
Facies Models 2. Turbidites and Associated Coarse Clastic ...

Taxonomy, facies relationships and palaeobiology of bakevelliidbivalves from the Lower Cretaceous of west-central Argentina
Darıo Gustavo Lazo*
Departamento de Ciencias Geologicas, Universidad de Buenos Aires, Ciudad Universitaria, Pabellon II, 1428 Buenos Aires, Argentina
Accepted 10 September 2003
Abstract
Cretaceous bakevelliids in Argentina are restricted to the Lower Cretaceous. They are recorded in shallow marine deposits in twoMesozoic basins located in the Andes foothills in Patagonia. In the Austral Basin (44(–55(S) there is only one genus, Gervillella,represented by a single specimen from the Barremian. In the Neuquen Basin (30(–40(S) two genera, Gervillaria and Gervillella,occur, ranging from the Berriasian to the Lower Barremian. In the Pilmatue Member of the Agrio Formation two species areidentified: Gervillaria alatior (Imlay) and Gervillella aviculoides (J. Sowerby). The former is also recorded in Mexico while the latteris more widespread, occurring in Europe, Asia, and Africa. In the Bajada del Agrio section they come from eight tabular shell bedslocated near and on top of shallowing-upwards cycles. They were abundant only in four of these levels. These shell beds areinterpreted as having been deposited in the mid-ramp (middle of cycles), and upper mid-ramp (top of cycles). A functional analysisof shells revealed two life habits: epibyssate and endobyssate.� 2003 Elsevier Ltd. All rights reserved.
Keywords: Bakevelliidae; Lower Cretaceous; Argentina; taxonomy; taphonomy; palaeobiology
1. Introduction
Bakevelliids are an extinct group of pteriomorphianbivalves with an extensive palaeobiogeographical distri-bution in the marine Mesozoic. This group ranged fromthe Late Permian to the Eocene (Cox, 1954, 1969). Theydiversified widely during the Triassic, reaching theirmaximum diversity by Early and Middle Jurassic times(Hayami, 1957; Muster, 1995; Aberhan and Muster,1997). They have a lot of potential for palaeobiogeo-graphic studies and plate reconstructions because theirdispersal pattern apparently was not facies-influencedand they show a low degree of endemism (Muster, 1995;Aberhan and Muster, 1997).
Bakevelliidae, Pterineidae, Pinnoidea+Pterioidea,and Ostreoida form a monophyletic group, even thoughtheir relationships are still unresolved (Waller, 1998).According to Waller (1998) bakevelliids share onlyone derived character: an alivincular or multivincular
ligament system with pits unevenly spaced and its ven-tral zone expanding through ontogeny. The weak byssalsinus, a reduced or absent anterior adductor muscle, anda shell composed of an outer calcitic and an inneraragonitic layer are plesiomorphic characters. The mul-tivincular ligament has evolved at least twice in pterio-morphian evolutionary history as it is also present in thePinnoidea+Pterioidea clade.
Information on Lower Cretaceous bivalves fromSouth America is scarce (Dhondt and Dieni, 1988),though data have been obtained from Argentina sincethe end of the nineteenth century with the taxonomy ofthe Order Trigonioida the most studied topic. Researchon other groups (e.g. heterodonts and pteriomorphians)or studies on bivalve taphonomy or palaeoecology arescarce.
Cretaceous bakevelliids from Argentina are particu-larly poorly known. The present paper reviews previousrecords and then considers in detail the taxonomy, faciesdependence and stratigraphic distribution of thebakevelliids of the Agrio Formation (Lower Cretaceous)of the Neuquen Basin (Fig. 1), west-central Argentina,
* Corresponding author.E-mail address: [email protected] (D. Gustavo Lazo).
Cretaceous Research 24 (2003) 765–788
0195-6671/03/$ - see front matter � 2003 Elsevier Ltd. All rights reserved.doi:10.1016/j.cretres.2003.09.001
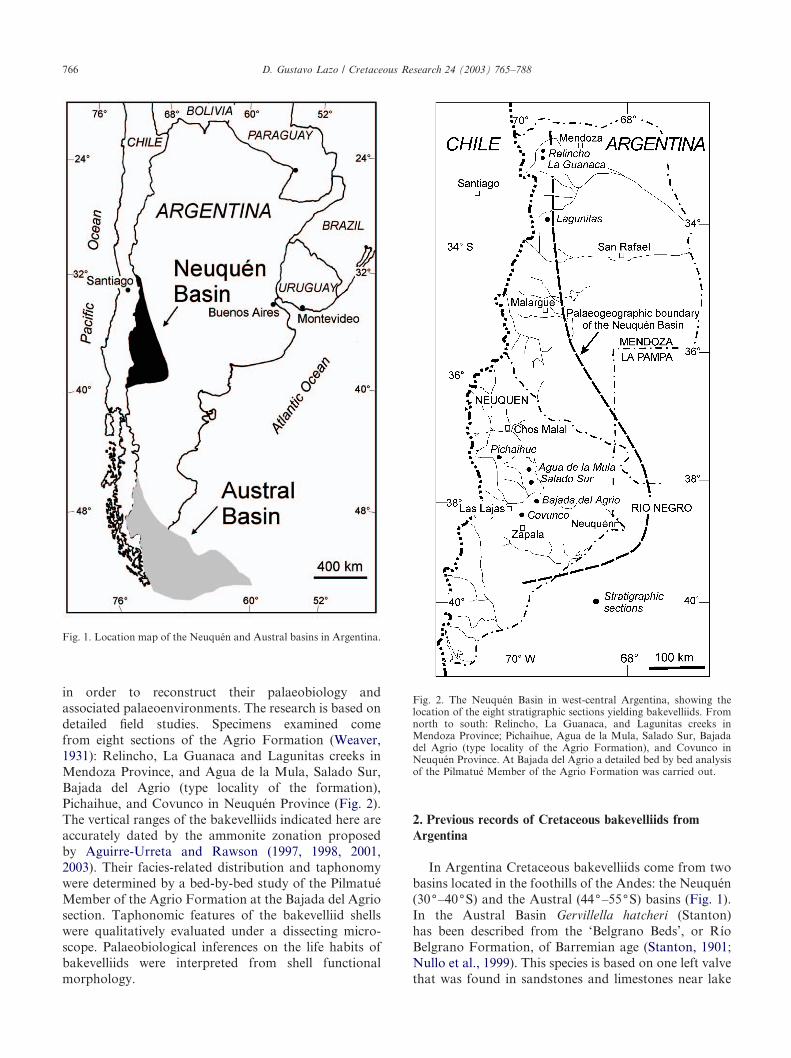
in order to reconstruct their palaeobiology andassociated palaeoenvironments. The research is based ondetailed field studies. Specimens examined comefrom eight sections of the Agrio Formation (Weaver,1931): Relincho, La Guanaca and Lagunitas creeks inMendoza Province, and Agua de la Mula, Salado Sur,Bajada del Agrio (type locality of the formation),Pichaihue, and Covunco in Neuquen Province (Fig. 2).The vertical ranges of the bakevelliids indicated here areaccurately dated by the ammonite zonation proposedby Aguirre-Urreta and Rawson (1997, 1998, 2001,2003). Their facies-related distribution and taphonomywere determined by a bed-by-bed study of the PilmatueMember of the Agrio Formation at the Bajada del Agriosection. Taphonomic features of the bakevelliid shellswere qualitatively evaluated under a dissecting micro-scope. Palaeobiological inferences on the life habits ofbakevelliids were interpreted from shell functionalmorphology.
2. Previous records of Cretaceous bakevelliids fromArgentina
In Argentina Cretaceous bakevelliids come from twobasins located in the foothills of the Andes: the Neuquen(30(–40(S) and the Austral (44(–55(S) basins (Fig. 1).In the Austral Basin Gervillella hatcheri (Stanton)has been described from the ‘Belgrano Beds’, or RıoBelgrano Formation, of Barremian age (Stanton, 1901;Nullo et al., 1999). This species is based on one left valvethat was found in sandstones and limestones near lake
Fig. 1. Location map of the Neuquen and Austral basins in Argentina.
Fig. 2. The Neuquen Basin in west-central Argentina, showing thelocation of the eight stratigraphic sections yielding bakevelliids. Fromnorth to south: Relincho, La Guanaca, and Lagunitas creeks inMendoza Province; Pichaihue, Agua de la Mula, Salado Sur, Bajadadel Agrio (type locality of the Agrio Formation), and Covunco inNeuquen Province. At Bajada del Agrio a detailed bed by bed analysisof the Pilmatue Member of the Agrio Formation was carried out.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788766

Pueyrredon, Santa Cruz Province. Since the pioneeringwork of Stanton (1901), no other data on this specieshave been published.
For the Neuquen Basin, bivalve (including bake-velliid) species lists and stratigraphical occurrences werepublished by Camacho and Riccardi (1978), Mancenido& Damborenea (1984), Riccardi (1988), and Riccardiet al. (1993, 1999). The Neuquen Basin is an importantback-arc depocentre which evolved from the LateTriassic until the Late Tertiary, primarily withinMendoza and Neuquen provinces (Vergani et al., 1995).Bakevelliids have been found in the Quintuco, Chachaoand Agrio formations.
The Quintuco Formation (Berriasian–Valanginian) isa mixed carbonate-siliciclastic unit mostly developed inNeuquen Province (Weaver, 1931). It is approximately300 m thick, and has been interpreted to reflect a basinalto littoral marine environment (Rodriguez Schelottoet al., 1981). Two species of bakevelliids have beenrecorded: Gervillaria alatior (Imlay) (see Weaver, 1931,pp. 201–203, pl. 15, figs. 62–63 as ‘Gervillia’ alaeformis)and Gervillaria militaris (Burckhardt) (Weaver, 1931,p. 211, pl. 15, fig. 65, as ‘Perna’ militaris).
The Chachao Formation (Valanginian) is a cal-careous unit developed in Mendoza Province(35(21#–37() (Uliana et al., 1977; Carozzi et al., 1981)where it overlies the Vaca Muerta Formation and con-formably underlies the Agrio Formation (see below). Itreaches a maximum thickness of 40 m, and towards thesouth becomes progressively thinner where it interfingerswith clastic deposits of the Mulichinco Formation(Valanginian). It is mostly composed of bioclastic pack-stones, wackestones and mudstones interpreted asdeposits of a low-energy sublitoral carbonate ramp(Mombru et al., 1978; Damborenea et al., 1979;Legarreta et al., 1981). Two species of bakevelliid occur:Gervillaria alatior (Imlay) and Gervillaria militaris(Burckhardt) (Damborenea et al., 1979); these formautochthonous associations with, and are dominated by,corals.
The Agrio Formation (Upper Valanginian–LowerBarremian) is a thick sequence of approximately 1600 moverlying the Chachao and Mulichinco formations(Weaver, 1931; Riccardi, 1988). This unit crops out inMendoza and Neuquen provinces and has been sub-divided into the Pilmatue (Lower), Avile (Middle) andAgua de la Mula (Upper) members (Weaver, 1931). Incentral Neuquen the Lower and Upper members com-prise shales, sandstones, and bioclastic limestones ofmarine origin. These have been interpreted as offshoreto shoreface deposits in a storm-dominated mixedcarbonate-siliciclastic ramp (Brinkmann, 1994; Spallettiet al., 2001). The Middle Member is a siliciclas-tic continental wedge (mainly aeolian to restrictedlacustrine environments) of 20–150 m thickness, whichaccumulated during a major regression that occurred in
the Early Hauterivian (Gulisano & Gutierrez Pleimling,1988; Vergani et al., 2002). Two species of bakevelliidhave been found in the Agrio Formation: Gervillariaalatior (Imlay) (see Weaver, 1931, pp. 201–203, pl. 15,fig. 61, as ‘Gervillia’ alaeformis) and Gervillella avicu-loides (J. Sowerby) (see Weaver, 1931, pp. 203–204, pl.14, figs. 58–59 as ‘Gervillia’ anceps). The former wasfound in Mendoza and Neuquen while the later has beenreported only from Neuquen (Weaver, 1931; Aguirre-Urreta and Lo Forte, 1996). Behrendsen (1892) brieflydescribed and figured Gervillaria sp. from the LowerCretaceous of Caryilauhue, 27 km south of Catan Lil inNeuquen (Camacho and Riccardi, 1978).
In summary, Cretaceous bakevelliids range from theBerriasian to the Lower Barremian in the NeuquenBasin, where they are relatively common in three litho-stratigraphic units (Fig. 3). From the Austral Basin onlyone valve has been recorded, from the Barremian. UpperCretaceous bakevelliid records are lacking from theseareas. All records come from shallow, subtidal marineenvironments. In contrast to the Lower Jurassic of theNeuquen Basin, from which six bakevelliid genera havebeen recorded (Aguilerella, Bakevellia, Gervillia, Gervil-laria, Gervillella, and Gervilleioperna), in the LowerCretaceous only two genera are found: Gervillaria andGervillella. In this basin both genera range from theUpper Sinemurian–Pliensbachian to Lower Barremian(Weaver, 1931; Damborenea, 1987; Aberhan, 1994).
3. Systematic palaeontology
The described and figured specimens are depositedin the collection of the Catedra de Paleontologıa,Departamento de Ciencias Geologicas, Facultad deCiencias Exactas y Naturales, Universidad de BuenosAires, Pabellon II, Ciudad Universitaria, 1428, BuenosAires, Argentina. Each catalogue number is prefixedCPBA- and refers to specimens from a given strati-graphic position and section. Suffix numbers separatedby a dash (e.g. .10) mean the number of specimensincluded in a given catalogue number.
Synonymy lists were prepared according to Matthews(1973). Only those publications with new and figuredspecimens of bakevelliids are listed. Descriptive termsused in this work are those listed and defined by Cox(1969) and Muster (1995). Ligament terminology fol-lows Crampton (1988). Measurements were made usinga digital caliper (accurate to 0.2 mm) and they wererecorded in mm. Abbreviations are as follows: L, length;H, height; W, width or inflation of single valves; 2W,width or inflation of both valves together; Wlig, width ofthe ligament area; D, length of central body; Wb,maximum width of central body perpendicular to diag-onal; ThLV, thickness of left valve at anterior margin;ThRV, thickness of right valve at anterior margin; Pits,
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 767

number of preserved ligament pits. Angular measure-ments are accurate to the nearest degree: O, obliquity(i.e. angle between the dorsal margin and the diagonal inthe central body). For details of terminology andmeasured characters, see Fig. 4; and for the measure-ments, see Appendix section. Shell size is said to besmall, medium or large by comparison with otherspecimens of the same species.
Class: Bivalvia von Linne, 1758Subclass: Autobranchia Grobben, 1894Superorder: Pteriomorphia Beurlen, 1944 (see Waller,
1978)Order: Pterioida Newell, 1965
Suborder: Pteriina Newell, 1965Superfamily: Pterioidea Gray, 1847Family: Bakevelliidae King, 1850 (see Cox, 1954)
Remarks. Bakevelliid shells are subequivalve toinequivalve, with the left valve larger than the right(Stanley, 1972). Valve shapes may fall into threemain types: ‘pteriiform’, having a rhomboidal to trap-ezoidal oblique outline (e.g. Bakevellia, Gervillaria, andAguilerella), ‘lanceolate’ or longitudinally elongated (e.g.Gervillia and Gervillella), and ‘intermediate’, representedby Hoernesia (Hayami, 1957; Cox, 1969, fig. 43,p. N306). ‘Pteriiform’ and ‘intermediate’ shells arecharacterized by shell torsion and umbonal thickening,
Fig. 3. Ranges of bakevelliid species in the Lower Cretaceous of the Neuquen Basin. Age and ammonite zonation from Aguirre-Urreta and Rawson(1997, 1998, 2001); stratigraphy, lithology and paleoenvironments from Uliana et al. (1977); unconformities from Zavala (2000).
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788768

while ‘lanceolate’ shells are variably elongate (McGhee,1978).
The hinge structure has two main patterns; hingeswith a discernible series of anterior and posterior trans-verse teeth (e.g. Bakevellia and Gervillella), and hingeswith numerous indistinguishable teeth (crenulation orpseudotaxodont dentition) along their entire margin(e.g. Gervillaria and Aguileria) (Hayami, 1957; Cox,1969; Muster, 1995).
Shell structure consists of both calcite and aragoniteas in other pteriomorphian groups such as the Pteriidae,Isognomonidae and Pinnidae (Taylor et al., 1969;Taylor, 1973). It shows an outer prismatic calcitic layerand nacreous inner layers (Waller, 1998). The ligamentarea is formed directly on the aragonitic inner layers asamong the Isognomonidae and Retroceramidae(Crampton, 1988; Waller, 1998). The ligament isexternal, opisthodetic, and multivincular. It showsmultiple ligament pits or resilifers on a rectangular flatligament area. Resilifers vary in number and support thefibrous layer during the bivalve’s lifetime. The fibrouslayer is stiffened by calcium carbonate crystals, and issubject to compression when the valves are closed. Theligament area between resilifers underlies the organiclamellar layer, which in turn suffers tension when thevalves are closed (Cox, 1969; Newell and Boyd, 1987;Muster, 1995). In addition, the ligament area showsgrowth lines formed by ventral migration of the hingeaxis. The functional components of the ligament occuronly in a narrow zone within the lowest part of it(Johnston and Collom, 1998).
Bakevelliids have been regarded as being unable toburrow actively. A byssate mode of life is assumedin most species (Stanley, 1972; McGhee, 1978;Damborenea, 1987; Freneix, 1995; Muster, 1995;Aberhan and Muster, 1997). Unfortunately, direct com-parison with Recent forms is hindered because they are
an extinct group. Hence, the presence of an adult byssuscan only be inferred from studies of shell functionalmorphology and life habits of closely related pterioi-deans. The byssus apparently was an important stabiliz-ing factor among Palaeozoic modioliform bakevelliidsand the diverse, soft bottom dwellers that appearedduring the Mesozoic (twisted recliners and mud stickersin Seilacher, 1984) were derived from that ancestralstock.
Genus Gervillaria (Cox, 1954)
Type species. Modiola? alaeformis J. Sowerby, 1819,p. 93, pl. 251.
Distribution. This genus has been recorded worldwidefrom the Middle Triassic–Upper Cretaceous (Muster,1995).
Gervillaria alatior (Imlay, 1940)Fig. 5, Fig. 6
1931 Gervillia alaeformis (Sowerby); Weaver, pp. 201–203, pl. 15, figs. 61–63.
*1940 Gervillia alatior Imlay, pp. 150–151, pl. 10, figs.1–2; pl. 11, figs. 1–6.
1979 Gervillaria alatior (Imlay, 1940); Damboreneaet al., pp. 27–28, pl. 5, figs. 6–7.
v. 1996 Gervillaria alaeformis (Sowerby, 1819); Aguirre-Urreta and Lo Forte, p. 201, fig. 12b.
v. 2002 Gervillaria alaeformis (Sowerby, 1819); Lazo,p. 3, fig. 2.
Material. Fifty-five specimens all in my collectionfrom the Pilmatue Member of the Agrio Formation incentral Neuquen Province: 32 articulated valves, fourleft valves and two right valves from Agua de la Mula(CPBA 19733.1–2 from Neocomites sp. Subzone, CPBA
Fig. 4. Descriptive morphology and measurements of bakevelliids. For abbreviations see text.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 769
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788770

19734.1–5 from Holcoptychites agrioensis Subzone,CPBA 19731.1–30, 19732.1–2 from Hoplitocriocerasgentilii Zone); two articulated valves and one left valvefrom Salado Sur (CPBA 19735.1, 19736.1 from H.agrioensis Subzone, CPBA 19737.1 from H. gentiliiZone); five articulated valves, six left valves and tworight valves from Bajada del Agrio (CPBA 19738.1–3from H. agrioensis, CPBA 19739.1–8, 19740.1–3 from H.gentilii Zone); and one left valve (CPBA 19741.1 fromH. gentilii Zone) from Pichaihue.
Additional material. Four specimens from thePilmatue Member of the Agrio Formation: one com-plete shell (CPBA 14585) collected by Marıa BeatrizAguirre-Urreta at La Guanaca creek, Pseudofavrellaangulatiformis Subzone, Mendoza Province; one leftvalve (CPBA 16208) collected by Graciela Lo Forte atRelincho creek, Pseudofavrella angulatiformis Subzone,Mendoza Province; one left valve (CPBA 18375) col-lected by Fernando Dazquez at Agua de la Mula,Neuquen Province; and one left valve collected bySebastian Saavedra at Lagunitas creek, Weavericerasvacaensis Zone, Mendoza Province (CPBA 19970).
Description. Highly inequivalve and inequilateralshell. Trapezoidal outline in left lateral view, and oval inanterior view. Commisural plane not straight, widelyconcave to the right since shell is slightly twisted anti-clockwise (in posterior view). Beak strongly pro-sogyrous. The left umbo turns over the dorsal margin,the right umbo reaches the dorsal margin withoutexceeding it. The dorsal margin is straight, the anteriormargin dorsally concave and ventrally convex, and theventral margin convex. The posterior margin is hardlypreserved but it can be reconstructed from the growthlines. It is concave with a posteriorly directed curvature(Fig. 5C–D, Fig. 6B, E).
The left valve is more convex than the right (Fig. 5B)and has three morphological elements: anterior auricle,main body, and posterior wing. The anterior auricle islocated forward of the umbo and is triangular in outline.It is limited by a depression at its posterior margin. Themain body is located between the anterior auricle andposterior wing. It is the most convex portion of the shell,forming an acute angle with the dorsal margin (meanobliquity, 55.7(; n, 11). The main body descends diag-onally from the umbo. It is limited by a thin depressionof 1 mm width at its posterior margin. The posteriorwing is widely concave with a triangular-concaveoutline.
The right valve also consists of three parts, thoughthey are less distinct than those of the left valve owing tothe lower convexity of the right main body. The anterior
auricle has a triangular form. The main body, locatedbetween the anterior and posterior wings, is slightlyconvex, and is limited by a gentle depression at itsposterior margin. The posterior wing is longer than theanterior, and triangular in outline.
Shell ornamentation includes concentric growth linesand scattered radial ribs on the left valve, which areclearly observed in well-preserved specimens (Fig. 5D,Fig. 6B, E). In small specimens, the radial ribs reach theventral margin (Fig. 6B) while in large specimens theyare restricted to the dorsal and median portions ofthe shell. Radial ribs are uniformly 1 mm wide, while theinterspace increases from 50 to 200 mm towards theventral region. There are differences in rib densitybetween the main body and the posterior wing, beinglower in the former.
The hinge includes a crenulated dentition withnumerous little teeth ‘imbricated’ towards the anteriormargin. The teeth are 1 mm wide and their lengthdecreases with their stage of development, from 10 to5 mm approximately (Fig. 6C, F, G). The ligament ismultivincular. Resilifers are ovate or elongate-ovate torectangular and sometimes multilobate (Fig. 6C, F). Theventral margin of the resilifers is crenulated (Fig. 6C).The ridges are flat and show a great variability of widthin the same hinge (2–16 mm). At least six pits have beenobserved. The ligament area is flat and ornamented bystraight growth lines parallel to the dorsal margin. Themargins of each ligament area meet ventrally at an angleof 90(. The width of the ligament increases with theontogenetic development, with the left one wider thanthe right (Fig. 7).
Internal characters are: (1) an ovate posterioradductor muscle scar with its maximum length parallelto the dorsal-ventral axis (Fig. 6A, D); (2) an entirepallial line marked by discontinuous little pits (Fig. 6A);and (3) an antero-dorsal triangular groove (Fig. 6C, F).
Distribution. Gervillaria alatior was originallydescribed from the Hauterivian of northern Mexico(Imlay, 1940). In Argentina it has been found in theQuintuco, Chachao, and Agrio formations in theNeuquen Basin (Fig. 3). In this work, it is recordedfrom the Pilmatue Member of the Agrio Formationin Neuquen Province, within the P. angulatiformisand Neocomites sp. subzones, and H. neuquensis, H.gentilii, W. vacaensis zones (Late Valanginian–EarlyHauterivian: Aguirre-Urreta and Rawson, 1997, 1998,2003).
Discussion. The type species of the genus, Gervillariaalaeformis (J. Sowerby), was originally described fromthe Lower Greensand (Aptian) of Great Britain. It is
Fig. 5. Gervillaria alatior (Imlay). A, B, CPBA 19735, right lateral and anterior views. C, CPBA 19740.3 left lateral view. D, CPBA 19739.6, leftlateral view. A and B from Salado Sur; C and D from Bajada del Agrio. All �1.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 771

D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788772

a very common species in the Lower Cretaceous ofEurope, and has been found in Central Asia, Japan,and Africa ranging from the Upper Triassic–LowerCretaceous (J. Sowerby, 1819, vol. 3, p. 93, pl. 251;d’Orbigny, 1845, pp. 484–485, pl. 395; Woods, 1905,pp. 79–83, pl. 11, figs. 9a–d, 10a–d, 11, text-figs. 9–14;Dhondt and Dieni, 1988, pp. 18–19, pl. 4, figs. 2–4,text-fig. 7; Muster, 1995, pl. 7, fig. 7). It differs from theArgentinian species in having a non-projecting posteriorwing and in the absence of radial ribs on large speci-mens. Small specimens of G. alaeformis (Woods, 1905,pl. 11, figs. 9a, 10a–b) are very similar to small and largespecimens of G. alatior (Imlay), which seems to be aneotenic form. These differences were noted in theoriginal description of G. alatior (Imlay, 1940, p. 151)and also by Damborenea et al. (1979), pp. 27–28) andDamborenea (1987, p. 18).
Gervillaria militaris (Burckhardt) from the LowerCretaceous Quintuco and Chachao formations of theNeuquen Basin (Burckhardt, 1903, p. 70, pl. 15, figs.3–5; Weaver, 1931, pp. 211–212, pl. 15, fig. 65;Damborenea et al., 1979, p. 28) has a smaller anteriorwing, more prominent radial ribs (2–3 mm in width),radial rib interspaces of constant width, and moreseparated resilifers on the hinge. This species is alsomore equivalve and has a greater angle between thedorsal margin and the central body. G. miyakoensis(Nagao), from the Lower Cretaceous of Japan andSardinia, is similar in general shape but with a higherobliquity (70–80() (Muster, 1995, pp. 58–60, text-fig.42). Gervillaria sp. (Behrendsen) from the LowerCretaceous of Caryilauhue in Neuquen (Argentina) is a
fragment of right valve. It has a finer and denser radialrib pattern (Behrendsen, 1892, p. 29, pl. 1, fig. 3). G.ashcroftensis (Crickmay) from the Lower and MiddleJurassic of Canada and Chile has a non-projectingposterior wing, narrower main shell body, and anextremely small anterior auricle (Aberhan, 1994, p. 17,pl. 3, figs. 3–4, text-figs. 10–11; Muster, 1995, pp. 53–55,pl. 9, figs. 1–2, text-fig. 39; Aberhan and Muster, 1997,p. 803, text-figs. 3a–d, 5a–b). The Jurassic species lacksradial ribs. G. hartmanni (Munster) from the MiddleTriassic–Upper Cretaceous of Europe, eastern Asia andAfrica, and Chile is more equivalve (Aberhan, 1994,pp. 17–18, pl. 3, figs. 5–7; Muster, 1995, pp. 56–58, pl. 9,figs. 3–5; pl. 10, figs. 1–2, text-fig. 40). It has a largeranterior auricle and a smaller posterior wing without aposterior projection. G. leufuensis (Weaver) from theCallovian of the Neuquen Basin (Weaver, 1931,pp. 201–203, pl. 15, fig. 60) differs from G. alatior(Imlay) in the absence of an anterior wing and in thelack of radial ornamentation. G. pallas (Leanza)from the Lower Jurassic (Upper Sinemurian–LowerToarcian) of Argentina and Chile is more twistedwith the right valve slightly concave (Leanza, 1942,pp. 155–156, pl. 4, fig. 1; Damborenea, 1987, pp. 135–138, pl. 2, figs. 1–5, text-fig. 7; Aberhan, 1994; pl. 4, figs.1–3; Muster, 1995, pp. 60–61, pl. 10, figs. 3–5, text-fig.43; Aberhan and Muster, 1997, pp. 803–805, text-fig.2a–c, h). The Jurassic species has a pair of carinae on theleft valve and a non-projecting posterior wing. It lacksthe anterior auricle and radial ribs.
Genus Gervillella (Waagen, 1907)
Type species. Perna aviculoides J. Sowerby, 1814,pp. 147–149, pl. 66.
Distribution. This genus has been recorded worldwidefrom the Middle Triassic–Upper Cretaceous (Hayami,1957; Muster, 1995).
Gervillella aviculoides (J. Sowerby, 1814)Fig. 8
*1814 Perna aviculoides J. Sowerby, vol. 1, pp. 147–149,pl. 66, figs. 1–4.
1842 Gervillia anceps Deshayes (Leymerie, 1842), p. 9,pl. 10, figs. 3a–3c.
1843 Gervillia anceps Deshayes; d’Orbigny, pp. 482–484,pl. 394.
1905 Gervillia sublanceolata (d’Orbigny, 1850); Woods,pp. 74–78, pl. 10, figs. 14–16, pl. 11, fig. 1, text-figs.7–8.
Fig. 6. Gervillaria alatior (Imlay). A, CPBA 19739.3 interior view of right valve. B, CPBA 19740.1, hinge on left valve. C, CPBA 19741, exterior viewof left valve, D, CPBA 19739.1, interior view of left valve, E, CPBA 19740.3, hinge on left valve, F, CPBA 19739.7, left lateral view. G, CPBA 19741,hinge on left valve. A, B, D–F from Bajada del Agrio; C and G from Pichaihue. All �1.
Fig. 7. Plot showing width of ligament area versus shell heightmeasured in left valves of Gervillaria alatior (Imlay). Data fromAppendix section. R2, R-squared; S, slope; *, significant Student’st test for slope (p<0.05).
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 773

D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788774

v. 1931 Gervillia anceps (Deshayes); Weaver, pp. 203–204, pl. 14, figs. 58–59.
1933 Gervillia aviculoides (Sowerby); Arkell, 1933,pp. 203–205, pl. 26, figs. 1–5.
1940 Gervillella kantkotensis sp. nov.; (Cox 1940),pp. 115–117, pl. 8, fig. 1–5.
1988 Gervillella anceps (Deshayes in Leymerie, 1842);Dhondt and Dieni, pp. 20–21, pl. 3, figs. 6a–6b.
1999 Gervillella sublanceolata (d’Orbigny); (Cecca et al.,p. 386, pl. 4, figs 1–2).
v. 2002 Gervillella sp.; Lazo, p. 3, fig. 2.
Material. Seventy-eight specimens in my collectionfrom the Pilmatue Member of the Agrio Formation incentral Neuquen Province: 11 articulated valves, one leftvalve and one right valve, both fragmented from Aguade la Mula (CPBA 19742.1–6, 19743.1–5 from Neo-comites sp. Subzone, CPBA 19744.1–2 from H. gentiliiZone); 14 articulated and fragmented specimens and oneincomplete left valve from Salado Sur (CPBA19745.1–7, 19746.1–7, 19747.1 from H. gentilii Zone); 42articulated and fragmented valves, three fragments ofleft valves and four fragments of right valves fromBajada del Agrio (CPBA 19749.1–7, 19750.1–2, 19752.2from Neocomites sp. Subzone, CPBA 19751.1 from H.agrioensis Subzone, CPBA 19748.1–38 from H. gentiliiZone).
Additional material. Two articulated and incompletespecimens and two fragments of left valves (CPBA 6997)collected by Horacio Harrington and Armando Leanzain the Lower Cretaceous of Covunco, NeuquenProvince.
Description. Subequivalve and very inequilateral shellwith ensiform outline. Straight and not twisted com-misural plane. Prosogyrous beak placed just above thedorsal margin. The left umbo turns over the dorsalmargin; the right umbo reaches the dorsal margin with-out exceeding it. The anterior margin is straight toslightly convex. An acute angle is formed between theanterior and dorsal margins (18–39(; mean, 28(; n, 36).The posterior margin is straight to slightly concave. Anobtuse angle is formed between the posterior and thedorsal margin (ca. 150(). The dorsal margin is straight,the ventral margin rounded (Fig. 8A, C, F, I).
The left valve is slightly more convex than the right.Three morphological elements characterize each valve:anterior auricle, main body, and posterior wing. Theanterior auricle is located forward of the umbo and istriangular in outline. In left lateral view a furrow sep-arates the anterior auricle from the main body. In the
right valve this furrow is absent and the auricle and mainbody are less differentiated. The main body is the mostconvex portion of the shell. An acute angle is formedbetween the dorsal margin and the diagonal (meanobliquity, 25(; n, 4). Shell convexity decreases ventrally.The main body is limited by a straight depressionparallel to the long axis of the body. The posterior wingis flat, triangular in outline, and larger than the anteriorauricle. It is limited by a straight depression. An acuteangle is formed between the straight depression and thedorsal margin (12–22(; mean, 15(; n, 13).
Valve thickness diminishes from the dorsal towardsthe ventral margin as can be seen from a series oftransverse sections across the long axis of the shell. Thesection outline changes in the same direction from arounded to elliptical central body.
Concentric growth lines ornament the shell. Thesebegin at the umbonal region and end on the dorsalmargin behind the umbo. The main body displays themaximum curvature of growth lines.
The hinge contains well-differentiated anterior andposterior teeth. There are two or three anterior teeth andone posterior tooth (Fig. 8D, G). The teeth are elon-gated (width: 1–2 mm) and ‘imbricated’ towards theanterior margin. The ligament is multivincular (Fig.8H). The resilifers are ovate to rectangular. They are3–8 mm in width. At least four pits have been observed.Ridges are flat, and 3–11 mm in width. A decreasingwidth is usually observed along a continual series ofpits-ridges when preserved. The relationship of the resil-ifers to the ventral margin of the ligament area iscrenulated. The ligament area is ornamented by straightgrowth lines parallel to the dorsal margin. The marginsof each half of the ligament area meet ventrally in anacute angle. The ligament is 6–12 mm in width.
One adductor muscle scar on a posterior positioncould be detected on disarticulated specimens. Due topoor preservation, no other internal characters havebeen observed.
Distribution. Gervillella aviculoides was originallydescribed from the Oxfordian of England. In Europe,it ranges from the Lower Jurassic to the LowerCretaceous. During Mid and Late Jurassic and EarlyCretaceous times the species extended its palaeobiogeo-graphic distribution to Asia, Africa and South America(Muster, 1995). In Argentina, G. aviculoides has beenrecorded from the Agrio Formation in the NeuquenBasin (Weaver, 1931). It is reported herein from thePilmatue Member of the Agrio Formation, from theNeocomites sp. Subzone and H. neuquensis and H.
Fig. 8. Gervillella aviculoides (J. Sowerby). A, CPBA 19748.13, left lateral view. B, CPBA 19748.15 left lateral view. C, CPBA 19744.1, left lateralview. D, CPBA 19745.3, left lateral view. E, CPBA 19742.5, interior view of right valve. F, CPBA 19748.16, dorsal view. G, CPBA 19750.1, rightlateral view. H, CPBA 19745.2, interior view of right valve. I, CPBA 19743.1, left lateral view. A, B, F, and G from Bajada del Agrio; C, E, and Ifrom Agua de la Mula; D and H from Salado Sur. All �1.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 775

gentilii Zones (Upper Valanginian–Lower Hauterivian:Aguirre-Urreta and Rawson, 1997, 1998, Aguirre-Urreta and Rawson, 1997, 1998, 2003).
Discussion. Gervillella aviculoides belongs to a groupof Gervillella characterized by thick non-twisted shellswith a pointed anterior end (Damborenea, 1987). It willbe compared here only with members of this group.
Gervillella hatcheri (Stanton) from the LowerCretaceous of the Austral Basin in Argentina is similarto G. aviculoides but has a narrower anterior end andwider postero-ventral sector (Stanton, 1901, pp. 15–16,pl. 3, figs. 1–2). G. alpina (Pictet and Roux) from theLower Cretaceous of Switzerland is based on a frag-mented specimen with a larger posterior wing (Pictet andRoux, 1849, p. 496, pl. 41, fig. 3). G. lanceolata (Munsterin Goldfuss) from the Jurassic of Germany has a convexanterior margin with a break in the middle of theshell (Goldfuss, 1833, vol. 2, pl. 115, fig. 9), while G.aviculoides has a straight to slightly convex anteriormargin. G. monotis (Eudes-Deslongchamps) from theMiddle Jurassic of Europe and eastern Africa has apointed posterior wing and more convex anteriormargin (Muster, 1995, pp. 69–71, pl. 14, figs. 3–6, text-fig. 52). G. siliqua (Eudes-Deslongchamps) from theMiddle and Upper Jurassic of Europe, eastern Africa,and India is very similar in shape, but has only 3–5 teethin the middle of the hinge axis and a smaller posteriorwing (Muster, 1995, pp. 71–73, pl. 14, figs. 7–8,text-fig. 55).
4. Facies relationships
4.1. General features of the Bajada del Agrio section
The Bajada del Agrio section was analyzed in detailin order to describe the bakevelliid-bearing facies, theirfaunal assemblage and taphonomic features (Fig. 9).The Pilmatue Member of the Agrio Formation is repre-sented here by a vertical stacking of coarsening-upwardand sometimes thickening-upward simple cycles. Theserange in thickness from 4 to 40 m. Complete cycles arecomposed of three facies:
1 The Basal facies are the thickest and comprise greyand dark grey shales and massive mudstones alternat-ing with fossiliferous limestones and calcareousnodules. Their maximum thickness is 30 m. Dispersedammonites and bivalves, e.g. Cucullaea gabrielisLeymerie, Steinmanella pehuenmapuensis (Leanza), S.transitoria (Steinmann), Modiolus cf. subsimplex(d’Orbigny) and Pinna (P.) robinaldina (d’Orbigny),can be preserved inside the nodules. Relatively thinshell beds and pavements of ammonites, bivalves,gastropods, and serpulid worm tubes can also occur.These fine sediments are interpreted as fall-outdeposits of a shallowing-upwards outer ramp, with
distal storm influence (Burchette and Wright, 1992).Occasional ammonites are interpreted as nektonicrain, while those deposits with infaunal or semi-infaunal bivalves may be related to colonizationevents as a result of episodic increases in bottomoxygen level and substrate consistency (Lazo, 2001a).
2 The Middle facies are coarsening upwards and com-posed of alternating thin yellowish, fine- to medium-grained sandstones and olive grey mudstones forminga lenticular to wavy heterolithic structure (Fig. 10A,C). The maximum thickness of the Middle facies is12 m. Individual beds have sharp, plane or erosionalbases and a thickness ranging from 5 to 30 cm. Thedominant sedimentary structures in the sand-stones are wave ripples, combined flow ripples, andhummocky cross-stratification. Mudstones aremassive or plane-laminated. Sandstone tops show adiverse trace fossil assemblage belonging to theCruziana ichnofacies (Lazo, 2001b). Occasionallyshell beds replace sandstones. Dispersed infaunalor semi-infaunal bivalves, such as S. transitoria(Steinmann) and P. (P.) robinaldina (d’Orbigny),occur within mudstones. The Middle facies are inter-preted as mid-ramp deposits, above the storm wavebase with periodic storm influence (Burchette andWright, 1992).
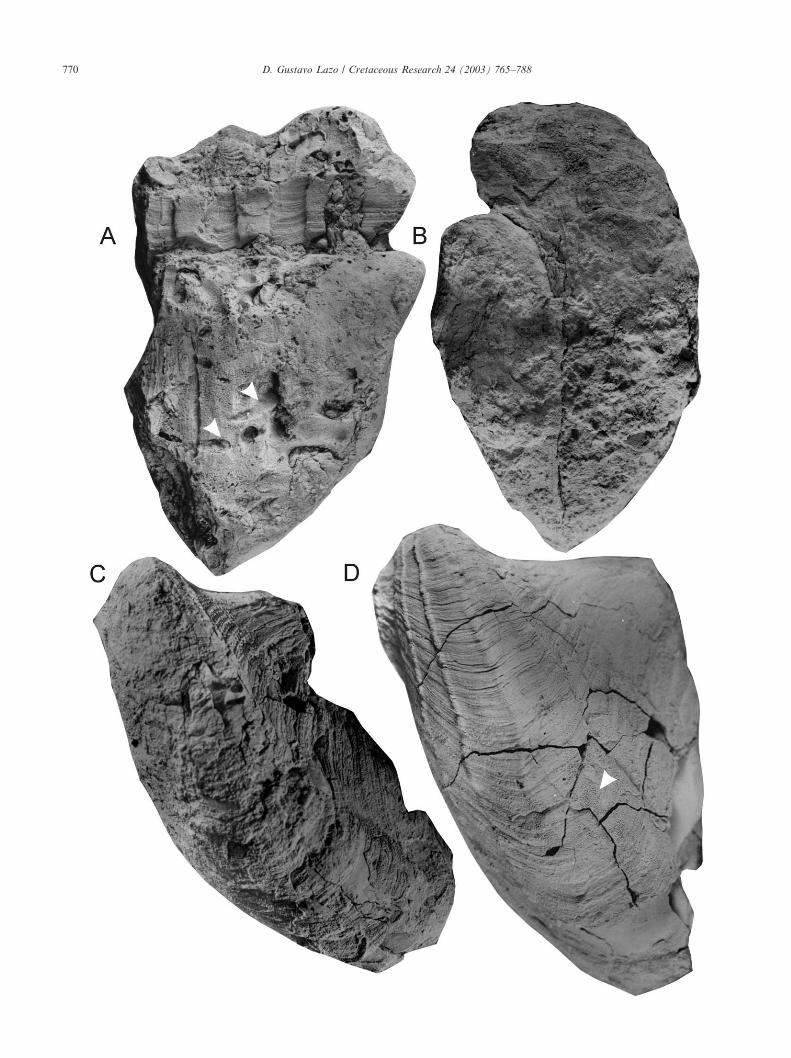
3 The Top facies are coarsening upwards and com-posed of amalgamated brownish fine- to coarse-grained sandstones alternating with abundant shellbeds (Fig. 10A, B). The maximum thickness of theTop facies is 8 m. Amalgamated sandstone beds showerosive bases, wave ripple cross stratification and abioturbation increasing upwards. Shell beds up to0.5 m thick are multi-event in character. They typi-cally comprise highly disarticulated and frag-mented, poorly size-sorted shells. Valves are ran-domly orientated and form a loosely to denselypacked biofabric. Species richness is high. The Topfacies are interpreted as mid-ramp deposits, but in ashallower location than the middle facies (uppermid-ramp).
4.2. Bakevelliid-bearing facies
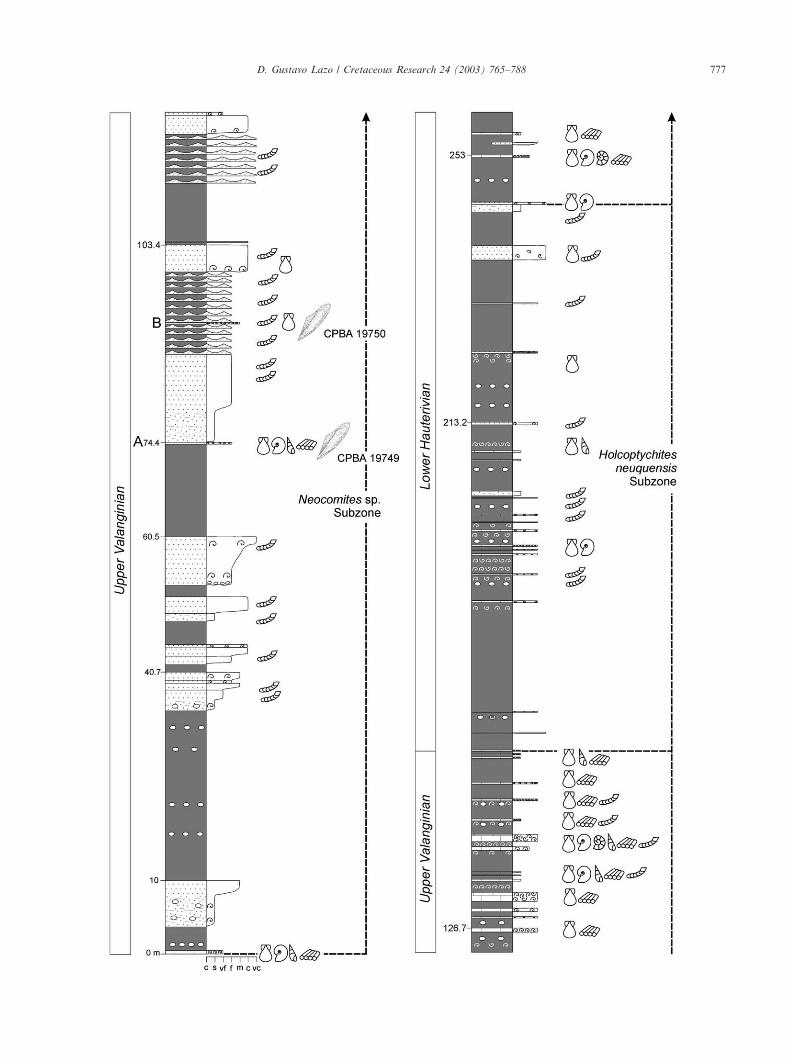
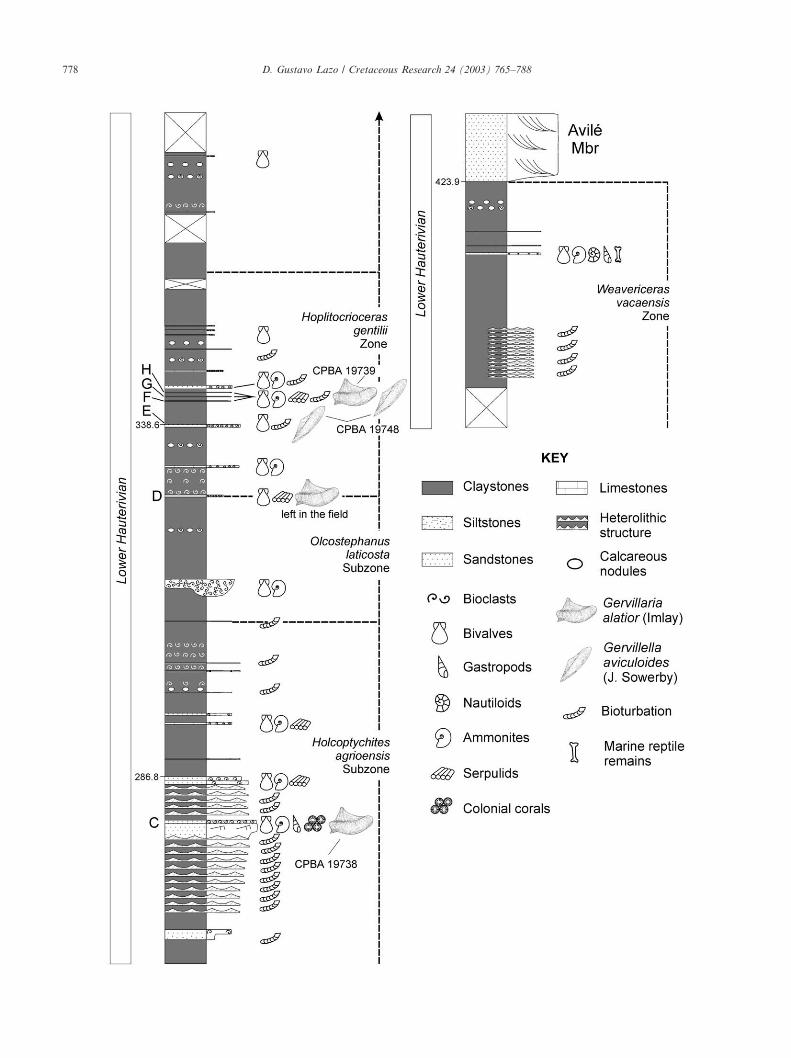
Bakevelliids appear discontinuously in the PilmatueMember of the Agrio Formation in Bajada del Agrio.They were found in eight tabular shell beds (0.05–0.5 mthick) belonging to the Top and Middle facies (Fig.9A–H). These shell beds can be correlated easily amongadjacent sections following both bio- and lithostrati-graphic criteria. A summary of features of bakevelliid-bearing beds and abundance of bakevelliids is given inTable 1.
Bakevelliid-bearing shell facies are described belowaccording to their occurrence in the Top and Middlefacies.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788776
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 777
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788778

4.2.1. Middle facies bakevelliid-bearing shell beds.Description: Shell beds B (Neocomites sp. Subzone),
F, G, and H (H. gentilii Zone) appear in the Middlefacies of cycles (Fig. 10C). These shell beds are thin(0.05–0.16 m), showing a sharp and irregular base,simple internal structure, low degree of amalgamation,loosely to densely packed biofabric and a fine tomedium-grained sandy matrix mixed with abundantshell debris. At the base, truncated vertical trace fossilsoccur (Lazo, 2001b). Large ammonites are concordantwhile articulated and fragmented bivalves are randomlyorientated in cross-sectional view.
Bakevelliids are rarely found in shell beds B and Hwhile they are abundant in the closely successive shellbeds F and G (Fig. 9). Trophic nuclei of shell beds F andG are given in the Appendix section. Suspension-feedersdominate both associations. The cemented bivalveAmphidonte (Ceratostreon) sp. and the serpulidParsimonia antiquata (Sowerby) are fragmented and outof life position. Amphidonte (C.) sp. was also found
encrusting shells. Epifaunal, semi-infaunal, and infaunalorganisms are found out of life position.
Bakevelliid shells are reasonably well preserved,articulated but fragmented, with a low degree of mech-anical abrasion and a medium degree of bioerosion andencrustation (Table 2). Both bioerosion and encrus-tation have patchy distributions on shells. Only micro-borings (less than 5 mm) are especially common on thecentral body of shells. Encrusters include Serpulidae sp.A, Amphidonte (C.) sp., and bryozoans. Many of theencrusting oysters are still articulated.
Interpretation: The good preservation of the faunaand the presence of articulated bivalves are evidence of anegligible amount of lateral transport. High degrees ofvalve fragmentation, chipped margins, and randomlyorientated valves in cross section suggest a high energyenvironment and turbulence. Irregular bases, shell lags,and eroded vertical trace fossils indicate periods oferosion before the shell beds were deposited. Moderate
Fig. 9. Detailed stratigraphic section of the Pilmatue Member of the Agrio Formation at Bajada del Agrio, central Neuquen Province. The positionof bakevelliid-bearing shell beds is indicated (A–H). Ammonite zonation is based on Aguirre-Urreta and Rawson (1997, 1998, 2001).
Fig. 10. Bakevelliid-bearing facies in the Pilmatue Member of the Agrio Formation at Bajada del Agrio. A, Top and Middle facies of ashallowing-upwards cycle, 286.8 m above the base, H. agrioensis Subzone. B, plan view of shell bed D, Top facies of cycles, H. gentilii Zone. C, planview of shell bed G yielding articulated specimens of Gervillella aviculoides (J. Sowerby), Middle facies of cycles, H. gentilii Zone.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 779

Table 1Taphonomic, sedimentological, and stratigraphic attributes of bakevelliid-bearing shell beds in the Pilmatue Member of the Agrio Formation at Bajada del Agrio section
Bed Position inparasequence
Thickness Internalcomplexity
Basal contact Type ofmatrix
Dominantfraction
Shell-packing Orientation incross-section
Bakevelliidabundance
A top facies 0.15 to0.2 m
lowamalgamation
sharp and irregular with reworkednodules and bivalves
fine sandstone valve fragments loose chaotic, concordant,and stacked
common
B middle facies 0.16 m lowamalgamation
sharp and irregular mediumsandstone
valve fragments dense chaotic rare
C top facies 0.45 m complex,amalgamation
gradational to sharp coarsesandstone
valve fragments,corals and bivalves
dense concordant common
D top facies 0.5 m complex,amalgamation
sharp and irregular silt to finesandstone
Ceratostreon sp. andits fragments
dense chaotic rare
E top facies 0.4 m complex,amalgamation
sharp and irregular mediumsandstone
valve fragments dense chaotic rare
F middle facies 0.05 m none sharp and irregular with trace fossils fine sandstone fragments ofCeratostreon sp.
loose concordant, andchaotic
abundant
G middle facies 0.05 m none sharp and irregular with trace fossils fine sandstone valve fragments loose concordant, andchaotic
abundant
H middle facies 0.1 m none sharp and irregular with reworkedammonites and bivalves
fine sandstone fragments ofCeratostreon sp.
dense concordant, andchaotic
rare
D.
Gustavo
Lazo
/C
retaceousR
esearch24
(2003)765–788
780
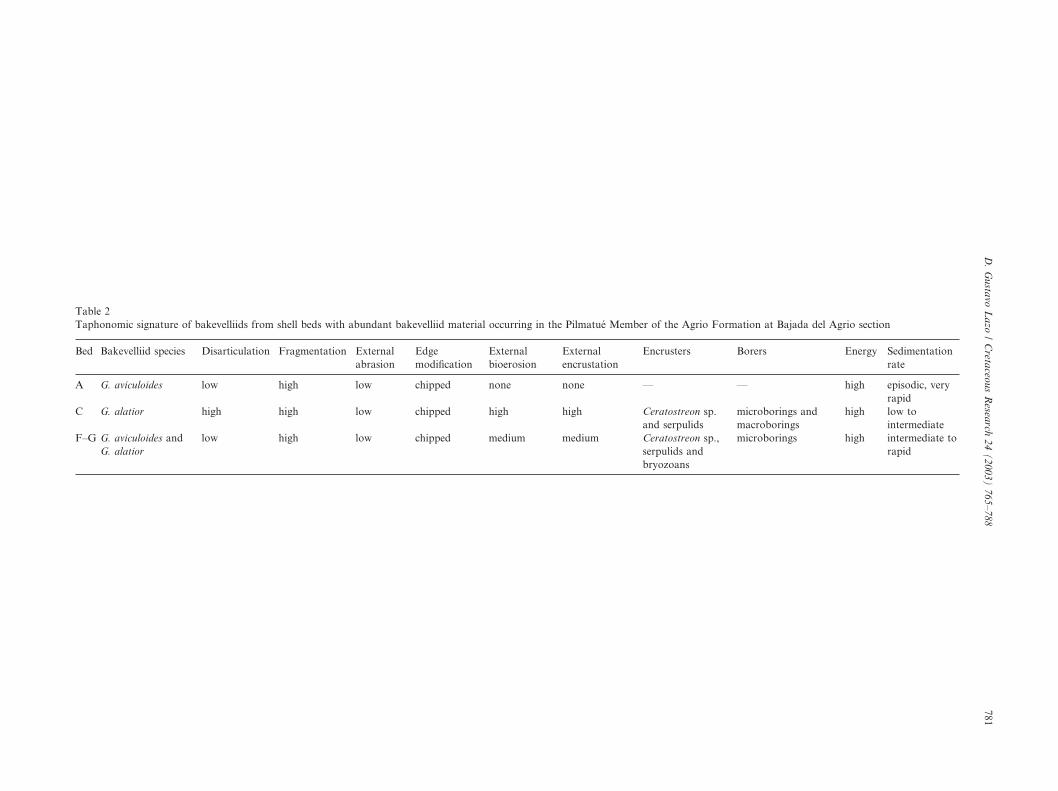
Table 2Taphonomic signature of bakevelliids from shell beds with abundant bakevelliid material occurring in the Pilmatue Member of the Agrio Formation at Bajada del Agrio section
Bed Bakevelliid species Disarticulation Fragmentation Externalabrasion
Edgemodification
Externalbioerosion
Externalencrustation
Encrusters Borers Energy Sedimentationrate
A G. aviculoides low high low chipped none none — — high episodic, veryrapid
C G. alatior high high low chipped high high Ceratostreon sp.and serpulids
microborings andmacroborings
high low tointermediate
F–G G. aviculoides andG. alatior
low high low chipped medium medium Ceratostreon sp.,serpulids andbryozoans
microborings high intermediate torapid
D.
Gustavo
Lazo
/C
retaceousR
esearch24
(2003)765–788
781

degrees of patchy bioerosion and encrustation suggest ashort period of exposure on the sea floor and rapid tovery rapid burial (Brett and Baird, 1986). A predomi-nance of suspension-feeders points to water energy levelhigh enough to keep nutrients in suspension. The highnumber of epifaunal and semi-infaunal bivalves suggesta fairly stable substrate, which was soft enough toallow colonization by burrowers. Bivalves adapted tonear-surface horizontal burrowing point to deep-water,subtidal sediments (McAlester and Rhoads, 1967)and probably confirm a depositional setting belowfairweather wave base.
Stratigraphic, lithologic, and taphonomic data indi-cate the action of alternating storm and fairweatherprocesses, the latter represented by suspension fall-outpelites, which is typical of the mid-ramp. Middle faciesshell beds occur only occasionally, probably because of areduced availability of shelly material.
4.2.2. Top facies bakevelliid-bearing shell beds.Description: Shell beds A (Neocomites sp. Subzone),
C (H. agrioensis Subzone), D, and E (H. gentilii Zone)appear in the Top facies of cycles (Fig. 9, Fig. 10B).They are thick beds (0.15–0.5 m) showing a complexinternal structure, a low to high degree of amalgama-tion, a loosely to densely packed biofabric with ran-domly orientated valves, and a fine to coarse-grainedsandy matrix.
Bakevelliids are abundant in shell beds A and C, butrarely found in shell beds D and E. Trophic nuclei of thecoquinas A and C are given in the Appendix section.Suspension-feeders dominate both associations. In shellbed A shallow and deep burrowers predominate while incoquina C, burrowers and byssates are equal in number.Two isognomonid species are recorded in shell bed C(Appendix A), the only bed in which they have beenfound in the Pilmatue Member. The cemented bivalveAmphidonte (C.) sp. and serpulid P. antiquata (Sowerby)are fragmented and out of life position. Amphidonte (C.)sp. was also found encrusting shells.
Bakevelliids from shell bed A are well preserved,articulated but fragmented, with low external abrasion,and lack bioerosion or encrustation (Table 2). Bake-velliids from shell bed C are disarticulated, fragmented,not abraded, and show a high degree of bioerosion andencrustation (Table 2). Both bioerosion and encrust-ation are usually widespread over shells. Micro- andmacroborings (e.g. Gastrochaenolites isp.) are present.Encrusters include Serpulidae sp. A, Amphidonte (C.)sp., and bryozoans. Most of the encrusting oysters aredisarticulated.
Interpretation: Bakevelliid-bearing shell beds in theTop facies are represented by two types with differenttaphonomic signatures. Shell bed A yields well-preserved shells. Their high articulation level, and
absence of bioerosion or encrustation, suggest a shorttime of exposure and episodic, very rapid burial. Emptyammonites also reflect rapid deposition and burial (Brettand Baird, 1986). A high degree of valve fragmentationand random orientation in cross section point to a highenergy environment. Irregular bases, reworked smallnodules and reworked bivalves indicate periods oferosion before deposition of the shell bed. Abundantsuspension-feeders suggest a water energy level highenough to keep nutrients in suspension. The predomi-nance of shallow burrowers over deep infaunal speciesconfirms a soft substrate in a deep subtidal environment(McAlester and Rhoads, 1967). The stratigraphic, litho-logic and taphonomic data indicate a high energy andmoderately deep subtidal setting influenced by stormsand high sedimentation rate processes (mid-rampdeposits).
Shell bed C has a poorly preserved fauna. The highdegrees of disarticulation and fragmentation andchipped shell margins point to a high energy environ-ment and turbulence. High degrees of bioerosion andencrustation suggest a low to intermediate sedimenta-tion rate and a slow burial (Brett and Baird, 1986).Widespread Gastrochaenolites isp. indicate very shallowwaters (Bromley, 1992). Ripple cross lamination, a highdegree of amalgamation, densely packed biofabric, thecoarse-grained sandy matrix, and shell debris back-ground are interpreted as the result of wave and currentaction. The record of herbivorous gastropods suggests ashallow sea bottom placed well within the photic zoneand colonized by benthic algae (Aberhan, 1993). Thetwo species of Isognomon from shell bed C are evidencefor a shallow setting though they are probably par-autochthonous because Recent species of this genus liveepifaunally fixed by a byssus to hard substrates inintertidal to shallow subtidal environments (Stanley,1972; Crampton, 1988; Bernard et al., 1993). Insummary, the fossil association characterizes a highenergy and shallow subtidal setting within the photiczone with strong influence of storm- and possibly fair-weather waves and a low to intermediate sedimentationrate located in the upper mid-ramp.
5. Palaeobiology
As bakevelliids are extinct, a direct ecological com-parison with recent forms is not possible. To elucidatetheir life habits a functional analysis of the shell mor-phology is of some help (Stanley, 1970; McGhee, 1978;Seilacher, 1984; Aberhan and Muster, 1997). Althoughfunctional morphology is a valid approach to auto-ecology, not all the shell characters can be seen asadaptive in origin since body plan and architecturalconstraints can be more important in delimiting
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788782

pathways of change than natural selection (Gould andLewontin, 1979).
Gervillaria alatior has its maximum width locatednear the anterior margin, as shown in serial cross-sections (Fig. 11A, B). The anterior margin is wide andflat (triangular cross shape). Such a flat anterior surfacewith the widest dimension of the shell lying near theanterior margin is typical of recent epifaunal mytiloids.It lowers the shell weight centre and provides a broadcontact surface with the substratum (Stanley, 1970). Areduced anterior region (anisomyarian or monomyariancondition) has been related to a firmer byssus attach-ment in epifaunal and semi-infaunal mytiloids (Stanley,1970, 1972). Aberhan and Muster (1997) have inter-preted an antero-dorsal triangular groove such as that inG. alatior, dividing the dentition from the anteriormargin of the shell (on the internal shell surface), as abyssal groove (Fig. 6C, F). During the ligament devel-opment, the maximum deposition of shell material wason the left umbo. This heavy umbo would have providedstability for substrate anchorage (McGhee, 1978). HenceG. alatior is here interpreted as an epibyssate bivalve(edgewise recliners in Seilacher, 1984) (Fig. 12). Thebroad anterior margin provided a wide anchorage sur-face. The presence of a byssus is suggested by thewell-developed byssal groove. The main stabilizing
factor in the adult stage seems to have been the weightyumbo, leaving the bivalve completely sessile. Owing toits epifaunal life habit G. alatior could have live on softor hard bottoms.
In Gervillella aviculoides, the maximum width liesapproximately midway between anterior and posteriormargins, as seen in serial cross-sections (Fig. 11C). Thereduced anterior margin is wedge-shaped. These featuresare typical of semi-infaunal mytiloids (Stanley, 1970,1972). Thus G. aviculoides is here interpreted as semi-infaunal (mud sticker in Seilacher, 1984) (Fig. 13),probably stabilized in the substrate by a byssus, whichallowed the penetration of the growing bivalve into asoft bottom. It has not been possible to estimate theexact angle formed between the shell height and thesubstrate or the penetration depth.
G. alatior shows clear affinities with small specimensof G. alaeformis, which suggests that G. alatior was aneotenic form that had a reduced rate of morphologicaldevelopment during the juvenile phase resulting ina morphologically retarded adult (see neoteny inMcNamara, 1986).
Both species are clearly facies-dependent in thePilmatue Member of the Agrio Formation, as they arerestricted to the mid-ramp and upper mid-rampdeposits. Their wide palaeobiogeographic distribution
Fig. 11. Gervillaria alatior (Imlay). A, transverse section of articulated shell, CPBA 19738.3. B, transverse section of articulated shell, CPBA19731.30. Gervillella aviculoides (J. Sowerby). C, transverse section of articulated shell, CPBA 19752.2. A and C from Bajada del Agrio; B from Aguade la Mula. All �1.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 783

may be explained by an extensive larval stage, whichcould have enhanced the oceanic dispersal of thesespecies. Their larvae probably had the ability to post-pone settling until they found a substrate suitable foradult life as in other marine benthic invertebrates(Thorson, 1957). The attractive substrates were prob-ably shallow marine sands or shell gravels located in themid-ramp.
Acknowledgements
Special thanks go to Marıa Beatriz Aguirre-Urretaand Silvia Lanes (Department of Geology, University ofBuenos Aires) whose comments and suggestions greatlyimproved the original manuscript and who gave con-stant encouragement. Julian Petrulevicius sent meimportant literature from the library of the MuseeNational d’Histoire Naturelle (Paris). David Batten,Noel Morris and Peter Rawson are acknowledged fortheir critical review of the manuscript, making it pub-lishable. Thanks are due to Marıa Beatriz Aguirre-Urreta, Peter Rawson, Andrea Concheyro, MarcelaCichowolski, Romina Sanci, Silvia Lanes, CeciliaRodriguez and Gaston Iovine for their help in thefield. This study was partially funded by UBACyTTX 155 and CONICET PIP 360/98 grants to MarıaBeatriz Aguirre-Urreta, and an IAS postgraduate grantto me.
Fig. 12. Life habit of Gervillaria alatior (Imlay) from the Lower Cretaceous of the Neuquen Basin. An epibyssate habit is interpreted for this species,which has a triangular cross-section. The reconstruction is based on analysis of functional morphology. Transverse sections are indicated by stripes.The arrows denote the position of the cross section. The angle between the shell and the sediment-water interface may be variable. Scale barrepresents 5 mm.
Fig. 13. Life habit of Gervillella aviculoides (Sowerby) from the LowerCretaceous of the Neuquen Basin. An endobyssate habit is proposedfor this species, which has a rounded cross-section. The reconstructionis based on analysis of functional morphology. Transverse sections areindicated by stripes. Arrows denote the position of cross section. Theangle between the shell and the sediment-water interface may bevariable. Scale bar represents 5 mm.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788784
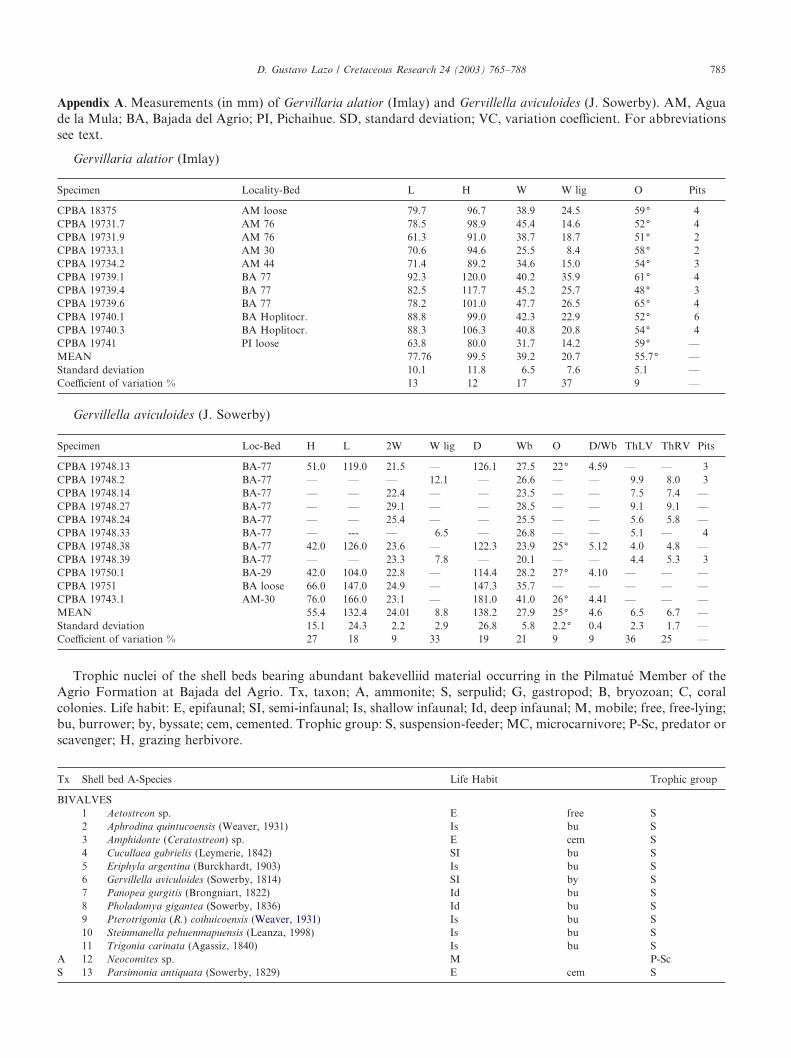
Appendix A. Measurements (in mm) of Gervillaria alatior (Imlay) and Gervillella aviculoides (J. Sowerby). AM, Aguade la Mula; BA, Bajada del Agrio; PI, Pichaihue. SD, standard deviation; VC, variation coefficient. For abbreviationssee text.
Gervillaria alatior (Imlay)
Specimen Locality-Bed L H W W lig O Pits
CPBA 18375 AM loose 79.7 96.7 38.9 24.5 59( 4CPBA 19731.7 AM 76 78.5 98.9 45.4 14.6 52( 4CPBA 19731.9 AM 76 61.3 91.0 38.7 18.7 51( 2CPBA 19733.1 AM 30 70.6 94.6 25.5 8.4 58( 2CPBA 19734.2 AM 44 71.4 89.2 34.6 15.0 54( 3CPBA 19739.1 BA 77 92.3 120.0 40.2 35.9 61( 4CPBA 19739.4 BA 77 82.5 117.7 45.2 25.7 48( 3CPBA 19739.6 BA 77 78.2 101.0 47.7 26.5 65( 4CPBA 19740.1 BA Hoplitocr. 88.8 99.0 42.3 22.9 52( 6CPBA 19740.3 BA Hoplitocr. 88.3 106.3 40.8 20.8 54( 4CPBA 19741 PI loose 63.8 80.0 31.7 14.2 59( —MEAN 77.76 99.5 39.2 20.7 55.7( —Standard deviation 10.1 11.8 6.5 7.6 5.1 —Coefficient of variation % 13 12 17 37 9 —
Gervillella aviculoides (J. Sowerby)
Specimen Loc-Bed H L 2W W lig D Wb O D/Wb ThLV ThRV Pits
CPBA 19748.13 BA-77 51.0 119.0 21.5 — 126.1 27.5 22( 4.59 — — 3CPBA 19748.2 BA-77 — — — 12.1 — 26.6 — — 9.9 8.0 3CPBA 19748.14 BA-77 — — 22.4 — — 23.5 — — 7.5 7.4 —CPBA 19748.27 BA-77 — — 29.1 — — 28.5 — — 9.1 9.1 —CPBA 19748.24 BA-77 — — 25.4 — — 25.5 — — 5.6 5.8 —CPBA 19748.33 BA-77 — --- — 6.5 — 26.8 — — 5.1 — 4CPBA 19748.38 BA-77 42.0 126.0 23.6 — 122.3 23.9 25( 5.12 4.0 4.8 —CPBA 19748.39 BA-77 — — 23.3 7.8 — 20.1 — — 4.4 5.3 3CPBA 19750.1 BA-29 42.0 104.0 22.8 — 114.4 28.2 27( 4.10 — — —CPBA 19751 BA loose 66.0 147.0 24.9 — 147.3 35.7 — — — — —CPBA 19743.1 AM-30 76.0 166.0 23.1 — 181.0 41.0 26( 4.41 — — —MEAN 55.4 132.4 24.01 8.8 138.2 27.9 25( 4.6 6.5 6.7 —Standard deviation 15.1 24.3 2.2 2.9 26.8 5.8 2.2( 0.4 2.3 1.7 —Coefficient of variation % 27 18 9 33 19 21 9 9 36 25 —
Trophic nuclei of the shell beds bearing abundant bakevelliid material occurring in the Pilmatue Member of theAgrio Formation at Bajada del Agrio. Tx, taxon; A, ammonite; S, serpulid; G, gastropod; B, bryozoan; C, coralcolonies. Life habit: E, epifaunal; SI, semi-infaunal; Is, shallow infaunal; Id, deep infaunal; M, mobile; free, free-lying;bu, burrower; by, byssate; cem, cemented. Trophic group: S, suspension-feeder; MC, microcarnivore; P-Sc, predator orscavenger; H, grazing herbivore.
Tx Shell bed A-Species Life Habit Trophic group
BIVALVES1 Aetostreon sp. E free S2 Aphrodina quintucoensis (Weaver, 1931) Is bu S3 Amphidonte (Ceratostreon) sp. E cem S4 Cucullaea gabrielis (Leymerie, 1842) SI bu S5 Eriphyla argentina (Burckhardt, 1903) Is bu S6 Gervillella aviculoides (Sowerby, 1814) SI by S7 Panopea gurgitis (Brongniart, 1822) Id bu S8 Pholadomya gigantea (Sowerby, 1836) Id bu S9 Pterotrigonia (R.) coihuicoensis (Weaver, 1931) Is bu S10 Steinmanella pehuenmapuensis (Leanza, 1998) Is bu S11 Trigonia carinata (Agassiz, 1840) Is bu S
A 12 Neocomites sp. M P-ScS 13 Parsimonia antiquata (Sowerby, 1829) E cem S
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 785

Tx Shell bed C-Species Life Habit Trophic group
BIVALVES1 Amphidonte (Ceratostreon) sp. E cem S2 Cucullaea gabrielis (Leymerie, 1842) SI bu S3 Eriphyla argentina (Burckhardt, 1903) Is bu S4 Gervillaria alatior (Imlay, 1940) E by-free S5 Isognomon (I.) lotenoensis (Weaver, 1931) E by S6 Isognomon (I.) ricordeanus (d’Orbigny, 1845) E by S7 Mimachlamys robinaldina (d’Orbigny, 1847) E by-free S8 Myoconcha transtlantica (Burckhardt, 1903) E free S9 Pterotrigonia (R.) coihuicoensis (Weaver, 1931) SI bu S10 Ptychomya vacaensis (Weaver, 1931) Is bu S11 Steinmanella transitoria (Steinmann, 1881) Is bu S12 Pinna (P.) robinaldina (d’Orbigny, 1844) SI by S
A 13 Holcoptychites compressum (Leanza and Wiedmann, 1980) M P-ScS 11 Serpulidae sp. A En cem SG 14 Pleurotomaria gerthi (Weaver, 1931) E free HC 15 Columastrea antiqua (Gerth, 1928) E cem MC
Tx Shell bed F-Species Life Habit Trophic group
BIVALVES1 Amphidonte (Ceratostreon) sp. E cem S2 Cucullaea gabrielis (Leymerie, 1842) SI bu S3 Gervillella aviculoides (Sowerby, 1814) SI by S4 Myoconcha transtlantica (Burckhardt, 1903) E free S5 Ptychomya koeneni (Behrendsen, 1892) Is bu S6 Trigonia carinata (Agassiz, 1840) SI bu S
A 7 Hoplitocrioceras sp. M P-ScS 8 Parsimonia antiquata (Sowerby, 1829) E cem S
9 Serpulidae sp. A En cem SB 10 Bryozoan sp. A En cem S
Tx Shell bed G-Species Life Habit Trophic group
BIVALVES1 Amphidonte (Ceratostreon) sp. E cem S2 Cucullaea gabrielis (Leymerie, 1842) SI bu S3 Gervillaria alatior (Imlay, 1940) E by-free S4 Gervillella aviculoides (Sowerby, 1814) SI by S5 Mimachlamys robinaldina (d’Orbigny, 1847) E by-free S6 Myoconcha transtlantica (Burckhardt, 1903) E free S7 Ptychomya koeneni (Behrendsen, 1892) Is bu S8 Steinmanella transitoria (Steinmann, 1881) Is bu S9 Trigonia carinata (Agassiz, 1840) SI bu S
A 10 Hoplitocrioceras sp. M P-ScS 11 Serpulidae sp. A En cem SB 12 Bryozoan sp. A En cem S
References
Aberhan, M., 1993. Benthic macroinvertebrate associations on acarbonate-clastic ramp in segments of the Early Jurassic back-arcbasin of northern Chile (26–29(S). Rev. Geol. Chile 20, 105–136.
Aberhan, M., 1994. Early Jurassic Bivalvia of northern Chile. Part I.Subclasses Palaeotaxodonta, Pteriomorphia, and Isofilibranchia.Beringeria 13, 3–115.
Aberhan, M., Muster, H., 1997. Palaeobiology of Early Jurassicbakevelliid bivalves from Western Canada. Palaeontology 40,799–815.
Aguirre-Urreta, M.B., Lo Forte, G.L. 1996. Los depositos tithoneo-comianos. In: Ramos, V.A., Aguirre-Urreta, M.B., Alvarez, P.P.et al. (Eds.). Geologıa de la region del Aconcagua. Provincias deSan Juan y Mendoza, Republica Argentina Direccion Nacional del
Servicio Geologico, Subsecretarıa de Minerıa de la Nacion Anales24, 179–229 (Buenos Aires).
Aguirre-Urreta, M.B., Rawson, P.F., 1997. The ammonite sequence inthe Agrio Formation (Lower Cretaceous), Neuquen Basin,Argentina. Geol. Mag. 134, 449–458.
Aguirre-Urreta, M.B., Rawson, P.F., 1998. The Early Cretaceous(Valanginian) ammonite Chacantuceras gen. nov. – a link betweenthe Neuquen and Austral basins. Rev. Asoc. Geol. Argentina 53,354–364.
Aguirre-Urreta, M.B., Rawson, P.F., 2001. Lower Cretaceousammonites from the Neuquen Basin, Argentina: a HauterivianOlcostephanus fauna. Cret. Res. 22, 763–778.
Aguirre-Urreta, M.B., Rawson, P.F., 2003. Lower Cretaceousammonites from the Neuquen Basin, Argentina: the Hauteriviangenus Holcoptychites. Cret. Res. 24, 589–613.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788786

Arkell, W.J., 1933. A monograph of British CorallianLamellibranchia. Palaeontographical Society Monographs 5,181–228, 8 pls.
Behrendsen, O., 1892. Zur Geologie des Ostabhanges derargentinischen Cordillere. II. Theil. Zeitschrift der Deutschengeologischen Gesellschaft 44, 1–42, 4 pls.
Bernard, F.R., Cai, Y.Y., Morton, B., 1993. Catalogue of livingmarine bivalve molluscs of China. Hong Kong University Press,Hong Kong, 146 pp.
Beurlen, K., 1944. Beitrage zur Stammesgeschichte der Muscheln.Bayerische Akademie der Wissenschaften. Sitzungsberichte 1–2,133–145.
Brett, C.E., Baird, G.C., 1986. Comparative taphonomy: a key topaleoenvironmental interpretation based on fossil preservation.Palaios 1, 207–227.
Brinkmann, H-D., 1994. Facies and sequences of the Agrio Formation(Lower Cretaceous) in the central and southern Neuquen Basin,Argentina. Zentralblatt fur Geologie und Palaeontologie Teil I(1/2), 309–317.
Bromley, R.G., 1992. Bioerosion: eating rocks for fun and profit. In:Maples, C.G., West, R.R. (Eds.). Trace Fossils. Short Courses inPaleontology, 5. Paleontological Society, Knoxville, Tenessee,pp. 121–129.
Burchette, T.P., Wright, V.P., 1992. Carbonate ramp depositionalsystems. Sedimen. Geol. 79, 3–57.
Burckhardt, C., 1903. Beirtrage zur Kenntnis der Jura- undKreideformation der Cordillere. Palaeontographica 50, 1–144,16 pls.
Camacho, H.H., Riccardi, A.C. 1978. Invertebrados. Megafauna.Relatorio del VII Congreso Geologico Argentino, Neuquen,pp. 137–146, 4 tabls.
Carozzi, A.V., Bercowski, F., Rodriguez, M., et al., 1981. Estudio demicrofacies de la Formacion Chachao (Valanginiano) provincia deMendoza. Actas del VIII Congr. Geol. Argentino, San Luis 2,545–565.
Cecca, F., Dhondt, A.V., Bogdanova, T.N., 1999. The Aptianstratigraphy of the southern Tuarkyr (NW Turkmenistan, CentralAsia). Riv. Italiana Paleontol. Stratigr. 105, 377–396.
Cox, L.R., 1940. The Jurassic lamellibranch fauna of Kachh (Cutch).Memoirs of the Geological Survey of India, Palaeontologia Indica(9) 3 (3), 1–157.
Cox, L.R., 1954. Taxonomic notes on Isognomonidae andBakevelliidae. Proc. Malacol. Soc. London 31 (2), 46–49.
Cox, L.R., 1969. Family Bakevelliidae. In: Moore, R.C., Teichert, C.(Eds.). Treatise on Invertebrate Paleontology. Part N, Mollusca 6,Bivalvia 1. Geological Society of America/University of KansasPress, Boulder/Lawrence, pp. N306–N310.
Crampton, J.S., 1988. Comparative taxonomy of the bivalvefamilies Isognomonidae, Inoceramidae, and Retroceramidae.Palaeontology 31, 965–996.
Damborenea, S.E., 1987. Early Jurassic Bivalvia of Argentina.Part 2: Superfamilies Pteriacea, Buchiacea and part of Pectinacea.Palaeontographica A 199, 113–216, 14 pls.
Damborenea, S.E., Mancenido, M.O., Riccardi, A.C. 1979. Estudiopaleontologico de la Formacion Chachao. YPF, UnpublishedInternal Report 1–152, 18 pls., 18 tabls.
Dhondt, A.V., Dieni, I., 1988. Early Cretaceous bivalves of easternSardinia. Mem. Sci. Geol. 40, 1–97.
Freneix, S., 1995. Bivalves cretaces du bassin de Tarfaya (Marocmeridional atlantique): signification ecologique et importancepaleobiogeographique. Bull. Mus. Nat. d’Hist. Nat., Paris, sectionC 2–4 (4( ser), 243–258.
Goldfuss, G.A. 1833–1841. Petrefacta Germaniae II, 1–312, 94 pls.(Dusseldorf).
Gould, S.J., Lewontin, R.C., 1979. The Spandrels of San Marco andthe Panglossian paradigm: a critique of the adaptationistprogramme. Proc. Royal Soc. London B 205, 581–598.
Gray, J.E., 1847. A list of genera of recent Mollusca, their synonymsand types. Proc. Zool. Soc. London 15, 129–219.
Grobben, C., 1894. Zur Kenntniss der Morphologie, der Verwandt-schaftsverhaltnisse und des Systems der Mollusken. Akad.Wissensch. Wien, Sitzungsber 103, 61–86.
Gulisano, C.A., Gutierrez Pleimling, A.R., 1988. Depositos eolicos delMiembro Avile (Cretacico Inferior) en el norte del Neuquen,Argentina. Act. II Reun. Argentina Sedimentol., Buenos Aires 1,120–124.
Hayami, I., 1957. Liassic Gervillia and Isognomon in Japan (Studieson the Liassic pelecypods in Japan 2). Japanese J. Geol. Geogr. 28,95–106, 2 pls.
Imlay, R.W., 1940. Neocomian faunas of northern Mexico. Bull. Geol.Soc. America 51, 117–190.
Johnston, P.A., Collom, C.J., 1998. The bivalve heresies–Inoceramidae are Cryptodonta, not Pteriomorphia. In: Johnston,P.A., Haggart, J.W. (Eds.). Bivalves: an Eon of Evolution. Paleo-biological Studies Honoring Norman D. Newell. University ofCalgary Press, Calgary, pp. 347–360.
King, W. 1850. A monograph of the Permian fossils of England.Palaeontographical Society Monographs 3(5), 1–258, 28 pls.
Lazo, D.G. 2001a. Myophorellids (Bivalvia, Trigonioidea) of theAgrio Formation (Lower Cretaceous), Neuquen Basin, Argentina.Actas XI Congreso Latinoamericano de Geologıa y III CongresoUruguayo de Geologıa, Montevideo, 10–8.
Lazo, D.G. 2001b. Distribucion ambiental de trazas fosiles en elMiembro Inferior de la Formacion Agrio, Cretacico de CuencaNeuquina. Actas IV Reunion Argentina de Icnologıa y II Reunionde Icnologıa del Mercosur, Tucuman, 52.
Lazo, D.G., 2002. Bakevellidos (Bivalvia, Pteriomorphia) delCretacico Inferior de la Cuenca Neuquina. Act. XV Congr. Geol.Argentino, Calafate 1, 508–512.
Leanza, A.F., 1942. Los pelecıpodos del Lıas de Piedra Pintada en elNeuquen. Rev. Mus. La Plata (Nueva Serie) Secc. Paleontol. 2,143–206, 19 pls.
Legarreta, L., Kozlowski, E., Bol, A., 1981. Esquema estratigrafico ydistribucion de facies del Grupo Mendoza en el ambito surmendo-cino de la Cuenca Neuquina. Act. VIII Congr. Geol. Argentino,San Luis 3, 389–409.
Leymerie, M.A., 1842. Suite du memoire sur le terrain Cretace dudepartement de l’Aube. Second Partie (Partie Paleontologique).Mem. Soc. Geol. France ser. 1 (5), 1–34, 18 pls.
Linne von, C., 1758. Systema naturae per tria regna naturae. Edit.decima, 1. Laurentius, Stockholm, 1–824.
Mancenido, M.O., Damborenea, S.E., 1984. Megafauna de inverte-brados Paleozoicos y Mesozoicos. Rel. IX Congr. Geol. Argentino,San Carlos de Bariloche 2, 413–465.
Matthews, S.C., 1973. Notes on open nomenclature and on synonymylists. Palaeontology 16, 713–719.
McAlester, A.L., Rhoads, D.C., 1967. Bivalves as bathymetricindicators. Mar. Geol. 5, 383–388.
McGhee, G.R. Jr, 1978. Analysis of the shell torsion phenomenon inthe Bivalvia. Lethaia 11, 315–329.
McNamara, K.J., 1986. A guide to nomenclature of heterochrony. J.Paleontol. 60, 4–13.
Mombru, C.A., Uliana, M.A., Bercowski, F., 1978. Estratigrafıa ysedimentologıa de las acumulaciones biocarbonaticas del CretacicoInferior surmendocino. Act. VII Congr. Geol. Argentino, Neuquen1, 685–700.
Muster, H., 1995. Taxonomie und palaobiogeographie derBakevelliidae. Beringeria 14, 3–161.
Newell, N.D., 1965. Classification of the Bivalvia. Am. Mus. Nov.2206, 1–25.
Newell, N.D., Boyd, D.W., 1987. Iteration of ligament structures inpteriomorphian bivalves. Am. Mus. Nov. 2875, 1–11.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788 787

Nullo, F.E., Panza, J.L., Blasco, G., 1999. Jurasico y Cretacico de lacuenca Austral. Geol. Argentina, Inst. Geol. Rec. Min., Anales 29(17), 528–535.
Orbigny, A. d’. 1843–1847. Paleontologie Francaise. Descriptionzoologique et geologique de tous les animaux mollusques etrayonnes fossiles de France. In: Bertrand, A. (Ed.). TerrainsCretaces, Tome troisieme (Lamellibranches), 1–807, 253 pls.(Paris).
Orbigny A. d’. 1850. Prodrome de Paleontologie stratigraphiqueuniverselle des animaux mollusques et rayonnes II, 1–428. (Paris).
Pictet, F.J. & Roux, W. 1847–1853. Description des mollusquesfossiles qui se trouvent dans les Gres Verts des environs de Geneve,558 pp, 51 pls. (Geneve).
Riccardi, A.C., 1988. The Cretaceous System of southern SouthAmerica. Geol. Soc. Am. Mem. 168.
Riccardi, A.C., Damborenea, S.E., Mancenido, M.O., 1993. Inverte-brados Mesozoicos. Geol. Rec. Nat. Mendoza, Relatorio 2 (12),329–340.
Riccardi, A.C., Damborenea, S.E., Mancenido, M.O., et al., 1999. 3.Bioestratigrafıa. Geol. Argentina. El Jurasico y Cretacico de laCordillera Principal y la Cuenca Neuquina. Inst. Geol. Rec. Min.Anales 29 (16), 419–432.
Rodriguez Schelotto, M.L., Orchuela, I., Balina, M., et al., 1981.Medios depositacionales y microfacies de la Formacion Quintuco(Berriasiano–Valanginiano) en el yacimiento Loma La Lata. Pro-vincia del Neuquen. Act. VIII Congr. Geol. Argentino, San Luis 2,503–520.
Seilacher, A., 1984. Constructional morphology of bivalves: evolution-ary pathways in primary versus secondary soft-bottom dwellers.Palaeontology 27, 207–237.
Sowerby, J., 1819. The Mineral Conchology of Great Britain, pp. 1–4.Meredith, London.
Spalletti, L., Poire, D., Pirrie, D., et al., 2001. Respuesta sedimento-logica a cambios en el nivel de base en una secuencia mixtaclastica-carbonatica del Cretacico de la Cuenca Neuquina,Argentina. Rev. Soc. Geol. Espana 14, 57–74.
Stanley, S.M., 1970. Relations of shell form to life habits of theBivalvia (Mollusca). Mem. Geol. Soc. America 125.
Stanley, S.M., 1972. Functional morphology and evolution of byssallyattached bivalve mollusks. J. Paleontol. 46, 165–212.
Stanton, T.W., 1901. The marine Cretaceous invertebrates. In: Scott,W.B. (Ed.). Reports of the Princeton University Expeditions toPatagonia, 1896–1899. 4 Palaeontology, 1, pp. 1–43, 10 pls.
Taylor, J.D., 1973. The structural evolution of the bivalve shell.Palaeontology 16, 519–534.
Taylor, J.D., Kennedy, W.J., Hall, A., 1969. The shell structure andmineralogy of the Bivalvia. Introduction. Nuculacea-Trigoniacea.Bull. Br. Mus. (Nat. Hist.) Zoology 3, 125 pp. 29 pls.
Thorson, G., 1957. Chapter 17. Bottom communities (sublittoral orshallow shelf). Geol. Soc. America Mem. 67 (1), 461–534.
Uliana, M.A., Dellape, D.A., Pando, G.A., 1977. Analisis estrati-grafico y evaluacion del potencial petrolıfero de las FormacionesMulichinco, Chachao y Agrio. Cretacico Inferior de las provinciasde Neuquen y Mendoza. Petrotecnia, Enero-Febrero 1977, 31–46,Marzo 1977, 25–33.
Vergani, G., Cafferata, A., Boggetti, D., et al., 2002. Modelo de faciesde un sistema eolico en transicion a un lago efımero en el MiembroAvile (Hauteriviano), noroeste de la Cuenca Neuquina. Act. IXReun. Argentina Sedimentol., Cordoba, 42.
Vergani, G.D., Tankard, A.J., Belotti, H.J., et al., 1995. Tectonicevolution and paleogeography of the Neuquen Basin, Argentina.In: Tankard, A.J., Suarez, S.R., Welsink, H.J. (Eds.). Petroleumbasins of South America. American Association of PetroleumGeologists Memoir, 6, pp. 383–402.
Waagen, L. 1907. Die Lamellibranchiaten der Pachycardientuffeder Seiser Alm nebst vergleichend palaontologischen und phylo-genetischen Studien. Abhanlungen der Kaiserlich-KoniglichenGeologischen Reichsanstalt 18, 1–180, 34 pls. (Wien).
Waller, T.R., 1978. Morphology, morphoclines and a new classifica-tion of the Pteriomorphia (Mollusca: Bivalvia). Phil. Trans. RoyalSoc. London B 284, 345–365.
Waller, T.R., 1998. Origin of the molluscan class Bivalvia and aphylogeny of major groups. In: Johnston, P.A., Haggart, J.W.(Eds.). Bivalves: an Eon of Evolution. Paleobiological StudiesHonoring Norman D. Newell. University of Calgary Press,Calgary, pp. 1–45.
Weaver, C.E., 1931. Paleontology of the Jurassic and Cretaceous ofwest central Argentina. Memoirs of the University of Washington1, 1–595.
Woods, H. 1899–1913. A monograph of the Cretaceous Lamelli-branchiata of England. Palaeontographical Society MonographsI–II, 1–705, 104 pls.
Zavala, C., 2000. New advances in the sequence stratigraphy andsedimentology of the Mulichinco Formation, Neuquen Basin. Bol.Inform. Petroleras, Tercera Epoca 63, 40–54.
D. Gustavo Lazo / Cretaceous Research 24 (2003) 765–788788
Copyright © 2022 FDOKUMEN




















