
the leaching behaviour of various zinc sulphide minerals with ...
347
THE LEACHING BEHAVIOUR OF VARIOUS ZINC SULPHIDE MINERALS WITH THREE THIOBACILLUS SPECIES THESIS SUBMITTED FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN THE SCHOOL OF BIOLOGICAL TECHNOLOGY UNIVERSITY OF NEW SOUTH WALES. BY AHMAD MUKHTAR KHALID JULY, 1978.
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of the leaching behaviour of various zinc sulphide minerals with ...

THE LEACHING BEHAVIOUR OF
VARIOUS ZINC SULPHIDE
MINERALS WITH THREE
THIOBACILLUS SPECIES
THESIS SUBMITTED FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
IN THE
SCHOOL OF BIOLOGICAL TECHNOLOGY
UNIVERSITY OF NEW SOUTH WALES.
BY
AHMAD MUKHTAR KHALID
JULY, 1978.

ACKNOWLEDGEMENT SUMMARY
1- INTRODUCTION
C O N T E N T S
1. 1- GENERAL INTRODUCTION
1.2- THE THIOBACILLI
1.2.1- Isolation and Taxonomical Characteristics
of Thiobacilli.
1.2.2- Isolation of Thiobacilli.
1
1
4
8
10
1.2.3- Taxonomy of Thiobacilli. 16
1.2.3.1- Morphology. 16
1.2.3.2- Biochemical Characteristics. 18
1.2.3.2- Submicroscopic Organisation of Thiobacilli. 27
1.3-BIODEGRADATION OF MINERALS BY MICROORGANISMS.36
1.3.1- Biodegradation of Minerals by
Heterotrophic Microorganisms. 36
Acidification or Alkalisation. 39
Chelation of Metal ions by Metabolites. 40
1.3.2- Biodegradtion of Minerals by Thiobacilli. 43
1.3.3- Factors Affecting the Microbial Leaching
Processes.
1.3.3.1- Nature of Substrate.
Metal distribution.
Properties of component minerals.
1.3.3.2- Effect of Physico-Chemical Factors.
Particle size.
Surface effects.
Temperature.
Redox potential and pH.
45
45
45
46
47
47
50
51
54

1.3.3.3- Nutritional Requirements of Microflora
Involved in Leaching.
Requirement for carbon.
Nitrogen source.
Phosphate.
Sulphate.
Trace elements.
Effect of dissolved ions on growth and
leaching behaviour of Thiobacilli.
Effect of organic compounds.
1.4-BIOGEOCHEMISTRY OF ZINC SULPHIDE.
1.4.1- Microbiological Oxidation of Zinc Sulphide.
1.4.2- Effect of Crystal and Lattice Structure on
Rates of Leaching of Zinc Sulphide.
1.5- AIMS OF THE CURRENT INVESTIGATION.
2- MATERIALS AND METHODS.
2.1- PREPARATION OF SULPHUR SUBSTRATES.
2.1.1- Polythionates.
2.1.2- Synthetic Zinc Sulphide.
2.1.3- Natural Zinc Sulphide Minerals.
2.1.4- Preparation of Zinc Sulphide Substrates.
2.2-ASSAY PROCEDURES.
2.2.1- Thiosulphate.
2.2.2- Polythionates.
2.2.3- Ferrous & Ferric Ions.
2.2.4- Emission Spectroscopic Analysis of Minerals.
2.2.5- Mineral Analysis by Atomic Absorption
Spectroscopy.
2.2.6- X-ray Diffraction Measurements.
57
57
61
62
63
64
65
70
75
79
85
87
88
88
88
89
89
90 90
92
92
94
94
96

2.3- ISOLATION OF THIOBACILLI.
2.3.1- Isolation of Non-Acidophilic Thiobacilli.
Source samples.
Media.
Isolation procedures.
2.3.2. Isolation of Acidophilic Thiobacilli.
Source samples.
Medium.
Isolation procedures.
2.3.3- Maintenance of cultures.
2.4-TAXONOMICAL INVESTIGATIONS.
2.4.1- Microscopic Examination.
2.4.2- Biochemical Tests.
Growth on nutrient agar.
Growth on different sulphur substrates.
Oxidation of ferrous ion.
Anaerobic growth.
Nitrate reduction.
Determination of DNA base composition.
Determination of FAME profile.
2.5-CULTIVATION OF THIOBACILLI.
2.5.1- Culture Vessels.
2.5.2- Preparation of Meida.
Batch culture.
Continuous culture.
2.5.3- Measurement of Dissolved oxygen.
2.5.4- Measurement of co2 in the Effluent Gas Stream.
2.5.5- Determination of Total Bacterial Population.
3- RESULTS AND DI SC USS I ON.
98
98
98
99
100
101
101
101
102
102
104
104
105
105
103
105
106
106
106
107
109
109
109
109
112
112
113
113
3.1- ISOLATION AND CHARACTERISATION OF TH10BACILLI.
3.1.1- Isolation of Non-:-,acidophilic sulphur oxidising
Bacteria. 115

3.1.2- Isolation of Acidophilic Iron-Oxidising Bacteria. 116
3.1.3- Isolation of Acidophilic Sulphur Oxidising
Bacteria. 117
3.1.4- Taxonomy of Isolates. 117
BJR-451. 117
BJR-Kl 128
BJR-K0l 133
3. 2- OPT IMI SATI ON OF I SO LA TES. 134
3.2.1- Selection of an Appropriate Growth Medium for
Non-acidophilic BJR-451. 135
3.2.2- Batch Growth of BJR-451. 137
3.2.3- Batch Growth of BJR-451 in Fermenter. 137
3.2.4- Determination of the Optimum pH for Growth of
BJR-451. 142
3.2.5- Determination of the Optimum Temperature for
the Growth of BJR-451. 14 7
3.2.6- Growth of BJR-451 in a Chemostat. 153
3.2.7- Effect of Carbon dioxide on Growth of BJR-451. 157
3.2.8- Toxicity of Zinc ions towards Thiobacillus
thioparus, Thiobacillus ferrooxidans & Thiobacillus
thiooxidans. 158.
3.3- LEACHING OF ZINC SULPHIDE BY THIOBACILLUS
FERROOXIDANS, THIOBACILLUS THIOOXIDANS AND
THIOBACILLUS THIOPARUS. 163
3.3.1- Leaching Capabilities of Thiobacillus thioparus,
BJR-451. 164
3.3.2- Leaching of Zinc Sulphides by Thiobacillus
ferrooxidans.
Leaching of marmatite.
Leaching of synthetic zinc sulphide.
Leaching of museum-grade zinc sulphide
minerals.
Discussion.
168
170
170
172
172

3.3.3- Leaching of Zinc Sulphides by Thiobacillus
thiooxidans.
Leaching of marmatite.
Leaching of synthetic zinc sulphide.
Leaching of museum grade zinc sulphide
minerals.
Discussion.
3.3.4- Leaching of Zinc Sulphide by Thiobacillus
176
176
178
178
178
thioparus. 182
Leaching of marmatite. 182
Leaching of synthetic zinc sulphide. 184
Leaching of natural zinc sulphide minerals. 184
Discussion. 184
3.3.5- Influence of Structure of Zinc Sulphide on
Microbial Leaching by Thiobacilli.
Comparative leaching of marmatite.
Comparative leaching of synthetic
zinc sulphide.
Comparative leaching of natural
188
189
192
zinc sulphide minerals. 194
Comparative leaching of natural wurtzite. 196
Discussion. 196.
3.3.6- Effect of Ferrous ion on Leaching of Synthetic
Zinc Sulphide by Thiobacillus ferrooxidans and
Thiobacillus thiooxidans.
Discussion.
4- C ONC LU S IONS
5- REFERENCES
215
218
223
229

6- APPENDICES
6.1- Isolation of Non-Acidophilic Sulphur
Oxidising Bacteria. (i)
6.2- Isolation of Acidophilic Iron Oxidising
Bacteria.
6.3- Isolation of Acidophilic Sulphur
Oxidising Bacteria.
6.4- Calculation of DNA base composition.
(xiv)
(xv)
(xvi)
6.5- Wave length, Slit Width, Lamp Current and
Gas Mixtures Used for Analysis by
Atomic Absorption Spectrophotometer. ( xxi) .
6.6- The Leaching Behaviour of Various Zinc
Sulphide Minerals with Three Thiobacillus
Species.
6.7- Microbial Ecology of Overburden Heaps
From Uranium Mining at Rum Jungle, N. T.
Australia.

A C K N O W L E D G E M E N T
I am greatly indebted to my supervisors, Professor
B. J. Ralph and Associate Professor Dr. P. A. D.
Rickard, for their continued guidance and criticism
throughout the course of this work. I am also
grateful to my consultant supervisors, Mr. B. Harris
(School of Metallurgy) and Professor R. Golding of
Physical Chemistry for their assistance and helpful
suggestions during experimentation.
I have benefited greatly from the advice, criticism,
and encouragement of my colleagues in the School of
Biological Technology.
I am also greatful to the Australian Development
Assistance Bureau of the Commonwealth of Australia,
for financial support.
I am thankful to my parents whose patience and
encouragement have always been a source of comfort.
Last, but not least, I wish to convey my sincere
gratitude and thanks to the Australian People, who
were very kind and friendly throughout my stay in
this country.

S U M M A R Y
Comparatively little data describing the role of the
physical and chemical characteristics of mineral substrates
in biodegradative processes is available. As a contribution
to this relatively unexplored area, the current investigation
was concerned with the characteristics of various forms of
zinc sulphide and their effects on biodegradation by three
different species of ThiobaciZZi namely, ThiobaciZZus ferro
oxidans ( BJR-Kl ), ThiobaciZZus thiooxidans ( BJR-K0l) & ThiobaciZZus thioparus ( BJR-451 ).
Isolates of the three species were purified and
characterised with respect to their principal biochemical and
physiological attributes. The ThiobaciZZus ferrooxidans strain
was isolated from a water sample from the Mount Lyell Mining and
Railway Company operating at Queenstown, Tasmania;it resembled
in most of its characteristics those ThiobaciZZus ferrooxidans
strains described in the literature.The ThiobaciZZus thiooxidans
strain, an isolate from' a sulphur heap at the Zinc Sulphide
Corporation, Boolaroo, N. S. W., was found to be closely related
to the type species previously described.
An isolate from road side near Tamworth, N. S. W., grew
at comparatively high pH values, ranging from 5.0 to 7.0. As
information regarding mineral leaching at pH values higher than
2.5 is sparse, this bacterium was considered suitable for study
ing microbial degradation of zinc sulphide at relatively high pH
values, i.e., those appropriate for this organism's prolifera
tion. It was extensively studied with respect to its biochemi-
cal and physiological characteristics and was found to be
identical to ThiobaciZZus thioparus. The optimum conditions
for its growth were precisely determined using a continuous
flow fermenter; the optimum pH and temperature for growth on
thiosulphate were found to be 5.5 and 30± 0.5° C respectively.

The ThiobaciZZus ferrooxidans strain was found to
tolerate zinc concentrations as high as 20,000 p.p.m.without any
significant deleterious effects on its growth rate being obser
ved. Concentrations of zinc higher than 15,000 p.p.m.were found
to inhibit the growth of the ThiobaciZZus thiooxidans strain.
The investigation involved the two crystallographic forms
of zinc sulphide, in the form of museum-grade specimens of
sphalerite and wurtzite. In addition, museum-grade marmatite
( a zinc iron sulphide Zn3.5FeS5) and synthetic zinc sulphide
were studied. The minerals were characterised by their X-ray
diffraction patterns and their purity was ascertained by
chemical analysis. Studies on their degradation by the three
pure cultures of ThiobaciZZi strains showed that the sphalerite
was degraded more rapidly and to a greater extent than was the
wurtzite: the latter could not be leached significantly by any
of the ThiobaciZZus species employed. The zinc sulphide with
iron substitution (marmatite) was the most amenable to bacterial
attack.
The ThiobaciZZus thioparus strain was the least active
in degrading any of the minerals and consequently showed the
lowest rates of metal release. The ThiobaciZZus thiooxidans
strain degraded synthetic zinc sulphide more easily than the
iron-oxidising ThiobaciZZus ferrooxidans, strain.
The results obtained indicated that:
1- The degradability of zinc sulphide minerals depends
upon the species of ThiobaciZZus used: usually it
is greater with the acidophilic members of the
genus. The rate and extent of metal release from
natural museum-grade mineral specimens decreased in
the following order :
T. ferrooxidans > T. thiooxidans > T. thioparus

and for the synthetic mineral the capabilities were
( in decreasing order) T. thiooxidans> T. ferroo
xidans > T. thioparus.
2- Ferrous iron concentrations higher than 100 p.p.m.
were found to inhibit the synthetic zinc sulphide
leaching by Thiobacillus thiooxidans strain.
3- The crytallographic form to which a substrate
belongs is of some importance. In the case of zinc
sulphides, the cubical form (sphalerite) was found
to be more readily leached than was the
form (wurtzite).
hexagonal
4- The release of zinc and iron from marmatite was
found to occur at different rates & the pattern
of release of these metals to be different in the
degradations catalysed by Thiobacillus thiooxidans
and Thiobacillus ferrooxidans. The implications
of these observations for the interpretation of
the release mechanisms is discussed.

'I INTRODUCTION

1
1.1 GENERAL INTRODUCTION
Winogradsky (1887), while working with Beggiatoa, envisaged an
ecological niche where microorganisms could procure all their energy
from the oxidation of inorganic compomds. In 1888, he isolated a
chemolithotrophic bacterium which could fulfil all its energy require
ments by the oxidation of exogenous iron salts. This discovery
introduced a second group of autotrophic organisms, the chemolithotrophs.
Until then the only autotrophs known were the photoautotrophs, viz.
plants and algae, characterised by their de nova synthesis of cellular
organic materials without requiring any preformed organic compomds.
As a result of the work of Winogradsky and others, it is now known that
both chemoautotrophs and phototrophs will grow on completely inorganic
media with carbon dioxide as the sole carbon source. Chemoautotrophs,
however, unlike phototrophs, obtain their energy for the synthesis of
cellular material, not from the electromagnetic radiations of visible
light, but from the oxidation of simple inorganic compounds. Thus, if
the chemoautotrophic bacterium is one of the nitrifiers, Nitrobaater or
Nitrosomonas, the inorganic compound would be either nitrite or annno
nilUil ion; if it were one of the non-photosynthetic sulphur bacteria,
the inorganic ion would be either sulphide, sulphur or thiosulphate;
hydrogen gas would be an equally good substrate for the growth of
Hydrogenomonas.
In Table 1.1 some chemoautotrophic bacteria, along with their
substrates, have been listed. It is evident from this table that
a common characteristic of these chemoautotrophic bacteria is the
oxidation of the exogenous inorganic substrates, regardless of their
chemical nature, which supplies the energy to the cellular machinery
for all synthetic activities including the assimilation of carbon

2
l11or~a11ic 1·11erl!,
, 11 I" trait·
II,
S"
F .. •
(:()
ll2 + ~02
TABLE 1.1
CHEMOLIIBOTROPHIC
Formal t'q11atio11 for rt'adio11
BACTERIA
Badni11111
---+;\;02 - + H20 + 2H· N11,·01orruHw,
,:l.(; 0 = -oo kcal
---+ :'\i O:, ,\' Urobnrtn
~(; 0 = -17 .. 5 kl'al
---+ H,<> H_wlrof!nwmona, ~( ;o = -.57 kl'al
S0 + ;o, + I 1,0 -----+H,SO. ThwhanJ/u., ~(; 0 = -118 kl'al
F,,2 --.F .. 3 ' +, T. ferrooxulans ~(; 0 = - 11 kcal
co+ (o, --+(:<>, Carho.,·wiomon"' ,l(;'' = -oo kl'al
Taken from: Doetsch & Cook- 1973. Introduction to Bacteria & Their Ecobiology.
Family
THIOBACTERIACEAE
BEOOIATOACl!AB
ACHROMATACEAE
TABLE 1. 2
THE LIIBOTROPHIC SULPHUR MICROORGANISMS
General characteristics Habitat
CHEMOLITHOTROPHS colourless, coccoid, straight or curved rod
shaped bacteria; polar flagellate when motile. Oxidi.te sulphur compounds and usually depo,it sulphur granules within or without the cells
colourless cells occurring in 1 rid1romcs within which they a,e arr.in!'cd in chains. The trichromcs shnw a gliding motion when in contact with a substrate. When grown in the presence or hydrogen sulphide the trichromes contain sulphur globules
large spherical, ovoid or short cylindric_al cells containing sulphur granules and sometimes inclusions of calcium carbonate
a wide variety of fresh-water and marine environments containing H,S. T/Jiobacil!us found in soil, mine waste-waters, sewage, industrial effluents
fresh water and marine environments containing H,S
fresh water and brackish mud containing H,S
Genera
Thiobacterium Macromonas Thio1•11/111n Thiospira Thiobacil/11s Beggiatoa Thiospiril/opsis Thioplaca Thiothrix
.Achromatium
Taken from: Roy & Trudinger - 1970. The Biochemistry of Inorganic Compounds of Sulphur.

3
dioxide.
Although a number of heterotrophs have also been found to oxidise
reduced sulphur compounds to sulphate in soils or other natural milieu,
the roles played by these transformations in the physiology of the micro
organisms are not yet fully understood. Nevertheless, most chemolitho
trophs of sulphur utilise the energy released by the oxidation of the
reduced compounds of sulphur for their synthetic activities. Table 1.2
lists the main groups of these chemolithotrophic bacteria of sulphur
and their main characteristics.
The not insignificant problems involved in the study of these
organisms are aggravated by the slow growth rates exhibited by some
of these genera. Therefore, for this reason very little information
pertaining to them is available at present. ThiobaciZZi, however, due
to their comparatively faster growth rates and the role played by them
in the mineral degradation and oxidation of various sulphur compounds,
have been far more extensively studied. They have been frequently
employed in various mineral mop-up operations for extracting metal
values from low grade ores and, due to their unique physiological
attributes, have found some novel applications, for example, the
removing of rust microbiologically (Iida et al., 1975) and the
recovery of hydrocarbons by the degradation of oil shales (Findley et
al., 1974; Meyer & Yen, 1976).
Since this study is mainly concerned with the degradative
activities of ThiobaciZZi on various zinc sulphide minerals, an
account of their principal characteristics follows.

4
1.2 THE THIOBACILLI
The credit for discovery of the ThiobaciZZi is attributed to
Nathansohn, who in 1902 succeeded in isolating small, Gram negative,
motile rods, which were capable of oxidising thiosulphate (Nathansohn,
1902). Since no carbon source was present in the medium, it was
correctly assumed that this bacterium could fix atmospheric carbon
dioxide, at the expense of energy obtained by the oxidation of thio
sulphate, thereby confirming the definition of a chemoautotroph, as
proposed by Winogradsky (1887). This bacterium was named ThiobaciZZus
thioparus a type species of the genus ThiobaciZZus.
ThiobaciZZi are Gram negative rods, generally motile, occurring
in various natural milieus, including mine waters, sewage water and
soils. Their common characteristic is the the capability of oxidising
elemental sulphur. They are usually aerobic, the exception being one
species, ThiobaciZZus denitrificans, which can proliferate anaerobically,
using nitrate instead of molecular oxygen as an electron acceptor.
The classification bases for the ThiobaciZZi have been bio
chemical and metabolic ones, like the oxidation of various compounds
of sulphur, thiocyanates, ferrous sulphate and the growth conditions
required. These bases of classification are fully debatable, particu
larly now that evidence for the existence of intermingled properties
in this genus has been amply provided. For example, when ThiobaciZZus
denitrificans, an anaerobe, is repeatedly plated in normal atmospheric
conditions, it loses its anaerobic trait and exhibits the characteris
tics of ThiobaciZZus thiopaPUs (Baalsrud & Baalsrud, 1954; Vishniac &
Santer, 1957). Similarly, ThiobaciZZus denitrificans & ThiobaciZZus
thiocyanoxidans were considered by Beijerinck (1904) and Happold et

5
al. (1954) to metabolise nitrate and thiocyanate respectively, and
were mainly distinguished from each other and from Thiobacillus thio
parus by these metabolic differences. With the accumulation of more
information, it has been noted that these metabolic characteristics are
commonly met in the genus Thiobacillus and therefore should no longer
be regarded as the basis of distinctionbetween the various species of
this genus ( Van der Walt & De Kruyff, 1955; De Kruyff et al., 1957;
Happold et al., 1958; Woolley et al., 1962).
Another interesting situation is the taxonomical position of the
bacteria oxidising ferrous ions. In the literature one generally comes
across three different names of microorganisms capable of oxidising
ferrous to ferric ions. These are Thiobacillus ferrooxidan,s, Ferro
bacillus ferrooxidans and Ferrobacillus sulfooxidan,s, which suggests
two different genera to be involved in this transformation. Leathen
et al. (1956) reported that the main difference between Thiobacillus
ferrooxidans and Ferrobacillus sulfooxidans was the capability of the
former to oxidise both sulphur as well as ferrous ions, whereas Fer
robooillus sulfooxidans could not oxidise sulphur at all when it was
present as the sole oxidisable substrate. However, Silverman & Lund
gren (1959-a) observed a slow but significant oxidation of elemental
sulphur by their strain of Ferrobacillus ferrooxidans, and Unz and
Lundgren (1961), studying the comparative nutritional patterns of
Ferrobooillus ferrooxidans, Thiobacillus ferrooxidan,s & Thiobacillus
thiooxidans, could not confirm Leathen's observations. Recently,
Silver (1970) has conclusively demonstrated that Ferrobacillus ferr
ooxidans can oxidise sulphur compounds concomitantly with fixing
carbon dioxide.
This leads to an important question, viz.,"Are the species of
ThiobaciUus distinct or simple variants of one another"? The use of

6
biochemical characteristics for the characterisation of the different
members of the
difficulties.
Thiobacillus group is not without some taxonomic
For example the variant components of some described
isolates in respect to the oxidation of iron, the adaptation of iso
lates to different substrates, the varying degrees of persistence to
the presence of organic matter and to heavy metals have led to some
taxonomic confusion. Some important points remain to be solved,
for example, the validity or otherwise of the novel species Thiobaci
llus acidophilus (Guay & Silver, 1975) with its capacity for the
oxidation of elemental sulphur and glucose but inability to oxidise
ferrous ion, even though de.rived by stepwise subcul turing from an
authentic Thiobacillus ferrooxida:ns strain. These continuing un
resolved problems are referred to in greater detail later in this
introduction.
I f
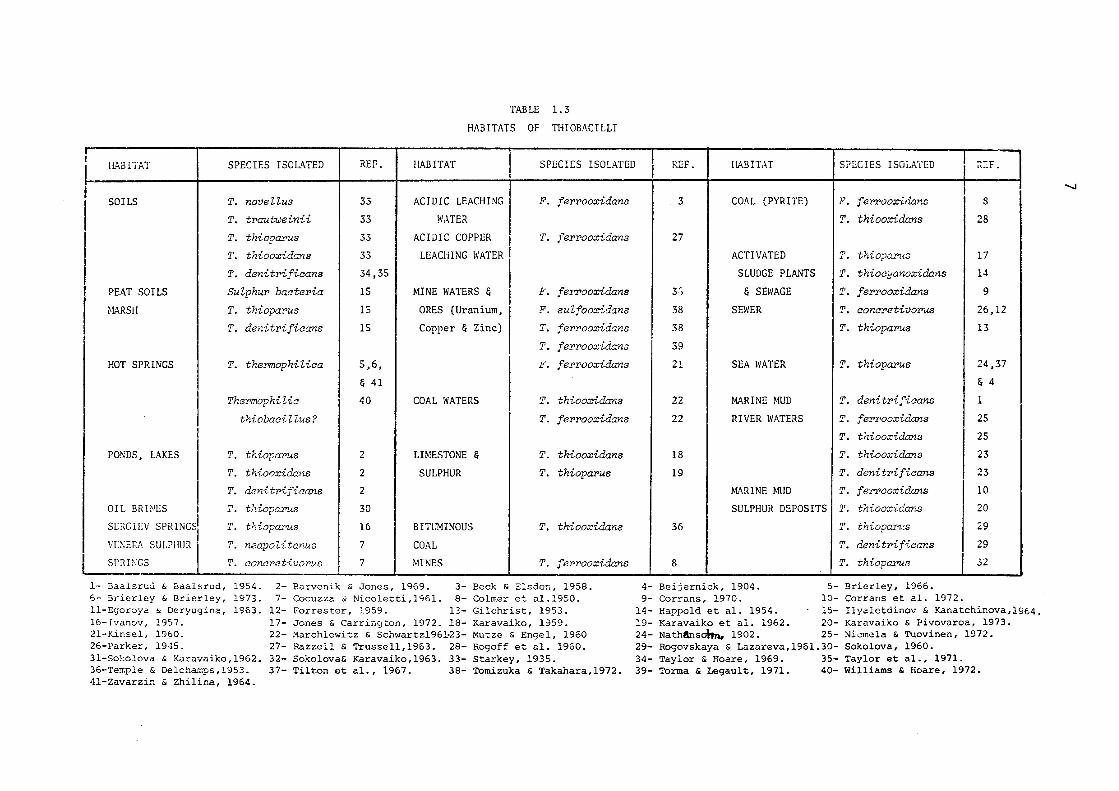
TABLE 1. 3
HABITATS OF T!-IIOBACILLI
HABITAT SPECIES ISOLATED REF. HABITAT SPECIES ISOLATED
~ SOILS T. novellus 33 ACIDIC LEACHING F. ferrooxidans
I T. traut-weinii 33 WATER
T. thiopar-us 33 ACIDIC COPPER T. ferrooxidans
T. thiooxidans 33 LEACHING WATER
T. denitrificans 34,35
PEAT SOILS Sulphur bacteria 15 MINE WATERS & F. f er'Y'ooxidans
MARSH T. thiopar-us 15 ORES (Uranium, F. sulfooxidans
T. denitrificans 15 Copper & Zinc) T. f errooxidans
T. f errooxidans
HOT SPRINGS T. thermophilica 5,6, F. ferrooxidans
& 41
ThermophiZic 40 COAL WATERS T. thiooxidans
thiobacilZus? T. f errooxidans
PONDS, LAKES T. thiopar-us 2 LIMESTONE & T. thiooxidans
T. thiooxidans 2 SULPHUR T. thiopar-us
T. denitrificans 2
OIL BRH!ES T. thiopo.rus 30
SERGIEV SPRINGS T. thiopar-us 16 BITUMINOUS T. thiooxidans
V[;-;ERA SULPHUR T. neapo Zi tanus 7 COAL SPRH;GS T. oonoretivorus 7 MINES T. ferrooxidans
1- Baalsrud & Baalsrud, 1954. 2- Barvenik & Jones, 1969. 3- Beck & Elsden, 1958. 6- Brierley & Brierley, 1973. 7- Cocuzza & Nicoletti,1961. 8- Colmer et al.1950. 11-Egoroya & Deryugina, 1963. 12- Forrester, 1959. 12- Gilchrist, 1953. 16-Ivanov, 1957. 17- Jones & Carrington, 1972. 18- Karavaiko, 1959. 21-Kinsel, 1960. 22- Marchlewitz & Schwartzl96~23- Mutze & Engel, 1960 26-Parker, 1945. 27- Razzell & Trussell,1963. 28- Rogoff et al, 1960. 31-Sol:olova & Karavaiko, 1962. 32- Sokolova& Karavaiko,1963. 33- Starkey, 1935. 36-Terr.ple & Delchamps, 1953. 37- Tilton et al., 1967. 38- •romizuka & Takahara,1972. 41-Zavarzin & Zhilina, 1964.
REF. HABITAT SPECIES ISOLA'fED ~!:F.
"'-J
3 COAL (PYRITE) F. fer'Y'ooxidans s T. thiooxidans 28
27
ACTIVATED T. thio?a:r>uB 17
SLUDGE PLANTS T. thiooyanoxidans 14
3r, & SEWAGE T. f errooxidans 9
38 SEWER T. concretivorus 26,12
38 T. thiopar-us 13
39
21 SEA WATER T. thiopar-us 24,37
& 4
22 MARINE MUD T. denitrificans 1
22 RIVER WATERS T. ferrooxidans 25
T. thiooxidans 25
18 T. thiooxidans 23
19 T. denitrificans 23
MARINE MUD T. feY'!'ooxidans 10
SULPHUR DEPOSITS T. thiooxidans 20
36 T. thiopar-us 29
T. denitrificans 29
8 T. thiopa:,'ur; 32
4- Beijernick, 1904. 5- Brierley, 1966. 9- Corrans, 1970. 10- Corrans et al. 1972.
14- Happold et al. 1954. 15- Ilyaletdinov & Kanatchinova,l964, 19- Karavaiko et al. 1962. 20- Karavaiko & Pivovaroa, 1973. 24- Nath6nsohn, 1902. 25- Niemela & Tuovinen, 1972. 29- Rogovskaya & Lazareva,1961.30- Sokolova, 1960. 34- Taylor & Hoare, 1969. 35- Taylor et al., 1971. 39- Torma & Legault, 1971. 40- Williams & Hoare, 1972.

3
1.2.1 ISOLATION AND TAXONOMICAL CHARACTERISTICS OF THIOBACILLI
Thiobcwilli are ubiquitous in nature, occurring in a wide
variety of natural environments including sewage water, hot springs
and soils. The acidophilic members of the genus have been detected in
acid mine waters, sewage holes, sulphur dumps and natural waters. A
large number of Thiobaeilli capable of growing at neutral pH have been
isolated from soils. In Table 1.3 the various habitats of Thiobaoilli
so far reported are summarised. The first detected member of the genus,
Thiobaoillus thioparus, was isolated from marine mud by Nathansohn
(1902) and has been frequently re-isolated from fresh waters and garden
soils. Starkey (1935) noticed that only 2 out of 29 soil samples, when
tested for the presence of the acidophilic Thiobaoillus thiooxidans,
showed positive results. However, he was able to isolate other species
of this genus from all these samples and concluded that the acidophilic
members occur sparsely in the soils. The isolation of Thiobaoillus
ferrooxidans, by Colmer et al. (1950), from bituminous coal mines.drainage
provided a ground to study the role of these microorganisms in mineral
leaching. It is evident from the Table 1.3 that most of the Thiobaoi-
llus ferrooxidans and Thiobaoillus thiooxidans strains have been iso
lated from the acidic waters of copper, uranium and bituminous coal
mines. The capability of these species to degrade sulphide minerals
and tolerate relatively high hydrogen ion concentrations suggests sul
phide ores as suitable preferred milieus for their proliferations.
Various workers have reported the isolation of Thiobaoillus ferrooxidans
and Thiobaoillus thiooxidans strains from the sulphide ores of uranium,
copper and nickel.
Thermophilic members of the genus Thiobaoillus have also been
isolated from hot springs containing sulphur by groups of scientists
working in Russia, United States of America & United Kingdom (Kaplan,
- I.O
'Tj -C") ;1~ 0 • ~ t--' (D
{ 1 THIOBACILLUS THIOOXIDANS
I THIOBACILUS FERROOXIDANS ] Ul
"O I-'• ::::: 5. 0 I THIOBACILLUS ACIDOPHILUS
·I -I-'· -0 () '~
I"'! Ill I -(D rt ~ '"O (D ;
~ I\) w ~ 0, 0) ....... 0) <O 0 I"'! ' (D '"O j::t> Ul ::i:: z (D '0
. ·o :I:
::, I"'! rt Ill :;i::,
::, ;:t> '"O IQ z ::i:: (D C")
,m 0 Hl
I THIOBACILLUS INTERMEDIUS I fTHIOBACILLUS DENITRIFICANS
'"O 0 'T1 rt I"'! 0 I-'· 8 IQ
:;i::, T. RUBELLUS Ill 11 C") . 0 :;i::, ....., ( 0
T. DELICATUS
rt ~ ::r :i Ill ::, 0 0, 'T1
Ul ~ 0 .... 8 I-'• 0, ~ .... ~ I-'• ::, t--< (D t--< Ul 1--.t
I THIOBACILLUS NEAPOLITANUS I I ] THIOBACILLUS NOVELLUS
- [ THIOBACILLUS PEROMETABOLIS I [ THIOBACILLUS THIOPARUS

J.O
1956; Schwartz & Schwartz, 1965; Williams & Hoare, 1972; Le Roux et al.
1977). Egorova & Deryugina (1963) were among the early investigators
to report the isolation from a hot spring of a spore forming, thermo
philic Thiobacillus, Thiobacillus thermophilica Imchenestskii, its
temperature optimum for growth being between 55° and 66°C. Recently
these thermophilic thionic bacteria have been studied for their role
in the leaching of sulphide minerals (Brierley & Murr, 1973).
1.2.2 ISOLATION OF THIOBACILLI
The Thiobacilli have been isolated from their environments by
enrichment techniques, employing suitable media and appropriate cultu
ral conditions.
The genus Thiobacillus includes a variety of microorganisms which
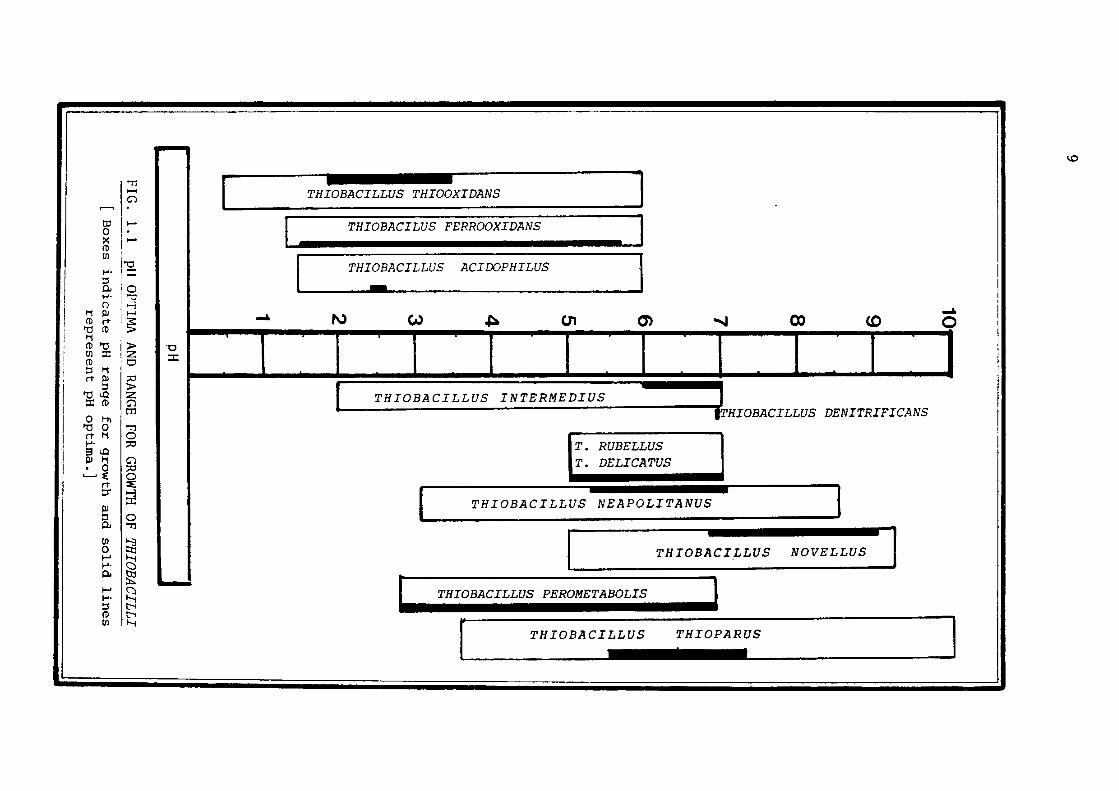
vary considerably in their pH optima for growth (Fig. 1.1). Thus,
ThiobaciUus thiooxidans & ThiobaciUus ferrooxidans grow luxuriantly
at pH 2.5, or even less, whereas Thiobacillus thiopa:r>us, Thiobacillus
denitrifioo.ns, and other species prefer a higher pH for their proli
feration. The Thiobacilli are pH-sensitive and are easily killed by
unfavourable pH environmental conditions. This characteristic of
these bacteria makes it very important that their growth media should
be properly buffered.
Vishniac & Santer (1957) in their review have listed a number of
media used for isolation of the various species of Thiobacilli and
Tables 1.4A & 1.4B describe the composition of the different media
usually employed for the isolation and growth of the individual species
of Thiobacilli.
Since these bacteria are autotrophic, bubbling Co2 through the
medium usually enhances the growth processes. Generally, K2HP04 and
KH2P04 , combined in buffers, control the pH and fulfil the phosphate
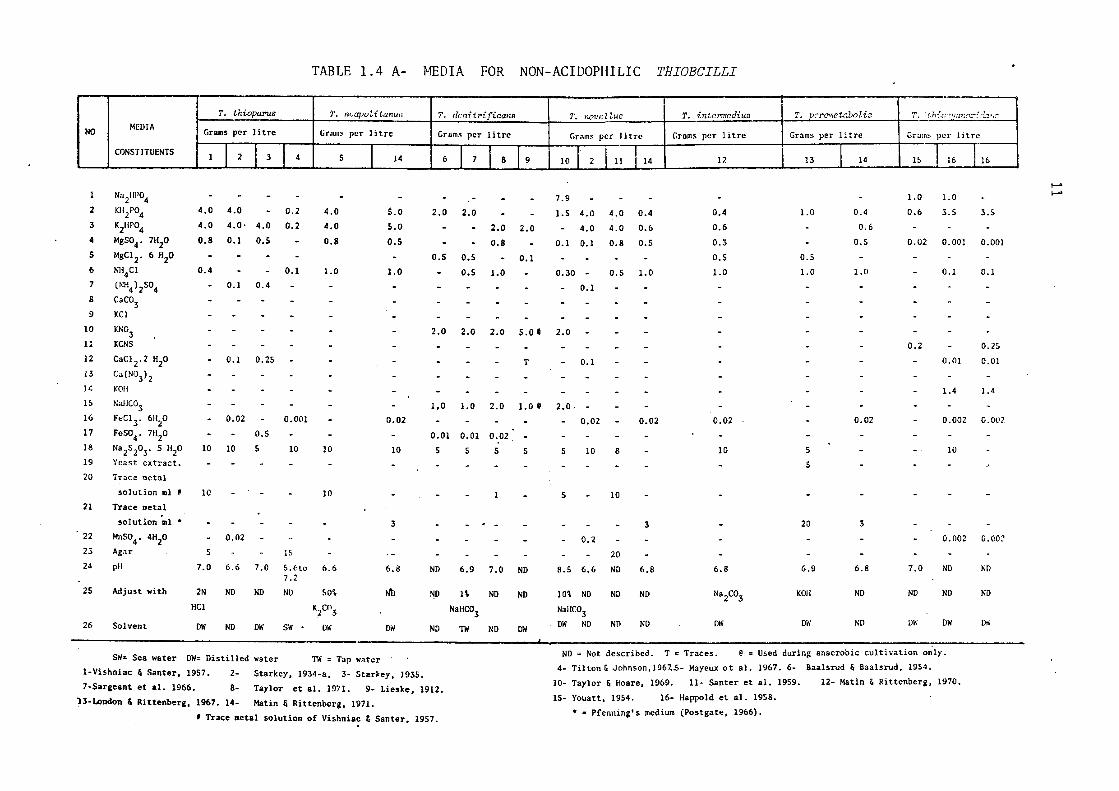
TABLE 1. 4 A- MEDIA FOR NON-ACIDOPHILIC THIOBCILLI
T. thioparus T. n,.apoZitanuo T. dc,dt.rificans 1', r.ovcUuo T. iritn,rtcdiuo T, DCT'C"netaboU/J T. ·thfo "!i1GWJ:rf_ d.:-:.,,r:
NO MEDIA Grams per litre Grams per litre Grams per litre Grams per Ji tn• Grnms per Ii trc Grams per litre Grains per litre
CONSTITU~NTS l I 2 l 3 I 4 s I 14 6 I 7 I I 9 10 I I 11
I 12 8 2 I 14 13 I 14 15 I 16 I 16
1--'
N,i/11'04 7.9 1.0 l. 0 I-'
2 KH2Po4 4.0 4.0 0.2 4.0 5,0 2,0 2,0 l.S 4,0 4.0 0.4 0.4 1.0 0.4 0.6 3.5 3.S 3 K2HP04 4.0 4.0 · 4.0 0.2 4.0 s.o 2.0 2.0 4.0 4.0 0.6 0.6 0.6 4 MgS04• 7H20 0.8 0.1 0.5 - 0.8 0.5 0.8 0.1 0.1 0.8 o.s 0.3 o.s 0.02 0,001 0.001 5 MgCJ 2• 6 H2o 0.5 0,5 0.1 0.5 o.s 6 Nli4CJ 0.4 0.1 1.0 1.0 o.s 1,0 0.30 0.5 1.0 1.0 1.0 1.0 0.1 C. I
7 (NH4)2S04 0.1 0.4 0.1 8 CaC03 9 KC!
10 KN03 2.0 2.0 2.0 5.0 I 2.0 11 KCNS - - 0.2 0. 25 12 CaCJ2.2 H2o 0.1 0.2S T - 0,1 - 0,01 0.01 13 Ca(NO3) 2 14 KOH - - 1.4 1.4
15 Nar1C03 - 1,0 1,0 2.0 1,0@ 2.0. 16 FeCJ 3. 6H20 0.02 0,001 0.02 - 0.02 0.02 0.02 0.02 0.002 0.002 17 FeS04. 7H20 - 0.5 - 0.01 0.01 0.02 18 Na2S203. 5 H2o 10 10 5 10 10 10 5 5 s s s 10 8 10 5 - 10
19 Ynast ext~act. 5 20 Trace metal
solution ml# 10 JO l 5 JO 21 Trace metal
solution ml• 3 3 20 3 22 MnS04• 4H20 0.02 0,2 0.002 o.oc~ 23 Agar s 15 20 24 pH 7.0 6.6 7.0 S.6to 6.6 6.8 ND 6.9 7.0 ND 8.5 6.6 ND 6.8 6.8 6.9 6.8 7.0 ND ND
7.2 25 Adjust with 2N ND ND ND SO% Nb 1-!D I\ ND ND 10\ ND ND ND Na2co3 KOH ND ND ND ND
HCl K2C03 NaHC03 NallC:03 26 Solvent ow ND DW DW ND ND ND ow DW ND DW DW D• SW OW DW ND TW ND DW
SW= Sea water DW= Distilled water ND= Not described. Tc Traces. @=Used during anaerobic cultivation only.
TW = Tap W!ltcr 4. Tilton & Johnson,1967 . .S- Mayeux et al. 1967. 6- Baalsrud & Baalsrud, 1954. 1-Vishniac & Santer, 1957, 2- Starkey, 1934-a, 3- Starkey, 1935.
JO- Taylor & Hoare, 1969. 11- Santer et al. 1959. 12- Matin & Rittcnberg, 1970. 7-Sargeant et al. 1966. 8- Taylor et al. 1971. 9- Lieske, 1912. 13-London & Rittenberg, 1967. 14- Matin & Rittenberg, 1971.
15- Youatt, 1954. 16- Happold et al. 1958.
f Trace metal solution of Vishniac & Santer, 1957. • • Pfenning's medium (Postgate, 1966).

12
demand at the same time. Ammonium salts provide the necessary nitrogen
for growth; either ammonium chloride or ammonium sulphate can be safely
used. ThiobaciZZus denitrificans, an anaerobic member of the genus,
requires nitrate as an electron acceptor. ThiobaciZZi can oxidise a
number of reduced sulphur compounds but sodium thiosulphate is most
commonly employed as the energy source.
Like all soil bacteria, they each have specific demands for the
trace minerals. For example, ThiobaciZZus thioparus has been shown
to require iron and manganese (Starkey, 1934) and Baalsrud & Baalsrud
(1954) reported augmentation in the growth rate of ThiobaciZZus
denitrificans in the presence of traces of ferrous ions. Neverthe~
less, a trace metal solution as described by Vishniac & Santer (1957)
meets the demands for trace elements for most species.
ThiobaciZZus thiocyanoxidans, which utilises CNS as a carbon and
sulphur source can also oxidise sodium thiosulphate in the presence
of ammonium ions plus carbon dioxide.
compounds, facultatively autotrophic
In addition to reduced sulphur
ThiobaciZZi can also obtain
their energy from organic compounds such as D-glucose, D-glactose,
sodium citrate, DL-aspartic acid and yeast extract (London, 1963;
London & Rittenberg, 1966; Guay & Silver, 1975; Mizoguchi et al.,
1976).
The acidophilic members of the genus do not require high concen
trations of phosphate and trace elements (Manning, 1975). They require
a low pH ( 2.0 to 2.5) and can oxidise either ferrous ions, elementary
sulphur or thiosulphate and other reduced sulphur compounds to obtain
their energy. Due to the lack of knowledge of appropriate growth media
and harvesting techniques, the work on the acidophilic ThiobaciZZi
was hindered until Silverman and Lundgren (1959) devised their, now
most widely used, medium known as '9K'. The composition of this medium
is shown in Table 1.4B.
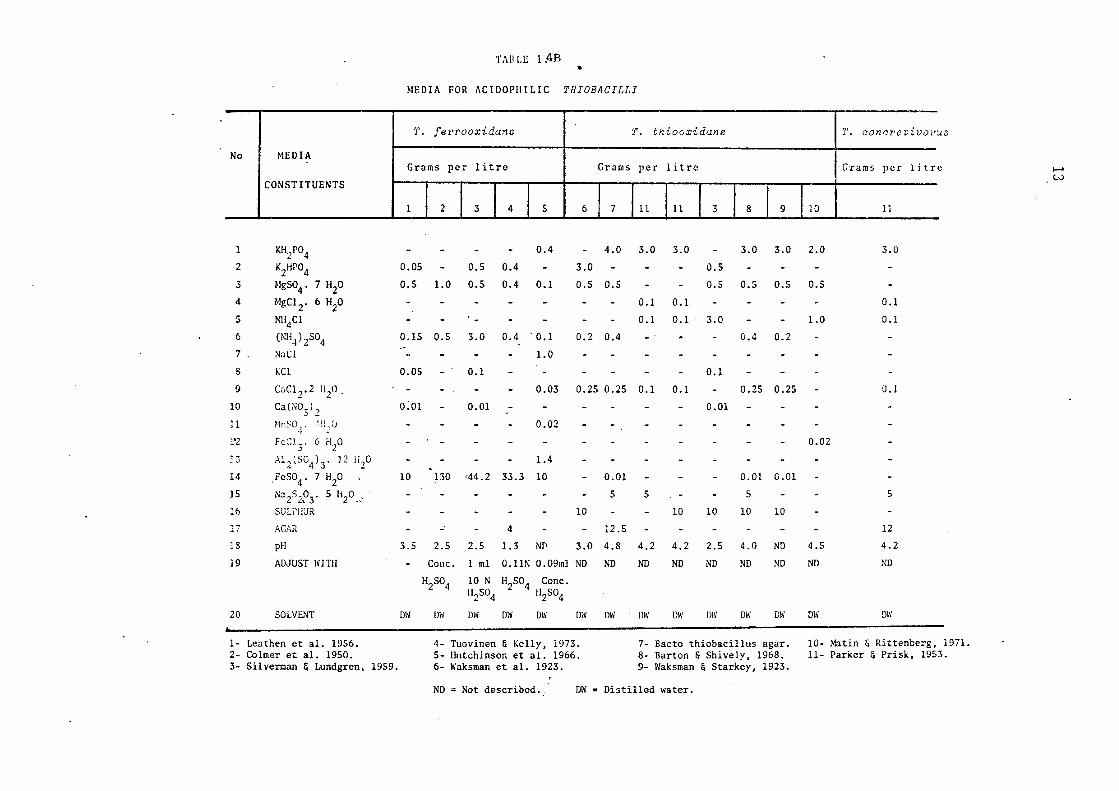
Ti\11 LE 1 :4B •
MEDIA FOR J\CJDOPIIILJC Tl!IOBACILLI
-T. f en•ooxidane I 'I'. thloc,xidane f T. conr:r•e t ivoi•u;,
No I MEDIA I Grams per litre I Grams per litre I Grams per litre ......
IT, (.-.)
CONSTITUENTS
I 3 I 4 I 5 I 6 I 7 111 111 I 3 I 8 I 9 1 10 I 11
l KH/04 - - - - 0.4 - 4.0 3.0 3.0 - 3.0 3.0 2.0 3.0
2 K2HP04 0.05 - o.s 0.4 - 3.0 - - 0.5
3 MgS04 . 7 H2o 0.5 1.0 0.5 0.4 0.1 0.5 0.5 - - 0.5 0.5 0.5 0.5
4 MgC1 2• 6 H2o - - - - - - 0.1 0.1 - - - - 0.1
5 NH4Cl - - - - - - - 0.1 0.1 3.0 - - 1.0 0.1
6 (NH4) 2so4 0.15 0.5 3.0 0.4 . 0.1 0.2 0.4 - - 0.4 0.2 .-7 NaCl - - - - 1.0
8 KCl 0.05 - 0.1 - - - - - - 0.1
9 CaCl 2" 2 1120 . - - - 0.03 0.25 0.25 0.1 0.1 - 0.25 0.25 0.1
10 Ca (NO.)? 0:01 - 0.01 - - - - - - 0.01 .) ~
11 MnS0_1• lll/l - - - 0.02
1'2 FeC]3. 6 H2o - - - - - - - - - - - 0.02
13 Al 2lSG4) 3. 12 Hz° - - - - 1.4
14 .FeS04 . 7 H2o 10 130 •44.2 33.3 10 - 0.01 - - - 0.01 0.01
15 Na 2s293. 5 H2o __ .· - - - - 5 5 - 5 - - 5
16 SULPHUR - - - - 10 - - 10 10 10 10
17 AGAR - _. - 4 - - 12.5 - - - - - - 12
18 pH 3.5 2.5 2.5 1.3 Nf\ 3.0 4.8 4.2 4.2 2.5 4.0 ND 4.5 4.2
19 ADJUST WITH - Cone. 1 ml 0. llN O .09ml ND ND ND ND ND ND ND ND ND
H2so4 10 N H2so4 Cone. Hz504 H2so4
20 SOLVENT ow DW DW DW ow ow DW DW ow ow DW ow ow DW
1- Leathen et al. 1956. 4- Tuovinen & Kelly, 1973. 7- Bacto thiobacillus agar. 10- Matin & Rittenberg, 1971. 2- Colmer et al. 1950. 5- Hutchinson et al. 1966. 8- Barton & Shively, 1968. 11- Parker & Prisk, 1953. 3- Silverman & Lundgren, 1959. 6- Waksman et al. 1923. 9- lfaksman & Starkey, 1923.
ND= Not described •. DW = Distilled water.

14
The purification of acidophilic members of the genus Thio"baciZZus
which are obligatelyautotrophic(ThiabaciZZus ferrooxida:ns, ThiobaciZZus
thiooxida:ns) on solid media is beset with problems. Agar cannot be
gelled properly because of its hydrolysis at the low pH required by the
organisms. A large number of investigators have tested and devised
modified agar media for the growth of Thio"baciZZus ferrooxidans and th~ir
reported L, inhibitory effects on the colony development (Bryner and
Jameson, 1958; Unz & Lundgren, 1961; Beck, 1967; McGoran et al., 1969;
Niemela & Tuovinen, 1972; Tuovinen et al., 1971-b). In attempts to
solve these problems workers (Leathen et al., 1951; Bryner & Jameson,
1958; Beck, 1960; Lapteva et al., 1971) have suggested the use of silica
gel for solidifying the medium. Gels prepared according to some pro
cedures widely in use at present are either too soft or too cloudy to
be really useful. A method claimed to be satisfactory for the prepara
tion of silica gel for growth of Nitrosomonas and Nitro"bacter species
has been described by Roslycky (1972). However, no attempts were made
to grow ThiobaciZZi on this medium. The absence of any suitable
method for colony development has made the conventional isolation of
low pH ThiobaciZZi from a single colony very difficult and has also
je.opardised performance of any classical genetic studies involving
these microorganisms. However, Tuovinen & Kelly (1973, 1974-c) have
reported a method for determining viable counts of Thio"baciZZus ferro-
oxida:ns using membrane filters placed on agar plates; specially
purified agar is used as the solidifying agent. According to these
workers, the purified agar does not inhibit the growth of Thio"baciZZus
ferrooxidans or ThiobaciZZus ne!X[JoZita,nus. That this technique suffers
from the drawback of lack of reproducibility is indicated from the
results of various workers (Unz & Lieberman,1973). This study showed
that the type of membrane filter as well as brand of agar used influ
enced the colony development of ThiobaciZZi. A low-ferrous, phosphate

15
free, modified form of 9K medium for isolating acidophilic iron-
oxidisers from acid mine drainage has been reported to give well deve
loped colonies on agar plates (Manning, 1975). No attempts to determ
ine the percentage recovery of ThiobaaiZZus ferrooxidans with the new
methods were made. However, with some modification, this technique
provides the means for the development of acidophilic cultures from
a single colony. Pure suspensions of ThiobaaiZZus fewooxidans have
been standardised by using a coulter counter (Shuler & Tsuchiya, 1975)
and Gormely & Dwcan (1974) developed a method for estimating bacterial
populations in association with mineral particles by determining the
nitrogen contents of such mixtures.
The purification of non-acidophilic ThiobaaiZZi is not so
difficult as that of acidophilic ThiobaaiZZi, though it does involve
some peculiar problems, including inhibition of colony development
on agar media due to the toxicity of agar and other associated material.
They will grow on solid media prepared from pure grade agar using
some special techniques. On solid plates these bacteria (Thiobaai
ZZus thioparus and others) need frequent transfer because of their
sensitivity to metabolically generated acid. They are best maintained
in liquid culture.

16
1.2.3 TAXONOMY OF THIOBACILLI
The taxonomy of ThiobaciZZi has been most confusing and
challenging since the beginning. As mentioned earlier, the basic
classification rests upon biochemical reactions, which in turn depend
upon various other factors. In the following, an endeavour has been
made to summarise the different traits of these microorganisms.
1.2.3.1 MORPHOLOGY
It has been generally accepted that all ThiobaciZZi are Gram
negative, motile rods of diameter varying from 0.3 ~ 0.4 µm, and
about 1 ~ 3 µm long. They are easily stained with Gram stain and a
few (unconfirmed) reports exist which describe them as Gram-positive
rods (Starkey, 1935). The mode of motility of these bacteria indicates.
the presence of a single polar flagellum, which is very fragile and
frequently cannot be observed by ordinary optical microscopy employ-
ing conventional staining techniques. The flagellum is usually lost
during preparatory procedures. However, electron micrographs have
clearly shown the presence of one polar flagellum. The only non
motile strain described is ThiobaciZZus noveZZus. The non-motility of
ThiobaciZZi depends upon a number of factors, viz. acidic environ
ments or the exhaustion of substrate. The reason may be the lack of
energy under the 1.lllfavourable conditions, when cells are 1.lllable to
spend their energy for motility.
With the exception of a strain ThiobaciZZus thermophiZica
Imschenetskii nov. sp., isolated by Egorova & Deryugina (1963), all
ThiobaciZZi are non-sporulating. The spores were terminal and able
to withstand high temperatures. However, the position of this
microorganism in the genus ThiobaciZZus has been seriously questioned
by some workers (Hutchinson et al., 1967).
SPECIES
T. cur.dophil~a (II
T. delicatus 121
. T. dsnit"rificans
T. fenooridane
I. ntrJeZZ.ue
T. r.eapolitanua
.. "'. :nter1r,edi~UJ
*
.
,,. p€.rametat:olis
T. :rubellus <2>
t'r.WCxidans .
T. thioparu11 .
.
SHAPE
Short rods
Short rods
Short rods
Short. rods with rounded edges.
Short rods, coccoidal or ellipsoidal cells
Short rods
Thin short rods.
Thin short rods
Single rods.
Short rods
Thin
SIZE µm
0,5 - 0.8 X
1.0 - 1.5
0.45-0.50 X
0.7 -1.3
0.5 X
1.0 - 3.0
o.sx 1.0
0,4 - 1.0 X
0,6 - 4.0
0,5 • 1.0-1.5
0.5 l< LO· 2.0
0.5 X
1.0 - 2.0
1.0 - 1.6 x 1.7 - 3,0
0.5 X
1.0 - 2.0
0.5•1. 7
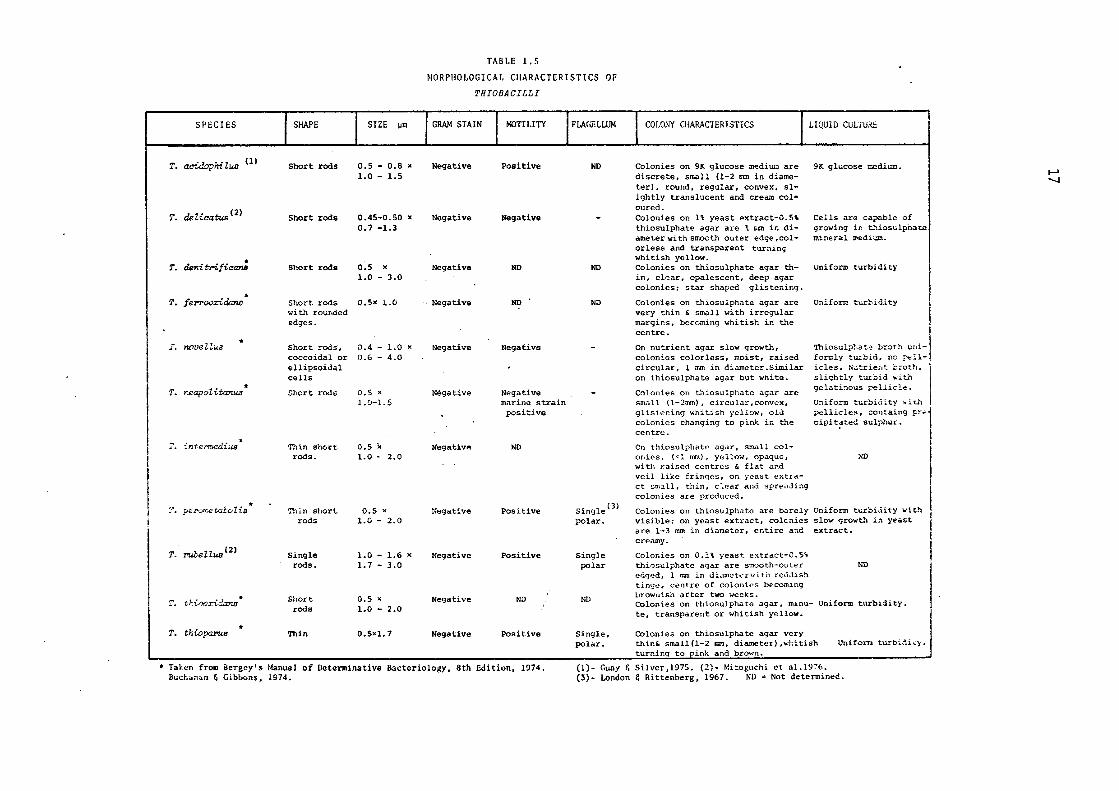
TABLE 1.5
MORPHOLOGICAL CHARACTERISTICS OF
THIOBACILLI
GRAM STAIN
Negative
Negative
Negative
Negative
Negative
Negative
NegativA
Negative
Negative
Negative
Negative
MOTILITY
Positive
Negative
ND
ND
Negative
Negative marine strain positive
ND
Positive
Positive
ND
Positive
FLAGELLUM
ND
ND
ND
Single (J)
polar.
Single polar
ND
Single, polar.
COLONY CHARAClERISTICS
Colonies on 9K glucose mediUll are discrete, small (1-2 mm in diameter), round, regular, convex, slightly translucent and cream coloured. Colonies on li yeast extract-0,5~ thiosulphate agar are 1 ncn ir. diameter with smoot..11 outer edge ,colorless and transparent· turning whitish yellow. Colonies on thiosulphate agar thin, clear, opalescent, deep agar colonies; star shaped glistening.
Colonies on thiosulphate agar are very thin & small with irregular margins, becoming whitish in the centre.
On nutrient agar slow growth, colonies colorless, moist, raised circular, 1 mm in diameter.Similar on thiosulphate agar but white.
Colonies on t~iosulphate agar are small {l-2rmn), circular ,convex, glistening whitish yellow, old colonies changing to pin~ in the centre.
On thiosulphate ngdr, small colonies, (<l mm), yellow, opaque, with raised centres & flat c'.}.nd veil like fringes, on yeast extra-ct small, thin, clear and spreading colonies are product1d.
LIQUID CULTUi\E
9K gh,cose :::edium.
Cells are capable of growing in thiosulphace mineral :l\E:diw:i..
Uniform turbidity
Unifurm turbid.ity
Th iasulpha t:e brot..li. wiif om.ly turbid, no f't,:.llicles. N;.itrier,t broth, slightly turbid ••i th gelati:1ous pellicle.
Uniform turbidity \\·i 't.h
pellicle~, containg pre cipit~ted sulphur.
)10
Colonies on lhiosulphate are barely Uniform turbidity wit..--i visible; on yeast extract, colcnies slow growth in yeast a"L"e 1·-3 mm in diameter, entire and extract. creamy.
Colonies on 0.1% yeast extract-0.5~ thiosulphate agar are smooth-outer edged, 1 mm in dL:unctt.."'rwiti1 reddish tinge, centre of colonies becoming brownish at'ter two weeks.
ND
Colonies on thiosulphatc agar, minu- Uniform turbidity. te, transparent or whitish yellow.
colonies on thiosulphate agar very thin& small(l-2 mm, diameter) ,whitish turnin~ to pink and brown.
Uniforn turbidic).
• Talen from Bergcy's Manual of Determinative Bacteriology, 8th Edition, 1974, Buchon~n & GibLons, 1974,
(1)- Cuay & Silvcr,1975. (2)- Mi:oguchi et ol.1976. (3)- London & Rittenberg, 1967. ND= Not determined.
I--' -..J

18
Cells occur singly or in pairs, and sometimes chains of cells
have been observed. Silverman & Rogoff (1961) reported alteration of
cellular morphology of ThiobaaiZZus ferrooxidans in response to exce
ssively vigorous aeration. According to these authors, the normal
cellular rods were transformed into a coccoidal form with a diameter
of 1.0 µm, when saturated with air and strongly agitated. These cells
regained the normal morphology when grown en masse or with less vigo
rous aeration. Cells grown in glucose have been reported to become
slightly rounder, with a tendency to form pairs (Lundgren et al., 1964).
ThiobaoiZZus ferrooxidans and ThiobaoiZZus thiooxidans were found
to possess two mechanisms of reproduction, by fission and by partition,
the former being the most common mode of cell division (Karavaiko &
Avakyan, 1970; 1971).
Table 1.5 summarises the morphological characteristics of the
genus ThiobaoiZZus as described in the latest edition of the Bergey's
Manual of Determinative Bacteriology (Buchanan & Gibbons,1974).
1.2.3.2 BIOCHEMICAL CHARACTF:R.ISTICS
Parker & Prisk (1953) carried out a comprehensive study of
ThiobaoiZZi whilst they were examining the utilisation of various red
uced compounds of sulphur and classified them into different species.
They found that thiosulphate was oxidised by all ThiobaoiZZi and Thio
baoiZZus thiooxidans, ThiobaoiZZus oonoPetivorus and ThiobaoiZZus X
converted it to tetrathionate and sulphite and then oxidised it
further to sulphate and free sulphuric acid. ThiobaoiZZus thiopaPUs
was found to convert thiosulphate to sulphite and sulphur followed
by partial oxidation of sulphur to sulphuric acid. Elementary sulphur
was found to be oxidised by ThiobaoiZZus thiooxidans, ThiobaoiZZus
oonoPetivorus, ThiobaoiZZus X, and ThiobaoiZZus thiopaPus; the rate of
oxidation was in that order. Hydrogen sulphide was oxidised by
THE GE~US THIOBACILLUS
~ OBLIGATELY AUTOTROPHIC FACULTATIVELY AUTOTROPHIC
AEROBIC
T. THIOPARUS
T. NEAPOLITANUS
T. FERROOXIDANS
T. THIOOXIDANS
FACULTATIVELY ANAEROBIC
T. DENITRIFICANS
T. ACIDOPHILUS
T. DELICATUS
T. NOVELLUS
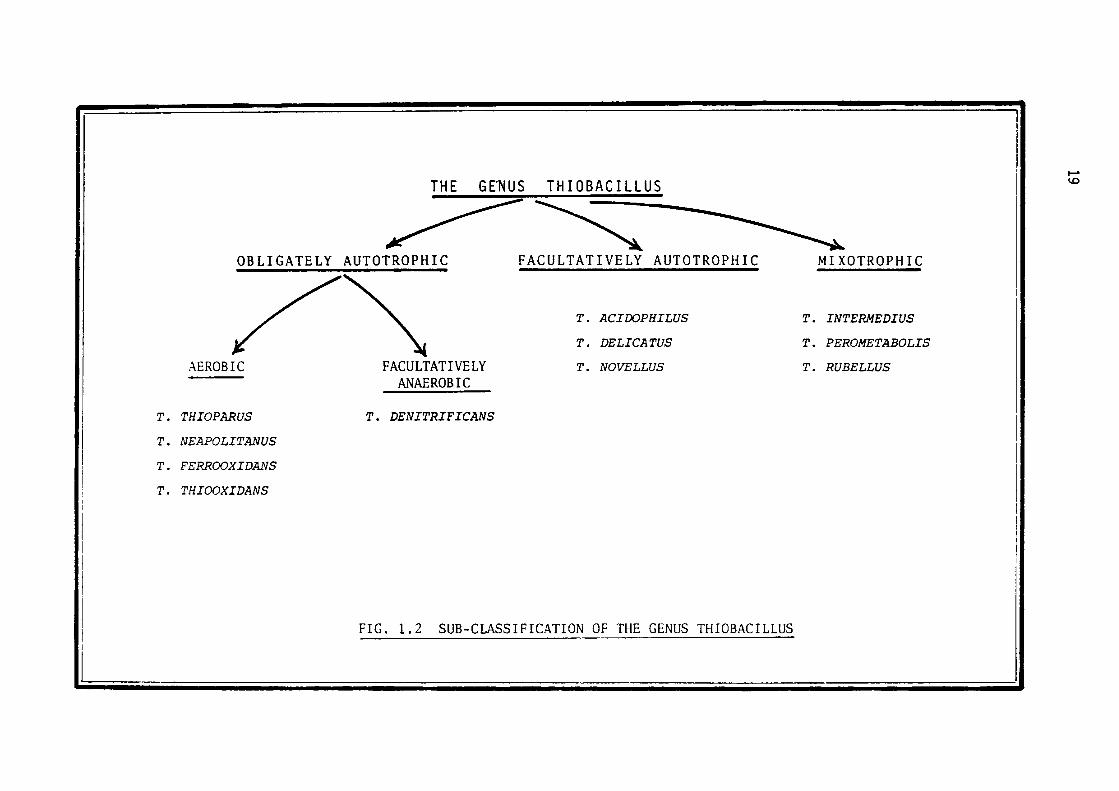
FIG. 1.2 SUB-CLASSIFICATION OF THE GENUS THIOBACILLUS
MIXOTROPHIC
T. INTERMEDIUS
T. PEROMETABOLIS
T. RUBELLUS
...... I.O

20
Thiobacillus concretivorus (Thiobacillus thiooxidans) and Thiobacillus
X. Therefore, in the early studies the basic differentiating charact
eristics serving to distinguish between the different forms of Thiobacilli
were considered to be the capacity of utilising certain specified
reduced compounds of sulphur as the substrate and the production of
certain metabolites. Both of these criteria have many disadvantages
and, indeed, have caused a good deal of confusion in the taxonomical
studies of the Thiobacilli. For example, in a number of recent inves
tigations it has been amply demonstrated that the utilisation of
various sulphur compounds and the formation of products other than
sulphate are dependent on such conditions of growth as pH, oxygen
transfer, substrate level and so on. Therefore, these biochemical
characteristics can no longer be regarded as definitive diagnostic
tests for taxonomic purposes.
Earlier, microorganisms capable of oxidising or otherwise meta
bolising thiosulphate were catalogued as Thiobacillus species; for
example, Thiobacillus coproliticus and Thiobacillus trau-tweinii
(Lipman & Mcless, 1940; Bergey et al., 1925). Those heterotrophs, now
known to possess the ability to oxidise thiosulphate without obtaining
energy from the process (Guittoneau, 1925; Starkey, 1935), are no
longer considered to be members of the genus Thiobacillus (Vainshtein,
1975). Those species which obtain energy from the oxidation of thios
ulphate can be divided into three subgroups, namely obligately auto
trophic, facultatively autotrophic and mixotrophic (Fig. 1.2). The
later subgroup includes heterotrophic microorganisms whichutilise compounds
sulphur and organiclsimultaneously (Rittenberg, 1969). The obligate
autotrophs are further divided into aerobic and facultatively anaero
bic sub-sub-groups. The aerobic sub-sub-group is comprised of
(a) species which are capable of oxidising only sulphur compounds in
order to fulfil their energy requirements (Thiobacillus thiooxidans~
I • OPTH~: SPECIES I OPTIMUM pH-
TEMPERATURE I
T. acidophilua<ll 3.0 25 - 30 [LS - 6.0] .
T. denitrificana 7.0 30
I'. d.alwatus(Z) 5.0 - 7.0 30
* 6.0 - 7.0# T. i nt,ermediua 30
[1.9 - 7.0]
* T. f en'ac:::id.ans 2.5 - 5.8 28 - 35
[J.4 - 6.0]
. 1'. ri.eapo'Zit,-;r..us 6.2 - 7.0 28
[3.0 - 8.5)
. . T. nct;,eZZus 7.8 - 9.0 30
[5.0 - 9.2]
* T. perometabolis 2.6 - 6.8 30
'
'I'. roeZZus (2) s.o - 7.0 30
. T. thioo::.damJ 2.0 - 3.5 28 - 30
[0.5 - 6.0]
* T. th{..oparus 6.6 -7.2 28 - 30
[up to ID J
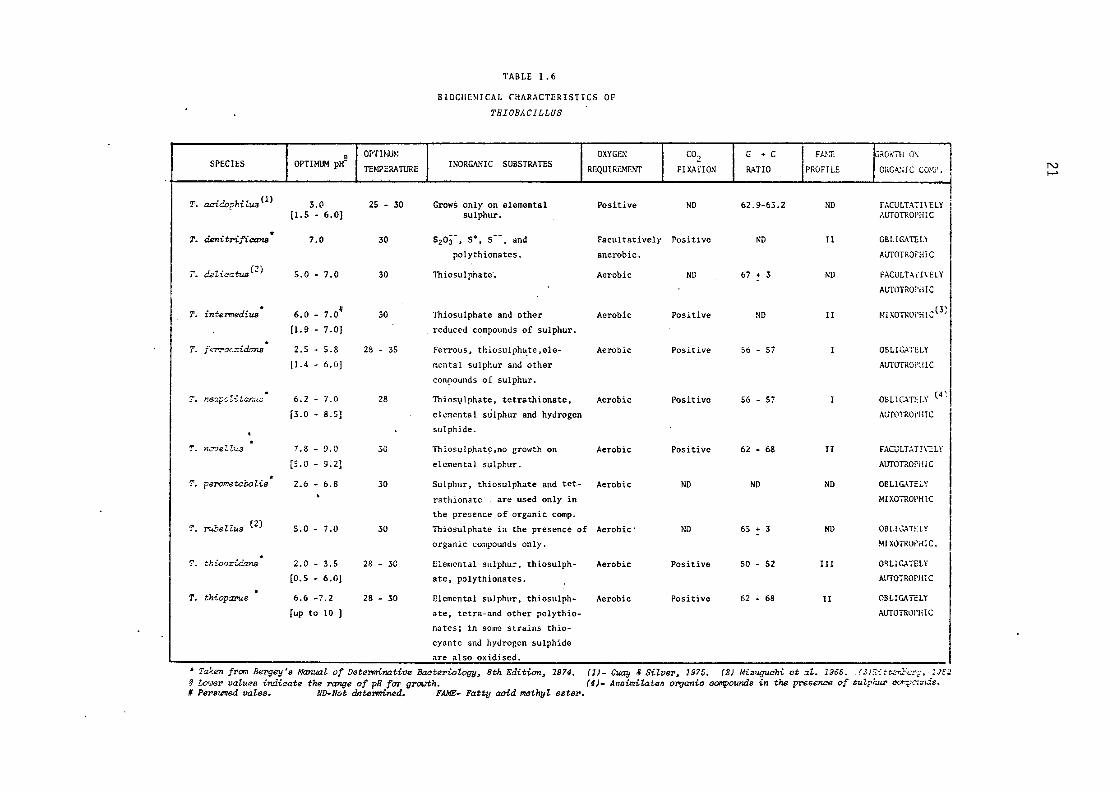
TABLE 1.6
BIOCHEMICAL CHARACTERISTICS OF
THIOBACILLUS
OXYGEN INORGANIC SUBSTRATES REQUIREMENT
Grows only on elemental Positive sulphur.
5203-, s•, s--. and Facultatively
polythionates. anerobic.
Thiosulphate·. Aerobic
Thiosulphate and other Aerobic
reduced compounds of sulphur.
Ferrous, thiosulpha,te ,ele- Aerobic
mental sulphur and other
compounds of sulphur.
Thiosvlphate, tetrathionate, Aerobic
elemental sulphur and hydrogen
sulphide .
Thiosulphate,no growth on Aerobic
elemental sulphur.
Sulphur, thiosulphate and tet- Aerobic
rathionatc are used only in
the presence of organic comp.
Thiosulphate in the presence of Aerobic·
organic compounds only .
Elemental sulphur, thiosulph- Aerobic
ate, polythionates.
Elemental sulphur, thiosulph- Aerobic
ate, tetra-and other polythio-
natcs; in some strains thio-
cyantc and hydrogen sulphide
are also oxidised.
C02 G + C FA.'<E ~RGi,'111 0\
FIXATION RATIO PROFILE ORGA.',lC Cu).(?.
ND 62.9-63.2 ND FACLILTATI\ELY AUTOTROPHIC
Positive ND II CBLIGATIL\
AITTOTROf'HiC
ND 67 + 3 ND FAGJLTATI\cl.Y
AUTOTROl'illC
Positive ND II MlXOTROrHI.::(J;
Positive 56 - 57 I OBLIGATELY
AlITOiROPl!lC
Positive 56 - 57 I OELlG.~Tii.Y (4~
AUTOTROi'lilC
Positive 62 - 68 II FACULTATI\1:LY
AUTOTROPH!C
ND ND ND OBLIGATEi.Y
MIXOTROPHIC
ND 65 + 3 ND OBLIGATHY
MIXOTROPHIC.
Positive so - 52 III OBLIGATELY
AlffOTROPHl C
Positive 62 - 68 II OBLIGATE LY
AlITOTROPHI C
• Ta:S:en from Bergey's Manual of Detel'minative Bacteriology, 8th Edition, 1974. (1}- Guay & SiZvez,, 1975. (2) Mizuguchi et at. 1966 .. (.li.'/:.tt.."r.for;;, 1Jc~ q W,]ql' values ir.::licate the :range of pH fo-r- gz,ovth. (4)- Aaaimil.ates organic compounds in the pNsence of sulp/;uz, =,;,l"..:n.is. fl Persumed vales. ND-Not dntennined. FAME- Fatty acid methyl estez,.
N I-"

22
ThiobaaiZZus thioparus etc.,) and (b) species which can oxidise either
ferrous ions or sulphur compounds ( ThiolxxciZZus ferrooxidans)
ThiobaciZZus denitrifieans is the only species known to oxidise
sulphur compounds with nitrate as the terminal electron acceptor in
place of oxygen, and it is consequently the only facultatively anaero
bic species in the genus. According to a recent investigation it has
been claimed that both ThiobaciZZus feYTooxidans and ThiobaciZZus thio
oxidans have the ability to grow anaerobically by oxidising elemental
sulphur in the presence of ferric ions, which actas achemical oxidant
(Brock & Gustafson, 1976).
The facultative autotrophs of the genus are differentiated on
the basis of the effect of organic compounds on the oxidation of in
organic sulphur compounds. In one species, ThiobaciZZus noveZZus,
organic matter represses the oxidation of thiosulphate while in the
other species, such as ThiobaeiZZus intemzedius, ThiolxxciZZus rubeZZus,
ThiobaeiZZus deZicatus, this oxidation is not affected by organic
matter; in fact the latter two organisms oxidise thiosulphate and
organic substrates simultaneously and are classified as mixotrophic
(Mizoguchi et al., 1976).
This classification lays much emphasis upon the autotrophic
nature or otherwise of the microorganisms, a criterion which itself
is liable to fall prey to many drawbacks, the more important being the
lack of a precise definition of autotrophy (Kelly, 1971; Schlegel,
1975; Whittenbury & Kelly, 1977). It has been reported by many workers
that it is possible, with special techniques, to grow even the most
putatively obligate autotrophs on glucose (Shafia & Wilkinson, 1969);
the possibility of growing ThiobaciZZus thiooxidans on glucose as the
sole energy source has been discussed by Borichewski & Umbreit (1966).
The biochemical characteristics of the genus ThiobaciZZus are
tabulated in Table 1.6.

23
In order to resolve the controversy on whether the Thiobaaillus
genus consists of a spectrum of types (Baalsrud, 1954), or is a genus
containing distinct species (Vishniac & Santer, 1957; Parker & Prisk,
1953), Hutchinson et al. (1965, 1966, 1967) studied the taxonomy of
the genus exhaustively. They extended the method of numerical taxonomy
pioneered by Sneath (1957) to this group of microorganisms and conclu
ded that the 'groups' and 'sub-groups' in the genus were well differ
entiated and that they were unable to detect any intermediate organisms
in this genus. While considering these conclusions, the critical dis
course on numerical taxonomy published by Pratt (1972) should be kept
in mind. According to this critique the fundamentals of numerical
taxonomy, namely' single character' and 'unit character' are inade
quately and sketchily drawn assumptions.
Another technique which is helpful in determining the groups or
sub-groups within a genus is the base composition of DNA, a technique
now frequently applied to the taxonomy of a wide range of microorgani-
sms. The technique is useful in supplementing the results of other
taxonomical studies ( viz., numerical taxonomy, FAME profile) and has
helped in a number of cases to arrange the groups into sub-groups.
An example which can be cited is that of confirmation of the subdiv
ision of the Staphyloaoaaus - Miaroaoaaus group into the two
distinct groups already indicated by the numerical taxonomical
results (Silvestri & Hill, 1965).
The DNA base composition of Thiobaailli was studied by
Jackson et al. (1968) and their results are summarised in Table 1.6.
Their analyses support to a certain extent the classification of
Thiobaailli into distinct species, as proposed by Hutchinson et al.
(1965-1967). A recent study, however, indicated some anomalies with
respect to DNA base composition of Thiobaaillus ferrooxida:ns when
grown on different substrates ( Guay et al., 1975; 1976).

24
Another criterion recently described for the classification of
Thiol>acilli is the fatty acid composition of the cells. These are
extracted, converted to methylesters, and their occurrence and relative
abundance determined by gas-liquid chromatography (Agate & Vishniac,
1973-a). It is possible to divide the Thiobacilli into three distinct
types, differing in their fatty acid content (FAME profiles). It has
been claimed by the authors that the method provides a rapid differen
tiation between such closely related species as Thiobacillus neapoli
tcmus and Thiobacillus thioparus. The three FAME-profiles are prese
nted in Table 1.6, Table 1.7 summarises the composition of these FAME
profiles. The FAME-profile obtained by Levin(l971) from a Thiobacillus
thiooxidans strain contained a predominance of c19 cyclopropanic acid
and was different from the FAME-profiles obtained from other strains
of Thiol>acillus thiooxidans. There are profound changes in the
compositions of phospholipids during various phases of growth (Agate
& Vishniac, 1969; 1973) and therefore the accuracy of FAME-profiles
depends upon the conditions of growth and age of the cultures. Since
in a microbiological system it is difficult to reproduce similar cond
itions the method should be used with some caution. Advantages and
disadvantages of chemotaxonomical methods employing FAME-profiles in
bacterial taxonomy have been recently discussed in an exhaustive
review by Lechevalier (1977).
The current status of the classification of Thiobacilli is
summarised in Table 1.8. As can be seen from this table, numerical
taxonomy recognizes eight independent species/groups existing distinc-
tly in the genus Thiobacillus. The results of chemotaxonomical
methods (viz., DNA base composition, FAME-profiles) tend to agree
closely with one another and in general with the numerical taxonomical
classification. The results obtained from the study of five Thiobaci
Zli have indicated that they can be grouped into three distinct groups.

TABLE 1.7
TYPES OF FATTY ACID METHYL ESTER (FAME) PROFILES
FOUND IN GENUS THIOBACILLUS#
25
* TOTAL EXTRACTED FATTY ACIDS IN: FATTY ACID TYPE I TYPE II TYPE
% % %
c6 7
Cs 6 4 6
Cg 7
C10 6 3
Cu 2 10
C12 4 14
C13 7
C14: 1 30 2 16
C14 12 3 21
C15 2 32
C16:1 15 2
C16 9 7 4
C17: 1 15
C17 20
III
# Taken from Bergey's Manual of Determinative Bacteriology,
8th Edition, 1974.
* Number indicates the length of carbon chain; number to
right of colon indicates number of double bonds.

NO. I
1-
2-
3-
4-
5-
6-
7-
8-
9-
10-
11-
TABLE 1.8
GROUPS OR SPECIES OF THIOBACILLI RECOGNISED BY
THREE INDEPENDENT METHODS OF TAXONOMY
NUMERICAL TAXONOMY I NUCLEOTIDE COMPOSITION LIPID COMPOSITION
(GC-RATIO) (FAME-PROFILE)
T. thioparus T. thioparus }
T. thiopa.rus} Type
T. noveUus T. noveUus T. noveHus 11 }
ND T. acidophilus 62-68% ND }
ND T. :rubeUus NO }
ND T. del,icatus tl)
T. neapo li tanus T. neapo li tanus 6 % }5 -57
T. neapo 7,i tan.us }Type
T. ferTOo:xidans T. f erroozidans T. f eZTOcxcidans I
T. thioo:r:idans T. thioo:r:idans 50-52% T. thioo:z:idans Type 111
T. denit'l'ificans ND N)
T. intermedius ND N)
T. t'l'auweinii. N) JI)
ND= Not described.
N O'I

27
All the three methods agree in the fact that ThiobaeiZZus
thiopa:r'us, ThiobaciUus neapoZitanus, ThiobaciUus thiooxidans and
Thio'ba,ciZZus ferrooxidans (all of which are grouped together in Fig.
1.2) are distinctly different species. Thus, al though these biochemical
techniques are relatively new to taxonomy, they have helped already
towards the shaping of a useful basis. for the classification of the
ThiobaciUi.
1.2.3.3 SUBMICROSCOPIC ORGANIZATION OF THIOBACILLI
Thio'ba,ciZZi are capable of thriving on inorganic substances
such as elemental sulphur and ferrous ions. Some members of the genus
are also known to withstand very high hydrogen ion concentrations. On
account of their peculiar substrates and sharply defined environmental
requirements, there has been a wide interest in the study of their
ultrastructure.
Umbreit & Anderson (1942) were among the first workers to study
the structure of ThiobaciZZus thiooxidans with an electron micro
scope. These workers could not confirm the 'dipolar' appearance of
the cell reported earlier by Umbreit et al. (1942) after examining
these cells mder optical microscope. They also could not recognize
a loose cell envelope, seen on some of their figures (viz., Figs. 4
& 6), which they regarded as either bacterial cell wall or an artifact
developed during the processing. Knaysi (1943) however, in an excell
ent paper, was the first to point out the existence of slime-coat or
loose cell-envelope around the cells of ThiobaciZZus thiooxidans. He
also considered the 'halo structures' in the electron micrographs of
ThiobaciZZus thiooxidans reported by Umbreit & Anderson to be the cell
envelope.
The fine structure of ThiobaciZZus ferrooxidans was most

28
probably studied and reported for the first time by Lundgren and his
colleagues in 1964 (Lundgren et al., 1964). This group undertook the
investigations in the hope of demonstrating a correlation between
structure and function and of finding some unique and novel structural
components which would explain the organisms' special physiological and
biochemical properties. However, they could not find anything unusual
about the structure of ThiobaciZZus ferrooxidans and described its
cellular organization as being comparable to that of ordinary Gram
negative bacteria, with a loose envelope which consisted of at least
two electron-dense areas, enclosing a non-electron-dense area in bet
ween them. Inside the cell envelope a unit membrane structure contained 0
the cytoplasmic material. Many particles of 70 - 150 A in size,
which were believed to be polyribosomes, were also reported.
In 1965, a working model for the oxidation of ferrous to ferric
salts by ThiobaciZZus ferrooxidans was proposed by Dugan & Lundgren
(1965). The model was based on assumption that a complex is formed at
the cell surface of the bacterium. The concept of involvement of the
cell envelope in the oxidation process prompted Remsen & Lundgren (1966)
to undertake rigorous investigations of the cell envelope structure;
they employed both freeze-etching and chemical-fixation techniques
during their studies. They found that the structure of chemically
fixed cells of FerrobaciZZi was similar to that of other Gram-negat
ive bacteria ( e.g., Escherichia coZi, Pseudomonas sp. ) as well as
to that of ThiobaciZZus thiooxidans as described by Mahoney & Edwards
(1966). They also demonstrated that the cell envelope of chemically
fixed cells comprised five separate layers, distinguishable by
their location and electron density. On the other hand, freeze-etched
cells revealed only three layers, measuring in thickness approximato
ely 100 A each, and identifiable as an outer lipoprotein-lipopoly
saccharide layer, a middle layer (containing globular protein

29
attached to fibrillar mucopeptide) and an innermost layer, the cyto-0
plasmic membrane, which was covered with particles (100 - 120 A
diameter) considered to be enzyme particles.
The cytomembranes observed in ultrasections of a marine Thiobaai
ZZus were less numerous than those found in other autotrophic micro-
organisms, like Nitrosomonas, Nitroaystis oaeanus and Nitrobaater
(Tilton et al., 1967-a). However, it was presumed that their presence
supported Murray's hypothesis, based on the ultrastructure of some
autotrophic bacteria, that a relationship exists between the degree of
energetic processes and the extent and complexitiesof cytomembranes
(Murray, 1963; Murray & Watson, 1965).
Recent studies demonstrate that the ultrastructure of a marine
ThiobaaiZZus, when examined by thin-sectioning and freeze-etching
techniques, is similar to that of terrestrial species of the Thiobaai
ZZus genus (Murphy et al., 1974).
The ultrastructure of the facultatively autotrophic members of
the genus ThiobaaiZZus ( ThiobaaiZZus noveZZus) was first studied
in Czechoslovakia by Kocur et al. (1968), who were concerned with
finding any structural differences that might exist between the
facultative and obligate autotrophs and with explaining the diffe
rences in terms of variations in their physiology. These workers
reported that ThiobaaiZZus noveZZus resembles the obligate auto-
troph, ThiobaaiZZus thiooxidans, in each detail of its structure, with
the exception of the presence of vacuoles in ThiobaaiZZus noveZZus.
However, they considered the vacuoles to be artifacts resulting from
plasmolysis of cells during preparations of the samples; this was
later confirmed by Van Caeseele & Lees (1969), who could find no
vacuoles in their samples. The autotrophically-grown cells of Thio
baaiZZus noveZZus were folfild to be devoid of the electron-dense layer
present in the cell envelope of the heterotrophically-grown cells; the

30
latter contained large inclusions of polysaccharides as well (Van
Caeseele & Lees, 1969).
A comparative study of the ultrastructure of Thiobacilli was
made by Shively et al. (1970) in which they attempted to pin-point
the characteristic structural features peculiar to each species.
Although the structure of the cell envelope was similar to that found
in most Gram-negative bacteria, obvious differences were noted in the
middle layers of the cell envelopes of the seven species studied: in
Thiobacillus thiooxidans and Thiobacillus A2 it was very prominent;
Thiobacillus thiopaPus & Thiobacillus intermedius contained less
conspicuous middle layers; in Thiobacillus noveiius this structure
was either absent or very diffuse. These workers also reported the
presence of lamellar bodies, resembling those present in photosynthetic
bacteria, in a few cells of Thiobacillus thiopaPUs. However, this
finding remains unconfirmed. Paracrystalline bodies of unknown
function were demonstrated in a few cells of Thiobacillus intermedius.
Polyhedral inclusions were seen in four of the species, namely,
ThiobaciUus thiopaPUs, ThiobaciUus neapolitanus, ThiobaciUus
intermedius and Thiobacillus thiooxidans.
Thus it can be seen that the studies of the ultrastructures of
Thiobacilli so far made have revealed no specific organelle possessed
by them, not even the presence of the cytomembranes which were thought
to be possessed by all autotrophic bacteria. In fact, in structural
architecture, they most closely resemble the heterotrophic Gram-negative
bacteria.
THIOBACILLI REVISITED
The findings that Thiobacilli are closely related to Gram-negative
heterotrophic bacteria in their submicroscopic organization could not
be agreed upon by Russian workers and some other groups. Although

31
there was much evidence accumulated from previous studies to support
the above conclusion, the very llllique physiology of ThiobaciZZi such
as tolerance to very low pH and very high metal ion concentrations
caused research workers to presume that these microorganisms possess
an ultrastructure distinct from other Gram-negative bacteria. Among
these groups, the most prominent are the Russian biologists who have
studied the ultrastructures of ThiobaciZZi once again and published a
series of very interesting research papers since 1971. In the
following the ultrastructure of these ThiobaciZZi as seen by these
investigators is briefly discussed.
ThiobaciZZus ferrooxidans was the first bacterium to be
re-examined by Avakyan & Karavaiko (1970). The bacterial cells were
fixed chemically for electron microscopy. They noted no significant
difference in cell wall structure between their strains and those
previously described by Remsen & Lundgren (1966). However, they
compared the cell wall structures of a number of bacteria and concluded
in contrast with the conclusions of the other workers that, although
the structure of this organelle is similar in ThiobaciZZus thiooxidans
and ThiobaciZZus ferrooxidans, it is definitely different from those
of certain other Gram-negative bacteria but is closely related to the
cell wall of Escherichia coZi, Nitrosomonas spp & Nitrobacter spp.
An LU1confirrned observation by the authors was the presence of
intracellular membrane structures, which they considered similar to
the lamellae of photosynthetic bacteria or the membrane structure of
nitric bacteria. According to them, the data indicated that the
intracellular structure of ThiobaciZZus ferrooxidans is more
sophisticated than it is in the heterotrophic Gram-negative bacteria
and a number of its features, especially the presence of highly deve
loped membrane structures, indicated that it more closely related to
chernotrophs like nitric bacteria.

32
The ultrastructure of ThiobaciZZus thiooxidans indicated that
the cell wall was similar to that of ThiobaciZZus feYTooxidans
(Karavaiko & Avakyan, 1971). The cells were fomd to contain intra
cellular membranes of the mesosome type which occur as complex invagi
nations of the cytoplasmic layers and which are covered with electron 0
dense particles of 40 - 70A diameter, possibly enzyme complexes. A
very important observation was that there were no lamella-like
membrane structures as detected in ThiobaciZZus thiooxidans previously
(Avakyan & Karavaiko, 1970).
Submicroscopic examination of ThiobaciZZus neapoZitanus revealed
that the cell wall was different from that of ThiobaciZZus thiooxidans
and ThiobaciZZus ferrooxidans (Karavaiko & Avakyan, 1971; Avakyan &
Karavaiko, 1970) as it lacked the middle dense layer and
was similar to that of many Gram-negative bacteria (Pivovarova &
Karavaiko, 1973). The cells were found to contain numerous membranous
structures representing loop-like invaginations of the cytoplasmic
membrane and were supposed to be concerned with the oxidation of
sulphur. The full mechanism of this oxidation is, however, still not
known. It is suggested by these workers that these intracellular
membranous structures are responsible for oxidation as well as
excretion of sulphur granules.
Unlike ThiobaciZZus neapoZitanus (Pivovarova & Karavaiko, 1973),
ThiobaciZZus thiocyanoxidans cell wall is complex and was found to
resemble in its organization that of ThiobaciZZus ferrooxidans and
ThiobaciZZus thiooxidans (Pivovarova & Karavaiko, 1973-a). The
ultrastructure of this ThiobaciZZus exhibited an abundance of
intracellular membranous structures, which differ in their shape and
structure from those of ThiobaciZZus neapoZitanus (Pivovarova &
Karavaiko, 1973). The presence of a middle layer in the cell wall and
the pecularity of the membrane structure are attributed to the sulphur

33
deposition. A general conclusion is that, in combination with the
membrane structures, the cell wall represents a unique mechanism for
the secretion of the colloidal sulphur produced within the cell.
However, it must be pointed out at this juncture that nature and
precise information about the mechanisms operating are still obscure.
Recently, the ultrastructure of ThiobaciZZus thiooxidans,
ThiobaciZZus thioparus, ThiobaciZZus interrnedius, ThiobaciZZus noveZ
Zus and ThiobaciZZus neapoZitanus has been studied again, employing
both chemical fixation and freeze-etching techniques (Holt et al.,
1974) with the hope of obtaining some information about the
relationship between the structural organization and physiological
function of these microorganisms. These workers have confirmed that
the architecture of ThiobaciZZi is typical of Gram-negative bacteria.
They also observed certain characteristic features of individual
ThiobaciZZus species, like the paracrystalline inclusions in
ThiobaciZZus intermedius and the lamellar body in ThiobaciZZus
thioparus:, which were reported earlier by Shively et al. (1970).
Ultrastructural organization of freeze-etched cells revealed
the multilayered cell envelope as a closely juxtaposed structure, each
layer having a peculiar texture. An outer layer of the cell envelope
could bedemonstratedin only a few species, for example, ThiobaciZZus
neapoZitanus and ThiobaciZZus thioparus, but was not present in each
species. The middle layer contained particles of about 80 A in
size, except in ThiobaciZZus ferrooxidans (Remsen & Lundgren, 1966)
where this layer was reported to be smooth textured. One important
observation made by these investigators was the presence of 120 A
diameter, membrane-botmd particle studs on the plasma membrane; these
studs possessed some degree of surface differentiation, for example
'hole or channel' configurations. It has been suggested by this
group that these structures might be the connections between the outer

34
envelope and the inner cell by providing the channels or holes between
these bolllldaries.
This study, like many others, has confirmed the presence of 0
various cytoplasmic inclusions, ranging from 2150 A diameter granules 0
to 1300 A diameter membrane bolllld polyhederal inclusions which have
been recently recognized in Thiobacillus neapolitanus as the structures
containing the ribulose diphosphate carboxylase, and which have been
named carboxysomes (Shively et al., 1973). Each carboxysome is bolllld 0
by a monolayer membrane approximately 35 A thick with an array of 0
100 A particles of ribulose-diphosphate carboxylase with a central hole
or depression (Shively et al., 1973-a). Similar polyhedral bodies have
also been reported in Thiobacillus intermedius(Purohit et al., 1976-a).
Dense bodies are generally seen and reported in all Thiobacilli.
Their fllllction and nature still remain obscure. Recently, Shively,
in a review has summarised the occurrence and functions of these
inclusions in the Thiobacilli (Shively, 1974).
There is now sufficient evidence to conclude that the utilisat
ion of inorganic sulphur compollllds by Thiobacilli is highly unlikely
to be related to the development of 'specialised' structures or
extensive membrane systems such as are known to occur in prokaryotic
phototrophs and ammonia- and methane-oxidising bacteria. As far as
the contradictory evidence for the presence or absence of cytomem-
branes in Thiobacilli is concerned this may be due to differences in
the strains which each of the workers has studied, or it may be the
result of the artifacts developed in the preparation of samples. However,
the idea of cytomembranes in Thiobacilli gets very little support from
the currently existing experimentar evidence and no evidence for stru
ctural involvement is available regarding the oxidation of sulphur and
sulphur compollllds. The puzzles relating to how Thiobacilli survive at
low pH, while maintaining the internal pH at physiological level, the

35
mode of attack on solid substrate, the transportation mechanisms
involved, and many other characteristics of the genus are enough to
keep cytologists busy for another decade.

36
1.3 BIODEGRADATION OF MINERALS BY MICROORGANISMS
A world wide depletion of mineral resources within the coming
decades has been frequently forcast by reliable authorities and it
is generally believed that the supply of ores of copper, mercury, zinc
and of many other vitally important metals for present day technology
is becoming more and more uncertain. According to one estimate, the
known workable ore bodies of copper, cobalt, nickel, vanadium and
lead may be depleted by the end of the year 2000 (Brooks, 1972; Harts,
1973). In order to maintain the present rate of inflow of various
metals for technological developments, future technology for metal
winning operations will possibly have to consider the following
sources or modifications in already existing operations:-
1- Development of efficient recycling metrods for metals
such as tin, lead, copper, zinc, etc.
2- Novel methods for recovering metals from the rocks in
which they exist at average abundance levels. Appli
cations of PNE (Peaceful Nuclear Energy Explosion) for
crushing the secondary ores under ground and leaching
them in situ had been suggested (Nordyke, 1971), but
the study is in its initial stages.
3- Metal Extraction From Sea.
Sea water can be a potential source of metals. This
is apparent from the following estimates of the total
amounts of certain industrial metals which are conta
ined in the oceans (Cloud, 1968).
( Calculations based on· total volume of
Sea= 3.3. 108 cubic miles.)

METALS MILLION TONS
Copper 5,000
Manganese 3,000
Mercury 50
Silver 500
Tin 5,000
Tungsten 150
Uranium 5,000
Zinc 15,000
Perhaps it is due to this huge magnitude of metals that
Warren (1973) has described the oceans as the " Mining
Frontiers of Future". The major difficulty in extract
ing metals from sea water is their low concentrations ;
this means that the very large volumes required to be
treated to obtain sufficient amounts of metals would
affect the economics adversely. However, development &
isolation of microbial strains which are resistant to
and capable of accumulating heavy metals can be helpful
in concentrating these metals (Zajic & Chiu, 1972).
4- Development of substitutes for metals.
37
Microorganisms play an important part in the formation as well
as degradation of minerals in this planet (Trudinger, 1971). They
can be used for enhancing metal production from the various sources
and operations described under headings 1 to 3 above. So far, however,
they have not been employed in the synthesis of metallic substitutes.
Both heterotrophic and autotrophic microorganisms are known to
degrade minerals and rocks. In the following sections, the action of
micro bes in the degradation of minerals is described.

38
1.3.1 BIODEGRADATION OF MINERALS BY HETEROTROPHIC MICROORGANISMS
There is ample evidence that heterotrophic microorganisms take
a very active part in weathering and degradation of rocks to form soil
in nature. During this process mineral components of rocks are solu
bilised. In a series of studies primarily concerned with the mechanism
of weathering, a number of workers have clearly demonstrated the abil
ity of soil bacteria, actinomycetes and fungi to decompose a number
of natural and synthetic silicate minerals (Duff & Webley, 1959; Webley
et al., 1960, 1963; Duff et al., 1963; Henderson & Duff, 1963). The
most active bacterial isolate was found to produce 2-keto-gluconic
acid, whilst citric and oxalic acids were the products of the fungal
action. Fungi were fotmd to be most effective in degrading silicate
minerals; in some cases dissolution was nearly complete (calcium
silicate 94%, magnesium silicate 74%, zinc silicate 96%). Bacteria and
actinomycetes have been found to give an average dissolution of
calcium silicate within the limits of 83 - 87 %. There is sufficient
evidence to conclude that Pseudomonas cultures can destroy the
crystalline structure of silicates like olivine [(Mg.Fe) 2Si04 ] or
wollastonite [Ca 3 (Si0 3) 2 ]. The possible mechanism has been described
as the acidic action of 2-keto-gluconic acid.
Decomposition of tri-calcium phosphate and rock phosphate incre
ase the fertility of agricultural land. Therefore, microorganisms
capable of solubilising rock phosphates are of special interest and
have been isolated from these rocks by a number of workers (Bardiya
& Gaur, 1974). Heterotrophic bacteria isolated from gold mines have
been found to dissolve gold (Pares~ 1964, 1964-a, b). Lyalikova and
Mokeicheva (1969) found that a culture isolated from the Zodsk gold
deposits, similar to Bacillus alvei, was able to dissolve 500 - 600
µg/t of gold in three weeks; consequently such microorganisms have

39
been presumed to play a vital role in the migration of gold deposits
in nature (Korobushkina et al., 1974).
Two mechanisms appear to be involved in the biodegradation of
minerals by these heterotrophic microflora:-
1- Acidification or alkalisation, and
2- Chelation of metals by organic metabolites.
Acidification or alkalisation.
Acidification may result either from secretion of acidic
metabolites or by a selective utilisation of alkaline components such
as ammonium ions; organic acids such as lactic, butyric, succinic and
gluconic are the usual end products of the carbohydrate metabolism in
these microorganisms. Alternatively, in certain cases the production
of ammonium ions by the utilisation of proteineous material increa-
ses the pH of the environment up to pH 10; at this pH certain minerals,
especially those of copper, are easily attacked and the process can be
compared to chemical leaching by ammonia (Le Roux, 1966).
Silverman & Munoz (1970) in a study involving Penicillum simpli
cissimum conclusively showed that the solubilising agent produced
during the degradation of various minerals (basalt, granite, grano
diorite, rhyotite, andesite, peridotites & quartzite) was citric acid.
Intracellular enzymes did not appear to be involved in the degradation,
since heated (80 °C, for 1 hour) and unheated spent acid media were
equally effective in solubilising basalt. That acid was involved in
this process was indicated by the change in the infrared spectra of
the rocks during the experiments; this was assumed to be due to the
disruption of the crystalline structure. The oxalic acid, produced by
metabolism of glucose by chemoorganotrophic microorganisms has been
considered responsible for the leaching of uranium ores in situ
(Magne et al., 1973). Agbim & Doxtader (1975) have shown that

40
solubilisation of synthetic zinc silicate and the minerals willemite
[Zn2(SiO4) ] and hemimorphite [Zn4Si2O7(OH)2 H2O] was facilitated
by soil inoculated glucose-mineral salt medium. Volatile and non
volatile organic acid fractions of the culture liquids were folllld to
solubilise substantial amounts of these minerals. These studies are
in agreement with the previous findings of Silverman & Munoz (1970)
claiming that heterotrophic solubilisation is a non-enzymic phenomenon.
In another investigation, mineralysis of silica quartz has been
reported to be dependent upon growth rate of Bacillus caldolytics
(Lauwers & Heinen, 1974).
Chelation of metal ions by metabolites.
Certain organic metabolites of bacteria and fungi have the
ability to chelate important divalent metallic ions such as iron,
copper, zinc, magnesium, calcium and manganese. These metabolites may
function as metal-transporting agents in cellular metabolism. Amino
acids (glycine, histidine, glutamic acid), phenols (pyrocatechol,
salicylic acid), phosphates (ATP, ADP), organic acids (citric, 2-keto
gluconic) and polyols (mannitol) are the usual bacterial metabolites
which have been found to have a very strong affinity for metabolically
important divalent metal ions (Bourne et al., 1959). During the disso
lution of gold by Bacillus megaterium and Bacillus mesentericus niger,
Korobushkina et al. (1974) demonstrated the significant role played by
amino acids by employing various mutants of above strains.
Pyrocatechol, which isrelated to compoW1ds occurring in hurnic
substances, chelates iron to form the following complex and thus
accomplishes solubilisation of iron for transportation in soils.
41
FERRIOXAMINE B
ENTEROCHEL IN
Chemical structures of iron-transporting
substances produced by bacteria.
[ Doetsch & Cook, 1973]

42
Phenolic acids (2,3-dihydrobenzoic acid, 2,3-dihydroxybenzoyl
glycine, 2,3-dihydroxybenzoyl serine, salicylic acid, 6-methyl
salicylic acid) are secreted by a number of bacteria and these acids
chelate iron in similar manner. These acids have been shown as
essential for growth of a mutant of Esaherichia coZi which was unable
to synthesise 2,3-dihydroxybenzoyl serine in the presence of iron.
A cyclic trimer of 2,3-dihydroxybenzoyl serine, known as"enterochelin",
produced by Escherichia coZi, has been regarded as the major factor
in iron transport in the coliform bacteria. Enterochelin chelates
iron as a hexadentate ligand through the phenolic hydroxyls.
Iron transport into bacterial cells can also be accomplished by
the so-called siderarnine groups of compounds which are complex hydro
xamic acid derivatives with a high affinity for iron, and are
recognized as growth factors for these organisms. They include terre
gene factors ( growth factors for Arthrobacter terregens ), coprogens
( growth factors for PhiZoboZus and also produced by Sarcina Zutea),
mycobactins ( growth factors for Mycobacterium pamtuber-euZosis) and
various ferrioxamines produced by Streptomyces species ( Doetsch &
Cook, 1973).
Arrieta & Grez (1971) isolated_ 10 soil fungi which solubilised
iron from minerals. They concluded that complex formation was the
most probable mechanism in this process, for even a very strong
cation-exchange resin was not able to extract the complexed iron from

43
the culture solution. Wenberg et al. (1969) have reported laboratory
scale leaching of copper minerals by PeniciUium species, under
neutral or slightly alkaline conditions. They too suggested a chelat
ion mechanism for solubilisation. Certain marine microorganisms
have been known to solubilise large amounts of iron from limonite and
geothite. There are several indications that iron oxide in the minerals
is reduced enzymatically to its soluble form (De Castro & Ehrlich, 1970;
Ottow & Vonklopotek, 1969).
The understanding of mineral leaching by heterotrophs is still in
its infancy and vital information regarding kinetics, environmental
effects and mechanisms of dissolution is scarce.
1.3.2 BIODEGRADATION OF MINERALS BY THIOBACILLI
The first report concerning the microbial degradation of sulp-
hide minerals was published in the first quarter of the ninteenth
century by Rudolfs & Helbronner (1922), who put forward the idea of
employing bacterial cultures for recovering metals from low grade,
sulphide-bearing ores and wastes (Rudolfs, 1922; Rudolfs & Helbronner,
1922). The idea was not pursued again until 1947, when Colmer and
Hinkle succeeded in isolating and describing a chemoautotrophic,
acidophilic, iron-oxidising bacterium from acid mine drainage; these
workers demonstrated that these bacterial cultures were associated
with, and actually played a fundamental role in, sulphide degradation
(Colmer & Hinkle, 1947). Similar microorganisms were isolated by
Razzell & Trussell (1963-a) from Rio Tinto (Spain) leach water. These
authors confirmed the biological nature of the process responsible for
the natural leaching of chalcopyrites reported by Taylor & Whelan (1943).
Since the identification of ThiobaciZZus ferrooxidans by
Temple and Colmer in 1951, the organism has been found to be involved

44
in leaching operations in Utah (Bryner et al., 1954; Bryner & Anderson,
1957), British Columbia (Razzell & Trussell, 1963), Arizona (Razzell &
Trussell, 1963; Corrick & Sutton, 1961), England (Ashmead, 1955),
Russia (Ivanov et al., 1961) the Congo (De Cuyper, 1964), Australia
(Andersen & Allman, 1968; Agate, 1973), Japan (Imai, 1971) and in many
other countries of the world.
Vishniac & Santer (1957) summarised the physiological and bio
chemical characteristics of the genus ThiobaciZZus as they were llllder
stood at that date. Later, the role of microorganisms in formation
and degradation of minerals was reviewed by Silverman & Ehrlich in
1964. More recently, an introductory review pertaining to microbial
processes capable of attacking and degrading minerals was published
by Le Roux (1971), and Fletcher (1971) reviewed briefly the bacterial
leaching of low-grade ores. Since then Tuovinen & Kelly have published
a number of excellent review articles dealing with the biology and
mineral oxidative activities of the genus ThiolxiciZZus (Tuovinen, 1972;
Tuovinen & Kelly, 1972; 1974-d). These authors have painstakingly
collected and evaluated the huge amount of work published on the
subject. Exhaustive as they are, these reviews all lack a detailed
account of leaching mechanisms. This gap has been filled by a
detailed account of the known oxidative mechanisms involved in mineral
degradation by bacteria (Ralph, 1978).
The microbiological leaching of metal sulphides is now a common
practice throughout the world for recovering copper and uranium from
low grade ores. Torma (1977) has critically reviewed the biogenic
role of bacteria in hydrometallurgical operations.

45
1.3.3 FACTORS AFFECTING THE MICROBIAL LEACHING PROCESS
Microbial leaching of sulphide minerals by ThiobaciZZus species
can be considered as a particular case of the general interaction of
microorganisms with solid, insoluble substrates; extension of some
aspects of the general rationale of such transformations [ as developed
by Nickerson (1969) while studying the breakdown of natural organic
polymers] to mineral leaching studies might be advantageous. The two
important components involved in microbial leaching are the solid
substrates and the microbial agencies. The efficiency of the process
will be dependent upon the degree of associations, contacts or inter
actions between these two components and will be influenced by the
nature of the substrate and microbial system present.
1.3.3.1 NATURE OF THE SUBSTRATE
Metal Distribution.
In an ore-body, sulphide minerals are generally associated with
other materials, such as quartz, silicates, carbonates and oxides.
The distribution pattern within the ore-body is one of the major
parameters determining procedures most suitable for extraction of the
metal from that ore-body. Generally, high grade ores are processed
by pyrometallurgical operations, whereas low grade ores are dumped at
a suitable place near the mine, and may subsequently be leached.
Metal leaching rates are enhanced by acidophilic members of the
ThiobaciZZus genus, namely, ThiobaciZZus ferrooxidans and ThiobaciZZus
thiooxidans, which proliferate actively in low pH conditions. When the
metal sulphide is associated with high proportions of alkaline matrix,
achievement of low pH becomes impossible due to'high consumption of
acid for neutralization of such alkaline materials. This may

46
effectively slow down or completely inhibit the leaching process.
Oxidative degradation of mineral sulphide depends upon the
access to it of water, various dissolved ions, oxygen and other rea
gents; and the breakdown may be accelerated by the closer proximity of
bacteria. Access of these various components of the degradative sys
tem will depend upon a number of factors including relative distri
bution of mineral sulphides and matrix ( for example in veins, as
massive aggregates, finely disseminated etc.) and upon the diffusion
characteristics of the matrix in respect to the various reacting
components.
Properties of Component Minerals.
About two hundred and fifty metallic sulphides have physical and
chemical characteristics which allow them to exist as individual mineral
species in geological environments (Ralph, 1978). Each mineral sulp-
hide hasuniqµe characteristics which affect its behaviour as a
substrate in bacterial leaching systems. For example copper exists in
a number of minerals containing a sulphidic moiety (chalcocite Cu2S;
chalcopyrite CuFeS2 ; covellite CuS; enargite Cu3AsS4 ; bornite Cu5FeS4;
digenite Cu9S5). The degree of leachability of these minerals with
bacteria varies to a considerable extent and the mineralogy of copper
sulphide ores is well known to affect its ease of leaching, Razzell
& Trussell (1963-a) found chalcopyrite to be relatively resistant to
chemical leaching but readily oxidisable by bacterial cultures.
Covellite was auto-oxidised less readily than chalcocite, and conversely
was reported to be more readily pron~ to biological oxidation (Ehrlich
& Fox, 1967). Copper minerals containing iron in association ( i.e.,
bornite Cu 5FeS4 and chalcopyrite CuFeS2 ) are reported to be almost
completely oxidised by bacteria (Duncan and Trussell, 1964). In
contrast, enargite, which contains arsenic, has been found to be only

47
11 - 20 % oxidisable by these bacteria (Hao et al., 1972). The
difference in leaching behaviour can be attributed to the modification
of structure of these minerals brought about by different compositions.
Compositional variations may be accompanied by changes in the crysta
lline form ( e. g. pyrrhotites) offering entirely different surfaces for
interaction with bacteria. However, these variations have been
little studied in relation to their possible effects on the rate of
biodegradation.
The difference in leaching behaviour exhibited by different
sulphide minerals is probably due to a complex of factors which include
the stoichiometry of the mineral, its crystallographic form, its degree
of substitution by other metallic ions, the basic chemistry of the
metal itself and perhaps more subtle characteristics of an electroche
mical kind. The effect of such factors may well be overlaid by the
grosser effect of experimental parameters such as pH, Eh, temperature,
particle size and so on.
1.3.3.2 EFFECT OF PHYSICO-CHEMICAL FACTORS
Particle Size.
Particle size influences the leaching rate in a number of ways.
The smaller the size of the mineral particles the greater is the inter
facial area between the solid and liquid phases and therefore the
higher the rate of transfer of materials and the greater the specific
area of interaction between microorganisms and substrate.-
Many investigators have reported an enhancement of bacterial
leaching rate on reduction of particle size of the mineral. For
example, Razzell & Trussell(l963) reported that doubling the surface
area of chalcopyrite (by reducing the particle size from a mean
diameter of 53µ to 44µ) doubled the rate of leaching. Duncan et al.

48
(1964), while demonstrating the effects of surfactants and shaking on
the leachability of chalcopyrite by Thiobacillus ferrooxidans, employed
mineral particles less than 44 µ in diameter; Ehrlich & Fox (1967)
observed an increase in the rate of oxidation of synthetic copper
sulphide when particles less than 62 µ instead of 177-125 µ were used.
MoS2 was fotmd to be oxidised bacteriologically at a faster rate in
the airlift column when particles smaller than 250 µ, rather than 600µ,
were employed. The highest rates of bacterial oxidation of pyrites
were achieved in airlift columns by using particles less than 44 µ
rather than larger particles (Malouf & Prater, 1961). About six times
more copper was microbiologically recovered from chalcopyrite using a
finely grotmd ore of less than 44 µ diameter (Razzell & Trussell, 1963-a).
A similar enhancement in the oxidation of pyrites by using particles
of less than 44 µ in a Warburg respirometer was reported by Silverman
et al. (1961). The effect of particle size on the extraction of copper
has also been reported by Bruynesteyn & Duncan (1971). Since reduction
of particle size essentially results in increasing the surface area
of substrate, Torma et al. (1970 & 1972) studied the effect of initial
specific surface area of zinc sulphide concentrates on the rate of
metal release by Thiobacillus ferrooxidans ( also see Torma & Subra
manian, 1974). These authors were able to demonstrate that at 16%
pulp density zinc extraction rate was increased significantly by
increasing the specific surface area from 0.5 m2 /g to 1.37 m2/g,
a further increase in the specific surface area by using particles
smaller than 40 µ produced no further dramatic changes in the rate of
leaching (Torma & Legault, 1973; Torma et al., 1974; Torma & Guay,
1976). This rather surprising result, which is not matched in studies
on the chemical leaching of very finely divided chalcopyrite, may be
accotmted for by the formation of particle aggregates between the
finer particles and the microorganism (Torma & Gabra, 1976). Data

49
obtained by Torma et al. (1972) suggest that the rate of zinc extract
ion at maximum surface area is limited by carbon dioxide.
A decrease in particle size, however, may not always result in
an enhancement of rate of metal release, especially in the case of
low-grade ores where the metallic compound (substrate) is sparsely
distributed in the host rock. Under such circumstances the properties
of the host rock itself may influence the leaching process and should
be considered carefully. Reduction of particle size in such cases may
not always expose new mineral surfaces. On the contrary, it can
increase host rock surfaces immensely with an adverse effect on the
leaching process. Attempts to simulate the natural gangue-mineral
situations, in laboratory conditions were made by 'diluting' pure
copper sulphide with quartz sand during airlift leaching experiments.
Results from such studies have indicated that the situation is a
complex one for the mineral-sand mixture did not behave as did the
low-grade ores (Bryner et al., 1954; Bryner & Anderson, 1957; Malouf
& Prater, 1961).
Interaction between size of particle and depth of column has been
successfully demonstrated by Audsley & Daborn (1963) while investiga
ting leaching of uranium ores in columns. Their results showed that
the rate of uranium extraction depended upon the depth and particle
size of the ore-beds; air permeability of the column was concluded to
be a significant factor. The maximum rate of oxidation from Bica ore
was obtained in a 2 feet deep column, containing ore particles of mean
diameter 0.25 inches, which was irrigated at the rate of 25 ml per
kg of ore per week. In deep beds the rate of oxygen transfer rather
than the diffusion of solutes was shown to be rate limiting.
So far no data are available for the effect of microfined powders
of minerals on the rate of leaching by bacteria. It will be very
interesting to examine the rate of leaching of mineral particles when

50
they are reduced to size smaller than microorganism itself. It should
be noted that reduction of particle size is a very expensive process
on a large scale and is probably only of relevance for a bacterial
leaching process when such methods are used for the recovery of metals
from flotation residues and other bye-products of conventional meta
llurgical techniques.
Surface Effects.
Bacterial activity in soil is reported to be enhanced by the
presence of solid surfaces (Stotsky & Rem, 1966) and an increase in the
oxidation rate of ferrous iron solution in the presence of an inert
solid surface was noticed by Malouf & Prater (1961). The explanation
put forward for this phenomenon was that nutrients accumulate in the
vicinity of bacteria due to surface adsorption. Recently Marshall
(1975) has reviewed various interactions and physico-chemical phenomena
observed during the contact of bacteria with solid particles.
The beneficial effects of surface active agents on the initial
rate of microbial leaching (Duncan et al., 1964) and isolation of
phosphatidylinositol, a surface active compound from the culture fluids
of ThiobaciZZus thiooxidans, by Schaeffer & Umbreit (1963) indicated
a possibility that wetting of surfaces by these compounds enabled the
bacteria to cover the mineral surfaces during the process of leaching.
However, data obtained in a recent investigation of leaching of chalco
pyrite with ThiobaciZZus ferrooxidans in the presence of externally
added wetting agents (Tween 20, 40 & 60) are in disagreement with the
previous reports (Torma, 1976; Torma & Gabra, 1975, 1976). According
to these workers the wetting agent reduced the chalcopyrite oxidation
ability of ThiobaciZZus fer.rooxidans by reducing the surface tension
of the medium and thus affecting the oxygen mass transfer.
Earlier work presents some evidence of bacterial attachment to

51
crystals of elemental sulphur (Schaeffer et al., 1963). Adherence of
Thiobaaillus ferrooxidans to solid surfaces like pyrite and inert
surfaces like polyvinyl chloride has also been frequently reported
(Mehta & Le Roux,, 1974; Tributsch, 1976). Suggestion of the occurre
nce of an organic complexing agent to bind the iron so that its redox
potential is decreased to a value enabling it to couple with the
electron-transport system of the cell during its oxidation (Blaylock
& Nason, 1963) implies that activity is concentrated on the solid
surface in a 'microenvironment' which is controlled by adsorption
properties as well as the capacity of the bacteria to modify their
microenvironment in such a way that energy transfer can occur most
efficiently.
Recent work has shown that the selective attachment of therrno
philic microorganisrns of the genus Sulfolobus to molybdenite (MoS2 ),
pyrite (FeS2 ), or chalcopyrite (CuFeS2) is generally by adhesion
and not by pili (Berry & Murr, 1975; Murr & Berry, 1976). These
results emerged from the studies of a very important aspect of
mineral/bacterial interaction, i.e., the mode and place of attachment.
Further detailed investigation in this direction would no doubt add
much to our understanding of the fundamental mechanism involved in
bacterial leaching.
Temperature.
The effect of temperature on microbial leaching has been studied
extensively (Ehrlich & Fox, 1967; Leathen et al., 1956; Silverman &
Lundgren, 1959; Kinsel, 1960; Marchlewitz & Schwartz, 1961; Bryner et
al., 1966). Some of the early workers tried to differentiate between
the temperature optima for growth of the associated microorganisms and
for leaching processes. Leathen et al. (1956) had reported the
optimal temperature for growth of Thiobaaillus ferrooxidans as being

52
between 20 and 25°C, but according to Silverman & Lundgren (1959)
the optimal temperature for growth of ThiobaciZZus ferrooxidans was
28°C whilst the maximum rate of oxidation of ferrous ion was at 37°C.
However, more recently, workers have demonstrated that oxidative pro
cesses carried out by ThiobaciZZi are growth-associated, having their
temperature limits between 30 and 40 °C with an optimum usually at 35 °C
(Landesman et al., 1966; MacDonald & Clark, 1970; Bryner et al., 1966;
Torma, 1971). Manometric experiments revealed somewhat higher tempe
rature optima for mineral leaching by cell suspensions of ThiobaciZZus
ferrooxidans, and sulphur oxidation was found to be more sensitive than
iron oxidation to higher temperatures and was completely inhibited at
45°C (Beck, 1969). Torma et al. (1970) studied the effect of tempera
ture on leaching of zinc sulphide concentrate by ThiobaciZZus ferro
oxidans and found that microbial activity was limited at temperatures
greater than 45°C, and had an optimum at 35°C. Some recent studies
using fully monitored apparatus and continuous culture techniques
have demonstrated an optimum temperature for leaching by ThiobaciZZus
ferrooxidans between 30 and 35 °C (Gormely et al., 1975; Tomizuka
et al., 1976). (1972)
Tuovinen & Kelly are of the opinion that absolute limits of
temperatures cannot be precisely expressed for ThiobaciZZus ferrooxi
dans, since, like other microorganisms, response . of these bacteria
to temperature depends upon their previous growth histories, Further,
in virtually all published work on the effect of physico-chemical
variables on leaching rates the assumption is made that there is no
interaction between the different variables. It might be deduced
from general considerations ( for example Brock's work on the environ
mental limits) that at least some of the physico-chemical factors are
in fact highly interdependent and temperature and pH might well be two
of the most significant factors in this respect (Brock & Darland, 1970).

53
Under natural environments, active leaching dumps usually become
hot and temperatures upto 60-80°C have been reported, whereas in other
instances reports describing kindling the heap exist (Lyalikova, 1960).
High temperatures in the interior of the heap are mainly due to the
low thermal conductance of the rock mass and production of heat by
oxidative processes, both biological and abiological, whose sequence
is mainly unknown at present. Evidence that the initial steps in the
oxidative train are due to the participation of ThiobaciUus ferroox
idans is presented by Lyalikova (1961) on the basis of direct observ
ation of heating of the Kransogvardeisk deposits in the Middle Ural.
Effluent water temperatures well above the ambient temperature of 30 °C
from leaching heaps have been noted by Beck (1967) who showed that the
bacterial load of iron-oxidisers in the waters was highest ( 5.0 104
cells per ml) at 27°C and was dramatically reduced to 50 cells· per ml
in waters with a temperature of 39°C. This low population of bacteria
at higher temperature was also noticed during a microbial survey of
overburden heaps at Rum Jungle Mine (Khalid & Ralph, 1976); it indic
ates abiological leaching at this temperature. Leaching studies by
Bryner et al. (1966) have shown a rapid rate of copper solubilisation ab
ove. 45°C in a pure chemical system, but biological leaching was
maximum at 35°C with minimal activity at 45°C. Recently Corrans &
Scholtz (1976) have reported that chemical leaching of pentlandite is
highly temperature dependent.
Existence of thermophilic ThiobaciZZi has not so far been recor
ded with certainty, although one unconfirmed and doubtful thermophile,
ThiobaciZZus thermophiZica Imchenestskii, has been reported by Egorova
& Deryugina (1963). Earlier attempts to isolate thermophilic Thiobaci
ZZi from high temperature habitats were unsuccessful ( Marchlewitz &
Schwartz, 1961).

54
Isolation of therrnophilic ThiobaciZZus -like bacteria, capable of
oxidising sulphur and ferrous salts, has been carried out by Le Roux
et al. (1977) and Brierley & Lockwood (1977). These isolates have been
reported to oxidise metallic sulphides within a wide range of pH values
(Brierley & Le Roux, 1977). Caldwell et al. (1976) have reported the
isolation of a high-pH, facultatively anaerobic, facultatively autotr
ophic therrnophile, Thermoth:rix thioparus, whose temperature optimum is
70 - 73°C. In recent years, therrnophilic sulphur oxidisers have been
isolated from hot spring waters (Brock et al., 1972; Brierley, 1966;
Brierley.&. Brierley, 1973; Williams & Hoare, 1972) and have been named
as SuZfoZobus species. Brierley & Murr (1973) have demonstrated their
capabilities for leaching sulphidic minerals and to facilitate the
solubilisation of molybdenum at 60°C (Brierley, 1974; 1977). It may
be possible that these species are associated with leaching operations
in the hot regions inside the heaps. However,there are no reports of
their isolation from a heap.
The lower limits of temperature for activity of ThiobaciZZi have
not been studied extensively. Their leaching capabilities appear to
be hindered drastically at lower temperatures; this is indicated by
unsuccessful heap leaching operations in the cold parts of the world,
for example in copper mines on the Kola Peninsula in the USSR which lie
north of the Arctic Circle with temperature as low as 4°C (Lyalikova,
1961); in low temperature Canadian mines higher recoveries of uranium
have been obtained by a process involving hot water.
Redox Potential a:nd pH.
Microorganisms in any environment are affected by the energy flow
processes mediated by electrons and hydrogen ions. Therefore, the
existence of microbial populations can be predicted in certain environ
ments provided the oxidation-reduction characteristics of that

55
milieu are know. That occurrence of specific microflora is controlled
by the redox potential and the hydrogen ion concentration of the loca
tion has been well doct.nnented (Baas-Becking et al., 1960).
Diagrams can be constructed, from the redox potentials (Eh) and
pH values, to represent the free energies of the chemical species
involved over a pH range; detailed methods for constructing such
diagrams are available (Garrels & Christ, 1965; Pourbaix, 1966). Eh-pH
diagrams, as these are referred to, indicate the precise conditions of
Eh and pH for effective energy transfer. These diagrams also contri
bute significantly towards the development of hydrometallurgical
operations for metal sulphides. The metal sulphide, a common substrate
during microbial oxidation of sulphide minerals, forms one of the
components of the metal-sulphur-water system which is not a well under
stood system thermodynamically. With the help of Eh-pH diagrams the
lack of thermodynamic data required for determining the mechanism of
oxidation of most of the sulphides has been overcome (Majima & Peters,
1969). Moss and Andersen (1968) extended the use of Eh-pH diagrams to
study the mechanism of heap-leaching at Rum Jungle.
Microbial activity itself is highly pH dependent. Although high
rates of leaching of metallic sulphides has been reported in aqueous
solutions at pH values between 2 and 4 with ThiobaciZZus ferrooxidans,
the maximum leaching rate of these sulphides was at pH 2.5 (Razzell,
1962; Razzell & Trussell, 1963-a). A large nt.nnber of investigations
tend to support the conclusion that the optimt.nn pH for microbial
leaching of sulphide minerals is 2.5, a value usually quoted as optimal
for their growth ( Gupta & Sant, 1970; Duncan & Teather, 1966; Moss &
Andersen, 1968).
There is evidence that ThiobaciZZus ferrooxidans and ThiobaciZZus
thiooxidans are capable of oxidising sulphur and fixing carbon dioxide

56
at pH values as high as 5.0 and the experiments of Rao and Berger
(1971) on ThiobaciZZus thiooxidans indicate the retention of oxidative
ability in respect of elemental sulphur and maintenance of via-
bility at pH as high as 7.0. This work noted that at these high
pH values there was an uncoupling of substrate oxidation and oxidative
phosphorylation and no growth. Although these microorganisms ( i.e.,
ThiobaciZZus ferrooxidans and ThiobaciZZus thiooxidans) were cap
able of oxidising sulphur at pH values as high as 5.0, they seldom
oxidised metallic sulphides at pH values higher than 3.5 (Duncan et al.
1966; Rao & Berger, 1971). These studies have relied on measurement
of solubilised metal as measurement of metal degradation and at pH
values above 4.5 this would not be expected to occur for purely
chemical reasons, the oxidation of the sulphidic moiety to sulphate
can occur at relatively high pH without metal release (Mizoguchi
et al., 1976).
Some Russian work indicates that the pH optimum for covellite
and pyrite oxidation was 2.0 - 1.7, and that bacterial activity was
also affected by the concentration of metallic ions and the temperature.
According to these authors, if concentrations of iron, copper and zinc
were less than 5,000, 4,000 & 8,000 p.p.m. respectively at temperat
ures below 50°C, the optimal pH value for microbial oxidation of the min
eral was less than 1.5 (Ivanov, & Nagirnayak, 1962; Golomzik &
Ivanov, 1965). Watanabe (1968) has shown that the pH optimum for
ThiobaciZZus ferrooxidans towards high grade ores of copper sulphide
is 2.5 whereas pH 1.5 was found more satisfactory for low-grade
ores.

57
1. 3. 3. 3 NUTRITIONAL REQUIREMENTS OF THE MICROFWRA INVOLVED
IN LEACHING
In the previous section, factors relating to the substrate and
the microenvironment have been delineated and now some of the other
important parameters controlling microbial activities during mineral
degradation will be discussed. Besides the appropriate environmental
conditions of temperature, pH and Eh as described earlier, microorganisms
have specific requirements for nutrients in order to fulfil their
demands for the carbon, nitrogen, oxygen and phosphorous, etc., needed
for building cellular components. Trace elements are also required for
optimal biological functions.
Requirement for Carbon.
Chemolithotrophic bacteria are capable of acquiring carbon for
their biosynthetic activities from carbon dioxide. Carbon dioxide
fixation in these bacteria is achieved at the expense of free energy
released during oxidation of inorganic compounds and trapped in the
form of high energy compounds. ATP has been shown to be the major
energy trapping system in chemolithotrophs ( Aleem & Nason, 1960; Cole
& Aleem, 1973). Substrate level phosphorylation during sulphite oxida
tion by APS-reductase has been well documented for ThiobaciZZus thiop
arus ( Peck, 1960; Peck et al., 1965; Peck & Fisher, 1961). Schlegel
(1975) has recently summed up various mechanisms used by chemoautotr
ophs to fulfil their carbon demands.
NADH is not produced by substrate oxidation in the chemolithotr
ophic bacteria but it has been shown that it is possible to obtain
reduced NAD by means of a reversed electron transport system which,
like that of mitochondria, is ATP - dependent. Thus, although

58
oxidation of inorganic substrates by chemolithotrophs is linked directly
to an electron transport system, the ATP produced during these oxidat
ive processes could be partially used for deriving a reverse electron
flow for producing the NADH required for Co2 fixation. The possession
of the enzyme necessary for such a complete respiratory chain by
Nitrosomonas, Nitrobacter & ThiobaciZZi was established by Aleem and
his coworkers (Aleem, 1966, 1966-a, b; 1969; 1970; Aleem et al., 1963;
Peeters & Aleem, 1970) in their demonstration of an energy dependent
NAO reduction by electron transport from reduced cytochromes. The
reports of cytochrome c reduction by NADH in ThiobaciZZus neapoZitanus
(Trudinger & Kelly, 1968) and by NADPH in ThiobaciZZus thiooxida,ns
(Tano & Imai, 1968) and of cytochromes c and a by NADH in extracts
of N. winogradski(Van Gool & Laudelou41967) have shown, however,
that normal electron flow pathways can also exist in obligate
lithotrophs.
There is a general agreement regarding the fixation of C02 by
chemolithotrophs via the reductive pentose phosphate pathway (Calvin
cycle) ( Elsden, 1962; Kelly, 1967, 1971). Evidence for operation of
this pathway consists of direct demon5tration of ribulose diphosphate
carboxylase ( carboxydismutase, EC 4.1.1.39) activity in extracts and
whole cells (by the fixation of 14co2 into 3-phosphoglyceric acid,
3PGA) and is available for ThiobaciZZus thiooxida,ns(Suzuki & Werkman,
1958, 1958-b; Iwatsuka et al., 1962; Iwatsuka, 1962), ThiobaciZZus
thioparus (Santer & Vishniac, 1955), ThiobaciZZus noveZZus (Aleem &
Haung, 1965), ThiobaciZZus neapoZitanus (MacElroy et al., 1968),
ThiobaciZZus ferrooxida,ns (Gale & Beck, 1967; Din et al., 1967) and
ThiobaciZZus denitrificans (Trudinger, 1955, 1956).
Three molecules of ATP and two molecules of reduced pyridine
nucleotide are required by the Calvin-Benson cycle for fixing one
molecule of carbon dioxide. The ATP is derived directly from the

59
oxidation of growth substrates, whereas pyridine nucleotides are
reduced by the reversal of the electron transport chain. That reduced
compounds of sulphur and ferrous ions can be used for this process has
been amply demonstrated by various workers (Aleem, 1977; Aleem et al.,
1963; Margalith et al., 1966; Silver, 1970; 1977; Silver & Torma, 1974).
An important feature of this system in the ThiobaciZZi is the
inhibition by AMP of ATP-dependent CO2 fixation via ribulose diphos
phate carboxylase (Gale & Beck, 1966; Johnson, 1966). AMP inhibition
was suggested as being due to allosteric modification of phosphoribulo
kinase (Johnson, 1966) and Gale & Beck (1966) fomd that in ThiobaciZZ
us ferrooxidans CO2 fixation inhibition was in fact competitive with
AMP. Studies with phosphoribulokinase isolated from ThiobaciZZus
thioparu.s indicated that the enzyme was cooperatively affected by ATP
suggesting that inhibition of this enzyme by AMP may not be solely
competitive (MacElroy et al., 1968-a). In another study of RuDP
carboxylase from ThiobaciZZus noveZZus, high levels of inhibition (43%)
obtained with ADP seems to be indicative of a regulatory role for this
compomd also during Co2 fixation (McCarthy & Charles, 1973). This
inhibitory mechanism may be an effective control for preventing the
unnecessary wastage of ATP in Co2 fixation; in the case of depletion
of ATP, AMP produced from ATP would tend to shut off CO2 fixation by
inhibiting the ATP-consuming phosphoribulokinase.
In some of the chemolithotrophs, CO2 may also be fixed by carb
oxylation of phosphoenolpyruvate (PEP) to produce oxaloacetate (Aubert
et al., 1957-a). PEP carboxylase (EC 4.1.1.31) activity has been
demonstrated in ThiobaciZZus thiooxida:ns (Suzuki & Werkman, 1957, 1958;
Howden et al., 1972), ThiobaciZZus thioparus (Hoban & Lyric, 1975),
ThiobaciZZus ferrooxida:ns (Din et al., 1967) and ThiobaciZZus noveZZus,
(McCarthy & Charles, 1974).

60
The significance of this pathway for C02 fixation during growth
of chemolithotrophs is not fully understood, for the Calvin-Benson
pathway appears to be the predominent pathway during Co2 fixation.
This is indicated by very heavy labelling of 3PGA ( >80%) compared
to only 8% labelling of aspartic acid during experiments performed
with cell extracts or whole cells. Furthermore, a close relationship
between the Calvin cycle and chemolithotrophy has been demonstrated by
repression of synthesis of ribulose diphosphate carboxylase during
growth of the facultative chemolithotroph Thiobacillus novellus on
organic substrates, whilst PEP carboxylase levels were relatively unaff
ected (McCarthy & Charles, 1974), and by the fact that ribulose diphos
phate carboxylase could not be detected in Thiobacillus intermedius
when grown heterotrophically on yeast extract (Purohit et al., 1976).
The presence of the cardinal enzyme of the Calvin cycle, ribu
lose-1,5-diphosphate carboxylase, has been demonstrated in the
'carboxysomes' of ThiobaciUus neapolitanus (Shively et al., 1970,
1973-a) and recently Purohit and coworkers (1976-a) have presented
unequivocal evidence for the presence of polyhedral carboxysomes in
Thiobacillus intermedius during autotrophic growth.
According to the Bunsen Absorption Coefficient, Co2 solubility
in water is not affected drastically by sulphuric acid (Tuovinen &
Kelly, 1972). However, electrolytes have been found to retard C02
solubility in water due to a 'salting out' effect. This suggests that
the toxicity of high concentrations of metals may be due to their imp
osition of a carbon dioxide limitation. The partial pressure of
carbon dioxide affects the solubility and more than 100% increase
in carbon dioxide fixation by resting cells of Thiobacillus ferroo
xidans was noticed when the carbon dioxide partial pressure was
increased from 0.15 to 2.4% (v/v) with respect to oxygen in Warburg
flasks (Beck & Shafia, 1964).

61
Nitrogen SoUX'ae.
Thiobaailli have a preference for ammonium nitrogen. Nitrates
and nitrites, when present as the sole nitrogenous source, have been
reported to be unassimilated (Lundgren et al., 1964; Corrick & Sutton,
1965). Some amino acids ( alanine, lysine, arginine, histidine and
glutamic acid) have been found to substitute totally or partially for
ammonium ions during growth of Thiobaaillus ferrooxidans (Lundgren et
al., 1964; Remsen & Lundgren, 1963).
It has been demonstrated that Thiobaailli, in particular Thioba
aillus ferrooxidans, have a very low nitrogen requirement (Tuovinen
et al., 1971) and this characteristic has been employed in designing
nitrogen-limiting medium for isolation of an iron-oxidising bacterium
(Beck & Elsden, 1958). This low requirement of nitrogen by acidophilic
iron-oxidisers has been explained as being due to the solubility of
atmospheric ammonia into acidic solutions (Tuovinen & Kelly, 1972).
This explanation is unconvincing when activities of microorganisms in
the natural milieu are compared with their nitrogen demand.
One of the possible means by which Thiobaailli can obtain nitro
gen essential for their growth can be through fixation of atmospheric
nitrogen. An early report (Mackintosh, 1971) suggested that some
Thiobaailli possess nitrogen fixing capabilities. Further data on
this observation have recently been reported using tracer techniques
employing 15N2 -enriched gaseous nitrogen as the sole nitrogen source
for Thiobaaillus ferrooxidans (Mackintosh,1976). The results obtained
by this experiment showed unequivocal accumulation of the
isotope in the bacteria. In the natural milieu, there is the possibi
lity that microorganisrns of different genera may exist together and
situationsin.which a nitrogen-fixing bacterium can coexist with
acidophilic Thiobaailli can be envisaged. In fact, recent studies

62
have shown that in a nitrogen-free medium ThiobaciZZus ferrooxidans is
capable of leaching nickel ores in the presence of the nitrogen
fixing Beijerinckia Zacticogenes, which apparently obtain its carbon
requirements from ThiobaciZZus ferrooxidans (Tsuchiya, 1977; Tsychiya
et al., 1974; Trividi & Tsuchiya, 1975). There is a great deal to be
learnt about suchmutualismeffects between the various species fowd
in a leaching heap.
Nitrogen-fixation could be one of the major agencies for provi
ding ammonium nitrogen and in a natural leaching system a number of
reactions, microbiological as well as abiological, leading to this
end can be envisaged (Schrauzer & Guth, 1976).
Phosphate.
The bulk supply of phosphorus required for phospholipids,
nucleotides and their derivatives is derived from inorganic phosphate,
and its limitation can seriously affect Co 2-fixation, growth and assi
milation of energy yielding substrates. Addition of phosphate to a
suspension of ThiobaciZZi enhanced substrate oxidation and Co2-fixat
ion (Beck & Shafia, 1964; Beck, 1969). The possibility of a further
role for phosphate arises from the suggestion by Dugan & Lundgren
(1965) that phosphate complexes with ferrous ion during initiation of
its oxidation to ferric ion by ThiobaciZZi.
However, inhibitory effects of phosphate on mineral degradation
have also been noticed (Silverman & Lundgren, 1959-a; Corrick & Sutton,
1965; Razzell & Trussell, 1963-a; Beck, 1969). Torma et al. (1970)
from their study on the effect of K2 HP04 on the leaching of ZnS concen
trate by ThiobaciZZus ferrooxidans concluded that concentrations of
K2HPO4higher than 0.5% did not increase the rate of leaching. One

63
of the important findings on the effect of phosphate on microbial
leaching is that the phosphate level influences the rate rather than
the total amomt of a metal released in a leaching process. Therefore,
in order to achieve a constantly fast rate in a leaching operation
constant addition of phosphate into the leaching system could be
profitable. However, the disadvantages of this replenishment in the
stream of nutrients during active leaching operations have been pointed
out by workers who observed that such additions in a system apparently
lowered the adsorption forces between the microorganisms and the ore
surfaces. They concluded that this effect could seriously affect
outputs ( Tuovinen et al., 1971; Ehrlich & Fox, 1967); but the results
of Le Roux et al. (1973) contradicted the earlier findings by showing
that contact between bacteria and minerals during leaching conditions
is very firm and could not possibly be interfered with under mild
conditions. The reason for the inhibitory effects of phosphate have
not be·en explained but it is conceivably due to the precipitation of
insoluble metal phosphates which render mineral surfaces passive to
bacterial action.
Sulphate.
Besides the biosynthesis of sulphur amino acids, sulphate has
been shown to play an important role in the growth of ThiobaciZZi and
in the activation of their enzyme systems (Lazaroff, 1963, 1975, 1977;
Beck, 1969; Tominaga & Mori, 1974). The demand for sulphate during
iron oxidation by ThiobaciZZus ferrooxidans has been confirmed by
investigations involving whole cells (Schnaitman et al., 1969; Tuovinen
et al., 1971) and cell envelopes (Barvinchak, 1975). Involvement of
sulphate as a complexing agent during the oxidation of ferrous iron
has been frequently demonstrated by a ntDnber of workers (Dugan &
Lundgren, 1965). Although certain other divalent ions, viz., HP04

64
HAs04 have been shown to be able to replace sulphate to some extent
(Schnaitman et al., 1969), latter studies suggest an obligatory requi
rement for sulphate ions by ThiobaciZZus ferrooxidans growing on
ferrous ions as the sole energy source (Bodo & Lundgren, 1971). In a
recent study on ThiobaciZZus feYTooxidans an uptake of sulphate was
observed in cell suspensions incubated in the presence of ferrous ion
(Tuovinen et al., 1975). Barbie (1977) has shown that ThiobaciZZus
ferrooxidans can withstand high concentrations of sulphate. as
compared to chloride and nitrate.
When ThiobaciZZus ferrooxidans is grown in energy sources other
than iron, such as sulphur, thiosulphate or metallic suphides, the
requirement for sulphate is no longer obligatory. It is thought that
in such cases they most probably fufil their sulphate requirement ·
from the sulphate produced .from the oxidation of sulphur or other oxi
disable sulphur compollllds.
No information regarding the sulphate demands of other ThiobaciZZi
is available in the literature.
Trace EZemen ts.
The need for trace elements by ThiobaciZZi has been demonstrated
in the case of non-acidophilic members of the genus. For acidophilic
members, such as ThiobaciZZus ferrooxidans & ThiobaciZZus thiooxidans,
it is a general belief that the trace elements needed can be derived
from the natural town-water supply or from the impurities of these
elements in the rocks which these microorganism usually degrade.
One trace element which has been investigated to some extent is magne
sium. It acts as a co-factor for many enzymatic systems of ThiobaciZZus
ferrooxidans ( Gale & Beck, 1967; Howard & Lundgren, 1970). It is
required in such low levels [usually l,OOOth of the sulphate

65
requirement (Tuovinen et al., 1971) that Tonna et al. (1970) were
unsuccessful in their attempts to demonstrate an additional need for
magnesilllD in the presence of zinc sulphide concentrate; the impurities
in the mineral were thought to supply the necessary level.
Effect of Dissolved Ions on the Growth and
Leaching Behaviour of ThiobaciZZi.
High concentrations of ions usually inhibit the normal activities
of microorganisms by affecting the water activity, but inhibition by
specific metal ions is also common. Mechanisms for inhibition by metal
ions have been reviewed by Sadler & Trudinger (1967). Generally,
ThiobaciZZi are more tolerant to high concentrations of metallic ions
than are the heterotrophic microflora and it is conceivable that their
inhibition mechanisms differ from those of other microorganisms.
ThiobaciUus ferrooxidans can tolerate exceptionally high
concentrations of metallic ions and the experimental data indicate that
highly metal resistant strains can be selected by simply transferring
a parent strain from lower to higher metal ion levels.
The substrates on which they are cultivated seem to affect the
tolerance response of ThiobaciZZi to metal inhibition. Thus, whilst
ThiobaciZZus ferrooxidans grown on sulphur, sulphide or ferrous ion,
is relatively tolerant towards a diversity of metal ions, low concen
trations of the same metal ions were found to be inhibitory when thio
sulphate was used as energy source (Tuovinen et al., 1971-a). The
sensitivity of CO rfixation to metal ions is greater than is
ferrous oxidation, according to the manometric observations of
Tuovinen and Kelly (1972). Some works have indicated that the
phenomenon of metal toxicity is complex and that metal salt
toxicity should be considered separately (Barbie & Bacilovic, 1975).
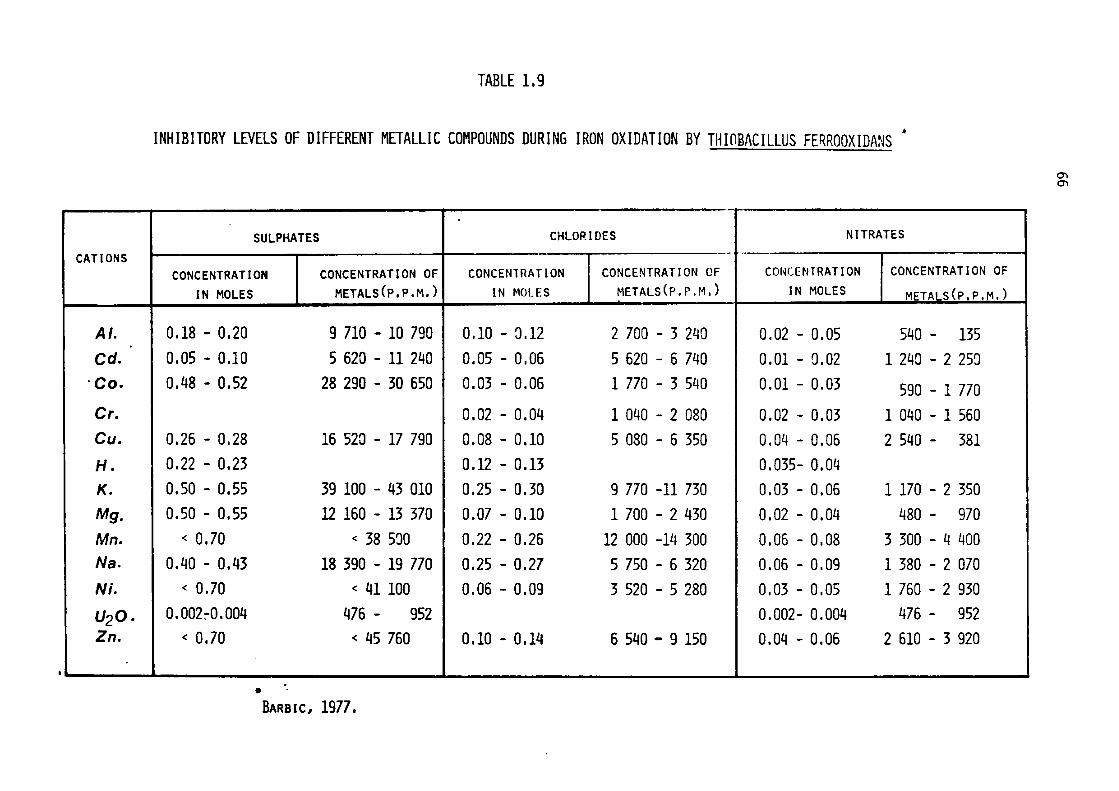
CATIONS
Al.
Cd.
·co.
Cr.
Cu.
H. K. Mg.
Mn.
Na.
Ni.
U20. Zn.
TABLE 1.9
INHIBITORY LEVELS OF DIFFERENT METALLIC COMPOUNDS DURING IRON OXIDATION BY THIOMCILLUS FER.ROOXIDANS ..
.. -
SULPHATES CHLORIDES NITRATES -
CONCENTRATION CONCENTRATION OF CONCENlRATION CONCENTRATION OF CONCENTRATION CONCENTRATION OF
IN MOLES METALS(P,P,M,) IN MOLES METALS(P,P,M,) IN MOLES METAi S(P,P,M,) -· 0.18 - 0.20 9 710 - 10 790 0.10 - :J.12 2 700 - 3 240 0,02 - 0.05 540 - 135 0.05 - 0.10 5 620 - 11 240 0.05 - 0.06 5 620 - 6 740 0.01 - 0.02 1 240 - 2 250 0.48 - 0.52 28 290 - 30 650 0.03 - 0.06 1 770 - 3 540 0.01 - 0.03 590 - 1 770
0.02 - 0.04 1 040 - 2 080 0.02 - 0.03 1 040 - 1 560 0.26 - 0.28 16 520 - 17 790 0.08 - 0.10 5 080 - 6 350 0.04 - 0.06 2 540 - 381 0,22 - 0.23 0.12 - 0.13 0.035- 0.04 a.so - o.s5 39 100 - 43 010 0.25 - 0.30 9 770 -11 730 0.03 - 0.06 1170 - 2 350 0,50 - 0.55 12 160 - 13 370 0,07 - 0.10 1 700 - 2 430 0,02 - 0.04 480 - 970
< 0.70 < 38 5::JO 0.22 - 0.26 12 000 -lll 300 0.06 - 0.08 3 300 - 4 400 0.40 - 0.43 18 390 - 19 770 0.25 - 0.27 5 750 - 6 320 0.06 - 0.09 1 380 - 2 070
< 0,70 < 41100 0.06 - 0,09 3 520 - 5 280 0,03 - 0.05 1 760 - 2 930 0.002:-0.004 476 - 952 0.002- 0,004 476 - 952
< 0.70 < 45 760 0.10 - 0,14 6 540 - 9 150 0.04 - 0,06 2 610 - 3 920
• BARBIC, 1977.
en en

67
The inhibitory levels of some metal ions and cations on iron
oxidation by ThiobaciZZus ferrooxidans are tabulated in Table 1.9.
The increased concentrations of metal ions which develop during
a natural leaching situation may be responsible for upgrading the metal
tolerance of ThiobaciZZi (Fletcher, 1970, 1971-a) and tolerance towards
a metal previously toxic has been found to develop rapidly in Thio
baciZZus ferrooxidans (Zinnnerley et al., 1958). Concentrations as
high as 72 g/t of zinc ion have been achieved in stirred continuous
leaching operations using ThiobaciZZus ferrooxidans, without any dele
terious effects upon microbial acitivity (Torma at al., 1970; Gormely
et al., 1975).
The effects of heavy metal ions on the growth & iron-oxidising
activities of ThiobaciZZus ferrooxidans have recently been studied by
Imai et al. (1975) who have found that high levels ( 10- 2 -10- 3 M) of
cupric, zinc, cadmium and chromium ions had no significant toxic effects;
lead was found to delay the lag period but no inhibition of growth was
evident. Levels of mercurous, mercuric and silver ions (l0-3M) have
been found to be toxic and to produce 100% inhibition of growth and
iron oxidation by ThiobaciZZus ferrooxidans by these workers. In an
endeavour to study the mechanism of metal inhibition, Imai et al.
(1975) employed cell free extracts and found that the activity of
cytochrome oxidase ( cytochrome a 597) of the iron oxidising system
was specifically inhibited by 5 x 10- 4 M mercuric ions. A similar
finding was reported by Lundgren (1975) after investigation of AgN0 3
inhibition of the iron oxidising system of ThiobaciZZus ferrooxidans.
Silver ion_toxicity towards ThiobaciZZus ferrooxidans has been
recently studied by Hoffman & Hendrix (1976) who showed that silver
ions when supplied as silver sulphate, were extremely toxic; concen-
trations greater than 1 p.p.m. of silver inhibited completely the

68
growth and iron oxidising capabilities of this species.
Tuovinen & Kelly (1974, 1974-a, b) noticed severe plasmolysis
in the culture of Thiobacillus feYTooxidans exposed to uranyl ions.
However, relief of uranyl ion inhibition by divalent ions led
Tuovinen& Kelly to assume that uranyl ions inhibited the transport
system of cells by binding to specific surfaces in the cells, as
has been the case with yeast cells (Rothstein, 1954; Maxwell et al.1971).
Tolerance of Thiobacillus ferrooxidans towards uranium is low
but selective subculturing has resulted in the development of strains
which are capable of withstanding a significant amount of uranium in
solution ( Duncan et al., 1967; Duncan & Bruynesteyn, 1971). Toxicity
of uranium ions towards Thiobacillus ferrooxidans can be ameliorated
by monovalent & divalent cations ( Tuovinen & Kelly, 1974-a, b).
Ebner & Schwartz (1974) reported a high population of a uranium-tole
rant strain of Thiobacillus ferrooxidans which was capable of proli
ferating in a maximum concentration of 1 g uranium per litre ( as
U0 2S0 4 ). The most interesting finding of these workers was that the
Thiobacillus thiooxidans could tolerate significantly higher amol.lllts
of uranium ( 12.5 g uranium per litre). Previously it was commonly
believed that Thiobacillus thiooxidans species was comparatively less
resistant to metallic ions than was Thiobacillus ferrooxidans.
Schwartz's group also has demonstrated for the first time that
each heavy metal ion, when present together with other heavy metal ions,
affects the bacterial activities as if it were present alone; and
that the maximum tolerance value for each ion is independent, i.e., a
combined toxicity effect is not observed. These findings have been
confirmed by Barbie (1977).
Generally, anions are more toxic than cations. Ferrous oxidation
by Thiobacillus ferrooxidans has been found to be hampered by a number

69
of anions. Fluoride, molybdate, cyanide and azide ions are most
deleterious, even in small concentrations (less than 0.25 mM)
(Andersen & Lundgren, 1969; Tuovinen & Kelly, 1972). Barbie (1977)
has shown that Thiobacillus ferrooxidans is most tolerant of
sulphate, less of chloride and least of nitrate; the relative toxicity
of sulphate, chloride and nitrate is 1:3:10.
Concentrations of chlorides above 6,000 p.p.m. and nitrate more
than 2,000 p.p.m. retarded iron oxidation by iron-oxidising bacteria.
It is worth noting, however, that Le Roux (1977- unpublished informa
tion made available to Professor B.J.Ralph) has shown that for other
strains of Thiobacillus ferrooxidans chloride inhibition is not shown
until levels of 6,000 p.p.m. chloride ions were reached and the bacte
rial population remained viable at concentrations of chloride ions up
to 8,000 - 9,000 p.p.m. Further recent work in Department of Micro
biology, University of Cardiff, have indicated that some Thiobacillus
ferrooxidans can be acclimatised to sea water media with reduced but
still substantial growth rates (Private communication to Professor
B. J. Ralph, 1977). Similar claims were made by Mayling (1966) in a
patent where iron-oxidising bacteria were grown on ferric chloride
solution. These findings also explain the anomalous behaviour of
Thiobacillus ferrooxidans strains during the covellite oxidation
observed by Vanselow (1976) and Golding et al. (1977) in a chloride
rich media. The iron- oxidising bacteria which these workers emplo
yed was isolated from a marine mud (Corrans, 1970).
Relatively little is known about anion toxicity in Thiobacilli
and the biochemical basis of both anion and cation inhibition in
Thiobacilli has not been studied extensively. Such studies should
be emphasised in the future, since the mechanism of metal toxicity
may open the door to our understanding of pollution control problems.

70
Effect of Organic Compounds.
Organic compounds normally capable of acting as nutrients for
heterotrophic microorganisms are known to inhibit growth and meta
bolic activity of several species of chemolithotrophic nitrifying
bacteria and some species of the genus ThiobaciZZus. Smith & Hoare
(1977) have recently reviewed the influence of organic compounds on
growth of lithotrophs. The acidophilic ThiobaciZZi, i.e., ThiobaciZZus
ferrooxida:ns and ThiobaciZZus thiooxidans are adversely affected
by the presence of organic matter in the medium (Tuttle & Dugan, 1969).
Pyruvate or oxaloacetate exceeding a level of 2 - 4 x 10- 4 Min a
medium containing elemental sulphur prevented the growth of ThiobaciZZus 2
thiooxidans (Borichewski, 1967; Rao & Berger, 1970). 10- M ~-alanine
has also been shown to inhibit growth of this species (Butler &
Umbreit, 1966).
Organic compounds such carbohydrates, carboxylic acids, anionic
detergents, cationic surfactants, quaternary ammonium compounds, yeast
extracts, peptone and amino acids have frequently been reported to have
deleterious effects upon iron oxidation or growth,or both, by Thio
baciZZus ferrooxidans (Dugan & Lundgren, 1964; Duncan et al., 1964;
Leathen et al., 1956; Silver et al., 1967; Silverman & Lundgren, 1959-a),
and inhibition of sulphur oxidation by primary alcohols, dicarboxylic
acids and protein hydrolysate has been reported by Kelly (1971) and
Landesman et al. (1966-a).
Conversely, there are reports that ThiobaciZZus thioparus and
ThiobaciZZus thiooxidans are not noticeably inhibited by organic matter.
In some instances, their growth is reported to be accelerated by the
addition of organic matter to their media (Butler & Umbreit, 1966;
Starkey, 1934; Matin & Rittenberg, 1971). Increased rates of oxidation
of sulphur by ThiobaciZZus thiooxida:ns in the presence of dicarboxylic

71
acids (Vogler et al., 1942) and an enhancement in the rate of oxida
tion of thiosulphate by ThiobaciZZus thioparus when TCA cycle inter
mediates, viz., malate, succinate and fumarate, were present in
concentrations ranging from 10- 3 - 10-1 M, have also been reported
(Vishniac & Santer, 1957).
Shafia & Wilkinson (1969) and Tabita & Lundgren (1970,1971) have
succeeded in cultivating ThiobaciZZus ferrooxidans in a glucose-ferrous
sulphate medium and they reported that glucose was dissimilated via
the Entner-Doudoroff pathway. The presence of enzymes of carbohydrate
metabolic pathways have also been detected and studied in other members
of the ThiobaciZZus genus, such as ThiobaciZZus perometaboZis, Thio
baciZZus intermedius, and ThiobaciZZus ferrooxida,ns (Matin & Rittenberg,
1970; Tabita & Lundgren, 1971-a, b). In order to investigate the
mechanism of metabolic regulation in facultative ThiobaciZZi, hetero
trophic growth of ThiobaciZZus A2 on labelled sugars and organic
acids has been studied; presence of enzymes essential to the operation
of three major catabolic pathways ( the Embden-Meyerhof glycolytic
pathway, the Entner-Doudoroff pathway and oxidative pentose phosphate
pathway) in ThiolxzciZZus A2 grown on glucose and other sugars has
been detected (Wood & Kelly, 1976; 1977; Wood et al. 1977).
Physiological and biochemical changes in the transition from
autotrophy to heterotrophy and vice versa may be explained in terms
of the induction,, repression and depression of enzymes demonstrated
with ThiobaciZZus noveZZus (McCarthy & Charles, 1974). Since the
identification of ThiobaciZZus feYTooxidans is invariably based on
its capability of oxidising ferrous-iron, the inability of heterotro
phic ThiobaciZZus ferrooxida,ns to grow autotrophically on ferrous
iron (Shafia & Wilkinson, 1969) casts doubts over its taxonomic
position. Earlier works have considered the possibility of original
heterogeneous cultures(Tuovinen & Kelly, 1972; Zavarzin, 1972) which

72
has been recently demonstrated by the isolation of Thiobacillus
acidophilus from a culture of Thiobacillus ferrooxidans (Guay & Sil
ver, 1975). A recent observation made by Tuovinen & Nicholas (1977)
suggests that the reputedly obligately heterotrophic Thiobacillus
ferrooxidans KG-4, cultured on glucose, contained a small proportion
of cells which grew autotrophically on ferrous iron. The current data
also suggest that some cultures areheterogeneous on the basis of their
DNA base composition depending on their growth substrate (Guay et
al. , 1975 & 1976). The close association of a mixed population indi
cates a very close metabolic coupling and interdependence of acidophi
lic Thiobacilli (Tuovinen & Kelly, 1978).
In practical leaching situations, organic levels are usually
very low and provide an ideal situation for the activities of autotro
phic microbial populations. In certain cases, however, where sewage
waters have been used for irrigating the heaps, leaching rates have
been found to decrease (Zajic & Ng, 1970), probably because of inhibi
tory effects of organic residues present in the sewage.
At present there is a growing trend towards the recovery of metals
from the leach-liquor by solvent extraction processes (Itzkovitch et
al., 1974) during which organic reagents dissolved in organic solvents
are mixed with leach solutions and the phases then separated. The
aqueous phase from which most of the metals have been removed are
recycled. Recently, it has been shown that residual organic solvents.
in these spent liquors exert an inhibitory effect upon the bacterial
leaching of chalcopyrite and should be removed by treatment with
activated charcoal prior to recycle (Itzkovitch & Torma, 1976; Torma
& Itzkovitch, 1976). These findings have been confirmed (Dr. John F.
Madgwick- personal communication).
So far there is no single common explanation for anomalies in
reports on the effects of organic compounds on acidophilic

73
ThiobaciZZi. However, it is suggested that a major factor contributing
to the inhibitory effects on iron oxidation may be the relative
electronegativity of the organic molecules (Tuttle & Dugan, 1976) and
that organic compotmds may inhibit iron oxidation in the following
manner:
1- Directly affecting the iron oxidising systems.
2- Reacting chemically with iron compounds.
3- Reacting with phosphate and sulphate and thus influencing
their roles in iron oxidation.
4- Interfering with the transport systems of cells by
disrupting cell membranes or envelopes; leakage of
cellular material from ThiobaciZZus ferrooxidans by
cell envelope disruption in the presence of organic acids
supports this hypothesis (Tuttle et al., 1977).

74
@
three.fold
@
Fig. 1.3 STRUCTURE AND AT(I.UC ARRANGEMENT OF ZnS.
(A)- SPHALERITE. (B)- WURTZITE.
(Berry & Mason, 1959)

75
1.4 BIOGEOCHEMISTRY OF ZINC SULPHIDE
Zinc sulphide (ZnS) minerals occur in nature in two polytypic
forms cubic (sphalerite) and hexagonal (wurtzite). Zinc, like cadmium
and mercury, always forms divalent sulphides. Zinc disulphide, ZnS2 ,
which is formed at very high pressures, also retains this divalent
nature of the cation (Bither, 1967; Bither et al., 1968).
The most common form in which zinc sulphide occurs in nature is
sphalerite,which has face-centered cubic stacking of the zinc sulphide
tetrahedral layers. Many specimens are mixtures of the two varieties
with predominance of the cubic type. Sphalerite is the stable form at
ordinary temperatures and, on heating to 1020°C, undergoes molecular
rearrangement resulting in the hexagonal stacking of ZnS layers to
form wurtzite:
1020 °C Sphalerite Wurtzite
The sphalerite - wurtzite inversion has been folllld to be pressure
and temperature dependent (Majumdar & Roy, 1965). The two types differ
only in the stacking pattern so it is possible to change from one to
the other by introducing a stacking fault. Several stacking sequences
longer than ABAB (hexagonal) or ABCABC (cubic) have been noted by
Smith (1955).
The main characteristics of these zinc sulphide forms are
described in the Table 1.10.
The structure of sphalerite (Fig. 1.3a) is analogous to that of
diamond, with four zinc atoms at the points of a face-centred cubic
lattice (comparable to four of the carbon atoms in diamond)and four
atoms of sulphur (comparable in position to the other four atoms of

76
carbon of diamond) at the points of a face-centred cubic lattice
displaced one quarter along the body diagonal of the first cube. In
sphalerite, each zinc atom is coordinated with four sulphur atoms and
each sulphur with four zinc. These tetrahedra of SZn4 are all oriented
in the same way, with a triangular face of the tetrahedra parallel to
the 111 face, and the ZnS4 tetrahedra are all oriented in the opposite 0
way. The atomic distance between adjacent cations, Zn2 +, is 5.406 A
Several minerals such as chalcopyrite, CuFeS2 , are structural
analogues of sphalerite.
TABLE 1.10
CHARACTERISTICS OF MAIN CRYSTALLOGRAPHIC FORMS
OF ZINC SULPHIDE#
PROPERTY
Crystal System
Space Group *
Crystal Class
Axial Elements
Lattice
Cell Dimensions (Pure ZnS)
Cell Content
Density
ZnS per cc.*
Zn - S *
s - s *
SPHALERITE
Cubic
216; 1 4 3 m
F 0
a= 5.406 A
Z = 4
4.096
2.52. 1022 0
2.34 A 0
3.82 A
WURTZITE
Hexagonal
186; c4 5V
6 mm
a:c = 1:1.638
p 0
a = 3 .82 A 0
C = 6 .26 A
Z = 2
4.086
2.52 • 1022 0
2.34 A 0
3.82 A
# Data taken from Berry & Mason, 1959, with the exception of*
* Data taken from Shuey, 1975.
Fig 1.3b shows the structure of wurtzite; the distance between
the adjacent zinc atoms in this crystallographic form of zinc sulphide
• 0
1s 3.82 A along a and b axes. In the hexagonal type the zinc and
sulphur atoms are on a similar lattice but zinc atoms are displaced

77
along the c axis. Along the c axis the distance between the adjacent 0
cations increases to 6.26 A.
In a purely ionic model, ZnS should consist of Zn2 + with a full
3d shell and s2 - with a full 3p shell. Direct covalence between
S - S atoms can be neglected due to the S - S distance being larger 0 0
than the ionic diameter (3.68 A). The Zn - S distance (2.34 A) is 0
distinctly less than the sum of ionic radii (2.58 A), and covalence
may occur by combination of Zn 4sp3 and S 3sp3 tetrahedral orbitals
reducing the effective charge on the atoms.
Due to the similarity in the ionic radii of zinc and iron atoms,
iron can occasionally replace zinc in the lattice of zinc sulphide
crystals resulting in the iron-containing sphalerite called' marmatite '
This substitution of iron in sphalerite has been found to be tempe
rature dependent and the iron contents in sphalerites are often used
as a geological thermometer, indicating the temperature of formation
of rocks (Korostelev & Chainikova, 1966; Kullerud & Neumann, 1953).
However, some discrepancies in the data relating percentage of iron in
sphalerite and temperature of formation have been pointed out, chall
enging the validity of the relationship (Sanzonov, 1961;
Cleveland, 1964).
Applications of ZnS in photoelectronic devices stimulated the
comprehensive study of atomic defects in ZnS. The results of these
investigations have been reviewed by Aven and Prener (1967), Ray (1969)
and Shuey (1975). The major effect of isoelectronic substitution is
a shift of the energy bonds. Thus when Cd replaces Zn, the conduction
band is lowered, whereas substitution of Se for S raises the valence
band. Copper, silver, gold, sodium and potassium in the lattice act
as acceptor defects. Transitional metal ions substituted for zinc
can in general act as donors or acceptors. Nickel, cobalt and iron

78
are known as 'killers' of the luminescence in ZnS commercial phosphors,
due to their similar behaviour when substituted in ZnS. Manganese is
apparently different and the paramagnetic resonance of Mn2+ is easily
observed and no evidence of another charged state of Mn in ZnS is ava
ilable . Cadmium and manganese are noticed to have no effect on elect
rical conductivity of ZnS whereas iron can produce free carriers.
The various proportions of these cations, especially iron, affect
the surface characteristics of the minerals and markedly influence the
flotation properties of sphalerites. It has been reported that marma
tite can be floated more easily than pure zinc sulphides (Chainikova,
1962). Hardness, reflectance, density and lattice parameters of
sphalerite crystals have been found to be influenced by chemical
composition of the minerals. Substitution by Fe, Mn, and Cd affects
the crystal dimensions as well as physical properties of the sphalerite.
It has been found that the one-unit cell edge size of synthetic sphale
rite bearing Fe, Mn and Cd is a linear function of the composition
expressed in mole% (Skinner, 1960; Hall, 1961; Takimoto et al.,1960).
Definite mathematical correlations between these properties and
mole percentages of Fe, Mn and Cd in these minerals have been proposed.
(Pudovkina et al., 1968). The lattice parameter 'a' of a substituted
crystal is given by the relationship: 0
a= ( 5.3985 + 0.0006 X + 0.00147 y + 0.00232 z) A
where x, y, and z are percentage mole fractions of Fe, Mn and Cd resp
ectively present in the sphalerite specimen. Similarly the hardness
(H) of sphalerite has been found to be decreased by substitution of
the elements in the lattice. The following relationship has been
found to hold for the hardness of sphalerite specimens:
H = ( 3 Zn - 2.5 Fe - 58 Cd) kg/nnn2
The presence of these cations in otherwise pure sulphide samples has

79
been fomd to affect the chemical leaching behaviour of the minerals
(Scott & Dyson, 1968).
1.4.1 MICROBIOLOGICAL OXIDATION OF ZINC SULPHIDE
Unlike copper, iron and manganese, zinc does not exhibit variable
valency states in its common compomds, e.g., sphalerite, wurtzite and
zincite, and is unable to provide energy to oxidative mcroorganisms.
Most probably, the divalent zinc ions found in its common minerals are
not attacked per se by these microorganisms. The sulphidic moiety
present in zinc sulphides ( sphalerites, wurtzites and marmatites),
however, is easily oxidisable by microorganisms to sulphate and at
low enough pH sulphate ions are released into solution.
Scott & Dyson (1968) reported that chemical oxidation of zinc
sulphides at high pressures ( ~1700 kPa) and at elevated temperatures
(113 °C) takes place according to the following equations:
ZnS
ZnS
Reaction [2] can take place in the absence of acid while an acidic
environment is necessary for reaction [l]. Both reactions proceed
extremely slowly with pure zinc sulphides.
[l]
[2]
An electrochemical model has been proposed by Pawlek (1969) in
which zinc sulphide reacts anodically according to the reaction:
ZnS -+ Zn2 + + S0 + 2 e [3]
The cathodic reduction of chemisorbed oxygen, the presence of which
may vary the number of charge carriers within ZnS owing to its semi
conduction properties, occurs according to the reaction:
[4]

80
The overall kinetics could not be explained on the basis of electronic
conduction alone, indicating that other rate controlling processes were
also involved. One of the factors involved was found to be H2S, which
is formed during the leaching of ZnS and its continual removal was
found to increase the leaching rate approximately 4-fold , while its
addition at a pressure of 2 atmospheres completely inhibits the leach
ing reaction. This is due to the equilibrium in the reaction:
[5]
The removal of H2 S from a system may proceed by one of the following
reactions; of which reaction [6] takes place in highly acidic solutions,
+ 2 S 0
+ so~-
[6]
[7]
[8]
and reaction [8] apparently occurs at higher temperatures. The middle
reaction [7] is very slow; this reaction can be catalyzed by activated
charcoal, thereby increasing the leaching rate of zinc sulphide by a
factor of 5 (Pawlek, 1969).
Sphalerite (ZnS) and marmatite are the only zinc sulphides of
any commercial significance and their dissolution behaviour in acidic
ferric solutions and in acidic bacterial solutions containing iron has
been widely investigated. Sphalerite is found to be leached at signi
ficant rates by acidic ferric sulphate solutions at temperatures
ranging from 80 - 100°C. The general equation for this dissolution is:
ZnS [9]
The reaction at high temperatures is rapid and the elemental sulphur
produced during oxidation does not impede the rates of dissolution.
The rate controlling step in ferric leaching appears to be the
availability of trivalent ferric ions since the dissolution rate of

81
sulphides is directly proportional to the amount of ferric ions pres
ent. Initial concentrations of zinc sulphate ( 0 - 80 g/i) and
ferrous sulphate ( 0 - 60 g/i) have no significant effect on the
leaching. However, acid concentrations are critical to the extent of
+ leaching and an increment in the acidic content from 2 to 20 g of H
per litre depressed the amount of zinc dissolved from 75 to 50 %
(Dutrizac & MacDonald, 1974).
When ferric ion is used for leaching zinc sulphide, ferrous
sulphate and elemental sulphur are produced in addition to ZnS0 4 , as
shown in equation [9]. Under acid conditions, oxygen can convert the
reduced ferrous species to ferric state where it can be reused in the
system:
[10]
Zinc sulphide was probably the firstsulphidic mineral to be
oxidised microbiologically in the laboratory for the recovery of zinc
metal (Rudolf & Helbronner, 1922). Since that time many workers have
studied the bacterial oxidation of sphalerite ores. Zimmerley et al.
(1958) claimed, in a patent issued to them, that with ThiobaciZZus
ferrooxidans zinc sulphide was solubilised to give a concentration of
17 g/t . Zinc concentrations up to 25 - 50 g/t have been obtained by
various workers (Marchlewitz & Schwartz, 1961; Silverman & Ehrlich,
1964; Moss & Andersen, 1968) and more recently semi-industrial studies
performed at the University of British Columbia with zinc sulphide
concentrate resulted in a very fast zinc extraction of about
1300 mg/(i.h) (Gormely et al., 1975) and a dissolved zinc concentration
as high as 120 g/i (Torma et al. 1972).
In the presence of the iron oxidising bacterium ThiobaciZZus
ferrooxidans, oxidation of ferrous iron at acidic pH is catalyzed and
proceeds at a much faster rate, consequently accelerating reaction [10]

82
in the forward direction. The elemental sulphur produced during
reaction [9] is also oxidised by these bacteria to produce sulphuric
acid:
so [11]
Since ferric sulphate contributes so significantly to the leach
ing of metallic sulphides it is possible that some of the early results
on bacterial leaching included an indirect chemical ferric leaching
component. Indeed a number of investigators, including Ivanov (1962,
1962-a) and Zirnrnerley et al. (1958), stated that the only part played
by the bacteria in the leaching process is the regeneration of ferric
iron from the ferrous iron produced during reaction [9].
It has been recently established that microorganisms are capable of
attacking zinc sulphide minerals directly according to the following
reaction:
ZnS Bacteria [12]
and significant rates of zinc leaching have been obtained by optimizing
the conditions of growth of bacterial populations (Duncan et al., 1966,
1967, 1967-a; Duncan & McGoran, 1971). Enhancement of leaching rates
by optimizing the microbial growth indicates that direct bacterial
attack on ZnS is significant. Duncan et al. (1967-b) also suggested
that attack on the sulphur moiety is the primary mechanism during
bacterial degradation. This view is also supported by Beck & Brown
(1968). Torma (1971), who leached pure synthetic zinc sulphide with
Thiobacillus ferrooxidans, reported reasonably high leaching rates
of 1150 mg/(t.h). The initial increase in the pH of the medium, noticed
by Torma during the bacterial leaching of the pure synthetic zinc
sulphide, was attributed to the consumption of acid as predicted from
the proposed oxidative mechanism on ZnS indicated in reaction [l].
Ferric iron, as stated earlier, is a good leaching agent, at

83
80-100°C, for sulphide minerals but its efficiency has been found to
be reduced in conditions which are necessary for rapid growth of
bacteria. It has been shown that addition of ferric ion at 35°C to
leaching experiments had no effect on the rate of release of copper
from covellite or zinc from marmatite (Duncan & Walden, 1972). The
data presented by these workers show that, during the microbial leach-
ing of chalcopyrite, ferric iron inhibited copper release. During
their investigations they noticed that iron precipitated out in spite
of the fact that the pH was low enough to prevent precipitation of
ferric hydroxide according to the equation [13].
+ -+ 2 Fe(OH) 3 + [13]
The iron was found to precipitate in the form of basic ferric
sulphates, frequently found in the form of jarosite-minerals (Jennings
et al., 1972; Torma & Legault, 1973; Rossi, 1974):
3 Fe (SO) + 12 H 0 2 4 3 2
-+ 2 HFe3(SO4) 2(OH) 6+5 H2so4 [14]
Jarosite
Hydronium ion in the jarosite can be replaced by the cations sodium,
potassium and ammonium etc.
After 60% of the metal in the leaching reactor has been leached,
copious amounts of these precipitates covered the mineral surfaces
thereby inhibiting the reaction. In certain cases these precipitates
were found to completely preclude the further leaching of the mineral
by forming a barrier between the mineral surface and bacteria ( March
lenwitz & Schwartz, 1961; Guay et al., 1975). Beck (1967) also reported
that if iron-rich solutions are used for dump leaching there exists the
danger that jarosite will precipitate locally and plug the dump ,thereby
greatly reducing the leaching efficiency. Exposure of new surfaces by
regrinding the leach-residues has frequently been suggested as an

84
effective remedy for the inhibitory effects of jarosite precipitation
in laboratory experiments (Torma, 1972; 1976). Methods involving one
stage(Torrna, 1975) and two-stage regrinding process have been described
(Duncan & Bruynesteyn, 1971).
Since the 9K basal salt medium of Silverman and Lundgren (1959)
contains only potassium and ammonium cations it is most probable that
jarosite formation can strip these from the medium. It has been
demonstrated that, during jarosite formation in the presence of
ThiobaciZZus ferrooxidans, both potassium and ammonium cations were
made insoluble (Duncan & Walden, 1972). These experiments indicated
that the ammonium concentration in solution was decreased from 800
mg/t to 100 mg/t, suggesting that medium may have become nitrogen
limiting for bacterial growth.
Inhibition of leaching by iron precipitation can be avoided by
using sulphur oxidising bacteria instead of the commonly employed
iron-oxidisers. The sulphur oxidising bacterium ThiobaciZZus thio
oxidans can oxidise the sulphidic moiety of zinc sulphide, producing
sulphuric acid. Since this bacterium is incapable of oxidising
ferrous iron this remains in solution; at pH values below about 4.5
the chemical oxidation of the ferrous iron is comparatively slow.
The only reason for not using this bacterium in commercial leaching
operations is its incapability of tolerating high concentrations of
metallic ions. However, recent work has indicated that ThiobaciZZus
thiooxidans can tolerate high amounts of zinc ions in solution
(20,000 - 30,000 mg/t)
At the present time, information on the leaching of zinc sulphide
concentrates by ThiobaciZZus ferrooxidans is available. Much of the
information describes the optimum cultural conditions and other related
parameters, such as particle size, surface area, nutrient level etc.,
likely to influence the extraction of zinc. This information has

been reviewed previously in section 1.3.3.
1.4.2 EFFECT OF CRYSTAL AND IATTICE STRUCTURE ON RATES
OF LEACHING OF ZINC SULPHIDE
85
It has been reported (Trussell et al., 1964) that marmatite
was completely and rapidly leached (100%) by ThiobaciZZus ferrooxidans
whereas sphalerite was relatively recalcitrant and leached only to the
extent of 7 - 11% There appears to be no systematic study reported
in the literature on the effects of crystallographic forms of zinc
sulphide upon leaching rate. How substitution of ions of different
species in the crystal lattice of ZnS affects its leaching behaviour
is also not known with certainty. The behaviour of the marmatite
forms of sphalerite differ markedly in their chemical leaching
behaviour and in flotation characteristics from pure zinc sulphide
mineral and it might be anticipated that similar effects would be
observed during bacterial leaching, as suported by an observation
by Trussell et al. (1964).
Scott & Dyson (1968), while investigating the effect of various
catalysts on the pressure leaching behaviour of zinc sulphide, found
that several metals in solution catalyzed the release of zinc, increa
sing in catalytic activity in the order Fe > Mo> Ru> Bi > Cu.
Copper and ruthenium were found to have appreciable catalytic activity
in the absence of acid. Although many other ions in addition to those
listed have the correct ionic radii to exchange with zinc in the zinc
sulphide lattice (e.g., Ag, Hg, Pb, Sn) these did not influence the
kinetics of dissolution. The effect of the catalysts was attributed to
the change in the electrical conductivity caused by penetration of

86
these ions into the lattice, thus pennitting electrochemical dissolu
tion to occur according to equation [3]. The specific catalytic
activity of Fe, Mo, Ru, Bi, and Cu was attributed to the effect of
the catalyst on the reduction of oxygen at active surface sites.
There is some limited evidence to suggest that the crystalline form of
minerals does significantly influence rates of oxidative breakdown
under purely chemical conditions. This fact is clearly shown with the
a and the Sforms of chalcopyrite (Ferreira & Burkin, 1975); microbiol
ogical analogies to this behaviour can be cited from the studies of
Silverman & Ehrlich (1964) on the behaviour of the two crystalline
forms of FeS2 (pyrite and marcasite).
Some of the work described in this thesis throws further light
on the influence of crystal structure and lattice substitution upon
leaching rates in the case of various zinc sulphide minerals with
different species of ThiobaciZZi.

87
1.5 AIMS OF THE CURRENT INVESTIGATION
It is well established that the acidophilic iron oxidiser,
ThiobaaiZZus feYTooxidans, can oxidise commercial and pure zinc
sulphides and there can be little doubt regarding the signific
ance of this ThiobaaiZZus species in bacterial leaching systems
at low pH. One of the aims of the current investigation was to
determine whether other ThiobaaiZZus species ( e.g., ThiobaaiZ
Zus thioparus and ThiobaaiZZus thiooxidans) can oxidise zinc
sulphides significantly and to establish a comparison, under
similar conditions of surface area, cell-density and temperatu
re, between the behaviour of such species. In order to study
these effects it was essential that the purity of species empl-
oyed be ascertained with utmost precision. Therefore the
three isolates were purified and characterised.
Nature of crystal structure and types of bonds existing
between different atoms in the crystals of minerals are princi
pal factors in controlling the properties and leaching behavio
ur of these minerals. Occurrence of zinc sulphide in two well
defined crystalline structures, i.e., wurtzite and sphalerite,
prompted investigation of leachability of these zinc sulphide
minerals with the three species of ThiobaaiZZus.

..
. I
I r.
..c • @·
r
.. !-,,
·, ~,
;-'
' , , ~ .
t~
)' .
I
~>
• .
. ... . ~.
"· : I~;
.......
• >\:
.. it,~
.,
•
;
. ' I\,
..
"
; '
' .. ' .
1 ~
•it : r
" f.,';
... -~ .
..~ ' ·~ • t • . ,
,._...:.wt, . . .•
'i
«, . !I
~
.. ... ,,,r.
.;; >.
' ....
!;_;> ..... ·r 1..
. ... . .,. j
' .. 1
,,
"' \
~:
' ,. .. ,¥' .. ~..,..·.
~ '
',
'f'
" I
·, J
. .. I·~~-' · .. , .;A;N.D
•• f',
"""
. .,. .. .....
' . -• r
-1,
. '~·
>.

88
2.1 PREPARATION OF SULPHUR SUBSTRATES
A number of sulphur compounds, viz. polythionates and sulphides
etc., were required in these investigations and most of them are not
commercially available. They were prepared as required.
2.1.1 POLYTHIO NATES
The polythionates were prepared as the potassium salts because
of ease of crystallisation and purification.
Potassium trithionates and pentathionates were prepared by the
method of Stamm & Goehring (1942) and Goehring & Feldman (1948)
respectively as modified by Roy & Trudinger (1970). Potassium tetra-
thionate was synthesised by the method of Trudinger (1961), using
iodine and sodium thiosulphate.
2.1.2 SYNTHETIC ZINC SULPHIDE
Attempts were made to prepare synthetic zinc sulphide by
direct combination of metallic zinc with a small stoichiometeric
excess of elemental sulphur. The reaction was carried out in an
evacuated tube at 800 °C but in a number of experiments completion of
the reaction was never achieved. It is now known that this method
requires the application of pressure for complete reaction and anneal
ing (Personal discussion with Mr W. M. B. Roberts, Bureau of Mineral
Resources, Canberra;and Roberts, 1965.)
Preparation of synthetic zinc sulphide was effected by precipi
tation, with gaseous hydrogen sulphide, from M/2 zinc sulphate
solution, which had been saturated with ammonium sulphate and made
alkaline with ammonia. The precipitate was thoroughly washed with
hot water and dried at 105° C overnight. The reaction was very slow

89
and gave extremely poor yields (10-15%). Zinc sulphide produced by
this method exerted enormous pressures and exploded the sealed tubes
during attempted annealing at 800° C.
In a view of these drawbacks, a synthetic zinc sulphide which was
prepared by Ajax & Co. was used instead. This zinc sulphide assayed
(AAS) at Zn 66.2% (theoretical 67.1%), and contained small amounts of
iron (0.01%), copper (0.005%) and nickel (0.005%) [See Table 3.3.2
section 3.3.2].
2.1.3 NATURAL ZINC SULPHIDE MINERALS
Two sphalerites (from Oklahoma and Spain) and a triboluminescent
wurtzite with some sphalerite (from Beaver County, Utah) were obtained
from Wards Natural Science Establishment Inc., and a marrnatite from
Zinc Corporation Ltd., Broken Hill, New South Wales.
2.1.4 PREPARATION OF ZINC SULPHIDE SUBSTRATES
The zinc sulphide mineral samples (moisture content 0.10%)
were ground in an agate mortar to pass 400 mesh screen. They were
stored in air-tight, screw-top jars until required.
The specific surface area of each sample was determined by the
B.E.T. method (Brunauer, Emmett & Teller, 1938) with the assistance of
Mr G. Roach of School of Metallurgy, University of New South Wales.
Krypton was used as absorbent gas and specific area was calculated by
a computer programme, supplied by Mr Roach, which analysed the experi
mental data using a least squares method. Specific surface area of
zinc sulphide minerals is presented in Table 3.3.3., section 3.3.5.
Each mineral sample was analysed for principal elements by
atomic absorption spectroscopy (see section 2.2.5) and for minor
components by semiquantitative emission spectroscopy (see section 2.2.4).

90
2.2 ASSAY PROCEDURES
2.2.1 THIOSULPHATE
The residual sodium thiosulphate in fermentation broths was
estimated either iodometrically or by the spectrophotometeric method
of Sorbo (1957).
Iodometeria Estimation of ThiosuZphate (Roy & Trudinger, 1970).
This well known method involves iodometric titration in 5% (v/v)
acetic acid, with thyodene (Purkis, Williams Ltd., 60 Brewery Road,
London, N. 7) as the indicator.
The sample was centrifuged immediately at 10,000. gin a
Sorval RC-2B centrifuge for 10 - 15 minutes. The cell-free superna
tant (1-2 ml) was pipetted into a 50 ml conical flask and 4% (w/v)
formaldehyde solution (0.5 ml) added to eliminate interference from
sulphite, if any. 10% acetic acid (1.0 ml) and thyodene (0.5-1.0 g)
were added and the mixture titrated with N/100 iodine solution,
previously standardised as described by Vogel (1962). Usually a
titration was carried out at least in triplicate. The following
relationship was used to calculate the amount of thiosulphate present
in the sample:
1 ml of N/100 Iodine 10 Micranoles of Thiosulphate.
Colorimetric Estirrution of ThiosuZphate (Sorbo, 1957).
The method involves the measurement of thiocyanate formed by the
interaction of thiosulphate with cyanide in the presence of copper ions,
which act as a catalyst. The procedure used was as follows:-
Centrifuged supernatant (1 ml) was added to a 25 ml volumetric

91
flask and the pH adjusted to 10 with 5% (w/v) sodium hydroxide solution
with thymolphthalein as indicator. The volume was adjusted to 25 ml
with distilled water. Aliquots of this solution (4.2 ml) were placed
in test tubes and 0.1 M potassium cyanide (0.5 ml) and 0.1 M cuprous
chloride (0.3 ml) were added while agitating the tubes to ensure rapid
mixing. Ferric nitrate reagent [ 100 g Fe(N0 3 ) 3 • 9 H2o+ 200 ml nitric
acid (65%, v/v), made up to 1000 ml with distilled water] (0.5 ml) was
added to dissolve the precipitate. A blank was also prepared, by add
ing ferric nitrate reagent and cuprous chloride solution to the samples
followed by the potassium cyanide reagent. The absorbance was measured
at 460 nm using a Hitachi Perkin-Elmer 139 UV-VIS Spectrophotometer.
Since phosphate is reported to interfere with the method, and
the growth medium contained phosphate, the method was standardised
with and without medium present. The similarity of the standard
curves (Fig. 2.1) indicated negligible interference from medium
components of Vishniac & Santer's medium (1957).
Distilled water (.) 0.4 Growth medium (o)
[Vishniac & Santer, 1957. ]
0.3 I= i::::
0
'° s::t"
0.2 FIG. 2.1 E-o
<i:: STANDARD CURVES FOR 1-Ll
u ESTIMATING THIOSUL- z
<i::
PHATE SPECTROPHOTO- ~ 0.1 0 Cl)
METRICALLY. a:i <i::
0
,/ /
,, I v,o'
.,, //
,P 'Y
k{'
i/''Y 0 40 80 120
Sodium thiosulphate
• /0
/
160 ( µg )

92
2.2.2 POLYTHIONATES
In order to obtain a general picture of by-products of thiosul
phate oxidation by the organism under study, polythionates were studi
ed and roughly estimated by the method suggested by Starkey (1934-a),
in which polythionates are hydrolysed, in the presence of potassium
hydroxide, to sulphite and thiosulphate both of which can reduce
iodine. The presence of polythionates was detected by the difference
in iodine titrations before and after alkali treatment of culture
broth.
Culture broth (5 ml), centrifuged at 10,000.g for 15 minutes
and filtered free of any suspended sulphur particles, was titrated
with N/100 iodine solution, as described previously under section
2.2.1., to obtain a measure of the amount of residual thiosulphate
present. To another aliquot (5.0 ml) 10% (w/v) potassitDil hydroxide
solution (5.0 ml) was added and thoroughly mixed while heating in a
boiling water bath for five minutes. After cooling, 10% (v/v) acetic
acid (10 ml) was added and the solution was titrated with N/100 iodine
solution, as described under section 2.2.1. An increase in the titra
tion over that initally performed for residual thiosulphate indicated
the presence of polythionate.
No attempts were made to identify or estimate individual polythio
nates produced in fermentation broths. The difference in iodine titres
before and after potassitDil hydroxide treatment was measured as such, as
a semi-quantitative measure of 'polythionates'
2.2.3 FERROUS AND FERRIC IONS
Ferrous iron was measured spectrophotometrically as described
by Welcher (1962).
Ferrous ions form a stable, orange-coloured complex with
o-phenanthroline, the absorption of which bears a reasonable

93
proportionality to the concentration of ferrous ions when measured at
520 nm. The reaction is so sensitive that ferrous concentrations bet
ween 1 and 7 p.p.m. can be accurately measured. The sample, if
turbid, was centrifuged at 5,000. g for 15 minutes, and diluted with
dilute sulphuric acid (pH 2.5) so that the concentration of ferrous
ions was within the limits of 1 - 7 p.p.m. An aliquot (10 ml) of
this solution was pipetted into a 25 ml volumetric flask and 0.25% (w/v)
o-phenanthroline solution (2.0 ml) was added. The volume was made up
to 25 ml with dilute sulphuric acid (pH 2.5). Absorbance was measured
at 520 nm in a 1 cm glass cell against a blank containing o-phenanthr
oline in dilute sulphuric acid (pH 2.5). A calibration curve was
constructed from standard ferrous solutions and the concentration of
samples read from this curve.
Although it was not necessary to reconstruct the standard curve
each time analyses were carried out, two concentrations of standard
ferrous solution ( 2 & 5 p.p.m.) were always included as a check on
the standard curve. In all these cases the deviations from the
standard curve were very small, i.e., within the limits of 2-5%.
The method was found to be specific for ferrous ions, but interference
from ferric ions can occur at very high ferric concentrations. Impor
tant factors for the reproducibility of results were found to be the
pH of the reaction mixture and the use of sulphuric acid (pH 2.5) for
dilution instead of water.
Ferric ions were estimated by a difference method. Total soluble
iron was determined by atomic absorption spectroscopy (see section
2.2.5 for procedure) and the amount of soluble ferric ion was then
determined by substracting the amowt of ferrous ions, estimated by the
Welcher (1962) method, from the total iron concentration.

94
2.2.4 EMISSION SPECTROSCOPIC ANALYSIS OF MINERALS
The semi-quantitative composition of minerals was determined
by emission spectroscopic analysis. With the assistance of Mr Finlay
son of the Department of Analytical Chemistry, the emission spectra of
minerals were recorded by using a BAIRD ATOMIC-3M grating spectrogr
aph, Model GX.
The zinc sulphide substrates were prepared as described tmder
section 2.1.4. An appropriate amotmt (9-11 mg) of the substrate was
mixed with an equal amount of spectroscopically pure carbon in a
sample bottle. The mixture was packed into the cavity (approximately
1/8 "diameter and 3/16" deep) of a pure carbon electrode and burned
in an electric arc.
Two 10" x 4" photographic plates [Ilford, No. 30] sensitive to 0 0
wavelenths of 2,300 A - 5,200 A were used to record the spectral lines
from these minerals. Using the iron spectrum as standard, spectral
lines for various elements were identified. Concentrations of elements
were determined from the intensities of their spectral lines when
compared with various standard concentrations.
Results of spectral analyses are shown in Table 3.3.1.
2.2.5 MINERAL ANALYSIS BY ATOMIC ABSORPTION SPECTROSCOPY
Atomic absorption spectroscopy was used to estimate quantitatively
the amounts of metallic ions leached by bacterial action on minerals
and also the composition of minerals used during these investigations.
Mineral Analysis.
An appropriate amount of finely divided mineral, prepared as
described under section 2.1.4 (0.5 - 0. 75 g), was weighed accurate,ly
in triplicate into clean dry test tubes. Concentrated hydrochloric

95
acid (10 ml) was added to each tube and the tubes were covered with
watch glasses. The acid was swirled around the test tubes so that the
sample was thoroughly wetted. The tubes were heated in an aluminium
block for 15-20 minutes. After digestion, the tubes were removed from
the aluminium block and concentrated nitric acid (2.5 ml) was poured
down the sides of the test tubes and the glass cover was replaced as
quickly as possible. When the reaction had lost its vigor, the tubes
were digested as before, by placing them in the heated aluminium block
for 15 minutes, until all the reaction had ceased, as indicated by the
losss of vigorous boiling. The residue at this stage should be light
brown coloured or, if the sample contained organic compounds, black.
If the residue was still black, a further addition of concentrated
nitric acid was followed by 16-20 minutes digestion and then the
glass cover was removed and the contents were allowed to evaporate to
a volume of approximately 5.0 ml. Care was taken that the samples were
not evaporated to dryness.
Concentrated hydrochloric acid (20 ml) was added to a 100 ml
volumetric flask and the contents of the tube were transferred to the
flask with distilled water. These samples were allowed to settle
for 3-4 hours at room temperature before flaming in the atomic absor
ption spectrophotometer.
Liquid Sa.mple Analysis.
Soluble metal contents of liquid samples obtained from fermenter
or shake-flask cultures were determined by spraying them directly into
atomic absorption spectrophotometer after suspending materials had
been removed from these liquid broths. In order to remove bacterial
and mineral suspensions, the solutions were either centrifuged at
10,000. g for 10 minutes and the supernatant collected or alter
natively they were filtered through a membrane filter removing

96
bacterial populations of the broth as well [ Sartorius membrane filter,
SM 11307].
It is important that the acid concentrations of standards and
samples are the same. Therefore, all dilutions were made with 3N
hydrochloric acid. Standard solutions were made from the stock solut
ion of 1000 p.p.m., purchased from B.D.H. The atomic absorption spectro-
photometer used was a Varian Techtron, Type AA3, fitted with digital
printer and digital indicator, built by Techtron Pty., Ltd., Melbourne,
Australia.
The spectral lines, wavelenths, gas mixtures and lamps used
for various elements during these analyses are shown in appendix 6.5.
2.2.6 X-RAY DIFFRACTION MEASUREMENTS
Representativesamples of minerals were finely grotmd in an
agate mortar. Mineral residues from shake flask experiments were
thoroughly washed with distilled water tmtil free from soluble sulphate
and dried overnight at 105°C.
A capillary tube loaded with sample was fixed in the centre
of a 6" camera loaded with Kodak X-ray film, 154, 8870. A copper
X-ray tube with nickel filter, operating at 30 kv and 25 mA, was used
to bombard the sample for 45-60 minutes. For marmatite, which
contained a higher percentage of iron, a chromium X-ray tube was used
and film was exposed for 90 minutes.
With the assistance of Mr F. Scott of the School of Metallurgy,
the repeat distances of the specimens were calculated and compared with
the standard values described in the Index Cards for sphalerite,
wurtzite and marmatite.
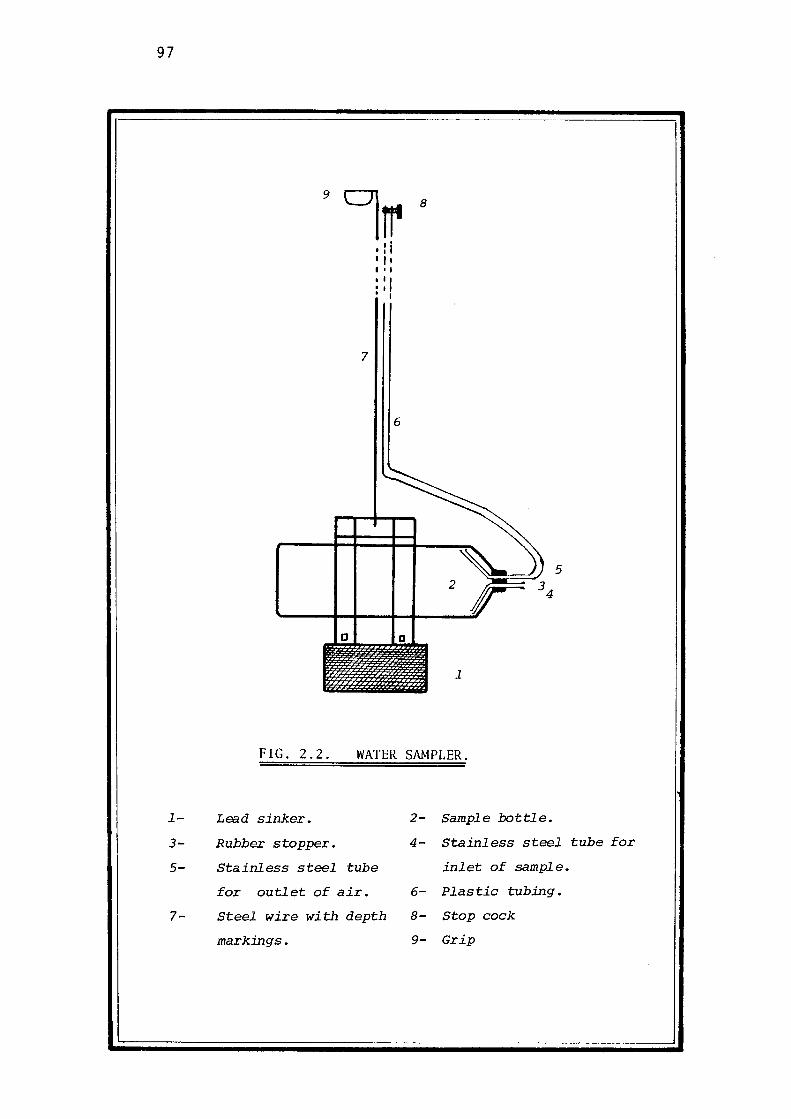
97
9
0 hr 8
I: I I I I I• I
I I I 'I ' I
7
6
~ 5 2
----+--+----+--ll-----'r C D
1
FIG. 2.2. WATER SAMPLER.
1- Lead sinker. 2- Sample bottle.
3- Rubber stopper. 4- Stainless steel tube for
5- Stainless steel tube inlet of sample.
for outlet of air. 6- Plastic tubing.
7- Steel wire with depth 8- Stop cock
markings. 9- Grip

98
2.3 ISOLATION OF THIOBACILLI
2.3.1 ISOLATION OF NON-ACIDOPHILIC THIOBACILLI
The isolation of Thiobacilli which are tolerant to pH values
between 4 & 9 (referred to herein as 'non-acidophilic Thiobacilli')
was carried out by employing enrichment techniques in both open and
closed systems.
Source Samples.
were:
The source materials used for the isolation of these bacteria
(i) Soil Samples.
About 300 soil samples from numerous areas of New South Wales.
(ii) Sewage Water Samples.
Water samples from various sewage treatment plants were collected
in pre-sterilised sample bottles and brought to the laboratory the same
day.
In order to get water samples from.various depths of oxidation
ponds, a special water sampler, designed and fabricated in the labo-
ratory with the assistance of Mr Thomas Babij, was used. It consisted
of a sample bottle (serum-bottle) fixed horizontally to a 'lead sinker'
with metal clamps. The wateFtight rubber stopper of the bottle was
fitted with two bores, one for inlet of the water sample and the other,
to which a sufficient length of plastic tubing was attached, for the
escape of air. The other end of the plastic tubing was connected to
a simple stop cock. This whole assembly was attached to a strong cord
marked for depth. For sampling at different depths, the stop cock was
closed and the sampler lowered to the desired depth. The air inside

99
the bottle was then released by opening the stop-cock, allowing the
water sample to rush in. When the bottle was filled, the stop-cock
was closed and sampler was removed from the water.
The water sample thus collected was poured into a sterile,glass
stoppered sample bottle and stored in a cold room ( 4° C ). The samp
ler bottle was washed with sodium hypochlorite ( 5% w/v) and then
thoroughly rinsed with sterile water before next sample collection.
Fig. 2.2 illustrates the sampler.
Media.
The following two media were used for isolating the non-acidophi-
lie Thiobaci Ui.
COMPONENTS
(NH4) 2so4
NH4Cl
K2HPo4
KH2Po4
MgS04.7H20
MgC12.6 H2o
CaC12
FeC13.6 H2o
MnS04 .4 H2o
NaHC0 3
Na2s2o3 .5 H2o
pH
Distilled Water
1- Starkey (1934-a).
2- Beijerinck (1904).
MEDIUM A1 MEDIUM B2
0.10 g
0.10 g
4.00 II 0.20 II
4.00 II II
0.10 II
2.50 II
0.10 II
0.02 II
0.02 II
1.00 II
10.00 II 5.00 II
6.6 9.1
1,000 ml 1~ 000 ml

100
Medium A.
The basal salts (i.e., all components except thiosulphate)
were dissolved in 900 ml of distilled water and the solution dispensed
into test tubes (4 ml per tube) and sterilised by autoclaving at 15
psi for 15 minutes. The sodium thiosulphate was dissolved separately
in 100 ml of distilled water and sterilised by filtration through a
membrane filter (0.2µ pore size, SM 11307 , Sartorius ). 0.5 ml of
sterilised sodium thiosulphate solution was added aseptically to
each tube of sterile basal salts medium.
Medium B.
The basal salts were dissolved in 900 ml of distilled water and
4-5 ml portions dispensed into test tubes. The tubes were then auto
claved at 15 psi for 15 minutes. The sodium thiosulphate and sodium
bicarbonate solutions were sterilised separately by filtration using
the same type of filter as described for medium A. 0.5 ml of this
solution was added aseptically to each test tube containing medium B
basal salt solution.
Solid media were prepared by adding 1% (w/v) 'Ionagar-2' to the
broths described above.
Isolation Procedu:res.
0.5 - 1.0 ml of each sewage water sample or 0.5 - 1.0 g of
each soil sample, withdrawn after thorough mixing, was aseptically
introduced into test tubes of medium A and B. The tubes were incubated
at 30°C. After 2 and 7 days the tubes were examined for evidence of
growth, decrease of pH and presence of residual sodium thiosulphate.
The pH was determined with narrow range pH papers (Whatman-BDH).
The samples showing any sign of growth were re-inoculated into
500 ml Erlenmeyer flasks containing the corresponding medium and

101
incubated on a reciprocal shaker at 30°C (5 cm stroke, 90 + 2 strokes
per minute). These flasks were also monitored for their residual
concentration of sodium thiosulphate and for pH.
Purification of the enriched cultures thus obtained was achieved
on solid medium plates, using conventional procedures.
2.3.2 ISOLATION OF ACIDOPHILIC THIOBACILLI
The isolation of Thiobacillus species tolerant to pH values
between 1.0 and 4.0 (referred to herein as 'acidophilic Thiobacilli'),
was achieved after enrichment in shake flasks.
Sou:t'ce Samples.
(i) Soil Samples.
Soil samples were taken from various mineral heaps, sulphur
stores and from the vicinity of eroding concrete structures.
(ii) Mineral Samples.
Mineral or rock samples obtained from various places.
(iii)Mine Drainage.
Acidic drainage solution from Mol.IDt Lyell Mine was fol.IDd to
be excellent source of these bacteria.
Mediwn.
The basal medium used for the isolation of acidophilic Thio
bacilli was the 9K medium devised by Silverman & Lundgren (1959); its
composition is shown in Table 1.4B. The basal salts were dissolved
in 900 ml of distilled water. 90 ml portions were then dispensed into
500 ml Erlenmeyer flasks and sterilised by autoclaving at 15 psi for
15 minutes.

102
Isolation ProcedUl'es.
For the isolation of ThiobaciZZus ferrooxida:ns, ferrous sulphate
solution (40%, w/v) was sterilised separately by filtration; (pH was
previously adjusted to 2.5 with dilute sulphuric acid to avoid its
rapid oxidation). 10 ml of this sterilised solution was added
to each flask aseptically. The flasks were then inoculated with a soil,
mineral or mine drainage water sample and incubated at 30° Con a
reciprocating shaker ( 5 cm stroke; 90 ~ 2 strokes per minute). Evid
ence of growth was obtained by monitoring the oxidation of ferrous ion.
Isolates were obtained by serial dilution of the enriched culture.
For the isolation of ThiobaciZZus thiooxida:ns, the 9K basal
medium was supplemented with 2% (w/v) elemental sulphur which had
previously been sterilised by Tyndallization over three consecutive days.
A decrease in pH, due to the production of sulphuric acid by the
oxidation of elemental sulphur, was taken as evidence for the presence
of ThiobaciZius thiooxidans. The purification was achieved on
thiosulphate agar, pH 4.0.
2.3.3 MAINTENANCE OF CULTURES
The preservation of ThiobaciZZi is still problematical. Therefore,
the cultures of 'non-acidophilic ThiobaciZZi' isolated during these
investigations were maintained on solid as well as liquid cultures.
The petri dishes with colonies were stored at 4°C, and they were
regularly transferred into fresh plates at fortnightly intervals. It
was found that, if there were delays of more than two weeks in the
transfer of these cultures onto fresh medium, the cultures lost
viability, most probably because of overproduction of acid. The same
was true of the shake flask cultures.

103
Preservation of the more acidophilic members of the genus was
carried out by the same method, with the exception that only liquid
cultures were preserved at 4°C. It was noticed that ThiobaciZZus
ferrooxidans cultures could be maintained viable for long periods
(6 - 8 weeks) by suspending them in dilute sulphuric acid ( pH 2.5)
instead of growth medium. ThiobaciZZus thiooxidans suspensions in
dilute sulphuric acid (pH 2.5) were also viable for 8-10 weeks.
The purity of each culture was checked by routine microscopic
examination of Gram-stained slides.

104
2.4 TAXONOMICAL INVESTIGATIONS
Taxonomical investigations were carried out according to
conventional microbiological procedures. However, due to the difficulty
in obtaining colonies on agar medium in the case of some species, some
tests had to be modified. The various tests performed to identify
these microorganisms are described in the following:
2. 4.1 MICROSCOPIC EXAMINATIONS
These microscopic tests were carried out according to the
general procedures described by Skennan, 1967.
1- Gram staining.
Actively growing cells were stained by Gram-stain and examined
under a microscope.
2- FZageZZurn staining.
The cells for this test were obtained from actively growing
liquid cultures. The culture was centrifuged and the cell pellet
suspended in deionised sterile water; the staining procedure of
Leifson was then applied ( Skerrnan, 1967).
3- MotiZi ty test.
For motility tests cells from the logarithmic growth phase of
liquid cultures were examined microscopically by phase contrast.
4- MorphoZogiaaZ s-t;udies.
The morphology of BJR-451 (ThiobaaiZZus thioparus) and BJR-KOl
(ThiobaaiZlus thiooxida:ns) was studied after growing them on thiosul
phate agar plates. For BJR-451, the solid medium of Vishniac & Santer
(1957) was used whereas BJR-KOl was grown on thiosulphate agar medium

105
made by adding 1% (w/v) sodium thiosulphate to the 9K medium of
Silverman & Lundgren (1959). ThiobaciZZus ferrooxidans (BJR-Kl)
was grown in the 9K medium with ferrous ion as the oxidisable energy
source.
Unfixed cells were examined microscopically under phase contrast.
This examination provided more reliable information regarding the shape
and size of the cells than would have obtained from fixed cells.
2.4.2 BIOCHEMICAL TESTS
The following biochemical tests were performed.
1- Growth on nutrient agar.
Cultures were streaked on the nutrient agar plates and were
incubated at 30°C for one week, after which they were examined for
development of colonies. The test was carried out to determine the
autotrophic nature of the isolates.
2- Growth on different sulphur substrates.
The ability of these isolates to oxidise various reduced sulphur
compollllds was tested by growing them in the presence of these substrates
as the sole energy source.
The sulphur salts were substituted for the usual substrates
(sodium thiosulphate for BJR-451, and elemental sulphur for BJR-KOl)
in liquid medium. Growth, indicated by the presence of turbidity or
increase in the acidity, or augmentation of the cell numbers measured
by direct enumeration under a microscope, was considered a positive
sign of the substrate having been utilised.
3- Oxidation of ferrous ion.
Distinction between ThiobaciZZus thiooxidans and Thiobaci
ZZus ferrooxidans was made by examining an isolate's capability of
oxidising ferrous salt. Each isolate was inoculated into the broth

106
containing ferrous sulphate ( 2%, w/v) and incubated at 30°C. Cultu
res showing oxidation of ferrous ion as indicated by the disappearance
of ferrous species from the medium and development of a brownish
coloration were classified as Thiobacillus ferrooxidans.
4- Anaerobic growth.
In order to examine the capabilities of the isolates to utilise
nitrate as a terminal electron acceptor in the absence of molecular
oxygen, they were grown anaerobically in the presence of nitrate and
a suitable energy substrate; (in most cases sodium thiosulphate was
used as energy source). The cultures were examined for growth after
2, 7 and 14 days' incubation at 30°C.
5- Nitrate reduction.
The reduction of nitrate in the presence of oxygen was tested standard bacteriological
by the L procedure. Tests for the accumulation of nitrite were
carried out as recommended by the standard procedure described by
Skerman, 1967.
6- Detemination of DNA base composition.
The DNA base composition of Thiooocillus thioparus was
determined by following the method described by Hill (1968).
Thiobacillus thioparus (BJR-451) cells were collected from the
outstream of a continuous flow fermenter and were centrifuged in
sterile centrifuge tubes in a Sorvall centrifuge operating at 12,000 .g
for 10-15 minutes. The cells were washed twice with the basal
salt medium in order to remove extracellular materials including
sulphur.
The nucleic acid was extracted from the washed cells by the
method described by Marmur (1961). Purified nucleic acids were
analysed by recording their UV-absorption spectrum, which is the
summation of the individual spectra of four bases; ( a highly purified

107
DNA isneeded, as traces of proteins with UV-absorption maxima at 280
nm interfere). Due to its relative simplicity and reliability, the
method described by Hirschman & Felsenfeld (1966) was used for spectrum
analysis of nucleic acids. The absorption of DNA solutions in phosph
ate buffer, pH 7.0, containing 0.01 M sodium chloride was read at 5 nm
intervals from 235 nm to 290 nm with a Hitachi-Perkin Elmer 139 UV-VIS
spectrophotometer.
The absorbance values for three different concentrations were
obtained and are shown in appendix 6.4a. The method of calculating
the [A+ T] ratio from the data is shown in appendix 6.4.
7- Determination of fatty acid composition.
The determination of fatty acid composition of the cells of
BJR-451 was carried out by the following method described by Babij
et al. (1969). Bacteria were grown in a chemostat at optimum growth
conditions with respect to pH and temperature and cells were harvested
by centrifuging the outflow at 10,000.g for 15 minutes. As recommended
by Agate and Vishniac (1973-b), the cell pellet was used as such
without washing.
Bacterial lipids were extracted by the technique used by Bloor
(1928), Floch et al. (1951) and Mcfarlane (1942). The cell pellet was
transferred to a sample tube and frozen to -20°C. The frozen cell
mass was transferred quantitatively into two volumes of ethanol:
diethyl ether [3:1 (v/v], and allowed to thaw at room temperature in
a nitrogen atmosphere. The solvent was decanted and the cells re
extracted with a ethanol:chloroform [2:1 (v/v] mixture for two hours.
All extractions were carried out under a nitrogen atmosphere in order
to avoid oxidation.
The combined solvent extracts, after filtration through
Whatrnan No. 1 filter paper, were evaporated at 50°C under vacuum. The
residue was refluxed for 30 minutes with 10 N methanolic potassium

108
hydroxide to saponify the lipids. The fatty acids were separated by
acidifying the contents to pH 2.0 with hydrochloric acid and were
methylated by refluxing for 30 minutes with 5% methanolic hydrochloric
acid. The methyl esters were concentrated by pouring the refluxed
mixture into cold saturated sodium chloride solution. They were
collected from the top of the sodium chloride solution and
at -20°C.
stored
The methylated esters of the fatty acids were analysed iso
thermally at 200°C in a Beckman GC-4 gas chromatograph fitted with a
column packed with 10% polyethylene glycol succinate on acid washed
Cromosorb W. The carrier gas pressure was 30 psi.

109
2.5 CULTIVATION OF THIOBACILLI
The ThiobaciZZi were cultivated in batch and continuous flow
systems. The degree of control of various parameters differed from
one system to another. In this section the apparatus, culture vessels
and control systems used during cultivation of Thio'baciZZi employed in
these studies are described.
2.5.1 CULTURE VESSELS
Erlenmeyer flasks of one litre capacity containing 200 ml medium
were frequently employed for growth of inoculum and during leaching
experiments. These flasks were shaken on a reciprocating shaker oper
ating at 5 cm stroke and 90: 2 strokes per minute. It was housed in
a temperature controlled room (30 ± 1 °C). One of the serious draw
backs of shake flasks is the lack of proper pH monitoring facilities
for the system. Therefore, the pH of the medium during these experi
ments was controlled manually. pH was adjusted with appropriate vol
umes of sterilised sodium hydroxide or sulphuric acid.
In order to achieve a better control of pH, temperature and
dissolved oxygen, a fermenter which could be operated both as a batch
reactor and as a continuous culture vessel was employed. The fermenter
with its control system is shown in Fig. 2.3.
2.5.2 PREPARATMNOFMEDM
Batch cuZtUPe.
Unless otherwise stated, the non-acidophilic ThiobaciZZus thiop
arus BJR-451 was cultivated in Vishniac & Santer medium (1957). Basal
salts of Vishniac & Santer medium was dissolved in distilled water.
The basal salt solution was dispensed in Erlenmeyer flasks of one litre
capacity (180 ml). The flasks were sterilised by autoclaving at

110
FIG. 2.3. A DIAGRAMATIC REPRESENTAION OF THE CULTURc
VESSEL AND THE AUXILLIARY EQUIPMENT.
1- Medium reservoir. 2- Mackley Air Filter, Microflow Ltd. 3- Medium pump (peristaltic finger pump). 4- Air filter packed with glass-wool. 5- Precision air pressure regulator valve,
(Negretti & Zambra, London). 6- Fine needle valve. 7- Air flow meter. 8- Air filter packed with glass-wool. 9- Drip feed.
10- Vibromix (El, Chemap, AG, Alte Landstr, 415, 8708 Mannedrof, ZH, Schweiz.).
11- Air outlet moisture trap. 12- Mackley Air Filter, Microflow Ltd. 13- pH-controller, (Dynaco, 21 A, England). 14- DOT meter (Scrool of Biological Technology). 15- Temperature controller (Ether, "MINI", Type 19-90 I, Pye). 16-17-} Acid and Alkali reservoirs.
18-} Acid and Alkali pumps (Delta Watson & Marlow Ltd.). 19-20- Infra red heating lamp (Osran, England). 21- Fermenter. (Volume 2 litres) 22- pH-electrode, combined, sterilisable. 2 3- Condenser. 24- Oxygen electrode (Borkowski & Johnson probe). 25- Hallow sparger of Vibromix. 26- Sterile neoprene seal. 27- Overflow tube. 28- Thermister. 29- Steam sterilisable outlet. 30- Mackley Air Filter, Microflow Ltd. 31- Culture collecting cylinder. 32- Steam sterilisable sample collector. 33- Mackley Air Filter, Microflow Ltd. 34- Sterilised culture collecting vessel.
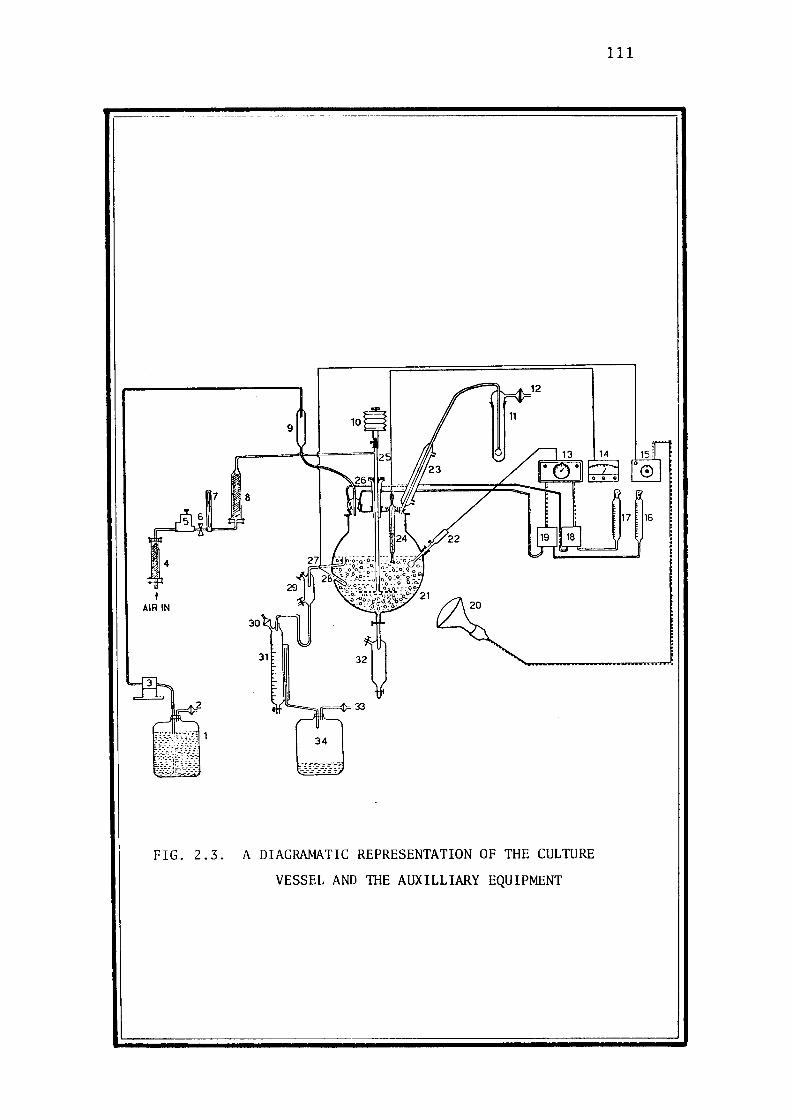
'· 4
t AIR IN
7
FIG. 2.3.
8
34
A DIAGRAMATIC REPRESENTATION OF THE CULTURE
VESSEL AND IBE AUXILLIARY EQUIPMENT
111

112
14 psi for 15 minutes. Sodium thiosulphate (10 g) was dissolved in
distilled water (100 ml) and sterilised by passing through a membrane
filter (0.2 µ pore size, SM 11307, Sartorius:). An appropriate
aliquot of this solution (20 ml) was added aseptically to each flask.
9K medium (Silverman & Lundgren, 1959) was used for culturing
acidophilic Thiobacilli. Thiobacillus ferrooxidans was grown in the
presence of ferrous ion as the sole energy source. The medium has been
described in section 2.3.2 which also details the growth medium for
Thiobacillus thiooxidans.
Continuous cultu.re.
The media for continuous culture investigations were the same
composition as those for batch culture and were prepared in bulk by
filtration through a membrane filter using the specially designed
apparatus described by Babij et al. (1971). The components were mixed
together by stirring in a stainless steel container and were then
passed through a pre-filter to remove insolubles. After prefiltration
the medium was pumped through a sterilised membrane filter into
sterilised storage bottles. Approximately 60 litres medium was pre
pared in one operation.
2.5.3 MEASUREMENT OF DI~SOLVED OXYGEN
The dissolved oxygen tension (DOT) in the culture broth
was measured by using a modified form of the oxygen electrode descri
bed by Johnson et al. (1964). The anode was a pure lead sheet
cylinder instead of the spiral as recommended by the above authors and
the silver anode was sealed in a glass capillary tube to protect it
from the corrosive action of the electrolyte. The output from the

113
probe was fed into a recorder.
The media used for cultivating ThiobaaiZZi in the fermenter
frequently contained thiosulphate and other sulphur compounds which
reacted chemically with the silver anode of the oxygen electrode and
lowered its sensitivity. The reaction between the silver anode and
sulphur compomds was prevented to some extent by covering the anode
with double layer of teflon membrane,which also prolonged the stabi
lity of the electrode.
2. 5. 4 MEASUREMENT OF CO 2 IN THE EFFLUENT GAS STREAM
The utilisation of co2 by ThiobaciZZi was determined by
measuring the residual co2 in the exit stream of gases from the
fermenter. A known amount of co2 was pumped into the fermenter at a
known rate, measured by the flowmeter and the co2 present in the out
let stream was determined by an infrared co2 analyzer (Hartman and
Braun, Germany).
The output from the co2 analyzer was transferred to a recorder
(Rikadenki-Multipen Recorder •. Model B-2021, Kogyo Co., Japan). The
instrument was previously calibrated with an accurate and pure mixture
of 0.49% co2 in pure nitrogen (CIG-pure mixture). The zero on the
recorder was calibrated with pure nitrogen (CIG). The amount of co2
utilised by the culture was detennined by difference.
2.5.5 DETERMINATION OF TOTAL BACTERIAL POPUIATION
The total number of bacteria in the fermentation broths was
determined by counting them in a bacterial counting chamber under a
microscope. Since the young cultures were actively motile, they were
killed with 5% (w/v) phenolic solution in water ( usually 2 ml
cell suspension was mixed with 1 ml of phenolic solution). The

114
mixture was thoroughly mixed on a vortex mixer. The counting chamber
was then filled with the bacterial suspension and the number of micro
organisms seen by phase-contrast microscopy corn1ted. The bacteria
present in at least 20 squares were corn1ted and their average was
calculated.

3 RESULTS & DISCUSSION

115
3.1 ISOLATION AND CHARACTERISATION OF THIOBACILLI
About 300 soil and water samples from sewage works and mines
were screeened for non-acidophilic sulphur oxidising bacteria (appendix
6.1). Endeavours were also made to isolate iron-oxidisers and acido
philic sulphur bacteria from soil, sewage works and mine water samples
(appendices 6.2 & 6.3).
3.1.1 ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING BACTF:R.IA
Appropriate amounts of soil or water samples (soil sample 1-2 g
or water sample 1 ml) were incubated in test tubes containing either
medium A or medium B. As already mentioned under section 2.3.1 medium
A was as devised by Starkey (1934-a), adjusted to pH 6.6, whereas
medium B was Beijerinck's medium used for isolating microorganisms
with pH optima near or above neutrality and was adjusted to an initial
pH of 9.1 (Beijerinck, 1904). Sodium thiosulphate was the sole source
of energy provided in these media. After 2 and 7 days' incubation at
30°C, pH and thiosulphate levles were monitored with narrow range pH
papers and a standard iodine solution. Production of any turbidity,
sediments or pellicles was also recorded (appendix 6.1). Those samples
showing any sign of growth were reinoculated into fresh media; only
one sample (451) gave good growth and complete utilisation of sodium
thiosulphate. The strain isolated from this sample was designated
BJR-451, and was selected for further study.
The fact that chemical changes occurring during growth of a mixed
population in a closed system are very complex and highly uncontrollable
makes the reproducibility of the classical enrichment techniques very
difficult. A new type of growth selection for studying the mutation

116
rates in microorganisms was discussed and practised with the introduc
tion of continuous cultivation techniques (Novick & Szilard, 1950-a).
In contrast to batch culture, continuous culture provided the facility
of selecting microorganisms in a controlled environment on the basis
of their different growth rates at different substrate concentrations.
Surprisingly, continuous culture has been used very sparingly for the
selection and enrichment of microorganisms. Veldkamp (1970) discussed
the use of continuous culture methods for a possible enrichment
technique for isolating prokaryotes and Harder et al. (1977) recently
published an excellent appraisal of the various methods employed for
microbiological selection by using continuous culture.
Starkey's medium ( Starkey, 1934-a) (2 litre) was dispensed in
the fermenter described in section 2.5.1 and was inoculated with 10-15
ml growth suspension obtained by incubating soil sample 451 as descr
ibed earlier. Temperature and pH were controlled at 30°C and 6.6
respectively. When growth occurred in the fermenter (after 3-4 days,
indicated by production of white suspension and depletion of sodium
thiosulphate from the culture broth) fresh medium was pumped in at a
definite flow rate lllltil a steady state was reached. The flow rate
was progressivley increased to washout. The culture from the outlet
was collected and plated out on Starkey agar plates and incubated at
30°C. The strain derived from the highest dilution rate (0.12 h-l)
was designated BJR-451 and was reinoculated into a shake flask and
further studied.
3.1.2 ISOLATION OF ACIDOPHILIC IRON OXIDISING BACTERIA
The isolation of iron oxidising bacteria was carried out by
inoculating samples into test tubes containing 5.0 ml of 9K medium
of Silverman and Lundgren (1959) with 4% (w/v) ferrous sulphate as the
sole energy source. The tubes were incubated at 30°C and examined for

117
oxidation of ferrous ion after 2,7 and 14 days.
Only one water sample (from Mount Lyell) oxidised the ferrous
ion to a significant extent (appendix 6.2). Further transfers were
carried out in liquid medium and finally the culture was purified
from a single colony developed on a silica gel plate. The culture
was maintained in dilute sulphuric acid (pH 2.5) and was designated by
the code number BJR-Kl.
3.1.3 ISOLATION OF ACIDOPHILIC SULPHUR OXIDISING BACTERIA
For isolating acidophilic sulphur oxidising bacteria, samples
were inoculated into test tubes containing 5.0 ml of 9K medium of
Silverman & Lundgren (1959) containing elemental sulphur as the
energy source.
According to growth measurement, only one sample was able to
oxidise elemental sulphur at a significant rate at the low pH of 2.5.
It presumably contained acidophilic sulphur oxidisers; the soil
sample was obtained from sulphur storage pad at Sulphide Corporation
Ltd., at Boolaroo, N.S.W. (appendix 6.3) . The enriched sample
was reinoculated into a flask and finally purified on a thiosulphate
agar plate (pH 4.0). The organism isolated was designated by the
code number BJR-KOl.
3.1.4 TAXONOMY OF THE ISOLATES
The taxonomical characteristics of the three isolates described
under sections 3.1.1, 3.1.2 & 3.1.3 are summarised in Table 3.1.
BJR-451
BJR- 451 is short Gram-negative, non-sporulating rod, 0.6-1.6 µ
long and 0.5-0.75 µ wide. It was actively motile when in contact
with substrate but motility was lost after exhaustion of sodium

118
0
c;
PLATE 1
ELECTRON MICROGRAPH OF BJR-451 GROWN IN VISHNIAC AND SANTER'S
MEDIUM .
0
0

119
thiosulphate from the medium. All attempts to stain the flagellum
were unsuccessful, apparently because of its extremely fragile nature.
However, electron micrographs of a young culture, treated with phospho-
tungstic acid, reveales a single polar flagellum (Plate 1). The
microbial cells appeared to be embedded in a thick slime.
Growth in thiosulphate broth was rapid, the medium becoming
turbid within 24 hours. As growth proceeded, white pellicles appeared
on the surface of the broth. Complete decomposition of thiosulphate
occurred within 30-40 hours, with decrease of pH from 6.5 to 4.0.
When grown on thiosulphate agar plates, small colonies, about
1.0 - 1.5 mm in diameter, appeared within 24 - 36 hours (Plate 2).
These colonies were white and circular with entire edges. Fresh
colonies were transparent to translucent but developed a whitish
hue, due to the precipitation of sulphur, with the passage of time.
One week old colonies were usually opaque due to precipitation of
elemental sulphur whereas 2-3 weeks old colonies acquired a brownish
tinge with ageing.
Since in pure culture it does not grow on nutrient agar, this
organism may be an obligate autotroph. Sometimes contaminants grew
on nutrient agar; however, when these were streaked back onto
thiosulphate agar no growth was noticeable, indicating the absence of
any facultative autotrophic bacteria in the culture.
When inoculated into liquid broth containing elemental sulphur
instead of sodium thiosulphate, the organism oxidised the sulphur
slowly as compared to thiosulphate. Uniform turbidity was obtained
after three weeks and pH was dropped to 4.5.
BJR-451 was also capable of oxidising a number of other reduced
sulphur compounds, for example, sodium sulphide, tetrathionate, tri
thionate and pentathionate. Sodium sulphite and sodium thiocyanate were
not oxidised.

PLATE 2 COLONIES OF BJR-451 ON VISHNIAC
AND SANTER'S MEDIUM.
120

121
TABLE 3 .1
CHARACTERISTICS OF BJR-451, BJR-Kl & BJR-KOl.
CHARACTERISTICS
MORPHOLOGY:
CELL SHAPE &
SIZE.
MOTILITY
BJR-451
Short rods.
0.6-1.6 µ long, 0.5-
0.75µ wide, single
cells, occasionally
occurring in pairs.
Young cultures were
actively motile but
tended to lose their
motility with age.
FLAGELLUM Single, polar.
GRAM-STAINING Gram-negative,
easily stained.
SPORE FORMATIO! Non-sporulating.
COLONY
CHARACTERISTIC/
LICUID
BROTH.
Colonies on thiosul
phate agar at pH 6.0
were small, circular
with complete edges;
1-1.5 mm in diameter.
Initially colonies
were whitish yellow
turning brown after
prolonged incubation
for 2-3 weeks.
Liquid medium with
thiosulphate became
uniformly turbid,
BJR-Kl
Short rods.
1-2 µ long, 0.5-0.75p
wide, single cells.
Non-motile.
Gram-negative,
easily stained.
Non-sporulating.
Colonies could not
be produced on agar
plates. probably due
to the toxicity of
agar towards this
BJR-KOl
Rods.
1.0-2.5 11 long, 0.5 -
1.0µ wide, single
cells.
Young cultures were
motile but motility
appeared to be lost
as the pH decreased
with growth.
Single, polar.
Gram-negative,
easily stained.
Non-sporulating.
Colonies on thiosu
lphate agar (PH 4.0)
were small, circular,
varying in size from
0.5-1.0 mm in diam-
bacterium. On silica eter. They were
gel-ferrous sulphate
plates tiny colonies
of brownish red
whitish yellow in
colour and sometimes
the edges of colonies
colour were obtained. were clear due to
The colonies were
circular, sometimes
developing serrated
edges.
No growth was
noticeable in thio
sulphate broth. 9K
oxidation of precip
itated sulphur.
Uniform turbidity
i•n sulphur broth was
obtained and medium

CHARACTERISTICS
PHYSIOLOGY:
GROWTH ON
NUTRIENT AGAR
PLATES.
UTILISATION OF
SULPHUR COMP-
OUNDS:
Thiosulphate.
Trithionate.
Tetrathionate.
Elemental
Sulphur.
Sodium
Sulphide.
TABLE 3.1 (Contd.)
CHARACTERISTICS OF BJR-451, BJR-Kl & BJR-KOl.
BJR-451
with pellicles and
occasionally contained
precipitated sulphur.
The pH dropped from
6.5 to 4.0.
BJR-Kl
medium with ferrous
sulphate supported
growth and produced
uniform turbidity.
After 2 days' incub-
122
BJR-KOl
became extremely acid
(pH 0.7). Growth in
thiosulphate broth
was sparse and produ
ced a uniform turbi-
ation ferrous ion dity.
No growth.
Uniform turbidity.
Uniform tubidity.
Uniform turbidity
was obtained after
one week'sincubation.
Uniform turbidity
was obtained after 3
three weeks, pH
dropped to 4.5.
Uniform turbidity
was obtained.
was oxidised to ... i.
ferric state turning
the broth from
bluish green to red-
dish brown.
No growth.
No growth.
No growth.
No Growth.
Initially slow
growth but after two
weeks adaptation
strain oxidised sul
phur at rates compa -
rable to that given
by B.JR-K0l.
Not determined.
No growth.
oxidises very slow
ly.
Very slow oxidation
occurred after prolo
nged period of 3 weeks
incubation.
Uniform turbidity
was obtained after
two weeks'incubation.
Uniform turbidity
was obtained after
two days,dropping the
pH to 0.5.
Not determined.

CHARACTERISTICS
Thiocyanate.
Ferrous
Sulphate.
Oxygen requi
rement.
DNA Base
Composition.
FANE Profile.
pH Range for
growth.
TABLE 3.1 (Contd.)
CHARACTERISTICS OF
BJR-451
No growth.
No growth.
0bligately aerobic.
G+c = 64.8%
Cl2, Cl6 & Cl8:l
were predominant fatty
acids.
7.0 to 4.0.
BJR-451, BJR-Kl f, BJk-t:01.
BJR-Kl
Not Determined.
oxidised ferrous
iron to ferric.
0bligately aerobic.
Not determined.
Not determined.
4.0 to 1.0
123
BJR-KOl
Not determined.
No growth or oxi
dation of ferrous iron
was observed.
0bligately aerobic.
Not determined.
Not determined.
4.0 to 0.5
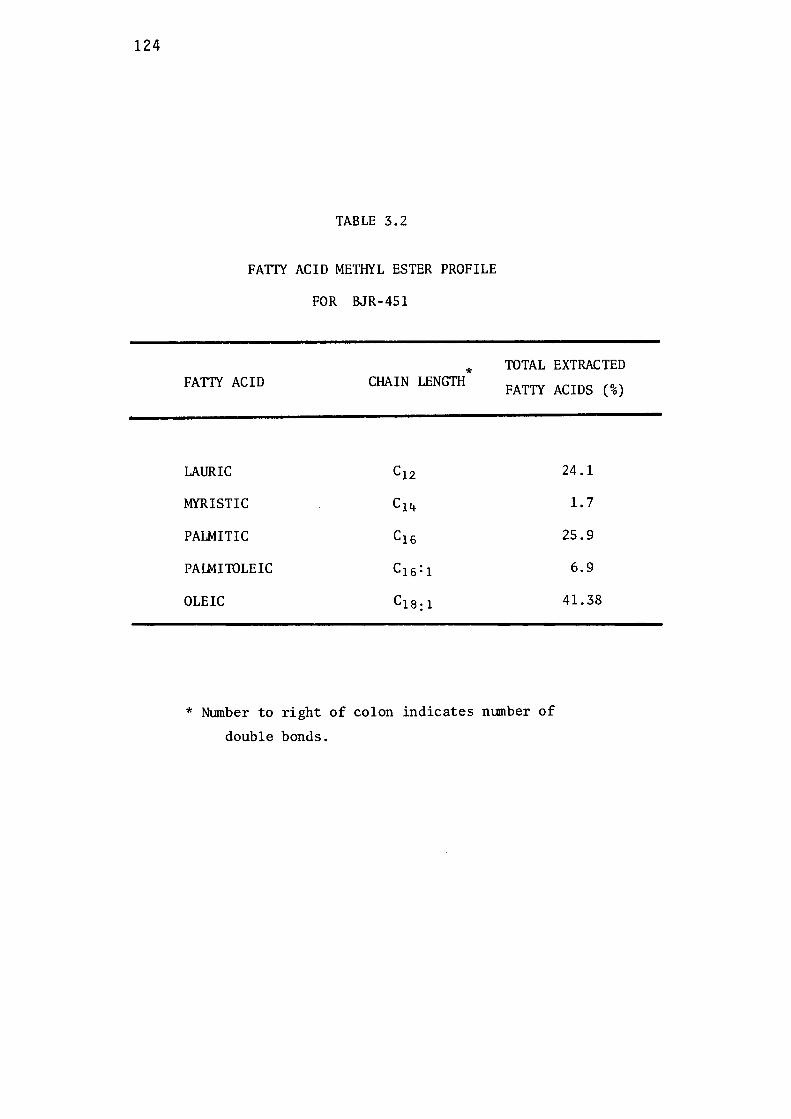
124
TABLE 3.2
FATIY ACID METHYL ESTER PROFILE
FOR BJR-451
* FATTY ACID CHAIN LENGTH
LAURIC C12
MYRISTIC C14
PALMITIC C16
PAIMITOLEIC C16:1
OLEIC C1a:1
TOTAL EXTRACTED
FATTY ACIDS (%)
24.1
1. 7
25.9
6.9
41.38
* Nl.llilber to right of colon indicates nl.llilber of
double bonds.

125
BJR-451 is aerobic and does not grow anaerobically in the
presence of sodium nitrate as an electron acceptor. It was also
unable to oxidise ferrous sulphate.
The DNA base composition for GC was found to be 64.8%. The
complete calculations have been described in appendix 6.4. Fig.3.1
is FAME (fatty acid methy ester) profile of BJR-451. The strain
was cultivated at its optimum pH and temperature (pH 5.5, temperature
30°C) in a chemostat and lipids were extracted with ethanol and
diethyl ether [3:1 (v/v)]with a final extraction by methanol chloroform
mixture [2:1 (v/v)] as described by Babij et al. (1969). Table 3.2
presents the percentage composition of the fatty acids detected and
identified from Fig. 3.1. Oleic acid was in predominance (41.38%),
followed by !auric and palmitic acids in almost equal proportions
(24.1 and 25.9% respectively) and myristic and palmitoleic acid were
present in smaller proportions ( 1.7 & 6.9 % respectively).
Since BJR-451 is a strict autotroph capable of oxidising
reduced sulphur compounds and its cells are motile rods without sulphur
granules, it presumbly belongs to the genus ThiobaciZZus, the known
species of which are ThiobaciZZus thioparus, ThiobaciZZus neapoZitanus,
ThiobaciZZus denitrificans, ThiobaciZZus noveZZus, ThiobaciZZus inter
medius, ThiobaciZZus perometaboZis, ThiobaciZZus thiooxidans and
ThiobaciZZus ferrooxida:ns (Buchanan & Gibbons, 1974). Recently some
workers have claimed the isolation of some new species of the genus
ThiobaciZZus, viz., ThiobaciZZus rubeZZus, ThiobaciZZus deZicatus
(Mizoguchi et al., 1976), and ThiobaciZZus acidophiZus (Guay &
Silver, 1975).
BJR-451 is a non-acidophilic sulphur oxidising organism as
acidic conditions (pH less than 4.0) were highly unfavourable for its
growth, therefore, the possibility of its being ThiobaciZZus

126
100-----------------r--------,-------,
80 I----+-----------·--·--------------
60 ---+--------~----~- - -t-------1
! I I
'
40 1-1--~------1----i
... . . >-- . 1---------i
20 ---- ---+-------·-------·-------··--·---- -- - -- - ·-----·---·------- -
Flg.3.1 FAME Profile Of BJR- 451.

127
thiooxida:ns, ThiobaciZZus ferrooxidans and ThiobaciZZus acidophiZus,
which are the acidophilic members of the genus ThiobaciZZus, is
eliminated .. ThiobaciZZus ferrooxidans is also an iron oxidiser,
obtaining its energy requirements from the oxidation of inorganic
ferrous ion to ferric ion; since BJR-451 could not oxidise ferrous
salts, it cannot resemble this species.
ThiobaciZZus denitrificans oxidises sodium thiosulphate anaero
bically in the presence of nitrate as hydrogen acceptor, producing
nitrogen and the incapability of BJR-451 to grow anaerobically in
the presence of nitrate distinguishes it from ThiobaciZZus denitri
ficans. BJR-451 is obli~atelv autotrophic which identifies it as facul tatively
different from the L autotrophic and mixotrophic species of
the genus ThiobaciZZus (i.e.,ThiobaciZZus deZicatus, ThiobaciZZus
rubeZZus, ThiobaciZZus perometaboZis, ThiobaciZZus intermedius and
ThiobaciZZus noveZZus). The only possibilities left are ThiobaciZZus
thiopa:rus and ThiobaciZZus neapoZitanus . The GC fraction of BJR-451 's
DNA was 64.8%, a value markedly different from that of ThiobaciZZus
neapoZitanus (56-57%), but comparable to ThiobaciZZus thioparus
(62-68%). Thus BJR-451 bears a close resemblance to this species in
its characteristics.
The fatty acid methyl ester (FAME) profile of ThiobaciZZus
thioparus contains a major fraction of higher fatty acids (Type II,
Table 1.2.6 & 1.2.7). The fatty acid methyl ester profile of BJR-451
as described in Table 3.2 differs from the standard profile for
ThiobaciZZus thiopa:rus described in Bergey's Manual of Determina-
tive Bacteriology (Buchanan & Gibbons, 1974); the major fatty acid
component was oleic C1 a: 1 instead of saturated c15 component.
Anomalies in the fatty acid methyl ester profile have previously
been reported for the FAME profile of a ThiobaciZZus thiooxidans
strain which was found to vary considerably from the standard

128
description (Levin, 1971). It is well established that variations in
the cultural conditions of many organisms are often reflected by qual
itative changes in the fatty acids produced by them ( O'Leary, 1962 &
1970). Fatty acid profiles of ThiobaciZZi have been found not to
be affected by changes in cultural parameters such as phosphate
concentration, temperature, agitation (Levin, 1972; Agate & Vishniac,
1973-a & b). The effect of changes of pH, oxygen tension or other
nutrients on the fatty acid composition of ThiobaciZZi have recently
been studied by Dunn et al. (1977).
Heterotrophically grown cells of ThiobaciZZus noveZZus have
been reported to contain more unsaturated ( C1a:1; C19 ) fatty eye
acids, the percentage of saturated fatty acids ( c18 & c19) was
considerably increased when the cells were grown autotrophically
(Levin, 1972). Although the fatty acid profile of BJR-451, bears
a resemblance to the pattern of fatty acids found in ThiobaciZZus
noveZZus, described by Levin (1972), in certain other characteristics,
especially its obligately autotrophic nature, it resembles more
closely ThiobaciZZus thioparus.
BJR-Kl
This a short, Gram-negative rod, about 1-2 µ in length and 0.5
to 0.75 µ wide. Cells occur singly and are non-motile. It
oxidises ferrous to ferric ion in liquid cultures and produces a brow
nish precipitation when cultivated at pH 2.5. When cultivated at pH 2.0
or below, the brownish precipitate does not appear and a culture with

129
uniform turbidity is produced.
All attempts to grow the culture on ferrous sulphate agar as
described by Tuovinen & Kelly (1973) failed, indicating the extreme
toxicity of organic matter. Small, circular, brownish colonies were
obtained on silica gel plates containing ferrous sulphate. These
colonies were initially brown and in the latter stages of growth, salts
precipitated around them.
When suspended in 9K medium of Silverman & Lundgren (1959) contain
ing sodium thiosulphate, at pH 4.0, the cells did not grow even if·
incubated for periods as long as three to five weeks. Elemental
sulphur could support growth of BJR-Kl; however, a lag of one week
occurs after the first transfer from mediwn with ferrous sulphate
to the medium containing elemental sulphur as the sole energy for~.
The organism was unable to grow on nutrient agar.
It is classified as Thiobaciiius ferrooxidans. Such classification
is decided upon after consideration of the following arguments which
have been put forward previously in the literature.
In 1947, Colmer & Hinkle reported an organism capable of partici
pating in the aerobic oxidation of ferrous ion in mine waters. The
organism was able to oxidise sulphur and thiosulphate and was mor
phologically very similar to the previously known sulphur oxidising
bacteria, Thiobaciiius thiooxidans. _
Further physiological studies of the organism showed it to be an auto
trophic, aerobic organism, capable of proliferation at low pH values
C Colmer et al., 1950; Temple & Colmer, 1951; Lyalikova,1959, 1960).
According to its morphological similarities to other ThiobaciZZus species
and its ability to grow on inorganic media containing ferrous ion as
the only oxidisable source of energy, it was classified as ThiobaciZZus
ferrooxidans (Colmer et al., 1950).
A similar organism was isolated from analogous environments by the

130
Leathen group in 1949 (Leathen & Madison, 1949; Leathen & Braley,
1954; Leathen et al., 1956). The organism was found to be capable of
oxidising ferrous ions only and not sulphur and its other reduced
compounds. The name Fer>robaaiUus ferrooxidans was suggested for it
by Leathen & Braley (1954).
Later in 1960, Kinsel isolated a new species Ferrobaaillus
sulfooxidans, which was very similar to the Thiobaaillus ferrooxidans
and Ferrobaaillus ferrooxidans species already mentioned. Kinsel's
organism was similar to the other two in that it could utilise ferrous
alone as the sole source of energy but differed from them in its ability
to use elemental sulphur as the sole source of energy i.n the presence of
ferrous ions. Thus, there appeared three descriptions of closely related
bacterial types, all capable of autotrophic growth on media containing
ferrous sulphate as the energy source but differing in their ability or
inability to utilise elemental sulphur or thiosulphate as a sole source
of energy. Lyalikova (1960) consequently raised doubts as to the
independent e.~istence of ThiobaaiUus ferrooxidans & FerrobaaiUus
ferrooxidans as separate species. Works of other investigators
describe these three. closely related, autotrophic microorganisms
(Beck & Elsden, 1958; Bryner & Jameson, 1958; Beck, 1960). Ivanov &
Lyalikova (1962) discussed the various characteristics of these three
strains and pointed out the following:-
1- The inability of Leathen's strain to utilise thiosulphate
was probably. due to the fact that it was an old culture which
had been repeatedly grown on ferrous ion, which causes a loss of
thiosulphate oxidising ability in Thiobaaillus ferrooxidans,
(Starkey,1935; Vishniac & Santer, 1957; Lyalikova, 1959).
2- Insufficient proof for inability of Fer>robaaillus ferrooxidans
to oxidise elemental sulphur was obtained by Leathen et al. ( 1956 ) .
3- Production of elemental sulphur by the spontaneous decompo-

131
sition of thiosulphate in sterile acid media, places in doubt the
statement about the inability of FerTobaciZZus ferrooxida:ns
to utilise elemental sulphur as the sole source of energy.
With these points in mind these three strains in question could
well be identical and Ivanov & Lyalikova (1962) suggested that the name
ThiobaciZZus ferrooxidans should be retained for all three. Recently,
Kelly & Tuovinen (1972) also suggested that the names FerrobaciZZus
ferrooxida:ns and FerrobaciZZus suZfooxida:ns should be regarded only
as subjective synonyms of ThiobaciZZus ferrooxidans, since the so
called differences in their abilities to oxidise sulphur or thiosulphate
are invalid in as much as all three can grow. on either sulphur or
thiosulphate. Consequently, BJR-Kl was named ThiobaciZZus ferrooxidans.
The pure strain of BJR-Kl was found to be capable of oxidising both iron
ferrous and elemental sulphur when present as the sole energy source.
It resembled the strain described by Colmer & Hinkle (1947) in its
inability to oxidise thiosulphate and its ability to oxidise sulphur
only slowly during its first transfer to sulphur medium. After the
second transfer the ability to oxidise sulphur was recovered markedly
and there was no prolonged lag period.
The strain was strictly autotrophic and could not be adapted to
grow on glucose; organic matter in traces was toxic and growth was
inhibited. Transition from autotrophic strains to organotrophic types
has been studied by Shafia & Wilkinson (1969) who reported that two
out of eight strains of ThiobaciZZus ferrooxidans were capable of
growing on sucrose. They also dernonstrate'd that putative facultatively autotro-
phic strain (KG-4) of ThiobaciUus ferrooxida:ns which was capable
of growing on glucose lost its ability to oxidise ferrous iron.
Oxidation of ferrous ion to ferric being the only real criterion for
ThiobaciZZus ferrooxidans, the taxonomic position of such organotrophic

132
strains becomes highly dubious. In .fact, various workers have suggested that
so called standard strains of ThiobaciZZus ferrooxidans may be a
heterogenous mixtures, associated with some other heterotrophic satellite
(Tuovinen & Kelly, 1972; Zavarzin, 1972). This idea gets further
support from a recent report describing the isolation of a facultative
autotroph, ThiobaciZZus acidophiZus from a standard culture of
ThiobaciZZus ferrooxidans (Guay & Silver, 1975). The strongly inter-
dependent nature of the various components of such heterogenous asso
ciations is further indicated by the report that an apparentorganotrophic
strains of ThiobaciZZus ferrooxidans (KG-4) also possess small iron
oxidising population (Tuovinen & Nicholas, 1977).
Attempts to isolate some heterotrophic satellites from a suspen
sion of BJR-Kl, by method used by Guay & Silver (1975) for isolating
ThiobaciZZus acidophiZus
of such heterotrophic form.
were unsuccessful, indicating the absence
Although this culture of BJR-Kl was developed
from a single colony, the purity of culture containing one strain
cannot be ascertained since the culture developed only from one cell can
be called as a pure culture.

133
BJR-K0l
This strain is a Gram-negative rod, approximately 1-2.5 µ in
length and 0.5-1.0 µ in diameter. It is motile and non-sporula-
ting. The flagellum could not be detected by ordinary flagella stains
but detached flagella were observed in the electron micrographs of young
cultures which had been negatively stained with phosphotungstic acid.
The cells were examined by electron microscopy at different growth
phases but in no case were flagella attached to the cells detected.
This suggests an extremely fragile nature.
The colonies on thiosulphate agar (pH 4.0) were very tiny varying
from 0.5 - 1.0 mm in diameter. Initially, colonies were whitish yellow
in colour due to the precipitation of elemental sulphur. As they became
old their edges were cleared due to the oxidation of precipitated sulphur.
Growth in thiosulphate broth with 9K medium of Silverman & Lundgren (1959)
was very slow and produced uniform turbidity. When inoculated into
9K basal salt solution containing elemental sulphur at pH 2.5, copious
growth was obtained. The pH of the broth was decreased to 0.5. When
inoculated into 9K basal salt solution at higher pH value ( 6.5),
in the presence of elemental sulphur, no growth could be detected.
Both nitrate and ammonium nitrogen were utilised as nitrogen source,
with preference towards the latter. Nutrient agar did not support
growth and the microorganism did not exhibit any ability to oxidise ferr
ous iron. Strain BJR-K0l is an acidophilic sulphur oxidising
bacterium which can grow profusely at low pH values, oxidising elemental
sulphur for its energy requirements. It was unable to oxidise ferrous
salts and in this and other respects resembles ThiobaeiZZus thiooxidans
description.

134
3.2 OPTIMISATION OF ISOLATES
The pure isolates obtained as described earlier (section 3.1)
were studied for their optimum growth conditions and nutrient requi
rements.
Erlenmeyer flasks of one litre capacity containing 200 ml medium
were frequently employed for growth studies of the isolates as well as
for growing inoculum. For batch studies, 190 ml of Vishniac & Santer
medium (1957) was distributed into Erlenmeyer flasks of one litre
capacity and sterilised at 15 psi for 15 minutes. Sodium thiosulp
hate (40%, w/v) solution was sterilised separately by filtration
through a membrane filter. 5 ml sterilised thiosulphate solution were
dispensed aseptically into the flask containing Vishniac & Santer med
ium. 5 ml inoculum was used and flasks were shaken on a reciprocal
shaker ( 5 cm stroke; 90 ± 2 strokes per minute) at an appropriate
temperature.
A fermenter, in which temperature and pH were controlled and
oxygen tension was measured continuously, was employed for determining
the optimum pH and temperature for BJR-451. At regular intervals
5 ml samples were withdrawn aseptically from the flasks/ fermenter and
total cell count was determined immediately as described in ,section
2.5.5. The sample was centrifuged at 10,000 . g for 15 minutes, and
residual sodium thiosulphate was determined in the supernatants by
the two procedures already described in section 2.2.1.

3.2.1 SELECTION OF AN APPROPRIATE GROWTH MEDIUM FOR
NON-ACIDOPHILIC BJR-451
135
The non-acidophilic sulphur oxidiser, BJR-451, had been isolated
on media_ designed by Beijerinck (1904) and Starkey (1934-a). As
mentioned earlier (section 3.1) relatively poor rates of thiosulphate
oxidation after 4-5 transfers on the same medium indicated that the
culture had lost its capacity for vigorous growth on this medium.
None of the trace elements required as enzyme-cofactors, which are
essential for growth (Pfenning, 1961), are added to Starkey's medium.
The medium of Vishniac & Santer (1957), which contains a wide spectrum
of trace elements, was therefore used in a comparative study of the
growth of BJR-451 in each medium concurrently.
Erlenmeyer flasks of one litre capacity containing 180 ml of
basal salts solution of Vishniac & Santer or Starkey medium,
were sterilised by autoclaving at 15 psi for 15 minutes. 20 ml of
sodium thiosulphate (10 %, w/v) previously sterilsed by filtration
through a membrane filter were added to each flask. The flasks were
then each inoculated with 10 ml of a freshly prepared inoculum of
BJR-451 and incubated at 30 ± 0.5°C on a reciprocating shaker ( 5 cm
stroke; 90 t 2 strokes per minute). Thiosulphate oxidation and changes
in pH were measured at regular intervals of time.
In the Vishniac & Santer medium 100% oxidation of thiosulphate
was achieved in 48 hours, as compared to 184 hours in the Starkey-
medium (Fig. 3.2.1). The stimulation was particularly apparent in
the fonner medium's ability to decrease the lag period before
thiosulphate consumption and acid production.( Maintenanace of the
pH at 5.0 for a relatively long duration in the Starkey medium
reflects the buffering capacity of this medium at pH 5.0. This pH
value was not achieved however, witil 18 hours after all the
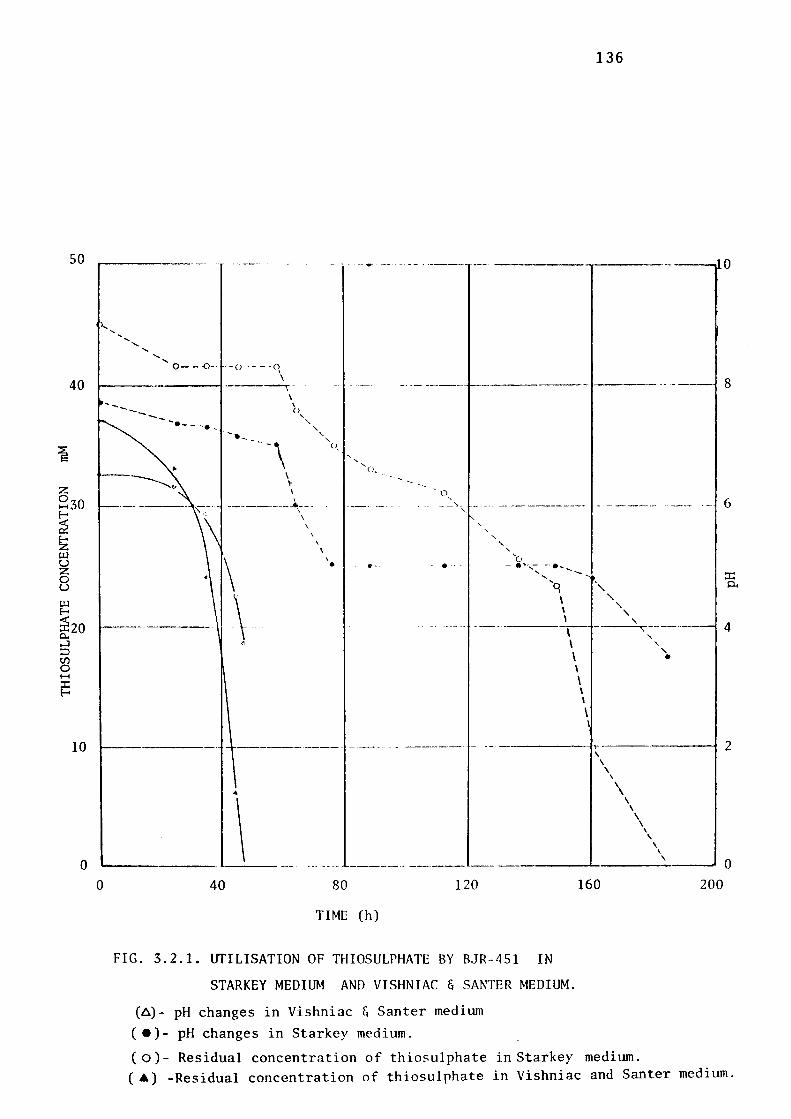
50
40
10
0
136
-------- - - ... -- --------···---·----------- --- ------------ --..... ~10
.... .... ......
...... ...... .....
0---0-· -o ----o \
---- -----1------,- .... -------1----·-··--- 8
0
------ ~-- - -· - ·-\
\
\ II
()
' ' ' '
___________ _., __ -\
\
\
\
\
' o_
\ • -•--
o, ---~- - ---- ---- -------- --------· ---- ·- 6
' '
.... ' ' '
'c., - •'"-,---- .... _
.... ...... -- - ~ '9 ·,
\ ' ' '
\ \ I
I -·- ·----- -----------'---'.:....._----------- 4
\ ' a \ \ I \ I I
\ \
' ' •
-------··---- ----. - - ------- - . -- --------',· ------~
..
\ 40
------- -----80
TIME (h)
120
FIG. 3.2.1. lITILISATION OF THIOSULPHATE BY BJR-451 IN
STARKEY MEDIUM AND VISHNIAC & SANTER MEDIUM.
(6)- pH changes in Vishniac & Santer medium
C•)- pH changes in Starkey medium.
\ \
\
' \ \
\ \
\
160
( o)- Residual concentration of thiosulphate in Starkey medium.
' \ \
\
' 200
2
0
( •) -Residual concentration of thiosulphate in Vishniac and Santer medium.

137
thiosulphate of the Vishniac & Santer medium had disappeared.)
The results clearly indicated the beneficial effects of the
trace elements in the Vishniac and Santer mediwn, which was therefore
adopted as the medium in which further growth optimising studies
were conducted.
3.2.2 BATCH GROWTH OF BJR-451 IN FLASK
pH, thiosulphate and cell number were monitored during growth,
under conditions identical with those described in section 3.2.1.
According to the pH curve, oxidation of thiosulphate involved
three stages (Fig. 3.2.2). During the first stage, which lasted over
the first 12 hours, the pH decreased gradually from 6.5 to 5.6, 18%
of the thiosulphate was oxidised and the bacterial count increased
slowly. These changes are typical of the lag-phase of a microbial
process.
In the next stage, which lasted over the next six hours, the
remaining 80% of the sodium thiosulphate was oxidised, and the cell
number increased whilst the pH remained stable at 5.5.
The third stage, which was the stationary phase according to
cell counts, was characterised by active production of hydrogen ions
(pH decreased from 5.5 to 3.7). During this phas~ the cultural broth
became turbid, due to precipitation of free elemental sulphur. If
allowed to continue for 60-80 hours, the culture became sterile,
preswnably as result of sensitivity to low pH.
3.2.3 BATCH GROWTH OF BJR-451 IN FERMENTER
Further information regarding the growth parameters of BJR-451
was obtained by cultivating it at 30°C in a fermenter. pH, dissolved
oxygen, cell number and residual thiosulphate were monitored over the
course of 300 hours. Fig. 3.2.3 reveals that the patterns of
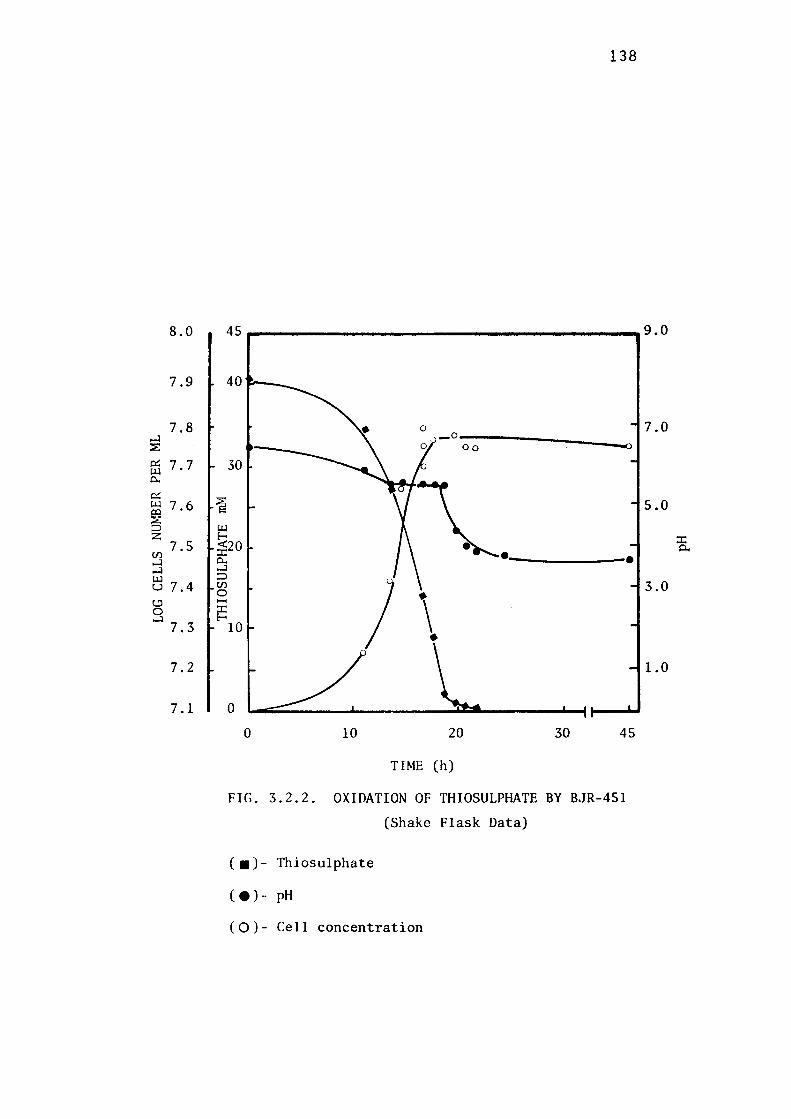
8.0
7.9
7.8 ,-J :E: 0:: 7.7 w 0..
0:: w 7.6 ~ :::i z
7.5 r.J) ,-J ,-J w u 7.4 c.:, 0 ,-J
7.3
7.2
7.1
138
45 ----------------------9.0
40
_o Oo
30
i w E--<20 ::c: 0.. ~- • ,-J :::i r.J) 0 ..... ::c: E-
10 .\ •
0
0 10 20
TIME (h)
30 45
FIG. 3.2.2. OXIDATION OF THIOSULPHATE BY BJR-451
(Shake Flask Data)
( • )- Thiosulphate
( e )- pH
( 0 )- Cell concentration
7.0
5.0
3.0
1.0
::c: 0..

139
utilisation of thiosulphate, changes in pH, and cell growth were very
similar to those recorded previously in Fig. 3.2.2. However the
experiment also revealed that the thiosulphate was initially converted
to polythionates(rnainly tetrathionate) and that during this phase,
which included phases 1 and 2 of the previous experiment, growth
occurred and· the pH changed little. Thiosulphate was completely
exhausted by 38 hour, after which the oxygen consumptjon rate
decreased markedly (as indicated by a sudden increase in the dissolved
oxygen tensiom from 57% to 93% saturation). During the next six hours
the dissolved oxygen tension decreased to 65% saturation as 30% of
the polythionate was consumed and acid was produced. By the time the
pH had decreased to 4.8, polythionate consumption had ceased and the
oxygen tension had begun to increase again. It continued to increase
to a final value of 98% saturation, whilst more acid was produced.
Cell growth continued, at a decelerated rate, during the period of
polythionate consumption, and then growth and polythionate consumtion
terminated concurrently. In the very late stages of culture (50-300
hour) cell lysis was accompanied by oxygen uptake (inidicated by a
decrease in oxygen tension) and continued acid production whilst the
polythionate concentration remained unaltered.
Since ThiobaciZZus thioparus is not acid tolerant, it was
presumed that cessation of biological activity was enforced by the
acidic environment produced by thiosulphate oxidation. Previous
reports also have indicated the lethal effects of low pH environments
on the growth and viability of ThiobaciZZus thioparus(Parker & Prisk,
1953). In order to confirm the deleterious effect of pH, ThiobaciZZ
us thioparus was grown in the fermenter tmder conditions which were
identical to those described above, except that, when the thiosulphate
had been totally depleted, the pH was automatically controlled at 5.5
by the addition of sterilised sodium hydroxide solution (2N). The
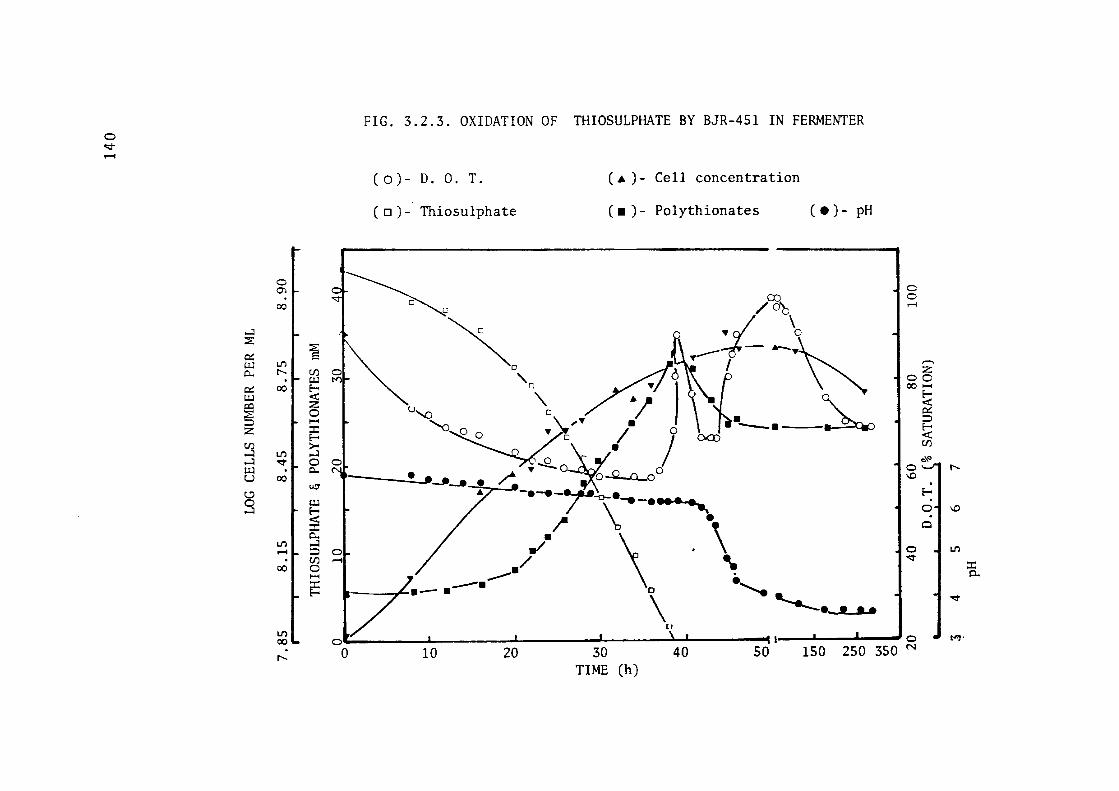
0 "::T ..-i
...:I ::E
0:: i:.u c. 0:: i:.u Q:l ~ ::) z Cl.) ...:I ...:I i:.u u C.!)
3
0 0)
00
1./) r--
00
1./)
-.:I"
00
1./) .-I . 00
1./) 00
r--
FIG. 3.2.3. OXIDATION OF THIOSULPHATE BY BJR-451 IN FERMENTER
(0)- D. 0. T.
( o ) - Thiosulphate
( .t. )- Cell concentration
( • ) - Polythionates ( • )- pH
~ /~
~ ~ ~ .--:I--:~ ~~ \, Ao ' E- ri • i <t: '\ &
5 "'° 0 / ) ~ 'o~o .)l.-• ;• o ·-•-~
>- ~: I 00) ~ o - A ®-"-AD / T -2..~ \ '.!;": Q 00
& -----~-~ /~~-~~ :2 ;- \ • c. • \ \ So ~ · ~ .-I / ...
\ "--•-t ....
C 0 .-I
,-.. z
00 00 .....
0
'°
0 -.:I"
E<t: 0:: ::)
~ Cl.)
0\0
. E-
0 . 0
~ 7·-.-----· 0 .'---.
[I
o-----'--------L-------IL...---.....l..--'-----... , _ __._ _ __,_ __ o
0 10 20 30 40 50 150 250 350 N
TIME (h)
r--
'° 1./)
~
t'-J·
:I: 0..

FIG. 3.2.4. OXIDATION OF THIOSULPHATE BY BJR-451 IN FERMENTER
( o )- Thiosulphate ( • ) - Polythionates ..--i
o:i- ( e) - D. 0. T. ( 6) - pH ..--i I i----------------------0 01"" O 100 ~\ ~
\ _.,.,__ r·· ..,,..:.,...-1:,._ q --0 "7.A~ e
~ I
o \ /~ i '-. IO ; r') ~{ '\. t • , 00 , l,C)
~ · r;-,___ . ___ I \. Z O • 'v" •-l - • \ 0 A,;, 1 ,-, ~ I -- -:-_ - I • ~
>- ~ " ~ I E-~ ...:I • / \ •• 0 0 • ~ \ C. N 'J / Cl \. • o;::,, ~ ut; \ \ ~ • .; '°~
...:J \ ·-· J C/)1 ::c: Ul I ~ a\ / \. f o,\'> 0-, E- ' . ~ E- 9.. . ......,,
! \K .. (\\ / 8 ~\,\ I ~ Cl) • r ::c:
~ ~ /• \ f 0.. \ j O o, N E- \ .. I, I~
/ lr
.,, . r! \ \ I
~ i ~ l 0 0 ___ ...._ __ __._ ___ ..__..;i..-&... __ __., ___ .._~ N - 0
0 20 40 60 80 90 100 110 120 130 140
TIME (h)

142
results,• shown in Fig. 3.2.4, revealed that oxygen consumption was
restored (as indicated by a decrease in oxygen tension) as soon as
the pH was restored to 5.5 and continued until the polythionates
were completely utilised. This confirms the observation of many
workers, including Parker & Prisk (1953) that tetrathionate was
completely oxidised by ThiobaciZZus X, when the pH of the cultural
broth was controlled manually.
Again, as in the test recorded in Fig. 3.2.3, cell growth
occurred during tetrathionate consumption but at a slower rate than
when sodium thiosulphate was present as the energy source.
The experiment confirmed that pH control is vitally important
to this organism and that pH values below 4.8 drastically reduce its
activity and viability.
3.2.4 DETERMINATION OF THE OPTIMUM pH FOR GROWTH OF
BJR-451
The pH optima for growth and thiosulphate utilisation were
determined by cultivating this bacterium at 30 ± 0.5°C, in a
fermenter, at various pH values. The effect of pH over the complete
growth cycle was monitored by analysis of 5 ml samples collected
aseptically at regular intervals from the fermenter.
pH 5.5 proved to be optimal for both growth and thiosulphate
utilisation. Figs. 3.2.5 - 3.2.8 demonstrate the greater sensitivity
of growth and thiosulphate utilisation to decrease, as compared to
increase in pH. The growth pH optimum was so well defined in this
respect that growth rate decrease to 1/l0th when the pH was decreased
from 5.5 to 5.0. Due to extremely slow growth rate, growth curve
for pH 5.0 in Fig. 3.2.7 could not be shown.
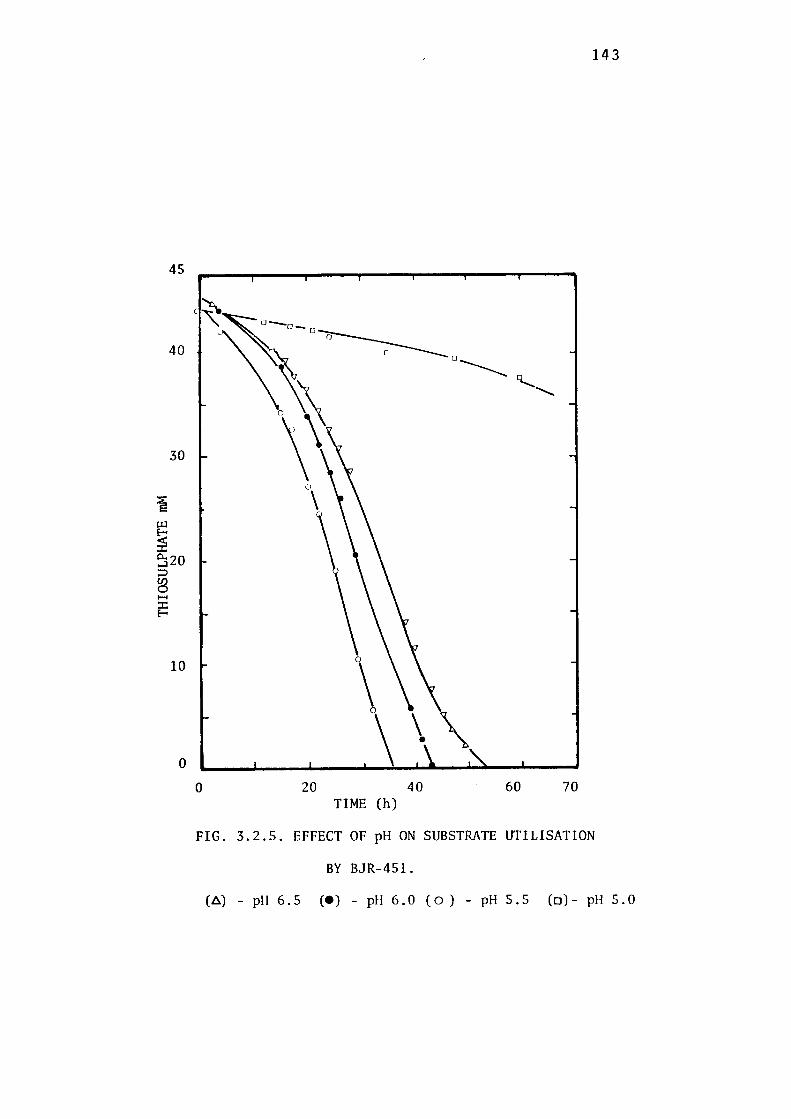
45
30
10
0
0 20 40 60 70 TIME (h)
FIG. 3.2.5. EFFECT OF pH ON SUBSTRATE UTILISATION
BY BJR-451.
143
(6) - pH 6. 5 (e) - pH 6. 0 ( o ) - pH 5. 5 (o) - pH 5. 0
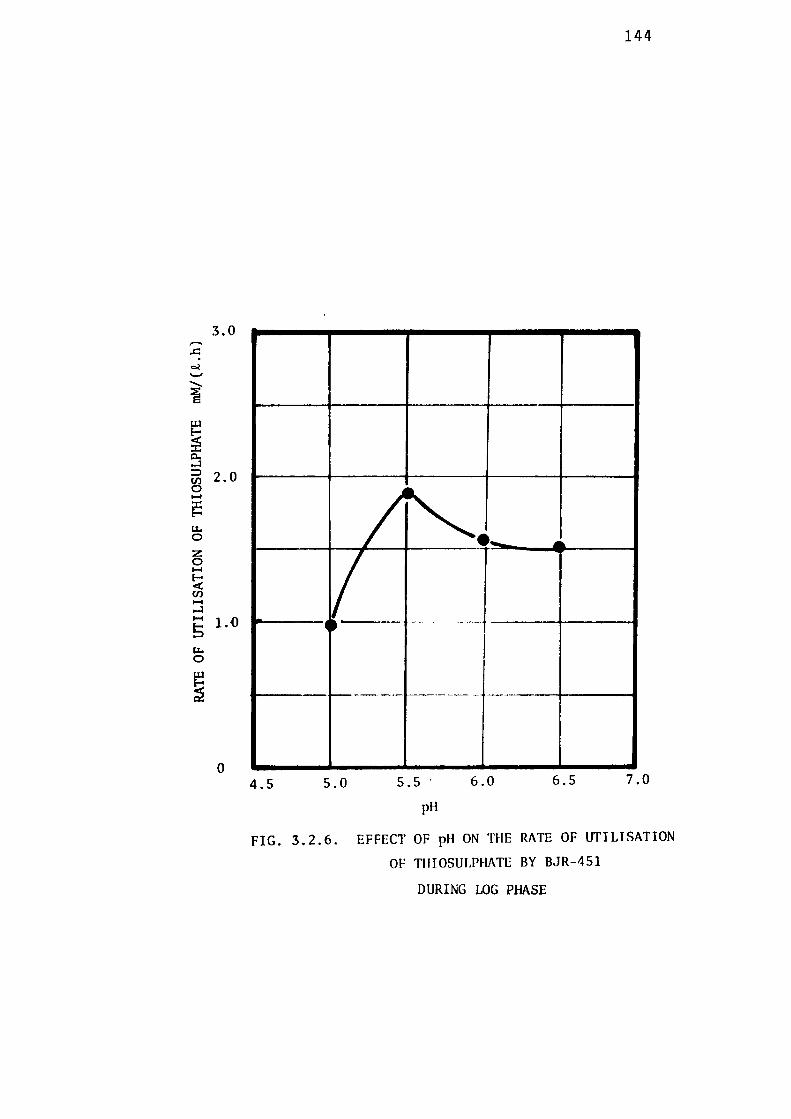
3.0 ,-.. ..c: o.l .._, -i IJ.l E-
~ 0. ,-J ~ 2.0 t/) 0 1-t :c E-u. 0
z 0 1-t E-< t/) 1-t ,-J 1-t 1.0 5 u. 0
5
0
144
~-- ...
I ----·-- -----~ ,
------- -----
4.5 5.0 5.5 6.0 6.5 7.0
pH
FIG. 3.2.6. EFFECT OF pH ON THE RATE OF UTILISATION
OF THIOSULPHATE BY BJR-451
DURING LOG PHASE
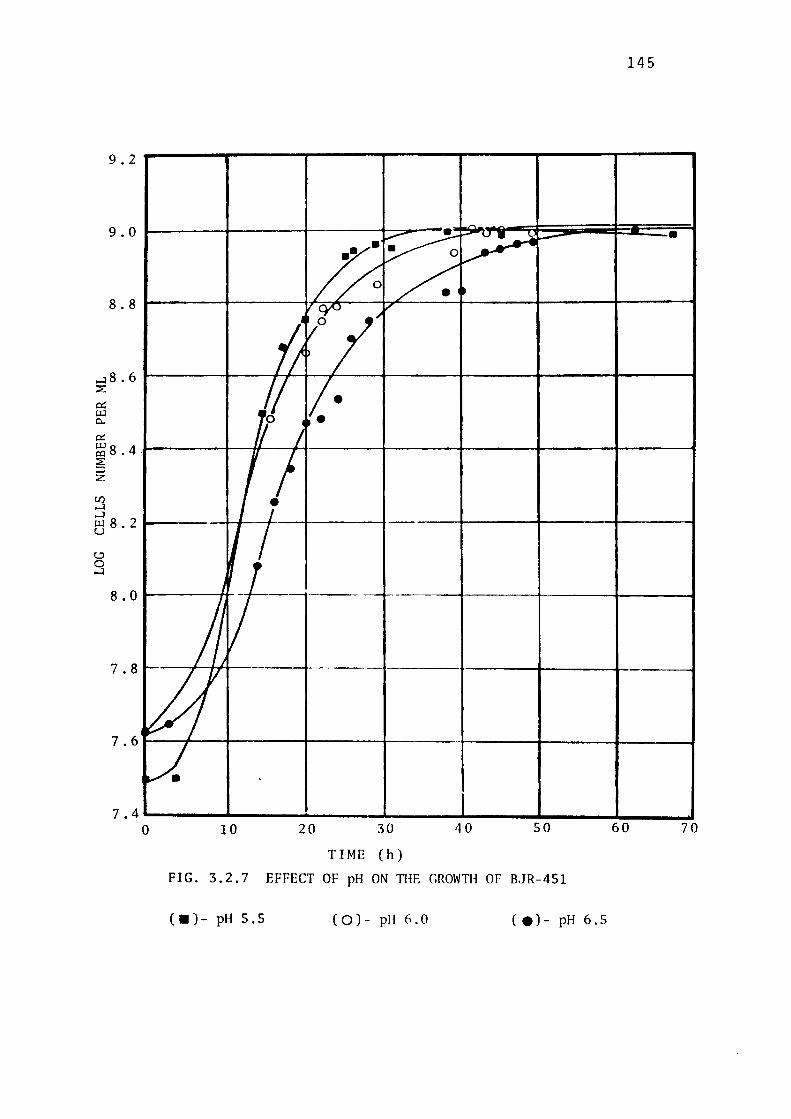
145
9.2
....:i 8 . 6 1------+----I-J~-~--~---+----+-----+-----t ~
0:: µ_J 0..
0:: ~ 8 . 4 --------+--H--+--+----1-----+-----+-----t-------,
~ z
7_4._ _______ ..._ ___ .,_ ___ .,_ __ __,,,.,_ __ ...,ii.... __ _,
0 10 20 30
TIME (h)
40 50
FIG. 3.2.7 EFFECT OF pH ON THE GROWTH OF BJR-451
( • )- pH 5 .5 ( 0 )- pH 6 .0 ( e) - pH 6 .5
60 70
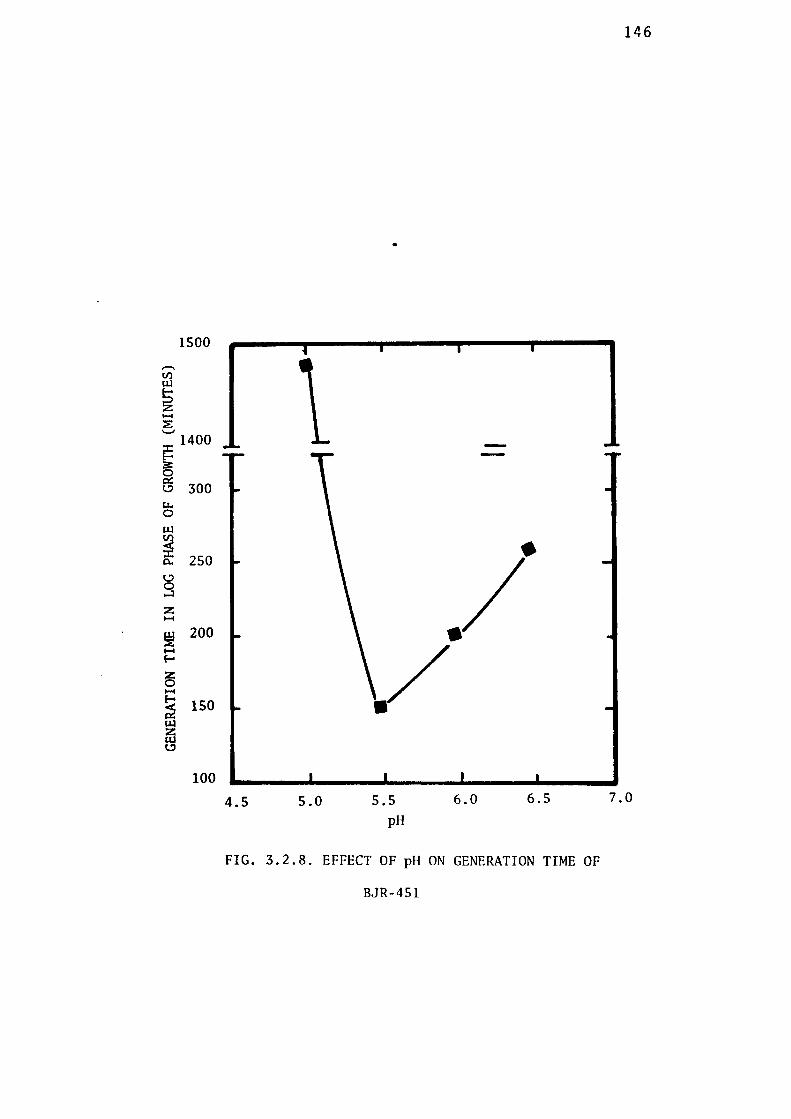
,....... U) riJ
Ei z 1-1 ~ '--,j
1500
~ 1400
~ (3 300 I.I-, 0
~ 250
§ z 1-1
100
4.5 5.0
•
5.5 pH
6.0 6.5
FIG. 3.2.8. EFFECT OF pH ON GENERATION TIME OF
BJR-451
146
7.0

147
3.2.5 DETERMINATION OF THE OPTIMUM TEMPERATURE FOR
GROWTH OF BJR-451
The effects of temperature variation on growth and sodium thios
ulphate utilisation by BJR-451 were studied over a temperature range
of 25-40°C in a fermenter. The results are plotted in Figs. 3.2.9 -
3.2.12.
Figure 3.2.9 reveals that the utilisation of thiosulphate was
facilitated by rise of temperature from 25°C to 30°C, above which the
utilisation rate of thiosulphate decreased. Up to 35°C, the duration
of lag phase was not significantly affected, but at 40°C, the lag phase
was prolonged.
The effect of temperature on the rates of growth and of sodium
thiosulphate utilisation during the logarithmic growth phase are shown
in Figs.3.2.10 and 3.2.11, respectively. The shapes of the curves are
typical of the temperature dependence of biological reation rates.
The maximum growth ( generation time 150 minutes) and sodium
thiosulphate utilisation rates were obtained when BJR-451 was cult
ivated at 30°C.
The optimum of 30°C recorded for both parameters is in
agreement with values reported by other workers who quote a range
from 28°- 35° C for growth and thiosulphate utilisation (Vishniac &
Santer, 1957). At 40°C thiosulphate utilisation rate had decreased
to less than half that recorded at the optimum temperature, 30°C
(see Table 3.2.1).
Using the experimental data in Fig. 3.2.11,values for the
temperature coefficient [ Q10 ], as defined by the expression below,
were calculated and tabulated in Table 3.2.1.
= K 1
10 [ ---=-------,=- ]
; - T1 (Miller-& Litsky, 1976).
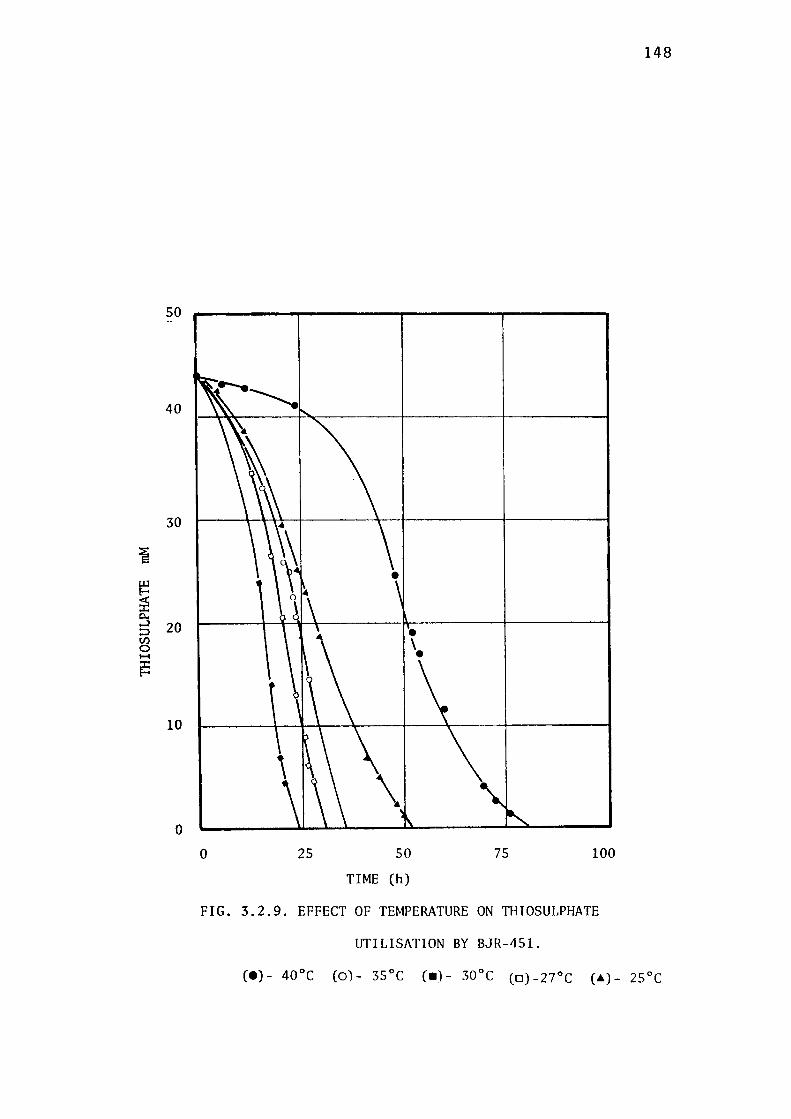
50
40
30
~
~ ~ 0.. ...l 20 :::> V) 0 1-4
~
10
0
0 25
•
so TIME (h)
'·
75 100
FIG. 3.2.9. EFFECT OF TEMPERATURE ON THIOSULPHATE
UTILISATION BY BJR-451.
148
(•)- 40°C (0)- 35°C (•)- 30°C (o)-27°C (•)- 25°C
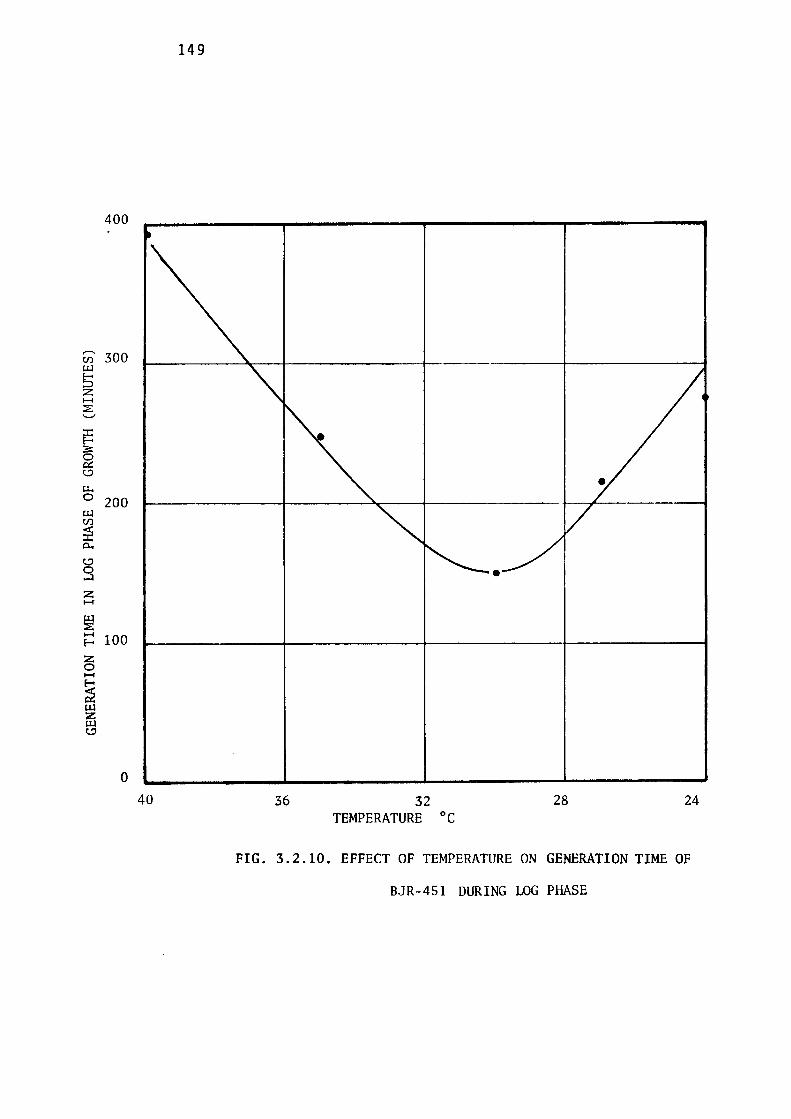
,-.. Cl) i:.u
~ z ~
~ ~
~ :3:: 0 ex: c.:,
'-1-. 0
i:.u Cl)
~ 0..
c.:, 0 ...J
z I-{
~ ~
E--
z 0 I-{
~ 1,.1.l z l'.-U c.:,
149
400
300
200
100
0 40 36 32
TEMPERATURE °C
•
28 24
FIG. 3.2.10. EFFECT OF TEMPERATURE ON GENERATION TIME OF
BJR-451 DURING LOG PHASE
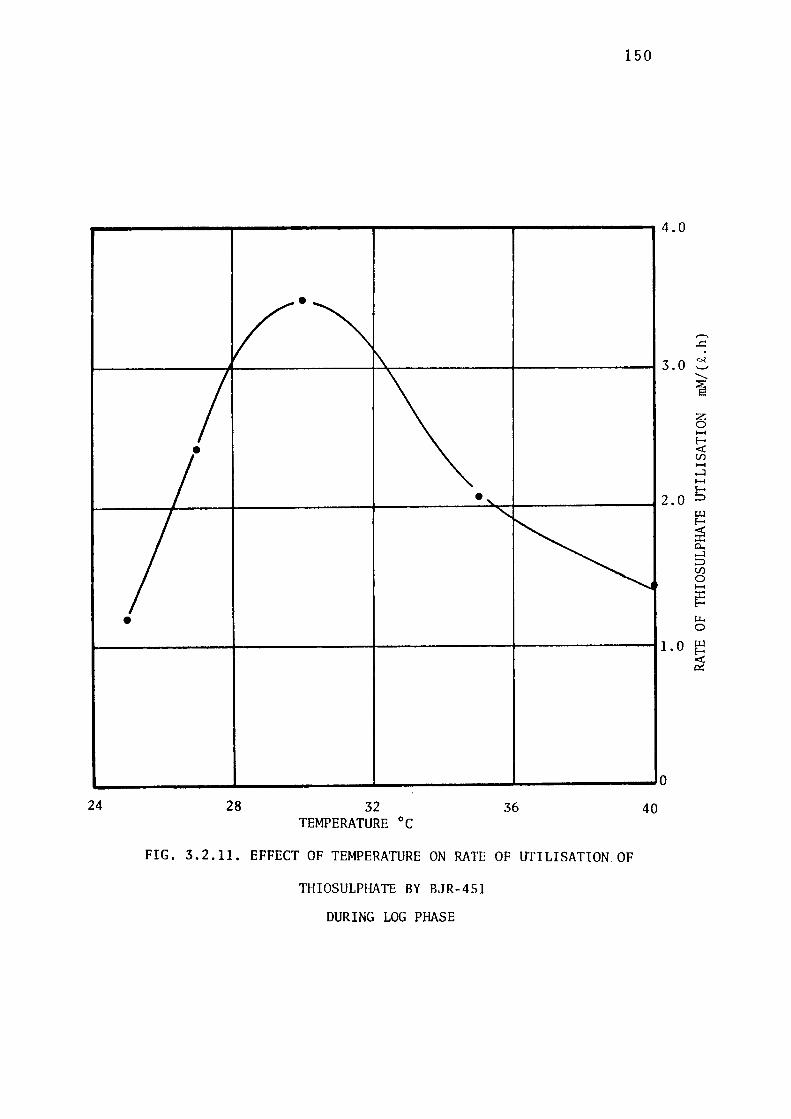
150
-------,-------r-------,-------,4.0
•
3.0
•
• 2.0
• 1.0
L..------L-----L--------'---------0 24 28 32
TEMPERATURE °C 36
FIG. 3.2.11. EFFECT OF TEMPERATURE ON RATE OF UTILISATION OF
THIOSULPHATE BY B,JR-451
DURING LOG PHASE
40
,-., ...c:: ~ .._, -~ z 0 ..... E-<i: en ..... ....J ..... ~ i:.u E-
~ 0.. ....J :::, en 0 ..... :I: E-
ii. 0
i:.u
~
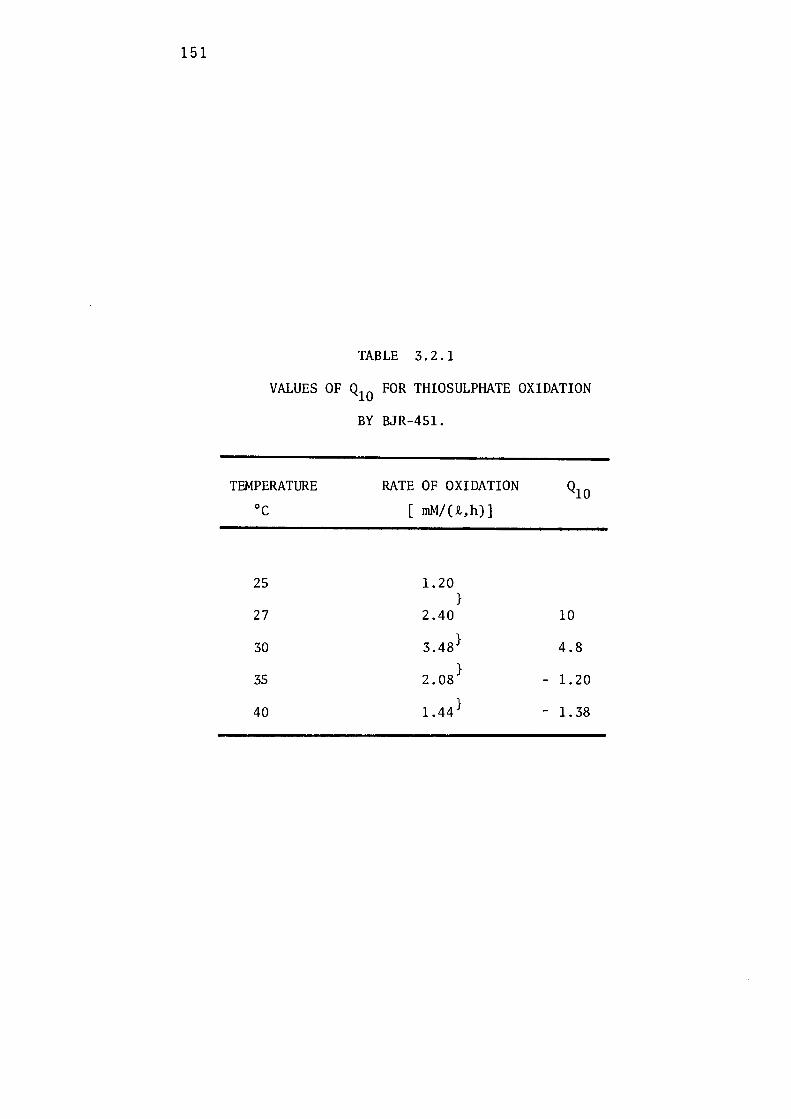
151
TABLE 3.2.1
VALUES OF Q10 FOR THIOSULPHATE OXIDATION
BY BJR-451.
TEMPERATURE oc
25
27
30
35
40
RATE OF OXIDATION
[ mM/(i,h)]
1.20 }
2.40
3.48}
2.08}
1.44}
10
4.8
- 1.20
- 1. 38

152
For chemical reaction Q10 values range from 2 to 4. Since
numerous chemical reactions are involved during the growth of bacteria,
this law is usually not upheld during bacterial growth except in the
middle of the active temperature range (Litchfield, 1976). Values
of Q10 obtained for utilisation of thiosulphate by ThiobaciZZus
thioparus are typical of bacterial behaviour, the Q10 value of 10
between 25 and 27° C decreasing to 4.8 as the optimum temperature
(30°C) was approached. Negative values were of course obtained
as the temperature was increased above its optimum value.

153
:3.2.6 GROWTH OF I3JR-451 IN A CHEMOSTAT
Organisms can be cultured continuously at specific growth rates
less thanµ in a chemostat, as first described by Novick & Szilard, max
1950. The maximum specific growth rate of the organism can be
calculated by growing it at a series of dilution rates. The exact
relationship between dilution rate (D) and the growth rate (µ)
of organisms in a chemostat can be calculated from the following:
Increase in bacterial concentration= Growth - Output
i.e., dx Dx dt = µx -
i.e., dx (µ D) [I] where x concentration of dt = X - = cells.
It follows from equation [I] that if dx µ > D, dt will be positive and
the concentration of organisms in the culture will increase with time.
On the other hand, if dx µ < D, then dt will be negative and the cell
concentration will decrease with time until the culture eventually
'washes out' of the fermenter. During steady state, the concentration
of microorganisms in the fermenter remain constant; therefore, dx dt is
zero, andµ will be equal to the dilution rate, D. The continuous
culture of organisms in a chemostat therefore depends on provid-
ing conditions in which the specific growth rate(µ) and the dilution
rate (D) are equal and invariant with time. Since the growth rate is
controlled by the dilution rate, it can be adjusted, up to an upper
limit, to any value desired. The upper limit is the critical dilution
rate (Dc), which approaches µrnax If the dilution rate is set to
a value greater than Dc, it will exceed the maximum growth rate (µmax)
and the culture will be progressivley washed out from the fermenter.
Determination of D is thus a measure of C
Thiobacillus thioparus, BJR-451, was grown continuously in the
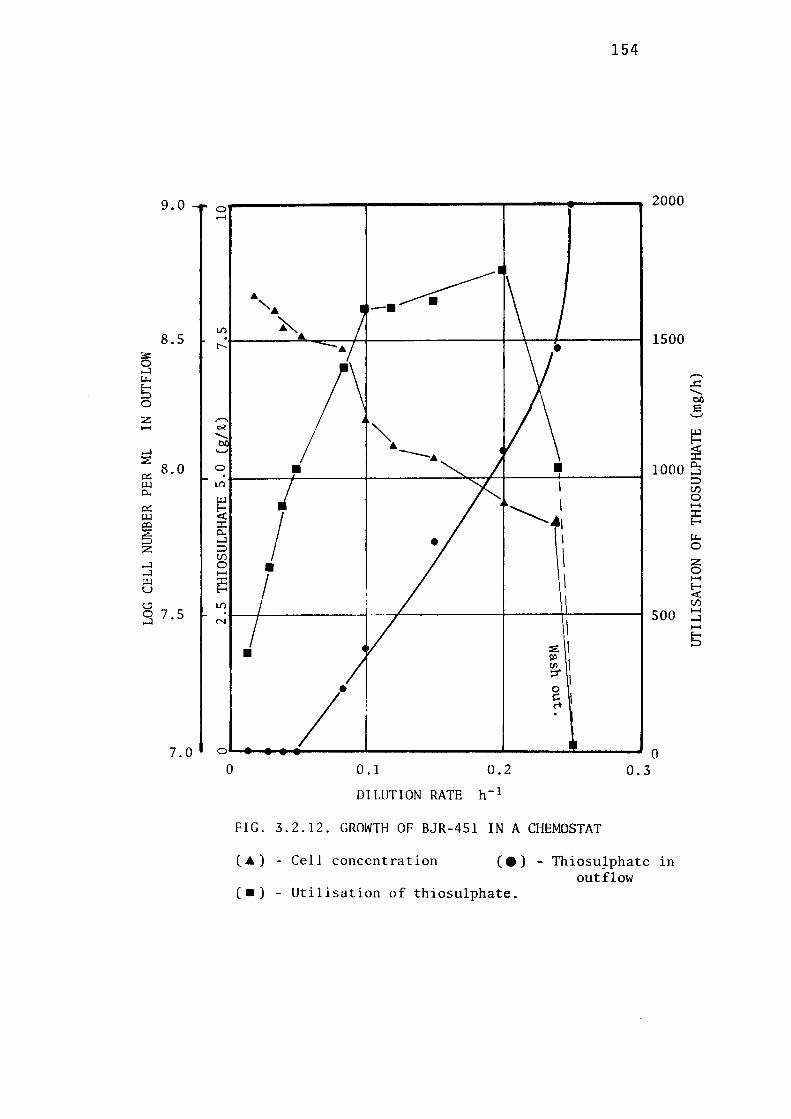
9.0
8.5
~ ...J u.. 5 0
z H
...J ~
8.0 i:i::: UJ Q..
i:i::: UJ ,:x:i
$ z ...J ...J UJ u C.:J 0 7.5 ...J
7.0
0 ......
L/'l . r--
,-.. ~ -._,
0
L/'l
UJ
~ ::r:: Q.. ...J :::> Cf) 0 1-1
::r:: E-
L/'l
N
0
0
154
2000
... --~ , ... ~
1500
,-... ..c:: -bi)
s ._,
UJ E-
:ii • • 1000 ~
:::> I Cf)
0 I H
I '---~, ::r:: E-
I u.. 0
\ z • 0 I H
E-<
\ Cf)
500 H ...J
I H
~1 5 • :1 ';j'I
•
I g I rt
0 0 .1 0.2 0.3
DILUTION RATE h-1
FIG. 3.2.12. GROWTH OF BJR-451 IN A CHEMOSTAT
( ... ) - Cell concentration (.) ThiosuJphate in outflow
( . ) - Utilisation of thiosulphate.

155
chemostat illustrated in Fig. 2.3 and described in section 2.5, with
sodiwn thiosulphate as the growth-limiting substrate. The pH was
controlled at 5.5, and the temperature at 30 ± 0.5°c, throughout the
cultivation. 1% (w/v) sodiwn thiosulphate was used in Vishniac &
Santer mediwn (1957) in the inflow (S ), and dissolved oxygen tension r
was maintained at approximately 50-60% saturation by passing sterilised
atmospheric air through the fermenter.
The cell number and substrate concentration in the outflow
during steady state at various dilution rates are shown in Fig. 3.2.12.
A maximum growth rate (dilution rate) of 0.24 h- 1 was recorded. A
steady state could not be obtained at a dilution rate greater than
o.24 h- 1 ; at a dilution rate of 0.25 h- 1 wash-out occurred. At low
dilution rates ( < 0.05 h- 1) there was a complete utilisation of
sodium thiosulphate. A conspicuous wall-film of elemental sulphur
containing microbial cells was developed at these dilution rates.
BJR-451, behaved· normally in the chemostat giving standard shapes of
its cultivation curves. During steady states, the cell population
remained constant, but as'wash-out' was approached the cell population
decreased with time.
ThiobaciZZus thiopa:rus and other ThiobaciZZi have been
frequently cultivated in continuous culture. For example, Kuenen &
Veldkamp (1973) cultivated a marine strain of Thio"baciZZus thiopa:rus
in a chemostat with thiosulphate as the growth-limiting substrate and
recorded aµ of more than 0.3. max
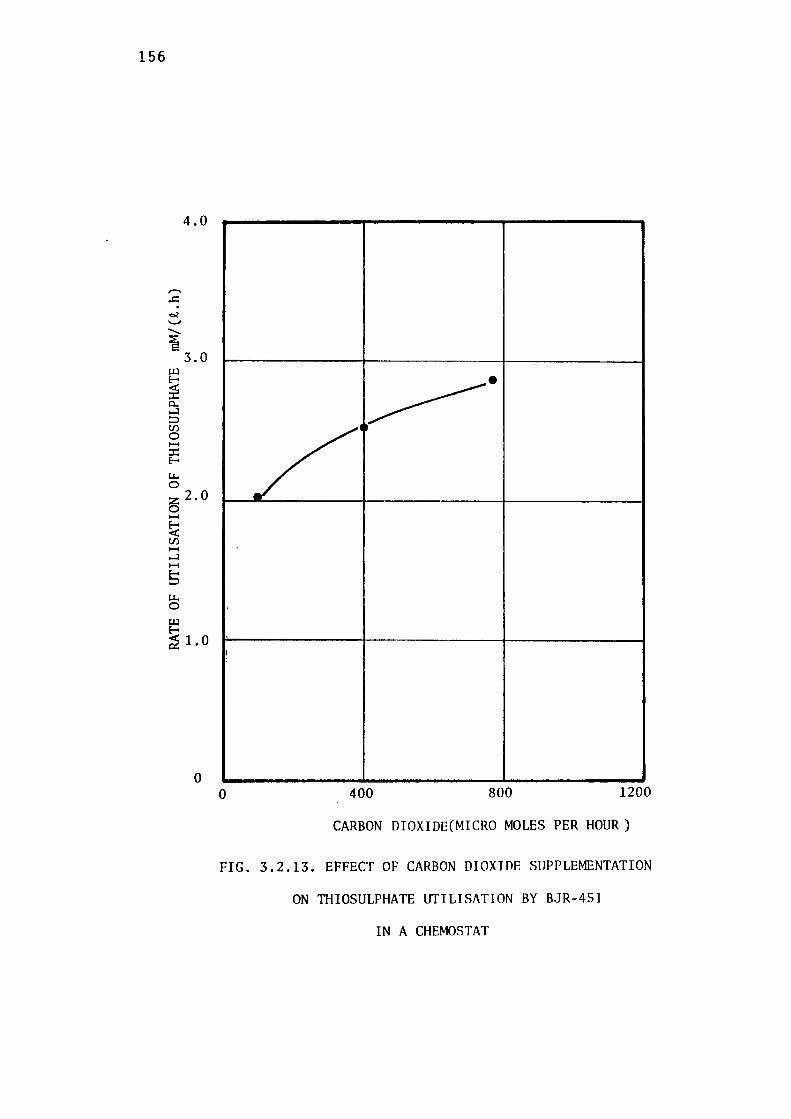
156
._,,
.........
~
4.0
3.0 U-l
~ 0.. ...J :::, U) 0 ..... :c: r' Lt. 0 :z 2. 0 0 ..... ~ U) ..... ...J ..... 5 Lt. 0
~ ~ 1.0
0
/' ~-
••
I
0 400 800 1200
CARBON DIOXIDE(MICRO MOLES PER HOUR)
FIG. 3.2.13. EFFECT OF CARBON DIOXIDE SUPPLEMENTATION
ON TIIIOSULPHATE UTILISATION BY BJR-451
IN A CHEMOSTAT

157
3.2.7 EFFECT OF CARBON DIOXIDE ON GROWTH OF BJR-451
ThiobaciZZi, like other autotrophic microorganisms, fulfil their
carbon requirements by fixing Co2 from the atmosphere. In this experi
ment the effect of carbon dioxide on the growth of BJR-451, while
growing in a chemostat, was studied.
Co2 present in atmospheric air was removed by passing it through
10% (w/v) solution of sodium hydroxide and scrubbing it with sterilised
water. C02 -free atmospheric air was mixed with known volumes of a
standard C0 2 mixture. BJR-451 was cultivated in a chemostat, with
thiosulphate as the growth-limiting substrate, at a dilution rate of
0.1 h- 1 (approximately 1/3 rd of itsµ ). max
At steady state, known amounts of C02 were fed into the fermenter,
and residual co2 in the gas-outflow was analysed by passing it
through an infrared gas analyser, as described in section 2.5.4.
The concentration of thiosulphate in the outflow was measured till a
new steady state was indicated by constant value.
Increased C02 in the chemostat up to 770 µmoles/hour resulted
in enhancement of thiosulphate utilisation, indicating that the system
was limited with respect to carbon source (Fig. 3. 213 ) . By increasing
the input of Co2 from 100 to 402 µmoles/hour, thiosulphate utilisa
tion rate was improved 2.0 to 2.5 rnM/(i.h), indicating that more
carbon was being fixed. Effect of carbon dioxide limitation during
leaching of zinc sulphide by acidophilic ThiobaciZZus ferrooxidans
have been studied by Torma et al. (1972), who showed that
with increased concentration of carbon dioxide, bacteria leached more
concentrate. However, it is generally suggested that carbon dioxide
contents in the air of a modern city are sufficient for supplying the
carbon dioxide demands of these bacteria.

3.2.8
158
TOXICITY OF ZINC IONS TOWARDS THIOBACILLUS THIOPARUS,
THIOBACILLUS FERROOXIDANS AND THIOBACILLUS THIOOXIDANS
Effect of zinc ion concentration on growth and metabolic activity
of three strains of ThiobaciZZi was tested in shake flasks. The
following procedure was used. For acidophilic members, the basal
salt solution of 9K medium (pH 2.5) of Silverman & Lundgren (1959)
was used. 180 ml of this basal salt solution were contained in
Erlenmeyer flasks of one litre capacity and sterilised by autoclaving
at 15 psi for 15 minutes. Ferrous sulphate solution (40%, w/v) was
sterilised separately by filtration. For ThiobaciZZus ferrooxida:ns,
10 ml of sterilised ferrous sulphate solution was added to the flasks
aseptically; and for ThiobaciZZus thiooxidans, 2% (w/v) sterilised
flowers of sulphur were added to the flasks aseptically. For Thiobac
iZZus thioparus, Vishniac & Santer medium (1957), (pH 5.5), (180 ml)
dispensed in to Erlenmeyer flasks of one litre capacity and the flasks
sterilised at 15 psi for 15 minutes. 10 ml sodium thiosulphate (20%,
w/v) solution previously sterilised by filtration, were added
aseptically. Sterilised zinc sulphate to provide different zinc ion
concentrations were added to these flasks which were then inoculated
with the respective species. The flasks were incubated at 30° Con
a reciprocating shaker ( 5 cm stroke; 90 ± 2 strokes per minute),
After regular intervals 5 ml samples were aseptically withdrawn from
the flasks and total bacterial cells were counted immediately. Resid
dual concentrations of the respective substrates were also analysed.
pH was manually adjusted during the experiment. (ThiobaciZZus thiooxi
dans and ThiobaciZZus ferrooxidans at pH 2.5;ThiobaciZZus thioparus
at pH 5.5).
ThiobaciZZus ferrooxidans was found to be tolerant to zinc ions
upto 20,000 p.p.m. The rate of oxidation of ferrous iron in the
presence of 20,000 p.p.m. zinc ion was not markedly different from that
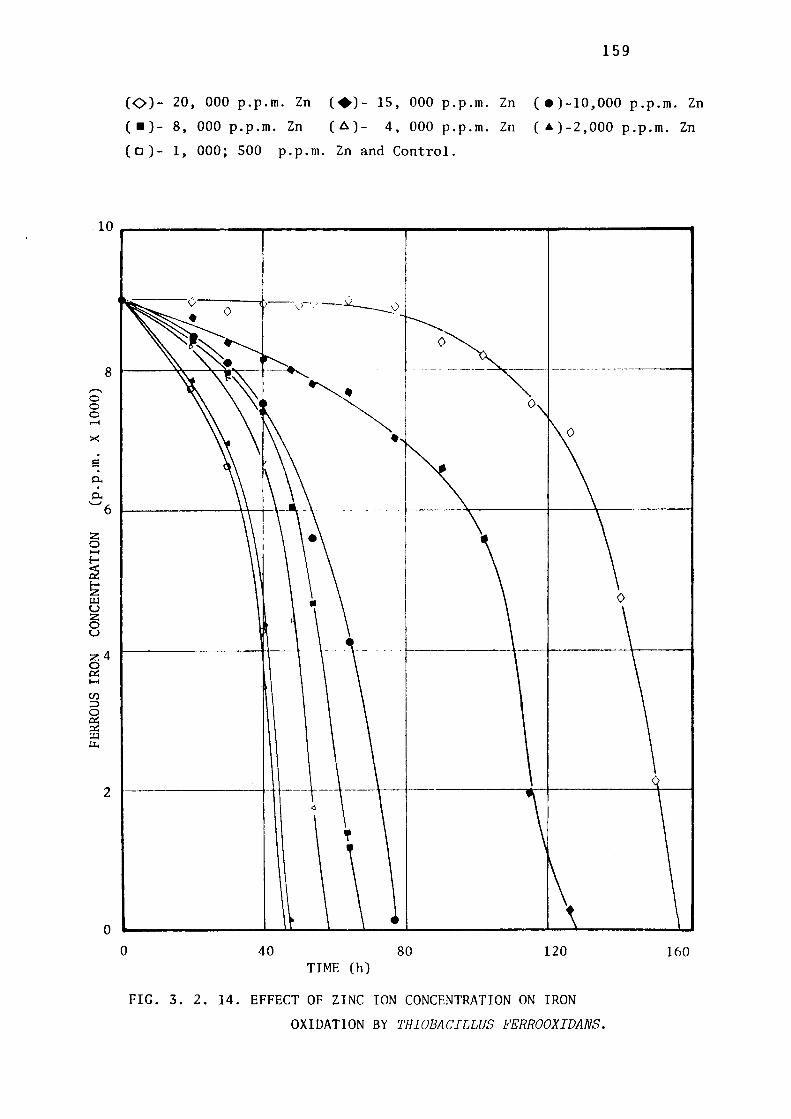
,--.. 0 0 0 ..... ><
e
0..
159
(0)- 20, 000 p.p.m. Zn (+)- 15, 000 p.p.m. Zn ( e)-10,000 p.p.m. Zn
( • ) - 8, 000 p. p. m. Zn ( 1::..) - 4, 000 p. p. m. Zn ( • ) - 2,000 p. p. m. Zn
( o )- 1, 000; 500 p.p.m. Zn and Control.
'-' 6 1-------~r--i--1
z 0 1-4
~ ~ Ul u z 0 u
:z:4 ~
2
0
0 40 80 120 TIME (h)
FIG. 3. 2. 14. EFFECT OF ZINC ION CONCENTRATION ON IRON
OXIDATION BY THIOBACILLUS FERROOXIDANS.
160
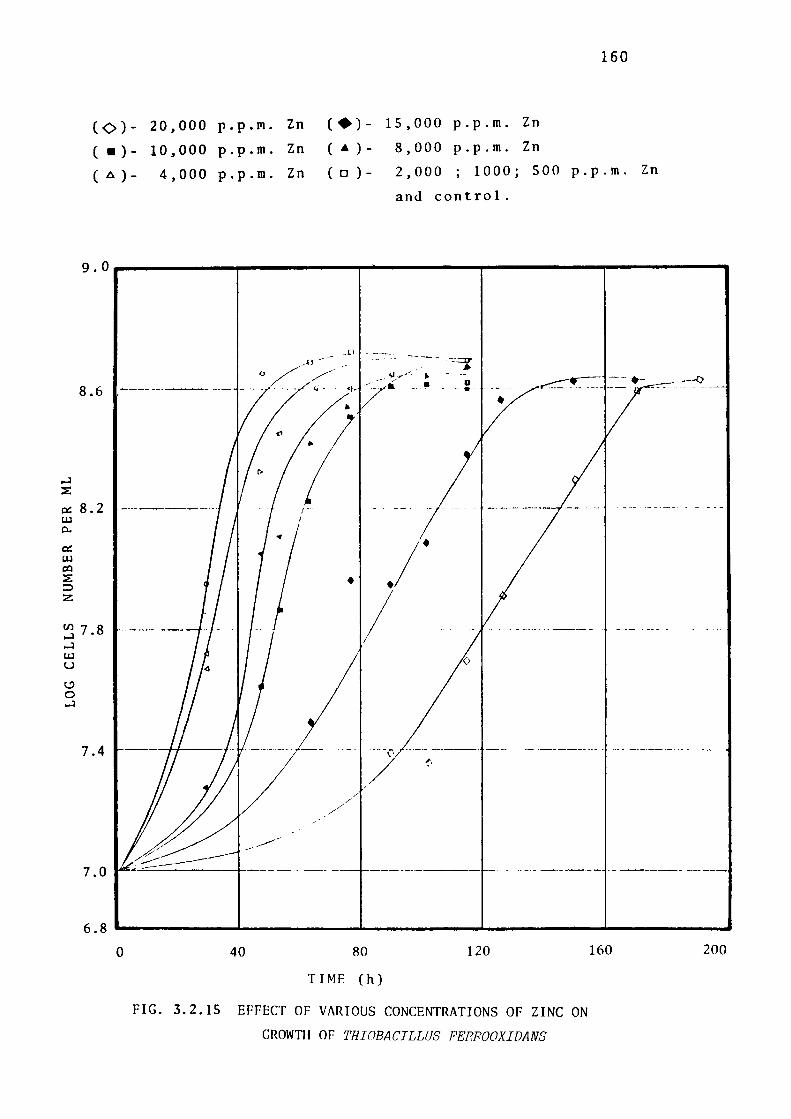
,-J
~
160
( 0 )- 20,000 p.p.Jll. Zn ( ·)- 15,000 p.p.m. Zn
( . ) - 10,000 p.p.m. Zn ( . ) - 8,000 p.p.m. Zn
( .6 ) - 4,000 p.p.m. Zn ( 0 ) - 2,000 '
1000; SOO p.p.m. Zn
and control.
9.0------,.-----.,...,-------,------,-------,
,.u - - _l) - -~::-:--_ --- - ---:.:~
/I>
8.6 D
- - -- ----··-- ~- --=---= -..- --=----- ---0_ ---
•
a: 8.2 ----------1.J,J
0..
0:: LU co ~ :::> z
5 7 .8 ,-J
LU u c.? 0 ,-J
7.4
7.0
• .1· I
0
----1--- - --------- -----
6.8 '--------J~-------L-------'------------------0 40 80
TIME (h)
120
FIG. 3.2.15 EFFECT OF VARIOUS CONCENTRATIONS OF ZINC ON
GROWTH OF THI()BACILLUS FERROOXIDANS
160 200
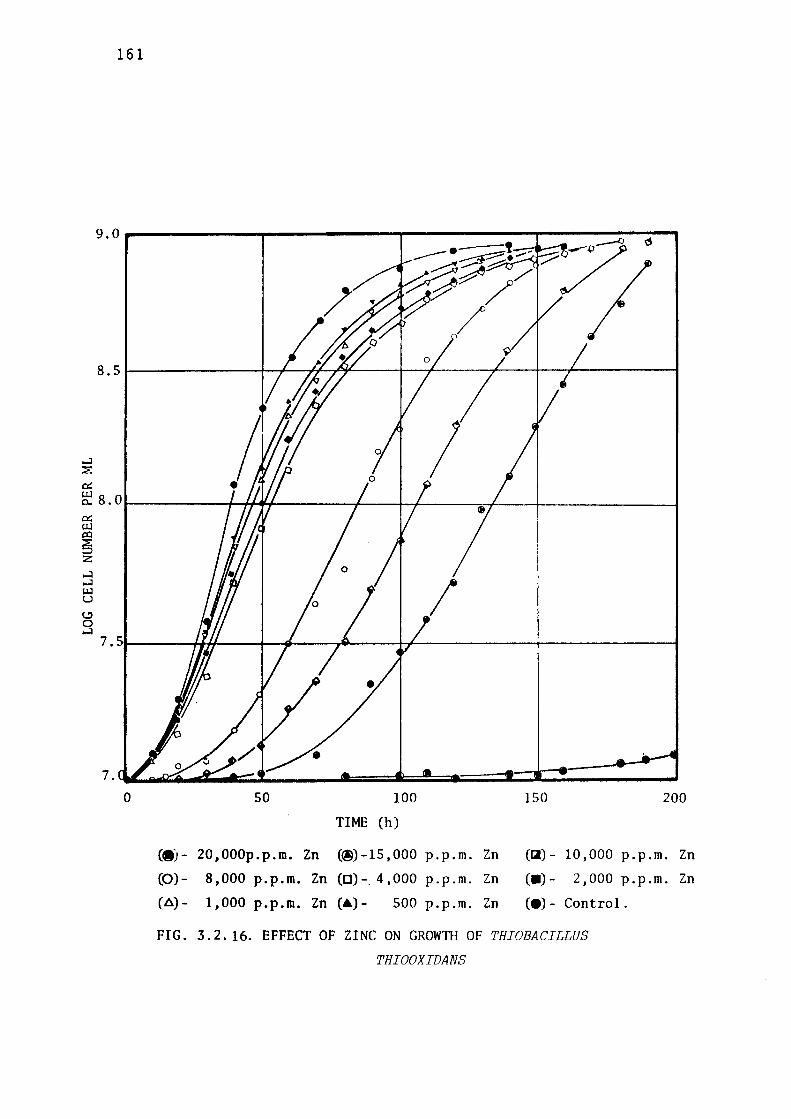
...:I ~
0::
161
~ 8. 01--------+-+,~.._----+----,r-+-----=-t'----+---------f 0:: UJ i:o
!5 z ...:I ...:I UJ u I:.:)
0 ...:I
0 50 100
TIME (h)
(·)- 20,000p.p.m. Zn (@)-15,000 p.p.m. Zn
(0)- 8,000 p.p.m. Zn (0)-_ 4,000 p.p.m. Zn
(8)- 1,000 p.p.m. Zn (a)- SOO p.p.m. Zn
150 200
(tl)- 10,000 p.p.m. Zn
C•)- 2,000 p.p.m. Zn
(e)-Control.
FIG. 3.2. 16. EFFECT OF ZINC ON GROWTH OF THIOBACILLUS
THIOOXIDANS

162
in the control. The effect of increased concentrations of zinc ion
on growth was to increase the duration of the lag phase and decrease
the growth rate, without significantly affecting the final cell mass
(Figs. 3.2.14 & 3.2.15). Similar effects were recorded in the case of
Thiobacillus thiooxidans (Fig. 3.2.16) which was comparatively less
tolerant to increased concentration of zinc ion; the maximum tolerable
concentration of zinc ion for this strain was 15,000 p.p.m. 20,000
p.p.m. severely retarded growthbutwas not lethal.
Zinc sulphate solution, when added to Vishniac & Santer medium
(pH 5.5), suddenly decreased the pH to 3.5. Neutralisation of this
solution produced copious amollllts of zinc hydroxide precipitate, which
associated with the dissociation products of sodium thiosulphate, pro
duced by acidification. Under these circumstances it was very diffic
ult to determine the toxic levels of zinc for Thio/Jacillus thiopa:r>us.
Generally, acidophilic members of the genus Thiobacilli are
tolerant to high metallic concentrations. Bacteria tolerant to very
high concentrations of zinc (120 g/i) have been reported (Gormely
et al., 1975) but molybdenum and silver are reported to be very toxic
to these bacteria (Imai et al., 1975; Hoffman & Hendrix, 1976).

3.3 LEACHING OF ZINC SULPHIDE BY THIOBACILLUS FERROXIDANS,
THIOBACILLUS THIOOXIDANS AND THIOBACILLUS THIOPARUS
163
Preliminary (sections 3.3.1, 3.3.2, 3.3.3 and 3.3.4) and
definitive (section 3.3.5) experiments were conducted with the aim
of comparing the abilities of three Thiobacillus species to leach
various zinc sulphides, and to explore the effects on leaching rates
of crystal structure, lattice substitution and other parameters, Two
sets of experiments were designed; in one set (section 3.3.2, 3.3.3,
& 3.3.4) leaching rates were studied in the presence of equal pulp
density of minerals whereas the other set (section 3.3.5) employed
equal surface area of minerals for comparison of leaching rates.
Unless otherwise stated, the following standard experimental
procedure was followed. For ThiobaciZZus thiooxidans and ThiobaciZZus
ferrooxidans the growth medium was the 9K basal salts medium of
Silverman & Lundgren(l959) adjusted to pH 2.5 with sulphuric acid
(2 N), lacking a soluble energy source; for ThiobaciZZus thioparus
the growth medium was identical to that of Vishniac and Santer (1957)
except that it lacked thiosulphate, and was adjusted to pH 5.5. 190
ml of the appropriate medium, contained in Erlenmeyer flasks of one
litre capacity, were sterilised by autoclaving at 15 psi for 15 minutes.
2 g of zinc sulphide, previously ground to pass through a 400 mesh
(37 micron) screen and sterilised by propylene oxide, were transferred
aseptically to each flask. 10 ml of inoculum (prepared as described
in individual test) were added to each flask .to give an initial
concentration of approximately 107 cells per ml. An uninoculated
flask, with otherwise identical contents to that of the test flask, was
prepared with each test and was incubated under identical conditions.

164
The flasks were shaken at 30 ± 0.5°C on a reciprocating shaker: ( 5 cm
stroke; 90 + 2 strokes per minute).
The pH was controlled manually at pH 2.5 or 5.5,as appropriate,
with sterile 2N sulphuric acid or 2N sodium hydroxide throughout the
course of each test. At appropriate intervals the flasks were removed
from the shaker and allowed to stand for 30 minutes to permit settling
of the zinc sulphide; 5 ml of culture were then withdrawn aseptically
from each flask and transferred to centrifuge tubes; the 5 ml were
replaced with 5 ml of fresh, sterile medium before returning the flasks
to the shaker. The clear supernatants, obtained by centrifugation at
10,000 g for 15 minutes, were analysed for zinc, ferrous and ferric
ions and their pH measured. Cell counts were determined on tennination
of each experiment.
3.3.1 LEACHING CAPABILITIES OF THIOBACILLUS THIOPARUS (BJR-451)
This preliminary experiment was designed to test the potential
of BJR-451 for leaching sulphide minerals of zinc, copper and nickel.
The three sulphides were tested concurrently by suspending each in a
shake flask containing a medium which was identical to that of Vishniac
and Santer (1957) except that it lacked thiosulphate. The initial pH
of the medium was adjusted to 5.5, the optimum pH for growth of
BJR-451. pH was not subsequently controlled. Controls containing the
medium and mineral but no microbial cells, were treated similarly
and incubated concurrently. The flasks were shaken on a reciprocating
shaker at 90 ± 2 strokes per minute with 5 cm stroke, at 30°C.
Every 3-4 days over a period of three weeks the flasks were removed
from the shaker, the mineral allowed to settle and 5 ml of culture
withdrawn aseptically and an equal volume of fresh sterile medium
added. The samples were centrifuged at 10,000 . g for 15 minutes and
the clear supernatant analysed for copper, zinc or nickel.
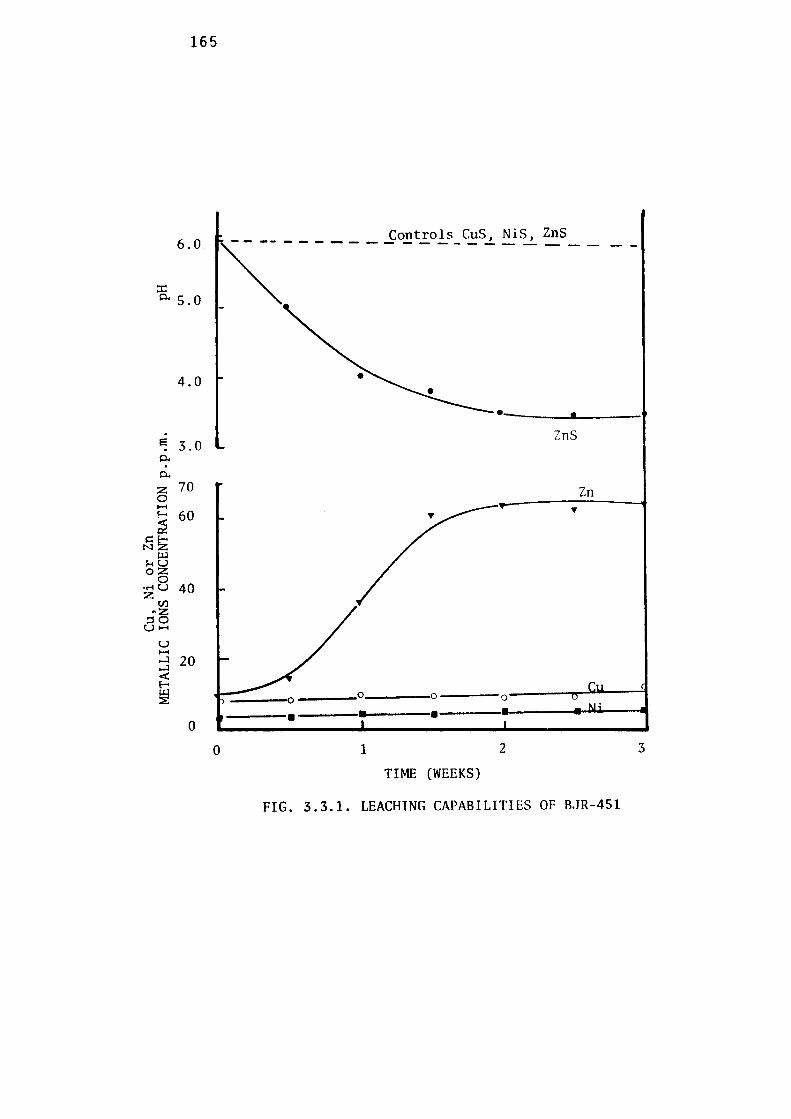
165
6.0
::r: i:i.. 5.0
4.0
• . ZnS
El 3.0 . p.. . p..
z 70 Zn 0 H
~ 60 •
~~ (J.J
r-iu oz
0 •..-tU 40 z
Cl) .,z
::so UH
u H ...l 20 ...l <I'. E-
~ 0
0 • • • 0 1 2 3
TIME (WEEKS)
FIG. 3.3.1. LEACHING CAPABILITIES OF BJR-451

166
The results of the metal releasing ability of BJR-451, illustra
ted in Fig. 3.3.1, revealed that in the presence of the bacterium,
zinc ions were released into solution. The copper and nickel ion
concentrations in the control and inoculated flasks were almost
identical and no significant changes in pH were discernable. It
was concluded that BJR-451 was incapable of degrading copper and nickel
sulphides under these conditions, but was capable of degrading zinc
sulphide. The pH of the medium containing zinc sulphide decreased
from 6.0 to 3.5 as the mineral was solubilised. During the first 14
days the zinc ion concentration increased progressively from 15 p.p.m.
to 65 p.p.m. Thereafter, there was no change in zinc ion concentration
and the pH remained constant at its minimum value of 3.5. Since the
bacterium had been shown to be inhibited at pH values lower than 4.5
(see section 3.2) it seems reasonable to assume that cessation of
activity was due to production of acid. Most probably the complex
reaction involving the oxidation of the sulphidic moiety of zinc
sulphide is represented stoichiometrically by:
s (Torma, 1972)
Because of its ability to leach zinc sulphide, ThiobaciZZus
thioparus was included in the comparative study of the leaching of
several zinc sulphide samples.
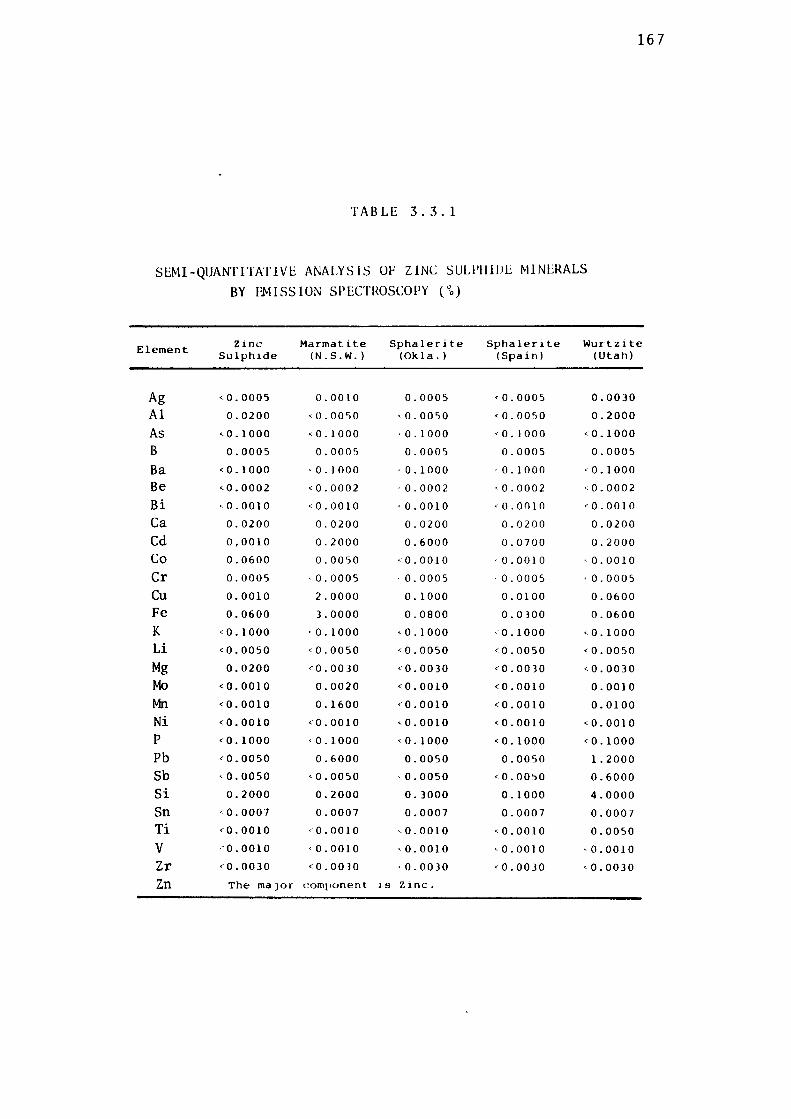
TABLE 3.3.1
SEMI-QUANT ITATlVE ANALYSIS OF ZINC SULl'IIIIJE Ml NERALS
BY EMISSION SPECTROSCOPY ( 0o)
167
Element Zinc Marmatite Sphalerite Sphalerite Wurtz1te Sulphide (N. S .W.) (Okla.) (Spain) (Utah)
Ag <0.0005 0.0010 0.0005 <0.0005 0.0030
Al 0.0200 <0.0050 <0.0050 <0.0050 0.2000
As ,0.1000 <0.1000 •0.1000 <0.1000 <0.1000
B 0.0005 0.0005 0.0005 0.0005 0.0005
Ba <0.1000 ,0.1000 ·0.1000 ·0.1000 <0.1000
Be <0.0002 <0.0002 ·0.0002 ,0.0002 <0.0002
Bi •0.0010 <0.0010 •0.0010 •0.0010 <0.0010
Ca 0.0200 0.0200 0.0200 0.0200 0.0200
Cd 0,0010 0.2000 0.6000 0.0700 0.2000
Co 0.0600 0.0050 ,0.0010 ·0.0010 ·0.0010
Cr 0.0005 ·0.0005 ·0.0005 ·0.0005 ·0.0005
Cu 0.0010 2.0000 0.1000 0.0100 0.0600
Fe 0.0600 3.0000 0.0800 0.0300 0.0600
K <0.1000 ·0.1000 <0.1000 <0.1000 <0.1000
Li <0.0050 <0.0050 <0.0050 <0.0050 <0.0050
Mg 0.0200 <0.0030 <0.0030 <0.0030 ,0.0030
Mo <0.0010 0.0020 <0.0010 <0.0010 0.0010
Mn <0.0010 0.1600 <0.0010 <0.0010 0.0100
Ni <0.0010 <0.0010 <0.0010 <0.0010 <0.0010 p <0.1000 ,0.1000 <0.1000 <0.1000 <0.1000
Pb <0.0050 0.6000 0.0050 0.0050 1.2000
Sb ,0.0050 <0.0050 ,0.0050 <0.00~0 0.6000
Si 0.2000 0.2000 o. 3000 0.1000 4.0000
Sn <0.0007 0.0007 0.0007 0.0007 0.0007
Ti <0.0010 •0.0010 ,0.0010 <0.0010 0.0050
V •'.0.0010 •0.0010 ,0.0010 ,0.0010 •0.0010
Zr <0.0030 <0.0030 ·0.0030 <0.0030 <0.0030
Zn The maJor component ls Zinc.

168
3.3.2 LEACHING OF ZINC SULPHIDES BY THIOBACILLUS FERROOXIDANS
ThiobaaiZZus ferrooxidans is well-known for its ability to
degrade zinc sulphide (Zimmerley et al., 1958; Torma et al., 1972;
Gormely et al., 1975). The aim of this experiment was to examine the
the leachability of different zinc sulphides by a strain of ThiobaciZ
Zus ferrooxidans isolated from a mine drainage at Mount Lyell. The
elemental compositions,, of the five zinc sulphides tested are given
in Tables 3.3.1 and 3.3.2. Semiquantitative analysis of these
minerals revealed that arsenic, cadmium, lead, iron and silicon were
present as impurities in them.
Quantitative analyses of these five minerals for
principal metals like zinc, iron, lead, copper and nickel as shown
in Table 3.3.2, which reveals that synthetic zinc sulphide, and
museum grade specimens of sphalerites from Spain and Oklahoma, are
almost identical in their mineral compositions, except that the latter
contained a higher iron content. Mannatite from Broken Hill, N.S.W.,
contained 12.2% iron and had a high percentage (1.35%) of lead
associated with it. The lowest zinc content was that of wurtzite from
Utah (46.9%). This sample also had a high percentage of lead (5.6%)
associated with it.
The experiment was performed in one litre flasks containing 190
ml of the 9K basal salt medium of Silverman & Lundgren (1959). The
experimental procedure as described earlier was followed (see section
3. 3 ) .· ThiobaciUus ferrooxidans grown on ferrous sulphate as the
sole source of energy was used as inoculum. Leaching results are
shown in Fig. 3.3.2. The curves in Fig. 3.3.2 were obtained from the
net value of microbial leaching, i.e., the differences between the zinc
concentrations in the inoculated flask and in the control flasks.
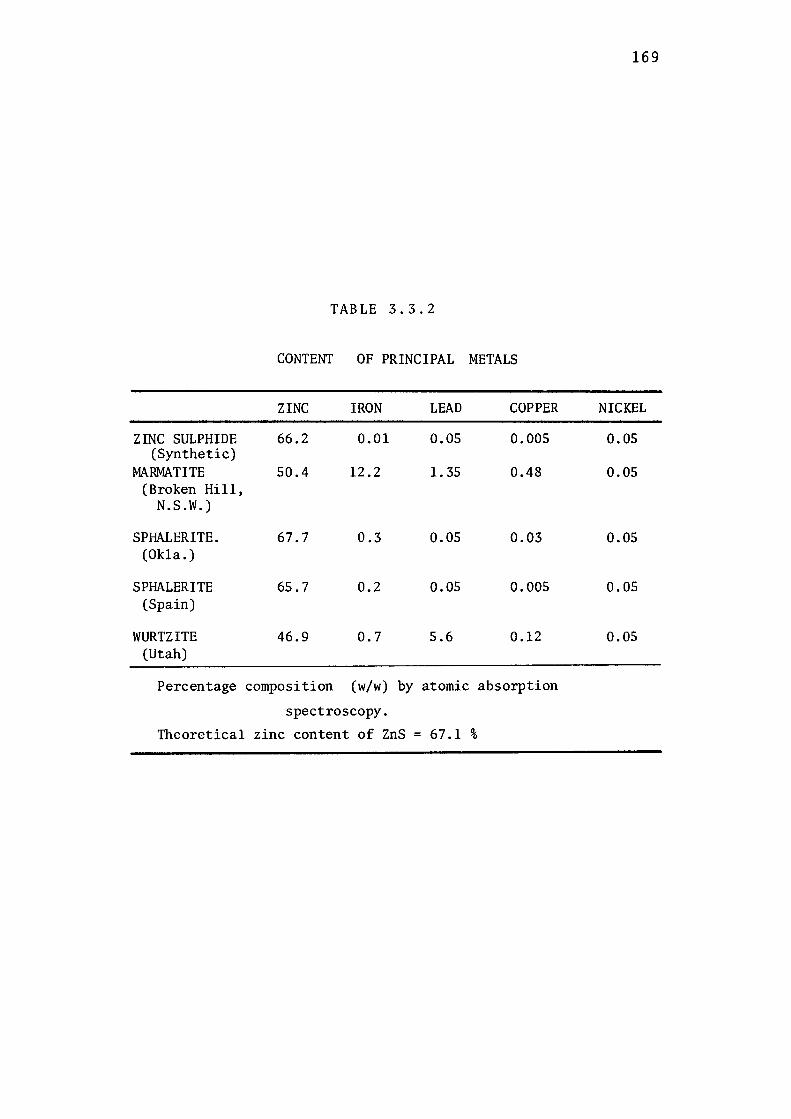
169
TABLE 3.3.2
CONTENT OF PRINCIPAL METALS
ZINC IRON LEAD COPPER NICKEL
ZINC SULPHIDE 66.2 0.01 0.05 0.005 0.05 (Synthetic)
MARMATITE 50.4 12.2 1.35 0.48 0.05 (Broken Hill,
N.S.W.)
SPHALERITE. 67.7 0.3 0.05 0.03 0.05 (Okla.)
SPHALERITE 65. 7 0.2 0.05 0.005 0.05 (Spain)
WURTZITE 46.9 0.7 5.6 0.12 0.05 (Utah)
Percentage composition (w/w) by atomic absorption
spectroscopy.
Theoretical zinc content of ZnS = 67.1 %

170
Leaahing of Marmatite.
Leaching of this mixed sulphide by Thiobacillus ferrooxidans was
revealed by a progressive increase in the concentration of solubilised
zinc ions in the inoculated flasks. A maximum concentration of 1,920
p.p.m. was recovered with 145 p.p.m. in control. The lag period was
approximately 2 - 2.5 days; zinc release then occurred up to the 12th
day of experiment, after which its rate of release decelerated. By
the 7th day of the experiment, precipitation had occurred and the
degree of precipitation was such that brown particles of marmatite
were hardly visible; they were embedded in the hard reddish brown
precipitate.
During these experiments endeavours were made to monitor the
soluble ferrous and ferric ions in the flasks. It was folllld that,
whilst the soluble ferrous content was negligible at all stages, the
soluble ferric contents progressively increased up to the 7th day
(see Fig. 3.3.2). By the 8th day it had begllll a progressive decrease
which was accompanied by the formation of the heavy precipitate
referred to above. The maximum concentration of ferric ions at 7th day
of experimentation was 80 p.p.m. (average of three experiments).
Insolubilisation of virtually all the ferric ion in the form of
precipitate, commonly reported as jarosite by other authors (Dllllcan
& Walden, 1972), was coincident with the cessation of zinc ion
solubilisation.
Leaahing of Synthetia Zina Sulphide.
Synthetic zinc sulphide was also leached by Thiobacillus ferroo
xidans grown on ferrous sulphate as energy source. The shape of the
curve was very similar to the marmatite leaching curve. The lag was
also 2-2.5 days and after the 12th day of the experiment the zinc solu
bilisation decelerated. During this leaching period a maximum

2000
1500
. ~ 1000 p..
p..
u z H N
SOO
0
171
----------------,,.--------,--------,100
I al I
' I
b
I
I I
I
~ I
I I
/,Q. / \
~ \ / \
D
'
\
I I • ~r;;,_,...
\ \ \
\
\ \
'o \
\
\ R: <1 --I> 0 <1- -l>-<1 - ....- .. - .. I>- \ .,._ -_..,..-~ \
\ 0
/ j ~ .,,.,,< ,b /• ------ ... \ ___ _....._ .- •--...
75
so
25
/ ~ ··----= i- • ·- -· -·--?9~ -•-•- 0-..- --o--O a::.;;--;.;;.--;.;;.,;:,..__ __ _;,•• ____ • _ _,1., _______ _._ _______ ..i,.. _______ ,.. 0
0 5 10
TIME (Days)
15
FIG. 3.3.2. LEACHING OF ZINC SULPHIDE MINERALS BY
THIOBACILLUS FERROOXIDANS
(. )- Marrnatite ( 0) - Synthetic zinc sulphide
c• )- Sphalerite (Spain) ( 6) - Sphalerite (Oklahoma)
(. )- Wurtzite ( D ) - Ferric iron (Total iron) (From rnarrnatite)
20
. E
0.. . 0..
z 0 0::: H

172
concentration of 2,000 p.p.m. zinc ion was recovered with 210 p.p.m.
in the control.
Leaching of Museum Grade Zinc SuZphid,e Minero.ls.
Comparatively pure museum grade specimens of zinc sulphide mine
rals in the form of sphalerite were also leached by ThiobaciZZus ferro
oxidans. Their leaching rate was slower than that of marmatite or the
relatively pure synthetic zinc sulphide. Wurtzite, a hexagonal form of
zinc sulphide was, however, very recalcitrant to leaching by the organism.
Maximum concentrations of 467 p.p.m. and 405 p.p.m. of zinc were obta
ined from sphalerite samples of Oklahoma and Spanish origins, respect
ively. The concentrations of solubilised zinc ions in the controls
were 30 and 32 p.p.m. The maximum concentration of solubilised zinc
from the wurtzite sample was 125 p.p.m. with 15 p.p.m. in the control.
Discussion.
As previously stated, zinc sulphide is among the earliest mineral
sulphides examined for microbial leachability. Rudolf & Helbronner
(1922) were the first to apply bacterial leaching to the extraction of
zinc from zinc blende. Subsequently, numerous studies on the microbial
leaching of zinc sulphide by ThiobaciZZus ferrooxida:ns have been made,
including those of Ivanov et al.(1961), Torma et al.(1970), Gormely et
al.(1975). It is now well documented that zinc sulphide concentrates
are amenable to microbial degradation. Most of the information has
been obtained from studies with marmatite as the zinc mineral, which
contains appreciable amotmts of iron in it. The so called 'indirect
mechanism' (which limits the role of the iron oxidising bacteria to
ferrous oxidation, with the ferric ions produced acting as the leachi
ng agents) had been considered as the main leaching mechanism during
earlier investigations. Generally all natural sulphides contain iron
as one of the impurities, and according to the indirect mechanism

173
of microbial leaching this iron can be constantly recycled.
The stationary phases reached after leaching of marmatite and
synthetic zinc sulphide in the current investigation preswnably resul
ted from either some nutrient or other cultural requirement becoming
limiting. The main energy sources for the bacteria in the case of
marmatite were the sulphide and ferrous components. At stationary phase
ample amounts of these components were still present in the medium and
available to the bacteria. It is conceivable that oxygen and carbon
dioxide, which are important nutrients for bacteria, may have become
limiting. However, in any consideration of limiting factors, it must
be born in mind that one of the most important factors in any solid
bacterial interaction is the availability of surfaces to the bacteria;
such access is improved by contact between the solid substrate and the
bacterial cells. Whether such contact changes the bacterial response
towards the solid substrate is still unknown but a number of reports
exist in which the importance of contact between solid surface and
bacteria has been suggested by the relationship between the rate of
oxidation of solid substrate (viz., sulphur) and shaking speed.
It has been observed, for example, that an increased shaking rate
sometimes decreases the oxidation rate of elemental sulphur by Thio
bacillus thiooxidans ( Starkey et al., 1956). Reaction rate can also
be limited by the precipitation of materials on surfaces, which inact
ivate the contact points between the bacteria and solid substrate.
The situation would simulate substrate limitation.
The formation of a complex ferric precipitate (jarosite) during
microbial leaching has been observed previously by many investigators
(Duncan & Walden, 1972; Torma & Legault, 1973; Rossi, 1974). The
current results indicate that, after the 7th day, the concentration
of soluble ferric ions in the medium starts decreasing and at the 14th

174
day almost all the ferric ions are precipitated out. This coincides
with the cessation of zinc release from marmatite. Thus it is most
likely that when marmatite is the zinc source the formation of ferric
precipitates could inhibit the rate of leaching by blocking the contact
between the bacteria and the mineral particles. Similar findings have
been reported by a number of workers, who have shown that by regrinding
the mineral particles more of the mineral can be extracted (Torma,
1977). Dtmcan & Walden (1972) demonstrated that during bacterial
leaching of marmatitic zinc sulphide, iron concentration was increased
for the first 100 hours of the process, after which iron was precipi-
tated out from the medium with a concomitant depletion of ammonium ion
from the medium.
The ability of Thiobacillus ferTooxidans to degrade pure, synth
tic minerals has been demonstrated by many workers including Torma
(1971). In this study the aim was to compare, under identical condit
ions ( e.g. bacterial concentrations, pulp density, temperature etc.)
the leaching pattern of relatively pure, synthetic zinc sulphide with
that of other natural zinc sulphide minerals. The leaching pattern
was surprisingly similar to that of marmatite considering that the
synthetic zinc sulphide had a low iron contentbut (presumably in
compensation) was very finely divided and hence presented a greater
surface area (w/w) to the bacterium than did the marmatite.
One of the most striking findings of this test was the relatively
slow leaching rates of the two natural museum grade sphalerite minerals
and the almost zero response of the organism to the wurtzite. An
early report by Trussell et al. (1964) also recorded the low leaching
rates of sphalerites by Thiobacillus ferrooxidans. They found that
almost 100% of the marmatite but only 4-9% of the sphalerite was
leached. The current experiment confirms that different sulphides of
the same metal do not respond identically to bacterial leaching.

175
That structural differences in the sulphides affect their leachability
is indicated by the fact that wurtzite, which belongs to a crystal
system which is different from that of sphalerite, is virtually resis-
tant to these bacteria. No previous reports on
ble for comparison with the current findings.
wurtzite are availa-
Iron contents of these minerals seemed to be overlapped by the
structural influence in determining the leaching behaviour. Thus,
although wurtzite contains a significant amotmt of iron in its lattice
(0.7%), it was not leached readily. The higher content of lead (5.6%)
present may be another inhibitory factor affecting the leachability
of wurtzite by these bacteria.

176
3.3.3 LEACHING OF ZINC SULPHIDES BY THIOBACILLVS THIOOXIDANS
As previously mentioned, ThiobaciZZus thiooxidans is generally
not considered a suitable leaching bacterium; consequently little work
has been reported previously on the leaching behaviour of this Thio
baciZZus species. In the current experiment, the inoculum was
prepared as follows:
ThiobaciZZus thiooxidans (BJR-KOl) was grown with elemental
sulphur as the sole energy source, at 30°C, pH 2.5, in a shake flask
culture; the residual sulphur was removed from the suspension by fil
tration through Whatman No. 1 filter paper and the filtrate centrifu
ged at 10,000. g for 15 minutes; the cell pellet, suspended in 9K
basal salt solution of Silvennan & Llllldgren (1959) was used to inocu
late the growth flasks at a level of approximately 107cells per ml.
The amollllts of zinc released from the various zinc sulphides are
illustrated in Fig. 3.3.3.
Leaching of Marmatite.
ThiobaciZZus thiooxidans released a total of 1,250 p.p.m. of
zinc from the marmatite during 18 days of incubation. Ferrous ion
concentration increased as the dissolution of marmatite proceeded.
The maximum concentration of ferrous ion was 395 p.p.m. at the maximum
total soluble iron concentration of 410 p.p.m.; the difference, which
was presumably a measure of ferric ion, is negligible and therefore
indicates the relatively pure state of the ThiobaciZZus thiooxidans in
terms of its freedom from iron oxidising bacteria. In contrast to the
tests with ThiobaciZZus ferrooxidans, no precipitati0n occurred in the
flasks, and the mineral particles were at all times clearly visible.
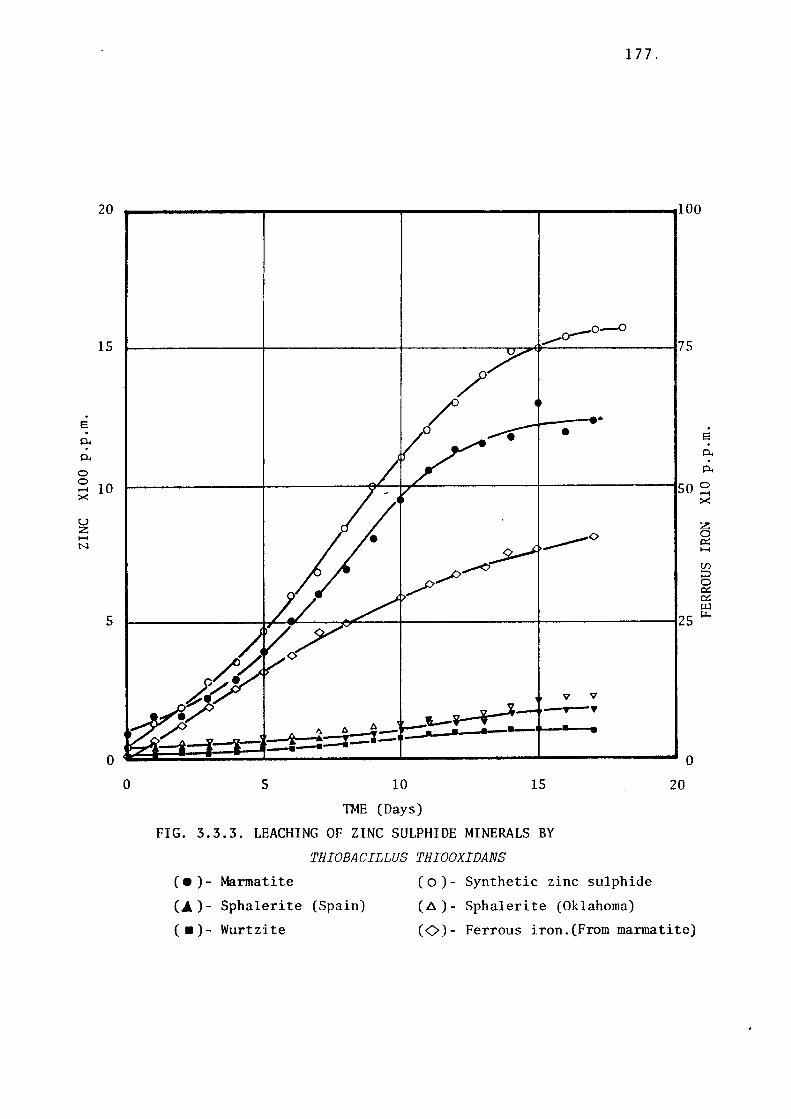
20
15
. e . p..
p..
0 0 10 .... ><
u z H N
5
17 7.
--------r--------,--------,.-------1100
o--°--0 1---------+--------+------....,....~~,..----------175
0 5 10
TME (Days)
. •
----0
V V
~ J-•---Y+-• 1--V--- .. - - . ------
15
FIG. 3.3.3. LEACHING OF ZINC SULPHIDE MINERALS BY
( • ) - Marrnati te
THIOBACILLUS THIOOXIDANS
( o) - Synthetic zinc sulphide
( 6.) - Sphaleri te (Oklahoma)
e . p.. . p..
50 ~ ><
z 0 IX H
U) :::i 0 IX IX tJ.l
25 i:.r..
20
(~) - Sphaleri te (Spain)
( • )- Wurtzite (0)- Ferrous iron.(From marmatite)

178
Leaching of Synthetic Zinc Sulphide.
The leaching curve for the relatively pure zinc sulphide was
steeper than that for marmatite and the maximum concentration of
leached zinc, 1,550 p.p.m. was greater than that obtained from marmat
ite (1,250 p.p.m.). The leaching rate had decelerated by the 15th
day and the leaching had ceased by the 18th day.
Leaching of Museum Grade Zinc Sulphide Minerals.
The museum grade specimens of sphalerite were not as effectively
leached as were the marmatite and synthetic zinc sulphide. The maximum
concentrations of zinc ion released from the Spanish and Oklahoma
sphalerites were 180 and 230 p.p.m. respectively. Wurtzite was
particularly recalcitrant to the action of Thiobacillus thiooxidans &
only 100 p.p.m. of zinc were solubilised.
The viable bacterial population at the end of experiments with
the two sphalerites and the wurtzite was 2 . 10 8 organisms per ml,
which was about one tenth of the population obtained after growth on
the marmatite and zinc sulphide samples.
Discussion.
The results indicate that strain BJR-KOl is capable of degrading
sulphide minerals. It released 1,250 p.p.m. zinc from marmatite into
solution in 18 days, compared with the 1,920 p.p.m. released by
Thiobacillus ferrooxidans under similar conditions. Therefore, the
marmatite leaching activity of Thiobacillus thiooxidans is about
65% of that of Thiobacillus ferrooxidans for leaching marmatite.
Since sulphur oxidisers can oxidise only the sulphur moiety in
the crystal lattice, it can be assumed that all metal released from
sulphides by Thiobacillus thiooxidans is due to direct bacterial
activity. In the case of iron oxidisers, such as Thiobacillus

179
ferrooxidans, which oxidise ferrous ions preferentially over sulphide
ions, the leaching of mannatite cannot be considered as being entirely
due to direct bacterial oxidation, since the ferric product of the
ferrous oxidation can serve as a good leaching agent and thus contrib
ute a degree of chemical leaching to the process. If we were to
consider that the sulphur oxidising capacities of ThiobaaiZZus
thiooxidans and ThiobaaiZZus ferrooxidans were equal, then the diff
rence between the maximum concentrations of zinc ions released by
ThiobaaiZZus thiooxidans (1,250 p.p.m.) and ThiobaaiZZus ferrooxidans
(1,920 p.p.m.) would be due to ferrous/ferric oxidative activity,
i.e., due to chemical leaching. However, such a simplistic concept
cannot be proposed when it is known that ThiobaaiZZus ferrooxidans
oxidises ferrous ions preferentially over sulphide ions. Account
must also be taken of the fact that the former reaction involves a
one electron transfer and the second (for complete oxidation to
suphate) an eight electron transfer._ Moreover, the relative availab
ilities and . rates of oxidation of ferrous and sulphide ions would
be extremely significant factors in the relative contributions of the
direct and indirect mechanism to leaching by ThiobaaiZZus ferrooxidans.
Vanselow (1976) has studied the relative contributions of the two
processes in detail by comparing rates of leaching of covellite
(CuS) by one strain of ThiobaaiZZus ferrooxidans in the presence and
absence of ferrous ions. A similar observation was made in the
current study (see section 3.3.6). Nevertheless it is worthy of n-0te
that ThiobaaiZZus feYTooxidans leached 35% more zinc from marmatite
than did ThiobaaiZZus thiooxidans and that the increase could perhaps
relate to its iron oxidation ability.
The iron (mainly in the ferrous form) released from mannatite
remained in solution and reached a maximum concentration of 415 p.p.m
(7.4 rnM) compared with a maximum soluble zinc concentration of

180
1,250 p.p.m. (19.12 mM), i.e., soluble iron: soluble zinc= 1:2.6.
The iron content of the original marmatite was 12.2 % and zinc content
50.4% (Table 3.3.2) which would give marmatite an empirical formula
of zn3 • 5Fe s5 • Therefore the molar ratio of zinc and iron is 4.1 : 1.
If metal release of iron and zinc had been proportional, they would
have maintained the same stoichiometric ratio in the solution, i.e.,
1:4.1 rather than the 1:2.6 observed.
The difference between the theoretical and observed stoichiometry
represents 37% deviation from proportional leaching of iron and zinc
at the concentration in which they occurred in the marmatite sample.
Conclusions regarding this deviation are difficult to draw. However,
a situation whereby the iron atoms are solubilised and extracted to
a greater proportional extent than are the zinc ions would leave
'holes' inside the crystal lattice and this could facilitate more
leaching. If this were so then marmatite should leach more readily
than synthetic zinc sulphide. The fact that it did not could however
relate to differences in the specific surface areas of the two
substrates. This variable was removed in the definitive experiments
in section 3.3.5.
The activity of Thioba.ciZZus thiooxidans towards marmatite had
virtually ceased by the 15th day. Metal toxicity and/or oxygen
limitations are among the possible causes of cessation of activity;
further work would be required before conclusions regarding termination
of the leaching process could be made.
A maximum concentration of 1,550 p.p.m. zinc was released from
synthetic zinc sulphide which was 19.4% greater than marmatite. A
high microbial population ( 5 • 109 bacteria per ml) after the end of
the experiment with synthetic zinc sulphide indicated the acceptiblity
of synthetic zinc sulphide as a suitable substrate for ThiobaciZZus
thiooxida:ns. A microbial population of 3.5 . 109 cells per ml was

181
recovered in the culture growing with mannatite.
In common with the ThiobaciZZus ferrooxidans tested, ThiobaciZZus
thiooxidans was unable to leach sphalerites or wurtzite to any
significant extent. The reason for their low leachability by these
bacteria may be due to a very stable crystal structure, or to a
stability induced by the natural fonnation of minerals; the data
collected are however an insufficient basis for more than very
tentative speculation.

182
3.3.4 LEACHING OF ZINC SULPHIDE BY THIOBACILLUS THIOPARUS
The study with ThiobaciZZus thioparus (BJR-451) was carried out
at pH 5.5 in shake flasks ( one litre Erlenmeyer flasks) containing
190 ml of the basal salt mediun of Vishniac & Santer (1957). After
sterilising the medium at 15 psi for 15 minutes, 2 g of zinc sulphide
minerals previously sterilised with propylene oxide gas for 12-18
hours, were aseptically added. The bacterial inoculum was prepared
as follows:
ThiobaciZZus thioparus (BJR-451) was cultivated in a chemostat
with thiosulphate as the growth limiting substrate, at a dilution rate
of 0.1- 1 h; the cells were collected after centrifugation at 10,000 g
for 15 minutes, washed with and resuspended in sterilised Vishniac
& Santer medium (1957). 10 ml of this inoculum was introduced into
each shake flask, so that the final concentration was approximately
107 cells per ml. The flasks were shaken in a reciprocal shaker (at
5 cm stroke and 90 ± 2 strokes per minute) and samples withdrawn and
analysed as described in section 3.3.2 and 3.3.3. The results are
illustrated in Fig. 3.3.4.
Leaching of Marmatite.
ThiolxiciZZus thioparus leached marmatite at a considerably
slower rate than did the acidophilic bacteria ThiolxiciZZus ferrooxi
dans and ThiolxiciZZus thiooxidans. The maximum soluble zinc concen
tration recorded was 90 p.p.m. after 19 days' incubation. The
bacterial cell collllt at the end of of experiment was 3 . 108 bacteria
per ml of which 90% viable.
The ferrous ion concentration was less than 0.1 p.p.m. and the
total soluble iron was 3 p.p.m.
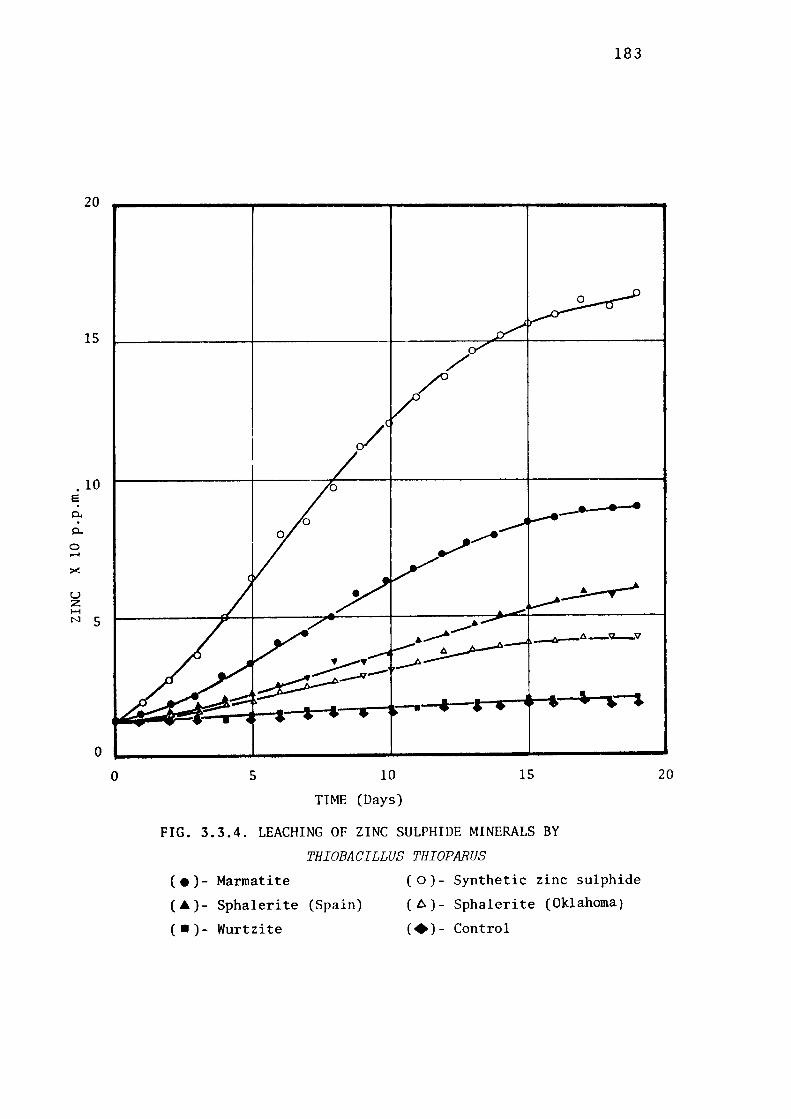
20
15
. 10 e 0..
0..
0 .-I
><
u z 1-1 N 5
0
0 5 10
TIME (Days)
15
183
_. ..
FIG. 3.3.4. LEACHING OF ZINC SULPHIDE MINERALS BY
THIOBACILLUS THIOPARUS
( • )- Marmatite
(A)- Sphaleri te (Spain)
( • )- Wurtzite
( o )- Synthetic zinc sulphide
( 6 )- Sphalerite (Oklahoma)
(+)- Control
20

184
Leaching of Synthetic Zinc sulphide.
The leachability of synthetic zinc sulphide was greater than
that of marmatite, the maximum recorded concentration of zinc ion in
the medium being 167 p .p .m. The lag phase was shorter and the leaching
curve was steeper than in the case of marmatite. The viable population
at the end of the experiment was between 3 and 4 . 108cells per ml,
a comparable figure to that recorded in the marmatite culture.
Leaching of Natural Zinc SUlphid,e Minerals.
The action of Thiobacillus thioparus on natural sphalerite was
not so significant. The leaching of two samples of sphalerite was
relatively slow, and the maximum recorded concentration of zinc ion
was 60 and 43 p.p.m. for Spanish and Oklahoma variety respectively.
No significant degree of leaching over that recorded in the control was
observed when the zinc sulphide was in the form of wurtzite, the
maximum recorded concentration of soluble zinc being 25 p.p.m.
Discussion.
The leaching capabilities of non-acidophilic members of the
genus Thio'bacillus have not been examined previously to any signifi
cant extent. The results obtained from current experiments suggest
that the non-acidophilic Thiobacillus thioparus is relatively slow
in its leaching activity towards the various zinc sulphide minerals
studied. However, the pattern of leaching which has emerged from the
current study is very similar to the one observed on the study of the
leaching of these minerals by Thiobacillus thiooxidans (see

185
section 3.3.3.). Synthetic zinc sulphide was leached at the fastest
rate followed by marmatite, and the natural sphalerites were partially
recalcitrant, whilst wurtzite, was almost completely recalcitrant.
Whenever marmatite is leached, its iron component would be expected
also to be leached out, as it was when marmatite was leached by the
acidophilic ThiobaciZZi. Negligible amounts of soluble iron were
recorded however when marmatite was leached by ThiobaciZZus thio
parus. The maximum being less than 3 p.p.m.It is believed that the
ferrous iron released at pH 5.5 was converted to insoluble ferric
complexes, such reactions being favoured at that pH. This phenomenon
may also have contributed to the relatively low leachability of marma
tite (compared with the leachability of synthetic zinc sulphide)
by passivating the surfaces of the zinc sulphide particles with
ferric hydroxide precipitates. However, a valid comparison can only
be made when samples of equal specific surface area are presented to the
organisms ( see section 3.3.5). The increase in viable counts of
ThiobaciZZus thioparus on synthetic zinc sulphide and marmatite from
107 to 3-4.108 bacteria per ml indicates a reasonable growth activity
of this member of the ThiobaciZZi on sulphide minerals. Whilst the
relatively low concentrations of solubilised zinc in the cultures
indicate that this species is not a good leaching agent, it must be
kept in mind that at pH 5.5 the solubility of metals is drastically
affected; equlibriurn shifts towards hydroxide formation can occur and
render the solubilized metal once more insoluble; in other words,
a conversion of metal sulphide to metal hydroxide could occur. By
analogy, when galena (PbS) is oxidised microbiologically, the concen
tration of lead in solution does not change significantly, since the
final product of bacterial oxidation, PbS04,is also practically
insoluble (Tomizuka, 1976). It is not unreasonable to assume

186
therefore that, under relatively high-pH conditions, a correct measure
of leaching of any metal sulphide cannot be obtained by determining
the soluble metal concentration alone. In order to have a reasonable
assessmentof mineral leaching under such conditions, other products
of bacterial oxidation, such as sulphate, should also be measured
and the presence of any new insoluble mineral in the medium should
be checked by X-ray crystallographic methods.

187
TABLE 3.3.3
SPECIFIC SURFACE AREA OF * ZINC SULPHIDE MINERALS
MINERAL SURFACE AREA (m2/g)
ZINC SULPHIDE 30.00 (Synthetic)
MARMATITE 9.14 (Broken Hill, N.S.W.)
SPHALERITE (Okla.) 7.60
SPHALERITE (Spain) 5.70
WURTZITE (Utah) 6.40
* Determined by B.E.T. Method.

188
3.3.5 INFLUENCE OF STRUCTURE OF ZINC SULPHIDE ON MICROBIAL
LEACHING BY THIOBACILLI
From the studies described earlier in this section it was evident
that our three strains of Thiobacillus species each leached the
different zinc sulphide minerals at different rates and to different
extents. It was however realized that (a) not only were two different
crystallographic forms, i.e., sphalerite (cubic) and wurtzite (hexa
gonal), represented in the zinc sulphide samples but (b) these samples
probably had different specific surface areas. In the current
experiment the second variable was removed in order to investigate
the effect of the first more definitely.
In the preliminary experiments equal pulp densities were used,
but since in solid-substrate microbiology it is the surface area which
effectively determines the substrate concentration, a comparison was
made of the surface area of these zinc sulphide samples. The speci
fic surface areas of the ground minerals used in the previous experi
ment were determined by the B.E.T. method (Brunauer, Eramet & Teller,
1938), and are recorded in Table 3.3.3. The synthetic zinc sulphide
was relatively fine, with a specific area of [ 30 m2/g], 3 to 5 times
that of the other minerals. Mineral weights were adjusted to provide
equal surface area of 100 m2/litre in the present experiment.
Thiobaaillus ferrooxidans and Thiobacillus thiooxidans cultures
were first acclimatised to the individual mineral substrates in 9K
basal salt solution (Silverman & Lundgren, 1959). In the case of
wurtzite, where growth rate was poor, these thiobacilli were grown
with the mineral plus elemental sulphur as the substrate.
Because of its relatively poor leaching capabilities, the non
acidophilic, Thiobacillus thioparus, bacterium was acclimatised only
to synthetic zinc sulphide. Acclimatisation in all cases was conducted

189
in shake flasks at 30°C for 4 - 5 weeks, and the cells harvested by
centrifugation of the filtrate after filtration through Whatman No.I
filter paper. ThiobaciZZus ferrooxidans and ThiobaciZZus thiooxidans
were then suspended in the 9K basal salt medium (pH 2.5) of Silverman
and Lundgren (1959), and ThiobaciZZus thioparus suspended in the basal
salt medium of Vishniac and Santer (1957). The cell suspensions
served as inocula for the leaching experiments.
For the leaching experiments, an appropriate weight of mineral,
providing a surface area of 20 m2 and previously sterilised by
propylene oxide, and 190 ml of sterile basal salt medium were added
aseptically to sterile Erlenmeyer flasks of one litre capacity. The
basal salt media used were those described above for suspension of
inocula. 10 ml of inoculum was added to each flask to give an initial
level of 107 bacteria per ml.
The flasks were incubated at 30°C on a reciprocating shaker
(5 cm stroke; 90 + 2 strokes per minute). The pH was manually adjusted
to its initial level ( 2.5 for ThiobaciZZus thiooxidans, ThiobaciZZus
fePPooxidans; 5.5 for ThiobaciZZus thioparus) at time intervals of
12 hours. At appropriate times (after every 24 hours) the flasks were
removed from the shaker and allowed to stand for 20-30 minutes at 30°C.
5 ml of mineral-free culture were withdrawn aseptically from each
flask, and replaced with appropriate sterile medium. The samples
withdrawn were centrifuged at 10,000 g for 10 minutes and the
supernatant was analysed for solubilised zinc, total iron and ferrous
iron. Results are illustrated in Figs. 3.3.5 - 3.3.9 and summarised
in Table 3.3.4.
Comparative Leaching of MaPmatite.
Fig. 3.3.5 represents the leaching of marmatite by ThiobaciZZus
ferrooxidans and ThiobaciZZus thiooxidans species. Under identical
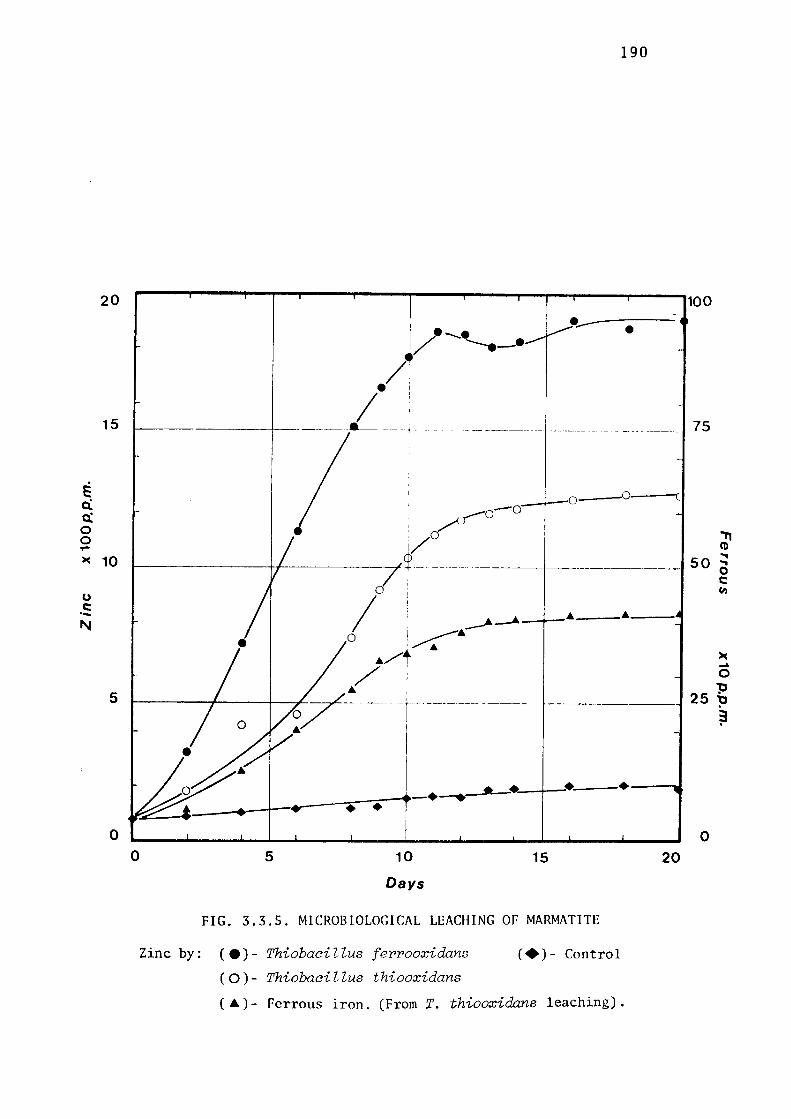
20
15
e a: a: 0 0 ,-)( 10
(J C: ·-~
5
0
I • • 1/ -----· • • /i / :
190
100
·--- . I----------+--------•- --- __ , - -· - - - ------- ------ ·- ·- _ -· -- _ ------ 7 5
0
•
•
---o----o) __ _,.. _,--i----o-o .,,
(b i/if 7f-- ------ ---- -----------1 so a
0 ,
/ : ~,.__.__. __ . 0 '
C: c,,
' . •/t ~ /' ~
{_-/-- I t--------~ 25 .;..• 0 / -- - t---------- ---- -- ~
• I
•0 • I • ..'--+-_ .. :...----._ ....... ---~~t .__.. ~-+-..- • •
I 5 10
Days
15
FIG. 3.3.5. MICROBIOLOGICAL LEACHING OF MARMATITE
Zinc by: ( •) - Thiobaci l lus f er>r>ooxidans
( O) - Thiobaci l lus thiooxidans
(+)- Control
( .&)- Ferrous iron. (From T. thiooxidans leaching).
20
0
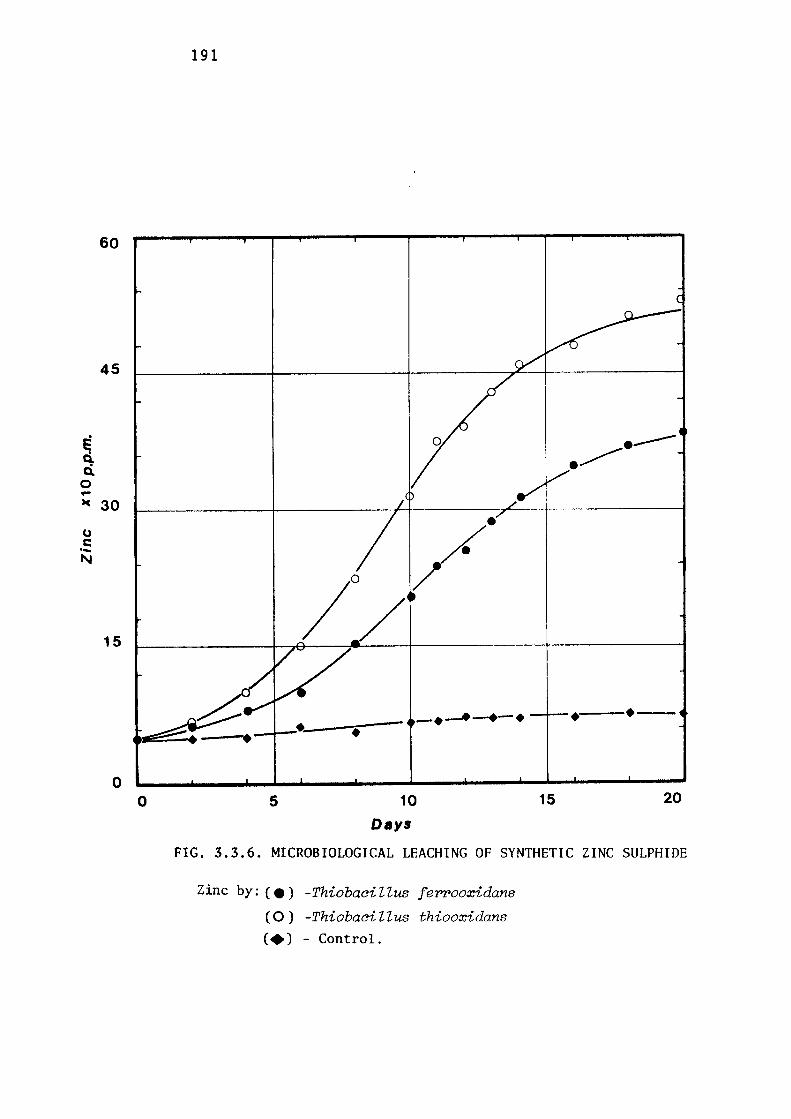
e q. a.
0 .....
60
45
)( 30
CJ C: ·-N
15
0 0
191
•
5 10
Days
15
.--.~
··--·-20
FIG. 3.3.6. MICROBIOLOGICAL LEACHING OF SYNTHETIC ZINC SULPHIDE
Zinc by: ( •) -Thiobaci Uus ferrooxidans
( 0 ) -Thiobaci llus thiooxidans
( +) - Control.

192
conditions of pH, temperature and initial specific surface area,
marmatite leaching behaviour was very similar to that observed before.
Marmatite was more rapidly leached by ThiobaciZZus ferrooxidans than
by ThiobaciZZus thiooxidans . The ThiobaciZZus ferrooxidans'lag
period was comparatively shorter and the maximum concentration of zinc
ions found in solution was 1,875 p.p.m. after 11 days' incubation;
the soluble .Fe2 + & Fe 3+concentrations were too low to be measured but
ferric salt precipitation was observed. The viable ThiobaciZZus
ferrooxidans at the termination of the experiment was found within
the range of 3 - 4. 109 cells per ml and its maximum leaching rate
of zinc was 230 mg/(i.day).
ThiobaciZZus thiooxidans leached marmatite at a maximum rate of
130 mg/(i.day) and a maximum concentration of 1,270 p.p.m. zinc
was obtained at the end of the experiment; the iron released was
substantially ferrous and its maximum concentration at the end of the
experiment was 425 p.p.m.
The leachability of marmatite at the higher pH value of 5.5 by
ThiobaciZZus thioparus was comparatively slow and the maximum concen
tration of leached zinc was 80 p.p.m. The rate of leaching was
about 4.9 mg/(i.day). Negligible amounts of soluble iron were found
in the medium. The viable counts of ThiobaciZZus thioparus at the
end of the experiment were in the range of 2 - 3. 108 cells per ml.
Comparative Leaching of Synthetic Zinc Sulphide.
Leaching of synthetic zinc sulphide by ThiobaciZZus ferrooxidans
and ThiobaciZZus thiooxidans is shown in Fig. 3.3.6. In contrast to
marmatite, ThiobaciZZus thiooxidans leached zinc sulphide at a
faster rate than did ThiobaciZZus ferrooxidans. The maximum concen
trations of solubilised zinc were 530 p.p.m. and 385 p.p.m. in the
ThiobaciZZus thiooxidans & ThiobaciZZus ferrooxidans cultures

20
15
I 0 10
5
0 0
193
• ---· •
--+-•------=.~--..
5 10
Days
15
FIG. 3.3.7. MICROBIOLOGICAL LEACHING OF SPl~LERITE (SPAIN).
Zinc by: ( •) - Thiobaci Uus f errooxidans
( 0 ) - Thiobaci Z Zus thiooxidans
(+)- Control.
20

194
respectively and the solubilisation rates were 40 and 24 mg/(i.day)
respectively. The soluble Fe2 +& Fe 3+ iron concentrations were too
low to be measured. The viable bacterial populations of Thiolxicillus
ferrooxidans and Thiobacillus thiooxidans at the end of the experiment
were 2. 109 and 3-4 . 10 9 cells per ml respectively. Total zinc ions
solubilised by Thiolxicillus thioparus were 59.5 p.p.m. and only
traces of iron ions could be detected. The maximum rate of zinc
solubilisation with this species was only 6.9 mg/(i.day) and at the
end of the experiment only 2-3 . 10 8 cells per ml were recorded.
Comparative Leaching of Natural Zinc Sulphide Minerals.
Leaching curves for sphalerite are shown in Figs. 3.3.7 and
3.3.8. Both sphalerites responded similarly towards the acidophiles,
Thiolxicillus ferrooxidans and Thiobacillus thiooxidans in that the
maximum concentration of zinc ions liberated by each organism was
less than that released from marmatite or synthetic zinc sulphide.
The higher purity of the Spanish .sphalerite, compared with the
Oklahoma sphalerite, was presumed to be the reason why it was less
degradable by either acidophilic Thiobacillus species than was the
latter variety.
The maximum solubilisation rate of zinc from the Oklahoma and
Spanish varieties by Thiobacillus ferrooxidans, was 22 and 15 mg/(i.day),
respectively (Table 3.3.4). On the other hand, Thiolxicillus thio
oxidans leached both sphalerites at an almost equal rate of 12 - 13
mg/(i.day).
Leaching with Thiobacillus thioparus produced the relatively
meagre soluble zinc ion concentration of 27-31 p.p.m. after a maximum
rate of solubilisation of 3.1 - 3.9 mg/(i.day). With such a small .
amotmt of zinc ions in solution, it was difficult to ascertain which,
if either, sphalerite variety was more amenable to degradation by
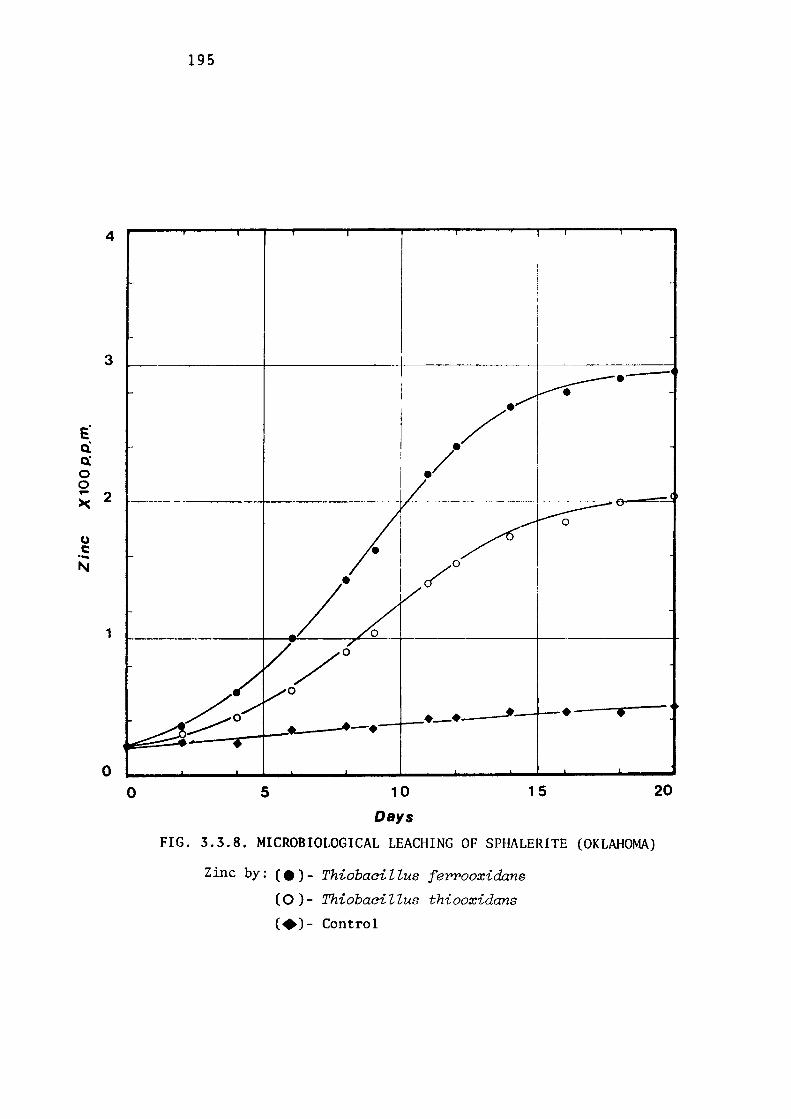
4
3
e· 0'. a: 0 0 ... 2 )(
CJ C: ·-"'
1
0 0
195
5 10
Days
/. /.
•
/ /0
JI·--· •
15
FIG. 3.3.8. MICROBIOLOGICAL LEACHING OF SPHALERITE (OKLAHOMA)
Zinc by: ( •) - Thiobaci Z Zus feY'Y'Ooxidczns
( 0 ) - Thiobaci Z Zus thiooxidans
C•)- Control
20

196
Thiobacillus thioparu.s. In the presence of either sphalerite, the
viable counts of Thiobacillus thioparus were 1.5 . 108 cells per ml
at the end of the experiment.
Comparative Leaching of Natural Wurtzite.
The result of the comparative study of wurtzite leaching by
three Thiobacilli are shown in Fig. 3.3.9 and Table 3.3.4. The maxi
mum concentration of zinc in the Thiobacillus thiooxidans and Thio
!Jacillus ferrooxidans flasks were 85 and 102 p.p.m. respectively and
the maximum leaching rates were 6.5 and 9.0 mg/(t.day) respectively.
The wurtzite sample containing 0.7% (w/w) iron as an impurity
(Table 3.3.2); a total of 25 p.p.m. of this was released as ferrous
iron into the solution by Thiobacillus .thiooxida:ns but only a total
of 2.5 p.p.m. iron could be found in the soluble form in the iron
oxidising Thiobacillus ferrooxida:ns culture. The viable count of
bacteria at the end of the experiment was within the range of 6. 107
cells per ml.
Negligible amounts of zinc ions were solublised from wurtzite
by the non-acidophilic species, Thio!Jacillus thioparus, at the end
of the experiment only slightly more than 3. 10 7 cells per ml were
found.
Discussion.
The results of this experiment suggest that the leaching
capability of Thiobacilli varies from species to species and that
both the structure and the composition of substrates may be factors
governing the leachability of the minerals.
The results demonstrate that the non-acidophile, Thio!Jacillus
thioparus (BJR-451), is the least active of the three species studied
in leaching zinc sulphides. None of the minerals studied, except
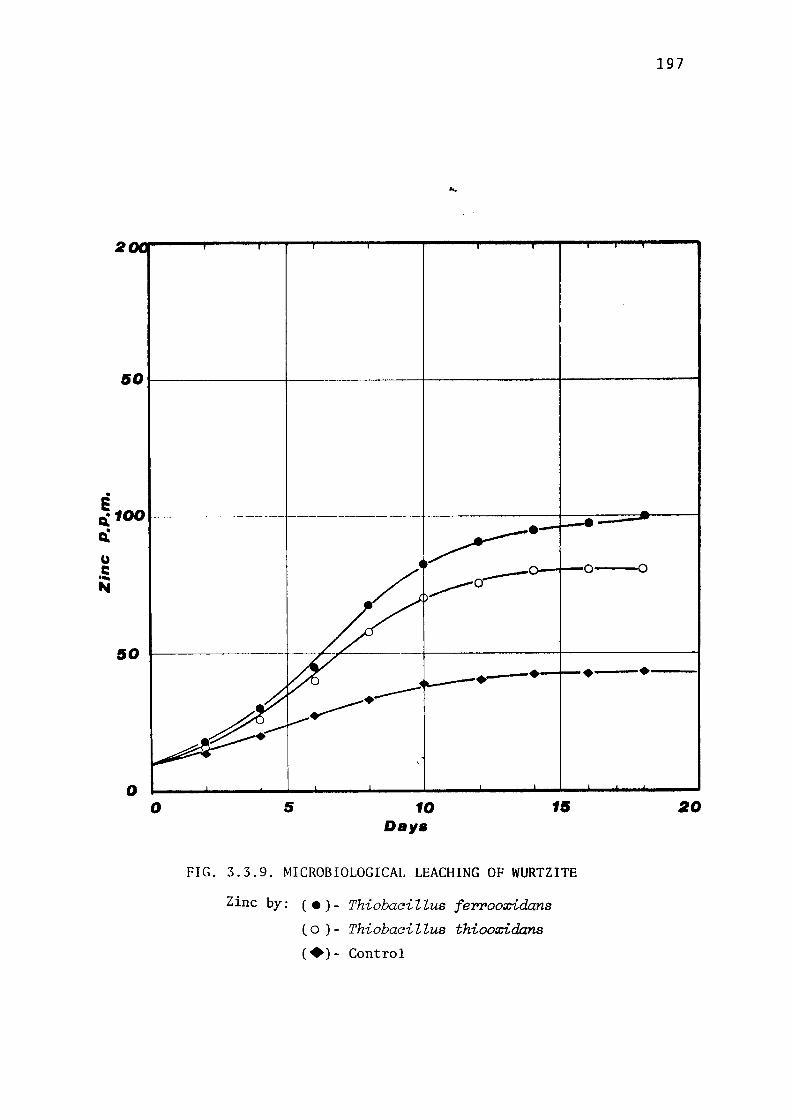
197
...
50l--------+-------+-------+-------t
e ~100--------+--~ u C: .... ...
~-0--+--0--0
50 1----------+
o o
·----•--ir--•--•---· .,----
5
.---10
Dav• 15
FIG. 3.3.9. MICROBIOLOGICAL LEACHING OF WURTZITE
Zinc by: ( • ) - Thiobaci Z Zus ferrooxidans
( o ) - Thiobaci Z Zus thiooxidans
(+)- Control
20
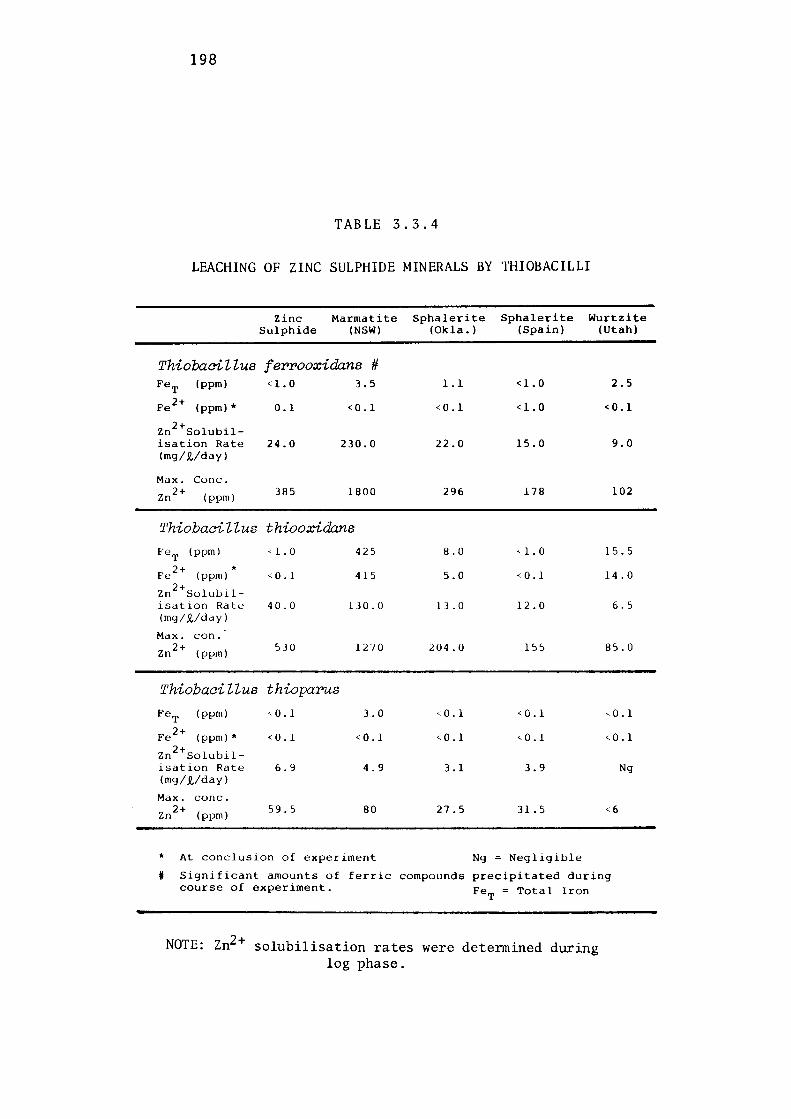
198
TABLE 3.3.4
LEACHING OF ZINC SULPHIDE MINERALS BY THIOBACILLI
Zinc Marmatite Sphalerite Sphalerite Wurtzite Sulphide (NSW) (Okla.) (Spain) (Utah)
Thiobaai l lus ferrooxidans # FeT (ppm) <1.0 3.5 1.1 <1.0 2.5
Fe 2+ (ppm)* 0.1 <0.1 <0.1 <1.0 <0.1
zn 2+solubil-isation Rate 24.0 230.0 22.0 15.0 9.0 (mg/Jl,/day)
Max. Cone.
Zn 2+ (ppm) 385 1800 296 178 102
Thiobaai l lus thiooxidans
f'eT (ppm) , 1. 0 425 8.0 '1. 0 15.5
Fe 2 + • (ppm) < 0. 1 415 5.0 < 0. 1 14.0
zn 2+solubil-isation Rate 40.0 130.0 13. 0 12.0 6.5 (mg/ Jl,/day)
Max. con.
Zn 2+ (ppm) 530 1270 204.0 155 85.0
Thiobaci Uus thioparus
FeT (ppm) < 0. 1 3.0 <0.1 <0.1 <0.1
Fe 2+ (ppm)* <0.1 <0.1 <0.1 <0.1 <0.1
zn 2+Solubil-isation Rate 6.9 4.9 3 .1 3.9 Ng (mg/R,/day)
Max. cone.
Zn 2+ (ppm) 59.5 80 27.5 31. 5 <6
* At conclusion of experiment Ng= Negligible
# Significant amounts of ferric compounds precipitated during course of experiment. FeT = Total Iron
NOTE: Zn2+ solubilisation rates were determined during log phase.

199
synthetic zinc sulphide, was degraded appreciably. However, the
strain was able to maintain its viability and some growth did occur
when these minerals were supplied as the only source of energy. The
degree of leachability of zinc sulphide minerals by ThiobaciZZus
thioparus varied between O and 2.69% (Table 3.3.5). Leachability of
wurtzite was negligible whereas 2.69% of synthetic zinc sulphide was
leached. Of all five minerals tested, synthetic zinc sulphide was
maximally leached with, ThiobaciZZus thioparus as well as with
ThiobaciZZus thiooxidans. Since synthetic zinc sulphide was almost
iron-free, the only possible mechanism of degradation can be the
direct oxidation of the sulphidic moiety of the minerals. The leaching
behaviour towards synthetic zinc sulphide of the two sulphur
oxidisers (ThiobaciZZus thioparus and ThiobaciZZus thiooxidans) was
very similar, except that a higher percentage of leachability was
obtained by the former (24 %); this can be attributed to the low pH
of the medium.
One of the obvious reasons for the low soluble metal content of
ThiobaciZZus thioparus cultures may be the relatively high pH (5.5)
of the leaching medium. At this pH the solubility of zinc ions might
have been drastically reduced because of formation of a new (insoluble)
complex. Consideration was given to clarifying the results by
determining the sulphate released into the medium as a product of
oxidation of the sulphide moiety. Attempts, however, to obtain a
meaningful determination of sulphate ion in this medium were seriously
affected by the presence of the large quantities of phosphate present
in the Vishniac & Santer medium (1957). It has been reported that
the, leaching capabilities of the newly isolated ThiobaciZZus
rubeZZus and ThiobaciZZus deZicatus species were also very small
compared with those of the acidophilic members of the
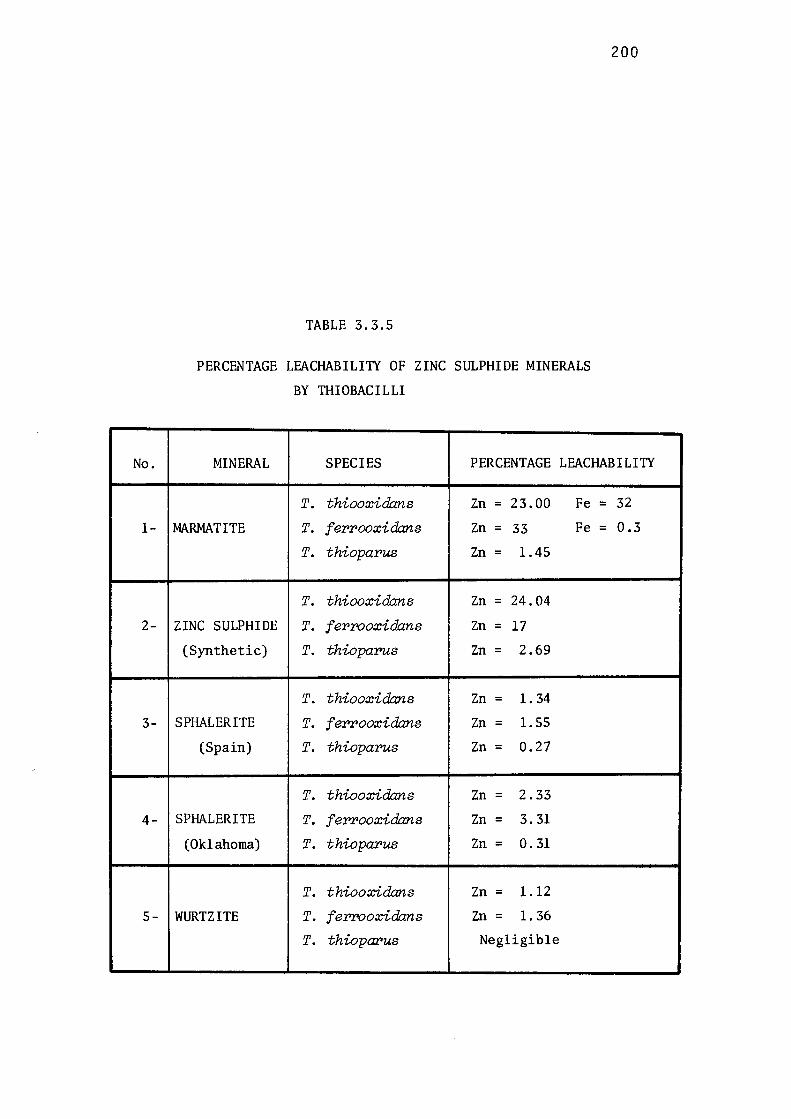
No.
1-
2-
3-
4-
5-
TABLE 3.3.5
PERCENTAGE LEACHABILI1Y OF ZINC SULPHIDE MINERALS
BY THIOBACILLI
200
MINERAL SPECIES PERCENTAGE LEACHABILI1Y
T. thiooxidans Zn = 23.00 Fe= 32
MARMATITE T. f errooxidans Zn = 33 Fe= 0.3
T. thioparus Zn = 1.45
T. thiooxidans Zn = 24.04
ZINC SULPHIDE T. ferrooxidans Zn = 17
(Synthetic) T. thioparus Zn = 2.69
T. thiooxidans Zn = 1. 34
SPHALERITE T. ferrooxidans Zn = 1.55
(Spain) T. thioparus Zn = 0.27
T. thiooxidans Zn = 2.33
SPHALERITE T. f errooxidans Zn = 3.31
(Oklahoma) T. thioparus Zn = o. 31
T. thiooxidans Zn = 1.12
WURTZITE T. ferrooxidans Zn = 1. 36
T. thioparus Negligible

201
genus Thiobacillus (Mizoguchi et al., 1976). Bacterial leaching of
various ores with an alkaline medium using Thiobacillus thioparus and
Thiobacillus thiooxida:ns has been studied by Mayling (1969), who has
suggested the use of a Thiobacillus thioparu.s strain at pH 8.4 for
leaching copper and zinc ores. But further study in this direction has
not yet been pursued, presumably because of the extremely slow
reactions and low solubilisation rates obtained.
In comparison with Thiobacillus thioparu.s both Thio"bacillus
thiooxidans and Thiobacillus ferrooxidans were relatively good
leaching agents. In the case of the natural minerals, Thiobacillus
ferrooxidans was more efficient in leaching zinc than was the sulphur
oxidiser, Thiobacillus thiooxidans. However, the relatively pure,
synthetic zinc sulphide was more effectively leached by
Thiobacillus thiooxidans than by Thiobacillus ferrooxidans. 41%more
zinc was released by the sulphur oxidising bacterium than by Thio
"bacillus ferrooxidans from the synthetic zinc sulphide, whilst the
zinc concentration released from marmatite by Thiobacillus ferroo
xidans was 43% greater than that released by Thio"bacillus thiooxid
ans. The most plausible reason for the increased activity of
Thio"bacillus ferrooxidans is that metal release by Thiobacillus
thiooxidans is mostly due to the direct attack of bacteria, whereas
both direct and indirect mechanisms are likely to operate in the
case of Thio"bacillus ferrooxidans. (It is notable in this context that
the marmatite samples contain 12.2% iron while the synthetic zinc
sulphide contained only 0.06%.) This was further studied in the next
experiment, where the role of iron on leaching was investigated.
Leachability of zinc sulphide minerals by different Thio"bacilli
is tabulated in Table 3.3.S. It is evident that marmatite was leached
to a maximum degree of 33% by Thiobacillus ferrooxidans, which leached
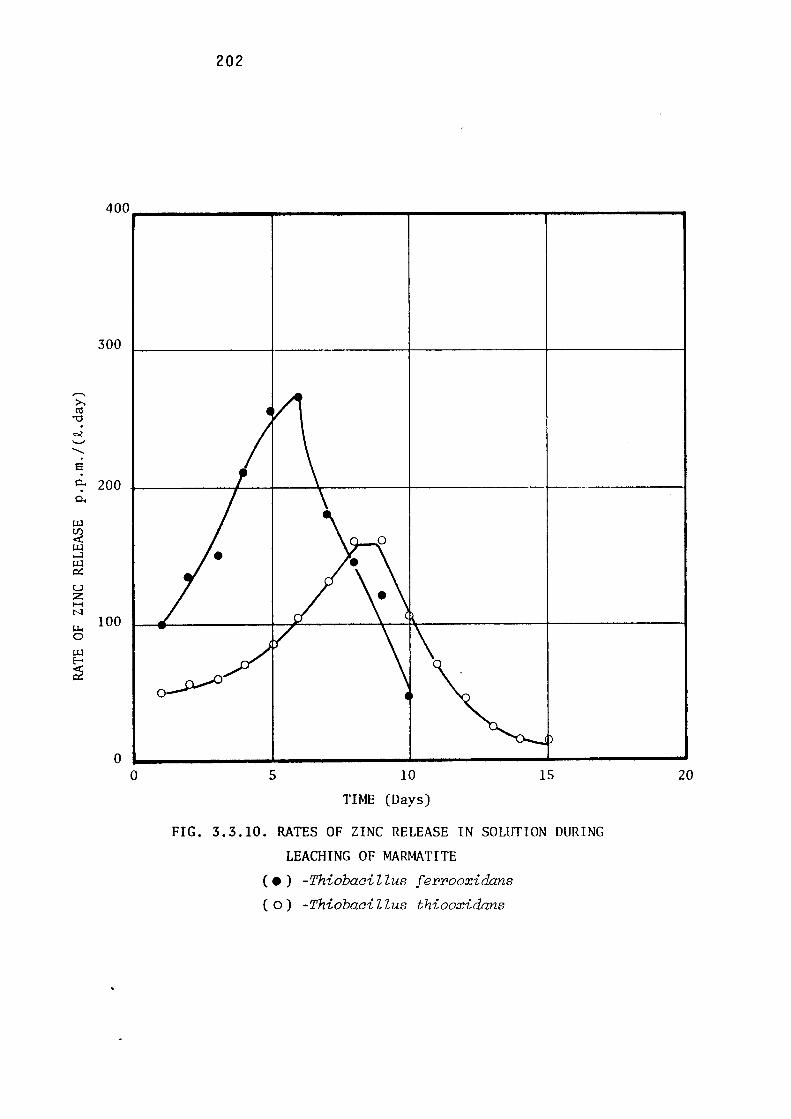
,......,_ >. c,:I
"C . ~ .._,
..........
s . 0-, . 0-,
UJ CJ) <!'. UJ ...:I UJ 0::
u z ..... N
u.. 0
UJ E--t
~
202
400-------~-------,---------,--------,
300
200
100
0 0 5 10
TIME (Days)
15
FIG. 3.3.10. RATES OF ZINC RELEASE IN SOLUTION DURING
LEACHING OF MARMATITE
( • ) -Thiobaci l lus f errooxidans
( o) -Thiobaci l lus thiooxidans
20

203
17% of synthetic zinc sulphide and 1.55 and 3.31% of museum grade
sphalerites. Wurtzite leachability was a minimum (1.36%). It is
interesting to note that even with a small increase in the iron content
of sphalerite from Oklahoma (Table 3.3.2), its leachability was increa
sed 114% when compared with the other sphalerite sample, from Spain.
Thiobaaillus thiooxidans was capable of releasing 23% zinc from
marmatite and its maximum leaching (24.04%) was observed in the case
of synthetic zinc sulphide. Wurtzite, again was attacked least (1.12%
leachability). Thiolxiaillus,thioparus was the least effective leac
hing agent. It is interesting to note that leachability of zinc from
synthetic zinc sulphide, by both acidophilic species (Thiobaaillus
thiooxidans and Thiolxiaillus ferrooxidans), differed signifi-
cantly (Table 3.3.5). However, leachability of marmatite zinc was
43% greater with Thiobacillus ferrooxidans than with the sulphur oxi
dising bacterium, whilst the leachability of marmatite iron was 100-fold
greater with the latter than with the former.
The rates of release of zinc during leaching of the different
zinc sulphides by the three Thiolxicilli are presented in Figs. 3.3.10-
3.3.14. These curves share some similarity in their pattern of rate
changes during the release of zinc ion into solution. During marmati
te leaching the rate of release of zinc by Thiolxicillus ferrooxidans
increased steeply until 20.3 % zinc was released, then the rate
decreased rapidly (Fig. 3.3.10). A similar pattern was observed
during leaching of marmatite with Thiobaaillus thiooxidans. The rate
curve (which was non-linear) reached a maximum by eight ~and started
decreasing after remaining constant for two days. Marmatite was
leached at a much higher rate by Thiobacillus ferrooxidans than by
Thiobacillus thiooxidans. Rates of zinc released during leaching of
synthetic zinc sulphide by Thiobacillus thiooxidans and Thiobacillus
ferrooxidans are shown in Fig. 3.3.11. Synthetic zinc sulphide was
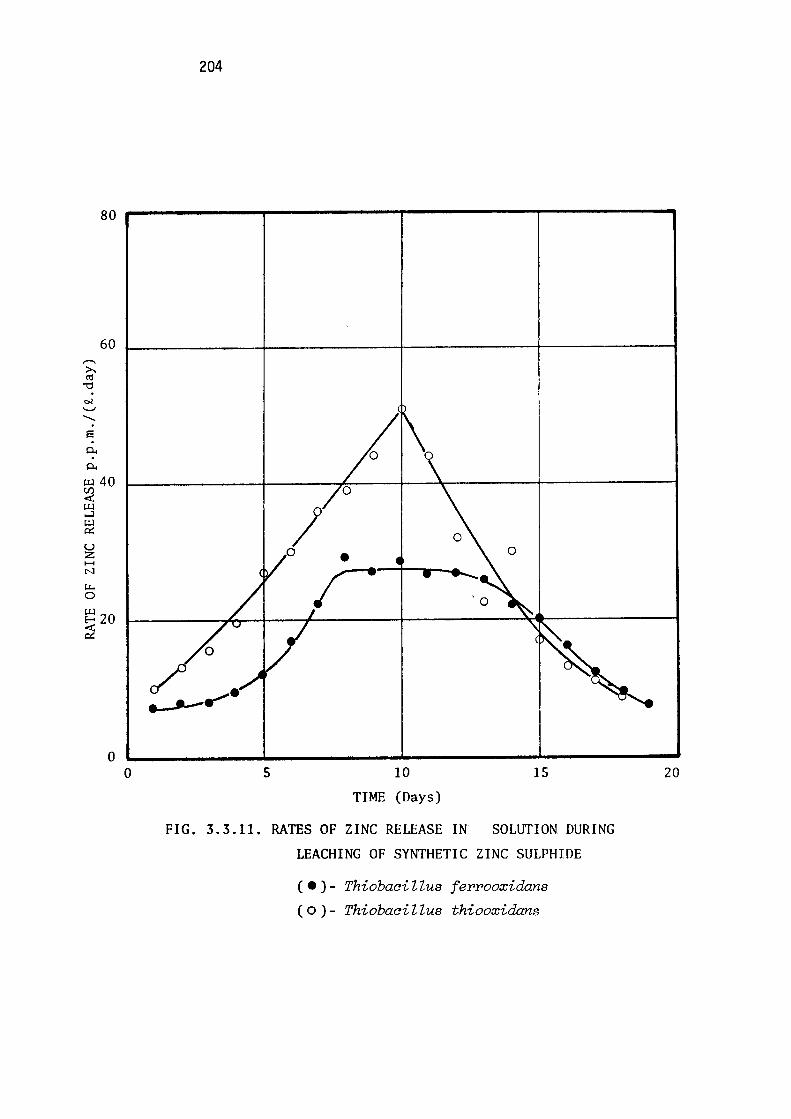
204
80 -------,-.------,---------,-------it
s 0..
0..
60
i:.u 40 r:f) c:i:: i:.u ,._J
~ u z ..... N
t.L. 0
~20 ~
0 0 5
I • r
10
TIME (Days)
15
FIG. 3.3.11. RATES OF ZINC RELEASE IN SOLlITION DURING
LEACHING OF SYNTHETIC ZINC SULPHIDE
( • ) - Thiobaci Uus f errooxidans
( o ) - Thiobaci Z Zus thiooxidans
20

205"
leached rapidly by ThiobaciZZus thiooxida:ns until 14% zinc was
released into solution after which it decreased rapidly. However,
pattern of the rate curve for zinc release by ThiobaciZZus ferrooxi
dans is different from that for marmatite. In this case the rate of
zinc release increased non-linearly over the first eight days of the
experiment, remained constant for the next six days and then decreased.
The total metal leached ( 17% ) in this case was less than that
leached by ThiobaciZZus thiooxida:ns (24%).
Rates of release of zinc into solution
during leaching of sphalerite samples from Spain and Oklahoma are
depicted in Figs. 3.3.12 & 3.3.13. The rates of leaching the
sphalerite sample containing a higher percentage of iron (Oklahoma)
was (like marmatite, but in contrast to synthetic ZnS) higher with
ThiobaciUus ferrooxida:ns than with ThiobaciUus thiooxidans.
Leaching rates of zinc from wurtzite and spanish sphalerite also
increased during the early stages of the experiment and then decrea
sed (Figs. 3.3.12 &' 3.3.14). There was little difference between
the rates of the two organisms . From all these rate curves no
unifying concept emerges which distinguishes the rate pattern of
ThiobaciUus ferrooxida:ns from that of ThiobaciUus thiooxidans.
With some minerals the curves were steeper for one organism but with
other minerals the reverse was the case.
It is apparent from these Figs (3.3.10 - 3.3.14) that the rate
of zinc release increased upto certain maximum and then declined.
The increases and the decreases in rates as well as their maximum
values varied with the mineral and with the organism. What caused the
rate to decline is not known. It cannot have been due to substrate
limitation per se, as maximum leachability achieved during these
experiments was 33% zinc from marmatite . With marmatite,
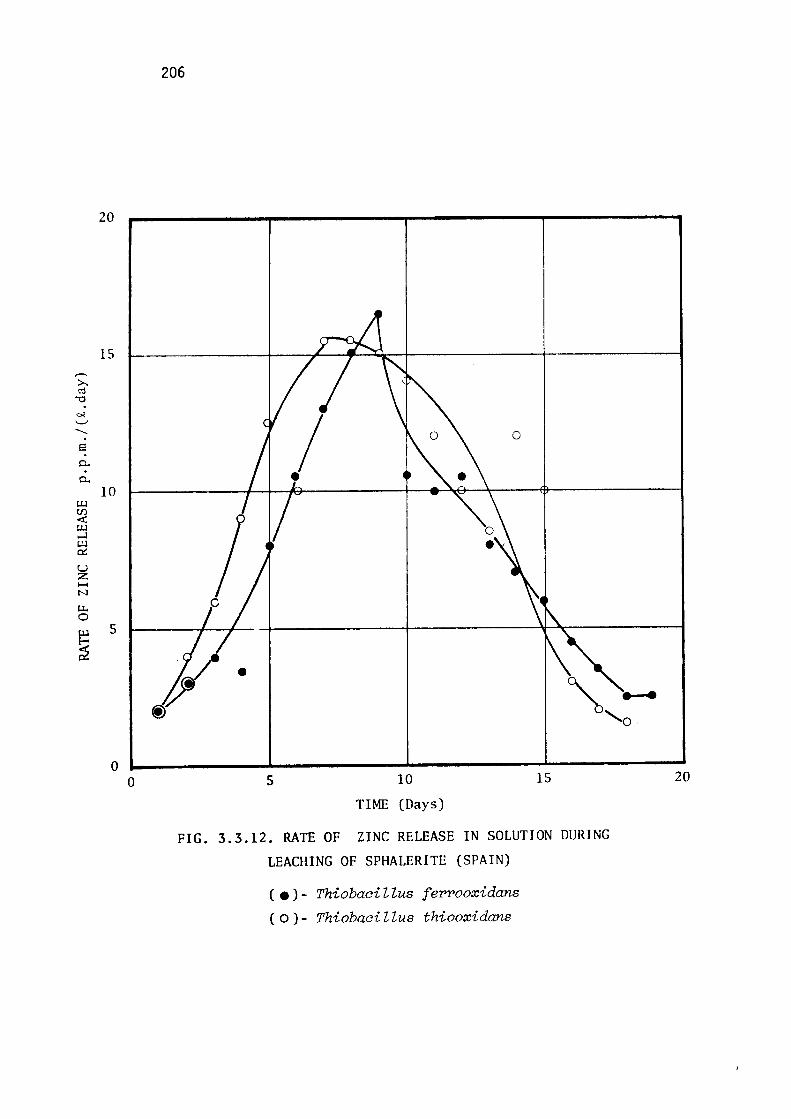
,--, >-. ('j
-0 . ~ '-' -E . p.. . p..
(.J..l rf) <i: (.J..l ~ (.J..l 0:::
u z ..... N
r.i.. 0 (.J..l
~
20
15
10
5
0 0
206
5 10
TIME (Days)
0
15
FIG. 3.3.12. RATE OF ZINC RELEASE IN SOLUTION DURING
LEACHING OF SPHALERITE (SPAIN)
( •) - Thiobaci "l "lus f err>ooxidans
( O ) - Thiobaci 7., "lus thiooxidans
20
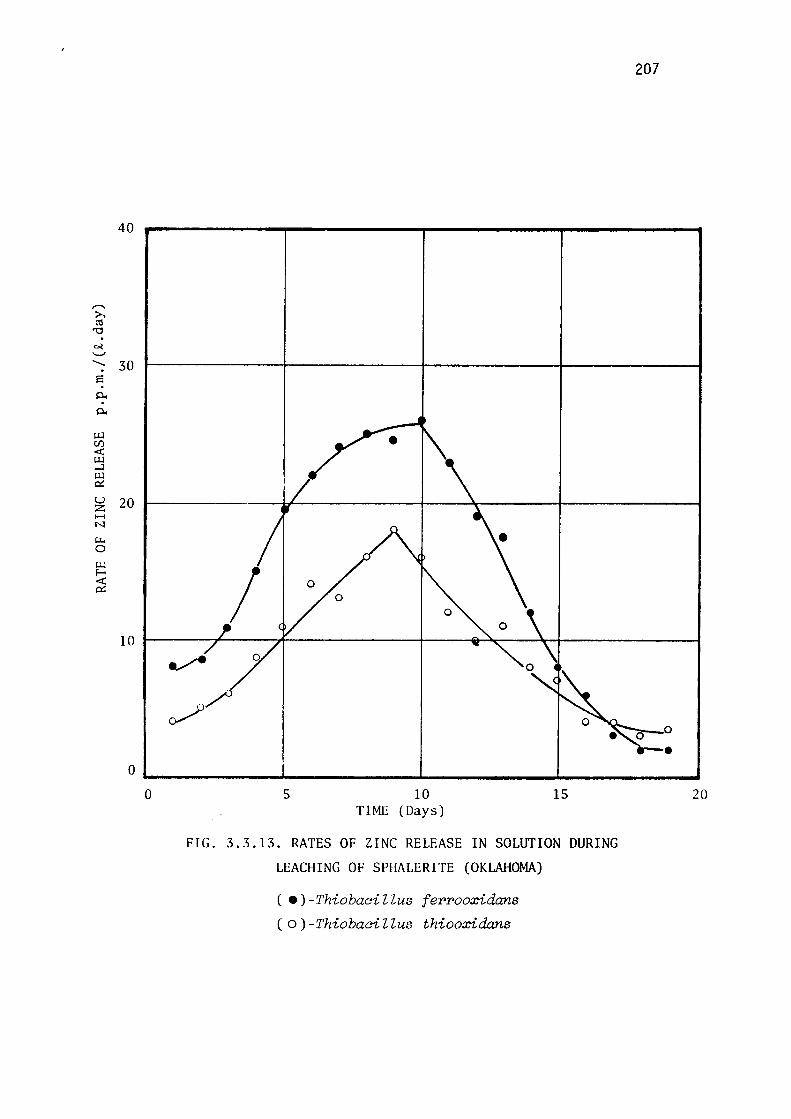
,-.. >.. ro
"O . ~ ~ -. s . 0..
0..
U.l U)
c:i:: U.l ..J U.l 0::
u z ..... N
LI-,
0
U.l E--o
~
207
40
30
20
10
o _______ __._ _______________________ ...,.
0 5 10 TIME (Days)
15
FIG. 3.3.13. RATES OF ZINC RELEASE IN SOLUTION DURING
LEACHING OF SPHALERITE (OKLAHOMA)
( • )-Thiobaci Uus ferrooxidans
( o ) -Thiobaci l lus thiooxidans
20
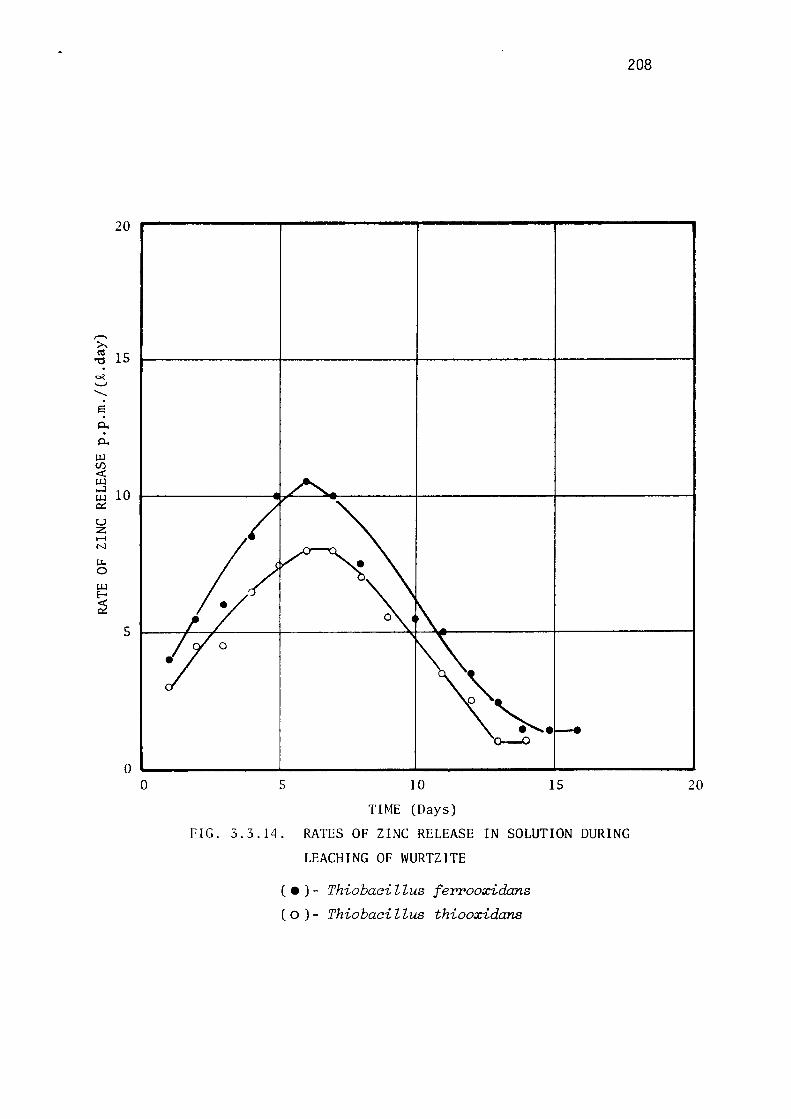
208
20
,...._ >. ~ 15 . ~ .._, -s p.. . p..
c.u U)
-ex: c.u ...J c.u 0:::
u z H N
i:.i.. 0
c.u E-o
~
10
I 5
0 '-, ______ _._ ______ ....,j, _______ ...... ______ __
0 5 10
TIME (Days)
15
FIG. 3.3.14. RATES OF ZINC RELEASE IN SOLUTION DURING
LEACHING OF WURTZlTE
( • ) - Thiobaci l lus f errooxidans
( o ) - Thiobaci l lus thiooxidans
20

209
fonnation of an insoluble precipitate may have been responsible for
deceleration in the rate of leaching by ThiobaciZZus ferrooxidans.
However, this explanation is not valid for ThiobaciZZus thiooxidans;
perhaps the ferrous concentrations released into solution, which have
been shown as being inhibitory to this microorganism (see section 3.3.6),
contributed to the slowing down of the leaching rate. The nature of
the limiting factors in the case of synthetic zinc sulphide remains
unresolved. There are apparently no other toxic metals or other comp
onents present in this mineral, and the maximum zinc contents released
into solution are definitely not injurious to these bacteria. The
inhibition might be due to the non-accessibility of oxygen, which is
one of the important nutrients for these bacteria or due to the forma
tion of new surface characteristics. The changes brought about at
the surfaces of the minerals could contribute to inhibition of mineral
leaching because of their associated changes in the surface-bacteria
relationship. The postulate of Corrans et al. (1972), re:garding the
formation of a passive sulphur layer over covellite and other copper
sulphide minerals during chemical leaching could conceivably be
extended to zinc sulphide system, where perhaps rate of formation of
a passive sulphur layer over the mineral exceeds the rate of its
removal by bacteria. It is obvious that further work is needed to
detennine the exact nature of the factors limiting microbial leaching
of zinc sulphides.
As previously stated (see section 3.3.3), zinc and iron leaching
rates are different when they are present together in mannatite.
Leachability of the iron in marmatite was 39% more than that of zinc in
the case of T.thiooxidans(Table 3.3.5) but was only 1% of the leachability
of zinc in the case of T.ferrooxidans. In order to study the comp-
arative leachability of iron and zinc from marmatite, the molar ratio
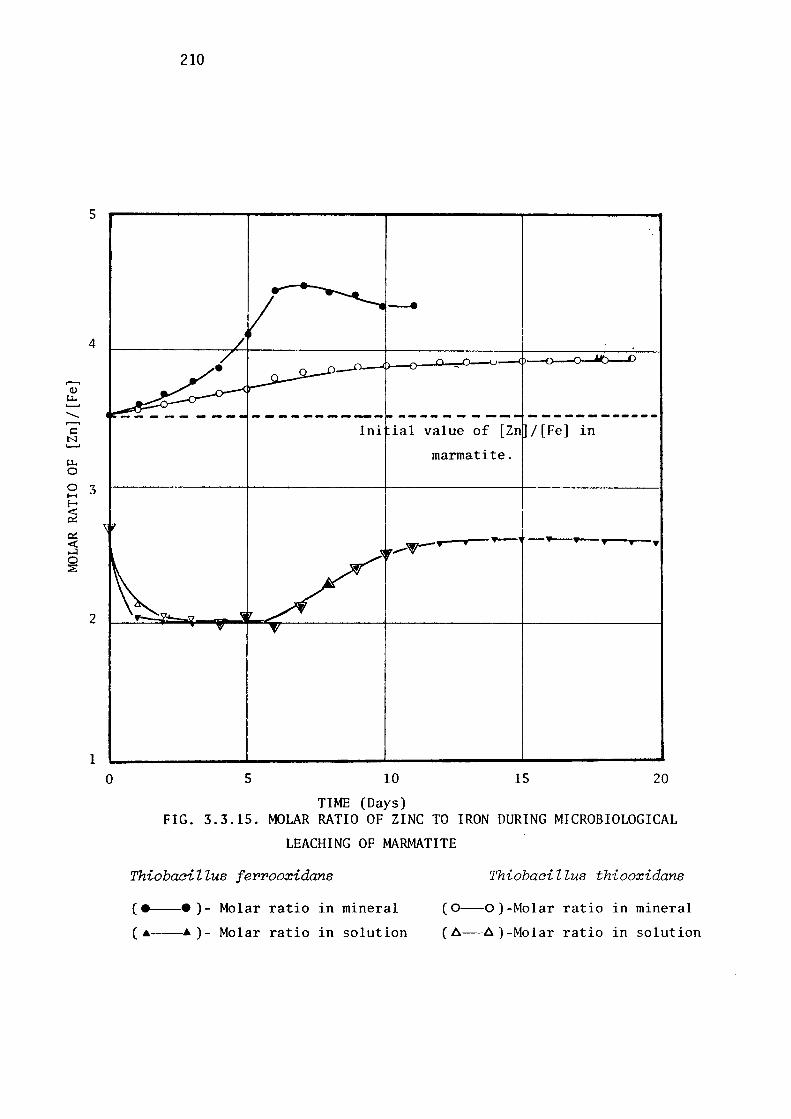
5
4
,....., Q)
LI. ......... -,....., i::: N .........
LI. 0
0 3 H E-
~ 0::: <: ~ 0 :E:
2
1 0
210
----------- --------- ------------
5
Ini ial value of [Zn /[Fe] in
marmatite.
_..,,--.---• -- ---,,--,,.--.
10 15 20
TIME (Days) FIG. 3.3.15. MOLAR RATIO OF ZINC TO IRON DURING MICROBIOLOGICAL
LEACHING OF MARMATITE
ThiobaciZZus ferrooxidans
( e-• ) - Molar ratio in mineral
( •--• )- Molar ratio in solution
Thiobacillus thiooxidans
( 0-0 )-Molar ratio in mineral
( 6-6 )-Molar ratio in solution

211
of [Zn]/[Fe] in solution was plotted in Fig. 3.3.15. These results
indicate that during leaching of marmatite by either of the acidophilic
species of Thiobacillus this ratio decreases abruptly from an initial
value of 2.75 to 2.0, and therefore suggest that in the initial stages
of leaching more iron molecules are released than zinc. The ratio
remained constant between the third and sixth days of the
experiment, the time which coincided with the active period of
bacterial leaching by Thiobacillus ferrooxidans (Fig. 3.3.10). The
ratio then increased and remained constant at a value similar to that
in the original solution. An interesting extrapolation of this observ
ation was to calculate the molar composition of the residual mineral.
The molar ratio of [Zn]/[Fe] in the marmatite (initially 3.52) incre
ased during bacterial leaching (Fig. 3.3.15), the increase being
slower and of lower maximum value when Thiobacillus thiooxidans repla
ced Thiobacillus ferrooxidans as the leaching catalyst. This is
consistent with the fact that Thiobacillus ferrooxidans oxidises
iron much more rapidly than does Thiobacillus thiooxidans. At the
end of the experiment the zinc to iron ratio in the marmatite subjected
to Thiobacillus ferrooxidans was 4.55 whilst that subjected to the
sulphur oxidising bacterium (Thiobacillus thiooxidans) was 3.98. The
third component, sulphur was not estimated and it was not possible to
determine the change in [Zn]/[S] ratio, a value helpful in-predicting
the mechanism of leaching.
These results have therefore indicated that the basic leaching
capacity· of Thiobacilli in decreasing order is Thiobacillus
ferrooxidans > Thiobacillus thiooxidans > Thiobacillus thioparus
in the case of natural minerals and Thiobacillus thiooxidans >
Thiobacillus ferrooxidans > Thiobacillus thioparus in the case of
synthetic, relatively pure zinc sulphides.

212
The results also provide a basis of comparison of the leaching
of the various zinc sulphides by each individual Thiobacillus. It was
obvious that wurtzite was the least amenable to attack by any of the
three Thio/Jacillus species employed. The hexagonal crystallographic
structure in which the component ions of wurtzite are arranged contr
asts with the cubical forms of sphalerite and marmatite, and this may
contribute to its difference in reactivity. However, it must be kept
in mind that results of the kind described can only be considered def
initive if obtained with properly annealed, high purity, synthetic
minerals. Such facilities were not available at the time of experi
mentation, and therefore natural minerals of known composition were
used instead.
A limited arnomt of published information suggests significant
variations in the behaviour of different crystalline forms towards
microbial attack and that further effects can arise from substitutions
in the lattice. A high degree of resistance to oxidation by museum
grade pyrite in the presence of acidophilic, iron oxidising bacteria
was noted by Leathen et al.(1953), who reported the relative ease of
degradation of high grade museum specimen of marcasite by the same
bacterial species. In more detailed investigations of these two iron
sulphide minerals, Silverman et al.(1961) confirmed the resistance of
coarsely crystalline pyrite to oxidative attack by Thiobacillus thioo
xidans and Thiobacillus ferrooxidans and observed that both these
species readily degraded marcasite. Leachability of sphalerite and
marmatite was investigated by Trussell et al.(1964), who reported that
the marmatitic form of zinc sulphide was rapidly and completely leached
by Thiobacillus ferrooxidans but that, mder similar conditions, iron
free sphaleriteshowed only a minor release of zinc ions. These findi
ngs are thus compatible with the results of thecurrent investigation
which indicates that leachability of minerals is dependent upon the

213
structure and composition of the mineral.
Further evidence that the composition of minerals has a major
effect on their leachability is seen in the work of Scott & Dyson
(1968). Zinc sulphide containing copper and iron were shown to be
more readily leached chemically than was the pure mineral. The
catalytic effect of some ions on leaching is so pronotmced that
even 0.05% copper in zinc sulphide is sufficient to affect the
leaching rate significantly (Scott & Dyson, 1968). The catalytic
effect of these ions has been attributed to the possibility of
activating the metal in its lattice and at the same time promoting
the reduction of dissolved oxygen at the cathodic sites on the zinc
sulphide mineral. Sufficient detailed work is not however available
for formulating the precise mechanism operating under such conditions.
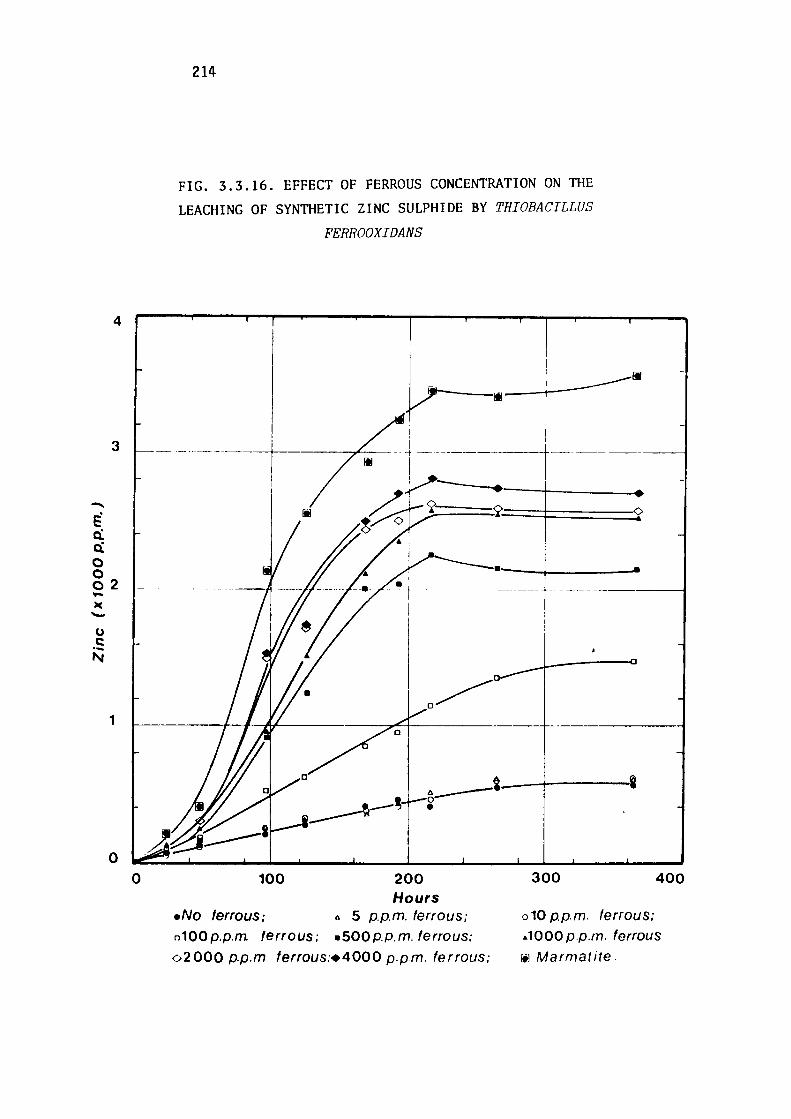
-e a: a:
4
3
0 0 02 ->c -u C:
N
1
0
0
214
FIG. 3.3.16. EFFECT OF FERROUS CONCENTRATION ON THE
LEACHING OF SYNTI-IETIC ZINC SULPHIDE BY THIOBACILLUS
FERROOXIDANS
fil
---------------+----- ----------1
---------1----------------- - ------------l
e.--1-----i "---· 0 •
100 200 Hours
eNo ferrous; " 5 p.p.m. ferrous; o100p.p.m ferrous; .soop,p.m. ferrous:
02000 p.p.m ferrous:+4000 p.p.m. ferrous;
300 400
o 10 p.p.m. ferrous: .. 1000 p.p.m. ferrous
r-1 Marmat ite.

215
3.3.6 EFFECT OF FERROUS IONS ON LE:ACHING OF SYNTHETIC ZINC
SULPHIDE BY THIOBACILLUS FERROOXIDANS & THIOBACILLUS
THIOOXIDANS
The effect of ferrous iron concentration on the leaching rate of
synthetic zinc sulphide was detennined on the presence of Thiobacillus
ferrooxida,ns and ThiobaciUus thiooxiclans. These experiments were
carried out according to the procedure described in section 3.3.S,
except that the pulp density was doubled.
Addition of low concentrations (5-10 p.p.m.) of ferrous iron had
no significant effect upon the leaching curve of the sulphide by Thio
baciZZus ferrooxidans (Fig. 3.3.16) but increasing ferrous concentra
tions between 100 and 2,000 p.p.m. caused a progressive increase in
leaching rate. The total amount of zinc ions leached was increased from
600 p.p.m. at 10 p.p.m. ferrous to 2,575 p.p.m. in the presence of
2,000 p.p.m. ferrous. Marmatite, under similar conditions but without
the addition of ferrous ions, was leached more rapidly than was synth
etic zinc sulphide in the presence of saturating concentrations of
ferrous ion.
The maximum. rates of leaching, and the total amount of zinc
leached in the presence of various concentrations of ferrous ion are
tabulated in the Table 3.3.6. These values indicate that the rate of
zinc release was proportional to the amount of iron present, but
that the system was saturated with respect to iron at concentrations
of 2,000 or greater p.p.m. Fig. 3.3.17 represents the Lineweaver and
Burk plot of values of ferrous ion versus rate of zinc leaching. The
straight line obtained indicates that the addition of ferrous ion to
synthetic zinc sulphide increases zinc leaching in accordance with the
Michaelis-Menton equation. The value of K determined by this plot m
was 555.S p.p.m. (9.92 mM Fe) which closely agreed with the value
obtained by plotting the Eadie-Hofstee plot (586 p.p.m.), (White,
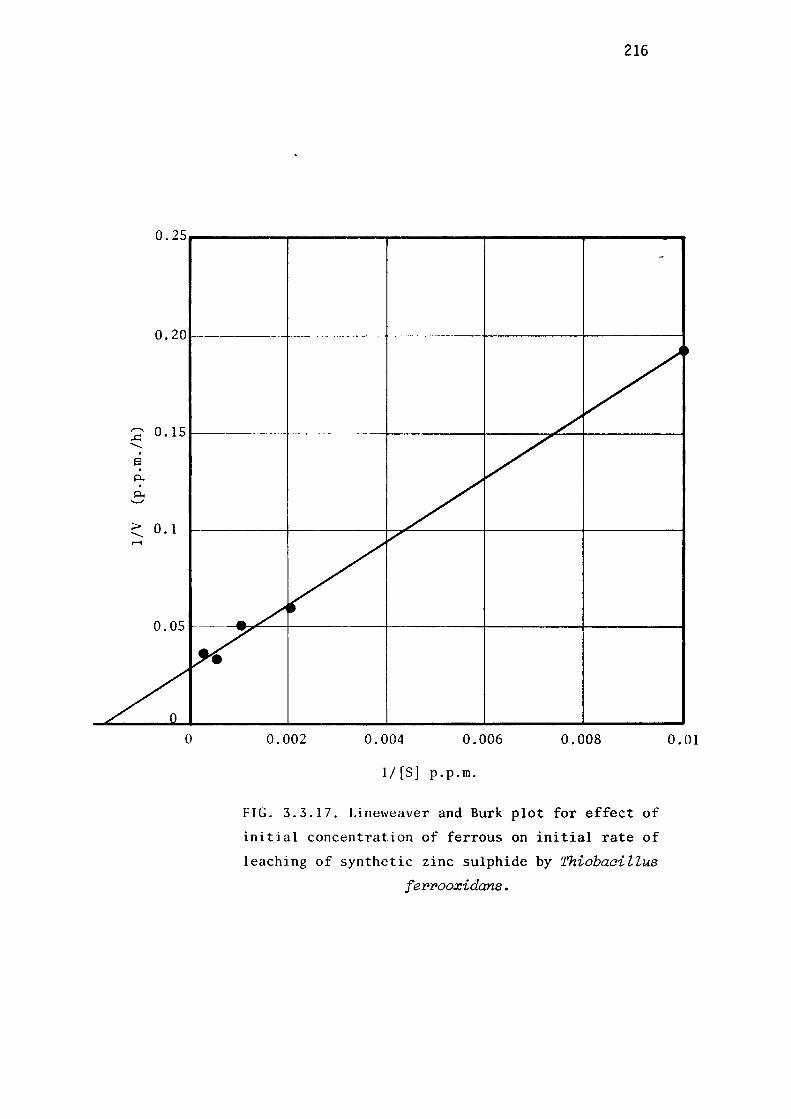
,......_ ..c: -. s . p., . p., '-'
> -...-t
216
0.25----------------------------
0.201-------~---------<~------------1-------t
0.15
0.1
0 0.002 0.004 0.006 0.008
1/ [S] p.p.m.
FIG. 3.3.17. Lineweaver and Burk plot for effect of
initial concentration of ferrous on initial rate of
leaching of synthetic zinc sulphide by Thiobacillus
fer>r>ooxidans.
0.01

217
TABLE 3. 3. 6
EFFECT OF FERROUS IRON CONCENTRATION ON THE LEACHING
OF IRON-FREE ZINC SULPHIDE BY THIOBACILLUS FERROOXIDANS
AND THIOBACILLUS THIOOXIDANS
Ferrous Iron T. ferrooxidans T. thiooxidans Concentration 2+ 2+ 2+ 2+
(ppm) Rate of Zn Tot-al Zn Rate of Zn Total Zn Solution Concn. Solution Concn. (ppm/hr) (ppm) (ppm/hr) (ppm)
0 1. 50 550 2.4 730
5 1. 65 570 2.45 790
10 1. 50 600 2.60 820
100 5.2 1475 2.00 670
500 15.8 2150 1.80 610
1000 20.8 2500 1.8 570
2000 29.0 2575 1. 75 550
4000 28.0 2700
Marmatite 31. 0 3575

218
Handler & Smith, 1973; Lehninger, 1970).
Low concentrations of ferrous ion stimulated leaching by Thio
baciZZus thiooxidans, but concentrations of 100 p.p.m. or more were
inhibitory (Fig. 3.3.18 and Table 3.3.6). For example there was 8%
increase in the leaching rate in the presence of 10 p.p.m. ferrous but
500 p.p.m. ferrous inhibited the rate of zinc release by 25%. Maximum
inhibition (27%) was observed in the presence of 2,000 p.p.m. ferrous
ions.
Discussion.
The environment in which leaching occurs normally contains
dissolved iron in either the ferric or ferrous form. The prominent
role played by iron species during chemical, as well as microbiological,
leaching of sulphide ores has been well documented previously (Scott
& Dyson, 1968; Dutrizac & MacDonald, 1974; Tuovinen & Kelly, 1974;
Forward, 1960; Duncan & Walden, 1972). The results reported in this
and the previous section revealed that, in the absence of additional
iron, the synthetic zinc sulphide was leached slightly more rapidly
by ThiobaciZZus thiooxuia:ns than by ThiobaciZZus ferrooxidans.
With a relatively pure zinc sulphide such as this, virtually the only
oxidisable source for the ThiobaciZZi is the sulphidic moiety on the
solid surface of the substrate and consequently all zinc release is
presumably due to the 'direct action' of the bacterial oxidation of
the sulphur atoms in the crystal lattice, with perhaps some contri
bution from the sulphuric acid produced in accordance with the
following equaion:
+ so + l½ 0 Bacteria
ThiobaciZZus ferrooxidans has been reported to obtain its energy by
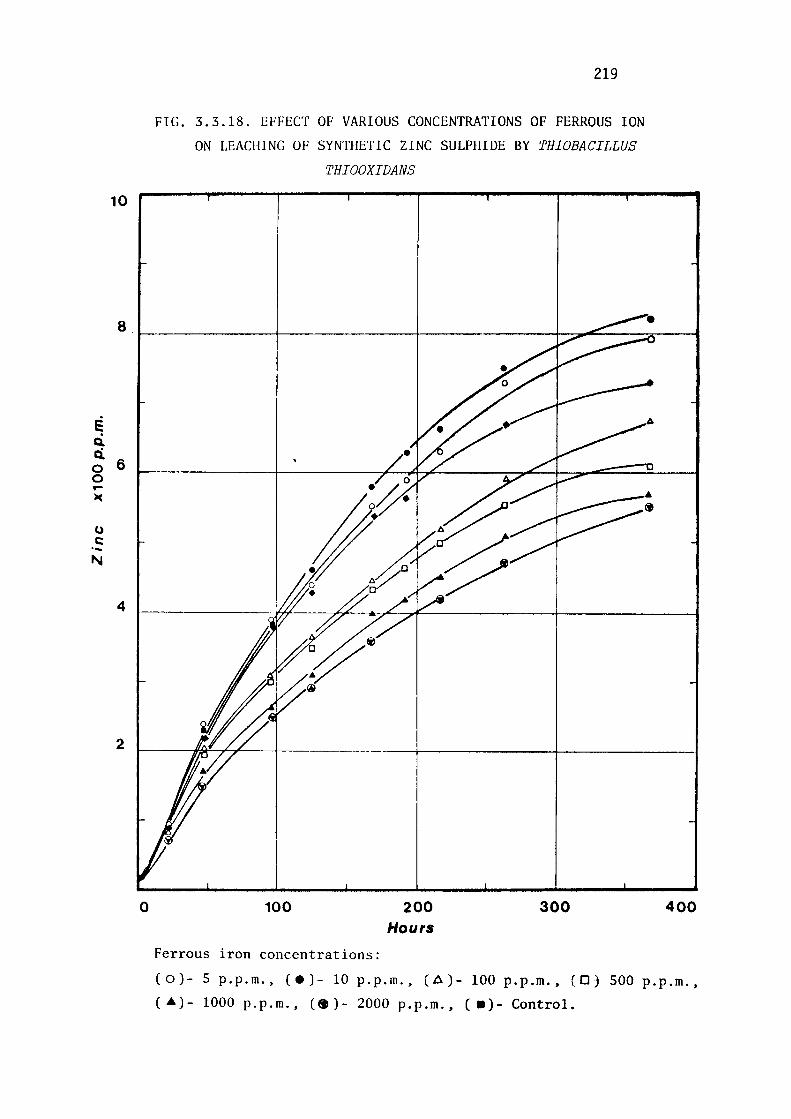
e Q. Q.
6 0 0 .... )(
(,)
C: ·-N
4
0
219
FIG. 3.3.18. EFFECT OF VARIOUS CONCENTRATIONS OF FERROUS ION
ON LEACHING OF SYNTHETIC ZINC SULPHIDE BY THIOBACILLUS
THIOOXIDANS
100
Ferrous iron concentrations:
200 Hours
300 400
(0)- 5 p.p.m., (e)- 10 p.p.m., (.6)- 100 p.p.m., (D) SOO p.p.m.,
( .a)- 1000 p.p.m., C• )- 2000 p.p.m., ( •)- Control.

220
oxidation of metallic ions per se occurring in the lattice,
especially in the case of copper minerals (Nielsen & Beck, 1972;
Golding et al., 1974). Since zinc does not exhibit variable oxidation
states in its minerals, there is very little probability of a direct
oxidation of the zinc species in the crystal lattice (Zajic, 1969).
Addition of ferrous ion to a Thio"baaiZZus ferrooxida:ns culture
provides another energy source for this organism. In the
current investigation its addition stimulated the leaching rate of
zinc from synthetic zinc sulphide by ThiobaaiZZus ferrooxida:ns to
values which exceeded those achieved by ThiobaaiZZus thiooxidans.
The question remaining is whether it did so directly or indirectly.
In any attempt to assess the possible mechanistic roles of ferrous
iron in this situation the following points require consideration:
(a) ferric ions produced by Thio"baaiZZus ferrooxidans' oxidation of
ferrous iron,are an efficientlixiviant and can oxidise sulphides
chemically to sulphates, regenerating ferrous ion again for recycling;
(b) since growth of ThiobaaiZZus ferrooxida:ns on ferrous ions is
luxurious as compared to that on insoluble substrates such as zinc
sulphide, larger population densities become available for direct
leaching of the sulphides. It is hard to differentiate between (a)
and (b), but the following do provide some guidance towards a logical
conclusion. It is a common observation. that the leaching rates of
sterile sulphides by ferric ion at temperatures (30-40°C) favourable
for bacterial growth are slow compared with rates in the presence of
ThiobaaiZZi. Such an observation provides a strong basis for the
assumption that the direct bacterial oxidation of sulphide in the
current investigation contributes more than did the chemical oxidation.
Even stronger evidence for this assumption lies in the fact that addi
tion of ferric sulphate to a leaching medium in the presence of Thio
baaiZZus ferrooxida:ns has been shown to exert no influence on the

leaching of commercial zinc sulphide
1972). Further evidence against
221
concentrates (Duncan & Walden,
a significant contribution
from direct chemical oxidation of the sulphide by ferric iron was
provided by the demonstration in the current results that only a small
concentration of soluble ferric ions were detectable in the culture
media. Once formed, most of the ferric form precipitated, presumably
as jarosite salts of ammonium. Although the formation of jarosite
would decrease the availability of ammonium ions, the only suitable
nitrogen source for the bacteria, the ammonium ion concentration
evidently remained sufficiently high to permit increased growth in
the presence of the iron supplement.
This was the first study to reveal inhibition of zinc leaching
by ThiolJaeiZZus thiooxidans in the presence of high ferrous ion
concentrations. Such inhibition, which could explain, at least
partly, why a comparatively low population of sulphur oxidising bacte
ria is found in natural milieu, could be due to the one or both of
the following possibilities; (a) adsorption of ferrous ions into the
surface of the zinc sulphide particles, which would thereby become
resistant to ThiolJaeiZZus thiooxidans' action, (b) inhibition of the
activities of ThiolJaeiZZus thiooxidans per se, by perhaps, affecting
the cell surfaces and transport mechanisms.
The zinc from marmatite (unsupplemented with additional iron)
was released at a faster rate than from synthetic zinc sulphide
by both species of ThiolJaeiZZi; this indicates the possibility of
some contribution from the interstitial positioning of the iron ions
to the leachability of minerals. The amount of iron released during
marmatite leaching by ThiobaeiZZus ferrooxidans is difficult to
assess, since most of it was precipitated as jarosite salts. However an
assessment can be made from the leaching behaviour of marmatite
with the ThiobaeiZZus thiooxidans, which does not oxidise ferrous

222
ions. In such experiments it was folllld that ferrous and zinc ions
were released in the approximate ratio of 1:2.99 (section 3.3.3) from
marmatite by ThiobaciZZus thiooxidans. If it is assumed that the
same proportion of zinc and ferrous ions would be released from
marmatite by ThiooociZZus ferrooxidans, then for the final zinc conc
entration of 3,575 p.p.m. recorded, the ferrous concentration would
be 1196 p.p.m.
The rate of release of zinc ions from synthetic zinc sulphide by
ThiobaciZZus ferrooxidans in the presence of 1196 p.p.m. is 22.2
mg/(i.h) compared to 31 mg/(i.h) from mannatite . 39% increase in
the rate of release of zinc might have been related to the fact that
the iron atoms were present in the solid marmatite, rather than in
solution. In this form they were released slowly into the solution
without adversly affecting the bacterial growth as the excess ferric
ion has been reported to be inhibitory to both ferrous ion and sulphide
oxidation(Landesman et al., 1966; Razzell & Trussell, 1963a; Dllllcan &
Walden, 1972; Wong et al., 1974). In the solid form they should
affect the electrochemical behaviour of zinc sulphide crystal lattice
or even perhaps the excitation state of the iron/oxygen complex at the
mineral interface. Another possible explanation could be that ferrous
iron leaching from the lattice proceeds that of zinc or sulphide and
leaves 'holes' in the crystal for further activation and penetration
of bacterial and chemical agents. Section 3.3.3 reports that in the
presence of ThiobaciZZus thiooxidans, at least, iron leaches out in
a greater proportion than that expected from its proportion in the
marmatite. This observation needs further investigation before a
more precise theory regarding the mechanism of leaching of minerals of
different composition can be proposed.

4 CONCLUSIONS

223
4. CONCLUSIONS
A comparison of the leaching action of three principal species
of ThiobaciZZi on different zinc sulphide minerals has been studied
with the intention of providing some information regarding leachability
of zinc sulphide within a pH spectrum of 2.0 - S.S. Two principal
species, isolated indigenously, were ThiobaciZZus thiooxidans and
ThiobaciZZus ferrooxidans, representing acidophiles, while ThiobaciZZ
us thioparus was selected to represent non-acidophilic members of the
genus. It was ascertained that isolated species are closely related
to the above mentioned principal species. Taxonomical studies revealed
no strikingly different characteristics of these species other than
those described in Bergey's Manual of Determinative Bacteriology,
(Buchanan & Gibbons, 1974). The FAME (fatty acid methyl ester) profile
of BJR-451 was foillld to be different from that of described by Agate
and Vishniac (1973-a); the F.AM:E profile of BJR-451, however, was later
studied by Dillln et al.(1977), employing more sophisticated and elabor
ate techniques, and was found to be identical to ThiobaciZZus thio
parus. These workers also confirmed, by FAME profile examination,
the identity of two other strains, which were identified as Thiobaci
ZZus thiooxidans and ThiobaciZZus ferrooxidans (section 3.1.4).
The ability of ThiobaciZZus ferrooxidans to utilise elemental
sulphur besides ferrous ion, and its possession of a pH optimum for
growth very similar to that of ThiobaciZZus thiooxidans, enhance the
possibility of its existence in acidophilic cultures of ThiobaciZZus
thiooxidans. Isolation of the new associate, ThiobaciZZus acidophiZus,
from a standard culture of ThiobaciZZus ferrooxidans indicates the
possibjle usage of 'mixed' cultures in the past (Guay & Silver, 1975)
and casts doubt on the validity of certain comparative. studies made.

224
Lack of availability of specific media for isolation of each disti-
nct ThiobaeiZZus species can lead to the isolation of mixed species
and a need to devise such media is very urgent for ecological surveys
of mine fields and exhausted dumps. However, in the current investig
ation, particular attention has been paid to purification of each
strain studied. Although micro-manipulation for single cell iso
lation was not employed, the isolated suspensions behaved homogeneou
sly and yielded reproducible results. Absence of high ferric ion
concentration in solution when mannatite was leached by ThiobaeiZZus
thiooxidans, even in the presence of ferrous ion,, indicated that the
sulphur-oxidiser was free from ThiobaeiZZus ferrooxidans.
BJR-451 was studied in detail and its pH and temperature optima
determined. Its pH curve was sharp and optimal at S.S. (Acidic
environments (pH below 4.5) were bacteriostatic, whereas lower pH
values (4.0 - 3.5) were bacteriocidal.
Tolerance of the three ThiobaeiZZi to zinc ion was determined,
and it was found that ThiolxieiZZus ferrooxidans could withstand
concentrations of zinc as high as 20,000 p.p.m. However, ThiobaeiZZus
thiooxidans was capable of tolerating zinc concentrations only up to
15,000 p.p.m. without deleterious effects on its growth being observed.
ThiobaeiUus thioparus' maximum zinc tolerance level could not be
determined, because of the rapid changes in pH resulting from the
addition of zinc sulphate solution. A most notable observation was
that high zinc concentrations influenced growth only by prolonging the
lag period which was followed by growth at a rate similar to that of
the control without zinc). This suggests a complete adaptation by
these bacteria to zinc. The low metal tolerance observed in the case
of ThiobaeiZZus thiooxidans might have been due to its growth on
sulphur; this would accord with the suggestion that ThiobaeiZZus
ferrooxida:ns grown on thiosulphate as the sole energy source is less

225
tolerant to metals than cells harvested from medium containing ferrous
ions (Tuovinen et al., 1971). Changes in metal tolerance constitute
another very important field in which further research should be
carried out before definitive statements regarding species differen
ces can be made. Metal resistant plasmids have been reported in
some bacteria (Summers et al., 1975) and, with the advent of modern
genetic engineering technology, a suitable metal tolerant plasmid
should provide a desirable ThiobaaiZZi for each particular assign
ment when introduced into the appropriate bacterium.
The observation made by Mayling (1969) for extracting alkaline
ores of zinc and copper could not be confirmed by using ThiobaciZ Zus
thioparus. The leaching capabilities of this species was found to
be minimwn when compared with the two acidophilic species (i.e.,
ThiobaciUus thiooxidans and ThiobaciUus fe:rTooxidans). Low metal
solubilisation by ThiobaciZZus thioparus might have been due to one or
more of the following possibilities: viz., low rate of oxidation of
sulphide; metal toxicity; the pH of the environment; the high concent
ration of phosphates in growth medium. At pH 5.5 zinc phosphates and
hydroxides might have been released into solution and have precipitated.
However, no direct evidence in support of this postulate is forwarded.
ThiobaciZZus thiooxidans leached synthetic zinc sulphide more
effectively than did ThiobaciZZus ferrooxidans.
An early observation made by Trussell et al.(1964), regarding
the i higher leachability by ThiobaciUus ferrooridans of marrnatite
compared with sphalerite, was confirmed. The work was extended to a
study of the behaviour of various zinc sulphides, differing in
their composition and structure. This was the first investigation
into the effects of structures and compositions of minerals on their

226
relative leachability. It was fotmd that, in addition to the physico
chemical parameters (which have been already discussed by many workers
and are summarised in section 1.3.3.), two other factors influence the
leaching rates: (a) the bacterial species involved; and (b) the
mineral used. It has been observed that, when leaching natural
museum-grade mineral specimens, the capabilities of Thio"baciZZi to
leach them are, (in decreasing order) Thio"baciZZus ferrooxida:ns >
ThiobaciZZus thiooxida:ns > ThiobaciZZus thioparus and for
synthetic mineral the capabilities are,(in decreasing order)
Thio"baciZZus thiooxidans > Thio"baciZZus ferrooxida:ns > Thiobac
iZZus thioparus. It would be most informative to determine the rela
tive behaviour of every species of the genus Thio"baciZZus in its
ability to leach other zinc ( and other metal) sulphides.
It was shown that the hexagonal species of ZnS (wurtzite) is
very inert towards leaching by all species of ThiobaciZZi; natural
sphalerite specimens were leached by these bacteria, but. not to a
significant degree. Maximum leachability, however, was found in
the case of synthetic zinc sulphide and marmatite. Present data
are insufficient for postulating a reason for wurtzite's inertness.
It may be due to the difference in the arrangement of sulphur tetra
hedra in the crystal of wurtzite. Further investigation involving
electrochemical and electron-probe studies of the wurtzite before and
after leaching are needed before any firm conclusion can drawn.
Marmatite ( Zn 3 • 5Fe S5) was released by both acidophilic species
to give a soluble zinc to iron ratio which progressively decreased in
the early stages of leaching as a result of a iron being released
at a rate which was faster than the rate of zinc release,consequently,
the ratio of zinc to iron in the residual solid mineral increased.
The general nature of atomic point defects in ZnS indicates that
Fe is inserted in 'holes' rather than in valency bands, and an

227
explanation for the higher leachability of marmatite can be based on
the reasoning that bacteria extract ' ferrous' from these holes,
and that subsequently zinc is leached as a result of the collapse of
unit cells. However, a rigorous study should be made involving the use of
electron probe and thin section techniques in an attempt to obtain
definitive evidence for pitting and changes in the relative concen
trations of the component metal atoms.
Zinc sulphide crystals can accommodate many different atomic
species in their 'holes' (Shuey, 1975), viz., noble elements (Cu, Ag,
Au) and alkali metals of group I (Li, Na) which produce acceptor
defects, as well as Br, Cl and I (as substituents for sulphur), which
produce donor effects. It is known that the presence of Cu, Cd, or Mn
in traces catalyzes the pressure leaching of zinc (Scott & Dyson,
1968). However no systematic study involving the effects of substi
tuents in the zinc sulphide lattice on ZnS leaching by bacteria is
known; such information could provide very useful guidelines for deter
mining the relationship between mineral structure and leaching.
It was shown that the presence of ferrous ions in the leaching
medium were inhibitory to the sulphur oxidising bacteria. (Previously,
no information regarding the effects of iron on the activities of these
bacteria was available.) On the other hand, enhanced rate of leaching
of zinc from synthetic zinc sulphide by ThiobaciUus ferrooxidans
was observed on the addition of ferrous ions to the system. The
effects of iron present in the marmatite crystals and that added exte
rnally to synthetic zinc sulphide were not comparable; the iron
present in the marmatite was shown to be more efficient than was added
iron.
A study employing marmatite of various ferrous contents would
be extremly useful in determiningthe definitive role of iron in
leaching. Such studies would provide a significant contribution to

228
the newly emerging field of biohydrometallurgy.


229
5. R E F E R E N C E S
AGATE, A. D., 1973. Survey of ThiobaciZZus ferrooxidans in solubil
isation of copper sulphide ores. NML Technical Journal 15: 25-27.
AGATE, A. D. and VISHNIAC,W.,1969. Changes in the lipid composition
of ThiobaciZZus neapolitanus. Bacteriological Proceedings P29.
AGATE, A. D. and VISHNIAC, W., 1973. Changes in phospholipids compo
sition of ThiobaciZZus neapolitanus during growth. Archives
fur Mikrobiologie 89: 247-256.
AGATE, A. D. and VISHNIAC, W., 1973-a. Characterization of Thiobaci
ZZus species by gas-liquid chromatography of cellular fatty
acids. Archives fur Mikrobiologie 89: 257-267.
AGBIM, N. N. and DOXTADER, K. G., 1975. Microbial degradation of
zinc silicates. Soil Biology and Biochemistry 7: 275-280.
ALEEM, M. I. H., 1966. Generation of reducing power in chemosynthesis.
II-Energy linked reduction of pyridine nucleotides in the chemo
autotroph Nitrosomonas europae. Biochimica et Biophysica Acta
113: 216-224.
ALEEM, M. I. H., 1966-a. Generation of reducing power in chemosyn
thesis. III- Energy linked reduction of pyridine nucleotides in
ThiobaciZZus noveZZus. Journal of Bacteriology 91: 729-736.
ALEEM, M. I. H., 1966-b. Generation of reducing power in chemosyn
thesis. IV- Energy linked reduction of pyridine nucleotides by
succinate in ThiobaciZZus noveZZus. Biochimica et Biophysica
Acta 128: 1-12.
ALEEM, M. I. H., 1969. Generation of reducing power in chemosynthesis.
VI- Energy linked reduction in the chemoautotroph ThiobaciZZus

230
VI- Energy linked reduction in the chemoauto troph ThiobaciZZus
neapoZitanus. Antonie van Leeuwenhoek, Journal of Microbiology
and Serology 35: 379-391.
ALEEM, M. I.H., 1970. Oxidation of inorganic nitrogen compounds.
Annual Review of Plant Physiology 21: 67-90.
ALEEM, M. I. H., 1975. Biochemical reaction mechanisms in sulfur
oxidation by chemosynthetic bacteria. Plant and Soil 43: 587-607.
ALEEM, M. I. H., 1977. Coupling of energy with electron transfer
reactions in chemolithotrophic bacteria. Symposia- Society for
General Microbiology 27: 351-381.
ALEEM, M. I. H. and HUANG,E., 1965. Carbon dioxide fixation and
carboxydismutase in ThiobaciZZus noveZZus. Biochemical and
Biophysical Research Communications 20: 515-520.
ALEEM, M. I. H. and NASON, A., 1960. Phosphorylation coupled to
nitrite oxidation by particles from the chemoautotroph Nitrobacter
agiZis. Proceedings of the National Academy of Sciences of the
United States of America 46: 763-769.
ALEEM, M. I. H., LEES, H. and NICHOLAS, D. J. D., 1963. ATP-dependent
reductase of nicotinamide adenine dinucleotide by ferro-cytochrome
c in chemoautotrophic bacteria. Nature 200: 759-761.
ANDERSEN, J.E. and ALLMAN, M. B., 1968. Some operational aspects of
heap leaching at Rum Jungle. Proceedings of the Australian
Institute of Mining and Metallurgy 225: 27-31.
ANDERSEN, K. J. and LUNDGREN, D.G., 1969. Enzymatic studies of the
iron-oxidizing bacterium FerrobaciZZus ferrooxidans : Evidence
for a glycolytic pathway and Krebs cycle. Canadian Journal of
Microbiology 15: 73-79.

231
ARRIETA, L. and GREZ, R., 1971. Solubilisation of iron-containing
minerals by soil microorganisms. Applied Microbiology 22: 487-490.
ASHMEAD, D., 1955. The influence of bacteria in the formation of acid
mine water. Colliery Guardian 190: 694-698.
AUBERT, J. P., MILHAUD, G. and MILLET, J., 1957. Carbon metabolism
in chemosynthetic autotrophism. Carbon dioxide fixation on
phosphoenol pyruvic acid. Comptes Rendus Des Travaux du Labora
toire Carlsberg 244: 398-401.
AUBERT, J. P., MILHAUD, G. and MILLET, J., 1957-a. The path of
carbon dioxide assimilation in chemoautotrophic bacteria.
Annales de l'Institut Pasteur 92: 515-524.
AUDSLEY, A. and DABORN, G. R., 1963. Natural leaching of uranium ores.
II- A study of experimental variable$. Transactions, Institute
of Mining and Metallurgy 72: 235-246.
AVAKYAN, A. A. and KARAVAIKO, G. I., 1970. Submicroscopic organization
of Thiobacillus ferrooxidans. Mikrobiologiya 39: 855-861.
AVEN, M. and PRENER, J. S., 1967. Physics and chemistry of II-VI
compounds. North Holland, Amsterdam, 844 pp.
BAALSRUD, K. 1954. Physiology of Thiobacilli. Symposia- Society
for General Microbiology 4 : 54- 67 •
BMLSRUD, K. and BAALSURD, K. S., 1954. Studies on Thiobacillus
denitrificans. Archives fur Mikrobiologie 20: 34-62.
BAAS-BECKING, L.G. M., KAPLAN, I. R. and MOORE, D., 1960. Limits
of the natural environment in terms of pH and oxidation-reduction
potentials. The Journal of Geology 68: 243-284.

232
BABIJ, T., DOBLE, R. and RALPH, B. J., 1971. Mobile equipment for
the sterilisation of fermentation media by filtration.
Chemistry and Industry pp. 1045-1046.
BABIJ, T., MOSS, F. J. and RALPH, B. J., 1969. Effects of oxygen
and glucose levels on lipid composition of yeast Candida utilis
grown in continuous culture. Biotechnology and Bioengineering
11: 593-603.
BARBIC, F. and BRACILOVIC, D., 1975. Effect of mineral acids on
iron bacteria. Zeitschrift fur Allgemeine Mikrobiologie 15:
307-313.
BARBIC, F. F., 1977 . Effects of different compounds of metals and
their mixtures on the growth and survival of Thiobacillus
ferrooxidans. Zeitschrift fur Allgemeine Mikrobiologie 17:
277-281.
BARDIYA, M. C. and GAUR, A. C., 1974. Isolation and screening of
microorganisms dissolving low grade rock phosphate.
Folia Microbiologica 19: 386-389.
BARTON, L. L. and SHIVELY, J.M., 1968. Thiosulfate utilization by
Thiobacillus thiooxidans ATCC 8085. Journal of Bacteriology
95: 720.
BARVENIK, F. W. and JONES, G. E., 1969. Chemosynthetic productivity
in a coastal pond. Bacteriological Proceedings G38, p. 124.
BARVINCHAK, G., 1975. Investigation of the mechanism of iron
oxidation by cells and cell envelopes of Thiobacillus ferrooxi
dans. Ph.D. Thesis, Syracuse University, U.S.A. [cited from

233
Lazaroff, 1977].
BECK, J. V., 1960. A ferrous iron oxidizing bacterium. I- Isolation
and some general physiological characteristics. Journal of
Bacteriology 79: 502-509.
BECK, J. V., 1967. The role of bacteria in copper mining operations.
Biotechnology and Bioengineering 9: 487-497.
BECK, J. V., 1969. Thiobacillus ferrooxidans and its relation to the
solubilization of ores of copper and iron. In: Perlman, D., Ed.,
Fermentation Advances pp. 747-771. Academic Press, London.
BECK, J. V. and BROWN, D.G., 1968. Direct sulfide oxidation in the
solubilization of sulfide ores by Thiobacillus ferrooxidans.
Journal of Bacteriology 96: 1433-1434.
BECK, J. V. and ELSDEN, S. R., 1958. Isolation and some characteris
tics of an iron oxidizing bacterium. Journal of General
Microbiology 19: i.
BECK, J. V. and SHAFIA, F. M., 1964. Effect of phosphate ion and
2:4-Dinitrophenol on the activity of intact cells of ThiobaciZZ
us ferrooxidans. Journal of Bacteriology 88: 850-857.
BEIJERINCK, M. W., 1904. Uber die bakterienwelche sich in dunkeln
mit kohleusaure als kohleusloffquelle ernahren konneu.Zentralbl
att fur Bakteriologie, Parasitenkunde. II Abt. 11: 593-599.
~ited from Roy & Trudinger, 1970, p.210.]
BERGEY, D. H., HARRISON, F. C., BREED, R. S., HAMMER, B. W. and
HUNTOON, F. H., 1925. Bergey's Manual of Determinative Bacteriology,
2nd. Edition. pp. 1-462. The Williams & Wilkins Co., Baltimore.
BERRY, L. G. and MASON, B., 1959. Mineralogy - pp. 308-317. W. H.
Freeman and Company, San Francisco & London.

234
BERRY, V. K. and MURR, L. E., 1975. Bacterial attachment to molybde
nite: An electron microscope study. Metallurgical Transactions B,
6B: 488-490.
BITHER, T. A., 1967. Second International Conference on Solid
Compounds. Trans. Elem. Enschede. p.26.
[ Cited from: JELLINEK, F., 1968].
BITHER, T. A., BOUCHARD, R. J., CLOUD, W. H., DONOHUE, P. C. and
SEIMONS, W. J., 1968. Transition metal pyrite dichalcogenides.
High-pressure synthesis and correlation of properties.
Inorganic Chemistry 7: 2208-2220.
BLAYLOCK, B. A.and NASON, A., 1963. Electron transport system of
the chemoautotroph Ferrobacillus ferrooxidans. Journal of
Biological Chemistry 238: 3453-3462.
BLOOR, W. R., 1928.
The determination of small amounts of lipids in blood plasma.
Journal of Biological Chemistry 77: 53-73.
BODO, C. A. and LUNDGREN, D. G., 1971. Uptake and oxidation of iron
Thiobacillus ferrooxidans. Bacteriological Proceedings P44. p.131.
BORICHEWSKI, R. G., 1967. Keto acids as growth limiting factors in
autotrophic growth of Thiobacillus thiooxidans. Journal of
Bacteriology 93: 597-599.
BORICHEWSKI, R. G. and UMBREIT, W.W., 1966. Growth of Thiobacillus
thiooxidans on glucose. Archives of Biochemistry and Biophysics

235
116: 97-102.
BOURNE, E. J., NERY, R. and WEIGEL, H., 1959. Metal chelates of
polyhydroxy compounds. Chemistry and Industry, August 1,
pp. 998-999.
BRIERLEY, J. A., 1966. Contribution of chemoautotrophic bacteria
to the acid thermal waters of the Geyser Spring Group in
Yellowstone National Park. Ph. D. 'l'hesis, Montana State Unive
rsity, Boseman, Montana, U.S.A.
BRIERLEY, C.L., 1974. Molybdenite leaching: use of a high-temperature
microbe. Journal of Less-Common Metals 36: 237-242.
BRIERLEY, J. A., 1977. Thermophilic microorganisms in extracion
of metals from ores. Developments in Industrial Microbiology
18: 273-284.
BRIERLEY, C. L. and BRIERLEY, J. A., 1973. A chemoautotrophic and
thermophilic microorganism isolated from an acid hot spring.
Canadian Journal of Microbiology 19: 183-188.
BRIERLEY, C. L. and MURR, L. E., 1973. Leaching: Use of a thermophi
lic and chemoautotrophic microbe. Science 179: 488-490.
BRIERLEY, J. A. and Le ROUX, N. W., 1977. A facultative thermophilic
Thiobacillus like bacterium: Oxidation of iron and pyrite.
GBF Monograph Series, No. 4 (August, 1977), 55-66, Conference
Bacterial Leaching 1977.
BRIERLEY, J. A. and LOCKWOOD, S. J., 1977. The occurrence of
thermophilic iron-oxidizing bacteria in a copper leaching system.

236
FEMS Microbiology Letters 2: 163-165.
BROCK, T. D. and DARLAND, G. K., 1970. Limits of microbial exiatence:
Temperature and pH. Science 169: 1316-1318.
BROCK, T. D. and GUSTAFSON, J., 1976. Ferric iron reduction by
sulfur and iron-oxidizing bacteria. Applied and Environmental
Microbiology 32: 567-571.
BROCK, T. D., BROCK, K. M., BELLY, R. T. and WEISS, R. L., 1972.
Sulfolobus: A new genus of sulfur-oxidizing bacteria living at
low pH and high temperature. Archives fur Mikrobiologie 84:
54-68.
BROOKS, H., 1972. Materials in a steady state world.
Metallurgical Transactions 3: 759-768.
BRUNAUER, S., EMMETT, P. H. and TELLER, E. J., 1938. Adsorption of
gases in multimolecular layers.
Society 60: 309-319.
Journal of American Chemical
BRUYNESTEYN, A, and DUNCAN, D. W., 1971. Microbiological leaching of
sulphide concentrates. Canadian Metallurgical Quarterly 10:57-63.
BRYNER, L. C. and ANDERSON, R., 1957. Microorganisms in leaching
sulfide minerals. Industrial and Engineering Chemistry 46:
1721-1724.
BRYNER, L. C. and JAMESON, A. K., 1958. Microorganisms in leaching
sulphide minerals. Applied Microbiology 6: 681-687.

237
BRYNER, L. C., BECK, J. V., DAVIS, D. B., and WILSON, D. G., 1954.
Microorganisms in leaching sulfide minerals. Indastrial and
Engineering Chemistry 46: 2587-2592.
BRYNER, L. C., WALKER, R. B. and PALMER, R., 1967. Some factors
influencing the biological and non-biological oxidation of
sulfide minerals. Transactions of Society of Mining Engineering
238: 56-61.
BUCHANAN, R. E. and GIBBONS, N. E. (ed.), 1974. Bergey's Manual of
Determinative Bacteriology, 8th Edition, pp. 456-461. The
Williams and Wilkins Company, Baltimore, U.S.A.
BITTLER, R. G., 1975. Pyruvate inhibition of the carbon dioxide
fixation of the strict chemolithotroph Thiobacillus thiooxidans.
Canadian Journal of Microbiology 21: 2089-2093.
BITTLER, R. G. and UMBREIT, W.W., 1966. Absorption and utilization of
organic matter by the strict autotroph Thiobacillus thiooxidans
with special reference to aspartic acid. Journal of Bacteriology
91: 661-666.
CALDWELL, D. E., CALDWELL, S. J. and LAYCOCK, J. P., 1976. ThePmo
thrix thioparus gen. et sp. nov. a facultatively anaerobic
facultative chemolithotroph living at neutral pH and high
temperature. Canadian Journal of Microbiology 22: 1509-1517.
CHAINIKOVA, N. A., 1962. Flotation properties of sphalerite varieties
having various iron contents (Maritime Territory Deposits).
Soobschch. Dal'nevost. Filiala. Sibirsk. Otd. Akad. Nauk. SSSR

238
No. 15, 47-51.[cited from Chemical Abstracts, 7425b : 62: 1965].
CLEVELAND, J. H., 1964. Variation of iron sulfide content in
sphalerites of Mississippi Valley Deposits. Proceedings of
Indiana Academy of Sciences 74: 271-277.
CLOUD, P., 1968. Mineral resources from the sea. In Resources and
Man, p. 135. W. H. Freeman & Co., San Francisco.
COCCUZZA, A. and NICOLETTI, G., 1961. Metabolic characteristics of
certain chemosynthesizing species of autotrophic Thiobacteria
isolate. Atti. Soc. Peloritana Sci. Fis. Mat. Nat., 7: 769-775.
COLES, J. S.III, and ALEEM, M. I. H., 1973. Electron transport linked
with proton induced ATP generation in Thiobacillus novellus.
Proceedings of National Academy of Sciences of the United
States of America 70: 3571-3575.
COLMER, A. R. and HINKLE, M. E., 1947. The role of microorganisms
in acid mine drainage- A preliminary report. Science 106: 253-256.
COLMER, A. R., TEMPLE, K. L. and HINKLE, M. E., 1950. An iron
oxidizing bacterium from the acid drainage of some bituminous
coal mines. Journal of Bacteriology 59: 317-328.
CORRANS, I. J., 1970. Studies on the bacterial leaching of natural
and synthetic iron copper sulfide minerals. M.Sc. Thesis,
University of New South Wales.
CORRANS, I. J., HARRIS, B. and RALPH, B. J., 1972. Bacterial
leaching: An introduction to its application and theory and a
study of its mechanism of operation. Journal of the South African
Institute of Mining and Metallurgy 72: 221-230.
CORRANS, I. J. and SCHOLTZ, M. T., 1976. A kinetic study of the
leaching of pentlandite in acidic ferric sulphate solutions.
Journal of the South African Institute of Mining and Metallurgy,

239
76: 403-411.
CORRICK, J. D. and SUTTON, J. A., 1961. Three chemosynthetic autotro
phic bacteria important to leaching operations at Arizona Copper
Mines. United States Bureau of Mines Investigation Report 5719.
CORRICK, J. D. and SUTTON, J. A., 1965. Copper extraction from a low
grade ore by Ferrobaaillus ferrooxidans. Effects of environmnetal
factors. United State Bureau of Mines Investigation Report 6717.
De CASTRO, A. F. and EHRLICH, H. L., 1970. Reduction of iron oxide
minerals by a marine Bacillus . Antonie van Leeuwenhoek Journal
of Microbiology and Serology 36: 317-327.
De CUYPER, J. A., 1964. Bacterial leaching of low-grade copper and
cobalt ores. In: Wadesworth, M. E. & Davis, F. T. (Eds.), Unit
Processes in Hydrometallurgy 24: 126-142.
De KRUYFF, C. D., Van DER WALT, J. P. and SCHWARTA, H. M., 1957.
The utilization of thiocyanate and nitrate by Thiobaailli. Antonie
van Leeuwenhoek Journal of Microbiology and Serology 23: 305-316.
DIN, G. A., SUZUKI, I. and LEES, H., 1967. Carbon dioxide fixation
and phosphoenol pyruvate carboxylase in Ferrobaaillus ferrooxidans.
Canadian Journal of Microbiology 13: 1413-1419.
DOESTSCH, R. N. and COOK, T. M., 1973. Bacterial attack on
minerals. In: Introduction to Bacteria and their Ecobiology
pp.342-247. University Park Press, Baltimore, Maryland, U.S.A.
DUFF, R. B. and WEBLEY, D. M., 1959. 2-ketogluconic acid as a natural
chelator produced by soil bacteria. Chenistry and Industry,
pp. 1376-1377.
DUFF, R. B., WEBLEY, D. M. and SCOTT, R. 0., 1963. The solubilisation

of minerals and related materials by 2-ketogluconic acid
producing bacteria. Soil Science 95: 105-114.
DUGAN, P.R. and LUNDGREN D. G., 1964. Acid production by Ferro
bacillus ferrooxidans and its relation to water pollution.
Developments in Industrial Microbiology 5: 250-257.
240
DUGAN, P. R. and LUNDGREN, D. G., 1965. Energy supply for the chemo
autotroph Ferrobacillus ferrooxidans. Journal of Bacteriology
89: 825-834.
DUNCAN, D. W. and BRUYNESTEYN, A., 1971. Enhancing bacterial activity
in an uranium mine. Canadian Mining and Metallurgical Bulletin
64: 32-36.
DUNCAN, D. W. and MCGORAN, C. J.M., 1971.Rapid bacteriological extr-
action from materials containing metallic sulphides,
u. s. Patent 3,607,235.
DUNCAN, D. W. and TEATI-IER, C. J., 1966. Method. of extracting metals
from sulphide ores using bacteria and an accelerating agent.
u. s. Patent 3,266,889.
DUNCAN, D. W. and TRUSSELL, P. C., 1964. Advances in microbiological
leaching of sulfide ores. Canadian Metallurgical Quarterly 3:
43-55.
DUNCAN, D. W. and WALDEN, C.C., 1972. Microbiological leaching in
the presence of ferric iron. Developments in Industrial Microb
iology 13: 66-75.
DUNCAN, D. W., TRUSSELL, P. C. and WALDEN, C.C., 1964. Leaching of
chalcopyrite with Thiobacillus ferrooxidans-Effects of surfacta
nts and shaking. Applied Microbiology 12: 122-126.
DUNCAN, D. W., WALDEN, C.C. and TRUSSELL, P. C., 1966. Biological
leaching of mill products. Canadian Mining and Metallurgical

241
Bulletin 59: 1075-1079.
DUNCAN, D. W., WALDEN, C. C. and TRUSSELL, P. C., 1967. Accelerated
microbiological ore extraction process. u. s. Patent 3,305,353.
DUNCAN, D. W., WALDEN, C. C., TRUSSELL, I'. C. and LOWE, E. A., 1967-a.
Recent advances in the microbiological leaching of sulfides.
Transactions of Society for Mining Engineers (AIME), 238: 122-128.
DUNCAN, D. W., TRUSSELL, P. C. and WALDEN, C. C., 1967-b. Role of
Thiobacillus ferrooxidans in the oxidation of sulfide minerals.
Canadian Journal of Microbiology 13: 397-403.
DUNN, N. W., RALPH, B. J. and RHODES, S. H., 1977. Further observa
tions on the FAME profiles of Thiobacillus species. GIAM v
Symposium, Bangkok, November, 1977. (To be published).
DITTRIZAC, J.E. and MacDONALD, R. J. C., 1974. Ferric ion as a
leaching medium. Minerals Science and Engineering 6: 59-100.
EBNER, H. G. and SCHWARTZ, W., 1974. Geomikrobiologische untersuch
ungen. XII- Verhalten von mikroorganismen auf uranhaltigen
gesteinen. Zeitschrift fur Allgemeine Mikrobiologie 14: 93-102.
EGOROVA, A. A. and DERYUGINA, A. P., 1963. The spore-forming thermo
philic thiobacterium, Thiobacillus thermophilic Imschenetskii.
Mikrobiologiya 32: 439-446.
EHRLICH, H. L. and FOX, S. I., 1967. Environmental effects on

242
bacterial copper extraction from low grade copper sulphide ores.
Biotechnology and Bioengineering 9: 471-485.
ELSDEN, S. R., 1962. Photosynthesis and lithotrophic carbon dioxide
fixation. In: Cunsalua, I. C. & Stanier, R. Y. (Eds.), The
Bacteria pp. 1-40. Academic Press, London.
FERREIRA, R. C. H. and BURKIN, A. R., 1975. Acid leaching of
chalcopyrite. In: Burkin, A. R. (Ed.), Leaching and Reduction
in Hydrometallurgy pp. 54-66. The Institute of Mining and Meta
llurgy, London.
FINDLEY, J., APPLEMAN, M. D. and YEN, T. F., 1974. Degradation of
oil-shale by sulfur-oxidizing bacteria. Applied Microbiology
28: 460-464.
FLETCHER, A. W., 1970. Metal winning from low grade ores by
bacterial leaching. Transactions of the Institute of Mining
and Metallurgy 79: C247-C252.
FLETCHER, A. W., 1971. Copper recovery from low grade ores by
bacterial leaching. In: Miller, J. D. A. (Ed.), Microbial
Aspects of Metallurgy pp. 183-194. MTP, Chiltern House
Aylesbury, England.
FLETCHER, A. W., 1971-a. Metal winning from low grade ores by
bacterial leaching. Discussion & contributions. Transactions
of the Institute of Mining and Metallurgy, BO: Cll4-Cll6.
FLOCH, J., ASCOL!, I., LEES, M., MEATH, J. A., and Le BARON, F. N.,
1951. Preparation of lipids extracts from brain tissue.
Journal of Biological Chemistry 191: 833-841.
FORRESTER, J. A., 1959. Destruction of concrete caused by sulfur

243
bacteria in purification plant. Surveyor 118: 881-884.
GALE, N. L. and BECK, J. V., 1966. Competitive inhibition of phospho-
ribulokinase by AMP. Biochemical and Biophysical Research
Communications 24:792-796.
GALE, N. L. and BECK, J. V., 1967. Evidence for the Calvin cycle and
hexose monophosphate pathw~y in ThiobaciZZus ferrooxidans. I
Journal of Bacteriology 94: 1052-1059.
GARRELS, R. S. M. and CHRIST, c.: L., 1965.Solutions, Minerals and
Equilibria. Harper and Ro~e. New York, Evanston and London.
GILCHRIST, F. M. C., 1953. Mic~obiological studies of the corrosion
'
of concrete sewers by sulfuric acid producing bacteria. South
African Industrial Chemistr:y 7: 214-215.
GOEHRING, M. and FELDMANN, U., 1!948. ,Nue Verfahren zur Darstellung van
Kaliumpentathionat und van Kaliumhexathionat. Zeitschrift fur
anorganische Chemie. 257: 223-226.
GOLDING, R. M., RAE, A. D., RAL~H, B. J. and SULLIGOI, L., 1974.
A new series of polynuclear copper Dithiocarbamate copper halide
polymers. Inorganic Chemis~ry 13: 2499-2504.
GOLDING, R. M., HARRIS, B., RAU~H, B. J., RICKARD, P.A. D. and
VANSELOW, D. G., 1977. The nature of the passivation film on
covellite exposed to oxygen. GBF Monograph Series No. 4,
191-200. Conference Bacterlial Leaching, 1977.
GOLOMIZK, A. I. and IVANOV, V. l., 1965. Izv. Vyssh. Ucheb. Zaned.

244
8: 33 [cited from Gupta & Sant, 1970].
GORMELY, L. S. and DUNCAN, D. W., 1974. Estimation of ThiobaciZZus
ferrooxidans concentrations. Canadian Journal of Microbiology
20: 1453-1455.
GORMELY, L. S., DUNCAN, D. W., BRANION, R. M. R. and PINDER, K. L.,
1975. Continuous culture of Thiobacillus ferrooxidans on a
zinc sulphide concentrate. Biotechnology and Bioengineering
17: 31-49.
GUAY, R. and SILVER, M., 1975. ThiobaciZZus acidophiZus sp nov.:
Isolation and some physiological chacteristics. Canadian
Journal of Microbiology 21: 281-288.
GUAY, R., SILVER, M. and TORMA, A. E. , 1975. DNA base composition
of some bacteria oxidizing ferrous iron and metal sulphides.
IRCS Medical Science: Biochemistry, Microbiology, Parasitology
and Infectious Diseases .3: 417.
GUAY, R., TORMA. A. E. and SILVER, M., 1975-a. Oxydation de l'ion
ferreux et mise en solution de !'uranium d'um rninerai par
ThiobaciZZus ferrooxidans. Annales Microbiologie (Institute
Pasteur) 126B: 209-216.
GUAY, R. SILVER, M. and TORMA, A. E., 1976. Base somposition of DNA
isolated from ThiobaciZZus ferrooxidans grown on different sub
strates. Revue Canadienne de Biologie 35: 61-67.

245
GUITTONNEAU, G., 1925. Sur la formation d'hyposulfites aux depens du
soufre par les microorganisms du sol. Comptes Rendus Hebbomadaires
des seances De L'Academie de Sciences. Paris. 180: 1142-1144.
GUPTA, R. C. and SANT, B. R.,1970. Beneficiationof low grade ores by
microbial leaching. Journal of Scientific and Industrial
Research (India) 29: 372-377.
HALL, W. E., 1961. Unit cell edges of cobalt and cobalt-iron bearing
sphalerites. u. s. Geological Survey Profess. Papers. No. 424-B
271-3. [ Cited from: Chemical Abstracts, 9541 i, 1962.]
HAPPOLD, F. C., JOHNSTONE, K. I., ROGERS, H.J. AND YOUATT, J.B.,
1954. The isolation and characteristics of an organism oxidizing
thiocyanate. Journal of General Microbiology 10: 261-266.
HAPPOLD. F. C. JONES, G. L. and PRATT, D. B., 1958. Utilization of
thiocyanate by ThiobaciZZus thiopa.rus and ThiobaciZZus thio
cyanoxidans. Nature 182: 266-267.
HAO. P. L. C., CHANG, W. S. and WANG, N. W., 1972. Microbiological
leaching of copper from low grade enargite. Proceedings of IV
International Fermentation Symposium, Fermentation Technology
Today 509-512.
HARDER, W. KUENEN, J. G. and MATIN, A., 1977. A review: Microbial
selection in continuous culture. Journal of Applied Bacteriology
43: 1-24.
HARTS, L. H., 1973. Mineral science and the future of metals.
Mining Engineering 25: (April issue) pp. 57-61.
HENDERSON, M. E. K. and DUFF, R. B., 1963. The release of metallic and
silicate ions from minerals, rocks and soils by fungal activity.
Journal of Soil Science 14: 236-246.
HILL, L. R., 1968. The determination of deoxyribunucleic acid base
composition and its application to bacterial taxonomy. In: Gibbs,
B. M. and Shamton, G. (Eds.), Identification Methods for

246
Microbiologists: Part B, pp. 177-186. Academic Press, New York.
HIRSCHMAN, S. Z., and FELSENFELD, D. G., 1966. Determination of DNA
composition and concentration by spectral analysis. Journal of
Molecular Biology 16: 347-358.
HOBAN, D. J. and LYRICS, R. M., 1975. Phosphoenolpyruvate carboxylase
of Thiobaeillus thioparus. I- General properties. Canadian
Journal of Biochemistry 53: 875-880.
HOFFMAN, L. E.and HENDRIX, J. L., 1976. Inhibition of Thiobacillus
ferrooxida:ns by soluble silver. Biotechnology and Bioengineering,
18: 1161-1167.
HOLT, S. C., SHIVELY, J.M. and GREENAWALT, J. W., 1974. Fine struct
ure of selected species of the genus Thiobacillus as revealed by
chemical fixation and freeze-etching. Canadian Journal of
Microbiology 20: 1347-1351.
HOWARD, A. and LUNDGREN, D. G., 1970. Inorganic phosphatase from
Ferrobacillus ferrooxidans. Canadian Journal of Microbiology
7: 381-396.
HOWDEN, R. L., LEES, H. and SUZUKI, I., 1972. Phosphoenolpyruvate
carboxylase of Thiobacillus thiooxida:ns. Kinetics and metabolic
control properties. Canadian Journal of Biochemistry 50: 158-165.
HUTCHINSON, M., JOHNSTONE, K. I. and WHITE, D., 1965. The taxonomy
of certain Thiobaeilli. Journal of General Microbiology, 41:
357-366.
HUTCHINSON, M., JOHNSTONE, K. I. and WHITE, D., 1966. Taxonomy of
acidophilic Thiobacilli. Journal of General Microbiology
44: 373-381.
HUTCHINSON, M., JOHNSTONE, K. I. and WHITE, D., 1967. Taxonomy of
anaerobic Thiobacilli. Journal of General Microbiology 47:17-23.

247
IMAI, K., 1971. Bacterial leaching in the mining industry. In:
Sakaguchi, K., Uemura, T. & Kinoshita, S.(Eds.), Biochemical and
Industrial Aspects of Fermentation- Recent Contributions From
Japa.n, pp. 329-336. Kodaush- Tokyo.
IMAI, K., SUGIO, T., TSUCHIDA, T. ans TANO, T., 1975. Effect of
heavy metal ions on the growth and iron-oxidizing activity of
ThiobaciZZus ferrooxidans. Agricultural and Biological Chem
istry 39: 1349-1354.
IIDA, T., KOZU, H. and USAMI, S., 1975. Rust removal by chemoautotr
ophic bacteria. Journal of Fermentation Technology 53: 347-353.
ILYALETDINOV, A. N. and KANATCHINOVA, M. K., 1964. Microbiological
transformation of sulfur compounds in periodically flooded soils
of Kzyl-Orda region. Mikrobiologiya 33: 118-125.
ITZKOVITCH, I. J. and TORMA, A. E., 1976. Influence of organic copper
extractants in activity of ThiobaciZZus ferrooxidans. IRCS Medi
cal Sciences: Biochemistry, Environmental Biology, Medical Micro
biology, Parasitology and Infectious Diseases 4: 155.
ITZKOVITCH, I. J., ETTEL, V. A. and GEUDRON, A. S., 1974. Copper
removal from Thompson nickel anolyte by solvent extraction.
CIM Bulletin 67: 92-96.
IVANOV, M. V., 1957. Role of microorganisms in depositing sulfur in
hydrogen sulphide springs of Sergleo Mineral Waters.
Mikrobiologiya 26: 338-345.
IVANOV, V. I., 1962. Role of sulphur bacteria in the leaching of
sulphide ores. Dokl. Akad. Nauk. SSr., 146: 447-449.
[Cited from Dutrizac & MacDonald, 1974.]
IVANOV, V. I., 1962-a. Iron oxidation by ThiobaciZZus ferrooxidans.
Mikrobiologiya 31: 795-799.

248
IVANOV, V. I., 1962b. Effect of some factors on iron oxidation by
cultures of Thiobacillus ferrooxidans. Mikrobiologiya 31: 645-648.
IVANOV, V. I. and LIALIKOVA, N. N., 1962. On the taxonomy of iron
oxidizing Thiobacilli. Mikrobiologiya 31: 468-469.
IVANOV, V. I. and NAGIRNYAK, F. I., 1962.Tsvet. Metally. Mosk. 35: 30.
[Cited from Gupta and Sant, 1970].
IVANOV, V. I., NAGIRNOYAK, F. I. and STEPANOV, B. A., 1961., Bacterial
oxidation of sulphide ore. I- The role of Thiobacillus ferrooxi
dans in the oxidation ofchalcopyrite and sphalerite. Mikrobiolo
giya, 30: 688-692.
IWATSUKA, H., 1962. Studies on the metabolism of a sulfur oxidising
bacterium. III- Postoxidative fixation of carbon dioxide.
Plant and Cell Physiology. 3: 167-173.
IWATSUKA, H., KUNO, M. & MARUYAMA, M., 1962. Studies in the metabol
ism of a sulfur oxidizing Bacterium. II- The system of carbon
dioxide fixation.in, T-hiobacillus. thiooxidans. Plant and
Cell Physiology 3; 157-166.
JACKSON, J. F., MORIARTY, D. J. W. and NICHOLAS, D. J. D., 1968.,
Deoxyribonucleic acid base composition and taxonomy of Thiobacilli
and some mitrifying bacteria. Journal of General Microbiology,
53: 53-60.
JELLINEK, F., 1968.,Sulphides. In: Nickless, G., (Ed.)Inorganic Sulp
hur Chemistry pp. 669-747. Elsevier Publishing Company,
Amsterdam, London, New York.

249
JENNINGS, P.H., TARASSOFF, P. and FORD, R. E., 1972. Heap tests on
Gaspe copper oxide ore. Paper presented at 101st Annual
Meeting of the American Institute of Mining Engineering,
San Francisco, 1972.
JOHNSON, E. J., 1966. Occurrence of the adenosine monophosphate
inhibition of carbon dioxide fixation in photosynthtic and chem
osynthetic autotrophs. Archives of Biochemistry and Biophysics
114: 178-183.
JOHNSON, M. J., BORKOWSKI, J. and ENGBLOM, C., 1964. Steam sterli
sable probes for dissolved oxygen measurements. Biotechnology
and Bioengineering 6: 575-467.
JONES, G. L. and CARRINGTON, E, G,, 1972. Growth of pure and mixed
cultures of microorganisms concerned in the treatment of
carbonization waste liquors. Journal of Applied Bacteriology
35: 395-404.
KAPLAN, L.B., 1956. Evidence of microbiological activity in some
of the geothermal regions of New Zealand.
nal of Science & Technology 37B: 639-662.
New Zealand Jour-
KARAVAIKO, G. I., 1959. Biological influence in oxidation of sulfur
compounds of Rozdolski Deposits. Mikrobiologiya 28: 846-849.
KARAVAIKO, G. I. and AVAKYAN, A. A., 1970. The mechanism of Thiobaci
llus ferrooxidans multiplication. Mikrobiologiya 39: 950-952.
KARAVAIKO, G. I. and AVAKYAN, A. A., 1971. Submicroscopic organization
of Thiobacillus thiooxidans. Mikrobiologiya 40: 278-283.
KARAVAIKO, G. I., IVANOV, .M. V. and SREBRODOLSKII, B. I., 1962.
Oxidation of stored sulfur ores. Soviet Geology 5: 133-139.

250
KARAVAIKO, G. I. and PIVOVAROA, T. A., 1973. Oxidation of elementary
sulfur by ThiobaeiZZus thiooxidans. Mikrobiologiya 42: 345-350.
KELLY, D. P., 1967. Incorporation of acetate by the chemoautotroph
ThiobaeiZZus neapolitanus strain C. Archives fur Mikrobiologie
58: 99-101.
KELLY, D. P., 1971. Autotrophy: Concepts of lithotrophic bacteria and
thie organic metabolis. Annual Review of Microbiology 25:177-210.
KELLY, D. P. and TUOVINEN, 0. H., 1972. Recommendation that the
mames FerrobaeiZZus ferrooxidans, Leathen & Braley and Ferroba
eiUus sulfooxidans, Kinsel, be recognized as synonyms of Thio
baeiZZus ferrooxidans, Temple and Colmer. International Journal of
Systematic Bacteriology 22: 170-172.
KHALID, A. M. and RALPH, B. J., 1976.Unpublished information on the
microbial ecology of the Rum Jungle Mine area, Northern Territory,
Australia. (Australian Atomic Energy Commission Research Contract
74/F/40).
KHALID, A. M. and RALPH, B. J., 1977. The leaching behaviour of
various zinc sulphide minerals with three ThiobaeiZZus species.
GBF Monograph Series No. 4 ( August, 1977). Conference Bacterial
Leaching pp. 165-173.
KINSEL, N. A., 1960. New sulphur oxidizing iron bacterium, Ferro
baei Z Zus suZ f ooxidans Sp . Nov. Journal of Bacteriology 80: ::- ·,'
628-632.
KNAYSI, G., 1943. A cytobiological and microbiological study of
ThiobaeiZZus thiooxidans. Journal of Bacteriology 46: 451-461.
KOCUR, M., MARTINEC, T. and MAZANCE, K., 1968. Fine structure of
ThiobaeiZZus noveZZus. Journal of General Microbiology 52:
343-345.

251
KOROBUSHKINA, E. D., CHERNIAK, A. S. and MINEEV, G. G., 1974. Dissolut
ion of gold by microorganisms and their metabolism. Mikrobiologiya
43: 49-54.
KOROSTELEV, P. G. and CHAINIKOVA, N. A., 1966. Possible use of
FeS-ZnS systems as geological thermometers. Vop. Geol. Sev.
Zap. Sekt., Tikhookean, poyasa, Valadivostok. 185-187.
[Cited from: Chemical Abstracts 67: 34748 y, 1967.]
KUENEN, J. G. and VELDKAMP, H., 1973. Effects of organic compounds on
growth of chemostat cultures of Thiomicrospira pelophila,
Thiobacillus thioparus and Thiobacillus neapolitanus.
Archives fur Mikrobiologie 94: 173-190.
KULLERUD, G. and NEUMANN, H., 1953. The temperature of granitization
in the Rendalsvik area, northern Norway. Norsk. Geol. Tidsskr.
32: 148-155. [Cited from: Chemical Abstracts 4377d, 1954.]
LANDESMAN, J., DUNCAN, D. W. and WALDEN, C.C., 1966. Iron oxidation by
washed cell suspensions of the chemoautotroph Thiobacillus ferro
oxidans. Canadian Journal of Microbiology 12: 25-33.
LANDESMAN, J., DUNCAN, D. W. and WALDEN, C. C., 1966-a. Oxidation of
inorganic sulphur compounds by washed cell suspensions of Thio
bacillus ferrooxidans. Canadian Journal of Microbiology 12:957-964.
LAPTEVA, A. M., KRIUCHKOV, V. A. and GOLOMZIK, A. I., 1971. Application
of gel plates impregnated with the medium 9K for quantitative
control and isolation of Thiobacillus ferrooxidans. Mikrobiologiya
40: 572-574.
LAUWERS, A. M. and HEINEN, W., 1974. Bio-degradation and utilization of
silica quartz. Archives fur Mikrobiologie 95: 67-78.

252
LAZAROFF, N., 1963. Sulfate requirement for iron oxidation by Thio
baaiZZus ferrooxida:ns. Journal of Bacteriology 85: 78-83.
LAZAROFF, N., 1975. Specificity of the anionic requirement for iron
oxidation by cells of ThiobaaiZZus ferrooxidans. Abstracts Annual
Meeting of American Society for Microbiology, I 119.
LAZAROFF, N., 1977. Specificity of anionic requirement for iron
oxidation by ThiobaaiZZus ferrooxida:ns. Journal of General
Microbiology 101: 85-91.
LEATHEN, W.W. and BRALEY, S. A., 1954. A new iron-oxidizing bacterium:
·FerrobaaiZZus ferrooxida:ns. Bacteriological Proceedings
G64, p 44.
LEATHEN, W.W., BRALEY, S. A. and MCINTYRE, L. D., 1953. The role of
bacteria in the formation of acid from certain sulfuric constitu
ents associated with bituminous coal. II- Ferrous irom oxidizing
bacteria. Applied Microbiology 1: 65-68.
LEATHEN, W.W. and MADISON, K. M., 1949. The oxidation of ferrous
iron by bacteria found in acid mine water.. Bactei:iological
Proceedings. p.64.
LEATHEN, W. W, KINSEL, N. A. and BRALEY, S. A. 1956. FerrobaaiZZus
ferrooxidans:A chemosynthetic, autotrophic bacterium. Journal of
Bacteriology 72: 700-704.
LEATHEN, W.W., MCINTYRE, L.D.and BRALEY, S. A., 1951. A medium for the
study of the bacterial oxidation of ferrous iron. Science 114:
280-281.
LECHEVALIER, M. P., 1977. Lipids in bacterial taxonomy- A taxonomist's
view. CRC Critical Reviews in Microbiology, 5: 109-210.
LEHNINGER, A. L., 1970., Biochemistry. Worth publishers, Inc.,

253
70 Fifth Avenues, New York, pp. 832.
Le ROUX, N. W., 1966.
Annual Report, Division of Plant Industry, Commonwealth Scientific
and Industrial Research Organization, Australia. pp.147-148.
Le ROUX, N. W., Mineral attack by microbiological processes.
In: Miller, J. D. A. (Ed.), Microbial Aspects of Metallurgy
pp. 173-182. MTP, Chiltern House, Aylesbyry, England.
Le ROUX, N. W., NORTI-1, A. A. and WILSON, J.C., 1973. Bacterial
oxidation of pyrite. Proceedings of the 10th International
Mineral Processing Congress, London, 2-14 April, Paper No. 10.
Institute of Mining and Metallurgy, London.
Le ROUX, N. W., WAKERLEY, D.S. and HUNT, S. D., 1977. Thermophilic
Thiobacillus-type bacteria from Iceland thermal areas.
Journal of General Microbiology 100: 197-201.
LEVIN, R. A., 1971. Fatty acids of Thiobacillus thiooxidans.
Journal of Bacteriology 108: 992-995.
LEVIN, R. A., 1972. Effect of cultural conditions on the fatty acid
composition of Thiobacillus novellus.
Journal of Bacteriology 12: 903-909.
LIESKE, R., 1912. Untersichungen uber die physiologie denitrifiziere
nder Schwefel bacterien. Ber. deut. bot. Gesell, 30: 12-22.
[ Cited from: Baalsrud & Baalsrud, 1954.]
LIPMAN, C. B. and McLESS, E., 1940.
Soil Science 40: 429.
[Cited from Roy & Trudinger, 1970. ]
LITCHFIELD, J. H., 1976. Food Microbiology. In: Miller, B.M.
and Litsky, W. (Eds.), Industrial Microbiology. pp. 257-308.

254
McGraw-Hill Book Company, 1976.
LONDON, J., 1963. ThiobaciZZus noveZZus nov. sp - A novel type of
facultative autotroph. Archives fur Mikrobiologie 46: 329-337.
LONDON, J. and RITTENBERG, S. C., 1966. Effects of organic matter on
the growth of ThiobaciZZus intermedius. Journal of Bacteriology
91: 1062-1069.
LONDON, J. and RITTENBERG, S. C., 1967. Thiobacillus perometabolis,
nov.sp. a non-autotrophic ThiobaciZZus. Archives fur Mikro-
biologie 59: 218-225.
LUNDGREN, D. G., 1975. Microbiological problems in strip mine
areas: Relationship to the metabolism of ThiobaciZZus ferro
oxidans. Ohio Journal of Science 75: 280-287.
LUNDGREN, D. G., ANDERSEN, K. J., REMSEN, C. C. and MAHONEY, R. P.,
1964. Cutlture. structure and physiology of the chemoautotroph
FerrobaciZZus ferrooxidans. Developments in Industrial
Microbiology 6: 250-259.
LYALIKOVA, N. N., 1959. Fiziologiya i Ekologiya ThiobaciZZus
ferrooxidans v Svyazi s Yego Rol'yu v Okinlenii Sul'fidnykh
Rud. [ The physiology and ecology of ThiobaciZZus ferrooxidans
in relation to its role in the oxidation of sulfide ores].
Diss. Bibl-ka Otd. Biol. Nauk AN SSSR.
[ Cited from: Kuznetsov, S. I., Ivanov, M. V., and Lyalikova,
N. N. (Eds.), Introduction to Geological Microbiology, pp. 252.
McGraw-Hill Book Company, 1963.]
LYALIKOVA, N. N., 1960. The part played by ThiobaciZZus ferrooxidans
in oxidizing the sulfide ores in the copper pyrite deposits of
the Middle Urals. Mikrobiologiya 29: 773.

255
LYALIKOVA, N. N., 1961. The role of bacteria in the oxidation of the
sulphide ores in the copper-nickel deposits of the Kola Pensisula.
Mikrobiologiya 30: 135-139.
LYALIKOVA, N. N. and MOKEICHEVA, L. Ya., 1969. The role of bacteria in
gold migration in deposits. Mikrobiologiya 38: 805-810.
MacDONALD, D. G., and CLARK, R.H., 1970.The oxidation of aqueous ferr
ous sulpate by Thiobaaillus ferrooxidan. Canadian Journal of
Chemical Engineering 48: 669-676.
MacELROY, R. D., JOHNSON, E. J. and JOHNSON, M. K., 1968. Characteri
zation of ribulose diphosphate carboxylase and phosphoribulo
kinase from Thiobaaillus thioparus & Thiobaaillus neapolitanus.
Archives of Biochemistry and Biophysics 127: 310-316.
MlcELROY, R. D., JOHNSON, E. J. and JOHNSON, M.K., 1968-a. Allosteric
regulation of phosphoribulokinase activity. Biochemical and
Biophysical Research Communications 30: 678-682.
MACKINSTOSH, M. E., 1971. Nitrogen fixation by Thiobaaillus ferroo
xidans. Journal of General Microbiology 66: i-ii.
MACKINTOSH, M. E., 1976. Fixation of 15N2 by Thiobaaillus
ferrooxidans. Proceedings of the Society of General Microbiology
4 (Part 1) 23.
MAGNEM, de MM. R., BERTHELIN, J. and DOMMERGIJES, Y., 1973. Solubilis
ation de l'uranium dans les roches, par des bacteries n'appatenant
pas au genne Thiobaaillus. Comptes Rendus Hebdomadaires des Seances
de l'Academie des Sciences de Paris t 276 (9 mai) series D-
2625-2628.

256
MAHONEY, R. R. and EDWARDS, M. R., 1966. Fine structure of Thiobaeillus
thiooxidans. Journal of Bacteriology 42: 487-495.
MAJIMA, H.and PETERS, E., 1968. Electrochemistry of sulfide dissolution
in hydrometallurgical systems. VIII International Mineral Process
ing Congress, Leningrad.
MAJUMDAR, A. J., and ROY, R., 1965. Pressure-temperature dependence of
the sphalerite and wurtzite inversion in ZnS. American Mineralo
gist 50: 1121-1125.
MALOUF, E. E. and PRATER, J. D., 1961. The role of bacteria in the
alteration of sulphide minerals. Journal of Metals 13: 353-356.
MANNING, H. L., 1975. New medium for isolating iron-oxidizing and
heterotrophic acidophilic bacteria from acid mine drainage.
Applied Microbiology 30: 1010-1016.
MARCHLEWITZ, B. and SCHWARTZ, W., 1961. Investigations of microbial
association of acid mine waters. Zeitschrift fur Allgemein
Mikrobiologie 1: 100-114.
MARGALITH. P., SILVER, M. and LUNDGREN, D. G., 1966.Sulfur oxidation
by the iron-bacterium Ferrobaeillus ferrooxida:ns. Journal of
Bacteriology 92: 1706-1709.
MARMUR, J., 1961. A procedure for the isolation of deoxyribonucleic
acid from microorganisms. Journal of Molecular Biology 3:
208-218.
MARSHALL, K. C., 1975. Clay mineralogy in relation to survival of
soil bacteria. Annual Review of Phytopathology 13: 357-373.
MATIN, A. and RITTENBERG, S. C., 1970. Utilization of glucose in
heterotrophic media by Thiobacillus intermedius. Journal of
Bacteriology 104: 234-238.

257
MATIN, A. RITTENBERG, S. C., 1971. Enzymes of carbohydrate metabolism
in ThiobaciZZus species. Journal of Bacteriology 107: 179-186.
MAXWELL, W. A., METZLER, R. and SPOERL, E., 1971. Uranyl nitrate
inhibition in Saccharomyces cerevisiae. Journal of Bacteriology
105: 1205-1206.
MAYEUX, J. V. and JOHNSON, E.J., 1967. Effect of AMP, ADP and
reduced NAO on ATP-dependent carbon dioxide fixation in the
the autotroph ThiobaciZZus neapoZitanus. Journal of Bacteriol
ogy 94: 409-414.
MAYLING, A. A., 1966. Cyclic~ leaching process employing iron
oxidizing bacteria. Canadian Patent 744, 701.
MAYLING, A. A., 1969. Bacterial leaching of basic metal sulfide
ores. u.s.Patent 3,455,679.
McCARTHY, J. T. and CHARLES, A. M., 1973. Purification and purine nuc
leotide regulation of ribulose-1,5, diphosphate carboxylase from
ThiobaciZZus noveZZus. FEBS Letters 37: 329-332.
McCARTHY, J. T., and CHARLES, A. M., 1974. Co2 fixation by the
facultative autotroph ThiobaciZZus noveZZus during autotrophy
heterotrophy interconversions. Canadian Journal of Microbiology
20: 1577-1587.
McFARLANE, A. S., 1942. Behaviour of lipoids in human serum.
Nature 149: p. 439.
McGORAN, C. J.M., DUNCAN, D. W. and WALDEN,C.C.,1969. Growth of Thio
baciZZus ferrooxidans on various substrates. Canadian Journal
of Microbiology 15: 135-138.
MEHTA, K. B. and Le ROUX, N. W., 1974. Effect of wall growth on conti
nuous biological oxidation of ferrous iron. Biotechnology and

258
Bioengineering 16: 559-563.
MEYER, C. W. and YEN, T. F., 1976. Enhanced dissolution of oil shale
by bioleaching with Thiobacilli. Applied and Environmental
Microbiology 32: 610-616.
MILLER, B. M. and LITSKY, W. (Eds.). 1976. Industrial Microbiology
MgGraw-Hill Book Company, Sydney. 465 pp.
MIZOGUCHI, T., SATO, T. and OKABE, T., 1976. New sulfur-oxidizing
bacteria capable of growing heterotrophically, Thiobacillus
rubellus nov sp & Thiobacillus delicatus nov. sp. Journal
of Fermentation Technology 54: 181-191.
MOSS, F. J. and ANDERSEN, J.E., 1968. The effect of environment on
bacterial leaching rates. Proceedings of the Australian Institute
of Mining and Metallurgy 225: 15-25.
MURPHY, J. R., GIRARD, A. E. and TILTON, R. C., 1974. Ultrastructure
of a marine Thiobacillus. Journal of General Microbiology 85:
130-138.
MURR, L. E. and BERRY, V. K., 1976. Direct observations of selective
attachment of bacteria on low-grade sulfide ores and other mineral
surfaces. Hydrometallurgy 2: 11-24.
MURRAY, R. G. E., 1963. The organelles of bacteria. In: Mazia, D. &
Tyler, A, (Eds.), General Physiology of Cell Specialization pp 28-
52. MgGraw-Hill Book Company, Inc. N.Y.
MURRAY, R. G. E. and WATSON, S. W., 1965. Structure of Nitrocystis
ocea:nus and comparison with Nitrosomonas & Nitrobacter.
Journal of Bacteriology 89: 1594-1609.
MUTZE, B. and ENGEL, H. 1960. Bacterial oxidation of sulfur in the
river Elbe. Archives fur Mikrobiologie 35: 303:309.

259
NATHANSOHN, A., 1902. Uber eine neu gruppe von schwefelbakterien und
ihren stoffwechsel. Mitt. z.ool. stn. Neapol. 15: 655-680.
NICKERSON, W. J., 1969. Microbial transformations of naturally
occurring polymers. In: Perlman, D. (Ed.), Fermentation Advances,
631-634.
NIELSEN, A. M. and BECK, J. V., 1972., Chalcocite oxidation and
coupled carbon dioxide fixation by ThiobaciZZus ferrooxida:ns.
Science 175: 1124-1126.
NIEMELA, S. I. and TUOVINEN, O.H., 1972. Acidophillic ThiobaciZZi in
river Sirppujoki. Journal of General Microbiology 73: 23-28.
NORDYKE, M. D., 1971. Other unique applications of PNE(Peaceful
Nuclear Explosions). In: Peaceful Nuclear Explosions II- The
Practical Applications pp 223-232. International Atomic
Energy Agency, Viena.
NOVICK, A., and SZILARD, L., 1950. Description of the chemostat.
Science 112: 715-716.
NOVICK, A., and SZILARD, L., 1950-a. Experiments with the chemostat on
spontaneous mutation of bacteria. Proceedings of the National
Academy of Sciences (Washington) 36: 708-719.
O'LEARY, W. M., 1962. The fatty acids of bacteria. Bacteriological
Reviews 26: 421-447.
O'LEARY, W. M., 1970. Bacterial lipid metabolism. In:Florkins, M. &
Stotz, E. H. (Eds.), Comprehensive Biochemistry 18: 229-264.
Elsevier Publishing Co. Amsterdam.
OTTOW, J.C. and VON KLOPOTEK, A., 1969. Enzymatic reduction of
iron oxide by fungi. Applied Microbiology 18: 41-43.

260
PARES, Y., 1964. Intervention des bacteries dans la solubilisation
du cuivre. Annales de L'Institut Pasteur 107: 132-135.
PARES, Y., 1964-a. Action de Serr'atia macescens dans le cycle biologi
que des m'etaux. Annales de L'Institut Pasteur 107: 136-141.
PARES, Y. 1964-b. Action d'Agrobacteriwn twnefaciens dans la mise
en solution de l'or. Annales de L'Institut Pasteur 107:141-143.
PARKER, C. D., 1945. The corrosion of concrete. I- The isolation of a
species of bacterium associated with the corrosion of concrete
exposed to atmospheres containg H2S. Australian Journal of
Experimental Biology and Medical Science 23: 81-90.
PARKER, C. D. and PRISK, J., 1953. The oxidation of inorganic compounds
of sulphur by various sulphur bacteria. Journal of General
Microbiology 8: 344-364.
PAWLEK, F. E., 1969. Research in pressure leaching. Journal of
_South African Institute of Mining and Me.tallurgy 69: 632-:-654.
PECK, H. D. Jr., 1960. Adenosine-5'-phosposulfate as an intermediate
in the oxidation of thiosulphate by Thiobacillus thioparus.
Proceedings of the National Academy of Sciences, U.S.A., 46:
1053-1057.
PECK, H. D. Jr., DEACON, T. E. and DAVIDSON, J. T., 1965. Studies on
APS-reductase from Desulphovibrio desulphuricans& Thiobacillus
thioparus. I- The assay and purification. Biochimica et Biophysica
Acta 96: 429-446.
PECK, H. D. Jr. and FISHER, E.Jr. 1961. The oxidation of thiosulfate
and phosphorylation in extracts of Thiobacillus thioparus.
Journal of Biological Chemistry 237: 190-197.
PEETERS, T. & ALEEM, M. I. H., 1970. Oxidation of sulfur compounds

261
and electron transport in ThiobaciZZus denitrificans. Archives
fur Mikrobiologie 71: 319-330.
PFENNIG, N., 1961. Eine vollsynthetische nahrlosung zur sele ktiven
a neicher ung einiger schwefelpur purbaketrien. Die Natur
wissenschaften 48: 136.
PIVORAROVA, T. A., and KARAVAIKO, G. I., 1973. Submicroscopic organi
zation and function of some structures in ThiobaciZZus neapoZitanus.
Mikrobiologiya 42: 672-676.
PIVORAROVA, T. A. and KARAVAIKO, G. I., 1973-a. Submicroscopic organi
zation of ThiobaciZZus thiocyanoxidans. Mikrobiologiya
42: 1078 - 1080.
POSTGATE, J. R., 1966. Media for sulphur bacteria.
Laboratory Practice 15: 1239-1244.
POURBAIX, M. J. N., 1966. (Ed.) Atlas of Electrochemical Equlibria
in Aqueous Solutions. Pergammon Press, Oxford.
PRATT, V., 1972. Numerical Taxonomy- A critique. Journal of Theoretical
Biology 36: 581-592.
PUDOVKINA, I. A., RYABEVA, E. G., AKSENOVA, E. K. and DUBAKINA, L.S.,
1968. Effect of chemical composition on some physical properties
of sphalerite ores. Miner. Syr'e. No. 18, 13-28.
[Cited from: Chemical Abstracts 134999 c: 72: 1970]
PUROHIT, K., McFADEN, B. A. and COHEN, A. L., 1976. Purification,
quaternary structure, composition and properties of D-ribulose-
1-5- bisphosphate carboxylase fron ThiobaciZZus intermedius.
Journal of Bacteriology 127: 505-515.
PUROHIT, K., M:FADEN, B. A. and SHAYKH, M. M., 1976-a. D-ribulose

262
1,5-bisphosphate carboxylase and polyhederal inclusions bodies
in Thiobacillus intePmedius. Journal of Bacteriology 127: 516-522.
RALPH, B. J., 1978. Oxidative processes. In: Biogeochemical Cycling
of Mineral-forming Elements. Elsevier Press. (In press).
RAO, G.. •. -~. _ 2nd BERGER, L. R., 1970. Basis of pyruvate inhibition
in Thiobacillus thiooxidans. Journal of Bacteriology 102: 462-466.
RAO, G. S. and BERGER, L. R., 1971. The requirement of low pH for growth
of Thiobacillus thiooxidans. Archive fur Mikrobiologie 79:338-344.
RAY, B., 1969. II-IV Compounds. Pergarnon Press, Oxford, 268 pp.
RAZZELL, W. E., 1962. Bacterial leaching of metallic sulphides.
Canadian Mining and Metallurgical Bulletin 55: 190-191.
RAZZELL, W. E. and TRUSSELL, P. C., 1963. Isolation and properties of
and iron-oxidizing Thiobacillus. Journal of Bacteriology 85:
595-603.
RAZZELL, W. E. and TRUSSELL, P. C., 1963-a. Microbiological leaching of
metallic sulphides. Applied Microbiology 11: 105-110.
REMSEN, C. C. and LUNDGREN, D. G., 1963. The heterotrophic growth of
the chemoautotroph Ferrobacillus ferrooxidans. Bacteriological
Proceedings 33.
REMSEN, C. C. and LUNDGREN, D. G., 1966. Electron microscopy of the
cell envelope of Ferrobacillus ferrooxidans prepared by freeze
etching and chemical fixation techniques. Journal of Bacteriology
92: 1765-1771.
RITTENBERG, S. C., 1969. The role of exogenous organic matter in the
physiology of chemolithtrophic bacteria. Advances in Microbial
Physiology 3: 159-196.
ROBERTS, W. M. B., 1965. Recrystallization and mobilization of

263
sulfides at 2,000 atmospheres and in the temperature range
50° - 145°. Economic Geology 60: 168-180.
ROGOFF, M. H., SILVERMAN, M. P. and WENDER, I., 1960. Elimination o,f, '.
sulfur from cpal by microbiological action. American Chemical
Society, Division of Gas & Fuel Chemistry 2: 25-36.
ROGOVSKAYA, T. I. and LAZAREVA, M. F., 1961.Intensification of bio
chemical purification of industrial sewage. III- Microbiologi
cal characteristics of the biofilm purifying H2S containing
sewage. Mikrobiologiya 30: 699-702.
ROSLYCKY, E. B., 1972. Reliable procedure for silica gel preparation.
Applied Microbiology 24: 844-845.
ROSSI, G., 1974., Microbial leaching of ores: Progress and prospects.
Bollettino della Associazione Mineraria subalpina. 11: 1-40.
ROTHSTEIN, A., 1954. Enzyme systems of the cell surface involved
in the uptake of sugars by yeasts. Symposium of Society for
Experimental Biology 8: 165-201.
ROY, A. B. and TRUDINGER, P.A., 1970. The Biochemistry of Inorganic
Compounds of Sulphur. Cambridge University Press, Cambridge. pp.400.
RUDOLFS, W., 1922. Oxidation of pyrite by microorganisms and their
use for making mineral phosphate available. Soil Science
14: 135-147.
RUDOLFS, W. and HELBRONNER, A., 1922. Oxidation of zinc sulphide
by microorganisms. Soil Science 14: 459-465.

264
SADLER, W.R. and TRUDINGER, P.A., 1967. The inhibition of microo
rganisms by heavy metals. Mineralium Deposita 2: 158-168.
SANTER, M. and VISHNIAC, W., 1955. co2 incorporation by extracts of
ThiobaaiZZus thioparus. Biochimica et Biophysica 18: 157-158.
SANTER, M., BOYER, J. and SANTER, U., 1959. ThiobaaiZZus noveZZus .
I- Growth on inorganic and organic media. Journal of Bacteriology
78: 197-202.
SANZONOV, V. D., 1961. Determination of temperature of sphalerite
from its iron content. Tr. Inst. Geol. Akad. Nauk. Tadzh. ssr.
4: 215-233. [ Cited from: Chemical Abstracts 1241h, 1963]
SARGEANT, K., BUCK, P. W., FORD, J. W. S. and YEO, R. G., 1966.
Anaerobic production of ThiobaaiZZus denitPifiaans for the
enzyme rhodanese. Applied Microbiology 14: 998-1003.
SCHAEFFER, W. I. and UMBREIT, W.W., 1963. Phosphotidylinositol as a
wetting agent in sulfur oxidation by ThiobaaiZZus thiooxidans.
Journal of Bacteriology 85: 492-493.
SCHAEFFER, W. I., HOLBERT, P. E. and UMBREIT, W.W., 1963. Attachment
of Thiobacillus thiooxidans to sulfur crystals. Journal of
Bacteriology 85: 137-140.
SCHLEGEL, H. G., 1975. Mechanism of chemo-autotrophy. In: Kinne, 0.
(Ed.), Marine Ecology, Volume II, Part I, pp 9-60. John Wiley
& Sons, New York.
SCHNAITMAN, C. A., KORCZYNYSKI, M. S. and LUNDGREN, 1969. Kinetic
studies of iron oxidation by whole cells of FePPobaaiZZus
fePPooxidans. Journal of Bacteriology 99: 552-557.
SCHRAUZER, G. N. and GUTH, T. D., 1976. Hydrogen evolving systems.
I- The formation of hydrogen from aqueous suspensions of Fe(OH) 2

265
and reactions with reducible substrates, including molecular
nitrogen. Journal of the American Chemical Society 98: 3508-3513.
SCHWARTZ, A. and SCHWARTZ, W., 1965. Geomikrobiologische Untersuchungen.
VII- Uber das Vorkommen von Mikroorganismen in Solfataren und
heiben Quellen. Zeitschrift fur Allgemeine Mikrobiologie 5:395-405.
SCOTT, T. R. and DYSON, N. F., 1968. The catalyzed oxidation of zinc
sulphide under acid pressure leaching conditions. Transactions of
the Metallurgical Society of AIME 242: 1815-1821.
SHAFIA, F. and WILKINSON, R. F., 1969. Growth of Ferrobacillus ferro
oxidans on organic matter. Journal of Bacteriology 97: 256-260.
SHIVELY, J.M., 1974. Inclusion bodies of prokaryotes. Annual Review
of Microbiology 28: 167-187.
SHIVELY, J.M., DECKER, G. L. and GREENWALT, J. W., 1970. Comparative
ultrastructure of Thiobacilli. Journal of Bacteriology 101:618-627.
SHIVELY, J.M., BALL, F. L., BROWN, D. H. and SAUNDERS, R. F.,1973.
Functional organelles in prokaryotes: Polyhederal inclusions
(carboxysomes) of Thiobacillus neapolitanus. Science 182:584-586.
SHIVELY, J.M., BALL, F. L. and KLINE, B. W., 1973-a. Electron
microscopy of carboxysomes (polyhederal bodies) of Thiobacillus
neapolitanus. Journal of Bacteriology 116:1405-1411.
SHUEY, R. T., 1975. Sphalerite-ZnS. In: Semiconducting Ore Minerals
319-327. Elsevier Scientific Publishing Company. Amsterdam,Oxford.
SHULER, M. L. and TSUCHIYA, H. M., 1975. Determination of cell numbers
and size for Thiobacillus ferrooxidans by electrical resistance
method. Biotechnology and Bioengineering 17: 621-624.
SILVER, M., 1970. Oxidation of elemental sulfur and sulfur compounds
and co2 fixation by Ferrobacillus ferrooxidans. Canadian Journal
of Microbiology 16: 845-849.

266
SILVER, M. 1977. Metabolic mechanisms of iron-oxidizing Thiobacilli.
In: Murr, L. E., Torma, A. E. & Brierley, J. A. (Eds.) Metallurgi
cal Applications of Bacterial Leaching and Related Microbiological
Phenonena. The Academic Press, N. Y. (In press).
SILVER, M. and TORMA, A. E., 1974. Oxidation of metal sulphides by
Thiobacillus ferrooxidans grown on different substrates.
Canadian Journal of Microbiology 20: 141-147.
SILVER, M. , MARGALITH, P. and LUNDGREN D. G., 1967. Effects of gluco
se on carbondioxide assimilation and substrate oxidation by
Ferrobacillus ferrooxidans. Journal of Bacteriology 93: 1765-1769.
SILVERMAN, M. P. and EHRLICH, H. L., 1964. Microbial formation and
degradation of minerals. Advances in Applied Microbiology 6:
153-206.
SILVERMAN, M. P. and LUNDGREN, D. G., 1959. Studies on the chemoauto
trophic bacterium Ferrobacillus ferrooxidans. I- An improved
medium and a harvesting procedure for screening high cell yields.
Journal of Bacteriology 77: 642-647.
SILVERMAN, M. P. and LUNDGREN, D. G., 1959-a. Studies on chemoautotro
phic iron bacterium Ferrobacillus ferrooxidans. II- Manometric
studies. Journal of Bacteriology 78: 326-331.
SILVERMAN, M. P. and MUNOZ, E. F., 1970. Fungal attack on rocks:
Solubilisation and altered infrared spectra. Science 169: 985-987.
SILVERMAN, M. P. and ROGOFF, M. H., 1961. Morphological variations in
Ferrobacillus ferrooxidans related to iron oxidation.
Nature 191: 1221-1222.
SILVERMAN, M. P., ROGOFF, M. H. and WENDER, I., 1961. Bacterial
oxidation of pyritic materials in coal. Applied Microbiology 9:
491-496.

267
SILVESTRI, L. G., HILL, L.R., 1965. Agreement between deoxyribonucleic
acid base composition and taxonometeric classification of Gram
positkve Cocci. Journal of Bacteriology 90: 136- 140.
SKERMAN, V. B. D., 1967. A Guide to the Identification of the
Genera of Bacteria. 2nd. Edition. The Williams & Wilkins Company,
Baltimore.
SKINNER, B. J., 1960. Unit-cell edges of natural and synthetic
sphalerites. American Mineralogist 46: 1399-1411.
SMITH, A. J. and HOARE, D.S., 1977. Specialist phototrophs, lithotro
phs and methylotrophs: a unity among a diversity of procaryotes?
Bacteriological Reviews 41: 419-448.
SMITH, F. G., 1955. Structure of zinc sulphide minerals. American
Mineralogist 40: 658-675.
SNEATH, P.H. A., 1957. The application of computer to taxonomy. Jour
nal of General Microbiology 17: 201-226.
SOKOLOVA, G. A., 1960. Microbiological production of sulfur from
sulfide water strata. Mikrobiologiya 29: 888- 893.
SOKOLOVA, G. A. and KARAVAIKO, G. I., 1962. Biogenic oxidation of
sulfur of the Rozdol ore under laboratory conditions.
Mikrobiologiya 31: 984-989.
SOKOLOVA, G. A. and KARAVAIKO, G. I., 1963. Biogenic oxidation of
sulfur in Rozdol ore in laboratory conditions.
Mikrobiologiya 31: 797-800.
SORBO, B., 1957. A colorimetric method for the determination of
thiosulphate. Biochimica et Biophysica Acta 23: 412-416.
STAMM, H. and GOEHRING, M., 1942. Zur kenntnis der polythionsauren und
ihren bildung. Neu verfahren zur darstellung von kaliumtrithionat
und von kallumtetrathionat. Zeitschrift fur anorganische und

268
allegmeine chemie 250: 226-228.
STARKEY, R. L., 1934. Cultivation of organisms concerned in the
oxidation of thiosulfate. Journal of Bacteriology 28: 365-386.
STARKEY, R. L., 1934-a. The production of polythionates fron thiosulfate
by microorganisms. Journal of Bacteriology 28: 387-400.
STARKEY, R. L., 1935. Isolation of some bacteria which oxidize
thiosufate. Soil Science 39: 197-219.
STARKEY, R. L., JONES, G. E. and FREDERICK, L. R., 1956. Effects of
medium agitation and wetting agents on oxidation of sulfur by
ThiobaciZZus thiooxidans. Journal of General Microbiology
15: 329-334.
STOTSKY, G. and REM, L.T., 1966. Influence of clay minerals on
microorganisms. I- Montmorillonite and kaolinite on bacteria.
Canadian Journal of Microbiology 12: 547-563.
SUMMERS, A. 0., CHOTTEL, J.,CLARK, D. and SILVERS, S., 1975. Plasmid-
borne Hg(II) and organomercurialresistance. In: Schlessinger,
D. (Ed.) Microbiology-1974 pp 219-226. The American Society for
Microbiology, Washington, D.C.
SUZUKI, I. and WERKMAN, C. H., 1957. Phosphoenolpyruvate carboxylase
in extracts of ThiobaciZZus thiooxidans, a chemoautotrophic
bacterium. Archives of Biochemistry and Biochemistry 72: 514-515.
SUZUKI, I., and WERKMAN, C. H., 1958. Chemoautotrophic fixation of co2
by ThiobaciZZus thiooxidans. Iowa State College Journal of
Science 32: 475-483.
SUZUKI, I. and WERKMAN, C. H. , 1958-b. Chemoautotrophic Co2 fixation
by extracts of ThiobaciZZus thiooxidans. Archives of Biochemistry
and Biophysics 77: 112-123.

269
TABITA, F. R. and LUNDGREN, D. G., 1970. Glucose metabolism in
FerrobaciZZus ferrooxidans. Bacteriological Proceedings Pl7, pl7.
TABITA, R. and LUNDGREN, D. G., 1971. Utilization of glucose and the
effect o f organic compounds on the chemoli thotroph ThiobaciUus
ferrooxidans. Journal of Bacteriology 108: 328-333.
TABITA, R. and LUNDGREN, r. G, 1971-a. Heterotrophic metabolism of the
chemolithotroph ThiobaciZZus ferrooxidans. Journal of Bacteriology
108: 334-342.
TABITA, R. and LUNDGREN, D. G., 1971-b. Glucose-6-phosphate dehydroge
nase from the chemolithotroph ThiobaciZZus ferrooxidans. Journal
of Bacteriology 108: 343-352.
TAKIMOTO, K., MINATO, T. and HIRONO, S., 1970. Relation between
the unit-cell dimensions and minor elements of sphalerite from the
Chichbu Mine, Sitama Prefacture, Japan. Suiyotaishi 14: 186-189.
[ Cited from: Chemical Abstracts 8060 g]
TANO, T. and IMAI, K., 1968. Physiological studies on ThiobaciZZi.
VI- Certain properties of NADPH-cytochrome C oxidoreductase of
ThiobaciZZus thiooxidans.
32: 401-404.
Agricultural and Biological Chemistry
TAYLOR, B. F. and HOARE, S. L. , 1969. Thiobacillus denitrificans
as an obligate chemolithotroph. Bacteriological Proceedings p63.
TAYLOR, B. F., HOARE, D.S. and HOARE, S. L., 1971 . ThiobaciZZus
denitrificans as an obligate chemolithotroph: Isolation and growth
studies. Archives fur Mikrobiologie 78: 193-204.
TAYLOR, J. H. and WHELAN, P. F., 1943. The leaching of cupreous pyrites
and the precipitation of copper at RioTinto, Spain. Transactions
of Institute of Mining and Metallurgy 52: 35-96.
TEMPLE, K. L. and COLMER, A., 1951. The autotrophic oxidation of iron

270
by a new bacterium, ThiobaciZZus ferrooxidans. Journal of
Bacteriology 62: 605-611.
TEMPLE, K. L. and DELCHAMPS, E.W., 1953. Autotrophic bacteria and the
formation of acid in bituminous coal mines. Applied Microbiology
1: 255-258.
TILTON, R. C., COBET, A. B. and JONES, G. E., 1967. Marine Thiobacilli.
I- Isolation and distribution. Canadian Journal of Microbiology
13: 1521-1528.
TILTON, R. C., COBET. A. B. and JONES, G. E., 1967-a. Marine
ThiobaciZZi. II-Culture and ultrastructure. Canadian Journal of
Microbiology 13: 1529-1534.
TOMINAGA, N. and MORI, T., 1974. A sulfate-dependent acid phosphatase
of ThiobaciZZus thiooxidans Its partial purification and some
properties. Journal of Biochemistry 76: 397-409.
TOMIZUKA, N. and TAKAHARA, Y., 1972. Bacterial leaching of uranium
from Ningyo-Toge ore. Proceedings of IV International Fermentation
Symposium- Fermentation Technology Today, pp 513-520.
TOMIZUKA, N, 1976. Microbiological oxidation of lead sulphide.
Report of the Fermentation Research Institute No. 48, pp 51-66.
TOMIZUKA, N., YAGISAWA, M., SOMEYA, J. and TAKAHARA, Y. 1976. Continuous
leaching of uranium by ThiobaciZZus ferrooxidans. Agricultural
and Biological Chemistry 40: 1019-1025.
TORMA, A. E., 1971. Microbiological oxidation of synthetic Co, Ni and
Zns by ThiobaciZZus ferrooxidans. Revue Canadienne de Biologie
30: 209-216.
TDRMA, A. E., 1972. Biohydrometallurgy of cobalt and nickel.
TMS-Paper section, paper No. A72-7.
TORMA, A. E., 1975. Microbiological extraction of cobalt and nickel

271
from sulphide ores and concentrates. Canadian Patent 960 463.
TORMA, A. E., 1976. Biodegradation of chalcopyrite. In: Shapley, J.M.,
Kaplan, A. M. (Eds.) Proceedings of 3rd International Bio
degradation Symposium pp. 937-946. Applied Science Publishers
Ltd., London.
TORMA, A. E., 1977. The role of ThiobaeiZZus ferrooxidans in
hydrornetallurgical processes. Advances in Biochemical Engineer
ing 6: 1-37.
TORMA, A. E. and GABRA, G. G., 1975. Effect of wetting agents on
chalcopyrite oxidation ability of ThiobaeiZZus ferrooxidans.
IRCS Medical Science, Biochemistry, Microbiology, Parasitology
and Infectious diseases 3: 228.
TORMA, A. E. and GABRA, G. G., 1976. Effects of surface active
agents on the oxidation of chalcopyrite by ThiobaeiZZus ferro
oxidans. Hydrometallurgy 1: 301-309.
TORMA, A. E. and GUAY, R., 1976. Effect of particle size on the
biodegradation of a sphalerite concnetrate. Le Naturaliste Canadien
103: 103: 133-138.
TORMA, A. E. and ITZKOVITCH, I. J., 1976. Influence of organic solvents
on chalcopyrite oxidation ability of ThiobaeiZZus ferrooxidans.
Applied and Environmental Microbiology 32: 102-107.
TORMA, A. E. and LEGAULT, G., 1971. Contribution to characterization of
a rnicroorganisrn- ThiobaeiZZus ferrooxidans. Paper presented at
54th Conference of Chemical Institute of Canada, Halifax, Nova
Scotia, May 30 - 2 June.
TORMA, A. E. and LEGAULT, G., 1973. Metal sulfide biodegradation by
ThiobaeiZZus ferrooxidans. Effect of their total surfaces.
Anneles Microbiologie l'Institut Pasteur 124A: 111-121.

272
TORMA, A. E., and SUBRAMANIAN, K. N., 1974. Selective bacterial leach
ing of lead sulphide concentrate. International Journal of
Mineral Processing 1: 125-134.
TORMA, A. E., WALDEN, C.C., and BRANION, R. M. R., 1970. Microbio
logical leaching of zinc sulphide concentrate. Bitechnology and
Bioengineering 12: 501-517.
TORMA, A. E., WALDEN, C. C., DUNCAN, D. W. and BRANION, R. M. R., 1972.
The effect of co2 and particle surface area on the microbiological
leaching of zinc sulphide concentrate. Biotechnology and Bioengi-
neering 14: 777-786.
TORMA, A. E., LEGAULT, G., KOUGIOMOUTZAKIS; D. and OUELLET, R., 1974.
Kinetics of biooxidation of metal sulphides. Canadian Journal of
Chemical Engineering 52: 515-517.
TRIBUTSCH, H. , 1976. The oxidative disintegration of sulfide
crystals by ThiobaciZZus ferrooxidans. Naturwissenschaften 63: 88.
TRIVEDI, N. C. and TSUCHIYA, H. M., 1975. Microbial mutualism in
leaching of Cu-Ni sulfide concentrate. International Journal
of Mineral frocessing 2: 1-14.
TRUDINGER, P.A., 1955. Phosphoglycerate formation from pentose
phosphate by extracts of ThiobaciZZus denitrificans. Biochimica
et Biophysica Act 18: 581-582.
TRUDINGER, P.A., 1956. Fixation of co2 by extracts of the strict
autotroph, ThiobaciZZus denitrifican. Biochemical Journal 64:
274-286.
TRUDINGER, P. A., 1961. Thiosulphate oxidation and cytochromes in
ThiobaciZZus X. I- Fractionation of bacterial extracts and
properties of cytochromes. Biochemical Journal 78: 673-680.
TRUDINGER, P.A., 1971. Microbes, Metals and Minerals. Minerals

273
and Engineering 3: 13-25.
TRUDINGER, P.A. and KELLY, D. P., 1968. Reduced nicotinarnide
adenine dinucleotide oxidation by ThiobaciZZus neapoZitanus &
ThiobaciZZus strain C. Journal of Bacteriology 95: 1962-1963.
TRUSSELL, P. C., DUNCAN, D. W. and WALDEN, C.C., 1964. Biological
mining. Canadian Mining Journal 83: 46-49.
TSUCHIYA, H. M., 1977. Leaching of Cu-Ni sulfide concentrate from
the Duluth Gabbro. GBF Monograph Series No. 4, pp 101-106.
Conference Bacterial Leaching.
TSUCHIYA, H. M., TRIVIDI, N. C. and SCHULLER, M. L., 1974. Microbial
mutualism in ore leaching. Biotechnology and Bioengineering 16:
991-995.
TU0VINEN, 0. H., 1972. Microbiological aspects in the leaching of
uranium by ThiobaciZZus ferrooxidans. Atomic Energy Review 10:
251-258. (International Atomic Energy Agency, Vienna)
TU0VINEN, 0. H. and KELLY, D. P., 1972. Biology of ThiobaciZZus
ferrooxidans in relation to the microbiological leaching of
sulphide ores. Zeitschrift fur Allgemeine Mikrobiologie 12:
311-346.
TU0VINEN, 0. H. and KELLY, D. P.1973. Studies on the growth of
ThiobaciZZus ferrooxidans. I- Use of membrane filters and ferrous
iron agar to determine viable numbers and comparison with 14co12 -
fixation and iron-oxidation as measures of growth. Archives
fur Mikrobiologie 88: 285-299.
TU0VINEN, 0. H. and KELLY, D. P., 1974. Studies on the growth of
ThiobaciZZus ferrooxidans. II- Toxicity of uranium growing
cultures and tolerance conferred by mutation, other metal cations
and EDTA. Archives fur Mikrobiologie 95: 153-165.

274
TUOVINEN, 0. H. and KELLY, D. P., 1974-a. Studies on the growth of
Thiobacillus ferrooxidans. III- Influence of uranium, other metal
ions and 2:4-Dinitrophenol on ferrous iron oxidation and co2
fixation by the cells suspension. Archives fur Mikrobiologie
95: 165-180.
TUOVINEN, O. H. and KELLY, D. P., 1974-b. Studies on the growth of
Thiobacillus ferrooxidans. IV- Influence of monovalent metal
cations on ferrous iron oxidation and uranium toxicity in growing
cultures. Archives fur Mikrobiologie 98: 167-174.
TUOVINEN, O. H. and KELLY, D. P., 1974-c. Studies on the growth of
Thiobacillus ferrooxidans. V- Factors affecting growth in liquid
cultures and development of colonies on solid media containing
inorganic sulphur compounds. Archives fur Mikrobiologie 98:
351-364.
TUOVINEN, 0. H. and KELLY D. P., 1974-d. Use of microorganisms for the
recovery of metals. International Metallurgical Reviews 19:
21-31.
TUOVINEN, O. H., and KELLY, D. P., 1978. Metabolic transitions in cult
ures of acidophilic Thiobacilli. In: Murr, L., Torma, A. E. &
Brierley, J. A. (Eds)Metallurgical Applications of Bacterial
Leaching and Related Phenomena. The Academic Press, N. Y. (In
Press).
TUOVINEN, 0. H. and NICHOLAS, D. J. D., 1977. Transition of Thiobacil
lus ferrooxidans KG-4 from heterotrophic growth on glucose to
autotrophic growth on ferrous iron. Archives of Microbiology
114: 193-195.
TUOVINEN, 0. H., NIEMELA, S. I. and GYLLENBERG, H. G., 1971. Effect of
mineral nutrients and organic substances on the development of
Thiobacillus ferrooxidans. Biotechnology and Bioengineering 13:

275
517-527
TUOVINEN, 0. H., NIEMELA, S. I. and GYLLENBERG, H. G., 1971-a.
Tolerance of Thiobacillus ferrooxidans to some metals. Antonie
van Leeuwenhoek Journal of Microbiology and Serology 37: 489-496.
TUOVINEN, 0. H., NIEMELA, S. I. and GYLLENBERG, H. G., 1971-b.
Toxicity of membrane filters to Thiobaaillus ferrooxid.ans.
Zeitschrift fur Allgemeine Mikrobiologie 11: 627-631.
TUOVINEN, 0. H., KELLEY, B. C. and NICHOLAS, D. J. D., 1975. The
uptake and assimilation of sulphate by Thiobaaillus ferrooxidans.
Archives for Microbiology 105: 123-127.
TUTTLE, J. H. and DUGAN, P.R., 1969. Inhibitory effect of organic
substances on autotrophic growth of Thiobacillus ferrooxidans.
Bacteriological Proceedings GP131, p.64.
TUTTLE, J. H. and"DUGAN, P. R., 1976., Inhibition of growth, iron and
sulfur oxidation in Thiobaaillus ferrooxidans by simple organic
compounds. Canadian Journal of Microbiology 22: 719-730.
TUTTLE, J. H., DUGAN, P.R. and APEL, W. A., 1977. Leakage of
cellular material from Thiobaaillus ferrooxid.ans in the presence
of organic acids. Applied and Environmental Microbiology
33: 459-469.
UMBREIT, W.W. and ANDERSON, T. F., 1942. A study of Thiobaaillus
thiooxid.ans with electron microscope. Journal of Bacteriology
44: 317-320.
UMBREIT, W.W., VOGEL, H. R. and VOGLER, K. G., 1942. The significance
of fat in sulfur oxidation by Thiobacillus thiooxid.ans. Journal of
Bacteriology 43: 141-148.

276
UNZ, R. F. and LIEBERMAN, M. T., 1973. The microbiology of acid mine
water treatment in packed bed columns. Research Publication No. 77.
Institute of Research, Land and Water Resources, the Pennsylvania
State University, Pennsylvaniz.
UNZ, R. F., and LUNDGREN, D. G., 1961. A comparative nutritional study
of three chemoautotrophic bacteria, FerrobaciZZus ferrooxidans,
ThiobaciZZus ferrooxidans & ThiobaciZZus thiooxidans.
Soil Science 92: 302-313.
VAINSHTEIN, M. B., 1976. Systematic position of ThiobaciZZus
trautweinii. Mikrobiologiya 45: 137-141.
Van CAESEELE, L. and LEES, H., 1969. The ultrastructure of autotrophic
ally grown ThiobaciZZus noveZZus. Canadian Journal of
Microbiology 15: 651-654.
Van der WALT, J. P. and De KRUYFF, C. D., 1955. Anaerobic metabolism
of thiocyanate by ThiobaciZZi. Nature 176: 310-311.
Van GOOL, A. and LAUDELOUT, H., 1967. Spectrophotometric and kinetic
study of nitrite and formate oxidation by Nitrobacter
winogradskyi. Journal of Bacteriology 93: 215-220.
VANSELOW, D. G., 1976. Mechanisms of bacterial oxidation of the copper
sulphide mineral covellite. Ph.D. Thesis. University of N.S.W.
VELDKAMP. H., 1970. Enrichment cultures of prokaryotic organisms. In:
Norris, J. R. & Ribbons, D.W., (Eds.), Methods in Microbiology
vol 3A: pp. 305-361. Academic Press, London & New York.
VISHNIAC, W. and SANTER, M., 1957. The ThiobaciZZi. Bacteriological
Reviews 21: 195-213.
VOGEL, A. I., 1962. A Text Book of Quantitative Inorganic Analysis.
3rd Edition, pp. 343-357. London, Longman.

277
VOGLER, K. G., LE PAGE, G. A. and UMBREIT, W.W., 1942. The
respiration of ThiobaciZZus ferrooxidans on sulfur. Journal
of General Physiology 26: 89-102.
WAKSMAN, S. A. and STARKEY, R. L., 1923. On the growth and respi
ration of sulfur-oxidizing bacteria. Journal of General
Physiology 5: 285-310.
WAKSMAN, S. A., WARK, C. H., JOFFE, J. S. and STARKEY, R. L., 1923.
Oxidation of sulfur by microorganisms in black alkali soils.
Journal of Agricultural Research 24: 297-305.
WARREN, K., 1973. Mineral resources for the future. In: Mineral
Resources, pp. 241-244. Penguin Books, Ltd., Harrnondsworth,
England.
WATANABE, A. 1968. Hakko Koyokaishi 26: 115.
f Cited from: Gupta & Sant, 1970 ]
WEBLEY, D. M. , DUFF, R. B. and MITCHELL, W. A., 1960. A plate method
for studying the breakdown of synthetic and natural silicates by
soil bacteria. Nature 188: 766-767.
WEBLEY, D, M,, HENDERSON, M. E. K. and TAYLOR, I. F., 1963. The micro
biology of rocks and weathered stones. Journal of Soil Science
14: 102-112.
WELCHER, F. J., 1962. Standard Methods of Chemical Analysis, vol. 3B.
Van Nostrand, New York.
WENBERG, G. M., ERBISCH, F. H. and VIOLIN, M. E., 1969. Leaching of
copper by fungi. American Institute of Mining and Metallurgical
Engineering Meeting Feb. 1969; Washington D. C., Reprint No. 69B43.
WHITE, A., HANDLER, P., and SMITH, E. L., 1973. Principles of

278
Biochemistry. 5th. edition. MgGraw-Hill Kogakusha Ltd. Tokyo.
WHITTENBURY, R. and KELLY, D. P., 1977. Autotrophy: A conceptual
phoenix. Symposium- Society for General Microbiology 27:
pp. 121-149. Microbial Energetics.
WILLIAMS, R. A. D. and HOARE, D.S., 1972. Physiology of a new
facultative autotrophic ThiobaciZZus. Journal of General
Microbiology 70: 555-566.
WINOGRADSKY, S., 1887. Uber schwefelbakterien. Botan. Zeitung 45:
489-526.
[ Cited from: Vishniac & Santer, 1957.]
WINOGRADSKY, S., 1888. Uber eisenbaketrien. Botan. Zeitung 46:
261-269.
[ Cited from: Tuovinen & Kelly, 1972.]
WONG, C., SCHARER, J.M. and REILLY, P. M., 1974. A discrimination
among microbial iron oxidation mechanisms, Canadian Journal
of Chemical Engineering 52: 645-653.
WOOD, A. P. and KELLY, D. P., 1976. Triple mechanism for glucose oxid
ation in ThiobaciZZus A2. Proceedings of Society for General
Microbiology vo. 4 (Part 1): pp.23-24.
WOOD, A. P. and KELLY, D. P., 1977. Heterotrophic growth of ThiobaciZZus
A2 in sugars and organic acids. Archives for Microbiology
113: 257-264.
WOOJ, A. P., KELLY, D. P. and THURSTON, C. F., 1977. Simultaneous
operations of three catabolic pathways in the metabolism of
glucose by ThiobaciZZus A2. Archives for Mikrobiology 113:
265-274.
WOOLLEY, D., JONES, G. L. and HAPPOLD, F. C., 1962. Some metabolic
differences between ThiobaciZZus thiopa.rus, ThiobaciZZus

279
denitrifiaans and Thiobaaillus thioayanoxidans. Journal of
General Microbiology 29: 311-316.
YOUATT, J. B., 1954. Studies on the metabolism of Thiobaaillus
thioayanoxidans. Journal of General Microbiology 11: 139-149.
ZAJIC, J.E., 1969. Microbial Biogeochemistry. Academic Press,
New York and London. 345 pp.
ZAJIC, J.E. and CHIU, Y. S., 1972. Recovery of heavy metals by
microbes. Developments in Industrial Microbiology 13: 91-100.
ZAJIC, J.E. and NG, K. S., 1970. Biochemical uranium leaching.
Developments in Industrial Microbiology 11: 413-419.
ZAVARZIN, G. A., 1972. A heterotrophic satellite of Thiobaaillus
ferrooxidans. Mikrobiologiya 41: 369-370.
ZAVARZIN, G. A. and ZHILINA, T. Z., 1964. Thione bacteria from
thermal springs. Mikrobiologiya 33: 753-759.
ZIMMERLEY, S. R., WILSON, D. G. and PRATER, J. D., 1958.
Cycling leaching process employing iron oxidizing bacteria.
u. s. Patent 2,829,964.

B APPENDICES

NO.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
I
APPENDIX 6.1
ISOLATION OF NON-AC IDOPH ILi C SULPHUR OXIDISING
BACTERI/1.
P = PELLICLE T = TlJRlllllTTY + = GROW111 nm l CA TED
MEDIUM A MEDIUM B
·------ - --------- -- -------------
SAMPLE 2 DAYS 7 DAYS 2 DAYS 7 DAYS CODE. ~-..---- --- . -· -·-- ----- - -~----, -· - - .. - --
% S203 t. s2o3 r-- -----pH GROHTH
% S203 pH GROWTH
% S203 OXIDN. pH OXIDN.tROWlH OX ION. pH I OX I ON. iROWTH
--·-
2 6.5 0 0 5.0 21 p 9.0 0 0 9.0 10 +
3 6.5 0 0 6.0 10 + 9.0 0 0 9.0 0 0
5 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
6 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
6a 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
7 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
9 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
9a 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
11 6.5 0 0 6.0 10 p 9.0 0 0 9.0 0 0
12 6.5 0 0 5.5 15 p 9.0 0 0 9.0 0 0
13 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
14 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
17 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
18 6.5 0 0 6.5 0 0 9.u 0 0 9.0 0 0
19 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
21 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
24 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
26 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
29 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
31 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
33 6.5 0 0 5.5 25 T 9.0 0 0 9.0 0 0
36 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
38 6.5 0 0 6.0 16 T 9.0 0 0 9.0 0 0
48 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
49 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
Enrichment medium A cont a incLI sodium thi!Vitil phatc and its jnitail pH was 6.(,. (Starkey, l!l:itl-a)
EnrichffiC'nt medium ll was devi~;cd by lleijcrnick (1904). It also contained sodium thiosulphatc and its initial pll was 9.1.
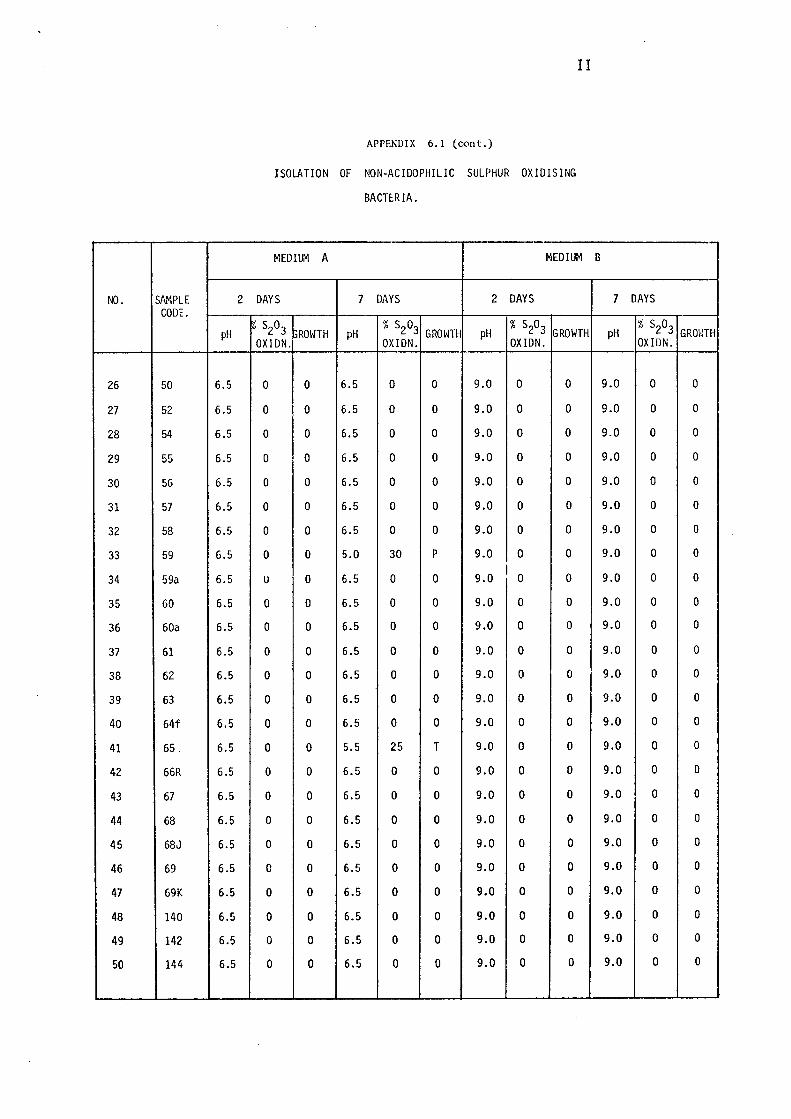
NO. SAMPLE 2 CODE.
pH
26 50 6.5
27 52 6.5
28 54 6.5
29 55 6.5
30 5G 6.5
31 57 6.5
32 58 6.5
33 59 6.5
34 59a 6.5
35 60 6.5
36 60a 6.5
37 61 6.5
38 62 6.5
39 63 6.5
40 64f 6.5
41 65. 6.5
42 66R 6.5
43 67 6.5
44 68 6.5
45 68J 6.5
46 69 6.5
47 69K 6.5
48 140 6.5
49 142 6.5
50 144 6.5
II
APPENDIX 6.1 (c-0nt.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM
DAYS 7 DAYS 2 DAYS
% S203 ,ROWTH pH % S203 GROWTH pH % S203 GROWTH
OXIDN. OXIDN. OXIDN.
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 5.0 30 p 9.0 0 0
u 0 6.5 0 0 9.0 I 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 5.5 25 T 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
B
7 DAYS
pH % S203 GROWTH OXIDN.
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
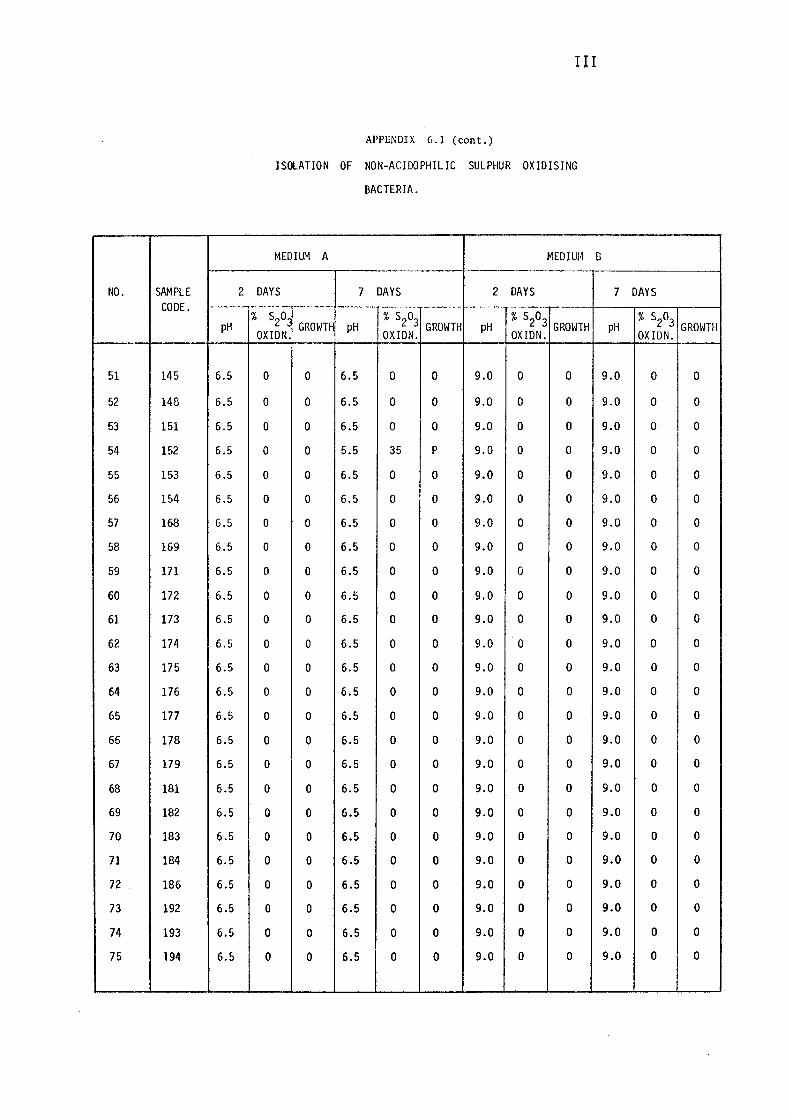
NO. SAMPLE 2 CODE.
pH
51 145 6.5
52 148 6.5
53 151 6.5
54 152 6.5
55 153 6.5
56 154 6.5
57 168 6.5
58 169 6.5
59 171 6.5
60 172 6.5
61 173 6.5
62 174 6.5
63 175 6.5
64 176 6.5
65 177 6.5
66 178 6.5
67 179 6.5
68 181 6.5
69 182 6.5
70 183 6.5
71 184 6.5
72 186 6.5
73 192 6.5
74 193 6.5
75 194 6.5
llI
APPENDIX 6.1 (cont.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM [3
DAYS 7 DAYS 2 DAYS
% s2o_ GROWTH pH
% S203 GROWTH pH
% S203 GROWTH
OXIDN. OXIDN. OXIDN.
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 5.5 35 p 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
7 DAYS
pH % S203
GROWTH OXIDN.
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
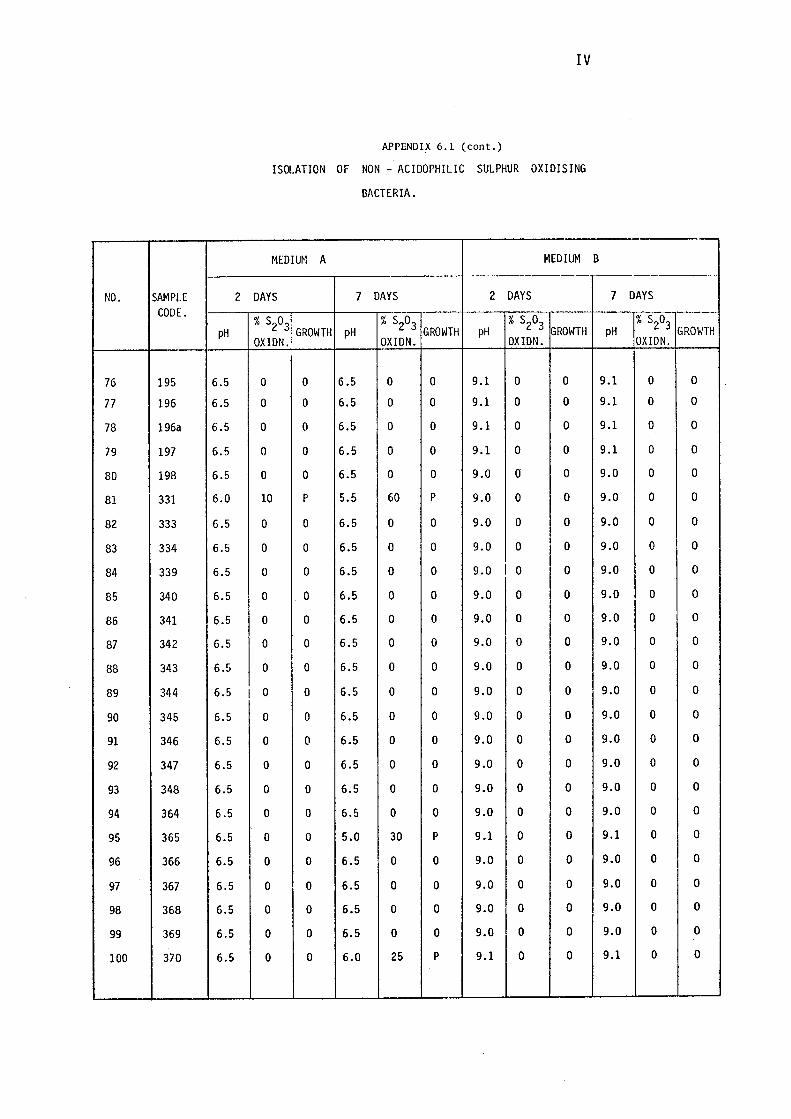
NO. SAMPLE 2 CODE.
pH
76 195 6.5
77 196 6.5
78 196a 6.5
79 197 6.5
80 198 6.5
81 331 6.0
82 333 6.5
83 334 6.5
84 339 6.5
85 340 6.5
86 341 6.5
87 342 6.5
88 343 6.5
89 344 6.5 I
90 345 6.5
91 346 6.5
92 347 6.5
93 348 6.5
94 364 6.5
95 365 6.5
96 366 6.5
97 367 6.5
98 368 6.5
99 369 6.5
100 370 6.5
IV
APPENDIX 6.1 (cont.)
ISOLATION OF NON - ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM
DAYS 7 DAYS 2 DAYS
% S203 GROWTH pH
% S203 GROWTH pH
% S203 GROWTH
OXIDN. OXIDN. OXIDN.
0 0 6.5 0 0 9.l 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 o 0
10 p 5.5 60 p 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 I 0 0
0 . 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 5.0 30 p 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.0 25 p 9.1 0 0
B
7 DAYS
pH % S203
GROWTH OXIDN.
9.1 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.0 0 o 9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.1 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.1 0 0
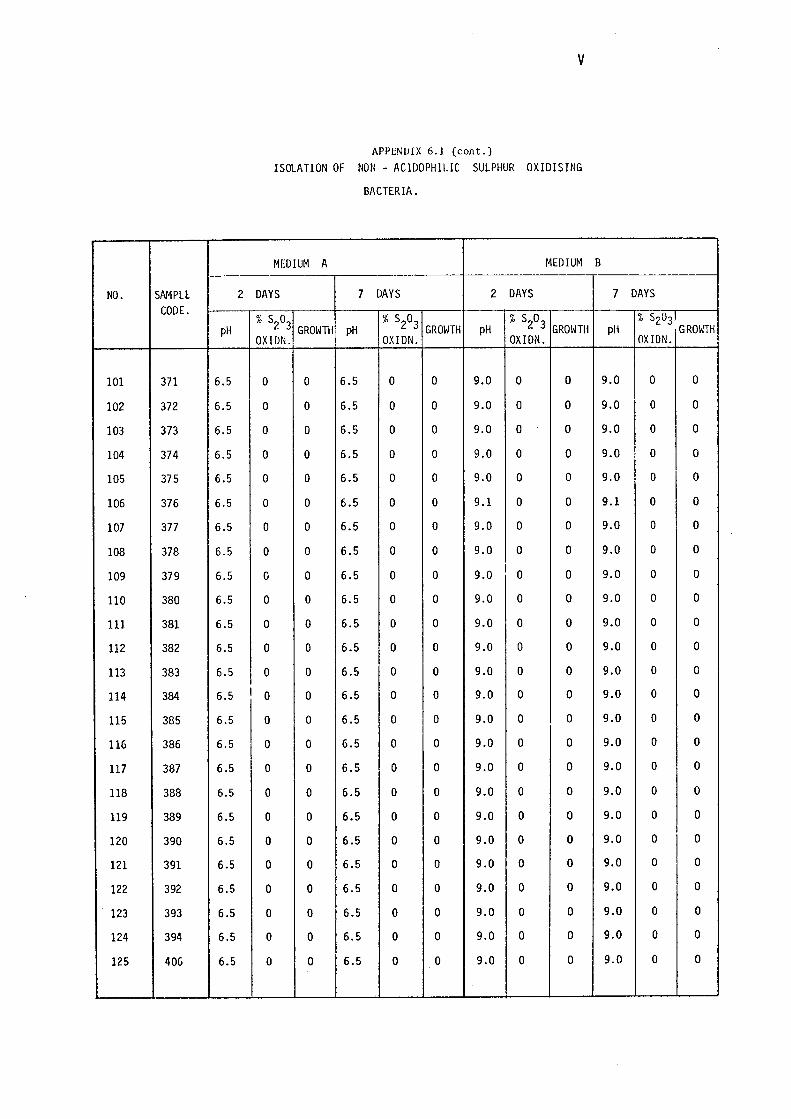
V
APPENUIX 6.1 (cont.)
ISOLATION OF NON - ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM B
NO. SAMPLE 2 DAYS 7 DAYS 2 DAYS 7 DAYS CODE.
pH % S203
GROWTH pH % S203
GROWTH pH % S203
GROWTH pH % S203I
GROh'TH OXIDN. OXIDN. OXIDN. OXIDN.
101 371 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
102 372 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
103 373 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
104 374 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
105 375 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
106 376 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
107 377 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
108 378 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
109 379 6.5 0 0 6.5 0 0 9.0 I 0 0 9.0 0 0
llO 380 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
111 381 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
112 382 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
113 383 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
114 384 6.5 I 0 0 6.5 0 0 9.0 0 0 9.0 0 0
115 385 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
116 386 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
117 387 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
118 388 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
119 389 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
120 390 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
121 391 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
122 392 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
123 393 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
124 394 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
125 406 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
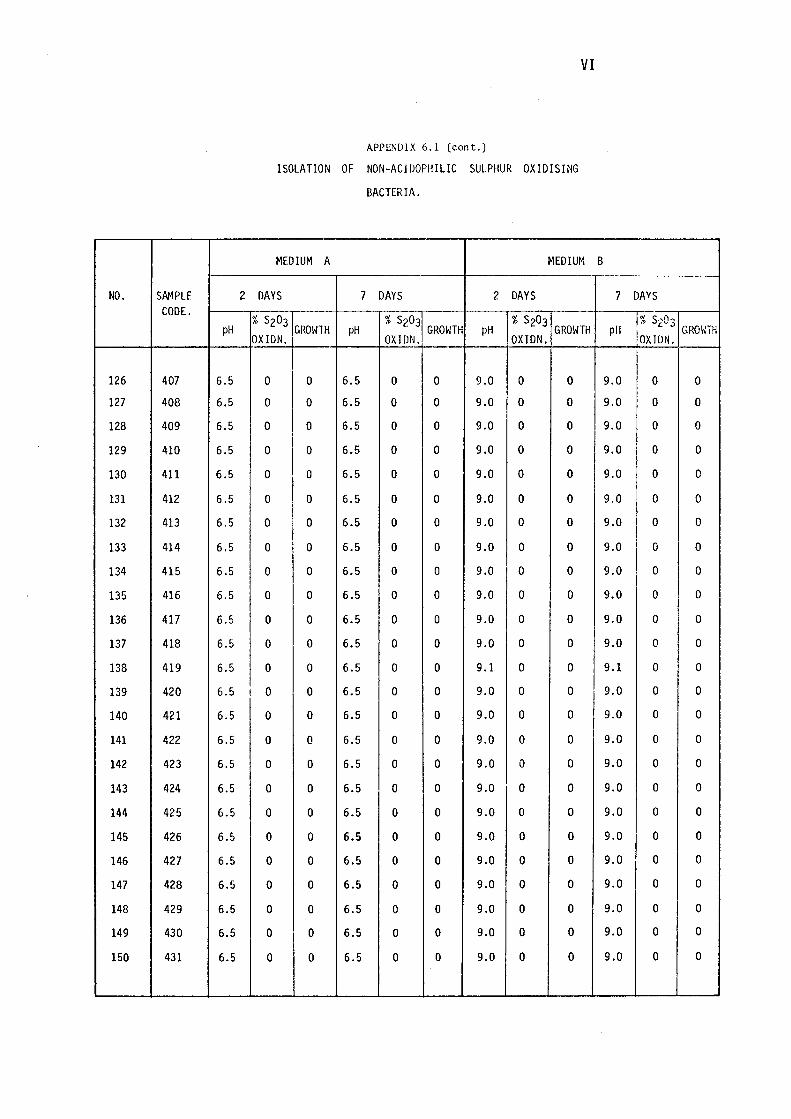
NO. SAMPLE 2 CODE.
pH
126 407 6.5
127 408 6.5
128 409 6.5
129 410 6.5
130 411 6.5
131 412 6.5
132 413 6.5
133 414 6.5
134 415 6.5
135 416 6.5
136 417 6.5
137 418 6.5
138 419 6.5
139 420 6.5 I
140 421 6.5
141 422 6.5
142 423 6.5
143 424 6.5
144 425 6.5
145 426 6.5
146 427 6.5
147 428 6.5
148 429 6.5
149 430 6.5
150 431 6.5
VI
APPENDIX 6.1 (cont.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM
DAYS 7 DAYS 2 DAYS
% S203 GROWTH pH
% S203 GROWTH pH
% S203 GROWTH
OXIDN. OXIDN. OXIDN.
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
B
7 DAYS
pH Ix s2o3
GROIHH OXIDN.
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.1 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
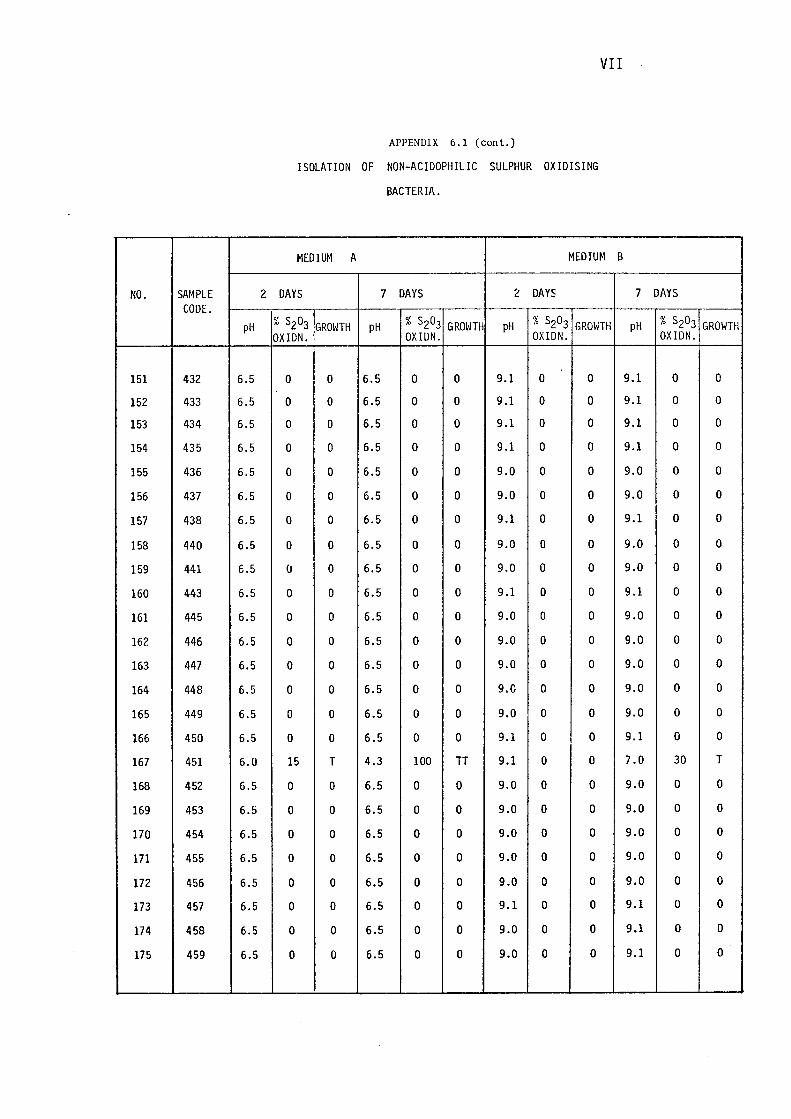
NO. SAMPLE 2 CODE.
pH
151 432 6.5
152 433 6.5
153 434 6.5
154 435 6.5
155 436 6.5
156 437 6.5
157 438 6.5
158 440 6.5
159 441 6.5
160 443 6.5
161 445 6.5
162 446 6.5
163 447 6.5
164 448 6.5
165 449 6.5
166 450 6.5
167 451 6.0
168 452 6.5
169 453 6.5
170 454 6.5
171 455 6.5
172 456 6.5
173 457 6.5
174 458 6.5
175 459 6.5
VII
APPENDIX 6.1 (cont.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM
DAYS 7 DAYS 2 DAYS
% S203 GROWTH pH % S203 GROWTH pH % S203 GROWTH OXIDN. OXIDN. OXIDN.
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.C 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
15 T 4.3 100 TT 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
B
7 DAYS
pH % S203 GROWTH OXIDN.
9.1 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.0 0 0
9.0 0 0
9.1 0 0
9.0 0 0
9.0 0 0
9.1 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.1 0 0
7.0 30 T
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.1 0 0
9.1 0 0
9.1 0 0
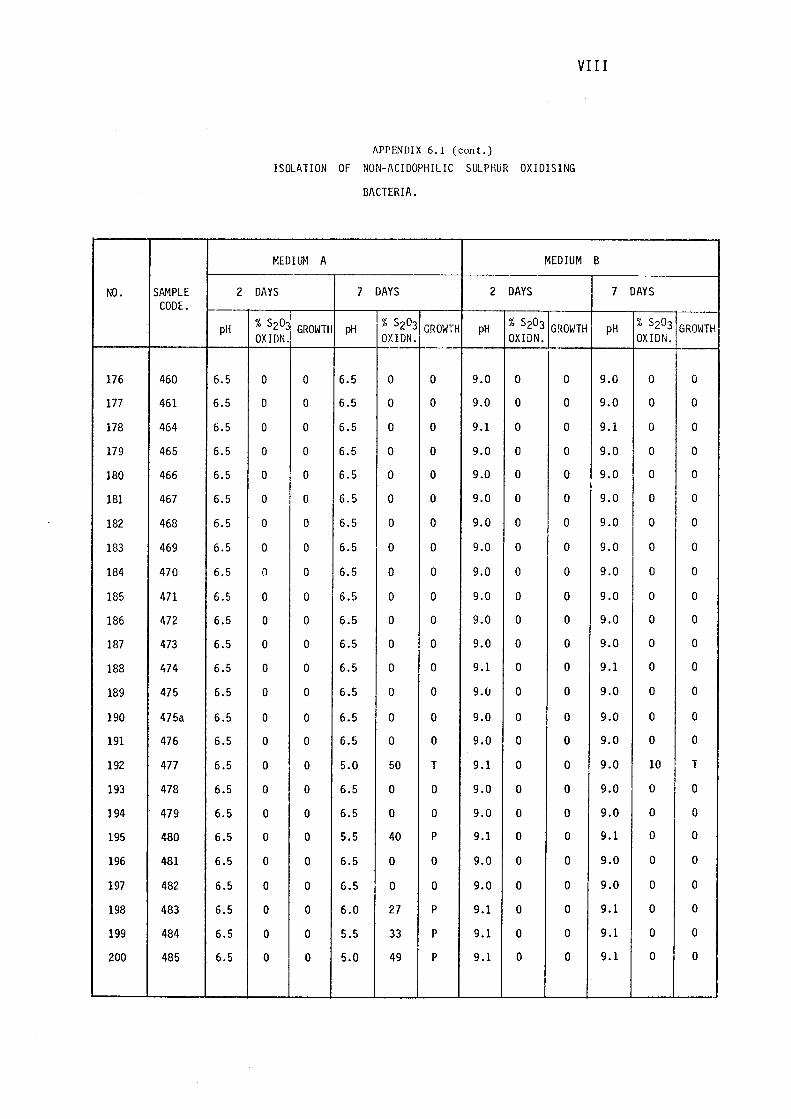
VIII
APPENDIX6.l (cont.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM B
NO. SAMPLE 2 DAYS 7 DAYS 2 DAYS 7 DAYS CODE.
pH % S203 GROWTH pH % S203 GROWTH pH % S203 GROWTH pH % S203 GROWTH OXIDN. OXIDN. OXIDN. OXIDN.
176 460 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
177 461 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
178 464 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
179 465 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
180 466 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
181 467 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
182 468 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
183 469 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
184 470 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
185 471 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
186 472 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
187 473 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
188 474 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
189 475 6.5 0 0 6.5 0 0 9.(J 0 0 9.0 0 0
190 475a 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
191 476 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
192 477 6.5 0 0 5.0 50 T 9.1 0 0 9.0 10 T
193 478 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
194 479 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
195 480 6.5 0 0 5.5 40 p 9.1 0 0 9.1 0 0
196 481 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
197 482 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
198 483 6.5 0 0 6.0 27 p 9.1 0 0 9.1 0 0
199 484 6.5 0 0 5.5 33 p 9.1 0 0 9.1 0 0
200 485 6.5 0 0 5.0 49 p 9.1 0 0 9.1 0 0
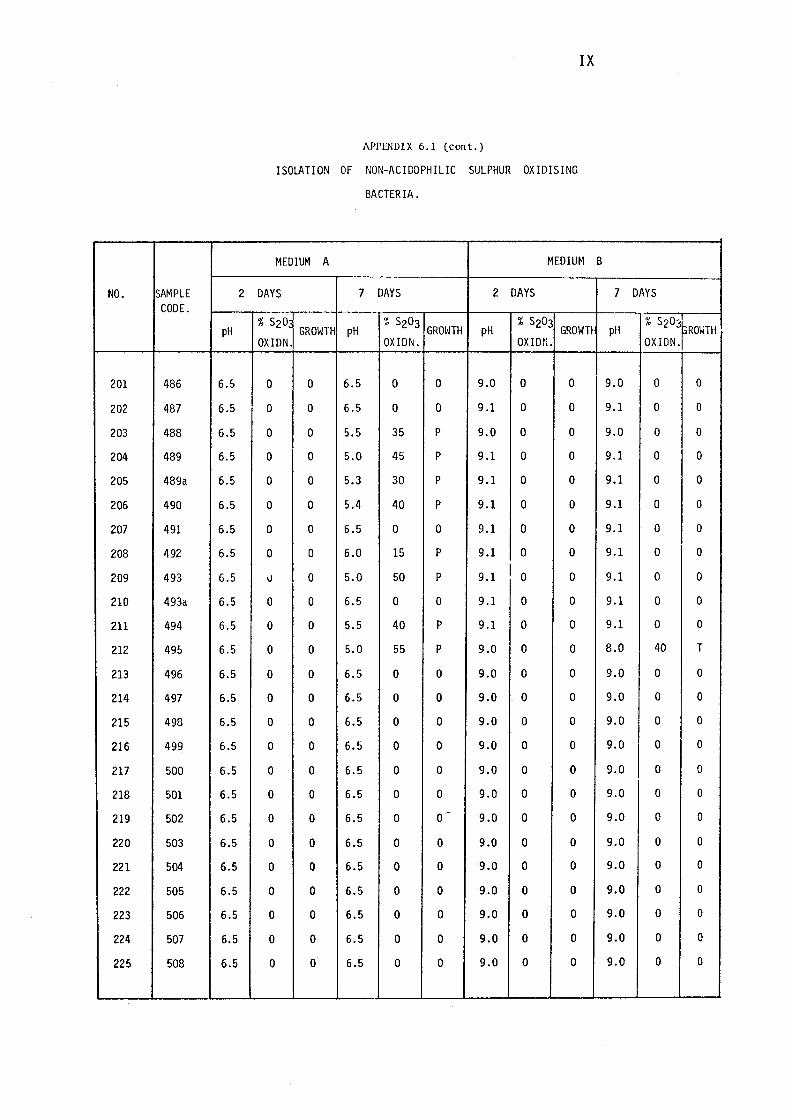
NO. SAMPLE 2 CODE.
pH
201 486 6.5
202 487 6.5
203 488 6.5
204 489 6.5
205 489a 6.5
206 490 6.5
207 491 6.5
208 492 6.5
209 493 6.5
210 493a 6.5
211 494 6.5
212 495 6.5
213 496 6.5
214 497 6.5
215 498 6.5
216 499 6.5
217 500 6.5
218 501 6.5
219 502 6.5
220 503 6.5
221 504 6.5
222 505 6.5
223 506 6.5
224 507 6.5
225 508 6.5
IX
APPENDIX 6.1 (cont.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM
DAYS 7 DAYS 2 DAYS
% S20 % S203 % S20 3 GROWH GROWTH pH GROWTH pH OXIDN. OXIDN. OXIDN.
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 5.5 35 p 9.0 0 0
0 0 5.0 45 p 9.1 0 0
0 0 5.3 30 p 9.1 0 0
0 0 5.4 40 p 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.0 15 p 9.1 0 0
u 0 5.0 50 p 9.1 I 0 0
0 0 6.5 0 0 9.1 0 0
0 0 5.5 40 p 9.1 0 0
0 0 5.0 55 p 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 -- 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
B
7 DAYS
% s 0-1
pH 2 3rROWTH OXIDN.
9.0 0 0
9.1 0 0
9.0 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.1 0 0
8.0 40 T
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
9.0 0 0
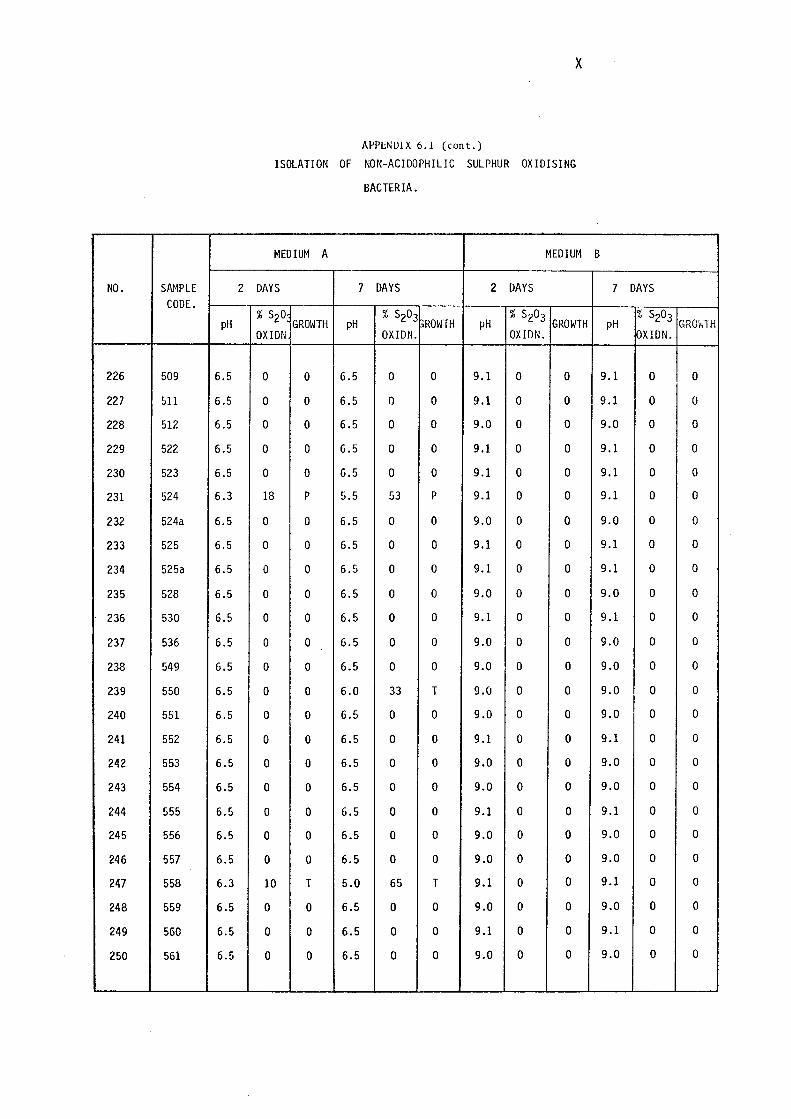
X
APPENDIX 6.1 (cont.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM B
NO. SAMPLE 2 DAYS 7 DAYS 2 DAYS 7 DAYS CODE.
pH % S20:
GROWTH pH % S203
GROWTH pH % S203
GROWTH pH % S203
GROviTH OXIDN OXIDN. OXIDN. OXIDN.
226 509 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
227 511 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
228 512 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
229 522 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
230 523 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
231 524 6.3 18 p 5.5 53 p 9.1 0 0 9.1 0 0
232 524a 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
233 525 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
234 525a 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
235 528 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
236 530 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
237 536 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
238 549 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
239 550 6.5 0 0 6.0 33 T 9.0 0 0 9.0 0 0
240 551 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
241 552 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
242 553 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
243 554 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
244 555 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
245 556 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
246 557 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
247 558 6.3 10 T 5.0 65 T 9.1 0 0 9.1 0 0
248 559 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
249 560 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
250 561 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
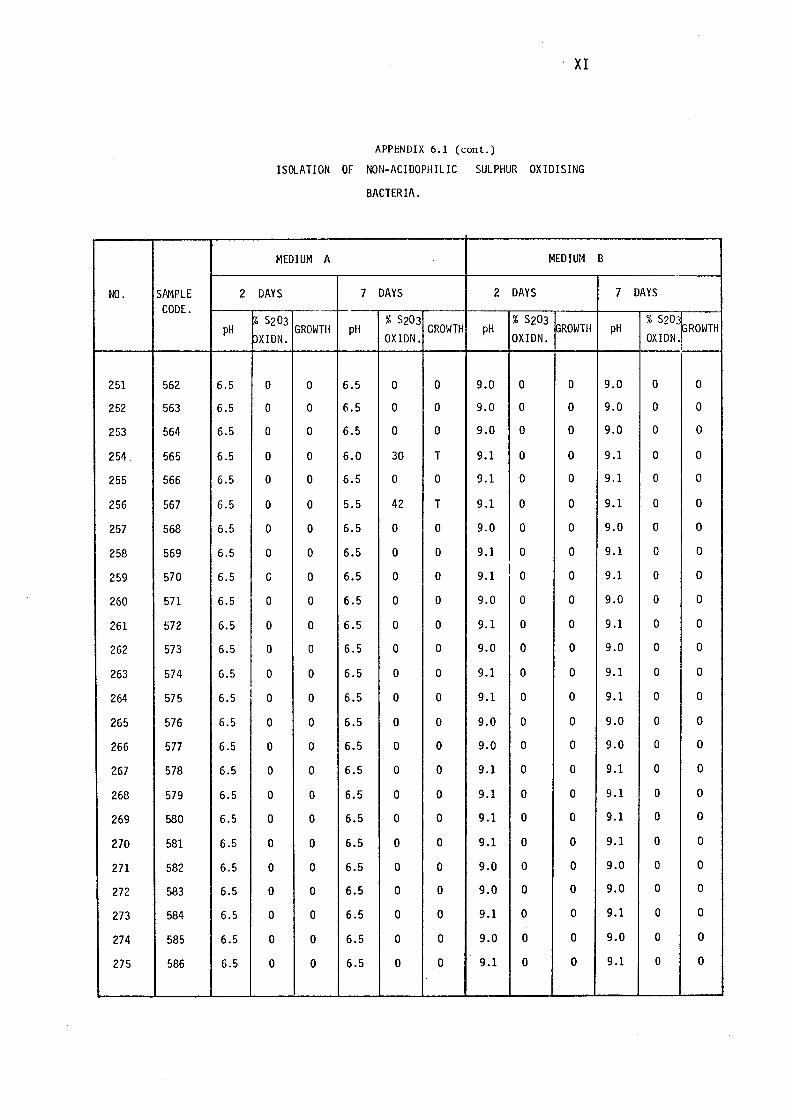
NO. SJ\MPLE 2 CODE.
pH
251 562 6.5
252 563 6.5
253 564 6.5
254 565 6.5
255 566 6.5
256 567 6.5
257 568 6.5
258 569 6.5
259 570 6.5
260 571 6.5
261 572 6.5
262 573 6.5
263 574 6.5
264 575 6.5 I
265 576 6.5
266 577 6.5
267 578 6.5
268 579 6.5
269 580 6.5
270 581 6.5
271 582 6.5
272 583 6.5
273 584 6.5
274 585 6.5
275 586 6.5
· XI
APPENDIX 6.1 (cont.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM
DAYS 7 DAYS 2 DAYS ·-,.._
r~ s203 GROWTH pH
% s203 GROWTH pH
% s203 GROWTH
OXIDN. OXIDN. OXIDN.
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.0 30 T 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 5.5 42 T 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
C 0 6.5 0 0 9.1 I 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
0 0 6.5 0 0 9.0 0 0
0 0 6.5 0 0 9.1 0 0
B
7 DAYS
pH % s203,
PROWTH OXIDN.J
9.0 0 0
9.0 0 0
9.0 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.0 0 0
9.1 0 0
9.1 0 0
9.0 0 0
9.1 0 0
9.0 0 0
9.1 0 0
9.1 0 0
9.0 0 0
9.0 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.1 0 0
9.0 0 0
9.0 0 0
9.1 0 0
9.0 0 0
9.1 0 0
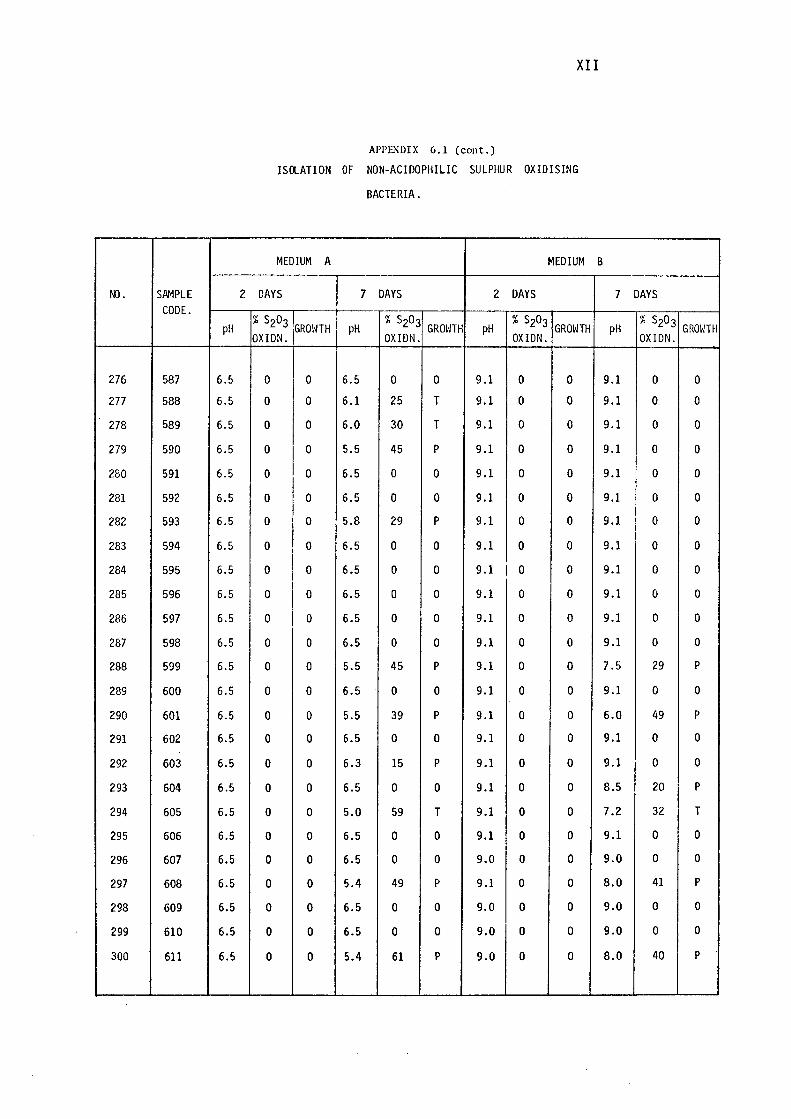
XII
APPEt~DIX 6.1 (cont.)
ISClATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM B
Ml. SAMPLE 2 DAYS 7 DAYS 2 DAYS 7 DAYS CODE.
% S O i % S203 % S203 % S203 pH 2 3 GROIHH pH GROWTH pH GROWTH pH GROWTH
OXIDN. OXIDN. OXIDN. OXIDN.
276 587 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
277 588 6.5 0 0 6.1 25 T 9.1 0 0 9.1 0 0
278 589 6.5 0 0 6.0 30 T 9.1 0 0 9.1 0 0
279 590 6.5 0 0 5.5 45 p 9.1 0 0 9.1 0 0
280 591 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
281 592 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
282 593 6.5 0 0 5.8 29 p 9.1 0 0 9.1 0 0
283 594 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
284 595 6.5 0 0 6.5 0 0 9.1 I 0 0 9.1 0 0
285 596 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
286 597 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
287 598 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
288 599 6.5 0 0 5.5 45 p 9.1 0 0 7.5 29 p
289 600 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
290 601 6.5 0 0 5.5 39 p 9.1 0 0 6.0 49 p
291 602 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
292 603 6.5 0 0 6.3 15 p 9.1 0 0 9.1 0 0
293 604 6.5 0 0 6.5 0 0 9.1 0 0 8.5 20 p
294 605 6.5 0 0 5.0 59 T 9.1 0 0 7.2 32 T
295 606 6.5 0 0 6.5 0 0 9.1 0 0 9.1 0 0
296 607 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
297 608 6.5 0 0 5.4 49 p 9.1 0 0 8.0 41 p
298 609 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
299 610 6.5 0 0 6.5 0 0 9.0 0 0 9.0 0 0
300 611 6.5 0 0 5.4 61 p 9.0 0 0 8.0 40 p
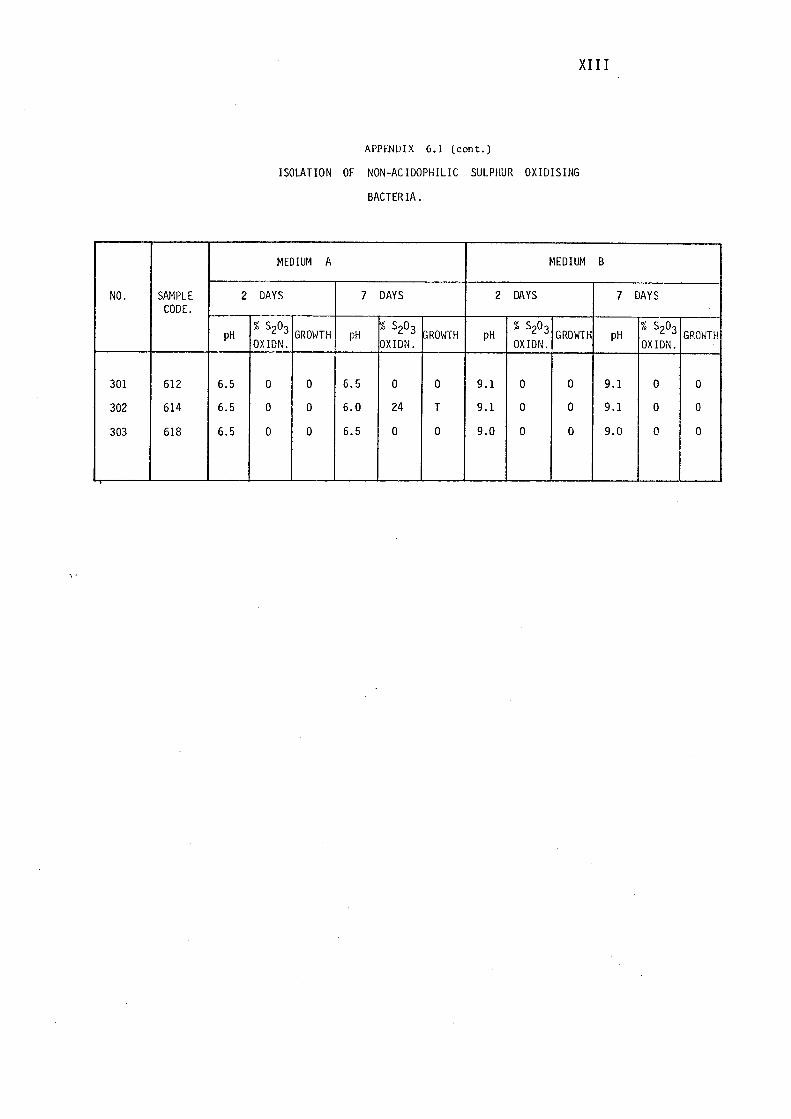
NO. SAMPLE 2 CODE.
pH
301 612 6.5
302 614 6.5
303 618 6.5
XII I
APPI:N!JIX 6.1 (cont.)
ISOLATION OF NON-ACIDOPHILIC SULPHUR OXIDISING
BACTERIA.
MEDIUM A MEDIUM
DAYS 7 DAYS 2 DAYS
% S203 GROIHH pH
% S203 1,ROWTH pH
% Sz03 GROWTH
OX IDN. OXIDN. OXIDN.
0 0 6.5 0 0 9.1 0 0
0 0 6.0 24 T 9.1 0 0
0 0 6.5 0 0 9.0 0 0
B
7
pH
9.1
9.1
9.0
DAYS
% Sz03 GROIHH
OXIDN.
0 0
0 0
0 0
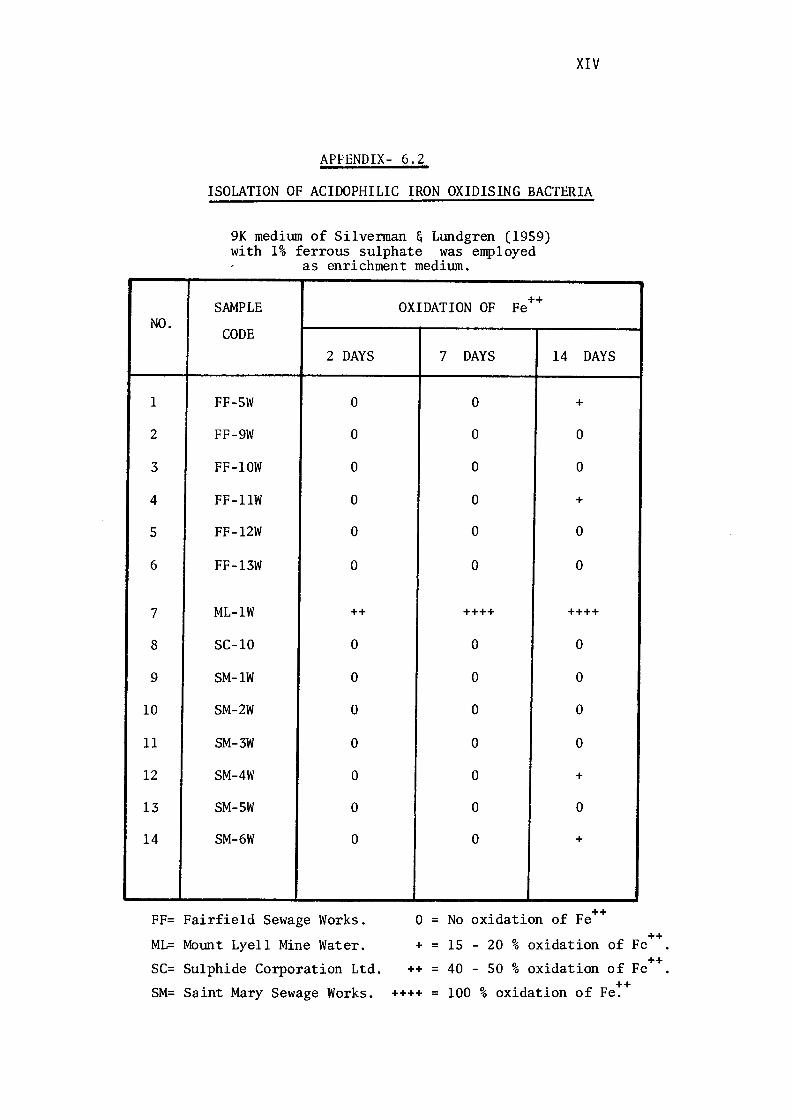
NO.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
FF=
ML=
SC=
SM=
XIV
APPENDIX- 6.2
ISOLATION OF ACIOOPHILIC IRON OXIDISING BACTERIA
9K medium of Silvennan & Lundgren (1959) with 1% ferrous sulphate was employed
as enrichment medium.
SAMPLE OXIDATION OF Fe ++
CODE 2 DAYS 7 DAYS 14 DAYS
FF-SW 0 0 +
FF-9W 0 0 0
FF-lOW 0 0 0
FF-llW 0 0 +
FF-12W 0 0 0
FF-13W 0 0 0
ML-lW ++ ++++ ++++
SC-10 0 0 0
SM-lW 0 0 0
SM-2W 0 0 0
SM-3W 0 0 0
SM-4W 0 0 +
SM-SW 0 0 0
SM-6W 0 0 +
Fairfield Sewage Works. 0 = "d . f F ++ No oxi ation o e
Mount Lyell Mine Water. + = 15 20 % oxidation ++
of Fe .
Sulphide Corporation Ltd. % ++ ++ = 40 so oxidation of Fe .
Saint Mary Sewage Works. ++++ = 100 % oxidation of Fe:+
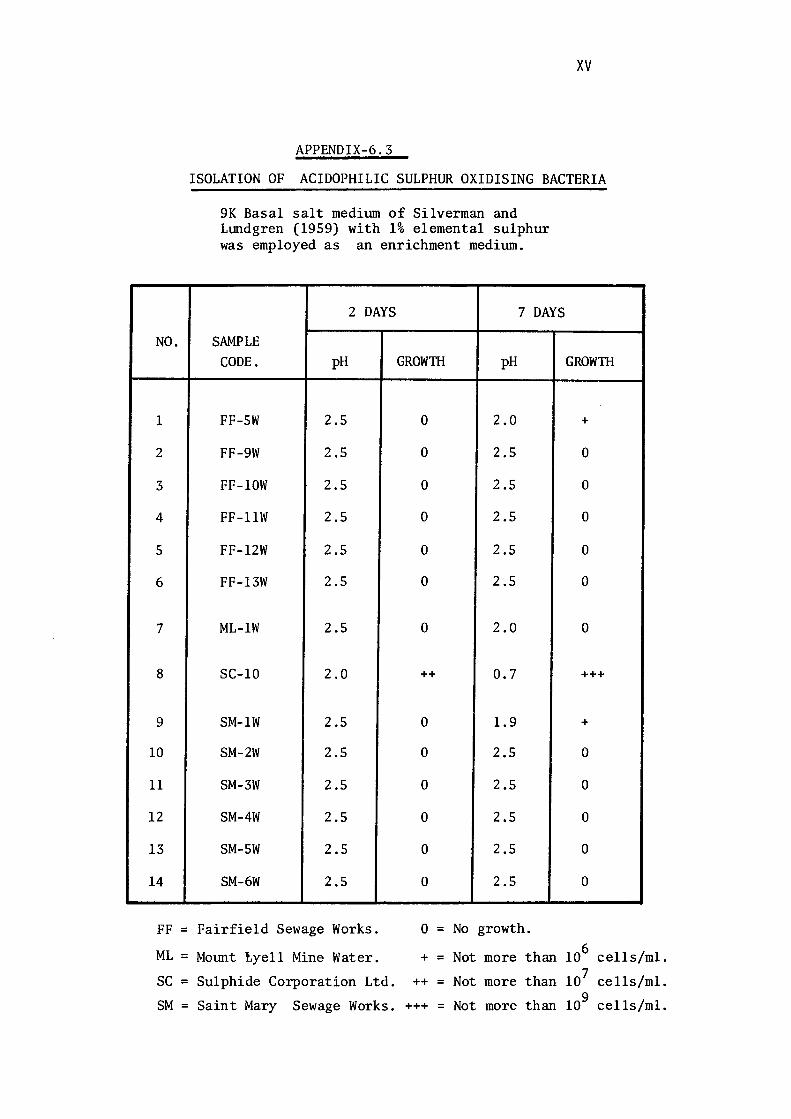
xv
APPENDIX-6.3
ISOLATION OF ACIDOPHILIC SULPHUR OXIDISING BACTERIA
NO.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
FF=
ML=
SC=
SM=
9K Basal salt medium of Silverman and Lmdgren (1959) with 1% elemental sulphur was employed as an enrichment medium.
2 DAYS 7 DAYS
SAMPLE CODE. pH GROWTH pH
FF-SW 2.5 0 2.0
FF-9W 2.5 0 2.5
FF-lOW 2.5 0 2.5
FF-llW 2.5 0 2.5
FF-12W 2.5 0 2.5
FF-13W 2.5 0 2.5
ML-lW 2.5 0 2.0
SC-10 2.0 ++ 0.7
SM-lW 2.5 0 1.9
SM-2W 2.5 0 2.5
SM-3W 2.5 0 2.5
SM-4W 2.5 0 2.5
SM-SW 2.5 0 2.5
SM-6W 2.5 0 2.5
Fairfield Sewage Works. 0 = No growth.
Momt tyell Mine Water. + = Not more than
Sulphide Corporation Ltd. ++ = Not more than
Saint Mary Sewage Works. +++ = Not more than
GROWTH
+
0
0
0
0
0
0
+++
+
0
0
0
0
0
106 cells/ml.
107 cells/ml.
109 cells/ml.

APPENDIX 6.4
CALCULATION OF DNA BASE COMPOSITION
(Hirschman & Felsenfield, 1966)
XVI
1- The absorbance at 5 mµ intervals from 235 mµ to 290 mµ was
measured. The absorbance values are set out in appendix 6.4a.
2- Appendix 6.4b was consulted for values of ai, Si, yi, and the
three sums were computed.
µl = I: Ai .ai = Ai235 ai235 + ----- + Ai290 . ai290
µ2 = I: Ai . Bi = Ai235 f3i235 + ----- + Ai290 . f3i290
µ3 = I: Ai .yi = Ai235 yi235 + ----- + Ai290 .yi290
Values of µ1, µ2, µ3, are set out in appendix 6.4d.
3- Appendix 6.4c was consulted and thevalues of the following
expressions computed:
ccp = µ151 + µ252 + µ353
C = µ1s4 + µ2 53 + µ3 5s
[l]
[2]
[3]
[4]
[5]
where cp is the AT fraction in the unknown DNA sample, of concentration
C, in mole nucleotides per litre and 51, s2, s3, s4 and 55 are
the constants shown in the appendix 6.4c.
4- From equations 4 & 5 the value of cp, the mole fraction of AT
was calculated as follows:
cp = ~ C
µ151 + µ252 + µ353 =
µ154 + µ253 + µ3 5s
5- Having calculated the mole fraction of AT the mole fraction and

XVII
and hence % of GC was calculated as
GC = 1 - qi
% GC = [1 - <P] • 100
Using the values of µ 1 , µ 2 ,& µ 3from appendix 6.4d, calculated the
values of Cqi as follows:
C<P = [(66.2 X 0.9329 10- 7) + (-7250.6x 2.0631 . 10-7)
+ (24441.1 X 0.6198 10-7 )]
= [(61.8 .10- 7) + (-14958.7 . 10-7) + (15148.6 . 10-7)]
= 251. 7 . 10-7
Cq, = 0.2517 . 10-4
Now calculated the value of C according to equation [5]:
C ~ [ (66.2 x 0.2792 . 10- 7 ) + (-7250.6 x 0.6198. 10- 7)
+ (24441.1 X 0.2124 • 10- 7 )]
= [ (18.5 10- 7) + (-4493.9. 10-7) + (5191.3 . 10-7)]
= 715.9 10 - 7
C = 0. 7159 10- 4
0 .2517 10-4 <P =
0. 7159 10-4
= o. 352
Therefore A+T molar ratio is 0.352
GC mole fraction ratio would be [ 1.000 - 0.352] . 100
c: 64.8%
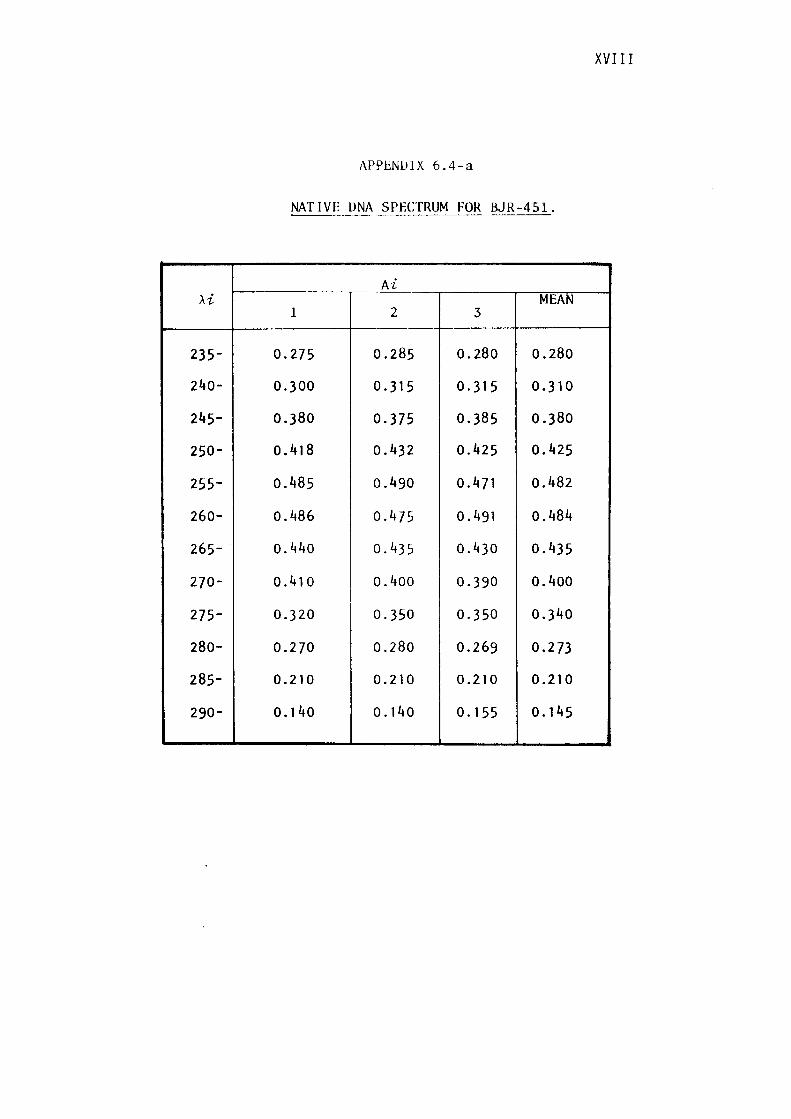
XVIII
1\PPENUIX 6.4-a
NATIVE ONA SPECTRUM FOR BJR-451.
Ai >..i
-- ..--- --· MEAN
1 2 3
235- 0.275 0.285 0.280 0.280
240- 0.300 0.315 0.315 0.31 O
245- 0.380 0.375 0.385 0.380
250- 0.418 0.432 o.425 0.425
255- 0.485 o.490 0.471 o.482
260- o.486 0.475 0. 491 0.484
265- 0.440 0.435 0.430 0.435
270- 0 .410 0.400 0. 390 0.400
275- 0.320 0.350 0.350 0.340
280- 0.270 0.280 0.269 0.273
285- 0.210 0.210 0.210 0.210
290- o. 140 0. 140 0. 155 0. 145

APP_ENDIX 6. 4-b
PARAMETERS FOR THE THREE-TERM ANALYSIS OF
NATIVE IJNA SPECTRA.
Purameters fur three-term aruilysis uf native DNA Spectra
~,(rnµ) «1 /J, Y1
~--~--·-·
235 -2026 -656 3052 240 -1889 -1251 5031 245 -1390 -1917 6338 250 43 -2830 7480 255 -319 -1807 7016 260 -608 --1141 7307 265 2515 -3370 7052 270 871 -- l 409 5740 275 - :l86 -154 4587 280 ll59 --1 ;i58 3938 285 1797 - 2424 31G4 200 1187 --209!) 2188
APPENDIX 6.4 -c
l\111lti[>ly eflch tm·rn hy 10- 7
SI 0 03'.!9 S-l 0·2702 S2 2·0fi31 S5 0·2124 S3 0·6198
XIX
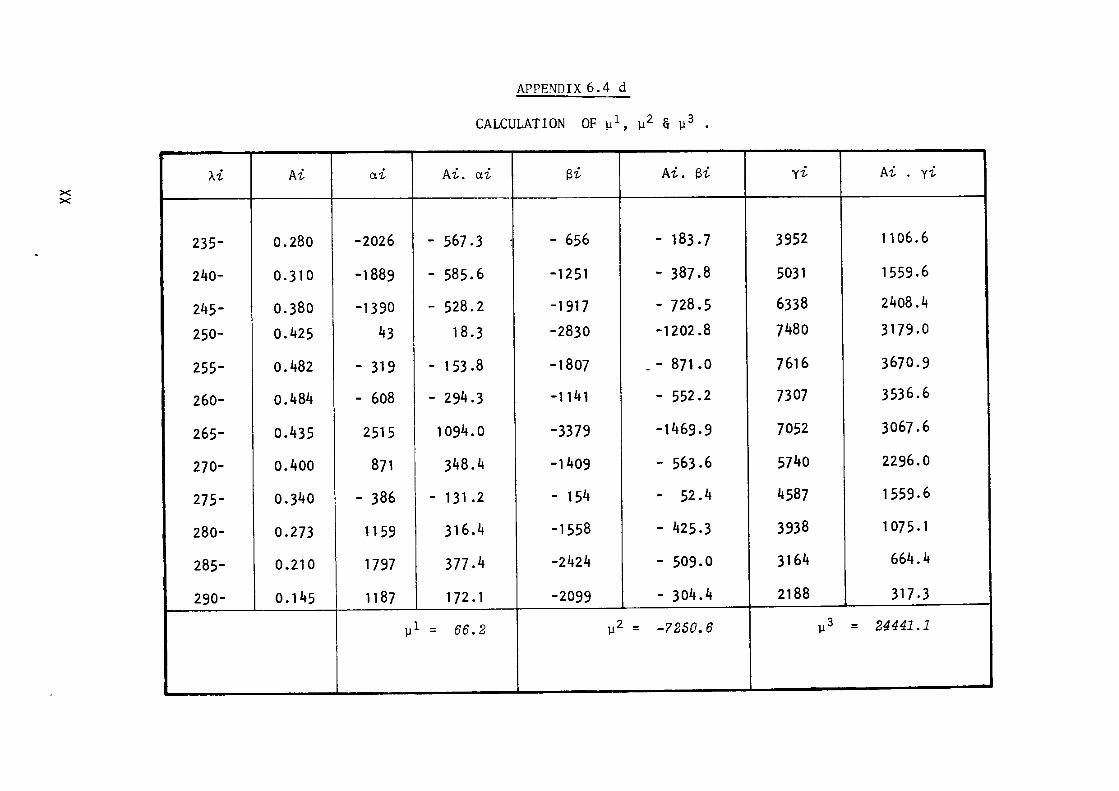
X X
)..i
235-
240-
245-
250-
255-
260-
265-
270-
275-
280-
285-
290-
Ai a1,
0.280 -2026
0.31 0 -1889
0.380 -1390
0.425 43
0.482 - 319
o.484 - 608
o.435 2515
o.4oo 871
0,340 - 386
0.273 1159
0.210 1797
0. 145 1187
APPENDIX 6. 4 d
CALCULATION OF µl, µ2 & µ3 .
Ai. ai Si Ai. Si
- 567 .3 - 656 - 183. 7
- 585.6 -1251 - 387.8
- 528.2 -1917 - 728.5
18. 3 -2830 -1202.8
- 153 .8 -1807 . - 871 .o
- 294.3 -1141 - 552.2
1094. 0 -3379 -1469.9
348.4 -1409 - 563.6
- 131.2 - 154 - 52.4
316.4 -1558 - 425,3
377 .4 -2424 - 509.0
172. 1 -2099 - 304. 4
µ 1 = 66.2 µ2 = -7250.6
y1, Ai. yi
3952 1106. 6
5031 1559,6
6338 2408.4
7480 3179.0
7616 3670.9
7307 3536.6
7052 3067.6
5740 2296.0
4587 1559. 6
3938 1075. 1
3164 664.4
2188 317.3
µ3 = 24441.1
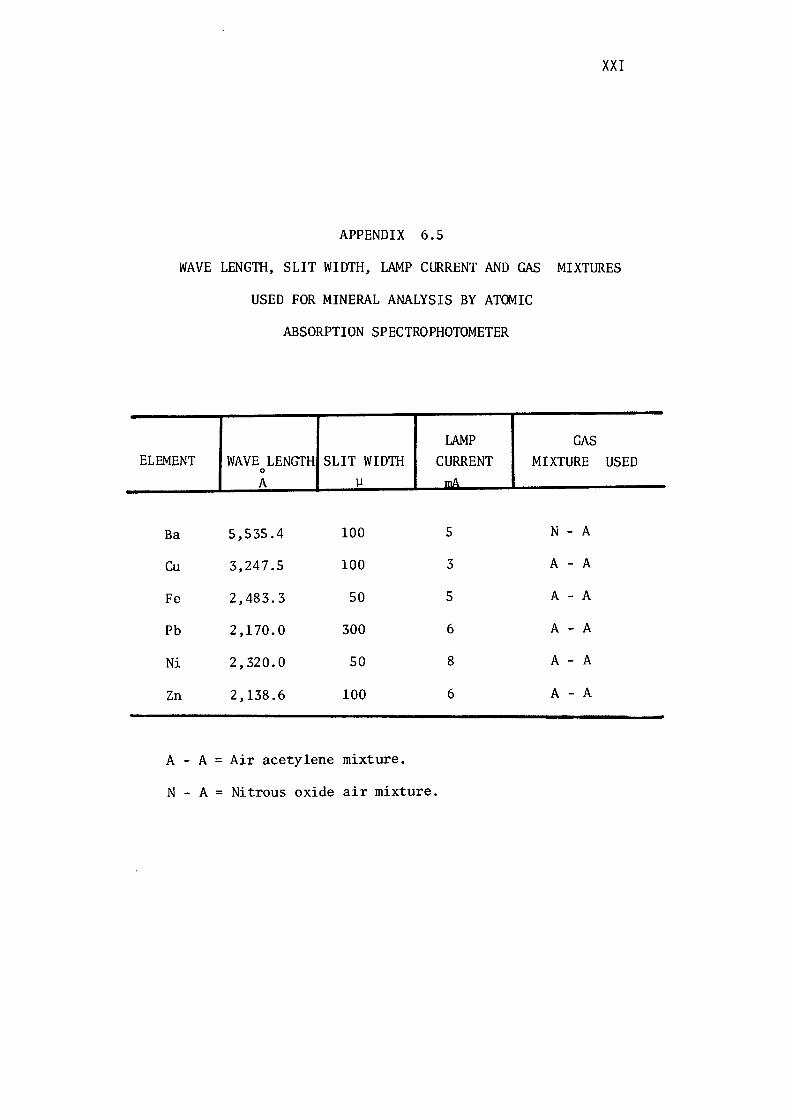
XXI
APPENDIX 6 . 5
WAVE LENGTH, SLIT WIDTH, LAMP CURRENT AND GAS MIXTURES
USED FOR MINERAL ANALYSIS BY ATOMIC
ABSORPTION SPECTROPHOTOMETER
LAMP GAS ELEMENT WAVE LENGTH SLIT WIDTH CURRENT MIXTURE USED
0
A µ mA
Ba 5,535.4 100 5 N - A
Cu 3,247.5 100 3 A - A
Fe 2,483.3 50 5 A - A
Pb 2,170.0 300 6 A - A
Ni 2,320.0 50 8 A - A
Zn 2,138.6 100 6 A - A
A - A= Air acetylene mixture.
N - A= Nitrous oxide air mixture.

APPENDIX- 6.6
Gesellschaft fur Biotechnologische Forschung mbH Braunschweig-Stockheim A. M. KHALID AND B. J. RALPH
THE LEACHING BEHAVIOUR OF
VARIOUS ZINC SULPHIDE
MINERALS WITH THREE
THIOBACILLUS SPECIES
Edited by W Schwartz
Conference Bacterial Leaching 1977
Verlag Chemie · Weinhe1m · New York 1977

THE LEACHING BEHAVIOUR OF VARIOUS ZINC SULPHIDE MINERALS
WITH THREE THIOBACILLUS SPECIES
A.M. Khalid and B.J. Ralph
School of Biological Technology, University of New South Wales, Kensington, New South Wales, Australia.
Abstract
The availability of high-grade specimens of sphalerite, wurtzite
and marmatite prompted a comparative study of the leaching rates of
these minerals in the presence of Thiobacillus ferrooxidans, !· thiooxidans and T. thioparus. In this preliminary study, samples
of the finely-ground minerals, of equivalent surface area, were sub
jected to attack by populations of equal magnitude of each of the
three bacterial species in shake flasks at 30 c. The microorganisms
were acclimatised to the particular substrates.
The results indicate that
(i) Wurtzite is much more slowly degraded than marmatite or
sphalerite by all three organisms.
(ii) Marmatite is leached more rapidly by T. ferrooxidans and
T. thiooxidans than sphalerite, in both the presence and
absence of soluble iron.
(iii) Iron-free synthetic zinc sulphide is leached more rapidly
by T. thiooxidans than by T. ferrooxidans or T. thioparus.
The hexagonal crystalline structure of zinc sulphide in wurtzite
appears to be more recalcitrant to microbial degradation than the
cubical form of sphalerite, and the substitution of iron for some of
the zinc in the marmatitic form of zinc sulphide appears to greatly
facilitate biodegradation.
Introduction
Interpretation of the behaviour of sulphide minerals as substrates
in biochemical reactions is frequently complicated by uncertainties
with respect to composition and the effects of some physical
characteristics. These minerals frequently do not display a simple
stoichiometry but exhibit a range of compositions, and their strict

166 A. M. Khalid and B. J. Ralph
chemical definition as substrates may be blurred by the presence of
other metallic ions in solid solution, by imperfections in the crystal
structure and by lattice substitution. Natural specimens of the
highest grade may contain inclusions of other minerals of sub-micron
dimensions, the detection of which can be difficult. Variations in
composition may be accompanied by changes in crystalline form as in
the pyrrhotites, and some sulphide minerals of identical chemical
composition exhibit more than one crystalline form (for example, iron
and zinc sulphides).
A limited amount of published information suggests significant
variations in the behaviour of different crystalline forms towards
microbial attack and further effects arising from substitutions in
the lattice. Leathen et al. (1) noted a high degree of resistance to
oxidation by museum-grade pyrite in the presence of acidophilic,
iron-oxidising bacteria whereas high-grade specimens of marcassite
were readily attacked. In a more detailed investigation of these two
iron sulphide minerals, Silverman et al. (2) confirmed the resistance
of coarsely-crystalline pyrite to oxidative attack by Ferrobacillus
ferrooxidans and Thiobacillus thiooxidans and observed that both these
species readily degraded marcassite. Trussell et al. (3) reported
that the marmatitic form of zinc sulphide was rapidly and completely
leached bacterially but that under similar conditions, iron-free
sphalerite showed only a minor release of zinc ions.
The availability of relatively high-grade specimens of sphalerite,
wurtzite and marmatite prompted a further investigation of the effect
upon leaching rate of variations in crystal structure and of the sub
stitution of some zinc by iron in the zinc sulphide lattice. A
further comparison was made by the use of three different species of
Thiobacillus, each operating within its optimum pH range for growth.
Materials and Methods
Two sphalerites and a triboluminescent wurtzite containing some
sphalerite were obtained from Ward's Natural Science Establishment
Inc., and a marmatite from Zinc Corporation Ltd. Chemically prepared
zinc sulphide was obtained from Ajax Co. Ltd. The content of
principal metals in each sample is shown in Table 1. Semi-quantitative
analysis by emission spectroscopy for 28 elements on the same samples
is shown in Table 2. X-ray diffraction examinations confirmed the de
scriptions of the minerals. Each sample was ground to pass 400 mesh
and specific surface areas were determined by the B.E.T. method (4).
Values are shown in Table 3.
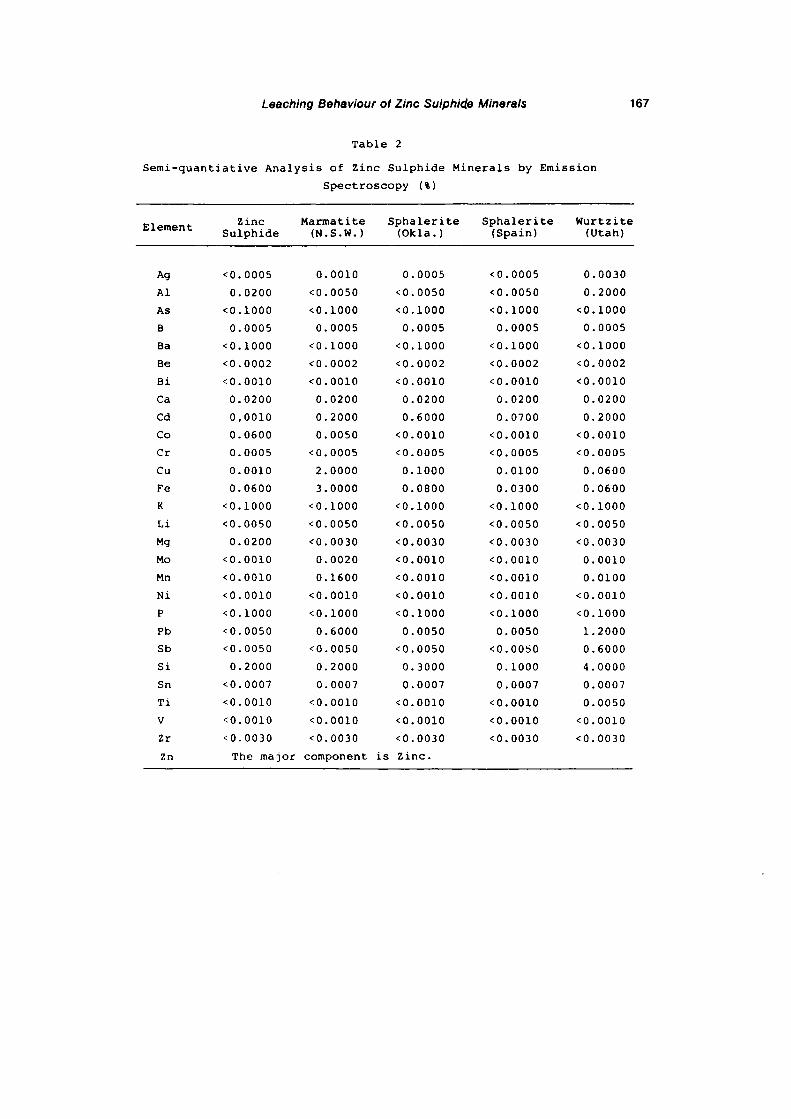
Leaching Behaviour of Zinc Sulphid.e Minerals 167
Table 2
Semi-quantiative Analysis of Zinc Sulphide Minerals by Emission
Spectroscopy (%)
Element
Ag
Al
As
B
Ba
Be
Bi
Ca
Cd
Co
er Cu
Fe
K
Li
Mg
Mo
Mn
Ni p
Pb
Sb
Si
Sn
Ti
V
Zr
Zn
Zinc Sulphide
<0.0005
0.0200
<0.1000
0.0005
<0.1000
<0.0002
<0.0010
0.0200
0,0010
0.0600
0.0005
0.0010
0.0600
<0.1000
<0.0050
0.0200
<0.0010
<0.0010
<0,0010
<0.1000
<0.0050
<0.0050
0.2000
<0.0007
<0.0010
<0.0010
<0.0030
Marmatite (N.s.w. l
0.0010
<0.0050
<0.1000
0.0005
<0.1000
<0.0002
<0.0010
0.0200
0.2000
0.0050
<0.0005
2.0000
3.0000
<0.1000
<0.0050
<0.0030
0.0020
0.1600
<0.0010
<0.1000
0.6000
<0.0050
0.2000
0.0007
<0.0010
<0.0010
<0.0030
Sphalerite (Okla.)
0.0005
<0.0050
<0.1000
0.0005
<0.1000
<0.0002
<0.0010
0.0200
0.6000
<0.0010
<0.0005
0.1000
0.0800
<0.1000
<0.0050
<0.0030
<0.0010
<0.0010
<0.0010
<0.1000
0.0050
<0.0050
0.3000
0.0007
<0.0010
<0.0010
<0.0030
The major component is Zinc.
Sphalerite (Spain)
<0.0005
<0.0050
<0.1000
0.0005
<0.1000
<0.0002
<0.0010
0.0200
0.0700
<0.0010
<0.0005
0.0100
0.0300
<0.1000
<0.0050
<0.0030
<0.0010
<0.0010
<0.0010
<0.1000
0.0050
<0.00~0
0.1000
0.0007
<0.0010
<0.0010
<0.0030
Wurtzite (Utah)
0.0030
0.2000
<0.1000
0.0005
<0.1000
<0.0002
<0.0010
0.0200
0.2000
<0.0010
<0.0005
0.0600
0.0600
<0.1000
<0.0050
<0.0030
0.0010
0.0100
<0.0010
<0.1000
1.2000
0.6000
4.0000
0.0007
0.0050
<0.0010
<0.0030
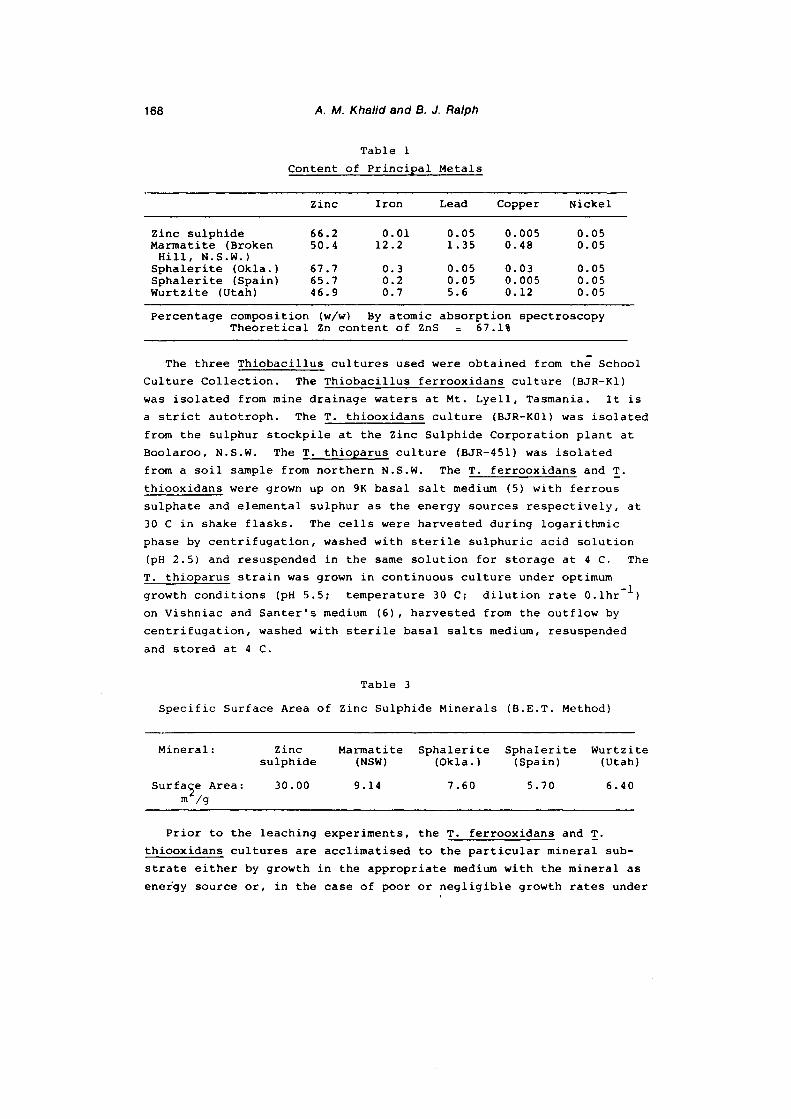
168 A. M. Khalid and B. J. Ralph
Table 1
Content of Principal Metals
Zinc Iron Lead Copper Nickel
Zinc sulphide 66.2 0.01 0.05 0.005 0.05 Marmatite (Broken 50.4 12.2 1.35 0.48 0.05
Hill, N.S.W.) Sphalerite (Okla.) 67.7 0.3 0.05 0.03 0.05 Sphalerite (Spain) 65.7 0.2 0.05 0.005 0.05 Wurtzite (Utah) 46.9 0.7 5.6 0.12 0.05
Percentage composition (w/w) By atomic absorption spectroscopy Theoretical Zn content of ZnS 67 .1%
The three Thiobacillus cultures used were obtained from the School
Culture Collection. The Thiobacillus ferrooxidans culture (BJR-Kl)
was isolated from mine drainage waters at Mt. Lyell, Tasmania. It is
a strict autotroph. The T. thiooxidans culture (BJR-K0l) was isolated
from the sulphur stockpile at the Zinc Sulphide Corporation plant at
Boolaroo, N.S.W. The T. thioparus culture (BJR-451) was isolated
from a soil sample from northern N.S.W. The T. ferrooxidans and T.
thiooxidans were grown up on 9K basal salt medium (5) with ferrous
sulphate and elemental sulphur as the energy sources respectively, at
30 C in shake flasks. The cells were harvested during logarithmic
phase by centrifugation, washed with sterile sulphuric acid solution
(pH 2.5) and resuspended in the same solution for storage at 4 C. The
T. thioparus strain was grown in continuous culture under optimum
growth conditions (pH 5.5; temperature 30 C; dilution rate 0.lhr-1 )
on Vishniac and Santer's medium (6), harvested from the outflow by
centrifugation, washed with sterile basal salts medium, resuspended
and stored at 4 C.
Table 3
Specific Surface Area of Zinc Sulphide Minerals (B.E.T. Method)
Mineral: Zinc Marmatite Sphalerite Sphalerite Wurtzite sulphide (NSW) (Okla.) (Spain) (Utah)
Surfa~e Area: 30.00 9.14 7.60 5.70 6.40 m /g
Prior to the leaching experiments, the T. ferrooxidans and T.
thiooxidans cultures are acclimatised to the particular mineral sub
strate either by growth in the appropriate medium with the mineral as
energy source or, in the case of poor or negligible growth rates under
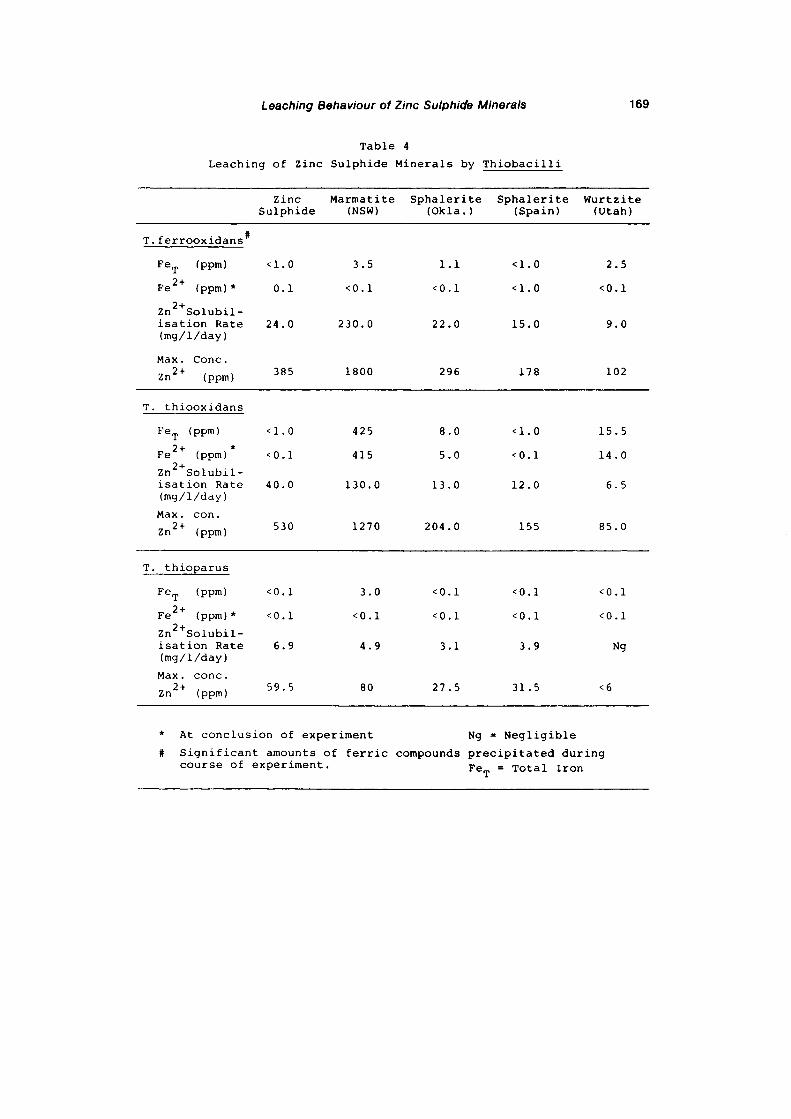
Leaching Behaviour of Zinc Sulphide Minerals 169
Table 4
Leaching of Zinc Sulphide Minerals by Thiobacilli
Zinc Sulphide
Marmatite (NSW)
Sphalerite (Okla.)
Sphalerite (Spain)
Wurtzite (Utah)
T.ferrooxidans#
FeT (ppm)
Fe 2+ (ppm)*
zn 2+solubilisation Rate (mg/1/day)
Max. Cone.
zn 2+ (ppm)
T. thiooxidans
FeT (ppm)
2+ * Fe (ppm)
zn 2+solubilisation Rate (mg/1/day)
Max. con. 2+ Zn (ppm)
T. thioparus
FeT (ppm)
2+ Fe (ppm)*
zn 2+solubilisation Rate (mg/1/day)
Max. cone. 2+ Zn (ppm)
<1.0
0.1
24.0
385
<1.0
<0.1
40.0
530
<0.1
<0.1
6.9
59.5
3. 5
<O.l
230.0
1800
425
415
130.0
1270
3.0
<0.1
4.9
80
* #
At conclusion of experiment
Significant amounts of ferric course of experiment.
1.1
<0.1
22.0
296
8.0
5.0
13. 0
204.0
<0.1
<0.1
3 .1
27.5
<1.0
<1.0
15.0
178
<1.0
<0.1
12.0
155
<0.1
<0.1
3.9
31. 5
2.5
<0.1
9.0
102
15.5
14.0
6.5
85.0
<0.1
<0.1
Ng
<6
Ng = Negligible
compounds precipitated during FeT = Total Iron

170 A. M. Khalid and B. J. Ralph
such conditions, with mineral plus the relevant soluble substrate.
The T. thioparus culture which had poor leaching capabilities against
all the natural minerals, was acclimatised against the zinc sulphide
sample. Acclimatisation was carried out in shake flasks at 30 C for
4-5 weeks, and the cells harvested by centrifugation after filtration
of the flask contents.
Determination of Leaching Rates
In the leaching experiments, an amount of mineral calculated to
provide 10 m2/100 ml. medium for surface area was added to a sterile
1000 ml. conical flask and the mineral sterilised by propylene oxide
vapour. Sterile basal salts medium (200 ml.) was added to the flask
(9K medium for T. ferrooxidans and Vishniac and Santer medium for!·
thioparus). Inoculation with acclimatised washed cells was at a level
to yield 10 7 cells/ml.
The inoculated shake flasks were incubated for 360 hours on a re
ciprocating shaking machine at 30 C. The pH of the suspending medium
was maintained at the initial level by manual adjustment from time to
time. Samples for analysis of solubilised metal ions (5 ml.) were
removed at appropriate time intervals and the volumes adjusted with
sterile medium. Zinc and total iron in the samples were determined
by atomic absorption spectroscopy; ferrous iron was estimated by the
o-phenanthroline method (7). The results of these experiments are
shown in Table 4.
A further experiment was carried out to explore the effect of
ferrous iron concentration on the leaching rate of the chemically-pure,
iron-free zinc sulphide sample, in the presence of T. ferrooxidans
and T. thiooxidans. Similar procedures were followed, except that the
pulp density was doubled. The results are shown in Table 5 and
graphically in Figure 1 (for T. ferrooxidans only).
Discussion
The results of experiments of the kind described can only be de
finitive if carried out with properly-annealed, high-purity synthetic
minerals. Nevertheless, the results obtained suggest that wurtzite
is much less amenable to attack by all three of the Thiobacillus
species employed than sphalerite, marmatite or chemically-pure zinc
sulphide. The hexagonal crystal structure in which its component ions
are arranged, in contrast to the cubical forms of sphalerite and
marmatite, may contribute to this difference in reactivity.
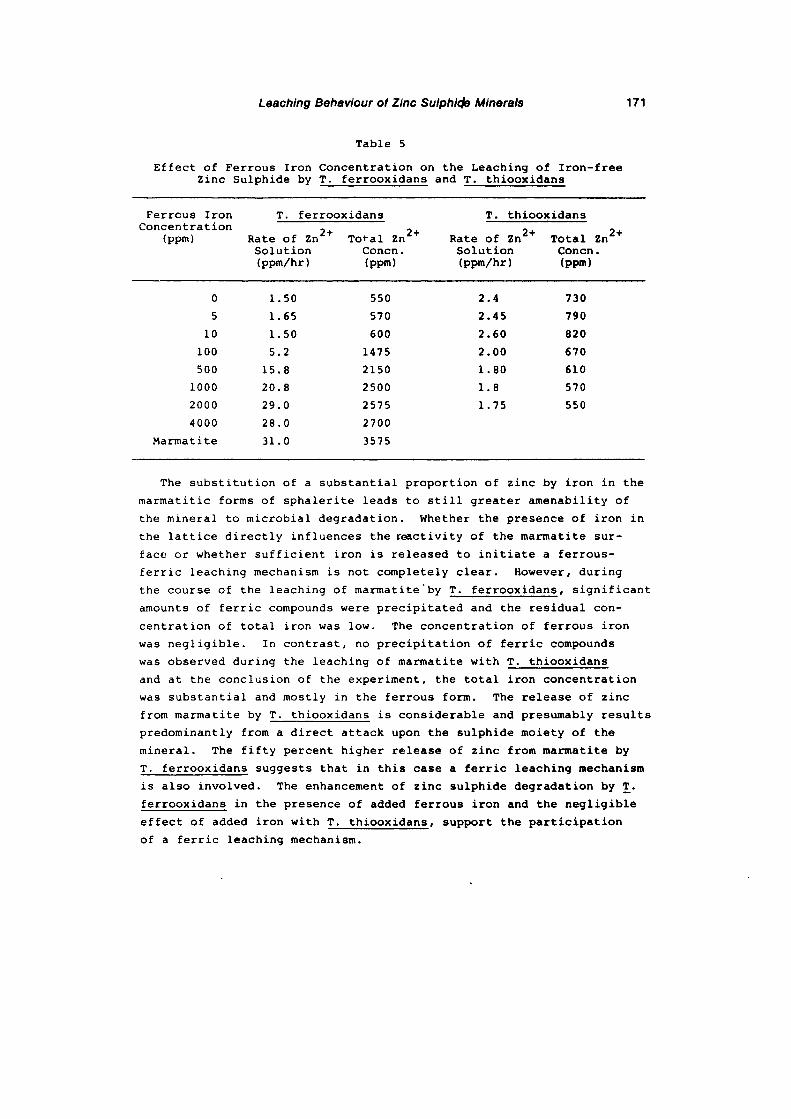
Leaching Behaviour of Zinc Sulphi</e Minerals
Table 5
Effect of Ferrous Iron Concentration on the Leaching of Iron-free Zinc Sulphide by T. ferrooxidans and T. thiooxidans
Ferrous Iron Concentration
(ppm)
0
5
10
100
SOO
1000
2000
4000
Marmatite
T. ferrooxidans
Rate of zn 2+ Solution (ppm/hr)
l. 50
1.65
l. so 5.2
15.8
20.8
29.0
28.0
31. 0
Tot-al zn 2+ Concn. (ppm)
550
570
600
1475
2150
2500
2575
2700
3575
T. thiooxidans
Rate of zn 2+ Solution (ppm/hr)
2.4
2.45
2.60
2.00
1.80
1.8
1.75
Total zn 2+ Concn. (ppm)
730
790
820
670
610
570
550
171
The substitution of a substantial proportion of zinc by iron in the
marmatitic forms of sphalerite leads to still greater amenability of
the mineral to microbial degradation. Whether the presence of iron in
the lattice directly influences the reactivity of the marmatite sur
face or whether sufficient iron is released to initiate a ferrous
ferric leaching mechanism is not completely clear. However, during
the course of the leaching of marmatite'by T. ferrooxidans, significant
amounts of ferric compounds were precipitated and the residual con
centration of total iron was low. The concentration of ferrous iron
was negligible. In contrast, no precipitation of ferric compounds
was observed during the leaching of marmatite with T. thiooxidans
and at the conclusion of the experiment, the total iron concentration
was substantial and mostly in the ferrous form. The release of zinc
from marmatite by T. thiooxidans is considerable and presumably results
predominantly from a direct attack upon the sulphide moiety of the
mineral. The fifty percent higher release of zinc from marmatite by
T. ferrooxidans suggests that in this case a ferric leaching mechanism
is also involved. The enhancement of zinc sulphide degradation by!·
ferrooxidans in the presence of added ferrous iron and the negligible
effect of added iron with T. thiooxidans, support the participation
of a ferric leaching mechanism.
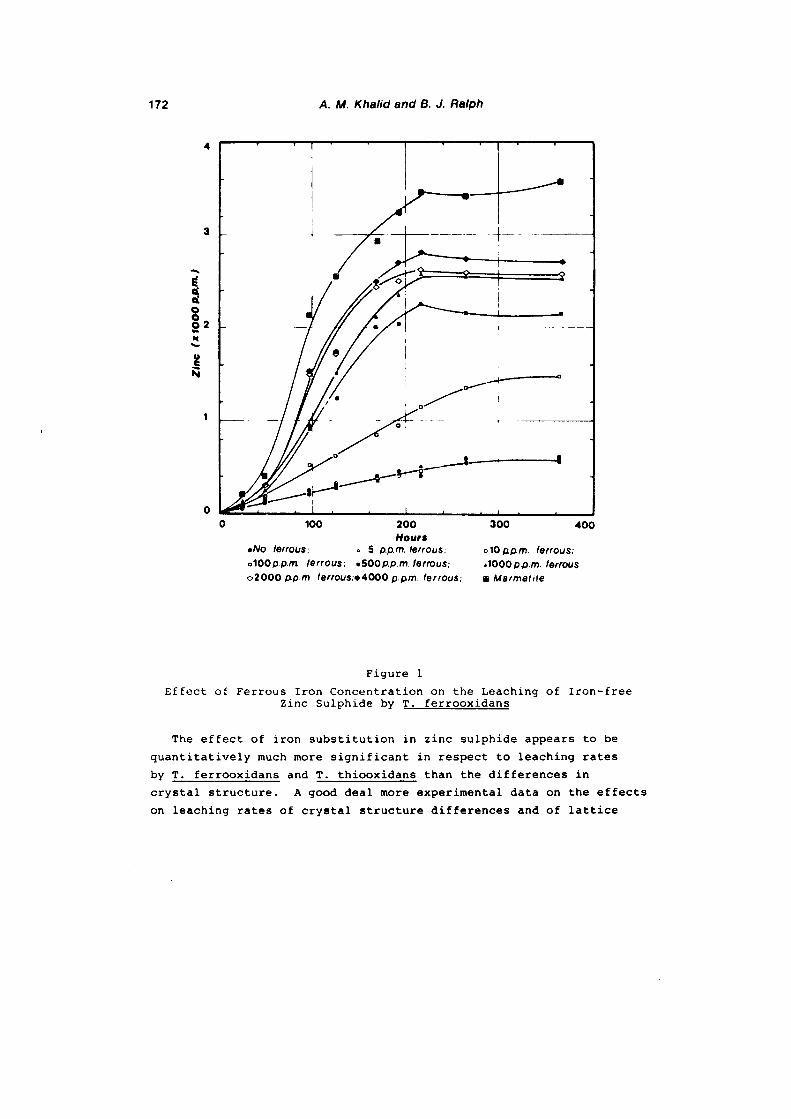
172
4
3
-i ~ 82 ... --!
0 0
A. M. Khalid and B. J. Ralph
•
100 200 Hour,
eNo ferrous; • 5 p.p.m. lerrous; a100p.p.m ferrous; .soop.p.m. ferrous; 02000 p.p.m ferrous:•4000 p.p.m. lerrous;
Figure 1
·-t- -
•
0
I
300 400
a 10 p.pm. lerrous; .1000 p.p.m. lerrous
• Marmatite
Effect of Ferrous Iron Concentration on the Leaching of Iron-free Zinc Sulphide by T. ferrooxidans
The effect of iron substitution in zinc sulphide appears to be
quantitatively much more significant in respect to leaching rates
by T. ferrooxidans and T. thiooxidans than the differences in
crystal structure. A good deal more experimental data on the effects
on leaching rates of crystal structure differences and of lattice

Leaching Behaviour of Zinc Sulphide Minerals 173
substitution is needed before generalisations can emerge. The avail
ability of a more extensive range of synthetic minerals would enable
such investigations to be carried out.
References
1. Leathen, W.W., Braley, S.A. and McIntyre, L.D. The Role of Bacteria in the Formation of Acid from Certain Sulphuric Constituents Associated with Bituminous Coal. II. Ferrous Iron Oxidising Bacteria. Appl. Microbial. ~: 65-68 (1953).
2. Silverman, M.P., Rogoff, M.H. and Wender, I. Bacterial Oxidation of Pyritic Materials in Coal. Appl. Microbial. 9 491-496 (1961).
3. Trussell, P.C., Duncan, D.W. and Walden, C.C. Biological Mining. Car.~d. Min. Jour. 85 : 46-49 (1964).
4. Brunauer, S.P., Emmett, P.H. and Teller, E.J. Adsorption of gases in multimolecular layers. J. Amer. Chem. Soc. 60 309-319 (1958).
5. Silverman, M.P. and Lundgren, D.G. Studies on the Chemoautotrophic Iron Bacterium Ferrobacillus ferrooxidans. I. An Improved Medium and a Harvesting Procedure for Securing High Cell Yield. J. Bact. 77 : 642-647 (1959).
6. Vishniac, W. and Santer, M. The Thiobacilli. Bact. Rev. 21 195-209 (1957).
7. Welcher, F.J. (Ed.) "Standard Methods of Chemical Analyses". Vol. 38; Van Nostrand, N.Y. (1962).

APPENDIX- 6.7 POSTER PAPER PRESENTE,'D AT: The Third Inter>national Syrrrposiwn on EnvirOY11T1ental Biogeoohemistry,March 27,19??,0ldenberg.
MICROBIAL ECOLOGY OF OVERBURDEN HEAPS FROM URANIUM MINING AT
RUM JUNGLE, N.T., AUSTRALIA
A.M. Khalid and B.J. Ralph
School of Biological Technoloyy, University of New South Wales,
Kensington, N.S.W., 2033, Australia.
Abstract
Microbiological studies have been undertaken to contribute
to the understanding of the basic physical, chemical and biological
mechanisms concerned in the continuing heavy metal (primarily copper and
zinc} pollution arising from uranium mining operations in the Rum Jungle
area, Northern Territory, Australia, over the period 1953-71. Release of
heavy metals from overburden heaps owing to leaching of low grade metal
sulphide ores is observed. Climatically, the area is distinguished by two
distinct seasons, an almost rainless Dry from May to September and a Wet
from November to March. The population levels of principal autotrophic
microorganism types and some heterotrophs occurring in effluent waters and
in overburden heaps have been determined and are discussed in relation to
the climatic cycle and other factors. The results of a preliminary study
of the distribution of microbial types within one of the larger overburden
heaps, at the end of a dry season, are recorded and discussed.
Introduction
Large scale uranium mining and processing is proposed for a
number of locations in Northern Australia and one proposal is the subject
of the comprehensive Ranger Uranium Environmental Inquiry at the present
time. The only major uranium mining operation hitherto carried out in the
Northern Territory was that in the Rum Jungle area which lies about 64 km
south of Darwin and 80 km east of the Joseph Bonaparte Gulf on the Timer Sea.
The area is highly mineralised and uranium, copper, lead, nickel, zinc and
cobalt ores occur (Roberts, 1960). Extensive sub-grade copper mineralisation
occurred with most of the uranium ores. The rich uranium ore deposits were
extracted by opencut operations and processed on site for'the recovery of
uranium oxide and some copper over the period 1953-71. Some copper was

2
recovered by the heap leaching of two small heaps constructed in 1966 from
low grade copper sulphide and oxide ores.
During the period of operation of the processing plant, con
siderable acid and metal pollution arose from the eventual release of process
effluents into the East Branch of the Finniss River system, owing to over
flows from holdings ponds and tailing dams during the wet season, and by 1966
further sources of pollution had arisen from the autogenous leaching of
sulphidic material in the overburden dumps and low grade ores in stockpiles.
The termination of commercial operations in 1961 left two stockpiles un
treated, an extensive tailings dam area containing considerable amounts of
residual minerals and, in the Rum Jungle Mine area proper, three substantial
overburden heaps and the leaching heaps used for copper recovery.
In 1973, the Australian Atomic Energy Commission, jointly
with the Department of Northern Australia, initiated the Rum Jungle En
vironmental Studies with the principal objectives of assessing precisely
the extent of environmental pollution in the area and of gaining under
standing of the mechanisms underlying the generation and spread of con
taminants. A secondary objective of the studies was the obtaining of
information which could be relevant to proposed developments of uranium
mining and processing in areas of similar climate in other parts of the
Northern Territory. An impressive body of information has already been
amassed by the A.A.E.C. Research Establishment in the course of the R.J.
Environmental Study (Davy, 1975). Microbiological studies were commenced
late in 1974 by the University of New South Wales, in order to assess the
nature and extent of the microbial associations and to evaluate various
procedures for the control and suppression of microbial activity. Rum
Jungle lies in a very sparsely populated area of Australia and is over
3000 km fro~ the investigating laboratories in Sydney; the geographical
remoteness has imposed some limitations on all aspects of the environmental
studies.
Some General Considerations
The relatively limited information on the microbial ass-

3
ociations in rock piles containing sulphide minerals and in the effluent
water systems suggests a considerable degree of complexity which matches
the heterogeneity of chemical composition and physical characteristics of
the milieu (Ralph, 1977). Further, the microbial populations are likely
to vary both in nature and in magnitude under the in~luence of a diversity
of factors which include the age of the heap and climatic characteristics
such as rainfall patterns and temperature. The situation in overburden
dumps is likely to be more subject to climatic factors than those in
leaching heaps with recycle of percolating solutions and other controls.
Relatively simple models of the mechanisms operating in
leaching heaps have been proposed (Harris, 1969) and have some utility
for metal recovery operations. More effective control of mineral de
gradation in rock piles might be possible if the intermeshing of geo
chemical and microbial populations in specific locations does provide
information on the physico-chemical characteristics of the milieu and
can yield clues to the events which generate such conditions.
The first steps in the studies here reported have been
(i) The enumeration of the autotrophic microorganism types known to
be associated with the degradation of sulphide minerals (acidophilic,
iron oxidisers such as Thiobacillus ferrooxidans; acidophilic sulphur
oxidisers such as T. thiooxidans; less-acidophilic sulphur oxidisers
such as T. thioparus), together with heterotrophic types.
(ii) The correlation of population level data with available in-
formation on the physico-chemical characteristics of the sample location
(pH, moisture content, water-solubles content, soluble metal ions,
organic carbon content, etc.), and with climatic factors.
(iii) The nature, distribution and magnitude of microbial populations
within the overburden heaps and their variation with climatic conditions.
The investigation to date has highlighted the difficulties
of adequately sampling rock piles and the uncertainties of current
techniques for the enumeration of some microbial types.
Methods
The layout of the Rum Jungle Mine area is shown in the map

(Figure 1). Sample sites are identifiable by the map co-ordinates.
Samples were collected in September, 1975 (Dry season), March, 1976
(Wet season) and September, 1976 (Dry season).
Water samples were either transferred directly at the
field location into pre-sterilised containers or concentrated onto
membrane filters by syringe. Water samples for chemical analysis were
pre-filtered and membrane-filtered and the filtrate stored in pre
sterilised containers. All samples were stored at 4 C as soon as
practicable. Solid materials were sampled with heat-sterilised imp
lements and immediately transferred into pre-sterilised containers.
4
Prior to examination, solid samples were ground with a sterilised mortar
and pestle, under clean air conditions, to a particle size of approximately
120 microns.
Standard methods were used for the analysis of solid samples
in respect of moisture content, water-soluble components and pH (Belly and
Brock, 1974). The content of zinc, copper and iron ions was determined in
water samples and the water-soluble components of solid samples by atomic
absorption spectroscopy.
For the microbiological examination of solid materials, a
weighed amount of the finely-ground sample was agitated with a known
volume of 9K basal medium (Silverman and Lundgren, 1959) and aliquots of
the supernatant, with appropriate dilution, used for enumeration of the
various rnicroorganism types. Water samples were either examined directly,
after appropriate dilution, or, in the case of samples concentrated by
membrane filtration in the field, the organisms were washed off the mem
brane and re-suspended.
Acidophilic, iron-oxidising organisms of the Thiobacillus
ferrooxidans type were enumerated by concentrating from the sample sus
pension ont~ a washed membrane filter to a silica gel plate impregnated
with 9K-ferrous sulphate medium (pH 2.5), and incubating at 30 C for 2-3
weeks. The reddish colonies developing from each cell were counted under
a stereo microscope. Nine plates were counted for each sample; the
results were reported as a most probable number (MPN) and standard
deviation recorded.

5 Ac:· aophilic, sulphur-oxidising organisms of T. thiooxidans
type were enume'rated by direct streaking of various dilutions of the sample
suspensions onto 9K-thiosulphate agar (pH 3.5), incubating at 30 C for one
week and counting with a colony counter. Sulphur-oxidising organisms of
the T. thioparus type were enumerated by similar procedures, using agar
plates impregnated with the thiosulphate medium (pH 6) described by
Vishniac and Santer (1957).
Heterotrophic populations were evoked and enumerated on
nutrient agar medium (pH 7.4), Czapek-Dox agar (pH 6.8), and glucose
Scott Yeast extract medium (pH 3.5), (Manning, 1975). For example:
T. denitrificans A modification of the method of Hutchinson
et al. (1965) •
Metallogenium spp. Medium & procedure recommended by Walsh &
Mitchell (1972) were used for isolating and
enumerating these microorganisms.
Anaerobic, sulphate-reducing spp. Method of Pankhurst (1971) involving
Baars medium was employed.
Nitrogen-fixing activity has been sought by use of the acetylene
reduction test {Burris, 1972).
Results and Discussions
The water and solid samples were collected in September, 1975
{dry season) and March, 1976 (wet season) from closely similar locations,and
enable comparisons to be made between some physico-chemical character
istics and the population levels of some groups of microorganisms. The
sampling of solid materials was restricted on these occasions to less
than a metre from the surface. The population levels of various micro
organism types in water samples collected during these wet and dry
seasons are-shown in Table 1, and similar data is shown in the same table
for rock samples from the 15-30 cm. depth zones. Table 2 contains in
formation on the variation of populations with depth during the wet season
(March, 1976) on White's Heap and the Sulphide Heap.
In general, the population levels are lower in the wet season
samples, the effect being more marked in the spring outflows than in the

opencuts. The reverse effect is the case of tne acidophilic hetero
trophs in Dyson's Opencut suggests an influx of organic nutrients in the
wet season. The reduction of population counts in spring samples by a
dilution effect is likely to be more marked than in opencut samples. The
reverse effect is shown in the counts for rock samples from the heaps at
depths of 15-30 cm., suggesting that at these near-surface depths during
the dry season, microbial activity is inhibited by lack of water.
In Table 2, the results of a first attempt to correlate
population levels and types with depth within the heaps are set out.
The samples were secured from holes dug with pick and shovel and pen
etration to depths greater than 70-80 ems. was not practicable by this
method. It is evident from the scatter of the counts that any depth
effect is over-ridden by the heterogeneity of the heap materials and that
aerobic conditions persist to at least the depths reached.
6
During the last dry season (September, 1976), a more in
tensive sampling of White's Heap became practicable, and mechanical equip
ment was used to cut sampling holes to a depth of 4 metres in the top of
the heap and at its base. The locations of these holes, which are
designated A to G, are shown in the map (Figure 2). Composite samples
were taken from the walls of these holes (from 15-20 cm. in), in a
vertical sequence from the bottom at 30 cm. intervals (Figure 3). Sub
sequent examination of these samples yielded the data shown in Tables
3.1 to 3.6. Direct sampling into appropriate media was carried out for
the detection of T. denitrificans and Metallogenium spp. These species
were not found in any sample, nor was any evidence for nitrogen-fixing
activity found in subsequent examinations. No growth was obtained from
any sample on the medium described by Manning (1975) for the different
iation of colony types of acidophilic iron oxidising bacteria, but·this
author's medium and method for acidophilic heterotrophs proved most use
ful.
The full evaluation of these results will not be possible
until similar sampling can be carried out at other times within the annual
climatic cycle, but some general comments can be made.
Moisture levels in top of heap samples (Holes D.E.G) are

7 ci1dracteristically variable but similar in pattern to those measured in
near-surface samples during the previous dry season. Wet season samples
are characteristically more uniform in water content with values generally
within the 15-20% range. The moisture levels of the base of heap samples
(Holes A,B,C) are clearly affected by the drainage patterns; the A hole
is near the point of e::it of a wet season spring and Band C holes are
near the highest points of the basement of the heap. As might be an
ticipated, the water-solubles content of all samples are comparatively
low; in fact, about an order of magnitude lower than in wet season
samples.
There is a rough correlation between the pH levels of samples
and the occurrence of various microorganism types. For example, the hole A
samples (av. pH 2.77) contains significant levels of acidophilic iron
oxidising and acidophilic heterotrophic organisms, and a similar situation
is evident in hole G samples (av. pH 3.0). There has been extensive
leaching in the vicinity of hole c, with considerable breakdown of rock
masses to clays and other finely divided material. The samples have a
much higher average pH (5.94) and the less acidophilic sulphur-oxidising
bacteria make up the largest microbial populations.
The complete absence of acidophilic sulphur-oxidising
organisms is an interesting feature, since earlier observations indicate
that this group is characteristically the most abundant of the auto
trophic populations in the wet season samples. The occurrence of
anaerobic sulphate-reducing organisms in the sampling holes at the base
of the heap, in the zone 70 ems. up from the basement, is of great
interest (Dugan, 1975) and suggests an anaerobic zone running through the
base of the heap. The overall distribution of aerobic microbial types,
however, suggests that the penetration of oxygen into the heap at the end
of the dry season is at least 4 metres deep over the higher parts and may
be more extensive.
Acknowledgments
The studies reported have been carried out under a research
contract with the Australian Atomic Energy Commission. The authors are
most appreciative of the financial support provided and the continuing
help and advice available from the staff of the AAEC Research Establishment.

References
Belly, R.T. & Brock, T.D., 1974. Ecology of Iron oxidising bacteria in pyritic metals associated with coal. J. Bact. 117 : 726-732.
Burris, R.H., 1972. Measurement of biological N2-fixation with 15N2 and acetylene. In : Sorokin & Kadota, Ed. Techniques for the assessment of microbial production and decomposition in fresh water, pp 3-14. Blackwell Scientific Publications, Oxford.
Davy, D.R., 1975. Run Jungle Environmental Studies. Australian Atomic Energy Commission Report, AAEC/E365.
Dugan, P.R., 1975. Bacterial ecology of strip mine areas and its relationship to the production of acidic mine drainage. Ohio Journal of Science 75 : 266-279.
Harris, J.A., 1969. Development of a theoretical approach to the heap leaching of copper sulphide ores. Proc. Aust. Inst. Min. Met. 230 81-92.
Hutchinson, M., Johnstone, I.I. & White, D., 1965. The taxonomy of certain thiobacilli. J. Gen. Microbial. 41 357-366.
Manning, H.L., 1975. New Medium for isolating iron oxidising and heterotrophic acidophilic bacteria from acid mine drainage. Appl. Microbial. 30 : 1010-1016.
Pankhurst, E.S., 1971. The isolation and enumeration of sulphate-reducing bacteria. pp 223-240. In: isolation of anaerobes. Ed. Shapton & Board. Soc. Appl. Bact. Tech. Ser. No. 5. Academic Press, London.
Ralph, B.J., 1977. Oxidative Processes, Chap. 6. To appear in : Biological Factors in Mineral Cycling. Eds. P.A. Trudinger, & D.J. Swaine. Elsevier.
Roberts, W.M.B., 1960. Mineralogy and genesis of White's ore body, Rum Jungle Uranium Fielq, Australia. Neus Jahrbuch f~r mineralogie, Abhanlungen. Bd. 94 : 868-889,
Silverman, M.P. & Lundgren, D.G., 1959. Studies on the chemoautotrophic iron bacterium Ferrobacillus ferrooxidans. 1. An improved medium and a harvesting procedure for
securing high cell yield. J. Bact. 77 : 642-647.
8

9
Vishniac, w~ & Sauter, M., 1957. The Thiobacilli. Bact. Rev. 21 195-209.
Walsh, F. & Mitchell, R.C., 1972. An acid-tolerant-iron-oxidising Metallogenium. J. Gen. Microbiol. 72 : 369-376.

):> C: ......... VI :;:o c+ (!)
-s "'O QI -s __. 0 -'• 0. PI C: ::I n
(!) ):> 0. c+ 0 ~ 3 -'• -'• c+ ...,, n ;:r ......
G) c+ C:
rn ':::T :;:o ::I (!) rn (!)
-s "'O -',
~ (!) -s 3 -'•
n VI 0 VI -3 -'• 3 0 -'• :::s VI VI -'• 0 0 -t) :::s ........
I : 5000 THE RUM JUNGLE HINE OIIAWN. II f . LOW SO'I AP'IUL 1914
~ ~ TO MO!("'' ,nc .. IIIINI ••• § §l
05 00 07 08 09 10 II 12 i, 14 15 16 17
! . T II , I
9l 94 9~ 90 i 98 _ 99! 1 oi _ 02 05
-- r j I
i-i
'1\J- I 1-- ! ':, . -+- -,- . ~~- --+-1-r-··1: -~--l--~--1---
I-
18 19 20 21 22 25 24 25 20 27 28 I - . I I
I
1 -I
I I
rH-l 29 lO
7 00 (']
) 9
57
50
55 I
§!soi-- -- __ :__ --
! ~
5
I ! . ~
~
48 >---------+-~ 41
ff
45
44
45 ,_ -
35 I I
34 1
' ni 32
51 I I
{!] 30 L
- , VI . . a1
n~1· -t-----+-+----J_ II
- 1 ·-;-I
I I
/
' 1 ·--·-- -t
- i---l- ---~ ,, t' / : . . -/ -- .
I
·1 I ••• •·~"., ... "M'" T"
I - , - - - ---+ -L-- -_, - I i
I I I -r::-:=::::
I .. , ussu , ,u" 1 Ill t ttSU ltl tUUS I I ., ,,,m " nun
"' tln)I
I ' i i I
__ -_ -~-.t,- _ --,I ~ 2~ ~ f!l I '
, 58
57
56
55
I 54
- .53
52
---- 51 I
--j 508
-----; 49
-4 48
47
46
145
- 44
-I 45 I
J 42
-- i 41
7•0B ---. 59
I __ __j 39
_I 57
36
35 .! - .. ,, .... , L' i 41· I , ,i.\f/ .... -i ,--, f I I '
--'---L--'----'---'-----'-----'-----'--...L...---'--'---'----'-----'--' _ ·· c..' _.,_-..J._....!1_...,______JI....--' _ : lli _l : . ,, ,,
- I 54
l l
l2
-1 51
90 91 92 Ol 96 97 98 99 00 01 02 05 04 05 06 07 08 09 10 If 12 ll 14 15 16 17 18 19 20 21 22 2l 24 25 26 27 28 29 50 E!J
50 8
flo•• o.,, llo••~T,.c;t.
,.,,. o .. ,,..." 1,1, ,11, .. , c .. , ..... n
@:) C)
~ ~ ~ ~
o, ...... u,_ loJ1.
S1Kwv P'oh1t
fl lr t Woll
l,..eN OlrtWol
-;. ;..n.,;1i.,·
"' ::.===---
Loou """ ' •: llwbtli. f'ii., $ ,',•;)!
llilcinl"" l""iln t. lloch 1!•
,.1.__.,,._n~ Our-"Q'-0
:~~ ... ~'1.,!!-=~-1 U -dlf~•;,.-,.,.,tr_ s,,1,., l•uMOf''l•lDrtf .,._ Holl
Lan, OoM, ..... d = C)
l ll • Nf111,rd '--111• ,: - . tOOO 'f•rd c;,i, + Oiu ,ti. Jt..-t
100 • 0 - MIO JOO ~ ......
.. 01111o•hL ,nu"' ,,!!'.~:·~~.H:\<'~~~~T~l~l.:,".~;!1~T:~, ~ .... 11' n ' ..... ,~O• to1UOI ,,, . . .. • • • . , ; l. L...._. 1110,C.,.T'( fHI IOO ,-.10 fl•W,\<'l•U llt..C:•11• 1111111, 100,1 • 1-..,,. , •• .,. HIIIO I, U>•• t .. ,, IP H H O ~
- l • ot .... • •tlto o f - . ... h4'0NO .. , _ ,,_
c-,· ·E~r-, : .. -.... ~=~·,5i,,,,,,,"~'-~ .. ,_., ....... u,,.... • ......... , •.• , ·--·--· . . • _ _ JtUiS•_ _ _ _ ___ , ___ _
....L
0
11
05 06 07 08 09 10 11 12 44 I \ 44
I I I - - - - - - ---.t '----- ---. - -- -· _.,.._,,
43 I 43 I
42 42 I I
I I I I
I
41 1 41 I
35 __________ '--______________ _.35 05 06 07 08 09 10 1'1 12
0 Indicates sampling positions . .
WHITES OVERBURDEN HEAP.
Figure 2
Top
Basement
~ Indicates sampling points.
WHITE"S OVERBURDEN HEAP
SAMPLING PATTERN.
( Not to scale J
Figure 3
12
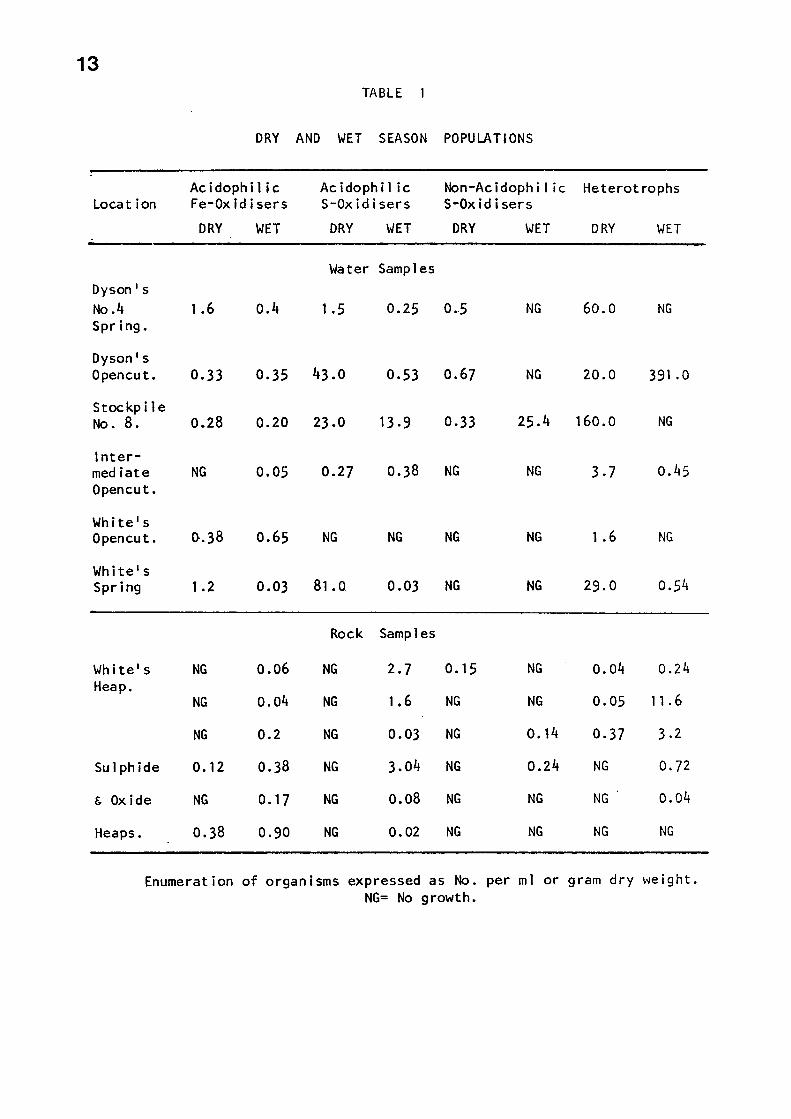
13 TABLE 1
DRY AND WET SEASON POPULATIONS
Acidophilic Acidophil ic Non-Ac i doph i l i c Heterot rophs Location Fe-Oxidisers S-Oxidisers S-Oxidisers
DRY WET DRY WET DRY WET DRY WET
Water Samples Dyson' s No .4 1.6 0.4 1.5 0.25 0.-5 NG 60.0 NG Spring.
Dyson's Opencut. 0.33 o. 35 43.0 0.53 0.67 NG 20.0 391 .o
Stockpile No. 8. 0.28 0.20 23.0 13.9 0.33 25.4 160.0 NG
Inter-mediate NG 0.05 0.27 0.38 NG NG 3.7 0.45 Open cut.
White's Opencut. 0-.38 0.65 NG NG NG NG 1.6 NG
White's Spring 1.2 0.03 81.0 0.03 NG NG 29.0 0.54
Rock Samp 1 es
White's NG 0.06 NG 2.7 o. 15 NG 0.04 O .24 Heap.
NG 0.04 NG 1.6 NG NG 0.05 11. 6
NG 0.2 NG 0.03 NG o. 14 0.37 3,2
Sulphide o. 12 0.38 NG 3.04 NG 0.24 NG 0.72
& Oxide NG o. 17 NG 0.08 NG NG NG 0.04
Heaps. 0.38 0.90 NG 0.02 NG NG NG NG
Enumeration of organisms expressed as No. per ml or gram dry \-Je i ght. NG= No growth.
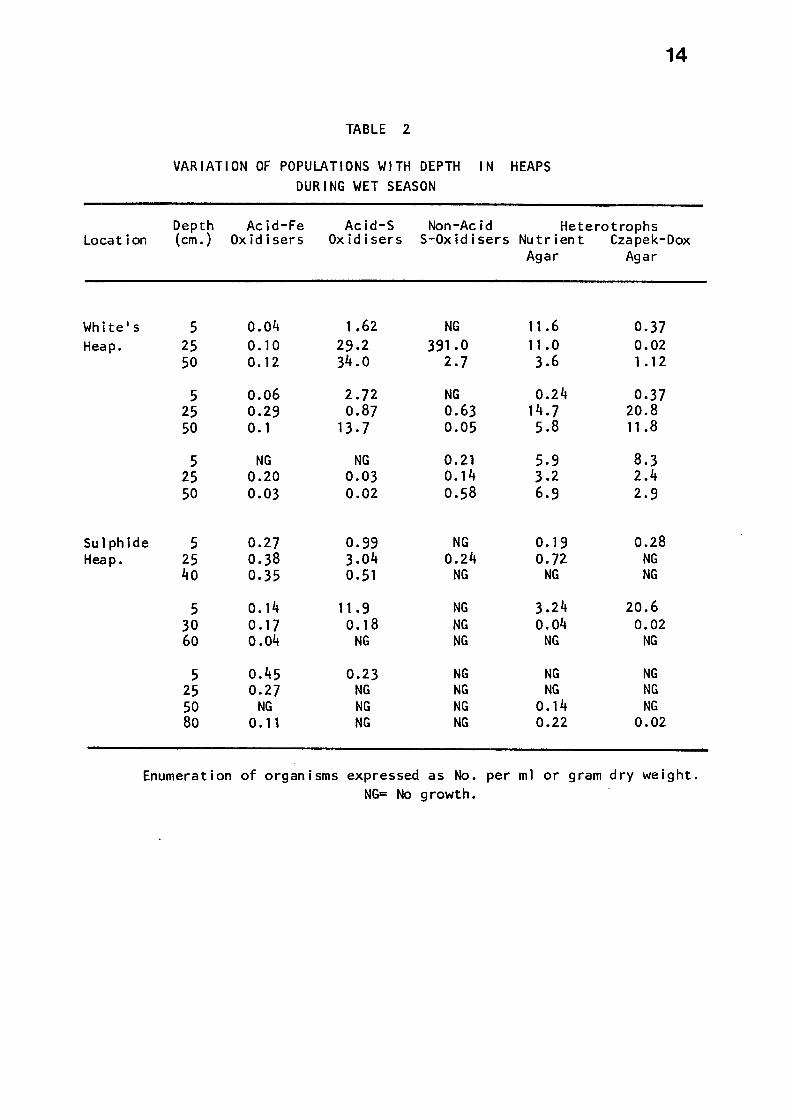
Location
White's Heap.
Sulphide Heap.
TABLE 2
VARIATION OF POPULATIONS WITH DEPTH IN HEAPS DURING WET SEASON
14
Depth Acid-Fe Acid-S Non-Acid Heterotrophs (cm.) Oxidisers Oxidisers $-Oxidisers Nutrient Czapek-Dox
Agar Agar
5 0.04 1.62 NG 11.6 0.37 25 o. 10 29.2 391.0 11.0 0.02 50 0.12 34 .0 2.7 3.6 1. 12
5 0.06 2.72 NG 0.24 0.37 25 0.29 0.87 0.63 14. 7 20.8 50 o. 1 13,7 0.05 5.8 11.8
5 NG NG 0.21 5,9 8.3 25 0.20 0.03 0. 14 3.2 2.4 50 0.03 0.02 0.58 6.9 2.9
5 0.27 0.99 NG o. 19 0.28 25 0.38 3.04 0.24 0.72 NG 40 0.35 0.51 NG NG NG
5 o. 14 11.9 NG 3.24 20.6 30 O .17 o. 18 NG 0.04 0.02 60 0.04 NG NG NG NG
5 0.45 0.23 NG NG NG 25 0.27 NG NG NG NG 50 NG NG NG o. 14 NG 80 0. 1 t NG NG 0.22 0.02
Enumeration of organisms expressed as No. per ml or gram dry weight. NG= No growth.
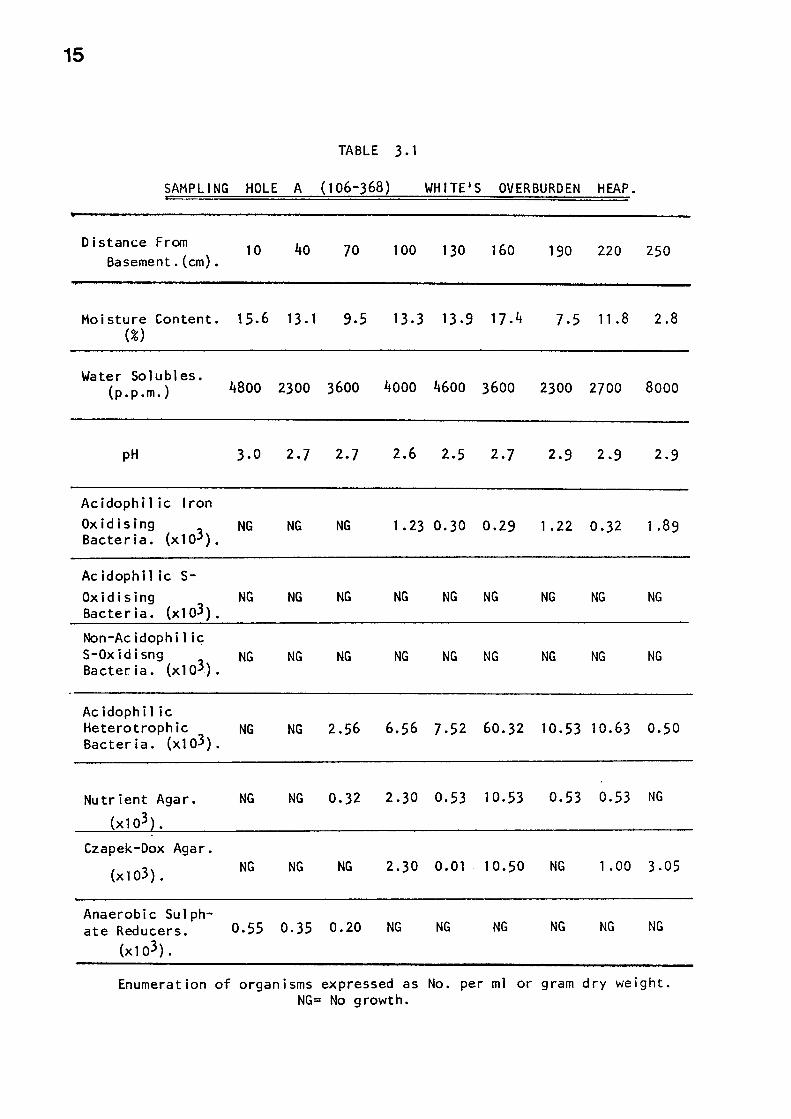
15
TABLE 3. 1
SAMPLING HOLE A (106-368) WHITE'S OVERBURDEN HEAP.
Distanc;e From Basement. (cm).
10 40 70 100 130 160
Moisture Content. 15.6 13.1 9.5 13.3 13.9 17.4 (%)
190 220 250
7.5 11.8 2.8
Water So 1 ubl es. ( ) 4800 2300 3600 4000 4600 3600 2300 2700 8000 p.p.m.
pH
Ac idoph i 1 ic Iron Oxidising
(x 103). Bacteria.
Acidophil ic s-Oxidising Bacteria. (x103).
Non-Acidophil ic S-Oxidisng
(x103). Bacteria.
Acidophil ic Heterotrophic Bacteria. (x103).
Nutrient Agar.
(x103 .
Czapek-Dox Agar.
(x l o3) .
Anaerobic Sulph-
3.0
NG
NG
NG
NG
NG
NG
2.7 2.7 2.6 2.5 2.7 2.9 2.9 2.9
NG NG 1.23 0.30 0.29 1.22 0.32 1.89
NG NG NG NG NG NG NG NG
NG NG NG NG NG NG NG NG
NG 2.56 6.56 7.52 60.32 10.53 10.63 0.50
NG 0.32 2.30 0,53 10.53 0.53 0.53 NG
NG NG 2.30 0.01 10.50 NG 1.00 3,05
ate Reducers. 0.55 0.35 0.20 NG NG NG NG NG NG
(xl o3).
Enumeration of organisms expressed as No. per ml or gram dry weight. NG= No growth.
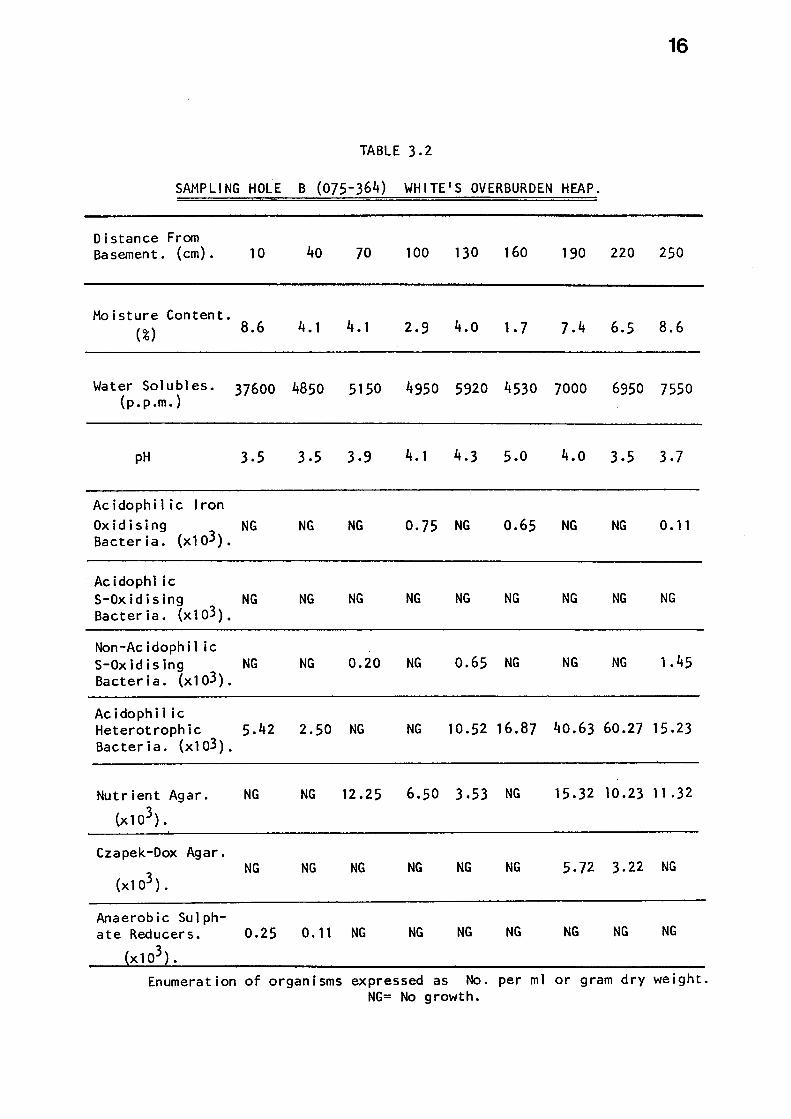
16
TABLE 3,2
SAMPLING HOLE B (075-364) WHITE'S OVERBURDEN HEAP.
Distance From Basement. (cm).
Moisture Content. (%)
Water Solubles. (p.p.m.)
pH
Acidophil ic Iron Oxidising
(xl o3). Bacteria.
Acidophl ic S-Oxidising Bacteria. (x 103).
Non-Acidophil ic S-Oxidising Bacteria. (x 1 o3).
Ac i do phi 1 i c Heterotrophic Bacteria. (xl o3).
Nutrient Agar.
(x103).
Czapek-Dox Agar.
(xl o3).
Anaerobic Sulph-ate Reducers.
X 103).
10 40
8.6 4. 1
37600 4850
3,5 3.5
NG NG
NG NG
NG NG
5,42 2.50
NG NG
NG NG
0.25 0.11
Enumeration of organisms
70 100 130 160 190 220 250
4. 1 2.9 4.o 1.7 7,4 6.5 8.6
5150 4950 5920 4530 7000 6950 7550
3.9 4. 1 4.3 5.0 4.o 3,5 3,7
NG 0.75 NG 0.65 NG NG 0. 11
NG NG NG NG NG NG NG
0.20 NG 0.65 NG NG NG t.45
NG NG 10.52 16.87 40.63 60.27 15.23
12.25 6.50 3,53 NG 15 .32 10.23 11 ,32
NG NG NG NG 5.72 3.22 NG
NG NG NG NG NG NG NG
expressed as No. per ml or gram dry weight. NG= No growth.

17
TABLE 3,3
SAMPLING HOLE C (071-368) WHITE'S OVERBURDEN HEAP.
Distance From Ba semen t • (cm) . 10 40 70 100 130 160 190 220 250
Moisture Content. 14.3 15.1 14.8 14.4 15.2 16.5 17,4 16.5 16. 1 (%) •
Water Solubles. (p.p.m.) 3500 5100 2500 3051 5183 3915 5261 5578 6098
pH 6.5 6.5 4.5 4.8 4.5 6.5 6.6 6.9 6.9
Ac idoph i 1 ic Iron Oxidising
(x 1 o3). NG NG NG
Bacteria. 0.03 NG NG NG 0.02 NG
Acidophil ic s-Oxidising
(x 103). NG NG NG NG NG NG NG NG NG
Bacteria.
Non-Ac i doph i I ic S-Oxidising
(x103). NG NG 0.57 1.65 2.57 3.75 NG 12.58 11.58
Bacteria.
Acidophil ic Heterotrophic Bacteria. (x103).
NG NG NG NG NG NG 0.01 NG NG
Nutrient Agar. NG NG NG 1.58 0.76 NG NG _o. 31 0.10
(x 1 o3) •
Czapek-Dox Agar. NG NG NG NG NG 0.03 0.02 0.21 NG (x103).
Anaerobic Sulph-ate Reducers. 0.01 0.23 NG NG NG NG NG NG NG
x103
Enumeration of organisms expressed as No. per ml or gram dry weight. NG= No growth.
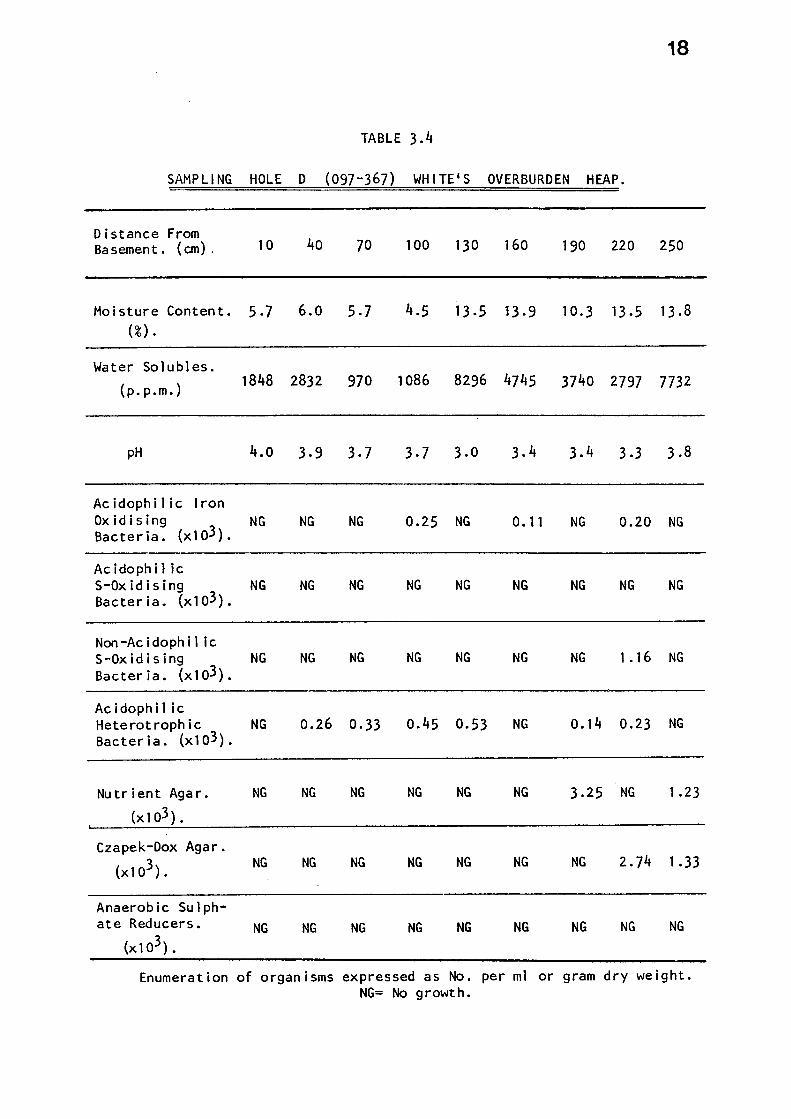
18
TABLE 3.4
SAMPLING HOLE D (097-367) WHITE'S OVERBURDEN HEAP.
Distance From Basement. (cm). 10 40 70
Moisture Content. 5.7 6.0 5-7 (%) •
100 130 160 190 220 250
4.5 13.5 13.9 10.3 13.5 13.8
Water Solubles.
(p. p.m.) 1848 2832 970 1086 8296 4745 3740 2797 7732
pH 4.0 3,9 3-7 3.7 3.0 3.4 3.4 3,3 3,8
Ac idophi 1 ic Iron Oxidising
(x103). NG NG NG 0.25 NG 0. 11 NG 0.20 NG
Bacteria.
Ac idoph i 1 ic S-Oxidising NG NG NG NG NG NG NG NG NG Bacteria. (x103).
Non-Ac idoph i 1 ic S-Oxidising NG NG NG NG NG NG NG 1.16 NG Bacteria. (x 103).
Acidophil ic Heterotrophic NG Bacteria. (x103).
0.26 0.33 o.45 0.53 NG o. 14 0.23 NG
Nutrient Agar. NG NG NG NG NG NG 3,25 NG 1.23
(x 103).
Czapek-Dox Agar.
(x103). NG NG NG NG NG NG NG 2.74 1.33
Anaerobic Sulph-ate Reducers. NG NG NG NG NG NG NG NG NG
(x 1 o3).
Enumeration of organisms expressed as No. per ml or gram dry weight. NG= No growth.
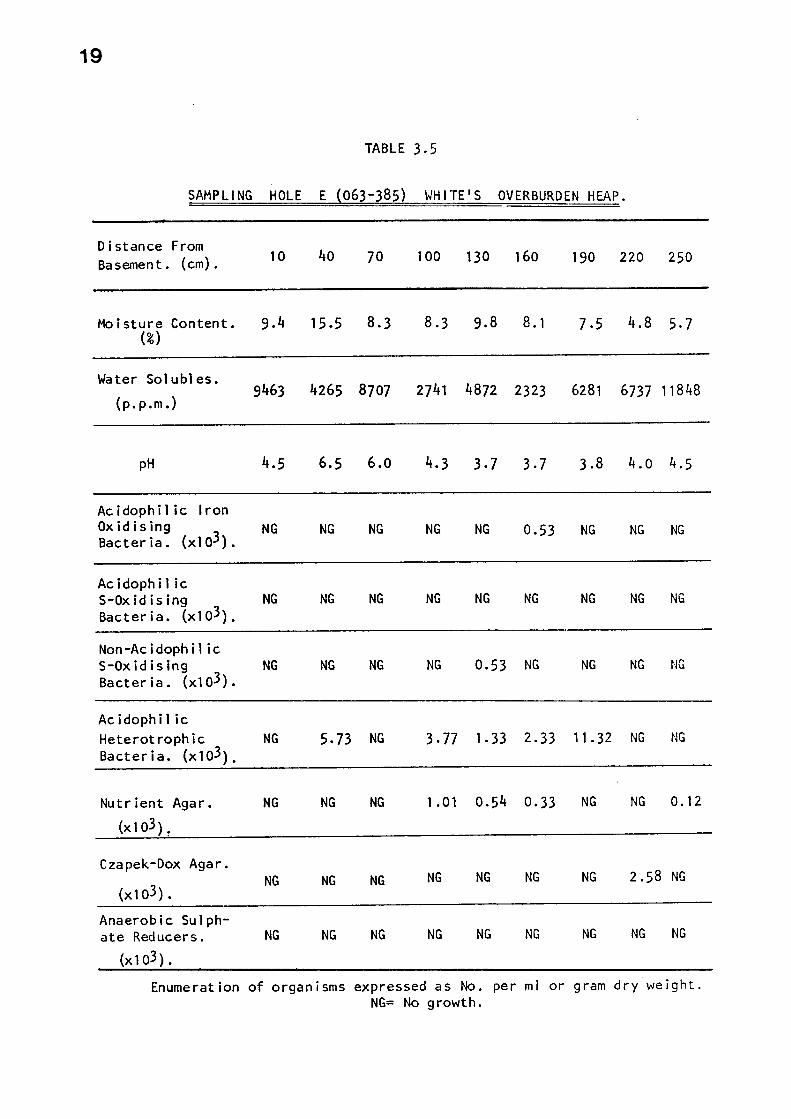
19
TABLE 3.5
SAMPLING HOLE E (063-385) WHITE'S OVERBURDEN HEAP.
Distance From Basement. (cm). 10 40 70 100 130 160 190 220 250
Moisture Content. 9.4 15.5 8.3 (%)
8.3 9.8 8.1 7.5 4.8 5.7
Water So 1 u b 1 es.
(p.p.m.)
pH
Acidophil ic Iron Oxidising
(x103). Bacteria.
Acidophil ic S-Oxidising Bacteria. (x103).
Non-Acidophil ic S-Oxidising Bacteria. (x103).
Acidophil ic Heterotroph ic Bacteria. (x103).
Nutrient Agar.
(x103).
C za pek-Dox Agar •
(x103).
Anaerobic Sulph-ate Reducers.
(x 1 o3).
Enumeration
9463 4265 8707 2741 4872 2323 6281 6737 11848
4.5 6.5 6.o 4.3 3.7 3.7 3.8 4.0 4.5
NG NG NG NG NG 0.53 NG NG NG
NG NG NG NG NG NG NG NG NG
NG NG NG NG 0.53 NG NG NG NG
NG 5,73 NG 3. 77 1.33 2.33 11. 32 NG NG
NG NG NG 1.01 0.54 0.33 NG NG 0. 12
NG NG NG NG NG NG NG 2 ,58 NG
NG NG NG NG NG NG NG NG NG
of organisms expressed as No. per ml or gram dry weight. NG= No growth.
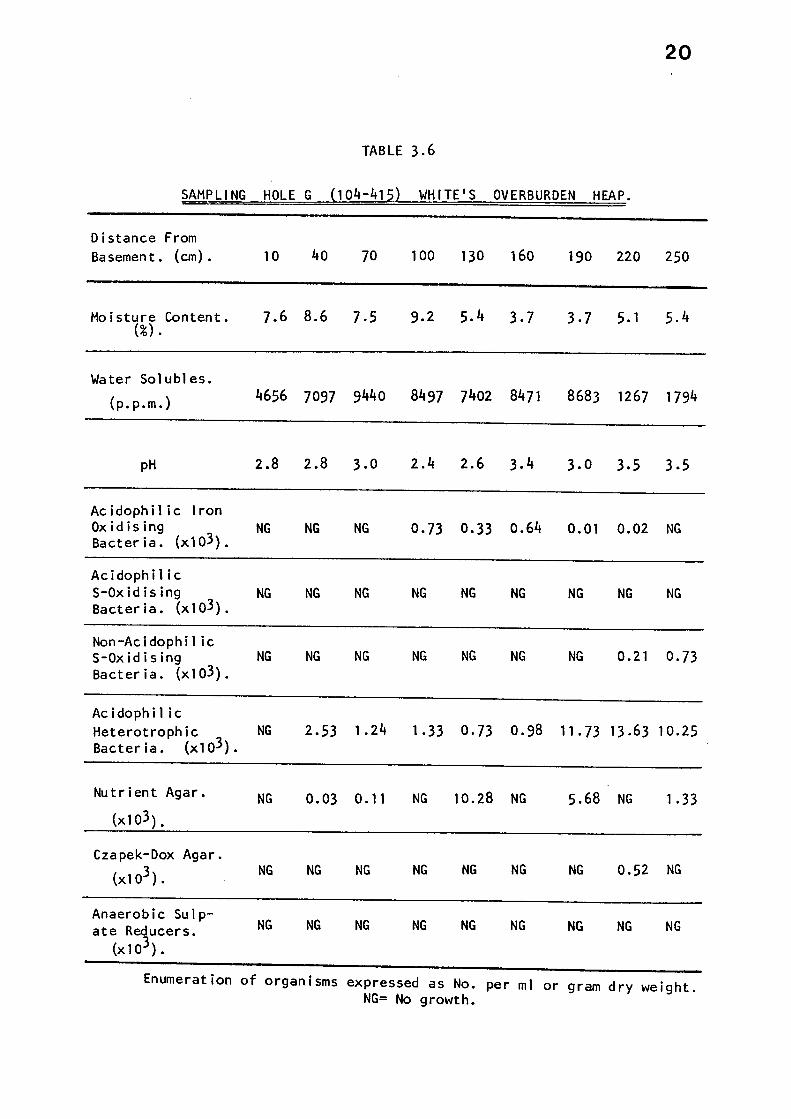
20
TABLE 3.6
SAMPLING HOLE G (104-415) WHITE'S OVERBURDEN HEAP.
Distance From Basement. (cm).
Moisture Content. (%) •
Water Solubles.
(p.p.m.)
pH
Acidophil ic Iron Oxidising Bacteria. (x103).
Ac i doph i l i c S-Oxidising Bacteria. (x 1 o3) •
Non-Acidophil ic $-Oxidising Bacteria. (x 1 o3).
Acidophil ic Heterotrophic 3 Bacteria. (x 10 ) •
Nutrient Agar.
(x103).
Czapek-Dox Agar.
(x103).
Anaerobic Su 1 p-ate Reducers.
(x 1 o3).
10 40 70 100 130 160 190 220 250
7.6 8.6 7.5 9.2 5.4 3,7 3.7 5.1 5.4
4656 7097 9440 8497 7402 8471 8683 1267 1794
2.8 2.8 3.0 2.4 2.6 3.4 3.0 3.5 3.5
NG NG NG 0.73 0.33 o.64 0.01 0.02 NG
NG NG NG NG NG NG NG NG NG
NG NG NG NG NG NG NG 0.21 0.73
NG 2.53 1.24 1.33 0.73 0.98 11.73 13.63 10.25
NG 0.03 0. 11 NG 10.28 NG 5.68 NG 1.33
NG NG NG NG NG NG NG 0.52 NG
NG NG NG NG NG NG NG NG NG
Enumeration of organisms expressed as No. per ml or gram dry weight. NG= No growth.




















