
The lipid dependence of diadinoxanthin de-epoxidation presents new evidence for a macrodomain...
16
Journal of Plant Physiology ] (]]]]) ]]]—]]] The lipid dependence of diadinoxanthin de-epoxidation presents new evidence for a macrodomain organization of the diatom thylakoid membrane Reimund Goss a, , Jana Nerlich a , Bernard Lepetit a , Susann Schaller a , Astrid Vieler b , Christian Wilhelm a a Institute of Biology I, University of Leipzig, Johannisallee 21-23, 04103 Leipzig, Germany b Department of Biochemistry and Molecular Biology, Michigan State University, 215 Biochemistry Building East Lansing, MI 48824-1319, USA Received 29 May 2009; accepted 29 May 2009 KEYWORDS Bilayer lipids; Diatom; Inverted hexagonal phase; Non-bilayer lipids; Xanthophyll cycle Summary The present study shows that thylakoid membranes of the diatom Cyclotella meneghiniana contain much higher amounts of negatively charged lipids than higher plant or green algal thylakoids. Based on these findings, we examined the influence of SQDG on the de-epoxidation reaction of the diadinoxanthin cycle and compared it with results from the second negatively charged thylakoid lipid PG. SQDG and PG exhibited a lower capacity for the solubilization of the hydrophobic xanthophyll cycle pigment diadinoxanthin than the main membrane lipid MGDG. Although complete pigment solubilization took place at higher concentrations of the negatively charged lipids, SQDG and PG strongly suppressed the de-epoxidation of diadinoxanthin in artificial membrane systems. In in vitro assays employing the isolated diadinoxanthin cycle enzyme diadinoxanthin de-epoxidase, no or only a very weak de-epoxidation reaction was observed in the presence of SQDG or PG, respectively. In binary mixtures of the inverted hexagonal phase forming lipid MGDG with the negatively charged bilayer lipids, comparable suppression took place. This is in contrast to binary mixtures of MGDG with the neutral bilayer lipids DGDG and PC, where rapid and efficient de-epoxidation was observed. In complex lipid ARTICLE IN PRESS www.elsevier.de/jplph 0176-1617/$ - see front matter & 2009 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2009.05.017 Abbreviations: DDE, diadinoxanthin de-epoxidase; Ddx, diadinoxanthin; DGDG, digalactosyldiacylglycerol; DGTS, 1,2-diacylglyceryl- 3-O-4 0 -(N,N,N-trimethyl)-homoserine; Dtx, diatoxanthin; FCP, fucoxanthin chlorophyll protein complex; H II phase, inverted hexagonal phase; LHC, light-harvesting complex; MGDG, monogalactosyldiacylgycerol; PC, phosphatidylcholine; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; PS, photosystem; SQDG, sulfoquinovosyldiacylglycerol; TLC, thin-layer chromatography; VDE, violaxanthin de-epoxidase; Vx, violaxanthin. Corresponding author. Tel.: +49341 9736873; fax: +49341 9736899. E-mail address: [email protected] (R. Goss). Please cite this article as: Goss R, et al. The lipid dependence of diadinoxanthin de-epoxidation presents new evidence for a macrodomain organization of the diatom thylakoid membrane. J Plant Physiol (2009), doi:10.1016/j.jplph.2009.05.017
Transcript of The lipid dependence of diadinoxanthin de-epoxidation presents new evidence for a macrodomain...

ARTICLE IN PRESS
Journal of Plant Physiology ] (]]]]) ]]]—]]]
0176-1617/$ - sdoi:10.1016/j.
Abbreviation3-O-40-(N,N,N-tphase; LHC, ligPG, phosphatidde-epoxidase;�CorrespondE-mail addr
Please cite tmacrodomain
www.elsevier.de/jplph
The lipid dependence of diadinoxanthinde-epoxidation presents new evidence for amacrodomain organization of the diatomthylakoid membrane
Reimund Gossa,�, Jana Nerlicha, Bernard Lepetita, Susann Schallera,Astrid Vielerb, Christian Wilhelma
aInstitute of Biology I, University of Leipzig, Johannisallee 21-23, 04103 Leipzig, GermanybDepartment of Biochemistry and Molecular Biology, Michigan State University, 215 Biochemistry Building East Lansing,MI 48824-1319, USA
Received 29 May 2009; accepted 29 May 2009
KEYWORDSBilayer lipids;Diatom;Inverted hexagonalphase;Non-bilayer lipids;Xanthophyll cycle
ee front matter & 2009jplph.2009.05.017
s: DDE, diadinoxanthinrimethyl)-homoserine;ht-harvesting complex;ylglycerol; PS, photosyVx, violaxanthin.ing author. Tel.: +49 341ess: [email protected]
his article as: Goss R,organization of the dia
SummaryThe present study shows that thylakoid membranes of the diatom Cyclotellameneghiniana contain much higher amounts of negatively charged lipids than higherplant or green algal thylakoids. Based on these findings, we examined the influenceof SQDG on the de-epoxidation reaction of the diadinoxanthin cycle and compared itwith results from the second negatively charged thylakoid lipid PG. SQDG and PGexhibited a lower capacity for the solubilization of the hydrophobic xanthophyllcycle pigment diadinoxanthin than the main membrane lipid MGDG. Althoughcomplete pigment solubilization took place at higher concentrations of thenegatively charged lipids, SQDG and PG strongly suppressed the de-epoxidation ofdiadinoxanthin in artificial membrane systems. In in vitro assays employing theisolated diadinoxanthin cycle enzyme diadinoxanthin de-epoxidase, no or only a veryweak de-epoxidation reaction was observed in the presence of SQDG or PG,respectively. In binary mixtures of the inverted hexagonal phase forming lipid MGDGwith the negatively charged bilayer lipids, comparable suppression took place. Thisis in contrast to binary mixtures of MGDG with the neutral bilayer lipids DGDG andPC, where rapid and efficient de-epoxidation was observed. In complex lipid
Elsevier GmbH. All rights reserved.
de-epoxidase; Ddx, diadinoxanthin; DGDG, digalactosyldiacylglycerol; DGTS, 1,2-diacylglyceryl-Dtx, diatoxanthin; FCP, fucoxanthin chlorophyll protein complex; HII phase, inverted hexagonalMGDG, monogalactosyldiacylgycerol; PC, phosphatidylcholine; PE, phosphatidylethanolamine;stem; SQDG, sulfoquinovosyldiacylglycerol; TLC, thin-layer chromatography; VDE, violaxanthin
9736873; fax: +49 341 9736899.g.de (R. Goss).
et al. The lipid dependence of diadinoxanthin de-epoxidation presents new evidence for atom thylakoid membrane. J Plant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
R. Goss et al.2
Please cite this article as: Goss R,macrodomain organization of the dia
mixtures resembling the lipid composition of the native diatom thylakoid membrane,we again found strong suppression of diadinoxanthin de-epoxidation due to thepresence of SQDG or PG. We conclude that, in the native thylakoids of diatoms, astrict separation of the MGDG and SQDG domains must occur; otherwise, the rapiddiadinoxanthin de-epoxidation observed in intact cells upon illumination would notbe possible.& 2009 Elsevier GmbH. All rights reserved.
Introduction
The diadinoxanthin (Ddx) cycle represents themost important photoprotection mechanism ofdiatoms (Lavaud et al., 2002; Goss et al., 2006).It is located in the chloroplast and consists of a de-epoxidation and epoxidation reaction (Stransky andHager, 1970). De-epoxidation comprises a one stepde-epoxidation from the mono-epoxy xanthophyllDdx to the epoxy-free diatoxanthin (Dtx), and takesplace when intact diatom cells are illuminated withhigh actinic light intensities. Low light stimulatesthe back reaction of the Ddx cycle, in which the de-epoxidized Dtx is epoxidized to Ddx (for a reviewsee Wilhelm et al., 2006).
The de-epoxidation step in the Ddx cycle iscatalyzed by the enzyme Ddx de-epoxidase (DDE),which is located in the thylakoid lumen andbecomes activated by a decreasing lumenal pHdue to light-driven photosynthetic electron trans-port. DDE exhibits significant differences whencompared with the violaxanthin (Vx) cycle enzymeof vascular plants, the Vx de-epoxidase (VDE). DDEactivation can be observed at almost neutral pHvalues (Jakob et al., 2001; Grouneva et al., 2006),whereas the threshold value for VDE activation isfound at more acidic pH values (Pfundel et al.,1994). Further, DDE exhibits a significantly lower KMvalue for its co-substrate ascorbate compared withthe VDE of vascular plants (Grouneva et al., 2006).
Like VDE activity, DDE activity is strongly en-hanced in the presence of non-bilayer lipids likesuch as the major thylakoid membrane lipidmonogalactosyldiacylgycerol (MGDG) (Latowskiet al., 2002; Goss et al., 2005, 2007). In the thylakoidmembranes of vascular plants, MGDG comprisesabout half of the total lipids. The second mostabundant thylakoid lipid is the bilayer-formingdigalactosyldiacylglycerol (DGDG), which amountsto about 30% of the thylakoid lipids (Joyard et al.,1998; Siegenthaler, 1998). The lipid classes, whichare present in the thylakoid membranes of vascularplants, are comparable to those found in intactdiatom cells, where MGDG also represents the mainlipid (Harwood, 1998; Vieler et al., 2007; Goss andWilhelm, 2009).
et al. The lipid dependence otom thylakoid membrane. J P
With regard to the concentration of chargedlipids, however, significant differences exist be-tween diatoms and vascular plants. Diatom cellscontain high amounts of the negatively chargedlipids sulfoquinovosyldiacylglycerol (SQDG) andphosphatidylglycerol (PG), and SQDG surpassesDGDG as the second most abundant lipid. Theoverall concentration of negatively charged lipids isabout twice as high in diatoms compared with thatof higher plants or green algae (Vieler et al., 2007).Diatoms contain about 40% of their total cell lipid inthe form of SQDG and PG, whereas in vascular plants,these two lipid species amount to only 20% or less.Significant differences in vascular plant and diatomlipids also exist with respect to the fatty acidscontained in the different lipid species. MGDG andDGDG from diatoms contain longer chain fatty acidswith up to 22 carbon atoms and as many as six doublebonds (Harwood, 1998; Goss and Wilhelm, 2009).
Due to strongly increased solubility of Ddx inMGDG compared with Vx, the MGDG concentrationsfor optimal DDE activity are significantly lowercompared to those needed for full activation of VDE(Goss et al., 2005, 2007). In addition to its role inpigment solubilization, MGDG provides anotherimportant feature for Ddx de-epoxidation. MGDGbelongs to the so-called non-bilayer lipids, whichdue to their small head-group area and criticalpacking parameter value higher than one, forminverted hexagonal structures (HII phases) in anaqueous medium (Shipley et al., 1973; Israelachviliand Mitchell, 1975). These three-dimensionalstructures promote efficient Ddx de-epoxidationin single lipid systems (Goss et al., 2005) orartificial membrane systems composed of bilayerlipids like phosphatidylcholine (PC) and MGDG (Gosset al., 2007). It has been proposed that the HII
phases enable the binding of DDE to the thylakoidmembrane after the pH-dependent activation ofthe enzyme has taken place. Recently, it has beenobserved that the transition from the gel (Lb) to theliquid-crystalline (La) phase in a membrane bilayerstrongly enhances both the solubilization of thexanthophyll cycle pigments and the activity of theVDE (Vieler et al., 2008). At higher temperaturesthe La phase seems to be even better suited to
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
Diadinoxanthin de-epoxidation and lipid organization in diatoms 3
promoting VDE activity than the HII phase, which isadvantageous at lower temperatures.
Although the existence of HII phases in thethylakoid membranes of vascular plants has beenindicated by various methods (Gounaris et al., 1983;Haranczyk et al., 1995), definitive evidence of theirpresence in the native membrane is still missing. It isalso unclear if these phases may exist in the plane ofa surrounding lipid bilayer or if the HII phases aresegregated from the membrane bilayer upon theirformation (Garab et al., 2000). For diatoms, thesituation is even less clear, as precise informationabout the lipid composition of thylakoid membranesis nonexistent, due to the complicated preparationprocedure for purified thylakoids. This means that,before information about the existence of, HII phasesin the thylakoid membranes of diatoms can begathered, a detailed determination of the lipid‘composition must be performed.
We attempted to perform such an analysis in thepresent study by determining the lipid compositionof the native thylakoid membrane of the diatomCyclotella meneghiniana. It was our aim to clarifywhether the high concentrations of SQDG and PGfound in intact cells of this alga (Vieler et al., 2007)are also reflected by the lipid composition of theisolated thylakoids. We further aimed to examinehow the negatively charged lipid SQDG influencesthe Ddx de-epoxidation reaction, which normallydepends on the presence of the HII phase forminggalactolipid MGDG (Goss et al., 2005, 2007). Toanalyze whether the effects of SQDG on DDEactivity are a general feature of negatively chargedlipids, we included the second negatively chargedthylakoid membrane lipid, PG, into our presentexperiments. The influence of SQDG and PG on theDdx de-epoxidation was tested in in vitro de-epoxidation assays with the isolated Ddx cycleenzyme DDE. Furthermore, the capacity of SQDGand PG for the solubilization of the hydrophobicsubstrate of the DDE, Ddx, was determined. SQDGand PG were added to the enzyme assay as singlelipids or in combination with the HII phase forminglipid MGDG. Finally, complex lipid mixtures, re-sembling the lipid composition of native diatomthylakoids, were tested in the enzyme assays withrespect to a possible enhancement or suppressionof the Ddx de-epoxidation reaction.
Materials and methods
Plant material
The diatom Cyclotella meneghiniana (strain SAG1020-1a) and the green alga Chlamydomonas
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
reinhardtii (strain SAG 11-32b) were obtained fromthe Culture Collection of Algae, Gottingen (SAG,Germany). Fresh spinach leaves were obtainedfrom the local market. C. meneghiniana cultureswere grown as air-lift cultures in artificial seawatermedium f/2 according to Guillard and Lorenzen(1972) with double silica and half salt content.C. reinhardtii was grown as batch cultures in basalES medium. The algae were cultivated at a lightintensity of 30 mmolm�2 s�1 (PAR) with a light:darkregime of 14:10 h. The temperature during growthwas held constant at 20 1C. For the preparation ofthylakoid membranes, the DDE and Ddx, the algaewere harvested from the late logarithmic growthphase at a chlorophyll content of 10mg Chl a+c L�1.
Preparation of thylakoid membranes and DDE
C. reinhardtii cells were harvested by a 10mincentrifugation at 2500g and 4 1C (Varifuge 3.0,Heraeus, Germany). The cells were resuspended inisolation medium A (330mM sorbitol, 10mM NaCl,1mM MgCl2, 1mM MnCl2, 2mM EDTA, 0.5mMKH2PO4, 44mM MES pH 6.1, 0.4% BSA) and disruptedin a French Pressure Cell (Thermo Spectronic,Cambridge, UK) at 4000 psi. The collected suspen-sion was then centrifuged at 500g and 4 1C for 5min(Allegra 64R, Beckman Coulter, USA) to removeunbroken cells. The supernatant was centrifugedfor 7min at 3000g and 4 1C. The resulting pellet wasresuspended in shock medium B (10mM NaCl, 5mMMgCl2, 10mM HEPES, pH 6.7) and centrifuged at10,000g and 4 1C for 10min. The resulting thyla-koids were stored in a small volume of storagemedium C (330mM sorbitol, 10mM NaCl, 1mMMgCl2, 1mM MnCl2, 2mM EDTA, 0.5mM KH2PO4,40mM HEPES pH 6.7, 0.4% BSA) until further use.The preparation of C. meneghiniana thylakoidsfollowed the protocol described by Lepetit et al.(2007) for the preparation of thylakoids from thediatom P. tricornutum.
To release the DDE from the thylakoid mem-branes of C. meneghiniana, the thylakoids werecentrifuged at 10,000g for 5min at 4 1C (Allegra64R, Beckman Coulter, USA) and the pellet wasincubated in DDE preparation medium (5mM MgCl2,20mM Tris pH 7.5) for 5min. After the incubation,the thylakoids were centrifuged at 40,000g and 1 1Cfor 40min (Allegra 64R, Beckman Coulter, USA),and the resulting supernatant, which contained theDDE, was collected and stored at �20 1C for use inthe enzyme assays.
Spinach thylakoid membranes were isolatedaccording to standard procedures as described byJensen and Bassham (1966).
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
R. Goss et al.4
Preparation of Ddx
Ddx was extracted from dark-adapted cells ofC. meneghiniana and purified by HPLC according toGoss et al. (2005). Ddx from several HPLC separa-tions was pooled, dried under nitrogen and storedat �20 1C until further use in absorption spectro-scopy and the enzyme assays.
In vitro de-epoxidation assays
The influence of different lipids on Ddx de-epoxidation was tested in in vitro de-epoxidationassays. The enzyme assays were performed in thepresence of single bilayer or non-bilayer lipids,binary mixtures of MGDG with different bilayerlipids, or complex mixtures of up to four lipidsresembling the lipid composition of native diatomthylakoids. The lipids were used at a concentrationof either 11.6 or 38.7 mM in the enzyme assays;details about the lipids and the composition of thelipid mixtures are presented in the Results sectionand the legends of the figures. In addition to thelipids, the standard enzyme assay contained 0.4 mMDdx and 30mM ascorbate. Ddx de-epoxidation wascarried out in a reaction medium adjusted to thepH optimum of DDE (10mM KCl, 5mM MgCl2 and40mM MES pH 5.2). The reaction mixture wasprepared by mixing the ethanolic pigment solutionwith lipid, and injecting the pigment/lipid mixtureinto the reaction buffer with a Hamilton syringe.After addition of the DDE solution (for a 3mL assay,100 mL of DDE solution were used), the reactionmixture was incubated at 30 1C for 5min before a500 mL control sample (corresponding to time point0 of the de-epoxidation kinetics) was taken. De-epoxidation of Ddx was started by addition ofascorbate to the reaction mixture, and sampleswere collected after de-epoxidation times of 3, 5,10 and 20min. In the samples, de-epoxidation wasstopped by injecting 500 mL of the reaction assayinto 500 mL of an extraction medium consisting of amixture of chloroform:methanol:NH3 with the ratio1:2:0.004 (v/v). After a phase separation had takenplace, the organic phase was collected, dried undera gentle stream of nitrogen and stored at �20 1Cuntil pigment analysis by HPLC was performed.
Ddx de-epoxidation is depicted as an increase inthe de-epoxidation state (DES) of the Ddx cyclepigment pool calculated as Dtx/(Ddx+Dtx).
Determination of Ddx solubility
Ddx solubility was determined by absorptionspectroscopy. 0.4 mM Ddx was mixed with single
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
lipids, binary mixtures of MGDG with bilayer lipidsor with complex lipid mixtures of up to fourthylakoid membrane lipids (for a detailed descrip-tion see the Results section). The lipid concentra-tion was varied within a range from 1.45 to 38.7 mM.The pigment/lipid mixture was then injected intothe reaction medium pH 5.2 (composition seeabove), which was adjusted to a temperature of30 1C. Absorption spectra of Ddx were recorded ona Specord M 500 photometer (Zeiss, Germany) in awavelength range between 350 and 550 nm with abandpass setting of 1 nm against reference sampleswithout Ddx, which contained the same amount oflipid dissolved in reaction medium pH 5.2. Thesolubility of Ddx in the different lipids and lipidmixtures is depicted as III/II values of the respec-tive Ddx spectra. The III/II values were calculatedas the ratio of the third absorption peak at 485 nmto the second absorption peak at 455 nm, with theminimum between the two peaks at 470 nm servingas the baseline. The III/II values of the Ddx spectraat different lipid concentrations were then normal-ized to the maximum III/II value achieved aftercomplete solubilization of Ddx in ethanol.
Pigment analysis by HPLC
The dried pigment samples from the in vitro de-epoxidation assays were dissolved in HPLC extrac-tion medium consisting of 90% methanol/0.2Mammonium acetate (9:1, v/v) and 10% ethylacetate. The pigments were analyzed on a Waters600-MS chromatography system with a Waters 996photodiode array detector (Waters, Germany)equipped with a Nucleosil ET 250/8/4, 300-5,C-18 column (Macherey & Nagel, Germany), usingthe eluents and gradient described by Goss et al.(2005). The detection wavelength for Ddx and Dtxwas set to 480 nm and the pigments were quantifiedaccording to Lohr and Wilhelm (2001).
Lipid extraction from thylakoid membranes
Total lipids were extracted from the isolatedthylakoid membranes by addition of CHCl3:CH3OH(2:1, v/v) to yield a final concentration ofCHCl3:CH3OH:H2O of 8:4:3 (v/v) according to Folchet al. (1957). After the separation of the organicand the aqueous phase had taken place, the organicphase was collected and washed with 5M NaCl at aratio of organic solvent to NaCl solution of 60/40.The water phase was re-extracted and the organicextracts were combined and dried under a gentlestream of nitrogen. The dried lipids were then
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
Diadinoxanthin de-epoxidation and lipid organization in diatoms 5
stored at �20 1C until lipid determination by thin-layer chromatography.
Separation and quantification of lipids
Concentrated lipid extracts were applied toHPTLC silica gel 60 plates (Merck, Germany) anddeveloped in thin-layer chromatography (TLC)chambers using two different eluent systems. Thefirst system was based on a method describedby Mock and Kroon (2002) with methylacetate:isopropanol:CHCl3:MeOH:KCl (0.25%) in the ratio25:25:25:25:10:4 as eluent. The second systemwas performed according to Ventrella et al. (2007)with CHCl3:MeOH:acetic acid:H2O in the ratio75:13:9:3 as eluent. The use of two differenteluent systems was necessary in order to separateall different lipid classes of the native thylakoidmembranes of vascular plants, green algae anddiatoms. After the separation, the lipids werestained with primuline (Direct Yellow 59) accordingto White et al. (1998). The resulting spots weresemi-quantitatively assessed using a Biometradigital imaging system in combination with the pro-gram BiodocAnalyze (Biometra, Germany). The con-centration of the different lipids was calculated aspercentage of lipid of the total lipid concentration.
Chemicals
MGDG, DGDG, SQDG and PG were purchased fromLipid Products (UK). PC was obtained from Sigma-Aldrich (USA).
Table 1. Lipid composition of isolated thylakoid membraneC. meneghiniana.
Lipid Spinach (% of total lipid)
MGDG 45DGDG 29SQDG 11PG 9PE –
DGTS –
PC 6
Lipid ratio neutral/negative
4
The lipid concentrations are depicted as % lipid of the total thylakoidis additionally depicted. This table shows the mean values of three ind10% and 20%. Note that, for the calculation of the lipid percentagesfatty acid composition of the different diatom lipid classes is presen
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
Results
Lipid composition of isolated thylakoidmembranes
Table 1 shows the lipid composition of isolatedthylakoid membranes of the diatom C. meneghini-ana and the green alga C. reinhardtii. For acomparison, the lipid composition of spinachthylakoids is also depicted. In the thylakoidmembranes of C. reinhardtii, the galactolipidMGDG was the main lipid and amounted to around40% of the total lipid content. In C. meneghiniana,the MGDG content was significantly lower andrepresented less than 30% of the thylakoid mem-brane lipid. In spinach thylakoid membranes, high-er concentrations of MGDG were present than inboth algal species and MGDG represented almosthalf of the total thylakoid lipid content. In bothspinach and C. reinhardtii, DGDG was the secondmost abundant lipid, whereas in the diatomC. meneghiniana, the DGDG concentration wassignificantly reduced. Interestingly, in C. meneghini-ana, the negatively charged sulfolipid SQDG wasfound to be the main thylakoid lipid, with a slightlyhigher concentration than MGDG. The SQDG con-centration in the diatom thylakoid membranes wasthree times higher than the SQDG concentration inspinach and more than two times higher than theSQDG content of C. reinhardtii. For the secondnegatively charged membrane lipid, PG, compar-able concentrations were observed in the thyla-koids of both algae and spinach. However, it isimportant to note that, in diatom thylakoids, thenegatively charged lipids amounted to more than40% of the total thylakoid lipid content, whereas in
s of spinach, the green alga C. reinhardtii and the diatom
C. reinhardtii(% of total lipid)
C. meneghiniana(% of total lipid)
38 2820 1214 338 125 –
15 –
– 15
3.5 1.2
membrane lipid. The ratio of neutral to negatively charged lipidsependent lipid determinations with standard deviations between, the mass of the respective lipids was used because the precisetly unknown.
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
R. Goss et al.6
C. reinhardtii and spinach, SQDG and PG repre-sented only around 20% of the total membranelipid. This resulted in drastic differences in theratio of neutral to negatively charged lipids (Table 1).While spinach and C. reinhardtii thylakoid mem-branes contained approximately four times moreneutral than negatively charged lipids, the thyla-koids of C. meneghiniana were characterized byalmost equal amounts of both lipid types. Thephospholipid phosphatidylethanolamine (PE) wasfound in traces in C. reinhardtii, but was comple-tely absent in spinach and C. meneghiniana. Thebetaine lipid 1,2-diacylglyceryl-3-O-40-(N,N,N-tri-methyl)-homoserine (DGTS) was a major compo-nent of the thylakoid membranes of C. reinhardtii,but was absent in spinach and C. meneghiniana.The phospholipid PC, on the other hand, was notpresent in the green alga, but was observed in thethylakoid membranes of spinach and C. meneghini-ana, where the highest PC concentrations werefound.
For C. meneghiniana, we also analyzed the lipidcontent of intact cells (data not shown). We foundthat the lipid composition was comparable to thatof the isolated thylakoid membranes depicted inTable 1, and that the slight differences corre-sponded to the error range of the lipid quantifica-tion. This observation indicates that the lipidcontent of the intact cells was dominated by thelipid content of the thylakoid membranes, which isin accordance with electron micrographs andrelative respiratory activities of diatoms. Thesedata indicate that the diatom thylakoid lipids mayrepresent up to 90% of the total phospho- andglycolipids of the cell. The data also show that theintact cells of C. meneghiniana that were used forthe present experiments did not contain significantamounts of triacylglycerides, which can be used asstorage lipids.
Solubility of Ddx in single lipids
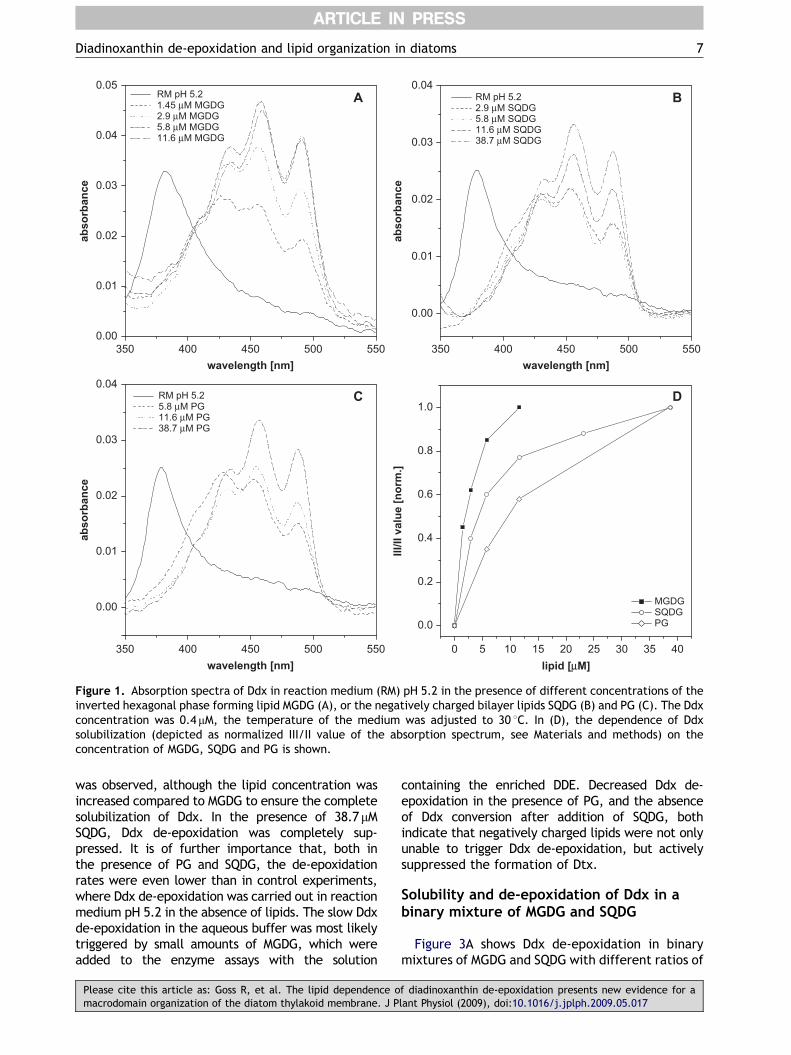
Figure 1 compares the solubility of Ddx in theinverted hexagonal phase forming lipid MGDG andthe two negatively charged bilayer lipids SQDG andPG. In the absence of lipids, i.e. when Ddx wasinjected into the aqueous reaction medium, theabsorption spectrum exhibited a single peak ataround 380 nm. Addition of increasing concentra-tions of MGDG (Figure 1A) restored the typical Ddxabsorption spectrum, which is normally found inorganic solvents, and shifted the absorption max-imum to 458 nm. At low MGDG concentrations, thethree peak structure of the Ddx spectrum wasalready visible, but the absorption spectrum was
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
still slightly distorted. In the present experiments,5.8 mM MGDG led to an almost complete solubiliza-tion of 0.4 mM Ddx, whereas a further increase inthe MGDG concentration only slightly enhanced thefine structure of the Ddx spectrum. The depen-dence of Ddx solubilization on the MGDG concen-tration can also be seen in Figure 1D, where thenormalized III/II values of the Ddx spectrum serveas a measure of Ddx solubility. Figure 1D shows thatcomplete Ddx solubilization took place at MGDGconcentrations between 5.8 and 11.6 mM MGDG,corresponding to a lipid per pigment ratio of about15. When SQDG (Figure 1B) and PG (Figure 1C) wereused to solubilize Ddx in the aqueous reactionmedium, higher lipid concentrations were neededto achieve the typical Ddx spectrum. At concentra-tions of 11.6 mM SQDG and PG, the spectrum stilllooked slightly distorted, although the three peakstructure was observable. Complete solubilizationof Ddx in SQDG and PG was found at lipidconcentrations of 38.7 mM, which corresponds to alipid per pigment ratio of more than 90. Althoughboth negatively charged lipids proved to be lessefficient in Ddx solubilization compared withMGDG, SQDG was better suited than PG. Especiallyat lower concentrations of SQDG, the fine structureof the Ddx spectrum was more defined than in theassays with PG (compare Figures 1B and C), and thenormalized III/II ratios reached higher values(Figure 1D). It is also noteworthy that SQDG andPG exhibited a slightly higher solubilization capa-city for Ddx than the uncharged bilayer lipid DGDGand the neutral bilayer lipid PC, which wereanalyzed in a previous study (Goss et al., 2005).
De-epoxidation of Ddx in the presence ofsingle lipids
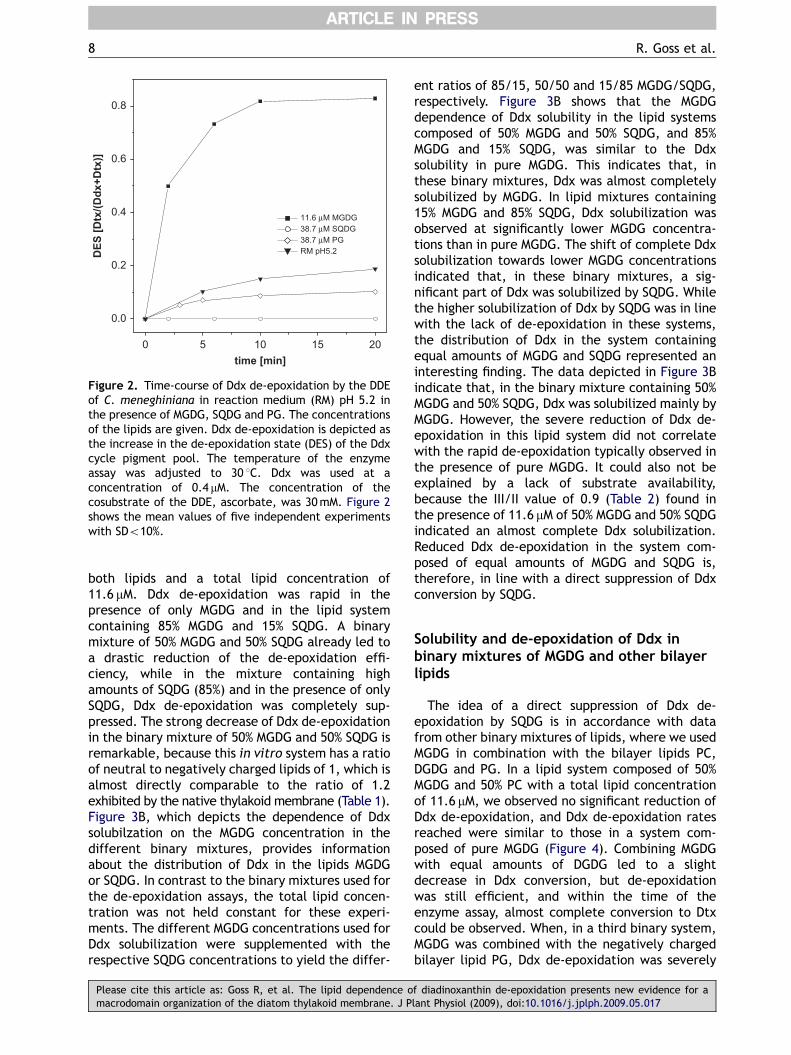
Figure 2 shows the dependence of Ddx de-epoxidation (depicted as an increase in the de-epoxidation state of the Ddx cycle pigment pool) onthe presence of different concentrations of MGDG,SQDG and PG. Ddx de-epoxidation was rapid in invitro de-epoxidation assays in the presence of11.6 mM MGDG, corresponding to the MGDG con-centration where complete Ddx solubilization wasachieved. After 10min of the enzyme assay, Ddxwas almost completely converted to Dtx, resultingin a de-epoxidation state of the Ddx cycle poolhigher than 0.8. Additional incubation for 10mindid not lead to further significant increases in thede-epoxidation state, indicating that the reactionwas completed after 10min and that a part of Ddxwas not convertible to Dtx. In the presence of38.7 mM PG, strongly reduced Ddx de-epoxidation
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017
ARTICLE IN PRESS
3500.00
0.01
0.02
0.03
0.04
0.05RM pH 5.21.45 µM MGDG2.9 µM MGDG5.8 µM MGDG11.6 µM MGDG
abso
rban
ce
wavelength [nm]350
0.00
0.01
0.02
0.03
0.04RM pH 5.22.9 µM SQDG5.8 µM SQDG11.6 µM SQDG38.7 µM SQDG
abso
rban
ce
wavelength [nm]
350
0.00
0.01
0.02
0.03
0.04RM pH 5.25.8 µM PG11.6 µM PG38.7 µM PG
abso
rban
ce
wavelength [nm]0
0.0
0.2
0.4
0.6
0.8
1.0
MGDGSQDGPG
III/II
val
ue [n
orm
.]
lipid [µM]400 450 500 550 5 10 15 20 25 30 35 40
400 450 500 550400 450 500 550
A B
C D
Figure 1. Absorption spectra of Ddx in reaction medium (RM) pH 5.2 in the presence of different concentrations of theinverted hexagonal phase forming lipid MGDG (A), or the negatively charged bilayer lipids SQDG (B) and PG (C). The Ddxconcentration was 0.4mM, the temperature of the medium was adjusted to 30 1C. In (D), the dependence of Ddxsolubilization (depicted as normalized III/II value of the absorption spectrum, see Materials and methods) on theconcentration of MGDG, SQDG and PG is shown.
Diadinoxanthin de-epoxidation and lipid organization in diatoms 7
was observed, although the lipid concentration wasincreased compared to MGDG to ensure the completesolubilization of Ddx. In the presence of 38.7mMSQDG, Ddx de-epoxidation was completely sup-pressed. It is of further importance that, both inthe presence of PG and SQDG, the de-epoxidationrates were even lower than in control experiments,where Ddx de-epoxidation was carried out in reactionmedium pH 5.2 in the absence of lipids. The slow Ddxde-epoxidation in the aqueous buffer was most likelytriggered by small amounts of MGDG, which wereadded to the enzyme assays with the solution
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
containing the enriched DDE. Decreased Ddx de-epoxidation in the presence of PG, and the absenceof Ddx conversion after addition of SQDG, bothindicate that negatively charged lipids were not onlyunable to trigger Ddx de-epoxidation, but activelysuppressed the formation of Dtx.
Solubility and de-epoxidation of Ddx in abinary mixture of MGDG and SQDG
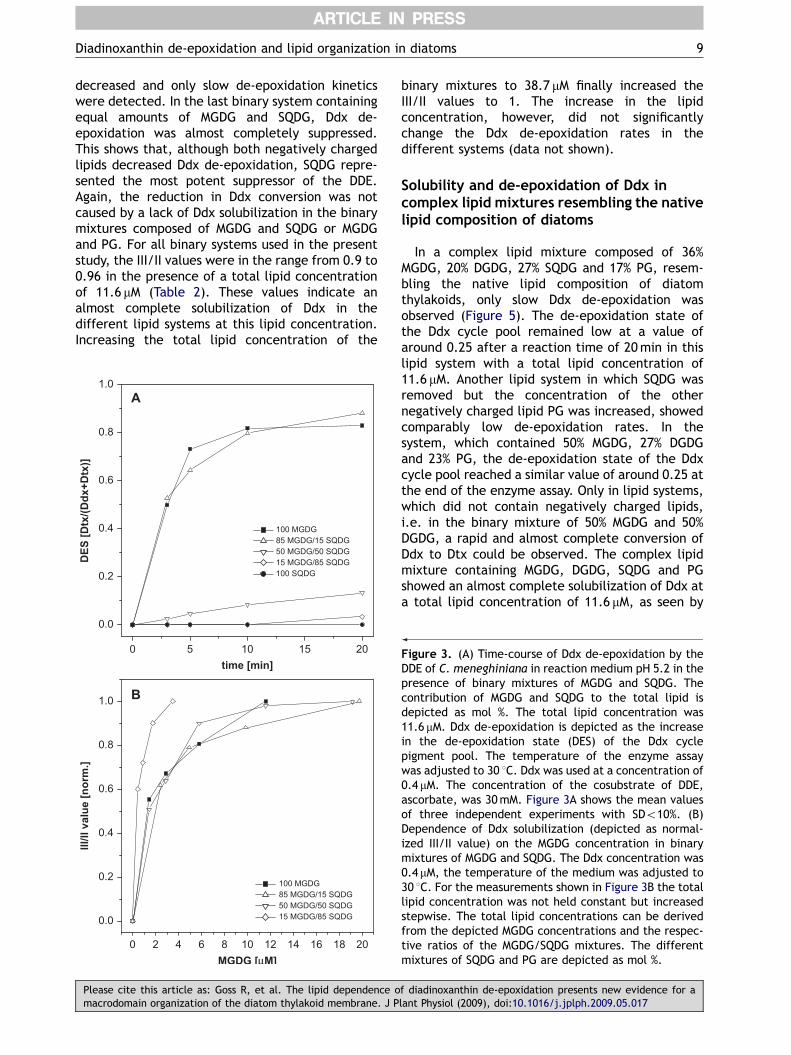
Figure 3A shows Ddx de-epoxidation in binarymixtures of MGDG and SQDG with different ratios of
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017
ARTICLE IN PRESS
0
0.0
0.2
0.4
0.6
0.8
11.6 µM MGDG38.7 µM SQDG38.7 µM PGRM pH5.2D
ES [D
tx/(D
dx+D
tx)]
time [min]5 10 15 20
Figure 2. Time-course of Ddx de-epoxidation by the DDEof C. meneghiniana in reaction medium (RM) pH 5.2 inthe presence of MGDG, SQDG and PG. The concentrationsof the lipids are given. Ddx de-epoxidation is depicted asthe increase in the de-epoxidation state (DES) of the Ddxcycle pigment pool. The temperature of the enzymeassay was adjusted to 30 1C. Ddx was used at aconcentration of 0.4 mM. The concentration of thecosubstrate of the DDE, ascorbate, was 30mM. Figure 2shows the mean values of five independent experimentswith SDo10%.
R. Goss et al.8
both lipids and a total lipid concentration of11.6 mM. Ddx de-epoxidation was rapid in thepresence of only MGDG and in the lipid systemcontaining 85% MGDG and 15% SQDG. A binarymixture of 50% MGDG and 50% SQDG already led toa drastic reduction of the de-epoxidation effi-ciency, while in the mixture containing highamounts of SQDG (85%) and in the presence of onlySQDG, Ddx de-epoxidation was completely sup-pressed. The strong decrease of Ddx de-epoxidationin the binary mixture of 50% MGDG and 50% SQDG isremarkable, because this in vitro system has a ratioof neutral to negatively charged lipids of 1, which isalmost directly comparable to the ratio of 1.2exhibited by the native thylakoid membrane (Table 1).Figure 3B, which depicts the dependence of Ddxsolubilzation on the MGDG concentration in thedifferent binary mixtures, provides informationabout the distribution of Ddx in the lipids MGDGor SQDG. In contrast to the binary mixtures used forthe de-epoxidation assays, the total lipid concen-tration was not held constant for these experi-ments. The different MGDG concentrations used forDdx solubilization were supplemented with therespective SQDG concentrations to yield the differ-
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
ent ratios of 85/15, 50/50 and 15/85 MGDG/SQDG,respectively. Figure 3B shows that the MGDGdependence of Ddx solubility in the lipid systemscomposed of 50% MGDG and 50% SQDG, and 85%MGDG and 15% SQDG, was similar to the Ddxsolubility in pure MGDG. This indicates that, inthese binary mixtures, Ddx was almost completelysolubilized by MGDG. In lipid mixtures containing15% MGDG and 85% SQDG, Ddx solubilization wasobserved at significantly lower MGDG concentra-tions than in pure MGDG. The shift of complete Ddxsolubilization towards lower MGDG concentrationsindicated that, in these binary mixtures, a sig-nificant part of Ddx was solubilized by SQDG. Whilethe higher solubilization of Ddx by SQDG was in linewith the lack of de-epoxidation in these systems,the distribution of Ddx in the system containingequal amounts of MGDG and SQDG represented aninteresting finding. The data depicted in Figure 3Bindicate that, in the binary mixture containing 50%MGDG and 50% SQDG, Ddx was solubilized mainly byMGDG. However, the severe reduction of Ddx de-epoxidation in this lipid system did not correlatewith the rapid de-epoxidation typically observed inthe presence of pure MGDG. It could also not beexplained by a lack of substrate availability,because the III/II value of 0.9 (Table 2) found inthe presence of 11.6 mM of 50% MGDG and 50% SQDGindicated an almost complete Ddx solubilization.Reduced Ddx de-epoxidation in the system com-posed of equal amounts of MGDG and SQDG is,therefore, in line with a direct suppression of Ddxconversion by SQDG.
Solubility and de-epoxidation of Ddx inbinary mixtures of MGDG and other bilayerlipids
The idea of a direct suppression of Ddx de-epoxidation by SQDG is in accordance with datafrom other binary mixtures of lipids, where we usedMGDG in combination with the bilayer lipids PC,DGDG and PG. In a lipid system composed of 50%MGDG and 50% PC with a total lipid concentrationof 11.6 mM, we observed no significant reduction ofDdx de-epoxidation, and Ddx de-epoxidation ratesreached were similar to those in a system com-posed of pure MGDG (Figure 4). Combining MGDGwith equal amounts of DGDG led to a slightdecrease in Ddx conversion, but de-epoxidationwas still efficient, and within the time of theenzyme assay, almost complete conversion to Dtxcould be observed. When, in a third binary system,MGDG was combined with the negatively chargedbilayer lipid PG, Ddx de-epoxidation was severely
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017
ARTICLE IN PRESS
Diadinoxanthin de-epoxidation and lipid organization in diatoms 9
decreased and only slow de-epoxidation kineticswere detected. In the last binary system containingequal amounts of MGDG and SQDG, Ddx de-epoxidation was almost completely suppressed.This shows that, although both negatively chargedlipids decreased Ddx de-epoxidation, SQDG repre-sented the most potent suppressor of the DDE.Again, the reduction in Ddx conversion was notcaused by a lack of Ddx solubilization in the binarymixtures composed of MGDG and SQDG or MGDGand PG. For all binary systems used in the presentstudy, the III/II values were in the range from 0.9 to0.96 in the presence of a total lipid concentrationof 11.6 mM (Table 2). These values indicate analmost complete solubilization of Ddx in thedifferent lipid systems at this lipid concentration.Increasing the total lipid concentration of the
0
0.0
0.2
0.4
0.6
0.8
1.0
100 MGDG85 MGDG/15 SQDG50 MGDG/50 SQDG15 MGDG/85 SQDG100 SQDG
DES
[Dtx
/(Ddx
+Dtx
)]
time [min]
0
0.0
0.2
0.4
0.6
0.8
1.0
100 MGDG85 MGDG/15 SQDG50 MGDG/50 SQDG15 MGDG/85 SQDG
III/II
val
ue [n
orm
.]
MGDG [µM]2 4 6 8 10 12 14 16 18 20
5 10 15 20
A
B
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
binary mixtures to 38.7 mM finally increased theIII/II values to 1. The increase in the lipidconcentration, however, did not significantlychange the Ddx de-epoxidation rates in thedifferent systems (data not shown).
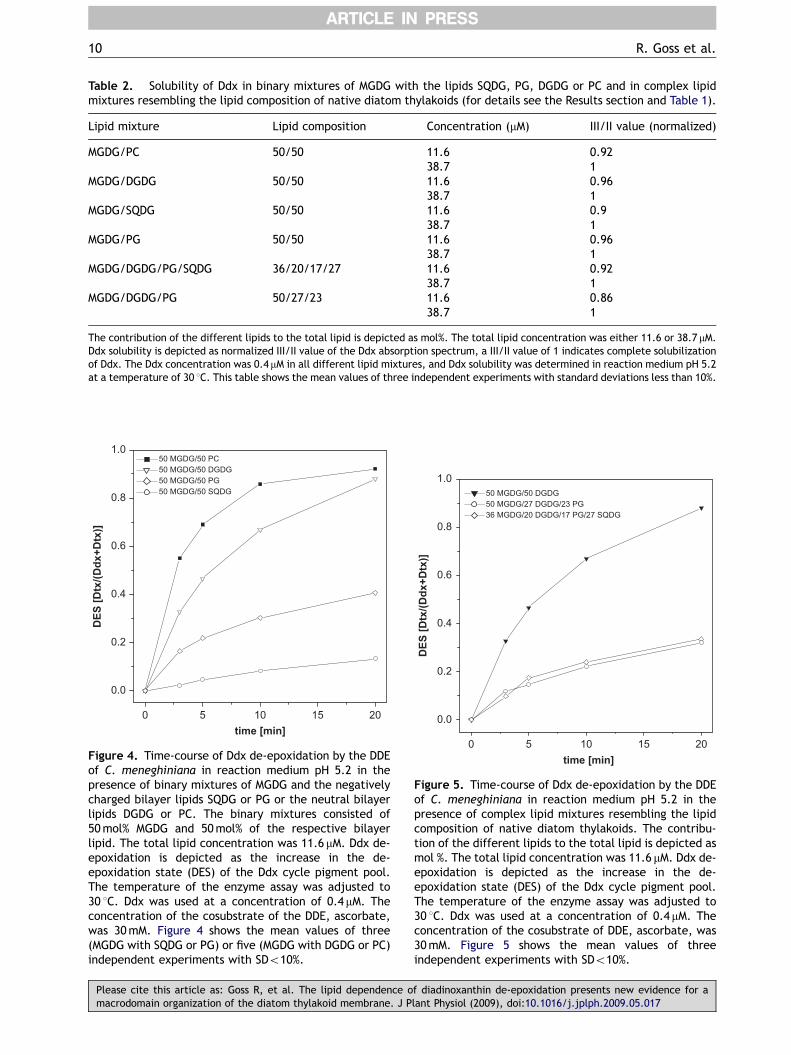
Solubility and de-epoxidation of Ddx incomplex lipid mixtures resembling the nativelipid composition of diatoms
In a complex lipid mixture composed of 36%MGDG, 20% DGDG, 27% SQDG and 17% PG, resem-bling the native lipid composition of diatomthylakoids, only slow Ddx de-epoxidation wasobserved (Figure 5). The de-epoxidation state ofthe Ddx cycle pool remained low at a value ofaround 0.25 after a reaction time of 20min in thislipid system with a total lipid concentration of11.6 mM. Another lipid system in which SQDG wasremoved but the concentration of the othernegatively charged lipid PG was increased, showedcomparably low de-epoxidation rates. In thesystem, which contained 50% MGDG, 27% DGDGand 23% PG, the de-epoxidation state of the Ddxcycle pool reached a similar value of around 0.25 atthe end of the enzyme assay. Only in lipid systems,which did not contain negatively charged lipids,i.e. in the binary mixture of 50% MGDG and 50%DGDG, a rapid and almost complete conversion ofDdx to Dtx could be observed. The complex lipidmixture containing MGDG, DGDG, SQDG and PGshowed an almost complete solubilization of Ddx ata total lipid concentration of 11.6 mM, as seen by
Figure 3. (A) Time-course of Ddx de-epoxidation by theDDE of C. meneghiniana in reaction medium pH 5.2 in thepresence of binary mixtures of MGDG and SQDG. Thecontribution of MGDG and SQDG to the total lipid isdepicted as mol %. The total lipid concentration was11.6 mM. Ddx de-epoxidation is depicted as the increasein the de-epoxidation state (DES) of the Ddx cyclepigment pool. The temperature of the enzyme assaywas adjusted to 30 1C. Ddx was used at a concentration of0.4 mM. The concentration of the cosubstrate of DDE,ascorbate, was 30mM. Figure 3A shows the mean valuesof three independent experiments with SDo10%. (B)Dependence of Ddx solubilization (depicted as normal-ized III/II value) on the MGDG concentration in binarymixtures of MGDG and SQDG. The Ddx concentration was0.4 mM, the temperature of the medium was adjusted to30 1C. For the measurements shown in Figure 3B the totallipid concentration was not held constant but increasedstepwise. The total lipid concentrations can be derivedfrom the depicted MGDG concentrations and the respec-tive ratios of the MGDG/SQDG mixtures. The differentmixtures of SQDG and PG are depicted as mol %.
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017
ARTICLE IN PRESS
Table 2. Solubility of Ddx in binary mixtures of MGDG with the lipids SQDG, PG, DGDG or PC and in complex lipidmixtures resembling the lipid composition of native diatom thylakoids (for details see the Results section and Table 1).
Lipid mixture Lipid composition Concentration (mM) III/II value (normalized)
MGDG/PC 50/50 11.6 0.9238.7 1
MGDG/DGDG 50/50 11.6 0.9638.7 1
MGDG/SQDG 50/50 11.6 0.938.7 1
MGDG/PG 50/50 11.6 0.9638.7 1
MGDG/DGDG/PG/SQDG 36/20/17/27 11.6 0.9238.7 1
MGDG/DGDG/PG 50/27/23 11.6 0.8638.7 1
The contribution of the different lipids to the total lipid is depicted as mol%. The total lipid concentration was either 11.6 or 38.7 mM.Ddx solubility is depicted as normalized III/II value of the Ddx absorption spectrum, a III/II value of 1 indicates complete solubilizationof Ddx. The Ddx concentration was 0.4mM in all different lipid mixtures, and Ddx solubility was determined in reaction medium pH 5.2at a temperature of 30 1C. This table shows the mean values of three independent experiments with standard deviations less than 10%.
0
0.0
0.2
0.4
0.6
0.8
1.050 MGDG/50 PC50 MGDG/50 DGDG50 MGDG/50 PG50 MGDG/50 SQDG
DES
[Dtx
/(Ddx
+Dtx
)]
time [min]5 10 15 20
Figure 4. Time-course of Ddx de-epoxidation by the DDEof C. meneghiniana in reaction medium pH 5.2 in thepresence of binary mixtures of MGDG and the negativelycharged bilayer lipids SQDG or PG or the neutral bilayerlipids DGDG or PC. The binary mixtures consisted of50mol% MGDG and 50mol% of the respective bilayerlipid. The total lipid concentration was 11.6 mM. Ddx de-epoxidation is depicted as the increase in the de-epoxidation state (DES) of the Ddx cycle pigment pool.The temperature of the enzyme assay was adjusted to30 1C. Ddx was used at a concentration of 0.4 mM. Theconcentration of the cosubstrate of the DDE, ascorbate,was 30mM. Figure 4 shows the mean values of three(MGDG with SQDG or PG) or five (MGDG with DGDG or PC)independent experiments with SDo10%.
0
0.0
0.2
0.4
0.6
0.8
1.050 MGDG/50 DGDG50 MGDG/27 DGDG/23 PG36 MGDG/20 DGDG/17 PG/27 SQDG
DES
[Dtx
/(Ddx
+Dtx
)]
time [min]5 10 15 20
Figure 5. Time-course of Ddx de-epoxidation by the DDEof C. meneghiniana in reaction medium pH 5.2 in thepresence of complex lipid mixtures resembling the lipidcomposition of native diatom thylakoids. The contribu-tion of the different lipids to the total lipid is depicted asmol %. The total lipid concentration was 11.6 mM. Ddx de-epoxidation is depicted as the increase in the de-epoxidation state (DES) of the Ddx cycle pigment pool.The temperature of the enzyme assay was adjusted to30 1C. Ddx was used at a concentration of 0.4 mM. Theconcentration of the cosubstrate of DDE, ascorbate, was30mM. Figure 5 shows the mean values of threeindependent experiments with SDo10%.
R. Goss et al.10
Please cite this article as: Goss R, et al. The lipid dependence of diadinoxanthin de-epoxidation presents new evidence for amacrodomain organization of the diatom thylakoid membrane. J Plant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
Diadinoxanthin de-epoxidation and lipid organization in diatoms 11
the III/II values of 0.92 (Table 2). Ddx solubility wasslightly lower in the lipid mixture containing MGDG,DGDG and PG (III/II value of 0.86), but was higher inthe binary system composed of MGDG and DGDG(III/II value 0.96). Increasing the lipid concentra-tion to 38.7 mM led to complete solubilization ofDdx and values of 1 were achieved. The almostcomplete solubilization of Ddx in the different lipidmixtures with a total lipid concentration of 11.6 mMsupports the idea that the slow Ddx de-epoxidationin the presence of SQDG and PG was not caused bylimitations in substrate availability. Furthermore,the slow Ddx de-epoxidation in the systemscontaining negatively charged lipids was not sig-nificantly enhanced in the presence of a total lipidconcentration of 38.7 mM (not shown).
Discussion
Lipid composition of diatom thylakoidmembranes
The data from the present study show thatisolated thylakoid membranes of the diatomC. meneghiniana are enriched in the negativelycharged lipid SQDG. SQDG, together with the othernegatively charged lipid PG, which is found in lowerconcentrations, amounts to more than 40% of thetotal lipid content. The concentration of SQDG, andnegatively charged lipids in general, is reduced ingreen algae and vascular plants, as shown by thecomparison with C. reinhardtii and spinach, whereSQDG and PG represent about 20% of the total lipidcontent. This is in line with other data on vascularplant thylakoid membranes that have shown thatthe charged lipids SQDG and PG contribute around15–20% to the total membrane lipids (Murata andSiegenthaler, 1998). It is also important that theincrease in SQDG in the diatom thylakoids isaccompanied by a concomitant decrease in thegalactolipids MGDG and DGDG, which are present insignificantly reduced amounts compared to thegreen alga and spinach. Taking into account that, inthe thylakoid membranes of the vascular plant,MGDG amounts to 45% of the total membrane lipid(see also Murata and Siegenthaler, 1998), thereduction in the MGDG content in diatoms becomeseven more obvious. MGDG and DGDG, whichnormally represent the most abundant thylakoidlipids, are both surpassed by SQDG in diatoms.
The lipid composition of the thylakoid mem-branes of C. meneghinina differed from the lipidcomposition of intact cells of the same algadetermined by Vieler et al. (2007). Both MGDG
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
and DGDG were found in reduced amounts in thethylakoid membranes in the present study com-pared to the galactolipid content of the cellsreported by Vieler et al. (2007). The PG concentra-tion was comparable to intact cells, whereas weobserved enrichment in the thylakoid membranesfor the phospholipid PC. SQDG was also found inhigher concentrations in the thylakoids whencompared with the SQDG content of the intactcell. The discrepancies between the lipid determi-nation of the present study and the previousexperiments (Vieler et al., 2007) are most likelycaused by the use of a new digital imaging systemand a different software algorithm for the lipidquantification. This assumption is corroborated bythe observation that the lipid analysis of intactcells with the new system (data not shown) shows ahigher similarity to the lipid composition of thethylakoid membranes of the present study than tothat of the intact cells determined by Vieler et al.(2007). We also observed that the light climateused for the growth of the algal cultures has astrong impact on the lipid composition of the cells.This means that differences in the growth lightconditions between the present experiments andthose in the study of Vieler et al. (2007) were atleast partially responsible for the differencesobserved in the lipid composition. Our finding thatthe MGDG and DGDG content of the diatomthylakoid membranes is lower than the content ofSQDG is, in our opinion, not caused by a contam-ination of the thylakoid lipids with lipids of othercell membranes. Although, due to the differentpreparation methods, the thylakoid membranes ofC. meneghiniana and C. reinhardtii are not as pureas those of spinach, contamination with mitochon-drial or plasma membranes would not haveaffected the MGDG/SQDG ratio, since these lipidsare only found in the lipid membranes of thechloroplast. Contamination with mitochondrial orplasma membranes, which are highly enriched inphospholipids (Joyard et al., 1998), would onlyhave resulted in a higher phospholipid content ofthe algal thylakoid preparations.
Apart from our knowledge about the structuralrole that SQDG and PG play in the oligomerizationof PSII, PSI and the LHCII (Liu et al., 2004; Jones,2007; Loll et al., 2007), the role of the negativelycharged lipids in membrane function is unclear atpresent. There is, however, evidence that theirpresence and concentration is essential for thethylakoid membrane. Under sulphate limitation,which strongly decreases the SQDG content of themembranes, there are complementary increases inPG, allowing the algae to keep the overall contentof negatively charged lipids at a constant level
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
R. Goss et al.12
(Sato et al., 2000). This observation indicates thatthe high concentration of SQDG in diatom thylakoidmembranes serves a specific, yet unknown, pur-pose. With regard to the possible role of SQDG andPG, it may prove useful to look at the function ofthe negatively charged lipid cardiolipin, which ishighly enriched in membranes designed to generatean electrochemical gradient for ATP synthesis otherthan the thylakoid membrane (for recent reviewssee Schlame et al., 2000; Joshi et al., 2009). Ineukaryotic cells, cardiolipin interacts with almostall of the protein complexes of the inner mitochon-drial membrane, including complex I, complex III,the cytochrome oxidase (complex IV) and the ATPsynthase (complex V). It has been suggested(Schlame et al., 2000) that cardiolipin is requiredfor maintaining the mitochondrial membrane po-tential, and that it couples the respiratory chainwith ATP synthase through an intramembraneproton transfer. It is of further interest that thenegatively charged PG can substitute, to someextent, for cardiolipin in keeping mitochondrialrespiration in a functional state. Our present datafurther show that the thylakoid membranes ofdiatoms contain significant concentrations of thephospholipid PC. The presence of PC in thethylakoids of C. meneghiniana does not necessarilyrepresent contamination by lipids of the chloro-plast envelope membrane (Williams, 1998), asearlier studies have shown that diatoms in generalcontain higher PC contents than vascular plants orgreen algae (Kates, 1987). PC was also found inpreparations of isolated fucoxanthin chlorophyllprotein complexes (FCPs) of C. meneghiniana andP. tricornutum (data not shown), a further indica-tion for the existence of significant amounts of PCin the native diatom thylakoid membrane.
Solubilization of Ddx by the negativelycharged lipids SQDG and PG
According to the present results, higher concen-trations of the negatively charged bilayer lipidsSQDG and PG are needed for the completesolubilization of Ddx compared with the invertedhexagonal structure forming lipid MGDG. On theother hand, previous results for PC and DGDG (Gosset al., 2005) indicate that, at lower lipid concen-trations, PG and especially SQDG exhibit higherefficiency for pigment solubilization than theuncharged or neutral bilayer lipids. However, asobserved for PC and DGDG (Goss et al., 2005),complete solubilization of Ddx in SQDG and PG isnot able to induce Ddx de-epoxidation. This showsthat another factor is needed for the de-epoxida-
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
tion reaction in addition to Ddx solubilization.According to our previous results, inverted hex-agonal phases (HII phases), which cannot beprovided by both the uncharged or the chargedbilayer lipids, are essential for efficient Ddx de-epoxidation.
Influence of the negatively charged lipidsSQDG and PG on Ddx de-epoxidation
Addition of the single lipids SQDG or PG to the invitro de-epoxidation assay leads to a completesuppression of Ddx de-epoxidation, as it observedpreviously for the uncharged bilayer lipid DGDG andthe neutral bilayer lipid PC (Goss et al., 2005). Thiscan easily be explained by the complete absence ofinverted hexagonal phases in the system usingsingle bilayer lipids. However, the situation is morecomplex in systems containing binary mixtures ofHII phase and bilayer-forming lipids. Here, ourpresent results show that binary lipid systemscomposed of MGDG and the uncharged bilayerlipids DGDG or PC are well suited for efficient Ddxde-epoxidation, given that equal amounts of HII
phase and bilayer-forming lipids are employed. Thisis consistent with data from earlier work (Gosset al., 2007). In binary systems, which contain MGDGand the negatively charged bilayer lipids SQDG orPG, the situation is completely different. Even atMGDG concentrations that allowed rapid de-epox-idation in the system with DGDG and PC, no or onlyvery slow Ddx de-epoxidation can be observed. Thealmost complete absence of de-epoxidation in thebinary systems in the presence of SQDG or PG is inline with the following assumptions: (i) It can behypothesized that the presence of the negativelycharged bilayer lipids inhibits the formation ofMGDG-induced inverted hexagonal phases in thebinary systems, which are needed for the binding ofthe DDE, thereby blocking the contact between theenzyme and its hydrophobic substrate, Ddx. Thiswould imply that SQDG and PG, due to theirnegative charge, have a strong capacity to stabilizea lipid membrane in the bilayer phase. That thepresence of DGDG and PC, on the other hand, doesnot seem to affect the MGDG HII phase may be dueto the absence of an overall negative charge. Thus,efficient Ddx de-epoxidation can be observed in abinary system consisting of uncharged or neutralbilayer lipids and HII phase forming lipids. (ii) Itcannot be excluded that, even in the presence ofSQDG and PG, an MGDG-induced HII phase isestablished. If this is the case, SQDG and PG mayalter the interaction of the DDE with the MGDGphase in such a way that efficient enzyme activity
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
Diadinoxanthin de-epoxidation and lipid organization in diatoms 13
cannot take place. (iii) SQDG and PG may furtheract as repulsion sites for the DDE due to theirnegative charges. For the VDE of higher plants, it isknown that the C-terminal region of the protein isenriched in negatively charged amino acids (for areview of the VDE structure see Hieber et al.,2000). A sequence comparison of the C-terminusfollowing the characteristic lipocalin motif of theVDE of Arabidopsis thaliana (Accession NumberQ39249) and the DDE of Thalassiosira pseudonana(Accession Number B8C775, UniProtKB, http://www.uniprot.org) revealed that the diatom DDEcontains a lower number of negatively chargedamino acids (1 aspartic acid and 20 glutamic acids)than the VDE of vascular plants (2 aspartic acidsand 28 glutamic acids). However, the C-terminus ofthe diatom DDE is still enriched in negativelycharged amino acids, making it likely that theDDE is repelled not only from the negativelycharged lipids, but from the lipid layer in general.This would suppress the interaction of the enzymewith its lipid-soluble substrate Ddx, thereby block-ing the de-epoxidation reaction. (iv) It is alsopossible that a stronger interaction of the chargedheadgroup of SQDG/PG with the ionon ring of Ddxleads to a significant deceleration of the Ddxdiffusion in the lipid phase. This would reduce theamount of Ddx at the peripheral DDE binding site,leading to a drastic reduction of Ddx de-epoxida-tion. (v) It must also be considered that negativelycharged lipids exhibit a stronger interaction in theheadgroup area in the presence of positivelycharged counter-ions (Cascales et al., 1995). Inour in vitro system, this may have led to a moretightly sealed membrane surface of the lipid layer,thereby depriving the DDE of insertion sites, whichare needed to access the hydrophobic substrateDdx. The importance of hydrophobic insertion sitesfor VDE/DDE activity has been proposed for boththe inverted hexagonal (HII) phase (Goss et al.,2007) and for a lipid bilayer in the liquid-crystalline(La) phase (Vieler et al., 2008). (vi) Furthermore, adirect inhibition of the DDE by the negativelycharged lipids must be considered. This assumptionis corroborated by reports that showed that a directinactivation of enzymes by SQDG or PG is possible.Isolated and purified chlorophyllase of the diatomP. tricornutum, which catalyzes the hydrolysis ofchlorophyll into chlorophyllide and phytol, hasbeen shown to associate with the negativelycharged SQDG and PG, leading to an inactivationof the enzyme in a cooperative way (Lambers andTerpstra, 1985). It is of special interest that, as inour present results, the uncharged bilayer lipidDGDG was unable to inhibit chlorophyllase activity.Like the DDE, ATPase activity of the CF0CF1
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
complex of the spinach thylakoid membrane isstimulated in the presence of MGDG, whereas thenegatively charged lipids SQDG and PG stronglyinhibit the hydrolysis of ATP (Pick et al., 1987). Incontrast to our present experiments, where evenlow concentrations of SQDG and PG in complex lipidmixtures suppressed Ddx conversion, optimal AT-Pase activity has been observed in the presence ofsmall amounts of SQDG and PG. An interactionbetween negatively charged lipids and specialprotein domains has been observed for the pre-cursor protein of plastidic ferredoxin (Horniaket al., 1993; van’t Hoff and de Kruijff, 1995). Thetransit sequence of precursor ferredoxin interactedstrongly with SQDG and PG, whereby its secondarystructure was changed. Addition of lipid vesiclescontaining negatively charged lipids led to a strongbinding of the transit peptide to the vesicles by aninsertion of the transit sequence into the hydro-phobic domain. Although MGDG was also able tobind the precursor protein, SQDG showed a stronglyenhanced capacity for protein attachment.
Separation of lipid domains in the nativediatom thylakoid membrane
Taking into account the strong suppression of Ddxde-epoxidation by the negatively charged lipidsSQDG and PG on the one hand, and the high SQDGconcentration in the thylakoid membranes on theother hand, a clear separation of the MGDG and theSQDG domains must exist in the native thylakoids.Otherwise, the highly efficient Ddx de-epoxidation,as is taking place in illuminated intact cells (Gosset al., 2006; Grouneva et al., 2008), would not bepossible. The present data from the in vitro lipidsystems consisting of binary mixtures of MGDG andSQDG or PG or complex lipid mixtures thatresemble the native lipid composition show thatseparation of the domains cannot be established inthe presence of lipids alone, as in these systemseven low concentrations of the negatively chargedlipids suppress Ddx de-epoxidation completely.Therefore, it is reasonable to believe that, in thenative thylakoid membrane, the separation ofMGDG and SQDG is realized by the proteincomplexes incorporated into the lipid membrane.Here, our first measurements show that the LHCs ofthe diatoms, the FCPs, are enriched in MGDG (datanot shown). This is in line with the observation thatthe FCPs represent the site where the majority ofthe Ddx cycle pigments are bound (Lavaud et al.,2003; Lepetit et al., 2007) and where the process ofnon-photochemical quenching of chlorophyll fluor-escence (NPQ) is taking place (Goss et al., 2006;
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
R. Goss et al.14
Grouneva et al., 2008). Based on the distributionand function of cardiolipin in the inner mitochon-drial membrane (Schlame et al., 2000), SQDG-enriched domains of the thylakoid membrane maythen represent the sites where the protein com-plexes of the electron transport chain are located.These domains may also represent the place wherethe photosynthetic electron transport is coupled toATP synthesis. Future measurements also mustshow whether the long chain fatty acids of diatoms(C22, C20), with their high degree of unsaturation,have a stronger capacity for lipid domain formationthan the commercially available lipids (with theirshorter fatty acids (C18, C16)) used in the in vitroassays in the present study. For this purpose nativediatom galactolipids and SQDG must be isolatedand, after determination of their fatty acidcomposition, tested in the respective Ddx de-epoxidation assays.
With regard to the existence of specialized lipiddomains, several attempts have been made toclarify whether lipid heterogeneities exist in thenative thylakoid membrane of vascular plants(reviewed by Siegenthaler, 1998). These studieshave shown that the acyl lipids may be asymme-trically distributed within the plane of the mem-brane, but that the lateral heterogeneity ofmembrane proteins is much more pronounced. Thismeans that the differentiation of the thylakoidmembrane of vascular plants into grana and stromamembranes depends on differences in the type andpercentage of lipids, but is modulated primarily bythe interplay of the respective photosyntheticprotein complexes with the lipids (Dekker andBoekema, 2005). As different lipids have differentstructural roles in the function and oligomerizationof photosynthetic protein complexes and theATPase (reviewed by Domonkos et al., 2008), thisinevitably also leads to heterogeneity of lipids inthe thylakoid membrane. Such requirements, how-ever, involve only a small number of particular lipidspecies and cannot account for the large scaleseparation of MGDG and SQDG that has to be calledfor, given the efficient Ddx de-epoxidation in nativediatom thylakoid membranes. Interestingly, it hasbeen shown that the outer layer of the thylakoidmembrane of vascular plants is rich in PG(Siegenthaler et al., 1989). Recent results have alsoindicated that changes in the PG content lead toperturbations of the surface charge of the thylakoidmembrane as well as alterations of the function ofthe photosynthetic complexes (Apostolova et al.,2008). With respect to the thylakoid membranes ofdiatoms, no experimental data exist showing thatthe different lipid species are heterogeneouslydistributed. Future measurements should show if
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
and how the uncharged, neutral, and negativelycharged lipids are separated and how efficient Ddxde-epoxidation is realized on a structural level.Taking into account that the diatom thylakoidmembrane does not show a differentiation intograna and stroma regions but, on the contrary,forms a regular arrangement of groups of three,makes the search for the possible mechanism oflipid domain formation in diatoms even moreinteresting. It will be of future interest to studyhow the extremely high concentration of negativelycharged lipids affects the organization and functionof the diatom thylakoid membrane, and if the highcontent of SQDG and PG must be seen in conjunc-tion with the different membrane protein andenzyme composition of the diatom chloroplast.Future experiments may also address the questionof whether SQDG and PG serve comparable func-tions in the thylakoid membrane of plants, asdescribed in detail for the negatively charged lipidcardiolipin in the inner membrane of the mitochon-drium.
Acknowledgements
Financial support from the Deutsche Forschungs-gemeinschaft (Go 818/6-1) is gratefully acknowl-edged.
References
Apostolova EL, Domonkos I, Dobrikova AG, Sallai A, BogosB, Wada H, et al. Effect of phosphatidylglyceroldepletion on the surface electric properties and thefluorescence emission of thylakoid membranes. JPhotochem Photobiol B: Biol 2008;91:51–7.
Cascales JJP, de la Torre JC, Marrink SJ, Berendsen HJC.Molecular dynamics simulation of a charged biologicalmembrane. J Chem Phys 1995;104:2713–20.
Dekker JP, Boekema EJ. Supramolecular organization ofthylakoid membrane proteins in green plants. BiochimBiophys Acta 2005;1706:12–39.
Domonkos I, Laczko-Dobos H, Gombos Z. Lipid-assistedprotein-protein interactions that support photosyn-thetic and other cellular activities. Prog Lipid Res2008;47:422–35.
Folch J, Lees M, Stanley GHS. A simple method for theisolation and purification of total lipids from animaltissues. J Biol Chem 1957;226:497–509.
Garab G, Lohner K, Laggner P, Farkas T. Self-regulation ofthe lipid content of membranes by non-bilayer lipids.Trends Plant Sci 2000;5:489–94.
Goss R, Lohr M, Latowski D, Grzyb J, Vieler A, Wilhelm C,et al. Role of hexagonal structure forming lipids in
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
Diadinoxanthin de-epoxidation and lipid organization in diatoms 15
diadinoxanthin and violaxanthin solubilization and de-epoxidation. Biochemistry 2005;44:4028–36.
Goss R, Pinto EA, Wilhelm C, Richter M. The importanceof a highly active and DpH-regulated diatoxanthinepoxidase for the regulation of the PS II antennafunction in diadinoxanthin cycle containing algae. JPlant Physiol 2006;163:1008–21.
Goss R, Latowski D, Grzyb J, Vieler A, Lohr M, Wilhelm C,et al. Lipid dependence of diadinoxanthin solubiliza-tion and de-epoxidation in artificial membrane sys-tems resembling the lipid composition of the naturalthylakoid membrane. Biochim Biophys Acta 2007;1768:67–75.
Goss R, Wilhelm C. Lipids in algae, lichens and mosses, In:Murata N, Wada H, editors. Lipids in photosynthesis:essential and regulatory functions. The Netherlands:Kluwer Academic Publishers; 2009, in press.
Gounaris K, Sen A, Brain APR, Quinn R, Williams WP. Theformation of non-bilayer structures in total polar lipidextracts of chloroplast membranes. Biochim BiophysActa 1983;728:129–39.
Grouneva I, Jakob T, Wilhelm C, Goss R. Influence ofascorbate and pH on the activity of the diatomxanthophyll cycle enzyme diadinoxanthin de-epoxi-dase. Physiol Plant 2006;126:205–11.
Grouneva I, Jakob T, Wilhelm C, Goss R. A new multi-component NPQ mechanism in the diatom Cyclotellameneghiniana. Plant Cell Physiol 2008;49:1217–25.
Guillard RRL, Lorenzen CJ. Yellow–green algae withchlorophyllide c. J Phycol 1972;8:10–4.
Haranczyk H, Strzalka K, Dietrich W, Blicharski JS. 31P-NMR observation of the temperature and glycerolinduced non-lamellar phase formation in wheatthylakoid membranes. J Biol Phys 1995;21:125–39.
Harwood JL. Membrane lipids in algae. In: Murata N,Siegenthaler P-A, editors. Lipids in photosynthesis.The Netherlands: Kluwer Academic Publishers; 1998.p. 53–64.
Hieber AD, Bugos RC, Yamamoto HY. Plant lipocalins:violaxanthin de-epoxidase and zeaxanthin epoxidase.Biochim Biophys Acta 2000;1482:84–91.
Horniak L, Pilon M, van’t Hoff M, de Kruijff B. Thesecondary structure of the ferredoxin transit sequenceis modulated by its interaction with negativelycharged lipids. FEBS Lett 1993;334:241–6.
Israelachvili JN, Mitchell DJ. A model for the packing oflipids in bilayer membranes. Biochim Biophys Acta1975;389:13–9.
Jakob T, Goss R, Wilhelm C. Unusual pH-dependence ofdiadinoxanthin de-epoxidase activation causes chlor-orespiratory induced accumulation of diatoxanthin inthe diatom Phaeodactylum tricornutum. J PlantPhysiol 2001;158:383–90.
Jensen RG, Bassham JA. Photosynthesis by isolatedchloroplasts. Proc Natl Acad Sci USA 1966;56:1095–101.
Jones MR. Lipids in photosynthetic reaction centres:structural roles and functional holes. Prog Lipid Res2007;46:56–87.
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
Joshi AS, Zhou J, Gohil VM, Chen S, Greenberg ML.Cellular functions of cardiolipin in yeast. BiochimBiophys Acta 2009;1793:212–8.
Joyard J, Marechal E, Miege C, Block MA, Dorne A-J,Douce R. Structure, distribution and biosynthesis ofglycerolipids from higher plant chloroplasts. In:Murata N, Siegenthaler P-A, editors. Lipids in photo-synthesis. The Netherlands: Kluwer Academic Publish-ers; 1998. p. 21–52.
Kates M. Lipids of diatoms and of halophilic Dunaliellaspecies. In: Stumpf PK, Mudd JD, Nes WD, editors. Themetabolism, structure and function of plant lipids.New York: Plenum; 1987. p. 613–21.
Lambers JWJ, Terpstra W. Inactivation of chlorophyllaseby negatively charged membrane lipids. BiochimBiophys Acta 1985;831:225–35.
Latowski D, Kruk J, Burda K, Skrzynecka-Jaskier M,Kostecka-Gugala A, Strzalka K. Kinetics of violaxanthinde-epoxidation by violaxanthin de-epoxidase, axanthophyll cycle enzyme, is regulated by membranefluidity in model lipid bilayers. Eur J Biochem2002;269:4656–65.
Lavaud J, Rousseau B, Etienne A-L. In diatoms, atransthylakoid proton gradient alone is not sufficientto induce a non-photochemical fluorescence quench-ing. FEBS Lett 2002;523:163–6.
Lavaud J, Rousseau B, Etienne A-L. Enrichment of thelight-harvesting complex in diadinoxanthin and im-plications for the nonphotochemical fluorescencequenching in diatoms. Biochemistry 2003;42:5802–8.
Lepetit B, Volke D, Szabo M, Hoffmann R, Garab G,Wilhelm C, et al. Spectroscopic and molecularcharacterization of the oligomeric antenna of thediatom Phaeodactylum tricornutum. Biochemistry2007;46:9813–22.
Liu Z, Yan H, Wang K, Kuang T, Zhang J, Gui L, et al.Crystal structure of spinach major light-harvestingcomplex at 2.72 (A resolution. Nature 2004;428:287–92.
Lohr M, Wilhelm C. Xanthophyll synthesis in diatoms:quantification of putative intermediates and compar-ison of pigment conversion kinetics with rate con-stants derived from a model. Planta 2001;212:382–91.
Loll B, Kern J, Saenger W, Zouni A, Biesiadka J. Lipids inphotosystem II: Interaction with protein and cofac-tors. Biochim Biophys Acta 2007;1767:509–19.
Mock T, Kroon BMA. Photosynthetic energy conversionunder extreme conditions. I. Important role of lipidsas structural modulators and energy sink under N-limited growth in Antarctic sea ice diatoms. Phyto-chemistry 2002;61:41–51.
Murata N, Siegenthaler P-A. Lipids in photosynthesis: anoverview. In: Murata N, Siegenthaler P-A, editors.Lipids in Photosynthesis. The Netherlands: KluwerAcademic Publishers; 1998. p. 1–20.
Pfundel E, Renganathan M, Gilmore AM, Yamamoto HY,Dilley RA. Intrathylakoid pH in isolated pea chloro-plasts as probed by violaxanthin de-epoxidation. PlantPhysiol 1994;106:1647–58.
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017

ARTICLE IN PRESS
R. Goss et al.16
Pick U, Weiss M, Gounaris K, Barber J. The role ofdifferent thylakoid glycolipids in the function ofreconstituted chloroplast ATP synthase. Biochim Bio-phys Acta 1987;891:28–39.
Sato N, Hagio M, Wada H, Tsuzuki M. Environmentaleffects on acidic lipids of thylakoid membranes. In:Harwood JL, Quinn PJ, editors. Recent advances in thebiochemistry of plant lipids. London: Portland PressLtd.; 2000. p. 912–4.
Schlame M, Rua D, Greenberg ML. The biosynthesis andfunctional role of cardiolipin. Prog Lipid Res 2000;39:257–88.
Shipley GG, Green JP, Nichols BW. The phase behaviour ofmonogalactosyl, digalactosyl and sulphoquinovosyldiglycerides. Biochim Biophys Acta 1973;311:531–44.
Siegenthaler P-A, Rawyler A, Smutny J. The phospholipidpopulation which sustains the uncoupled non-cyclicelectron flow activity is localized in the innermonolayer of the thylakoid membrane. BiochimBiophys Acta 1989;975:104–11.
Siegenthaler P-A. Molecular organization of acyl lipids inphotosynthetic membranes of higher plants. In:Murata N, Siegenthaler P-A, editors. Lipids in photo-synthesis. The Netherlands: Kluwer Academic Publish-ers; 1998. p. 119–44.
Stransky H, Hager A. Das Carotinoidmuster und dieVerbreitung des lichtinduzierten Xanthophyllzyklus inverschiedenen Algenklassen. Arch Mikrobiol 1970;73:315–23.
van’t Hoff M, de Kruijff B. Transit sequence-dependentbinding of the chloroplast precursor protein ferredox-
Please cite this article as: Goss R, et al. The lipid dependence omacrodomain organization of the diatom thylakoid membrane. J P
in to lipid vesicles and its implication for membranestability. FEBS Lett 1995;361:35–40.
Ventrella A, Catucci L, Mascolo G, Corcelli A, AgostianoA. Isolation and characterization of lipids associatedto PSII complexes: focus on cardiolipin structural andfunctional role. Biochim Biophys Acta 2007;1768:1620–7.
Vieler A, Wilhelm C, Goss R, Suß R, Schiller J. The lipidcomposition of the unicellular green alga Chlamydo-monas reinhardtii and the diatom Cyclotella mene-ghiniana investigated by MALDI-TOF MS and TLC. ChemPhys Lipids 2007;150:143–55.
Vieler A, Scheidt HA, Schmidt P, Montag C, Nowoisky JF,Lohr M, et al. The influence of phase transitions inphosphatidylethanolamine models on the activity ofviolaxanthin de-epoxidase. Biochim Biophys Acta2008;1778:1027–34.
White T, Bursten S, Frederighi D, Lewis RA, Nudelman E.High-resolution separation and quantification of neu-tral lipid and phospholipid species in mammalian cellsand sera by multi-one-dimensional thin-layer chroma-tography. Anal Biochem 1998;10:109–17.
Wilhelm C, Buchel C, Fisahn J, Goss R, Jakob T, LaRocheJ, et al. The regulation of carbon and nutrientassimilation in diatoms is significantly different fromgreen algae. Protist 2006;157:91–124.
Williams WP. The physical properties of thylakoidmembrane lipids and their relation to photosynthesis.In: Murata N, Siegenthaler P-A, editors. Lipids inphotosynthesis. The Netherlands: Kluwer AcademicPublishers; 1998. p. 103–18.
f diadinoxanthin de-epoxidation presents new evidence for alant Physiol (2009), doi:10.1016/j.jplph.2009.05.017




















