
Plasmodium vivax Sporozoite Challenge in Malaria-Naïve and Semi-Immune Colombian Volunteers
Upload
khangminh22Category
view
4download
0
ava i l ab l e a t www.sc i enced i r ec t . com
C l i n i ca l Immuno logy
www.e l sev i e r . com/ l o ca t e / y c l im
Clinical Immunology (2011) 141, 253–267
REVIEW
The role of naïve T-cells in HIV-1 pathogenesis:An emerging key playerGabriela Khoury a, d, Reena Rajasuriar a, e,Paul U. Cameron a, b, c, d, Sharon R. Lewin a, c, d,⁎
a Department of Medicine, Monash University, 85 Commercial Road, Melbourne Victoria, 3004, Australiab Department of Immunology, Monash University, 85 Commercial Road, Melbourne Victoria, 3004, Australiac Infectious Diseases Unit, Alfred Hospital, 85 Commercial Road, Melbourne, Victoria, 3004, Australiad Centre for Virology, Burnet Institute, 85 Commercial Road Melbourne, Victoria 3004, Australiae Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia
Received 8 July 2011; accepted with revision 8 September 2011Available online 17 September 2011
⁎ Corresponding author at: Infectious9076 2431.
E-mail address: [email protected]
1521-6616/$ - see front matter © 2011doi:10.1016/j.clim.2011.09.002
KEYWORDSNaïve T-cells;HIV-1;IL-7;Immune reconstitution;Antiretroviral therapy;Reservoirs
Abstract Functional naïve T-cells are critical for an effective immune response to multiplepathogens. HIV leads to a significant reduction in CD4+ naïve T-cell number and impaired func-tion and there is incomplete recovery following combination antiretroviral therapy (cART). Herewe review the basic homeostatic mechanisms that maintain naïve CD4+ T-cells and discuss re-cent developments in understanding the impact of HIV infection on naïve CD4+ T-cells. Finallywe review therapeutic interventions in HIV-infected individuals aimed at specifically enhancingrecovery of naïve CD4+ T-cells.© 2011 Elsevier Inc. All rights reserved.
Diseases Unit, Alfred Hospital
.au (S.R. Lewin).
Elsevier Inc. All rights reserve
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2542. Naïve T-cell homeostasis: insights from murine models . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 254
2.1. Homeostatic expansion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2542.2. Homeostatic proliferation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 255
3. Naïve T-cell homeostasis, HIV infection and the impact of cART . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2563.1. Naïve T-cell number . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2563.2. Naïve T-cell function . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 257
, 85 Commercial Road, Melbourne, Victoria, 3004, Australia. Fax: +61 3
d.

254 G. Khoury et al.
3.2.1. Response to IL-7 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2573.2.2. TCR diversity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2573.2.3. Response to neo-antigens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 257
4. HIV infection of naïve T-cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2574.1. Infection of naïve T-cells in vitro . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 257
4.1.1. Role of chemokines . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2584.1.2. Role of IL-7 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2584.1.3. Tissue microenvironment and abortive infection . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2584.1.4. DC-naïve T-cell interactions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 258
4.2. Infection of thymocyte subsets in vitro . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2584.3. Infection of naïve T-cells in vivo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 258
4.3.1. Total and integrated HIV DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2584.3.2. Viral compartmentalization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2594.3.3. Viral co-receptor usage . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 259
5. Therapeutic approaches to enhance naïve T-cell recovery . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2605.1. Exogenous IL-2 therapy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2605.2. Exogenous IL-7 therapy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 260
5.2.1. Animal preclinical studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2605.2.2. Human clinical trials: HIV-infected patients . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2605.2.3. IL-7 Receptor polymorphisms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 260
5.3. Other novel therapeutic approaches . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2616. Outstanding research questions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2617. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261Role of the funding source . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261Conflict of interest statement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261
1. Introduction
HIV infection is characterized by substantial depletion of CD4+T-cells including recent thymic emigrants, naïve T-cells andmemory T-cells. Following control of HIV replication witheffective combination antiretroviral therapy (cART), CD4+T-cells recover to normal levels, in most but not all patients,and impaired CD4+ T-cell recovery has been associated withnon-AIDS events including cardiovascular disease, liver diseaseand malignancy [1–4]. Therefore despite the substantialreduction in morbidity and mortality from cART, life expec-tancy has still not returned to normal [5,6]. A detailed under-standing of naïve CD4 T-cell homeostasis is required in order todevelop novel strategies to enhance the quality of immune re-constitution following cART.
2. Naïve T-cell homeostasis: insights frommurinemodels
Naïve T-cells are characterized by the expression of surfacemarkers CD45RA, CD27, CD28, CD62L, CCR7 and the IL-7receptor [7,8]. Naïve T-cells exit the thymus following matu-ration and are enriched for T-cell receptor excision circles(TREC) and express the surface marker platelet endothelialcell adhesion molecule-1 (PECAM-1), also known as CD31 [9].Naïve T-cells circulate between the blood and the lymphoidtissue driven by cell surface markers CD62L and CCR7 [10].The number of naïve T-cells in blood remains relativelyconstant throughout adult life despite continuous stimulationwith foreign antigens and a dramatic reduction in thymic out-put with age. Maintenance of naïve T-cells in adults is largelydependent on homeostatic proliferation which leads to a
decrease in CD31 expression and TREC concentration result-ing in an increase in the proportion of CD31- naïve T-cellswith aging [9,11]. The mechanisms which drive homeostaticproliferation of naïve T-cells in humans are not fully under-stood, however extensive studies in murine models havedemonstrated multiple factors are involved in both post-thymic survival and proliferation of naïve T-cells. Exposureto the cytokine IL-7 and contact with major histocompati-bility complex (MHC) molecules presenting self-peptidesthrough the T-cell receptor (TCR) within secondary lym-phoid tissue are both critical for naïve T-cell homeostasis[12–15]. (See Fig. 1).
2.1. Homeostatic expansion
In lymphopenic murine models, adoptive transfer of naïve T-cells results in a biphasic increase in naïve T-cells with bothrapid and slow proliferation phases. Rapid proliferation (or ho-meostatic expansion) was associated with conversion of naïveT-cells into an effector/memory phenotype [16,17]. There arediverse views on whether IL-7 is required for homeostatic ex-pansion which is thought to be dependent on the strength ofbinding between antigen and the TCR. Proliferative responsesfollowing low-affinity binding, as seen with self-peptides, isthought to be more dependent on IL-7 [12,18] compared tohigh-affinity cognate antigen binding (reviewed in [19]).
Naïve T-cell homeostatic expansion has also been shown tobe associated with exposure to microbial products. Followingadoptive transfer of T-cells from immunocompetent BALB/cThy1.1 mice to mice with severe combined immunodeficien-cy (SCID) that have been raised in a germ-free environment,there was little naïve T-cell proliferation, activation or
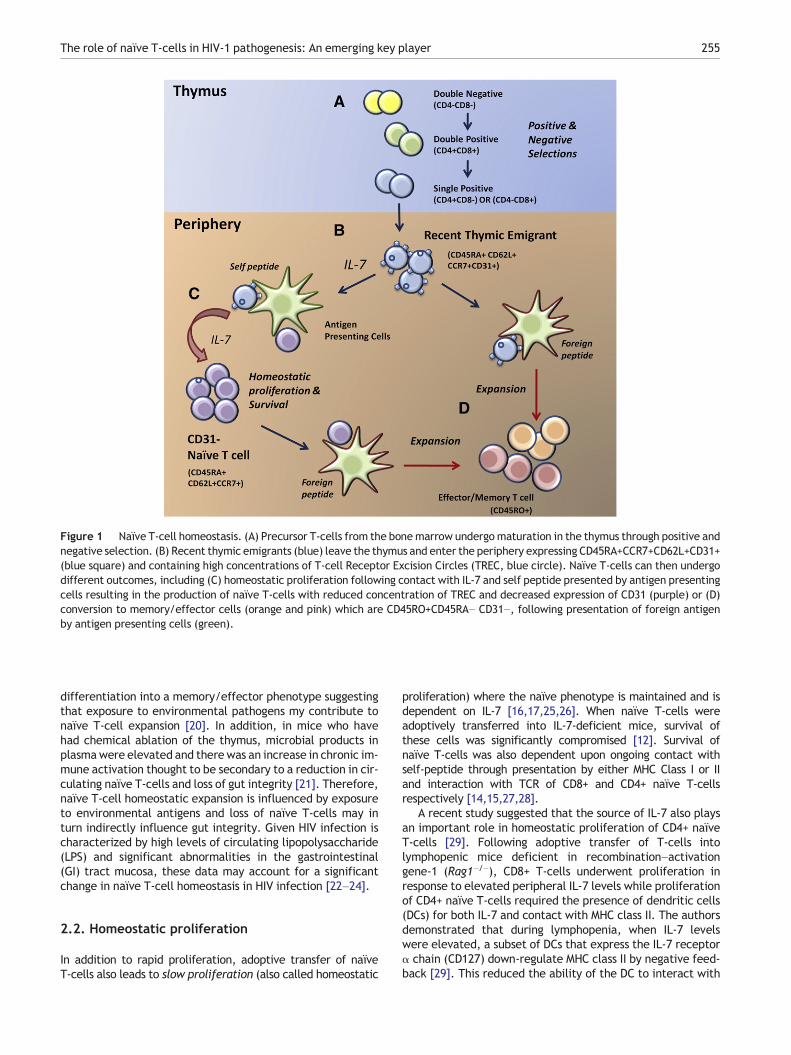
Figure 1 Naïve T-cell homeostasis. (A) Precursor T-cells from the bonemarrow undergomaturation in the thymus through positive andnegative selection. (B) Recent thymic emigrants (blue) leave the thymus and enter the periphery expressing CD45RA+CCR7+CD62L+CD31+(blue square) and containing high concentrations of T-cell Receptor Excision Circles (TREC, blue circle). Naïve T-cells can then undergodifferent outcomes, including (C) homeostatic proliferation following contact with IL-7 and self peptide presented by antigen presentingcells resulting in the production of naïve T-cells with reduced concentration of TREC and decreased expression of CD31 (purple) or (D)conversion to memory/effector cells (orange and pink) which are CD45RO+CD45RA− CD31−, following presentation of foreign antigenby antigen presenting cells (green).
The role of naïve T-cells in HIV-1 pathogenesis: An emerging key player 255
differentiation into a memory/effector phenotype suggestingthat exposure to environmental pathogens my contribute tonaïve T-cell expansion [20]. In addition, in mice who havehad chemical ablation of the thymus, microbial products inplasmawere elevated and there was an increase in chronic im-mune activation thought to be secondary to a reduction in cir-culating naïve T-cells and loss of gut integrity [21]. Therefore,naïve T-cell homeostatic expansion is influenced by exposureto environmental antigens and loss of naïve T-cells may inturn indirectly influence gut integrity. Given HIV infection ischaracterized by high levels of circulating lipopolysaccharide(LPS) and significant abnormalities in the gastrointestinal(GI) tract mucosa, these data may account for a significantchange in naïve T-cell homeostasis in HIV infection [22–24].
2.2. Homeostatic proliferation
In addition to rapid proliferation, adoptive transfer of naïveT-cells also leads to slow proliferation (also called homeostatic
proliferation) where the naïve phenotype is maintained and isdependent on IL-7 [16,17,25,26]. When naïve T-cells wereadoptively transferred into IL-7-deficient mice, survival ofthese cells was significantly compromised [12]. Survival ofnaïve T-cells was also dependent upon ongoing contact withself-peptide through presentation by either MHC Class I or IIand interaction with TCR of CD8+ and CD4+ naïve T-cellsrespectively [14,15,27,28].
A recent study suggested that the source of IL-7 also playsan important role in homeostatic proliferation of CD4+ naïveT-cells [29]. Following adoptive transfer of T-cells intolymphopenic mice deficient in recombination–activationgene-1 (Rag1−/−), CD8+ T-cells underwent proliferation inresponse to elevated peripheral IL-7 levels while proliferationof CD4+ naïve T-cells required the presence of dendritic cells(DCs) for both IL-7 and contact with MHC class II. The authorsdemonstrated that during lymphopenia, when IL-7 levelswere elevated, a subset of DCs that express the IL-7 receptorα chain (CD127) down-regulate MHC class II by negative feed-back [29]. This reduced the ability of the DC to interact with

256 G. Khoury et al.
the CD4+ naïve T-cell and therefore prevented proliferation. Itis unknown whether similar mechanisms involving CD127-pos-itive DCs exist in humans.
3. Naïve T-cell homeostasis, HIV infection and theimpact of cART
3.1. Naïve T-cell number
Following HIV infection there is a significant decline in numberand function of naïve CD4+ T-cells in the blood and lymphnode compared to healthy individuals [30–34]. (summarized inTable 1) The reduction in naïve T-cells is likely multi-factorialand secondary to reduced thymic function, increased naïve T-cell proliferation, enhanced immune activation and direct HIVinfection [30–32,35]. Collagen deposition in the paracorticalT-cell zones of the lymphoid tissue also plays a critical role inlimiting naïve T-cell homeostasis [36,37]. Recent work in bothSIV-infected macaques and HIV-infected humans demonstratedthat following SIV/HIV infection, increased production of TGF-βin lymphoid tissue resulted in procollagen production and depo-sition of collagen fibrils [38]. This resulted in restricted T-cellaccess to the fibroblastic reticular cell (FRC) network which isthemajor source of the survival factor IL7, resulting in apoptosisand depletion of naïve CD4+ T-cells. The loss of naïve T-cells inturn, removed a major source of lymphotoxin-β, a survival fac-tor for FRCs [38]. The resulting loss of FRCs and the loss of IL-7produced by FRCs may thus perpetuate a vicious cycle of
Table 1 Naïve T-cell abnormalities in the setting of HIV infectio
Compartment assessed Abnormality in naïve T-cells in H
Blood↓ Numbers of naïve T-cells and R↑ CMV responses associated with↑ Expansion proliferation (Ki67)↓ Cell cycle entry and proliferati↓ Co-stimulatory response (CD27↑ Signaling (Basal STAT5 phospho↓ Survival (↓ Bcl-2)↑ Phosphorylated STAT5 in cytop↓ Phosphorylated STAT5 nuclear
Lymph node↓ Number of naïve T-cells (TREC↑ Number of naïve T-cells (CD45RCollagen deposition in•Paracortical T zones•High endothelial venules•Fibroblastic reticular cell netw
↑ Distance between IL-7 source a↓ Access to IL-7↑ Naïve T-cell apoptosis (TUNEL+
Gastro-intestinal tract↓ CD4+ T cells (naïve, effector a↑ Microbial translocation (↑ LPS,↑ Fibrosis
depletion of naïve CD4+ T-cells and the FRC network in the set-ting of untreated HIV infection.
HIV infection leads to significant depletion of total muco-sal CD4+ T-cells in the GI tract which allow for migration ofmicrobial products into the systemic circulation [22,24]. Mi-crobial products, specifically LPS, can drive DC activation vialigation of toll like receptors (TLR) leading to elevated pro-inflammatory cytokines and enhanced T-cell proliferation[22,39,40]. Microbial products may also be responsible forenhanced naïve T-cell proliferation as demonstrated inmouse models but not yet confirmed in human studies[20,21]. More recently, the contribution of cytomegalovirus(CMV) co-infection to naïve T-cell proliferation and deple-tion has been explored. HIV-infected patients with strongCMV-specific T-cell responses had fewer naïve T-cells [41].The authors suggested that the loss of naïve T cells couldbe due to movement of naïve T-cells into the large pool ofCMV-specific memory T-cells leading to an adverse effecton total CD4+ T-cell immune recovery following cART.
Following initiation of cART, naïve T-cell proliferation de-creases; thymic output increases and the total number ofnaïve T-cells significantly increases [30,32,33,35]. However,the numbers of naïve T-cells rarely reach normal levels inadults even after more than 7 years of cART [42]. In contrast,in HIV-infected children, naïve T-cells reached normal levelsfollowing cART, and recovery of thymic output was indepen-dent of age and time of initiation of cART [43]. A recentstudy showed that naïve T-cell count at initiation of cARTcould predict successful immune reconstitution [44]. In thiscase, immune reconstitution was defined as an increase of
n.
IV (measurement) Reference
TE (CD45RA+, CD45RA+ CD62L+, TREC) [30–32,34]reduced number of naïve T-cells [41]
[31]on (Ki67, CFSE) [48,49]+ CD28) [49]rylation) [50,52]
[50]lasm [52]localisation [52]
) [32]A+ CD62L+, TREC) [34]
[37][36]
ork [38]nd naïve T-cells (CD45RA+) [38]
[38]) [38]
nd memory) [23,137,138]16s rDNA) [22,139]
[140]

The role of naïve T-cells in HIV-1 pathogenesis: An emerging key player 257
100–200 CD4+ T-cells after 2 years on therapy. There is a needto further understand why recovery of naïve T-cells is im-paired following cART. The role of age, nadir CD4 T-cellcount or other biological factors that may enhance lymphnode fibrosis or impair thymic output remains to be furtherexplored.
3.2. Naïve T-cell function
Abnormalities in naive.
3.2.1. Response to IL-7The dramatic depletion of CD4+ T-cells in HIV infection is alsoassociated with significant functional defects in naïve T-cells(summarised in Table 1). HIV infection is associated with in-creased systemic levels of IL-7, low surface expression of theα chain of the IL-7 receptor (lL7Rα), CD127, and impaired re-sponse to IL-7 [45–47]. Naïve T-cells from HIV-infected indi-viduals had a reduced capacity to expand followingstimulation with TCR ligation and IL-7 [48]. In addition,CD31+ naïve T-cells, a subset of naïve T-cells that have re-cently left the thymus and not undergone proliferation[9,11], failed to express the co-stimulatory molecules CD28and CD27 after TCR stimulation [49]. Following TCR engage-ment of CD31- naïve T-cells, there was an increase in co-stim-ulatory molecules but the cells were unable to enter cell cycle[49]. These data demonstrate discreet functional defects inresponse to IL-7 and TCR engagement in the setting of HIV in-fection, in addition to reduced total number of naïve T-cells.
IL-7 signaling which occurs through the STAT5 pathway hasalso been shown to be altered during HIV infection. The basallevel of STAT5 phosphorylation was elevated in both naïve andmemory CD4+ T-cell subsets in patients receiving cART, whowere viremic and aviremic [50]. Expression of CD127 onnaïve T-cells and the ability of IL-7 to bind to the IL-7R onnaïve T-cells in patients on cART were similar to healthycontrols [50]. Recent analysis of signaling pathways in CD4+T-cells from viremic and aviremic patients also suggestedthat naïve CD4+ T-cells, in contrast to memory CD4+ T-cells,were able to sustain CD127-STAT5 signaling [51]. However,despite normal IL7 binding and sustained IL7R signaling inHIV-infected patients, the expression of the survival proteinBcl-2 was reduced in naïve T-cells from both viremic andcART-treated patients, consistent with a post-receptor blockthat prevented the induction of downstream pro-survivalsignals [50]. A study of viremic patients attributed this out-come to an increased accumulation of phosphorylated STAT5within the cytoplasm due to the inability to enter the nucleus[52]. The multiple defects in signaling pathways important forsurvival of naïve and memory T-cells could therefore play animportant role in limiting the capacity for CD4+ T-cell recov-ery following cART.
Following long-term cART in HIV-infected adults [53], andchildren [54], the concentration of IL-7 reduces to normallevels. One multivariate analysis showed that an increase inthe mean CD4+ T-cell count in patients was associated withincreased IL-7 responsiveness and this was independent ofT-cell activation, PD-1 expression, CCR5 expression and IL-7plasma levels [55]. In addition, patients on suppressivecART with a complete immunological response (≥500 CD4+T-cells/mm3 ≥5 years after initiation of ART), demonstrated
higher functional response to IL-7 (measured by STAT5 phos-phorylation), which correlated with increased total andnaïve T-cell levels compared to patients with incompleteresponses (CD4 b500 cells/μl) [55]. Other studies have shownthat defects in IL-7 responsiveness were only partiallycorrected with cART while levels of IL-7 remained elevatedand IL-7 receptor signaling defects were still present in manypatients following cART [50,56–58].
3.2.2. TCR diversityTCR diversity is also altered in patient with HIV infection. TCRdiversity is quantified by the length of the hypervariable CDR3Vβ regions of the TCR [59–62]. In healthy individuals the mul-tiple CDR3 lengths follow a Gaussian distribution but in HIV in-fection this distribution is skewed and is associated withdisease progression and accelerated loss of total CD4+ T-cellsin both adults and children [59–62]. Translocation of microbialproducts as well as the HIV glycoprotein (gp)160 which can in-duce superantigen responses in specific Vβ groups can alsolead to a skewed TCR repertoire [63]. In individuals whohave a poor immunological response to cART (b20% increasein CD4+ T-cells from baseline or do not reach 200 cells/μl)there was an increased expansion in some Vβ groups and thiswas associated with reduced naïve T-cell number and thymicoutput [58]. Two longitudinal studies have shown that despitean increase in total CD4+ T-cell count on long-term cART,CD4+ T-cell subsets remain skewed [64,65].
3.2.3. Response to neo-antigensA reduction in functional responses to new antigens and vacci-nations has also been associated with late initiation of cARTand was most likely due to skewing of the proportion of CD4+T-cell populations and depletion of naïve T-cells [66]. Otherstudies have shown that despite increases in CD4+ T-cells tolevels >500 CD4+ T-cells with long-term cART, responses toneoantigens through vaccinations were lower in HIV-infectedindividuals compared to healthy controls [67]. Lack of cellularresponse to neoantigens has also been observed in childrenand adolescents vaccinated during their first year on cART[68]. One possible explanation for these findings is a defectin naïve T-cell proliferation in response to neoantigen [69].The implications of long-term impaired naïve T-cell recoveryare unclear but it is highly likely that impaired naïve T-cellnumber and functionwill contribute to ongoing immune dysre-gulation in patients on cART.
4. HIV infection of naïve T-cells
One potential mechanism for depletion and impaired functionof naïve T-cells is that HIV can directly infect naïve T-cellsboth in vivo and in vitro. However the extent of infection,the impact of HIV infection on function and the effect ofcART on the reservoir of infected naïve T-cells is less clear.
4.1. Infection of naïve T-cells in vitro
Naïve CD4+ T-cells are relatively resistant to HIV infection invitro but in some conditions, direct infection can occur[70–72]. Infection of naïve T-cells with both CXCR4-using (X4)and CCR5-using (R5) viruses can lead to integration but the effi-ciency of infection of naïve CD4+ T-cells was significantly lower

258 G. Khoury et al.
than in resting memory CD4+ T-cells [70]. In vitro infection ofnaïve T-cells was associated with accumulation of partial or in-complete viral transcripts and delayed viral integration but themajor block to direct infection of naïve T-cells appears to be de-creased efficiency of viral fusion [70,73]. Viral fusion in naïve T-cells can be significantly enhanced in viruses that co-express ve-sicular stomatitis virus (VSV) and R5 envelope protein, but fu-sion was inefficient with VSV or R5 env protein alone [71]. Themolecular mechanisms for this observation are currentlyunclear.
4.1.1. Role of chemokinesOur laboratory recently demonstrated that direct infection ofresting CD4+ T-cells can be established following incubationwith chemokines that bind to chemokine receptors highlyexpressed on the surface of resting T-cells, including CCL19and CCL21 which both bind to CCR7 which is highly expressedon both naïve and memory T-cells [74,75]. However, thesechemokine mediated changes only facilitated infection ofresting memory and not naïve T-cells, [75]. Taken togetherthese data demonstrate that there are clear differences inthe ability of HIV to infect resting naïve and memory T-cellsand that the major block to infection of naïve T-cells occursearly in the viral life cycle, most likely at the level of viralfusion.
4.1.2. Role of IL-7The role of IL-7 in enhancing infection of naïve T-cells has alsobeen explored [76–79]. Naïve T-cells become more suscepti-ble to infection following culture with IL-7 [79]. Followingtreatment with IL-7, cord blood naïve T-cells and adult mem-ory T-cells, but not adult naïve T-cells, enter G1b phase ofcell cycle and become susceptible to infection with bothVSV-pseudotyped HIV and wild type HIV [76–78]. Total restingCD4+ T-cells treated with IL-7 together with the phosphatidy-linositol 3-kinase (PI3K) inhibitor, wortmannin, led to nochange in HIV infection suggesting that pathways other thanPI3K may be involved [78].
4.1.3. Tissue microenvironment and abortive infectionThe microenvironment may also have an important role insusceptibility of naïve T-cells to HIV infection. HIV infectionof resting naïve CD4+ T-cells was significantly enhancedwhen infection was performed using tonsil lymphoid tissueex vivo [80,81]. In contrast, when naive CD4+ T-cells werepurified from tonsil and cultured alone in vitro, HIV infectiondid not occur [80]. Differences between susceptibility to HIVinfection of naïve T-cells from lymphoid tissue and those iso-lated from the blood may be due to differences in the ex-pression of active and inactive forms of the cellular proteinAPOBEC3G [82].
When human lymphoid aggregated cultures (HLACs) fromtonsil were infected with X4-using virus, incomplete reversetranscription products accumulated within resting CD4+T-cells [72]. These incomplete transcripts triggered pro-apoptotic and pro-inflammatory cellular responses resultingin the death of resting CD4+ T-cells and may potentiallyexplain the significant CD4+ T-cell loss in HIV infection [72].These experiments however didn't distinguish betweenmemo-ry and naïve T-cells, so it is currently unknown if this occurs inboth T-cell subsets [72]. Enhanced apoptosis of resting CD4+T-cells isolated from the blood and infected in vitro has also
been described [83]. This newly describedmechanism of accu-mulation of incomplete reverse transcripts and abortive infec-tion could be an important contributing factor to naïve CD4+T-cell depletion in HIV-infected patients.
4.1.4. DC-naïve T-cell interactionsDCs located within the lymphoid tissue may play a role in en-hancing infection of naïve T-cells. One group has demonstrat-ed that in vitro co-culture of naïve T-cells with DCs cansignificantly enhance infection of naïve CD4+ T-cells [84],while others have shown no effect of DCs [70]. A study usingmonocyte-derived DCs (MDDCs) explored the efficiency ofMDDCs to transmit HIV to different T-cell subsets includingnaïve, central and effector memory subsets [84], The studyshowed that MDDC-mediated transmission of virus was depen-dent on virus co-receptor usage and co-receptor expression onthe target T-cell. MDDCs were only able to transfer X4 strainsto CD4+ naïve T-cells [84]. In other experiments, when restingT-cells were infected in the presence of DCs and then sortedinto naïve and memory cells, infection of naïve T-cells wasnot enhanced by the presence of DCs [70]. Other groups havedemonstrated that physical contact of total resting CD4+ T-cells with uninfected autologous immature-MDDCs increasedinfection of resting CD4+ T-cells [85]. This was not simply a re-sult of a mixed lymphocytic reaction as, an increase in activa-tion markers was only observed when a higher ratio of DC:T-cell was used [85].
Combined, these studies demonstrate that HIV infection ofnaïve T-cells requires additional signals, mediated by eithercytokines such as IL-7 or cell-cell contact via DC-T-cell con-tact, and that these signals are more likely to be present with-in the endogenous microenvironment of lymphoid tissue.
4.2. Infection of thymocyte subsets in vitro
HIV infection of thymocyte subsets could be themain source ofHIV-infected naïve T-cells in the periphery. All thymocytepopulations predominately express high levels of CXCR4 andlow levels of CCR5 making these cells more susceptible to in-fection with X4-using viruses [86,87]. However, HIV is able toenter thymocytes and undergo reverse transcription after in-fection with either X4 or R5-using viruses [88]. Our lab hasdemonstrated that thymic plasmacytoid dendritic cells playan important role in the infection of thymocytes throughtransfer of R5-using virus to both immature andmature thymo-cytes [89]. Various cytokines have differential effects on thy-mocyte infection. IL-7 alone or in combination with IL-4increased expression of CXCR4 and enhanced infection withX4-using viruses [87,90]. Thymocytes cultured with IL-2 andIL-4 expanded, the expression of both co-receptors increasedand thymocytes were more permissive to X4 and R5-using vi-ruses [90]. In addition, mature thymocytes cultured withTNF-α and IL-7weremore likely to become infected as a resultof IL-7 enhancing cell survival of mature cells [91].
4.3. Infection of naïve T-cells in vivo
4.3.1. Total and integrated HIV DNAIn contrast to findings in vitro, naïve T-cells are clearly infectedwith HIV in vivo, although at low levels compared with other T-cell subsets such as CD4+ memory T-cells [92–100];
The role of naïve T-cells in HIV-1 pathogenesis: An emerging key player 259
(summarized in Table 2). Accurate quantification of HIV in naïveT-cells in vivo can be complicated by the surface markers usedto define naïve T-cells. If insufficient surface markers are usedto define a naïve T-cells (b3 specific markers) [7] then it is pos-sible that memory T-cell contamination may be contributing tothe assessment of naïve T-cells [101–103]. In addition, manystudies only assess total HIV DNA rather than integrated DNAwhich again may provide an over estimation of the number ofinfected naïve T-cells [92–97,99,100]. However, to furtherstrengthen the case that naïve T-cells are indeed infected invivo, replication competent virus has been detected in naïveT-cells from patients receiving cART [93,94,98,99].
There is conflicting data on whether specific naïve T-cellsubsets are preferentially infected. In one study, there was asignificantly higher concentration of HIV DNA in CD31- naïveT-cells compared with the CD31+ naive T-cells [92]. Thesefindings suggested that infection of naïve T-cells did notoccur in the thymus; but that infection potentially occurredfollowing naïve T-cell homeostatic proliferation in the pe-riphery. However, the samples in this study were fixedprior to sorting and this could have had an effect on the ef-ficiency of HIV DNA detection [92]. In a more recent longitu-dinal study of patients initiating cART, our group showedthat both CD31+ and CD31- naïve T-cell subsets wereinfected at similar frequency [95]. In addition we showedthat the contribution of naïve T-cells to the total reservoirincreased following cART [95]. Overall, even though the ab-solute number of infected naïve T-cells in patients on cART issignificantly smaller than infected memory T-cells, naïve T-cells expand significantly following cART and therefore mayrepresent a significant persistent reservoir in patients oncART.
Table 2 Detection of HIV infection of naïve T-cells in vivo. Studiepatients who are either on or off cART (upper rows without shading)
[92]
Reference
[93]
[94][95]
[96]
[97][98]
[99][100]
Patient Number (naïve/cART)
22 (11/11)
13 (13/0)
11 (4/7)10 (10/10)
26 (11/15)
102 (0/102)34 (0/34)
11 (0/11)3 (0/3)
CD4+ cells/µl(Median)
526
610
547143b
561
271581
582305
Plasma HIV RNA copies/ml (Median)
15,989
17,090
1,08565,150b
36,541(viremic)<40 (aviremic)
6,908<50
<20<20
Phenotype naïve T-cell
CD11a10
CD45RO—CDCD27+ CD31CD27+CD45RCD11a+CD57CD45RA+CDCD45RO—CDCD31+/—CD45RA+CCCD31+/—
CD45RA+CDCD45RA+ CCCD27+CD45RA+CDCD45RA+CD
–Not assessed.a Phylogenetic analysis.b Baseline sample at the beginning of longitudinal study.
Amore recent cross-sectional study looking at both viremicand aviremic patients showed that both CD31+ and CD31-naïve T-cell subsets were infected with HIV at similar frequen-cies and in some patients, infected naïve T-cells made a sub-stantial contribution to the overall pool of infected T-cells[96]. This study also showed that in CD31+ naïve T-cells fromboth viremic and aviremic patients, the concentration of HIVDNA positively correlated with IL-7 plasma levels, suggestinga possible function of IL-7 in the infection of naïve T-cells.
4.3.2. Viral compartmentalizationIn order to better understand the origin of infection of naïveT-cells, several groups have examined the phylogenetic rela-tionship between virus isolated from naïve T-cell subsets,memory cells and plasma. We recently showed that therewas no compartmentalisation of virus envelope sequences inCD31+ and CD31- naïve and memory T-cells subsets from pa-tients prior to and following cART [95]. Similar findings havebeen described in another study using sorted samples from pa-tients naïve to cART [93]. In a smaller cohort of aviremic pa-tients on cART for seven years, compartmentalization ofvirus in naïve T-cells was observed in two of the three patientsstudied [100]. In order to better understand the origin of infec-tion of naïve T-cells, these studies should be performed inlarger prospective studies of patients on long term cART.
4.3.3. Viral co-receptor usageBoth R5-using and X4 using virus has been isolated from naïve T-cells from HIV-infected patients [93–95,100]. In all of thesestudies naïve T-cells were predominantly infected with R5-using virus. However, two of the studies did detect some X4-using viruses within the naïve T-cell compartment [94,100]. It
s that have assessed HIV infection of naïve T-cells in HIV-infectedor on cART (lower rows, gray shading) are shown.
s
57—+/—O——
62L+28+
R7+
62L+R7+
62L+62L+
Total HIV DNA
Yes (CD31 neg >CD31pos)
Yes (total)
Yes (total)Yes (CD31+ = CD31 neg)Yes (CD31+ = CD31 neg)
Yes (total)—
Yes (total)Yes (total)
Integrated HIV DNA
—
—
Yes—
Yes CD31+ (10 cART—patients) —Yes
——
Replication competentvirus
—
Yes
Yes—
—
—Yes
Yes—
Compartment-alisationa
—
No
—No
—
——
—Yes (2/3)

260 G. Khoury et al.
still remains unclear how naïve T-cells become infected withR5-using viruses given the low expression of CCR5 on the surfaceof naïve T-cells. We have recently demonstrated in vitro thatthymic plasmacytoid DC are able to efficiently transfer R5-using virus to both immature and mature thymocytes providinga potential mechanism to explain R5-using infection of naïveT-cells [89].
5. Therapeutic approaches to enhance naïveT-cell recovery
In addition to cART, there is now increasing interest to de-velop immunotherapeutic agents that specifically enhanceCD4+ T-cell recovery, including naïve T-cells.
5.1. Exogenous IL-2 therapy
Administration of IL-2 to lymphopenic mice increased totalCD4+ T-cells including naïve, resting memory and activatedCD4+ T-cells [104]. Similar responses were seen in HIV-infected patients on cART who received IL-2 [105–107].Both thymic output and peripheral homeostatic proliferationcontributed to the increase in naïve CD4+ T-cells [104–109].In patients treated with cART and IL-2 there was a reductionin apoptosis of total and naïve CD4+ T-cells [106,110].
Two large, randomized international clinical trials of IL-2in patients on cART, Evaluation of Subcutaneous Proleukin ina Randomized International Trial (ESPRIT) [111] and Subcu-taneous, Recombinant Human Interleukin-2 in HIV-InfectedPatients With Low CD4+ Counts Receiving Active Antiretrovi-ral Therapy (SILCAAT) [112] showed no additional clinicalbenefit following administration of IL-2 to patients receivingcART. These studies demonstrated that IL-2 led to a clear in-crease in total CD4+ T-cell counts, including naïve CD4+ T-cells, but this did not lead to a reduction in opportunisticinfections or mortality, showing that IL-2 therapy had noadditional clinical benefit for patients on cART [111–113].Similar findings were observed in the Study of ALdesleukinWith and without AntiRetroviral Therapy, (STALWART),which tested intermittent IL-2 combined with peri-cyclecART in HIV-infected individuals who were cART-naïve andasymptomatic [114]. These studies demonstrated that anincrease in CD4+ T-cell number secondary to IL-2 did nottranslate to effective function and improved clinical out-comes and further work is needed to understand the biolog-ical basis for these findings.
5.2. Exogenous IL-7 therapy
5.2.1. Animal preclinical studiesAdministration of IL-7 to SIV-infected and uninfected cynomo-logus monkeys, baboons and macaques, induced proliferationof naïve CD4+ T-cells and a decline in the frequency of TRECconsistent with enhanced homeostatic proliferation [115–119]. Administration of IL-7 induced migration of T-cells tothe lymph node where they entered cell cycle [120] and great-er TCR diversity was observed consistent with enhanced thy-mic output [119]. Importantly, administration of IL-7 to SIV-infected macaques resulted in no increase in SIV viral load[116,117].
5.2.2. Human clinical trials: HIV-infected patientsThere have been two completed trials of recombinant human(rh) IL-7 in cART-treatedHIV-infected patients. In one study, pa-tients with CD4+ T-cell counts 100–400 cells/μl and HIV RNAb50 copies/ml had eight subcutaneous injections three timesper week over a 16 day period and two doses of rhIL-7 were ex-amined [121]. There was a preferential increase in both naïveand central memory T-cells following rhIL-7. Increases inCD4+ naïve T-cells peaked at day 14 and started to decline14 days after completion of rhIL-7. At the higher dose,four patients also experienced a transient increase in plas-ma HIV RNA [121].
The second trial was performed in HIV-infected patients withCD4+ T-cell counts >100 cells/μl, in patients with a viral load ofb50 or 50–50,000 copies/ml HIV RNA [122]. A transient increasein HIV RNAwas demonstrated in 6 out of 11 patients who hadHIVRNA b50 copies/ml at baseline. On day 1 following rhIL-7, therewas a significant decrease in circulating CD4+ and CD8+ T-cells.By day 14, the levels had increased but mainly consisted of cen-tral memory T-cells, with minimal increase in naïve T-cells. Thenumber of proliferating naïve T-cells increased compared tobaseline, however these were mainly CD8+ T-cells [122].
Aside from the effects of IL-7 on CD4+ T-cell number, IL-7can also have an impact on latently infected cells and possi-bly co-receptor usage of virus in vivo. In both clinical trials ofIL-7, some patients experienced a transient increase in plas-ma HIV RNA at the time of peak T-cell proliferation, whicheventually returned to b50 copies/ml [121,122]. There wasno significant increase in the concentration of cell-associatedHIV DNA in either PBMC or CD4+ T-cells following IL-7 [121].However, this study did not correct for the overall increasein total T-cell number following rhIL-7, meaning that the abso-lute number of infected cells (expressed as HIV copies/mlblood) may have increased as a result of proliferation of la-tently infected naïve, central memory or transitional memoryT-cells, as has recently been demonstrated ex vivo [98].Therefore, it is possible that IL-7 may induce proliferation ofboth uninfected and latently infected naïve and memory T-cells and it remains unclear if IL-7 will expand or activate la-tently infected cells. Clinical trials are currently underway totest this (ERAMUNE; www.clinicaltrials.gov).
IL-7 may also potentially lead to a change in HIV co-recep-tor usage. Following long-term culture of HIV-infected PBMCwith IL-7 in vitro, there was an increase in the expression ofCXCR4 on CD4+ T-cells, which led to a change in co-receptorusage from R5-using to X4-using virus [123]. The same groupalso suggested that increases in CXCR4 expression on CD4+ T-cells from HIV-infected patients could lead to the emergenceof R4-using viruses [124]. Using IL-7 as an immunotherapycould potentially have the same effect in vivo although todate this has not been demonstrated.
5.2.3. IL-7 Receptor polymorphismsIt is possible that response to endogenous and exogenous rhIL-7 is dependent upon differences in the IL-7R which are deter-mined by genetic polymorphisms. The IL-7R is composed of anα and common γ chain [125]. Four haplotypes of the gene cod-ing for IL-7Rα (CD127) have been described in Caucasians[126]. These haplotypes are defined by single nucleotide poly-morphisms located throughout the IL-7Rα gene and lead to al-ternative splicing ofmRNA resulting in varying ratios of solubleand membrane bound CD127 [127–129]. Using a multivariate

Table 3 Outstanding research questions on naïve CD4+ T-cells and HIV pathogenesis.
Outstanding research questions
1. Are naïve T-cells infected in all patients or only a selectproportion?
2. At what stage of disease do naïve T-cells become infected?3. Are naïve T-cells preferentially infectedwith X4-using or R5-using
viruses?4. Does viral compartmentalization occur in naïve T-cell subsets?5. How do R5-using viruses infect naïve T-cells?6. How does self-peptide and microbial products drive naïve T-
cell proliferation in humans?7. Which surrogate markers of immune function should be used
in future clinical trials of immune based therapies?8. What genetic factors contribute to CD4 T-cell immune re-
constitution following cART?
The role of naïve T-cells in HIV-1 pathogenesis: An emerging key player 261
analysis, we have recently shown that the IL-7Rα haplotype 2was significantly associated with more rapid recovery of CD4+T-cells >500 cells/μl following cART [53]. In addition, solubleCD127 (sCD127) levels in cART-treated HIV-infected patientswere significantly lower in individuals who were homozygousfor haplotype 2 than non-haplotype 2 carriers. A potential ex-planation for these findings is that high levels of sCD127“mops” up circulating IL-7 allowing for less efficient IL-7-related cell signaling and impaired CD4+ T-cell recovery. Eval-uation of these and other polymorphisms should be performedin other patient cohorts. In addition these polymorphisms maypotentially be important in determining how effectively a pa-tient responds to exogenous IL-7, although these studies havenot been performed to date.
5.3. Other novel therapeutic approaches
Other immunotherapies like Growth Hormone (GH), Keratino-cyte Growth Factor (KGF), and androgen blockade which areknown to improve thymopoiesis and thymic architecture regen-eration are also being considered for evaluation in HIV-infectedpatients. Other approaches could also include anti-fibroticagents that could potentially reduce lymph node fibrosis [38].
A randomized control trial of GH in a cohort of 46 HIV-infected patients receiving cART showed promising resultswith significant increases in thymopoiesis as measured byTREC concentration [130]. Studies in macaques have shownthat KGF targets epithelial cells, improves both naïve T-cellnumbers and TCR diversity [131] and also activates the pro-duction of IL-7 within the thymus [132]. At puberty when sexsteroid levels increase, the thymus decreases in size and thy-mic output significantly reduces. Therefore androgen block-ade may improve thymic output of HIV patients as shownfollowing castration in mouse models [133]. In addition, hypo-gonadal men have been shown to have an increased number ofRTE and following treatment with androgens, the levels of RTEdecline [134]. Sex steroid blockade using luteinizing-hormonereleasing hormone antagonists is currently used in the treat-ment of prostate cancer [133] and has also been trialed in pa-tients undergoing stem cell transplantations [135]. In both ofthese scenarios, thymic function and production of naïveT-cells were enhanced. No trials have yet been performed inthe setting of HIV infection.
6. Outstanding research questions
Many outstanding research questions remain about naive T-cells and HIV infection (Table 3). Most of our detailed under-standing of the mechanisms of naïve T-cell homeostasis is de-rived from mouse models. The effects of self-peptide andmicrobial translocation in driving enhanced naïve T-cell prolif-eration should be further explored in ex vivo human models.New interventions targeting immune reconstitution like exog-enous IL-7 have shown promise in clinical trials. However, amajor challenge for the field is to determine how best to eval-uate efficacy of immunomodulators such as IL-7 given the ob-servation of enhanced CD4+ T-cell recovery but noimprovement in clinical endpoints in the recent IL-2 studies.The efficacy of adjunct immunomodulatory therapy withcART can no longer be evaluated on the basis of CD4+ T-cellnumber alone and large randomised trials with clinical
endpoints with better surrogate markers of immune functionother than the number of CD4+ T-cells alone will be needed.The varying responses in immune reconstitution between indi-viduals may potentially be predicted by genetic differencesand larger genomewide association studies similar to those re-cently conducted on HIV controllers [136] need be conductedto assess the determinants of CD4+ T-cell reconstitution.These studies will be important in identifying novel therapeu-tic targets to enhance immune recovery.
7. Conclusion
Successful immune reconstitution requires an increase intotal CD4+ T-cells as well as the recovery of CD4+ naïve T-cell number and function. Understanding the multiple fac-tors involved in naïve T-cell homeostasis and proliferationwill allow for the development of new strategies to enhanceimmune reconstitution.
Role of the funding source
G.K is a recipient of the National Health and Medical ResearchCouncil (NHMRC) biomedical postgraduate scholarship.
S.R.L is funded by the NHMRC and Alfred Foundation and isan NHMRC Practitioner Fellow
P.U.C and S.R.L were funded by an NHMRC program grant forthis work.
R.R is a recipient of the King Scholarship from the Malaysiangovernment.
Conflict of interest statement
The author(s) declare that there are no conflicts of interest.
References
[1] J.V. Baker, G. Peng, J. Rapkin, D.I. Abrams, M.J. Silverberg, R.D.MacArthur, W.P. Cavert, W.K. Henry, J.D.f. Neaton, the TerryBeirn Community Programs for Clinical Research on AIDS., CD4+count and risk of non-AIDS diseases following initial treatmentfor HIV infection, AIDS 22 (2008) 841–848.

262 G. Khoury et al.
[2] J.V. Baker, G. Peng, J. Rapkin, D. Krason, C. Reilly, W.P.Cavert, D.I. Abrams, R.D. MacArthur, K. Henry, J.D. Neaton,For the Terry Beirn Community Programs for clinical researchon AIDS., Poor initial CD4+ recovery with antiretroviral therapyprolongs immune depletion and increases risk for AIDS and non-AIDS diseases, J. Acquir. Immune Defic. Syndr. 48 (2008)541–546.
[3] J. Reekie, C. Kosa, F. Engsig, A.d.A. Monforte, A. Wiercinska-Drapalo, P. Domingo, F. Antunes, N. Clumeck, O. Kirk, J.D.Lundgren, A. Mocroft, S.S.G. for the Euro, relationship betweencurrent level of immunodeficiency and non-acquired immuno-deficiency syndrome-defining malignancies, Cancer 116 (2010)5306–5315.
[4] K.A. Lichtenstein, C. Armon, K. Buchacz, J.S. Chmiel, K. Buck-ner, E.M. Tedaldi, K. Wood, S.D. Holmberg, J.T. Brooks, LowCD4+ T Cell count is a risk factor for cardiovascular diseaseevents in the HIV outpatient study, Clin. Infect. Dis. 51 (2010)435–447.
[5] N. Lohse, A.B. Hansen, G. Pedersen, G. Kronborg, J. Gerstoft,H.T. Sorensen, M. Vaeth, N. Obel, Survival of persons with andwithout HIV infection in Denmark, 1995–2005, Ann. Intern.Med. 146 (2007) 87–95.
[6] T.A.T.C. Collaboration, Causes of death in HIV-1—infectedpatients treated with antiretroviral therapy, 1996–2006:collaborative analysis of 13 HIV cohort studies, Clin. Infect.Dis. 50 (2010) 1387–1396.
[7] S.C. De Rosa, L.A. Herzenberg, L.A. Herzenberg, M. Roederer,11-color, 13-parameter flow cytometry: Identification ofhuman naive T cells by phenotype, function, and T-cell receptordiversity, Nat. Med. 7 (2001) 245–248.
[8] L. Swainson, E. Verhoeyen, F.-L. Cosset, N. Taylor, IL-7Rα geneexpression is inversely correlated with cell cycle progression inIL-7-stimulated T lymphocytes, J. Immunol. 176 (2006)6702–6708.
[9] S. Kimmig, G.K. Przybylski, C.A. Schmidt, K. Laurisch, B.Möwes, A. Radbruch, A. Thiel, Two subsets of naive T helpercells with distinct T cell receptor excision circle content inhuman adult peripheral blood, J. Exp. Med. 195 (2002)789–794.
[10] F. Sallusto, D. Lenig, R. Forster, M. Lipp, A. Lanzavecchia,Two subsets of memory T lymphocytes with distinct homingpotentials and effector functions, Nature 401 (1999) 708–712.
[11] S. Junge, B. Kloeckener-Gruissem, R. Zufferey, A. Keisker, B.Salgo, J.-C. Fauchere, F. Scherer, T. Shalaby, M. Grotzer, U.Siler, R. Seger, T. Güngör, Correlation between recent thymicemigrants and CD31+ (PECAM-1) CD4+ T cells in normal individ-uals during aging and in lymphopenic children, Eur. J. Immunol.37 (2007) 3270–3280.
[12] J.T. Tan, E. Dudl, E. LeRoy, R. Murray, J. Sprent, K.I. Weinberg,C.D. Surh, IL-7 is critical for homeostatic proliferation andsurvival of naïve T cells, Proc. Natl. Acad. Sci. U. S. A. 98(2001) 8732–8737.
[13] U. von Freeden-Jeffry, P. Vieira, L.A. Lucian, T. McNeil, S.E.Burdach, R. Murray, Lymphopenia in interleukin (IL)-7 gene-deleted mice identifies IL-7 as a nonredundant cytokine,J. Exp. Med. 181 (1995) 1519–1526.
[14] T. Brocker, Survival of mature CD4 T lymphocytes is dependenton major histocompatibility complex class II–expressingdendritic cells, J. Exp. Med. 186 (1997) 1223–1232.
[15] J. Kirberg, A. Berns, H.v. Boehmer, Peripheral T cell survivalrequires continual ligation of the T cell receptor to majorhistocompatibility complex-encoded molecules, J. Exp. Med.186 (1997) 1269–1275.
[16] B. Min, H. Yamane, J. Hu-Li, W.E. Paul, Spontaneous andhomeostatic proliferation of CD4 T cells are regulated by differ-ent mechanisms, J. Immunol. 174 (2005) 6039–6044.
[17] T. Onoe, H. Kalscheuer, M. Chittenden, G. Zhao, Y.-G. Yang, M.Sykes, Homeostatic expansion and phenotypic conversion of
human T cells depend on peripheral interactions with APCs, J.Immunol. 184 (2010) 6756–6765.
[18] K.S. Schluns, W.C. Kieper, S.C. Jameson, L. Lefrancois, Inter-leukin-7 mediates the homeostasis of naive and memory CD8 Tcells in vivo, Nat. Immunol. 1 (2000) 426–432.
[19] T.J. Fry, C.L. Mackall, The many faces of IL-7: from lymphopoi-esis to peripheral T cell maintenance, J. Immunol. 174 (2005)6571–6576.
[20] W.C. Kieper, A. Troy, J.T. Burghardt, C. Ramsey, J.Y. Lee, H.-Q.Jiang, W. Dummer, H. Shen, J.J. Cebra, C.D. Surh, Cutting edge:recent immune status determines the source of antigens thatdrive homeostatic T cell expansion, J. Immunol. 174 (2005)3158–3163.
[21] C. Bourgeois, Z. Hao, K. Rajewsky, A.J. Potocnik, B. Stockinger,Ablation of thymic export causes accelerated decay of naïveCD4 T cells in the periphery because of activation by environ-mental antigen, Proc. Natl. Acad. Sci. U. S. A. 105 (2008)8691–8696.
[22] J.M. Brenchley, D.A. Price, T.W. Schacker, T.E. Asher, G.Silvestri, S. Rao, Z. Kazzaz, E. Bornstein, O. Lambotte, D.Altmann, B.R. Blazar, B. Rodriguez, L. Teixeira-Johnson, A.Landay, J.N. Martin, F.M. Hecht, L.J. Picker, M.M. Lederman,S.G. Deeks, D.C. Douek, Microbial translocation is a cause ofsystemic immune activation in chronic HIV infection, Nat.Med. 12 (2006) 1365–1371.
[23] J.M. Brenchley, T.W. Schacker, L.E. Ruff, D.A. Price, J.H.Taylor, G.J. Beilman, P.L. Nguyen, A. Khoruts, M. Larson, A.T.Haase, D.C. Douek, CD4+ T cell depletion during all stages ofHIV disease occurs predominantly in the gastrointestinal tract,J. Exp. Med. 200 (2004) 749–759.
[24] S. Mehandru, M.A. Poles, K. Tenner-Racz, A. Horowitz, A.Hurley, C. Hogan, D. Boden, P. Racz, M. Markowitz, PrimaryHIV-1 infection is associated with preferential depletion ofCD4+ T lymphocytes from effector sites in the gastrointestinaltract, J. Exp. Med. 200 (2004) 761–770.
[25] A.W. Goldrath, C.J. Luckey, R. Park, C. Benoist, D. Mathis, Themolecular program induced in T cells undergoing homeostaticproliferation, Proc. Natl. Acad. Sci. U. S. A. 101 (2004)16885–16890.
[26] B.K. Cho, V.P. Rao, Q. Ge, H.N. Eisen, J. Chen, Homeostasis-stimulated proliferation drives naive T cells to differentiate di-rectly into memory T cells, J. Exp. Med. 192 (2000) 549–556.
[27] C. Viret, F.S. Wong, C.A. Janeway, Designing and maintainingthe mature TCR repertoire: the continuum of self-peptide:Self-MHC complex recognition, Immunity 10 (1999) 559–568.
[28] B. Ernst, D.-S. Lee, J.M. Chang, J. Sprent, C.D. Surh, The pep-tide ligands mediating positive selection in the thymus controlT cell survival and homeostatic proliferation in the periphery,Immunity 11 (1999) 173–181.
[29] M. Guimond, R.G. Veenstra, D.J. Grindler, H. Zhang, Y. Cui, R.D.Murphy, S.Y. Kim, R. Na, L. Hennighausen, S. Kurtulus, B. Erman,P. Matzinger, M.S. Merchant, C.L. Mackall, Interleukin 7 signalingin dendritic cells regulates the homeostatic proliferation andniche size of CD4+ T cells, Nat. Immunol. 10 (2009) 149–157.
[30] L. Zhang, S.R. Lewin, M. Markowitz, H.-H. Lin, E. Skulsky, R.Karanicolas, Y. He, X. Jin, S. Tuttleton, M. Vesanen, H. Spiegel,R. Kost, J. van Lunzen, H.-J. Stellbrink, S. Wolinsky, W.Borkowsky, P. Palumbo, L.G. Kostrikis, D.D. Ho, Measuringrecent thymic emigrants in blood of normal and HIV-1-infectedindividuals before and after effective therapy, J. Exp. Med. 190(1999) 725–732.
[31] M.D. Hazenberg, S.A. Otto, J.W.T.C. Stuart, M.C.M. Verschuren,J.C.C. Borleffs, C.A.B. Boucher, R.A. Coutinho, J.M.A. Lange, T.F.R. de Wit, A. Tsegaye, J.J.M. van Dongen, D. Hamann, R.J. deBoer, F. Miedema, Increased cell division but not thymicdysfunction rapidly affects the T-cell receptor excision circlecontent of the naive T cell population in HIV-1 infection, Nat.Med. 6 (2000) 1036–1042.

The role of naïve T-cells in HIV-1 pathogenesis: An emerging key player 263
[32] D.C. Douek, R.D. McFarland, P.H. Keiser, E.A. Gage, J.M.Massey, B.F. Haynes, M.A. Polis, A.T. Haase, M.B. Feinberg,J.L. Sullivan, B.D. Jamieson, J.A. Zack, L.J. Picker, R.A.Koup, Changes in thymic function with age and during thetreatment of HIV infection, Nature 396 (1998) 690–695.
[33] B. Autran, G. Carcelain, T.S. Li, C. Blanc, D. Mathez, R.Tubiana, C. Katlama, P. Debré, J. Leibowitch, Positive effectsof combined antiretroviral therapy on CD4+ T cell homeostasisand function in advanced HIV disease, Science 277 (1997)112–116.
[34] M.A. Nokta, X.-D. Li, L. Al-Harthi, J. Nichols, A. Pou, D. Asmuth,A. Landay, R.B. Pollard, Entrapment of recent thymic emigrantsin lymphoid tissues fromHIV-infected patients: association withHIV cellular viral load, AIDS 16 (2002) 2119–2127.
[35] M. Di Mascio, I. Sereti, L.T. Matthews, V. Natarajan, J.Adelsberger, R. Lempicki, C. Yoder, E. Jones, C. Chow, J.A.Metcalf, I.A. Sidorov, D.S. Dimitrov, M.A. Polis, J.A. Kovacs,Naive T-cell dynamics in human immunodeficiency virus type 1infection: Effects of highly active antiretroviral therapy provideinsights into the mechanisms of naive T-cell depletion, J. Virol.80 (2006) 2665–2674.
[36] T.W. Schacker, J.M. Brenchley, G.J. Beilman, C. Reilly, S.E. Pam-buccian, J. Taylor, D. Skarda, M. Larson, D.C. Douek, A.T. Haase,Lymphatic Tissue fibrosis is associated with reduced numbers ofnaive CD4+ T cells in human immunodeficiency virus type 1 infec-tion, Clin. Vaccine Immunol. 13 (2006) 556–560.
[37] T.W. Schacker, P.L. Nguyen, G.J. Beilman, S. Wolinsky, M.Larson, C. Reilly, A.T. Haase, Collagen deposition in HIV-1infected lymphatic tissues and T cell homeostasis, J. Clin.Invest. 110 (2002) 1133–1139.
[38] M. Zeng, A.J. Smith, S.W. Wietgrefe, P.J. Southern, T.W.Schacker, C.S. Reilly, J.D. Estes, G.F. Burton, G. Silvestri, J.D. Lifson, J.V. Carlis, A.T. Haase, Cumulative mechanisms oflymphoid tissue fibrosis and T cell depletion in HIV-1 and SIVinfections, J. Clin. Invest. 121 (2011) 998–1008.
[39] R.L. Sabado, M. O'Brien, A. Subedi, L. Qin, N. Hu, E. Taylor, O.Dibben, A. Stacey, J. Fellay, K.V. Shianna, F. Siegal, M. Shodell,K. Shah, M. Larsson, J. Lifson, A. Nadas, M. Marmor, R. Hutt, D.Margolis, D. Garmon, M. Markowitz, F. Valentine, P. Borrow, N.Bhardwaj, Evidence of dysregulation of dendritic cells in primaryHIV infection, Blood 116 (2010) 3839–3852.
[40] N. Sachdeva, V. Asthana, T.H. Brewer, D. Garcia, D. Asthana,Impaired restoration of plasmacytoid dendritic cells in HIV-1-infected patients with poor CD4 T cell reconstitution is associ-ated with decrease in capacity to produce IFN-α butnot proinflammatory cytokines, J. Immunol. 181 (2008)2887–2897.
[41] V. Appay, S. Fastenackels, C. Katlama, H. Ait-Mohand, L.Schneider, A. Guihot, M. Keller, B. Grubeck-Loebenstein, A.Simon, O. Lambotte, P.W. Hunt, S.G. Deeks, D. Costagliola, B.Autran, D. Sauce, Old age and anti-CMV immunity are associat-ed with altered T cell reconstitution in HIV-1 infected patients,AIDS 25 (2011) 1813–1822.
[42] N. Vrisekoop, R. van Gent, A.B. de Boer, S.A. Otto, J.C.C.Borleffs, R. Steingrover, J.M. Prins, T.W. Kuijpers, T.F.W.Wolfs, S.P.M. Geelen, I. Vulto, P. Lansdorp, K. Tesselaar,J.A.M. Borghans, F. Miedema, Restoration of the CD4 T cellcompartment after long-term highly active antiretroviraltherapy without phenotypical signs of accelerated immuno-logical aging, J. Immunol. 181 (2008) 1573–1581.
[43] A. De Rossi, A.S. Walker, N. Klein, D. De Forni, D. King, D.M.Gibb, Increased thymic output after initiation of antiretroviraltherapy in human immunodeficiency virus type 1-Infected Chil-dren in the Paediatric European Network for Treatment of AIDS(PENTA) 5 Trial, J. Infect. Dis. 186 (2002) 312–320.
[44] T.W. Schacker, R.J. Bosch, K. Bennett, R. Pollard, G.K. Robbins,A.C. Collier, R.M. Gulick, J. Spritzler, D. Mildvan, For the AIDSclinical trials group., Measurement of naive CD4 cells reliably
predicts potential for immune reconstitution inHIV, J. Acquir. Im-mune Defic. Syndr. 54 (2010) 59–62.
[45] L.A. Napolitano, R.M. Grant, S.G. Deeks, D. Schmidt, S.C. DeRosa, L.A. Herzenberg, B.G. Herndier, J. Andersson, J.M.McCune, Increased production of IL-7 accompanies HIV-1-mediated T-cell depletion: implications for T-cell homeostasis,Nat. Med. 7 (2001) 73–79.
[46] S.A. Koesters, J.B. Alimonti, C. Wachihi, L. Matu, O. Anzala, J.Kimani, J.E. Embree, F.A. Plummer, K.R. Fowke, IL-7Rα expres-sion on CD4+ T lymphocytes decreases with HIV disease progres-sion and inversely correlates with immune activation, Eur. J.Immunol. 36 (2006) 336–344.
[47] B. Rethi, C. Fluur, A. Atlas, M. Krzyzowska, F. Mowafi, S. Grütz-meier, A. De Milito, R. Bellocco, K.I. Falk, É. Rajnavölgyi, F.Chiodi, Loss of IL-7R[alpha] is associated with CD4 T-cell deple-tion, high interleukin-7 levels and CD28 down-regulation in HIVinfected patients, AIDS 19 (2005) 2077–2086.
[48] D.A. Bazdar, S.F. Sieg, Interleukin-7 enhances proliferationresponses to T-cell receptor stimulation in naive CD4+ Tcells from human immunodeficiency virus-infected persons,J. Virol. 81 (2007) 12670–12674.
[49] A.A. Luciano, M.M. Lederman, A. Valentin-Torres, D.A. Bazdar,S.F. Sieg, Impaired induction of CD27 and CD28 predicts naiveCD4 T cell proliferation defects in HIV disease, J. Immunol.179 (2007) 3543–3549.
[50] O. Juffroy, F. Bugault, O. Lambotte, I. Landires, J.-P. Viard, L.Niel, A. Fontanet, J.-F. Delfraissy, J. Theze, L.A. Chakrabarti,Dual mechanism of impairment of interleukin-7 (IL-7) responsesin human immunodeficiency virus infection: decreased IL-7binding and abnormal activation of the JAK/STAT5 pathway,J. Virol. 84 (2010) 96–108.
[51] D.A. Bazdar, M. Kalinowska, S.F. Sieg, Interleukin-7 receptorsignaling is deficient in CD4+ T cells from HIV-infected personsand is inversely associated with aging, J. Infect. Dis. 199(2009) 1019–1028.
[52] I. Landires, F. Bugault, O. Lambotte, P.D. Truchis, L. Slama, A.Danckaert, J.F. Delfraissy, J. Theze, L.A. Chakrabarti, HIVinfection perturbs IL-7 signaling at the step of STAT5 nuclearrelocalization, AIDS 25 (2011) 1843–1853.
[53] R. Rajasuriar, D. Booth, A. Solomon, K. Chua, T. Spelman, M.Gouillou, T.E. Schlub, M. Davenport, S. Crowe, J. Elliott, J.Hoy, C. Fairley, G. Stewart, P. Cameron, S.R. Lewin, Biologicaldeterminants of immune reconstitution in HIV-infected patientsreceiving antiretroviral therapy: the role of interleukin 7 andinterleukin 7 receptor α and microbial translocation, J. Infect.Dis. 202 (2010) 1254–1264.
[54] R. Correa, S. Resino, M.A. Muñoz-Fernández, Increased inter-leukin-7 Plasma levels are associated with recovery of CD4+ Tcells in HIV-infected children, J. Clin. Immunol. 23 (2003)401–406.
[55] Jose F. Camargo, H. Kulkarni, Brian K. Agan, Alvaro A. Gaitan,Lisa A. Beachy, S. Srinivas, W. He, S. Anderson, Vincent C. Mar-coni, Matthew J. Dolan, Sunil K. Ahuja, Responsiveness of Tcells to interleukin‐7 Is associated with higher CD4+ T cellcounts in HIV‐1-positive individuals with highly active antiretro-viral therapy-induced viral load suppression, J. Infect. Dis. 199(2009) 1872–1882.
[56] J.-H. Colle, J.-L. Moreau, A. Fontanet, O. Lambotte, J.-F. Del-fraissy, J. Thèze, The correlation between levels of IL-7R[alpha] expression and responsiveness to IL-7 is lost in CD4 lym-phocytes from HIV-infected patients, AIDS 21 (2007) 101–103.
[57] J.-H. Colle, J.-L. Moreau, A. Fontanet, O. Lambotte, M. Jousse-met, S. Jacod, J.-F. Delfraissy, J. Thèze, Regulatory dysfunctionof the interleukin-7 receptor in CD4 and CD8 lymphocytes fromHIV-infected patients-effects of antiretroviral therapy, J. Acquir.Immune Defic. Syndr. 42 (2006) 277–285.
[58] M. Marziali, W. De Santis, R. Carello, W. Leti, A. Esposito, A.Isgrò, C. Fimiani, M.C. Sirianni, I. Mezzaroma, F. Aiuti, T-cell

264 G. Khoury et al.
homeostasis alteration in HIV-1 infected subjects with low CD4T-cell count despite undetectable virus load during HAART,AIDS 20 (2006) 2033–2041.
[59] S. Pahwa, V. Chitnis, R.M.Mitchell, S. Fernandez, A. Chandrase-kharan, C.M. Wilson, S.D. Douglas, CD4+ and CD8+ T cell recep-tor repertoire perturbations with normal levels of T cellreceptor excision circles in HIV-infected, therapy-naive adoles-cents, AIDS Res. Hum. Retroviruses 19 (2003) 487–495.
[60] L. Yin, Z.C. Kou, C. Rodriguez, W. Hou, M.M. Goodenow, J.W.Sleasman, Antiretroviral therapy restores diversity in theT-cell receptor V{beta} repertoire of CD4 T-Cell subpopula-tions among human immunodeficiency virus type 1-infectedchildren and adolescents, Clin. Vaccine Immunol. 16 (2009)1293–1301.
[61] G. Gorochov, A.U. Neumann, A. Kereveur, C. Parizot, T. Li, C.Katlama, M. Karmochkine, G. Raguin, B. Autran, P. Debre,Perturbation of CD4+ and CD8+ T-cell repertoires duringprogression to AIDS and regulation of the CD4+ repertoire duringantiviral therapy, Nat. Med. 4 (1998) 215–221.
[62] U. Malhotra, C. Huntsberry, S. Holte, J. Lee, L. Corey, M.J.McElrath, CD4+ T cell receptor repertoire perturbations inHIV-1 infection: association with plasma viremia and diseaseprogression, Clin. Immunol. 119 (2006) 95–102.
[63] P.N. Akolkar, B. Gulwani-Akolkar, N. Chirmule, S. Pahwa, V.S.Kalyanaraman, R. Pergolizzi, S. Macphail, J. Silver, The HIVglycoprotein gp160 has superantigen-like properties, Clin.Immunol. Immunopathol. 76 (1995) 255–265.
[64] K. Sakai, H. Gatanaga, H. Takata, S. Oka, M. Takiguchi,Comparison of CD4+ T-cell subset distribution in chronicallyinfected HIV+ patients with various CD4 nadir counts, MicrobesInfect. 12 (2010) 374–381.
[65] G.K. Robbins, J.G. Spritzler, E.S. Chan, D.M. Asmuth, R.T.Gandhi, B.A. Rodriguez, G. Skowron, P.R. Skolnik, R.W. Shafer,R.B. Pollard, A.C.T.G. Team, Incomplete reconstitution of T cellsubsets on combination antiretroviral therapy in the AIDS clinicaltrials group protocol 384, Clin. Infect. Dis. 48 (2009) 350–361.
[66] C.G. Lange, M.M. Lederman, K. Medvik, R. Asaad, M. Wild, R.Kalayjian, H. Valdez, Nadir CD4+ T-cell count and numbers ofCD28+ CD4+ T-cells predict functional responses to immuniza-tions in chronic HIV-1 infection, AIDS 17 (2003) 2015–2023.
[67] L.B. Gelinck, C.M. Jol-van der Zijde, A.M. Jansen-Hoogendijk,D.M. Brinkman, J.T. van Dissel, M.J. van Tol, F.P. Kroon, Resto-ration of the antibody response upon rabies vaccination in HIV-infected patients treated with HAART, AIDS 23 (2009)2451–2458.
[68] M. Rigaud, W. Borkowsky, P. Muresan, A. Weiberg, P. LaRussa,T. Fenton, J.S. Read, P. Jean-Philippe, E. Fergusson, B. Zim-mer, D. Smith, J. Kraimer, Impaired immunity to recall antigensand neoantigens in severely immunocompromised children andadolescents during the first year of effective highly active anti-retroviral therapy, J. Infect. Dis. 198 (2008) 1123–1130.
[69] B. Rodriguez, H. Valdez, C.G. Lange, R. Asaad, K. Medvik, S.F.Sieg, In vitro naïve T cell proliferation failure predicts poorpost-immunization responses to neoantigen, but not recall anti-gens, in HIV-infection, Clin. Immunol. 136 (2010) 400–408.
[70] J. Dai, L.M. Agosto, C. Baytop, J.J. Yu, M.J. Pace, M.K. Liszewski,U. O'Doherty, Human immunodeficiency virus integrates directlyinto naive resting CD4+TCells but enters naive cells less efficient-ly than memory cells, J. Virol. 83 (2009) 4528–4537.
[71] M.J. Pace, L. Agosto, U. O'Doherty, R5 HIV env and vesicularstomatitis virus G protein cooperate to mediate fusion tonaive CD4+ T cells, J. Virol. 85 (2011) 644–648.
[72] G. Doitsh, M. Cavrois, K.G. Lassen, O. Zepeda, Z. Yang, M.L.Santiago, A.M. Hebbeler, W.C. Greene, Abortive HIV infectionmediates CD4 T cell depletion and inflammation in human lym-phoid tissue, Cell 143 (2010) 789–801.
[73] J.A. Zack, S.J. Arrigo, S.R. Weitsman, A.S. Go, A. Haislip, I.S.Y.Chen, HIV-1 entry into quiescent primary lymphocytes:molecular
analysis reveals a labile, latent viral structure, Cell 61 (1990)213–222.
[74] P.U. Cameron, S. Saleh, G. Sallmann, A. Solomon, F.Wightman,V.A. Evans, G. Boucher, E.K. Haddad, R.-P. Sekaly, A.N.Harman, J.L. Anderson, K.L. Jones, J. Mak, A.L. Cunningham,A. Jaworowski, S.R. Lewin, Establishment of HIV-1 latency inresting CD4+ T cells depends on chemokine-induced changesin the actin cytoskeleton, Proc. Natl. Acad. Sci. U. S. A. 107(2010) 16934–16939.
[75] S. Saleh, A. Solomon, F. Wightman, M. Xhilaga, P.U. Cameron,S.R. Lewin, CCR7 ligands CCL19 and CCL21 increase permis-siveness of resting memory CD4+ T cells to HIV-1 infection: anovel model of HIV-1 latency, Blood 110 (2007) 4161–4164.
[76] V. Dardalhon, S. Jaleco, S. Kinet, B. Herpers, M. Steinberg, C.Ferrand, D. Froger, C. Leveau, P. Tiberghien, P. Charneau, N.Noraz, N. Taylor, IL-7 differentially regulates cell cycle progres-sion and HIV-1-based vector infection in neonatal and adultCD4+ T cells, Proc. Natl. Acad. Sci. U. S. A. 98 (2001) 9277–9282.
[77] D. Unutmaz, V.N. KewalRamani, S. Marmon, D.R. Littman, Cy-tokine signals are sufficient for HIV-1 infection of restinghuman T lymphocytes, J. Exp. Med. 189 (1999) 1735–1746.
[78] O. Ducrey-Rundquist, M. Guyader, D. Trono, Modalities ofinterleukin-7-induced human immunodeficiency virus permissive-ness in quiescent T lymphocytes, J. Virol. 76 (2002) 9103–9111.
[79] C.M. Steffens, E.Z. Managlia, A. Landay, L. Al-Harthi, Inter-leukin-7-treated naive T cells can be productively infectedby T-cell-adapted and primary isolates of human immunodefi-ciency virus 1, Blood 99 (2002) 3310–3318.
[80] A. Kinter, A. Moorthy, R. Jackson, A.S. Fauci, Productive HIVinfection of resting CD4+ T cells: role of lymphoid tissue mi-croenvironment and effect of immunomodulating agents,AIDS Res. Hum. Retroviruses 19 (2003) 847–856.
[81] D.A. Eckstein, M.L. Penn, Y.D. Korin, D.D. Scripture-Adams,J.A. Zack, J.F. Kreisberg, M. Roederer, M.P. Sherman, P.S.Chin, M.A. Goldsmith, HIV-1 actively replicates in naive CD4+T cells residing within human lymphoid tissues, Immunity 15(2001) 671–682.
[82] J.F. Kreisberg,W. Yonemoto,W.C. Greene, Endogenous factorsenhance HIV infection of tissue naive CD4 T cells by stimulatinghigh molecular mass APOBEC3G complex formation, J. Exp.Med. 203 (2006) 865–870.
[83] Y. Zhou, H. Zhang, J.D. Siliciano, R.F. Siliciano, Kinetics ofhuman immunodeficiency virus type 1 decay following entryinto resting CD4+ T cells, J. Virol. 79 (2005) 2199–2210.
[84] F. Groot, T. van Capel, J. Schuitemaker, B. Berkhout, E. deJong, Differential susceptibility of naive, central memory andeffector memory T cells to dendritic cell-mediated HIV-1 trans-mission, Retrovirology 3 (2006) 52.
[85] C. Barat, C. Gilbert, M.J. Tremblay, Efficient replication ofhuman immunodeficiency virus type 1 in resting CD4+ T lympho-cytes is induced by coculture with autologous dendritic cells inthe absence of foreign antigens, J. Virol. 83 (2009) 2778–2782.
[86] R. Zamarchi, P. Allavena, A. Borsetti, L. Stievano, V. Tosello, N.Marcato, G. Esposito, V. Roni, C. Paganin, G. Bianchi, F. Titti, P.Verani, G. Gerosa, A. Amadori, Expression and functional activ-ity of CXCR-4 and CCR-5 chemokine receptors in human thymo-cytes, Clin. Exp. Immunol. 127 (2002) 321–330.
[87] N. Schmitt, L. Chene, D. Boutolleau, M.T. Nugeyre, E. Guille-mard, P. Versmisse, C. Jacquemot, F. Barre-Sinoussi, N. Israel,Positive regulation of CXCR4 expression and signaling by inter-leukin-7 in CD4+ mature thymocytes correlates with their ca-pacity to favor human immunodeficiency X4 virus replication,J. Virol. 77 (2003) 5784–5793.
[88] K.B. Gurney, C.H. Uittenbogaart, Human immunodeficiencyvirus persistence and production in T-cell development,Clin. Vaccine Immunol. 13 (2006) 1237–1245.
[89] V.A. Evans, L. Lal, R. Akkina, A. Solomon, E.Wright, S.R. Lewin,P.U. Cameron, Thymic plasmacytoid dendritic cells are

The role of naïve T-cells in HIV-1 pathogenesis: An emerging key player 265
susceptible to productive HIV-1 infection and efficiently trans-fer R5 HIV-1 to thymocytes in vitro, Retrovirology 8 (2011) 43.
[90] L. Pedroza-Martins, W.J. Boscardin, D.J. Anisman-Posner, D.Schols, Y.J. Bryson, C.H. Uittenbogaart, Impact of cytokines onreplication in the thymus of primary human immunodeficiencyvirus type 1 isolates from infants, J. Virol. 76 (2002) 6929–6943.
[91] E. Guillemard, M.T. Nugeyre, L. Chene, N. Schmitt, C. Jacque-mot, F. Barre-Sinoussi, N. Israel, Interleukin-7 and infection itselfby human immunodeficiency virus 1 favor virus persistence inma-ture CD4(+)CD8(−)CD3(+) thymocytes through sustained induc-tion of Bcl-2, Blood 98 (2001) 2166–2174.
[92] J.M. Brenchley, B.J. Hill, D.R. Ambrozak, D.A. Price, F.J. Gue-naga, J.P. Casazza, J. Kuruppu, J. Yazdani, S.A. Migueles, M.Connors, M. Roederer, D.C. Douek, R.A. Koup, T-Cell subsetsthat harbor Human Immunodeficiency Virus (HIV) in vivo: impli-cations for HIV pathogenesis, J. Virol. 78 (2004) 1160–1168.
[93] E.J. Heeregrave, M.J. Geels, J.M. Brenchley, E. Baan, D.R.Ambrozak, R.M. van der Sluis, R. Bennemeer, D.C. Douek, J.Goudsmit, G. Pollakis, R.A. Koup, W.A. Paxton, Lack of invivo compartmentalization among HIV-1 infected naïve andmemory CD4+ T cell subsets, Virology 393 (2009) 24–32.
[94] M. Ostrowski, T. Chun, S. Justement, I. Motola, M. Spinelli, J.Adelsberger, L. Ehler, S. Mizell, C. Hallahan, A. Fauci, Bothmemory and CD45RA+/CD62L+ naive CD4(+) T cells areinfected in human immunodeficiency virus type 1-infected in-dividuals, J. Virol. 73 (1999) 6430–6435.
[95] F. Wightman, A. Solomon, G. Khoury, J.A. Green, L. Gray, P.R.Gorry, Y.S. Ho, N.K. Saksena, J. Hoy, S.M. Crowe, P.U. Camer-on, S.R. Lewin, Both CD31+ and CD31- Naive CD4+ T cells arepersistent HIV Type 1-infected reservoirs in individuals receiv-ing antiretroviral therapy, J. Infect. Dis. 202 (2010) 1738–1748.
[96] V. Fabre-Mersseman, J. Dutrieux, A. Louise, S. Rozlan, A.Lamine, R. Parker, M. Rancez, H. Nunes-Cabaco, A.E. Sousa,O. Lambotte, R. Cheynier, CD4+ recent thymic emigrants areinfected by HIV in vivo, implication for pathogenesis, AIDS 25(2011) 1153–1162.
[97] F. Baldanti, S. Paolucci, R. Gulminetti, R. Maserati, G. Miglior-ino, A. Pan, F. Maggiolo, G. Comolli, A. Chiesa, G. Gerna, Higherlevels of HIV DNA in memory and naive CD4+ T cell subsets of vi-remic compared to non-viremic patients after 18 and 24monthsof HAART, Antiviral Res. 50 (2001) 197–206.
[98] N. Chomont, M. El-Far, P. Ancuta, L. Trautmann, F.A. Procopio,B. Yassine-Diab, G. Boucher, M.-R. Boulassel, G. Ghattas, J.M.Brenchley, T.W. Schacker, B.J. Hill, D.C. Douek, J.-P. Routy,E.K. Haddad, R.-P. Sekaly, HIV reservoir size and persistenceare driven by T cell survival and homeostatic proliferation,Nat. Med. 15 (2009) 893–900.
[99] O. Lambotte, A. Demoustier, M.G. de Goër, C. Wallon, J. Gas-nault, C. Goujard, J.F. Delfraissy, Y. Taoufik, Persistence ofreplication-competent HIV in both memory and naive CD4 Tcell subsets in patients on prolonged and effective HAART,AIDS 16 (2002) 2151–2157.
[100] P. Delobel, K. Sandres-Sauné, M. Cazabat, F.-E. L'Faqihi, C.Aquilina, M. Obadia, C. Pasquier, B. Marchou, P. Massip, J. Izo-pet, Persistence of distinct HIV-1 populations in blood mono-cytes and naive and memory CD4 T cells during prolongedsuppressive HAART, AIDS 19 (2005) 1739–1750.
[101] J.W. Sleasman, L.F. Aleixo, A. Morton, S. Skoda-Smith, M.M.Goodenow, CD4+ memory T cells are the predominant popula-tion of HIV-1-infected lymphocytes in neonates and children,AIDS 10 (1996) 1477–1484.
[102] H. Blaak, A.B. van't Wout, M. Brouwer, B. Hooibrink, E. Hoven-kamp, H. Schuitemaker, In vivo HIV-1 infection of CD45RA(+)CD4(+) T cells is established primarily by syncytium-inducingvariants and correlates with the rate of CD4(+) T cell decline,Proc. Natl. Acad. Sci. U. S. A. 97 (2000) 1269–1274.
[103] S. McBreen, S. Imlach, T. Shirafuji, G.R. Scott, C. Leen, J.E.Bell, P. Simmonds, Infection of the CD45RA+ (naive) subset
of peripheral CD8+ lymphocytes by human immunodeficiencyvirus type 1 in vivo, J. Virol. 75 (2001) 4091–4102.
[104] A. Foussat, L. Bouchet-Delbos, J. Couderc, D. Berrebi, M.German-Fattal, M.-C. Maillot, I. Durand-Gasselin, P. Galanaud,J.P.D. Santo, D. Emilie, Effects of exogenous IL-2 administrationon the homeostasis of CD4+ T lymphocytes, J. Clin. Immunol. 24(2004) 503–514.
[105] G. Carcelain, P. Saint-Mézard, H.K. Altes, R. Tubiana, P.Grenot, C. Rabian, R. de Boer, D. Costagliola, C. Katlama, P.Debré, B. Autran, IL-2 therapy and thymic production of naiveCD4 T cells in HIV-infected patients with severe CD4 lymphope-nia, AIDS 17 (2003) 841–850.
[106] F. Pandolfi, M. Pierdominici, M. Marziali, M. Livia Bernardi, G.Antonelli, V. Galati, G. D'Offizi, F. Aiuti, Low-dose IL-2 reduceslymphocyte apoptosis and increases naive CD4 cells in HIV-1 pa-tients treated with HAART, Clin. Immunol. 94 (2000) 153–159.
[107] G. Marchetti, L. Meroni, C. Molteni, A. Bandera, F. Franzetti,M. Galli, M. Moroni, M. Clerici, A. Gori, Interleukin-2 immuno-therapy exerts a differential effect on CD4 and CD8 T cell dy-namics, AIDS 18 (2004) 211–216.
[108] V. Natarajan, R.A. Lempicki, I. Sereti, Y. Badralmaa, J.W.Adelsberger, J.A. Metcalf, D.A. Prieto, R. Stevens, M.W. Baseler,J.A. Kovacs, H.C. Lane, Increased peripheral expansion of naiveCD4+ T cells in vivo after IL-2 treatment of patients with HIVinfection, Proc. Natl. Acad. Sci. U. S. A. 99 (2002) 10712–10717.
[109] P. De Paoli, M.T. Bortolin, S. Zanussi, A. Monzoni, C. Pratesi,M. Giacca, Changes in thymic function in HIV-positive patientstreated with highly active antiretroviral therapy and interleu-kin-2, Clin. Exp. Immunol. 125 (2001) 440–446.
[110] I. Sereti, K.B. Anthony, H. Martinez-Wilson, R. Lempicki, J.Adelsberger, J.A. Metcalf, C.W. Hallahan, D. Follmann, R.T.Davey, J.A. Kovacs, H.C. Lane, IL-2-induced CD4+ T-cell expan-sion in HIV-infected patients is associated with long-term de-creases in T-cell proliferation, Blood 104 (2004) 775–780.
[111] M. Losso, D. Abrams, I.E.S. Group, Effects of Interleukin-2 onclinical outcomes in patients with a CD4+ cell count of 300/mm3: primary results of the ESPRIT Study, Conference on Retro-viruses and Opportunistic Infections, Montreal, Canada, 2009.
[112] Y. Levy, S.S. Committee, Effect of interleukin-2 on clinicaloutcomes in patients with CD4+ cell count 50 to 299/mm3;Primary results of SILCAAT study, Conference on Retrovirusesand Opportunistic Infections., Montreal, Canada, 2009.
[113] D. Abrams, Y. Levy, M.H. Losso, A. Babiker, G. Collins, D.A. Coo-per, J. Darbyshire, S. Emery, L. Fox, F. Gordin, H.C. Lane, J.D.Lundgren, R. Mitsuyasu, J.D. Neaton, A. Phillips, J.P. Routy, G.Tambussi, D. Wentworth, Interleukin-2 therapy in patients withHIV infection, N. Engl. J. Med. 361 (2009) 1548–1559.
[114] J.A. Tavel, I.S.S. Group, Effects of intermittent IL-2 Alone orwith Peri-cycle antiretroviral therapy in early HIV infection:The STALWART Study, PLoS One 5 (2010) e9334.
[115] T.J. Fry, M. Moniuszko, S. Creekmore, S.J. Donohue, D.C.Douek, S. Giardina, T.T. Hecht, B.J. Hill, K. Komschlies, J.Tomaszewski, G. Franchini, C.L. Mackall, IL-7 therapy dramati-cally alters peripheral T-cell homeostasis in normal and SIV-infected nonhuman primates, Blood 101 (2003) 2294–2299.
[116] S. Beq, M.T. Nugeyre, R. Ho Tsong, D. Fang, R. Gautier, N.Legrand, J. Schmitt, F. Estaquier, B. Barre-Sinoussi, R. Hurtrel,N. Cheynier, Israel, IL-7 induces immunological improvement inSIV-infected rhesus macaques under antiviral therapy, J. Immu-nol. 176 (2006) 914–922.
[117] M.T. Nugeyre, V. Monceaux, S. Beq, M.C. Cumont, R. Ho Tsong, L.Fang, M. Chene, F. Morre, B. Barre-Sinoussi, N. Hurtrel, Israel,IL-7 stimulates T cell renewal without increasing viral replicationin simian immunodeficiency virus-infected macaques, J. Immu-nol. 171 (2003) 4447–4453.
[118] A. Leone, M. Rohankhedkar, A. Okoye, A. Legasse, M.K.Axthelm, F. Villinger, M. Piatak Jr., J.D. Lifson, B. Assouline,M. Morre, L.J. Picker, D.L. Sodora, Increased CD4+ T cell levels

266 G. Khoury et al.
during IL-7 administration of antiretroviral therapy-treatedsimian immunodeficiency virus-positive macaques are not de-pendent on strong proliferative responses, J. Immunol. 185(2010) 1650–1659.
[119] R. Parker, J. Dutrieux, S. Beq, B. Lemercier, S. Rozlan, V.Fabre-Mersseman, M. Rancez, C. Gommet, B. Assouline, I.Rance, A. Lim, M. Morre, R. Cheynier, Interleukin-7 treatmentcounteracts IFN-alpha therapy-induced lymphopenia and stimu-lates SIV-specific cytotoxic T lymphocyte responses in SIV-infected rhesus macaques, Blood 116 (2010) 5589–5599.
[120] S. Beq, S. Rozlan, D. Gautier, R. Parker, V. Mersseman, C.Schilte, B. Assouline, I. Rance, P. Lavedan, M. Morre, R. Chey-nier, Injection of glycosylated recombinant simian IL-7 provokesrapid and massive T-cell homing in rhesus macaques, Blood 114(2009) 816–825.
[121] Y. Levy, C. Lacabaratz, L. Weiss, J.-P. Viard, C. Goujard, J.-D.Lelièvre, F. Boué, J.-M. Molina, C. Rouzioux, V. Avettand-Fénoêl,T. Croughs, S. Beq, R. Thiébaut, G. Chêne, M. Morre, J.-F. Del-fraissy, Enhanced T cell recovery in HIV-1-infected adults throughIL-7 treatment, J. Clin. Invest. 119 (2009) 997–1007.
[122] I. Sereti, R.M. Dunham, J. Spritzler, E. Aga, M.A. Proschan, K.Medvik, C.A. Battaglia, A.L. Landay, S. Pahwa, M.A. Fischl,D.M. Asmuth, A.R. Tenorio, J.D. Altman, L. Fox, S. Moir, A.Malaspina, M. Morre, R. Buffet, G. Silvestri, M.M. Lederman,IL-7 administration drives T cell-cycle entry and expansion inHIV-1 infection, Blood 113 (2009) 6304–6314.
[123] N. Brieu, P. Portalès, M.-J. Carles, P. Corbeau, Interleukin-7 in-duces HIV type 1 R5-to-X4 switch, Blood 117 (2011) 2073–2074.
[124] A.L. Fiser, T. Vincent, N. Brieu, Y.L. Lin, P. Portales, C. Mettling,J. Reynes, P. Corbeau, High CD4+ T-cell surface CXCR4 density asa risk factor for R5 to X4 switch in the course of HIV-1 infection,J. Acquir. Immune Defic. Syndr. 55 (2010) 529–535.
[125] M. Noguchi, Y. Nakamura, S.M. Russell, S.F. Ziegler, M. Tsang,X. Cao, W.J. Leonard, Interleukin-2 receptor gamma chain: afunctional component of the interleukin-7 receptor, Science262 (1993) 1877–1880.
[126] S.M. Teutsch, D.R. Booth, B.H. Bennetts, R.N.S. Heard, G.J.Stewart, Identification of 11 novel and common single nucleo-tide polymorphisms in the interleukin-7 receptor-[alpha] geneand their associations with multiple sclerosis, Eur. J. Hum.Genet. 11 (2003) 509–515.
[127] D. Booth, A. Arthur, S. Teutsch, C. Bye, J. Rubio, P. Armati, J.Pollard, R. Heard, G. Stewart, Gene expression and genotypingstudies implicate the interleukin 7 receptor in the pathogenesisof primary progressivemultiple sclerosis, J. Mol. Med. 83 (2005)822–830.
[128] S.G. Gregory, S. Schmidt, P. Seth, J.R. Oksenberg, J. Hart, A.Prokop, S.J. Caillier, M. Ban, A. Goris, L.F. Barcellos, R. Lincoln,J.L. McCauley, S.J. Sawcer, D.A.S. Compston, B. Dubois, S.L.Hauser, M.A. Garcia-Blanco, M.A. Pericak-Vance, Interleukin 7receptor [alpha] chain (IL7R) shows allelic and functional asso-ciation with multiple sclerosis, Nat. Genet. 39 (2007)1083–1091.
[129] T. Rose, O. Lambotte, C. Pallier, J.-F. Delfraissy, J.-H. Colle,Identification and biochemical characterization of humanplasma soluble IL-7R: lower concentrations in HIV-1-infectedpatients, J. Immunol. 182 (2009) 7389–7397.
[130] B.R. Hansen, L. Kolte, S.B. Haugaard, C. Dirksen, F.K. Jensen,L.P. Ryder, A.L. Sørensen, A. Flyvbjerg, S.D. Nielsen, O. Ander-sen, Improved thymic index, density and output in HIV-infectedpatients following low-dose growth hormone therapy: a placebocontrolled study, AIDS 23 (2009) 2123–2131.
[131] R. Seggewiss, K. Loré, F.J. Guenaga, S. Pittaluga, J. Mattapallil,C.K. Chow, R.A. Koup, K. Camphausen, M.C. Nason, M. Meier-Schellersheim, R.E. Donahue, B.R. Blazar, C.E. Dunbar, D.C.Douek, Keratinocyte growth factor augments immune reconsti-tution after autologous hematopoietic progenitor cell trans-plantation in rhesus macaques, Blood 110 (2007) 441–449.
[132] D. Min, P.A. Taylor, A. Panoskaltsis-Mortari, B. Chung, D.M.Danilenko, C. Farrell, D.L. Lacey, B.R. Blazar, K.I. Weinberg,Protection from thymic epithelial cell injury by keratinocytegrowth factor: a new approach to improve thymic and periph-eral T-cell reconstitution after bone marrow transplantation,Blood 99 (2002) 4592–4600.
[133] J.S. Sutherland, G.L. Goldberg, M.V. Hammett, A.P. Uldrich, S.P. Berzins, T.S. Heng, B.R. Blazar, J.L. Millar, M.A. Malin, A.P.Chidgey, R.L. Boyd, Activation of thymic regeneration in miceand humans following androgen blockade, J. Immunol. 175(2005) 2741–2753.
[134] N.J. Olsen, W.J. Kovacs, Evidence that androgens modulatehuman thymic T cell output, J. Investig. Med. 59 (2011) 32–35.
[135] J.S. Sutherland, L. Spyroglou, J.L. Muirhead, T.S. Heng, A.Prieto-Hinojosa, H.M. Prince, A.P. Chidgey, A.P. Schwarer, R.L. Boyd, Enhanced Immune system regeneration in humans fol-lowing allogeneic or autologous hemopoietic stem cell trans-plantation by temporary sex steroid blockade, Clin. CancerRes. 14 (2008) 1138–1149.
[136] F. Pereyra, X. Jia, P.J. McLaren, A. Telenti, P.I. de Bakker, B.D.Walker, S. Ripke, C.J. Brumme, S.L. Pulit, M. Carrington, C.M.Kadie, J.M. Carlson, D. Heckerman, R.R. Graham, R.M. Plenge,S.G. Deeks, L. Gianniny, G. Crawford, J. Sullivan, E. Gonzalez,L. Davies, A. Camargo, J.M. Moore, N. Beattie, S. Gupta, A.Crenshaw, N.P. Burtt, C. Guiducci, N. Gupta, X. Gao, Y. Qi, Y.Yuki, A. Piechocka-Trocha, E. Cutrell, R. Rosenberg, K.L.Moss, P. Lemay, J. O'Leary, T. Schaefer, P. Verma, I. Toth, B.Block, B. Baker, A. Rothchild, J. Lian, J. Proudfoot, D.M. Alvino,S. Vine, M.M. Addo, T.M. Allen, M. Altfeld, M.R. Henn, S. LeGall, H. Streeck, D.W. Haas, D.R. Kuritzkes, G.K. Robbins, R.W.Shafer, R.M. Gulick, C.M. Shikuma, R. Haubrich, S. Riddler,P.E. Sax, E.S. Daar, H.J. Ribaudo, B. Agan, S. Agarwal, R.L.Ahern, B.L. Allen, S. Altidor, E.L. Altschuler, S. Ambardar,K. Anastos, B. Anderson, V. Anderson, U. Andrady, D. Anto-niskis, D. Bangsberg, D. Barbaro, W. Barrie, J. Bartczak, S.Barton, P. Basden, N. Basgoz, S. Bazner, N.C. Bellos, A.M.Benson, J. Berger, N.F. Bernard, A.M. Bernard, C. Birch, S.J.Bodner, R.K. Bolan, E.T. Boudreaux, M. Bradley, J.F. Braun,J.E. Brndjar, S.J. Brown, K. Brown, S.T. Brown, J. Burack,L.M. Bush, V. Cafaro, O. Campbell, J. Campbell, R.H. Carl-son, J.K. Carmichael, K.K. Casey, C. Cavacuiti, G. Celestin,S.T. Chambers, N. Chez, L.M. Chirch, P.J. Cimoch, D.Cohen, L.E. Cohn, B. Conway, D.A. Cooper, B. Cornelson, D.T.Cox, M.V. Cristofano, G. Cuchural Jr., J.L. Czartoski, J.M. Dah-man, J.S. Daly, B.T. Davis, K. Davis, S.M. Davod, E. DeJesus,C.A. Dietz, E. Dunham, M.E. Dunn, T.B. Ellerin, J.J. Eron, J.J.Fangman, C.E. Farel, H. Ferlazzo, S. Fidler, A. Fleenor-Ford,R. Frankel, K.A. Freedberg, N.K. French, J.D. Fuchs, J.D. Fuller,J. Gaberman, J.E. Gallant, R.T. Gandhi, E. Garcia, D. Garmon,J.C. Gathe Jr., C.R. Gaultier, W. Gebre, F.D. Gilman, I. Gilson,P.A. Goepfert, M.S. Gottlieb, C. Goulston, R.K. Groger, T.D.Gurley, S. Haber, R. Hardwicke, W.D. Hardy, P.R. Harrigan,T.N. Hawkins, S. Heath, F.M. Hecht, W.K. Henry, M. Hladek,R.P. Hoffman, J.M. Horton, R.K. Hsu, G.D. Huhn, P. Hunt, M.J.Hupert, M.L. Illeman, H. Jaeger, R.M. Jellinger, M. John, J.A.Johnson, K.L. Johnson, H. Johnson, K. Johnson, J. Joly, W.C.Jordan, C.A. Kauffman, H. Khanlou, R.K. Killian, A.Y. Kim,D.D. Kim, C.A. Kinder, J.T. Kirchner, L. Kogelman, E.M.Kojic, P.T. Korthuis, W. Kurisu, D.S. Kwon, M. LaMar, H. Lam-piris, M. Lanzafame, M.M. Lederman, D.M. Lee, J.M. Lee, M.J.Lee, E.T. Lee, J. Lemoine, J.A. Levy, J.M. Llibre, M.A. Liguori,S.J. Little, A.Y. Liu, A.J. Lopez, M.R. Loutfy, D. Loy, D.Y. Mo-hammed, A. Man, M.K. Mansour, V.C. Marconi, M. Markowitz,R. Marques, J.N. Martin, H.L. Martin Jr., K.H. Mayer, M.J. McEl-rath, T.A. McGhee, B.H. McGovern, K. McGowan, D. McIntyre,G.X. McLeod, P. Menezes, G. Mesa, C.E. Metroka, D. Meyer-Olson, A.O. Miller, K. Montgomery, K.C. Mounzer, E.H. Nagami,I. Nagin, R.G. Nahass, M.O. Nelson, C. Nielsen, D.L. Norene,

The role of naïve T-cells in HIV-1 pathogenesis: An emerging key player 267
D.H. O'Connor, B.O. Ojikutu, J. Okulicz, O.O. Oladehin, E.C.Oldfield III, E.C. Oldfield III, S.A. Olender, M. Ostrowski, W.F.Owen Jr., E. Pae, J. Parsonnet, A.M. Pavlatos, A.M. Perlmut-ter, M.N. Pierce, J.M. Pincus, L. Pisani, L.J. Price, L. Proia,R.C. Prokesch, H.C. Pujet, M. Ramgopal, A. Rathod, M.Rausch, J. Ravishankar, F.S. Rhame, C.S. Richards, D.D. Rich-man, B. Rodes, M. Rodriguez, R.C. Rose III, E.S. Rosenberg, D.Rosenthal, P.E. Ross, D.S. Rubin, E. Rumbaugh, L. Saenz, M.R.Salvaggio, W.C. Sanchez, V.M. Sanjana, S. Santiago, W.Schmidt, H. Schuitemaker, P.M. Sestak, P. Shalit, W. Shay,V.N. Shirvani, V.I. Silebi, J.M. Sizemore Jr., P.R. Skolnik, M.Sokol-Anderson, J.M. Sosman, P. Stabile, J.T. Stapleton, S.Starrett, F. Stein, H.J. Stellbrink, F.L. Sterman, V.E. Stone,D.R. Stone, G. Tambussi, R.A. Taplitz, E.M. Tedaldi, W. Thei-sen, R. Torres, L. Tosiello, C. Tremblay, M.A. Tribble, P.D.Trinh, A. Tsao, P. Ueda, A. Vaccaro, E. Valadas, T.J. Vanig,I. Vecino, V.M. Vega, W. Veikley, B.H. Wade, C. Walworth,C. Wanidworanun, D.J. Ward, D.A. Warner, R.D. Weber, D.Webster, S. Weis, D.A. Wheeler, D.J. White, E. Wilkins, A.Winston, C.G. Wlodaver, A. van't Wout, D.P. Wright, O.O.Yang, D.L. Yurdin, B.W. Zabukovic, K.C. Zachary, B. Zeeman,M. Zhao, The major genetic determinants of HIV-1 control
affect HLA class I peptide presentation, Science 330 (2010)1551–1557.
[137] S. Mehandru, M. Poles, K. Tenner-Racz, A. Horowitz, A. Hur-ley, C. Hogan, D. Boden, P. Racz, M. Markowitz, PrimaryHIV-1 infection is associated with preferential depletion ofCD4+ T lymphocytes from effector sites in the gastrointestinaltract, J. Exp. Med. 200 (2004) 761–770.
[138] M. Guadalupe, E. Reay, S. Sankaran, T. Prindiville, J. Flamm, A.McNeil, S. Dandekar, Severe CD4+ T-cell depletion in gut lym-phoid tissue during primary human immunodeficiency virus type1 infection and substantial delay in restoration following highlyactive antiretroviral therapy, J. Virol. 77 (2003) 11708–11717.
[139] W. Jiang, M.M. Lederman, P. Hunt, S.F. Sieg, K. Haley, B.Rodriguez, A. Landay, J. Martin, E. Sinclair, A.I. Asher, S.G.Deeks, D.C. Douek, J.M. Brenchley, Plasma levels of bacterialDNA correlate with immune activation and the magnitude of im-mune restoration in persons with antiretroviral-treated HIV infec-tion, J. Infect. Dis. 199 (2009) 1177–1185.
[140] J.D. Estes, A.T. Haase, T.W. Schacker, The role of collagen de-position in depleting CD4+ T cells and limiting reconstitution inHIV-1 and SIV infections through damage to the secondary lym-phoid organ niche, Semin. Immunol. 20 (2008) 181–186.
Copyright © 2022 FDOKUMEN




















