
Kip's Role as Mediator in Michael Ondaatje's The English ...
Upload
independentCategory
view
3download
0
The structure of human collapsin response mediator protein 2,a regulator of axonal growth
Pal Stenmark,* Derek Ogg,* Susanne Flodin,* Alex Flores,* Tetyana Kotenyova,* Tomas Nyman,*Par Nordlund* and Petri Kursula�
*Structural Genomics Consortium, Department of Medical Biochemistry and Biophysics, Karolinska Institute, Stockholm, Sweden
�Department of Biochemistry, Oulu University, Oulu, Finland
Abstract
Axonal growth cone guidance is a central process in nervous
system development and repair. Collapsin response mediator
protein 2 (CRMP-2) is a neurite extension-promoting neuronal
cytosolic molecule involved in the signalling of growth inhibi-
tory cues from external stimuli, such as semaphorin 3A and
the myelin-associated glycoprotein. We have determined the
crystal structure of human tetrameric CRMP-2, which is
structurally related to the dihydropyriminidases; however, the
active site is not conserved. The wealth of earlier functional
mapping data for CRMP-2 are discussed in light of the three-
dimensional structure of the protein. The differences in
oligomerisation interfaces between CRMP-1 and CRMP-2 are
used to model CRMP-1/2 heterotetramers.
Keywords: collapsin response mediator protein 2, crystal
structure, growth cone guidance, neuronal regeneration, tetr-
amer, TIM barrel.
J. Neurochem. (2007) 101, 906–917.
During the development and function of the nervous system,a key process is the coordinated growth of neuronal axons totheir targets. Both attractive and repulsive signals are used inorder for the neuronal growth cone to find its appropriatetarget. Semaphorins are a large family of axon guidanceproteins. The best characterised one, semaphorin 3 (alsocalled collapsin), is a secreted protein acting as a chemore-pellent for axonal growth.
The collapsin response mediator proteins [CRMP; thefamily is also known as Unc-33-like proteins (Ulip), dihydro-pyriminidase-related proteins (DRP), TUC (TOAD/Ulip/DRP) and dihydropyriminidase-like proteins] are a family ofcytosolic proteins, CRMP-1 to CRMP-5 in humans, related todihydropyriminidases (DHPs) (Hamajima et al. 1996; WangandStrittmatter 1996).Out of theCRMP family, CRMP-2 is byfar the most extensively characterised member. It has beenshown to be O-glycosylated (Cole and Hart 2001) and to havespecific functions within the nervous system (see below)(Arimura et al. 2004). The expression of CRMP-2 in thenervous system is developmentally regulated (Kamata et al.1998; Inagaki et al. 2000). Overexpression of CRMP-2induces the growth of numerous axons; it is also involved inthe maturation of neurites and pre-existing dendrites to axons(Yoshimura et al. 2005). It plays an important role in axonalguidance, being a mediator of semaphorin 3A signalling(Brown et al. 2004). CRMP-2 is phosphorylated by GSK-3
(Cole et al. 2004) and Rho kinase (Arimura et al. 2000; Guet al. 2000). The phosphorylation by GSK-3 is primed byphosphorylation of CRMP-2 by Cdk5 (Cole et al. 2006).
A specific case of axonal pathfinding is neuronal injury, inwhich case a major inhibitory factor for growth coneextension is comprised of the proteins of the myelin sheath.CRMP-2 is involved in the regulation of growth conecollapse by myelin-related inhibitors, and its activation canbe induced by the binding of the myelin-associated glyco-protein (MAG) on the axonal surface (Mimura et al. 2006). Itis also expressed in oligodendrocytes, the myelinating cellsof the CNS, in a developmentally regulated manner (Ricardet al. 2000), but its exact functions during myelinationremain to be characterised. A number of studies have alsosuggested a role for CRMP-2 in the aetiology of neurologicaldisorders, including Alzheimer’s disease (Cole et al. 2004;
Received October 16, 2006; revised manuscript received November 14,2006; accepted November 16, 2006.Address correspondence and reprint requests to Petri Kursula,
Department of Biochemistry, FIN-90014 University of Oulu, Oulu,Finland. E-mail: [email protected] used: DHP, dihydropyriminidase; DRP, dihydropyri-
minidase-related proteins; CRMP, collapsin response mediator protein;MAG, myelin-associated glycoprotein; SDS–PAGE, sodium dodecylsulfate–polyacrylamide gel electrophoresis; TIM, triosephosphate iso-merase; Ulip, Unc-33-like proteins.
Journal of Neurochemistry, 2007, 101, 906–917 doi:10.1111/j.1471-4159.2006.04401.x
906 Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917� 2007 The Authors

Czech et al. 2004; Kanninen et al. 2004; Sultana et al.2006). Roles for CRMP-2 have also been suggested in non-neuronal cells (Tahimic et al. 2006).
Collapsin response mediator protein 2 has 58% sequenceidentity with the human enzyme DHP, which catalyses thesecond step in pyrimidine degradation; however, no catalyticactivity has been observed for any member of the CRMPfamily, and key active site residues of DHP are not conservedin the CRMPs. It, thus, appears that the CRMPs constitute anexample of enzymes that have lost their catalytic functionand gained a regulatory role instead. In this respect, it is ofinterest that CRMP-2 can interact with DHP (Wang andStrittmatter 1997). CRMP-2 has two splice isoforms, differ-ing by the presence of an additional N-terminal sequence inthe CRMP-2A isoform. The expression pattern and functionof the two isoforms are different (Yuasa-Kawada et al. 2003;Bretin et al. 2005).
Collapsin response mediator protein 2 is able to regulatemicrotubule dynamics (Gu and Ihara 2000), and it directlybinds to tubulin heterodimers (Fukata et al. 2002). Thephosphorylation of CRMP-2 by Rho kinase in turn inhibitsits binding to tubulin; however, CRMP-2 can bind actinirrespective of its phosphorylation status (Arimura et al.2005). Rho phosphorylation of CRMP-2 is an event down-stream ofMAG binding to the neuronal surface (Mimura et al.2006). CRMP-2 also interacts with Numb, an endocytosis-related protein, possibly indicating a role in the endocytoticrecycling of the adhesion molecule L1 at the neuronal growthcone (Nishimura et al. 2003). Attempts to map the bindingsites for both tubulin andNumb have beenmade using deletionexperiments (Fukata et al. 2002; Nishimura et al. 2003). Thephosphorylation sites, and most likely also the glycosylationsites, lie within the disordered C terminus of the protein.
We have determined the crystal structure of humanCRMP-2. The structure is used to analyse differencesbetween members of the CRMP family and other homolog-ous proteins. Furthermore, existing data on CRMP-2 andits interactions are discussed based on the structure. Aplausible model, based on the crystal structures of CRMP-1and CRMP-2, for a heterotetramer between CRMP-1 andCRMP-2 is also presented.
Materials and methods
Cloning and expression
The coding sequence for human CRMP-2 (residues 13–490),
excluding the C-terminal region predicted to be disordered, was
subcloned into the expression vector pNIC-Bsa4, resulting in a
construct with an N-terminal hexahistidine tag and an integrated TEV
protease cleavage site. The plasmid was transformed into Escherichiacoli BL21(DE3) cells. For expression, cells were grown in 1500 mL
Terrific Broth (containing 7% glycerol, 50 lg/mL kanamycin and
100 lLBREOX) in bubble flasks in the large scale expression system
(developed at Structural Genomics Consortium, Toronto).
Cells were grown at +37�C until an optical density of 2.5 at
600 nm was reached. The cultures were down-tempered to +18�Cfor 1 h in a water bath. The expression of CRMP-2 was induced by
the addition of 0.5 mmol/L IPTG, and expression was allowed to
continue overnight at +18�C.Cells were harvested by centrifugation, and the pellets were
suspended in IMAC buffer 1 (50 mmol/L sodium phosphate pH 7.5,
500 mmol/L NaCl, 10% glycerol), supplemented with Complete
EDTA-free protease inhibitors (Roche, Basel, Switzerland), and
frozen at )80�C until used.
The cells were briefly thawn in warm water, and 2000 U of
benzonase were added. The disruption of the cells was performed by
three rounds of high-pressure homogenisation at 10 000 PSI, the
samples were centrifuged for 20 min at 40 000 g, and the soluble
fraction was filtered.
Collapsin response mediator protein 2 purification
Purification was conducted automatically on an AKTA Xpress
system at a flow of 0.8 mL/min. Prior to purification, the HisTrap
HP and Superdex 200 columns were equilibrated with IMAC buffer
1 (50 mmol/L HEPES pH 7.5, 10 mmol/L imidazole, 500 mmol/L
NaCl, 10% glycerol, 0.5 mmol/L TCEP) and gel filtration buffer
(20 mmol/L HEPES pH 7.5, 300 mmol/L NaCl, 10% glycerol,
0.5 mmol/L TCEP), respectively. The protein sample was loaded on
the HisTrap HP column that was washed with IMAC buffer 1
followed by IMAC buffer 2 (50 mmol/L HEPES pH 7.5, 50 mmol/
L imidazole, 500 mmol/L NaCl, 10% glycerol, 0.5 mmol/L TCEP).
Bound protein was eluted from the IMAC columns with 7.5 mL of
IMAC elution buffer (50 mmol/L HEPES pH 7.5, 400 mmol/L
imidazole, 500 mmol/L NaCl, 10% glycerol, 0.5 mmol/L TCEP)
and loaded onto the gel filtration column. The chromatogram from
gel filtration showed one major protein peak that consisted of highly
pure CRMP-2. TCEP was added to the pooled protein peak to a final
concentration of 2 mmol/L. The protein was concentrated to 33 mg/
mL and stored at )80�C.
Table 1 Data collection and structure refinement. The values in
parentheses correspond to the high-resolution shell
Space group P21
Unit cell (a,b,c,b) 86.4 A, 126.1 A, 102.9 A, 113�Resolution (A) 30–2.4 (2.6–2.4)
Optical resolution (A) 1.76
Rsym (%) 14.6 (46.6)
<I/rI> 9.4 (3.3)
Completeness (%) 99.7 (99.7)
Redundancy 4.3 (4.3)
Rcryst (%) 17.0
Rfree (%) 24.5
r.m.s. deviation bond length (A) 0.015
r.m.s. deviation bond angle (�) 1.6
Average B factors (A2)
Protein (A,B,C,D) 23,23,23,22
Solvent 22
Calcium ions 35
Structure of human CRMP-2 907
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917
Crystallisation and data collection
Collapsin response mediator protein 2 was crystallised with the
hanging drop method at +20�C, by mixing 2 lL of the well solution
(0.2 mol/L CaCl2, 0.1 mol/L Tris pH 8.5, 14–18% PEG10000) and
1 lL of protein (17 mg/mL) and equilibrating against 0.5 mL of the
well solution.
(a) (b)
(d)(c)
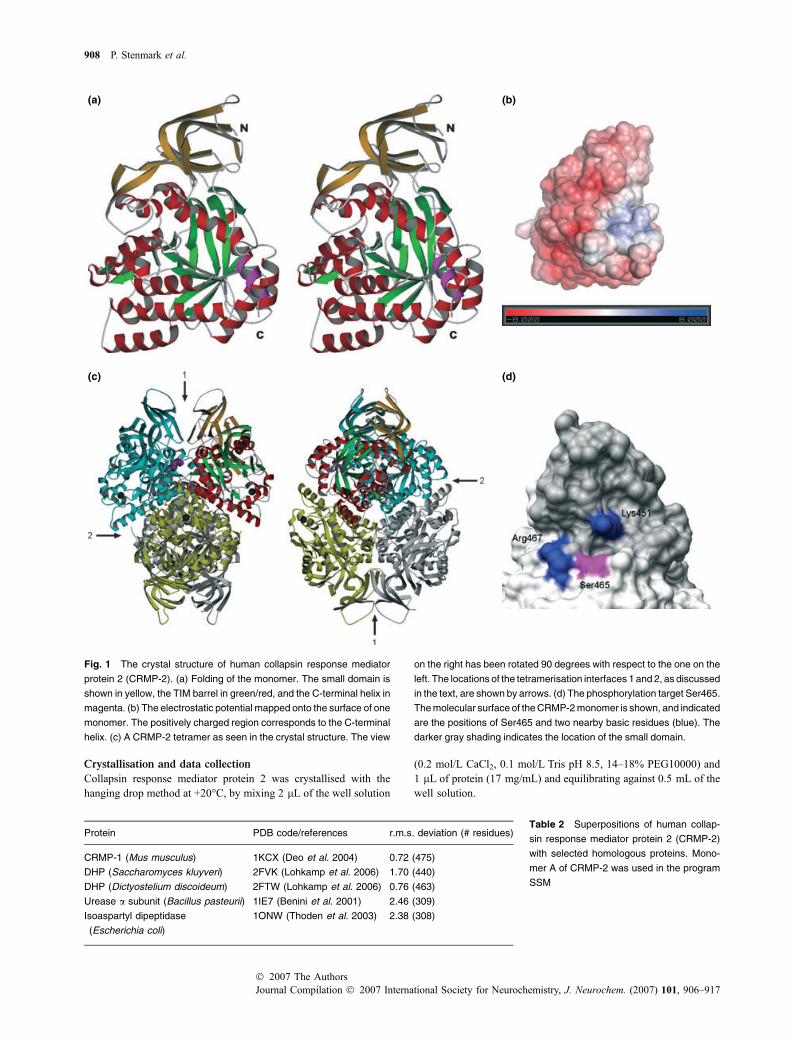
Fig. 1 The crystal structure of human collapsin response mediator
protein 2 (CRMP-2). (a) Folding of the monomer. The small domain is
shown in yellow, the TIM barrel in green/red, and the C-terminal helix in
magenta. (b) The electrostatic potential mapped onto the surface of one
monomer. The positively charged region corresponds to the C-terminal
helix. (c) A CRMP-2 tetramer as seen in the crystal structure. The view
on the right has been rotated 90 degrees with respect to the one on the
left. The locations of the tetramerisation interfaces 1 and 2, as discussed
in the text, are shown by arrows. (d) The phosphorylation target Ser465.
The molecular surface of the CRMP-2 monomer is shown, and indicated
are the positions of Ser465 and two nearby basic residues (blue). The
darker gray shading indicates the location of the small domain.
Table 2 Superpositions of human collap-
sin response mediator protein 2 (CRMP-2)
with selected homologous proteins. Mono-
mer A of CRMP-2 was used in the program
SSM
Protein PDB code/references r.m.s. deviation (# residues)
CRMP-1 (Mus musculus) 1KCX (Deo et al. 2004) 0.72 (475)
DHP (Saccharomyces kluyveri) 2FVK (Lohkamp et al. 2006) 1.70 (440)
DHP (Dictyostelium discoideum) 2FTW (Lohkamp et al. 2006) 0.76 (463)
Urease a subunit (Bacillus pasteurii) 1IE7 (Benini et al. 2001) 2.46 (309)
Isoaspartyl dipeptidase
(Escherichia coli)
1ONW (Thoden et al. 2003) 2.38 (308)
908 P. Stenmark et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917� 2007 The Authors
A crystal was briefly soaked in a cryoprotectant solution (15%
glycerol, 0.1 mol/L Tris pH 8.5, 18% PEG 10000, 0.2 mol/L CaCl2,
0.3 mol/L NaCl, 20 mmol/L HEPES pH 7.5, 2 mmol/L TCEP) prior
to data collection. X-ray diffraction data were collected at 100 K on
the ESRF beamline ID14.2 (Grenoble, France). The data were
processed using XDS (Kabsch 1993) and XDSi (Kursula 2004), and
5% of the data were randomly selected for the calculation of the free
R factor. The data processing statistics are shown in Table 1.
Structure solution and refinement
The structure was solved using molecular replacement in Molrep
(Vagin and Teplyakov 1997) with a monomer from the mouse
CRMP-1 structure (PDB entry 1KCX) (Deo et al. 2004) as
template, and four monomers of CRMP-2 were found within the
asymmetric unit, arranged into a tetrameric assembly. The structure
was refined with data between 30 and 2.4 A using Refmac
(Murshudov et al. 1997), employing loose non-crystallographic
symmetry restraints between the four monomers, and iterative model
building between refinement rounds was carried out in coot (Emsley
and Cowtan 2004). Water molecules were added into difference
electron density maps using ARP/wARP (Perrakis et al. 1999). Inthe final model, 87.7% of all residues were in the most favoured
region of the Ramachandran plot, and no residues were in the
disallowed regions. Proline residues 305 and 396 were modelled as
cis-prolines. The final refinement statistics are summarised in
Table 1. The coordinates and structure factors were deposited in the
Protein Data Bank under the accession code 2GSE.
The structure was analysed using the programs ProFunc
(Laskowski et al. 2005a), PDBsum (Laskowski et al. 2005b),
PQS (Henrick and Thornton 1998), ElNemo (Suhre and Sanejouand
2004) and SSM (Krissinel and Henrick 2004). Figures were made
using Dino, POV-ray, POVscript+ (Fenn et al. 2003), ESPript
(Gouet et al. 1999), APBS (Baker et al. 2001), PyMol and UCSF
Chimera (Pettersen et al. 2004).
Covalent cross-linking
The oligomeric status of purified CRMP-2 was analysed by covalent
cross-linking and sodium dodecyl sulfate–polyacrylamide gel
electrophoresis (SDS–PAGE). In brief, 1.3 mg/mL CRMP-2 was
incubated at 22�C for 15 min in 100 mmol/L HEPES (pH 7.3)
containing either 50, 150 or 500 mmol/L NaCl. Then, glutaralde-
hyde was added to a final concentration of 10, 25 or 100 mmol/L,
and incubation was continued at 22�C for 30 min. The reaction was
stopped by the addition of SDS–PAGE sample buffer, and the
samples were analysed by electrophoresis.
Results
The overall structure of human collapsin response
mediator protein 2
The crystal structure of human CRMP-2 was determined at2.4-A resolution. The C-terminus of the CRMPs has beenshown to be cleaved off by partial proteolysis (Deo et al.2004), and thus, this region was not included in the expressionconstruct. The largest fraction of the monomer structure isfolded into a triosephosphate isomerase (TIM) barrel; anotherwell-defined domain is the small b-sheet domain formed bythe N-terminal residues and segments close to the C terminusof the folded CRMP-2 (Fig. 1). As expected from sequencehomology, the fold of CRMP-2 is closely related to that of theenzymes related to DHPs (Table 2). They all have in common
440
mAU
(a)
(b)
(c)
1200
1000
800
600
400
200
050.0
C10 C12 D11D10D9 D8 D7 D6 D5 D4 D3 D2 D1 E1 E2 E3 E4 E5 E6 E7 E8
60.0 70.0 80.0 mL
198
62
49
38
2 3 4 5 6 7 8
1
190
12085605040
2 3 4 5 6 7 8 9
****
**
*
232158 67 43 25
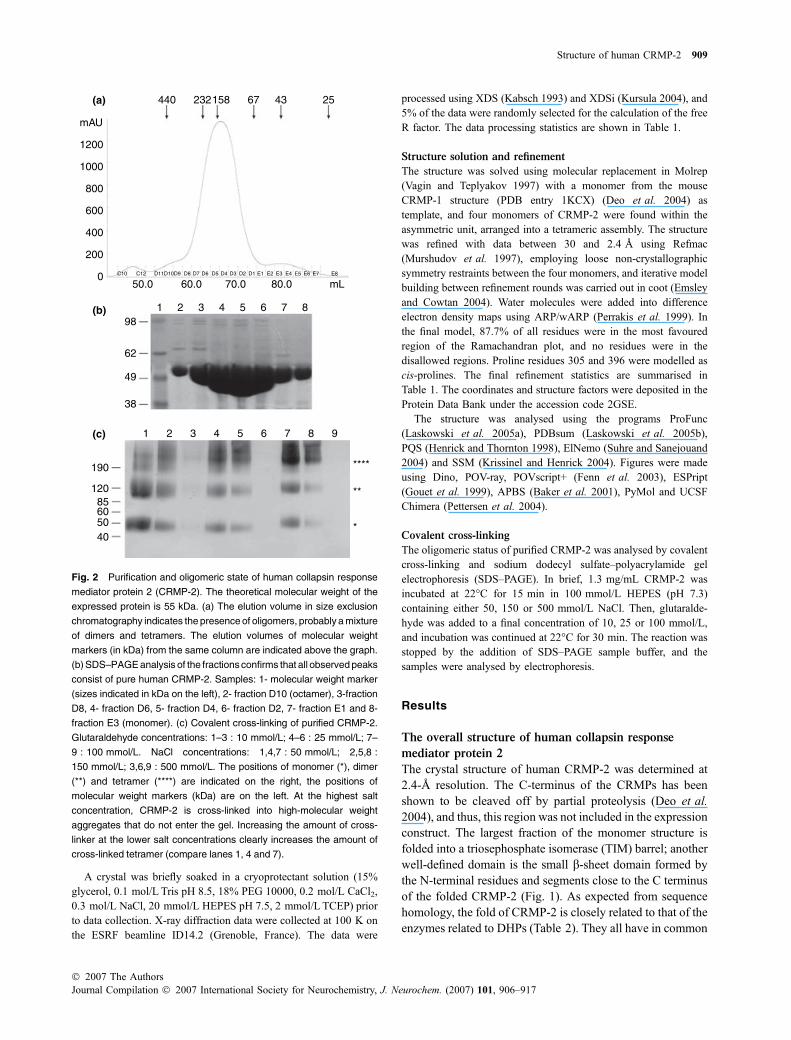
Fig. 2 Purification and oligomeric state of human collapsin response
mediator protein 2 (CRMP-2). The theoretical molecular weight of the
expressed protein is 55 kDa. (a) The elution volume in size exclusion
chromatography indicates the presence of oligomers, probably a mixture
of dimers and tetramers. The elution volumes of molecular weight
markers (in kDa) from the same column are indicated above the graph.
(b) SDS–PAGE analysis of the fractions confirms that all observed peaks
consist of pure human CRMP-2. Samples: 1- molecular weight marker
(sizes indicated in kDa on the left), 2- fraction D10 (octamer), 3-fraction
D8, 4- fraction D6, 5- fraction D4, 6- fraction D2, 7- fraction E1 and 8-
fraction E3 (monomer). (c) Covalent cross-linking of purified CRMP-2.
Glutaraldehyde concentrations: 1–3 : 10 mmol/L; 4–6 : 25 mmol/L; 7–
9 : 100 mmol/L. NaCl concentrations: 1,4,7 : 50 mmol/L; 2,5,8 :
150 mmol/L; 3,6,9 : 500 mmol/L. The positions of monomer (*), dimer
(**) and tetramer (****) are indicated on the right, the positions of
molecular weight markers (kDa) are on the left. At the highest salt
concentration, CRMP-2 is cross-linked into high-molecular weight
aggregates that do not enter the gel. Increasing the amount of cross-
linker at the lower salt concentrations clearly increases the amount of
cross-linked tetramer (compare lanes 1, 4 and 7).
Structure of human CRMP-2 909
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917
the TIM-barrel domain and the small domain. Normal modeanalysis (Suhre and Sanejouand 2004) of the structureindicates that it is likely that the small b-sheet domain ofCRMP-2 is flexible with respect to the rest of the monomer(not shown). As the active site in the related DHPs lies on theother side of the TIM barrel, there is little clue as to thefunction of the small domain. Most probably it is involved inprotein–protein interactions (see below).
The asymmetric unit consists of a homotetramer withnearly 222 symmetry (Fig. 1c). In the tetramer, the totalburied solvent-accessible surface area is over 9400 A2,heavily suggesting the presence of a physiologically
important tetramer. A superposition of the four monomerson each other gives root mean square (r.m.s.) deviations forthe Ca positions of 0.2–0.3 A between the differentmonomers, indicating strong similarity between the mono-mers in the asymmetric unit. In line with the homotetramericstructure, common to the structural family, the majority ofCRMP-2 eluted from gel filtration as a broad peak with amolecular weight of 150 kDa, corresponding to a trimer,with a minor fraction of monomer detectable (Fig. 2). Theresult is reproducibile between different batches of pureCRMP-2 protein as well as between protein constructsdiffering in length by a few amino acids (unpublished data).
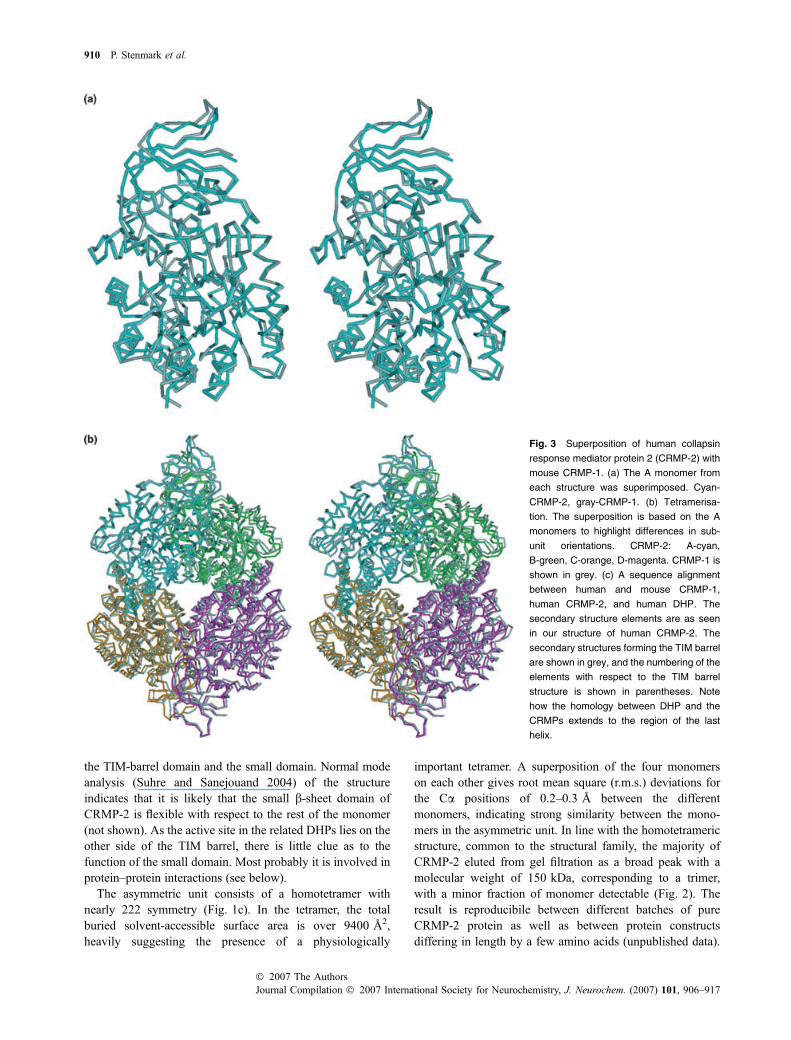
Fig. 3 Superposition of human collapsin
response mediator protein 2 (CRMP-2) with
mouse CRMP-1. (a) The A monomer from
each structure was superimposed. Cyan-
CRMP-2, gray-CRMP-1. (b) Tetramerisa-
tion. The superposition is based on the A
monomers to highlight differences in sub-
unit orientations. CRMP-2: A-cyan,
B-green, C-orange, D-magenta. CRMP-1 is
shown in grey. (c) A sequence alignment
between human and mouse CRMP-1,
human CRMP-2, and human DHP. The
secondary structure elements are as seen
in our structure of human CRMP-2. The
secondary structures forming the TIM barrel
are shown in grey, and the numbering of the
elements with respect to the TIM barrel
structure is shown in parentheses. Note
how the homology between DHP and the
CRMPs extends to the region of the last
helix.
910 P. Stenmark et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917� 2007 The Authors

All peaks from the gel filtration consisted of CRMP-2,which is a clear indication of the presence of various homo-oligomeric states (Fig. 2b). It is possible that the differencefrom the expected tetramer molecular weight reflects a rapiddynamic equilibrium between dimeric and tetrameric statesof CRMP-2, also supported by the width of the peakcovering both the dimeric and tetrameric molecular weights.One of the best-characterised such equilibria is that seen inhaemoglobin; in fact, gel filtration has been used to performdetailed studies on haemoglobin dimer-tetramer kinetics, theobserved peak corresponding roughly to the size of a trimer(Manning et al. 1996). Furthermore, a trimer cannot berationally explained by our structure, which clearly is a‘dimer of dimers’. It is possible that the equilibrium resultsfrom different oligomerisation kinetics between the full-length CRMP-2 and our truncated version, lacking theC-terminal unstructured tail. In addition, a minor peakcorresponding to a size of an octamer was observed. Incovalent cross-linking (Fig. 2c), the main species of CRMP-2 that were observed corresponded to monomers, dimers andtetramers.
Recently, Ser465 was identified as a CRMP-2 phos-phorylation site in mouse brain (Vosseller et al. 2005). This
is the only characterised phosphorylation site lying withinthe structure determined here, i.e. outside the C-terminaltail. Ser465 is easily accessible in the tetramer, lying in anexposed loop leading to the last helix of each monomer.The phosphorylation of serine 465 is unique to CRMP-2, asthis residue is not conserved in the other four CRMPproteins. CRMP-5 has a threonine in this position thatpossibly could be phosphorylated. In human DHP, thecorresponding residue is an aspartic acid. Nearby Ser465,two basic residues are found, one from the same loop andone from the small domain, which could interact with thephosphorylated residue (Fig. 1d). This could, for example,generate a stabilising salt bridge between the small domainand the rest of the CRMP-2 molecule. It is also possiblethat Ser465 phosphorylation could affect interactionsbetween the folded core of the protein and the positivelycharged C-terminal tail.
Comparison of CRMP-1 and -2 and
heterotetramerisation
The structure of murine CRMP-1 was solved recently (Deoet al. 2004), thus permitting a detailed structural compar-ison between these two closely related proteins. Despite
Fig. 3 Continued
Structure of human CRMP-2 911
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917

the high overall similarity of the structures (Fig. 3),differences may also be found that could have functionalimplications.
Collapsin response mediator protein 1 and CRMP-2 areable to form heterotetramers with each other (Wang andStrittmatter 1997), and we, thus, compared the monomer–monomer interfaces within the CRMP-1 and CRMP-2homotetramers to shed further light on these interactions(Fig. 4). This analysis was, to some extent, carried out basedon the structure of CRMP-1 and a sequence alignment forCRMP-2 (Deo et al. 2004), but having both experimentallydetermined structures at hand, we bring novel aspects to thiscomparison.
Interface 1 is formed between monomers A–B and C–D,generating a dimerwith a dyad axis.Most of the residues in thisinterface are conserved between CRMP-1 and CRMP-2,indicating that this interface probably can be used to makehetero-oligomers. The differences in this region betweenCRMP-1 and CRMP-2 are shown in Fig. 4a. The mostsignificant differences concern residues 266 and 316, Leu andTyr in CRMP-1 and Gln and Phe in CRMP-2, respectively.
Interface 2, between the two dimers related by interface 1, isalso required for tetramer formation. Interestingly, several ofthe non-conserved residues between CRMP-1 and CRMP-2are found at this interface (Fig. 4c). This is likely to adverselyaffect hetero-oligomerisation via interface 2. This interface isalsomuchmore hydrophobic inCRMP-1 than inCRMP-2. Forexample, in CRMP-1, Phe237 from monomers A and C faceeach other at a distance of 3.3 A; in CRMP-2, the correspond-ing residue is Asn237, being 3.5 A away from Asn237 on theopposing monomer. In addition, the above-mentioned Leu266in CRMP-1 contributes to both interfaces 1 and 2, but Gln266in CRMP-2 points towards interface 1. From the differences, itwould seem logical that a CRMP-1/CRMP-1 interaction at thisinterface should be stronger than a CRMP-1/CRMP-2 inter-action (or a CRMP-2/CRMP-2 interaction).
The pocket corresponding to the active site region
of DHP
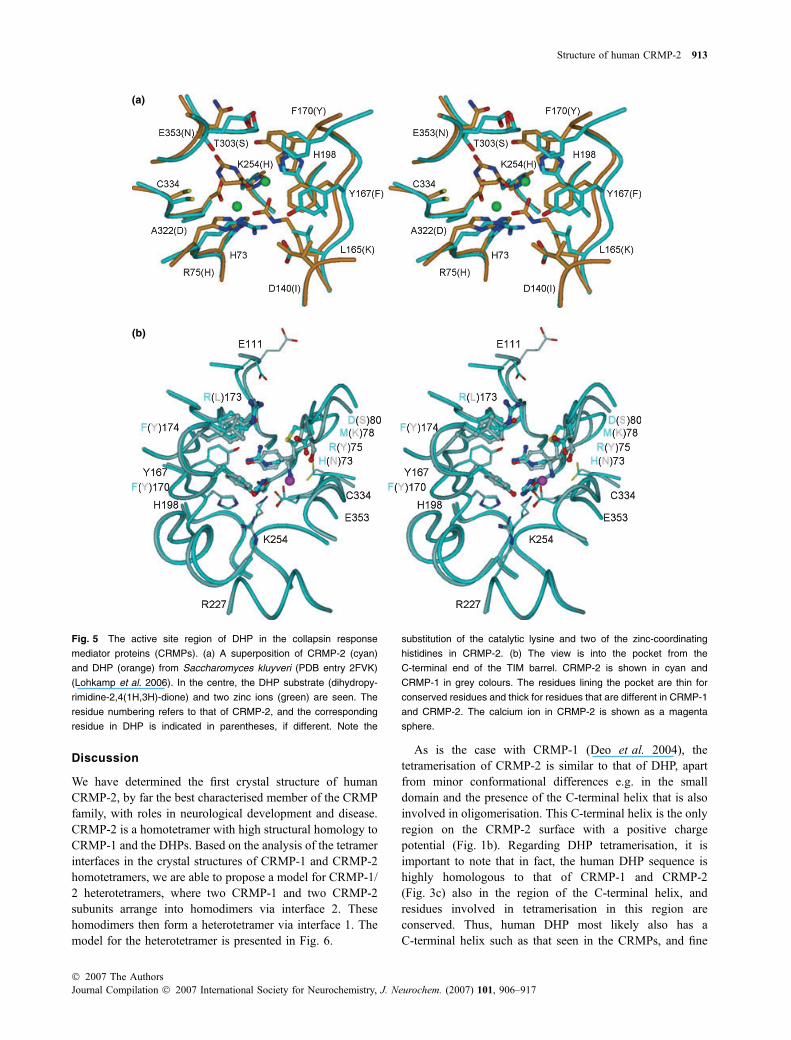
The active sites ofDHP and related enzymes liewithin the TIMbarrel, such that the cavity opens towards the C-terminal end ofthe barrel. In CRMP-1 and CRMP-2, a cavity also exists at thisposition. The volume of this cavity in CRMP-2 is 1400 A3 andthe depth 16 A. Catalytic residues of DHP have not beenconserved in CRMP-2; for example, the catalytic lysine andtwo of the Zn-coordinating histidine residues are not present(Fig. 5). However, it is possible that this site can be used tobind small molecules; an indication of the presence of a ligand-binding potential in the cavity is the presence of a well-definedCa2+ ion inCRMP-2 in the cavity. On the other hand, one of themost divergent regions between CRMP-1 and CRMP-2 isformed by thewalls and immediate surroundings of this cavity,which may either indicate no evolutionary pressure forconservation or the formation of different binding cavities inthe different CRMPs. For example, CRMP-1 has three tyrosineresidues in thewalls of this pocket, none ofwhich are present inCRMP-2 (Fig. 5b).
(a)
(b)
(c)
Fig. 4 The tetramerisation interfaces in human collapsin response
mediator protein 2 (CRMP-2). Differences in the oligomerisation inter-
faces between CRMP-1 and CRMP-2 are also pointed out. (a) A central
region of interface 1, with three sequence differences. In CRMP-2,
Gln266 forms part of a hydrophilic interface, while in CRMP-1, Leu266
points away from interface 1. (b) The interaction of the C-terminal helix
with the opposing monomer, also part of interface 1. The main difference
concerns Lys480 of CRMP-2 (glutamine in CRMP-1). Arg485 interacts
with backbone carbonyl groups from both monomers, having no neg-
ative counter-charge nearby. (c) Features of interface 2 and main dif-
ferences between CRMP-1 and CRMP-2. In the centre of the interface,
residue 237 is not conserved, and the relative properties of the sequence
differences in the vicinity result in a much more hydrophobic interface for
CRMP-1. At the edge of the hydrophobic patch, CRMP-1 has an inter-
subunit salt bridge, which is missing in CRMP-2.
912 P. Stenmark et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917� 2007 The Authors
Discussion
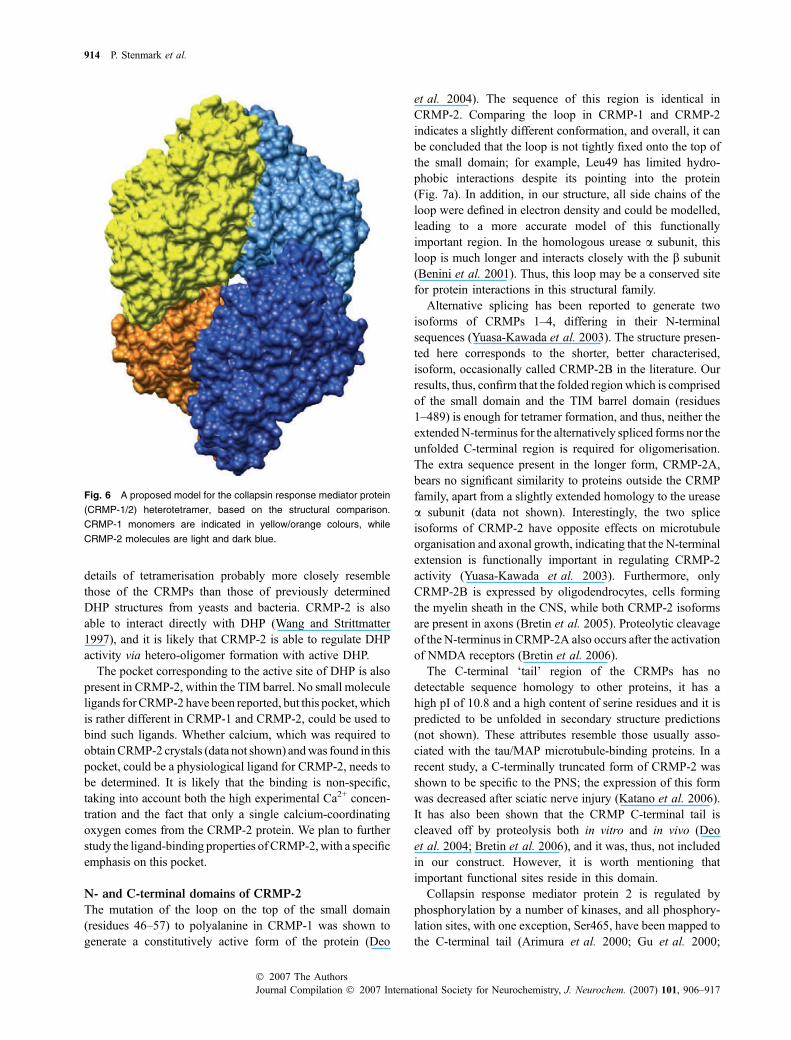
We have determined the first crystal structure of humanCRMP-2, by far the best characterised member of the CRMPfamily, with roles in neurological development and disease.CRMP-2 is a homotetramer with high structural homology toCRMP-1 and the DHPs. Based on the analysis of the tetramerinterfaces in the crystal structures of CRMP-1 and CRMP-2homotetramers, we are able to propose a model for CRMP-1/2 heterotetramers, where two CRMP-1 and two CRMP-2subunits arrange into homodimers via interface 2. Thesehomodimers then form a heterotetramer via interface 1. Themodel for the heterotetramer is presented in Fig. 6.
As is the case with CRMP-1 (Deo et al. 2004), thetetramerisation of CRMP-2 is similar to that of DHP, apartfrom minor conformational differences e.g. in the smalldomain and the presence of the C-terminal helix that is alsoinvolved in oligomerisation. This C-terminal helix is the onlyregion on the CRMP-2 surface with a positive chargepotential (Fig. 1b). Regarding DHP tetramerisation, it isimportant to note that in fact, the human DHP sequence ishighly homologous to that of CRMP-1 and CRMP-2(Fig. 3c) also in the region of the C-terminal helix, andresidues involved in tetramerisation in this region areconserved. Thus, human DHP most likely also has aC-terminal helix such as that seen in the CRMPs, and fine
(a)
(b)
Fig. 5 The active site region of DHP in the collapsin response
mediator proteins (CRMPs). (a) A superposition of CRMP-2 (cyan)
and DHP (orange) from Saccharomyces kluyveri (PDB entry 2FVK)
(Lohkamp et al. 2006). In the centre, the DHP substrate (dihydropy-
rimidine-2,4(1H,3H)-dione) and two zinc ions (green) are seen. The
residue numbering refers to that of CRMP-2, and the corresponding
residue in DHP is indicated in parentheses, if different. Note the
substitution of the catalytic lysine and two of the zinc-coordinating
histidines in CRMP-2. (b) The view is into the pocket from the
C-terminal end of the TIM barrel. CRMP-2 is shown in cyan and
CRMP-1 in grey colours. The residues lining the pocket are thin for
conserved residues and thick for residues that are different in CRMP-1
and CRMP-2. The calcium ion in CRMP-2 is shown as a magenta
sphere.
Structure of human CRMP-2 913
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917
details of tetramerisation probably more closely resemblethose of the CRMPs than those of previously determinedDHP structures from yeasts and bacteria. CRMP-2 is alsoable to interact directly with DHP (Wang and Strittmatter1997), and it is likely that CRMP-2 is able to regulate DHPactivity via hetero-oligomer formation with active DHP.
The pocket corresponding to the active site of DHP is alsopresent in CRMP-2, within the TIM barrel. No small moleculeligands forCRMP-2 have been reported, but this pocket, whichis rather different in CRMP-1 and CRMP-2, could be used tobind such ligands. Whether calcium, which was required toobtainCRMP-2 crystals (data not shown) andwas found in thispocket, could be a physiological ligand for CRMP-2, needs tobe determined. It is likely that the binding is non-specific,taking into account both the high experimental Ca2+ concen-tration and the fact that only a single calcium-coordinatingoxygen comes from the CRMP-2 protein. We plan to furtherstudy the ligand-binding properties of CRMP-2,with a specificemphasis on this pocket.
N- and C-terminal domains of CRMP-2
The mutation of the loop on the top of the small domain(residues 46–57) to polyalanine in CRMP-1 was shown togenerate a constitutively active form of the protein (Deo
et al. 2004). The sequence of this region is identical inCRMP-2. Comparing the loop in CRMP-1 and CRMP-2indicates a slightly different conformation, and overall, it canbe concluded that the loop is not tightly fixed onto the top ofthe small domain; for example, Leu49 has limited hydro-phobic interactions despite its pointing into the protein(Fig. 7a). In addition, in our structure, all side chains of theloop were defined in electron density and could be modelled,leading to a more accurate model of this functionallyimportant region. In the homologous urease a subunit, thisloop is much longer and interacts closely with the b subunit(Benini et al. 2001). Thus, this loop may be a conserved sitefor protein interactions in this structural family.
Alternative splicing has been reported to generate twoisoforms of CRMPs 1–4, differing in their N-terminalsequences (Yuasa-Kawada et al. 2003). The structure presen-ted here corresponds to the shorter, better characterised,isoform, occasionally called CRMP-2B in the literature. Ourresults, thus, confirm that the folded regionwhich is comprisedof the small domain and the TIM barrel domain (residues1–489) is enough for tetramer formation, and thus, neither theextendedN-terminus for the alternatively spliced forms nor theunfolded C-terminal region is required for oligomerisation.The extra sequence present in the longer form, CRMP-2A,bears no significant similarity to proteins outside the CRMPfamily, apart from a slightly extended homology to the ureasea subunit (data not shown). Interestingly, the two spliceisoforms of CRMP-2 have opposite effects on microtubuleorganisation and axonal growth, indicating that the N-terminalextension is functionally important in regulating CRMP-2activity (Yuasa-Kawada et al. 2003). Furthermore, onlyCRMP-2B is expressed by oligodendrocytes, cells formingthe myelin sheath in the CNS, while both CRMP-2 isoformsare present in axons (Bretin et al. 2005). Proteolytic cleavageof the N-terminus in CRMP-2A also occurs after the activationof NMDA receptors (Bretin et al. 2006).
The C-terminal ‘tail’ region of the CRMPs has nodetectable sequence homology to other proteins, it has ahigh pI of 10.8 and a high content of serine residues and it ispredicted to be unfolded in secondary structure predictions(not shown). These attributes resemble those usually asso-ciated with the tau/MAP microtubule-binding proteins. In arecent study, a C-terminally truncated form of CRMP-2 wasshown to be specific to the PNS; the expression of this formwas decreased after sciatic nerve injury (Katano et al. 2006).It has also been shown that the CRMP C-terminal tail iscleaved off by proteolysis both in vitro and in vivo (Deoet al. 2004; Bretin et al. 2006), and it was, thus, not includedin our construct. However, it is worth mentioning thatimportant functional sites reside in this domain.
Collapsin response mediator protein 2 is regulated byphosphorylation by a number of kinases, and all phosphory-lation sites, with one exception, Ser465, have been mapped tothe C-terminal tail (Arimura et al. 2000; Gu et al. 2000;
Fig. 6 A proposed model for the collapsin response mediator protein
(CRMP-1/2) heterotetramer, based on the structural comparison.
CRMP-1 monomers are indicated in yellow/orange colours, while
CRMP-2 molecules are light and dark blue.
914 P. Stenmark et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917� 2007 The Authors

Brown et al. 2004; Cole et al. 2004; Yoshimura et al. 2005).Phosphorylation inhibits the binding of CRMP-2 to tubulinand prevents its effect on tubulin polymerisation (Arimuraet al. 2005). It is likely that the C-terminal tail plays a majorrole in interactions between CRMP-2 and tubulin. Further-more, the interaction between CRMP-2 and the tetratricopep-tide repeat (TPR) domain of kinesin seems to be mediated byresidues 440–572, i.e. the C-terminal tail (Kimura et al. 2005).
Collapsin response mediator protein 2 is also O-glycosyl-ated (Cole and Hart 2001; Kanninen et al. 2004), andprediction of glycosylation sites using the NetOGlyc server(Hansen et al. 1998) strongly suggests that the glycosylationsite(s) also are in the C-terminal tail (not shown). The effectsof glycosylation on CRMP-2 function have not been defined,but it is possible that glycosylation regulates the interactionsof CRMP-2 with its ligands and/or its phosphorylation.
Interestingly, a recent screen for brain calmodulin-bindingproteins identified CRMP-2 as a putative CaM target (Zhang
et al. 2006). Searching for putative CaM-binding sequences(Yap et al. 2000) of CRMP-2, the most likely region wouldbe the last helix of the current structure, residues 475–489.The helix is involved in the tetramerisation interface, notbeing completely buried, however. Another putative CaM-binding region could be the C-terminal tail. While furtherstudies on CRMP-2–CaM interactions are clearly required, itcan be speculated that CaM binding could affect CRMP-2tetramerisation, its interactions with other proteins, and/or itspost-translational modifications.
Previously mapped functional domains and their relation
to the three-dimensional structure of CRMP-2
A number of studies have been carried out to find interactionpartners for CRMP-2. These studies have, to a large extent,used truncation mutants in cell culture experiments, whichhave also allowed the mapping of binding domains (Fig. 7b).Apart from a short region around residue 320, none of the
Fig. 7 Previously mapped interaction domains of collapsin response
mediator protein 2 (CRMP-2) in the structure. (a) The loop (residues
46–57) in the small N-terminal domain is well defined in the electron
density. Cyan-CRMP-2, grey-CRMP-1. The electron density is that of
the final 2Fo–Fc map, contoured a 1 r. For CRMP-1, most side chains
are not present in the model, and for example Leu49 was modelled to
point outwards from the loop. In CRMP-2, Leu49 is well-defined,
together with the rest of the loop. (b) Overall view with coloured
mapped domains for protein interactions, such as determined mainly
by using truncation mutants in previous studies. Note how the previ-
ously suggested minimal binding sites for both Numb (275–322, blue)
and tubulin (323–381, red) are buried within the CRMP-2 fold.
The loop on top of the small domain (residues 46–57) is shown in
orange.
Structure of human CRMP-2 915
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917

studied segments are involved in tetramerisation interfaces.The interaction data can now be discussed in light of thecurrent structural information.
Collapsin response mediator protein 2 binds tubulin andregulates microtubule assembly (Gu and Ihara 2000; Fukataet al. 2002). The functional domain in this case was mappedto the region 323–381, based on a series of truncationmutants. This region was the most effective in promotingmicrotubule assembly in vitro; it was not enough for in vivofunction, however (Fukata et al. 2002). Judging from ourstructure (Fig. 7b), this region is mostly buried within thefold of CRMP-2 (indeed forming an a/b unit of the TIMbarrel), and it is likely that the effects observed with eitheronly this domain present or with CRMP-2 lacking thisregion, are affected by folding defects.
Collapsin response mediator protein 2 also interacts withNumb, a protein involved in endocytosis (Nishimura et al.2003). It was suggested that the region 275–322 is required forthe binding, based on the behaviour of various truncationmutants. On the basis of the crystal structure, the inactive 1–275 mutant used in the interaction study is most likelymisfolded, as it contains a truncated TIM domain, and therecorded lack of activity should therefore be interpreted withcaution. The active 1–381mutant, however, contains the entireTIM barrel and most of the small domain. A direct interactionhas also been characterised between CRMP-2 and phosphol-ipase D2, requiring residues 243–300 of CRMP-2 (Lee et al.2002). This region overlaps with that suggested to bind Numb,and forms a central part of the folded TIM barrel domain.
In summary, while the actual interactions of full-lengthCRMP-2 are not brought into question by the crystalstructure of human CRMP-2 presented in this article, thedetailed mapping data from truncation experiments are verydifficult to rationalise while analysing the three-dimensionalstructure. This is, of course, a general problem whilemapping binding sites by using linear truncation mutants,when the three-dimensional structure of the studied protein isnot accurately known. Based on our work, structure-basedexperiments can now be carried out on CRMP-2 and itsinteraction properties.
Acknowledgements
The Structural Genomics Consortium is a registered charity (number
1097737) funded by VINNOVA, the Knut and Alice Wallenberg
Foundation, the Swedish Foundation for Strategic Research, the
Karolinska Institute, the Wellcome Trust, GlaxoSmithKline,
Genome Canada, the Canadian Institutes of Health Research, the
Ontario Innovation Trust, the Ontario Research and Development
Challenge Fund, and the Canadian Foundation for Innovation. The
work was also supported by the Swedish Cancer Society and the
Swedish Research Council (PN). PK is an Academy Research
Fellow (Academy of Finland).
References
Arimura N., Inagaki N., Chihara K., Menager C., Nakamura N.,Amano M., Iwamatsu A., Goshima Y. and Kaibuchi K. (2000)Phosphorylation of collapsin response mediator protein-2 by Rho-kinase. Evidence for two separate signaling pathways for growthcone collapse. J. Biol. Chem. 275, 23 973–23 980.
Arimura N., Menager C., Fukata Y. and Kaibuchi K. (2004) Role ofCRMP-2 in neuronal polarity. J. Neurobiol. 58, 34–47.
Arimura N., Menager C., Kawano Y. et al. (2005) Phosphorylation byRho kinase regulates CRMP-2 activity in growth cones. Mol. Cell.Biol. 25, 9973–9984.
Baker N. A., Sept D., Joseph S., Holst M. J. and McCammon J. A.(2001) Electrostatics of nanosystems: application to microtubulesand the ribosome. Proc. Natl Acad. Sci. USA 98, 10 037–10 041.
Benini S., Rypniewski W. R., Wilson K. S., Ciurli S. and Mangani S.(2001) Structure-based rationalization of urease inhibition byphosphate: novel insights into the enzyme mechanism. J. Biol.Inorg. Chem. 6, 778–790.
Bretin S., Reibel S., Charrier E. et al. (2005) Differential expressionof CRMP1, CRMP2A, CRMP2B, and CRMP5 in axons ordendrites of distinct neurons in the mouse brain. J. Comp.Neurol. 486, 1–17.
Bretin S., Rogemond V., Marin P., Maus M., Torrens Y., Honnorat J.,Glowinski J., Premont J. and Gauchy C. (2006) Calpain product ofWT-CRMP2 reduces the amount of surface NR2B NMDA receptorsubunit. J. Neurochem. 98, 1252–1265.
Brown M., Jacobs T., Eickholt B., Ferrari G., Teo M., Monfries C., QiR. Z., Leung T., Lim L. and Hall C. (2004) Alpha2-chimaerin,cyclin-dependent Kinase 5/p35, and its target collapsin responsemediator protein-2 are essential components in semaphorin3A-induced growth-cone collapse. J. Neurosci. 24, 8994–9004.
Cole R. N. and Hart G. W. (2001) Cytosolic O-glycosylation is abundantin nerve terminals. J. Neurochem. 79, 1080–1089.
Cole A. R., Knebel A., Morrice N. A., Robertson L. A., Irving A. J.,Connolly C. N. and Sutherland C. (2004) GSK-3 phosphorylationof the Alzheimer epitope within collapsin response mediator pro-teins regulates axon elongation in primary neurons. J. Biol. Chem.279, 50 176–50 180.
Cole A. R., Causeret F., Yadirgi G., Hastie C. J., McLauchlan H.,McManus E. J., Hernandez F., Eickholt B. J., Nikolic M. andSutherland C. (2006) Distinct priming kinases contribute to dif-ferential regulation of collapsin response mediator proteins byglycogen synthase kinase-3 in vivo. J. Biol. Chem. 281, 16 591–16 598.
Czech T., Yang J. W., Csaszar E., Kappler J., Baumgartner C. and LubecG. (2004) Reduction of hippocampal collapsin response mediatedprotein-2 in patients with mesial temporal lobe epilepsy. Neuro-chem. Res. 29, 2189–2196.
Deo R. C., Schmidt E. F., Elhabazi A., Togashi H., Burley S. K. andStrittmatter S. M. (2004) Structural bases for CRMP functionin plexin-dependent semaphorin3A signaling. EMBO J. 23, 9–22.
Emsley P. and CowtanK. (2004) Coot: model-building tools for moleculargraphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132.
Fenn T. D., Ringe D. and Petsko G. A. (2003) POVScript+: a programfor model and data visualization using persistence of vision ray-tracing. J. Appl. Cryst. 36, 944–947.
Fukata Y., Itoh T. J., Kimura T. et al. (2002) CRMP-2 binds to tubulinheterodimers to promote microtubule assembly. Nat. Cell Biol. 4,583–591.
Gouet P., Courcelle E., Stuart D. I. and Metoz F. (1999) ESPript: analysisof multiple sequence alignments in PostScript. Bioinformatics 15,305–308.
916 P. Stenmark et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917� 2007 The Authors

Gu Y. and Ihara Y. (2000) Evidence that collapsin response mediatorprotein-2 is involved in the dynamics of microtubules. J. Biol.Chem. 275, 17 917–17 920.
Gu Y., Hamajima N. and Ihara Y. (2000) Neurofibrillary tangle-associ-ated collapsin response mediator protein-2 (CRMP-2) is highlyphosphorylated on Thr-509, Ser-518, and Ser-522. Biochemistry39, 4267–4275.
Hamajima N., Matsuda K., Sakata S., Tamaki N., Sasaki M. and NonakaM. (1996) A novel gene family defined by human dihydropyri-midinase and three related proteins with differential tissue distri-bution. Gene 180, 157–163.
Hansen J. E., Lund O., Tolstrup N., Gooley A. A., Williams K. L. andBrunak S. (1998) NetOglyc: prediction of mucin type O-glycosy-lation sites based on sequence context and surface accessibility.Glycoconj. J. 15, 115–130.
Henrick K. and Thornton J. M. (1998) PQS: a protein quaternarystructure file server. Trends Biochem. Sci. 23, 358–361.
Inagaki H., Kato Y., Hamajima N., Nonaka M., Sasaki M. and Eimoto T.(2000) Differential expression of dihydropyrimidinase-relatedprotein genes in developing and adult enteric nervous system.Histochem. Cell Biol. 113, 37–41.
Kabsch W. (1993) Automatic processing of rotation diffraction data fromcrystals of initially unknown symmetry and cell constants. J. Appl.Cryst. 26, 795–800.
Kamata T., Subleski M., Hara Y., Yuhki N., Kung H., Copeland N. G.,Jenkins N. A., Yoshimura T., Modi W. and Copeland T. D. (1998)Isolation and characterization of a bovine neural specific protein(CRMP-2) cDNA homologous to unc-33, a C. elegans geneimplicated in axonal outgrowth and guidance. Brain Res. Mol.Brain Res. 54, 219–236.
Kanninen K., Goldsteins G., Auriola S., Alafuzoff I. and Koistinaho J.(2004) Glycosylation changes in Alzheimer’s disease as revealedby a proteomic approach. Neurosci. Lett. 367, 235–240.
Katano T., Mabuchi T., Okuda-Ashitaka E., Inagaki N., Kinumi T. andIto S. (2006) Proteomic identification of a novel isoform of col-lapsin response mediator protein-2 in spinal nerves peripheral todorsal root ganglia. Proteomics. 6, 6085–6094.
Kimura T., Watanabe H., Iwamatsu A. and Kaibuchi K. (2005) Tubulinand CRMP-2 complex is transported via Kinesin-1. J. Neurochem.93, 1371–1382.
Krissinel E. and Henrick K. (2004) Secondary-structure matching(SSM), a new tool for fast protein structure alignment in threedimensions. Acta Crystallogr. D Biol. Crystallogr. 60, 2256–2268.
Kursula P. (2004) XDSi – a graphical interface for the data processingprogram XDS. J. Appl. Cryst. 37, 347–348.
Laskowski R. A., Watson J. D. and Thornton J. M. (2005a) ProFunc: aserver for predicting protein function from 3D structure. NucleicAcids Res. 33, W89–W93.
Laskowski R. A., Chistyakov V. V. and Thornton J. M. (2005b) PDBsummore: new summaries and analyses of the known 3D structures ofproteins and nucleic acids. Nucleic Acids Res. 33, D266–D268.
Lee S., Kim J. H., Lee C. S., Kim J. H., Kim Y., Heo K., Ihara Y.,Goshima Y., Suh P. G. and Ryu S. H. (2002) Collapsin responsemediator protein-2 inhibits neuronal phospholipase D(2) activityby direct interaction. J. Biol. Chem. 277, 6542–6549.
Lohkamp B., Andersen B., Piskur J. and Dobritzsch D. (2006) Thecrystal structures of dihydropyrimidinases reaffirm the close rela-tionship between cyclic amidohydrolases and explain their sub-strate specificity. J. Biol. Chem. 281, 13 762–13 776.
Manning L. R., Jenkins W. T., Hess J. R., Vandegriff K., Winslow R. M.and Manning J. M. (1996) Subunit dissociations in natural andrecombinant hemoglobins. Protein Sci. 5, 775–781.
Mimura F., Yamagishi S., Arimura N., Fujitani M., Kubo T., Kaibuchi K.and Yamashita T. (2006) Myelin-associated glycoprotein inhibits
microtubule assembly by a Rho-kinase-dependent mechanism.J. Biol. Chem. 281, 15 970–15 979.
Murshudov G. N., Vagin A. A. and Dodson E. J. (1997) Refinement ofmacromolecular structures by the maximum-likelihood method.Acta Crystallogr. D Biol. Crystallogr. 53, 240–255.
Nishimura T., Fukata Y., Kato K., Yamaguchi T., Matsuura Y.,Kamiguchi H. and Kaibuchi K. (2003) CRMP-2 regulates polar-ized Numb-mediated endocytosis for axon growth. Nat. Cell Biol.5, 819–826.
Perrakis A., Morris R. and Lamzin V. S. (1999) Automated proteinmodel building combined with iterative structure refinement. Nat.Struct. Biol. 6, 458–463.
Pettersen E. F., Goddard T. D., Huang C. C., Couch G. S., GreenblattD. M., Meng E. C. and Ferrin T. E. (2004) UCSF Chimera–avisualization system for exploratory research and analysis.J. Comput. Chem. 25, 1605–1612.
Ricard D., Stankoff B., Bagnard D. et al. (2000) Differential expressionof collapsin response mediator proteins (CRMP/ULIP) in subsetsof oligodendrocytes in the postnatal rodent brain. Mol. Cell.Neurosci. 16, 324–337.
Suhre K. and Sanejouand Y. H. (2004) ElNemo: a normal mode webserver for protein movement analysis and the generation of tem-plates for molecular replacement. Nucleic Acids Res. 32, W610–W614.
Sultana R., Boyd-Kimball D., Poon H. F., Cai J., Pierce W. M., KleinJ. B., Merchant M., Markesbery W. R. and Butterfield D. A. (2006)Redox proteomics identification of oxidized proteins in Alzhei-mer’s disease hippocampus and cerebellum: An approach tounderstand pathological and biochemical alterations in AD.Neurobiol. Aging 27, 1564–1576.
Tahimic C. G., Tomimatsu N., Nishigaki R., Fukuhara A., Toda T.,Kaibuchi K., Shiota G., Oshimura M. and Kurimasa A. (2006)Evidence for a role of Collapsin response mediator protein-2 insignaling pathways that regulate the proliferation of non-neuronalcells. Biochem. Biophys. Res. Commun. 340, 1244–1250.
Thoden J. B., Marti-Arbona R., Raushel F. M. and Holden H. M. (2003)High-resolution X-ray structure of isoaspartyl dipeptidase fromEscherichia coli. Biochemistry 42, 4874–4882.
Vagin A. and Teplyakov A. (1997) MOLREP: an automated program formolecular replacement. J. Appl. Cryst. 30, 1022–1025.
Vosseller K., Hansen K. C., Chalkley R. J., Trinidad J. C., Wells L.,Hart G. W. and Burlingame A. L. (2005) Quantitative analysis ofboth protein expression and serine / threonine post-translationalmodifications through stable isotope labeling with dithiothreitol.Proteomics 5, 388–398.
Wang L. H. and Strittmatter S. M. (1996) A family of rat CRMP genes isdifferentially expressed in the nervous system. J. Neurosci. 16,6197–6207.
Wang L. H. and Strittmatter S. M. (1997) Brain CRMP forms hetero-tetramers similar to liver dihydropyrimidinase. J. Neurochem. 69,2261–2269.
Yap K. L., Kim J., Truong K., Sherman M., Yuan T. and Ikura M. (2000)Calmodulin target database. J. Struct. Funct. Genomics 1, 8–14.
Yoshimura T., Kawano Y., Arimura N., Kawabata S., Kikuchi A. andKaibuchi K. (2005) GSK-3beta regulates phosphorylation ofCRMP-2 and neuronal polarity. Cell 120, 137–149.
Yuasa-Kawada J., Suzuki R., Kano F., Ohkawara T., Murata M. andNoda M. (2003) Axonal morphogenesis controlled by antagonisticroles of two CRMP subtypes in microtubule organization. Eur. J.Neurosci. 17, 2329–2343.
Zhang Z., Ottens A. K., Golden E. C., Hayes R. L. and Wang K. K. W.(2006) Using calmodulin-affinity capture to study the rat braincalmodulin binding proteome and its vulnerability to calpain andcaspase proteolysis. Calcium Binding Proteins 1, 125–134.
Structure of human CRMP-2 917
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 906–917
Copyright © 2022 FDOKUMEN




















