
21 OSTRACODA - SEDICI
26
OSTRACODA 599 OSTRACODA LOS INVERTEBRADOS FÓSILES 21 Cecilia Laprida Sara Ballent OSTRACODA INTRODUCCIÓN Los ostrácodos constituyen una clase de crus- táceos de hábitos acuáticos y morfológicamen- te muy conservadores, fácilmente identificables por su pequeña talla y la presencia de un exoesqueleto o caparazón bivalvo, de naturale- za quitinoso-calcárea, que cubre completamen- te las partes blandas. El tamaño de un ostrácodo adulto oscila entre los 0,15 y los 2-3 mm, si bien algunas especies de agua dulce alcanzan los 8 mm; las de Gigantocypris miden hasta 32 mm y algunas paleozoicas del género Leperditia, llega- ron a los 50 mm. El caparazón está compuesto por carbonato de calcio en forma de calcita con proporciones variables de magnesio, recubierto externamen- te por una capa de quitina. Lo integran piezas o valvas laterales, ubicadas a ambos lados del cuerpo y articuladas dorsalmente mediante una charnela. En caso de agresión o perturbación, el individuo se retrae completamente en el capa- razón, el cual se cierra mediante la acción de músculos aductores. En la actualidad son organismos frecuentes en todos los medios acuáticos. Se calcula que exis- ten cerca de 33000 especies entre vivientes y fósiles, aunque algunos autores elevan esta ci- fra a 55000. De ellas, 5000 fueron descriptas vi- viendo en lagos, lagunas, humedales, charcos temporarios, ambientes subterráneos, margi- nales, marinos costeros y de mar abierto, hasta los lagos de los altiplanos de todo el mundo. Los ostrácodos son los artrópodos más abun- dantes del registro fósil. Dados la calcificación y el pequeño tamaño, las valvas poseen un alto potencial de fosilización, y su hallazgo en las rocas sedimentarias y los sedimentos de grano fino (arenas y limos) es relativamente frecuen- te. El primer registro fehaciente de ostrácodos fósiles corresponde al Ordovícico, o sea, aproxi- madamente hace unos 500 m.a. En el Cámbrico Superior existen registros de artrópodos con un caparazón bivalvo de naturaleza fosfática, los Phosphatocopina y los Bradoriida que han sido relacionados con los ostrácodos, pero los estudios morfológicos de las partes blandas de ejemplares preservados excepcionalmente, per- mitirían afirmar que no serían verdaderos os- trácodos (Hou et al., 1996; Siveter et al., 2001). De- bido a sus reducidas dimensiones, los ostráco- dos son objeto de estudio de la Micropaleontolo- gía. Esto implica técnicas especiales de muestreo, preparación y observación. Dentro de esta disci- plina, conforman uno de los grupos de mayor importancia, por lo que numerosos investigado- res están dedicados a su estudio. Históricamen- te, la importancia de estos artrópodos se basó en sus aplicaciones bioestratigráficas asociadas a la industria del petróleo pero, en la actualidad, la mayoría de las investigaciones se desarrollan en centros académicos y se hallan más relaciona- das con estudios paleoecológicos, paleogeográfi- cos y paleoclimáticos. ANATOMÍA PLANO CORPORAL El cuerpo de los ostrácodos es alargado y com- primido lateralmente. Su pared consta de una capa externa quitinosa y otra interna epidér- mica. No hay tagmatización evidente aunque sí un pequeño estrechamiento por detrás de la ca- beza que marca el límite entre el céfalo y el tórax (Figura 21. 1). Los apéndices del céfalo y el tó- rax, constituidos por podómeros, son birrámeos (con endopodito y exopodito) a excepción de la anténula que es unirrámea. El abdomen, com- pletamente reducido y fusionado al tórax, cul- mina mediante un par de furcas no articuladas, de gran importancia sistemática. SISTEMA DIGESTIVO Consta de la cavidad bucal flanqueada por un par de mandíbulas, el esófago que desem-
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of 21 OSTRACODA - SEDICI

OST
RA
CO
DA
599OSTRACODALOS INVERTEBRADOS FÓSILES
21
Cecilia LapridaSara Ballent
OSTRACODA
INTRODUCCIÓN
Los ostrácodos constituyen una clase de crus-táceos de hábitos acuáticos y morfológicamen-te muy conservadores, fácilmente identificablespor su pequeña talla y la presencia de unexoesqueleto o caparazón bivalvo, de naturale-za quitinoso-calcárea, que cubre completamen-te las partes blandas. El tamaño de un ostrácodoadulto oscila entre los 0,15 y los 2-3 mm, si bienalgunas especies de agua dulce alcanzan los 8mm; las de Gigantocypris miden hasta 32 mm yalgunas paleozoicas del género Leperditia, llega-ron a los 50 mm.
El caparazón está compuesto por carbonatode calcio en forma de calcita con proporcionesvariables de magnesio, recubierto externamen-te por una capa de quitina. Lo integran piezas ovalvas laterales, ubicadas a ambos lados delcuerpo y articuladas dorsalmente mediante unacharnela. En caso de agresión o perturbación, elindividuo se retrae completamente en el capa-razón, el cual se cierra mediante la acción demúsculos aductores.
En la actualidad son organismos frecuentes entodos los medios acuáticos. Se calcula que exis-ten cerca de 33000 especies entre vivientes yfósiles, aunque algunos autores elevan esta ci-fra a 55000. De ellas, 5000 fueron descriptas vi-viendo en lagos, lagunas, humedales, charcostemporarios, ambientes subterráneos, margi-nales, marinos costeros y de mar abierto, hastalos lagos de los altiplanos de todo el mundo.
Los ostrácodos son los artrópodos más abun-dantes del registro fósil. Dados la calcificacióny el pequeño tamaño, las valvas poseen un altopotencial de fosilización, y su hallazgo en lasrocas sedimentarias y los sedimentos de granofino (arenas y limos) es relativamente frecuen-te. El primer registro fehaciente de ostrácodosfósiles corresponde al Ordovícico, o sea, aproxi-madamente hace unos 500 m.a. En el CámbricoSuperior existen registros de artrópodos conun caparazón bivalvo de naturaleza fosfática,los Phosphatocopina y los Bradoriida que han
sido relacionados con los ostrácodos, pero losestudios morfológicos de las partes blandas deejemplares preservados excepcionalmente, per-mitirían afirmar que no serían verdaderos os-trácodos (Hou et al., 1996; Siveter et al., 2001). De-bido a sus reducidas dimensiones, los ostráco-dos son objeto de estudio de la Micropaleontolo-gía. Esto implica técnicas especiales de muestreo,preparación y observación. Dentro de esta disci-plina, conforman uno de los grupos de mayorimportancia, por lo que numerosos investigado-res están dedicados a su estudio. Históricamen-te, la importancia de estos artrópodos se basó ensus aplicaciones bioestratigráficas asociadas ala industria del petróleo pero, en la actualidad, lamayoría de las investigaciones se desarrollan encentros académicos y se hallan más relaciona-das con estudios paleoecológicos, paleogeográfi-cos y paleoclimáticos.
ANATOMÍA
PLANO CORPORAL
El cuerpo de los ostrácodos es alargado y com-primido lateralmente. Su pared consta de unacapa externa quitinosa y otra interna epidér-mica. No hay tagmatización evidente aunque síun pequeño estrechamiento por detrás de la ca-beza que marca el límite entre el céfalo y el tórax(Figura 21. 1). Los apéndices del céfalo y el tó-rax, constituidos por podómeros, son birrámeos(con endopodito y exopodito) a excepción de laanténula que es unirrámea. El abdomen, com-pletamente reducido y fusionado al tórax, cul-mina mediante un par de furcas no articuladas,de gran importancia sistemática.
SISTEMA DIGESTIVO
Consta de la cavidad bucal flanqueada porun par de mandíbulas, el esófago que desem-

600 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
boca en un estómago alargado, al que siguen elintestino largo y musculoso y un recto que cul-mina en el ano. El alimento ingresa a la cavi-dad bucal luego de haber sido procesado físi-camente por mandíbulas y maxilas. La mayorparte del proceso digestivo ocurre en el estó-mago, que recibe las secreciones de una glán-dula, el hígado, ubicado en la zona de duplica-dura, en estrecha relación con las valvas.
SISTEMAS CIRCULATORIO YRESPIRATORIO
La mayoría de los ostrácodos carece de un sis-tema circulatorio definido y tampoco presen-tan un sistema respiratorio diferenciado. El in-tercambio gaseoso se efectúa a través de la pa-red del cuerpo por simple difusión. Los exopo-ditos de las maxilas han sido interpretadoscomo superficies de intercambio respiratorio.Algunos autores consideran que los tejidos blan-dos que ocupan el vestíbulo, un espacio interiorsituado cerca del margen de las valvas, partici-parían del proceso de intercambio gaseoso.
SISTEMA NERVIOSO
El órgano más conspicuo del sistema nervio-so es un cerebro de posición subcentral.Adicionalmente existe un anillo circunesofá-gico y una cadena ventral de pares de gangliosfusionados que recorre longitudinalmente el
cuerpo. Ganglios más pequeños, conectados alsistema central por nervios secundarios,inervan a todos los apéndices cefálicos.
ÓRGANOS DE LOS SENTIDOS
Gran número de ostrácodos carece por com-pleto de ojos, pero la mayoría posee 1 ojo sim-ple de posición mediana, el ojo nauplius, si-tuado entre las anténulas, justo por debajo delmargen dorsal de las valvas. Además, todo elcuerpo está cubierto por pequeñas sedas cor-tas sensoriales que cruzan las valvas a travésde poros y que son las encargadas de mantenerel contacto con el medio exterior, aun cuandoel caparazón se halle cerrado. Estas sedas sonparticularmente abundantes en la región an-terior del cuerpo, es decir, en la extremidadcefálica.
APÉNDICES
El rasgo morfológico más prominente, y el quepresenta mayor afinidad con el resto de loscrustáceos, es la presencia de apéndices paresarticulados (Figura 21. 1). Los ostrácodos por-tan apéndices tanto en el céfalo como en el tó-rax. Con excepción de la anténula, todos sonbirrámeos, aunque el exopodito puede estar re-ducido. La cabeza lleva 4 pares de apéndices:antenas I o anténulas (A1), antenas II (A2),mandíbulas (Md) y maxilas (Mx).
Figura 21. 1. Morfología interna de Cypridopsis vidua (Podocopida, Cypridoidea), no marino. Vista lateral interna de ejemplar femenino,valva izquierda removida (modificado de Van Morkhoven, 1959).

OST
RA
CO
DA
601OSTRACODALOS INVERTEBRADOS FÓSILES
Las anténulas o primer par de apéndicescefálicos son estructuras largas y acintadas quecumplen funciones sensoriales y motoras, uti-lizándolas en la natación o en la reptación, de-pendiendo del hábito de vida. Las antenas IIconstituyen el órgano de locomoción más im-portante. Son empleadas coordinadamente conlas A1 para nadar, o con el primer par de patastorácicas para reptar. Eventualmente, partici-pan en la alimentación. En los machos, la A2puede actuar como órgano de sujeción de lahembra en el proceso de cópula. Las mandíbu-las se ubican a ambos lados de la cavidad bu-cal y tienen funciones exclusivamente masti-catorias. El podómero basal del protopoditoestá muy quitinizado, y suele presentar fuer-tes crenulaciones que intervienen en la parti-ción mecánica del alimento. El cuarto par deapéndices cefálicos, las maxilas, yace por de-trás de las mandíbulas. Son apéndices fuerte-mente modificados: los artejos correspondien-tes al endopodito forman un palpo de funcio-nes masticatorias, mientras que el exopoditocumple funciones respiratorias. Por ello estáexpandido como una amplia placa branquialcon numerosas sedas en el margen posterior,que se extienden lateralmente al cuerpo y estánconstantemente en movimiento, asegurando lacirculación del agua dentro del caparazón.
El tórax lleva 1 a 4 pares de patas, de funcio-nes y morfologías variadas. El abdomen, fu-sionado al tórax, carece de apéndices y culmi-na en un par de furcas.
El primer par de patas torácicas presentamorfologías muy variadas en los distintos gru-pos. En algunos actúa como órgano de locomo-
ción. Otras veces se modifica para cumplir fun-ciones alimenticias (maxilipedios) o, en losmachos de algunas especies, para participaren la reproducción como órgano de sujeción dela hembra durante la cópula. El segundo parde patas es muy variable: puede ser idéntico alprimer par y participar activamente en la lo-comoción, adoptar forma de una placa bran-quial interviniendo en la respiración, e inclusotener funciones sensoriales asociadas a la re-producción. En algunos grupos el segundo parde patas falta por completo. Del mismo modo,el tercer par de patas puede ser morfológica-mente idéntico al segundo par, actuando igual-mente en la locomoción pero, en otros grupospuede estar altamente modificado y contribuiren la limpieza del interior del caparazón. Enalgunos casos, al igual que el segundo par depatas, está completamente ausente. Un cuartopar de patas es identificable en un único grupoviviente, la superfamilia Puncioidea.
MÚSCULOS ADUCTORES
En la mitad anterior del cuerpo, existe un pa-quete de musculatura estriada que lo atravie-sa de lado a lado: son los músculos aductores.Las fibras musculares son perpendiculares alplano de unión de las valvas y se unen a sucara interna por lo que, cuando se contraen, elcaparazón se cierra (Figura 21. 2 A). Aldistenderse los músculos, las valvas tienden aabrirse como consecuencia de la contracciónde un ligamento elástico dorsal. En el punto deunión entre cada fibra muscular y el capara-
Figura 21. 2. A. esquema del corte transversal de un ostrácodo al nivel de los músculos aductores; B. esquema de detalle de la regiónperiférica anterior de la valva izquierda (modificado de Van Morkhoven, 1959).

602 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
zón, existe una pequeña hendidura que se co-noce con el nombre de impresión muscular. Elconjunto de las impresiones musculares delaductor, ubicado en la parte ántero-medianavalvar, presenta un diseño característico querefleja el número y disposición de las fibrasmusculares, constituyendo un rasgo de granimportancia sistemática y para la orientaciónde las valvas (Figura 21. 3).
REPRODUCCIÓN Y DESARROLLO
Los ostrácodos poseen sexos separados y fe-cundación interna. Los órganos reproductores
se hayan duplicados. En las hembras existendos aberturas vaginales ubicadas por delantede las furcas y por detrás del tercer par de pa-tas torácicas. Los órganos internos incluyenun par de ovarios y oviductos que concentranlos huevos fecundados y los dirigen hacia unpar de aberturas uterinas, a través de las cua-les son eliminados al exterior. Los ovarios pue-den observarse en el interior de las valvas,donde suelen dejar débiles impresiones. En losmachos existe un complejo órgano copulatorioo pene de naturaleza quitinosa, ubicado entreel tercer par de patas torácicas. El pene es degran tamaño y ocupa la mitad posterior delcaparazón. Está formado por dos mitades idén-
Figura 21. 3. Representantes de los órdenes de Ostracoda. Esquemas del aspecto general de los caparazo-nes indicando su orientación y detalle de las impresiones musculares.

OST
RA
CO
DA
603OSTRACODALOS INVERTEBRADOS FÓSILES
ticas, llamadas hemipenes, cuya compleja es-tructura es altamente diagnóstica. La conexiónentre el órgano copulatorio y los testículos esun grupo complejo de espermiductosespiralados. En los Cypridoidea, los ductoseyaculatorios se modifican y forman una es-tructura musculosa, el órgano de Zenker, queactúa como una bomba de esperma. En otrosgrupos (Cytheroidea), la bomba espermáticaestá directamente incorporada a la estructurade los hemipenes.
Existen tres estrategias reproductivas en os-trácodos. La reproducción bisexual implicacópula y fertilización de las hembras por par-te de los machos de una misma población. Lamayor parte de las especies que habitan losambientes marinos se reproducen exclusiva-mente de este modo. Esta reproducción tienelugar también en ambientes continentales, enespecial estables, tales como en la zona másprofunda de los grandes lagos. La reproduc-ción partenogenética (sin la participación demachos) es relativamente frecuente, y existeun grupo (Darwinuloidea) en el cual virtual-mente se desconoce la existencia de individuosmasculinos. La partenogénesis es una estrate-gia reproductiva común, aunque no exclusiva,en ambientes no marinos inestables. Un tercertipo de estrategia es la reproducción mixta, enla cual una especie puede tener reproduccióntanto bisexual como partenogenética, depen-diendo de las condiciones imperantes.
Muchos ostrácodos depositan en el medio loshuevos fecundados, los que quedan a mercedde predadores y expuestos a cambios del am-biente hasta que eclosionan. En otros casos, lashembras retienen los huevos e incluso los pri-meros estadios larvales. En las formaspospaleozoicas, la retención se produce en laparte posterior del caparazón, por lo que enlas hembras dicha región está marcadamenteensanchada. En los extintos Beyrichoidea, lashembras de algunos géneros presentanabultamientos o lóbulos marcados en la parte
ántero-ventral de cada valva, conocidos con elnombre de crúminas, que han sido interpreta-dos como cámaras de cría (Figura 21. 4).
Cuando el huevo eclosiona da lugar a una lar-va nauplius, que ya posee un caparazónbivalvo escasamente calcificado, de menor ta-maño que el adulto. Luego sigue un crecimien-to discontinuo; el caparazón es eliminado pe-riódicamente y reemplazado por otro mayor,conforme a las nuevas dimensiones del cuer-po. Este proceso se denomina muda o ecdisis.Se han descripto hasta 8 estadios larvales en-tre la eclosión y la madurez sexual (Figura 21.5). Dado que existen 8 caparazones juveniles (es-tadios A-8 a A-1) por cada individuo que llega aadulto, la presencia de valvas juveniles es rela-tivamente frecuente en sedimentos de granofino asociados a ambientes de baja energía.
En los medios no marinos, ciertas especieshabitan charcas temporarias, sometidas a de-secación periódica. Estas formas producen hue-vos resistentes capaces de soportar largos pe-ríodos (hasta años) de sequía, eclosionando solo
Figura 21. 4. Dimorfismo sexual en Palaeocopida (Beyrichoidea).Esquemas de caparazones masculino y femenino (con crúminaántero-ventral) (modificado de Van Morkhoven, 1959).
Figura 21. 5. Dimorfismo sexual y ontogenia en Cyprideisriograndensis (Podocopida, Cytheroidea), no marino. Cuaternario,cuenca del Salado. El estadio juvenil A-8 permanece retenido en elinterior de la hembra.
604 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
después de que el ambiente se haya inundadonuevamente y la disponibilidad de agua estéasegurada. La formación de huevos de resis-tencia favorece la colonización de nuevoshábitats y la dispersión de los ostrácodos nomarinos. Pueden ser transportados por el vien-to o por animales.
La existencia de huevos de resistencia y elcuidado de los estadios larvales en linajes ex-clusivamente partenogenéticos, representaríanlas estrategias de dispersión y colonizaciónmás significativas en hábitats no marinos. Entérminos generales, especies con reproducciónpartenogenética pueden colonizar nuevos cuer-pos de agua con mayor facilidad que aquéllasque poseen exclusivamente reproducción bi-sexual. Sin embargo, la colonización involucrano solo capacidad de dispersión, sino tambiéncompetencia con otras especies, circunstanciaen la que la reproducción bisexual puede seruna ventaja.
CAPARAZÓN
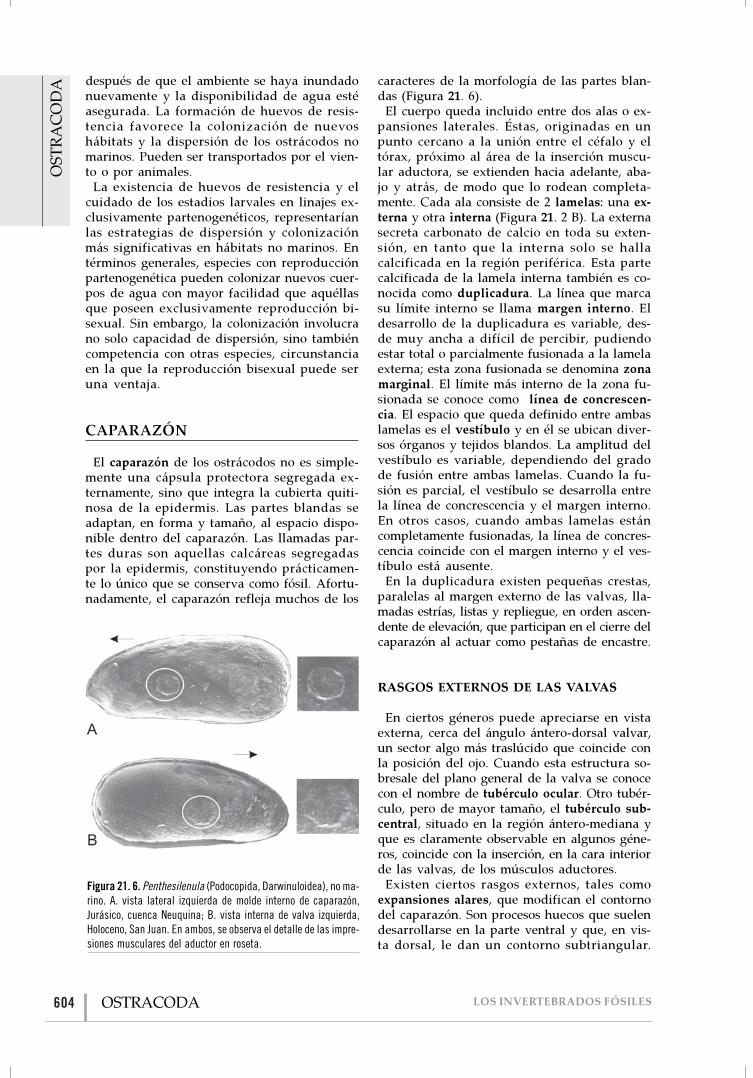
El caparazón de los ostrácodos no es simple-mente una cápsula protectora segregada ex-ternamente, sino que integra la cubierta quiti-nosa de la epidermis. Las partes blandas seadaptan, en forma y tamaño, al espacio dispo-nible dentro del caparazón. Las llamadas par-tes duras son aquellas calcáreas segregadaspor la epidermis, constituyendo prácticamen-te lo único que se conserva como fósil. Afortu-nadamente, el caparazón refleja muchos de los
caracteres de la morfología de las partes blan-das (Figura 21. 6).
El cuerpo queda incluido entre dos alas o ex-pansiones laterales. Éstas, originadas en unpunto cercano a la unión entre el céfalo y eltórax, próximo al área de la inserción muscu-lar aductora, se extienden hacia adelante, aba-jo y atrás, de modo que lo rodean completa-mente. Cada ala consiste de 2 lamelas: una ex-terna y otra interna (Figura 21. 2 B). La externasecreta carbonato de calcio en toda su exten-sión, en tanto que la interna solo se hallacalcificada en la región periférica. Esta partecalcificada de la lamela interna también es co-nocida como duplicadura. La línea que marcasu límite interno se llama margen interno. Eldesarrollo de la duplicadura es variable, des-de muy ancha a difícil de percibir, pudiendoestar total o parcialmente fusionada a la lamelaexterna; esta zona fusionada se denomina zonamarginal. El límite más interno de la zona fu-sionada se conoce como línea de concrescen-cia. El espacio que queda definido entre ambaslamelas es el vestíbulo y en él se ubican diver-sos órganos y tejidos blandos. La amplitud delvestíbulo es variable, dependiendo del gradode fusión entre ambas lamelas. Cuando la fu-sión es parcial, el vestíbulo se desarrolla entrela línea de concrescencia y el margen interno.En otros casos, cuando ambas lamelas estáncompletamente fusionadas, la línea de concres-cencia coincide con el margen interno y el ves-tíbulo está ausente.
En la duplicadura existen pequeñas crestas,paralelas al margen externo de las valvas, lla-madas estrías, listas y repliegue, en orden ascen-dente de elevación, que participan en el cierre delcaparazón al actuar como pestañas de encastre.
RASGOS EXTERNOS DE LAS VALVAS
En ciertos géneros puede apreciarse en vistaexterna, cerca del ángulo ántero-dorsal valvar,un sector algo más traslúcido que coincide conla posición del ojo. Cuando esta estructura so-bresale del plano general de la valva se conocecon el nombre de tubérculo ocular. Otro tubér-culo, pero de mayor tamaño, el tubérculo sub-central, situado en la región ántero-mediana yque es claramente observable en algunos géne-ros, coincide con la inserción, en la cara interiorde las valvas, de los músculos aductores.
Existen ciertos rasgos externos, tales comoexpansiones alares, que modifican el contornodel caparazón. Son procesos huecos que suelendesarrollarse en la parte ventral y que, en vis-ta dorsal, le dan un contorno subtriangular.
Figura 21. 6. Penthesilenula (Podocopida, Darwinuloidea), no ma-rino. A. vista lateral izquierda de molde interno de caparazón,Jurásico, cuenca Neuquina; B. vista interna de valva izquierda,Holoceno, San Juan. En ambos, se observa el detalle de las impre-siones musculares del aductor en roseta.

OST
RA
CO
DA
605OSTRACODALOS INVERTEBRADOS FÓSILES
Ciertos ostrácodos paleozoicos (Beyrichoidea)se caracterizan por la presencia de lóbulos ysurcos (Figura 21. 7). Los lóbulos son elevacio-nes marcadas del caparazón que reflejan laanatomía interna. Existen hasta 4 que se desig-nan L
1, L
2, L
3, L
4 desde la parte anterior a la
posterior, los que se hallan separados por sur-cos, depresiones elongadas denominadas S
1, S
2,
S3. El surco S
2 corresponde al área de inserción
de los músculos aductores y es el único queestá siempre presente. El velo es una estructu-ra amplia que se desarrolla a lo largo del mar-gen libre de la valva.
Adicionalmente a la presencia de tubérculos,lóbulos, surcos, expansiones alares o velos, lasvalvas pueden ser lisas u ornamentadas. Laornamentación consiste de costillas, estrías,surcos, tubérculos, reticulaciones, pequeñasdepresiones puntuales llamadas punctas, o
Figura 21. 7. Esquema de morfología externa de un Palaeocopida,marino, valva izquierda, amplete. Se observan el borde dorsal recto,ventral ampliamente convexo y superficie externa con lóbulos ysurcos. A. vista dorsal; B. vista lateral (modificado de Brasier, 1980).
Figura 21. 8. Esquemas de morfología interna en Podocopida. A. valva derecha de Loxoconcha paranensis (Cytheroidea) marino. Holoceno,Buenos Aires; B. valva derecha de Sclerocypris exserta (Cypridoidea), no marino. Holoceno, Namibia, África (A. modificado de Bertels yMartínez, 1997; B. modificado de Horne et al., 2002).

606 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
combinación de ellas. En las valvas de un mis-mo individuo suele ser idéntica, y es un carác-ter específico.
RASGOS INTERNOS DE LAS VALVAS
La superficie interna de las valvas tambiénpresenta gran cantidad de rasgos que si bienno tan conspicuos como aquéllos de la cara ex-terna, son de gran importancia sistemática. Setrata de una serie de pequeñas marcas obser-vables por transparencia (Figura 21. 8). En elárea subcentral, en ubicación levemente ante-rior, se destacan las impresiones del músculoaductor, marcas pequeñas que corresponden ala inserción de las fibras musculares. Cada fi-bra, al unirse a la cara interna de la valva, dejauna pequeña muesca o cicatriz. También existeotro tipo de impresiones asociadas a la inser-ción de músculos que mueven las mandíbulas(impresiones mandibulares) y las antenas (im-presiones musculares dorsales).
ORIENTACIÓN, FORMA Y DIMENSIONESDE LAS VALVAS
En los ostrácodos se definen una valva dere-cha y otra valva izquierda. Cada una presenta4 márgenes (dorsal, ventral, anterior y poste-rior), los que coinciden con la ubicación del or-ganismo dentro del caparazón (Figura 21. 1).Las valvas se articulan a lo largo del margendorsal, el cual es levemente convexo. Allí seubican un ligamento interior elástico, de na-turaleza orgánica, que las mantiene abiertas,y la charnela, que las articula e impide su des-plazamiento relativo. El margen ventral, quesuele ser flexuoso y levemente cóncavo en suparte media, es subparalelo al margen dorsal.El margen anterior, generalmente redondeado,corresponde a la ubicación de la cabeza. Cuan-do el caparazón está abierto, las antenas se pro-yectan al exterior a través de él. El margen pos-terior, variablemente redondeado o acumina-do, corresponde a la culminación del abdomen.
El alto de la valva es la máxima distancia queexiste entre el margen dorsal y el margen ven-tral, medida perpendicularmente al largo; ellargo es la máxima distancia entre el margenanterior y el posterior, perpendicularmente alalto. Las valvas suelen ser más largas que al-tas, presentando un contorno subelíptico,reniforme o subrectangular, aunque las formasplanctónicas son, en vista lateral, típicamentesubcirculares.
Como regla, si una valva, en vista lateral ex-terna, presenta el borde dorsal hacia la partesuperior y la región anterior hacia la izquier-da, se considera valva izquierda. Si la parteanterior está situada hacia la derecha, es unavalva derecha. La correcta ubicación e identi-ficación de las mismas es de enorme importan-cia para la adecuada descripción y clasifica-ción sistemática.
Los ostrácodos paleozoicos presentan mor-fología levemente diferente, por lo que una ter-minología y criterios especiales son usados enla orientación de las valvas. A diferencia de losostrácodos pospaleozoicos, el borde dorsal sue-le ser recto y el ventral ampliamente convexo.Las valvas se caracterizan por poseer ánguloscardinales bien marcados y pueden distinguir-se por la posición de la mayor altura respectoa la longitud de las mismas. Se las denominapreplete, posplete o amplete según que la máxi-ma altura esté por delante, detrás o sea coinci-dente con la mitad de la longitud, respectiva-mente. Muchos taxones paleozoicos presentanlóbulos y surcos marcados. En estos casos, elsurco S, que indica la posición del músculoaductor, tiene una posición media anterior ysu parte inferior está dirigida hacia adelante;el lóbulo más desarrollado indica generalmen-te la parte posterior del caparazón, mientrasque el velo es siempre ventral (Figura 21. 7).
En las formas pospaleozoicas fósiles, la parteanterior del caparazón es generalmente redon-deada; las impresiones musculares del aductorestán situadas en la parte media anterior de lavalva, y las impresiones mandibulares (cuan-do se observan) tienen una posición ántero-ventral; el tubérculo ocular es ántero-dorsal ylas espinas destacadas y expansiones alaresventro-laterales están dirigidas hacia atrás;asimismo, en vista dorsal, el caparazón es máscomprimido hacia la parte anterior.
En el material fósil, sobre todo en caparazo-nes donde no pueden observarse los rasgos in-ternos, la distinción entre valva derecha e iz-quierda es a veces difícil, por lo cual se recurreexclusivamente a los rasgos externos.
CANALES DE POROS MARGINALESY POROS NORMALES
La zona marginal suele estar atravesada poruna serie de canales largos a través de los cua-les se proyectan hacia el exterior sedas sensiti-vas. Los mismos, desarrollados en el plano defusión de ambas lamelas, reciben el nombre decanales de los poros marginales. Si las lamelasno están fusionadas, los canales de los poros

OST
RA
CO
DA
607OSTRACODALOS INVERTEBRADOS FÓSILES
marginales son muy cortos y se desarrollan enel margen mismo de las valvas pudiendo serrectos o curvos, simples o bifurcados, de diá-metro constante o ensancharse en algún puntode su recorrido (Figura 21. 8). En el margenanterior, donde la duplicadura generalmentees más ancha, los canales de los poros margi-nales suelen ser más largos y numerosos, loque constituye una ayuda adicional para ladeterminación del margen anterior.
Existe otro tipo de poros que atraviesa lalamela externa: los poros normales, que soncanales cortos perpendiculares al plano de lasvalvas. A través de ellos se libera la secreciónde ciertas glándulas y se proyectan hacia elexterior sedas sensitivas cortas que el animalposee en la superficie del cuerpo.
CHARNELA
Las valvas están articuladas dorsalmentemediante una charnela. Ésta consta de una se-rie de elementos calcificados diferenciados (ba-
rras y surcos, dientes y fosetas) dispuestos a lolargo del margen dorsal, que actúan como bi-sagra. Los elementos charnelares de ambasvalvas son complementarios: si en la izquier-da hay una foseta y una barra, en la mismaubicación de la derecha habrá un diente y unsurco. Se reconocen tres tipos básicos de char-nela (Figura 21. 9), de acuerdo al número y na-turaleza de elementos que la conformen. En lacharnela adonta, existe un único elemento alar-gado, una barra o surco, que puede o no estarcrenulado. La charnela merodonta lleva 3 ele-mentos: anterior (diente o foseta), mediano alar-gado (barra o surco) y posterior (diente ofoseta), los cuales pueden ser lisos o crenulados.En la charnela anfidonta se reconocen 4 ele-mentos constitutivos, como consecuencia de lamodificación del extremo anterior del elemen-to mediano. Charnelas más complejas puedenconsiderarse variantes de esta última.
En algunos grupos (por ejemplo Myodocopida)la articulación se realiza simplemente por unligamento elástico situado en el margen dorsalde las valvas.
DIMORFISMO SEXUAL
En muchas especies, los caparazones de ma-chos y hembras son diferentes en forma y ta-maño. Estas diferencias, notables ya en los úl-timos estadios ontogenéticos, se deben al de-sarrollo de los órganos reproductores en losmachos, y a la presencia de cámaras o espaciosde cría en las hembras. En algunos ostrácodospaleozoicos, como los Beyrichoidea, este dimor-
Figura 21. 9. Esquemas en vista lateral y dorsal de diferentes tiposde charnela (modificado de Brasier, 1980).
Figura 21. 10. Dimorfismo sexual en Podocopida. A. Huantraiconellaprima (Cytheroidea), marino, vista externa de valvas derechas.Paleoceno, cuenca Neuquina; B. Amphicypris argentinensis(Cypridoidea), no marino, vista externa de valvas izquierdas. Holo-ceno, Buenos Aires.

608 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
fismo sexual es muy notorio debido a la pre-sencia de crúminas en las hembras (Figura 21.4). En las especies en las que no existen cáma-ras de cría diferenciadas, los huevos y los pri-meros estadios larvales son retenidos en laparte posterior del caparazón de las hembras,por lo que dicha zona es más abultada y anchaque en los machos. Ésto es particularmente evi-dente en vista dorsal. Por su parte, los capara-zones machos suelen ser más elongados quelos femeninos, como consecuencia del gran ta-maño relativo de los hemipenes (Figura 21. 10).
VARIABILIDAD INTERESPECÍFICAE INTRAESPECÍFICA
La mayor parte de las especies de ostrácodosson identificables a partir de los rasgos morfo-lógicos del caparazón. Sin embargo, algunasmuestran gran plasticidad fenotípica. Este tipode variabilidad morfológica, asociada a condi-ciones ambientales, se expresa básicamentecomo variaciones en la intensidad de la orna-mentación, la presencia de tubérculos y el de-sarrollo de reticulación. Las especies polimór-ficas suelen ser euritópicas, habitantes deaguas salobres en ambientes de transición,donde las condiciones, en especial la salinidad,suelen ser muy variables.
Existe adicionalmente una variabilidad intra-específica relacionada con el desarrollo onto-genético. El crecimiento se realiza mediante 8mudas sucesivas, siendo el noveno estadio eladulto. Cuando el huevo eclosiona, el primerestadio larval ya está provisto de un capara-zón subtriangular. Los ejemplares juvenilestempranos, que son de menor tamaño, poseenvalvas delgadas, lisas, escasamente calcifica-das. Las impresiones musculares son poco evi-dentes, la charnela es rudimentaria, y la lamelainterna no se calcifica o está poco desarrolla-da. El tamaño y la calcificación de las valvas,asi como el grado de la ornamentación, vanaumentando a medida que se suceden las mu-das. Las valvas de los últimos estadios suelenreflejar los patrones generales de la ornamen-tación del adulto. Los órganos sexuales no sedesarrollan hasta los últimos estadios, por locual el dimorfismo sexual no es apreciable sinohasta ese momento (Figura 21. 5).
SISTEMÁTICA
Las partes blandas suelen destruirse comoconsecuencia de los procesos bioestratinómi-cos, preservándose casi exclusivamente valvas
y caparazones. Existen casos de preservaciónde partes blandas, como el de un myodocópidodel Silúrico del Reino Unido (Siveter et al., 2003)o de los apéndices fosfatizados de un cypridoi-deo cretácico del Brasil (Bate, 1972), pero ellosson casos excepcionales. Dado que la sistemá-tica de las formas actuales se basa principal-mente en la morfología de las partes blandas,en particular de los apéndices pares, esto re-presenta un inconveniente en la confección deuna sistemática natural para fósiles sin repre-sentación actual. Sin embargo, las valvas queconforman el caparazón están integradas conlas partes blandas del organismo, y muchos desus rasgos reflejan la morfología de estas últi-mas. Por ello, el conocimiento de las partes blan-das y de su relación con las duras permite in-terpretar filogenéticamente ciertos rasgosmorfológicos que se encuentran en las valvasfósiles.
El primero en utilizar el término Ostracodafue Latreille (1806) para reunir a un grupo deorganismos hoy ubicados en la clase Ostraco-da, y que Van Morkhoven (1959) caracterizópor la posesión de un caparazón más o menoscalcificado formado por 2 valvas perforadas,articuladas dorsalmente y sin líneas de creci-miento.
Los ostrácodos han sido considerados unasubclase de la clase Maxillopoda por la pre-sencia y morfología del ojo nauplius, lo que hasido motivo de crítica por otros autores, ba-sándose en interpretaciones de índole morfo-lógica y molecular. También fueron incluidosen los Entomostraca.
Actualmente, los ostrácodos son considera-dos una clase del subphylum Crustacea, tenien-do en cuenta las características del cuerpo y elcaparazón. Adicionalmente, la forma generalde este último y la posición y diseño de las im-
Phylum ArthropodaSubphylum Crustacea
Clase OstracodaOrden LeperditicopidaOrden PalaeocopidaOrden Podocopida
Suborden MetacopinaSuborden Podocopina
Superfamilia DarwinuloideaSuperfamilia CypridoideaSuperfamilia CytheroideaSuperfamilia BairdioideaSuperfamilia Terrestricytheroidea
Orden PlatycopidaOrden Myodocopida
Cuadro 21. 1. Sistemática de la Clase Ostracoda

OST
RA
CO
DA
609OSTRACODALOS INVERTEBRADOS FÓSILES
presiones musculares sirven para subdividir-los en 5 órdenes, que se detallan a continua-ción (Cuadro 21. 1).
PHYLUM ARTHROPODA
SUBPHYLUM CRUSTACEA
CLASE OSTRACODA
Crustáceos de cuerpo lateralmente compri-mido, contenido en un caparazón bivalvo máso menos calcificado y perforado; cabezaindiferenciada, presencia de 4 pares de apén-dices cefálicos, 1 a 3 (excepcionalmente 4) pa-res de apéndices torácicos y 1 par de furcas.Ordovícico-Holoceno.
Orden LeperditicopidaCaparazón grande, grueso y fuertemente cal-
cificado, generalmente liso. Margen dorsal ho-rizontal, largo y recto, con ángulos cardinalesbien marcados. Charnela adonta. Tubérculoocular en posición ántero-dorsal frecuente-mente bien desarrollado. Margen ventral con-vexo. Borde anterior de menor altura que elposterior. Diseño de las impresiones muscula-res consistente en un complejo compacto denumerosas cicatrices redondeadas (más de 200)ubicadas en la parte media anterior de la val-va. Tubérculo subcentral o un surco puedenindicar su ubicación en vista externa. Valvasfuertemente desiguales, la derecha recubriendoa la izquierda. El caparazón liso, margen ven-tral ampliamente convexo y destacado tubér-culo ocular sugieren un modo de vida pelági-co. Asimismo su gran tamaño indica una dietacarnívora como los recientes Myodocopida.Exclusivamente marinos. Ordovícico-Devóni-co (Figura 21. 3).
Orden PalaeocopidaCaparazón grande (entre 0,5 y 50 mm) fuer-
temente calcificado. Margen dorsal largo y rec-to, y el ventral ampliamente convexo podíapresentar adicionalmente un lóbulo ventral oun velo. Tubérculo ocular prominente y usual-mente con un profundo surco vertical (S
2) en
posición media-anterior, que corresponde alárea de inserción muscular. Impresiones mus-culares desconocidas. Lisos u ornamentadospor lóbulos y surcos. Pronunciado dimorfis-mo sexual, incluyendo el desarrollo decrúminas en posición ántero-ventral. Lamelainterna no calcificada. Charnela, cuando cono-cida, adonta. Una superfamilia de palaeocópi-dos, los Beyrichoidea (Figura 21. 4), fueron na-
dadores, probablemente cerca del fondo, enaguas poco profundas y con hábitos detritívo-ros o filtradores en su alimentación. Otros fue-ron reptadores y cavadores en fondos areno-sos y fangosos. Exclusivamente marinos. Or-dovícico-Pérmico, ?Triásico-Holoceno.
Las formas lisas y robustas, carentes de sur-cos y lóbulos y con el borde dorsal recto y muycorto en relación al largo total del caparazón,han sido separados en el orden Leiocopida (Or-dovícico-Pérmico) por Schallreuter (1973). Asi-mismo, Adamczak (1961) reunió en Eridostra-coda a formas primitivas del Ordovícico convalvas multilaminadas, es decir, integradaspor la superposición de numerosas láminasretenidas después de la muda (aunque esta ca-racterística puede no estar presente, segúnSchallreuter, 1993). Actualmente la posición deestos ostrácodos es controversial; algunos au-tores los incluyen en los Palaeocopida mien-tras que otros los consideran branquiópodosmarinos extinguidos.
Los Palaeocopida constituyen el grupo domi-nante entre los ostrácodos paleozoicos y estándiversamente representados en el Ordovícicode la Precordillera argentina.
Se incluye en este orden a los vivientesPuncioidea (Mioceno-Holoceno, Figura 21. 3)(Horne et al., 2002), una superfamilia de ostrá-codos marinos, extremadamente raros con ca-racterísticas exclusivas: poseen valvas con elmargen dorsal recto y el ventral ampliamenteconvexo con velo, y las valvas son capaces deextenderse lateralmente (amplia hiancia) demodo que sus márgenes se apoyan horizontal-mente sobre el sustrato. Asimismo sus prime-ros estadios ontogenéticos son univalvos.
Orden PodocopidaCaparazón generalmente inequivalvo, con el
borde dorsal variablemente convexo y el ven-tral ligeramente convexo, cóncavo o recto. Val-vas lisas o diversamente ornamentadas. Di-morfismo sexual a veces presente. Charnela dediferentes tipos. Lamela interna calcificada devariable desarrollo; zona marginal con cana-les de poros marginales. Su separación en ca-tegorías sistemáticas menores (subórdenes, su-perfamilias) se basa en el diseño de las impre-siones musculares. Este orden agrupa a la ma-yoría de los ostrácodos actuales bentónicos,ocupando una amplia gama de hábitats. Lamorfología de los apéndices, la forma de lasvalvas, su ornamentación y el tipo de charnelaestán en relación directa con el hábito de vida.Ordovícico-Holoceno.
El suborden Metacopina (Figura 21. 3), un gru-po marino extinguido, posee impresiones mus-

610 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
culares en un conjunto circular compacto deaproximadamente 20-25 cicatrices; su hábitofiltrador les permitió poblar ambientes mari-nos con niveles mínimos de oxígeno. Silúrico-Jurásico Temprano (Toarciano).
Dentro del suborden Podocopina, los Darwinu-loidea de hábitos caminadores, tienen valvaslisas o débilmente ornamentadas, generalmen-te charnela adonta e impresiones muscularesen una roseta de 9-12 cicatrices. Típicos de aguadulce, aunque se encuentran excepcionalmenteen ambientes de mezcla. Carbonífero (?Devóni-co)-Holoceno (Figuras 21. 3 y 6).
Los Cypridoidea incluyen formas no mari-nas y unas pocas marinas. A menudo tienencaparazón débilmente calcificado, liso o conuna tenue ornamentación, charnela adonta oraramente merodonta y las impresiones mus-culares forman un grupo de 5-6 cicatrices alar-gadas e irregulares. Son caminadores o acti-vos nadadores a escasos centímetros del sus-trato. En este grupo se incluye el género Cypridea(Jurásico Tardío-Cretácico), de utilidad bioes-tratigráfica, que posee un característico «pico»ántero-ventral. ?Devónico, Triásico-Holoceno(Figura 21. 3).
Los Cytheroidea son los más variables mor-fológicamente, habitando ambientes marinosy de transición, y más raramente de agua dul-ce. Poseen un característico diseño de 4-5 im-presiones aductoras en una hilera vertical, ypor delante de las mismas, impresionesmandibulares y dorsales. La charnela esadonta, merodonta o anfidonta, y la duplica-dura y zona marginal están bien desarrolla-das, con canales de los poros marginales des-tacados. Ordovícico-Holoceno (Figura 21. 3).
Los Bairdioidea son exclusivamente marinos,lisos o variablemente ornamentados, con char-nela usualmente robusta y simple, y músculosaductores en un grupo de 7-12 impresionesalargadas e irregulares. Ordovícico-Holoceno(Figura 21. 3).
Los Terrestricytheroidea comprenden a losostrácodos con hábitos semiterrestres. Holo-ceno (Figura 21. 3).
Los Podocopida están ampliamente represen-tados en Argentina a través de los distintosperíodos geológicos.
Orden PlatycopidaCaparazón liso o con costillas, fuertemente
inequivalvo; valva derecha usualmente mayorque la izquierda. Carecen de ojos y de lamelainterna calcificada. Filtradores, con apéndicesanteriores adaptados a hábitos cavadores. Di-morfismo sexual marcado. Impresiones mus-
culares con numerosas cicatrices en las formaspaleozoicas; menor número, aproximadamen-te 11, en dos hileras verticales (diseño «en plu-ma») en las pospaleozoicas (Figura 21. 3). Dosgéneros vivientes de platycópidos, Cytherella yCytherelloidea, forman asociaciones altamentediversificadas en mares de aguas templado-cálidas cercanas a la costa; asimismo, el grupose adapta a vivir en ambientes con niveles mí-nimos de oxígeno debido a su hábito alimenta-rio filtrador. Exclusivamente marinos. Ordo-vícico-Holoceno.
Orden MyodocopidaCaparazón grande, débilmente calcificado.
Charnela adonta o reemplazada por un liga-mento elástico. Impresiones musculares varia-bles en número, forma y diseño, y con menorvalor taxonómico que en otros grupos de os-trácodos. Antenas birrámeas adaptadas a lanatación. Formas paleozoicas, como losentomozoideos, poseen caparazón oblongo,ornamentación dactilar (fingerprint) y un pro-fundo surco dorsal medio. Los recientes(Cypridina) tienen el rostro anterior con una in-cisión ántero-ventral, por donde salen las an-tenas, llamada incisión rostral . Formasplanctónicas y pelágicas de mar abierto. Or-dovícico-Holoceno (Figura 21. 3).
Los myodocópidos están escasamente repre-sentados en el registro fósil dada su débil cal-cificación, lo que disminuye el potencial de fo-silización.
ECOLOGÍA Y PALEOECOLOGÍA
HÁBITATS Y HÁBITOS
Los ostrácodos habitan todo tipo de ambien-tes acuáticos, marinos y no marinos (ríos, arro-yos, lagos y lagunas). Como casos extremos,podemos mencionar la existencia de ostráco-dos en acuíferos confinados y otros que presen-tan hábitos semiterrestres, pudiendo vivir enlos microambientes húmedos que se generan enel humus y entre las hojas del follaje caído enselvas y bosques subtropicales.
Dado que los ostrácodos muestran claras pre-ferencias ambientales, en cada tipo de ambien-te va a existir una asociación característica:mientras que los darwinuloideos y cypridoi-deos dominan los ambientes no marinos, loscytheroideos, platycópidos y myodocópidos lohacen en los marinos (Figura 21. 11).
Los ostrácodos bentónicos pueden ser tantoinfaunales como epifaunales, e incluso compor-

OST
RA
CO
DA
611OSTRACODALOS INVERTEBRADOS FÓSILES
tarse como bentos vagabundo, viviendo partedel tiempo dentro del sustrato, pero nadandocortas distancias por encima de la interfaseagua-sedimento. Los infaunales se desplazanutilizando las antenas y las patas para abrir-se camino y propulsarse entre los granos delsedimento, que a veces son mayores que elpropio caparazón. Las especies epifaunalesviven apoyadas sobre la interfase, pudiendonadar cortas distancias con ayuda de las an-tenas, o trepando a los objetos con las ante-nas y las patas. Suelen presentar el margenventral plano y ensanchado e incluso proce-sos alares (Cytheropteron) que constituyen unsoporte adicional en fondos fangosos.
Algunas formas epifaunales son fitales: vi-ven sobre la superficie de algas y plantasacuáticas, de las cuales succionan alimento através de los apéndices cefálicos altamentemodificados. Aquéllos que habitan en ambien-tes marinos de alta energía cercanos a la cos-ta, suelen poseer valvas fuertemente calcifi-cadas y ornamentadas, y un prominente tu-bérculo ocular (Cornucoquimba). Las formaspsicrosféricas viven a profundidades entre2000 y 3000 m y a temperaturas constantesmenores a 4°C, son ciegas, poseen superficieexterna comúnmente reticulada, aunque tam-bién puede ser lisa o híspida, con el borde dor-sal recto, alerones laterales y proceso caudalposterior.
ALIMENTACIÓN
Los ostrácodos son omnívoros oportunistas,consumiendo, en función de la oferta alimenti-cia, protistas, algas, crustáceos de pequeño ta-maño, larvas o materia orgánica disuelta. Tam-bién se han descripto especies comensales yparásitas. Poseen hábitos filtradores, detrití-voros y depositívoros. Otros suelen tomar elalimento a partir de conductas activas de cazao ramoneo en las que participan la mayor par-te de los apéndices cefálicos. Los ostrácodosfiltradores se alimentan pasivamente de par-tículas que ingresan al caparazón con las co-rrientes generadas por el movimiento sincro-nizado de los apéndices cefálicos.
PARÁMETROS AMBIENTALES
Los parámetros con mayor influencia en ladistribución de los ostrácodos son la estabili-dad del ambiente, la salinidad, la naturalezadel sustrato, la temperatura y el contenido deoxígeno. Sin embargo, dado que los mismosestán interrelacionados, es difícil establecervinculaciones univalentes entre ellos y los pa-trones de distribución y riqueza.
Estabilidad. Los ambientes marinos son másestables que los continentales, con la excepción
Figura 21. 11. Diagrama de la distribución de ostrácodos vivientes en distintos ambientes marinos y no marinos (modificado de Brasier,1980).

612 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
de algunos lagos profundos de origen tectónico.Su estabilidad, en combinación con otros fac-tores tales como disponibilidad de nutrientesy saturación de oxígeno, crean ambientes pro-picios para el establecimiento de asociacionesricas y diversas. En los ambientes marinos mar-ginales suelen darse importantes gradientes.Incluso ambientes temporalmente muy esta-bles, tales como un estuario, con frecuencia soninestables en términos de los parámetros fisi-coquímicos.
Salinidad. Constituye uno de los factores quemayor control ejerce sobre la distribución delos ostrácodos. Es una de las propiedades másestables en el ambiente marino, y allí las aso-ciaciones alcanzan su máxima diversidad. Latransición entre las condiciones marinas y nomarinas se refleja en una disminución de ladiversidad.Los ambientes marinos marginales mixoha-linos (estuarios y lagunas costeras), son habi-tados por géneros típicos de aguas salobres,como también por géneros de afinidades ma-rinas y no marinas que toleran respectiva-mente reducción e incrementos notables en lasalinidad. En los ambientes salobres, las es-pecies suelen desarrollar poblaciones super-numerarias y las asociaciones son poco di-versas.En ambientes acuáticos continentales, el nú-mero de especies vuelve a incrementarse, aun-que asociaciones pobres en número de espe-cies y de individuos se verifican en aguas áci-das con escasos nutrientes. En las proximida-des del punto de precipitación de la calcita(aproximadamente 1400 mg/l), la diversidadalcanza valores elevados, declinando con elaumento de la salinidad. En lagos y lagunascontinentales, la relación entre la evaporacióny el aporte de aguas meteóricas determina lapermanencia y estabilidad del hábitat, lo quese refleja en la abundancia y diversidad delas asociaciones (Figura 21. 12). El límite su-perior de tolerancia para los ostrácodos es300g/l, por lo que, ambientes hipersalinos noson aptos para su desarrollo. Cabe destacarque la composición iónica del agua y no solola salinidad total, debe ser considerada en losambientes no marinos como parámetro de-terminante en la composición taxonómica delas asociaciones.
Naturaleza del sustrato. Este parámetro juegaun papel primario en la distribución de los os-trácodos bentónicos. Los condicionamientosmayores se relacionan con la energía del me-dio, la circulación del agua intersticial y la dis-
ponibilidad de materia orgánica. Habitan se-dimentos arenosos, limosos y limo arcillosos,pero su abundancia se reduce drásticamenteal aumentar la cohesividad del sustrato, lo queimplica problemas adicionales de desplaza-miento, limitación en la circulación intersti-cial y la difusión del oxígeno. Los ostrácodosinfaunales suelen vivir en los 2-3 cm inmedia-tamente por debajo de la interfase agua-sedi-mento. Más profundamente, el establecimien-to de ambientes anóxicos y la disminución denutrientes son factores limitantes para el de-sarrollo de la meiofauna en su conjunto.
Temperatura. También es un factor importanteen términos de distribución, aunque no parecedeterminar los patrones de riqueza y diversi-dad. Hay especies euritérmicas capaces de vi-vir en ambientes que presentan una gran am-plitud térmica, tales como ambientes no mari-nos de latitudes medias y altas, o en áreasdesérticas. Sin embargo, la mayoría de las es-pecies posee claras preferencias por rangos re-lativamente estrechos de temperatura. En losambientes marinos de plataforma, el gradientelatitudinal de la temperatura superficialoceánica es el principal factor que determinala existencia de provincias faunísticas. Estegradiente limita la expansión hacia el ecuadorde las especies criofílicas, o sea aquéllas conpreferencia por las aguas frías, asi como tam-bién la expansión hacia latitudes polares delas especies termofílicas, aquéllas con preferen-cia por aguas cálidas. Por ello, el provincialis-mo biogeográfico de ostrácodos en ambientesde plataforma guarda íntima relación con laszonas climáticas, pudiéndose definir asociacio-nes polares, subpolares, subtropicales y tropi-cales.La distribución vertical está determinada porla temperatura pero también por el contenidode oxígeno. Las especies psicrosféricas habitanel talud continental y las planicies abisales, endonde la temperatura es menor a 4°C. El esta-blecimiento de la psicrósfera a partir del Eocenoy de la circulación oceánica mundial favoreciósu migración entre las distintas cuencasoceánicas, dando origen a las actuales faunaspanabisales de distribución mundial.
Oxígeno. La presión parcial del oxígeno cons-tituye un parámetro fundamental en el con-trol de la distribución de los ostrácodos. Enambientes marinos y continentales, el oxígenodisuelto en la interfase agua-sedimento está enequilibrio con el de la columna de aguasuprayacente, pero pocos centímetros por de-bajo de la interfase el oxígeno disuelto descien-

OST
RA
CO
DA
613OSTRACODALOS INVERTEBRADOS FÓSILES
Figura 21. 12. Diagrama de las diferencias entre lagos bajo dos regímenes climáticos diferentes. A. laprecipitación es mayor que la evaporación (P>E); B. la evaporación es mayor que la precipitación (E>P). Enambos casos, esta relación controla la salinidad del agua del lago, la naturaleza del sedimento y ladisponibilidad de nutrientes, y por ende, la composición de la fauna de ostrácodos.

614 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
de abruptamente. Como consecuencia de ello,la mayoría de los ostrácodos bentónicos habi-ta en los primeros 2-3 cm del sedimento. Sinembargo, algunos ostrácodos son capaces desobrevivir con niveles sorprendentemente ba-jos de oxigenación. En los ostrácodos de hábi-tos filtrantes, como los Platycopida, las placasbranquiales están particularmente bien desa-rrolladas y mantienen una corriente de aguapermanente en el interior del caparazón, porlo cual este tipo de hábito alimenticio favoreceel intercambio gaseoso y la oxigenación. Comoconsecuencia de ello, asociaciones poco diver-sas de Platycopida son indicadoras de ambien-tes disaeróbicos.
UTILIDAD DE LOS OSTRÁCODOS
Las características ecológicas de los ostráco-dos han motivado que, con frecuencia, seanutilizados en interpretaciones paleoambienta-les, paleoclimáticas, paleooceanográficas y aunen problemas vinculados con la contaminaciónambiental y la tafonomía.
RECONSTRUCCIÓN PALEOAMBIENTAL
Desde el punto de vista de sus aplicaciones enla interpretación de los ambientes fósiles, elpotencial de los ostrácodos se basa en tres pi-lares: habitan todo tipo de ambientes acuáti-cos, abarcan casi todo el Fanerozoico y suelenser ecológicamente muy sensibles. Como con-secuencia, constituyen excelentes fósiles facia-les, permitiendo la caracterización de lospaleoambientes de depositación y de sus prin-cipales parámetros ecológicos a nivel local(paleosalinidad, paleotemperatura, paleobati-metría), el reconocimiento de eventos ecológi-cos de gran escala y la reconstrucción paleo-geográfica.
En los ambientes lacustres, la interpretaciónpaleoecológica de la fauna de ostrácodos con-tinentales tiene importantes aplicaciones pa-leoclimáticas. Los ostrácodos reflejan la sali-nidad, la composición iónica, el pH y la tem-peratura del agua entre otras variables cli-máticamente controladas. La naturaleza quí-mica del agua depende del equilibrio entre laprecipitación y la evaporación. Un cuerpo deagua en un ambiente donde la evaporación ex-cede a la precipitación, sufrirá un incrementode la salinidad. A medida que la salinidad au-menta, tendrá lugar la precipitación de ciertosminerales, lo que originará variaciones en lacomposición iónica relativa del agua como re-
sultado de la remoción preferencial de ciertosiones (Figura 21. 12). La calcita precipita enprimer lugar, lo que provocará un aumento enla relación Mg/Ca por remoción del Ca+. Dadoque la composición de los solutos y no solo lasalinidad determinan la presencia de ciertosostrácodos lacustres, el estudio de la natura-leza y evolución de las asociaciones permitecaracterizar químicamente las aguas y laevolución de la cuenca. Análisis de la rela-ción Mg/Ca, Sr/Ca o de otros elementos tra-za, constituyen estudios de rutina en inves-tigaciones paleolimnológicas ya que permi-ten caracterizar paleohidrológicamente lasaguas.
Los ostrácodos son una fuente importante decarbonato autigénico, por lo que su potencialcomo indicadores paleoambientales se ha vis-to incrementado a partir del desarrollo de es-tudios geoquímicos e isotópicos (principalmen-te δ18O y δ13C). La relación 18O/16O de las valvas,controlada principalmente por la temperatu-ra del agua y la composición isotópica en lainterfase agua-sedimento, puede ser interpre-tada en términos paleoclimáticos y como va-riaciones en la relación precipitación/evapo-ración, mientras que la relación 13C/14C ,13C/12Cpuede ser analizada en términos de paleopro-ductividad.
PALEOOCEANOGRAFÍA
En el ambiente marino, el reconocimiento deasociaciones de plataforma, batiales y abisalespermite la reconstrucción paleobatimétrica yde las variaciones relativas del nivel del marasociadas a fenómenos eustáticos, tectónicos ysedimentarios.
Las diferentes masas de agua de los océanosestán caracterizadas por su temperatura, sali-nidad y contenido de oxígeno. Siendo los os-trácodos particularmente sensibles a estosparámetros, hay cierta relación entre la distri-bución de los bentónicos y la de las masas deagua que componen la estructura hidrológicade los océanos.
Los ostrácodos son utilizados también comoindicadores de las condiciones de oxigenaciónde los fondos oceánicos. Ya mencionamos lacapacidad de los filtradores para sobreviviren ambientes disaeróbicos, de modo que la do-minancia de Platycopida, aunque con baja di-versidad en una asociación, estaría indicandoeste tipo de ambientes. Adicionalmente, cier-tos ostrácodos de hábitos no filtradores hansido utilizados como indicadores de oxigena-ción de las aguas intersticiales. Así, según al-

OST
RA
CO
DA
615OSTRACODALOS INVERTEBRADOS FÓSILES
gunos autores, existiría una relación positivaentre el tamaño del vestíbulo en Krithe, un gé-nero cosmopolita común en las profundidadesbatiales, y las condiciones de oxigenación de lainterfase.
CONTAMINACIÓN AMBIENTAL
Los ostrácodos son utilizados en estudios decontaminación ambiental, dado que las espe-cies euritópicas se tornan dominantes a medi-da que la polución se incrementa. No obstante,cabe aclarar que, al igual que el resto de lameiofauna, no soportan niveles de alta polu-ción, y son incapaces de vivir en medios fuer-temente contaminados. Un claro ejemplo loconstituyen las faunas del Mar Aral, que hansufrido un incremento de salinidad de 10‰ a35‰ en solo 30 años como consecuencia delimpacto antropogénico y de las condicionesclimáticas que determinan una fuerte evapo-ración. Bajo estas condiciones, de las 11 espe-cies presentes en 1960 solo Cyprideis torosa so-brevive en la actualidad. En Argentina, Kaesleret al. (1979) analizaron el efecto de la poluciónpor hidrocarburos sobre la fauna de ostráco-dos en el Estrecho de Magallanes.
TAFONOMÍA
También puede destacarse el valor de los os-trácodos en estudios de índole tafonómica. Lamorfología de valvas y caparazones, que faci-lita su transporte aun por corrientes débiles, yel crecimiento discontinuo, que da lugar a lapresencia de juveniles, les otorga un gran po-tencial para los estudios bioestratinómicos. Lapresencia de selección por tamaño y el estudiode la relación entre adultos y juveniles permi-te inferir la presencia de transporte aun cuan-do no existan otras evidencias que lo indiquen(Browers, 1988; Whatley, 1988). La distinción,algunas veces dificultosa, entre sedimentospelágicos y turbidíticos puede ser dilucidadaen base a la abundancia y preservación de losostrácodos. Asimismo, se ha sugerido que lalongitud de las valvas puede ser usada paraestimar el tamaño de grano original en rocassedimentarias recristalizadas. También se hademostrado su utilidad en la estimación develocidad de sedimentación y energía y direc-ción de las paleocorrientes.
LOS OSTRÁCODOS Y LAPALEOBIOGEOGRAFÍA DE AMÉRICADEL SUR
La Paleobiogeografía, establecida con inde-pendencia de los datos geológicos y geofísicos,ayuda no solamente a la reconstrucción paleo-geográfica, sino también a la datación de lasetapas de desplazamiento de las masas conti-nentales, al conocimiento de la evolucióntopográfica de los fondos marinos, de los cam-bios climáticos y al establecimientos de corrien-tes oceánicas.
Las microfaunas de ostrácodos ordovícicosde la Precordillera de San Juan evidencian cier-to grado de endemismo, al que no son ajenosotros grupos de invertebrados. Sin embargo,la presencia de los géneros Berdanellina,Saccelatia, Ningulella y Ordovizona indica afini-dad con las regiones boreales de Báltica yLaurentia, mientras que los registros de Pilla yEodominina muestran una relación con Austra-lia, afinidad observada también en trilobites ybraquiópodos. La ocurrencia de un importan-te número de taxones pandémicos entre lospodocópidos podría estar relacionado con lapreferencia de este grupo, durante el Paleozoi-co Temprano, por las aguas de mayor profun-didad de la plataforma y del talud (Siveter,1984).
En el Mesozoico, las profundidades oceánicasconstituyeron una barrera física prácticamenteinfranqueable para los ostrácodos; el factortectónico tuvo asimismo una influencia de pesoen la distribución del grupo. Acompañando aldesmembramiento de Gondwana se produje-ron sistemas de fracturas afectando, por unlado, el nordeste de América del Sur y el cen-tro-oeste de África y por el otro, los extremosaustrales de América del Sur (en particularArgentina) y África (en particular Sudáfrica).En el norte la fracturación inicial produjo es-tructuras tipo graben paralelas a las actualescostas; en el extremo austral, las fracturasmayores son perpendiculares a los márgenesy de rumbo este-oeste. Como consecuencia deello los continentes migraron a lo largo de lafractura de Malvinas de manera tal que, lacomunicación física entre ambas regionesaustrales continuó hasta tiempos albianos, noexistiendo, hasta ese momento, barrerasoceánicas entre ambos (Dingle, 1984). Sin em-bargo, la dorsal de Río Grande-Walvis repre-sentó en algunos casos una barrera física efec-tiva para impedir la migración de los ostrá-codos. Así, la distribución de Procytherura du-rante el Jurásico Temprano y Medio (Ballent yWhatley, 2000) con registros boreales y austra-

616 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
les, incluidos aquellos de Argentina, indican elcarácter cosmopolita del taxón, avalando diver-sas rutas de migración hacia el sur a través delTethys y del Corredor Hispánico (víaProtoatlántico, que comunicaba el Tethys occi-dental y el Pacífico oriental). Este género se vol-vió eminentemente austral durante el JurásicoTardío y Cretácico Temprano con registros envarias localidades gondwánicas, evidencian-do una conexión, a través de Sudáfrica, entreel este de África e India y el extremo australde América del Sur. La presencia de Procytheruraen el oeste de África (Gabón) en tiemposposalbianos indica una conexión entre el At-lántico sur y norte a partir del Albiano, endirección sur-norte, señalando también quela dorsal de Río Grande-Walvis, aunque efec-tiva para faunas de aguas profundas, permi-tió el pasaje de las de aguas someras.
Las similitudes entre las faunas de ostráco-dos del Jurásico Tardío y Cretácico Tempranode varias áreas gondwánicas, tales como elextremo sur de América del Sur, la Platafor-ma de Malvinas, Sudáfrica, Tanzania,Madagascar, India y el oeste de Australia, re-fuerzan la teoría de la cercanía de estas ma-sas continentales, ya que los ostrácodos invo-lucrados fueron habitantes de plataformasmarinas poco profundas
Durante el Jurásico Tardío y el CretácicoTemprano, en el nordeste del Brasil (cuencaSergipe/Alagoas) y en África occidental(Gabón), las cuencas petrolíferas contienenasociaciones con taxones no marinos comu-nes (Cypridea y otros cypridoideos), lo que evi-dencia la contigüidad de ambos continentes(Colin y Dépêche, 1997). En las mismas regio-nes, las similitudes en base a especies mari-nas del Cretácico alto estudiadas por Vivierset al. (2000) avalan la migración a través delAtlántico sur.
En el Cretácico alto (Campaniano), los regis-tros de las mismas especies de Majungaella enlas cuencas del oriente brasileño, Patagoniaargentina y Antártida (Fauth et al., 2002) apo-yan la presencia de vías de comunicación ha-cia el norte demostrando la existencia del océa-no Atlántico sur, y hacia el sur, vía el Pasaje deDrake. Por otro lado, Majungaella fue un taxóntermófilo exclusivamente de distribución aus-tral, de manera que su presencia en el Cretáci-co terminal del continente antártico indicatemperaturas más elevadas que las actualesen esas altas latitudes.
La distribución de las especies del género ma-rino bentónico Brachycythere, característico delCretácico, constituye otro ejemplo de la utili-dad de los ostrácodos en paleogeografia. El re-
gistro de B. sapucariensis y formas afines en cali-zas del Cretácico Superior bajo (Cenomaniano-Turoniano) en el oriente ecuatoriano, noroesteperuano y sudoeste boliviano evidencia unaconexión marina desde el norte – a través delactual territorio peruano – por medio de un bra-zo de mar de poca profundidad y aguas tem-pladas, que permitió la formación de calizas(Rodrigo et al., 1990). La presencia de Brachycythe-re cf. rhomboidalis en Cuba central y en las cuen-cas africanas y brasileñas vinculadas al domi-nio atlántico, está indicando una comunicaciónmarina durante el Cretácico Tardío entre el Ca-ribe y los mares que se desarrollaron a partir dela apertura del Atlántico sur (Díaz-Collell et al.,1998).
Las similitudes de ostrácodos de ambientesmarinos neógenos profundos y del psicrosféri-co Abyssocythere en Cuba, Trinidad, norte de Ita-lia y Cárpatos occidentales muestran la pre-sencia de facies profundas en ambas márgenesdel Tethys y su comunicación con la región delCaribe hacia fines del Mesozoico.
BIOESTRATIGRAFÍA
El hecho de que los ostrácodos sean orga-nismos en su gran mayoría bentónicos y sen-sibles a los cambios ambientales, limita encierta medida el potencial bioestratigráficode los mismos. Sin embargo, la amplia dis-tribución espacio-temporal que los caracte-riza permite su desarrollo en casi todo tipode facies marinas y continentales, incluso enaquéllas en las que otros tipos de fósiles no seencuentran.
Es así como, en ausencia de organismosplanctónicos, los ostrácodos constituyen unabuena herramienta bioestratigráfica, útilpara establecer esquemas zonales que se hanaplicado con éxito en la resolución de pro-blemas paleobiogeográficos, de tectónica deplacas y exploración de hidrocarburos, en-tre otros. Como ejemplo podemos mencionaral cypridoideo Cypridea, clásicamente utiliza-do en la delimitación estratigráfica de lasentidades salobres y dulceacuícolas del nor-te de Europa conocidas bajo los nombres dePurbeck (Jurásico Superior) y Wealden (Cretá-cico Inferior) (Anderson, 1985).
Otra aplicación de ostrácodos marinos ben-tónicos a la estratigrafía es aquella referida aespecies del género Majungaella a través delCretácico en las cuencas Neuquina de Argen-tina y Austral del sur de Argentina y Chile(Figura 21. 13).

OST
RA
CO
DA
617OSTRACODALOS INVERTEBRADOS FÓSILES
BIOESTRATIGRAFÍA DE LOSOSTRÁCODOS EN LA ARGENTINA YOTROS PAÍSES DE AMÉRICA DEL SUR
Los primeros registros de ostrácodos para laArgentina corresponden al Ordovícico de laPrecordillera de San Juan. Los mismos han sidoobjeto de estudios taxonómicos por parte deRossi de García (1975) y Rossi de García yProserpio (1978), entre otros. Posteriormente,fueron revisados y actualizados (Schallreuter,1996, Salas 2002 a, 2002 b, 2003) demostrandola gran diversidad del grupo en esos tiempos yaportando a su vez nuevos elementos de juicioen estudios paleogeográficos (Figura 21. 14). Sehan descripto especies de Palaeocopida yPodocopida que componen dos asociacionesbien diferenciadas: la inferior se correlacionabioestratigráficamente con la zona de Paraglo-ssograptus tentaculatus (parte superior) de edadllanvirniana temprana, y la superior con lazona de Nemagraptus gracilis del Caradocianoinferior. Las mismas permiten afinadas corre-
laciones entre secciones estratigráficas de lacuenca precordillerana (Salas, 2002 c). Los es-tudios sobre ostrácodos del Paleozoico Supe-rior de la cuenca Tepuel-Genoa en Patagoniason escasos, aun cuando es frecuente su men-ción en trabajos sistemáticos de otros gruposde invertebrados. Se cuenta con el registro pio-nero de Rossi de García (1972) y más reciente-mente con el de Díaz Saravia y Jones (1999),pero su estudio es todavía incipiente, siendoun campo de investigación muy interesantecon excelentes perspectivas futuras.
Para el Mesozoico (Triásico terminal, Jurásicoy Cretácico) de la cuenca Neuquina, las faunasde ostrácodos marinos están compuestas bási-camente por cytheroideos y cypridoideos su-bordinados. En el Jurásico Inferior (Hettangia-no) se mencionan faunas de metacopinos. Exis-te una refinada calibración entre las microfau-nas provenientes de secciones estratigráficas desuperficie y niveles de subsuelo con zonasamonitíferas, de otros invertebrados y de fora-miníferos bentónicos (Riccardi et al., 1999).
Figura 21. 13. Distribución estratigráfica de Majungaella (Podocopida, Cytheroidea), marino. Cretácico, cuenca Neuquina, Argentina ycuenca Austral del sur de Argentina y Chile (modificado de Ballent et al., 1998).

618 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES

OST
RA
CO
DA
619OSTRACODALOS INVERTEBRADOS FÓSILES
Las microfaunas marinas del Cretácico Supe-rior y Terciario de Patagonia son numerosas yhan sido estudiadas ininterrumpidamente des-de 1970, estableciéndose correlaciones con zo-nas bioestratigráficas de foraminíferos planc-tónicos y nanofósiles. Estos datos fueron resu-midos por Echevarría (1984). Los ostrácodos dellímite Cretácico-Terciario se hallan exhaustiva-mente estudiados (Bertels, 1995), distinguién-dose taxones endémicos tales como Neoveenia,Rocaleberis y Wichmannella y otros que reflejanclaras conexiones gondwánicas, como Togoina,Nigeria y Afranticythereis. Delimitan el pasaje Cre-tácico-Terciario, entre otras, las especies Togoinacretacea, T. semiinornata, Nigeria punctata, Wichman-nella cretacea, W. magna, Rocaleberis araucana yAfranticythereis venusta del Maastrichtiano, yTogoina argentinensis, Wichmannella meridionalis,Rocaleberis nascens y Afranticythereis schilleri delDaniano inferior (Figura 21. 14).
Marengo (2000) caracterizó los depósitos ter-ciarios relacionados con la transgresiónEntrerriense-Paranense (Mioceno Medio-Tar-dío) en la cuenca Chacoparanense y noroesteargentino, en base a microfósiles calcáreos,incluídos los ostrácodos.
Es abundante la bibliografía sobre los ostrá-codos cuaternarios y recientes de la plataformacontinental atlántica del Brasil, Argentina e is-las Malvinas (Whatley y Cusminsky, 2002;Whatley et al., 1997, 1998). Los ostrácodos deambientes transicionales recuperados de depó-sitos cuaternarios en la costa de la provincia deBuenos Aires han permitido la caracterizaciónpaleoecológica de los ambientes asociados a los
últimos eventos transgresivos. Bertels yMartínez (1997) y Bertels-Psotka y Laprida(1998) estudiaron los depósitos relacionados conlas transgresiones Querandinense y Platense enel ámbito del estuario de Bahía Blanca y la cuen-ca del Salado, respectivamente.
A lo largo del borde atlántico brasileño lasvariadas microfaunas de ostrácodos cretácicostambién han demostrado ser elementos útilespara caracterizar las asociaciones bentónicas(Viviers et al., 2000).
Los ostrácodos provenientes de las cuencasintracratónicas del Cretácico y Terciario Infe-rior del centro-oeste de América del Sur (Boli-via, Ecuador, Perú), también han sido objetode estudio, en muchos casos estableciéndosecorrelaciones locales con los foraminíferosplanctónicos acompañantes (Durán y Salazar,1990; Rodrigo et al., 1990, Musacchio, 1990).Respecto a las áreas caribeñas, cuyos ostráco-dos del Cretácico Tardío y Neógeno son cono-cidos desde mediados del siglo pasado merceda los trabajos de Van der Bold, merecen espe-cial atención las recientes investigaciones so-bre los ostrácodos de Cuba, los que tienen im-portantes implicancias paleogeográficas debi-do a sus afinidades con otras cuencas caribeñasy del nordeste del Brasil y África occidental.
Una mención aparte merecen los ostrácodosno marinos dada la ubicuidad de los mismos.Hasta el momento no hay registros de éstos enel Paleozoico de Argentina, siendo las micro-faunas del Paleozoico Superior de la cuenca deParaná las mejor representadas en territoriosdel Brasil (Sohn y Rocha Campos, 1990), Para-
Figura 21. 14. Ostrácodos marinos. En todos los casos se ilustran en vista lateral externa. A-C. Palaeocopida, Ordovícico, Precordillera.A. Chattertonella corrugata, valva izquierda, amplete, unisurcada, borde dorsal recto, dos nódulos, superficie corrugada; B. Cribobolbinacf. C. cribata, valva derecha, amplete, ángulos cardinales bien definidos, velo, superficie con débil reticulación; C. Cribobolbina cribata,valva izquierda, levemente preplete, unisurcada, borde dorsal recto, superficie reticulada y con pequeñas papilas. D-F. Platycopida,cuenca Neuquina. D. Cytherelloidea circunscripta, Jurásico Inferior, caparazón, vista lateral derecha, costilla paralela a los bordesdorsal, posterior y ventral; E. Cytherella australoamericana, Paleoceno, valva derecha, superficie con punctuaciones; F. Cytherellaspirocostata, Paleoceno, valva derecha, costilla en espiral; G. Bairdiodea. Bythocypris sp., Paleoceno, cuenca Neuquina, caparazón,vista lateral izquierda, superficie lisa; H-U. Cytheroidea. H. Procytherura transversiplicata, Jurásico Medio, cuenca Neuquina, valvaderecha, costilla longitudinal media, reticulado intercostal, tubérculo ocular; I. Sondagella theloides, Cretácico Inferior, cuenca Neuquina,caparazón, vista lateral izquierda, costillas longitudinales, pequeñas papilas, tubérculo ocular; J. Echinocythereis sp., Holoceno,Atlántico sudoccidental, valva izquierda, superficie con pequeñas espinas; K. Buntonia rocanortensis, Paleoceno, cuenca Neuquina,valva izquierda, costillas longitudinales; L. Trachyleberis weiperti, Paleoceno, cuenca Neuquina, caparazón, vista lateral izquierda,superficie con tubérculos y tubérculo ocular; M. Ambostracon bertelsae, Plioceno, Atlántico sudoccidental, valva izquierda, tubérculosubcentral y ocular; N. Austroaurila theeli, Plioceno, Atlántico sudoccidental, valva derecha, superficie con retículo y tubérculosubcentral; O. Cativella bensoni, Plioceno, Atlántico sudoccidental, valva derecha, costillas longitudinales, retículo y tubérculo ocular;P-S. Holoceno, cuenca del Salado. P. Hemicytherura chuiensis, valva derecha, tubérculo ocular, costilla submarginal ventral marcada,superficie ornamentada por hoyuelos y punctas; Q. Cytheretta punctata, valva derecha con costillas longitudinales chatas y anchas,espacios intercostales con reticulación, tubérculo subcentral marcado; R. Cornucoquimba lutziana, valva izquierda con tubérculosubcentral, tubérculo ocular y costilla marginal que recorre los extremos anterior y ventral; S. Oculocytheropteron micropunctatum, valvaderecha, superficie ornamentada por costillas cortas que forman un reticulado subpoligonal, costillas marginales subparalelas a losmárgenes ventral y anterior, espacios intercostulares con punctas; T-U. Holoceno, golfo de Vizcaya, Francia, T. Cytheropteron alatum,proceso alar subtriangular muy marcado y proceso caudal medial agudo; U. Krithe sp., proceso caudal ventral subredondeado.

620 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES

OST
RA
CO
DA
621OSTRACODALOS INVERTEBRADOS FÓSILES
guay (Zabert, 1985) y Uruguay (Díaz Saravia yHerbst, 2001).
Durante el Cretácico, los depósitos continen-tales, a veces con leve influencia marina, estánampliamente distribuidos en el territorio ar-gentino, ya como cuerpos de agua dulce o salo-bre desligados de acontecimientos marinos, ocomo una etapa preliminar o remanente deingresiones marinas, que se inician o finalizancon depósitos hipohalinos. Se citan ostrácodosen las cuencas del noroeste, del Neuquén yMendoza y de San Jorge. Especies de Cypridea,Ilyocypris y cytheroideos junto a algas caráceashan sido utilizadas para caracterizar bioestra-tigráficamente y correlacionar las cuencas con-tinentales mesozoicas de la Argentina(Musacchio, 1989, 1990) (Figura 21. 15). Espe-cies de cypridoideos son utilizadas en China yBrasil para establecer correlaciones localesentre las extensas cuencas continentales meso-zoicas en cada uno de esos países (Pang Qiqingy Whatley, 1990; Dias Brito et al., 2001).
Más escasos son los registros de ostrácodoscontinentales en el Terciario argentino, si bienfaunas del Oligoceno (Bertels-Psotka yCusminsky, 1999) y del Mioceno (Bertels-Psotka, 2000) fueron descriptas para la regiónpatagónica (Figura 21. 15).
En el Cuaternario, cabe mencionar los traba-jos pioneros de Zabert (1981) y Zabert y Herbst(1986) sobre ostrácodos pleistocenos del nores-te argentino; mientras que los cuaternarios deambientes continentales están recibiendo cre-ciente atención, como consecuencia de suaplicabilidad en estudios paleoclimáticos, enparticular en las regiones patagónica(Cusminsky y Whatley, 1996; Whatley yCusminsky, 1999) y pampeana (Laprida et al.,2004) (Figura 21. 15).
EVOLUCIÓN
Las primeras valvas y caparazones calcifica-dos asignables indudablemente a ostrácodosdatan del Ordovícico. A partir de ese momentoy a lo largo de su historia evolutiva estos ar-trópodos han sufrido una espectacular radia-ción y su registro en la columna geológica hasido constante.
El Ordovícico Temprano se caracteriza poruna gran diversidad de ostrácodos probable-mente asociada con un período transgresivo,el desarrollo de las plataformas continentalesy la consecuente aparición de nichos apropia-dos. Es así como ya en estos momentos, haceunos 500 m.a., están presentes los grandesleperditicópidos y aparecen los primeros pa-laeocópidos, que van a dominar hasta el De-vónico, además de los primeros podocópidosy myodocópidos pelágicos (entomozoideos).A fines del Paleozoico disminuyen los pa-laeocópidos y aparecen los primeros ostráco-dos no marinos (darwinuloideos y cypridoi-deos). Hacia el Triásico se extinguen casi porcompleto los Palaeocopida, y algunos podocó-pidos sobreviven a la extinción permo-triásica. Aparecen entonces nuevos linajes depodocópidos, más precisamente de cytheroi-deos, constituyéndose en el grupo dominantedurante el Mesozoico y todo el Cenozoico. Lasasociaciones marinas del Triásico Tardío yJurásico Temprano son poco diversificadas yestán representadas por los extinguidosmetacopinos. Las facies lacustres y salobresmesozoicas, ampliamente distribuidas comoconsecuencia de la regresión triásica, contie-nen mayormente cypridoideos no marinos,darwinuloideos y cytheroideos no marinos.La preferencia de estos grupos por tales am-bientes se mantiene aún en la actualidad. Los
Figura 21. 15. Ostrácodos no marinos de la República Argentina. En todos los casos se ilustran en vista lateral externa. A-B. CretácicoInferior, Patagonia. A. Cypridea craigi, valva izquierda, espinas cónicas, papilas y punctuaciones, pico bien desarrollado; B. Cyprideaaustralis, valva izquierda, punctuaciones fuertes, subpoligonales y papilas de tamaños variados, pico fuerte y moderadamente agudo; C-E.Paleógeno, cuenca Neuquina. C. Ilyocypris sp., superficie reticulada con tubérculos expandidos; D. Limnocythere sp., valva izquierda,superficie con reticulado subpoligonal; E. Cypridopsis (Prioncypris) sp., caparazón, vista lateral derecha, mayor altura en la parte media; F-H. Oligoceno. F. Candona? mapuche, caparazón, vista lateral derecha, margen dorsal muy convexo, anterior muy oblicuamente redondeado;G. Eucandona? patagonica, caparazón, vista lateral derecha, margen dorso-posterior muy oblicuo; H. Alicenula? sp., caparazón elongado,vista lateral derecha, borde charnelar subrecto, ventral sinuoso; I-K. Mioceno, Patagonia. I. Candona? sp. aff. C.? mapuche, caparazón, vistalateral derecha, mayor altura coincidente con la mitad del largo; J. Cyprinotus australis, molde interno de caparazón, vista lateral derecha,margen dorsal muy convexo; K. Limnocythere colloncurense, molde interno, vista lateral derecha, margen dorsal recto, superficie reticulada;L-M. Pleistoceno, cuenca del Salado. L. Cyprideis multidentata, valva derecha, superficie externa con punctas poco marcadas y denticulaciónen el margen ántero-ventral; M. Perissocytheridea krommelbeini, valva derecha, superficie tuberculada, con tubérculo ocular bien diferen-ciado e incipiente proceso alar, superficie externa con hoyuelos subcirculares marcados y finas costillas marginales subparalelas al margenanterior; N-Q. Holoceno, provincia de Buenos Aires. N. Candonopsis brasiliensis, valva izquierda; O. Heterocypris sp., valva izquierda; P.Chlamydotheca sp., valva derecha con espina póstero-ventral; Q. Cytheridella ilosvayi, valva izquierda con marcado surco ántero-dorsal ysuperficie con punctas marcadas.

622 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
myodocópidos decayeron durante el Mesozoi-co y Cenozoico, siendo actualmente habitan-tes pelágicos de mar abierto. Las faunaspsicrosféricas de ambientes marinos profun-
dos se han desarrollado a partir del CretácicoTardío coincidente con una etapa de expansiónde fondos oceánicos y la apertura del océanoAtlántico.
BIBLIOGRAFÍA
Adamczak, F. 1961. «Eridostraca»- A new Suborder ofostracods and its phylogenetic significance. ActaPalaeontologica Polonica 6: 29-94.
Anderson, F. W. 1985. Ostracod faunas in the Purbeckand Wealden of England. Journal of Micropalaeontology4: 1-68.
Ballent, S. C. y Whatley, R. 2000. The distribution of theMesozoic ostracod genus Procytherura Whatley:palaeogeographical implications with specialreference to Argentina. Alcheringa 24: 229-242.
Ballent, S.C., Ronchi, D.I. y Whatley, R. 1998. Theostracod genus Majungaella Grekoff in Argentina.Revista Geológica de Chile 25: 45-55
Bate, R. 1972. Phosphatyzed ostracods with appendagesfrom the Lower Cretaceous of Brazil. Palaeontology16: 379-393.
Bertels, A. 1995. The Cretaceous-Tertiary boundary inArgentina and its ostracodes. En: J. Ríha (ed.), Ostra-coda and Biostratigraphy, Balkema, Rotterdam,Brookfield. Proceedings of the Twelfth InternationalSymposium on Ostracoda (Praga, 1994). pp. 163-170.
Bertels, A. y Martínez, D. 1997. Ostrácodos holocenosde la desembocadura del Arroyo Napostá Grande,sur de la provincia de Buenos Aires, Argentina. Re-vista Española de Micropaleontología 29: 29-69.
Bertels-Psotka, A. 2000. Ostrácodos (Arthropoda,Crustacea) de hábitat límnico de la Formación Collón-Curá, provincia del Chubut. Ameghiniana 37 (1): 39-45.
Bertels-Psotka, A. y Cusminsky, G.C. 1999. Nuevas es-pecies de ostrácodos de la Formación Ñirihuau (Oli-goceno) en su área tipo (alrededores de San Carlosde Bariloche), provincia de Río Negro, RepúblicaArgentina. Ameghiniana 36 (1): 71-81.
Bertels-Psotka, A. y Laprida, C. 1998. Ostrácodos ypaleoambientes holocenos del nordeste de la pro-vincia de Buenos Aires. Ameghiniana 35 (2): 151-162.
Brasier, M. 1980. Microfossils. Allen & Unwin, Londres.194 pp.
Browers, E. M. 1988. Palaeobathymetry on the conti-nental shelf based on examples using ostracods fromthe Gulf of Alaska. En: P. De Dekker y J-P.Peypouquet (eds.), Ostracoda in the Earth Science.Elsevier. pp. 55-76.
Colin, J. P. y Dépêche, F. 1997. Faunes d’ostracodeslacustres des bassins intra-cratoniques d’âge albo-aptien en Afrique de l’Ouest (Cameroun, Tchad) et auBrésil: considerátions d’ordre paléoécologique etpaléobiogéographique. Africa Goescience Review 4:431-450.
Cusminsky, G. C. y Whatley, R. 1996. Quaternary non-marine ostracods from lake beds in Northern Patago-nia. Revista Española de Paleontología 11: 143-154.
Díaz Brito, D., Musacchio, E. A., de Castro, J. C.,Maranhão, M.S., Suárez, J. M. y Rodrigues, R. 2001.Grupo Bauru: uma unidade continental do Cretáceono Brasil- concepções basadas en dados micropa-leontológicos, isotópicos e estratigráficos. Revue dePaléobiologie 20: 245-304.
Díaz-Collell, M. L., Viviers, M. C. y Sánchez-Arango, J.P. 1998. Estudios taxonómicos y bioestratigráficosde las asociaciones de ostrácodos de formacionesdel Cretácico tardío en Cuba central. Reunión con-junta de los proyectos PICG/UNESCO 362 y 381,Correlaciones Mesozoicas en el Atlántico sur, Cre-tácico Boreal y Tethysiano (La Habana, 1998). Geolo-gía y Minería’98, Memorias 2: 277-279.
Díaz Saravia, P. y Herbst, R. 2001. Ostrácodos dulcea-cuícolas de la Formación Yaguarí, Pérmico Superior,R. O. del Uruguay. Ameghiniana 38 (2): 213-218.
Díaz Saravia, P. y Jones, P. J. 1999. New Carboniferous(Namurian) glaciomarine ostracods from Patagonia,Argentina. Journal of Micropalaeontology 18: 97-109.
Dingle, R. V. 1984. Mid-Cretaceous Ostracoda fromSouthern Africa and the Falkland Plateau. Annals ofthe South African Museum 93: 97-211.
Durán, G. y Salazar, E. 1990. Estudio bioestratigráficodel Cretácico Superior en el Ecuador. En: W.Volkheimer (ed.), Bioestratigrafía de los sistemas regio-nales del Jurásico y Cretácico de América del Sur,(Mendoza) 2: 553-567.
Echevarría, A. E. 1984. B-Ostrácodos. En Capítulo II-7,Los microfósiles. Relatorio del Noveno Congreso Geo-lógico Argentino (S. C. de Bariloche, 1984): 502-513.
Fauth, G., Seelig, J., Luther, A. y Bengtson, P. 2002.Ostracodes do Cretáceo Superior da Bacia de JamesRoss, Antártica. Boletim do 6º Simpósio sobre o Cretáceodo Brasil/2º Simpósio sobre el Cretácico de América delSur: 187-190.
Horne, D.J., Cohen, A. y Martens, K., 2002. Taxonomy,morphology and biology of quaternary and livingOstracoda. En: J. Holmes y A. R. Chivas (eds.), TheOstracoda Applications in Quaternary Research. Ame-rican Geophysical Union, Washington D.C.Geophysical Monograph 131: 5-36.
Hou, X., Siveter, D.J., Williams, M., Waloszek, D. yBergström, J. 1996. Appendages of the ArthropodKumingella from the Early Cambrian of China: itsbearing on systematic position of the Bradoriida andthe fossil record of Ostracoda. PhilosophycalTransactions Royal Society of London B 351: 1131-1145.
Kaesler, R. L., Smith S. y Whatley R. C. 1979. Ostracodaand petroleum pollution in the Strait of Magellan. En:N. Krstic (ed.), Taxonomy, biostratigraphy anddistribution on Ostracoda. Proceedings 7º Internatio-nal Symposium on Ostracoda. Serbian Geological SocietyBelgrade. pp. 237-247.

OST
RA
CO
DA
623OSTRACODALOS INVERTEBRADOS FÓSILES
Laprida, C., Gómez, A., Rosemblat, A. y Castañeda, E.2004. Cambios ambientales en la Laguna de Chascomúsmediante el análisis de ostrácodos y datos históricos:estudio preliminar. 22ª Reunión Científica de la Asocia-ción Argentina de Geofísicos y Geodestas (Buenos Aires,Septiembre 2004). Resúmenes: 82-83.
Latreille, P. A. 1806. Genera crustaceorum et insectorum.Volumen 1. Paris et Argentorati, Amand Koenig,bibliopolarumum. 303 pp.
Marengo, H. G. 2000. Rasgos micropaleontológicosde los depósitos de la trasgresión Entrerriense-Paranense en la cuenca Chaco-Paranense y Noroes-te Argentino. En: F. G. Aceñolaza y R. Herbst (eds.),El Neógeno de Argentina. Instituto Superior de Corre-lación Geológica (INSUGEO), Serie de CorrelaciónGeológica 14. pp. 29-45.
Moore, R.C. y Pitrat C. W. (eds.). 1961. Treatise onInvertebrate Paleontology. Part Q Arthropoda 3.Crustacea. Ostracoda. Geological Society of Americaand University of Kansas Press. 442 pp.
Musacchio, E. A. 1989. Biostratigraphy of the non-marineCretaceous of Argentina based on calcareousmicrofossils. En: J. Wiedmann (ed.), Cretaceous ofthe Western Tethys. Proceedings of the Third Internatio-nal Cretaceous Symposium (Tübingen, 1987). pp. 812-850.
Musacchio, E. A. 1990. Non-marine Cretaceous ostra-cods from Argentina and their palaeobiogeographicalrelationships. En: R. Whatley y C. Maybury (eds.),Ostracoda and Global Events, British Micropalaeonto-logical Society Publication Series and Chapman &Hall. pp. 557-569.
Pang Qiqing y Whatley, R. 1990. The biostratigraphicalsequence of Mesozoic non-marine ostracodassemblages in northern China. En: R. Whatley y C.Maybury (eds.), Ostracoda and Global Events. BritishMicropalaeontological Society Publication Seriesand Chapman & Hall. pp. 239-250.
Riccardi, A.C., Damborenea, S. E., Manceñido, M. O. yBallent, S. C. 1999. El Jurásico y Cretácico de la Pre-cordillera Principal y la cuenca Neuquina. 3. Bioes-tratigrafía. En: R. Caminos (ed.), Geología Argentina.Subsecretría de Minería de la Nación, Servicio Geo-lógico Minero Argentino, Instituto de Geología yRecursos Minerales, Anales 29. pp. 419-432.
Rodrigo, L. A., Branisa, L. y Musacchio, E. A. 1990. Lascuencas de los Grupos Puca y Tacurú (Cretácico-Terciario Inferior) de Bolivia. En: W. Volkheimer(ed.), Bioestratigrafía de los sistemas regionales delJurásico y Cretácico de América del Sur, (Mendoza) 2.pp. 569-623.
Rossi de García, E. 1972. Ostrácodos del Carboníferodel Sistema de Tepuel. Revista Española de Micropa-leontología 4: 27-29.
Rossi de García, E. 1975. Cuyanocopina, un nuevosuborden de Ostracoda paleozoico. Primer Congresode Paleontología y Bioestratigrafía (Tucumán, 1975),Actas 1: 59-76.
Rossi de García, E. y Proserpio, C. 1978. Ostrácodosordovícicos de la Precordillera de San Juan (Hoja18c, San José de Jáchal). Ameghiniana 15 (3-4): 391-405.
Salas, M. J. 2002 a. Ostrácodos binodicopas del Ordoví-cico de la Precordillera de San Juan, Argentina.Ameghiniana 39 (1): 41-58.
Salas, M. J. 2002 b. Ostrácodos podocopas del Ordoví-cico de la Precordillera de San Juan, Argentina.Ameghiniana 39 (2): 129-149.
Salas. M. J. 2002 c. Análisis bioestratigráficode la faunade ostrácodos llanvirnianos y caradocianos de la Pre-cordillera de San Juan, Argentina. En: C. A. Cingolani,N. Cabaleri, E. Linares, M. G. López de Luchi, H. O.Ostera y H. O. Panarelo (eds), Actas 15 Congreso Geo-lógico Argentino (Calafate, 2002), Artículo breve, tomo1: 566-571.
Salas, M. J. 2003. Ostrácodos palaeocopas, leicopas yeridostracas del Ordovícico de la Precordillera deSan Juan, Argentina. Ameghiniana 40 (3): 345-360.
Schallreuter, R. E. L. 1973. Die OstracodengattungHyperchilarina und das Aparchites-Problem. GeologiskaFöreningens Stockholm Förhandlingar 95: 37-49.
Schallreuter, R. E. L. 1993. Ostrakoden ausordovizischen Geschieben II. Geologie undPaläontologie Westfalen 27 (Beiträge Zur GeschiebekundeWestfalens II). 273 pp.
Schallreuter, R. E. L. 1996. Ordovizische OstracodenArgentiniens II. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 79.pp. 139-169.
Siveter, D. 1984. Habitats and mode of life of Silurianostracodes. Special Paper of Palaeontology 32: 71-85.
Siveter D. J, Williams, M. y Waloszek D. 2001. Aphosphatocopid crustacean with appendages from theLower Cambrian. Science 293: 479-481.
Siveter, D. J., Sutton, M., Briggs, D.E.G. y Siveter, D. J.2003. An ostracode crustacean with soft parts fromthe Lower Silurian. Science 302: 1747-1751.
Sohn, I. G. y Rocha Campos, A. G. 1990. Late Palaeozoic(Gondwanan) ostracodes in the CorumbataíFormation, Paraná Basin, Brazil. Journal of Paleontology64 (1): 116-128.
Van Morkhoven, F.P.F.M. 1959. The morphology,taxonomy and economic use of post-Paleozoic ostracoda.Part 1: General. Bataafse Internationale PetroleumMaatschappij N.V..Exploration and Production.Paleontological Section. Shell. La Haya. 204 pp.
Viviers, M. C., Koutsoukos, E. A. M., da Silva-Telles Jr.,A. y Bengtson, P. 2000. Stratigraphy andbiogeographic affinities of the late Aptian-Campanianostracods of the Potiguar and Sergipe basins innortheastern Brazil. Cretaceous Research 21: 407-455.
Whatley R. C. 1988. Population structure in ostracods:some general principles for the recognition ofpalaeoenvironments. En: P. De Dekker y J-P.Peypouquet (eds.), Ostracoda in the Earth Science.Elsevier. pp. 245-256.
Whatley, R. C. y Cusminsky, G. C. 1999. LacustrineOstracoda and late Quaternary palaeoenvironmentsfrom the Lake Cari-Laufquen region, Río Negroprovince, Argentina. Palaeogeography, Palaeoclima-tology, Palaeoecology 151: 229-239.
Whatley, R. C. y Cusminsky, G. 2002. Upper PlioceneOstracoda from the Burdwood Bank, SW Atlantic.Revista Española de Micropaleontología 34: 53-80
Whatley, R., Moguilevsky, A., Chadwick, J., Toy, N. yFeijó Ramos, M. I. 1998. Ostracoda from the southwest Atlantic. Part III. The Argentinian, Uruguayanand southern Brazilian Continental Shelf. Revista Es-pañola de Micropaleontología 30: 89-116.

624 OSTRACODA
OST
RA
CO
DA
LOS INVERTEBRADOS FÓSILES
Whatley, R., Moguilevsky, A., Toy, N., Chadwick, J. yFeijó Ramos, M. I. 1997. Ostracoda from the southwest Atlantic. Part II. The littoral fauna from betweenTierra del Fuego and the Río de La Plata. RevistaEspañola de Micropaleontología 29: 5-83.
Zabert, L. L. 1981. Ostrácodos cuartarios de Taco Pozo(provincia del Chaco, Argentina) con algunas con-sideraciones paleoecológicas. FACENA 4: 77-87.
Zabert, L. L. 1985. Ostrácodos de agua dulce del Pérmi-co Superior de Colonia Independencia (DeptoGuaira), Paraguay. Ameghiniana 22 (1-2): 121-131.
Zabert, L. L. y Herbst, R. 1986. Ostrácodospleistocénicos del arroyo Perucho Verna, provinciade Entre Ríos, Argentina. Ameghiniana 23 (3-4): 213-224.




















