
The small mammals of Mt. Banahaw-San Cristobal National Park, Luzon, Philippines: elevational...
16
ABSTRACT An intensive survey of the small non-volant mammals along the full elevational gradient of Mt. Banahaw - San Cristobal National Park in 1995-1996, 2004-2005, and 2012, using standardized procedures, produced records of ten native and three exotic small mammals. Four species discovered during our survey appear to be endemic to the park (Apomys banahao, A. magnus, Musseromys gulantang, and Rhynchomys banahao), a remarkable number for such a small area; the other native species are more widespread. Species richness in forest on Mt. Banahaw (peak = 2169 m) increased from five at 620 m to seven at 1465 m, and declined to three at 2030 m. Mixed agricultural areas and exotic grassland with shrubby second growth supported up to three exotic species and up to three native species. Exotic species were apparently absent from secondary or primary forest. Relative abundance of native species was significantly correlated with increasing elevation, with abundance about five times greater in montane and mossy forest than in the lowlands. Trap success using live earth- worms as bait increased significantly with increasing ele- vation, and small mammals showed an overall preference for earthworm bait, but a few were arboreal and a few pre- ferred coconut bait. Most species were significantly more active on the ground than in trees, and most were signifi- cantly more active during the night than during daylight. Mt. Banahaw - San Cristobal National Park is a premier example of a highly localized center of mammalian en- demism; along with its important role as a religious and recreational center and crucial function as a watershed, it clearly deserves continued and active protection. The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines: Elevational Distribution and Ecology of a Highly Endemic Fauna Lawrence R. Heaney 1 , Danilo S. Balete 1 , R. Grace B. Rosell Ambal 2 , Maria Josefa S. Veluz 3 , and Eric A. Rickart 4 1 Field Museum of Natural History, 1400 S Lake Shore Drive, Chicago, IL 60605 USA; 2 3G Sunvar Condominium, 2135 A. Luna St., Pasay City, Philippines; 3 National Museum of the Philippines, Rizal Park, Manila, Philippines; 4 Natural History Museum of Utah, 301 Wakara Way, Salt Lake City, UT 84108 USA INTRODUCTION Rising from the fertile, heavily utilized agricultural plains south of Manila and Laguna de Bay emerge a number of peaks, all of relatively recent volcanic origin, and usual- ly separated from the others by extensive lowlands. e tallest of these is Mt. Banahaw, reaching to 2169 m. Mt. Banahaw has long been known as a place of great natu- ral beauty, and has served as a place of recreation and re- ligious observance for many generations. Together with Mt. San Cristobal, which lies a short distance to the west (Fig. 1), Mt. Banahaw was declared a national park in 1941 and redesignated in 1966 (Mallari et al., 2001), and it has been listed as a priority site for conservation (Ambal et al., 2012; Mallari et al., 2001; Ong et al., 2002; see also Heaney et al., 2002). A few early naturalists visited the mountain to study its wildlife, principally birds (e.g., Zimmer, 1918; see also Dickerson, 1928). Despite its proximity to Manila, there was no effort to systematically document the mam- mal fauna of Mt. Banahaw until the 1970s, when research for a Master’s thesis was conducted by Miranda (1978) but remained unpublished. Miranda documented the presence of three native non-volant mammals, Apomys sp., Phloeomys cumingi, and Rattus everetti, plus four commensal, non-na- tive species: Suncus murinus, Mus musculus, Rattus exu- lans and R. rattus (= R. tanezumi). Since P. cumingi and R. everetti are widespread, only the mention of Apomys sp., generally known as forest mice, gave a hint of the po- tential for locally endemic mammals, especially since the genus was quite poorly known at the time (Musser, 1982; Heaney et al., 1998, 2011; Steppan et al., 2003). Mallari and Diesmos (1995) reported in an unpublished
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The small mammals of Mt. Banahaw-San Cristobal National Park, Luzon, Philippines: elevational...

The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines
ABStrActAn intensive survey of the small non-volant mammals along the full elevational gradient of Mt. Banahaw - San Cristobal National Park in 1995-1996, 2004-2005, and 2012, using standardized procedures, produced records of ten native and three exotic small mammals. Four species discovered during our survey appear to be endemic to the park (Apomys banahao, A. magnus, Musseromys gulantang, and Rhynchomys banahao), a remarkable number for such a small area; the other native species are more widespread. Species richness in forest on Mt. Banahaw (peak = 2169 m) increased from five at 620 m to seven at 1465 m, and declined to three at 2030 m. Mixed agricultural areas and exotic grassland with shrubby second growth supported up to three exotic species and up to three native species. Exotic species were apparently absent from secondary or primary forest. Relative abundance of native species was significantly correlated with increasing elevation, with abundance about five times greater in montane and mossy forest than in the lowlands. Trap success using live earth-worms as bait increased significantly with increasing ele-vation, and small mammals showed an overall preference for earthworm bait, but a few were arboreal and a few pre-ferred coconut bait. Most species were significantly more active on the ground than in trees, and most were signifi-cantly more active during the night than during daylight. Mt. Banahaw - San Cristobal National Park is a premier example of a highly localized center of mammalian en-demism; along with its important role as a religious and recreational center and crucial function as a watershed, it clearly deserves continued and active protection.
the Small Mammals of Mt. Banahaw - San cristobal national Park, Luzon, Philippines: elevational
distribution and ecology of a highly endemic Fauna
Lawrence R. Heaney1, Danilo S. Balete1,R. Grace B. Rosell Ambal2, Maria Josefa S. Veluz3,
and Eric A. Rickart4
1 Field Museum of Natural History, 1400 S Lake Shore Drive, Chicago, IL 60605 USA; 2 3G Sunvar Condominium, 2135 A. Luna St., Pasay City, Philippines; 3 National Museum of the Philippines, Rizal Park, Manila, Philippines; 4 Natural History Museum of Utah, 301 Wakara Way, Salt Lake City, UT 84108 USA
IntroductIon Rising from the fertile, heavily utilized agricultural plains south of Manila and Laguna de Bay emerge a number of peaks, all of relatively recent volcanic origin, and usual-ly separated from the others by extensive lowlands. The tallest of these is Mt. Banahaw, reaching to 2169 m. Mt. Banahaw has long been known as a place of great natu-ral beauty, and has served as a place of recreation and re-ligious observance for many generations. Together with Mt. San Cristobal, which lies a short distance to the west (Fig. 1), Mt. Banahaw was declared a national park in 1941 and redesignated in 1966 (Mallari et al., 2001), and it has been listed as a priority site for conservation (Ambal et al., 2012; Mallari et al., 2001; Ong et al., 2002; see also Heaney et al., 2002). A few early naturalists visited the mountain to study its wildlife, principally birds (e.g., Zimmer, 1918; see also Dickerson, 1928). Despite its proximity to Manila, there was no effort to systematically document the mam-mal fauna of Mt. Banahaw until the 1970s, when research for a Master’s thesis was conducted by Miranda (1978) but remained unpublished. Miranda documented the presence of three native non-volant mammals, Apomys sp., Phloeomys cumingi, and Rattus everetti, plus four commensal, non-na-tive species: Suncus murinus, Mus musculus, Rattus exu-lans and R. rattus (= R. tanezumi). Since P. cumingi and R. everetti are widespread, only the mention of Apomys sp., generally known as forest mice, gave a hint of the po-tential for locally endemic mammals, especially since the genus was quite poorly known at the time (Musser, 1982; Heaney et al., 1998, 2011; Steppan et al., 2003).
Mallari and Diesmos (1995) reported in an unpublished
45

National Museum of the Philippines: Journal of Natural History
manuscript the presence of Crocidura grayi, one small Apomys (A. microdon), three species of large Apomys (ten-tatively identified as A. abrae, A. datae, and A. sacobianus), Bullimus luzonicus, Phloeomys cumingi, and Rattus ever-etti, plus several non-native species; the voucher specimens were subsequently lost, and so the identifications cannot be verified. As described below, our studies of the mam-malian fauna of the mountain began when Rosell (1996) conducted an extensive survey of small mammals on Mts. Banahaw and San Cristobal and noted the presence of what appeared to be endemic species of Apomys and Rhynchomys. That study has remained unpublished also, aside from brief summary by Dans and Gonzalez (2010). The bats of Mt. Banahaw, however, have been the subject of a recent in-tensive study (Sedlock et al., 2011).
By 2004, it had become apparent to us that the diversi-ty of non-volant small mammals on Luzon was probably much higher than previously estimated. The limited data available suggested that each high mountain or mountain range that is surrounded (and isolated) by extensive low-lands would support a set of small mammal species that live only on that highland area, with species richness generally increasing with increasing elevation along a given transect. This observation led us to the hypothesis that the number of endemic species is a function of the height of the moun-tain (Balete et al., 2009; Heaney, 2001; Heaney & Rickart, 1990; Rickart et al., 1991, 2011a). The adjacent peaks of Mts. Banahaw and San Cristobal appeared to be a prime example of just such an isolated montane region, and there-fore an area highly suitable for testing this hypothesis, and, more generally, for examining patterns of small mammal distribution and endemism. Based on comparison with
Mt. Isarog, a volcanic peak of similar elevation that lies in southern Luzon (Heaney et al., 1999; Rickart et al., 1991), we predicted that at least three species of locally endemic mammals would be present on Mt. Banahaw. We began the second phase of our study in 2004, continued it in 2005, and completed it in 2012 (with the extensive delay due to temporary closure of the highest elevations in the park for administrative reasons). Our findings confirmed our predictions, but did so in some surprising ways. Our tax-onomic studies confirmed that the Rhynchomys first cap-tured by Rosell (1996) is indeed a species endemic to Mt. Banahaw, R. banahao (Balete et al., 2007). These studies also revealed not one but two species of large forest mice that are endemic (Apomys banahao and A. magnus; Heaney et al., 2011). Most remarkably, we captured a tiny arboreal mouse that initially could not be identified, and that un-officially became known as the “mystery mouse” (Heaney et al., 2004). Upon close study, it was found to be a previ-ously unknown genus and species (Musseromys gulantang Heaney et al., 2009) most closely related to the cloud rats (Batomys, Carpomys, Crateromys, and Phloeomys). Thus, four species of mammals are currently known only from Mt. Banahaw, a remarkable number of endemic species for so small an area; indeed, it is a level of endemism that exceeds that of many entire countries.
The purpose of this paper is to provide the first detailed information on the ecology of the small mammals that live on Mts. Banahaw and San Cristobal, based on our studies in 1995-96 and 2004-12. We present data on elevational range, abundance, habitat associations, degree of arbore-al activity, primary dietary preferences, and diel activity patterns of twelve species (nine native and three exotic), including the four endemic species.
Study AreA And MethodS Location and GeologyMt. Banahaw (Fig. 2A) is a stratovolcano located in the southernmost part of the Luzon Central Valley Basin, which is part of the Central Philippine Physiographic Province (Bureau of Mines and Geo-Sciences, Ministry of Natural Resources, 1982). This basin formed by in-filling of a ma-rine basin between the Zambales Range and the Southern Sierra Madre, with dry land that connected the two first appearing in the Pliocene; the deepest parts of the basin contain up to 6000 meters of sediment (Ku et al., 2009; Vogel et al., 2006). Mt. Banahaw, and the closely associat-ed Mts. San Cristobal and Banahaw de Lucban, are com-posed of volcanic materials, especially tuff, lying on top of the older marine sediments; the oldest volcanic materials on Mt. Banahaw date from about 1.98 Ma, and extend to
Fig. 1. Map of the study area, showing the locations of all sampling localities. See Study Area for details.
46

The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines
the present. Mt. San Cristobal is dated to a maximum of about 1.3 Ma, and Banahaw de Lucban is the youngest, per-haps Holocene in age (Bureau of Mines and Geo-Sciences, Ministry of Natural Resources, 1982; Geronimo-Catane, 1994; Ku et al., 2009; Vogel et al., 2006). These volcanoes are part of the Macolod Corridor, which probably origi-nated as a pull-apart rift zone in the late Pliocene; other major features of the Macolod Corridor include two vol-cano-tectonic depressions (Taal and Laguna de Bay), two additional stratovolcanos (Makiling and Cristo), and more than 200 small volcanic features including domes (Vogel et al., 2006). All three peaks in the park were originally smoothly cone-shaped, but two massive avalanches of de-bris, possibly simultaneous with magmatic eruptions, re-moved large portions of the eastern flank of Mt. Banahaw and flowed to Tayabas Bay. Mt. Banahaw de Lucban sub-sequently developed from small eruptions within the am-phitheater formed by the first avalanche, and is currently the most active of the three volcanoes (Geronimo-Catane, 1994). Thus, the set of three peaks dates to the late Pliocene (ca. 2 Ma), and have formed a volcanically active highland area since that time (see also Hall, 1998, 2002; Mitchell et al., 1986; Yumul et al., 2009). Other nearby volcanoes, sep-arated by extensive lowlands, include Taal (ca. 35 km to the west) and Mt. Makiling (ca. 30 km to the northwest).
Study Localities The 1995-1996 survey included eleven localities, four on Mt. Banahaw, four on Banahaw de Lucban, and three on Mt. San Cristobal. Our surveys in 2004-2005 included the same four localities on Mt. Banahaw, plus an additional one, and in 2012 included three additional localities on Mt. Banahaw. The descriptions below of the Mt. Banahaw localities that were sampled both times are based primarily on field notes from the 2004-2005 field seasons.
The population center of Barangay Lalo, which served as our base in 1995 and 2004-2005, lies at about 350 m in heavily utilized agricultural land on the south side of Mt. Banahaw, beside the Alataaw (Alitao) River. Patches of secondary lowland forest first appear at ca. 550 m el-evation, and mixed agriculture and secondary forest are interdigitated to about 750 m (Fig. 2A). Lowland forest (secondary and old growth) occurs from 750 m to 1100 m, with a transition to montane forest by about 1200 m.
Fig. 2. (A), Mt. Banahaw as seen from 620 m (Locality 1) on 13 May 2004, showing agricultural fields and second-growth lowland forest in the foreground; photo by E. A. Rickart. (B), mature montane forest at 1465 m (Locality 5) on 5 May 2004; photo by D. S. Balete. (C), mature montane forest at 1625 m (Locality 6) on 5 April 2012; photo by D. S. Balete.
A
B
c
47

National Museum of the Philippines: Journal of Natural History
A transition between montane and mossy forest occurs between 1700 m and 1800 m, with mossy forest from ca. 1800 m to the peak. The overall pattern of habitat and dis-turbance is similar on the north side in Barangay Bukal, but the population center lies at about 430 m, agricultural areas have less remnant forest, and there is a fairly rapid transition from mixed agriculture to secondary forest at about 750 m. Localities 1, 3 - 5, and 7 lie within Barangay Lalo, Tayabas Municipality, Quezon Province. Localities 2, 6, and 8 lie within Barangay Bukal, Majayjay Municipality, Laguna Province.
Locality 1. 5.1 km E, 2 km S Mt. Banahaw peak, 620 m elevation, 14o 3’ 6.5” N, 121o 32’ 22.5” E (July 1995 and 13 - 16 May 2004). This locality (Fig. 2A), locally known as Camp 1, lay adjacent to a small, enclosed tree nursery. We sampled in regenerating lowland forest interspersed with cropland (sweet potato, taro, cassava, bananas, and coco-nut) and pasture for horses and water buffalo in which tall grasses and erect bamboos were common. The terrain was hilly, with shallow slopes. Most traps were set within the disturbed forest near the Alataaw River, though some were set in shrubby habitat in the agricultural area. The canopy of the forest averaged ca. 25 m, with some emergents reach-ing 35 - 40 m; the largest had bole diameter (above the but-tresses) of up to ca. 1 m. Dipterocarps (Dipterocarpaceae) were the dominant trees, with figs (Ficus spp.), Elaeocarpus (Elaeocarpaceae) and some myrtles (Syzygium) also com-mon; most large trees had substantial buttresses. Canopy vines included lianas, climbing bamboo (Dinochloa), jade vines (Strongylodon macrobotrys), and climbing pandans (Freycinetia). Epiphytic ferns, orchids, and vines (Medinilla) were common on the large trees. Ground cover included saplings, small palms, ferns, tree ferns, and gingers. A thin layer of leaf litter sparsely covered the ground; humus was thin or absent. In shrubby areas around agricultural plots, saplings, Melastoma, ferns, and tall grasses predominated. Ants were abundant, and earthworms were uncommon. We noted temperatures as low as 19o C and as high as 27o C.
Locality 2. 4.0 km N, 1.75 km W Mt. Banahaw peak, 660 m elevation, 14.10581o N, 121.47307o E (17 - 19 April 2012). This locality was situated in an intensively cultivated area in Barangay Bukal. A few trees of the original lowland dipterocarp forest remained on the flatter slopes, mainly as boundary markers of vegetable farms. The strip of riparian vegetation restricted to the steep riverbank on the western side of this site was the only remnant of the natural vegeta-tion. Figs (Ficus spp.), Erythrina, Mangifera, dipterocarps (Shorea), and myrtles (Syzygium) were some of the cano-py and emergent trees of the riparian vegetation; the more
common understory vegetation consisted of saplings of the canopy trees, tree ferns (Cyathea), giant ferns (Angiopteris), rattans (Calamus), Pandanus, and sugar palm (Arenga pin-nata). Climbing bamboos (Dinochloa) and Tetrastigma were the common vines. Epiphytes included ferns, orchids, Begonia, Philodendron, and Freycinetia. Cultivated crops included bananas, beans, bitter melon, chayote, coconut, coffee, eggplant, pepper, taro, and tomatoes.
Locality 3. 1.25 km S, 4.75 km E Mt. Banahaw peak, 765 m elevation, 14o 3’ 26.9” N, 121o 31’ 57” E (July 1995 and 14 - 18 February 2005). This locality was known as Idoro among residents of Barangay Lalo. It is near to Locality 1, but had fewer forest clearings. The forest was regenerat-ing lowland dipterocarp forest, with species composition and structure similar to the forested portion of Locality 1.
Locality 4. 0.7 km S, 3 km E Mt. Banahaw peak, 1250 m elevation, 14o 3’ 44”N, 121o 31’ 8” E (April - May 1995 and 21 - 26 February 2005). We surveyed at this locality, known as Hasa-an to residents of Barangay Lalo, a portion of what appeared to be transitional lowland-montane forest, though elements of montane forest were more common. Canopy trees reached 20 m, commonly up to 15 m on ridgetops, with emergents up to about 25 m. Myrtles (Syzygium) and oaks (Lithocarpus) were common, along with some podo-carps (Podocarpus) and figs (Ficus). Pandanus and tree ferns (Cyathea) were common in the understory, along with palms (Pinanga) and rattan (Calamus). Buttressed trees were rare. Lianas and viney pandans (Freycinetia) were also common. Ephiphytes including orchids, Medinilla, ferns, mosses and liverworts were abundant. Leaf litter was common on the flatter forest floor, but scarce along steeper slopes except those caught in root tangles or piled behind the base of tree trunks. The humus layer was thin overall.
Locality 5. 0.6 km S, 1.75 km E Mt. Banahaw peak, 1465 m elevation, 14o 3’ 59.4” N, 121o 30’ 30.9” E (April - May 1995 and 30 April - 11 May 2004). This sampling area lay within old-growth montane forest (Fig. 2B), with only lo-calized human disturbance associated with “treasure hunt-ing”; it is locally known as “Camp 2”. Two small streams ran through the sampling area; the terrain was rough and steep, producing a rather uneven and broken canopy. Emergents reached about 20 m, but most large trees reached 12 - 15 m; these were predominantly Podocarpus (Podocarpaceae), Lithocarpus (Fagaceae), and Syzygium (Myrtaceae), although some figs (Ficus) and tree ferns (Cyathea) also reached the canopy. Dipterocarps were absent, and we noted no trees with buttresses. Rattan (Calamus) was common, and viney pandans (Freycinetia) and lianas were fairly common. Large trees supported heavy loads of ephiphytes, including moss,
48

The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines
ferns, liverworts, orchids, and some Medinilla. Understory plants included palms (Pinanga), tree ferns, ferns, rattan, and saplings. Little moss grew on the ground; leaf litter covered most of the forest floor, but the humus layer was rather thin over the generally steep, rocky soil. Ants were uncommon, and earthworms only moderately common. Temperatures on 4 days at ca. 07:00 hr averaged about 18.3o C, and at noon averaged about 20.7o C.
Locality 6. 1.0 km N, 0.4 km E Mt. Banahaw peak, 1625 m elevation, 14.07706o N, 121.49262o E (3 - 15 April 2012). This locality lay in old-growth montane forest (Fig. 2C), on slopes that averaged about 45o (range ca. 20-60o). The tallest trees reached ca. 20 m, with DBH of 75 cm, but the canopy was generally 12 - 15 m, with DBH of 30 - 75 cm. Canopy trees had small leaves, 20 - 40 mm, few of which were emarginate; podocarps (Podocarpus and Dacrycarpus), myrtles (Syzygium), laurels (Neolitsea, Persea, and Phoebe), and oaks (Lithocarpus) were most common. Figs (Ficus) were rare but present. Ferns, orchids, moss, Medinillia, and lipstick plants (Aeschynanthus) were all common epiphytes; vines included Freycinetia, which was abundant, Tetrastigma, viney bamboos (Dinochloa), and Piper. Understory and ground-cover plants includ-ed saplings, ferns, gingers, Melastoma shrubs, tree ferns (Cyathea), a few small palms (Pinanga), woody pandans (Pandanus), and Begonia; sedges were uncommon, and grasses appeared to be absent. We noted no wild banana (Musa spp.) or pitcher plants. Moss was common on the lower trunks of trees, fallen logs, and on large branches in the canopy, but little grew on the ground. Leaf litter cov-ered virtually the entire ground surface, but was thin (ca. 2 - 4 cm), and humus covered nearly the entire ground, but also was thin (ca. 2-5 cm), lying on top of weathered volcanic soil. Fallen logs were common; earthworms were common in these logs, but uncommon in the soil. Small ants were present but uncommon. Over 12 days, daily high temperatures averaged 19.0±1.1o C (16.6 - 20.5o C), and overnight lows averaged 13.8±1.8o C (10.6 - 16.3o C); evening and overnight fog was common.
Locality 7. 0.5 km S, 1.2 km E Mt. Banahaw peak, 1750 m elevation, 14o 3’ 49.7” N, 121o 30’ 12.5” E (7 - 11 May 2004). We surveyed at this locality in transitional montane-mossy forest on a steep slope. Canopy trees reached up to 12 - 15 m, with emergents up to 17 m, many with gnarled tree trunks. Common trees included myrtles (Syzygium) and oaks (Lithocarpus) as well as podocarps (Podocarpus and Dacrycarpus) and laurels (Lauraceae). Epiphytes, includ-ing ferns, orchids, mosses, and liverworts, were moderate-ly abundant on tree trunks and branches; ground orchids,
ferns, mosses and liverworts on the ground were also mod-erately abundant. Freycinetia, Smilax, and other vines were common. Leaf litter and humus covered the ground, but thickness was moderate on the steep slope. There were not many earthworms in the soil, but large ones were common-ly found inside rotting logs.
Locality 8. 0.05 km N, 0.15 km E Mt. Banahaw peak, 2030 m elevation, 14.06907o N, 121.49014o E (9 - 15 April 2012). This sampling locality included areas near the peak (2000-2050 m), but not including the peak itself, which is rocky, dry, and sparsely vegetated. The ground was moderately steep, averaging ca. 30o (range 15 - 60o). The vegetation was old-growth mossy forest; the tallest trees were 12 - 15 m, but the canopy, which was broken and uneven, was usually 5 - 7 m, with the larger trees ranging from 30 - 80 cm DBH. Podocarps (especially Podocarpus and Dacrycarpus) dominated, but Elaeocarpus, Syzygium, Lithocarpus, and members of the Theaceae (Eurya and Gordonia) were common; aside from feathery podocarp leaves, emarginate leaves were uncommon. Canopy epi-phytes included moss, orchids, ferns, and Medinilla; can-opy vines were uncommon, but included Tetrastigma and Freycinetia (with no bamboos). Understory plants includ-ed saplings, Helicia, Astronia, Melastoma, and Psychotria; ground cover included ferns, orchids, a few small shrubs, and a few sedges. Ficus, Musa, and pitcher plants appeared to be absent. Moss was generally abundant on trees and fallen logs, and some grew on the ground. Leaf litter and humus covered all of the ground surface, with leaf litter 2-6 cm thick and humus 5-100 cm thick, lying on top of weathered volcanic ash. Fallen logs were common, with earthworms common in the logs but not in the ground; ants appeared to be absent.
Localities 9 - 12 lie on the northeastern slope of Mt. Banahaw de Lucban (peak at ca. 1875 m), in Lucban Municipality, Quezon Province. This peak is connected to the main peak of Mt. Banahaw by a saddle at ca. 1550 m, which lies well within montane forest (Fig. 1). Habitat descriptions and data for Localities 9 - 15 are based on Rosell (1996).
Locality 9. Southern Luzon Polytechnic College refor-estation area, 1.45 km N, 2.3 km E Banahaw de Lucban peak, ca. 700 m elevation, 14.090159o N, 121.532437o E (July 1995). This sampling area lay in a mixture of cul-tivated fields, grassland, and secondary lowland rain for-est, similar to the vegetation at Localities 1 and 2. Some traps were set in a building, where many commensal rats were trapped.
Locality 10. 1.95 km N, 0.9 km W Banahaw de Lucban
49

National Museum of the Philippines: Journal of Natural History
peak, elevation 900 m, 14.094856o N, 121.520094o E, (October 1995). Our sampling area lay within a mixture of secondary and old-growth lowland forest, similar to that at Locality 3. Fig trees (Ficus) were especially common, as was rattan (Calamus).
Locality 11. 1.5 km N, 0.6 km W Banahaw de Lucban peak, 1100 m elevation, 14.090725o N, 121.516978o E (October 1995). We sampled in mature montane forest, similar to Locality 4. Rocky ridges, steep ravines, and dry creek beds were common. Moss was common on exposed rocks on a ridge, and viney pandans (Pandanus) and rat-tan reached into the canopy.
Locality 12. 0.5 km N, 0.25 km W Banahaw de Lucban peak, 1500 m elevation, 14.081912o N, 121.514417o E (October 1995). We sampled in mature montane forest with vegetation similar to that at Locality 5. Epiphytic plants, including orchids, ferns, and moss, were common, as were viney pandans (Freycinetia). A trail frequented by mountaineers led through the area.
Localities 13 - 15 lay on Mt. San Cristobal, which has a peak elevation of 1470 m (14o 4’ N, 121o 25’ E); all of the localities are within Dolores Municipality, Quezon Province. This peak is connected to Mt. Banahaw by a saddle at ca. 820 m (Fig. 1).
Locality 13. 1.4 km S, 0.6 km E Mt. San Cristobal peak, 900 m elevation, 14.05391o N, 121.43180o E (March - April 1996). The lower slopes of Mt. San Cristobal had been mostly cleared and converted to agriculture; evidence of logging was common, and fire-maintained grassland was widespread. Small patches of lowland forest, dominated by dipterocarps, remained in some deep ravines.
Locality 14. 0.95 km S, 0.25 km E Mt. San Cristobal peak, 1100 m elevation, 14.05751o N, 121.42833o E (March - April 1996). This sampling area was located within ma-ture montane forest with elements of lowland forest; there was little evidence of logging. The vegetation was gener-ally similar to that at Locality 4; epiphytes were common but moss was scarce, and bamboo was commonly present. The understory was dominated by saplings, small palms, and herbaceous growth.
Locality 15. 0.8 km S, 0.1 km W Mt. San Cristobal peak, 1300 m elevation, 14.05858o N, 121.42504o E (March - April 1996). This study area was located in mixed montane forest and grassland; disturbance was associated with a shallow volcanic vent and a camping area used by hikers. Trees were short in stature, with some moss present; the vegeta-tion was generally similar to that at Locality 4.
Field MethodsOur surveys conducted in 2004, 2005, and 2012 followed the standard field methods employed in our previous sur-veys of Philippine small mammals, so that direct compari-son is possible (Alviola et al., 2011; Balete et al., 2009, 2011; Duya et al., 2011; Heaney et al., 1989, 1999, 2006b; Rickart, 1993; Rickart et al., 1991, 2011a); the following description of those methods is based on Heaney et al., 2013. When procedures differed between 1995-1996 and 2004-2012, comments about procedures during the former are based on Rosell (1996).
At each locality we limited sampling to an area within a 50 m elevational band above and below the central el-evation, as listed above (see Study Area). Elevation was recorded with an altimeter. Latitude and longitude were determined with a hand-held GPS device utilizing the 1911 Philippine Luzon Datum (2004-2012) or from topographic maps (1995-1996). We took note of features of the vege-tation at each locality, as described below, and noted the relative abundance of earthworms and ants (scarce, un-common, common, abundant). Temperatures were mea-sured in the shade, ca. 1.5 m above ground; in 2004-2005, we took temperatures with a hand-held thermometer at the stated times (see “Study Areas”); in 2012, we used a digital minimum-maximum thermometer that recorded daily high and low temperatures.
For the 2004-2012 inventories, traps were baited with either fried coconut coated with peanut butter, or with live earthworms obtained at or near the sampling locality. Victor rat traps were set along runways, under root tan-gles, and beside fallen logs. Museum Special snap traps were set on horizontal branches of trees, overhanging lia-nas, and vines at heights up to 5 m. The location (ground or arboreal), type of trap, and type of bait were recorded for each trap; details are summarized below. Trap lines were maintained for three consecutive days before being transferred to another site.
In 1995-1996, about 90% of the trap-nights represent-ed Victor snap traps, and 10% were Sherman and cage live traps. We set 80-90% on the ground, and the rest on horizontal logs or branches above ground. The same two baits were used.
At each locality, traps were set on ridge tops and along slopes and ridge-sides to sample all available microhabi-tats. Traps were serviced at least twice each day, in the ear-ly morning (ca. 07:00 h) and in the late afternoon (before sundown, ca. 17:00 h), to retrieve captures and replace bait. Animals retrieved at 07:00 h were considered nocturnal/
50

The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines
crepuscular captures whereas animals retrieved from 07:00 h to ca. 17:00 h were diurnal captures.
All individuals captured during the 2004-2012 surveys were preserved as voucher specimens. A tissue sample taken from the thigh was placed in DMSO buffer solution prior to specimen preservation. Fluid-preserved specimens were initially injected with saturated formalin solution, then placed in 10% formalin and later transferred to 70% eth-anol. Other specimens were prepared as complete skele-tons in the field and subsequently cleaned using dermestid beetles followed by soaking in a weak ammonia solution. Skulls from some fluid-preserved specimens were later re-moved and similarly cleaned. Specimens were deposited at the Field Museum of Natural History, Chicago (FMNH) for study; half will be returned to the National Museum of the Philippines, Manila (NMP) after completion of our stud-ies. The identifications were verified at the Field Museum.
Some voucher specimens from 1995-1996 were pre-served in formalin, and some skulls were removed and cleaned. Identifications here are based on information in Rosell (1996), as discussed below; no voucher specimens were available for study.
Nomenclature used here is based on Heaney et al. (1998, 2010, 2011). The capture and handling of animals in the field were conducted in accordance with all relevant Philippine laws and regulations.
Data Analysis Because data from 1995-1996 were recorded differently than in 2004-2012, we do not include them in most of our statistical analyses; exceptions are noted below.
For 2004-2012 data, we used a “range-through” assump-tion of elevational distribution where the number of spe-cies at each locality was estimated as the number recorded directly, plus the number inferred to be present because of their presence at localities at both higher and lower eleva-tions along the gradient (e.g., Rickart et al., 1991, 2011a). To measure the adequacy of sampling, we used species accumulation curves that plotted the cumulative num-ber of species captured against the number of trap-nights (defined as one trap set for 24 hours). We plotted accu-mulation curves for individual localities and also plotted a curve for all eight sampling areas combined, including all types of traps and baits, beginning at the lowest eleva-tion and proceeding to the highest. This method has been used previously in the Philippines to assess the complete-ness of sampling effort for mammalian species richness
(e.g., Balete et al., 2009, 2011; Heaney et al., 1999, 2006a; Rickart, 1993; Rickart et al., 1991, 2011a).
Because the sampling efforts at 660 m and 1750 m were limited (less than 500 trap-nights), we did not include these localities in our tests of elevational patterns, but we did include the data in our assessment of bait preference, diel activity patterns, and trap position.
Relative abundance for each species at each sampling locality was derived by dividing the number of individu-als of each species captured times 100 by the number of relevant trap-nights (i.e., the number of captures per 100 trap-nights). Bait preference of each species was derived in a similar manner, expressed as the number of captures per bait type times 100, divided by the relevant total of trap-nights for the bait. Weighted trap success at each locality was calculated as the number of individuals cap-tured times 100, divided by the total number of ground traps set. The equalized (unweighted) trap success was expressed as the average of each species’ trap success for the two bait types used.
Chi-squared tests were used to determine if differenc-es for each species were significant for bait type, diel ac-tivity pattern, trap position, and number captured at each locality, with expected values calculated on the basis of the number of relevant trap-nights. For diel activity, the expected frequency was computed as the ratio 10:14, the proportion (in 24 hours) of diurnal and nocturnal/crepus-cular activity time periods. For all χ2 tests, we followed the convention of conducting tests only on total samples of 20 or more individuals, with five or more individuals per “treatment” in at least 80% of the treatments (e.g., per elevation or bait type). For sample sizes of less than 20, we used a binomial distribution test when appropriate. The relationship of elevation with species richness, rela-tive abundance (number of individuals/trap-night), and percent individuals diurnal was assessed using Spearman’s coefficient of rank correlation (rs).
reSuLtS We report the results from Mt. Banahaw (Localities 1 - 8) first, including results from 2004-2012, which permit de-tailed analysis, followed by results from 1995-1996 from Mt. Banahaw, Mt. Banahaw de Lucban, and Mt. San Cristobal.
We reached a total of 7,640 trap-nights on Mt. Banahaw during 2004-2012, and 1,362 trap-nights during 1995, for a total of 9,002 trap-nights. During 2004-2012, arboreal traps accounted for 28% of the total, and ground traps for 72%.
51
National Museum of the Philippines: Journal of Natural History
Of the ground traps, 3,595 were baited with fried coconut (65%), and 1,902 (35%) were baited with live earthworms.
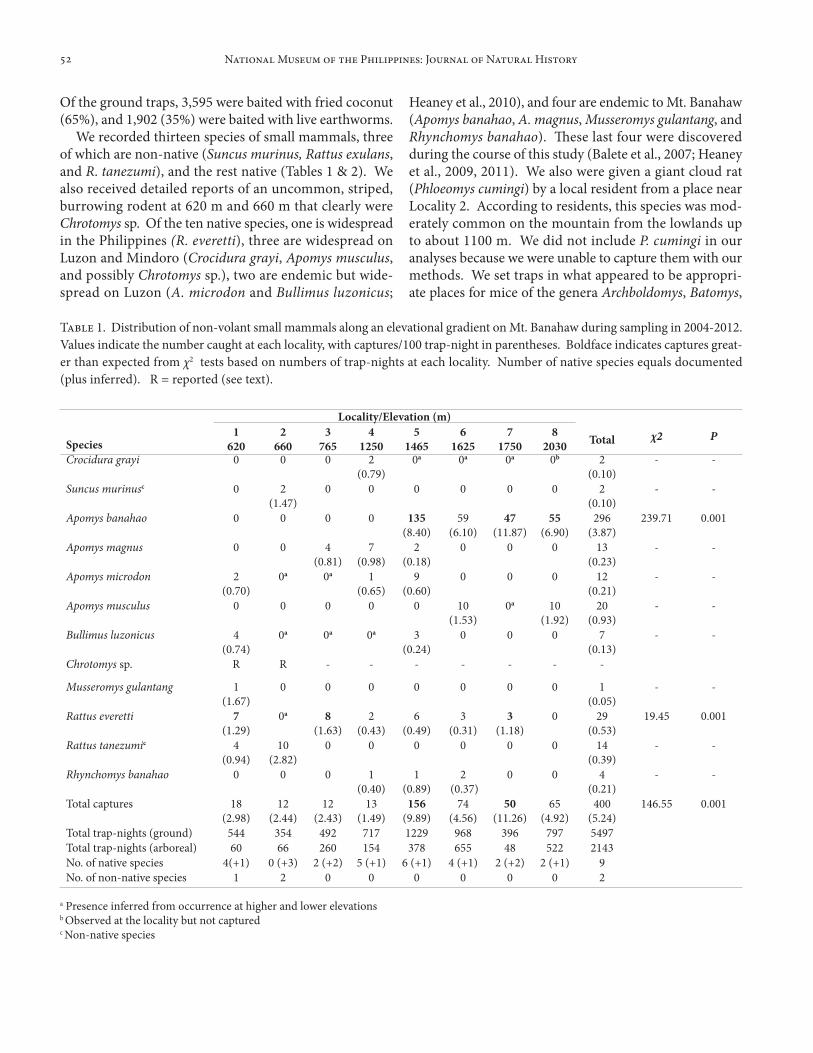
We recorded thirteen species of small mammals, three of which are non-native (Suncus murinus, Rattus exulans, and R. tanezumi), and the rest native (Tables 1 & 2). We also received detailed reports of an uncommon, striped, burrowing rodent at 620 m and 660 m that clearly were Chrotomys sp. Of the ten native species, one is widespread in the Philippines (R. everetti), three are widespread on Luzon and Mindoro (Crocidura grayi, Apomys musculus, and possibly Chrotomys sp.), two are endemic but wide-spread on Luzon (A. microdon and Bullimus luzonicus;
Heaney et al., 2010), and four are endemic to Mt. Banahaw (Apomys banahao, A. magnus, Musseromys gulantang, and Rhynchomys banahao). These last four were discovered during the course of this study (Balete et al., 2007; Heaney et al., 2009, 2011). We also were given a giant cloud rat (Phloeomys cumingi) by a local resident from a place near Locality 2. According to residents, this species was mod-erately common on the mountain from the lowlands up to about 1100 m. We did not include P. cumingi in our analyses because we were unable to capture them with our methods. We set traps in what appeared to be appropri-ate places for mice of the genera Archboldomys, Batomys,
Species
Locality/Elevation (m)
Total χ2 P1620
2660
3765
41250
51465
61625
71750
82030
Crocidura grayi 0 0 0 2 (0.79)
0a 0a 0a 0b 2 (0.10)
- -
Suncus murinusc 0 2 (1.47)
0 0 0 0 0 0 2(0.10)
- -
Apomys banahao 0 0 0 0 135 (8.40)
59 (6.10)
47 (11.87)
55 (6.90)
296 (3.87)
239.71 0.001
Apomys magnus 0 0 4 (0.81)
7 (0.98)
2 (0.18)
0 0 0 13 (0.23)
- -
Apomys microdon 2 (0.70)
0a 0a 1 (0.65)
9 (0.60)
0 0 0 12 (0.21)
- -
Apomys musculus 0 0 0 0 0 10 (1.53)
0a 10 (1.92)
20 (0.93)
- -
Bullimus luzonicus 4 (0.74)
0a 0a 0a 3 (0.24)
0 0 0 7 (0.13)
- -
Chrotomys sp. R R - - - - - - -
Musseromys gulantang 1 (1.67)
0 0 0 0 0 0 0 1 (0.05)
- -
Rattus everetti 7 (1.29)
0a 8 (1.63)
2 (0.43)
6 (0.49)
3 (0.31)
3 (1.18)
0 29 (0.53)
19.45 0.001
Rattus tanezumic 4 (0.94)
10 (2.82)
0 0 0 0 0 0 14 (0.39)
- -
Rhynchomys banahao 0 0 0 1 (0.40)
1(0.89)
2 (0.37)
0 0 4 (0.21)
- -
Total captures 18 (2.98)
12 (2.44)
12 (2.43)
13 (1.49)
156 (9.89)
74 (4.56)
50 (11.26)
65(4.92)
400 (5.24)
146.55 0.001
Total trap-nights (ground) 544 354 492 717 1229 968 396 797 5497Total trap-nights (arboreal) 60 66 260 154 378 655 48 522 2143No. of native species 4(+1) 0 (+3) 2 (+2) 5 (+1) 6 (+1) 4 (+1) 2 (+2) 2 (+1) 9No. of non-native species 1 2 0 0 0 0 0 0 2
a Presence inferred from occurrence at higher and lower elevationsb Observed at the locality but not capturedc Non-native species
Table 1. Distribution of non-volant small mammals along an elevational gradient on Mt. Banahaw during sampling in 2004-2012. Values indicate the number caught at each locality, with captures/100 trap-night in parentheses. Boldface indicates captures great-er than expected from χ2 tests based on numbers of trap-nights at each locality. Number of native species equals documented (plus inferred). R = reported (see text).
52

The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines
Carpomys, and Soricomys, but did not capture any and saw no sign of them.
Species richness (measured plus inferred) of native spe-cies in forested habitats ranged from five at 620 m, four at 765 m, six at 1250 m, seven at 1465 m, five at 1625 m, four at 1750 m, to three at 2030 m; only at 620 m, which included some agricultural plots, did we capture a non-na-tive species (Rattus tanezumi; Table 1). Our inadequately sampled, heavily disturbed agricultural site (660 m) had two non-native species (Suncus murinus and R. tanezumi) and no native species, though one native species (Chrotomys sp.) was reported and two others might be inferred by their presence at lower and higher elevations (Apomys microdon and Bullimus luzonicus; Table 1).
Our trapping effort at 620 m and 660 m (604 and 420 trap-nights, respectively), was less than the estimated min-imum requirement of 800 trap-nights to effectively sample small mammals on Luzon (Balete et al., 2009; Heaney et al., 1989, 2006a, b; Rickart et al., 1991, 2011a). As such, we may not have captured all species that were present at these localities, and indeed, as noted above, Chrotomys sp. was reported to be present but was not captured. At 765 m, a plateau was reached at two species on the first night of sampling and none was added by the final total of 752 trap-nights (Fig. 3A). At 1250 m, two species were added on the last night of 871 trap-nights. At 1465 m, we had over 1600 trap-nights, and reached a plateau at six species by about 900 trap-nights (Fig. 3A). With over 1,600 trap-nights at
Table 2. The number of small non-volant mammals trapped at study sites on Mt. Banahaw in 1995-1996. The num-bers of captures per 100 trap-nights are given in parentheses.
Mt. Banahaw Mt. Banahaw de Lucban Mt. San Cristobal
Locality Number 1620m
3765m
41250m
51465m
9700m
10900m
111100m
121500m
13900m
141100m
151300m
Species
Crocidura grayi 0 0 1(0.35)
0 0 0 0 0 0 0 0
Suncus murinus 0 0 0 0 0 0 0 1(1.43)
0 0 10(2.52)
Apomys banahao 0 0 0 60(13.04)
0 0 0 30(12.98)
0 0 0
Apomys magnus 0 2(0.42)
16(5.53)
0 0 1(0.62)
6(2.06)
0 0 2(0.58)
0
Apomys microdon 0 1(0.21)
0 0 0 0 0 0 0 0 0
Bullimus luzonicus 0 0 0 0 0 0 1(0.29)
0 0 1(0.29)
0
Rattus everetti 0 3(0.62)
0 1(0.22)
2(0.99)
1(0.62)
0 5(2.16)
0 0 1(0.25)
Rattus exulans 0 0 0 0 0 0 0 0 0 0 1(0.25)
Rattus tanezumi 0 0 0 0 7(3.48)
0 0 0 0 0 0
Rhynchomys banahao 0 0 1(0.35)
0 0 0 0 0 0 0 0
Total captures 0 6 18 61 9 2 7 36 0 3 12
Total trap-nights 137 476 289 460 201 162 340 231 209 342 396
Total mammals per 100 trap-nights 0 1.26 6.23 13.26 4.48 1.23 2.06 15.58 0 0.88 3.03
Number of Species 0 4 3 2 2 2 3 3 0 2 3
53
National Museum of the Philippines: Journal of Natural History
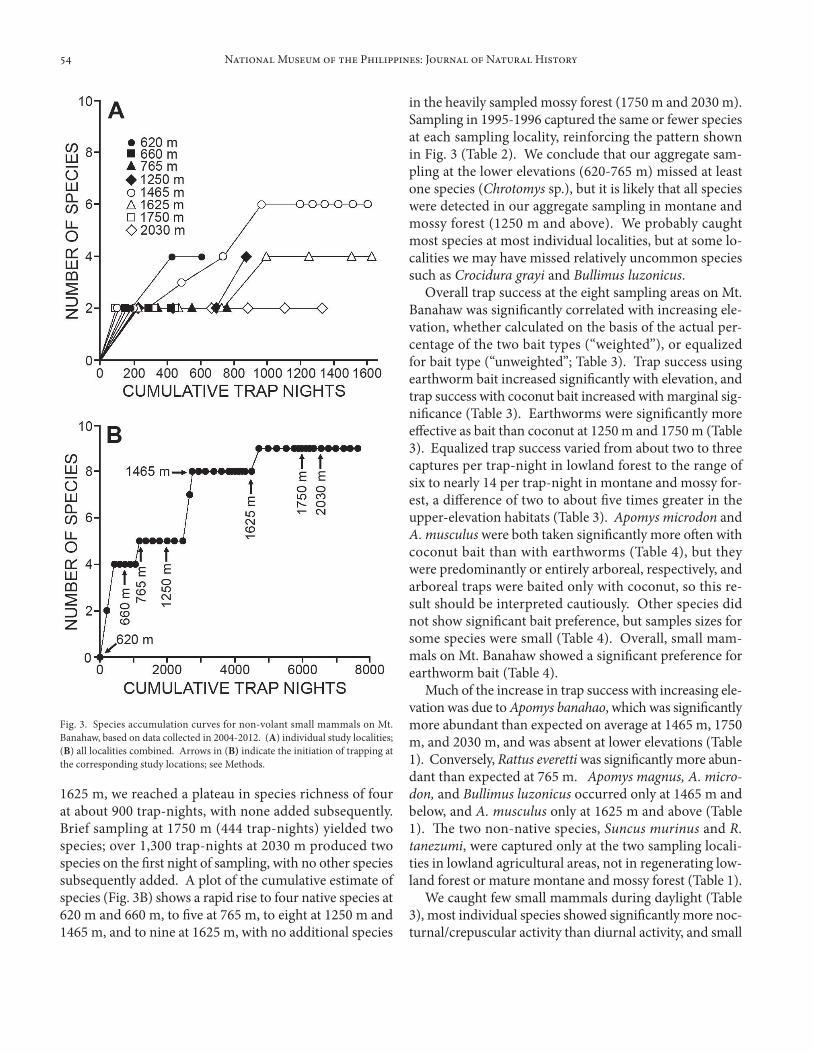
1625 m, we reached a plateau in species richness of four at about 900 trap-nights, with none added subsequently. Brief sampling at 1750 m (444 trap-nights) yielded two species; over 1,300 trap-nights at 2030 m produced two species on the first night of sampling, with no other species subsequently added. A plot of the cumulative estimate of species (Fig. 3B) shows a rapid rise to four native species at 620 m and 660 m, to five at 765 m, to eight at 1250 m and 1465 m, and to nine at 1625 m, with no additional species
in the heavily sampled mossy forest (1750 m and 2030 m). Sampling in 1995-1996 captured the same or fewer species at each sampling locality, reinforcing the pattern shown in Fig. 3 (Table 2). We conclude that our aggregate sam-pling at the lower elevations (620-765 m) missed at least one species (Chrotomys sp.), but it is likely that all species were detected in our aggregate sampling in montane and mossy forest (1250 m and above). We probably caught most species at most individual localities, but at some lo-calities we may have missed relatively uncommon species such as Crocidura grayi and Bullimus luzonicus.
Overall trap success at the eight sampling areas on Mt. Banahaw was significantly correlated with increasing ele-vation, whether calculated on the basis of the actual per-centage of the two bait types (“weighted”), or equalized for bait type (“unweighted”; Table 3). Trap success using earthworm bait increased significantly with elevation, and trap success with coconut bait increased with marginal sig-nificance (Table 3). Earthworms were significantly more effective as bait than coconut at 1250 m and 1750 m (Table 3). Equalized trap success varied from about two to three captures per trap-night in lowland forest to the range of six to nearly 14 per trap-night in montane and mossy for-est, a difference of two to about five times greater in the upper-elevation habitats (Table 3). Apomys microdon and A. musculus were both taken significantly more often with coconut bait than with earthworms (Table 4), but they were predominantly or entirely arboreal, respectively, and arboreal traps were baited only with coconut, so this re-sult should be interpreted cautiously. Other species did not show significant bait preference, but samples sizes for some species were small (Table 4). Overall, small mam-mals on Mt. Banahaw showed a significant preference for earthworm bait (Table 4).
Much of the increase in trap success with increasing ele-vation was due to Apomys banahao, which was significantly more abundant than expected on average at 1465 m, 1750 m, and 2030 m, and was absent at lower elevations (Table 1). Conversely, Rattus everetti was significantly more abun-dant than expected at 765 m. Apomys magnus, A. micro-don, and Bullimus luzonicus occurred only at 1465 m and below, and A. musculus only at 1625 m and above (Table 1). The two non-native species, Suncus murinus and R. tanezumi, were captured only at the two sampling locali-ties in lowland agricultural areas, not in regenerating low-land forest or mature montane and mossy forest (Table 1).
We caught few small mammals during daylight (Table 3), most individual species showed significantly more noc-turnal/crepuscular activity than diurnal activity, and small
Fig. 3. Species accumulation curves for non-volant small mammals on Mt. Banahaw, based on data collected in 2004-2012. (A) individual study localities; (B) all localities combined. Arrows in (B) indicate the initiation of trapping at the corresponding study locations; see Methods.
54

The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines
mammals as a whole were significantly more nocturnal/crepuscular than diurnal (Table 4). There was not a sig-nificant correlation between frequency of diurnal activity and elevation (Table 3).
Trap success was significantly greater on the ground than above ground, based on the numbers of traps set in each position (Table 4). Seven species were captured only on the ground. Only Apomys musculus was trapped sig-nificantly more often than expected in arboreal traps (all 20 captures were arboreal), although a majority of A. mi-crodon were also captured above ground as was the single capture of Musseromys. Of 396 A. banahao, only three were captured above ground (Table 4) and these were less than 1.5 m above ground on slanted trees or fallen logs.
Our results from 1995-1996 on Mt. Banahaw were en-tirely consistent with those from 2004-2012. At 620 m, we captured no small mammals in 137 trap-nights, a reflection of low overall abundance at this locality (Tables 1 and 2).
At 765 m (476 trap-nights), we captured two large Apomys (probably A. magnus) and three Rattus everetti, the same species we found to be most common in 2005 (Table 1). We captured one Crocidura grayi, 16 large Apomys (probably A. magnus), and one Rhynchomys banahao at 1250 m (289 trap-nights), the same species and similar relative abun-dances as in 2005. At 1465 m in 1995 (460 trap-nights), we captured 60 large Apomys (probably A. banahao) and one Rattus everetti, again in similar relative abundance to our captures in 2004 (Tables 1 and 2).
On Mt. Banahaw de Lucban in 1995 - 1996, with limit-ed trap-nights at each locality (Table 2), we captured two Rattus everetti (a native species) and seven R. tanezumi (a non-native species) in mixed agriculture and shrubby sec-ond growth at 700 m. In secondary lowland forest at 900 m, we captured one individual each of two species (Apomys magnus and R. everetti). In montane forest at 1100 m, we captured six individuals of large-bodied Apomys (probably
Locality/Elevation (m)rs1
6202
6603
7654
12505
14656
16257
17508
2030 Totals
Coconut bait (ground)Trap-nights 287 218 234 464 1117 542 254 479 3595Captures 11 8 6 3 127 35 25 35 250Captures/ 100 trap-nights 3.83 3.67 2.56 0.64 11.37 6.46 9.84 7.30 6.95 0.595*
Earthworm bait (ground)Trap-nights 257 136 258 253 112 426 142 318 1902Captures 6 4 6 9 18 29 25 20 117Captures/100 trap-nights 2.33 2.94 2.32 3.56 16.07 6.81 17.60 6.29 6.15 0.762**χ2 0.97 0.13 0.03 8.29 1.91 0.04 4.35 0.29 1.20P 0.324 0.717 0.866 0.004** 0.167 0.834 0.037** 0.592 0.273
Overall trap success (%)Weighted 3.12 3.39 2.43 1.67 11.80 6.61 12.63 6.90 6.68 0.667**Unweighted 3.08 3.30 2.44 2.10 13.72 6.63 13.72 6.80 6.55 0.647**
Coconut bait (arboreal) Trap-nights 60 66 260 154 378 665 48 522 2143Captures 1 0 0 1 11 10 0 10 33Captures/100 trap-nights 1.67 0 0 0.65 2.91 1.53 0 1.92 1.54 0.244
Diel periodDiurnal 0 0 0 1 4 2 0 0 7Nocturnal/crepuscular 18 12 12 12 152 72 50 65 393Percent diurnal 0 0 0 8.33 2.56 2.70 0 0 1.75 0.136
* = 0.1>P>0.05** = P< 0.05
Table 3. Summary of overall capture frequencies by bait type and diel period of non-volant mammals along an elevational gradi-ent on Mt. Banahaw. Captures greater than expected from χ2 tests are given in boldface. Correlation between elevation and cap-tures or overall trap success is indicated by Spearman’s correlation, rs. Brackets indicate data that were not included in statistical tests due to low sampling effort (see Methods).
55

National Museum of the Philippines: Journal of Natural History
A. magnus) and one Bullimus luzonicus. In mature mossy forest at 1500 m, somewhat disturbed by a campsite fre-quented by campers, we captured one Suncus murinus, 30 large Apomys (probably A. banahao), and five R. everetti (Table 2).
On Mt. San Cristobal in 1995-1996, in 209 trap-nights in mixed agriculture and exotic grassland, we captured no small mammals (Table 2). In mature montane forest at 1100 m, we captured two Apomys (probably A. magnus) and one Bullimus luzonicus. In montane forest mixed with exotic grassland and disturbed by a volcanic vent, at 1300 m, we captured ten Suncus murinus, one Rattus everetti, and one Rattus exulans (Table 2).
Although this paper focuses on small mammals, we learned from local residents that all of the large mam-mals, including monkeys (Macaca fascicularis), Malay civet (Viverra tangalunga), palm civet (Paradoxurus hermaphro-ditus), wild pig (Sus philippensis), and deer (Cervus marian-nus) are heavily hunted. According to our sources, the first three are now less common than in the recent past (roughly 10-20 years), wild pigs are much less common, and deer
are now very rare on the mountain. Large bats (Pteropus vampyrus) and giant cloud rats (Phloeomys cumingi) are hunted and have also declined, but perhaps less than the other species. Similar but more detailed observations on hunting were provided by Scheffers et al. (2012).
dIScuSSIonThis study constitutes the first comprehensive description of the small mammals of Mt. Banahaw - San Cristobal National Park, and confirms that four of its nine native small mammals are local endemic species. The presence of so many locally restricted species seems exceptional from a global perspective; indeed, many entire countries have fewer endemic mammals than Mts. Banahaw - San Cristobal. However, we now recognize that this level of local endemism is almost commonplace on Luzon, where it appears that a majority of the mammal species occur on only a single isolated mountain or mountain range, and many species have very limited distributions (e.g., Balete et al., 2007, 2012; Heaney, 2001; Heaney et al., 2009, 2010, 2011; Rickart et al., 1991, 1998). In this respect, Mt.
Species Total trapped
Bait type Diel period Trap position
Coconut Earthworm Nocturnal/crepuscular Diurnal Ground
surfaceAbove
groundCrocidura grayi 2 0 2 (0.10) 2 0 2 0Suncus murinus a 2 0 2 (0.10) 2 0 2 0Apomys banahao 296 210 (3.66) 86 (4.52) 291f 5 293 f 3Apomys magnus 13 5 (0.14) 8 (0.42) 13 d 0 13 d 0Apomys microdon 12 12 e, f (0.21) 0 (0) 12 d 0 3 9Apomys musculus 20 20 c,f (0.93) 0 (0) 20 b 0 0 20 b
Bullimus luzonicus 7 5 (0.14) 2 (0.10) 6 1 7 d 0Musseromys gulantang 1 1 (0.05) 0 (0) 1 0 0 1
Rattus everetti 29 19 (0.53) 10 (0.53) 29 b 0 29 b 0
Rattus tanezumia 14 11 (0.31) 3 (0.16) 14 d 0 14 d 0
Rhynchomys banahao 4 0 (0) 4 (0.21) 3 1 4 0TOTAL 400 283 117 b 393 f 7 367 f 33
a Non-native species b P < 0.05c P < 0.01d P < 0.001e P < 0.05, from binomial distribution f Only coconut bait was used on arboreal traps
Table 4. Summary of captures of the non-volant small mammals by bait type, diel period, and trap position on Mt. Banahaw. Captures per 100 trap-nights are indicated in parentheses. Captures greater than expected from χ2 test are shown in boldface.
56

The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines
Banahaw - San Cristobal National Park is deserving of recognition as a premier example of localized mammali-an endemism, along with the Central Cordillera (Balete et al., 2012; Heaney et al., 2011; Rickart et al., 2011a,b), the northern Sierra Madre Range (including Mts. Cetaceo and Anacuao; Duya et al., 2008, 2011; Heaney et al., 2013), the Mingan Mountains (Balete et al., 2011), the Zambales Range (including Mts. Tapulao and Natib; Balete et al., 2009; Rickart et al., 2013), the southern Sierra Madre Range (including Mt. Irid; Balete et al., 2013), Mt. Labo (Balete et al., 2013), and Mt. Isarog (Rickart et al., 1991; Heaney et al, 1999). Four species of Platymantis frogs and two species of the giant, parasitic plants of the genus Rafflesia are currently considered to be endemic to Mt. Banahaw - San Cristobal (Ambal et al., 2012; Barcelona et al., 2009; Barcelona, pers. comm.), further emphasizing the distinc-tiveness of the biota as a whole.
As we have found elsewhere on Luzon (e.g., Balete et al., 2009; Rickart, 1993; Rickart et al., 1991, 2011b), we captured non-native species of small mammals (Suncus murinus, Rattus exulans, and R. tanezumi) only in heav-ily disturbed habitats where few native small mammals were present (Tables 1 and 2). The near or total absence of non-native species in either secondary or old-growth forest is striking, in comparison with the abundance and species richness of native small mammals in those forests.
As on nearly all other mountains on Luzon, we found that species richness of native small mammals was low in the lowlands, increased steadily with increasing elevation to its highest level at a moderately high elevation (on Mt. Banahaw, at 1465 m, Locality 5), and declined thereafter. Mt. Banahaw is somewhat unusual in that the maximum number of species occurred within montane forest, rath-er than at the more usual area of transition from montane to mossy forest, which occurs on Banahaw at ca. 1750 m (e.g., Heaney, 2001; Heaney et al., 2013; Rickart et al., 1991, 2011a). This may be associated with the absence on Banahaw of genera that occur only at high elevation else-where on Luzon (e.g., Archboldomys, Batomys, Carpomys, and Soricomys, and highland Chrotomys).
As on most other mountains on Luzon, we found that overall abundance of small mammals was significantly cor-related with increasing elevation, by a factor of between three and six. This appears to be associated with a shift from lowland forest with abundant ants and little leaf litter and humus, to montane and mossy forest with progressively fewer ants and increasingly abundant fungi and soil inver-tebrates (including earthworms) living in deep leaf litter and humus, in forest with abundant oaks (Lithocarpus).
This shift in soil and food resources is clearly evident in the significant increase in the attractiveness of live earthworms as bait with increasing elevation (Table 4). The increase in abundance on Banahaw was also generally associated with the increase in species richness, though not in a simple, nearly linear fashion, as seen on some other Luzon moun-tains (e.g., Balete et al., 2009, 2011; Heaney, 2001; Heaney et al., 2013; Rickart et al., 1991, 2011a).
From work elsewhere on Luzon we predicted that spe-cies occurring at high elevations would be confined to mountains or mountain ranges that are isolated from other high elevation areas by lowland areas below 400 m (Heaney et al., 2013; Rickart et al., 2011a). The species of small mammals that are restricted to Mt. Banahaw and San Cristobal are generally consistent with our prediction. We found Apomys banahao only above about 1400 m, on both Banahaw and Banahaw de Lucban (Tables 1 and 2), where it is quite isolated from all other mountains on Luzon, and even A. magnus and Rhynchomys banahao, which extend down to 765 m and 1250 m, respectively, occur far above the broad lowland plains. Only Musseromys gulantang occurs in what is clearly lowland forest of a type that is similar to what would have occurred broadly in central Luzon; in that respect, its seemingly limited distribution is anomalous. Further research on this enigmatic species is clearly warranted.
We have previously documented that the number of species of native small mammals on Luzon mountains is correlated with the peak elevation of the mountain. With ten species, Mt. Banahaw follows this pattern quite clear-ly (Rickart et al., 2011a; Heaney et al., 2013). Overall, our data from Mt. Banahaw further demonstrate the consis-tency of patterns of small mammal diversity, distribution, and abundance throughout Luzon Island.
As noted above, the larger species of mammals on the three mountains have declined, especially wild pigs and deer. These species, when present in undisturbed conditions, may have significant impact on the ecology of the small mammals, through predation (the civets) or through habi-tat modification from digging up the forest floor in search-ing for earthworms, other invertebrates, fungi, etc. (pigs). The impact of the decreased abundance of these species on the small mammals is unknown but worthy of study.
concLuSIonWith four species of mammals that occur nowhere else, Mt. Banahaw is a classic example of localized endemism of small mammals; it is, in effect, an island of forested habi-tat, with each of the three major types of forest (lowland,
57

National Museum of the Philippines: Journal of Natural History
montane, and mossy) supporting one or two unique spe-cies of mammals, surrounded by a sea of people and in-tensively used anthropogenic habitat. This study provides basic information on the distribution and ecology of these unique species, and of the other small mammals that oc-cur in the Mt. Banahaw - San Cristobal National Park. However, much further research is needed to understand this remarkable fauna. For example, a study of the ecol-ogy of how the species interact with each other and use resources would tell us much about how this set of close-ly related species have co-evolved since their ancestors arrived on Luzon 12-15 million years ago. Comparing their ecology to small mammal communities in similarly species-rich but compositionally different places such as Mt. Isarog, or to communities in more species-rich plac-es such as the Central Cordillera, would be equally inter-esting. Studies addressing the ecological interactions of small mammals with larger mammals and the potential impact of the loss of large species on habitat and commu-nity structure would be of great value, particularly as pop-ulations of pig and deer continue to plummet throughout the Philippines (Schaffers et al., 2012).
Data provided here demonstrate that Mt. Banahaw - San Cristobal National Park is clearly a jewel in the system of protected areas on Luzon. Its function as a crucial water-shed for the heavily populated surrounding lowlands, its great natural beauty as seen from the surrounding plains, and its role as a center for religious observance, a place for enjoyment of nature, and habitat for a remarkable number of unique mammal species (plus other groups as well), to-gether offer strong justification for its continued protection.
AcknowLedgMentSWe sincerely thank everyone who offered support and as-sistance during this project. In Tayabas and Majayjay, we acknowledge the cooperation of the barangay officials of Lalo and Bukal, respectively, and local residents who en-thusiastically served as cooks, guides, and porters. In the Department of Environment and Natural Resources, we are grateful for the continued support and encouragement of the Protected Areas and Wildlife Bureau, including T. M. Lim, J. de Leon, M. Mendoza, and A. Tagtag. We also thank Protected Area Superintendant S. Pangan for her assistance in the issuance of permits.
For the 1995-1996 field work, we acknowledge the support and guidance of P. L. Alviola, A. T. L. Dans, W. Dee, and P. P. Ocampo. We thank D. Afan, P. A. Alviola, G. Bueser, A. Diesmos, A. Dimapilis, A. Duya, J. C. T. Gonzales, and M. Pedregosa for their assistance in the field
data collection. We also thank Luksang Alyansa Bundok Banahaw, Southern Polytechnic College, the John D. and Catherine T. MacArthur Foundation and the University of the Philippines, Los Baños for their support.
We thank J. Sarmiento, R. Buenviaje, and P. Plutado for their assistance in the field in 2004-2012. The figures were prepared by A. Niedzielski; A. Goldman and W. Stanley provided welcome help with specimen preparation and care. We gratefully acknowledge support for this project provid-ed by the Negaunee Foundation, Grainger Foundation, and the Field Museum’s Brown Fund for Mammal Research and Ellen Thorne Smith Fund.
LIterAture cItedAlviola, P. A., Duya, M. R. M., Duya, M. V., Heaney, L. R.,
& Rickart, E. A. (2011). Mammalian diversity patterns on Mt. Palali, Caraballo Mountains, Luzon. Fieldiana Life and Earth Sciences, 2, 61-74.
Ambal, R. G. R., Duya, M. V., Cruz, M. A., Coroza, O. G., Vergara, S. G., de Silva, N., Molinyawe, N., & Tabaranza, Jr, B. R. (2012). Key biodiversity areas in the Philippines: priorities for conservation. Journal of Threatened Taxa, 4, 2788-2796.
Balete, D. S., Heaney, L. R., Veluz, M. J., & Rickart, E. A. (2009). Diversity patterns of small mammals in the Zambales Mts., Luzon, Philippines. Mammalian Biology, 74, 456-466.
Balete, D. S., Alviola, P. A., Duya, M. R. M., Duya, M. V., Heaney, L. R., & Rickart, E. A. (2011). The mammals of the Mingan Mountains, Luzon: evidence for a new center of mammalian endemism. Fieldiana Life and Earth Sciences, 2, 75-87.
Balete, D. S., Rickart, E. A., Heaney, L. R., Alviola, P. A., Duya, M. R. M., Duya, M. V., Sosa, T., & Jansa, S. (2012). Archboldomys (Muridae: Murinae) reconsidered: A new genus and three new species of shrew mouse from Luzon Island, Philippines. American Museum Novitates, 3754: 1-60.
Balete, D. S., Rickart, E. A., Rosell-Ambal, R. G. B., Jansa, S. A., & Heaney, L. R. (2007). Descriptions of two new species of Rhynchomys Thomas (Rodentia: Muridae: Murinae), from Luzon Island, Philippines. Journal of Mammalogy, 88, 287-301.
Balete, D. S., Heaney, L. R., & Rickart, E. A. (2013) The mammals of Mt. Irid, Southern Sierra Madre, Luzon Island. National Museum of the Philippines Journal of Natural History, 1, 15-29.
Barcelona, J. F., Pelser, P. B., Balete, D. S., & Co, L. L. (2009). Taxonomy, ecology, and conservation status of
58

The Small Mammals of Mt. Banahaw - San Cristobal National Park, Luzon, Philippines
Philippine Rafflesia (Rafflesiaceae). Blumea, 54, 77-93.Bureau of Mines and Geo-sciences, Ministry of Natural
Resources. (1982). Geology and Mineral Resources of the Philippines. Vol. 1, Geology. 406 pp.
Dans, A. T. L., & Gonzales, J. C. T. (2010). Birds and mammals of Mount Banahaw de Tayabas, Luzon Island, Philippines with notes on their distribution along elevational gradients. Asia Life Sciences Supplement 4, 161-201.
Dickerson, R. E. (1928). Distribution of life in the Philippines. Monograph, Bureau of Science, Manila, 2, 1-322 + 42 pls.
Duya, M. R. M., Duya, M. V., Alviola, P. A., Balete, D. S., & Heaney, L. R. (2007). Report on a Survey of the Mammals of the Sierra Madre Range, Luzon Island, Philippines. Banwa, 4, 41-68.
Duya, M. R. M., Duya, M. V., Alviola, P. A., Balete, D. S., & Heaney, L. R. (2011). Diversity of small mammals in montane and mossy forests on Mount Cetaceo, Cagayan Province, Luzon. Fieldiana Life and Earth Sciences, 2, 88-95.
Geronimo-Catane, S. (1994). Mode of emplacement of two debris-avalanche deposits at Banahao Volcano, Southern Luzon, Philippines. Bulletin of the Volcanological Society of Japan, 39, 113-127.
Hall, R. (1998). The plate tectonics of Cenozoic SE Asia and the distribution of land and sea. In Hall, R. & Holloway, J. D. (Eds.), Biogeography and geological evolution of SE Asia (pp. 99-131). Backhuys Publishers, Leiden.
Hall, R. (2002). Cenozoic geological and plate tectonic evolution of SE Asia and the SW Pacific: computer-based reconstructions, model, and animations. Journal of Asian Earth Sciences, 20, 353-431.
Heaney, L. R. (1986). Biogeography of mammals in Southeast Asia: estimates of rates of colonization, extinction, and speciation. Biological Journal of the Linnean Society, 28, 127-165.
Heaney, L. R. (2001). Small mammal diversity along elevational gradients in the Philippines: an assessment of patterns and hypotheses. Global Ecology and Biogeography, 10, 15-39.
Heaney, L. R., & Rickart, E. A. (1990). Correlations of clades and clines: geographic, elevational, and phylogenetic distribution patterns among Philippine mammals. In G. Peters & R. Hutterer, (Eds.), Vertebrates in the Tropics (pp. 321-332). Museum Alexander Koenig, Bonn.
Heaney, L. R., Balete, D. S., Dolar, M. L., Alcala, A. C., Dans, A. T. L., Gonzales, P. C., Ingle, N. R., Lepiten, M. V., Oliver, W. L. R., Ong, P. S., Rickart, E. A., Tabaranza,
Jr., B. R., & Utzurrum, R. C. B. (1998). A synopsis of the mammalian fauna of the Philippine Islands. Fieldiana Zoology new series, 88, 1-61.
Heaney, L. R., Balete, D. S., Rickart, E. A., Utzurrum, R. C. B., & Gonzales, P. C. (1999). Mammalian diversity on Mt. Isarog, a threatened center of endemism on southern Luzon Island, Philippines. Fieldiana Zoology new series, 95, 1-62.
Heaney, L. R., Balete, D. S., Rickart, E. A., Veluz, M. J., & Jansa, S. (2009). A new genus and species of small “tree mouse” (Rodentia, Muridae) related to the Philippine giant cloud-rats. In R. S. Voss and M. D. Carleton (Eds.), Systematic Mammalogy, Contributions in Honor of Guy G. Musser. Bulletin of the American Museum of Natural History, 331, 205-229.
Heaney, L. R., Dolar, M. L., Balete, D. S., Esselstyn, J. A., Rickart, E. A., & Sedlock, J. L. (2010). Synopsis of Philippine Mammals. http://www.fieldmuseum.org/philippine_mammals/
Heaney, L. R., Balete, D. S., Rickart, E. A., Alviola, P. A., Duya, M. R. M., Duya, M. V., Veluz, M. J., VandeVrede, L., & Steppan, S. (2011). Seven new species and a new subgenus of forest mice (Rodentia: Muridae: Apomys) from Luzon Island. Fieldiana Life and Earth Sciences, 2, 1-60.
Heaney, L. R., Heideman, P. D., Rickart, E. A., Utzurrum, R. B., & Klompen, J. S. H. (1989). Elevational zonation of mammals in the central Philippines. Journal of Tropical Ecology, 5, 259-280.
Heaney, L. R., Tabaranza, Jr., B. R., Balete, D. S., & Rigertas, N. (2006a). Synopsis and biogeography of the mammals of Camiguin Island, Philippines. Fieldiana Zoology new series, 106, 28-48.
Heaney, L. R., Tabaranza Jr., B. R., Balete, D. S., Rickart, E. A., & Ingle, N. R. (2006b). The mammals of Mt. Kitanglad Nature Park, Mindanao, Philippines. Fieldiana: Zoology new series, 112, 1-63.
Heaney, L. R., Walker, E. K., Tabaranza, Jr., B. R., & Ingle N. R. (2000). Mammalian diversity in the Philippines: an assessment of the adequacy of current data. Sylvatrop, 10, 6-27.
Heaney, L. R., Balete, D. S., Rickart, E. A., Veluz, M. J., & Sarmiento, J. (2004). Welcome surprises from Mt. Banahaw. Haring Ibon, 16, 10-16.
Heaney, L. R., Balete, D. S., Alviola, P. A., Duya, M. R. M., & Rickart, E. A. (2013). The mammals of Mt. Anacuao, NE Luzon Island, Philippines: A test of predictions of Luzon small mammal biodiversity patterns. National Museum of the Philippines Journal of Natural History, 1, 1-13.
59

National Museum of the Philippines: Journal of Natural History
Ku, Y., Chen, C., Song, S., Iizuka, Y., & Shen, J. J.. (2009). A 2 Ma record of explosive volcanism in southwestern Luzon: implications for the timing of subducted slab steepening. Geochemistry, Geophysics, Geosystems, 10, doi:10.1029/2009GC002486.
Mallari, N. A. D., & Diesmos, A. C. (1995). Faunal diversity of Mt. Banahaw and implications for conservation priorities. Unpubl. ms, Department of Wildlife Biology, University of the Philippines, Los Baños, Laguna. Pp. 1-28.
Mallari, N. A. D., Tabaranza Jr., B. R., & Crosby, M. J. (2001). Key Conservation Sites in the Philippines. Bookmark Inc., Makati City.
Miranda, F. C. (1978). Altitudinal distribution of birds and mammals based on vegetative cover on Mt. Banahaw - San Cristobal National Park. Unpublished M.Sc. Thesis, University of the Philippines, Los Baños, Laguna. Pp 67.
Mitchell, A. H. G., Hernandez, F., & dela Cruz, A. P. (1986). Cenozoic evolution of the Philippine Archipelago. Journal of Southeast Asian Earth Sciences, 1, 3-22.
Musser, G. G. (1982). Results of the Archbold Expedition. No. 108. The definition of Apomys, a native rat of the Philippine Islands. American Museum Novitates, 2746, 1-43.
Ong, P., Afuang, L. E., & Rosell-Ambal, R. G. (Eds.). (2002). Philippine Biodiversity Conservation Priorities: A Second Iteration of the National Biodiversity Strategy and Action Plan. Philippine Department of the Environment and Natural Resources, Quezon City.
Rickart, E. A. (1993). Diversity patterns of mammals along elevational and disturbance gradients in the Philippines: implications for conservation. Asia Life Sciences 2, 251-260.
Rickart, E. A., Heaney, L. R., Balete, D. S., Alviola, P. A., Duya, M. R. M., Duya, M. V., Rosell-Ambal, G., & Sedlock, J. L. (2013) The mammals of Mt. Natib, Bataan Province, Luzon, Philippines. National Museum of the Philippines Journal of Natural History, 1, 31-44.
Rickart, E. A., Heaney, L. R., & Utzurrum, R. B. (1991). Distribution and ecology of small mammals along an elevational transect in southeastern Luzon, Philippines. Journal of Mammalogy, 72, 458-469.
Rickart, E. A., Heaney, L. R., Heideman, P. D., & Utzurrum, R. C. B. (1993). The distribution and ecology of mammals on Leyte, Biliran, and Maripipi islands, Philippines. Fieldiana Zoology new series, 72, 1-62.
Rickart, E. A., Heaney, L. R., Balete, D. S., & Tabaranza, Jr., B. R. (1998). A review of the genera Crunomys and Archboldomys (Rodentia, Muridae, Murinae) with
descriptions of two new species from the Philippines. Fieldiana Zoology new series, 89, 1-24.
Rickart, E. A., Heaney, L. R., Balete, D. S., & Tabaranza, Jr., B. R. (2011a). Small mammal diversity along an elevational gradient in northern Luzon, Philippines. Mammalian Biology, 76, 12-21.
Rickart, E. A., Balete, D. S., Rowe, R. J., & Heaney, L. R. (2011b). Mammals of the northern Philippines: tolerance for habitat disturbance and resistance to invasive species in an endemic fauna. Diversity and Distributions, 17, 530-541.
Rosell, R. G. B. (1996). Diversity patterns of small non-volant mammals along elevational gradients of Mt. Banahaw, Luzon Island, Philippines. Unpublished M. Sc. Thesis, University of the Philippines, Los Baños, Laguna. Pp. 1-79.
Scheffers, B. R., Corlett, R. T., Diesmos, A., & Laurance, W. F. (2012). Local demand drives a bushmeat industry in a Philippine forest preserve. Tropical Conservation Science, 5, 121-129.
Sedlock, J. L., Ingle, N. R., & Balete, D. S. (2011). Enhanced sampling of bat assemblages: a field test on Mount Banahaw, Luzon. Fieldiana: Life and Earth Sciences, 2, 96-102.
Steppan, S., Zawadski, C., & Heaney, L. R. (2003). Molecular phylogeny of the endemic Philippine rodent Apomys (Muridae) and the dynamics of diversification in an oceanic archipelago. Biological Journal of the Linnean Society, 80, 699-715.
Vogel, T. A., Flood, T. P., Patino, L. C., Wilmot, M. S., Maximo, R. P. R., Arpa, C. B., Arcilla, C. A., & Stimac, J. A.. (2006). Geochemistry of silicic magmas in the Macolod Corridor, SW Luzon, Philippines: evidence of distinct, mantle-derived, crustal sources for silicic magmas. Contributions to Mineral Petrology, 151, 267-281.
Yumul, G., Jr., Dimalanta, C., Queano, K., & Marquez, E. (2009). Philippines, Geology. In R. G. Gillespie & D. A. Clague (Eds.), Encyclopedia of Islands (pp. 732-738). University of California Press, Berkeley.
Zimmer, J. T. (1918). A few rare birds from Luzon and Mindoro. Philippine Journal of Science, 13, 223-233.
60




















