
The weak, fine-tuned binding of ubiquitous transcription factors to the II-2 enhancer contributes to...
9
Nucleic Acids Research, Vol. 20, No. 11 2657-2665 The weak, fine-tuned binding of ubiquitous transcription factors to the 11-2 enhancer contributes to its T cell- restricted activity Bernd Hentsch, Athanasia Mouzaki', Isolde Pfeuffer, Duri Rungger2 and Edgar Serfling* Institute of Virology and Immunobiology, University of Wurzburg, Versbacher StraBe 7, D-87 WOrzburg, Germany, 1Haematology, Geneva University Hospital, 25 rue Micheli-du-Crest, CH-1211 Geneva 4 and 2Station de Zoologie Experimentale, University of Geneva, 154 route de Malagnou, CH-1224 Chene-Bougeries, Switzerland Received April 8, 1992; Accepted April 30, 1992 ABSTRACT The T lymphocyte-specific enhancers of the murine and human Interleukin 2 (11-2) genes harbour several binding sites for ubiquitous transcription factors. All these sites for the binding of AP-1, NF-kB or Oct-1 are non- canonical sites, i.e. they differ in one or a few base pairs from consensus sequences for the optimal binding of these factors. Although the factors bind weakly to these sites, the latter are functionally important because their mutation to non-binding sites results in a decrease of inducible activity of the 11-2 enhancer. Conversion of three sites to canonical binding sites of Octamer factors, AP-1 and NF-kB results in a drastic increase in enhancer activity and the induction of the 11-2 enhancer in non-T cells, such as B cell lines, murine L cells and human HeLa cells. The introduction of two or three canonical sites into the enhancer leads to a further increase of its activity. 11-2 enhancer induction is also observed in B cells when the concentration of AP-1 and Oct factors increases as a result of co- transfections with FosB and Octamer expression plasmids. When 11-2 enhancer constructs carrying canonical factor binding sites were injected into Xenopus oocytes the strong binding of ubiquitous factors substantially overcomes the silencing effect of negatively acting factors present in resting primary T lymphocytes. These results suggest a fine-tuned interplay between ubiquitous and lymphoid-specific factors binding to and transactivating the 11-2 enhancer and show that the binding affinity of ubiquitous factors to the enhancer contributes to its cell-type specific activity. Moreover, we believe that a dramatic increase of transcriptional activity brought about by single point mutations at strategic important factor binding sites may also have relevance to the activation of nuclear oncogenes. INTRODUCTION A key event of tissue-specific expression of eukaryotic genes is the control of their transcription which is mediated by numerous transcription factors (see Ref. 1 for a review). Transcription factors are nuclear proteins which bind to DNA and/or to other factors and exert a positive or negative influence on the transcriptional activity of RNA polymerase II (2). Over the last years numerous eukaryotic transcription factors have been isolated and analysed in detail (3). Cell-type restricted as well as ubiquitous factors are involved in the transcriptional control of tissue-specific genes. However, the exact way in which these factors interact is not yet known. The complexity of transcriptional control regions of eukaryotic genes hampers the study of the interactions of transcription factors involved in the control of eukaryotic genes. Very often the gene control regions appear to be widely distributed, divided into promoters, located just upstream of the transcription start and enhancers which can be located several kilobases DNA away, either upstream or downstream from the gene's transcriptional start site or within the transcription unit of the gene (4). Even in situations where the gene's control region spans only one hundred or fewer base pairs of DNA, a large number of factors can interact with these short DNA segments, and even a single protein binding site consisting of a few base pairs DNA can be recognized by multiple factors. One example in this respect is the so-called Octamer motif ATGCAAAT which was first detected as a conserved sequence motif within the promoters of Immunoglobulin (Ig) genes and later also within the promoters and enhancers of many other genes, such as in those controlling the activity of such divergent genes as histone genes, small nuclear RNA genes and viral genes (5). In lymphoid cells two factors interact with the Oct motif, the ubiquitous factor Oct-I and the lymphoid-specific factor Oct-2. The genes encoding both factors have been cloned and their DNA binding and transacting properties have been analysed (5, 6). Mainly due to its lymphoid-specific occurrence, Oct-2 * To whom correspondence should be addressed .. 1992 Oxford University Press
Transcript of The weak, fine-tuned binding of ubiquitous transcription factors to the II-2 enhancer contributes to...

Nucleic Acids Research, Vol. 20, No. 11 2657-2665
The weak, fine-tuned binding of ubiquitous transcriptionfactors to the 11-2 enhancer contributes to its T cell-restricted activity
Bernd Hentsch, Athanasia Mouzaki', Isolde Pfeuffer, Duri Rungger2 and Edgar Serfling*Institute of Virology and Immunobiology, University of Wurzburg, Versbacher StraBe 7, D-87WOrzburg, Germany, 1Haematology, Geneva University Hospital, 25 rue Micheli-du-Crest, CH-1211Geneva 4 and 2Station de Zoologie Experimentale, University of Geneva, 154 route de Malagnou,CH-1224 Chene-Bougeries, Switzerland
Received April 8, 1992; Accepted April 30, 1992
ABSTRACTThe T lymphocyte-specific enhancers of the murine andhuman Interleukin 2 (11-2) genes harbour several bindingsites for ubiquitous transcription factors. All these sitesfor the binding of AP-1, NF-kB or Oct-1 are non-canonical sites, i.e. they differ in one or a few base pairsfrom consensus sequences for the optimal binding ofthese factors. Although the factors bind weakly to thesesites, the latter are functionally important because theirmutation to non-binding sites results in a decrease ofinducible activity of the 11-2 enhancer. Conversion ofthree sites to canonical binding sites of Octamerfactors, AP-1 and NF-kB results in a drastic increasein enhancer activity and the induction of the 11-2enhancer in non-T cells, such as B cell lines, murineL cells and human HeLa cells. The introduction of twoor three canonical sites into the enhancer leads to afurther increase of its activity. 11-2 enhancer inductionis also observed in B cells when the concentration ofAP-1 and Oct factors increases as a result of co-transfections with FosB and Octamer expressionplasmids. When 11-2 enhancer constructs carryingcanonical factor binding sites were injected intoXenopus oocytes the strong binding of ubiquitousfactors substantially overcomes the silencing effect ofnegatively acting factors present in resting primary Tlymphocytes. These results suggest a fine-tunedinterplay between ubiquitous and lymphoid-specificfactors binding to and transactivating the 11-2 enhancerand show that the binding affinity of ubiquitous factorsto the enhancer contributes to its cell-type specificactivity. Moreover, we believe that a dramatic increaseof transcriptional activity brought about by single pointmutations at strategic important factor binding sitesmay also have relevance to the activation of nuclearoncogenes.
INTRODUCTIONA key event of tissue-specific expression of eukaryotic genes isthe control of their transcription which is mediated by numeroustranscription factors (see Ref. 1 for a review). Transcriptionfactors are nuclear proteins which bind to DNA and/or to otherfactors and exert a positive or negative influence on thetranscriptional activity of RNA polymerase II (2). Over the lastyears numerous eukaryotic transcription factors have been isolatedand analysed in detail (3). Cell-type restricted as well asubiquitous factors are involved in the transcriptional control oftissue-specific genes. However, the exact way in which thesefactors interact is not yet known.The complexity of transcriptional control regions of eukaryotic
genes hampers the study of the interactions of transcription factorsinvolved in the control of eukaryotic genes. Very often the genecontrol regions appear to be widely distributed, divided intopromoters, located just upstream of the transcription start andenhancers which can be located several kilobases DNA away,either upstream or downstream from the gene's transcriptionalstart site or within the transcription unit of the gene (4). Evenin situations where the gene's control region spans only onehundred or fewer base pairs of DNA, a large number of factorscan interact with these short DNA segments, and even a singleprotein binding site consisting of a few base pairs DNA can berecognized by multiple factors.One example in this respect is the so-called Octamer motif
ATGCAAAT which was first detected as a conserved sequencemotif within the promoters of Immunoglobulin (Ig) genes andlater also within the promoters and enhancers of many othergenes, such as in those controlling the activity of such divergentgenes as histone genes, small nuclear RNA genes and viral genes(5). In lymphoid cells two factors interact with the Oct motif,the ubiquitous factor Oct-I and the lymphoid-specific factorOct-2. The genes encoding both factors have been cloned andtheir DNA binding and transacting properties have been analysed(5, 6). Mainly due to its lymphoid-specific occurrence, Oct-2
* To whom correspondence should be addressed
.. 1992 Oxford University Press

2658 Nucleic Acids Research, Vol. 20, No. 11
has been implicated in the control of the activity of lymphoid-specific genes, e.g. the activity of Ig genes, whereas Oct-i isthought to control genes which are ubiquitously expressed, e.g.the snRNA genes. Although direct functional tests havedemonstrated a preference of Oct-I for the transactivation ofsnRNA promoters and of Oct-2A for mRNA promoters (7),several other parameters play an additional role in the gene controlmediated by Oct-I and Oct-2.One such additional parameter is the interaction of Oct-I with
other transcription factors which help to enhance the relativelypoor transacting potency of Oct-1. The interaction of Oct-I withVP-16, a very strong viral transactivator, and an additional hostcell factor has been studied in detail (see Ref. 8 for a review)and it has been shown that the interaction with VP-16 alters theregulatory capacity of Oct-1. VP-16 acts as a transcriptionalmodulator which allows Oct-I to stimulate a new set ofpromoters.
A.
Further parameters which affect the transcriptional control ofOctamer factors are the binding affinity of their target sites andthe factor concentration in a given cell type. Functional tests ofOctamer DNA containing promoters in transient transfectionstudies revealed a clear correlation between the binding affinityof the site and its cell type-specific activity (9). Promoterscontaining high affinity sites were found to direct transcriptionin B cells as well as non-B cells, whereas the activity of promoterswith low affinity sites appeared to be restricted to B cells. Theactivity of promoters with low affinity octamer sites could beactivated in non-B cells by co-transfection with Oct-i or Oct-2expression plasmids, i.e. by an enhancement of factorconcentration (9).One of the most prominent factor binding sites within the
transcriptional control (enhancer) region of Interleukin 2 (11-2)gene is a non-canonical Octamer binding site (10, I1). This siteplays an important role in the T cell-specific induction of 11-2
Zt- 15>0 ~-,10i
r. Ih : G(Ai-f(i6\(l TCA.P|: l (iA AAIAi| iA; AK[i<,: AA\1:: TGiTTA\1tIKp:AGAGTCA: ATG(7AAAl
NFATI- P,!\ A3 NFAT-I \v._~~~~~~~~~~~~~~~~ ......_T A ' rA T
AP lp: IAGA.(TC>A
B.exti-tit E14
j;.., rLr .r It
:...: e
S-__-_-.
C.extrac:t E14irldction + +
I)IOo RE, 1SAT TRFp I53T
W. AP I.ct2B21:r2A
D.ext1,ac: E14induction + +probe T0 All v Aj
as.!-. 0 ~-,M
-
4~2 3 4
Fig. 1. Scheme of the murine Interleukin 2 enhancer, the sequences and factor binding of non-canonical sites for ubiquitous factors. A. The murine 11-2 enhancer.The most prominent protein binding sites are indicated by boxes. The two TATA boxes, TATA-l and TATA-2, are shown as stippled boxes, the protein bindingsites for which no function has been detected so far (the proximal T cell element, TCEp, and the upstream sites, US-I and US-2/3) by open boxes. The filled boxesindicate protein binding sites which are involved in the establishing of inducible enhancer activity in E14 T lymphocytes. These are the Mpstream promoter site,UPS, the two Purine boxes, Pu-bp and Pu-bd, the proximal TPA responsive element, TREp, and the distal T cell element, TCEd (see Refs. 13, 23-25). The positionsand sequences of four non-canonical binding sites for ubiquitous factors are shown. Nucleotides which differ from canonical consensus sequences are underlined.B. Binding of Oct factors to UPS DNA. EMSA. A radioactively labelled UPS probe (spanning the nucleotides from position -64 to -94 of 11-2 enhancer)(lanes1 and 3) or an UPSO probe carrying a canonical Octamer binding site (see Materials and methods)(lanes 2 and 4) were incubated with 4 iLg protein of crude nuclearextracts from uninduced (-) or induced E14 cells (+) treated for 4 h with TPA/ConA (10 ng/2.5 isg per ml) on ice followed by electrophoresis trough a non-denaturing5% polyacrylamide gel. The positions of Oct complexes are indicated. C. Binding of AP- I like factors to TREp DNA. A TREp-WT probe spanning the nucleotidesfrom -143 to -157 (lanes 1 and 3) or a 153T-TREp probe carrying a canonical TRE (lanes 2 and 4) were incubated with nuclear extracts of uninduced (-) orinduced E14 cells (+) and treated as described above. The position of AP-1 complex is indicated. D. Binding of NF-kB like factors to the WT-TCEd spanningthe nucleotides from -191 to -215 (lanes 1 and 3) and the TCEd A/C probe containing a canonical kB-binding site (lanes 2 and 4). Note the strong binding ofNF-kB to the TCEd A/C DNA.

Nucleic Acids Research, Vol. 20, No. 11 2659
gene because mutations which impair the binding of Octamerfactors drastically decrease the 11-2 enhancer activity (11;I.Pfeuffer et al., submitted). Recently, we have identified a non-canonical AP-1 binding site which is closely linked to the Octamersite within the enhancer's so-called upstream promoter site (UPS).We will show elsewhere that the cooperative binding of Octamerand AP-l like factors to the UPS DNA contributes to the overallactivity of 11-2 enhancer (I.Pfeuffer et al., submitted). In thiscommunication we will show that the affinity of Octamer factorsbinding to this site also contributes to the inducible 11-2 enhanceractivity since the conversion of non-canonical Octamer site to
A.extr actinduction ____ 4 + +
probe UPS, + + +
OCT + + +
comP(etitor: UPSr +
OCT +
OCT -_
OCT 2BOCT 2A
2 3 4 5 6
C.extricl _ ___induction + + + +probe: TCEd A/C + + +
xB + + +competitor: TCEd A/C +
kB +
NF-xB
a canonical site leads to a drastic increase of the 11-2 enhanceractivity in T cells and to its activity in non-T cells. Very similarresults were obtained when two other non-canonical binding siteswere converted to canonical sites which are bound by theubiquitous factors AP-1 and NF-kB. Moreover, in B cells theactivity of the otherwise non-active 11-2 enhancer could bestimulated by co-transfections of factors recognizing the 11-2enhancer, such as Fos B and Oct-2. These and furtherexperimental results demonstrate that the binding affinity andconcentration of ubiquitous factors interacting with the 11-2enhancer contribute to its cell type-specific activity.
Bextract E14induction + + + +
probe: 153T-TREp + + +TREcoll + + +
competitor: f53T-TREp +TREcoII +
AP-1----- 2 3 4
1 2 34 5 6
4'a
5 1 4 5 6
Fig. 2. The factor binding to the 'converted' sites of ubiquitous factors within the 11-2 enhancer is very similar, if not identical to those of well-characterized bindingsites of ubiquitous factors. EMSA. A. The binding of Oct factors to the UPSO motif (lanes 1, 3 and 4) and the Oct motif of Ig heavy chain enhancer (lanes 2,5 and 6). Radioactively labelled UPSO DNA (carrying an Oct consensus sequence within the 11-2 sequence from position -64 to -94: see Materials and methods)or the 51 bp Hinfl-DdeI fragment of Ig heavy chain enhancer (47) were incubated with 4 ug protein of crude nuclear extracts from uninduced (lanes 1 and 2) andinduced E14 cells (lanes 3-6) treated for 4 h by TPA/Con A (10 ng/2.5 lAg per ml). In lanes 4 and 6, an 100 fold molar excess of UPSO DNA or Oct DNA(corresponding to a 15 bp oligonucleotide containing an Oct consensus sequence) was added to the incubation for competition. B. Binding of AP-1 to the 153T-TREpand the TREcoll motifs. A 153T-TREp probe (see Fig. 1 and Materials and methods)(lanes 1, 3 and 4) or a TREcoll DNA probe containing the AP-1 site of humancollagenase promoter (48) (lanes 2, 5 and 6) were incubated with nuclear proteins from uninduced (lanes 1 and 2) or induced E14 cells (lanes 3-6). In lanes 4and 6, an 100 fold molar excess of 153T-TREp or TREcoll DNA were added for competition. C. Binding of NF-kB like factors to the TCEd A/C DNA and thecanonical kB-site of Ig kappa enhancer (49). A TCEd A/C DNA probe (spanning the nucleotides from -191 to -215 with a point mutation at position -199)(lanes 1, 3 and 4) and the kB-site of Ig kappa enhancer (lanes 2, 5 and 6) were incubated with nuclear proteins from uninduced (lanes 1 and 2) and induced E14cells (lanes 3-6). In lanes 4 and 6, an 100 fold excess of TCEd A/C DNA or of kB-DNA of Ig kappa enhancer were added for competition.

2660 Nucleic Acids Research, Vol. 20, No. 11
MATERIALS AND METHODS
Cell culture, DNA transfections and CAT assaysMurine E14 lymphoma cells, 70Z pre-B lymphocytes and A20JB lymphoma cells were grown in RPMI medium supplementedwith 5% fetal calf serum to a density of about 2 x I05 cells perml. About 2 xI07 cells were transfected with 10 itg DNA(purified by two CsCl/ethidium bromide density gradientcentrifugations) in a final volume of 1.2 ml using the DEAEdextran protocol (12). After transfection, the cells were shockedwith 15% dimethylsulfoxide for 2 min and, 20 h later, they weredivided. One half of E14 cells was induced with the phorbolesterTPA (12-O-tetradecanoylphorbol-13-acetate; 10 ng per ml) and
1$1,w,W'.100 E14-T
cI
1
*00
1u
100 ,
X 0-g 101
0
-jiSr I
the plant lectin Con A (Concanavalin A; 2.5 tg per ml), the otherhalf was used as uninduced control. (In some E14 cell experimentsthe Ca++-ionophore lonomycin (10 FM) was used instead ofConA). The B cell lines were induced by 50 ng per ml TPA.The cells were incubated for another 20 h, harvested, sonicatedand their CAT activities were measured as described earlier (13).Murine LMTK- and human HeLa cells were grown inDulbecco's modified Eagle's medium containing 5% fetal calfserum. They were transfected using the calcium phosphate co-precipitation method (14) and shocked with 25%dimethylsulfoxide for 3 min. 40 ug DNA was usually transfectedinto two plates of 10 mm diameter each. The cells of one platewere induced by TPA (50 ng per ml), the cells of the other were
'A
- 0+ - + - + - + - 4 +
+ ,- + _+ - + -
4.
100
C
0
100
X2 10
l10
II
+ -4. - + - + - + - +I
HO + -+ - + - . +
J1.1
Ci' -l
C9, 45 cp
4p 'A v
Fig. 3. The conversion of non-canonical binding sites of UPS-Oct, TREp and TCEd DNAs to canonical factor binding sites leads to a strong increase of enhancer'sactivity in E14 T cells and to its inducible activity in non-T cells. 10 jig DNA of the construct pIICAT 2/1 + containing the wild type enhancer in front of the tkpromoter ofCAT construct pBLCAT2 (13), designated as WT, or of constructs carrying a NF-kB consensus sequence (NF-kB (- 199C)), an AP-I consensus sequence(AP-1 (- 153T)) and an Oct-consensus sequence (Oct (-72T/-76T)), or two (AP-1 +Oct) and three (NF-kB+AP-l +Oct) consensus sequences were transfectedinto 2 x 1 E14 cells, 70 Z pre-B lymphocytes or A20J B lymphoma cells using the DEAE dextran tranfection protocol (12). 20 h after transfection, the cells weredivided. One half was used as uninduced control (-), the other half of cells was induced by TPA/ConA (10 ng/2.5 isg per ml) for a further 20 h period. The cellswere harvested, and their CAT activities (corresponding to the%-acetylation) were determined as described (see Ref. 13 and Materials and methods). Note that forthe determination of enhancer activity in E14 cells only one fifth of the protein amount was used in conrast to that of other cells. 40 j&g of each consuct was transfectedinto two 100 mm plates of LMTK- cells or HeLa cells (left column diagram) using the calcium phosphate co-precipitation technique (14). In the HeLa cells ofthe right column diagram, 5 iLg DNA were transfected per plate. The cells of one plate were used as uninduced control (-), the cells of the other plate were inducedby TPA (50 ng per ml) for 20 h (+). The diagrams show the mean results of at least three independent transfection experiments.
I
Nucleic Acids Research, Vol. 20, No. 11 2661
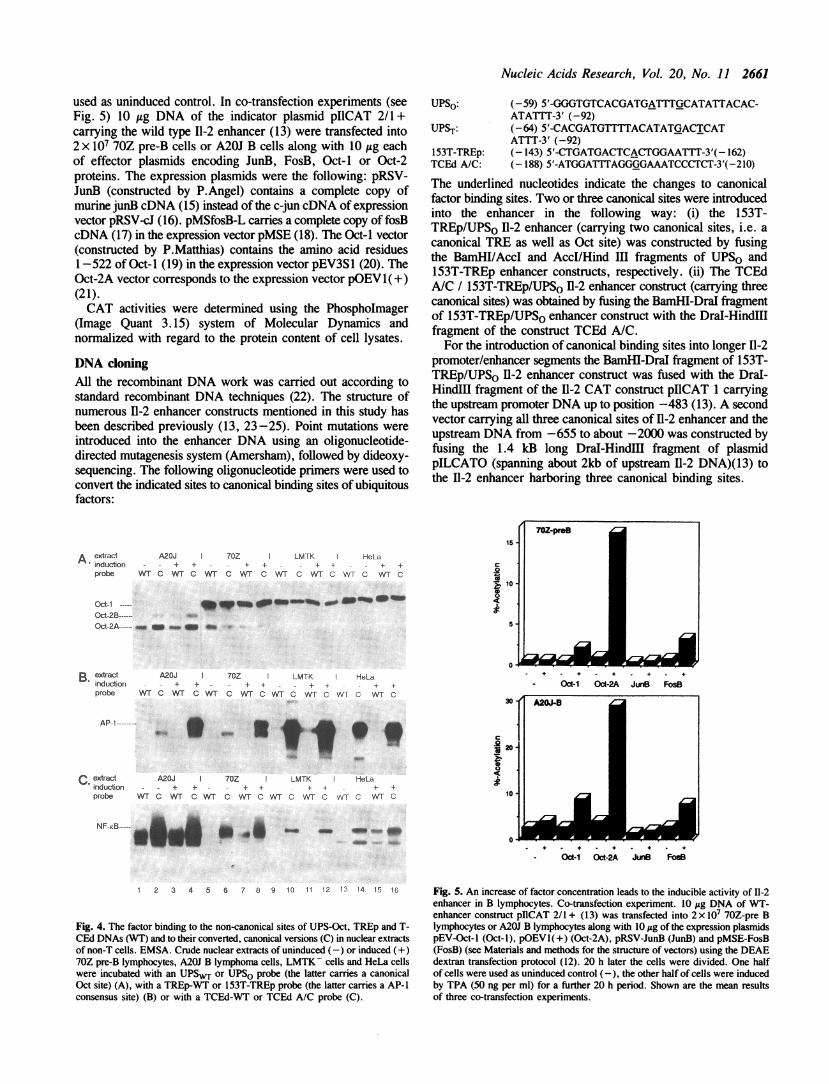
used as uninduced control. In co-transfection experiments (seeFig. 5) 10 itg DNA of the indicator plasmid pIlCAT 2/1 +carrying the wild type 11-2 enhancer (13) were transfected into2 x 107 70Z pre-B cells or A20J B cells along with 10 ug eachof effector plasmids encoding JunB, FosB, Oct-I or Oct-2proteins. The expression plasmids were the following: pRSV-JunB (constructed by P.Angel) contains a complete copy ofmurine junB cDNA (15) instead of the c-jun cDNA of expressionvector pRSV-cJ (16). pMSfosB-L carries a complete copy of fosBcDNA (17) in the expression vector pMSE (18). The Oct-I vector(constructed by P.Matthias) contains the amino acid residues1-522 of Oct-I (19) in the expression vector pEV3S1 (20). TheOct-2A vector corresponds to the expression vector pOEV1( +)(21).CAT activities were determined using the PhosphoImager
(Image Quant 3.15) system of Molecular Dynamics andnormalized with regard to the protein content of cell lysates.
DNA cloningAll the recombinant DNA work was carried out according tostandard recombinant DNA techniques (22). The structure ofnumerous 11-2 enhancer constructs mentioned in this study hasbeen described previously (13, 23-25). Point mutations wereintroduced into the enhancer DNA using an oligonucleotide-directed mutagenesis system (Amersham), followed by dideoxy-sequencing. The following oligonucleotide primers were used toconvert the indicated sites to canonical binding sites of ubiquitousfactors:
A extract A2OJ 7OZ 1 LMTK HeLaWinducton- - + + - - + + + + + +probe WT C wT C VVT C WVT C wT C wr7 C WT C WT C
UPSo: (-59) 5'-GGGTGTCACGATGATTTGCATATTACAC-ATATTT-3' (-92)
UPST: (-64) 5'-CACGATGTTTTACATATGACTCATATTT-3' (-92)
153T-TREp: (-143) 5'-CTGATGACTCACTGGAATTT-3'(- 162)TCEd A/C: (-188) 5'-ATGGATTTAGGGGAAATCCCTCT-3'(-210)
The underlined nucleotides indicate the changes to canonicalfactor binding sites. Two or three canonical sites were introducedinto the enhancer in the following way: (i) the 153T-TREp/UPSo 11-2 enhancer (carrying two canonical sites, i.e. acanonical TRE as well as Oct site) was constructed by fusingthe BamHI/AccI and AccI/Hind III fragments of UPSO and153T-TREp enhancer constructs, respectively. (ii) The TCEdA/C / 153T-TREp/UPSo 11-2 enhancer construct (carrying threecanonical sites) was obtained by fusing the BamHI-DraI fragmentof 153T-TREp/UPSo enhancer construct with the Dral-HindIIIfragment of the construct TCEd A/C.For the introduction of canonical binding sites into longer 11-2
promoter/enhancer segments the BamHI-DraI fragment of 153T-TREp/UPSo 11-2 enhancer construct was fused with the DraI-HindI fragment of the I1-2 CAT construct pUICAT 1 carryingthe upstream promoter DNA up to position -483 (13). A secondvector carrying all three canonical sites of 11-2 enhancer and theupstream DNA from -655 to about -2000 was constructed byfusing the 1.4 kB long Dral-Hindli fragment of plasmidpILCATO (spanning about 2kb of upstream 11-2 DNA)(13) tothe 11-2 enhancer harboring three canonical binding sites.
.2
6ot-t ---- *wtor _f_0_W fd_ so_f_ quo
Oct-2B----- Aw
Oct-2A-- m_ _ *__
B extract A20J 70Z LMTKinducton - - + + - + + + +probe WT C wr C wr C WT C wr C WT C
HeLa
wT c wrT c
+ - + . + . + . +
Octu-I Od J6jl FosB
AP-1 --------
a- _ _ ~~~~~ _
c80
Cextract A2OJ 70Z + LMTK HeLa +induction - - + + - - + + + + + +probe WTC WTTC CC WrC W' C WT C W C WT C
NF-+B--- " *£ _ _ _ U
w~~~4--.
- Oct-1 Oct-2A JunB FosB
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Fig. 4. The factor binding to the non-canonical sites of UPS-Oct, TREp and T-CEd DNAs (WT) and to their converted, canonical versions (C) in nuclear extractsof non-T cells. EMSA. Crude nuclear extracts of uninduced (-) or induced (+)70Z pre-B lymphocytes, A20J B lymphoma cells, LMTK- cells and HeLa cellswere incubated with an UPSWT or UPSO probe (the latter carries a canonicalOct site) (A), with a TREp-WT or 153T-TREp probe (the latter carries a AP-1consensus site) (B) or with a TCEd-WT or TCEd A/C probe (C).
Fig. 5. An increase of factor concentration leads to the inducible activity of 11-2enhancer in B lymphocytes. Co-transfection experiment. 10 Fg DNA of WT-enhancer construct pIICAT 2/1 + (13) was transfected into 2 x I 70Z-pre Blymphocytes or A20J B lymphocytes along with 10 yg of the expression plasmidspEV-Oct-l (Oct-i), pOEVl(+) (Oct-2A), pRSV-JunB (JunB) and pMSE-FosB(FosB) (see Materials and methods for the structure of vectors) using the DEAEdextran transfection protocol (12). 20 h later the cells were divided. One halfof cells were used as uninduced control (-), the other half of cells were inducedby TPA (50 ng per ml) for a further 20 h period. Shown are the mean resultsof three co-transfection experiments.

2662 Nucleic Acids Research, Vol. 20, No. 11
To avoid read-through from the vector's plasmid DNA we alsocloned the 11-2 enhancers with canonical sites into the constructpTKCAT containing two SV40 termination sequences in frontof the pBLCAT2 polylinker (26, 27). In these plasmids we alsoreplaced the long thymidine kinase (tk) promoter (reaching upto -105) by a shorter tk promoter (reaching to -32).
Preparation of nuclear protein extracts and electrophoreticmobility shift assays (EMSAs)The crude nuclear protein extracts used in this study wereprepared according to a published protocol (28). In EMSAexperiments 4 itg protein was incubated with 5000 cpm(equivalent to about 0.2 ng) of a 32P-labelled oligonucleotideprobe and 2 ytg poly dI dC as unspecific competitor as described(29). After incubation for 20-30 min on ice, the samples werefractionated on nondenaturing 5% polyacrylamide gels at200V/15 cm at room temperature followed by autoradiographyof the dried gel.
Xenopus Oocyte injectionsThe oocyte injections were done as described previously (30).5 ng of 300 mM KCl-extracted S100 proteins from resting splenicT cells (see 30 for the extraction procedure) were injected intothe cytoplasm of oocytes (15 per group). 3 h later, 1 ng DNAwas injected into the nuclei of oocytes; i.e. a different DNAconstruct per oocyte group. In parallel, batches of sibling oocytesreceived the construct alone without T cell proteins, in order tomeasure the constitutive transcriptional activity of I1-2promoter/enhancer region in the oocytes. After an incubation for16 h at 20°C, the oocytes were lysed and CAT assays wereperformed as described (30).
RESULTSThe T cell-specific 11-2 enhancer harbours numerous non-canonical, but functionally important binding motifs ofubiquitous factorsThe murine and human 11-2 enhancers span about 275 bp whichharbour numerous binding sites for transacting factors (13,23-25, 31-35). The sequences of these sites are highlyconserved between both enhancers. The most prominent factorbinding sites of the murine I1-2 enhancer are shown in Fig. 1.These are the upstream promoter site, UPS, the two Purine boxes,Pu-bp and Pu-bd, the proximal TPA responsive element, TREp,and the distal T cell element, TCEd.The UPS carries two closely linked non-canonical binding sites
for Octamer and AP-1 like factors (Fig. IA and B). When oneor both sites are mutated and factors are unable to bind, the 11-2enhancer activity decreases to about one third or less of its wildtype activity (I.Pfeuffer et al., submitted; see also 11). Althoughin extracts of T and B lymphocytes the same proteins seem tointeract with UPS DNA, CAT constructs carrying five copiesof UPS DNA (5 x UPS) were inactive in B cells but stronglyinducible in T cell lines, such as in murine E14 lymphoma cellsand human Jurkat lymphoma cells (24 and results not shown).The TREp is a weak binding site of AP-1 or AP-1 like factors.
The affinity of those to TREp DNA is at least 10fold less thanto a canonical AP-1 site, such as the AP-l site of humancollagenase gene (Fig. IC). The weak binding of AP-l to thissite is, however, of functional relevance because introducing a
binding results in a distinct decrease of inducible enhancer activityto about one fourth of the wild type level (S.Muller-Deubert etal., submitted). Very similar results have been reported for theTREp of the human I1-2 enhancer (36).The kB-like element of the 11-2 enhancer which we originally
designated as TCEd (for distal T cell element, since its activitybehaves in a T cell-restricted manner: 13, 25) is a poor bindingsite of authentic NF-kB. In nuclear protein extracts of E14 Tlymphoma cells we detected the binding of three factors to thiselement, one of which, designated as TCF-1, appeared to containthe p5O component of NF-kB (25). The introduction of pointmutations into the TCEd which abolished TCF-l binding led tothe inactivity of the 11-2 enhancer whereas the conversion of theTCEd to a perfect NF-kB binding site resulted in a drastic increaseof enhancer activity in E14 lymphoma cells (Fig. ID) and to itsactivity in HeLa cells (25).
The conversion of non-canonical binding sites to canonicalsites results in a drastic increase of In-2 enhancer activity inT cels and to its activity in non-T cels
In order to investigate whether the conversion of weak bindingsites for ubiquitous factors leads, as reported for the TCEd (25),to an increase of enhancer activity in T cells and its activity innon-T cells, we converted the TREp and Octamer-UPS sequences
to canonical sites in the context of the I1-2 enhancer (Fig. lA).Then we compared the protein binding of 'converted' sites withthat of well-characterized, canonical sites in electrophoreticmobility shift assays (EMSAs) and tested the activity ofcorresponding CAT constructs in transient transfection assays.Furthermore, we constructed 1-2 enhancers carrying two or threeof the 'perfect' mutations, the activity of which was tested inthe same way.As shown in Fig. 2, in EMSAs the converted Octamer-UPS
(UPSO), TREp (153T-TREp) and TCEd (TCEd A/C) probesgenerated the same complexes as canonical binding sites uponincubation with nuclear protein extracts ofE14 T lymphoma cells.This implies that the converted sites are recognized by verysimilar, if not the same factors as canonical binding site, i.e. theUPSO DNA by the Octamer factors Oct-i, Oct-2A and Oct-2B(as well as AP-1: see below), the 153T-TREp by AP-1 and theTCEd A/C site by NF-kB.The conversion of binding sites had a dual effect on the activity
of 11-2 enhancer in E14 cells. As shown in Fig. 3, the presenceof canonical binding sites within the enhancer resulted in anincrease of its constitutive and inducible activity. Approximatelythe same effect was observed for enhancers carrying two or allthree mutations. The strong binding sites also induced activityof the enhancer in B lymphocytes and in non-lymphoid cells, aftertransfection of 11-2 enhancer CAT constructs into 70Z pre-B cells,A20J B cells, L cells and human HeLa cells. Although in thetwo B cell lines the constructs with a converted enhancer showeda high constitutive activity, the activity of several of the constructswas also induced by TPA. This is particularly the case forconstructs carrying two or three canonical sites. Similarly, theseconstructs were found to be induced in L cells and, using differentDNA amounts, in HeLa cells (Fig. 3).
Incubation of non-canonical sites in parallel with theirconverted, canonical versions with nuclear protein extracts fromB cells, L cells and HeLa cells revealed in EMSAs mainlyquantitative differences in factor binding. This is true for all threesites, although, as shown in Fig. 4, the affinity of Oct factors
point mutation (at position -152 of TREp) which abolishes AP-l to the UPS and UPSO differed by a factor of two, whereas that

Nucleic Acids Research, Vol. 20, No. 11 2663
of AP-l to the TREp and 153T-TREp sites by at least one orderof magnitude (see also Figs. lB-ID).
In order to test whether the observed increase of enhanceractivity is not only a particular feature of the enhancer but ageneral property of I1-2 regulatory region, we introduced thecanonical sites into larger promoter fragments, spanning about0.5 and 2kB of upstream DNA. We and others observed anegative effect of upstream DNA sequences on the constitutiveactivity of enhancer (13, 37). Thus, one could expect that thesesequences also exert a negative influence on the enhanced,'unbalanced' activity of enhancers carrying canonical bindingsites. However, in transient transfections, CAT constructscarrying two or three canonical binding sites behave like theircorresponding enhancer CAT constructs (not shown). Theseobservations suggest that the upstream DNA sequences exert atmost a marginal effect on the stronger activity of enhancerscarrying canonical factor binding sites.We also tested the activity of 11-2 enhancer with canonical sites
after their cloning in CAT vectors carrying two SV40 poly Aaddition signals (in front of the tk promoter) and a truncatedversion of tk promoter. Using such vectors, we intent to avoidread-through transcription starting within the plasmid DNA anda possible cooperation of 11-2 enhancer factors with the factorsbinding to the tk promoter. However, no distinct differences inthe CAT activities were observed between such 11-2 CATconstructs and those used in the former experiments (not shown).
An increase of FosB and Octamer factor concentrations leadsto an increase of 11-2 enhancer activity in B cellsWe were also interested to assess whether a higher concentrationof 11-2 enhancer factors leads, like the higher affinity of factor
A. B.
binding sites, to the activity of 11-2 enhancer in non-T cells. Forthis purpose, we co-transfected expression plasmids coding forthe AP-1 proteins JunB and FosB, Oct-I or Oct-2A along withwild type enhancer CAT constructs into 70Z and A20J B cells(we chose JunB and FosB vectors because their expressionresulted in a strong transactivation of 11-2 enhancer in E14 cells:S.Muller-Deubert et al., submitted). As shown in Fig. 5, theexpression of all but the JunB coding plasmids had a stimulatoryeffect on the I1-2 enhancer activity in both cell lines. The co-transfection with the Oct-2A vector led to a dramatic increaseof I1-2 enhancer activity, an observation which supportspreviously published data on the strong transactivating potencyof Oct-2A (38).
The strong binding of NF-kB and Octamer factors to the II-2enhancer also overrides the inhibitory effect of repressorproteins from primary T lymphocytesCo-injections of 11-2 promoter constructs with proteins fromresting T lymphocytes into Xenopus oocytes have revealed theexistence of repressor-like factors in resting, ex-vivo T cells.These factors were found to act through the distal purine boxes,Pu-bd. They are absent in unstimulated T cell lines and non-Tcells and seem, therefore, to be involved in the repression ofthe I1-2 enhancer in primary T lymphocytes (30; A.Mouzaki etal., in prep.). These findings raise the question as to whetherthe increase in I1-2 enhancer activity, caused by the strongerbinding of ubiquitous factors to the I1-2 enhancer, reflects theartificial situation of cultured cell lines which lack the repressorsof primary T lymphocytes. The results of injections using I1-2CAT constructs with canonical binding sites show, however, thatthe tighter binding of NF-kB to the I1-2 enhancer is able to
100 -
80
4, c
X 60
. 40
4W
Pro,c,r
DNA
+ZNl',C vc C> ' c{
.-\ KMQLOIf ,. X
lb0 d )
OI
Fig. 6. The tight binding of NF-kB and Octamer factors to the 11-2 enhancer overrides the silencing activity of repressors in protein extracts from resting splenicT lymphocytes. Xenopus oocyte injections. Oocytes were injected with 5 ng SES100 proteins from resting splenic cells into their cytoplasm and, 3 h later, withI ng DNA of the various CAT constructs (labelled by +). As controls (labeled by -), similar injections were carried out with sibling oocytes which received the11-2 constructs alone (i.e. without the proteins from splenic T cells), to assess the basal activity of the 11-2 enhancer constructs in the oocytes. After an incubationfor 16 h the oocytes were lysed and their CAT activities determined as described (30). A. Autoradiography of a CAT experiment which shows that the tight bindingof NF-kB to the 11-2 enhancer carrying a NF-kB consensus sequence (NF-kB (- 199C)) overrides the silencing activity of factors present in resting splenic T cells.-, constitutive activity of I1-2 enhancer CAT constructs in Xenopus oocytes; +, the effect of proteins from resting splenic T cells (which were injected into Xenopusoocytes prior the injections of DNA constructs) on the activity of constructs. B. The effect of stronger binding of NF-kB, AP- 1 and Octamer factors on the silencingeffect of factors from resting splenic T cells. Shown are the mean values of injections into two different batches of oocytes.

2664 Nucleic Acids Research, Vol. 20, No. 11
substantially override the silencing effect of repressors fromprimary T lymphocytes. The tighter binding of Octamer factorswas less effective in overcoming the silencing effect of repressors,while no effect was observed for the 153T-TREp constructcarrying a converted AP-l binding site (Fig. 6).
DISCUSSION
The results of numerous transient transfection studies have shownthat the enhancers of murine and human I1-2 genes span about275 bp DNA which harbour the most important DNA sequencemotifs for the establishment of inducible and T cell-restrictedtranscription of the 11-2 gene (13, 31-35). Moreover, intransgenic mice the 11-2 enhancer (plus about 250 bp upstreamDNA) appeared to control the expression of a linked indicatorgene in a T cell-specific manner, i.e. the activity of the indicatorgene has only been detected in activated T cells (39). Takentogether, these results show that the T cell-specific transcriptionof the 11-2 gene is controlled by the transacting factors bindingto this immediate upstream (enhancer) region of about 300 bp.The 11-2 enhancer is recognized by a multiplicity of both
positively and negatively acting transcription factors. Among thepositively acting factors are several ubiquitously distributedfactors, such as Oct-l, AP-1 and NF-kB (10, 11, 13, 36, 40-42),but also lymphoid-specific factors, such as Oct-2 (10), NFAT-1(23, 43) and additional factors binding to the kB-like (TCEd)motif of the enhancer (25). Among the negatively factors arefactors interacting with the Purine boxes, i.e. the binding sitesof NFAT-1 (30), and a Zn+ + finger containing factor which hasrecently been described to interact with DNA sequences locatedjust upstream of the UPS motif (44). Thus, it appears likely thatthe T lymphocyte-restricted, inducible and transient expressionof the I1-2 gene is mediated at the level of transcription by a fine-tuned interplay of positively and negatively acting protein factorsbinding to the enhancer DNA. When the equilibrium of factorsbinding to and transactivating the 11-2 enhancer is changed theproper regulation of enhancer activity becomes impaired. Suchchanges can be brought about in several ways, such as by changesof concentration or affinity of factors binding to enhancer DNA.Both situations were artificially created in experiments in thisstudy. By co-transfections of Oct and FosB expression plasmidsinto B cell lines the activity of the 11-2 enhancer could be induced(Fig. 5). A similar effect was observed when the binding affinityof three poor binding sites for ubiquitous factors was enhancedby the conversion of non-canonical binding sites to canonicalones. This was shown for a kB-like sequence element (the T-CEd), an AP- I site (the TREp) and an Octamer binding site (theUPS).Not in every case did the conversion of a non-canonical binding
site for ubiquitous factors to a canonical site lead to an increaseof I1-2 enhancer activity. One exception was the non-canonicalUPS-TRE sequence (Fig. IA). When this site was convertedwithin the enhancer to a perfect TRE by the introduction of twopoint mutations (as shown in Fig. IA) and tested in transienttransfections, the activity of the 11-2 enhancer remained the sameas the wild type enhancer or was even reduced in its inducibleactivity in E14 cells (I.Pfeuffer et al., submitted). The reductionof 11-2 enhancer activity is probably due to a weaker binding ofOct factors to the converted UPS DNA. We will show elsewherethat the Oct factors not only contact the nucleotides within the
Oct-like motif ATGTAAAA of UPS DNA but also severalnucleotides of flanking DNA, including a T residue which islocated at the opposite DNA strand and complementary to theunderlined A of the TRE-like sequence TGTGTAA at position-82 of enhancer DNA. When the UPS-TRE was converted toa canonical TRE the A/T base pair was substituted by a C/Gbase pair, which is not longer contacted by the Oct factors butby AP-1. Thus, in this case the conversion of a non-canonicalAP-1 binding site does not only result in a stronger binding ofAP-1 but, in addition, to a decrease of the binding of Octamerfactors and 11-2 enhancer activity (I.Pfeuffer et al., submitted).A dual effect on the factor binding has also been observed when
the kB-like TCEd sequence was converted to a canonical NF-kB binding site (25). In crude nuclear extracts of induced E14cells (and of other cells) NF-kB binds much more strongly toa 'converted' TCEd carrying a canonical NF-kB site than to thewild type TCEd (Fig. ID). In contrast, in enriched preparationsof TCEd factors from E14 cells two DNA binding factors,designated as TCF-2 and TCF-3, were unable to bind to a NF-kB site but specifically bound to TCEd DNA (25). A third, kB-like factor, designated as TCF-1, was able to bind to bothsequences. Since the binding of TCF-2 and TCF-3 to a DNAsequence element was found to correlate with the loss of itsactivity, we concluded that both factors might exert a repressor-like function (25). Thus, in the case of the TCEd motif, itsconversion to a canonical NF-kB site seems to result not onlyin a tighter binding of a positively acting factor, but also in theloss of binding of repressor-like factors.We believe that the phenomenon described here for the murine
I1-2 enhancer, i.e. the dramatic increase of its activity by onepoint mutation at a non-canonical binding site for transactingfactors, might play an important role in the activation of nuclearoncogenes, which code for transcription factors (see 45 for arecent review). The occurrence of non-canonical binding sitesfor ubiquitous factors is no special peculiarity of the 11-2 enhancer.Instead, such sites exist within the promoters and enhancers ofmany eukaryotic genes (see the Refs. 5 and 46 for compilationson the occurence of non-canonical Octamer and NF-kB sites,respectively) and their conversion to canonical sites might alsolead to a similar enhanced activity as we observed for the 11-2enhancer. If one anticipates that such 'up' mutations occur withinthe promoters of (nuclear) proto-oncogenes during thedifferentiation of cells, cells will appear which contain a muchhigher level of a certain transcription factor than normal cells.The overproduction of such important factor proteins as Fos, Mycetc. can lead to the activation (or repression) of novel sets ofgenes and, in combination with other events, to profound changesin cell growth and differentiation and, finally, in carcinogenesis.
ACKNOWLEDGEMENTSFor excellent technical assistance we wish to thank Elke Schorr.We are indebted to Dr. Catherine Phillips for critical readingthe manuscript. For gifts of DNA constructs we thank Drs.P.Angel (Karlsruhe), Iris Kemler (Zurich), P.Matthias (Basel),W.Schaffner (Zurich) and M.Schuermann (Marburg). This workwas supported by grants of the Deutsche Forschungsgemeinschaft(DFG), SFB 165 (University Wurzburg), the Bundesgesund-heitsamt, research programme AIDS, and the Swiss NationalFoundation grant 3.12 86 54.90.

Nucleic Acids Research, Vol. 20, No. 11 2665
REFERENCES1. Renkawitz,R. (Ed.)(1989) Tissue specific gene expression. VCH
Verlagsgesellschaft Weinheim.2. Mitchell,P.J. and Tjian,R. (1989) Science 245, 371 -378.3. Faisst,S. and Meyer,S. (1992) Nucleic Acids Res. 20, 3-26.4. Serfling,E., Jasin,M. and Schaffner,W. (1985) Trends in Gen. 1, 224-230.5. Schreiber,E., Muller,M.M., Schaffner,W. and Matthias,P. (1989) in Tissue
specific gene expression. R.Renkawitz (Ed.). VHC VerlagsgesellschaftWeinheim, pp. 33-54.
6. Scholer,H.R. (1991) Trends in Gen. 7, 323-329.7. Tanaka,M., Lai,J.-S. and Herr,W. (1992) Cell 68, 755-767.8. Stern,S. and Herr,W. (1991) Gen. Dev. 5, 2555-2566.9. Kemler,I., Bucher,E., Seipel,K., Muller-Immergluck,M.M and Schaffner,W.
(1991) Nucleic Acids Res. 19, 237-242.10. Kamps,M.P., Corcoran,L., LeBowitz,J.H. and Baltimore,D. (1990)
Mol.Cell.Biol. 10, 5464-5472.11. Ullman,K.S., Flanagan,W.M., Edwards,C.A. and Crabtree,G.R. (1991)
Science 254, 558-562.12. Luthman,H. and Magnusson,G. (1983) Nucleic Acids Res. 11, 1295-1308.13. Serfling,E., Barthelmas,R., Pfeuffer,I., Schenk,B., Zarius,S., Swoboda,R.,
Mercurio,F. and Karin,M. (1989) EMBO J. 8, 465-473.14. de Villiers,J. and Schaffner,W. (1983) in Flavell,R.A. (Ed.), Techniques
in Nucleic Acid Biochemistry, Vol. B5, Elsevier, Ireland, pp. 1-20.15. Ryder,K., Lan,L.F. and Nathans,D. (1988) Proc.Natl.Acad.Sci. USA 85,
1487- 1491.16. Angel,P., Hattori,K., Smeal,T. and Karin,M. (1988) Cell 55, 875-885.17. Mumberg,D., Lucibello,F.C., Schuermann,M. and Muller,R. (1991) Gen.
Dev. 5, 1212-1223.18. Schuermann,M. (1990) Nucleic Acids Res. 18, 4945.19. Sturm,R.A., Das,G. and Herr,W. (1988) Gen.Dev. 2, 1582-1599.20. Matthias,P., Muller,M.M., Schreiber,E., Rusconi,S. and Schaffner,W. (1989)
Nucleic Acids Res. 17, 6418.21. Muller,M.M., Ruppert,S., Schaffner,W. and Matthias,P. (1988) Nature 336,
544-551.22. Sambrook,J., Fritsch,E.F. and Maniatis,T. (1989) Molecular Cloning. A
laboratory Manual. 2.Edition. Cold Spring Harbor Laboratory Press. ColdSpring Harbor, N.Y.
23. Randak,C., Brabletz,T., Hergenrother,M., Sobotta,I. and Serfling,E. (1990)EMBO J. 9, 2529-2536.
24. Brabletz,T., Pietrowski,I. and Serfling,E. (1991) Nucleic Acids Res. 19,61-67.
25. Briegel,K., Hentsch,B., Pfeuffer,I. and Serfling,E. (1991) Nucleic AcidsRes. 19, 5929-5936.
26. Luckow,B. and Schutz,G. (1987) Nucleic Acids Res. 15, 5490.27. Scholer,H.R., Balling,R., Hatzopoulos,A.K., Suzuki,N. and Gruss,P. (1989)
EMBO J. 8, 2551-2557.28. Schreiber,E., Matthias,P., Muller,M.M. and Schaffner,W. (1989) Nucleic
Acids Res. 17, 6419.29. Schreiber,E., Matthias,P., Muller,M.M. and Schaffner,W. (1988) EMBO
J. 7, 4221-4229.30. Mouzaki,A., Weil,R., Muster,L. and Rungger,D. (1991) EMBO J. 10,
1399-1406.31. Fujita,T., Shibuya,H., Ohashi,T., Yamanishi,K. and Taniguchi,T. (1986)
Cell 46, 401-407.32. Durand,D.B., Bush,M.R., Morgan,J.G., Weiss,A. and Crabtree,G. (1987)
J.Exp.Med. 165, 395-407.33. Durand,D.B., Shaw,J.-P., Bush,M.R., Replogle,R.E., Belagaje,R. and
Crabtree,G.R. (1988) Mol.Cell.Biol. 8, 1715-1724.34. Williams,T.M., Eisenberg,L., Burlein,J.E., Norris,C.A., Pancer,S., Yao,D.,
Burger,S., Kamoun,M. and Kant,J.A. (1988) J.Immunol. 141, 662-666.35. Brunvand,M.W., Schmidt,A. and Siebenlist,U. (1988) J.Biol.Chem. 263,
18904-18910.36. Jain,J., Valge-Archer,V.E., Sinskey,A.J. and Rao,A. (1992) J.Exp.Med.
175, 853-862.37. Novak,T.J., White,P.M. and Rothenberg,E.V. (1990) Nucleic Acids Res.
18, 4523-4533.38. Tanaka,M. and Herr,W. (1990) Cell 60, 375-386.39. Crabtree,G.R. (1989) Science 243, 355-361.40. Lenardo,M.J., Kuang,A., Gifford,A. and Baltimore,D. (1988)
Proc.Natl.Acad.Sci. USA 85, 8825-8829.41. Hoyos,B., Ballard,D.W., B6hnlein,E., Siekevitz,M. and Greene,W.C. (1989)
Science 244, 4567-460.42. Shibuya,H., Yoneyama,M. and Taniguchi,T. (1989) Internat. Immunol. 1,
43-49.
43. Shaw,J.-P., Utz,P.J., Durand,D.B., Toole,J.J., Emmel,E.A. andCrabtree,G.R. (1988) Science 241, 202-205.
44. Williams,T.M., Moolten,D., Burlein,J., Romano,J., Bhaerman,R.,Godillot,A., Mellon,M., Rauscher III,F.J. and Kant,J.A. (1991) Science254, 1791-1794.
45. Lewin,B. (1991) Cell 64, 303-312.46. Zabel,U., Schreck,R. and Baeuerle,P.A. (1991) J.Biol.Chem. 266, 252-260.47. Gerster,T., Matthias,P., Thali,M., Jiricny,J. and Schaffner,W. (1987) EMBO
J. 6, 1323-1330.48. Angel,P., Imagawa,M., Chiu,R., Stein,P., Imbra,R.J., Rahmsdorf,H.J.,
Jonat,C., Herrlich,P. and Karin,M. (1987) Cell 49, 729-739.49. Pierce,J.W., Lenardo,M. and Baltimore,D. (1988) Proc.Natl.Acad.Sci. USA
85, 1482-1486.




















