
The Late Miocene hominoids Ouranopithecus and Graecopithecus. Implications about their relationships...
14
Article original The Late Miocene hominoids Ouranopithecus and Graecopithecus. Implications about their relationships and taxonomy Les hominoïdes du Miocène supérieur Ouranopithecus et Graecopithecus. Implications de leurs relations et taxonomie George D. Koufos a, *, Louis de Bonis b a Laboratory of Geology and Palaeontology, Department of Geology, Aristotle University of Thessaloniki, 54124 Thessaloniki, Greece b Laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine, Université de Poitiers, 40, avenue du recteur Pineau, 86022 Poitiers, France Received 15 October 2004; accepted 27 May 2005 Abstract Two Late Miocene hominoids are known from Greece. The first, Graecopithecus freybergi, is known by a single mandible with the worn m2 from the locality of Pyrgos Vassilissis, near Athens. The other, Ouranopithecus macedoniensis, is known from Axios Valley and Chalkidiki (Macedonia, Greece) by a partial skull and a set of maxillary and mandibular remains. Some authors consider these two hominoids as synonyms and in the present article a detailed comparison of them is given. The morphology and size of the symphysis, the more robust mandible, the more open dental arcade of Ouranopithecus distinguishes it clearly from Graecopithecus. Moreover, the incompleteness of the mandible of Graecopithecus with the doubtful morphology and size, the limited material and the uncertain geological age of the locality cannot allow precise and clear comparisons with the rest extant and extinct hominoids. Thus in our opinion there are not enough data to support the similarity, and therefore, the synonymy of the two genera. The Pyrgos mandible must remain as a separate and isolated genus with one species, which only includes this sole mandible. © 2005 Elsevier SAS. All rights reserved. * Corresponding author. E-mail address: [email protected] (G.D. Koufos). Annales de Paléontologie 91 (2005) 227–240 http://france.elsevier.com/direct/ANNPAL/ 0753-3969/$ - see front matter © 2005 Elsevier SAS. All rights reserved. doi:10.1016/j.annpal.2005.05.001
Transcript of The Late Miocene hominoids Ouranopithecus and Graecopithecus. Implications about their relationships...

Article original
The Late Miocene hominoids Ouranopithecusand Graecopithecus. Implications about their
relationships and taxonomy
Les hominoïdes du Miocène supérieurOuranopithecus et Graecopithecus. Implications de
leurs relations et taxonomie
George D. Koufos a,*, Louis de Bonis b
a Laboratory of Geology and Palaeontology, Department of Geology, Aristotle University of Thessaloniki,54124 Thessaloniki, Greece
b Laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine, Université de Poitiers,40, avenue du recteur Pineau, 86022 Poitiers, France
Received 15 October 2004; accepted 27 May 2005
Abstract
Two Late Miocene hominoids are known from Greece. The first, Graecopithecus freybergi, isknown by a single mandible with the worn m2 from the locality of Pyrgos Vassilissis, near Athens.The other, Ouranopithecus macedoniensis, is known from Axios Valley and Chalkidiki (Macedonia,Greece) by a partial skull and a set of maxillary and mandibular remains. Some authors considerthese two hominoids as synonyms and in the present article a detailed comparison of them is given.The morphology and size of the symphysis, the more robust mandible, the more open dental arcadeof Ouranopithecus distinguishes it clearly from Graecopithecus. Moreover, the incompleteness ofthe mandible of Graecopithecus with the doubtful morphology and size, the limited material and theuncertain geological age of the locality cannot allow precise and clear comparisons with the restextant and extinct hominoids. Thus in our opinion there are not enough data to support the similarity,and therefore, the synonymy of the two genera. The Pyrgos mandible must remain as a separate andisolated genus with one species, which only includes this sole mandible.© 2005 Elsevier SAS. All rights reserved.
* Corresponding author.E-mail address: [email protected] (G.D. Koufos).
Annales de Paléontologie 91 (2005) 227–240
http://france.elsevier.com/direct/ANNPAL/
0753-3969/$ - see front matter © 2005 Elsevier SAS. All rights reserved.doi:10.1016/j.annpal.2005.05.001

Résumé
Deux genres de primates hominoïdes ont été décrits dans le Miocène supérieur de Grèce. L’un,Graecopithecus freybergi, provient de la localité de Pyrgos Vassilissis, près d’Athènes, dont l’âgen’est pas connu avec précision. Il n’est représenté que par une mandibule portant seulement unem2 complètement usée et les racines d’autres dents. L’autre, Ouranopithecus macedoniensis, pro-vient de gisements situés dans la basse vallée de l’Axios et en Chalcidique (Macédoine, Grèce) quisont datés du Vallésien supérieur. Il est représenté par une portion de crâne et par tout un ensemble demaxillaires, de mandibules et de dents isolées. Certains auteurs ont estimé que les deux genres étaientsynonymes. Le but de cet article est de comparer avec soin le matériel appartenant à ces deux taxons,tout au moins pour les parties présentes sur la mandibule de Pyrgos Vassilisssis. On remarque que lamorphologie de la symphyse est différente, que la mandibule d’Ouranopithecus est plus robuste avecune arcade dentaire plus ouverte et que les deux ensembles se distinguent clairement. Étant donnél’état de conservation de la mandibule de Graecopithecus il n’est pas possible de pousser les inves-tigations plus loin. Cependant, il nous semble que les données pour rassembler les deux genres sontinsuffisantes et que leur rapprochement par certains auteurs découle surtout de leur positiongéographique en Grèce bien que le site de Pyrgos-Vassilis soit situé à plusieurs centaines de kilo-mètres des gisements de Macédoine. Il est donc préférable de conserver Graecopithecus comme ungenre séparé contenant une seule espèce fondée sur une unique mandibule incomplète.© 2005 Elsevier SAS. All rights reserved.
Keywords: Ouranopithecus; Graecopithecus; Hominoids; Greece; Relationships; Taxonomy
Mots clés : Ouranopithecus ; Graecopithecus ; Hominoïdes ; Grèce ; Relations ; Taxonomie
1. Introduction
The hominoid primate Ouranopithecus is widely known from Macedonia, Greece by aquite rich sample of cranial and dental remains. It was originally found in the locality“Ravin de la Pluie” (RPl) of Axios Valley; later it was recognized in the localities“Xirochori-1” (XIR) of Axios Valley too and “Nikiti-1” (NKT), Chalkidiki Peninsula (deBonis et al., 1974, 1990; Koufos, 1993, 1995). All Ouranopithecus bearing localities havebeen dated to Late Vallesian, MN 10 with minor age differences between them. The mag-netostratigraphic data suggest an age ~9.6 Ma for XIR and ~9.3 Ma for RPl (de Bonis andKoufos, 1999; Sen et al., 2000). The faunal data from NKT indicate that it is younger thanthe previous two with an age between 9.3 and 8.7 Ma (Koufos, 2000). The study and analy-sis of these faunas indicate a relatively more open environment (de Bonis et al., 1992; deBonis and Koufos, 1999; Koufos, in press). The thick enamel and the dental microwear ofOuranopithecus suggest a hard-food eater (Ungar, 1996).
A hominoid mandible has been found in the Greek locality “Pyrgos Vassilissis”, nearAthens. It was originally referred to the cercopithecoid Mesopithecus pentelicus but later itwas described under the name Graecopithecus freybergi (von Koenigswald, 1972). Theassociated faunule includes the following species: Choerolophodon pentelici, Hipparionmediterraneum? “Dicerorhinus orientalis”, Bohlinia attica, Helladotherium duvernoyi?Tragoportax amalthea, Gazella? deperdita, Gazella? cf. gaudryi. This fauna cannot allowa more precise age than Late Miocene (Koufos, 1995; de Bonis and Koufos, 1999). The
228 G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

Pyrgos mandible lacks the external symphysis, the inferior limits of the mandibular bodiesare damaged and the anterior dentition is absent. The cheek teeth are almost absent; theright worn m2, the lingual surface of the right p4 and the base of the crown of the rightm1 are preserved. The occlusal border of the right m2 is also partially damaged.
The first unearthed mandible from RPl was described as Dryopithecus macedoniensis n.sp., following the nomenclature of that time and distinguishing the RPl mandible from theknown hominoid species (de Bonis et al., 1974). The material found during the later yearsallowed the erection of the new genus Ouranopithecus (de Bonis and Melentis, 1977), aname used by the authors until now. The species Ouranopithecus macedoniensis and Grae-copithecus. freybergi have been synonymized and included both to Ankarapithecus meteai;the genus Ankarapithecus was considered as synonym of Sivapithecus and thus all thesetaxa included to Sivapithecus meteai (Martin and Andrews, 1984). Later on, Ouranopith-ecus and Graecopithecus were separated from Sivapithecus and referred as G. freybergi(Andrews et al., 1996), while Cameron (1997) distinguished two species: G. freybergi and“G”. macedoniensis. Few years later the name Ankarapithecus became valuable againseparated from Sivapithecus (Alpagut et al., 1996; Begun and Gülec, 1998). Thus, there isa puzzle of the generic and specific names of the Miocene hominoids of Greece and Turkey.We believe that the Pyrgos mandible is so incomplete that it cannot provide the availablemetrical and morphological characters, in order to synonymize it with any of the knownhominoids. In fact, the Ouranopithecus material was found in Greece, as Graecopithecus,but this does not mean that any Greek hominoid must belong to the latter genus. The lim-ited morphology, size and age of the Pyrgos mandible cannot allow certain comparisonsand results about this. In the present article we shall try to show the limited morphologicalcharacters and comparisons of Graecopithecus, as well as to find the differences betweenthe two genera. This article has been presented in the 5th International Congress on EasternMediterranean Geology, Thessaloniki, April 2004.
2. Comparison of Graecopithecus and Ouranopithecus
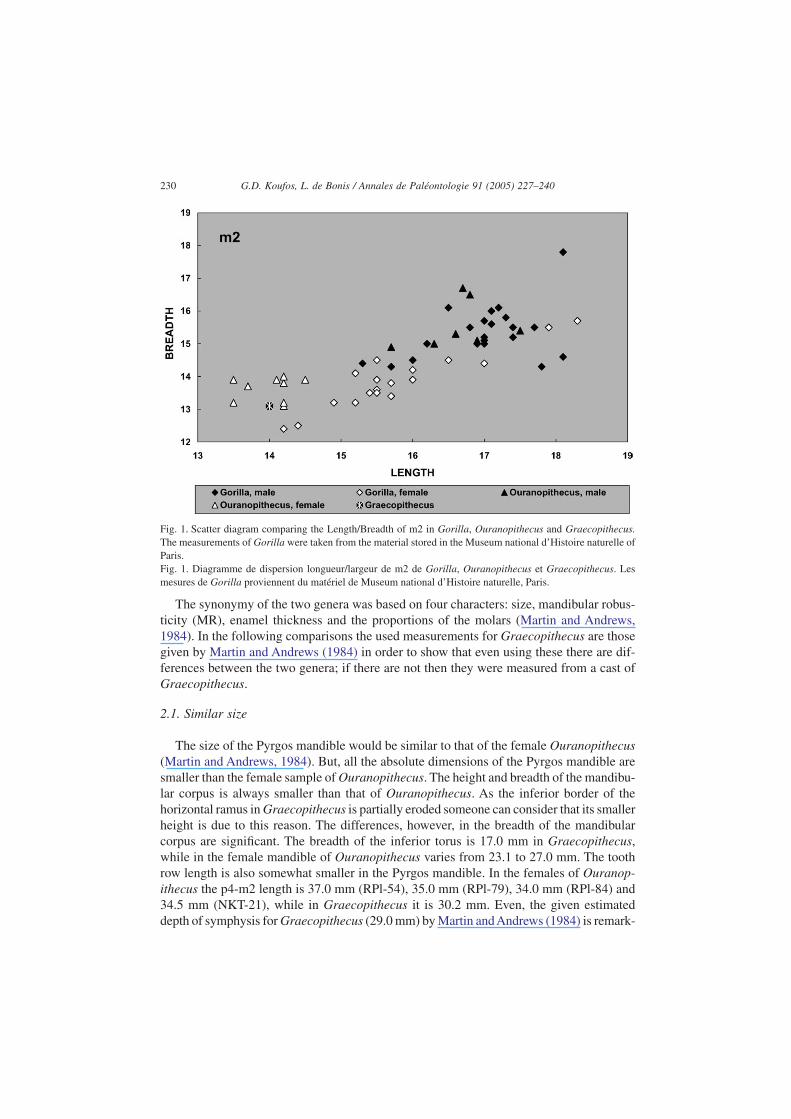
The Pyrgos mandible is referred as female from the size of its premolars and the roots ofthe canine (Martin and Andrews, 1984) but it is impossible to be sure from a single speci-men. The root of the canines corresponds to a relatively small canine and this is an indica-tion for a female individual. But, this is not clear, as we did not know the size of the malecanine. The more complete m2 can probably provide other indications about the sex of thePyrgos mandible. In the recent gorillas there is a clear sexual dimorphism in the size of themolars, which is also observed in Ouranopithecus sample (Fig. 1). The m2 of Graecopith-ecus is very close to the female group of both Gorilla and Ouranopithecus, but this doesnot mean that it is female as the male’s size is unknown. If Graecopithecus has smaller sizethan Ouranopithecus then the known mandible could belong to a male individual withsmaller canines and m2. Although, there are some indications that the Pyrgos mandiblebelongs to a female, its sex is uncertain. Besides this doubtful assumption, we shall con-tinue to consider the Pyrgos mandible as female in the following comparison in order toshow that even if this is true there are significant differences from the female mandible ofOuranopithecus. Otherwise, its remarkable size difference from the male Ouranopithecusmandible is quite enough indication for the distinction of the two genera.
229G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240
The synonymy of the two genera was based on four characters: size, mandibular robus-ticity (MR), enamel thickness and the proportions of the molars (Martin and Andrews,1984). In the following comparisons the used measurements for Graecopithecus are thosegiven by Martin and Andrews (1984) in order to show that even using these there are dif-ferences between the two genera; if there are not then they were measured from a cast ofGraecopithecus.
2.1. Similar size
The size of the Pyrgos mandible would be similar to that of the female Ouranopithecus(Martin and Andrews, 1984). But, all the absolute dimensions of the Pyrgos mandible aresmaller than the female sample of Ouranopithecus. The height and breadth of the mandibu-lar corpus is always smaller than that of Ouranopithecus. As the inferior border of thehorizontal ramus in Graecopithecus is partially eroded someone can consider that its smallerheight is due to this reason. The differences, however, in the breadth of the mandibularcorpus are significant. The breadth of the inferior torus is 17.0 mm in Graecopithecus,while in the female mandible of Ouranopithecus varies from 23.1 to 27.0 mm. The toothrow length is also somewhat smaller in the Pyrgos mandible. In the females of Ouranop-ithecus the p4-m2 length is 37.0 mm (RPl-54), 35.0 mm (RPl-79), 34.0 mm (RPl-84) and34.5 mm (NKT-21), while in Graecopithecus it is 30.2 mm. Even, the given estimateddepth of symphysis for Graecopithecus (29.0 mm) by Martin andAndrews (1984) is remark-
Fig. 1. Scatter diagram comparing the Length/Breadth of m2 in Gorilla, Ouranopithecus and Graecopithecus.The measurements of Gorilla were taken from the material stored in the Museum national d’Histoire naturelle ofParis.Fig. 1. Diagramme de dispersion longueur/largeur de m2 de Gorilla, Ouranopithecus et Graecopithecus. Lesmesures de Gorilla proviennent du matériel de Museum national d’Histoire naturelle, Paris.
230 G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

ably shorter than that of the female mandible of Ouranopithecus; in RPl-54 the internal oneis 40.1 mm and the external 43.0 mm, while in RPl-79 it is 41.2 and 45.0 mm, respectively.No more measurements can be taken from the Pyrgos mandible for further size compari-son. Someone can consider that the Pyrgos mandible seems to have similar size to thefemale Ouranopithecus, but there are some significant size differences in the symphysisand tooth row length. These size differences can be considered as intra-specific variations,but there are other clearer morphological differences. The comparison of the size of the twomandibles is quite doubtful too, as the Pyrgos mandible cannot provide certain measure-ments because it is damaged or eroded.
2.2. Mandibular Robusticity (MR)
The MR (thickness of body/depth of body) of the two genera would be similar (Martinand Andrews, 1984). This similarity cannot be proved, as the MR of Graecopithecus can-not be calculated accurately, as the height of the horizontal ramus is not reliable. Althoughthis problem exists, we shall try to give a comparison using the measurements given byMartin and Andrews (1984). The indices of the MR were calculated below each tooth andthey are given in Fig. 2. In this figure the RPl material is given, by three lines (male, female
Fig. 2. Comparison of MR in various late Miocene and recent hominoids, as well as A. afarensis. The sample ofSivapithecus includes the specimens YPM-9564 (male), YPM-4857 (female), GSI-18039 (male), AMNH-19413(female), and GSP-15000 (male) of S. indicus, and YPM-13811 (sex unknown) of S. sivalensis.Fig. 2. Comparaison de la robustesse des mandibules chez quelques hominoïdes miocènes et actuels et chez A.afarensis. Dans l’échantillon Sivapithecus sont placés les spécimens YPM-9564 (mâle), YPM-4857 (femelle),GSI-18039 (mâle), AMNH-19413 (femelle), et GSP-15000 (mâle) de S. indicus, alors que YPM-13811 (sexeindéterminé) de S. sivalensis.
231G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

and mixed) in order to be possible a comparison with the rest hominoids. The Pyrgos man-dible has smaller robusticity indices than Ouranopithecus, being remarkably smaller fromthe female ones. This means that the Pyrgos mandible is very gracile. It is worth mentionedthat the indices of the Pyrgos mandible must be somewhat smaller than the given ones asthe depth of the mandibular body is somewhat larger. Because the inferior border of thebody is damaged and weathered and no accurate measurements can be taken (Martin andAndrews, 1984; Cameron, 1997). In the right horizontal ramus the inferior border is pre-served in a small part below m3 (Von Koenigswald, 1972), a feature, which is clear in thecast of the Pyrgos mandible. The MR at m3, which is probably more reliable, is clearlysmaller than that of Ouranopithecus (Fig. 2). The female mandible of Ankarapithecus seemsto be very robust, especially below m3, and it is separated from the others (Fig. 2). Thismandible is female and thus it is very close to the RPl female one. Dryopithecus mandibleis slightly slender than that of Ouranopithecus. The mandible of Proconsul is quite robustand in this feature it is close to Ouranopithecus mean. Sivapithecus has quite robust man-dible in front, but it is being slenderer in its posterior part and in this feature it is close to therecent gorilla and chimp. The indices of the MR for Australopithecus afarensis are largerthan the mean of Ouranopithecus indicating a more robust mandible.
The comparison of the MR of the Pyrgos mandible suggests that: (a) it has an isolatedposition among the rest Miocene hominoids, and (b) it is separated from Ouranopithecusand cannot be included in the same taxon. The mandible of Graecopithecus seems to bequite gracile, but as the measurements of its dimensions are doubtful the comparison of itsMR leads to questionable assumptions which cannot help to its systematic determination.
2.3. Enamel thickness
The enamel thickness of Graecopithecus can be seen in the lingual surface of the solepreserved m2, and it seems to be rather thick. Its thickness varies from 1.7 to 2.0 mm. In thefemale m2 of Ouranopithecus the enamel thickness is 1.2–1.6 mm (RPl-79), 1.3–1.8 mm(RPl-84) and 1.6–1.7 mm (NKT-21), while in the male m2 it is 2.1–2.3 mm (RPl-74) and2.2–2.3 mm (RPl-76). Concerning this character the Pyrgos mandible is close to Ourano-pithecus. But, such a character is expected for Graecopithecus as the palaeoenvironment inEastern Mediterranean during Late Miocene was relatively open and the available foodwas hard (de Bonis et al., 1992; Koufos, in press). Thus, all the known Late Miocene homi-noids in Southeastern Eurasia (Ankarapithecus, Sivapithecus, Ouranopithecus, Graecop-ithecus) have thick-enameled teeth.
2.4. Proportions of the molars
The enlarged m3 is counted as synapomorphy and its shared presence in the two Greekgenera indicates relationship; the massiveness of the m2 crown of Graecopithecus is relatedwith an extremely gracile mandible (Martin and Andrews, 1984). However, some old homi-noids like Proconsul have large m3. The enlargement of the Graecopithecus m3 is esti-mated from the root, as both teeth are broken. So, the result is questionable as no possibilityto measure the real tooth’s dimensions. However, the m2 index breadth × 100/length forGraecopithecus is closer to the mean value for Ouranopithecus and smaller than the female’s
232 G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

value (Table 1). But, this index is also into the ranges of variation and close to the meanvalue of Ankarapithecus and Sivapithecus, as well as near the minimum values of maleGorilla. The Pyrgos m2 can proportionally fit with several genera and consequently theproportions of the molars cannot be used to estimate similarities or differences between thetwo genera. On the other hand the Pyrgos mandible is the more gracile one among thehominoids (Fig. 2).
Besides the above-mentioned characters there are some more which reinforce the dis-tinction of the two genera.
2.5. Shape of the internal symphysis
The internal symphysis of Graecopithecus is partially preserved; its border near theincisors is absent (Fig. 3). Martin and Andrews (1984) reported that it is “relatively verticaland straight as in Ouranopithecus and unlike Sivapithecus, and there was apparently nostrong torus developed”. In Ouranopithecus, however, the internal symphysis is not straightand vertical but it preserves a wide planum alveolare, which inclines roughly downwards,while a strong superior torus is developed. Below the torus there is a clear and large fossagenioglossa. This feature is quite clear in all known male and female mandibles of Oura-nopithecus from RPl and NKT (Fig. 3). In RPl-75 the fossa genioglossa is not so clear andthis is possibly due to a lateral compression of the mandible at the symphysis level. On thecontrary there is no fossa genioglossa in Graecopithecus and the superior torus is veryweak. Moreover, the internal symphysis of the Pyrgos mandible is narrower than that ofOuranopithecus. The breadth of the internal symphysis between the internal alveoles ofp3 and in the p4/m1 contact is 12.5 mm and 17.5 mm respectively for Graecopithecus,while in the female mandibles of Ouranopithecus varies from 23.5 to 24.4 mm, mean24.0 mm and in the males from 28.5 to 29.0 mm, mean 28.7 mm, respectively. Thus, Grae-copithecus and Ouranopithecus have completely different morphology and size of the inter-nal symphysis. One can support that the sole mandible of Graecopithecus cannot give acomplete idea of the symphysial morphology and may be new material will provide morecertain data. This is true but no possibility for new material at the moment, while the lim-ited morphology and size of the Pyrgos mandible is a fact.
Table 1Dental proportions of m2 and m3 in various late Miocene and recent hominoids, as well as A. afarensisProportions de m2 et m3 chez quelques hominoïdes miocènes et actuels et chez A. afarensis.
Taxon Bm2/Lm2 Bm3/Lm3Gorilla, male (94) 102 (104) (93) 102 (108)Gorilla, female (102) 104 (108) (100) 102 (104)Graecopithecus 94Ouranopithecus, male (88) 94 (100) (79) 85 (91)Ouranopithecus, female (92) 97 (103) (81) 85 (90)Ankarapithecus (90) 96 (98)Proconsul 86 80Dryopithecus 87 85A. afarensis (99) 100 (108) (87) 93 (98)
233G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

234 G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

2.6. Occlusal shape of the mandible
In occlusal view the shape of the Graecopithecus mandible is different than that of Oura-nopithecus being narrower (de Bonis and Koufos, 1993). The internal alveolar breadth inthe middle of the teeth of the Pyrgos mandible is compared to that of Ouranopithecus(Fig. 4). These measurements of the Pyrgos mandible are the most certain as the specimenhas not lateral distortion, which could affect the occlusal breadth between the two man-dibular rami. The Pyrgos mandible is clearly narrower from both males and females ofOuranopithecus. The internal alveolar breadth is very narrow between the canines and thepremolars but then it became relatively larger. This form of the mandible gives to the dentalarcade a more closed form with more parallel tooth rows. On the contrary in Ouranopith-ecus the dental arcade is open anteriorly with diverging tooth rows (more parabolic) indi-cating probably more evolved feature. Proconsul has more closed arcade than Ouranopith-ecus but more open comparatively to Graecopithecus (Fig. 4). In this feature the Pyrgosmandible is very close to Ankarapithecus; both retain primitive characters with closed arcadeand more parallel tooth rows. Dryopithecus and Sivapithecus are also closer to Proconsuland Graecopithecus (Fig. 4) On the other hand the extant hominoids Gorilla and Pan arecloser to Ouranopithecus with more open arcade. The mandible of Australopithecus is veryclose to Ouranopithecus and in this feature the latter seems to be more derived than theother extinct hominoids. In the diagram of Fig. 4 there are two groups: the one has moreclosed and parallel tooth rows and the other more diverged ones. Graecopithecus togetherwith Proconsul, Ankarapithecus and Sivapithecus belongs to the first group with moreprimitive characters, while Ouranopithecus and Australopithecus in the second with morederived occlusal shape of the mandible.
2.7. Dental dimensions
Concerning the dental dimensions the sole measurable tooth is the right m2 (14.0 × 13.1;Martin and Andrews, 1984). Its dimensions are very close to the minimum values for femaleOuranopithecus (Fig. 1). However, the advanced wear of the tooth obliterated all its mor-phological characters, making impossible the comparison. The other teeth are broken andthe root’s remains cannot provide reliable data for a certain comparison with the otherhominoids in order to find their relationships. Thus, once more the limited features of thePyrgos mandible are negative for a certain comparison.
3. Discussion
The systematic position of Ouranopithecus has been discussed since a long time fromthe middle of 1980s. Martin and Andrews (1984) discussed the phyletic position of Grae-
Fig. 3. Occlusal view of the known mandibles of Ouranopithecus (RPI, NKT) and Graecopithecus. f.g. = fossagenioglossa, p.a. = planum alveolare.Fig. 3. Vue occlusale des mandibules de Ouranopithecus (RPI, NKT) et de Graecopithecus. f.g. = fossa genio-glossa ; p.a. = planum alveolare.
235G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

copithecus and for the first time synonymized the two genera. Later on, Andrews et al.(1996) and Cameron (1997) discussed again the position of the two genera and proposedsynonymy, while the latter author suggested a separate species for the Ouranopithecussample. At the same time we gave several data, indicating the differences between the twogenera (de Bonis and Koufos, 1993; Koufos, 1995). In many cases the name Ouranopith-ecus was used by the people having a different opinion than us, but then came back to thesynonymy with Graecopithecus. This provided sometimes confusion to the palaeontolo-gists as several morphological characters of Ouranopithecus are referred as those of Grae-copithecus. In the present article we are making an effort to put the main arguments togetherand to discuss them again in order to give an answer for the synonymy or not of these twohominoids. The enlargement of Ouranopithecus sample during the last 5 years providedbetter comparative and statistical data. During the last 20 years several times we referredthat we do not insist for imposing our opinion and our genus Ouranopithecus, but we aretrying to show the real data we have about these two hominoids and their validity. If it willbe possible for anyone to find to morrow some new material from Pyrgos, which can pro-vide all the available data proving the similarity of the two genera, we are ready to acceptthe synonymy.
The Pyrgos locality cannot be excavated any more because the area was built. Thus, nonew material can be provided for certain comparisons at the moment, in order to give newevidences for the synonymy of the two genera. Concerning the age of Graecopithecus it isreferred “The type ...... of uncertain age, probably MN 10...” (Andrews et al., 1996). But,let see the biochronological or other data we have from Pyrgos locality. The accompanyingfauna is very poor and the designations were based on fragmentary material, e.g. T. amaltheais determined by a mandibular fragment with m1-m3 preserved; “D. orientalis” (possibly
Fig. 4. Comparison of the occlusal shape of the mandible in various Late Miocene and recent hominoids, as wellas A. afarensis.Fig. 4. Comparaison de la morphologie occlusale chez quelques hominoïdes miocènes et actuels et chez A. afa-rensis.
236 G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

Ceratotherium neumayri) on some bone fragments. Even the hominoid mandible was origi-nally assigned to Mesopithecus (von Freyberg, 1951; von Koenigswald, 1972). On the otherhand none of the recorded taxa can provide certain evidences for a Vallesian or Turolianage. C. pentelici lived during the whole Late Miocene. “D. orientalis” cannot providecertain age as it is known throughout Late Miocene. The giraffids Helladotherium andBohlinia are also known from the Vallesian and Turolian of Eastern Mediterranean (Kou-fos, 2003). The questionable presence of the two gazelles, are not age indicable as they areknown from the whole Late Miocene. Tragoportax (if the attribution is right) lived through-out Late Miocene. Finally, the assumption of von Koenigswald (1972) that the two hippa-rions of different size suggest an age later than Pikermi is not valuable, as hipparions ofdifferent size are known from entire Late Miocene and Pliocene. Thus the age of the Pyrgoslocality cannot be precisely determined as the fauna is very poor with limited attributions.The certain species are not so characteristic and cannot provide an age more precise thanLate Miocene. In this case the factor of the precise age of the Pyrgos mandible is negativefor its comparison and extraction of phylogenetic results. In fact the locality of PyrgosVassilissis is located in Attica, where Pikermi is also situated. But, this does not mean thatit is of similar age, as it was proposed several times. Several times mammalian localitiesvery close each other include different faunas and have different ages, e.g. Nikiti localities,Greece (Koufos, 2000) or Kemiklitepe localities, Turkey (de Bonis et al., 1994).
Cameron (1997: p. 244), argues that “in accepting Ouranopithecus and simply ignoringGraecopithecus, we are indirectly accepting two genera or large bodied hominoid in Greece(i.e. Ouranopithecus from Macedonia and Graecopithecus from southern Greece”. First ofall nobody ignores Graecopithecus specimen, after its appearance (von Koenigswald, 1972)as no reference before, except that of von Freyberg (1951), who refers the presence ofMesopithecus. On the other hand, nowadays, large hominoid primates, like gorillas andchimps, do live close each other in Africa. Continuing his analysis and looking on thedifferences between the northern and southern specimens, Cameron (1997) reported... “sug-gest that the southern Greek hominid Graecopithecus can be distinguished from the Mace-donian hominid Ouranopithecus by the following characteristics:• the internal face of the mandibular symphysis of Graecopithecus lacks the large supe-
rior torus seen in Ouranopithecus (see above);• and in occlusal view, the shape is very different as the anterior part is narrower and the
distance between the lingual faces of the two lower first molars reaches 16 mm in Grae-copithecus and 27.8–30.5 for the smallest mandibles of Ouranopithecus (see above)”.But the same author continued and referred “....the suggested significance of these char-
acters as being distinctive is open to question.” For us the question is closed insofar as thedifferences between the samples allow to see that they are very different and that there is noreason to put them together in the same genus. Ahead in the text Cameron (1997) writes“The Pyrgos mandible is moderately smaller in all values compared to the Macedonianspecimen..... the southern and northern Greek specimens are at least specifically distinct”.Here we agree and we insist on the words ‘at least’, which means that it is possible to begenerically distinct too. Andrews et al. (1996) reported that “Other apparent differences(between the two taxa) may be the result of deformation of the Pyrgos jaw”. In our opinionthe deformation can affect not only the differences but also the similarities between the twotaxa.
237G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

The Pyrgos mandible is a very peculiar specimen badly preserved and with very fewmorphological features. The weathering or the bad collection damaged the specimen andsignificant morphological characters have been destroyed (mandibular ramus, symphysialarea, teeth). Andrews et al. (1996) refers that the Pyrgos mandible is not so incomplete aswe believe. But, the same authors at the beginning of the corresponding chapter wrote,“The type is a fragmentary mandible....” and for us this means incomplete. Moreover, in theoriginal description of the mandible after its final cleaning, it is referred (Martin andAndrews,1984):• “.. it was damaged during excavation and most of the symphysis was destroyed....”;• “the right body is slightly distorted by crushing, but it is still measurable, whereas the
left body is more distorted”;• “The inferior border of the left body is partly missing and partly flattened..... it is not
possible to obtain accurate measurements of this side”;• “the lateral borders of both mandibular bodies have suffered superficial bone loss as a
result of pre-burial weathering....”;• “the weathering has also affected the inferior and alveolar borders.... The inferior mar-
gin is missing...”, etc.We think that all these damages, weatherings, distortions, absence of morphologies,
lack of the teeth cannot characterize the mandible complete. Cameron (1997) refers “with-out additional material of G. freybergi....... there is no evidence to support the recognitionof two large bodied hominid genera within the Miocene of southern and northern Greece”.It means that the fragmentary and incomplete mandible of Graecopithecus is not enough todefine a certain taxon, but then he puts Ouranopithecus in synonymy with Graecopithecus.If “...these samples should be considered specifically distinct” (Cameron, 1997) then twoquestions arise for us: (a) why not generically, and (b) how we can be sure about theirsimilarities with this limited material in order to be led to the synonymy. Our opinion is thatthe Pyrgos mandible is incomplete, with limited morphology and cannot be synonymizedwith any of the Miocene hominoids. The proposed similarities of Graecopithecus and Oura-nopithecus either they are not reliable or they are limited and cannot prove certainly thesynonymy. Another argument about the incompleteness of the mandible is that there aresome taxa whose type material is very few or single, e.g. Griphopithecus (Andrews et al.,1996). This is true, the type material of the later taxon consists of four teeth, but these teethare in a good condition and can provide all the available data for a comparison with othersamples. In Graecopithecus there is a badly preserved mandible with a very worn andfragmentary m2, whose morphology is almost unknown (except the enamel thickness); therest dental remains on the mandible are the roots of the p4 and m1.
Another problem with the comparison of Graecopithecus is that there is an one-sidedcomparison (Martin and Andrews, 1984; Andrews et al., 1996; Cameron, 1997). It is onlycompared with the Ouranopithecus sample. The question is why not with the almost iso-chronous Ankarapithecus or the slightly older Griphopithecus? The reason that the latertwo taxa were found in different countries is not strong, as the mammals could move fromAsia Minor to Greece and vice versa. In the above-mentioned comparisons Graecopith-ecus seems to have some similarities with Ankarapithecus, which can allow someone tosynonymize the two taxa. For us this is impossible especially for the incompleteness of thePyrgos mandible and for the uncertain morphological and metrical data that it can provide.
238 G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

The extensive comparison of the Pyrgos mandible with those of Ouranopithecus suggestsclearly that it is different in several characters (shape of the internal symphysis, MR, occlusalshape of the mandible, size of the mandible, and proportions of the teeth). The incomplete-ness of the Pyrgos mandible provides few comparative characters and dimensions, whilemost of them are uncertain and cannot provide reliable results. Thus, Graecopithecus man-dible is difficult to be compared with the other extinct and extant hominoids and as it is notpossible to find more material from this hominoid (the area where it was found is built),either it is impossible to determine its relationships with the other hominoids or to getlimited and doubtful results.
4. Conclusions
The above given comparison of Graecopithecus and Ouranopithecus shows that the twogenera cannot be synonymized. The sole known mandible of Graecopithecus is so incom-plete that cannot provide enough and reliable data for certain comparisons to the otherextinct and extant hominoids. The incompleteness of the mandible provides limited mea-surements and thus the metric comparisons are not valuable, in several cases. However,some morphological and metrical characters of the Pyrgos mandible are reliable (morphol-ogy of the internal symphysis, enamel thickness, internal alveolar breadth) and differentbetween the two genera. On the other hand the four characters, proposed by Martin andAndrews (1984) for the synonymy of the two genera, cannot clearly support this as it isproved in the above comparisons. As a conclusion the Pyrgos mandible must be separatedfrom all Miocene hominoids and remain as an isolated genus with only one species G.freybergi. O. macedoniensis constitutes a quite rich sample of cranial remains with clearmorphology and size, accurate age and it can be compared to all known extinct and extanthominoids.
Aknowledgements
Many thinks to Dr. Peter Andrews for providing us the cast Graecopithecus. Thanks arealso due to an anonymous reviewer for his usefull comments. The excavations in Axiosvalley and Nikiti have been financed by the French CNRS, the Leakey Foundation forAnthroplogical Research, and the National Science Fondation of USA (program RHOI).
References
Alpagut, B., Andrews, P., Fortelius, M., Kappelman, J., Temizsoy, I., Celebi, H., Lindsay, W., 1996. A newspecimen of Ankarapithecus meteai from the Sinap Formation of central Anatolia. Nature 382, 349–351.
Andrews, P., Harrison, T., Delson, E., Bernor, R.L., Martin, L., 1996. Distribution and Biochronology of Europeanand Southwest Asian Miocene catarrhines. In: Bernor, R.L., Fahlbusch, V., Mittmann, H.-W. (Eds.), TheEvolution of Western Eurasian Neogene Mammal Faunas. Columbia University Press, New York, pp. 168–207.
Begun, D., Gülec, E., 1998. Restoration of the type and palate of Ankarapithecus meteai; taxonomic andphylogenetic implications. American Journal of Physical Anthropology 105, 279–314.
239G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240

de Bonis, L., Bouvrain, G., Geraads, D., Koufos, G.D., 1990. New hominid skull material from the late Mioceneof Macedonia in Northern Greece. Nature 345, 712–714.
de Bonis, L., Bouvrain, G., Geraads, D., Koufos, G.D., 1992. Diversity and palaeoecology of Greek late Miocenemammalian faunas. Palaeogeography, Palaeoclimatology, Palaeoecology 19, 99–121.
de Bonis, L., Bouvrain, G., Geraads, D., Koufos, G., Sen, S., Tassy, P., 1994. Les gisements de mammifères duMiocène supérieur de Kemiklitepe, Turquie : 11. Biochronologie, paléoécologie et relations paléo-biogéographiques. Bulletin du Muséum National d’Histoire naturelle sér 4, 225–240 (16).
de Bonis, L., Bouvrain, G., Geraads, D., Melentis, J., 1974. Première découverte d’un primate hominoïde dans leMiocène supérieur de Macédoine (Grèce). Comptes Rendus de l’Académie des Sciences de Paris (D) 278,3063–3066.
de Bonis, L., Koufos, G.D., 1993. The face and the mandible of Ouranopithecus macedoniensis: description ofnew specimens and comparisons. Journal of Human Evolution 24, 469–491.
de Bonis, L., Koufos, G.D., 1999. The Miocene large mammal succession in Greece. In: Agustí, J., Rook, L.,Andrews, P. (Eds.), Hominoid Evolution and Climatic Change in Europe.Volume 1: The Evolution of NeogeneTerrestrial Ecosystems in Europe. Cambridge University Press, London, pp. 205–237.
de Bonis, L., Melentis, J., 1977. Les Primates hominoïdes du Vallésien de Macédoine (Grèce). Étude de lamâchoire inférieure. Geobios 10, 849–885.
Cameron, D.W., 1997. The taxonomic status of Graecopithecus. Primates 38, 293–302.von Freyberg, B., 1951. Die Pikermi-fauna von tour la Reine (Attica). Annales géologiques des Pays Helléniques
3, 7–10.von Koenigswald, G.H.R., 1972. Ein Unterkiefer eines fossilen Hominoiden aus dem Unterpliozan Griechen-
lands. Proceedings of the Koninklijke Nederlandse Akademie Wettenschaffen (B) 75, 385–394.Koufos, G.D., 1993. A mandible of Ouranopithecus macedoniensis from a new late Miocene locality of
Macedonia (Greece). American Journal of Physical Anthropology 91, 225–234.Koufos, G.D., 1995. The first female maxilla of the hominoid Ouranopithecus macedoniensis from the late
Miocene of Macedonia, Greece. Journal of Human Evolution 29, 385–399.Koufos, G.D., 2000. The hipparions of the late Miocene locality “Nikiti-1” Chalkidiki, Macedonia, Greece. Revue
Paléobiologique 19, 47–77.Koufos, G.D., 2003. Late Miocene mammal events and biostratigraphy in the Eastern Mediterranean. Deinsea 10,
343–371.Koufos, G.D., 2005. Palaeoecology and chronology of the Vallesian (late Miocene) in the Eastern Mediterranean.
Palaeogeography, Palaeoclimatology, Palaeoecology (in press).Martin, L., Andrews, P., 1984. The phyletic position of Graecopithecus freybergi Koenigswald. Courrier
Forschung Institute Senckenberg 69, 25–40.Sen, S., Koufos, G.D., Kondopoulou, D., de Bonis, L., 2000. Magnetostratigraphy of the late Miocene continental
deposits of the lower Axios valley, Macedonia, Greece. In: Koufos, G.D., Ioakim, C. (Eds.), MediterraneanNeogene Cyclostratigraphy in Marine–Continental Deposits. Bulletin of the Geological Society of Greece(special publication) 9. pp. 197–206.
Ungar, P., 1996. Dental microwear of European Miocene catarrhines: evidence for diet and tooth use. Journal ofHuman Evolution 55, 307–322.
240 G.D. Koufos, L. de Bonis / Annales de Paléontologie 91 (2005) 227–240




















