
Multi-antibody composition in lupus nephritis: Isotype and antigen specificity make the difference
Upload
independentCategory
view
3download
0
Immunogenetics (2003) 55:605–614DOI 10.1007/s00251-003-0620-7
O R I G I N A L P A P E R
Jan Salomonsen · Denise Marston · David Avila ·Nat Bumstead · Brit Johansson · Helle Juul-Madsen ·Gerda D. Olesen · Patricia Riegert · Karsten Skjødt ·Olli Vainio · Michael V. Wiles · Jim Kaufman
The properties of the single chicken MHC classical class IIa chain (B-LA) gene indicate an ancient origin for the DR/E-like isotypeof class II moleculesReceived: 29 August 2003 / Revised: 9 October 2003 / Published online: 8 November 2003� Springer-Verlag 2003
Abstract In mammals, there are MHC class II moleculeswith distinctive sequence features, such as the classicalisotypes DR, DQ and DP. These particular isotypes havenot been reported in non-mammalian vertebrates. Wehave isolated the class II (B-L) a chain from outbred
chickens as the basis for the cloning and sequencing ofthe cDNA. We found only one class II a chain transcript,which bears the major features of a classical class II asequence, including the critical peptide-binding residues.The chicken sequence is more similar to human DR thanto the DQ, DP, DO or DM isotypes, most significantly inthe peptide-binding a1 domain. The cDNA and genomicDNA sequences from chickens of diverse origins showfew alleles, which differ in only four nucleotides and oneamino acid. In contrast, significant restriction fragmentlength polymorphism is detected by Southern blot anal-ysis of genomic DNA, suggesting considerable diversityaround the gene. Analysis of a large back-cross familyindicates that the class II a chain locus (B-LA) is locatedroughly 5.6 cM from the MHC locus, which encodes theclassical class II b chains. Thus the chicken class II achain gene is like the mammalian DR and E isotypes inthree properties: the presence of the critical peptide-binding residues, the low level of polymorphism andsequence diversity, and the recombinational separationfrom the class II b chain genes. These results indicate thatthe sequence features of this lineage are both functionallyimportant and at least 300 million years old.
Keywords Major histocompatibility complex · B locus ·Class II · DR isotype · Chicken
Introduction
The major histocompatibility complex (MHC), originallydiscovered as the predominant locus responsible for rapidtissue allograft rejection, is now known to encode class Iand class II molecules that bind peptides for presentationas antigen to T lymphocytes of the immune system.Classical class II molecules bind peptides derivedpredominantly from proteins acquired from outside thecell (and from the topological equivalent, intra-cellularvesicles) for presentation to CD4-bearing T cells that
The nucleotide sequence data reported in this paper have beensubmitted to the EMBL/GenBank nucleotide sequence databasesand have been assigned the accession numbers AY357253 toAY357256
J. SalomonsenDepartment of Virology and Immunology,Royal Veterinary and Agricultural University,Stigbøjlen 7, C 1870 Frederiksberg, Denmark
D. Avila · B. Johansson · P. RiegertBasel Institute for Immunology,Grenzacherstrasse 487, 4005 Basel, Switzerland
D. Marston · N. Bumstead · J. Kaufman ())Institute for Animal Health, Compton, Berkshire, RG20 7NN, UKe-mail: [email protected].: +44-1635-577234Fax: +44-1635-577263
H. Juul-MadsenNational Institute of Animal Science, Foulum Research Center,P.O. Box 39, 8830 Tjele, Denmark
G. D. OlesenDepartment of Life Sciences and Chemistry, Roskilde University,P.O. Box 260, 4000 Roskilde, Denmark
K. SkjødtDepartment of Immunology and Medical Microbiology,University of Odense,Winslovparken 19, C 5000 Odense, Denmark
O. VainioDepartment of Medical Microbiology, Turku University,Kiinamyllynkatu 13, 20520 Turku, Finland
M. V. WilesMax Planck Institute for Molecular Genetics,Ihnestrasse 73, 14195 Berlin, Germany

regulate antibody production, among other actions. Theclassical class II molecules are highly polymorphic, withmost variation located in the peptide-binding domain andselected, at least in part, by the molecular arms race withpathogens. In addition, there are non-classical class IImolecules that are not highly polymorphic and areexpressed inside the cell, where they are involved inloading peptides onto the classical class II molecules(Marsh et al. 2000).
Class II molecules consist of two structurally relatedtransmembrane glycoproteins, the a and b chains, in non-covalent association. Each of these chains has an N-terminal signal sequence which is cleaved upon entry intothe lumen of the endoplasmic reticulum, followed by apeptide-binding a1 domain, an immunoglobulin-like a2domain, an acidic connecting peptide, hydrophobictransmembrane region and a short cytoplasmic tail. Manysequence features have been noted and located on three-dimensional structures, including residues involved ininteraction between the two chains, with the boundpeptide, with the T-cell receptor, with the co-receptorCD4 and between class II dimers. The a and b chains areencoded by A and B genes which are generally locatednext to each other in the MHC (Brown et al. 1993; Sternet al. 1994; Kaufman et al. 1995c; Hennecke et al. 2000;Marsh et al. 2000).
All jawed vertebrate groups examined have class IIgenes (Flajnik and Kasahara 2001), and typically expressa number of different classical class II molecules (so-called isotypes). In humans, there are three classical classII isotypes: one (or two, depending on haplotype) DRmolecules encoded by one (or two) DRA gene(s) and one(or two) neighboring DRB gene(s), one DQ moleculeencoded by adjacent DQA1 and DQB1 genes, and one DPmolecule encoded by adjacent DPA1 and DPB1 genes(Beck and Trowsdale 1999; Marsh et al. 2000). Allmammals examined express one or more of these sameclassical isotypes (along with the so-called DY isotype insome ruminants; Wright et al. 1994). For instance,laboratory mice express the DR-like I-E molecule andthe DQ-like I-A molecule (Steinmetz et al. 1982). Incontrast to mammals, the multiple classical class II-likegenes of non-mammalian vertebrates (including birds,frogs, bony fish and sharks) do not appear to be morerelated to one human isotype than to another (Kasahara etal. 1992; Ono et al. 1992; Sultmann et al. 1993; Zoorob etal. 1993; Dixon et al. 1995; Grimholt et al. 2000; Murrayet al. 2000; Liu et al. 2002).
Classical A and B genes are generally both polymor-phic, presumably because both a and b chains contributeto the peptide-binding site (Stern et al. 1994). The notableexceptions are the DRA-like genes, which are monomor-phic (or oligomorphic with very few sequence differ-ences). As pointed out long ago (Germain et al. 1985),infrequent recombination between the mouse Aa and Ab-chain genes means that the gene pair can co-evolve withinteracting alleles staying together, whereas the frequentrecombination between the Ea- and Eb-chain genesmeans that the Ea-chain gene must evolve to an “average
best-fit” with all of the Eb-chain alleles. Whether this istrue for the DRA genes of humans and other mammals isnot yet clear. It is also not clear whether the DRA-likegenes are selected to be monomorphic because they areseparated by recombination from their partner DRB-likegenes, or the recombination is selected because DR-like achain genes need to be monomorphic for optimalfunction.
In chickens, two genomic regions contain class II Bgenes: the B-F/B-L region of the B locus and the Rfp-Ylocus (Guillemot et al. 1988; Miller et al. 1994). Theseloci are genetically unlinked, but both are located on thesame microchromosome, presumably separated by thehighly repetitive (and therefore highly recombinogenic)nucleolar organiser region (Fillon et al. 1996; Miller et al.1996). The B-F/B-L region contains the highly polymor-phic classical class II B (so-called B-LB) genes anddetermines MHC-related functions such as rapid allograftrejection, whereas the Rfp-Y region contains non-poly-morphic and therefore non-classical class II B (so-calledY-LB) genes (Zoorob et al. 1990, 1993). The B-F/B-Lregion is thus the chicken MHC, but the organizationdiffers significantly from the typical mammalian MHC(Guillemot et al. 1988; Kaufman et al. 1995c, 1999a,1999b). The B-F/B-L region is small and simple, withonly 11 genes located in a 44 kb region that spans theclassical class II B to classical class I A genes. Moreover,different B haplotypes determine significant differencesin resistance to certain infectious pathogens as well asresponse to vaccines. We have attempted to relate thesestriking structural and functional features of the B-F/B-Lregion, suggesting that it functions as a “minimal essentialMHC” (Kaufman and Salomonsen 1997; Kaufman et al.1995c, 1999a, 1999b).
While B-LB cDNAs and genes were readily identifiedin chickens (Bourlet et al. 1987; Xu et al. 1989; Zoorob etal. 1990, 1993; Sung et al. 1993; Li et al. 1997, 1999;Pharr et al. 1998; Jacob et al. 2000), much effort has beenexpended in the unsuccessful search for the classical classII A gene (Kroemer et al. 1990). We found two classicalclass II B genes in the sequence of the chicken MHC, butwere unable to find a classical class II A gene nearby(Kaufman et al. 1999a, 1999b). Based on the patternsfound in one- and two-dimensional gel analysis ofimmunoprecipitates using alloantisera and monoclonalantibodies, it was clear that such a class II A gene mustexist. Interestingly, the 2-D gels showed arrays ofpolymorphic spots corresponding to class II b chains,but an invariant array of spots was taken to represent the achain (Guillemot et al. 1986). We therefore set out toidentify and characterize the a chain of the classical classII molecules of chicken.
606

Materials and methods
Protein chemistry
Spleens from 30 outbred chickens (obtained from a commercialslaughterhouse in Odense, Denmark) were homogenized in lysisbuffer containing NP-40, essentially as described previously(Kaufman et al. 1990). Class II molecules were isolated by affinitychromatography using monoclonal antibody 2G11 (IgG1k), whichrecognizes a monomorphic epitope on B-Lb (Kaufman et al. 1990;J. Salomonsen, unpublished). The a chain was separated from theheterogenous b chains by hydroxylapatite chromatography (Kauf-man 1983) and further purified by HPLC reverse phase chroma-tography. Tryptic peptides (generated by digestion at 37oCovernight under argon) were separated by HPLC reverse phasechromatography. The sequences of 30 amino acids (aa) from the N-terminus and 40 aa from a tryptic peptide were using an AppliedBiosystems 475A sequencer with an on-line PTH analyzer.
Isolation of the original probe
Total RNA was isolated from the chicken B cell line RP9 using thestandard guanidine isothiocyanate method (Sambrook et al. 1989)and total cDNA was made using poly-dT primer and a cDNAsynthesis kit (Pharmacia). The cDNA was amplified (30 cycles of94�C for 10 s, 45�C for 10 s and 72�C for 3 min, with a finalextension at 72�C for 9 min) on a Biometra Trio Thermocycler(Biometra, G�ttingen, Germany) using Taq polymerase (GeneAmpkit, Perkin Elmer) and degenerate oligonucleotides based on theprotein sequences including EcoRI sites: 2917 (50-CAYGRN-GAATTCARGCNGARTTYTAYCARMG-30), 2918 (50-TAYCA-RGAATTCGARGGNCCNGAYAARGC-30), 2921 (50-CNGGN-GTNACGAATTCYTGYTGNG-30) and 2922 (50-CNGCYTC-NGCNGGGAATTCNGCNARYTC-30). Gel-purified products ofthe expected size were digested with EcoRI and a resulting 110 bpfragment was cloned into the Bluescript vector (Stratagene).Double-stranded DNA was sequenced using the chain-terminationmethod with Sequenase (United States Biochemicals) and 35S-a-dATP.
Isolation of cDNA clones
Poly(A)+ RNA was isolated from bursa of CB chickens (kept at theBasel Institute for Immunology, Basel, Switzerland) using aFastTrack mRNA kit (Invitrogen). The mRNA preparation wastreated with DNase before cDNA synthesis and library constructionusing the Lambda ZAP cloning system (Stratagene). For eachscreen, approximately 500,000 plaques were lifted onto nitrocel-lulose filters, prehybridized (0.1 mg/ml single-stranded DNA, 35%formamide, 10% dextran sulfate, 5% SSPE, 5% Denhardt’s solutionand 0.1% SDS at 42�C) and then hybridized using probes labeledwith 32P-a-dCTP by random priming (Oligo-labeling kit, Pharma-cia).
Amplification from cDNA
mRNA was isolated from the spleens of line 72, C.B4, WL and Nchickens (kept at the Institute for Animal Health, Compton, UK.)using oligo-dT magnetic beads (Dynal) according to the manufac-turer’s instructions. Reverse transcriptase-polymerase chain reac-tion (RT-PCR) was performed using an oligo-dT primer andSuperscript II reverse transcriptase (Gibco BRL). After reversetranscription, the cDNA was amplified (96�C for 1 min, then onecycle of 96�C for 30 s, 50�C for 20 s, 72�C for 40 s followed by 22cycles of 96�C for 30 s, 72�C for 1 min) on a Biometra TrioThermocycler using Pwo proof-reading polymerase (Hybaid),forward primer c1010 (50-GGGCGTCCATATGGTCCTGAAGC-CGCAC-30) and reverse primer c1001 (50-AACGGATCCTCAC-CACATCCTCT GGAC-30). These primers flank exons 2 and 3
(corresponding to the a1 and a2 domains) with added NdeI andBamHI enzyme sites, respectively (underlined). The products weregel-purified and approximately 500 ng of each were sequenceddirectly using 3.2 pmol of the forward and reverse primers with theBig Dye version 1 sequencing kit on a 377 sequencer (ABI).
Amplification from genomic DNA
Genomic DNA was prepared by the standard SDS/proteinase Kmethod (Sambrook et al. 1989) from erythrocytes of individualchickens from the CB, CC, GB2, H.B2, H.B14, H.B15, H.B19 andH.B21 White Leghorn lines (kept at Foulum Research Centre,Denmark), the New Hampshire line (bought from the Danish StateSerum Institute), the IsaBrown line (courtesy of Tøttrup/Rasmussenfamily of Løvspringsvej, Charlottenlund) as well as a chicken fromrural Tanzania (courtesy of Dr. A. Permin, Network for Small-holder Poultry Development, Royal Veterinary and AgriculturalUniversity, Denmark) and a Red Jungle fowl (courtesy of theCopenhagen Zoo). The DNA samples were amplified (94�C for2 min, followed by 40 cycles of 94�C for 1 min, 50�C for 1 min and72�C for 1 min, with a final extension of 72�C for 10 min) on aGenius Thermocycler (Techne) using Taq polymerase (GeneAmpkit), jsa1 primer (50-CCGTCCTGAAGCCGCACGTGC-30) andjsa7 primer (50-CTGCTGGGAGCGGTTGGAG-30). The productswere gel-purified, cloned into the TA-vector (Invitrogen) andsequenced using the chain-termination method with Sequenase and35S-a-dATP.
Genomic DNA was also prepared by a modified SDS/proteinaseK method (Miller et al. 1988) from erythrocytes of individualchickens from lines 61, 72, 15I and N (kept at the Institute forAnimal Health, Compton, UK.). The DNA samples were amplified(96�C for 1 min, followed by 30 cycles of 96�C for 30 s, either66�C for exon 3 or 62�C for exon 4 for 20 s followed by 72�C for30 s) on a Biometra Trio Thermocycler using Pwo proof-readingpolymerase, and primers c1011 (50-ACCCCCGAGCTGGCTCT-GTT-30) and c1228 (50-CCTCGGCCCCCCAATGCCT-3’) forexon 3 (a2 domain) and primers c1030 (50-AGCCGGAAGTTC-CGGAGCCG-3’) and c1031 (50- GAGCAGCCCCGGTTGGCGG-30) for exon 4 (transmembrane and cytoplasmic domains). Theproducts were gel-purified using a MinElute kit (Qiagen) andapproximately 500 ng were sequenced directly using 5 pmol of theforward and reverse primers with the DYEnamic ET TerminatorCycle Sequencing kit (Amersham) on a 377 sequencer.
Southern blots
Genomic DNA from 50 individuals of 13 inbred White Leghornlines and sublines (CB, CC, 61, GB2, H.B14A, H.B14B, H.B14Cand H.B14D, H.B15, H.B19ov+, H.B19ov-, H.B21A and H.B21B;then available at the Basel Institute for Immunology) was prepared(Miller et al. 1988), and digested with SacI, KpnI or MspI. Afterelectrophoresis and transfer to nylon filters (Zeta-Probe, Bio-Rad)in 0.4 n NaOH, the blots were prehybridized and hybridized usingthe jsa probe as described above. The jsa probe (covering nearly thewhole coding region) was amplified from the cDNA clones 9 or 11using primers jsa1 and jsa4 (50-CCCCGGTTGGCGGTTGGCGGC-30) and purified using a GlassMax DNA isolation spin cartridge(Gibco/Life Technologies).
Mapping by Southern blot and sequence analysis
Genomic DNA from the parents and 56 offspring of the Comptonreference cross (a back-cross family between a line N sire and anF1 hen of line N and line 15I; Bumstead and Palyga 1992) was usedfor mapping by two methods. First, these samples were digestedwith SacI, MluI or BamHI and analyzed by Southern blots using thejsa probe as described above, as well as the probes F3 (chickenclass I cDNA) and pRA5 (chicken b2m cDNA) which have beendescribed previously (Kaufman et al. 1992). Second, fragments
607

corresponding to exon 3 (a2 domain) were amplified from thesesamples using the primers and conditions described above, and thengel-purified and sequenced directly as described above. Thefrequency of recombination was analyzed using the programMAP MANAGER (Manly 1993). Map distances were calculatedwith the Haldane map function (cM).
Sequence alignments, linear comparisonsand three-dimensional comparisons
Jotan Hein and Clustal algorithms (Hein 1990; Higgins and Sharp1989) from the DNAStar/Lasergene package were used forcomparisons. Alignments for sequence figures were modified byinspection. Coordinates from the structure of human class IImolecule DR1 (PDB entry 1DLH, Stern et al. 1994) and SwissPdbViewer v3.7b2 (http://www.expasy.ch/spdbv) were used as thebasis to display similarities and differences between B-LA, DRAand Ea chains.
Results
We isolated chicken class II molecules from outbredchickens by affinity chromotography using a monoclonalantibody, and then separated the two chains by SDS/hydroxylapatite chromotography. N-terminal sequencingby Edman degradation showed that one broad peak
contained many sequences similar to class II b chains(mammalian and particularly chicken B-LB), while theother broad peak consisted of just one sequence (NH2-VLKPHVLLQAEFYQRSEGPDKAxAQFGFWF) withsimilarities to mammalian class II a chains, indicatingthat it was B-LA. A tryptic peptide of B-LA was alsosequenced (NH2-SQQDFVTPELALFPAEAVSLEEPN-VLICYADKFDPPVVxM), and oligonucleotide primersbased on these two peptides were used to amplify afragment, which was then used to screen cDNA libraries.Four different clones were eventually isolated from a CBline bursa library, of which clones 9 and 11 are near full-length, while clones JP4 and C40 start at the same site inthe middle of the coding region. All four clones haveidentical sequence in the coding region, but differentsequences in the 30-untranslated region, suggesting alter-native splicing of exons. In addition, clones 9 and 11 havecompletely different sequences in the 50-untranslatedregion (Fig. 1, see database entries).
Over the whole coding region, both the nucleotide (nt)and amino acid (aa) sequences show strong similarity tomammalian, frog and fish class II A sequences (Fig. 1).The major features of all class II a chain sequences arepresent, with a signal sequence followed by a non-immunoglobulin-like a1 domain, an immunoglobulin C
Fig. 1 B-LA amino acid (aa) sequence compared to humanDRA*0101, DQA1*0101 and DPA1*01031 aa sequences. The firstblock includes the signal sequence (B-LA aa �22 to �1, DRA aa�23 to �1), the second block includes the a1 domain (1–89, 1–85),the third block includes the a2 domain (90–182, 86–178), and thefourth block includes the connecting peptide, transmembrane andcytoplasmic regions (183–232, 179–229). Identities are indicatedby a dash; gaps by a dot. Numbers to the left of the B-LA sequenceindicate %G+C in the wobble bases of codons; numbers to the leftof DR, DQ and DP sequences indicate % nt identity with B-LA (ascalculated by Jotan Hein method); numbers to the right of DR, DQand DP sequences indicate the number of aa identities and % aaidentities with B-LA. Structural features from HLA-DR1 (Brown etal. 1993; Stern et al. 1994; Hennecke et al. 2000; Wang et al. 2001)as follows: = b-strand; + a-helix; n N-linked glycan attachmentsite; p peptide contact; t T-cell receptor contact; 4 potential CD4
contact, d “dimer-of-dimers” contact. The transmembrane region isoverlined. S above the sequence of the a1 domain indicates the S/Gallelic residue (with AGC for serine in all inbred lines at theInstitute for Animal Health, but with GGC for glycine found in theC line originating from Reaseheath and kept at Compton as well asdescendents such as CB and CC kept at Basel Institute forImmunology and elsewhere; see Jacob et al. 2000 for descriptionand citation of lines). Asterisks above the sequence indicatepositions with silent nucleotide variation, with GTC for valine 169,CAT for histidine 171 and GGG for glycine 176 found in all lineswith two exceptions. The C line and derivatives have GTG forvaline 169, CAC for histidine 171 and GGA for glycine 176. The Nline chickens in the Compton reference family (but not other N linechickens examined) had GGA for glycine 176 (without the otherdifferences), which was the character used as the basis for mappingby sequence comparison
608

(IgC) region-like a2 domain with a 56 aa disulfide loop,an acidic connecting peptide, a transmembrane regionwith a free sulfhydryl and spaced glycines, and a shortcytoplasmic tail. Certain features of the classical class II achains of mammals are also present, including criticalpeptide-binding residues, a potential glycosylation site atB-LA residue 82 (equivalent to DRA residue 78) and nodisulfide bridge in the a1 domain. The residues thought tocontact CD4 are either identical or very similar in size andchemical nature, suggesting that class II a chain interac-tion with CD4 is conserved. However, most of thecontacts implicated in the formation of potential class II“dimer-of-dimers” are not conserved. Another interestingdifference is the loop between the first two b-strands (B-LA residues 15–22, equivalent to DRA residues 15–18)which is four or five amino acids longer in B-LAcompared to mammalian or fish sequences. In contrast tothe coding region, the sequence of the leader peptide andthe untranslated regions are not strongly related to anysequence in the database, including class II sequences.
There is a high G+C bias in the sequence (only 14%A+T wobble bases in the coding sequence compared tothe approximately 50% expected). This property is incommon with many other chicken genes, includingchicken class II B, b2-microglobulin and some exons ofthe class I genes (Kaufman et al. 1992).
Overall, the sequence of B-LA is more similar to DRAthan to other human class II A isotypes, when comparingthe whole coding sequence by two independent methods.The Clustal algorithm gave roughly 41% aa and 47% ntsimilarity with DRA*0101, 38% and 42% withDQA1*0101, 41% and 46% with DPA1*01031, 20%and 29% with DMA. The Jotan Hein algorithm gave 47%aa and 58% nt similarity with DRA*0101, 42% and 53%with DQA1*0101, 44% and 56% with DPA1*01031, 27%and 47% with DMA. Thus, both methods show the totalB-LA sequence to be slightly more similar to DRA thanDPA (which is very closely related to DRA), andconsiderably more similar to DRA than DQA or DMA.
The similarity between B-LA and DRA is not evenlydistributed through the sequence, but is concentrated incertain parts (a property also found in the relationship ofthe chicken and mammalian class I molecules; Kaufman
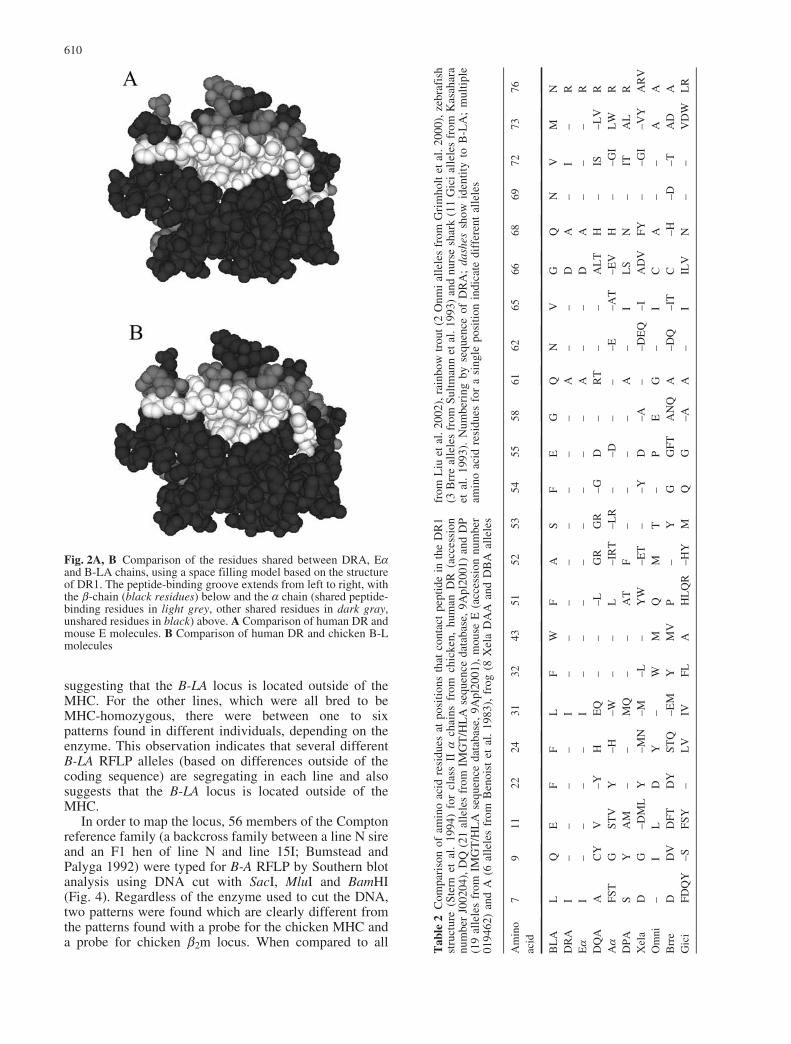
et al. 1992). The sequences of the signal sequence,connecting peptide, transmembrane region and cytoplas-mic tail compared individually show little or no differ-ences in their similarity to any particular human isotype.In contrast, the a1 and a2 domains are both more similarto DRA than the other isotypes, with the differentialsimilarity in the a1 domain being particularly marked(Fig. 1, Table 1). Indeed, the peptide-binding residues ofB-LA are virtually identical to DRA sequences fromvarious mammals, but not to mammalian DQA or DPAsequences, nor to frog and fish A sequences (Table 2).This point is particularly evident from views of thepeptide-binding domain from the perspective of the T-cellreceptor. Virtually all of the residues on this face of theclass II a chain that are shared between human DRA andmouse Ea are also shared with chicken B-LA (Fig. 2).
Polymorphism and variation in B-LA were examinedby sequence analysis of cDNA and genomic DNA.Fragments corresponding to the a1, a2, transmembraneand cytoplasmic domains were amplified from six inbredexperimental lines of White Leghorn chickens kept atCompton, and fragments corresponding to the a1 domainwere amplified from 12 individual chickens of widelyvarying origins (including Red Jungle fowl, the presumedwild ancestor of domestic chickens). Only four nucleotidedifferences were found, with three silent changes in the a2domain (valine 169, histidine 171, glycine 176) and oneproductive change in the a1 domain (see Fig. 1). Theamino acid difference (serine/glycine in B-LA aa 79corresponding to DRA aa 75, Fig. 1) between the twoputative alleles is found on the a-helix pointing up out ofthe groove, and the equivalent residue in DR moleculesdoes not affect peptide-binding or participate as a T-cellreceptor contact.
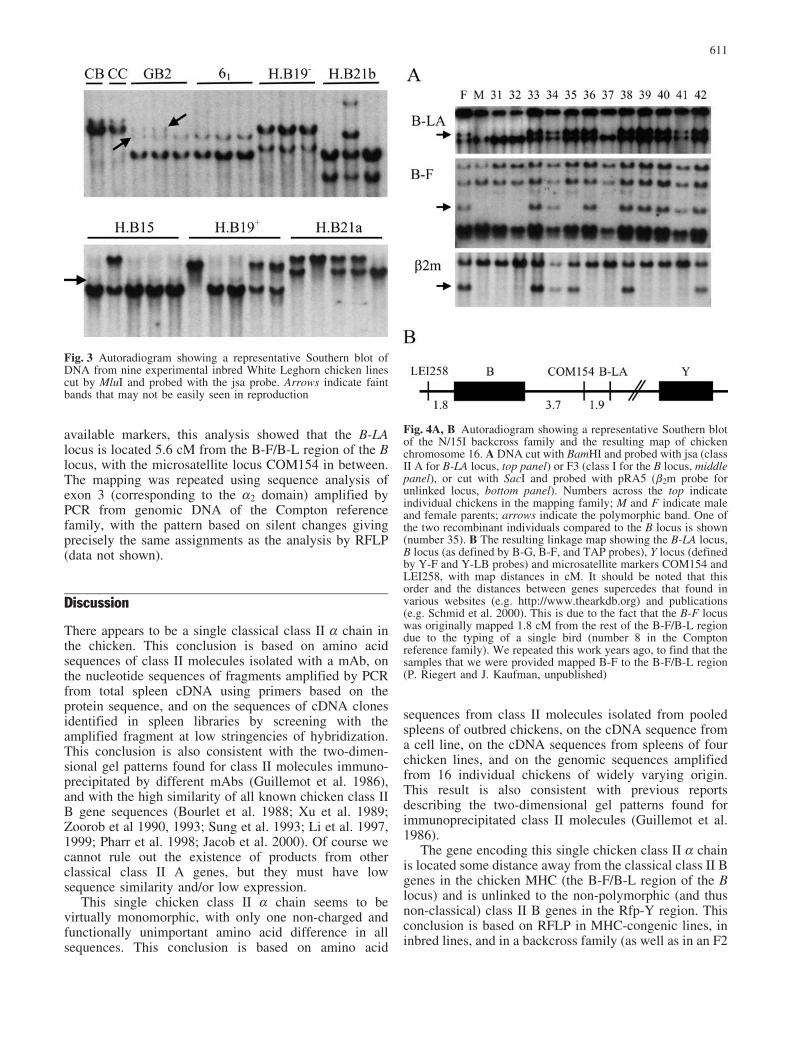
In contrast to the near monomorphism found for thesequence, Southern blot analysis of genomic DNA usingthe whole B-LA sequence as a probe shows substantialvariation (Fig. 3). Altogether, 50 individuals from 13chicken lines were examined, and restriction fragmentlength polymorphisms (RFLP) were found for all threerestriction enzymes employed. Individuals from the twohighly inbred MHC-congenic lines (CB and CC) showedthe same pattern with all restriction enzymes employed,
Table 1 Sequence pair differ-ences between human class IIisotypes (DRA*0101,DQA1*0101 and DPA1*01031;DRB*0101, DQB1*0101 andDPB1*01031) and chickenclass II sequences, as deter-mined using the Clustal methodwith PAM250 residue weight-ing table. PBD Peptide-bindingdomain; PBR peptide-bindingresidues; non-PBR residues inthe PBD which do not contactpeptide; non-PBD coding re-gion other than the PBD
a Chain b Chain
– BL DR DQ – BL DR DQ
PBD DR 50.0 – – DR 53.9 – –DQ 34.9 48.8 – DQ 48.3 62.1 –DP 39.3 56.0 47.6 DP 49.9 59.1 59.1
PBR DR 92.0 – – DR 7.9 – –DQ 82.0 84.0 – DQ 4.5 10.5 –DP 88.0 88.0 82.0 DP 5.6 8.6 12.9
non-PBR DR 33.1 – – DR 46.1 – –DQ 29.1 40.5 – DQ 43.8 51.6 –DP 27.4 44.0 41.7 DP 39.3 50.5 48.4
non-PBD DR 43.8 – – DR 48.6 – –DQ 40.3 59.3 – DQ 50.7 70.2 –DP 41.0 62.8 65.5 DP 52.2 65.4 69.4
609
suggesting that the B-LA locus is located outside of theMHC. For the other lines, which were all bred to beMHC-homozygous, there were between one to sixpatterns found in different individuals, depending on theenzyme. This observation indicates that several differentB-LA RFLP alleles (based on differences outside of thecoding sequence) are segregating in each line and alsosuggests that the B-LA locus is located outside of theMHC.
In order to map the locus, 56 members of the Comptonreference family (a backcross family between a line N sireand an F1 hen of line N and line 15I; Bumstead andPalyga 1992) were typed for B-A RFLP by Southern blotanalysis using DNA cut with SacI, MluI and BamHI(Fig. 4). Regardless of the enzyme used to cut the DNA,two patterns were found which are clearly different fromthe patterns found with a probe for the chicken MHC anda probe for chicken b2m locus. When compared to all
Fig. 2A, B Comparison of the residues shared between DRA, Eaand B-LA chains, using a space filling model based on the structureof DR1. The peptide-binding groove extends from left to right, withthe b-chain (black residues) below and the a chain (shared peptide-binding residues in light grey, other shared residues in dark gray,unshared residues in black) above. A Comparison of human DR andmouse E molecules. B Comparison of human DR and chicken B-Lmolecules
Tab
le2
Com
pari
son
ofam
ino
acid
resi
dues
atpo
siti
ons
that
cont
act
pept
ide
inth
eD
R1
stru
ctur
e(S
tern
etal
.19
94)
for
clas
sII
ach
ains
from
chic
ken,
hum
anD
R(a
cces
sion
num
ber
J002
04),
DQ
(21
alle
les
from
IMG
T/H
LA
sequ
ence
data
base
,9A
pl20
01)
and
DP
(19
alle
les
from
IMG
T/H
LA
sequ
ence
data
base
,9A
pl20
01),
mou
seE
(acc
essi
onnu
mbe
r01
9462
)an
dA
(6al
lele
sfr
omB
enoi
stet
al.
1983
),fr
og(8
Xel
aD
AA
and
DB
Aal
lele
s
from
Liu
etal
.200
2),r
ainb
owtr
out
(2O
nmi
alle
les
from
Gri
mho
ltet
al.2
000)
,zeb
rafi
sh(3
Brr
eal
lele
sfr
omS
ultm
ann
etal
.199
3)an
dnu
rse
shar
k(1
1G
ici
alle
les
from
Kas
ahar
aet
al.
1993
).N
umbe
ring
byse
quen
ceof
DR
A;
dash
essh
owid
enti
tyto
B-L
A;
mul
tipl
eam
ino
acid
resi
dues
for
asi
ngle
posi
tion
indi
cate
diff
eren
tal
lele
s
Am
ino
acid
79
1122
2431
3243
5152
5354
5558
6162
6566
6869
7273
76
BL
AL
QE
FF
LF
WF
AS
FE
GQ
NV
GQ
NV
MN
DR
AI
––
––
I–
––
––
––
–A
––
DA
–I
–R
Ea
I–
––
–I
––
––
––
––
A–
–D
A–
––
RD
QA
AC
YV
–YH
EQ
––
–LG
RG
R–G
D–
RT
––
AL
TH
–IS
–LV
RAa
FS
TG
ST
VY
–H–W
––
L–I
RT
–LR
––D
––
–E–A
T–E
VH
––G
IL
WR
DP
AS
YA
M–
–M
Q–
–A
TF
––
––
A–
IL
SN
–IT
AL
RX
ela
DG
–DM
LY
–MN
–M–L
–Y
W–E
T–
–YD
–A–
–DE
Q–I
AD
VF
Y–
–GI
–VY
AR
VO
mni
–I
LD
Y–
WM
QM
T–
PE
G–
IC
A–
–A
AB
rre
DD
VD
FT
DY
ST
Q–E
MY
MV
P–
YG
GF
TA
NQ
A–D
Q–I
TC
–H–D
–TA
DA
Gic
iF
DQ
Y–S
FS
Y–
LV
IVF
LA
HL
QR
–HY
MQ
G–A
A–
IIL
VN
––
VD
WL
R
610
available markers, this analysis showed that the B-LAlocus is located 5.6 cM from the B-F/B-L region of the Blocus, with the microsatellite locus COM154 in between.The mapping was repeated using sequence analysis ofexon 3 (corresponding to the a2 domain) amplified byPCR from genomic DNA of the Compton referencefamily, with the pattern based on silent changes givingprecisely the same assignments as the analysis by RFLP(data not shown).
Discussion
There appears to be a single classical class II a chain inthe chicken. This conclusion is based on amino acidsequences of class II molecules isolated with a mAb, onthe nucleotide sequences of fragments amplified by PCRfrom total spleen cDNA using primers based on theprotein sequence, and on the sequences of cDNA clonesidentified in spleen libraries by screening with theamplified fragment at low stringencies of hybridization.This conclusion is also consistent with the two-dimen-sional gel patterns found for class II molecules immuno-precipitated by different mAbs (Guillemot et al. 1986),and with the high similarity of all known chicken class IIB gene sequences (Bourlet et al. 1988; Xu et al. 1989;Zoorob et al 1990, 1993; Sung et al. 1993; Li et al. 1997,1999; Pharr et al. 1998; Jacob et al. 2000). Of course wecannot rule out the existence of products from otherclassical class II A genes, but they must have lowsequence similarity and/or low expression.
This single chicken class II a chain seems to bevirtually monomorphic, with only one non-charged andfunctionally unimportant amino acid difference in allsequences. This conclusion is based on amino acid
sequences from class II molecules isolated from pooledspleens of outbred chickens, on the cDNA sequence froma cell line, on the cDNA sequences from spleens of fourchicken lines, and on the genomic sequences amplifiedfrom 16 individual chickens of widely varying origin.This result is also consistent with previous reportsdescribing the two-dimensional gel patterns found forimmunoprecipitated class II molecules (Guillemot et al.1986).
The gene encoding this single chicken class II a chainis located some distance away from the classical class II Bgenes in the chicken MHC (the B-F/B-L region of the Blocus) and is unlinked to the non-polymorphic (and thusnon-classical) class II B genes in the Rfp-Y region. Thisconclusion is based on RFLP in MHC-congenic lines, ininbred lines, and in a backcross family (as well as in an F2
Fig. 3 Autoradiogram showing a representative Southern blot ofDNA from nine experimental inbred White Leghorn chicken linescut by MluI and probed with the jsa probe. Arrows indicate faintbands that may not be easily seen in reproduction
Fig. 4A, B Autoradiogram showing a representative Southern blotof the N/15I backcross family and the resulting map of chickenchromosome 16. A DNA cut with BamHI and probed with jsa (classII A for B-LA locus, top panel) or F3 (class I for the B locus, middlepanel), or cut with SacI and probed with pRA5 (b2m probe forunlinked locus, bottom panel). Numbers across the top indicateindividual chickens in the mapping family; M and F indicate maleand female parents; arrows indicate the polymorphic band. One ofthe two recombinant individuals compared to the B locus is shown(number 35). B The resulting linkage map showing the B-LA locus,B locus (as defined by B-G, B-F, and TAP probes), Y locus (definedby Y-F and Y-LB probes) and microsatellite markers COM154 andLEI258, with map distances in cM. It should be noted that thisorder and the distances between genes supercedes that found invarious websites (e.g. http://www.thearkdb.org) and publications(e.g. Schmid et al. 2000). This is due to the fact that the B-F locuswas originally mapped 1.8 cM from the rest of the B-F/B-L regiondue to the typing of a single bird (number 8 in the Comptonreference family). We repeated this work years ago, to find that thesamples that we were provided mapped B-F to the B-F/B-L region(P. Riegert and J. Kaufman, unpublished)
611

family; Kaufman et al. 1995a). In contrast to the sequencedata for a single non-polymorphic gene, the RFLPpatterns were complicated and highly polymorphic,suggesting considerable sequence variation in neighbor-ing non-coding DNA. Such data argues for caution inusing RFLP to infer polymorphism of the coding regionof a gene. One explanation may be that there are severalgene fragments including exon 1 located near theauthentic gene, as found by amplification from genomicDNA using PCR (Salomonsen et al. 2001). We wereunable to isolate the authentic B-LA gene from a numberof genomic libraries (including the East Lansing, Wa-geningen and RPMI BAC libraries). However, Southernblots indicate that there is only one copy of exon 3 in thechicken genome (which is why we re-mapped the geneusing polymorphisms in exon 3). In any case, therecombinational separation between class II A and Bgenes is consistent with a physical separation of at least50 kb (with many genes in between) determined fromcloned cosmid DNA (Guillemot et al. 1988).
This large physical separation of a class II A gene andthe class II B genes whose products are known to formheterodimers (chicken B-LA and B-LB genes) is unprece-dented. For instance, in humans, the DRA gene is locatedbeside the DRB genes and pseudogenes, DQA1 besideDQB1, DQA2 beside DQB2, DMA beside DMB, DPA1beside DPB1 and DPA2 beside DPB2; and only DNA/DZA is separated from DOB by the apparent insertion ofLMP, TAP, DM and RING3 genes (Beck and Trowsdale1999). However, recombinational separation is wellknown for the mouse E alpha-chain and E beta-chaingenes, and is predicted for DRA and DRB genes (Germainet al. 1985). As originally pointed out by Germain, geneswhose products interact and are separated by frequentrecombination are not both polymorphic in the interactingportions; one gene evolves towards a non-polymorphic“average best fit” to the polymorphic partner gene(s)whose alleles switch frequently. Thus, the lack ofpolymorphism of the B-LA gene is consistent with thefact that the B-LA gene is 5.6 cM away from thepolymorphic B-LB genes, just as the b2m gene ismonomorphic and unlinked to the polymorphic class Igenes.
While the classical class II isotypes (DR, DQ and DP)are well-conserved among mammals, it has often beenremarked that these isotypes are not found in othervertebrate groups. Our results show that the class II achain associated with the B-LA chains in chicken has thesequence, lack of polymorphism and recombinationalseparation expected for a DR-like class II a chain.Significantly, chicken class II B genes have beeninterpreted to be no more similar to any one particularhuman class II isotype, based on comparisons of thewhole sequence (Zoorob et al. 1993). However, we findthat the peptide-binding domains of the chicken class II bchains (with or without the polymorphic residues incontact with the peptide) are clearly more like DRBsequences than the other human isotypes (Table 1). Thissuggests that both the B-LA and B-LB genes are ortho-
logues of the DR genes of mammals. The non-polymor-phic (and therefore non-classical) Y-LB genes located inthe Rfp-Y locus may also encode partners for the B-LAchain, given that the B-LA chain is apparently the onlywell-represented classical class II a chain in the chicken.
The presence of a DR-like A gene in the chickenimplies that the DR-like lineage is ancient; at least 300million years old based on divergence from the lineageleading to mammals (Kumar and Hedges 1998). In fact,N-terminal sequencing of class II a chains from asalamander, the Mexican axolotl, also reveals a DR-likesequence (D. Avila and J. Kaufman, unpublished),implying that the DR-lineage may be over 350 millionyears old. Most importantly, these results indicate that theconserved sequence features of the a chain (found in thepeptide-binding domain) are important in the function ofsuch DR-like molecules, rather than the lack of polymor-phism. This may indicate that the particular sequence ofpeptide-binding residues in the a chain of DR-likemolecules is selected for some as yet unknown reason.
Finally, our results provide further support for theconcept of a “minimal essential MHC” in the chicken(Kaufman and Salomonsen 1997; Kaufman et al. 1995c,1999a, 1999b). This view argues that many genes foundin the mammalian MHC are not present in the chickenMHC, so the MHC-determined immune response of anindividual chicken is dominated by those few remaininggenes. The fact that the classical class II A gene inchicken is located far away, both in recombinational andphysical distance, is consistent with this view. Moreover,the fact that there is a single monomorphic class II A geneleaves all variation of chicken class II molecules to theclass II B genes located in the MHC. While there are twoB-LB genes, both of which are polymorphic, only onegene is strongly expressed at the level of RNA and protein(Pharr et al. 1998; Jacob et al. 2000).
Thus, as in chicken class I molecules, there should be asingle dominantly-expressed class II molecule expressedby any given chicken MHC haplotype, the peptide-binding specificity of which should be important in theMHC-determined level of class II-dependent responses.This prediction is consistent with the importance of thechicken MHC in the antibody response to killed viralvaccines and other relatively large immunogens (Bacon etal. 1987; Heller et al. 1991; N. Bumstead and J. Kaufman,unpublished). From this point of view, the genomicorganization of the class II genes in the chicken hassignificant functional ramifications.
Acknowledgements This work was begun at the Basel Institute forImmunology, and some of the authors would like to thankHoffmann-LaRoche for many years of financial support. Thiswork was also supported by a grant from Danish Natural ScienceResearch Council (11-0093-1) to J.S., and by core funding from theBiotechnology and Biological Sciences Research Council to J.K.The authors gratefully acknowledge Sally Rogers, Brian Walker,Kathryn Crouch, Pete Kaiser, and Shirley Ellis and an anonymousreviewer for valuable comments on this manuscript.
612

References
Bacon LD, Ismail N, Motta JV (1987) Allograft and antibodyresponses of 15I5 B congenic chickens. Prog Clin Biol Res238:219–233
Beck S, Trowsdale J (1999) Sequence organisation of the class IIregion of the human MHC. Immunol Rev 167:201–210
Benoist CO, Mathis DJ, Kanter MR, Williams VE 2nd, McDevittHO (1983) Regions of allelic hypervariability in the murine Aalpha immune response gene. Cell 34:169–177
Bourlet Y, Behar G, Guillemot F, Frechin N, Billault A, Chausse A-M, Zoorob R, Auffray C (1988) Isolation of chicken majorhistocompatibility complex class II (B-L) b chain sequences.EMBO J 7:1031–1039
Brown JH, Jardetzky TS, Gorga JC, Stern LJ, Urban RG,Strominger JL, Wiley DC (1993) Three-dimensional structureof the human class II histocompatibility antigen HLA-DR1.Nature 364:33–39
Bumstead N, Palyga J (1992) A preliminary linkage map of thechicken genome. Genomics 13:690–697
Dixon B, van Erp SH, Rodrigues PN, Egberts E, Stet RJ (1995)Fish major histocompatibility complex genes: an expansion.Dev Comp Immunol 19:109–133
Fillon V, Zoorob R, Yerle M, Auffray C, Vignal A (1996) Mappingof the genetically independent chicken major histocompatibilitycomplexes B and RFP-Y to the same microchromosome bytwo-color fluorescent in situ hybridization. Cytogenet CellGenet 75:7–9
Flajnik MF, Kasahara M (2001) Comparative genomics of theMHC: glimpses into the evolution of the adaptive immunesystem. Immunity15:351–362
Germain RN, Bentley DM, Quill H (1985) Influence of allelicpolymorphism on the assembly and surface expression of classII MHC (Ia) molecules. Cell 43:233–242
Grimholt U, Getahun A, Hermsen T, Stet RJ (2000) The majorhistocompatibility class II alpha chain in salmonid fishes. DevComp Immunol 24:751–763
Guillemot F, Turmel P, Charron D, Le Dourin NM, Auffray C(1986) Structure, polymorphism and biosynthesis of chickenMHC class II (B-L) antigens and associated molecules. JImmunol 137:1251–1257
Guillemot F, Billault A, Pourquie O, Behar G, Chausse A-M,Zoorob R, Kreibich G, Auffray C (1988) A molecular map ofthe chicken major histocompatibility complex: the class II bgenes are closely-linked to the class I genes and the nucleolarorganizer. EMBO J 7:2775–2785
Hein JJ (1990) Unified approach to alignment and phylogenies.Methods Enzymol 183:626–645
Heller ED, Uni Z, Bacon LD (1991) Serological evidence for majorhistocompatibility complex (B complex) antigens in broilersselected for humoral immune response. Poultry Sci 70:726–732
Hennecke J, Carfi A, Wiley DC (2000) Structure of a covalentlystabilized complex of a human alpha beta T-cell receptor,influenza HA peptide and MHC class II molecule, HLA-DR1.EMBO J 19:5611–5624
Higgins DG, Sharp PM (1989) Fast and sensitive multiple sequencealignments on a microcomputer. CABIOS 5:151–153
Jacob JP, Milne S, Beck S, Kaufman J (2000) The major and aminor class II beta-chain (B-LB) gene flank the Tapasin gene inthe B-F /B-L region of the chicken major histocompatibilitycomplex. Immunogenetics 51:138–147
Kasahara M, Vazquez M, Sato K, McKinney EC, Flajnik MF(1992) Evolution of the major histocompatibility complex:isolation of class II A cDNA clones from the cartilaginous fish.Proc Natl Acad Sci USA 89:6688–6692
Kasahara M, McKinney EC, Flajnik MF, Ishibashi T (1993) Theevolutionary origin of the major histocompatibility complex:polymorphism of class II alpha chain genes in the cartilaginousfish. Eur J Immunol 23:2160–2165
Kaufman J (1983) Proteolytic dissection of MHC class II antigenstructure. Dissertation, Harvard University
Kaufman J, Salomonsen J (1997) The “minimal essential MHC”revisited: both peptide-binding and cell surface expression levelof MHC molecules are polymorphisms selected by pathogens inchickens. Hereditas 127:67–73
Kaufman J, Ferrone S, Flajnik M, Kilb M, Volk H, Parisot R (1990)MHC-like molecules in some nonmammalian vertebrates canbe detected by some cross-reactive monoclonal antibodies. JImmunol 144:2273–2280
Kaufman J, Andersen R, Avila D, Engberg J, Lambris J,Salomonsen J, Welinder K, Skjødt K (1992) Different featuresof the MHC class I heterodimer have evolved at different rates:chicken B-F and b2-microglobulin sequences reveal invariantsurface residues. J Immunol 148:1532–1546
Kaufman J, Bumstead N, Miller M, Riegert P, Salomonsen J(1995a) The chicken class II a gene is located outside of the Bcomplex. In: Davison TF, Bumstead N, Kaiser P (eds)Advances in avian immunology research. Carfax, Abingdon,pp 119–127
Kaufman J, Salomonsen J, Flajnik MF (1995b) Evolutionaryconservation of Mhc class I and class II molecules: different yetthe same. Semin Immunol 6:411–424
Kaufman J, V�lke H, Wallny H-J (1995c) A “minimal essentialMHC” and an “unrecognized MHC”: two extremes in selectionfor polymorphism. Immunol Rev 143:63–88
Kaufman J, Jacob J, Shaw I, Walker B, Milne S, Beck S,Salomonsen J (1999a) Gene organization determines evolutionof function in the chicken MHC. Immunol Rev 167:101–117
Kaufman J, Milne S, G�bel TWF, Walker BA, Jacob JP, Auffray C,Zoorob R, Beck S (1999b) The chicken B locus is a minimalessential major histocompatability complex. Nature 401:923–925
Kroemer G, Bernot A, Behar G, Chausse A-M, Gastinel, L-N,Guillemot F, Park I, Thoraval P, Zoorob R, Auffray C (1990)Molecular genetics of the chicken MHC: current status andevolutionary aspects. Immunol Rev 113:119–146
Kumar S, Hedges SB (1998) A molecular timescale for vertebrateevolution. Nature 392:917–920
Li L, Johnson LW, Ewald SJ (1997) Molecular characterisation ofmajor histocompatibiilty complex (B) haplotypes in broilerchickens. Anim Genet 28:258–267
Li L, Johnson LW, Livant EJ, Ewald EJ (1999) The MHC of abroiler chicken line: serology, B-G genotypes and B-F/B-LBsequences. Immunogenetics 49:215–224
Liu Y, Kasahara M, Rumfelt LL, Flajnik MF (2002) Xenopus classII A genes: studies of genetics, polymorphism, and expression.Dev Comp Immunol 26:735–750
Manly KF (1993) A Macintosh program for storage and analysis ofexperimental genetic-mapping data. Mamm Genome 4:303–313
Marsh SGE, Parham P, Barber LD (2000) The HLA facts book.Academic Press, London
Miller MM, Goto R, Bernot A, Zoorob R, Auffray C, Bumstead N,Briles WE (1994) Two MHC class I and two MHC class IIgenes map to the chicken Rfp-Y system outside the B complex.Proc Natl Acad Sci USA 91:4397–4401
Miller MM, Goto RM, Taylor RL, Zoorob R, Auffray C, BrilesWE, Briles RW, Bloom SE (1996) Assignment of Rfp-Y to thechicken histocompatibility complex/NOR microchromosomeand evidence for high-frequency recombination associated withthe nucleolar organizer region. Proc Natl Acad Sci USA93:3958–3962
Miller SA, Dykes DD, Polesky HF (1988) A simple salting outprocedure for extracting DNA from nucleated cells. NucleicAcids Res 16:215
Murray BW, Shintani S, Sultmann H, Klein J (2000) Majorhistocompatibility complex class II A genes in cichlid fishes:identification, expression, linkage relationships, and haplotypevariation. Immunogenetics 51:576–586
Ono H, Klein D, Vincek V, Figueroa F, O’Huigin C, Tichy H,Klein J (1992) Major histocompatibility complex class II genesof zebrafish. Proc Natl Acad Sci USA 89:11886–11890
613

Pharr GT, Dodgson JB, Hunt HD, Bacon LD (1998) Class II MHCcDNAs in 15I5 B-congenic chickens. Immunogenetics 5:350–354
Salomonsen J, Kaufman J, Laursen C, Skjødt K, Vachek PD (2001)The chicken class II alpha locus BLA. In: Schat KA (ed)Current progress in avian immunology research. Proceedings ofthe 6th Avian Immunology Research Group meeting, Ithaca, 8–10 October 2000. Am Assoc Avian Pathol, Pennsylvania, pp192–195
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: alaboratory manual. Cold Spring Harbor Press, New York
Schmid M, Nanda I, Guttenbach M, Steinlein C, Hoehn H, SchartlM, Haaf T, Weigend S, Fries R, Buerstedde J-M, Wimmers K,Burt DW, Smith J, O’Hara S, Law A, Griffin DK, Bumstead N,Kaufman J, Thomson PA, Burke TA, Groenen MAM, Crooij-mans RPMA, Vignal A, Fillion V, Morrison M, Pitel F, Tixier-Boichard M, Ladjali-Mohammedi K, Hillel J, Maki-Tanila A,Cheng HH, Delany ME, Burnside J, Mizuno S (2000) Firstreport on chicken genes and chromosomes. Cytogenet CellGenet 90:169–218
Steinmetz M, Minard K, Horvath S, McNicholas J, Srelinger J,Wake C, Long E, Mach B, Hood L (1982) A molecular map ofthe immune response region from the major histocompatibilitycomplex of the mouse. Nature 300:35–42
Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG,Strominger JL, Wiley DC (1994) Crystal structure of thehuman class II MHC protein HLA-DR1 complexed with aninfluenza virus peptide. Nature 368:215–221
Sultmann H, Mayer WE, Figueroa F, O’Huigin C, Klein J (1993)Zebrafish Mhc class II alpha chain-encoding genes: polymor-phism, expression, and function. Immunogenetics 38:408–20
Sung AM, Nordskog AW, Lamont SJ, Warner CM (1993) Isolationand characterization of cDNA clones for chicken majorhistocompatibility complex class II molecules. Anim Genet24:227–233
Wang J-h, Meijers R, Xiong Y, Liu J-h, Sakihama T, Zhang R,Joachimiak A, Reinherz EL (2001) Crystal structure of thehuman CD4 N-terminal two-domain fragment complexed to aclass II MHC molecule. Proc Natl Acad Sci USA 98:10799–10804
Wright H, Ballingall KT, Remond J (1994) The D-Y subregion ofthe sheep MHC contains an A/B gene pair. Immunogenetics40:230–234
Xu Y, Pitcovski J, Peterson L, Auffray C, Bourlet Y, Gerndt BM,Nordskog AW, Lamont SJ, Warner CM (1989) Isolation andcharacterization of three class II MHC genomic clones from thechicken. J Immunol 142:2122–2132
Zoorob R, Behar G, Kroemer G, Aufray C (1990) Organization of afunctional chicken class II B gene. Immunogenetics 31:179–187
Zoorob R, Bernot G, Renoir DM, Choukri F, Auffray C (1993)Chicken major histocompatibility complex class II B genes:analysis of interallelic and inter-locus sequence variance. Eur JImmunol 23:1139–1145
614
Copyright © 2022 FDOKUMEN




















