
The role of CYP2E1 and 2B1 in metabolic activation of benzene derivatives
12
METABOLIC ACTIVATION / INACTIVATION Ivan Gut · Vessela Nedelcheva · Pavel Souc ˇek · Pavel Stopka Pavel Vodic ˇka · Harry V. Gelboin · Magnus Ingelman-Sundberg The role of CYP2E1 and 2B1 in metabolic activation of benzene derivatives Received: 12 March 1996/Accepted: 5 July 1996 Abstract CYP2B1 and 2E1 oxidized toluene, aniline and monochlorobenzene (MCB) to water-soluble meta- bolites and to products covalently binding to micro- somal proteins from male Wistar rats at high efficiency. Oxidation of benzene to covalently binding metabolites was catalysed by CYP2B1 and 2E1 more effectively than the formation of water-soluble metabolites, especially at low benzene levels. Thus, the formation of covalently binding products was inversely related but formation of soluble metabolites was proportional to benzene con- centration. 1,4-Benzoquinone was responsible for the majority of covalent binding to microsomal proteins, being suppressed by ascorbate; 1,4-semiquinone was not important, since a-tocopherol did not inhibit the cova- lent binding and ESR showed its rapid decay, if NADPH was available. Specific antibodies and in- hibitors confirmed the role of CYP2B1 and 2E1 induc- tion. Covalent binding of benzene to DNA was largely due to benzene oxide; ∼50% was due to N-7 guanine adduct. CYP2E1 oxidizing benzene via phenol to 1,4- hydroquinone appeared to mediate its further oxidation to 1,4-benzoquinone, which also occurred sponta- neously, but was reversed in a reducing environment of microsomes with NADPH. Production of OH radicals in microsomes with NADPH was greatly stimulated by HQ and less by BQ, especially in CYP2E1 induced mi- crosomes, although the quinones themselves failed to produce OH radicals. The quinones could act by sti- mulation of the CYP futile cycle. Therefore, CYP2B1 and 2E1 in rats appeared essential for metabolic acti- vation of benzene derivatives to potentially genotoxic products; BQ dominated the covalent binding of ben- zene to proteins, whereas DNA adducts were largely due to benzene oxide. Abbreviations BQ 1,4-Benzoquinone · SQ 1,4-Semiquinone · CYP cytochrome P450 · ESR electron spin resonance · HQ 1,4-hydroquinone · MCB monochlorobenzene · PB phenobarbital Key words Cytochrome P450 · Benzene · Metabolic activation · Covalent binding · DNA adducts Introduction The carcinogenicity of benzene, aniline and certain chlorobenzenes (IARC 1987) is believed to be due to their oxidation by cytochrome P450 enzymes (Souc ˇek and Gut 1992; Nedelcheva and Gut 1994). Although toluene is probably not carcinogenic, recent studies have reassesed this possibility (NIOSH 1987). CYP oxidizes toluene mainly to benzylalcohol, less to o- and p-cresol. Rat liver CYP2E1 had a lower K m and higher V max than CYP2B1 for oxidation to benzylalcohol, but CYP2B1 formed markedly more reactive cresols (Wang and Na- kajima 1991; Nakajima et al. 1991), which could bind covalently to biomacromolecules, as suggested by Pa- thiratne et al.(1986). Monochlorobenzene (MCB) is oxidized by CYP directly or via a ketone intermediate to MCB oxide (Korzekwa et al. 1989), which forms 2-, 3- and 4-chlorophenol, 4-chlorocatechol or 4-chlor- ophenylmercapturic acid (Yoshida and Hara 1985). MCB was effectively oxidized by rat CYP2E1 and 2B1 (Gut et al. 1993), but only water-soluble metabolites were studied. Aniline is oxidized via phenylhydroxyl amine to p-aminophenol in decreasing efficiency by CYP2E1, 1A1/2 and 2B1 (Souc ˇek and Gut 1992). Sub- Arch Toxicol (1996) 71: 45–56 Springer-Verlag 1996 Financial support: Internal Grant Agency, Ministry of Health of the Czech Republic, Swedish Medical Research Council I. Gut (&) · V. Nedelcheva · P. Souc ˇek · P. Stopka · P. Vodic ˇka National Institute of Public Health, 10042 Prague, S ˇ roba ´rova 48, Czech Republic H.V. Gelboin Laboratory of Molecular Carcinogenesis, National Cancer Institute, Bethesda, MD 20892, USA M. Ingelman-Sundberg Berzelius Laboratory, Karolinska Institutet, Stockholm, Sweden
-
Upload
ingenieria110 -
Category
Documents
-
view
0 -
download
0
Transcript of The role of CYP2E1 and 2B1 in metabolic activation of benzene derivatives

METABOLIC ACTIVATION/INACTIVATION
Ivan Gut · Vessela Nedelcheva · Pavel Soucek · Pavel StopkaPavel Vodicka · Harry V. Gelboin · Magnus Ingelman-Sundberg
The role of CYP2E1 and 2B1 in metabolic activationof benzene derivatives
Received: 12 March 1996/Accepted: 5 July 1996
Abstract CYP2B1 and 2E1 oxidized toluene, anilineand monochlorobenzene (MCB) to water-soluble meta-bolites and to products covalently binding to micro-somal proteins from male Wistar rats at high efficiency.Oxidation of benzene to covalently binding metaboliteswas catalysed by CYP2B1 and 2E1 more effectively thanthe formation of water-soluble metabolites, especially atlow benzene levels. Thus, the formation of covalentlybinding products was inversely related but formation ofsoluble metabolites was proportional to benzene con-centration. 1,4-Benzoquinone was responsible for themajority of covalent binding to microsomal proteins,being suppressed by ascorbate; 1,4-semiquinone was notimportant, since a-tocopherol did not inhibit the cova-lent binding and ESR showed its rapid decay, ifNADPH was available. Specific antibodies and in-hibitors confirmed the role of CYP2B1 and 2E1 induc-tion. Covalent binding of benzene to DNA was largelydue to benzene oxide; ∼ 50% was due to N-7 guanineadduct. CYP2E1 oxidizing benzene via phenol to 1,4-hydroquinone appeared to mediate its further oxidationto 1,4-benzoquinone, which also occurred sponta-neously, but was reversed in a reducing environment ofmicrosomes with NADPH. Production of OH radicalsin microsomes with NADPH was greatly stimulated byHQ and less by BQ, especially in CYP2E1 induced mi-crosomes, although the quinones themselves failed toproduce OH radicals. The quinones could act by sti-
mulation of the CYP futile cycle. Therefore, CYP2B1and 2E1 in rats appeared essential for metabolic acti-vation of benzene derivatives to potentially genotoxicproducts; BQ dominated the covalent binding of ben-zene to proteins, whereas DNA adducts were largely dueto benzene oxide.
Abbreviations BQ 1,4-Benzoquinone ·SQ 1,4-Semiquinone · CYP cytochrome P450 ·ESR electron spin resonance · HQ 1,4-hydroquinone ·MCB monochlorobenzene · PB phenobarbital
Key words Cytochrome P450 · Benzene · Metabolicactivation · Covalent binding · DNA adducts
Introduction
The carcinogenicity of benzene, aniline and certainchlorobenzenes (IARC 1987) is believed to be due totheir oxidation by cytochrome P450 enzymes (Soucekand Gut 1992; Nedelcheva and Gut 1994). Althoughtoluene is probably not carcinogenic, recent studies havereassesed this possibility (NIOSH 1987). CYP oxidizestoluene mainly to benzylalcohol, less to o- and p-cresol.Rat liver CYP2E1 had a lower Km and higher Vmax thanCYP2B1 for oxidation to benzylalcohol, but CYP2B1formed markedly more reactive cresols (Wang and Na-kajima 1991; Nakajima et al. 1991), which could bindcovalently to biomacromolecules, as suggested by Pa-thiratne et al.(1986). Monochlorobenzene (MCB) isoxidized by CYP directly or via a ketone intermediate toMCB oxide (Korzekwa et al. 1989), which forms 2-, 3-and 4-chlorophenol, 4-chlorocatechol or 4-chlor-ophenylmercapturic acid (Yoshida and Hara 1985).MCB was effectively oxidized by rat CYP2E1 and 2B1(Gut et al. 1993), but only water-soluble metaboliteswere studied. Aniline is oxidized via phenylhydroxylamine to p-aminophenol in decreasing efficiency byCYP2E1, 1A1/2 and 2B1 (Soucek and Gut 1992). Sub-
Arch Toxicol (1996) 71: 45–56 Springer-Verlag 1996
Financial support: Internal Grant Agency, Ministry of Healthof the Czech Republic, Swedish Medical Research Council
I. Gut (&) · V. Nedelcheva · P. Soucek · P. Stopka · P. VodickaNational Institute of Public Health, 10042 Prague,Srobarova 48, Czech Republic
H.V. GelboinLaboratory of Molecular Carcinogenesis,National Cancer Institute, Bethesda, MD 20892, USA
M. Ingelman-SundbergBerzelius Laboratory, Karolinska Institutet, Stockholm, Sweden
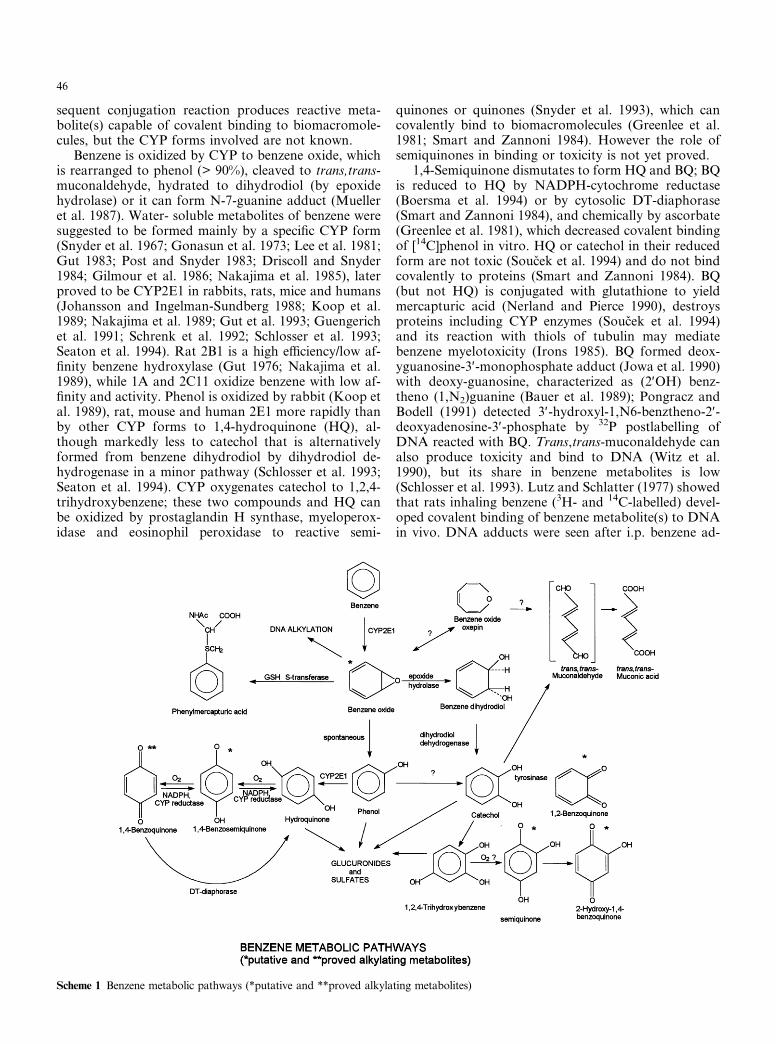
sequent conjugation reaction produces reactive meta-bolite(s) capable of covalent binding to biomacromole-cules, but the CYP forms involved are not known.
Benzene is oxidized by CYP to benzene oxide, whichis rearranged to phenol (> 90%), cleaved to trans,trans-muconaldehyde, hydrated to dihydrodiol (by epoxidehydrolase) or it can form N-7-guanine adduct (Muelleret al. 1987). Water- soluble metabolites of benzene weresuggested to be formed mainly by a specific CYP form(Snyder et al. 1967; Gonasun et al. 1973; Lee et al. 1981;Gut 1983; Post and Snyder 1983; Driscoll and Snyder1984; Gilmour et al. 1986; Nakajima et al. 1985), laterproved to be CYP2E1 in rabbits, rats, mice and humans(Johansson and Ingelman-Sundberg 1988; Koop et al.1989; Nakajima et al. 1989; Gut et al. 1993; Guengerichet al. 1991; Schrenk et al. 1992; Schlosser et al. 1993;Seaton et al. 1994). Rat 2B1 is a high efficiency/low af-finity benzene hydroxylase (Gut 1976; Nakajima et al.1989), while 1A and 2C11 oxidize benzene with low af-finity and activity. Phenol is oxidized by rabbit (Koop etal. 1989), rat, mouse and human 2E1 more rapidly thanby other CYP forms to 1,4-hydroquinone (HQ), al-though markedly less to catechol that is alternativelyformed from benzene dihydrodiol by dihydrodiol de-hydrogenase in a minor pathway (Schlosser et al. 1993;Seaton et al. 1994). CYP oxygenates catechol to 1,2,4-trihydroxybenzene; these two compounds and HQ canbe oxidized by prostaglandin H synthase, myeloperox-idase and eosinophil peroxidase to reactive semi-
quinones or quinones (Snyder et al. 1993), which cancovalently bind to biomacromolecules (Greenlee et al.1981; Smart and Zannoni 1984). However the role ofsemiquinones in binding or toxicity is not yet proved.
1,4-Semiquinone dismutates to form HQ and BQ; BQis reduced to HQ by NADPH-cytochrome reductase(Boersma et al. 1994) or by cytosolic DT-diaphorase(Smart and Zannoni 1984), and chemically by ascorbate(Greenlee et al. 1981), which decreased covalent bindingof [14C]phenol in vitro. HQ or catechol in their reducedform are not toxic (Soucek et al. 1994) and do not bindcovalently to proteins (Smart and Zannoni 1984). BQ(but not HQ) is conjugated with glutathione to yieldmercapturic acid (Nerland and Pierce 1990), destroysproteins including CYP enzymes (Soucek et al. 1994)and its reaction with thiols of tubulin may mediatebenzene myelotoxicity (Irons 1985). BQ formed deox-yguanosine-3′-monophosphate adduct (Jowa et al. 1990)with deoxy-guanosine, characterized as (2′OH) benz-theno (1,N2)guanine (Bauer et al. 1989); Pongracz andBodell (1991) detected 3′-hydroxyl-1,N6-benztheno-2′-deoxyadenosine-3′-phosphate by 32P postlabelling ofDNA reacted with BQ. Trans,trans-muconaldehyde canalso produce toxicity and bind to DNA (Witz et al.1990), but its share in benzene metabolites is low(Schlosser et al. 1993). Lutz and Schlatter (1977) showedthat rats inhaling benzene (3H- and 14C-labelled) devel-oped covalent binding of benzene metabolite(s) to DNAin vivo. DNA adducts were seen after i.p. benzene ad-
Scheme 1 Benzene metabolic pathways (*putative and **proved alkylating metabolites)
46

ministration in rats (Mazzullo et al. 1989), rabbits(Bauer et al. 1989; 1994) and mice (Snyder et al. 1989),but were absent in similar experiments in mice (Reddyet al. 1989; 1994). HQ reacts with O2 forming semi-quinone and superoxide anion; although Boersma et al.(1994) claimed this reaction to require pH 9.85, Lean-derson and Tagesson (1990) observed HQ to form 8-hydroxydeoxyguanosine adduct in vivo and inhibitionby catalase in vitro suggested the role of H2O2.
In CYP active site xenobiotics are oxygenated byoxene, but O2
) or H2O2 leaking CYP may oxidize themoutside the CYP active site. Superoxide anion did notoxidize benzene to phenol, but oxidized phenol to HQand to covalently binding species (Tunek and Oesch1980), apparently due to 2B1 in 2B1-induced but not incontrol microsomes (Smart and Zannoni 1984), whileOH radicals could mediate ∼ 20% of benzene oxidationin 2E1-induced microsomes (Johansson and Ingelman-Sundberg 1988). CYP2E1 forms and leaks unusuallyhigh levels of O2
) and H2O2 (Ekstrom and Ingelman-Sundberg 1989), a mechanism that seems negligible inother CYP forms. Iron redox cycling activates dioxygento superoxide anion and catalyses the Haber-Weiss re-action of O2
) with H2O2 and Fenton reaction withH2O2, both yielding OH radicals. However HQ was re-ported to chelate iron and could therefore inhibit oxygenactivation by iron similarly to EDTA (Tichavska andGut 1994; Singh et al. 1994).
As noted above benzene oxide reportedly binds toguanine N-7 and not to proteins, while BQ binds to allDNA bases and efficiently alkylates proteins. The role ofsemiquinones of di- and trihydroxymetabolites in cova-lent binding is less clear. Although the main CYP formsproducing catechol, HQ and 1,2,4-trihydroxy-benzeneare known, whether they oxidize them further to cova-lently binding semiquinones or quinones in liver (whichperoxidases, absent in liver, oxidize in bone marrow) isnot clear. Moreover, the above-mentioned conflictingdata about oxygen activation by benzene quinones at-tracted our attention, since they mutually interact, pro-duce toxicity and DNA adducts. It is of interest toestimate their importance in benzene toxicity. Our ex-periments were therefore aimed to investigate: (1) themain CYP forms involved in covalent binding of aniline,toluene, MCB and benzene to proteins and to DNA(benzene, MCB); (2) role of semiquinones in covalentbinding of benzene; (3) generation of OH radicals bybenzene quinonic metabolites; and (4) the relation ofcovalent binding to the formation of soluble metabolitesrelevant in health risk assessment, since soluble meta-bolites of benzene derivatives are used to evaluate ex-posure in biological exposure tests, whereas protein orDNA adducts of reactive metabolites are assayed inbiological monitoring to estimate the risk of exposure togenotoxic chemicals (Dashwood and Combes 1987).Benzene was investigated in more detail, since some re-sults showed unusual relation between production ofsoluble and covalently binding products as compared tothe three other chemicals.
Materials and methods
Materials
The chemicals used were of Analar grade and were either fromLabora, Praha, Czech Republic or Sigma Chemical Company, St.Louis, Mo., USA. [14C]Benzene (121 MBq/mmol), [14C-methyl]-toluene, (458 MBq/mmol), [14C]chlorobenzene (202 MBq/mmol)were obtained from Isocommerz, Germany. [14C]Aniline.HCl(490 MBq/mmol) was from Techsnab-export, Russia.
Animals and treatments
Male Wistar rats, SPF, 220–320 g body wt., from Velaz BreedingCo. Prague, were fed ad libitum on Velaz pellet chow and water for1 week before use. CYP2B1 was induced by sodium phenobarbital(PB), 80 mg/kg/day, given i.p. in water (2 ml/kg) for 4 days; ani-mals were killed 24 h after the last dose. CYP2E1 was induced byinhalation of benzene, 4 mg/l, 20 h a day for 4 days in a dynamicinhalation device (Frantik and Kratochvile 1976) as described byGut et al. (1993). Rats were killed immediately after the 4th ex-posure.
Microsomal preparations
The rats were killed by decapitation, livers were rapidly excised,chilled, washed and kept in ice-cold saline until homogenization ina motor-driven Teflon/glass homogenizer in 0.15 M KCl/50 mMTRIS-HCl buffer, pH 7.4. The S9 fraction (9 000 g, 20 min) of liverhomogenate (33.3%) was centrifuged for 60 min at 105 000 g,pellets were resuspended in the KCl/TRIS-HCl buffer and re-cen-trifuged under the same conditions. The pellets were finally re-suspended in the KCl/TRIS buffer with 20% (v/v) glycerol and thesamples stored at )50oC. The microsomal protein was measured(Lowry et al. 1951) using bovine serum albumin as standard. CYPconcentration was estimated according to Omura and Sato (1964).
Immunochemical analysis of CYP levels
Electrophoresis in the presence of SDS was performed as inLaemmli (1970) with 10% acrylamide slab gels and 3 lg protein perwell. Electroblotting was done after Towbin et al. (1979). VariousCYP levels were detected using a polyclonal antiserum (Johanssonet al. 1988; Tindberg and Ingelman-Sundberg 1989). The specificityof antisera was determined by (i) recognition of only one band inWestern blots of microsomes after various pretreatments; and (ii)absence of cross reactivity in Western blot analysis with other CYPforms (1A1/2, 2A1, 2B1/2, 2C3, 2C7, 2C11, 3A1/2) (Ingelman-Sundberg et al. 1988). Anti-rabbit IgG coupled with alkalinephosphatase or horseradish peroxidase was used and data werequantified by densitometry.
Microsomal incubations
In vitro metabolism of toluene, MCB, aniline and benzene wasmeasured in isolated liver microsomes. Incubation mixtures (2 ml),were prepared on ice and included 0.15 M KCl/50 mM TRISpH 7.4, 1 mg/ml microsomal protein, NADPH-generating system(6 mM MgSO4, 4 mM glucose 6-phosphate, 0.6 mM NADP and0.5 I.U./ml glucose 6-phosphate dehydrogenase). Substrate levelswere: aniline or [14C]aniline, 4 mM; [14C]benzene, 0.35 to 2.63 mM;[14C]MCB, 0.6 mM; [14C]toluene, 0.6–0.63 mM. The incubationswere carried out at 37oC in glass-stoppered Erlenmeyer flasks for30 min in a shaking water bath; saturated solutions of the sub-strates in KCl/TRIS buffer were added by a glass-steel syringe, after2 min preincubations of all other components at 37oC. Substratelevels were mostly at enzyme near-saturation conditions. The in-
47

cubations were terminated by chilling the flasks in an ice-cold waterbath.
Determination of water-soluble metabolites
One millilitre of the incubate with [14C]toluene, MCB, aniline orbenzene was made alkaline to a final concentration of 0.25 MNaOH; the remaining substrate was extracted with heptane and[14C]-labelled water-soluble metabolites of benzene, toluene, orMCB were assayed as described (Gut et al. 1993). Radioactivity ofthe substrates (n-heptane extracts) and metabolites (water phases)was assayed by liquid scintillation except the aniline metabolitep-aminophenol, which was assayed photometrically according toGram et al. (1967).
Estimation of covalently-bound metabolites to microsomal proteins
A 1 ml aliquot of the incubate was extracted by ice-cold mixture of97% ethanol/acetone/ethylacetate (1:1:1, v/v). The precipitate wasvortexed, centrifuged (3000 rpm, 10 min) and resuspended fourtimes: no radioactivity above background level was detected insubsequent washes. About 25% protein was lost in the first pre-cipitation step and 1% protein was lost in each of the followingsteps; 99.5% of extractable radioactivity was extracted in the firststep. This procedure gave better results than the paper-filter ortrichloroacetic acid precipitation procedure.
DNA-bound radioactivity
The incubation mixture, described above, was used with 1 mg calfthymus DNA and incubated with [14C]benzene or MCB. DNA waspurified by phenol/chloroform/isoamylalcohol extraction (24:24:1,v/v; Hemminki 1985), precipitated by cold ethanol and diluted withdistilled water. An aliquot was taken for measuring of absorbance;a value at 260:280 nm of 1.75–1.90 showed that DNA was suffi-ciently purified. DNA recovery was 36–46%. Radioactivity of ex-tracted microsomes or purified DNA was assayed with scintillationcocktail SLD31 (Spolana, CR) in Betaszint BF5000 (Berthold-Frieseke, Germany).
Inhibition studies
Monoclonal antibodies (MAb) used were against CYP1A1/2 (clone1-7-1; Park et al. 1982), 2B1 (clone 2-66-3 and 4-29-5; Park et al.1984) and 2C11 (clone 1-68-11, which cross-reacts with 2C12; Parket al. 1989) and a control monoclonal antibody (Hy-Hel) againstchicken lysozyme (used to determine nonspecific reactions). TheMAbs were from the Laboratory of Molecular Carcinogenesis,National Cancer Institute, USA and prepared by a modification ofthe method of Koehler and Milstein (1975).
Chemical inhibition studies
The role of CYP2E1 in benzene oxidation to soluble metabolitesand products binding covalently to proteins was studied usingdiethyldithiocarbamate (DEDTC), which specifically inhibitedCYP2E1 at 0.1–0.3 mM and another 2E1 substrate acrylonitrile.Ascorbate preventing HQ oxidation during incubations (Guen-gerich et al. 1991) enabled us to assess the role of HQ and BQ incovalent binding of benzene metabolites to proteins. A radicalscavenger a-tocopherol helped to estimate the role of benzenesemiquinones.
Analysis of N-7 alkylguanine adducts
For benzene and other chemicals, N-7 alkylguanine (Hemminki1983, 1985) appears to be the main product of interaction with
DNA. Alkylation at the 7-position of guanine labilizes the base fordepurination or imidazole ring-opening (Lawley and Brookes 1963;Moschel et al. 1984; Muller and Eisenbrandt 1985). Neutral hy-drolysis was used for liberating N-7 alkylated guanines (Lawleyand Brookes 1963; Lawley 1976; Vodicka and Hemminki 1988;Hemminki et al. 1989). A 1 ml aliquot of purified alkylated DNA(∼ 0.1 mg) was boiled for 15 min in 10 mM sodium phosphatebuffer, pH 7.0. 7-Alkylguanines were cleaved and remained in thesupernatant after the precipitation of the denatured alkylated DNAby ethanol. The supernatant was evaporated by blowing under N2and diluted with 1 ml distilled water. The above-mentioned scin-tillation assay was used. Results were expressed in nmol/mg DNAper h and as a percentage of total radioactivity covalently bound toDNA.
Estimation of semiquinone radical and hydroxyl radicals
HQ or BQ were dissolved in water and immediately added to mi-crosomal incubates described above according to the conditionsspecified in the figure legend. HQ dissolved in 5 min BQ required30 min to dissolve. ESR spectra were recorded on a ERS-220spectrometer (Academy of Sciences, Berlin, Germany), magneticfield was measured on a H-NMR magnetometer (Radiopan, Po-land), microwave frequency on a frequency counter C3-54 (Unionof Independent States) with the following conditions: microwavepower 60 mW, modulation amplitude 0.02 mT, time constant 0.5 s,scan speed 0.3 mT/min, diphenyl-dipicryl-hydrazyl calibrationstandard (g = 2.0037), temperature 25oC. ESR spectra were re-corded as a 1st (in some cases 2nd) derivation and main parametersas g-factor values, hyperfine coupling constant A, linewidth deltaHpp (peak-to-peak distance) and delta App (peak-to-peak ampli-tude) were calculated according to Kirmse and Stach (1985). Itbecame obvious that semiquinone radical formed signal preventedthe possibility to measure oxygen radical by ESR and anothermethod was used to measure OH radicals; these were assayed bymeasuring formaldehyde formed from dimethyl sulphoxide as de-scribed (Klein et al. 1981; Khan et al. 1990).
Statistical analysis
Statistical analysis was made by the Student’s t-test or by regressionanalysis (formation of 1,4-semiquinone from HQ or BQ). The levelof significance was taken as P < 0.05.
Results
Immunochemically determined CYP2B1and 2E1 levels in liver microsomes
Inhalation of benzene was shown to significantly induceCY2E1 and very slightly 2B1, but decreased theCYP1A1/1A2, 2A1 and 3A1 level in rat liver (Gut et al.1993). Thus, only CYP2B1 and 2E1 were assayed.CYP2B1 was not detectable (Table 1) in controls, wasslightly induced by benzene and markedly by PB.CYP2E1 level was not significantly influenced by PB,but markedly induced by benzene.
48

Effects of CYP2B1 or 2E1 induction on the metabolismof [14C]-labelled toluene, aniline, MCB and benzeneto soluble metabolites and products covalently boundto proteins (non-radioactive anilineto the soluble p-aminophenol)
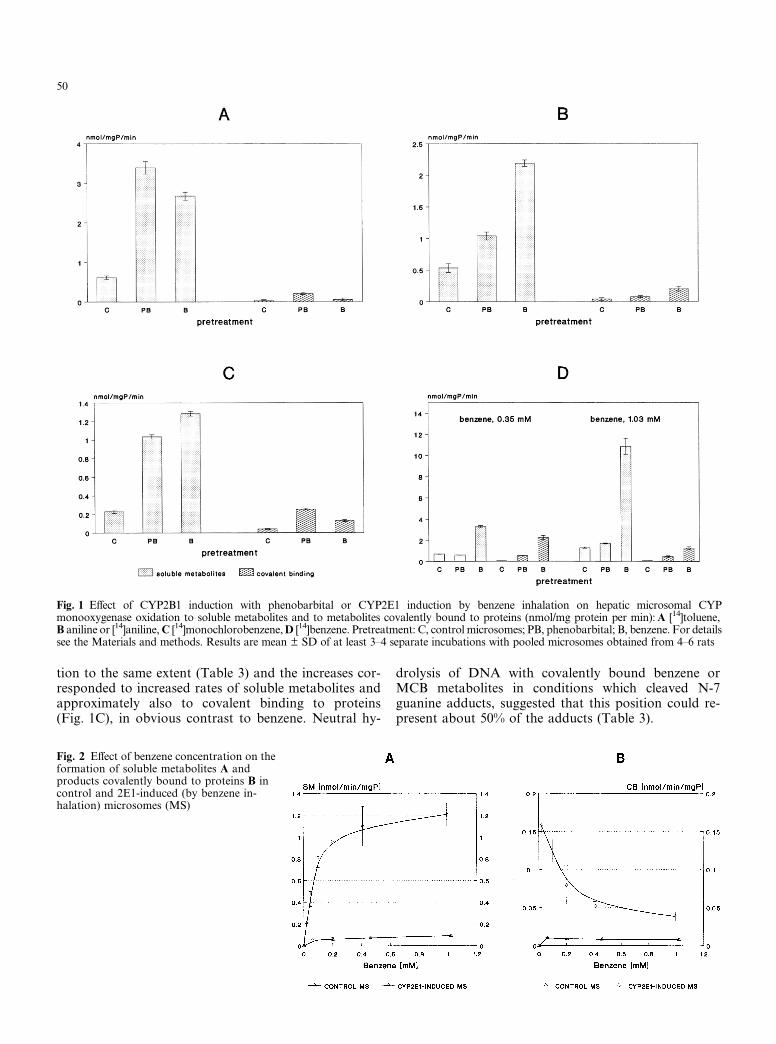
Both CYP2B1 or 2E1 induction caused large increases inthe rates of toluene, aniline and MCB oxidation(Fig. 1A–C, respectively). The proportion of covalentlybound products in total metabolites of aniline was un-changed, but the proportion of reactive metabolites oftoluene in controls (4%) slightly increased by 2B1 in-duction by PB pretreatment (6%) and decreased by 2E1induction by benzene pretreatment (2%). The corre-sponding proportions of reactive metabolites for MCBwere 15%, 20% and 9%, respectively. In contrast,CYP2B1 or 2E1 induction increased the proportion ofbenzene (Fig. 1D) protein-bound products more thantenfold indicating a shift to reactive products. In fact,2B1 induction did not increase the rate of production ofsoluble metabolites at 0.35 or 1.03 mM benzene, whileformation of protein-bound adducts increased tenfold.At these benzene levels, CYP2E1 induction increasedoxidation of benzene to soluble metabolites four- to fivefold, but protein-bound products were 16- to 43-foldhigher (Fig. 1D). Moreover, increasing the benzene leveldecreased the proportion of protein-bound metabolitesfrom 41% at 0.35 mM to 11% at 1 mM benzene(Fig. 1D). This effect of concentration was evident in2E1-induced (by benzene) but not in control microsomes(Fig. 2).
Estimation the role of CYP2B1 and 2E1 in [14C]benzeneand [14C]MCB oxidation by specific antibodiesand specific chemical inhibitors
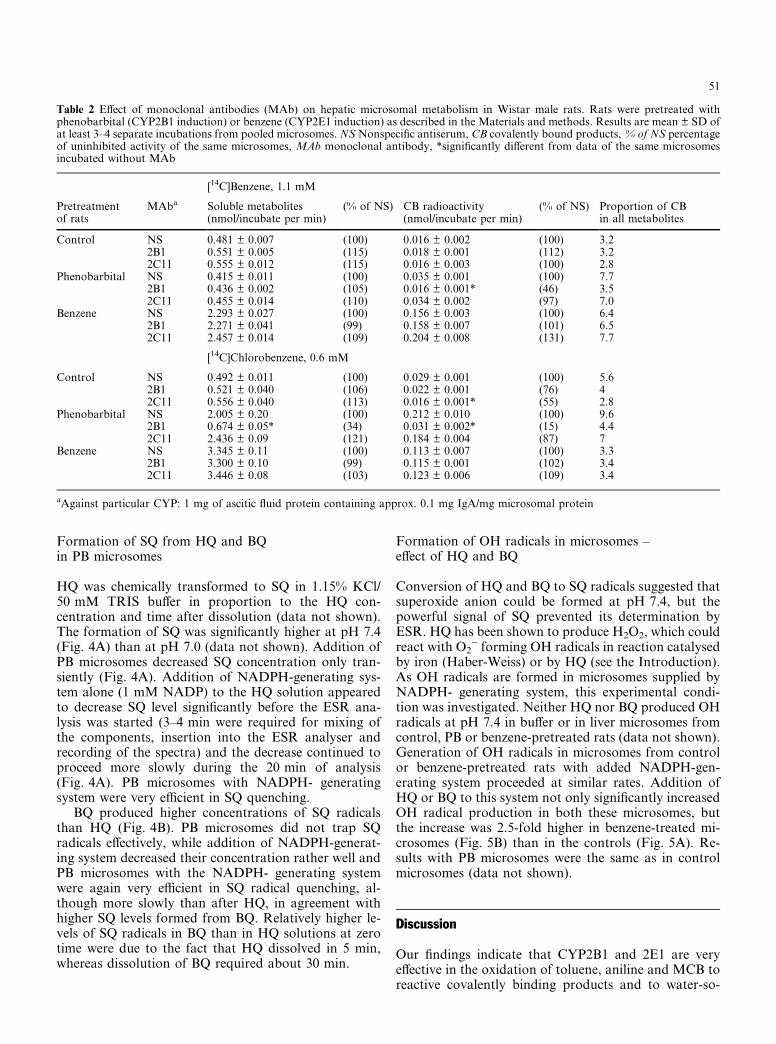
Anti-2B1 MAb did not inhibit benzene oxidation tosoluble or protein-bound metabolites in controls, where2B1 is absent or in 2E1-induced (by benzene) micro-somes, with a low 2B1 level (Table 2). In PB-pretreatedmicrosomes, anti-2B1 MAb did not inhibit the forma-
tion of soluble metabolites (which was not increased by2B1 induction see Table 2, Fig. 1), but decreased theformation of protein-bound products (which 2B1 in-duction increased). Anti-2C11 MAb did not inhibitbenzene oxidation to soluble or covalently bound me-tabolites in the control, 2B1 or 2E1-induced microsomesindicating an insignificant role of this quantitativelyimportant male CYP (Table 2) in benzene oxidation.
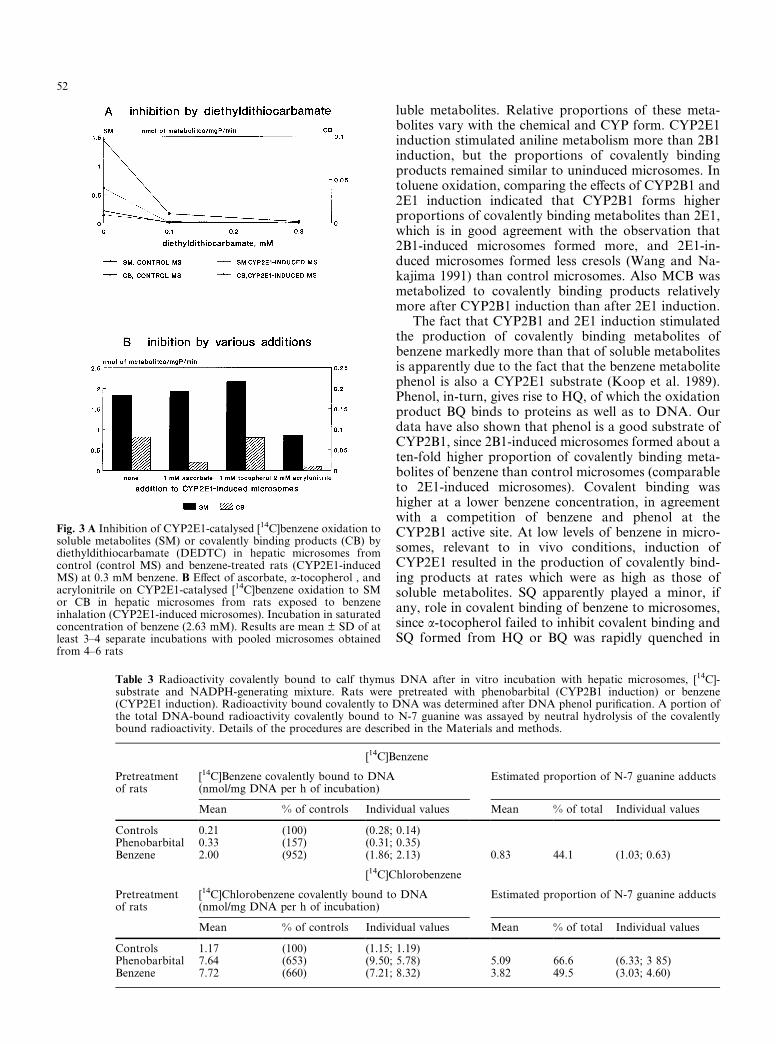
Diethyldithiocarbamate, which specifically inhibitsCYP2E1 at concentrations of 0.1–0.3 mM, almost sup-pressed benzene oxidation to both soluble and protein-bound metabolites in control and benzene-induced mi-crosomes (Fig. 3A) at 0.3 mM benzene, which is satur-ating for CYP2E1, and the inhibition therefore indicatedthat 2E1 was decisive in benzene oxidation at this level.Acrylonitrile (2 mM), a 2E1 substrate, significantly in-hibited benzene oxidation to soluble and even more ef-ficiently to reactive metabolites at a high benzeneconcentration (2.63 mM), indicating that even at thislevel of benzene, CYP2E1 played a decisive role in theproduction of covalently binding metabolites (Fig. 3B).
Inhibition by MAb of [14C]MCB metabolism is in-dicated by the data in Table 2. In PB-treated micro-somes, MAb against CYP2B1 inhibited MCB oxidationto soluble metabolites by 66% and to protein-boundproduct by 85% indicating an essential role of 2B1 inMCB oxidation in PB microsomes. No inhibition wasseen in control or 2E1-induced microsomes by anti-2B1.The anti-2C11 MAb did not inhibit MCB oxidation incontrol, 2B1 or 2E1 induced microsomes indicating aminor role of 2C11 in benzene oxidation.
Estimation of the role of quinonic metabolitesof benzene
Ascorbate, preventing HQ oxidation to BQ, markedlyinhibited oxidation of benzene to covalently boundproducts without changing the formation of solublemetabolites. a-Tocopherol influenced neither covalentlybound products nor soluble metabolites (Fig. 3B), in-dicating that BQ was predominant, whereas 1,4-semi-quinone played a negligible role in covalent binding ofbenzene.
Binding of [14C]benzene and [14C]MCBto calf thymus DNA and estimationof N-7 guanine adducts by neutral hydrolysisof purified DNA
The level of benzene covalent binding to calf-thymusDNA was proportional to the formation of solublemetabolites: no increase after PB-induced 2B1 level anda significant increase after benzene-induced 2E1 (Table 3,and Fig. 1D). This is in contrast to markedly increasedcovalent binding to microsomal proteins after 2B1 in-duction. Microsomal activation of MCB to DNA-boundproducts was elevated by 2B1 induction or 2E1 induc-
Table 1 Immunochemical characterization of CYP1A2, 2B1 and2E1 in liver microsomes, after various pretreatments of rats, usingspecific antibodies, in Western blots. The rats were pretreated withphenobarbital (CYP2B1 induction) or benzene (CYP2E1 induc-tion). The results are mean ± SD of assays in 3–5 different micro-some preparations for each treatment. Controls values, 100% forCYP2E1 and 1A2; phenobarbital-pretreatment microsomes, 100%for CYP2B1. Details of the pretreatments and immunochemicalassays are described in the Materials and methods (N.D. Not de-tectable)
Pretreatment Relative cytochrome CYP level
CYP1A2 CYP2B1 CYP2E1
Control 100 ± 31 N.D. 100 ± 45Phenobarbital 68 ± 24 100 ± 15 78 ± 36Benzene 58 ± 53 24 ± 4 511 ± 196
49
tion to the same extent (Table 3) and the increases cor-responded to increased rates of soluble metabolites andapproximately also to covalent binding to proteins(Fig. 1C), in obvious contrast to benzene. Neutral hy-
drolysis of DNA with covalently bound benzene orMCB metabolites in conditions which cleaved N-7guanine adducts, suggested that this position could re-present about 50% of the adducts (Table 3).
Fig. 1 Effect of CYP2B1 induction with phenobarbital or CYP2E1 induction by benzene inhalation on hepatic microsomal CYPmonooxygenase oxidation to soluble metabolites and to metabolites covalently bound to proteins (nmol/mg protein per min): A [14]toluene,B aniline or [14]aniline, C [14]monochlorobenzene, D [14]benzene. Pretreatment: C, control microsomes; PB, phenobarbital; B, benzene. For detailssee the Materials and methods. Results are mean ± SD of at least 3–4 separate incubations with pooled microsomes obtained from 4–6 rats
Fig. 2 Effect of benzene concentration on theformation of soluble metabolites A andproducts covalently bound to proteins B incontrol and 2E1-induced (by benzene in-halation) microsomes (MS)
50
Formation of SQ from HQ and BQin PB microsomes
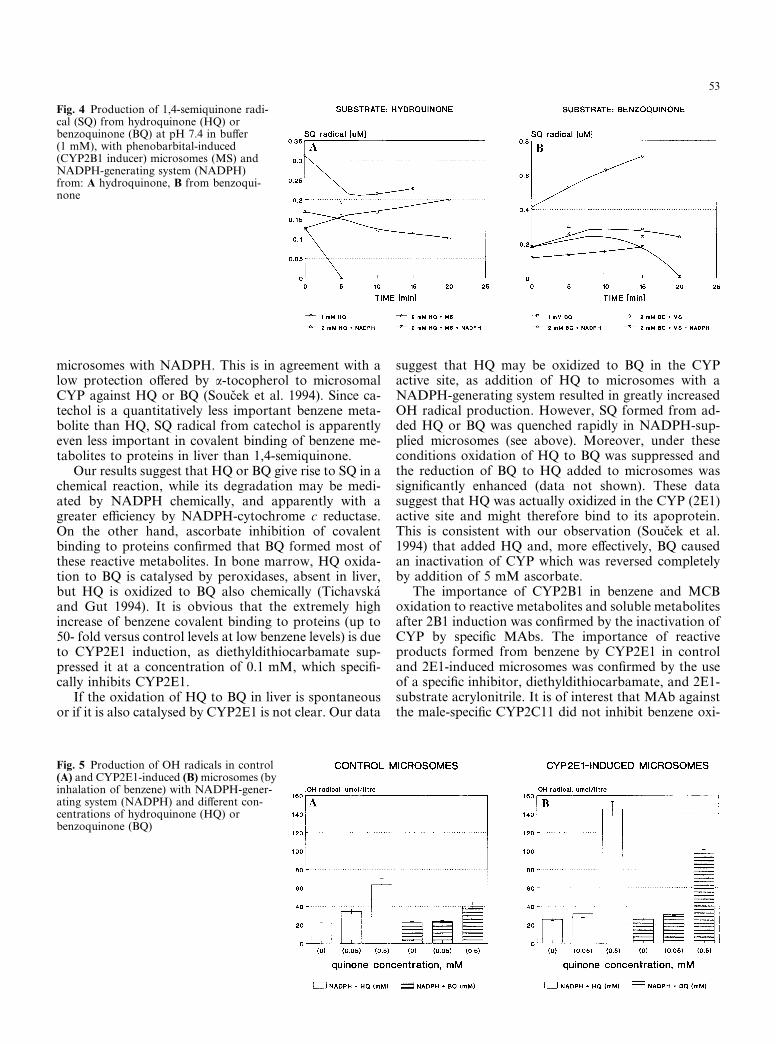
HQ was chemically transformed to SQ in 1.15% KCl/50 mM TRIS buffer in proportion to the HQ con-centration and time after dissolution (data not shown).The formation of SQ was significantly higher at pH 7.4(Fig. 4A) than at pH 7.0 (data not shown). Addition ofPB microsomes decreased SQ concentration only tran-siently (Fig. 4A). Addition of NADPH-generating sys-tem alone (1 mM NADP) to the HQ solution appearedto decrease SQ level significantly before the ESR ana-lysis was started (3–4 min were required for mixing ofthe components, insertion into the ESR analyser andrecording of the spectra) and the decrease continued toproceed more slowly during the 20 min of analysis(Fig. 4A). PB microsomes with NADPH- generatingsystem were very efficient in SQ quenching.
BQ produced higher concentrations of SQ radicalsthan HQ (Fig. 4B). PB microsomes did not trap SQradicals effectively, while addition of NADPH-generat-ing system decreased their concentration rather well andPB microsomes with the NADPH- generating systemwere again very efficient in SQ radical quenching, al-though more slowly than after HQ, in agreement withhigher SQ levels formed from BQ. Relatively higher le-vels of SQ radicals in BQ than in HQ solutions at zerotime were due to the fact that HQ dissolved in 5 min,whereas dissolution of BQ required about 30 min.
Formation of OH radicals in microsomes –effect of HQ and BQ
Conversion of HQ and BQ to SQ radicals suggested thatsuperoxide anion could be formed at pH 7.4, but thepowerful signal of SQ prevented its determination byESR. HQ has been shown to produce H2O2, which couldreact with O2
) forming OH radicals in reaction catalysedby iron (Haber-Weiss) or by HQ (see the Introduction).As OH radicals are formed in microsomes supplied byNADPH- generating system, this experimental condi-tion was investigated. Neither HQ nor BQ produced OHradicals at pH 7.4 in buffer or in liver microsomes fromcontrol, PB or benzene-pretreated rats (data not shown).Generation of OH radicals in microsomes from controlor benzene-pretreated rats with added NADPH-gen-erating system proceeded at similar rates. Addition ofHQ or BQ to this system not only significantly increasedOH radical production in both these microsomes, butthe increase was 2.5-fold higher in benzene-treated mi-crosomes (Fig. 5B) than in the controls (Fig. 5A). Re-sults with PB microsomes were the same as in controlmicrosomes (data not shown).
Discussion
Our findings indicate that CYP2B1 and 2E1 are veryeffective in the oxidation of toluene, aniline and MCB toreactive covalently binding products and to water-so-
Table 2 Effect of monoclonal antibodies (MAb) on hepatic microsomal metabolism in Wistar male rats. Rats were pretreated withphenobarbital (CYP2B1 induction) or benzene (CYP2E1 induction) as described in the Materials and methods. Results are mean ± SD ofat least 3–4 separate incubations from pooled microsomes. NS Nonspecific antiserum, CB covalently bound products, % of NS percentageof uninhibited activity of the same microsomes, MAb monoclonal antibody, *significantly different from data of the same microsomesincubated without MAb
[14C]Benzene, 1.1 mM
Pretreatmentof rats
MAba Soluble metabolites(nmol/incubate per min)
(% of NS) CB radioactivity(nmol/incubate per min)
(% of NS) Proportion of CBin all metabolites
Control NS 0.481 ± 0.007 (100) 0.016 ± 0.002 (100) 3.22B1 0.551 ± 0.005 (115) 0.018 ± 0.001 (112) 3.22C11 0.555 ± 0.012 (115) 0.016 ± 0.003 (100) 2.8
Phenobarbital NS 0.415 ± 0.011 (100) 0.035 ± 0.001 (100) 7.72B1 0.436 ± 0.002 (105) 0.016 ± 0.001* (46) 3.52C11 0.455 ± 0.014 (110) 0.034 ± 0.002 (97) 7.0
Benzene NS 2.293 ± 0.027 (100) 0.156 ± 0.003 (100) 6.42B1 2.271 ± 0.041 (99) 0.158 ± 0.007 (101) 6.52C11 2.457 ± 0.014 (109) 0.204 ± 0.008 (131) 7.7
[14C]Chlorobenzene, 0.6 mM
Control NS 0.492 ± 0.011 (100) 0.029 ± 0.001 (100) 5.62B1 0.521 ± 0.040 (106) 0.022 ± 0.001 (76) 42C11 0.556 ± 0.040 (113) 0.016 ± 0.001* (55) 2.8
Phenobarbital NS 2.005 ± 0.20 (100) 0.212 ± 0.010 (100) 9.62B1 0.674 ± 0.05* (34) 0.031 ± 0.002* (15) 4.42C11 2.436 ± 0.09 (121) 0.184 ± 0.004 (87) 7
Benzene NS 3.345 ± 0.11 (100) 0.113 ± 0.007 (100) 3.32B1 3.300 ± 0.10 (99) 0.115 ± 0.001 (102) 3.42C11 3.446 ± 0.08 (103) 0.123 ± 0.006 (109) 3.4
aAgainst particular CYP: 1 mg of ascitic fluid protein containing approx. 0.1 mg IgA/mg microsomal protein
51
luble metabolites. Relative proportions of these meta-bolites vary with the chemical and CYP form. CYP2E1induction stimulated aniline metabolism more than 2B1induction, but the proportions of covalently bindingproducts remained similar to uninduced microsomes. Intoluene oxidation, comparing the effects of CYP2B1 and2E1 induction indicated that CYP2B1 forms higherproportions of covalently binding metabolites than 2E1,which is in good agreement with the observation that2B1-induced microsomes formed more, and 2E1-in-duced microsomes formed less cresols (Wang and Na-kajima 1991) than control microsomes. Also MCB wasmetabolized to covalently binding products relativelymore after CYP2B1 induction than after 2E1 induction.
The fact that CYP2B1 and 2E1 induction stimulatedthe production of covalently binding metabolites ofbenzene markedly more than that of soluble metabolitesis apparently due to the fact that the benzene metabolitephenol is also a CYP2E1 substrate (Koop et al. 1989).Phenol, in-turn, gives rise to HQ, of which the oxidationproduct BQ binds to proteins as well as to DNA. Ourdata have also shown that phenol is a good substrate ofCYP2B1, since 2B1-induced microsomes formed about aten-fold higher proportion of covalently binding meta-bolites of benzene than control microsomes (comparableto 2E1-induced microsomes). Covalent binding washigher at a lower benzene concentration, in agreementwith a competition of benzene and phenol at theCYP2B1 active site. At low levels of benzene in micro-somes, relevant to in vivo conditions, induction ofCYP2E1 resulted in the production of covalently bind-ing products at rates which were as high as those ofsoluble metabolites. SQ apparently played a minor, ifany, role in covalent binding of benzene to microsomes,since a-tocopherol failed to inhibit covalent binding andSQ formed from HQ or BQ was rapidly quenched in
Fig. 3 A Inhibition of CYP2E1-catalysed [14C]benzene oxidation tosoluble metabolites (SM) or covalently binding products (CB) bydiethyldithiocarbamate (DEDTC) in hepatic microsomes fromcontrol (control MS) and benzene-treated rats (CYP2E1-inducedMS) at 0.3 mM benzene. B Effect of ascorbate, a-tocopherol , andacrylonitrile on CYP2E1-catalysed [14C]benzene oxidation to SMor CB in hepatic microsomes from rats exposed to benzeneinhalation (CYP2E1-induced microsomes). Incubation in saturatedconcentration of benzene (2.63 mM). Results are mean ± SD of atleast 3–4 separate incubations with pooled microsomes obtainedfrom 4–6 rats
Table 3 Radioactivity covalently bound to calf thymus DNA after in vitro incubation with hepatic microsomes, [14C]-substrate and NADPH-generating mixture. Rats were pretreated with phenobarbital (CYP2B1 induction) or benzene(CYP2E1 induction). Radioactivity bound covalently to DNA was determined after DNA phenol purification. A portion ofthe total DNA-bound radioactivity covalently bound to N-7 guanine was assayed by neutral hydrolysis of the covalentlybound radioactivity. Details of the procedures are described in the Materials and methods.
[14C]Benzene
Pretreatmentof rats
[14C]Benzene covalently bound to DNA(nmol/mg DNA per h of incubation)
Estimated proportion of N-7 guanine adducts
Mean % of controls Individual values Mean % of total Individual values
Controls 0.21 (100) (0.28; 0.14)Phenobarbital 0.33 (157) (0.31; 0.35)Benzene 2.00 (952) (1.86; 2.13) 0.83 44.1 (1.03; 0.63)
[14C]Chlorobenzene
Pretreatmentof rats
[14C]Chlorobenzene covalently bound to DNA(nmol/mg DNA per h of incubation)
Estimated proportion of N-7 guanine adducts
Mean % of controls Individual values Mean % of total Individual values
Controls 1.17 (100) (1.15; 1.19)Phenobarbital 7.64 (653) (9.50; 5.78) 5.09 66.6 (6.33; 3 85)Benzene 7.72 (660) (7.21; 8.32) 3.82 49.5 (3.03; 4.60)
52
microsomes with NADPH. This is in agreement with alow protection offered by a-tocopherol to microsomalCYP against HQ or BQ (Soucek et al. 1994). Since ca-techol is a quantitatively less important benzene meta-bolite than HQ, SQ radical from catechol is apparentlyeven less important in covalent binding of benzene me-tabolites to proteins in liver than 1,4-semiquinone.
Our results suggest that HQ or BQ give rise to SQ in achemical reaction, while its degradation may be medi-ated by NADPH chemically, and apparently with agreater efficiency by NADPH-cytochrome c reductase.On the other hand, ascorbate inhibition of covalentbinding to proteins confirmed that BQ formed most ofthese reactive metabolites. In bone marrow, HQ oxida-tion to BQ is catalysed by peroxidases, absent in liver,but HQ is oxidized to BQ also chemically (Tichavskaand Gut 1994). It is obvious that the extremely highincrease of benzene covalent binding to proteins (up to50- fold versus control levels at low benzene levels) is dueto CYP2E1 induction, as diethyldithiocarbamate sup-pressed it at a concentration of 0.1 mM, which specifi-cally inhibits CYP2E1.
If the oxidation of HQ to BQ in liver is spontaneousor if it is also catalysed by CYP2E1 is not clear. Our data
suggest that HQ may be oxidized to BQ in the CYPactive site, as addition of HQ to microsomes with aNADPH-generating system resulted in greatly increasedOH radical production. However, SQ formed from ad-ded HQ or BQ was quenched rapidly in NADPH-sup-plied microsomes (see above). Moreover, under theseconditions oxidation of HQ to BQ was suppressed andthe reduction of BQ to HQ added to microsomes wassignificantly enhanced (data not shown). These datasuggest that HQ was actually oxidized in the CYP (2E1)active site and might therefore bind to its apoprotein.This is consistent with our observation (Soucek et al.1994) that added HQ and, more effectively, BQ causedan inactivation of CYP which was reversed completelyby addition of 5 mM ascorbate.
The importance of CYP2B1 in benzene and MCBoxidation to reactive metabolites and soluble metabolitesafter 2B1 induction was confirmed by the inactivation ofCYP by specific MAbs. The importance of reactiveproducts formed from benzene by CYP2E1 in controland 2E1-induced microsomes was confirmed by the useof a specific inhibitor, diethyldithiocarbamate, and 2E1-substrate acrylonitrile. It is of interest that MAb againstthe male-specific CYP2C11 did not inhibit benzene oxi-
Fig. 4 Production of 1,4-semiquinone radi-cal (SQ) from hydroquinone (HQ) orbenzoquinone (BQ) at pH 7.4 in buffer(1 mM), with phenobarbital-induced(CYP2B1 inducer) microsomes (MS) andNADPH-generating system (NADPH)from: A hydroquinone, B from benzoqui-none
Fig. 5 Production of OH radicals in control(A) and CYP2E1-induced (B) microsomes (byinhalation of benzene) with NADPH-gener-ating system (NADPH) and different con-centrations of hydroquinone (HQ) orbenzoquinone (BQ)
53

dation in control microsomes, despite the fact that thisCYP form makes up about 50% of all liver P450s(Guengerich et al. 1991). CYP2C11 also did not catalyseMCB oxidation to soluble metabolites, but played somerole in MCB activation to covalently binding products(probably by further MCB oxidation to reactive qui-nones). Benzene oxide apparently alkylated calf thymusDNA, as indicated by neutral hydrolysis of DNA-boundradioactivity, in agreement with the observation that N-7 guanine adduct of benzene oxide was found in urine(Mueller et al. 1987). Our data (relative effects of 2B1and 2E1 induction and of inhibitors on soluble meta-bolites, products covalently bound to proteins and toDNA) indicated that BQ formed most of the proteincovalent adducts of benzene, whereas a large part ofDNA adducts came from benzene oxide. Benzene oxideis the obligatory intermediate binding apparently toDNA (Mueller et al. 1987), but not to proteins (Tunek etal. 1978). BQ binds to proteins as well as to DNA as wasalso demonstrated. The other potentially covalentlybinding benzene metabolites appear to play a minor rolein liver, since they are formed at much lower rates (ca-techol, muconaldehyde, 1,2,4-trihydroxybenzene, Sch-losser et al. 1993; Seaton et al. 1994).
Proportional increases in covalent binding of MCB toDNA and formation of soluble metabolites after in-duction of 2B1 or 2E1 suggest that the same metabolitesare involved. 4-Chlorocatechol is a minor (1.6%) meta-bolite of MCB in rats, whereas MCB-oxide could beresponsible for the binding (by analogy with benzene-oxide) as its products constitute 70% of MCB dose invivo (Yoshida and Hara 1985; Sullivan et al. 1983). Inaddition, the hydrolysis of radioactivity of MCB-alky-lated DNA shows N-7 guanine adduct to representabout 50% of the adducts.
The high proportion of reactive products in CYP2B1and 2E1-catalysed oxidation of benzene (particularly atlow substrate levels) suggest that phenol elimination inurine, used as a biological exposure test, may not reflectactual levels of reactive metabolites in the body. The factthat covalent binding of benzene to proteins is >100-foldhigher than to DNA complies with difficulties of de-termining DNA adducts of benzene in tissues and withthe view that benzene is an epigenetic carcinogen (IARC1987). On the other hand the MCB covalent binding toproteins was only >10-fold higher than to DNA.
The interactions between HQ, BQ and NADPH arevery complex, but some conclusions may be drawn. Thespontaneously formed SQ from HQ or BQ was some-what quenched by microsomes themselves, and veryeffectively after addition of NADPH. Although Boers-ma et al. (1994) claimed that SQ formation from HQrequired alkaline pH (9.85), we observed SQ formationat pH 7.4. However, effective SQ quenching in micro-somes with NADPH suggests that the SQ role in HQ orBQ toxicity is low. Boersma et al. (1994) also reportedthat HQ did not produce O2
) at physiological pH, butLeanderson and Tagesson (1990) report that HQ re-acted with DNA formed 8-OH-deoxyguanosine adduct
in vitro. Since this reaction was greatly inhibited byaddition of catalase, H2O2 (the O2
) product) was alsocertainly formed. Our observation that HQ and BQthemselves did not form .OH radicals in buffer or mi-crosomes suggested that although H2O2 may have beenformed, its subsequent (Haber-Weiss) reaction with O2
)
did not proceed to form significant amounts of .OHradicals. However, the NADPH-mediated .OH radicalproduction in microsomes was greatly stimulated byHQ and BQ indicating that with the physiologicallyavailable NADPH and at physiological pH, HQ or BQcould enhance activation of oxygen. It could be due toHQ-induced stimulation of the CYP-futile cycle (Man-nering 1980). Since .OH radicals effectively oxidizephenol (see the Introduction), HQ, by inducing .OHradical production in microsomes (endoplasmic re-ticulum) with NADPH, could contribute to oxidation ofphenol to HQ, whose oxidation product BQ alkylatesDNA and proteins. The greatly enhanced bone marrowtoxicity of phenol and HQ administered simultaneouslyto animals (Eastmond et al. 1980) could be due to thismechanism.
Acknowledgements The specific anti-P450 antibodies for im-munodetection were prepared in Berzelius Laboratory, KarolinskaInstitutet, Stockholm, where the immunodetection of individualP450 enzymes in Western blots was also made. This part of thework was supported by the Swedish Medical Research Council.Other financial support was obtained from the Internal GrantAgency and Ministry of Health of the Czech Republic. The authorswish to thank Martina Janouskova for drawing the metabolicscheme of benzene oxidation. Acknowledgement is also made tothe elaboration of a special programme and the calculation of theregression equations of semiquinone concentrations in Fortran byZdenek Roth.
References
Bauer H, Dimitriados EA, Snyder R (1989) An in vivo study ofbenzene metabolite DNA adduct formation in liver of maleNew Zealand rabbits. Arch Toxicol 63: 209–213
Boersma MG, Balvers WG, Boeren S, Vervoort J, Rietjens IM(1994) NADPH- cytochrome reductase catalyzed redox cyclingof 1,4-benzoquinone; hampered at physiological conditions,initiated at increased pH values. Biochem Pharmacol 47: 1949–1955
Dashwood RH, Combes RD (1987) Deficiencies in the covalentbinding index (CBI) for expressing in vivo binding to DNA withrespect to predicting chemical carcinogenicity. A proposal for atarget-organ binding index. Mutat Res 190: 173–175
Driscoll KE, Snyder CA (1984) The effect of ethanol ingestion andrepeated benzene exposures on benzene pharmacokinetics.Toxicol Appl Pharmacol 73: 525–532
Eastmond DA, Smith MT, Irons RD (1987) An interaction ofbenzene metabolites reproduces the myelotoxicity observedwith benzene exposure. Toxicol Appl Pharmacol 91: 85–95
Ekstrom G, Ingelman-Sundberg M (1989) Rat liver microsomalNADPH- supported oxidase activity and lipid peroxidationdependent on ethanol- inducible cytochrome P-450 (P-450IIE1). Biochem Pharmacol 38: 313–319
Frantık E, Kratochvıle K (1976) In: Horvath M (ed) Adverse ef-fects of environmental chemicals and psychotropic drugs.Elsevier, Amsterdam, p 321
Gilmour SK, Kalf GF, Snyder R (1986) Comparison of the me-tabolism of benzene and its metabolite phenol in rat liver. AdvExp Med Biol 197: 223
54

Gonasun LM, Witmer C, Kocsis JJ, Snyder R (1973) Benzenemetabolism in mouse liver microsomes. Toxicol Appl Phar-macol 26: 398–406
Gram TE, Rogers LA, Fouts JR (1967) Further studies on themetabolism of drugs by subfractions of hepatic microsomes. JPharmacol Exp Ther 155: 479–93
Greenlee WF, Sun JD, Bus JS (1981) A proposed mechanism ofbenzene toxicity: formation of reactive intermediates frompolyphenol metabolites. Toxicol Appl Pharmacol 59: 187–195
Guengerich FP, Kim D-H, Iwasaki M (1991) Role of human cy-tochrome P450 IIE1 in oxidation of low molecular weightsuspects. Chem Res Toxicol 2: 168–179
Gut I (1976) Effect of phenobarbital pretreatment on in vitro en-zyme kinetics and in vivo biotransformation of benzene in therat. Arch Toxicol 35: 195–206
Gut I (1983) Influence of frequently used industrial solvents andmonomers of plastics on xenobiotic metabolism. ZentralblPharm 122: 1139–1162
Gut I, Terelius Y, Frantık E, Linhart I, Soucek P, Filipcova B,Kluckova H (1993) Exposure to various benzene derivativesdifferently induces cytochromes P450 2B1 and P450 2E1 in ratliver. Arch Toxicol 67: 237–243
Hemminki K (1983) Nucleic acid adducts of chemical carcinogensand mutagens. Arch Toxicol 52: 249–285
Hemminki K (1985) Binding of metabolites of cyclophosphamideto DNA in rat liver microsomal system and in vivo in mice.Cancer Res 45: 4237–4243
Hemminki K, Peltonen K, Vodicka P (1989): Depurination fromDNA of 7-methylguanine, 7-(2-aminoethyl)-guanine and ring-opened 7-methylguanines. Chem Biol Interact 70: 289–303
IARC monographs on the evaluation of carcinogenic risk to hu-mans. An updating of IARC monographs (1987) vol 1–42,Suppl 7, Lyon, France
Ingelman-Sundberg M, Johansson I, Pentilla K, Glaumann H,Lindros KO (1988) Centrilobular expression of ethanol-in-ducible cytochrome P-450 (IIE1) in rat liver. Biochem BiophysRes Commun 157: 55–60
Ingelman-Sundberg M, Johansson I, Ekstrom G, Cronholm T,Norsten C, Eliasson E, Terrelius Y, Tindberg N, Ronis MJJ,Handler JA, Thurman RF, Hohansson A, Hagbjork AL (1989)Ethanol-inducible cytochrome P450 (IIE1) in alcohol-mediatedtoxicity. Adv Biosci 76: 255–280
Irons RD (1985) Quinones as toxic metabolites of benzene.J Toxicol Environ Health 16: 673–678
Johansson I, Ingelman-Sundberg M (1988) Benzene metabolism byethanol, acetone and benzene-inducible cytochrome P-450(IIE1) in rat and rabbit liver. Cancer Res 48: 5387–5390
Johansson I, Ekstrom G, Scholte B, Puzycki D, Jornvall H, In-gelman- Sundberg M (1988) Ethanol-, fasting-, and acetone-inducible cytochrome P-450 in rat liver: regulation and char-acteristics of enzymes belonging to the IIB and IIE gene sub-families. Biochemistry 27: 1925–1934
Jowa L, Witz G, Snyder R, Sinkle S, and Kalf GF (1990): Synthesisand characterization of deoxyguanosine-benzoquinone adducts.J Appl Toxicol 10: 47–54
Khan S, Krishnamurthy R, Padnya KP (1990) Generation of hy-droxyl radicals during benzene toxicity. Biochem Pharmacol 39:1393–1395
Kirmse R, Stach S (1985) ESR Spektroskopie. Akademie Verlag,Berlin Germany, p 123
Klein SM, Cohen G, Cederbaum AI (1981) Production of for-maldehyde during metabolism of dimethylsulfoxide by hydro-xyl radical generating systems. Biochemistry 20: 6006–6012
Koehler D, Milstein C (1975) Continuous cultures of fused cellssecreting antibody of predefined specificity. Nature 265: 495–497
Koop DR, Laethem CL, Schnier GG (1989) Identification ofethanol-inducible P450 isozyme 3a (P450IIE1) as a benzene andphenol hydroxylase. Toxicol Appl Pharmacol 98: 278–288
Korzekwa KR, Swinney DC, Trager WF (1989) Isotopically la-beled chlorobenzenes as probes for the mechanism of cyto-chrome P450 catalyzed aromatic hydroxylation. Biochemistry28: 9019–9027
Laemmli UK (1970): Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227: 680–685
Lawley PD (1976) Screening tests in chemical carcinogenesis.IARC Sci Publ 12: 181–208
Lawley PD, Brookes P (1963) Further studies on the alkylation ofnucleic acids and their constituent nucleotides. Biochem J 89:127–138
Leanderson P, Tagesson C (1990) Cigarette smoke-induced DNAdamage: role of hydroquinone and catechol in the formation ofthe oxidative DNA adduct 8-hydroxydeoxyguanosine. ChemBiol Interact 74: 71–81
Lee EW, Kocsis JJ, Snyder R (1981) The use of ferrokinetics in thestudy of experimental anemia. Environ Health Perspect 39: 29–37
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Proteinmeasurement with the Folin phenol reagent. J Biol Chem 193:265–275
Lutz WK, Schlatter CH (1977) Mechanism of the carcinogenicaction of benzene: irreversible binding to liver DNA. Chem BiolInteract 18: 241–245
Mannering GJ (1980) Hepatic cytochrome P-450-linked drug-me-tabolizing systems. In: Jenner P, Testa B (eds) Concepts in drugmetabolism, Part B. Marcel Dekker, New York, pp 53–166
Mazzullo M, Bartoli S, Bonora B, Colacci A, Grilli S, Lattanzi G,Niero A, Turina MP, Parodi S (1989) Benzene adducts with ratnucleic acids and proteins: dose-response relationship aftertreatment in vivo. Environ Health Perspect 82: 259–266
Moschel RC, Hudgins WR, Dipple A (1984) Substituent-inducedeffects on the stability of benzylated guanosines: model systemfor factors influencing stability of carcinogen-modified nucleicacid. J Org Chem 49: 363–372
Mueller G, Koelbel M, Heger M, Norpoth K (1987) Urinary S-phenyl-mercapturic acid and phenylglucuronide as indicators ofbenzene exposure. In: Ho MH, Dillon HK (eds) Biologicalmonitoring of exposure to chemicals. organic compounds. JohnWiley and Sons, New York, pp 91–98
Muller N, Eisenbrandt G (1985) The influence of N-7 substituentson the stability of N-7 alkylated guanosines. Chem Biol Interact53: 173–181
Nakajima T, Okuyama S, Yonekura I, Sato A (1985) Effects ofethanol and phenobarbital administration on the metabolismand toxicity of benzene. Chem Biol Interact 55: 23–38
Najakima T, Elovaara E, Park SS, Gelboin HV, Hietanen E,Vainio H (1989) Immunochemical characterization of cyto-chrome P450 isozyme responsible for benzene oxidation in ratliver. Carcinogenesis 10: 1713–1717
Nakajima T, Wang R-S, Elovaara E, Park SS, Gelboin HV, Hie-tanen E, Vainio H (1991) Monoclonal antibody-directed char-acterization of cytochrome P450 isozymes responsible fortoluene metabolism in rat liver. Biochem Pharmacol 41: 395–404
Nedelcheva V, Gut I (1994) Cytochromes P450 in rats and humans:methods of investigation, substrate specificities and relevance tocancer. Xenobiotica 24: 1151–1175
Nerland DE, Pierce WM (1990) Identification of N-acetyl-S-(di-hydroxy-hexyl)-L-cysteine as a urinary metabolite of benzene.Drug Metab Dispos 18: 958–961
NIOSH (1987) Registry of toxic effects of chemical substances,1985–1988 edn (ed Seet DW). USA PHS Center for DiseaseControl, Washington
Omura T, Sato R (1964) The carbon-monoxide-binding pigment ofliver microsomes. II. Solubilization, purification, properties.J Biol Chem 239: 2379–2385
Park SS, Fujino T, West D, Guengerich FP, Gelboin HV (1982)Monoclonal antibody that inhibits enzyme activity of 3-me-thylcholanthrene induced cytochrome P-450. Cancer Res 42:1798–1808
Park SS, Fujino T, Miller H, Guengerich FP, Gelboin HV (1984)Monoclonal antibodies against phenobarbital-induced rat livercytochrome P-450. Biochem Pharmacol 33: 2071–2081
Park SS, Waxman DJ, Lapenson DP, Schenkman JB, Gelboin HV(1989) Monoclonal antibodies to rat liver cytochrome P-450 2c/RLM5 that regiospecifically inhibit steroid metabolism. Bio-chem Pharmacol. 38: 3067–3074
55

Pathiratne A, Puyear RL, Brammer JD (1986) Activation of 14C-toluene to covalently binding metabolites by rat liver micro-somes. Drug Metab Dispos 14: 386–391
Pongracz K, Bodell WJ (1991) Detection of 3′-hydroxyl-1,N6-benztheno-2′-deoxyadenosine 3′-phosphate by 32P postlabelingof DNA reacted with p-benzoquinone. Chem Res Toxicol 4:199–202
Post GB, Snyder R (1983) Effect of enzyme induction on micro-somal benzene metabolism. J Toxicol Environ Health 11: 811–825
Reddy MV, Blackburn GR, Schreiner CA, Mehlman MA, Mack-erer CR (1989) 32P analysis of DNA adducts in tissues ofbenzene-treated rats. Environ Health Perspect 82: 253–268
Reddy MV, Schultz SC, Blackburn GR, Mackerer CR (1994) Lackof DNA adduct formation in mice treated with benzene. MutatRes 325: 149–155
Schlosser PM, Bond JA, Medinsky MA (1993) Benzene and phenolmetabolism by mouse and rat liver microsomes. Carcinogenesis14: 2477–2486
Schrenk D, Ingelman-Sundberg M, Bock KW (1992) Influence ofP-450 2E1 induction on benzene metabolism in rat hepatocytesand on biliary metabolite excretion. Drug Metab Dispos 20:137–141
Seaton MJ, Schlosser PM, Bond JA, Medinsky MA (1994) Benzenemetabolism by human liver microsomes in relation to cyto-chrome P450 2E1 activity. Carcinogenesis 15: 1799–1806
Singh V, Almad S, Rao GS (1994) Prooxidant and antioxidantproperties of iron-hydroquinone and iron-1,2,4-benzenetrilcomplex. Toxicology 89: 25–33
Smart RC, Zannoni VC (1984) DT-Diaphorase influences thecovalent binding of the metabolites of phenol, the major me-tabolite of benzene. Mol Pharmacol 26: 105–111
Snyder R, Uzuki D, Gonasun L, Bromfeld E, Wells A (1967) Themetabolism of benzene in vitro. Toxicol Appl Pharmacol 11:346–360
Snyder R, Dimitriadis E, Guy R, Hu P, Cooper K, Bauer H,Witz G, Goldstein BD (1989) Studies on the mechanism ofbenzene toxicity. Environ Health Perspect 82: 31–35
Snyder R, Witz G, Goldstein BD (1993) The toxicology of benzene.Environ Health Perspect 100: 293–306
Soucek P, Gut I (1992) Cytochromes P450 in rats: structures,functions and relevant human forms. Xenobiotica 22: 83–103
Soucek P, Tichavska B, Gut I (1994) Cytochrome P450 destructionand radical scavenging by benzene and its metabolites. Evidencefor the key role of quinones. Biochem Pharmacol 47: 2233–2242
Sullivan TM, Born GS, Carlson GP, Kessler WV (1983) Thepharmacokinetics of inhaled chlorobenzene in the rat. ToxicolAppl Pharmacol 91: 194–203
Tichavska B, Gut I (1994) Metabolism of reactive oxygen species[in Czech]. Chemicke Listy 88: 580–590
Tindberg N, Ingelman-Sundberg M (1989) Cytochrome P-450 andoxygen toxicity. Oxygen-dependent induction of ethanol-in-ducible cytochrome P-450 in rat liver and lung. Biochemistry28: 4499–4504
Towbin H, Stachelin T, Gordon J (1979) Electrophoretic transferof proteins from polyacrylamide gels to nitrocellulose: proce-dure and some applications. Proc Natl Acad Sci USA 76: 4350–4354
Tunek A, Oesch F (1980) Multi-step metabolic activation of ben-zene in rat liver microsomes. In: Snyder R et al. (eds) Biologicalreactive intermediates II. Chemical mechanisms and biologicaleffects, Part A. Plenum, New York London, pp 319–329
Tunek A, Platt KL, Bentley P, Oesch F (1978) Microsomal meta-bolism of benzene to species irreversibly binding to microsomalprotein and effect of modifications of this metabolism. MolPharmacol 14: 920–929
Vodicka P, Hemminki K (1988) Identification of alkylation pro-ducts of styrene oxide in single and double-stranded DNA.Carcinogenesis 9: 1657–1660
Wang R-S, Nakajima T (1991) Kinetic studies on toluene meta-bolism in ethanol- and phenobarbital-induced rat liver micro-somes in vitro. Arch Toxicol 65: 39–44
Witz G, Gad SC, Tice RR, Oshiro Y, Piper CE, Goldstein BD(1990) Genetic toxicity of the benzene metabolite trans,trans-muconaldehyde in mammalian and bacterial cells. Mutat Res240: 295–306
Yoshida M, Hara I (1985) Composition of urinary metabolites andvariation of urinary taurine levels in rats injected with chlor-obenzene. Indust Health 23: 239–243
56




















