
ORP2, a homolog of oxysterol binding protein, regulates cellular cholesterol metabolism
Upload
independentCategory
view
4download
0
10.1128/JB.187.7.2439-2447.2005.
2005, 187(7):2439. DOI:J. Bacteriol. Lau-Bonilla, William B. Davis and Anthony G. GarzaWhiteford, Sophia A. Pacheco, Olaus J. Walsh, Dalia John L. Dahl, Kriti Arora, Helena I. Boshoff, Danelle C. Viability, and Gene Expression
Affects Cell Appearance,smegmatisMycobacterium Homolog of relAThe
http://jb.asm.org/content/187/7/2439Updated information and services can be found at:
These include:
REFERENCEShttp://jb.asm.org/content/187/7/2439#ref-list-1at:
This article cites 36 articles, 25 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
on June 12, 2014 by guest
http://jb.asm.org/
Dow
nloaded from

JOURNAL OF BACTERIOLOGY, Apr. 2005, p. 2439–2447 Vol. 187, No. 70021-9193/05/$08.00�0 doi:10.1128/JB.187.7.2439–2447.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
The relA Homolog of Mycobacterium smegmatis Affects CellAppearance, Viability, and Gene Expression
John L. Dahl,1* Kriti Arora,1 Helena I. Boshoff,2 Danelle C. Whiteford,1Sophia A. Pacheco,1 Olaus J. Walsh,1 Dalia Lau-Bonilla,1
William B. Davis,1 and Anthony G. Garza3
School of Molecular Biosciences, Washington State University, Pullman, Washington1; Tuberculosis ResearchSection, National Institute of Allergy and Infectious Disease, Rockville, Maryland2; and
Department of Biology, Syracuse University, Syracuse, New York3
Received 17 November 2004/Accepted 14 December 2004
The modification of metabolic pathways to allow for a dormant lifestyle appears to be an important featurefor the survival of pathogenic bacteria within their host. One regulatory mechanism for persistent Mycobac-terium tuberculosis infections is the stringent response. In this study, we analyze the stringent response of anonpathogenic, saprophytic mycobacterial species, Mycobacterium smegmatis. The use of M. smegmatis as a toolfor studying the mycobacterial stringent response was demonstrated by measuring the expression of two M.tuberculosis genes, hspX and eis, in M. smegmatis in the presence and absence of relMsm. The stringent responseplays a role in M. smegmatis cellular and colony formation that is suggestive of changes in the bacterial cell wallstructure.
The ability of Mycobacterium tuberculosis to persist in thehuman host is a major challenge for both vaccine- and drug-based strategies for controlling the spread of tuberculosis (TB)(17). Persistent M. tuberculosis cells are capable of initiatingactive growth in the host, a condition known as reactivationTB. Although it is somewhat controversial, it is generally be-lieved that the site of viable M. tuberculosis in the host is insidecaseous, necrotic, granulomatous lesions in the lungs. It hasbeen reported that M. tuberculosis cells persisting inside gran-ulomas lack the acid-fast staining characteristic of bacteriarecovered from sputa and lesions of patents with active disease,indicating that changes occur in the bacterial cell wall duringmetabolic adaptation to a state of dormancy in the host (18,22). Various models have been developed to create in vitrogrowth conditions that mimic the presumed state of M. tuber-culosis inside granulomas, including oxygen limitation (32, 33)and nutrient starvation (1, 20). For example, when M. tuber-culosis cultures are suspended in distilled water, they appear tolose their acid-fast staining ability, but the cells can remainviable for over 2 years in this extreme nutrient-deprived envi-ronment (22). This implies that in vitro observations of myco-bacterial physiology may provide important insights into un-derstanding how pathogenic mycobacteria survive in hosts. Inaddition, several mutants of M. tuberculosis that are impairedin de novo biosynthesis of various amino acids and vitamins(14) are attenuated in mouse models of tuberculosis infection,suggesting that the organism resides in a nutrient-poor envi-ronment. This supports the significance of in vitro nutrientstarvation models for understanding in vivo persistence.
Recently, the stringent response of M. tuberculosis has been
shown to play an important role in the in vitro and in vivosurvival of this bacterium (9, 29). Escherichia coli has twohomologous genes that are responsible for initiating the strin-gent response, namely, relA and spoT. Collectively, RelA andSpoT can sense nutrient deprivation and respond by synthe-sizing guanosine tetraphosphate (ppGpp) and guanosine pen-taphosphate (pppGpp), which can alter the promoter specific-ity of RNA polymerase. In mycobacteria, however, there isonly one homologous gene, which by convention is referred toas relA. Deleting relMtb renders the tubercle bacillus less capa-ble of surviving nutrient deprivation, and the mutant is alsounable to persist in a mammalian host, in contrast to theisogenic wild-type strain (9, 29).
This report characterizes the stringent response of Mycobac-terium smegmatis, a nonpathogenic, fast-growing species whichis widely recognized as an excellent model system for studyingvarious aspects of M. tuberculosis biology, such as gene expres-sion (16), cell structure (7), and persistence in the face ofnutrient starvation (31). The M. smegmatis stringent responsewas previously analyzed by overexpression of the relA gene ofE. coli or relMtb in M. smegmatis (23). These strains producedelevated intracellular levels of ppGpp that led to altered cellmorphologies and a modest decrease in the cell doubling time.In this study, we examine the effects of deleting the native relAhomolog, relMsm, from M. smegmatis.
MATERIALS AND METHODS
Strains, media, and growth conditions for bacteria. M. smegmatis (strainmc2155) was grown in 7H9 (Difco) medium supplemented with 0.2% glyceroland 0.05% Tween 80. M. tuberculosis cells (strains H37Rv and H37Rv �relMtb)(29) were grown in 7H9 with 10% Middlebrook ADC (VWR), 0.2% glycerol, and0.05% Tween 80. M. smegmatis �relMsm was generated by allelic exchange.Briefly, M. smegmatis genomic DNA was digested with EcoRI, and fragments inthe size range of 4 to 4.5 kb were cloned into pGEM3Zf(�). Colony blottingidentified a clone (pGmsrelA) with a 4,229-bp EcoRI fragment containing the M.smegmatis relMsm gene. An internal BamHI-BglII fragment was replaced with theTn903 aph cassette, removing 387 bp from the central region of the relMsm gene
* Corresponding author. Mailing address: School of Molecular Bio-sciences, Washington State University, Science Hall, Room 301, Pull-man, WA 99164. Phone: (509) 335-7719. Fax: (509) 335-1907. E-mail:[email protected].
2439
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

and leaving 991 and 1,021 bp of the open reading frame on the 5� and 3� ends ofthe insertion, respectively. A PAg85-lacZ Phsp60sacB cassette (25) was cloned intothe XbaI site of this plasmid to create pGmsrelAKO. M. smegmatis was electro-porated, and mutants were selected as previously described (3). DNA sequenceanalysis of the M. smegmatis genome indicated that the gene immediately down-stream of relMsm is oriented in the opposite direction with respect to relMsm
transcription, suggesting that it is highly unlikely that the insertion in relMsm
inhibits transcription of this downstream gene via a polar effect. Therefore, thechances of a polar effect resulting from this deletion of relMsm are minimal.
Southern blot analysis. Southern blot analysis of the M. smegmatis relMsm genewas performed as previously described (30). Genomic DNAs were isolated fromwild-type M. smegmatis and from the candidate �relMsm strain as previouslydescribed (2). Genomic DNAs were first digested with ApaI for 24 h before thebuffer was adjusted and they were further digested with EcoRV for 24 h. The1-kb region downstream of the M. smegmatis relMsm gene was amplified by a PCRwith 5�-CATCTTAAGCGCACTGTTCGTCGTGTGGGC-3� and 5�-TGCTCTAGAAGCGCGGCCTGATCGAGCG-3� as forward and reverse primers, re-spectively. This PCR product was gel purified and used as a template in a PCRinvolving random oligonucleotide primers and [32P]CTP.
RNA extraction and analysis. Fifty-milliliter quantities of M. smegmatis strainswere grown to stationary phase (4 days in 7H9 medium with 0.05% Tween 80)before centrifugation. Cell pellets were resuspended in 1 ml of Trizol and lysedby the use of 0.1-mm-diameter glass beads (Biospec Products, Inc.) and a Fast-Prep FP120 bead-beating device (Bio 101). The cells were disrupted three timesat a speed setting of 6.5 for 45 s, with intermittent incubation on ice for 5 min.Lysed samples were centrifuged at 12,000 � g for 10 min to remove cellulardebris, and the supernatant was transferred to RNase-free 1.5-ml tubes contain-ing 300 �l of chloroform-isoamyl alcohol (24:1) and phase-lock gel (Sigma). Thesolution was mixed by inversion for 2 min at room temperature before beingcentrifuged at 12,000 � g for 10 min. The upper aqueous layer was removed andadded to 600 �l of isopropanol in a fresh tube, which was inverted several timesand incubated at 4°C overnight. Nucleic acids were pelleted by centrifugation at12,000 � g for 10 min and were washed with 70% ethanol. After the pellets wereair dried for 10 min, they were redissolved in 70 �l of RNase-free water. RNAsamples were mixed with an ethidium bromide-containing loading dye and sep-arated in 1.5% formaldehyde–agarose.
Measurement of (p)ppGpp. Three strains were examined for the induction ofradiolabeled (p)ppGpp synthesis, namely, E. coli JM105, wild-type M. smegmatis,and M. smegmatis �relMsm. Cells were inoculated into 3 ml of minimal medium(MOPS [morpholinepropanesulfonic acid] buffer, 7.4 �M vitamin B12, 2% glu-cose; pH 7.2) and incubated overnight. These cultures were used to seed 25-mlcultures in minimal medium that were allowed to grow to an optical density at600 nm (OD600) of 0.3 before each culture was split in half; one part was used tocontinue monitoring changes in the OD600, and one part received radiolabeledphosphoric acid at a final concentration of 100 �Ci/ml. The cultures were al-lowed to go through two doublings before the stringent response was induced bythe addition of either 1 mg of D,L-serine hydroxamate/ml to E. coli (4) or 1 mgof D,L-norvaline/ml to the M. smegmatis strains (11). Both E. coli and M. smeg-matis cells were induced for 6 h. Cells were harvested by centrifugation andwashed twice in phosphate-buffered saline to remove the unincorporated label.Cell pellets were resuspended in 1 ml of 2 M formic acid, disrupted three timeswith glass beads in a bead beater, and centrifuged at 12,000 � g for 10 min toremove cellular debris. Supernatants were spotted onto 20- by 20-cm, 100-�m-thick cellulose polyethyleneimine thin-layer chromatography plates (Selecto Sci-entific). Thin-layer chromatography plates were equilibrated by soaking in dis-tilled water and then drying at room temperature. Plates were spotted withsamples 1 cm from the bottom of the plates. Commercially available radiolabeledATP was spotted at a 1:1,000 dilution to identify the relative positions of radio-labeled spots. The samples were resolved with a 1.5 M potassium phosphaterunning buffer until the solvent front migrated 15 cm (about 2.5 h). The plateswere air dried and exposed to phosphorimager screens for 7 days. These exper-iments were performed independently two times.
Electron microscopy. Cells were grown to stationary phase (4 days growth) andpelleted before being fixed with 2% glutaraldehyde and 2% paraformaldehyde in0.1 M cacodylate buffer with 6 M sucrose. Samples were prepared and analyzedfor scanning electron microscopy (SEM) and transmission electron microscopy(TEM) as previously described (8).
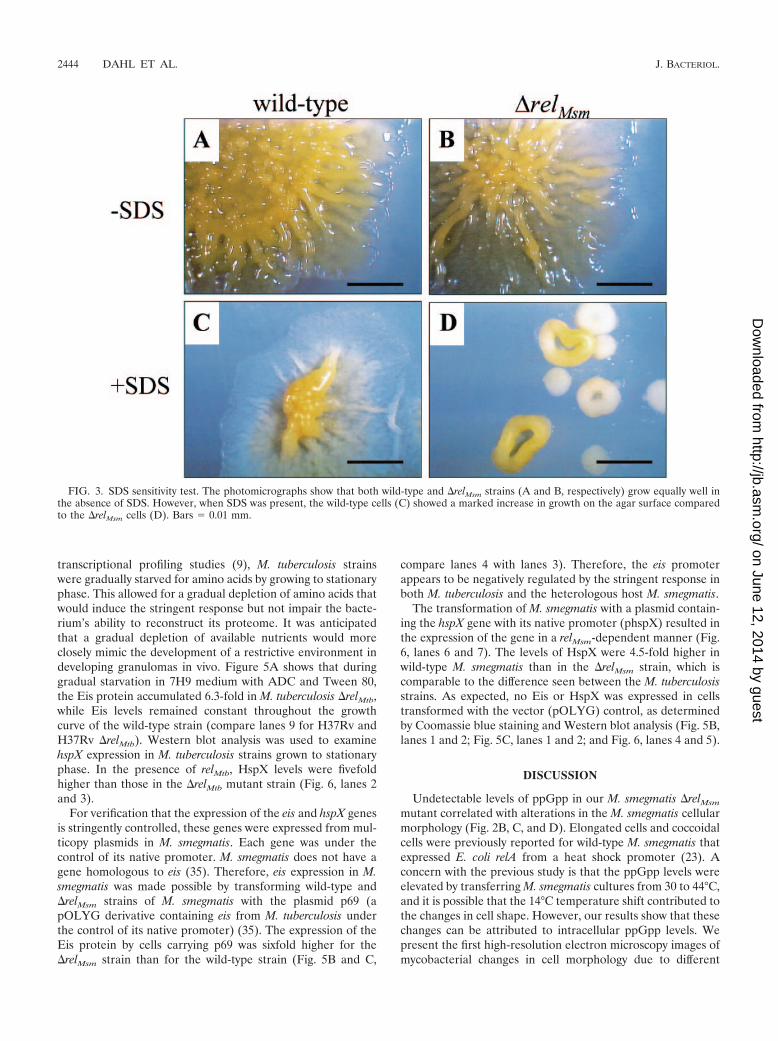
SDS sensitivity assay. Wild-type and �relMsm cells were compared for theirability to grow in the presence of 0.005% sodium dodecyl sulfate (SDS), aspreviously described (19). Cells were grown in 7H9 medium with Tween 80 to anOD600 of 0.5 and then serially diluted 10-fold onto Luria-Bertani agar plates,with or without 0.005% SDS. The plates were scored after 4 days of growth at37°C.
Competition starvation assays. Comparisons of wild-type M. smegmatis trans-formed with pMV306 (hygromycin-resistant integrating plasmid) and M. smeg-matis �relMsm (kanamycin resistant) for survival under nutrient and oxygendeprivation were performed as described previously (29). Briefly, each strain wasgrown to mid-log phase before being subjected to the following conditions. Foroxygen starvation conditions, the mid-log strains were kept in their originalmedium, mixed in a 1:1 ratio, and divided into aliquots in a series of 1.5-mlSarstedt screw-cap tubes containing gas-impermeable seals, with a 300-�l head-space for the cultures. A control culture containing 1.5 �g of methylene blue/mlindicated that oxygen was depleted within 36 h, as measured by the completedecolorization of the dye.
Western blot analysis of Eis. Detection of the Eis protein (Rv2416c) wasperformed as previously described, with a slight modification (10). M. tuberculosisstrains H37Rv and H37Rv �relMtb were grown from early log phase to stationaryphase, with aliquots removed at various time points to determine culture densi-ties (OD600) and prepare protein lysates. For analyses of Eis levels in M. smeg-matis, strains transformed with either pOLYG (12) or p69 (35) were grown in7H9 medium with 0.05% Tween 80 for 4 days before the cells were lysed andanalyzed for Eis protein as previously described (10). In addition, M. smegmatislysates separated by SDS–12% polyacrylamide gel electrophoresis were stainedwith Coomassie brilliant blue as previously described (35). A comparative quan-titation of band intensities in Western blots was made by densitometry with anAlphaImager 2000 (Alpha Innotech Corp.).
Western blot analysis of HspX. An analysis of HspX (�-crystallin) (Rv2031c)production was performed as previously described (10). H37Rv and H37Rv�relMtb cells were grown to an OD600 of 1.6 before being harvested and used toprepare protein lysates as described above. M. smegmatis strains were grown for4 days (OD600, 1.8) before the cells were harvested and lysed. For the expressionof hspX in M. smegmatis, the gene was amplified by PCR from H37Rv chromo-somal DNA with the primers 5�-CCCAAGCTTGACGGTGGCCCTCGGTGAC-3� and 5�-GCTCTAGAGCCGCTGCGGTCATCAGCAC-3�. The 976-bpPCR product containing hspX and its promoter was digested with HindIII andXbaI and ligated into HindIII- and XbaI-digested pOLYG to generate theplasmid phspX. Both pOLYG and phspX were electroporated into wild-type and�relMsm strains of M. smegmatis.
RESULTS
Deletion of relMsm from M. smegmatis. The relMsm gene wasinactivated in M. smegmatis by allelic replacement of the wild-type copy with one containing an internal, in-frame deletionand an internal kanamycin resistance marker (aph). Southernblot analysis and PCR analysis (results not shown) confirmedthat the M. smegmatis �relMsm strain lacked a 387-bp internalfragment and contained a 1.27-kbp insertion (aph) in relMsm.
The disruption of the relMsm gene caused a loss of RelAfunction, as shown by the loss of a radiolabeled spot for aformic acid extract of M. smegmatis �relMsm that correspondedwith the D,L-serine hydroxamate-induced ppGpp spot seen forE. coli (Fig. 1A, arrowhead). D,L-Serine hydroxamate was notused to induce the M. smegmatis stringent response since thisamino acid analog has previously been shown to be ineffectiveat inducing ppGpp synthesis in M. tuberculosis (29), possiblydue to impermeability of the mycobacterial cell wall to thisanalog. Therefore, D,L-norvaline was chosen to induce thestringent response since it was previously shown to initiate astringent response in Bacillus subtilis (11) and M. tuberculosis(our unpublished data). D,L-Norvaline induced the synthesis ofa molecule (Fig. 1A, lane 5) that was present in the wild-typestrain of M. smegmatis but was significantly reduced in the�relMsm strain (lane 6). The position of this spot matched thatfor radiolabeled ppGpp produced in E. coli. The identity of anadditional 32P-labeled spot migrating below ppGpp in M. smeg-matis formic acid extracts is unknown. This spot is not presentin E. coli extracts, but it has been previously seen in extracts ofM. smegmatis radiolabeled with 32P (23).
Further evidence for the inactivation of relMsm was seen by
2440 DAHL ET AL. J. BACTERIOL.
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

the accumulation of RNAs in stationary-phase M. smegmatis�relMsm (Fig. 1B), which was analogous to that reported for E.coli (6). Stable RNAs (23S and 16S rRNAs) were clearly visiblewith the M. smegmatis �relMsm strain induced for the stringentresponse (Fig. 1B, arrowheads).
The stringent response alters M. smegmatis morphology.Several structural differences were observed between the wild-type M. smegmatis and M. smegmatis �relMsm strains. Cultured�relMsm cells clumped significantly more than wild-type cells inliquid cultures (data not shown). This same phenotypic differ-
ence was also noticed between H37Rv �relMtb and H37Rv(data not shown). It has previously been demonstrated thatrelMtb controls several genes that are associated with biogenesisor the structure of the cell wall (9), which may correlate withthe apparent changes in cell wall characteristics. These genesinclude those known to play roles in cell invasion, peptidogly-can synthesis, mycolic acid biosynthesis, and transport as wellas those encoding a number of putative lipoproteins of un-known function and PE/PGRS family members. The Chatterjilaboratory recently reported that M. smegmatis can alter itsprofile of surface-associated glycopeptidolipids (GPLs) duringcarbon starvation (24). To analyze if GPLs were responsiblefor the clumping of the M. smegmatis �relMsm strain, we iso-lated and compared GPLs from the mutant and parentalstrains as previously described (24). There was no detectabledifference between GPLs of the wild-type and �relMsm strains(data not shown), which indicates that the GPLs are not likelyresponsible for the clumping phenotype.
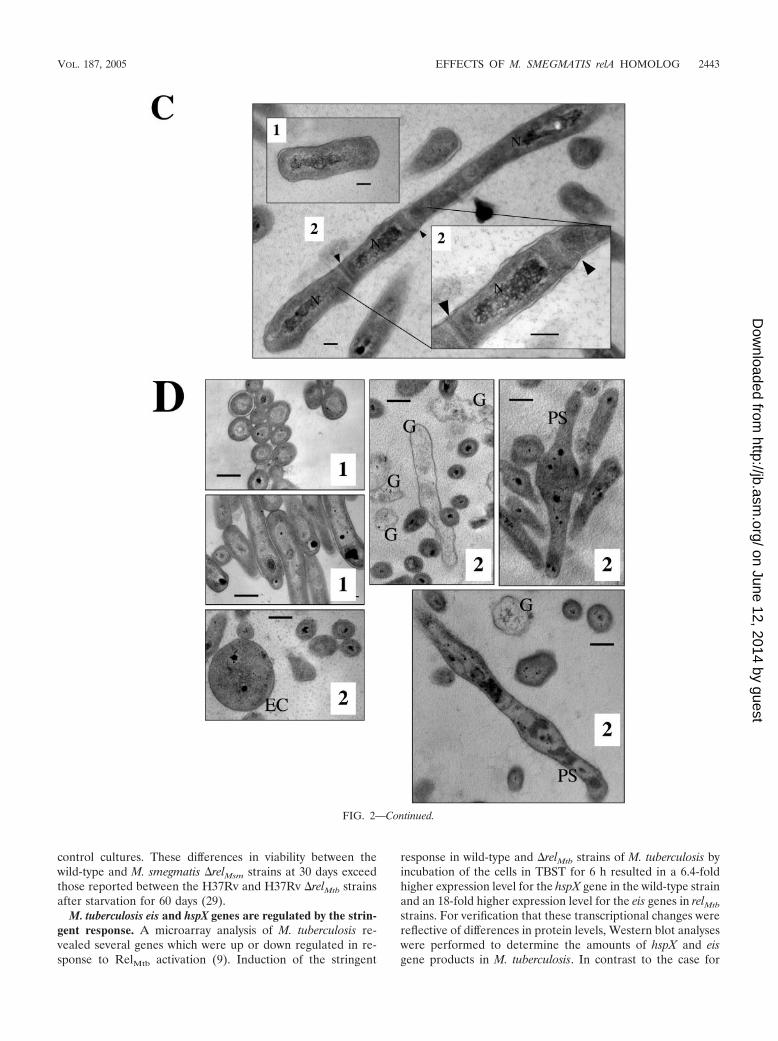
Individual colony morphologies also varied, as shown in Fig.2A. The wild-type strain formed colonies characteristic of M.smegmatis, with protruding edges on the perimeter and a rel-atively uniform orange pigmentation (Fig. 2A, panel 1). Whenviewed from the angle of the agar surface, these wild-typecolonies were relatively flat. Colonies of the M. smegmatis�relMsm strain, however, had smoother perimeter edges andless pigmentation in the perimeter of the colonies and formedcentral, elevated peaks when viewed from the side (Fig. 2A,panel 2).
Individual cell morphologies also varied between the two M.smegmatis strains. Wild-type cells viewed by SEM appeared asshort rods (Fig. 2B, panel 1) (average length, 1.2 �m), while�relMsm cells appeared significantly longer (Fig. 2B, panel 2)(average length, 3.5 �m).
TEM analysis of the two strains confirmed the short lengthsof wild-type cells compared to �relMsm cells (Fig. 2C). Numer-ous elongated M. smegmatis �relMsm cells contained multipledivision septa per cell (Fig. 2C, panel 2). These results aresimilar to those reported for ppGpp-free E. coli cells (36).Notably, the M. smegmatis �relMsm strain also produced roundenlarged cells (EC) as well as cells with a pear-shaped (PS)appearance that diverged from the typical bacillus shape (Fig.2D). TEM analysis of the M. smegmatis �relMsm strain alsorevealed a large number of cell wall “ghosts” (G) that ap-peared to be remnants of cells lacking cytoplasm (Fig. 2D).These ghosts were not seen in wild-type cells grown for anequivalent length of time. Despite the differences in cellclumping (data not shown) and cell shape (Fig. 2B, C, and D),there was no detectable difference in cell wall structure be-tween the two strains, as observed by TEM at a magnificationof �200,000 (data not shown).
The stringent response does not affect the M. smegmatisgrowth rate. It was previously shown that eliminating the strin-gent response in M. tuberculosis results in decreased growth at37°C and an inability to grow at 42°C (29). M. smegmatis�relMsm grown at 37 or 42°C showed no significant differencefrom the wild-type parental strain (data not shown). Both ofthese observations are in contrast to those for M. tuberculosis.
SDS sensitivity. Sensitivity to SDS has previously beenshown for M. smegmatis strains with altered cell surfaces (19).The presence of 0.005% SDS in Luria-Bertani agar reduced
FIG. 1. Inactivation of relMsm in M. smegmatis. (A) Thin-layer chro-matography of radiolabeled (p)ppGpp. E. coli strain JM105 (lane 1)was induced for the stringent response by D,L-serine hydroxamate(lane 2). The appearance of labeled (p)ppGpp is indicated with anarrowhead. Wild-type M. smegmatis (lane 3) and M. smegmatis �relMsm(lane 4) were induced for the stringent response by D,L-norvaline(lanes 5 and 6, respectively). Radiolabeled (p)ppGpp is present in lane5 (wild-type strain) but absent from lane 6 (�relMsm strain). (B) Aga-rose gel electrophoresis of RNAs obtained from either wild-type M.smegmatis (lane 1) or M. smegmatis �relMsm (lane 2) cells that had beengrown to stationary phase (3-day-old cultures). A 1-kb DNA ladder ispresent in lane L. rRNA bands are indicated (arrowheads).
VOL. 187, 2005 EFFECTS OF M. SMEGMATIS relA HOMOLOG 2441
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

the growth of both the wild-type and �relMsm strains, but thereduction was especially apparent with the �relMsm cells (Fig.3). The �relMsm colonies were much smaller and had edges thatwere raised. These colonies seemed to grow in a way thatminimized contact with the agar surface, while the wild-typecolonies were seen to be spreading out.
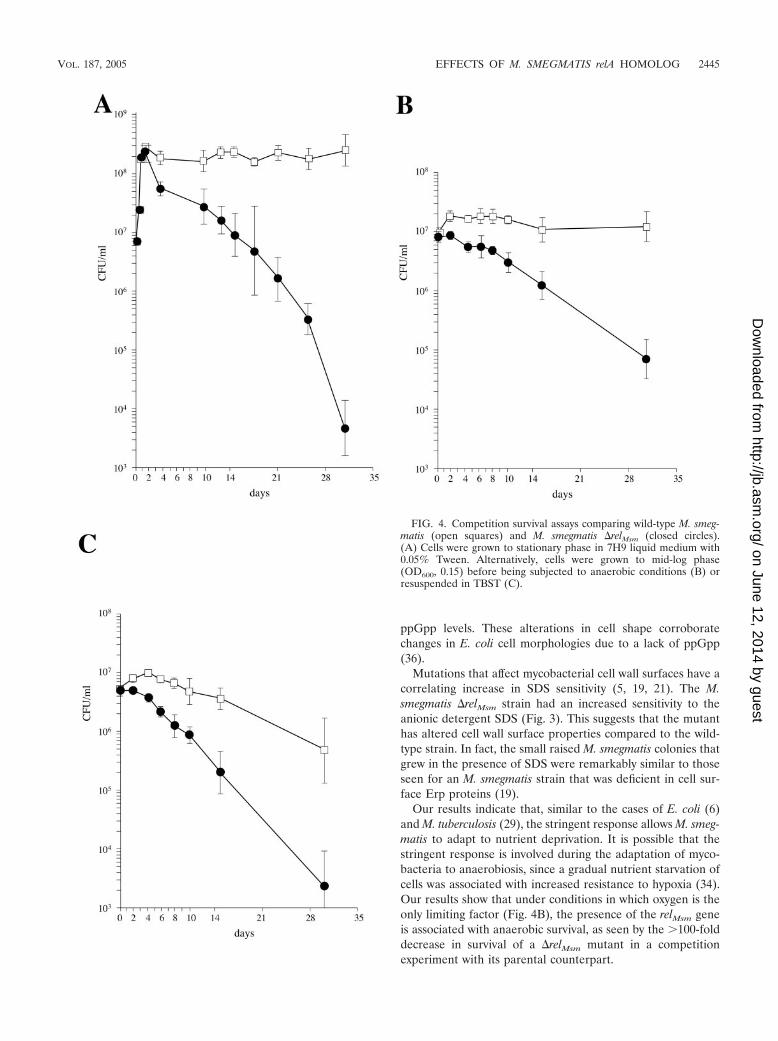
The stringent response is required for long-term survivalunder conditions of nutrient deprivation. Based on previousstudies of the stringent response in M. tuberculosis (29), wepredicted that the relMsm function would contribute to M. smeg-matis persistence in vitro under conditions of prolonged nutri-ent or oxygen starvation. Wild-type M. smegmatis containing anintegrated copy of pMV306 (Hygr) and M. smegmatis �relMsm
(Kanr) were grown separately to mid-log phase, mixed at a 1:1ratio, and subjected to various starvation conditions. After 30days of starvation, there were dramatic decreases in the viabil-ity of the M. smegmatis �relMsm strain compared to the wild-type strain. These included a 4.5-log decrease in viability whencells were grown to stationary phase in rich medium (Fig. 4A),a �2-log decrease when cells were subjected to oxygen limita-tion only (Fig. 4B), and a 2.5-log decrease when cells weresuddenly and completely starved for nutrients in Tris-bufferedsaline with Tween 80 (TBST) (Fig. 4C). The cessation ofgrowth of M. smegmatis after 36 h in gas-impermeable tubeswas due to the consumption of all available oxygen, as dem-onstrated by the complete decolorization of methylene blue in
FIG. 2. Effects of stringent response upon the appearance of M. smegmatis. Wild-type M. smegmatis (panels 1) and M. smegmatis �relMsm(panels 2) are shown. (A) Colonies grown on 7H9 agar for 2 weeks. Bars � 5 mm. (B) SEM analysis of cells grown for 3 days in 7H9 liquid medium.Bars � 3.8 �m. (C) TEM analysis of cells grown for 3 days in 7H9 liquid medium. “N” indicates a nucleoid region, and arrowheads indicate twosepta in a single cell. The inset shows a magnification of the two septa. Bars � 200 nm. (D) TEM analysis showing an enlarged cell (EC) comparedto the smaller (in diameter) wild-type cells. Ghost cells (G) and pear-shaped cells (PS) were seen routinely for M. smegmatis �relMsm but wereabsent from the wild-type cells. Bars � 500 nm.
2442 DAHL ET AL. J. BACTERIOL.
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
control cultures. These differences in viability between thewild-type and M. smegmatis �relMsm strains at 30 days exceedthose reported between the H37Rv and H37Rv �relMtb strainsafter starvation for 60 days (29).
M. tuberculosis eis and hspX genes are regulated by the strin-gent response. A microarray analysis of M. tuberculosis re-vealed several genes which were up or down regulated in re-sponse to RelMtb activation (9). Induction of the stringent
response in wild-type and �relMtb strains of M. tuberculosis byincubation of the cells in TBST for 6 h resulted in a 6.4-foldhigher expression level for the hspX gene in the wild-type strainand an 18-fold higher expression level for the eis genes in relMtb
strains. For verification that these transcriptional changes werereflective of differences in protein levels, Western blot analyseswere performed to determine the amounts of hspX and eisgene products in M. tuberculosis. In contrast to the case for
FIG. 2—Continued.
VOL. 187, 2005 EFFECTS OF M. SMEGMATIS relA HOMOLOG 2443
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
transcriptional profiling studies (9), M. tuberculosis strainswere gradually starved for amino acids by growing to stationaryphase. This allowed for a gradual depletion of amino acids thatwould induce the stringent response but not impair the bacte-rium’s ability to reconstruct its proteome. It was anticipatedthat a gradual depletion of available nutrients would moreclosely mimic the development of a restrictive environment indeveloping granulomas in vivo. Figure 5A shows that duringgradual starvation in 7H9 medium with ADC and Tween 80,the Eis protein accumulated 6.3-fold in M. tuberculosis �relMtb,while Eis levels remained constant throughout the growthcurve of the wild-type strain (compare lanes 9 for H37Rv andH37Rv �relMtb). Western blot analysis was used to examinehspX expression in M. tuberculosis strains grown to stationaryphase. In the presence of relMtb, HspX levels were fivefoldhigher than those in the �relMtb mutant strain (Fig. 6, lanes 2and 3).
For verification that the expression of the eis and hspX genesis stringently controlled, these genes were expressed from mul-ticopy plasmids in M. smegmatis. Each gene was under thecontrol of its native promoter. M. smegmatis does not have agene homologous to eis (35). Therefore, eis expression in M.smegmatis was made possible by transforming wild-type and�relMsm strains of M. smegmatis with the plasmid p69 (apOLYG derivative containing eis from M. tuberculosis underthe control of its native promoter) (35). The expression of theEis protein by cells carrying p69 was sixfold higher for the�relMsm strain than for the wild-type strain (Fig. 5B and C,
compare lanes 4 with lanes 3). Therefore, the eis promoterappears to be negatively regulated by the stringent response inboth M. tuberculosis and the heterologous host M. smegmatis.
The transformation of M. smegmatis with a plasmid contain-ing the hspX gene with its native promoter (phspX) resulted inthe expression of the gene in a relMsm-dependent manner (Fig.6, lanes 6 and 7). The levels of HspX were 4.5-fold higher inwild-type M. smegmatis than in the �relMsm strain, which iscomparable to the difference seen between the M. tuberculosisstrains. As expected, no Eis or HspX was expressed in cellstransformed with the vector (pOLYG) control, as determinedby Coomassie blue staining and Western blot analysis (Fig. 5B,lanes 1 and 2; Fig. 5C, lanes 1 and 2; and Fig. 6, lanes 4 and 5).
DISCUSSION
Undetectable levels of ppGpp in our M. smegmatis �relMsm
mutant correlated with alterations in the M. smegmatis cellularmorphology (Fig. 2B, C, and D). Elongated cells and coccoidalcells were previously reported for wild-type M. smegmatis thatexpressed E. coli relA from a heat shock promoter (23). Aconcern with the previous study is that the ppGpp levels wereelevated by transferring M. smegmatis cultures from 30 to 44°C,and it is possible that the 14°C temperature shift contributed tothe changes in cell shape. However, our results show that thesechanges can be attributed to intracellular ppGpp levels. Wepresent the first high-resolution electron microscopy images ofmycobacterial changes in cell morphology due to different
FIG. 3. SDS sensitivity test. The photomicrographs show that both wild-type and �relMsm strains (A and B, respectively) grow equally well inthe absence of SDS. However, when SDS was present, the wild-type cells (C) showed a marked increase in growth on the agar surface comparedto the �relMsm cells (D). Bars � 0.01 mm.
2444 DAHL ET AL. J. BACTERIOL.
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
ppGpp levels. These alterations in cell shape corroboratechanges in E. coli cell morphologies due to a lack of ppGpp(36).
Mutations that affect mycobacterial cell wall surfaces have acorrelating increase in SDS sensitivity (5, 19, 21). The M.smegmatis �relMsm strain had an increased sensitivity to theanionic detergent SDS (Fig. 3). This suggests that the mutanthas altered cell wall surface properties compared to the wild-type strain. In fact, the small raised M. smegmatis colonies thatgrew in the presence of SDS were remarkably similar to thoseseen for an M. smegmatis strain that was deficient in cell sur-face Erp proteins (19).
Our results indicate that, similar to the cases of E. coli (6)and M. tuberculosis (29), the stringent response allows M. smeg-matis to adapt to nutrient deprivation. It is possible that thestringent response is involved during the adaptation of myco-bacteria to anaerobiosis, since a gradual nutrient starvation ofcells was associated with increased resistance to hypoxia (34).Our results show that under conditions in which oxygen is theonly limiting factor (Fig. 4B), the presence of the relMsm geneis associated with anaerobic survival, as seen by the �100-folddecrease in survival of a �relMsm mutant in a competitionexperiment with its parental counterpart.
FIG. 4. Competition survival assays comparing wild-type M. smeg-matis (open squares) and M. smegmatis �relMsm (closed circles).(A) Cells were grown to stationary phase in 7H9 liquid medium with0.05% Tween. Alternatively, cells were grown to mid-log phase(OD600, 0.15) before being subjected to anaerobic conditions (B) orresuspended in TBST (C).
VOL. 187, 2005 EFFECTS OF M. SMEGMATIS relA HOMOLOG 2445
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

A noticeable difference exists between the stringent re-sponses of E. coli and mycobacteria. Recently, DksA wasshown to act in synergy with ppGpp during the regulation ofRNA polymerase activity (28). DksA is essential for the strin-gent control of rRNA in E. coli (27), but no DksA homologsexist in the known genomes of mycobacteria. Therefore, thetight regulation of rRNA by M. smegmatis (Fig. 1B) and M.tuberculosis (9) suggests that there is some DksA-independentmechanism.
Our study corroborates previous findings that the expressionof the hspX and eis genes is under opposite control by thestringent response, as evidenced by the increased expression ofHspX and Eis proteins in the wild-type and �relMtb mutants ofM. tuberculosis, respectively (Fig. 5 and 6). Eis was previouslyshown to be immunogenic in TB patients (10). This is similarto the case for several other immunodominant M. tuberculosisantigens which have been shown to be relMtb dependent, in-cluding the 19-kDa antigen Esat6 and members of the antigen85 complex (9). Although the function of the Eis protein isunknown, reverse position-specific BLAST, three-dimensionalposition-specific scoring matrix, and multiple EM for motifelicitation programs have revealed that Eis is a member of theGCN5 superfamily of N-acetyltransferases (Richard Friedman,personal communication). This family of proteins regulatesseveral different cellular functions, including transcriptionalactivity and antibiotic resistance.
In contrast to the unknown function of Eis, the HspX pro-tein has been shown to act as a heat shock protein (Hsp) andcan prevent the thermal denaturation of alcohol dehydroge-nase (37). HspX is also involved in Mycobacterium bovis BCGand M. tuberculosis cell wall thickening under prolonged an-
aerobic growth (7). The hspX gene is a member of the DosR-regulated dormancy regulon (26), and its expression has beenshown to increase during stationary-phase growth (15), duringhypoxia (38), or when cultures are exposed to nitric oxide (13).We provided evidence that hspX additionally belongs to theRelA regulon in mycobacteria. We believe that this regulationis independent of sigF since the expression of this alternativesigma factor in M. tuberculosis is not regulated by the stringentresponse (unpublished data). The inability of the H37Rv�relMtb strain to grow at 42°C (29) may be linked to the de-creased expression of HspX. The absence of temperature sen-sitivity in the M. smegmatis �relMsm strain may indicate thatheat shock proteins with a similar role in protection againstthermal denaturation are not under stringent control in thisorganism. RelMtb-mediated expression of HspX during nutri-ent restriction in the host environment may assist in persistentsurvival since this protein has been shown to play a role in vitroin structural adaptations to a persistent phenotype (7).
Our results have shown that the key elements of stringentcontrol are conserved between M. tuberculosis and M. smeg-matis and that M. smegmatis is a useful heterologous host forinvestigating critical elements of this bacterial response. Inaddition, we have shown that the stringent response plays arole in adaptation to both nutrient and oxygen starvation. Un-derstanding the mechanisms that allow M. smegmatis to entera state of apparent dormancy is essential for identifying targetsthat are necessary for the persistent survival of organisms inpatients harboring latent disease. However, the genome size ofM. smegmatis (about 7 Mb) is considerably larger than that ofM. tuberculosis (4.4 Mb), and thus it is likely that studying thestringent response of this nonpathogenic species may produceinteresting relA-dependent mechanisms that are not seen in thepathogen.
ACKNOWLEDGMENTS
This research was supported by a grant from the American LungAssociation (RG-022-N) and by a new investigator matching grantfrom the National Foundation for Infectious Diseases. Purified HspXprotein was provided by Colorado State University, courtesy of NIH,NIAID contract no. AI-75320, entitled “Tuberculosis Research Mate-rials and Vaccine Testing.”
We are appreciative of Christine Davitt and Valerie Lynch-Holm ofWashington State University for their assistance with electron micros-copy analysis. We thank Linoj Samuel and Richard Friedman fordiscussions about the function of Eis, William Louie and Nancy Tac-coni for helpful comments, and Robert Kadner for proofreading themanuscript.
FIG. 5. Regulation of eis (Rv2416c) by mycobacterial stringent re-sponse. (A) M. tuberculosis strains H37Rv and H37Rv �relMtb weregrown from early log (lanes 1 and 2) to mid-log (lanes 3 to 6) andstationary (lanes 7 to 9) phase. Protein lysates were equilibrated (22�g/lane) and examined for eis expression by Western blot analysis.(B) Coomassie-stained proteins (25 �g/lane) are shown for M. smeg-matis/pOLYG (lane 1), M. smegmatis �relMsm/pOLYG (lane 2), M.smegmatis/p69 (lane 3), and M. smegmatis �relMsm/p69 (lane 4).(C) Western blot analysis of Eis with the same protein lysates as thoseused for panel B.
FIG. 6. Regulation of hspX (Rv2031c) by mycobacterial stringentresponse. Western blot analysis was performed with an anti-HspXantibody. Lane 1, 3 �g of purified HspX; lane 2, M. tuberculosisH37Rv; lane 3, H37Rv �relMtb; lane 4, M. smegmatis/pOLYG; lane 5,M. smegmatis �relMsm/pOLYG; lane 6, M. smegmatis/phspX; and lane7, M. smegmatis �relMsm/phspX. Lanes 2 to 7 contained 22 �g ofprotein/lane.
2446 DAHL ET AL. J. BACTERIOL.
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

REFERENCES
1. Betts, J. C., P. T. Lukey, L. C. Robb, R. A. McAdam, and K. Duncan. 2002.Evaluation of a nutrient starvation model of Mycobacterium tuberculosispersistence by gene and protein expression profiling. Mol. Microbiol. 43:717–731.
2. Bose, M., A. Chander, and D. H. Das. 1993. A rapid and gentle method forthe isolation of genomic DNA from mycobacteria. Nucleic Acids Res. 21:2529–2530.
3. Boshoff, H. I., and V. Mizrahi. 2000. Expression of Mycobacterium smegmatispyrazinamidase in Mycobacterium tuberculosis confers hypersensitivity to pyr-azinamide and related amides. J. Bacteriol. 182:5479–5485.
4. Brown, L., D. Gentry, T. Elliott, and M. Cashel. 2002. DksA affects ppGppinduction of RpoS at a translational level. J. Bacteriol. 184:4455–4465.
5. Camacho, L. R., P. Constant, C. Raynaud, M. A. Laneelle, J. A. Triccas, B.Gicquel, M. Daffe, and C. Guilhot. 2001. Analysis of the phthiocerol dimy-cocerosate locus of Mycobacterium tuberculosis. J. Biol. Chem. 276:19845–19854.
6. Cashel, M., D. R. Gentry, V. J. Hernandez, and D. Vinella. 1996. Thestringent response, p. 1458–1496. In F. C. Neidhardt, R. Curtiss III, J. L.Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, W. S. Reznikoff, M. Riley,M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella:cellular and molecular biology, vol. 1. ASM Press, Washington, D.C.
7. Cunningham, A. F., and C. L. Spreadbury. 1998. Mycobacterial stationaryphase induced by low oxygen tension: cell wall thickening and localization ofthe 16-kilodalton alpha-crystallin homolog. J. Bacteriol. 180:801–808.
8. Dahl, J. L. 2004. Electron microscopy analysis of Mycobacterium tuberculosiscell division. FEMS Microbiol. Lett. 240:15–20.
9. Dahl, J. L., C. N. Kraus, H. I. Boshoff, B. Doan, K. Foley, D. Avarbock, G.Kaplan, V. Mizrahi, H. Rubin, and C. E. Barry III. 2003. The role ofRelMtb-mediated adaptation to stationary phase in long-term persistence ofMycobacterium tuberculosis in mice. Proc. Natl. Acad. Sci. USA 100:10026–10031.
10. Dahl, J. L., J. Wei, J. W. Moulder, S. Laal, and R. L. Friedman. 2001.Subcellular localization of the intracellular survival-enhancing Eis protein ofMycobacterium tuberculosis. Infect. Immun. 69:4295–4302.
11. Eymann, C., G. Homuth, C. Scharf, and M. Hecker. 2002. Bacillus subtilisfunctional genomics: global characterization of the stringent response byproteome and transcriptome analysis. J. Bacteriol. 184:2500–2520.
12. Garbe, T. R., J. Barathi, S. Barnini, Y. Zhang, C. Abou-Zeid, D. Tang, R.Mukherjee, and D. B. Young. 1994. Transformation of mycobacterial speciesusing hygromycin resistance as selectable marker. Microbiology 140:133–138.
13. Garbe, T. R., N. S. Hibler, and V. Deretic. 1999. Response to reactivenitrogen intermediates in Mycobacterium tuberculosis: induction of the 16-kilodalton alpha-crystallin homolog by exposure to nitric oxide donors. In-fect. Immun. 67:460–465.
14. Hingley-Wilson, S. M., V. K. Sambandamurthy, and W. R. Jacobs, Jr. 2003.Survival perspectives from the world’s most successful pathogen, Mycobac-terium tuberculosis. Nat. Immunol. 4:949–955.
15. Hu, Y., and A. R. Coates. 1999. Transcription of the stationary-phase-asso-ciated hspX gene of Mycobacterium tuberculosis is inversely related to syn-thesis of the 16-kilodalton protein. J. Bacteriol. 181:1380–1387.
16. Jacobs, W. R., Jr., M. Tuckman, and B. R. Bloom. 1987. Introduction offoreign DNA into mycobacteria using a shuttle phasmid. Nature 327:532–535.
17. Kaufmann, S. H. E., and I. D. A. van Embden. 1993. Tuberculosis: a ne-glected disease strikes back. Trends Microbiol. 1:2–5.
18. Khomenko, A. G. 1987. The variability of Mycobacterium tuberculosis inpatients with cavitary pulmonary tuberculosis in the course of chemotherapy.Tubercle 68:243–253.
19. Kocincova, D., B. Sonden, L. de Mendoca-Lima, B. Gicquel, and J.-M.Reyrat. 2004. The Erp protein is anchored at the surface by a carboxy-
terminal hydrophobic domain and is important for cell-wall structure inMycobacterium smegmatis. FEMS Microbiol. Lett. 231:191–196.
20. Lobel, R. O., E. Shorr, and H. B. Richardson. 1933. The influence of adverseconditions upon the respiratory metabolism and growth of human tuberclebacilli. J. Bacteriol. 26:167–200.
21. Nikaido, H., and M. Vaara. 1985. Molecular basis of bacterial outer mem-brane permeability. Microbiol. Rev. 49:1–32.
22. Nyka, W. 1974. Studies on the effect of starvation on mycobacteria. Infect.Immun. 9:843–850.
23. Ojha, A. K., T. K. Mukherjee, and D. Chatterji. 2000. High intracellular levelof guanosine tetraphosphate in Mycobacterium smegmatis changes the mor-phology of the bacterium. Infect. Immun. 68:4084–4091.
24. Ojha, A. K., S. Varma, and D. Chatterji. 2002. Synthesis of an unusual polarglycopeptidolipid in glucose-limited culture of Mycobacterium smegmatis.Microbiology 148:3039–3048.
25. Parish, T., and N. G. Stoker. 2000. Use of a flexible cassette method togenerate a double unmarked Mycobacterium tuberculosis tlyA plcABC mutantby gene replacement. Microbiology 146:1969–1975.
26. Park, H. D., K. M. Guinn, M. I. Harrell, R. Liao, M. I. Voskuil, M. Tompa,G. K. Schoolnik, and D. R. Sherman. 2003. Rv3133c/dosR is a transcriptionfactor that mediates the hypoxic response of Mycobacterium tuberculosis.Mol. Microbiol. 48:833–843.
27. Paul, B. J., M. M. Barker, W. Ross, D. A. Schneider, C. Webb, J. W. Foster,and R. L. Gourse. 2004. DksA: a critical component of the transcriptioninitiation machinery that potentiates the regulation of rRNA promoters byppGpp and the initiating NTP. Cell 118:311–322.
28. Perederina, A., V. Svetlov, M. N. Vassylyeva, T. H. Tahirov, S. Yokoyama, I.Artsimovitch, and D. G. Vassylyev. 2004. Regulation through the secondarychannel—structural framework for ppGpp-DksA synergism during tran-scription. Cell 118:297–309.
29. Primm, T. P., S. J. Andersen, V. Mizrahi, D. Avarbock, H. Rubin, and C. E.Barry III. 2000. The stringent response of Mycobacterium tuberculosis isrequired for long-term survival. J. Bacteriol. 182:4889–4898.
30. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
31. Smeulders, M. J., J. Keer, R. A. Speight, and H. D. Williams. 1999. Adap-tation of Mycobacterium smegmatis to stationary phase. J. Bacteriol. 181:270–283.
32. Wayne, L. G. 1994. Dormancy of Mycobacterium tuberculosis and latency ofdisease. Eur. J. Clin. Microbiol. Infect. Dis. 13:908–914.
33. Wayne, L. G., and L. G. Hayes. 1996. An in vitro model for sequential studyof shiftdown of Mycobacterium tuberculosis through two stages of nonrepli-cating persistence. Infect. Immun. 64:2062–2069.
34. Wayne, L. G., and K. Lin. 1982. Glyoxylate metabolism and adaptation ofMycobacterium tuberculosis to survival under anaerobic conditions. Infect.Immun. 37:1042–1049.
35. Wei, J., J. L. Dahl, J. W. Moulder, E. A. Roberts, P. O’Gaora, D. B. Young,and R. L. Friedman. 2000. Identification of a Mycobacterium tuberculosisgene that enhances mycobacterial survival in macrophages. J. Bacteriol.182:377–384.
36. Xiao, H., M. Kalman, K. Ikehara, S. Zemel, G. Glaser, and M. Cashel. 1991.Residual guanosine 3�,5�-bispyrophosphate synthetic activity of relA nullmutants can be eliminated by spoT null mutations. J. Biol. Chem. 266:5980–5990.
37. Yuan, Y., D. D. Crane, and C. E. Barry III. 1996. Stationary-phase-associatedprotein expression in Mycobacterium tuberculosis: function of the mycobac-terial alpha-crystallin homolog. J. Bacteriol. 178:4484–4492.
38. Yuan, Y., D. D. Crane, R. M. Simpson, Y. Zhu, M. J. Hickey, D. R. Sherman,and C. E. Barry III. 1998. The 16-kDa �-crystallin (Acr) protein of Myco-bacterium tuberculosis is required for growth in macrophages. Proc. Natl.Acad. Sci. USA 95:9578–9583.
VOL. 187, 2005 EFFECTS OF M. SMEGMATIS relA HOMOLOG 2447
on June 12, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
Copyright © 2022 FDOKUMEN
















![Pulmonary Inflammation Induced by a Recombinant Brugia malayi [gamma]-glutamyl transpeptidase Homolog: Involvement of Humoral Autoimmune Responses](https://static.fdokumen.com/doc/165x107/631e10e40ff042c6110c2b14/pulmonary-inflammation-induced-by-a-recombinant-brugia-malayi-gamma-glutamyl-transpeptidase.jpg)



