
The POT1 gene for yeast peroxisomal thiolase is subject to three different mechanisms of regulation
10
Molecular Microbiology (1992) 6(14), 1867-1875 The POT1 gene for yeast peroxisomal thiolase is subject to three different mechanisms of regulation J. Carlos Igual, Carmen Gonzalez-Bosch, Luis Franco and Jose E. P6rez-0rtin* Departamento de Bioquimiea y Biologia Moleeutar, Faeultades de Cieneias, Universitat de Valeneia, Spain. Summary The Saccharomyces cerevisiae P0T1 gene is, as are other yeast peroxisomal protein genes, inducible by tatty acids and repressible by glucose. We have now found that it is also induced during the stationary phase of the culture. To investigate these three regu- latory circuits, we have studied the mRNA levels of regulatory mutants as well as the changes in chro- matin structure upon gene activation. We conclude that the regulation of transcriptional activity in glu- cose repression, oleate induction, and stationary phase induction follow different molecular mecha- nisms. We suggest that this multiplicity of regulatory mechanisms may represent a general rule for fhe yeast peroxisomal protein genes. Introduction Peroxisomes are single-membrane-bound organelles present in almost all eukaryotic cells. They are involved in many biochemical functions, and both their abundance and metabolic activity depend on the cell type (Lazarow and Fujiki, 1985; Borst. 1989). Among the peroxisomal functions, the p-oxidafion of fatty acids has been described as occurring in hepatocytes (Osumi and Hashimoto, 1984), germinating seeds (Cooper and Beev- ers, 1969), plants (Gerhardt, 1983), and yeasts (Kunau et ai. 1988). Yeasts may be the organisms of choice for studying peroxisomes. not only because of tfie possibility of using recombinant DNA technology, but also because peroxiso* mal development and the metabolic functions of the organelle can be modified by the growth conditions (Veenhuis and Harder, 1987), It is known that some yeast species are able to grow on unusual carbon and nitrogen sources such as metfianol, alkanes, long-chain fatty acids, amines, D-amino acids, and uric acid. These Received 28 February, 1992; revised and accepted 6 April, 1992, *For correspondence, Tel. (6) 386 4385; Fax (6) 386 4372. compounds induce proliferation of peroxisomes, which contain the essential enzymatic activities required for the catabolism of those substances (Veenhuis and Harder, 1987). It was thought that Saccharomyces cerevisiae consti- tuted an exception to this general behaviour and that its peroxisomes were scarce and small (Avers and Feder- man. 1968: Parish, 1975). However, it has been shown recently that oleate induces peroxisomal proliferation in S. cerevisiae {Veenhuis etai, 1987). This fatty acid also induces the peroxisomal enzymatic activities, including those involved in the |i-oxidation pathway (Veenhuis et ai., 1987; Skoneczny ef ai. 1988), as it does in Candida species (Tanaka et ai, 1982). and Neurospora crassa (Kionka and Kunau. 1985). so that the effects of the pres- ence of oleate in the culture medium suggests co-regula- tion involving the induction of peroxisomal [i-oxidation enzymes and the proliferation of the organelle. Therefore. the peroxisomal [i oxidation system may be very useful for studying the mechanisms of peroxisome biogenesis and gene regulation in higher eukaryotes. We have recently identified and sequenced the S. cere- visiae POTI gene for the peroxisomal 3-oxoacyl CoA thi- olase (Igual et at.. 1991), We showed that it is repressible by glucose, as are other peroxisomal protein genes (Dmochowska etat., 1990; Erdmann etal., 1991). Thus, this group of genes seems to be reguiated by two mecha- nisms at least, namely glucose-repression and fatty acid induction. In this work we have studied the regulation of POT1 gene, showing that its expression is controlled by three different circuits, including the growth state. The study of gene expression in glucose-regulation mutants and the analysis of the chromatin structure under several physio- logical conditions allowed us to show that those circuits follow different molecular mechanisms. Results POTI is influenced by three different regulatory mechanisms Some peroxisomal enzymatic activities are subject to a dual mechanism of regulation: glucose repression, and fatty acid induction (Veenhuis and Harder. 1987; Skoneczny et ai, 1988; McCammon et ai, 1990) and it
Transcript of The POT1 gene for yeast peroxisomal thiolase is subject to three different mechanisms of regulation

Molecular Microbiology (1992) 6(14), 1867-1875
The POT1 gene for yeast peroxisomal thiolase issubject to three different mechanisms of regulation
J. Carlos Igual, Carmen Gonzalez-Bosch, LuisFranco and Jose E. P6rez-0rtin*Departamento de Bioquimiea y Biologia Moleeutar,Faeultades de Cieneias, Universitat de Valeneia, Spain.
Summary
The Saccharomyces cerevisiae P0T1 gene is, as areother yeast peroxisomal protein genes, inducible bytatty acids and repressible by glucose. We have nowfound that it is also induced during the stationaryphase of the culture. To investigate these three regu-latory circuits, we have studied the mRNA levels ofregulatory mutants as well as the changes in chro-matin structure upon gene activation. We concludethat the regulation of transcriptional activity in glu-cose repression, oleate induction, and stationaryphase induction follow different molecular mecha-nisms. We suggest that this multiplicity of regulatorymechanisms may represent a general rule for fheyeast peroxisomal protein genes.
Introduction
Peroxisomes are single-membrane-bound organellespresent in almost all eukaryotic cells. They are involved inmany biochemical functions, and both their abundanceand metabolic activity depend on the cell type (Lazarowand Fujiki, 1985; Borst. 1989). Among the peroxisomalfunctions, the p-oxidafion of fatty acids has beendescribed as occurring in hepatocytes (Osumi andHashimoto, 1984), germinating seeds (Cooper and Beev-ers, 1969), plants (Gerhardt, 1983), and yeasts (Kunau etai. 1988).
Yeasts may be the organisms of choice for studyingperoxisomes. not only because of tfie possibility of usingrecombinant DNA technology, but also because peroxiso*mal development and the metabolic functions of theorganelle can be modified by the growth conditions(Veenhuis and Harder, 1987), It is known that some yeastspecies are able to grow on unusual carbon and nitrogensources such as metfianol, alkanes, long-chain fattyacids, amines, D-amino acids, and uric acid. These
Received 28 February, 1992; revised and accepted 6 April, 1992, *Forcorrespondence, Tel. (6) 386 4385; Fax (6) 386 4372.
compounds induce proliferation of peroxisomes, whichcontain the essential enzymatic activities required for thecatabolism of those substances (Veenhuis and Harder,1987).
It was thought that Saccharomyces cerevisiae consti-tuted an exception to this general behaviour and that itsperoxisomes were scarce and small (Avers and Feder-man. 1968: Parish, 1975). However, it has been shownrecently that oleate induces peroxisomal proliferation inS. cerevisiae {Veenhuis etai, 1987). This fatty acid alsoinduces the peroxisomal enzymatic activities, includingthose involved in the |i-oxidation pathway (Veenhuis etai., 1987; Skoneczny ef ai. 1988), as it does in Candidaspecies (Tanaka et ai, 1982). and Neurospora crassa(Kionka and Kunau. 1985). so that the effects of the pres-ence of oleate in the culture medium suggests co-regula-tion involving the induction of peroxisomal [i-oxidationenzymes and the proliferation of the organelle. Therefore.the peroxisomal [i oxidation system may be very usefulfor studying the mechanisms of peroxisome biogenesisand gene regulation in higher eukaryotes.
We have recently identified and sequenced the S. cere-visiae POTI gene for the peroxisomal 3-oxoacyl CoA thi-olase (Igual et at.. 1991), We showed that it is repressibleby glucose, as are other peroxisomal protein genes(Dmochowska etat., 1990; Erdmann etal., 1991). Thus,this group of genes seems to be reguiated by two mecha-nisms at least, namely glucose-repression and fatty acidinduction.
In this work we have studied the regulation of POT1gene, showing that its expression is controlled by threedifferent circuits, including the growth state. The study ofgene expression in glucose-regulation mutants and theanalysis of the chromatin structure under several physio-logical conditions allowed us to show that those circuitsfollow different molecular mechanisms.
Results
POTI is influenced by three different regulatorymechanisms
Some peroxisomal enzymatic activities are subject to adual mechanism of regulation: glucose repression, andfatty acid induction (Veenhuis and Harder. 1987;Skoneczny et ai, 1988; McCammon et ai, 1990) and it

1868 J. C. Igual. C. GonzAlez-Boseh, L Franco and J. E. Perez-Ortfn
has been reported that this kind of regulation occurs atthe transcriptional level (Dmochowska etal., 1990; Erd-mann et al., 1991). To establish whether P0T1 is underthe same type of regulation, we studied the thiolase enzy-matic activity and the level of POTI mRNA in wild-typeMCY1389 and PO7"/-disrupted BQS20 strains grown inculture media containing different carbon sources. Pre-liminary experiments showed that thiolase activity inyeast was dependent on the growth state of the cell.Therefore, we determined the thiolase activity and POTImRNA level in extracts obtained from both exponentialand stationary phase cultures.
The results shown in Table 1 are compatible with theidea that POTI expression is activated by oleate andrepressed by glucose. Thiolase activity was only detectedin low-glucose media (YPg) and it clearly increased inoleic-acid-supplemented media. It is noteworthy that thespecific activity of thiolase was strongly increased byentering stationary phase. Under these conditions, thio-lase activity also increased in the presence of oleate. butthe effect of the fatty acid is less pronounced than in theexponentially growing cells.
The enzymatic activity assays were performed by usingacetoacetyl-CoA as substrate. Acetoacetyl-CoA offersthe obvious advantage of its ready accessibility, but it is asomewhat poor substrate for 3-oxoacyl-CoA thiolasebecause other enzymatic activities are able to split it.Therefore, we used the BQS20 strain, which possesses adisrupted POTI gene, as a control for the presence ofother thiolase activities. No thiolase activity was detectedin any sample of this strain under our assay conditions(results not shown), indicating that these conditions areactually specific for the peroxisomal thiolase studied inthis work.
The analysis of POTI mRNA confirmed that the regula-tion of enzymatic activity observed in the experiment inTable 1 takes place at the mRNA level (Fig. 1). Actually.in exponentially growing cells, the 1.6 kb transcript ofPOTI was not detected in the presence of glucose, evenwhen oieate was present. POTI transcript was present
Table 1. Thiolase specific activity (U mg"') in MCY1389 cell extracts.^
L-xpttncnlial phase sialionary phase
Meditim
YPgYPgtOYPGYPG+0YPGalYPGa).O
Exponential phase
<0.11.600
<0.10.8
Stationary phase
4JS10.0<G.1<0.1
1 ^4.3
a. The delermiration of enzymatic aciivity (in units per milligfam of pro-tein) and the composition of culture media are described in the Experi-mefJtal procedures.
POTl
ACTI
Fig. 1. Northern blot analysis of the transcriptional regulation ot the POTIgene. Total yeast RNA (60 tjg) was loaded on a i% agarose gel. electro-phoresed and transferred lo a nitrocellulose (ilter. which was hybridized loa radiolabelled probe, consisting of a 1,6 kb H;ndlll fragment from thePOTf 5'region. RNA was extracted from MCY1389 cells grown on mediacontaining different carbon sources: YPg or YPG (G - or +, respectively)in the absence or presence of oleate (O - or +. respectively). The cellswere harvested at exponential or stationary phase as indicated. The fillerwas rehybridized with an ACTI probe as a control.
only as a faint band in cells from low-glucose media with-out oleic acid, and its presence was clearly increased(over 9 times) when the glucose-deficient medium wassupplemented with oleic acid. RNA analysis also showsthat the expression of P0T1 gene increased at stationaryphase, and that oleate-induction in this case was less pro-nounced (about two- to threefold).
To study in further detail the induction of POTT whenthe cells enter stationary phase, we assayed the thiotaseactivity and mRNA level at different stages of growth inYPg medium. In the absence of oleate the enzymaticactivity remained below the detection limit until the earlystationary phase was reached and then increased,passed a maximum, and slowly decreased along the sta-tionary phase (Fig. 2, upper panel). The presence ofoleate in the culture medium resulted in an increase ofenzymatic activity, but no qualitative differences werenoticeable. Similar results were obtained in YPGalmedium (not shown). The mRNA analysis showed thatthe P0T1 transcript foliows a similar pattern (Fig, 2. lowerpanel), although, reasonably enough, the maximum ofPOTI mRNA preceded that of thiolase activity. The levelsof actin (ACTI) and URA3 mRNA were determined as acontrol, and it is clear that, in both cases, the amount oftranscript (high during the exponential phase), was signif-icantly reduced when the culture reached the stationaryphase. We also estimated as a control, the mRNA level ofFBP1. a glucose-repressible gene. It behaved as URA3and ACT1 showing that the accumulation of POTI mRNAat stationary phase is a peculiar feature of the gene andnot a general property of glucose-repressed genes.

Mechanisms of yeast POJ^ regulation 1869
10 20 30
culture time (h)
culture time (h)S 6 8 9 11 12 14 15 16 17 18 29 30 31 33
P0T1
ACT1
URA3
FBP1
Fig, 2. Growth dependence of thiolase activity and POT1 mRNA. Theupper part of the Fig. displays the ihiolase activity (•, O) and growth pro-tile (thin lines) of MCY1389 strain grown on YPg media with (•)(—•) orwithout (O) (..,) oleate. The Norlhero blots of RNA extracted at differentlimas during growth of MCY1389 cells on a YPg medium supplementedwith oleate are shown below. Electrophoresis and blotting conditions wereas in Fig. 1. The same filter was successively hybridized with P0T1,ACT1. URA3. and FBP1 probes.
SNFI and SSNQ but not M\G^ gene products arerequired for giucose regulatory mechanism of P0T1 gene
Glucose repression is a complex process that affects theactivity and synthesis ot enzymes of numerous metabolicpathways (Entian, 1986). Several mutants defective inglucose repression have been identified. Two of the best-characterized groups are: snf (sucrose non-fermenting)mutants, with defects in the derepression, and ssn (sup-pressors cf sn/mutations), incapable of repression in thepresence of glucose. Some of these genes have beenisolated, e.g. the SNFI and SSN6 genes (Celenza andCarlson, 1986; Schultz and Carlson, 1987). We wonderedwhether these two genes were also related to the tran-scriptional regulation of glucose-repressible peroxisomal
proteins. To address this question we analysed the tran-scription of POT1 gene in snf1 and ssn6 mutants.
The effects of these mutations on the transcription ofPOT1 gene are similar to those observed in other glu-cose-repressible genes. The transcription of the gene insnf1 mutants, as opposed to the wild-type strain, wasrepressed even in the absence of glucose (comparelanes 5-6 and 1-2 in Fig. 3). On the other hand. ssn6mutants failed to repress P0T1 in the presence of glu-cose, and significant amounts of transcript were ob-served, whereas POT1 mRNA was not detected in thewild-type strain under the same conditions (lanes 7 and 8and 3 and 4, respectively). Therefore, the glucose-regula-tion of POTI gene is mediated by the products of SNFIand SSN6 genes. It is interesting to note that glucoserepression and oleate induction seem to be independentmechanisms, the former being dominant over the latter. Inthe snfi mutant the presence of oleate in the mediumincreased the level of transcript detected, but the amountof mRNA detected was still very low (lane 6), whereas inthe ssn6 mutant oleate induction was not affected by thepresence of high concentrations of glucose in the medium(lane 8).
We also examined whether the MIG1 gene, a repressorof the glucose-repressible gene SUC2 (Nehlin andRonne, 1990) and G>4/. genes (Nehlin etai, 1991), affectPOTI transcription. Our results indicate that this does notoccur in the present instance, since P0T1 mRNA was notdetected in the migi mutant grown in YPG medium (lanes11 and 12).
w i l d t y pe s nfl xsii6 migl
POTI
ACT1
7 8 9 10 1 1 1 2
Fig. 3, Analysis ot POTI mRNA In mutant strains. The amount of POTImRNA was estimated in the wild type MCY1389 cells, as well as in Itiesnfi. ssn6. and mig1 mutant strains. Celts were harvested at mid-expo-nential phase. Symbols, electrophoresis. and Wotting conditions were asin Fig. 1. The Illter was rehybridized to an ACT1 gene probe as a control,

1870 J. C. Igual. C. Gonzalez-Bosoh, L. Franco and J. E. Perez-Orti'n
( 8 )V
SB
Fig. 4. Map of the yeas! chromosomal region conlained in pRB58. Thethick Ime represenis yeast DNA. and the thin line represents p8R322DNA. The relevant restnction sites are shown: B. BamHI; H, H/ndlll; S,Sail, St, Stu\. Below the map the strategic approaches for mapping IheDNase I and micrococcal nuclease cutting siies by indirecl end-labelling(Wu. 1980) are Indicated. Briefly, digested DNA was reslricted lo comple-tion with either Stu\ or Sal\. electrophoresed, and Southern-blotted. Theblots were then hybridized to a probe abutting on the reslnction site, Ihusallowing mapping of the original nuclease cutting sites. The thick lineslabelled SH and SB stand lor ihe probes, the iatler corresponding topBR322 DNA, and the thm arrows stand for the analysed regions follow-ing both strategic approaches. SH and SB.
Chromatin structure of P0T1 iocus is affected duringglucose starvation and stationary phase but not by oieateinduction
Changes in the chromatin structure in the SUC2 3" flankupon glucose deprivation led us to identify the P0T1 gene(Igual et ai., 1991). Upon further sequencing, an ORF of705 bp was also found between both genes. This ORF isconstitutively transcribed from the strand opposite toP0T1, only 57 bp away from the end of P0T1 codingsequence (our unpublished results). The changesobserved in chromatin occur at the intergenic areabetween this ORF and the P0T1 gene, in the middle of aregion characterized by a strict positioning of 13 nucleo-somes between two strong nuclease hypersensitive sites(Perez-Ortfn et ai., 1986: 1987). Therefore, we studiedthe chromatin structure in an attempt to cast some light onthe molecular mechanisms of the regulatory circuits.
To this end, indirect end-labelling experiments (Wu,1980) were used to localize the cutting sites of bothmicrococcat nuclease and DNase 1. The strategicapproaches used are shown in Fig. 4.
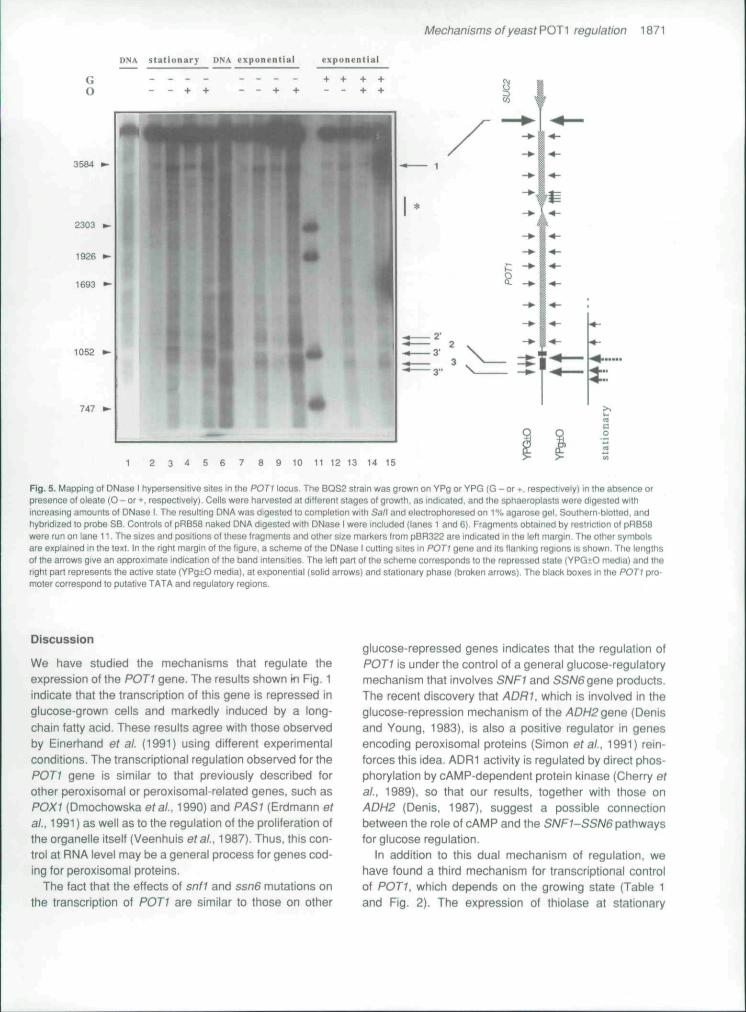
The DNase I experiments following strategy SB areshown in Fig. 5. Apart from many weakly sensitive sites,two main cutting sites were found in chromatin. Band 1corresponds to a constitutive hypersensitive site at -f2200(relative to the transiational start of P0T1). This site waspreviously detected in our studies on the SUC2 locus(P6rez-Ortin et al., 1986). The second zone, only sensi-tive in active cells, is located at the 5' flank of POTI and ismade up of several bands. In glucose-repressed samples(lanes 12-15) this region is not particularly sensitive andthe intensities of bands 2 (at -165), 2' (at -130). and 3 (at-290) were similar fo those of the ladder of bands alongthe POTI gene. In glucose-derepressed samples fromthe exponential phase (lanes 7-10), there is a general
increase in sensitivity and band no. 2' becomes moreexposed than 2. When the cells entered the stationaryphase, band 2' remained unaltered whereas band 3 wassplit into bands 3', at -250, and 3", at -340. The regionbetween the latter two bands, which corresponds to thepreviously observed band 3, is partially protected. In allthe cases a clearly protected zone of 35 bp centred at-220 is seen. The presence of oieate in the culturemedium had no noticeable effects on DNase I sensitivity.
The above-mentioned ladder of bands extendsbetween sites 1 and 2. These bands probably result fromDNase I digestion in the linkers of positioned nucleo-somes (Perez-Ortin etai., 1986: Lee and Garrard, 1991).An increase in DNase I sensitivity in the region between+1340 and -^1430 (marked with an asterisk in Fig. 5) wasobserved in transcriptionally active cells (lanes 2-5 and7-10).
The two strategic approaches indicated in Fig. 4 wereused to map the micrococcai nuclease cutting sites. Theresults with strategy SB are shown in Fig. 6A. In samplesfrom repressed cells (lanes 11-14), a prominent band(which is, in fact, a doublet a/b: bands 27/28 in Perez-Ortin et al., 1987), was found far downstream of POTI.just overlapping DNase I hypersensitive site 1. Betweenthis site and the promoter of POTI, there can be foundclear chromatin-specific bands (c, d, e, f, i, j) and otherbands that, though present too in naked DNA (a/b. g, h, k,m, n, 0, p, q), may correspond to linkers of positionednucleosomes. As will be shown in the model given in Fig.7, the spacing between putative linker bands is compati-ble with the 160-165 bp repeat length of yeasl nucleo-somes (reviewed by Perez-Ortin etal., 1989). Moreover,some DNA-characteristic bands between bands m/n andn/o were protected in chromatin, reinforcing the hypothe-sis that positioned nucleosomes exist along the region.
Some distinctive features could be observed in going tothe transcriptionally active state (Fig. 6A). The sensitivityof bands f and g to micrococcal nuclease increased(lanes 1-8), and the pattern became more like nakedDNA (see lanes 9-10) in this region. In all the activestates, some new chromatin-characterlstic bandsappeared: n", o' and. less clearly, !'. Some of these find-ings were more obvious when strategy SH was used (Fig.6B). In this instance, the splitting of band m is also ob-vious. The shift of band o to band o' was detectable inexponentially growing low-glucose cells, but it was evenmore pronounced in stationary cells (lanes 15-18), inwhich band o was absent and a new band (o"), whichapparently coincides with a naked DNA band, was seen.Therefore small, but precise, differences between expo-nential and stationary cells are seen in the micrococcalnuclease patterns. However, no oleate-characteristicchanges were detected either by micrococcal nuclease orDNase I digestions.
Mechanisms of yeast POT 1 regulation 1871
DNA stationar;p DNA exponential expunenl ial
747 p-
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Fig. 5. Mapping o( DNase I hypersensitive sites in the POTI locus. The BQS2 strain was grown on YPg or YPG (G - or •, respectively) in the absence orpresence of oleate (O - or +, respectively). Cells were harvested at different stages of growth, as indicaSed, and the sphaeroplasis were digested withincreasing amounts of DNase I, The resulting DNA was digested to compleiion with Sa/I and eiectrophoresed on 1% agarose gel. Southern-blotted, andhybridized to probe SB, Controls ot pRB58 naked DNA digested with DNase I were included (lanes 1 and 6). Fragments obtained by restriction of pRB58were run on lane 11. The sizes and positions of these fragments and olher size markers from p8R322 are indicated in the left margin. The other symbolsare explained in the text. In the right margin of Ihe figure, a scheme of the DNase I cutting sites in POTI gene and its flanking regions is shown. The lengthsof the arrows give an approximate indication of the band intensities. The left pan of the scheme corresponds lo Ihe repressed stale (VPG-tO media) and theright part represents the active state (YPg±O media), at exponential (solid arrows) and stationary phase (broken arrows). The black boxes in the POTI pro-moter correspond to putative TATA and regulatory regions.
Discussion
We have studied the mechanisms that regulate theexpression of the P0T1 gene. The results shown fn Fig. 1indicate that the transcription cf this gene is repressed inglucose-grown cells and markedly induced by a long-chain fatty acid. These results agree with those observedby Einerhand etat. (1991) using different experimentalconditions. The transcriptional regulation observed for thePOTI gene is similar to that previously described forother peroxisomal or peroxisomal related genes, such asPOXI {Dmochowska etai. 1990) and PAS1 {Erdmann efai. 1991) as well as to the regulation cf the proliferation ofthe organelle itself (Veenhuis et ai, 1987). Thus, this con-trol at RNA level may be a general process for genes cod-ing for peroxisomal proteins.
The fact that the effects of snf1 and ssn6 mutations onthe transcription of POTI are similar to those on other
glucose-repressed genes indicates that the regulation ofPOTI is under the control cf a general glucose-regulatorymechanism that involves SNF1 and SSN6 ger\e products.The recent discovery that ADRI, which is involved in theglucose-repression mechanism of the ADH2 gene {Denisand Young. 1983), is also a positive regulator in genesenccding percxisomal proteins {Simon etai. 1991) rein-forces this idea, ADRI activity is regulated by direct phcs-phorytation by cAMP-dependent protein kinase (Cherry efat.. 1989), so that our results, together with those cnADH2 {Denis, 1987), suggest a possible connectionbetween the role of cAMP and the SA/Fy-SSW6pathwaysfor glucose regulation.
In addition to this dual mechanism of regulation, wehave found a third mechanism for transcriptional controlcf P0T1. which depends on the growing state (Table 1and Fig. 2). The expression of thiolase at stationary

1872 J. C. Iguai, C. Gonzalez-Boseh. L. Franco and J. E P^rez-Ortin
slal . exp. UNA exp.
3584 •-
2303
1926 •-
t693
747
1 2 3 4 5 6 7 8 9 10 11 12 13 14
B
exp. UNA exp.
GO
- 3584
1 2 3 4 5 G 7 8 9 10 11 12 13
Fig, 6. Mapping of the micrococcal nuclease cutting sites In the POTI locus.A. Results obtained by Sal\ restfiction and SB probing.B. Results with Stu\ restnction and SH probing. All the conditions and symbols are as m Fig. 5. Bands are marked with letters as described in the text andthe migration of size markers (either from pRB58 or pBR322) is depicted as arrowheads at the outer sides of both panels.
phase is not due to a reversion of catabolite repression,because celis were grown on a medium that prevents thiseffect, and because the time-course of expression ofFBP1. repressed also by glucose, is markedly different(Fig. 2). In spite of the above comments, the possibilitythat changes in stability of mRNA and/or the protein itselfmay be also involved cannot be excluded. On the otherhand, the growth-dependent regulatory mechanismseems not to be related to oleate, because it is observedboth in the presence and absence of the fatty acid. How-ever, at stationary phase the induction by oleate is lowerthan in exponentially growing cells, which may be due toan effect of 'saturation' of gene transcription. The activa-tion of thiolase in stationary phase is not surprisingbecause depletion of nutrients may activate secondarymetabolic pathways to use alternative energy and carbonsources. One of these alternative pathways is fatty aciddegradation via p-oxidation. It has been shown for other
peroxisomal enzymes that their activities increase duringculture (Thieringer et 3l., 1991). It is possible, therefore,that the mechanism of transcriptional control by entry intostationary phase may be universal for peroxisomalenzymes.
The amounts of several yeast proteins increase byentry into stationary phase (Boucherie. 1985). Most oftheir genes are induced by heat shock (Tanaka et al.,1988; Werner-Washburne et al., 1989; Praekelt and Mea-cock, 1990). Nevertheless. POTI is not heat-shock-inducible (J. C, Igual, unpublished results) and its pro-moter does not contain any heat-shock element (HSE),as revealed by analysis of its sequence.
The physiological state of stationary phase cells isqualitatively different from that of cycling cells. The prop-erties of stationary cells are also adopted by the cde25mutant at restrictive temperatures, even in the presenceof nutrients, and aiso by mutation in CYRI gene, which

Mechanisms of yeast POT ̂ regulation 1873
encodes adenylate cyctase (Drebot etai. 1990). Thesemutations decrease cAMP levels and would thus beexpected to decrease the activity of cAMP-dependentprotein kinases. Therefore the physiological changescharacteristic of stationary phase can result from definedmolecular changes in the nutrient signalling pathwaymediated by cAMP (see Broach. 1991). POTI is activatedin stationary cells, as shown in this paper, and this factallows us to postulate that this induction might be relatedto the cAMP regulatory pathway. In view of the fact thatthe product of ADR1 is regulated by cAMP, it may be thatADRI has a role in the activation of transcription relatedto nutrient depletion. Glucose depletion, a situation inwhich ADRI is known to act (Simon et al., 1991). mayrepresent a particular type of nutrient depletion thatresults in entry into the stationary phase. Additional stud-ies of POT1 expression are in progress in order to checkthese hypotheses.
The different nature of the three regulatory mecha-nisms of POTI transcription has been further checked atchromatin level. The chromatin structure in the 5" regionof many yeast genes depends on the transcriptionat state(Perez-Ortin et al., 1989), In the present instance, thechanges during glucose derepression suggest that threepositioned nucleosomes close to the initiation site are dis-placed downstream (see Fig. 7). The DNase I sensitivebands 2 and 3 flank a protected zone of about 35 bp cen-tred at -220, which may be the binding site for regulatoryfactors. Upon gene activation, significant changes in both
the intensity and position of bands 2 and 3 are seen. Thedisplacement of DNase I sensitivity towards the cap siteof POTI gene (located at -19. Einerhand ef al., 1991)coincides with the proposed displacement of nucleo-somes, which may be produced, in turn, by the binding ofspecific activators (ADRI?) to that region. The displace-ment of the first nucleosome would completely uncover aDNA sequence between -153 and -203, with UAS (up-stream activating sequence) activity, responsible for theglucose repression and oieate induction (Einerhand etal.,1991). Further upstream, the nuclease digestion patternsare consistent with the existence of a constitutive nucleo-some-less region. The proposed displacement of nucleo-somes may only occur in some copies of the gene, andour results are also compatible with the existence of twophases of nucleosomes, as suggested by the perma-nence of the bands o-l together with the transcription-specific bands o'-l". Multiple phases of nucleosomal posi-tioning have been previously demonstrated, both in vitro(Dong ef at.. 1990) and in vivo (Ponder and Crawford,1977),
In stationary phase cells, additional changes in nucle-ase sensitivity were observed (see the Resuits). indicat-ing that additional protein factors are added to or substi-tute for those previously bound. Particularly interesting isthe disappearance of DNase I band 3. now replaced bytwo neighboring bands (3' and 3"), which flank a some-what protected zone. This may be a footprint effectcaused by some stationary phase-specific factor(s).
A)YPG±O
a b c d e f g
SUC2
B) YPg ± 0
sues
k l m n o p q r s
C) Stationary
r m' n' 0"
Fig. 7. A model lor the chromatin struciure ofPOTt gene and its flanks.A. Repressing conditions. Lettered, solid anowsrepresent Ihe micrococcal nuclease cutting sitesin chfomalin samples. Their lengths reflect Iheiniensity ol the band detected (see Fig. 6), Arrow-heads mark the micrococcaj nudease cuttingsiles <n naked DNA The positioned nucleosomesare depicted as circles covering about 140 bpDNA Numbered, broken arrows below ihe genepoint to the DNase I hypersensitive siles. Blackboxes stand for TATA (verdcal rectangle) andUAS (horizontal reclangle) elements.B Partially ad've conditions Only Ihe distinctivebands are lettered The asterisk marks a zonewith increased sensitivity to nucleases. Two alter-native phases tor the iirsi three nucleosomesdownstream the UAS are proposed, as describedin the text.
C. Fully active conditions Only the charactensticbands in the promoter region are shown.
2- 3, 3,,

1874 J. C. Igual. C. Gonzalez-Bosch. L. Franco and J. E, P§rez-Onin
Moreover, the changes in the micrococcal nuclease pat-
terns (see bands o" and n', Fig. 6) are suggestive of
changes in the nucieosomal organization of the promoter,
hypothesized in Fig. 7 as the loss of the first nucleosome.
Oleate induction of the gene does not produce any
detectable change in the DNase I and micrococcal nucle-
ase patterns, but subtle changes in the organization of
POTI promoter chromatin would not be detected by the
methodology used. The dominance of glucose repression
upon oleate induction can be explained according to the
chromatin model proposed above assuming that the bind-
ing of the oleate-dependent activating factors to the UAS
is blocked by a nucleosome which is only displaced dur-
ing Ihe derepression process after glucose deprivation.
At the 3" end of the gene, in a region placed between
P0T1 and the ORF of unknown function, interesting
changes in both micrococcal nuclease and DNase I sen-
sitivity occur when POTI is switched on. These changes
include a diffuse increase in DNase I sensitivity and the
disappearance of the micrococcal nuciease cutting sites g
and h. which are replaced by DNA-characteristic bands,
suggesting that two nucleosomes are lost. Lee and Gar-
rard (1991) have proposed that nucleosomes may split in
the positively supercoiled DNA that forms just ahead of
RNA poiymerase when DNA is anchored to the nuclear
matrix. Given that transcription of the open reading frame
is constitutive (our unpublished observations) the coinci-
dence of two opposite waves of positive supercoiling
when POT1 transcription is activated may have a similar
effect and destabilize these nucleosomes (see Thoma,
1991). This hypothesis is now being tested.
Experimental procedures
Yeast strains
The following haploid strains of S. cerevisiae were used inthese experiments; SEY2101 (a, ura3-52. Ieu2-3J12, ade2-101. SUC2A9. poti-M, MCY1389 {a. ura3-52. leu2::HIS3.SUC2. POTI). MCY1484 (a. lys2'B01. ade2'10ioc. snh-13.SUC2. POTI). MCY1371 (a. his4-539am. ade2-101oc. ssn6-1.SUC2, POTI). H222 (a. teu2::HIS3. Iys2-8O1. ura3-52. migi-&2::LEU2. SUC2. POTI). BQS2 (SEY2101 transformed withplasmid pRB58) (P^rez-Ortfn etaL 1986) and BQS20 (a. ura3-52. ieu2::HIS3. SUC2. pot1::URA3} (Igual etal.. 1991).
Growth of yeast
Yeast cells were grown on YP medium ( 1 % yeast extract. 2%bactopeptone) containing 2% glucose (YPG). 0.1% glucose{YPg) or 2% galactose (YPGal). When appropriate, the samemedia were supplemented with 0.1% (v/v) oleic acid (markedas +0). Celis were precultured In the corresponding medium,transferred to the same fresh medium, and harvested at mid-exponential phase or after 10 h in stationary phase.
Enzyme assay
Yeast cells grown on the different media were sphaeroplastedas described (Pere2-Ortin et al.. 1986) except thai zymolyasewas used as the lytic enzyme. The washed sphaeroplasts werelysed in 75 mM Tris-HCi. pH 7,9. 1 mM MgCI^., 0.25 mM EDTAand 5 mM [S-mercaptoethanol. After centrifugation at 5000 x gfor 5 min, supernatanis were saved for enzyme measure-ments.
Thiolase activity was assayed by a modification of themethod of Middleton (1973), by monitoring, at 303 nm, thestimulation of acetoacetyl-CoA breakdown caused by (he addi-tion of CoA, The assay was carried out in 100 mM Tris-HCI. pH8.1, 25 mM MgCI^, 50 mM NaCI. 10 yM acetoacetyl-CoA and50 [iM CoA. The specific activity of thiolase was expressed asnanomoles of acetoacetyl-CoA removed per minute per mil-ligram of protein. Protein concentration was determined by themethod of Bradford (1976). using bovine serum albumin asstandard.
Yeast RNA isolation and anaiysis
RNA from yeast cells was prepared and analysed as describedpreviously (Igual etal.. 1991).
Nuclease digestion and localization of cutting sites
Digestions with DNase I and micrococcal nudeass ar>d local-ization o( the cutting sites were carried out as previouslydescribed (Perez-Ortin etal.. 1986; 1987). Random-primed^^ probes were used.
Acknowledgements
J.C.I is a Fellow ol the Spanish FPI Programme. This workwas supported by grant PB88-352 from DGICYT {Spain) toL.F. We thank Drs F. Estruch and E. Matallana for their criticalreview of the manuscript. Dr H. Tabak for communicating hisresults before publication, and Drs M. Carlson, H. Ronne andS. Emr for the gift of yeast strains.
References
Avers. C.J., and Federman, M, (1968) The occurrence in yeastSaccharomyces cerevisiae of cytoptasmic granules whichresemble microbodies. JCellBiolZ7: 555-559.
Borst, P. (1989) Peroxisome biogenesis revisited. Biochim Bio-phys Acta ^00B•.^-^ 3.
Boucherie, H, (1985) Protein synthesis during transition andstationary phases under glucose limitation in Saccha-romyces cerevisiae. J Bacterioi ^6'\: 385-392.
Bradford. M. (1976) A rapid and sensitive method for the quan-titatlon of microgram amounts of protein utilizing the princi-ple of protein dye binding. Anal Biochem 72: 248-254.
Broach. JR. (1991) flAS genes in Saccharomyces cerevisiae:signal transduction in search of pathway. Trends Genet 7:28-33.
Celenza, J.L., and Carlson. M. (1986) A yeast gene that Isessential for release from glucose repression encodes a pro-tein kinase. Science 233:1175-1180.

MeciianismsofyeastPO^^ reguiation 1875
Cherry, J.R.. Johnson, T.R.. Dollard. C. Shuster, J.R.. andDenis, C.L. (1989) Cyclic AMP-dependent protein kinasephosphorylates and inactivates the yeast transcriptional acti-vator ADR1 Ce//56:409-419.
Cooper. T.G.. and Beevers. H. (1969) p-oxidation enzymes inglyoxysomes from castor bean endosperm. J Biol Chem244:3514-3520.
Denis. C,L. (1987) The effects of ADR1 and CCR1 genedosage on the regulation of the glucose-repressible alcoholdehydrogenase from Sacciiaromyces eerevisiae. Moi GenGene/208: 101-106.
Denis. C.L., and Young. E.T. (1983) Isolation and characteriza-tion of the positive regulatory gene ADR1 from Saeeha-romyees eerevisiae. Mol Cell Biol3:360-370.
Dmochowska, A.. Dignard. D.. Male2ska. R.. and Thomas,D.Y. (1990) Structure and transcriptional control of the Sae-eharomyces eerevisiae P0X1 gene encoding acyi-coen-zyme A oxidase. Gene 88: 247-252.
Dong. F., Hansen. J.C. and van Hoide, K.E. (1990) DNA andprotein determinants of nucleosome positioning on seaurchin 5S ribosomal RNA gene sequences in vitro. Proc NatIAead Sei USA 87: 5724-5728,
Drebot, M.A.. Barnes. C.A., Singer, R.A.. and Johnston. G.C.(1990) Genetic assessment of stationary phase for cells ofthe yeast Saeeharomyees eerevisiae. J Baeteriol 172:3584-3589.
Einerhand. A.W.C, Voorn-Brouwer. T.M., Erdman, K.. Kunau,W.H., and Tabak. H.F. (1991) Regulation of transcription ofthe gene coding for peroxisomal 3 oxoacyl-CoA thiolase ofSaeeharomyees eerevisiae. Eur J Bioehem 200:113-122.
Entian. K.D. (1986) Glucose repression: a complex regulatorysystem in yeast. Miero Sc/3: 366-371.
Erdmann. R.. Wiebel. F.F.. Flessau, A , Rytka, J.. Beyer. A..Frohlich. K.U., and Kunau. W.H. (1991) P-4Sr, a yeast generequired for peroxisome biogenesis, encodes a member of anovel family of putative ATPases. Ce//64:499-510.
Gerhardt, B. (1983) Localization of p-oxidation enzymes in per-oxisomes isolated from nonfatty piant tissues. Pianta 159:238-246.
Igual. J.C. Matallana. E., Gonz^lez-Bosch. C, Franco. L.. andPerez-Ortin. J.E, (1991) A new glucose-repressible geneidentified from the analysis of chromatin structure in deletionmutants of yeast SUC2 locus. Yeast 7: 379-389.
Kionka, C . and Kunau, W.H. (1985) Inducible p-oxidation path-way in Neurospora erassa. J Baeterioi ^6'\: 153-157.
Kunau. W.H.. Buhne. S.. Moreno de la Garza. M., Kionka, C .Mateblowski. M., Schultz-Borchard. U., and Thieringer. R,(1988) Comparative enzymology of [i-oxidation. BioehemSoc Trans 16: 418-420.
Lazarow. P.B , and Fujiki, Y. (1985) Biogenesis of peroxi-somes. Annu Rev Celi Biol 1: 489-530.
Lee, M.S.. and Garrard. W.T, (1991) Transcription-inducednudeosome splitting': an underlying structure for DNase Isensitive chromatin. Ei\4B0 J10:607-615.
McCammon. M.T.. Veenhuis, M.. Trapp. S.B.. and Goodman,J.M. (1990) Association of glyoxylate and beta-oxidationenzymes with peroxisomes of Saeeharomyees eerevisiae. JBaeteriol ^72: 5816-5827.
Middleton, B. (1973) The oxoacyl-CoA thiolases of animal tis-sues. Bioehem J132: 717-730.
Nehlin. J.O.. and Ronne. H. (1990) Yeast MIGi repressor isrelated to the mammalian early response and Witms* tumorfinger proteins. E/WeOJ9:2891-2898.
Nehlin. J.O.. Carlberg. M.. and Ronne. H. (1991) Control of
yeast GAL genes by MIG1 repressor: a transcriptional cas-cade in the glucose response. EMBO J10: 3373-3377.
Osumi T.. and Hashimoto. T. (1984) The inducible fatty acidoxidation system in mammalian peroxisomes. Trends Bio-ehem Sei 9: 3^7-3^9.
Parish, R.W. (1975} The isolation and characterization ol per-oxisomes (microbodias) from baker's yeast Saeeharomyeeseerevisiae. Areh Mierobiefi05:187-192.
Perez-Ortrn. J,E . Estruch, P.. Matallana, E , and Franco L.(1986) DNase I sensitivity ol the chromatin of the yeastSUC2Qene for invertase. Mol Gen Gene/205: 422-427.
P6rez-0rtin, J.E.. Estruch, F., Matallana, E.. and Franco. L,(1987) Fine analysis of the chromatin structure of the yeastSUC2 gene and of its changes upon derepression. Compar-ison between the chromosomal and plasmid-insertedcopies, NueiAeids Res 15:6937-6954.
P6rez-0rtin. J.E,. Matallana. E., and Franco L. (1989) Chro-matin structure of yeast genes. Yeast 5: 219-238.
Ponder. B.A.J.. and Crawford. L.V. (1977) The arrangement ofnucieosomes in nucleoprotein complexes from polyomavirus and SV40. Cell 11: 35-49.
Praekeit, U.M and Meacock, P.A. (1990) HSP12, a new smallheat shock gene of Saeeharomyees eerevisiae: analysis ofstructures, regulation and function. Moi Gen Genet 223:97-106.
Schultz. J.. and Carlson, M. (1987) Molecular analysis olSSN6. a protein functionally related to the SNF1 proteinkinase of Saeeharomyees eerevisiae Mol Ceil Biol 7:3637-3645.
Simon. M.. Adam, G., Rapatz, W.. Spevak. W., and Ruis. H.(1991) The Saeeharomyees eerevisiae ADR 1 gene is a pos-itive regulator of transcription of genes encoding peroxiso-mal proteins. Mol Celi Biol 11: 699-704.
Skoneczny, M.. Chelstowska. A . and Rytka. J. (1988) Study ofthe coinduction by fatty acids of catalase A and acyl-CoAoxidase in standard and mutant Saeeharomyees eerevisiae.Eur J Bioehem 174: 297-302.
Tanaka, K., Matsumolo, K., and Toh-e, A, (1988) Dual regula-tion of the expression of the polyubiquitin gene by cyclicAMP and heat shoch in yeast. EMBOJ7: 495-502.
Tanaka, A.. Osumi, M., and Fukui. S. (1982) Peroxisomes ofalkane-grown yeast: fundamental and practical aspects. AnnNYAeadSei3B6:183-199.
Thieringer, R., Shio, H.. Cohen, G., and Lazarow, P.B. (1991)Peroxisomes in Saeeharomyees eerevisiae. immunofluores-cence analysis and import of catalase A into isolated peroxi-somes. Mo/Ce/yefo/11: 510-522.
Thoma, F. (1991) Structural changes in nucieosomes duringtranscription: strip, split or flip? Trends Genet 7:175-177,
Veenhuis. M.. and Harder. W. (1987) Metabolic significanceand biogenesis of microbodies in yeast. In Peroxisomes inBiology and Medieine. Fajimi, H.D.. and Sies, H. {eds),Berlin: Springer-Verlag. pp. 435-457.
Veenhuis, M.. Mateblowski, M.. Kunau, W.H.. and Harder, W.
(1987) Proliferation of microbodies in Saeeharomyees eere-visiae. YeastZ: 77-84.
Werner-Washburne. M., Becker, J.. Kosic-Smithers, J., andCraig, E.A. (1989) Yeast Hsp70 RNA levels vary in responseto the physiological status of the cell. J Baeterioi 171:2680-2688.
Wu, C (1980) The 5' ends of Drosophiia heat shock genes inchromatin are hypersensitive to DNase I. Nature 286:854-660.





















