealis has trapezoidal valves. Victor and
Fernando (1979) recorded only a parthenogenetic popu-lation, so details of the hemipenis remain unknown. It seems that the Indian species has spines on both furcal claws, although this character is not properly described by the authors. The carapace shape and details of the furca resemble C. (C.) navicula, described from Kenya (Daday 1910). However, C. putealis cannot be added to the synonymy of the later species until the original mate-rial is checked. On the other hand, after rechecking the type material of two Tanganyikan species (Rome 1962), Candonopsis (C.) depressa Rome, 1962 and C. (C.) dor-sorecta Rome, 1962, they are both synonymized with C. navicula. Three main features of this species are: 1. anterior furcal claw with a very small, claw-like seta; 2. very short Te, Tf, and Tg setae on the third thoracopod; 3. widely rounded posterior margin on carapace, and dorsal margin sloping down towards the anterior one. The first feature was probably overlooked by Daday (1910), because he drew the furca without an anterior seta, while Rome (1962) erroneously drew long setae in both Tanganyikan species. Unfortunately, Daday (1910) reported only juvenile males from Kenya, while they are never recorded in the Lake Tanganyika.
The genera Candonopsis, Cubacandona Brood-bakker, 1983, and Caribecandona Broodbakker, 1983 have very similar appearance of Zenkerʼs organ, Fu, T3, T2, T1, which makes them very closely related and, on the other hand, distinct from the rest of the Candoninae. The three genera are included in the tribe Candonopsini trib. nov., which is mainly distributed in the Southern Hemisphere. Candonopsini is, according to its primitive morphology, probably one of the oldest living tribes in the subfamily. Among the three genera, Candonopsis is the most diverse one and, after the revision done in this paper, the following 24 species belong to it (in alpha-betical order): Candonopsis (Abcandonopsis) aula, C. (A.) indoles, C. (A.) williami, C. (Candonopsis) africana Klie, 1944, C (C.) anteroarcuata Rome, 1962, C. (C.) boui Danielopol, 1978, C. (C.) bujukensis, C. (C.) dani Karanovic and Mamronier, 2002, C. (C.) hummelincki, C. (C.) kingsleii (Brady and Roberston, 1870), C. (C.) kimberleyi, C. (C.) mareza Karanovic and Petkovski, 1999, C. (C.) murchisoni Karanovic and Marmoneir, 2002, C. (C.) nama, C. (C.) navicula, C. (C.) putealis, C. (C.) scourfieldi Brady, 1910; C. (C.) solitaria Vávra, 1895; C. (C.) sumatrana, C. (C.) tenuis, C. (C.) thien-emanni, C. (C.) trichota Schäffer, 1945, C. (C.) urmilae, C. (C.) westaustraliensis. The genus Cubacandona has the following species: Cubacandona cubensis (Danielo-pol, 1978), C. anisitsi (Daday, 1905) new combination, C. columbiensis (Méhes, 1913) new combination, and C. falclandica (Vávra, 1898) new combination. Only three species belong to the genus Caribecandona: Caribecan-dona trapezoidea Broodbakker, 1983, C. auricularia Broodbakker, 1983, and C. ansa Broodbakker, 1983. Caribecandona should be, according to Krstic and Guan (2000), a member of the tribe Lineocyprini Schneider, 1960 together with Mixtacandona Sywula, 1970 and

106 I. Karanovic A revision of Candoninae (Ostracoda): on the genus Candonopsis 107
Danielocandona Broodbakker, 1983 (beside two fossil genera). However, the three living genera have so differ-ent soft parts that this systematic position of the genus Caribecandona cannot be accepted. Krstic and Guan (2000) also placed the genus Candonopsis into so-called tribe “C”. They gave a new proposal for the systemat-ics of the subfamily Candoninae, dividing it into the 20 tribes. However, without assigning a type genus for each tribe, and naming it according to the International Code of the Zoological Nomenclature, this revision cannot be further discussed.
ACKNOWLEDGEMENTS
My sincere thanks are extended to Ms Judith Price (Canadian Museum of Nature), Dr Karel Wouters (Belgium Institute of Natural Science) and Mr Andrew Cabrinovic (British Museum), who kindly loaned me the material of various Candonopsis species deposited in the collections of their Institutions. I am thankful to Dr Mark Hervey (Western Australian Museum) for read-ing the first version of the manuscript, Dr Claude Meisch (Natural History Museum, Luxembourg) and another anonymous referee for reviewing the manuscript and suggesting important improvements. The Australian Biological Resources Study is acknowledged for the grant for the project “Revision of the subfamily Can-doninae (Crustacea, Ostracoda, Candonidae)”, of which the present paper is a part.
REFERENCES
Baird, W. 1845. Arrangements of the British Entomost-raca, with a list of species particularly noticing those which have as yet been discovered within the bounds of the club. Transactions of the Berwickhire Natural-ists ̓Club 2 : 145-158.
Brady, G. S. 1886. Notes on freshwater Entomostraca from South Australia. Proceedings of the Scientific Meetings of the Zoological Society of London 1886 (1): 82-93.
Brady, G. S., D. Robertson. 1870. The Ostracoda and Foraminifera of tidal rivers. The Annals and Maga-zine of Natural History 4 : 1-33.
Broodbakker, N. W. 1983. The subfamily Candoninae (Crustacea, Ostracoda) in the West Indies. Bijdragen tot de Dierkunde 53: 287-326.
Broodbakker, N. W., D. L. Danielopol, D. L. 1982. The chaetotaxy of Cypridacea (Crustacea, Ostracoda) limbs: proposal for a descriptive model. Bijdragen tot de Dierkunde 52: 103-120.
Daday, J. 1905. Untersuchungen uber die Süsswasser-Mikrofauna Paraguays. Zoologica 44: 1-374.
Daday, J. 1910. Die Süsswasser-Mikrofauna Deutsch-Ost-Afrikas. Zoologica 59: 1-314.
Daday, J. 1913. Cladoceren und Ostracoden aus Sud-
und Sudwestafrika. Denkschriften der Medizinisch-Naturwissenschaftlichen Gesellschaft zu Jena 70: 92-102.
Danielopol, D. L. 1969. Recherches sur la morphologie de lʼorgane copulateur mâle chez quelques ostra-codes du genre Candona Baird (fam. Cyprididae Baird). Pp. 136-153 in J. W. Neale, ed. The taxon-omy, morphology and ecology of recent Ostracoda. Edinbourgh: Oliver & Boyd Ltd.
Danielopol, D. L. 1980. Deux espèces hypogées du genre Candonopsis (Ostracoda, Candoninae) du Sud de la France et de Cuba. Vie et Milieu 30: 315-323.
Gupta, L. P. 1988. Candonopsis urmilae a new species of subterranean Crustacea (Ostracoda: Candonidae) from India. Records of the Zoological Survey of In-dia 85: 419-427.
Harding, J. P. 1962. Mungava munda and four other new species of Ostracod Crustaceans from fish stomachs. The Natural History of Rennell Island, British Solo-mon Islands 4: 51-62.
Karanovic, I., P. Marmonier. 2002. On the genus Cando-nopsis (Crustacea, Ostracoda, Candoninae) in Aus-tralia, with a key to the world recent species. Annales de Limnologie 38: 1999-240.
Klie, W. 1932. Die Ostracoden der Deutschen Limnolo-gischen Sunda-Expedition. Archiv fur Hydrobiol-ogie, Suppl.-Bd. 11: “Tropische Binnengewasser”, Band 3: 447-502.
Klie, W. 1944. Ostracoda. Exploration du Parc National Albert, Mission H. Damas. Institut des Parcs Nation-aux du Congo Belge 12: 1-62.
Krstic, N., S.-z. Guan. 2000. A proposal for the system-atics of the subfamily Candoninae (Ostracoda) with the description of the Macedocandona, new genus. Geologica Macedonica 14: 25-48.
Löffler H. 1968. Die Crustaceenfauna der Binnenge-wasser Ostafrikanischer Hochberge. Hochgebirgs-forschung 1: 107-170.
Martens, K. 1987. Homology and functional morphol-ogy of the sexual dimorphism in the antenna of Sclerocypris Sars, 1924 (Crustacea, Ostracoda, Megalocypridinae). Bijdragen tot de Dierkunde 57: 183-190.
Martens, K. 1998. General morphology of non-marine ostracods. Pp. 57-75 in K. Martens, ed. Sex and Par-thenogenesis: evolutionary ecology of reproductive modes in non-marine ostracods. Leiden: Backhuys Publishers.
Méhes, G. 1913. Süsswasser-Ostracoden aus Columbien und Argentinien. Mémoires de la Société neuchate-loise des Sciences naturelles 5: 639-663.
Meisch, C. 1996. Contribution to the taxonomy of Pseudocandona and four related genera, with the description of Schellencandona nov. gen., a list of the Candoninae genera, and a key to the European genera of the subfamily (Crustacea, Ostracoda). Bul-letin de la Société des Naturalistes luxembourgeois 97: 211-237.

108 I. Karanovic
Rome, R. 1962. Ostracodes. - Exploration Hydro-biologique du Lac Tanganika (1946-1947). Resultats Scientifique 3: 1-305.
Sars, G. O. 1896. On fresh-water Entomostraca from the neighbourhood of Sydney, partly raised from dried mud. Archiv for Mathematik og Naturvidenskab 18: 1-81.
Schäfer, H. W. 1945. Grundwasser-Ostracoden aus Griechenland. Archiv fur Hydrobiologie 40: 847-866.
Vávra, W. 1891. Monographie der Ostracoden Bohm-ens. Archiv der Naturwissenschaftl Landesdurchfor-
schung von Böhmen, 8: 1-116. Vávra, V. 1895. Die von Dr F. Stuhlmann gesammelten.
Süsswasser-Ostracoden Zanzibar s̓. Jahrbuch der Ham-burgischen Wissenschaftlichen Anstalten 12: 1-23.
Vávra, V. 1898. Süsswasser-Ostracoden. Hamburger Magalhaensische Sammelreise: 1-25.
Victor, R., C.H. Fernando. 1978. Systematics and eco-logical notes on Ostracoda from container habitats of some South Pacific Islands. Canadian Journal of Zoology 56: 414-422.
Victor, R., C.H. Fernando. 1979. The freshwater ostra-cods (Crustacea, Ostracoda) of India. Records of the Zoological Survey of India 74: 147-242.




















