
The Mg/Ca-temperature relationship of benthic foraminiferal calcite: New core-top calibrations in...
8
The Mg/Ca-temperature relationship of benthic foraminiferal calcite: New core-top calibrations in the b 4 °C temperature range Stephanie L. Healey a, ⁎, Robert C. Thunell a , Bruce H. Corliss b a Department of Geological Sciences, University of South Carolina, Columbia, South Carolina 29208, USA b Department of Earth and Ocean Sciences, Nicholas School of the Environment and Earth Sciences, Duke University, Durham, North Carolina 27706, USA ABSTRACT ARTICLE INFO Article history: Received 21 April 2008 Accepted 15 May 2008 Available online 7 July 2008 Editor: M.L. Delaney Keywords: paleoceanography Mg/Ca paleothermometry benthic foraminifera carbonate ion Calcite magnesium/calcium ratios were determined for four species of benthic foraminifera (Cibicidoides wuellerstorfi, Cibicidoides mundulus, Oridorsalis umbonatus and Pyrgo murrhina) from core-top samples collected from the Atlantic, Indian, and Pacific Oceans where modern bottom water temperatures are less than 4 °C. Mg/Ca of all four species increase with increasing temperature, and the Mg/Ca:temperature relationship does not appear to be necessarily exponential. Mg/Ca of C. mundulus is consistently higher than C. wuellerstorfi, implying differences in the uptake of Mg by different species within this genus. The observed Mg/Ca for C. wuellerstorfi at temperatures less than 3 °C is lower than those predicted from published Mg/Ca: temperature relationships derived for Cibicidoides spp. at higher temperatures. Previous studies have attributed this change in the Mg/Ca-temperature relationship of C. wuellerstorfi to the effect of decreased carbonate ion saturation (Δ[CO 3 2- ]) on Mg/Ca at low temperatures. A relationship between the Δ[CO 3 2- ] and Mg/Ca of 0.0083 ± 0.002 (mmol/mol)/(μmol/kg) has been established for C. wuellerstorfi from this study. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Ocean circulation and temperature, along with continental ice volume are important elements of the global climate system and reconstructing past changes in these parameters are basic objectives of paleoclimatology. The change in δ 18 O composition of seawater due to the melting of continental ice sheets since the last glacial maximum (LGM) has been estimated using fossil coral reef terraces (Fairbanks, 1989) and sediment pore-fluid chemistry (Adkins and Schrag, 2001; Schrag et al., 2002, 1996). Although δ 18 O changes in benthic foraminiferal calcite in deep-sea cores have been used as a first- order approximation of Late Pleistocene glacial–interglacial sea-level changes (e.g. Shackleton, 1967), most of these records also contain glacial–interglacial variability of a higher amplitude than what is expected based on independent sea-level estimates (e.g. Labeyrie et al., 1987; Martin et al., 2002; Skinner et al., 2003), suggesting that bottom water temperatures also vary significantly on these timescales. For example, it has been estimated that bottom waters were about 1.5–3 °C colder in the Pacific (Chappell and Shackleton, 1986; Shackleton, 1987; Martin et al., 2002) and 3–4 °C colder in the Atlantic (Schrag et al., 2002; Adkins and Schrag, 2001; Martin et al., 2002) during glacial times than today. The oxygen isotope composition (δ 18 O) of benthic foraminiferal calcite is a function of the temperature and the δ 18 O composition of sea water at the time of calcification. Likewise, the δ 18 O composition of sea water is a function of global ice volume and local variations in salinity. Thus, benthic foraminiferal δ 18 O measurements alone cannot provide quantitative estimates of past changes in any of these parameters. However, with the development of Mg/Ca paleothermo- metry, it is now possible to use benthic foraminiferal Mg/Ca to directly estimate bottom water temperatures (Rosenthal et al., 1997; Rathburn and De Deckker, 1997; Martin et al., 2002; Lear et al., 2002). Thus, paired Mg/Ca and δ 18 O measurements of benthic foraminifera offer the potential to identify how much of the total δ 18 O change of benthic foraminiferal calcite is due to changes in the isotopic composition of seawater. The majority of paleoclimate records utilizing benthic foraminiferal δ 18 O and/or Mg/Ca are based on sediment cores retrieved from water depths greater than 2000m. At present, over 95% of the global ocean deeper than 2000m has bottom water temperatures that are b 5 °C (Fig. 1), and it is reasonable to assume that in most regions temperatures below this depth were colder during glacial periods. Thus, it is important that the relationship between temperature and the Mg/Ca of benthic foraminiferal calcite be well constrained at the low end of the temperature spectrum. Existing benthic foraminiferal Mg/Ca-tempera- ture calibrations are based on data sets spanning bottom water temperatures of 0–18 °C and combine different species of benthic foraminifera, including species that live primarily in waters warmer than Earth and Planetary Science Letters 272 (2008) 523–530 ⁎ Corresponding author. E-mail address: [email protected] (S.L. Healey). 0012-821X/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.epsl.2008.05.023 Contents lists available at ScienceDirect Earth and Planetary Science Letters journal homepage: www.elsevier.com/locate/epsl
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The Mg/Ca-temperature relationship of benthic foraminiferal calcite: New core-top calibrations in...

Earth and Planetary Science Letters 272 (2008) 523–530
Contents lists available at ScienceDirect
Earth and Planetary Science Letters
j ourna l homepage: www.e lsev ie r.com/ locate /eps l
The Mg/Ca-temperature relationship of benthic foraminiferal calcite: New core-topcalibrations in the b4 °C temperature range
Stephanie L. Healey a,⁎, Robert C. Thunell a, Bruce H. Corliss b
a Department of Geological Sciences, University of South Carolina, Columbia, South Carolina 29208, USAb Department of Earth and Ocean Sciences, Nicholas School of the Environment and Earth Sciences, Duke University, Durham, North Carolina 27706, USA
⁎ Corresponding author.E-mail address: [email protected] (S.L. Healey).
0012-821X/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.epsl.2008.05.023
A B S T R A C T
A R T I C L E I N F OArticle history:
Calcite magnesium/calcium Received 21 April 2008Accepted 15 May 2008Available online 7 July 2008Editor: M.L. Delaney
Keywords:paleoceanographyMg/Ca paleothermometrybenthic foraminiferacarbonate ion
ratios were determined for four species of benthic foraminifera (Cibicidoideswuellerstorfi, Cibicidoides mundulus, Oridorsalis umbonatus and Pyrgo murrhina) from core-top samplescollected from the Atlantic, Indian, and Pacific Oceans where modern bottom water temperatures are lessthan 4 °C. Mg/Ca of all four species increase with increasing temperature, and the Mg/Ca:temperaturerelationship does not appear to be necessarily exponential. Mg/Ca of C. mundulus is consistently higherthan C. wuellerstorfi, implying differences in the uptake of Mg by different species within this genus. Theobserved Mg/Ca for C. wuellerstorfi at temperatures less than 3 °C is lower than those predicted frompublished Mg/Ca: temperature relationships derived for Cibicidoides spp. at higher temperatures. Previousstudies have attributed this change in the Mg/Ca-temperature relationship of C. wuellerstorfi to the effectof decreased carbonate ion saturation (Δ[CO3
2−]) on Mg/Ca at low temperatures. A relationship betweenthe Δ[CO3
2−] and Mg/Ca of 0.0083 ± 0.002 (mmol/mol)/(μmol/kg) has been established for C. wuellerstorfifrom this study.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Ocean circulation and temperature, along with continental icevolume are important elements of the global climate system andreconstructing past changes in these parameters are basic objectivesof paleoclimatology. The change in δ18O composition of seawater dueto themelting of continental ice sheets since the last glacial maximum(LGM) has been estimated using fossil coral reef terraces (Fairbanks,1989) and sediment pore-fluid chemistry (Adkins and Schrag, 2001;Schrag et al., 2002, 1996). Although δ18O changes in benthicforaminiferal calcite in deep-sea cores have been used as a first-order approximation of Late Pleistocene glacial–interglacial sea-levelchanges (e.g. Shackleton, 1967), most of these records also containglacial–interglacial variability of a higher amplitude than what isexpected based on independent sea-level estimates (e.g. Labeyrieet al., 1987; Martin et al., 2002; Skinner et al., 2003), suggesting thatbottomwater temperatures also vary significantly on these timescales.For example, it has been estimated that bottom waters were about1.5–3 °C colder in the Pacific (Chappell and Shackleton, 1986;Shackleton, 1987; Martin et al., 2002) and 3–4 °C colder in theAtlantic (Schrag et al., 2002; Adkins and Schrag, 2001; Martin et al.,2002) during glacial times than today.
l rights reserved.
The oxygen isotope composition (δ18O) of benthic foraminiferalcalcite is a function of the temperature and the δ18O composition ofsea water at the time of calcification. Likewise, the δ18O compositionof sea water is a function of global ice volume and local variations insalinity. Thus, benthic foraminiferal δ18O measurements alone cannotprovide quantitative estimates of past changes in any of theseparameters. However, with the development of Mg/Ca paleothermo-metry, it is now possible to use benthic foraminiferal Mg/Ca to directlyestimate bottomwater temperatures (Rosenthal et al., 1997; Rathburnand De Deckker, 1997; Martin et al., 2002; Lear et al., 2002). Thus,paired Mg/Ca and δ18O measurements of benthic foraminifera offerthe potential to identify howmuch of the total δ18O change of benthicforaminiferal calcite is due to changes in the isotopic composition ofseawater.
The majority of paleoclimate records utilizing benthic foraminiferalδ18O and/or Mg/Ca are based on sediment cores retrieved from waterdepths greater than 2000m. At present, over 95% of the global oceandeeper than 2000m has bottom water temperatures that are b5 °C(Fig.1), and it is reasonable to assume that inmost regions temperaturesbelow this depthwere colderduring glacial periods. Thus, it is importantthat the relationship between temperature and the Mg/Ca of benthicforaminiferal calcite be well constrained at the low end of thetemperature spectrum. Existing benthic foraminiferal Mg/Ca-tempera-ture calibrations are based on data sets spanning bottom watertemperatures of 0–18 °C and combine different species of benthicforaminifera, including species that liveprimarily inwaterswarmer than
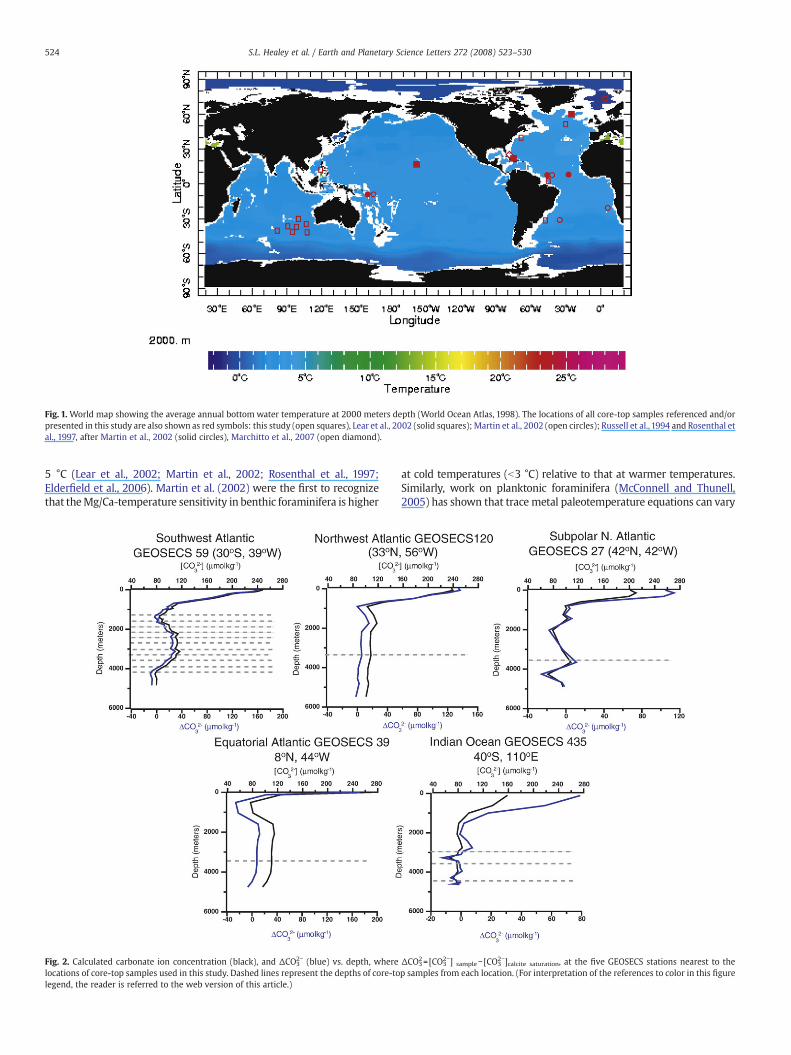
Fig. 1. World map showing the average annual bottom water temperature at 2000 meters depth (World Ocean Atlas, 1998). The locations of all core-top samples referenced and/orpresented in this study are also shown as red symbols: this study (open squares), Lear et al., 2002 (solid squares); Martin et al., 2002 (open circles); Russell et al., 1994 and Rosenthal etal., 1997, after Martin et al., 2002 (solid circles), Marchitto et al., 2007 (open diamond).
524 S.L. Healey et al. / Earth and Planetary Science Letters 272 (2008) 523–530
5 °C (Lear et al., 2002; Martin et al., 2002; Rosenthal et al., 1997;Elderfield et al., 2006). Martin et al. (2002) were the first to recognizethat theMg/Ca-temperature sensitivity in benthic foraminifera is higher
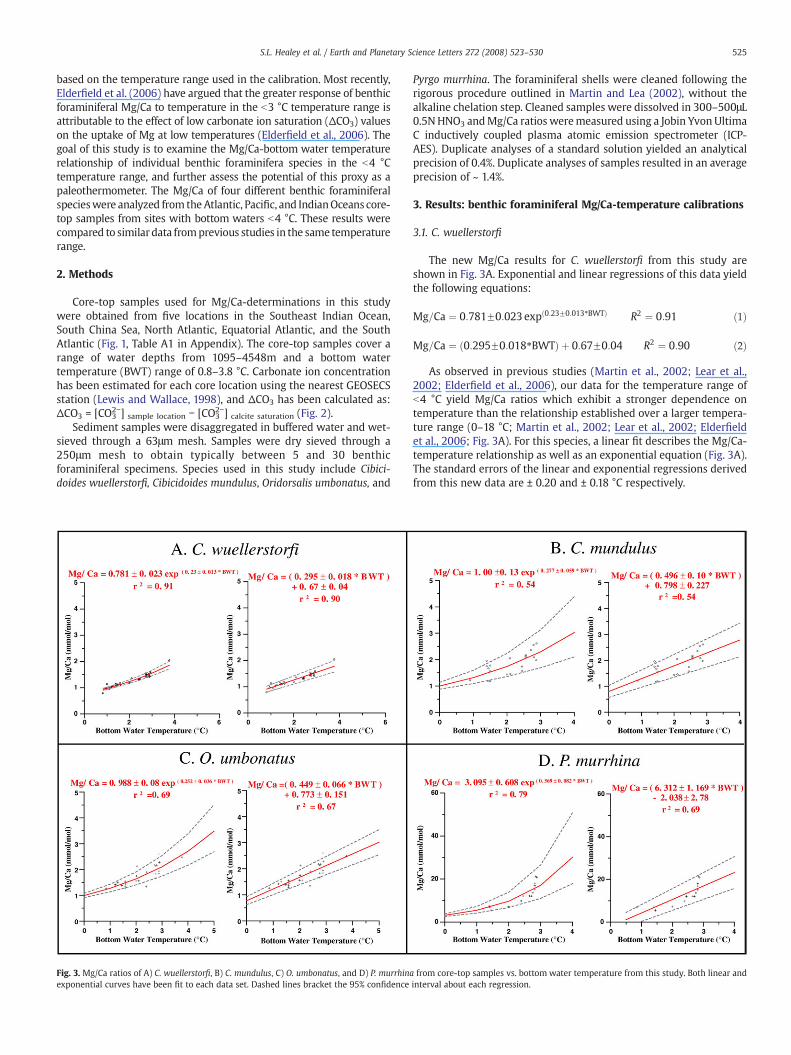
Fig. 2. Calculated carbonate ion concentration (black), and ΔCO32− (blue) vs. depth, where
locations of core-top samples used in this study. Dashed lines represent the depths of core-tolegend, the reader is referred to the web version of this article.)
at cold temperatures (b3 °C) relative to that at warmer temperatures.Similarly, work on planktonic foraminifera (McConnell and Thunell,2005) has shown that trace metal paleotemperature equations can vary
ΔCO32=[CO3
2−] sample− [CO32−]calcite saturation, at the five GEOSECS stations nearest to the
p samples from each location. (For interpretation of the references to color in this figure
525S.L. Healey et al. / Earth and Planetary Science Letters 272 (2008) 523–530
based on the temperature range used in the calibration. Most recently,Elderfield et al. (2006) have argued that the greater response of benthicforaminiferal Mg/Ca to temperature in the b3 °C temperature range isattributable to the effect of low carbonate ion saturation (ΔCO3) valueson the uptake of Mg at low temperatures (Elderfield et al., 2006). Thegoal of this study is to examine the Mg/Ca-bottom water temperaturerelationship of individual benthic foraminifera species in the b4 °Ctemperature range, and further assess the potential of this proxy as apaleothermometer. The Mg/Ca of four different benthic foraminiferalspecieswere analyzed from theAtlantic, Pacific, and IndianOceans core-top samples from sites with bottom waters b4 °C. These results werecompared to similardata fromprevious studies in the same temperaturerange.
2. Methods
Core-top samples used for Mg/Ca-determinations in this studywere obtained from five locations in the Southeast Indian Ocean,South China Sea, North Atlantic, Equatorial Atlantic, and the SouthAtlantic (Fig. 1, Table A1 in Appendix). The core-top samples cover arange of water depths from 1095–4548m and a bottom watertemperature (BWT) range of 0.8–3.8 °C. Carbonate ion concentrationhas been estimated for each core location using the nearest GEOSECSstation (Lewis and Wallace, 1998), and ΔCO3 has been calculated as:ΔCO3 = [CO3
2−] sample location − [CO32−] calcite saturation (Fig. 2).
Sediment samples were disaggregated in buffered water and wet-sieved through a 63μm mesh. Samples were dry sieved through a250μm mesh to obtain typically between 5 and 30 benthicforaminiferal specimens. Species used in this study include Cibici-doides wuellerstorfi, Cibicidoides mundulus, Oridorsalis umbonatus, and
Fig. 3. Mg/Ca ratios of A) C. wuellerstorfi, B) C. mundulus, C) O. umbonatus, and D) P. murrhinaexponential curves have been fit to each data set. Dashed lines bracket the 95% confidence
Pyrgo murrhina. The foraminiferal shells were cleaned following therigorous procedure outlined in Martin and Lea (2002), without thealkaline chelation step. Cleaned samples were dissolved in 300–500μL0.5NHNO3 andMg/Ca ratios weremeasured using a Jobin YvonUltimaC inductively coupled plasma atomic emission spectrometer (ICP-AES). Duplicate analyses of a standard solution yielded an analyticalprecision of 0.4%. Duplicate analyses of samples resulted in an averageprecision of ~ 1.4%.
3. Results: benthic foraminiferal Mg/Ca-temperature calibrations
3.1. C. wuellerstorfi
The new Mg/Ca results for C. wuellerstorfi from this study areshown in Fig. 3A. Exponential and linear regressions of this data yieldthe following equations:
Mg=Ca ¼ 0:781F0:023exp 0:23F0:0134BWTð Þ R2 ¼ 0:91 ð1Þ
Mg=Ca ¼ 0:295F0:0184BWTð Þ þ 0:67F0:04 R2 ¼ 0:90 ð2Þ
As observed in previous studies (Martin et al., 2002; Lear et al.,2002; Elderfield et al., 2006), our data for the temperature range ofb4 °C yield Mg/Ca ratios which exhibit a stronger dependence ontemperature than the relationship established over a larger tempera-ture range (0–18 °C; Martin et al., 2002; Lear et al., 2002; Elderfieldet al., 2006; Fig. 3A). For this species, a linear fit describes the Mg/Ca-temperature relationship as well as an exponential equation (Fig. 3A).The standard errors of the linear and exponential regressions derivedfrom this new data are ± 0.20 and ± 0.18 °C respectively.
from core-top samples vs. bottom water temperature from this study. Both linear andinterval about each regression.
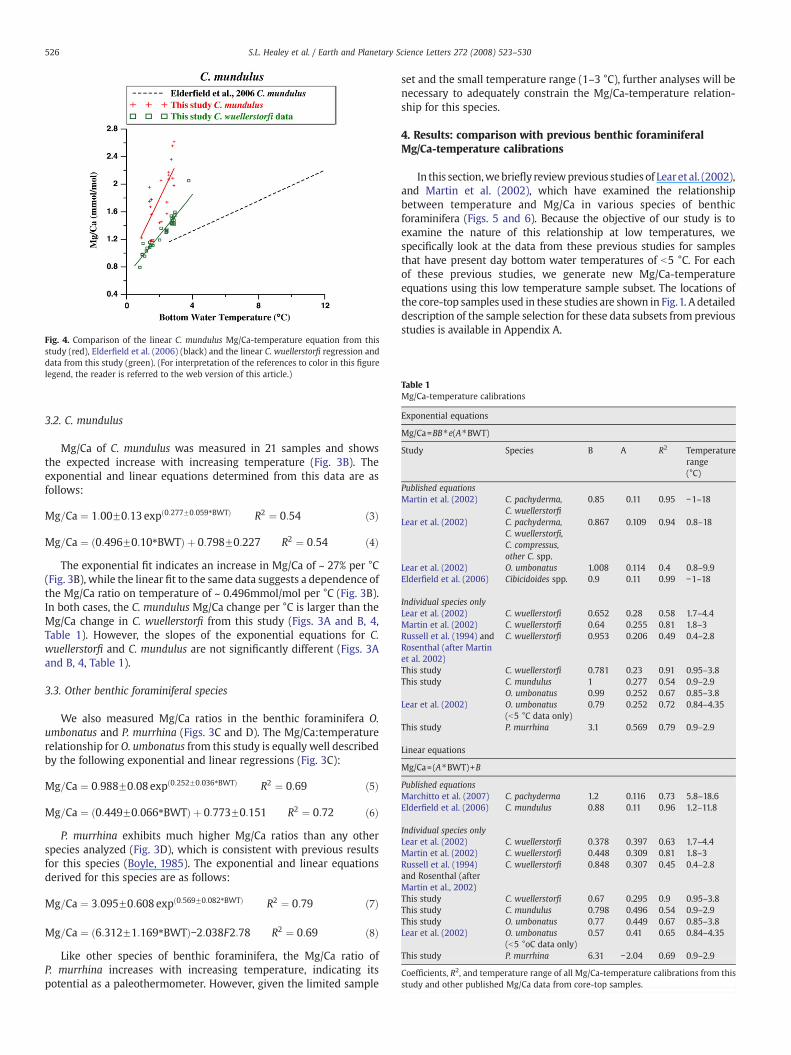
Fig. 4. Comparison of the linear C. mundulus Mg/Ca-temperature equation from thisstudy (red), Elderfield et al. (2006) (black) and the linear C. wuellerstorfi regression anddata from this study (green). (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of this article.)
Table 1Mg/Ca-temperature calibrations
Exponential equations
Mg/Ca=BB⁎e(A⁎BWT)
Study Species B A R2 Temperaturerange(°C)
Published equationsMartin et al. (2002) C. pachyderma,
C. wuellerstorfi0.85 0.11 0.95 −1–18
Lear et al. (2002) C. pachyderma,C. wuellerstorfi,C. compressus,other C. spp.
0.867 0.109 0.94 0.8–18
Lear et al. (2002) O. umbonatus 1.008 0.114 0.4 0.8–9.9Elderfield et al. (2006) Cibicidoides spp. 0.9 0.11 0.99 −1–18
Individual species onlyLear et al. (2002) C. wuellerstorfi 0.652 0.28 0.58 1.7–4.4Martin et al. (2002) C. wuellerstorfi 0.64 0.255 0.81 1.8–3Russell et al. (1994) andRosenthal (after Martinet al. 2002)
C. wuellerstorfi 0.953 0.206 0.49 0.4–2.8
This study C. wuellerstorfi 0.781 0.23 0.91 0.95–3.8This study C. mundulus 1 0.277 0.54 0.9–2.9
O. umbonatus 0.99 0.252 0.67 0.85–3.8Lear et al. (2002) O. umbonatus
(b5 °C data only)0.79 0.252 0.72 0.84–4.35
This study P. murrhina 3.1 0.569 0.79 0.9–2.9
Linear equations
Mg/Ca=(A⁎BWT)+B
Published equationsMarchitto et al. (2007) C. pachyderma 1.2 0.116 0.73 5.8–18.6Elderfield et al. (2006) C. mundulus 0.88 0.11 0.96 1.2–11.8
Individual species onlyLear et al. (2002) C. wuellerstorfi 0.378 0.397 0.63 1.7–4.4Martin et al. (2002) C. wuellerstorfi 0.448 0.309 0.81 1.8–3Russell et al. (1994)and Rosenthal (afterMartin et al., 2002)
C. wuellerstorfi 0.848 0.307 0.45 0.4–2.8
This study C. wuellerstorfi 0.67 0.295 0.9 0.95–3.8This study C. mundulus 0.798 0.496 0.54 0.9–2.9This study O. umbonatus 0.77 0.449 0.67 0.85–3.8Lear et al. (2002) O. umbonatus
(b5 °oC data only)0.57 0.41 0.65 0.84–4.35
This study P. murrhina 6.31 −2.04 0.69 0.9–2.9
Coefficients, R2, and temperature range of all Mg/Ca-temperature calibrations from thisstudy and other published Mg/Ca data from core-top samples.
526 S.L. Healey et al. / Earth and Planetary Science Letters 272 (2008) 523–530
3.2. C. mundulus
Mg/Ca of C. mundulus was measured in 21 samples and showsthe expected increase with increasing temperature (Fig. 3B). Theexponential and linear equations determined from this data are asfollows:
Mg=Ca ¼ 1:00F0:13 exp 0:277F0:0594BWTð Þ R2 ¼ 0:54 ð3Þ
Mg=Ca ¼ 0:496F0:104BWTð Þ þ 0:798F0:227 R2 ¼ 0:54 ð4Þ
The exponential fit indicates an increase in Mg/Ca of ~ 27% per °C(Fig. 3B), while the linear fit to the same data suggests a dependence ofthe Mg/Ca ratio on temperature of ~ 0.496mmol/mol per °C (Fig. 3B).In both cases, the C. mundulus Mg/Ca change per °C is larger than theMg/Ca change in C. wuellerstorfi from this study (Figs. 3A and B, 4,Table 1). However, the slopes of the exponential equations for C.wuellerstorfi and C. mundulus are not significantly different (Figs. 3Aand B, 4, Table 1).
3.3. Other benthic foraminiferal species
We also measured Mg/Ca ratios in the benthic foraminifera O.umbonatus and P. murrhina (Figs. 3C and D). The Mg/Ca:temperaturerelationship for O. umbonatus from this study is equally well describedby the following exponential and linear regressions (Fig. 3C):
Mg=Ca ¼ 0:988F0:08 exp 0:252F0:0364BWTð Þ R2 ¼ 0:69 ð5Þ
Mg=Ca ¼ 0:449F0:0664BWTð Þ þ 0:773F0:151 R2 ¼ 0:72 ð6Þ
P. murrhina exhibits much higher Mg/Ca ratios than any otherspecies analyzed (Fig. 3D), which is consistent with previous resultsfor this species (Boyle, 1985). The exponential and linear equationsderived for this species are as follows:
Mg=Ca ¼ 3:095F0:608exp 0:569F0:0824BWTð Þ R2 ¼ 0:79 ð7Þ
Mg=Ca ¼ 6:312F1:1694BWTð Þ−2:038F2:78 R2 ¼ 0:69 ð8Þ
Like other species of benthic foraminifera, the Mg/Ca ratio ofP. murrhina increases with increasing temperature, indicating itspotential as a paleothermometer. However, given the limited sample
set and the small temperature range (1–3 °C), further analyses will benecessary to adequately constrain the Mg/Ca-temperature relation-ship for this species.
4. Results: comparison with previous benthic foraminiferalMg/Ca-temperature calibrations
In this section,webriefly reviewprevious studies of Learet al. (2002),and Martin et al. (2002), which have examined the relationshipbetween temperature and Mg/Ca in various species of benthicforaminifera (Figs. 5 and 6). Because the objective of our study is toexamine the nature of this relationship at low temperatures, wespecifically look at the data from these previous studies for samplesthat have present day bottom water temperatures of b5 °C. For eachof these previous studies, we generate new Mg/Ca-temperatureequations using this low temperature sample subset. The locations ofthe core-top samples used in these studies are shown in Fig.1. A detaileddescription of the sample selection for these data subsets from previousstudies is available in Appendix A.
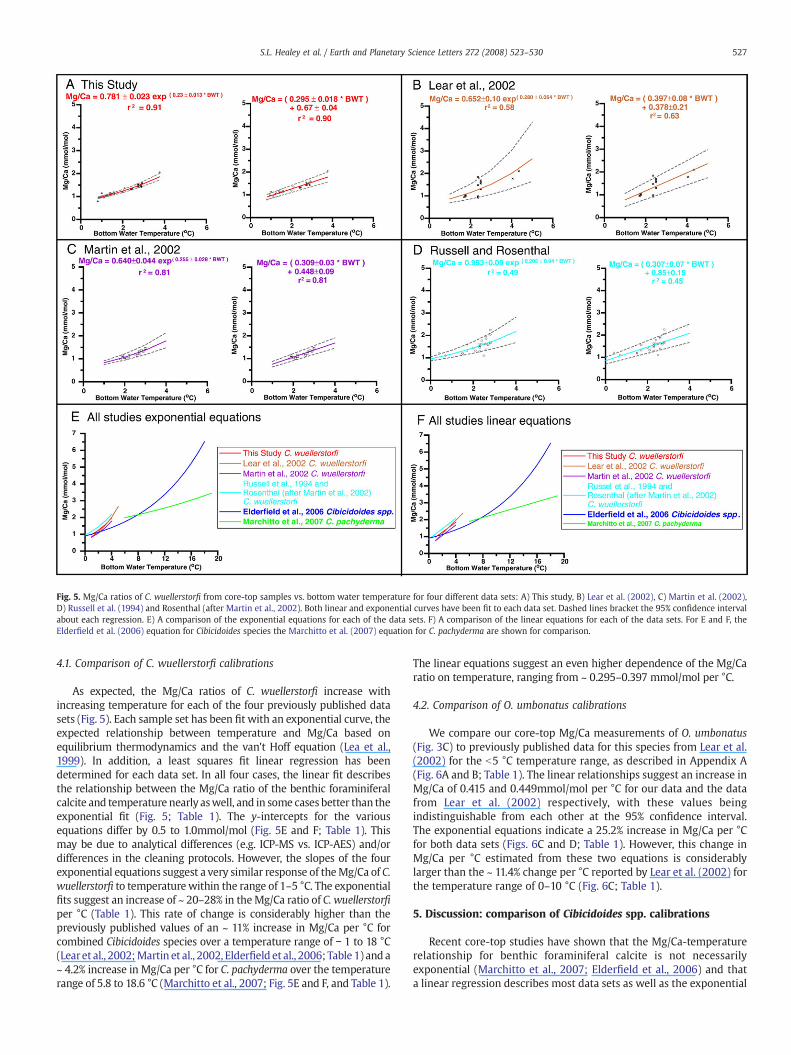
Fig. 5. Mg/Ca ratios of C. wuellerstorfi from core-top samples vs. bottom water temperature for four different data sets: A) This study, B) Lear et al. (2002), C) Martin et al. (2002),D) Russell et al. (1994) and Rosenthal (after Martin et al., 2002). Both linear and exponential curves have been fit to each data set. Dashed lines bracket the 95% confidence intervalabout each regression. E) A comparison of the exponential equations for each of the data sets. F) A comparison of the linear equations for each of the data sets. For E and F, theElderfield et al. (2006) equation for Cibicidoides species the Marchitto et al. (2007) equation for C. pachyderma are shown for comparison.
527S.L. Healey et al. / Earth and Planetary Science Letters 272 (2008) 523–530
4.1. Comparison of C. wuellerstorfi calibrations
As expected, the Mg/Ca ratios of C. wuellerstorfi increase withincreasing temperature for each of the four previously published datasets (Fig. 5). Each sample set has been fit with an exponential curve, theexpected relationship between temperature and Mg/Ca based onequilibrium thermodynamics and the van't Hoff equation (Lea et al.,1999). In addition, a least squares fit linear regression has beendetermined for each data set. In all four cases, the linear fit describesthe relationship between the Mg/Ca ratio of the benthic foraminiferalcalcite and temperature nearly aswell, and in some cases better than theexponential fit (Fig. 5; Table 1). The y-intercepts for the variousequations differ by 0.5 to 1.0mmol/mol (Fig. 5E and F; Table 1). Thismay be due to analytical differences (e.g. ICP-MS vs. ICP-AES) and/ordifferences in the cleaning protocols. However, the slopes of the fourexponential equations suggest a very similar response of theMg/Ca of C.wuellerstorfi to temperaturewithin the range of 1–5 °C. The exponentialfits suggest an increase of ~ 20–28% in theMg/Ca ratio of C. wuellerstorfiper °C (Table 1). This rate of change is considerably higher than thepreviously published values of an ~ 11% increase in Mg/Ca per °C forcombined Cibicidoides species over a temperature range of − 1 to 18 °C(Learet al., 2002;Martin et al., 2002, Elderfield et al., 2006; Table 1) and a~ 4.2% increase in Mg/Ca per °C for C. pachyderma over the temperaturerange of 5.8 to 18.6 °C (Marchitto et al., 2007; Fig. 5E and F, and Table 1).
The linear equations suggest an even higher dependence of the Mg/Caratio on temperature, ranging from ~ 0.295–0.397 mmol/mol per °C.
4.2. Comparison of O. umbonatus calibrations
We compare our core-top Mg/Ca measurements of O. umbonatus(Fig. 3C) to previously published data for this species from Lear et al.(2002) for the b5 °C temperature range, as described in Appendix A(Fig. 6A and B; Table 1). The linear relationships suggest an increase inMg/Ca of 0.415 and 0.449mmol/mol per °C for our data and the datafrom Lear et al. (2002) respectively, with these values beingindistinguishable from each other at the 95% confidence interval.The exponential equations indicate a 25.2% increase in Mg/Ca per °Cfor both data sets (Figs. 6C and D; Table 1). However, this change inMg/Ca per °C estimated from these two equations is considerablylarger than the ~ 11.4% change per °C reported by Lear et al. (2002) forthe temperature range of 0–10 °C (Fig. 6C; Table 1).
5. Discussion: comparison of Cibicidoides spp. calibrations
Recent core-top studies have shown that the Mg/Ca-temperaturerelationship for benthic foraminiferal calcite is not necessarilyexponential (Marchitto et al., 2007; Elderfield et al., 2006) and thata linear regression describes most data sets as well as the exponential
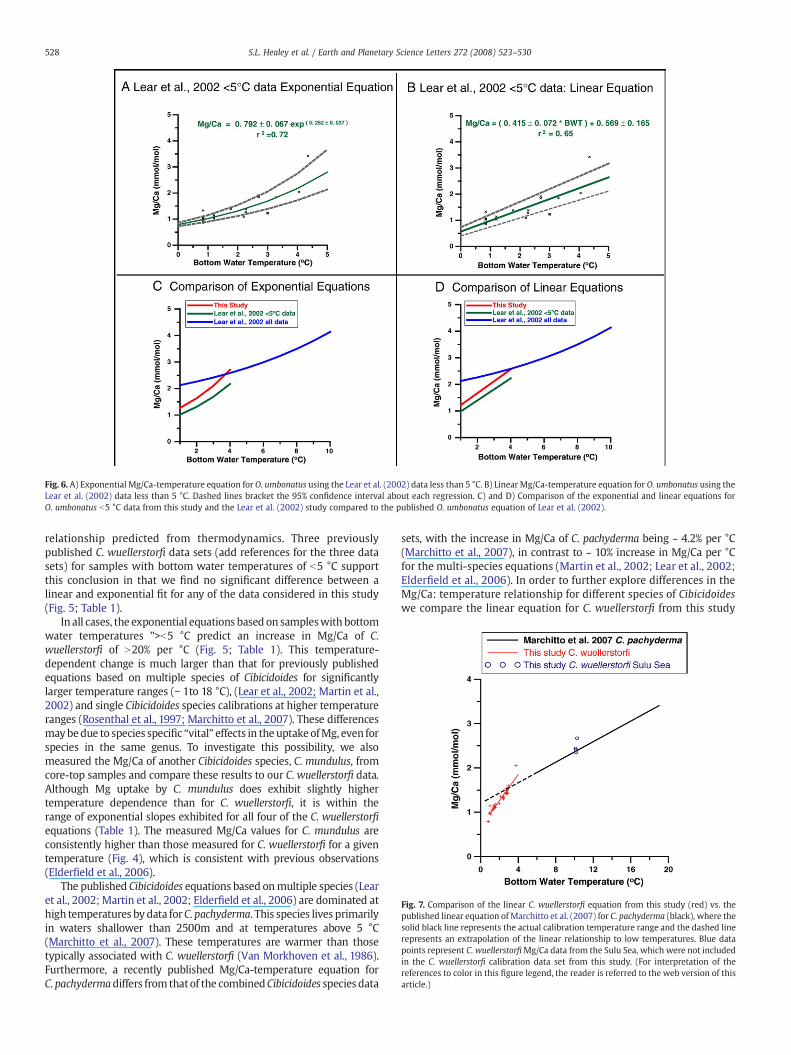
Fig. 6. A) Exponential Mg/Ca-temperature equation for O. umbonatus using the Lear et al. (2002) data less than 5 °C. B) Linear Mg/Ca-temperature equation for O. umbonatus using theLear et al. (2002) data less than 5 °C. Dashed lines bracket the 95% confidence interval about each regression. C) and D) Comparison of the exponential and linear equations forO. umbonatus b5 °C data from this study and the Lear et al. (2002) study compared to the published O. umbonatus equation of Lear et al. (2002).
Fig. 7. Comparison of the linear C. wuellerstorfi equation from this study (red) vs. thepublished linear equation of Marchitto et al. (2007) for C. pachyderma (black), where thesolid black line represents the actual calibration temperature range and the dashed linerepresents an extrapolation of the linear relationship to low temperatures. Blue datapoints represent C. wuellerstorfiMg/Ca data from the Sulu Sea, which were not includedin the C. wuellerstorfi calibration data set from this study. (For interpretation of thereferences to color in this figure legend, the reader is referred to the web version of thisarticle.)
528 S.L. Healey et al. / Earth and Planetary Science Letters 272 (2008) 523–530
relationship predicted from thermodynamics. Three previouslypublished C. wuellerstorfi data sets (add references for the three datasets) for samples with bottom water temperatures of b5 °C supportthis conclusion in that we find no significant difference between alinear and exponential fit for any of the data considered in this study(Fig. 5; Table 1).
In all cases, the exponential equations based on sampleswith bottomwater temperatures ">b5 °C predict an increase in Mg/Ca of C.wuellerstorfi of N20% per °C (Fig. 5; Table 1). This temperature-dependent change is much larger than that for previously publishedequations based on multiple species of Cibicidoides for significantlylarger temperature ranges (− 1to 18 °C), (Lear et al., 2002; Martin et al.,2002) and single Cibicidoides species calibrations at higher temperatureranges (Rosenthal et al., 1997; Marchitto et al., 2007). These differencesmaybedue to species specific “vital” effects in the uptakeofMg, even forspecies in the same genus. To investigate this possibility, we alsomeasured the Mg/Ca of another Cibicidoides species, C. mundulus, fromcore-top samples and compare these results to our C. wuellerstorfi data.Although Mg uptake by C. mundulus does exhibit slightly highertemperature dependence than for C. wuellerstorfi, it is within therange of exponential slopes exhibited for all four of the C. wuellerstorfiequations (Table 1). The measured Mg/Ca values for C. mundulus areconsistently higher than those measured for C. wuellerstorfi for a giventemperature (Fig. 4), which is consistent with previous observations(Elderfield et al., 2006).
The published Cibicidoides equations based onmultiple species (Learet al., 2002; Martin et al., 2002; Elderfield et al., 2006) are dominated athigh temperatures by data forC. pachyderma. This species lives primarilyin waters shallower than 2500m and at temperatures above 5 °C(Marchitto et al., 2007). These temperatures are warmer than thosetypically associated with C. wuellerstorfi (Van Morkhoven et al., 1986).Furthermore, a recently published Mg/Ca-temperature equation forC. pachydermadiffers from thatof the combinedCibicidoides species data
sets, with the increase in Mg/Ca of C. pachyderma being ~ 4.2% per °C(Marchitto et al., 2007), in contrast to ~ 10% increase in Mg/Ca per °Cfor the multi-species equations (Martin et al., 2002; Lear et al., 2002;Elderfield et al., 2006). In order to further explore differences in theMg/Ca: temperature relationship for different species of Cibicidoideswe compare the linear equation for C. wuellerstorfi from this study

529S.L. Healey et al. / Earth and Planetary Science Letters 272 (2008) 523–530
with the linear C. pachyderma equation based on samples fromthe Florida Straights (Marchitto et al., 2007; Fig. 7). The equationsfor these two species are significantly different, with C. wuellerstorfiexhibiting a much greater change in Mg/Ca per °C thanC. pachyderma. However, the C. wuellerstorfi data points above 3 °Ccould just as reasonably be described by the C. pachyderma equationas the b4 °C equation for C. wuellerstorfi (Fig. 7). To examine thisissue further, we measured Mg/Ca on C. wuellerstorfi from five core-top samples from the Sulu Sea with bottom water temperatures of10.5–11 °C (Fig. 7). These samples are unusual in that C. wuellerstorfirarely inhabit waters above 5 °C. The Mg/Ca values for these fivesamples (2.25–2.75mmol/mol) fall close to or on the line describingthe C. pachyderma Mg/Ca-temperature relationship and do notexhibit the higher Mg/Ca values that would be predicted (3.77–3.92mmol/mol) by our low temperature C. wuellerstorfi equation.This suggests that the different C. wuellerstorfi and C. pachydermaMg/Ca-temperature relationships are not species specific, but dueto some other factor. This does not imply that the two speciesexhibit identical responses to temperature, but rather that thelarge difference observed in the two Mg/Ca-temperature relation-ships cannot be attributed simply to interspecies differences in Mguptake.
It has been suggested that the strong decrease in Mg/Ca inC. wuellerstorfi at low temperatures is the result of an enhanced changein carbonate ion concentration per °C in the low temperature range(Martin et al., 2002; Lear et al., 2002; Elderfield et al., 2006; Marchittoet al., 2007; Rosenthal et al., 2006). The decrease in ΔCO3
2− per °Csteepens tenfold at temperatures below 3 °C and it is at ~ 3 °C wherethere is an apparent change in the C. wuellerstorfi Mg/Ca-temperaturerelationship (Elderfield et al., 2006). Elderfield et al. (2006) used twoapproaches to isolate the ΔCO3
2− effect on Mg/Ca in the b3 °Ctemperature range. One method used a multi-species Cibicidoidesequation (Mg/Ca = 0.90e (0.11⁎BWT)) to approximate the true tempera-ture-Mg/Ca relationship and then subtracted the predicted Mg/Ca forthe observed temperatures from the measured Mg/Ca of C. wuellerstorfifor each sample. This residual Mg/Ca (Mg/Ca CO3) was attributed to thecarbonate ion saturation effect and when plotted versus the ΔCO3
2− ofambient waters yields the equation:
Mg=CaCO3 ¼ 0:00864ΔCO2−3 −0:15 ð9Þ
Elderfield et al., 2006In order to test this relationship,we carriedout a similar exercisewith
our C. wuellerstorfi data from b3 °C. Because the Mg/Ca: temperature
Fig. 8. Mg/CaCO3 vs. ΔCO3 for the C. wuellerstorfi calibration data set from this study(black). Mg/CaCO3=Mg/Cameasured Cameasured− (0.116⁎T+1.2), the Mg/Ca-temperaturerelationship defined by Marchitto et al. (2007) for C. pachyderma. The relationshipbetween Mg/CaCO3 and ΔCO3 for C. wuellerstorfi proposed by Elderfield et al. (2006)(red) is shown for comparison. (For interpretation of the references to color in thisfigure legend, the reader is referred to the web version of this article.)
relationship for C. wuellerstorfi at high temperatures (i.e. Sulu Sea data) issimilar to theC. pachyderma equation ofMarchitto et al. (2007; Fig. 7),weuse this equation (Mg/Ca = 0.116T + 1.20) as an approximation for thefirst-order response of C. wuellerstorfi Mg/Ca to temperature. We thencalculate the difference between our measured Mg/Ca values and theMg/Ca predicted by this relationship to estimate the Mg/Ca CO3. PlottingMg/Ca CO3 versus the ΔCO3
2− at our sample locations yields the followingequation:
Mg=CaCO3 ¼ 0:0083F0:0024ΔCO2−3 −0:28� 0:028 ð10Þ
This equation has a sensitivity that is identical to that determined byElderfield et al. (2006; Fig. 8), suggesting a relatively uniform response ofMg/Ca to the carbonate ion saturation of waters b3 °C. Both O.umbonatus and C. mundulus Mg/Ca data for temperatures b3 °C alsoshow a departure (larger decrease in Mg/Ca per °C) from the calibrationcurves established for these species over a larger temperature range(Figs. 4 and 6, Table 1), further supporting a more observable carbonateion saturation effect in waters b3 °C.
6. Conclusions
We have produced new Mg/Ca measurements on C. wuellerstorfifrom core-top samples from five locations in the Atlantic, Pacific, andIndian Oceans, spanning a depth range of 1095–4548m and a bottomwater temperature range of 0.8–3.8 °C. A linear regression of the datayields a relationship between the Mg/Ca of C. wuellerstorfi and bottomwater temperature of:
Mg=Ca ¼ 0:295F0:0184Tð Þ þ 0:67F0:04 R2 ¼ 0:90
This is similar to equations derived using previously publishedresults for this species at low temperatures (b4 °C), but is a muchlarger decrease inMg/Ca per °C than suggested by eithermulti-speciesCibicidoides equations or single species equations calibrated overlarger temperature ranges (Rosenthal et al., 1997; Lear et al., 2002;Martin et al., 2002; Marchitto et al., 2007; Elderfield et al., 2006). Thisapparent “greater temperature dependence” is most likely due to acombined effect of carbonate ion concentration and temperature onMg uptake by benthic foraminifera that changes significantly at low(b3 °C) temperatures (Elderfield et al., 2006; Rosenthal et al., 2006). Apreliminary quantification of the carbonate ion saturation effect onMg/Ca ratios of C. wuellerstorfi yields the relationship:
Mg=CaCO3 ¼ 0:00834ΔCO2−3
� �−0:28 R2 ¼ 0:51
This equation is identical to the relationship proposed recentlyby Elderfield et al. (2006). This increased slope of the Mg/Ca-temperature relationship at low temperatures is also observed for O.umbonatus and C. mundulus, with the threshold being at ~ 3 °C. C.mundulus yield higher Mg/Ca ratios than C. wuellerstorfi over thesame temperature range, indicating interspecies differences in theuptake of Mg.
Acknowledgments
We would like to thank T. Marchitto for useful discussions, as wellas E. Tappa, M. Luc, and R.McCroan for laboratory assistance.We thankS. Carey from the University of Rhode Island for providing the samplesfrom site TR163-14. Funding for the curation of TR163-14 is providedby NSF grant OCE-0002226.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.epsl.2008.05.023; Boyle andKeigwin, 1985.

530 S.L. Healey et al. / Earth and Planetary Science Letters 272 (2008) 523–530
References
Adkins, J.F., Schrag, D.P., 2001. Pore fluid constraints on deep ocean temperature andsalinity during the Last Glacial Maximum. Geophys. Res. Lett. 28 (5), 771–774.
Boyle, E.A., 1985. Manganese carbonate overgrowths on foraminifera tests. Geochim.Cosmochim. Acta 47, 1815–1819.
Boyle, E.A., Keigwin, L.D., 1985. Comparison of Atlantic and Pacific paleochemicalrecords for the last 215,000years: changes in deep ocean circulation and chemicalinventories. Earth Planet. Sci. Lett. 76, 135–150.
Chappell, J., Shackleton, N.J., 1986. Oxygen isotopes and sea level. Nature 324, 137–140.Elderfield, H., Anand, Yu, J., Kiefer, T., Nyland, B., 2006. Calibrations for benthic
foraminiferal Mg/Ca paleothermometry and the carbonate ion hypothesis. EarthPlanet. Sci. Lett. 250, 633–649.
Fairbanks, R.G., 1989. A 17,000-year glacio-eustatic sea-level record: influence of glacialmelting rates on the Younger Dryas event and deep ocean circulation. Nature 342,637–642.
Labeyrie, L., Duplessy, J.C., Blanc, P.L., 1987. Variations in mode of formation andtemperature of oceanic deep waters over the past 125, 000years. Nature 327,477–482.
Lea, D.W., Mashiotta, T.A., Spero, H.J., 1999. Controls on magnesium and strontiumuptake in planktonic foraminifera determined by live culturing. Geochim.Cosmochim. Acta 63 (16), 2369–2379.
Lear, C.H., Rosenthal, Y., Slowey, N., 2002. Benthic foraminiferal Mg/Ca paleothermo-metry: a revised core-top calibration. Geochim. Cosmochim. Acta 66 (19),3375–3387.
Lewis, E., Wallace, D., 1998. Program developed for CO2 system calculations, Rep. ORNL/CDIAC-105, Carbon Dioxide Inf. Anal. Cent., Oak Ridge Natl. Lab., U.S. Dept. of Energy,Oak Ridge Tenn. (Available at http://cdiac.esd.ornl.gov/oceans/co2rprt.html).
Marchitto, T.M., Bryan, S.P., Curry, W.B., McCorkle, D.C., 2007. Mg/Ca temperaturecalibration for the benthic foraminifer Cibicidoides pachyderma. Paleoceanography22.
Martin, P.A., Lea, D.W., 2002. A simple evaluation of cleaning procedures on fossilbenthic foraminiferal Mg/Ca. Geochem. Geophys. Geosyst. 3 (10), 8401.
Martin, P.A., Lea, D.W., Rosenthal, Y., Shackleton, N.J., Sarnthein, M., Papenfuss, T., 2002.Quaternary deep sea temperature histories derived frombenthic foraminiferalMg/Ca.Earth Planet. Sci. Lett. 222, 275–283.
McConnell, M.C., Thunell, R.C., 2005. Calibration of the planktonic foraminiferal Mg/Capaleothermometer: Sediment trap results from the Guaymas Basin, Gulf ofCalifornia. Paleoceanography 20.
Rathburn, A.E., De Deckker, P., 1997. Magnesium and strontium compositions of recentbenthic foraminifera from the Coral Sea, Australia and Prydz Bay, Antarctica. Mar.Micropaleontol. 32, 231–248.
Rosenthal, Y., Boyle, E.A., Slowey, N., 1997. Temperature control on the incorporation ofmagnesium, strontium, fluorine, and cadmium into benthic foraminiferal shellsfrom Little Bahamas Bank: Prospects for thermocline paleoceanography. Geochim.Cosmochim. Acta 61 (17), 3633–3643.
Rosenthal, Y., Lear, C.H., Oppo, D.W., Linsley, Braddock, 2006. Temperature andcarbonate ion effects on Mg/Ca and Sr/Ca ratios in benthic foraminifera: Aragoniticspecies Hoeglundina elegans. Paleoceanography 21.
Russell, A.D., Emerson, S., Nelson, B.K., Erez, J., Lea, D.W., 1994. Uranium in foraminiferalcalcite as a recorder of seawater uranium concentrations. Geochim. Cosmochim.Acta 58, 671–681.
Schrag, D.P., Hampt, G., Murray, D.W., 1996. Pore fluid constraints on the temperatureand oxygen isotopic composition of the glacial ocean. Science 272, 1930–1932.
Schrag, D.P., Adkins, J.F., McIntyre, K., Alexander, J.L., Hodell, D.A., Charles, C.D.,McManus, J.F., 2002. The oxygen isotopic composition of seawater during the LastGlacial Maximum. Quat. Sci. Rev. 21, 331–342.
Shackleton, N.J., 1967. Oxygen isotope analysis and Pleistocene temperature reassessed.Nature 215, 15–17.
Shackleton, N.J., 1987. Oxygen isotopes, ice volume, and sea level. Quat. Sci. Rev. 6,183–190.
Skinner, L.C., Shackleton, N.J., Elderfield, H., 2003. Millennial-scale variability of deepwater temperature and d18Odw indicating deep-water source variations in theNortheast Atlantic, 0–34cal ka BP. Geochem. Geophys. Geosyst. 4, 1–17.
Van Morkhoven, F.P.C.M., Berggren, W.A., Edwards, A.S., 1986. Cenozoic CosmopolitanDeep-Water Benthic Foraminifera. Bull. Centres Rech. Explor. -Prod. Elf-Aquitaine,Mem. 11, 48–50 Pau.




















