
The influence of the structural orientation of amide linkers on the serum compatibility and lung...
10
The influence of the structural orientation of amide linkers on the serum compatibility and lung transfection properties of cationic amphiphiles Marepally Srujan a , Voshavar Chandrashekhar a , Rakesh C. Reddy a , Rairala Prabhakar a , Bojja Sreedhar b , Arabinda Chaudhuri a, * a Division of Lipid Science and Technology, Indian Institute of Chemical Technology, Hyderabad 500607, India b Inorganic and Physical Chemistry Division, Indian Institute of Chemical Technology, Hyderabad 500607, India article info Article history: Received 11 March 2011 Accepted 25 March 2011 Available online 17 April 2011 Keywords: Serum compatible lipofectins Selective lung transfection Linker orientation effect Fluorescence resonance energy transfer Biomembrane fusogenicity abstract Understanding the structural parameters of cationic amphiphiles which can influence gene transfer efficiencies of cationic amphiphiles continues to remain important for designing efficient liposomal gene delivery reagents. Previously we demonstrated the influence of structural orientation of the ester linker (widely used in covalently tethering the polar head and the non-polar tails) in modulating in vitro gene transfer efficiencies of cationic amphiphiles. However, our previously described cationic amphiphiles with ester linkers failed to deliver genes under in vivo conditions. Herein we report on the development of a highly serum compatible cationic amphiphile with circulation stable amide linker which shows remarkable selectivity in transfecting mouse lung. We also demonstrate that reversing structural orientation of the amide linker adversely affects both serum compatibility and the lung selective gene transfer property. Dynamic laser light scattering and atomic force microscopic studies revealed smaller average hydrodynamic sizes of the liposomes of transfection efficient lipid than those for the liposomes of transfection incompetent analog (148 1 nm vs 214 4 nm). Average surface potential of the liposomes of transfection competent amphiphiles were found to be significantly higher than that for the liposomes of transfection incompetent analog (10.7 5.4 mV vs 2.8 1.3 mV, respectively). Findings in fluorescence resonance energy transfer and dye entrapment experiments support lower rigidity and higher biomembrane fusogenicity of the liposomes of the transfection efficient amphiphiles. Importantly, cationic lipoplexes of the novel amide-linker based amphiphile exhibited higher mouse lung selective gene transfer properties than DOTAP, one of the widely used commercially available liposomal lung transfection kits. In summary, the present findings demonstrate for the first time that amide linker structural orientation profoundly influences the serum compatibility and lung transfection efficiencies of cationic amphiphiles. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Understanding the structural parameters of cationic amphi- philes which can influence their gene transfer properties is important for designing efficient liposomal gene delivery reagents. To this end, the focus of many prior structureeactivity investiga- tions has been centered around probing the influence of each of the three main structural components of cationic amphiphiles namely, head-groups, hydrophobic tails and the nature of the linker functionality used to covalently tether the polar head- groups with the hydrophobic tails [1e3]. For instance, a number of prior reports have demonstrated that the gene transfer efficiencies of cationic amphiphiles critically depends on their molecular architectures including hydrophobic alkyl chain-lengths [3e9], nature of head-groups [10e12] as well as on the nature of linker and spacer functionalities used in covalent tethering of the polar head-groups and the non-polar tails of cationic amphiphiles [11e 16]. More recently, the in vitro gene transfer efficiency of a cati- onic amphiphile with asymmetric hydrocarbon chains, namely oleoyldecanoyl-ethylphosphatidylcholine (C18:1/C10-EPC), has been demonstrated to be about 50 fold superior to that of its structurally very similar saturated asymmetric counterpart Abbreviations: Chol, cholesterol; DCM, dichloromethane; DMEM, Dulbecco’s modified Eagle’s medium; MEM, minimum essential medium; DMF, N,N-dimethyl formamide; FBS, fetal bovine serum; ONPG, o-nitrophenyl-b-D-galactopyranoside; PBS, phosphate buffered saline; GFP, green fluorescent protein.. * Corresponding author. Tel.: þ91 40 27193201; fax: þ91 40 27193370. E-mail address: [email protected] (A. Chaudhuri). Contents lists available at ScienceDirect Biomaterials journal homepage: www.elsevier.com/locate/biomaterials 0142-9612/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.biomaterials.2011.03.059 Biomaterials 32 (2011) 5231e5240
-
Upload
praveenlab -
Category
Documents
-
view
1 -
download
0
Transcript of The influence of the structural orientation of amide linkers on the serum compatibility and lung...

lable at ScienceDirect
Biomaterials 32 (2011) 5231e5240
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
The influence of the structural orientation of amide linkers on the serumcompatibility and lung transfection properties of cationic amphiphiles
Marepally Srujan a, Voshavar Chandrashekhar a, Rakesh C. Reddy a, Rairala Prabhakar a,Bojja Sreedhar b, Arabinda Chaudhuri a,*aDivision of Lipid Science and Technology, Indian Institute of Chemical Technology, Hyderabad 500607, Indiab Inorganic and Physical Chemistry Division, Indian Institute of Chemical Technology, Hyderabad 500607, India
a r t i c l e i n f o
Article history:Received 11 March 2011Accepted 25 March 2011Available online 17 April 2011
Keywords:Serum compatible lipofectinsSelective lung transfectionLinker orientation effectFluorescence resonance energy transferBiomembrane fusogenicity
Abbreviations: Chol, cholesterol; DCM, dichlorommodified Eagle’s medium; MEM, minimum essentialformamide; FBS, fetal bovine serum; ONPG, o-nitropPBS, phosphate buffered saline; GFP, green fluorescen* Corresponding author. Tel.: þ91 40 27193201; fax
E-mail address: [email protected] (A. Chaudhuri
0142-9612/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.biomaterials.2011.03.059
a b s t r a c t
Understanding the structural parameters of cationic amphiphiles which can influence gene transferefficiencies of cationic amphiphiles continues to remain important for designing efficient liposomal genedelivery reagents. Previously we demonstrated the influence of structural orientation of the ester linker(widely used in covalently tethering the polar head and the non-polar tails) in modulating in vitro genetransfer efficiencies of cationic amphiphiles. However, our previously described cationic amphiphileswith ester linkers failed to deliver genes under in vivo conditions. Herein we report on the developmentof a highly serum compatible cationic amphiphile with circulation stable amide linker which showsremarkable selectivity in transfecting mouse lung. We also demonstrate that reversing structuralorientation of the amide linker adversely affects both serum compatibility and the lung selective genetransfer property. Dynamic laser light scattering and atomic force microscopic studies revealed smalleraverage hydrodynamic sizes of the liposomes of transfection efficient lipid than those for the liposomesof transfection incompetent analog (148 � 1 nm vs 214 � 4 nm). Average surface potential of theliposomes of transfection competent amphiphiles were found to be significantly higher than that for theliposomes of transfection incompetent analog (10.7 � 5.4 mV vs 2.8 � 1.3 mV, respectively). Findings influorescence resonance energy transfer and dye entrapment experiments support lower rigidity andhigher biomembrane fusogenicity of the liposomes of the transfection efficient amphiphiles. Importantly,cationic lipoplexes of the novel amide-linker based amphiphile exhibited higher mouse lung selectivegene transfer properties than DOTAP, one of the widely used commercially available liposomal lungtransfection kits. In summary, the present findings demonstrate for the first time that amide linkerstructural orientation profoundly influences the serum compatibility and lung transfection efficiencies ofcationic amphiphiles.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Understanding the structural parameters of cationic amphi-philes which can influence their gene transfer properties isimportant for designing efficient liposomal gene delivery reagents.To this end, the focus of many prior structureeactivity investiga-tions has been centered around probing the influence of each ofthe three main structural components of cationic amphiphiles
ethane; DMEM, Dulbecco’smedium; DMF, N,N-dimethylhenyl-b-D-galactopyranoside;t protein..: þ91 40 27193370.).
All rights reserved.
namely, head-groups, hydrophobic tails and the nature of thelinker functionality used to covalently tether the polar head-groups with the hydrophobic tails [1e3]. For instance, a number ofprior reports have demonstrated that the gene transfer efficienciesof cationic amphiphiles critically depends on their moleculararchitectures including hydrophobic alkyl chain-lengths [3e9],nature of head-groups [10e12] as well as on the nature of linkerand spacer functionalities used in covalent tethering of the polarhead-groups and the non-polar tails of cationic amphiphiles[11e16].
More recently, the in vitro gene transfer efficiency of a cati-onic amphiphile with asymmetric hydrocarbon chains, namelyoleoyldecanoyl-ethylphosphatidylcholine (C18:1/C10-EPC), hasbeen demonstrated to be about 50 fold superior to that ofits structurally very similar saturated asymmetric counterpart

M. Srujan et al. / Biomaterials 32 (2011) 5231e52405232
stearoyldecanoyl-ethylphosphatidylcholine (C18:0/C10-EPC) [17].In 2007, we reported on another important structural parameter,linker orientation, which can dramatically influence the trans-fection efficiencies of cationic amphiphiles [18]. We designed &synthesized two structurally isomeric remarkably similar cationicamphiphiles 1 & 2 (Fig.1, Part A) bearing the same hydrophobic tailsand the same polar head-groups connected by the same ester linkergroup [18]. The only structural difference between the two cationicamphiphiles 1 & 2 is the orientation of their linker ester function-ality. Despite having such striking structural similarity, only lipid 1could efficiently deliver plasmid DNA encoding b-galactosidaseenzyme into a number of cultured mammalian cells [18]. Con-trastingly, lipid 2 was found to be essentially incompetent indelivering b-galactosidase reporter gene into any of these cells [18].This prior work demonstrated for the first time the profoundinfluence of linker orientation in cationic lipid mediated genedelivery under in vitro conditions. However, in spite of its high invitro gene transfer activity, lipid 1 turned out to be essentiallyincompetent in delivering genes under in vivo conditions. Forexample, the mouse lung transfection efficiency of lipid 1 wasfound more than 100 fold less than that of DOTAP/chol, a widelyused commercially available liposomal in vivo transfection kit(Fig. S1, Supporting Information).We envisaged that lipid 1 failed toexhibit in vivo transfection efficiency presumably due to hydrolyticassault from systemic esterases. Stated differently, we envisagedthat demonstrating existence of linker orientation influence inliposomal gene delivery under in vivo conditions would requiredesigning cationic amphiphiles with more circulation stable linkerfunctionality. Based on such rationale, herein we report on thedevelopment of a highly serum compatible cationic amphiphilewith circulation stable amide linker which shows remarkableselectivity in transfecting mouse lung. Taken together, the presentfindings demonstrate for the first time that the structural orienta-tion of the amide linker profoundly influences the serum compat-ibility and lung transfection efficiencies of cationic amphiphiles.
2. Materials and methods
2.1. General procedures and reagents
Mass spectra data were acquired by using a commercial LCQ ion trap massspectrometer (ThermoFinnigan, SanJose, CA, USA) equipped with an ESI source ormicromassQuatro LC triple quadrupole mass spectrometer for ESI analysis. 1H NMRspectra were recorded on a Varian FT 400 MHz or AV 300 MHz NMR Spectrometer.Myristic acid, Benzylamine, 10% Palladium/charcoal were purchased from Lancaster(Morecambe, UK) and Tridecylamine, 2-Bromoethanol, 2-Iodo ethanol, Amberlite-IRA 400 Cl� were procured from SigmaeAldrich (USA). Unless otherwise stated,all other reagents were purchased from local suppliers including Rankem (India),sd fine chem-ltd (India), Loba chemie (India), Spectrochem (India) and HiMedia(India) and were used without further purification. Column chromatographywas performed with silica gel (Acme Synthetic Chemicals, India, 60e120 mesh).
N
O O
C C
C13H27C13H27
OO
OHH3C H3C
+
Cl-
Lipid 1
N
C C
O O
OH+
Lipid 2
OO
A
C13H27C13H27
Cl-
Fig. 1. Structures of lipids 1 & 2 with different orientation of ester linke
p-CMV-SPORT-b-gal plasmid was a generous gift from Dr. Nalam MadhusudhanaRao of Centre for Cellular and Molecular Biology, Hyderabad, India. CAL-B enzymewas a generous gift from Dr. Sanjit Kanjilal of Lipid Science and Technology, IICT,Hyderabad, India. Cell culture media, fetal bovine serum, 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT), polyethylene glycol 8000, o-nitro-phenyl-b-D-galactopyranoside and cholesterol were purchased from Sigma, St.Louis, USA. NP-40, antibiotics and agarose were purchased from Hi-media, India.HepG2 (Human hepatocarcinoma cells) and B16F10 (Murine melanoma cells),MCF-7 (Human breast cancer cells), CHO (Chinese hamster ovary cells), cells wereprocured from the National Centre for Cell Sciences (NCCS), Pune, India. Cells weregrown at 37 �C either in Dulbecco’s modified Eagle’s medium (DMEM) or MinimumEssential Medium (MEM) with 10% FBS in a humidified atmosphere containing 5%CO2/95% air. Elemental analysis (C, H, N) for lipids 3 & 4 was conducted atUniversity of Hyderabad.
2.2. Syntheses
The synthetic route used for preparing the lipids 3 & 4was shown in Scheme 1Aand B respectively.
2.2.1. Synthesis of (2-hydroxyethyl) (methyl) di [2-(tetradecanoylamino) ethyl]ammonium chloride (lipid 3, Scheme 1A)
Step i. Synthesis of N1-(2-[2-(tetradecanoylamino)ethyl]aminoethyl) tetra dec-anamide (Intermediate IA, Scheme 1A)
In a 100 mL single-neck round-bottom flask, diethylenetriamine (0.1 g,0.97 mmol) and tetradecanoic acid (0.49 g, 2.32 mmol) were dissolved in drytoluene. CAL-B enzyme (10% w/w) was added and the reaction mixture was stirredat 50 �C for 70 h. The reaction mixture was filtered off to remove the enzyme. To thefiltrate CAL-B enzyme (10% w/w) was added again and reaction continued at 50 �Cfor a further period of 48 h. The reaction mixture was filtered and concentrated. Theresidue upon column chromatographic purification (using 60e120 mesh size silicagel and 5:95 methanol:chloroform as eluent) afforded intermediate IA as a whitesolid (0.24 g, 62% yield, Rf ¼ 0.2, 5:95 methanol:chloroform, v/v).
1H NMRof (300MHz, CDCl3): d/ppm¼ 0.9 [t, 6H, CH3eCH2eC11H23e]; 1.2e1.4 [m,40H, CH3e(CH2)10e]; 1.6 [m, 4H, CH3e(CH2)10eCH2eCH2]; 2.1[t, 4H, CH3e(CH2)10eCH2eCH2]; 2.8 [t, 4H, NeCH2eCH2eNHeCOe]; 3.4[t, 4H, NeCH2eCH2eNHeCOe].
ESI-MS m/z: calcd 523.884 (for C32H65N3O2), found 524 [M]þ.
Step ii. Synthesis of N1-(2-(2-hydroxyethyl)[2-(tetradecanoylamino)ethyl]ami-noethyl) tetra decanamide (Intermediate IIA, Scheme 1A)
Intermediate IA (100 mg, 0.19 mmol) prepared in step i above was dissolved in10 mL of ethyl acetate in a 25 mL round-bottom flask. 2-bromoethanol (24 mg,0.19 mmol), K2CO3 (53 mg, 0.38 mmol) were added to the above solution. Reactionwas stirred at 80 �C for 48 h. The reaction mixturewas diluted with 100mL of CHCl3,and washed with water (2 � 50 mL). The organic layer was separated, dried overanhydrous sodium sulfate, and concentrated by rotary evaporator. The residue, uponcolumn chromatographic purification over 60e120mesh size silica gel column using4% methanol/chloroform (v/v) as eluent, afforded the pure title compound IIA asa white solid. (2.03 g, 80.3% yield, Rf ¼ 0.3, in 5% methanol:chloroform).
1HNMRof (300MHz, CDCl3): d/ppm¼0.9 [t, 6H, CH3eCH2eC11H23e]; 1.2e1.4 [m,40H, CH3e(CH2)10e]; 1.6 [m, 4H, CH3e(CH2)10eCH2eCH2]; 2.2[t, 4H, CH3e(CH2)10eCH2eCH2]; 2.6 [t, 2H, NeCH2eCH2eOH]; 2.7 [t, 4H, NeCH2eCH2eNHeCOe]; 3.3[t,4H, NeCH2eCH2eNHeCOe]; 3.6 [t, 2H, NeCH2eCH2eOH].
ESI-MS m/z: calcd 567.936(for C34H69N3O3), found 568 [Mþ], 569 [M þ H]þ.
N
HN NH
C C OO
OH+
Lipid 3
N
C C
NH NH
OH+
Lipid 4
OO
H3CH3CCl-
B
C13H27C13H27 C13H27C13H27
Cl-
r (A) and lipids 3 & 4 with different orientation of amide linker (B).
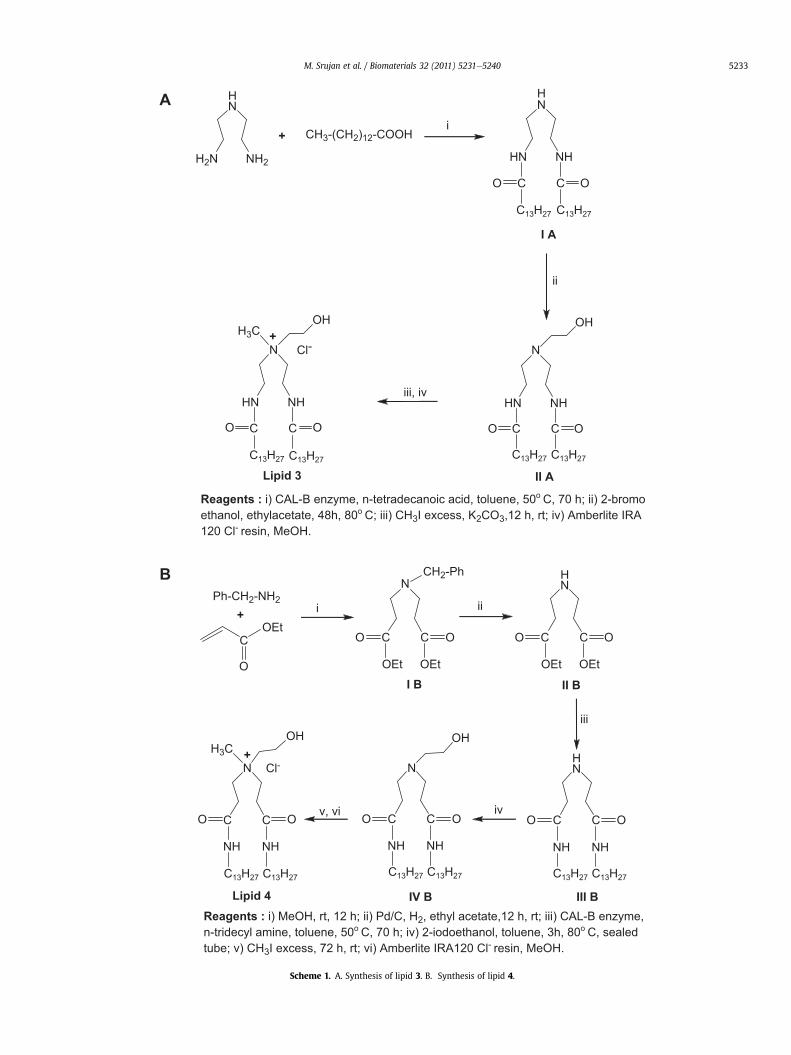
Reagents : i) CAL-B enzyme, n-tetradecanoic acid, toluene, 50o C, 70 h; ii) 2-bromoethanol, ethylacetate, 48h, 80o C; iii) CH3I excess, K2CO3,12 h, rt; iv) Amberlite IRA120 Cl- resin, MeOH.
HN
H2N NH2
HN
HN NH
C C
C13H27C13H27
OO
+ CH3-(CH2)12-COOH
N
HN NH
C C OO
OH
N
HN NH
C C OO
OHH3C
i
ii
iii, iv
+
Cl-
Lipid 3 II A
I A
A
B
Reagents : i) MeOH, rt, 12 h; ii) Pd/C, H2, ethyl acetate,12 h, rt; iii) CAL-B enzyme,n-tridecyl amine, toluene, 50o C, 70 h; iv) 2-iodoethanol, toluene, 3h, 80o C, sealedtube; v) CH3I excess, 72 h, rt; vi) Amberlite IRA120 Cl- resin, MeOH.
Ph-CH2-NH2
C
O
OEt
N
C CO O
NH NH
N
C CO O
OEt OEt
CH2-Ph HN
C CO O
OEt OEt
N
C CO O
NH NH
OHOHH3C
Cl-
I B II B
IV BLipid 4
HN
C CO O
NH NH
III B
i ii
iii
ivv, vi
+
+
C13H27C13H27C13H27C13H27
C13H27C13H27C13H27C13H27C13H27C13H27
Scheme 1. A. Synthesis of lipid 3. B. Synthesis of lipid 4.
M. Srujan et al. / Biomaterials 32 (2011) 5231e5240 5233

M. Srujan et al. / Biomaterials 32 (2011) 5231e52405234
Step iii. Synthesis of (2-hydroxyethyl)(methyl)di[2-tetradecanoylamino) ethyl]ammonium chloride (Lipid 3, Scheme 1A)
Intermediate IIA (80 mg, 0.14 mmol) prepared in step ii above was dissolvedin 1 mL of DCM in a 25 mL round-bottom flask, and 1 mL of methyl iodide wasadded to the solution. Catalytic amount of K2CO3 was added and the reactionmixture was stirred at room temperature for 12 h and concentrated. The crudecompound obtained was filtered and purified by column chromatography using60e120 mesh silica gel in chloroform/methanol 95:5 (v/v). The pure compoundupon chloride ion exchange on Amberlite-IRA 120 resin (using methanol aseluent) afforded pure lipid 3 as a white solid (53 mg, 63% yield, Rf ¼ 0.2 in 5%methanol in chloroform).
1H NMR of (300 MHz, CDCl3): d/ppm ¼ 0.9 [t, 6H, CH3eCH2eC11H23e]; 1.2e1.4[m, 40H, CH3e(CH2)10e]; 1.6 [m, 4H, CH3e(CH2)10eCH2eCH2]; 2.2[t, 4H,CH3e(CH2)10eCH2eCH2]; 3.9 [s, 3H, CH3eNþe(CH2eCH2e)3]; 3.4[t, 4H,NeCH2eCH2eNHeCOe]; 3.5 [t, 4H, NeCH2eCH2eNHeCOe]; 3.9 [t, 2H,NeCH2eCH2eOH]; 4.1 [t, 2H, NeCH2eCH2eOH].
ESI-MS m/z: calcd 582.971(for C35H72N3O3), 584 found [M þ H]þ.Elemental analysis data (C, H, N):Calcd for C35H72ClN3O3: C, 67.98; H, 11.73; N, 6.79.Found: C, 68.09; H, 11.68; N, 6.71.
2.2.2. Synthesis of (2-hydroxyethyl) (methyl)di[3-oxo-3-(tridecylamino) propyl]ammonium chloride (Lipid 4, Scheme 1B)
Step i. Synthesis of ethyl 3-[benzyl (3-ethoxy-3-oxopropyl) amino] propanoate(Intermediate IB, Scheme 1B)
A solution of benzylamine (1 g, 9.34mmol) in 25mL of drymethanolwas taken ina 50mL single-neck round-bottom flask and ethyl acrylate (1.84 mL, 23.3 mmol) wasadded to the stirring solution. The reactionmixturewas stirred at room temperaturefor 12 h. Solvent methanol and unreacted ethyl acrylate were removed on a rotaryevaporator by repeated chasingwith dryDCM. The reactionmixturewas dilutedwith100 mL of CHCl3, and washed with water (2 � 50 mL). The organic layer was sepa-rated, dried over anhydrous sodium sulfate, and concentrated by rotary evaporator.The residue, upon short columnchromatographicpurificationover 60e120mesh sizesilica gel column using 5% ethyl acetate/hexane (v/v) as eluent, afforded the pure titlecompound IB as a yellow liquid (2.03 g, 80.3% yield, Rf ¼ 0.5 in 20% ethyl acetate:hexane).
1H NMR of (300 MHz, CDCl3): d/ppm ¼ 1.3 [t, 6H, CH3eCH2eO]; 2.4[t, 4H, eNeCH2eCH2eCOeOeC2H5]; 2.7 [t, 4H, NeCH2eCH2eCOeOeC2H5]; 3.6[s, 2H, NeCH2eC6H5]; 4.1[q, 4H, eCOeOeCH2eCH3]; 7.3[m, 2H, NeCH2eC6H5]; 7.2[m, 3H, NeCH2eC6H5].
ESI-MS m/z: calcd 307.387 (for C17H25NO4), found 308 [M þ H]þ.
Step ii. Synthesis of ethyl 3-[(3-ethoxy-3-oxopropyl)amino] propanoate (Inter-mediate IIB, Scheme 1B)
In a 25 mL round bottomed flask, intermediate IB (0.6 g, 1.9 mmol), preparedabove in step i dissolved in ethyl acetate (10mL) was added to the 10% Pd/C (200mg)suspended in ethyl acetate (5mL) and the reactionmixturewas stirred at room tempfor 12 h under H2 (2 atm). The mixture was filtered using Sartorius filter paper. Thefiltratewas dried over anhydrous Na2SO4 and concentrated to dry ness to afford purecompound II as a colorless gummy liquid (0.33 g, 78.5% yield, Rf ¼ 0.6, 5% methanolin chloroform). The intermediate IIB obtained was used directly in next reactionwithout further purification.
1H NMR of (300 MHz, CDCl3): d/ppm ¼ 1.2 [t, 6H, CH3eCH2eO]; 2.4[t, 4H, eNeCH2eCH2eCOeOeC2H5]; 2.7 [t, 4H, NeCH2eCH2eCOeOeC2H5]; 4.1[q, 4H, eCOeOeCH2eCH3].
ESI-MS m/z: calcd 217.263 (for C10H19NO4), found 218 [M þ H]þ.
Step iii. Synthesis of N1-tridecyl-3-[3-oxo-3-(tridecylamino) propyl] aminopropanamide (Intermediate IIIB, Scheme 1B)
In a 100 mL single-neck round-bottom flask, intermediate IIB (0.1 g, 0.46 mmol)prepared above in step ii and tridecylamine (0.202 g,1.1mmol) were dissolved in drytoluene. CAL-B enzyme (10% w/w) was added and the reaction mixture was stirredat 50 �C for 70 h. The reaction mixture was filtered off to remove the enzyme. To thefiltrate CAL-B enzyme (10% w/w) was added again and reaction continued at 50 �Cfor further 48 h. The reaction mixture was filtered and concentrated. The residueupon column chromatographic purification (using 60e120 mesh size silica gel and5:95 methanol:chloroform as eluent) afforded intermediate IIIB as a white solid(0.204 g, 84.6% yield, Rf ¼ 0.2, 5:95 methanol:chloroform).
1H NMR of (300 MHz, CDCl3): d/ppm ¼ 0.9 [t, 6H, CH3eCH2eC11H23e]; 1.2e1.4[m, 40H, CH3e(CH2)10e]; 1.5 [m, 4H, CH3e(CH2)10eCH2eCH2eNH]; 2.0 [t, 4H,NeCH2eCH2eCOeNH]; 2.7 [t, 4H, NeCH2eCH2eCOeNHe]; 3.1 [t, 4H,CH3e(CH2)10eCH2eCH2eNH].
ESI-MS m/z: calcd 523.884 (for C32H65N3O2), found 525 [M þ H]þ.
Step iv. Synthesis of N1-tridecyl-3-(2-hydroxyethyl)[3-oxo-3-tridecylamino)propyl] amino propanamide (Intermediate IVB, Scheme 1B)
To intermediate IIIB (0.22 g, 0.38 mmol, prepared above in step iii) in sealedtube, 2-Iodo ethanol (0.12 g, 0.76 mmol) was added in dry toluene. The reactionmixture was stirred at 80 �C for 3 h. Solvent toluene was removed by chasing withmethanol on rotary evaporator. The residue upon column chromatographic puri-fication (using 60e120 mesh size silica gel and 5:95 methanol:chloroform, v/v, aseluent) afforded intermediate IVB as a white solid (0.11 g, 75% yield, Rf ¼ 0.2, 5:95methanol:chloroform).
1H NMR of (300 MHz, CDCl3) : d/ppm ¼ 0.9 [t, 6H, CH3eCH2eC11H23e]; 1.2e1.4[m, 40H, CH3e(CH2)10e]; 1.5 [m, 4H, CH3e(CH2)10eCH2eCH2eNH]; 2.6 [t,4H, NeCH2eCH2eCOeNH]; 2.8 [t, 4H, NeCH2eCH2eCOeNHe]; 2.9 [t, 2H,NeCH2eCH2eOH]; 3.1 [t, 4H, CH3e(CH2)10eCH2eCH2eNH]; 3.8 [t, 2H,NeCH2eCH2eOH].
ESI-MS m/z: calcd 567.936(for C34H69N3O3), found 568 [M]þ, 569 [M þ H]þ, 570[M þ 2H]þ.
Step v. Synthesis of (2-hydroxyethyl) (methyl)di[3-oxo-3-(tridecylamino)propyl] ammonium chloride (Lipid 4, Scheme 1B)
Intermediate IVB (80 mg, 0.14 mmol) prepared in step iv above was dissolved in1mL of DCM in a 25mL round-bottom flask, and 1mL of methyl iodide was added tothe solution. The reaction mixture was stirred at room temperature for 36 h andconcentrated. The crude compound obtained was purified by column chromatog-raphy using 60e120 mesh silica gel in chloroform/methanol 95:5 (v/v). The purecompound upon chloride ion exchange on Amberlite-IRA 120 resin (using methanolas eluent) afforded pure lipid 4 as a white solid (48 mg, 58.8% yield, Rf ¼ 0.2 in 5%methanol in chloroform).
1H NMR of (300 MHz, CDCl3): d/ppm ¼ 0.9 [t, 6H, CH3eCH2eC11H23e]; 1.2e1.4[m, 40H, CH3e(CH2)10e]; 1.5 [m, 4H, CH3e(CH2)10eCH2eCH2eNH]; 2.8 [t, 4H,NeCH2eCH2eCOeNH]; 3.1 [t, 4H, CH3e(CH2)10eCH2eCH2eNH]; 3.3 [s,3H, CH3eNþe(CH2eCH2e)3]; 3.9 [t, 2H, þNeCH2eCH2eOH]; 4.2 [t, 4H, þNeCH2eCH2eCOeNHe]; 4.3 [t, 2H, þNeCH2eCH2eOH].
ESI-MS m/z: calcd 582.971(for C35H72N3O3), found 583 [M]þ, 584 [M þ H]þ, 585[Mþ 2H]þ.
Elemental analysis data (C, H, N):Calcd for C35H72ClN3O3: C, 67.98; H, 11.73; N, 6.79.Found: C, 67.85; H, 11.79; N, 6.86.
2.3. Transfection assay
Cells were seeded at a density of 10,000 cells (for B16F10) and 15,000 cells (forA-549, CHO, HepG2) per well in a 96-well plate 12e18 h before the transfection.0.3 mg of plasmid DNA was complexed with varying amounts of lipids(0.45e7.2 nmol) in plain DMEM/MEMmedium (total volumemade up to 100 mL) for30 min. The lipid:DNA (�) charge ratios were from 8:1 to 0.5:1 over these ranges ofthe lipids. The complexes were then added to the cells. For serum compatibilitystudy with 2:1 lipid:DNA charge ratios, the final lipid and DNA concentrations usedin each well were 18.2 and 9.1 mM, respectively. For preparing lipoplexes in mediacontaining increasing amounts of added serum (10e90%), the volume of the stocklipoplex used was 4.8 mL in each well and the volumes were made up to 100 mL byaddition of necessary volumes of DMEM and FBS so that the final serum concen-trations were in the range 10e90% (for instance, for 10% added serum, 10 mL of FBSand 85.2 mL of DMEM were used). After 3 h of incubation, DMEM/MEM wasremoved, 10% complete medium was added to the cells. The reporter gene activitywas estimated between 36 and 48 h. The cells were washed with PBS (2 � 100 mL)and lysed with 50 mL lysis buffer [0.25 M TriseHCl pH 8.0, 0.5% NP40]. Care was takento ensure complete lysis. The b-galactosidase activity per well was estimated byadding 50 mL of 2�-substrate solution [1.33 mg/mL of ONPG, 0.2 M sodium phos-phate (pH 7.3) and 2 mM magnesium chloride] to the lysate in a 96-well plate.Absorption at 405 nm was converted to b-galactosidase units using a calibrationcurve constructed with pure commercial b-galactosidase enzyme. The values ofb-galactosidase units in triplicate experiments assayed on the same day varied byless than 10%. The transfection experiment was carried in duplicate and the trans-fection efficiency values shown in Fig. 2 (AeD) are the average of triplicate experi-ments performed on the same day. The day to day variation in average transfectionefficiency was found to be within 2-fold. The transfection profiles obtained ondifferent days were identical.
2.4. Cytotoxicity assay
The cytotoxicities of the lipids 3 & 4 were evaluated in representative CHO(Chinese hamster ovary) cells across the lipid:DNA charge ratios of 8:1e0.5:1 usingMTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) based reduc-tion assay as described earlier [18]. The cytotoxicity assay was performed in 96-wellplates bymaintaining the same ratio of number of cells to amount of cationic lipid, asused in the previously described transfection experiments. Briefly, 4 h after the
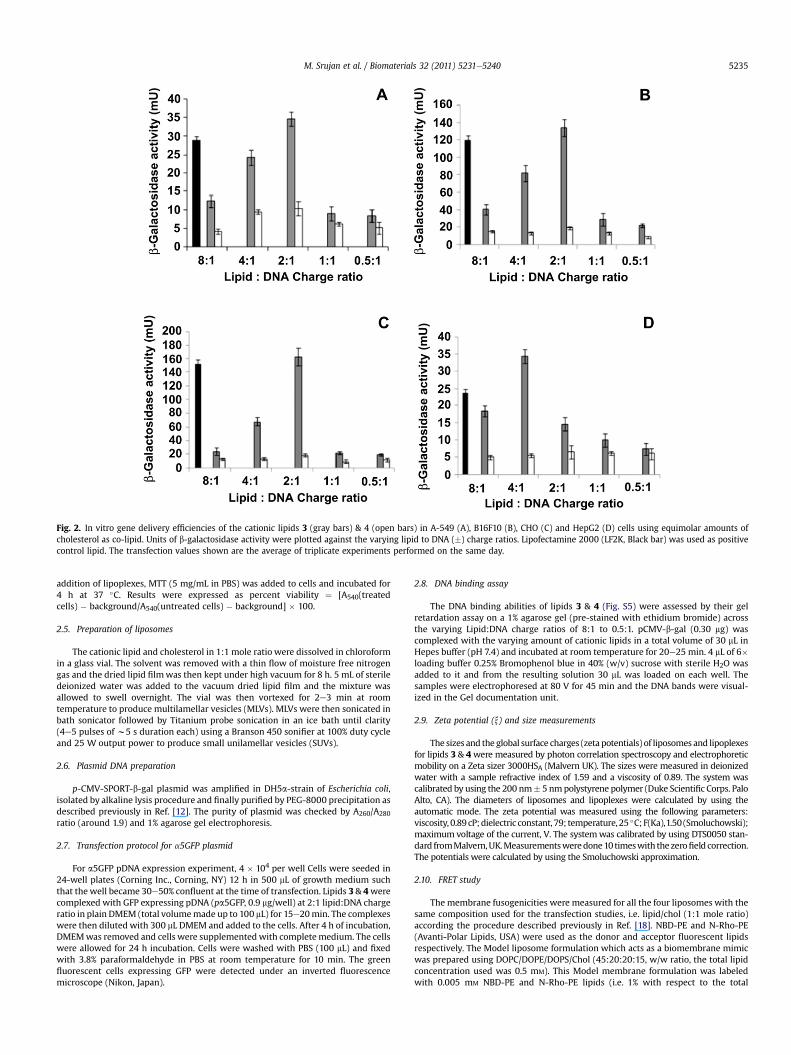
Fig. 2. In vitro gene delivery efficiencies of the cationic lipids 3 (gray bars) & 4 (open bars) in A-549 (A), B16F10 (B), CHO (C) and HepG2 (D) cells using equimolar amounts ofcholesterol as co-lipid. Units of b-galactosidase activity were plotted against the varying lipid to DNA (�) charge ratios. Lipofectamine 2000 (LF2K, Black bar) was used as positivecontrol lipid. The transfection values shown are the average of triplicate experiments performed on the same day.
M. Srujan et al. / Biomaterials 32 (2011) 5231e5240 5235
addition of lipoplexes, MTT (5 mg/mL in PBS) was added to cells and incubated for4 h at 37 �C. Results were expressed as percent viability ¼ [A540(treatedcells) � background/A540(untreated cells) � background] � 100.
2.5. Preparation of liposomes
The cationic lipid and cholesterol in 1:1 mole ratio were dissolved in chloroformin a glass vial. The solvent was removed with a thin flow of moisture free nitrogengas and the dried lipid filmwas then kept under high vacuum for 8 h. 5 mL of steriledeionized water was added to the vacuum dried lipid film and the mixture wasallowed to swell overnight. The vial was then vortexed for 2e3 min at roomtemperature to produce multilamellar vesicles (MLVs). MLVs were then sonicated inbath sonicator followed by Titanium probe sonication in an ice bath until clarity(4e5 pulses of w5 s duration each) using a Branson 450 sonifier at 100% duty cycleand 25 W output power to produce small unilamellar vesicles (SUVs).
2.6. Plasmid DNA preparation
p-CMV-SPORT-b-gal plasmid was amplified in DH5a-strain of Escherichia coli,isolated by alkaline lysis procedure and finally purified by PEG-8000 precipitation asdescribed previously in Ref. [12]. The purity of plasmid was checked by A260/A280
ratio (around 1.9) and 1% agarose gel electrophoresis.
2.7. Transfection protocol for a5GFP plasmid
For a5GFP pDNA expression experiment, 4 � 104 per well Cells were seeded in24-well plates (Corning Inc., Corning, NY) 12 h in 500 mL of growth medium suchthat the well became 30e50% confluent at the time of transfection. Lipids 3 & 4werecomplexed with GFP expressing pDNA (pa5GFP, 0.9 mg/well) at 2:1 lipid:DNA chargeratio in plain DMEM (total volumemade up to 100 mL) for 15e20min. The complexeswere then diluted with 300 mL DMEM and added to the cells. After 4 h of incubation,DMEMwas removed and cells were supplementedwith completemedium. The cellswere allowed for 24 h incubation. Cells were washed with PBS (100 mL) and fixedwith 3.8% paraformaldehyde in PBS at room temperature for 10 min. The greenfluorescent cells expressing GFP were detected under an inverted fluorescencemicroscope (Nikon, Japan).
2.8. DNA binding assay
The DNA binding abilities of lipids 3 & 4 (Fig. S5) were assessed by their gelretardation assay on a 1% agarose gel (pre-stained with ethidium bromide) acrossthe varying Lipid:DNA charge ratios of 8:1 to 0.5:1. pCMV-b-gal (0.30 mg) wascomplexed with the varying amount of cationic lipids in a total volume of 30 mL inHepes buffer (pH 7.4) and incubated at room temperature for 20e25 min. 4 mL of 6�loading buffer 0.25% Bromophenol blue in 40% (w/v) sucrose with sterile H2O wasadded to it and from the resulting solution 30 mL was loaded on each well. Thesamples were electrophoresed at 80 V for 45 min and the DNA bands were visual-ized in the Gel documentation unit.
2.9. Zeta potential (x) and size measurements
The sizes and the global surface charges (zeta potentials) of liposomes and lipoplexesfor lipids 3 & 4were measured by photon correlation spectroscopy and electrophoreticmobility on a Zeta sizer 3000HSA (Malvern UK). The sizes were measured in deionizedwater with a sample refractive index of 1.59 and a viscosity of 0.89. The system wascalibrated by using the 200 nm� 5 nmpolystyrene polymer (Duke Scientific Corps. PaloAlto, CA). The diameters of liposomes and lipoplexes were calculated by using theautomatic mode. The zeta potential was measured using the following parameters:viscosity, 0.89cP;dielectric constant, 79; temperature,25 �C;F(Ka),1.50 (Smoluchowski);maximum voltage of the current, V. The systemwas calibrated by using DTS0050 stan-dard fromMalvern,UK.Measurementsweredone10 timeswith thezerofield correction.The potentials were calculated by using the Smoluchowski approximation.
2.10. FRET study
The membrane fusogenicities were measured for all the four liposomes with thesame composition used for the transfection studies, i.e. lipid/chol (1:1 mole ratio)according the procedure described previously in Ref. [18]. NBD-PE and N-Rho-PE(Avanti-Polar Lipids, USA) were used as the donor and acceptor fluorescent lipidsrespectively. The Model liposome formulation which acts as a biomembrane mimicwas prepared using DOPC/DOPE/DOPS/Chol (45:20:20:15, w/w ratio, the total lipidconcentration used was 0.5 mM). This Model membrane formulation was labeledwith 0.005 mM NBD-PE and N-Rho-PE lipids (i.e. 1% with respect to the total

M. Srujan et al. / Biomaterials 32 (2011) 5231e52405236
biomembrane mimicking formulation content). The total lipid concentrations usedfor all the 2 liposomes were same as that of the biomembrane mimicking lipidformulation (0.5 mM). Labeled model biomembrane liposomal formulations wereplaced in a FLX 800 Microplate Fluorescence Reader (BioTek Instruments Inc. UK) atroom temperature and equimolar amounts of liposomes of the lipids 3 & 4 withChol/lipid (1:1 mole ratio) were added. Fluorescence intensities were recorded asa function of time with excitation at 485 nm and emission at 595 nm. Fusion (100%)was determined from the Rho-PE fluorescence intensity of the labeled bio-membrane liposomal formulation in presence of 1% Triton X-100.
2.11. Dye entrapment experiment
The protocols followed for liposomal entrapment of 5/6-CF and for monitoring itssubsequent release fromthe liposomeswere asdescribedpreviously inRef. [18]. Briefly,equimolar cholesterol containing liposomal suspensions of lipids 3& 4 (1mM total lipidconcentration; 0.5mM in both cationic lipid and cholesterol)were prepared inTriseHClbuffer (25 mM, pH 8.0) containing 25 mM CF. The CF-loaded liposomes were separatedfrom the unentrapped free CF dye by passing through a sephadex G-50 columnwith anelution buffer of 50 mM TriseHCl buffer (pH 8.0). Transmembrane permeation of theliposomally loaded CFwasmeasured at 25 �C by following the changes in fluorescenceintensityof theemitted light (due to leakageofCF fromliposomes) in30min ina inaFLX800 Microplate Fluorescence Reader (BioTek Instruments Inc. UK). The excitation andemission wavelengths used for this experiment were 485 and 528 nm, respectively.Percentage CF releases were calculated using the equation: Permeation of CF(%)¼ [(Ft� Fi)/(Ff� Fi)]� 100where Fi, Ft and Ff are the fluorescence emission values attime t¼ 0, at t¼ 30min and thefinal fluorescence on complete disruption of liposomalstructures with the final concentration of 1% (v/v) Triton X-100.
2.12. Cellular uptake studies by epifluorescence microscopy
For fluorescence microscopy experiments, 10,000 cells were seeded in each well ofa96-wellplate (Corning Inc.,Corning,NY)12h in500mLofgrowthmediumsuchthat thewell became30e50% confluent at the time of transfection. Rhodamine-PE labeled lipids3 & 4 were complexed with pCMV-SPORT-b-gal (0.3 mg/well) at 2:1 lipid:DNA chargeratio in a total volume of 100 mL DMEM for 15e20min. The complexeswere then addedto the cells. After 4e6hof incubation, cellswerewashedwithPBS (2�100 mL) andfixedwith 3.8%paraformaldehyde in PBS at roomtemperature for 10min. The redfluorescentcells were detected under an inverted fluorescence microscope (Nikon, Japan).
2.13. AFM analysis of liposomes
Atomic forcemicroscopy (AFM) was employed to analyze themorphology of theliposomes. Briefly, glass piece (0.5 � 0.5 mm) is placed on to one side of a doublestick tape and on to the other side of the tape a mica sheet is attached. The micasheet with a cellophane tape stuck on it is pulled out to peel it and removal of layerof mica sheet was repeated till several times to obtain a smooth surface of micasheet. A 10 mL of the 0.5 mM liposome sample prepared using equimolar ratio ofindividual lipids and cholesterol was uniformly dispersed on to freshly cleaved micaand the mica surface was then dried with a thin stream of nitrogen followed byaerial drying in a dust free zone for 12 h. The AFM images were observed in thetapping mode in air using Digital Nanoscope IV (Veeco Instruments, Santa Barbara,CA). The imaging was carried out with the vibration damped microscope.Commercial phosphorous (n) doped silica tips on a I-tape cantilever with a length of115e135 mm and resonance frequency of about 260 kHz were used.
2.14. In vivo studies
All the in vivo transfection experiments were performed in accordance with theInstitutional Bio-Safety and Ethical Committee Guidelines using an approved animalprotocol. Each of the 6 weeks-old male Balb/C mice (w20 g) was injected in the tailvein with 300 mL of the liposome:DNA complexes in 5% w/v glucose solution usinga 26.5 gauge syringe needle. Micewere sacrificed 8 h post-injection and organs wereharvested and washed in cold saline. 500 mL of lysis buffer (0.1 M TriseHCl, 2 mM
ETDA and 0.2% Triton X-100 pH 7.8) was added to each organ and homogenizedusing a mechanical homogenizer. The homogenates were centrifuged at 14,000 rpmfor 10 min at 4 �C, 5 mL supernatant was assayed using the Promega Luciferase assaykit (Madson,Wisconsin, USA) in an FLx 800microplate Luminescence Reader (BioTekInstruments Inc. UK). Protein concentration of each tissue extract was determinedby the modified Lowry procedure [19] and the Luciferase activity in each organ wasexpressed as the relative light unit (RLU) per mg of extracted protein.
3. Results and discussion
3.1. Chemistry
The key intermediates namely, IA for lipid 3 and IIIB for lipid 4(Scheme 1) were synthesized by employing an enzymatic reaction.
The intermediate IA was synthesized by coupling myristic acidwith Bis(2-aminoethyl)amine (diethylenetriamine) in presenceof 10% CAL-B enzyme (w/w). Intermediate IA upon reaction with2-bromoethanol provided the tertiaryamine intermediate IIAwhichupon quarternization with methyl iodide followed by chloride ionexchange chromatography over Amberlite-IRA 400 Cl� resin affor-ded pure lipid 3 (Scheme 1, Part A). To synthesize lipid 4, first benzylamine was reacted with ethyl acrylate under Michael additioncondition toprovide the tertiaryamine intermediate IB (Scheme1B).The tertiary amine intermediate IB, upon benzyl deprotection withH2/Pd followed by reactionwith n-tridecylamine in presence of 10%CAL-B enzyme (w/w) afforded the crucial di-amide intermediate IIIB(Scheme 1B). The di-amide intermediate IIIB upon reaction with2-iodo ethanol in a sealed tube afforded the tertiary amine inter-mediate IVB (Scheme 1B). Intermediate IVB upon quarternizationwith excess methyl iodide followed by chloride ion exchange overAmberlite-IRA-120 resin afforded the pure target lipid 4 (Scheme1B). The structures of all the intermediates shown in Scheme 1were confirmed by 1H NMR spectral analysis and the structures ofthe final target lipids 3 & 4 were confirmed by NMR, Mass and IRspectral analysis. The purity of the final lipids were confirmed byelemental analysis (C, H, N) as well as by analytical HPLC analysisin two different mobile phases (100% methanol and 95:5 meth-anol:water, v/v).
3.2. In vitro transfection biology
First wemeasured the in vitro gene delivery efficacies of lipids 3and 4 in four cultured mammalian cells including A-549 (humanlung carcinoma cells), B16F10 (murine melanoma cells), CHO(Chinese hamster ovary cells) and HepG2 (Human hep-atocarcinoma) using p-CMV-SPORT-b-gal plasmid DNA as thereporter gene encoding the enzyme b-galactosidase across thelipid:DNA charge ratio (�) range 8:1e0.5:1. Despite the oppositeorientation of the linker (amide) functionality being the solestructural differences between lipids 3 & 4, only lipid 3 wascompetent in delivering genes into all the four cells particularly athigher charge ratios above 2:1 and lipid 4 was found to be essen-tially transfection incompetent (Fig. 2, Parts AeD). Importantly, thein vitro gene delivery efficiencies of lipid 3 were found to becomparable or better than that for the widely used commerciallyavailable Lipofectamine 2000 (Fig. 2, Parts AeD). Equimolaramounts of cholesterol (with respect to cationic lipids) were usedas the co-lipid. Similarly contrasting transfection properties forlipids 3 & 4were observed when the plasmid DNA encoded a greenfluorescence protein. A number of fluorescently labeled cells werevisiblewhen the cells were treatedwith the lipoplex of lipid 3whilehardly any fluorescently labeled cells could be detected when cellswere treated with lipoplex of lipid 4 (Fig. S2, SupportingInformation). Thus, consistent with our previous findings [18], thefindings summarized in Fig. 2 and Fig. S2 demonstrated thedramatic influence of linker orientation in liposomal gene deliveryunder in vitro conditions. Toward probing whether any inherentinstability of the liposomes of lipid 4 was at the origin of itsessentially transfection incompetent nature, HPLC analysis wasconducted for both the liposomes of lipids 3 & 4 on day 1 and day 3after the liposome preparation. As was revealed in the HPLCchromatograms, both the liposomes were equally stable (Fig. S3,Supporting Information). Clearly, inherent chemical stabilitydifferences of the liposomes of lipids 3 & 4were ruled out as one ofthe causative factors behind the remarkably higher efficiencies oflipid 3 (with respect to lipid 4) in delivering genes into culturedcells. MTT-based cell viability assays in representative CHO cellsrevealed lipoplexes of lipids 3 & 4 to be equally non-cytotoxic.Percent cell viabilities were found to be remarkably high (80e100%)

Table 1Hydrodynamic Diameters (A) Zeta Potentials (B, mV) of the 1 mM plain liposomesprepared using equimolar ratio of Chol/Lipids 3 & 4 in DI water.
A. Hydrodynamic diametersa
Liposomes of Size on Day 1 (nm) Size on Day 3 (nm)Lipid 3 148 � 1 143 � 1Lipid 4 213 � 4 214 � 7
B. Zeta potentialsa
Liposomes of Zeta potentials (mV)Lipid 3 10.7 � 5.4Lipid 4 2.8 � 1.3
a The sizes and the surface potentials of the liposomes prepared in Deionizedwater were measured by the laser light scattering technique using Zeta sizer 3000A(Malvern Instruments, U.K.) as described in the text. Values shown are the averagesobtained from three (sizes) and ten (zeta potentials) measurements.
M. Srujan et al. / Biomaterials 32 (2011) 5231e5240 5237
when cells were treated with the lipoplexes of both lipids 3 & 4within the lipid:DNA charge ratio 4:1e0.5:1 and somewhat low(50e70%) for cells treated with lipoplexes containing lipid:DNAcharge ratio of 8:1 (Fig. S4, Supporting Information). Thus, the poorin vitro transfection efficiency of lipid 4 is unlikely to originate fromthe relatively higher cytotoxicities of lipid 4 lipoplexes than that oflipid 3 lipoplexes.
3.3. Physico-chemical characterizations
The average hydrodynamic diameters of the freshly preparedliposomes of transfection efficient lipid 3 (measured by dynamiclaser light scattering technique) were found to be significantlylower than those for the transfection incompetent liposomes oflipid 4 (148 � 1 nm vs 214 � 4 nm, respectively; Table 1, Part A).Toward checking the stability of these liposomal systems, liposomesizes were also measured on day 3 after preparation. As shown inTable 1, Part A, the liposomal sizes remained essentially unchangedthereby indicating that both the liposomes of lipids 3 & 4 are stable.Similarly, lipoplex sizes for lipid 3 were found to be lower thanthose for the lipoplexes of lipid 4 across the entire lipid:DNA chargeratios 8:1e2:1 (Table 2, Part A). While the lipoplex sizes for bothlipids 3 & 4 decreased with increasing lipid contents in the lip-oplexes, the extent of size decrease was more pronounced for thelipoplexes of lipid 3 than that for the lipoplexes of lipid 4 (Table 2,Part A). This finding is consistent with better DNA condensingefficacies of the liposomes of lipid 3 compared to that for theliposomes of lipid 4. The gel patterns observed in gel retardationassay were also consistent with higher DNA condensation effi-ciencies of the liposomes of lipid 3 compared to the liposomes oflipid 4. While lipid 3 was able to completely inhibit the electro-phoretic movement of DNA from its lipoplexes even at the lowestlipid:DNA charge ratios of 1:1 and 0.5:1, significant bands for freeDNA were found to be present for the lipoplexes of lipid 4 at theselowest lipid:DNA charge ratios (Fig. S6, Supporting Information). It
Table 2Hydrodynamic Diameters (A) and Zeta Potentials (B, mV) of the lipoplexes across lipid:D
Lipoplexes of Lipid:DNA8:1
Lipid:DNA4:1
A. Hydrodynamic diameters (nm)a
Lipid 3 146 � 7 165 � 14Lipid 4 351 � 8 395 � 18B. Zeta potentials (mV)a
Lipid 3 21.4 � 6.3 13.5 � 3.8Lipid 4 9.7 � 2.4 6.3 � 2.2
a The sizes and the surface potentials of the lipoplexes prepared in plain DMEM wereInstruments, U.K.) as described in the text. Values shown are the averages obtained from
is worth mentioning at this point of discussion that these averagehydrodynamic diameters are calculated from the cumulant (mon-omodal) analysis software provided with the light scatteringinstrument (Zeta sizer 3000HSA, Malvern, UK) and the repeatmeasurements are usually within one or two percent of oneanother. The software also provides the actual liposomes sizedistributions. The actual size distribution profiles revealed exis-tence of liposomes with varying hydrodynamic diameters acrossthe range 50e500 nm for both the liposomes of lipids 3 & 4 withthe number of large liposomes (w500 nm) being slightly higher forlipid 4 (Fig. S5, Supporting Information). Intensities of scatteredlight for spherical particles with radius r is proportional to r6 andthus scattering by large particles, even if their population is small inthe size distribution profiles, contributes heavily to the hydrody-namic diameter obtained by the dynamic light scattering methodwhereas the scattered intensity from small particles gets lost inthe background signal [20]. Thus, the observed slightly higherpopulation of large size (w500 nm) liposomes of lipid 4 (Fig. S5,Supporting Information) might be the reason behind their largeraverage hydrodynamic diameters observed in the light scatteringstudy. To this end, AFM images are more informative in the sensethat they actually show the presence of particles with varying sizes.Toward confirming the lower sizes of liposomes of lipid 3, AFMimages were taken on mica sheet. AFM images of liposomesprovided further evidence for the smaller sizes of the liposomes oflipid 3 than that for the liposomes of lipid 4 (Fig. 3). However, thehydrodynamic diameters of the liposomes of both lipids 3 & 4measured by light scattering method (Table 1, Part A) were found tobe bigger when compared to their sizes observed in AFM images(Fig. 3). This is not unusual. AFM scans cannot always be used toaccurately calculate the ratio of nanoparticle sizes present asdeposition methods may skew the size distribution [20].
The global surface potential of liposomes of lipid 3was found tobe higher than that for the liposomes of lipid 4 (10.7 � 5.4 mV vs2.8 � 1.3 mV, respectively, Table 1, Part B). Previously it has beenreported that hydration of ester carbonyl groups (throughhydrogen bonding with water) present near the polar head-groupregion of amphiphilic molecules contributes to the head-grouphydration of liposomal aggregates [21]. Since the covalent bonddistance of the amide carbonyl group from the positively chargednitrogen in lipid 4 is less than that in lipid 3, it may not be unlikelythat the degree of interfacial hydration in lipid 4 liposomes ishigher than that in liposomes of lipid 3. Such enhanced interfacialhydration in the liposomes of lipid 4 may play an important role insignificantly shielding the surface positive charge in the liposomesof lipid 4 thereby reducing the global positive charge on the lipo-some surface compared to that in liposomes of lipid 3 (Table 1,Part B). The larger sizes of the liposomes of lipid 4 compared tothose for lipid 3 (Table 1, Part A) may also be related to the liposomeswelling due to such possible enhanced interfacial hydration. Theglobal surface potentials of lipid 3 lipoplexes were also found to bemore positive than those for lipid 4 lipoplexes in the lipid:DNA
NA charge ratios 8:1e0.5:1 in presence of plain DMEM.
Lipid:DNA2:1
Lipid:DNA1:1
Lipid:DNA0.5:1
266 � 8 568 � 8 689 � 6496 � 26 602 � 17 722 � 28
4.7 � 1.3 �1.8 � 1.1 �10.4 � 4.42.8 � 0.7 �2.8 � 1.2 �12.36 � 8.4
measured by the laser light scattering technique using Zeta sizer 3000A (Malvernthree (sizes) and ten (zeta potentials) measurements.
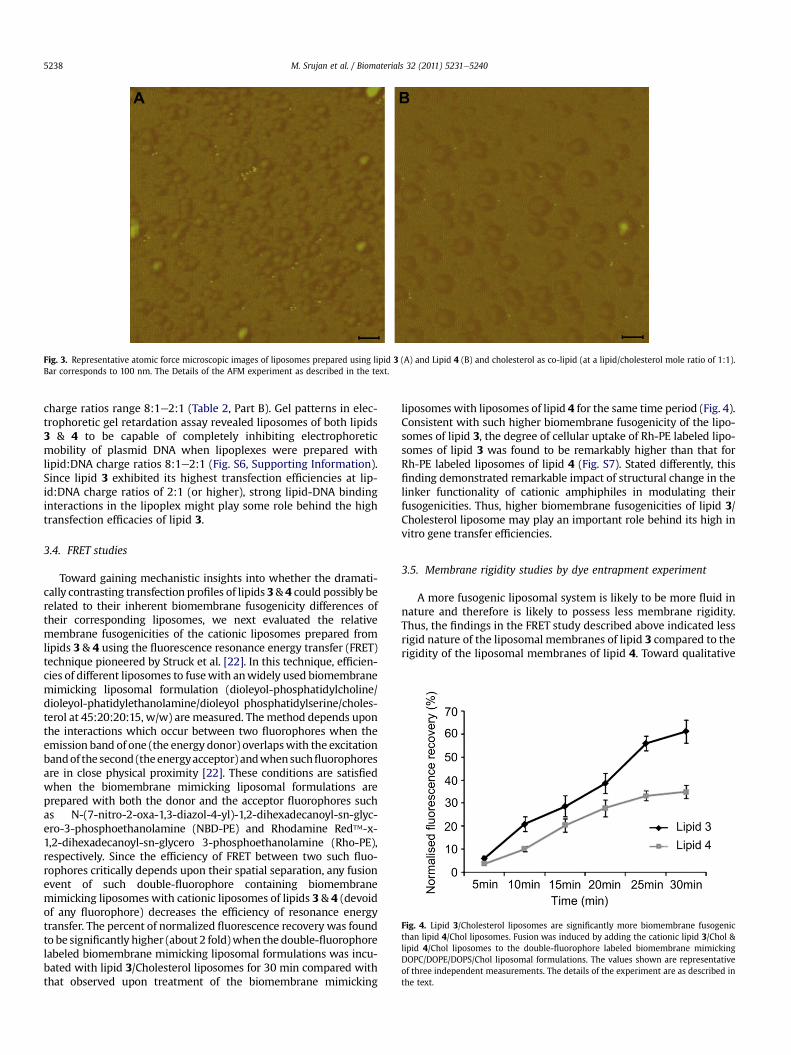
Fig. 3. Representative atomic force microscopic images of liposomes prepared using lipid 3 (A) and Lipid 4 (B) and cholesterol as co-lipid (at a lipid/cholesterol mole ratio of 1:1).Bar corresponds to 100 nm. The Details of the AFM experiment as described in the text.
Fig. 4. Lipid 3/Cholesterol liposomes are significantly more biomembrane fusogenicthan lipid 4/Chol liposomes. Fusion was induced by adding the cationic lipid 3/Chol &lipid 4/Chol liposomes to the double-fluorophore labeled biomembrane mimickingDOPC/DOPE/DOPS/Chol liposomal formulations. The values shown are representativeof three independent measurements. The details of the experiment are as described inthe text.
M. Srujan et al. / Biomaterials 32 (2011) 5231e52405238
charge ratios range 8:1e2:1 (Table 2, Part B). Gel patterns in elec-trophoretic gel retardation assay revealed liposomes of both lipids3 & 4 to be capable of completely inhibiting electrophoreticmobility of plasmid DNA when lipoplexes were prepared withlipid:DNA charge ratios 8:1e2:1 (Fig. S6, Supporting Information).Since lipid 3 exhibited its highest transfection efficiencies at lip-id:DNA charge ratios of 2:1 (or higher), strong lipid-DNA bindinginteractions in the lipoplex might play some role behind the hightransfection efficacies of lipid 3.
3.4. FRET studies
Toward gaining mechanistic insights into whether the dramati-cally contrasting transfection profiles of lipids 3& 4 could possibly berelated to their inherent biomembrane fusogenicity differences oftheir corresponding liposomes, we next evaluated the relativemembrane fusogenicities of the cationic liposomes prepared fromlipids 3 & 4 using the fluorescence resonance energy transfer (FRET)technique pioneered by Struck et al. [22]. In this technique, efficien-cies of different liposomes to fusewith anwidely used biomembranemimicking liposomal formulation (dioleyol-phosphatidylcholine/dioleyol-phatidylethanolamine/dioleyol phosphatidylserine/choles-terol at 45:20:20:15, w/w) are measured. The method depends uponthe interactions which occur between two fluorophores when theemission band of one (the energy donor) overlapswith the excitationbandof the second (theenergyacceptor) andwhensuchfluorophoresare in close physical proximity [22]. These conditions are satisfiedwhen the biomembrane mimicking liposomal formulations areprepared with both the donor and the acceptor fluorophores suchas N-(7-nitro-2-oxa-1,3-diazol-4-yl)-1,2-dihexadecanoyl-sn-glyc-ero-3-phosphoethanolamine (NBD-PE) and Rhodamine Red�-x-1,2-dihexadecanoyl-sn-glycero 3-phosphoethanolamine (Rho-PE),respectively. Since the efficiency of FRET between two such fluo-rophores critically depends upon their spatial separation, any fusionevent of such double-fluorophore containing biomembranemimicking liposomes with cationic liposomes of lipids 3 & 4 (devoidof any fluorophore) decreases the efficiency of resonance energytransfer. The percent of normalized fluorescence recovery was foundto be significantly higher (about 2 fold)when the double-fluorophorelabeled biomembrane mimicking liposomal formulations was incu-bated with lipid 3/Cholesterol liposomes for 30 min compared withthat observed upon treatment of the biomembrane mimicking
liposomes with liposomes of lipid 4 for the same time period (Fig. 4).Consistent with such higher biomembrane fusogenicity of the lipo-somes of lipid 3, the degree of cellular uptake of Rh-PE labeled lipo-somes of lipid 3 was found to be remarkably higher than that forRh-PE labeled liposomes of lipid 4 (Fig. S7). Stated differently, thisfinding demonstrated remarkable impact of structural change in thelinker functionality of cationic amphiphiles in modulating theirfusogenicities. Thus, higher biomembrane fusogenicities of lipid 3/Cholesterol liposome may play an important role behind its high invitro gene transfer efficiencies.
3.5. Membrane rigidity studies by dye entrapment experiment
A more fusogenic liposomal system is likely to be more fluid innature and therefore is likely to possess less membrane rigidity.Thus, the findings in the FRET study described above indicated lessrigid nature of the liposomal membranes of lipid 3 compared to therigidity of the liposomal membranes of lipid 4. Toward qualitative
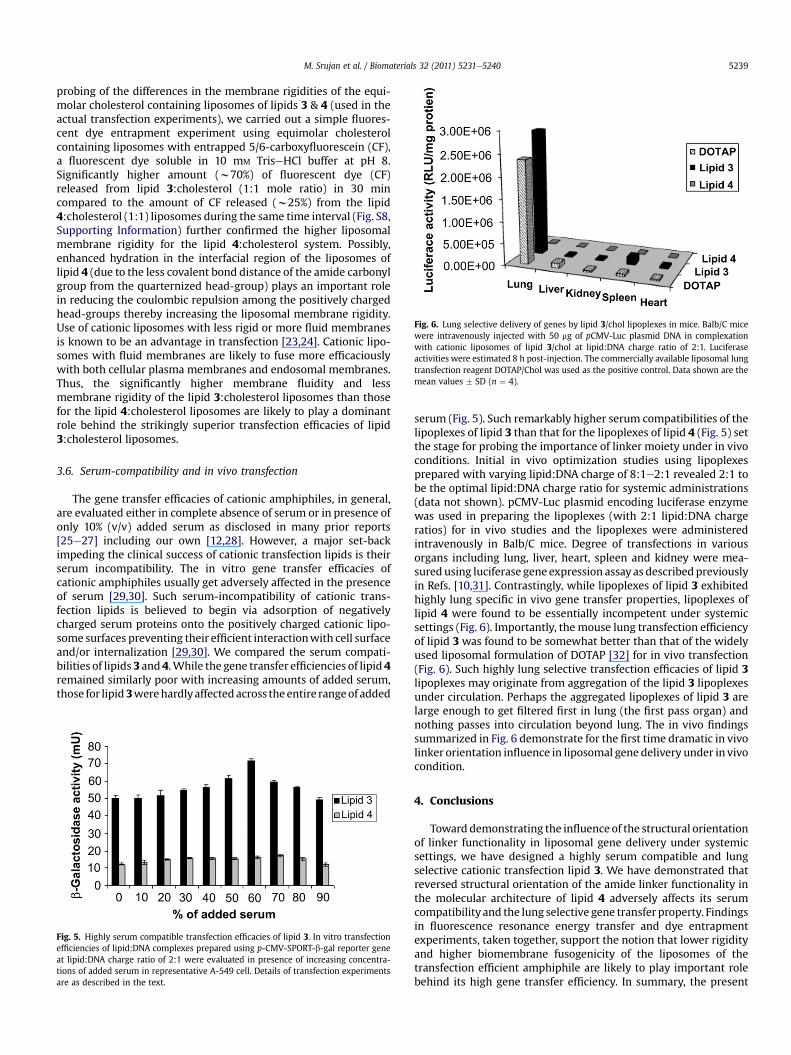
Fig. 6. Lung selective delivery of genes by lipid 3/chol lipoplexes in mice. Balb/C micewere intravenously injected with 50 mg of pCMV-Luc plasmid DNA in complexationwith cationic liposomes of lipid 3/chol at lipid:DNA charge ratio of 2:1. Luciferaseactivities were estimated 8 h post-injection. The commercially available liposomal lungtransfection reagent DOTAP/Chol was used as the positive control. Data shown are themean values � SD (n ¼ 4).
M. Srujan et al. / Biomaterials 32 (2011) 5231e5240 5239
probing of the differences in the membrane rigidities of the equi-molar cholesterol containing liposomes of lipids 3 & 4 (used in theactual transfection experiments), we carried out a simple fluores-cent dye entrapment experiment using equimolar cholesterolcontaining liposomes with entrapped 5/6-carboxyfluorescein (CF),a fluorescent dye soluble in 10 mM TriseHCl buffer at pH 8.Significantly higher amount (w70%) of fluorescent dye (CF)released from lipid 3:cholesterol (1:1 mole ratio) in 30 mincompared to the amount of CF released (w25%) from the lipid4:cholesterol (1:1) liposomes during the same time interval (Fig. S8,Supporting Information) further confirmed the higher liposomalmembrane rigidity for the lipid 4:cholesterol system. Possibly,enhanced hydration in the interfacial region of the liposomes oflipid 4 (due to the less covalent bond distance of the amide carbonylgroup from the quarternized head-group) plays an important rolein reducing the coulombic repulsion among the positively chargedhead-groups thereby increasing the liposomal membrane rigidity.Use of cationic liposomes with less rigid or more fluid membranesis known to be an advantage in transfection [23,24]. Cationic lipo-somes with fluid membranes are likely to fuse more efficaciouslywith both cellular plasma membranes and endosomal membranes.Thus, the significantly higher membrane fluidity and lessmembrane rigidity of the lipid 3:cholesterol liposomes than thosefor the lipid 4:cholesterol liposomes are likely to play a dominantrole behind the strikingly superior transfection efficacies of lipid3:cholesterol liposomes.
3.6. Serum-compatibility and in vivo transfection
The gene transfer efficacies of cationic amphiphiles, in general,are evaluated either in complete absence of serum or in presence ofonly 10% (v/v) added serum as disclosed in many prior reports[25e27] including our own [12,28]. However, a major set-backimpeding the clinical success of cationic transfection lipids is theirserum incompatibility. The in vitro gene transfer efficacies ofcationic amphiphiles usually get adversely affected in the presenceof serum [29,30]. Such serum-incompatibility of cationic trans-fection lipids is believed to begin via adsorption of negativelycharged serum proteins onto the positively charged cationic lipo-some surfaces preventing their efficient interactionwith cell surfaceand/or internalization [29,30]. We compared the serum compati-bilities of lipids 3 and 4.While the gene transfer efficiencies of lipid 4remained similarly poor with increasing amounts of added serum,those for lipid3werehardlyaffectedacross theentire rangeof added
Fig. 5. Highly serum compatible transfection efficacies of lipid 3. In vitro transfectionefficiencies of lipid:DNA complexes prepared using p-CMV-SPORT-b-gal reporter geneat lipid:DNA charge ratio of 2:1 were evaluated in presence of increasing concentra-tions of added serum in representative A-549 cell. Details of transfection experimentsare as described in the text.
serum (Fig. 5). Such remarkably higher serum compatibilities of thelipoplexes of lipid 3 than that for the lipoplexes of lipid 4 (Fig. 5) setthe stage for probing the importance of linker moiety under in vivoconditions. Initial in vivo optimization studies using lipoplexesprepared with varying lipid:DNA charge of 8:1e2:1 revealed 2:1 tobe the optimal lipid:DNA charge ratio for systemic administrations(data not shown). pCMV-Luc plasmid encoding luciferase enzymewas used in preparing the lipoplexes (with 2:1 lipid:DNA chargeratios) for in vivo studies and the lipoplexes were administeredintravenously in Balb/C mice. Degree of transfections in variousorgans including lung, liver, heart, spleen and kidney were mea-sured using luciferase gene expression assayas describedpreviouslyin Refs. [10,31]. Contrastingly, while lipoplexes of lipid 3 exhibitedhighly lung specific in vivo gene transfer properties, lipoplexes oflipid 4 were found to be essentially incompetent under systemicsettings (Fig. 6). Importantly, the mouse lung transfection efficiencyof lipid 3 was found to be somewhat better than that of the widelyused liposomal formulation of DOTAP [32] for in vivo transfection(Fig. 6). Such highly lung selective transfection efficacies of lipid 3lipoplexes may originate from aggregation of the lipid 3 lipoplexesunder circulation. Perhaps the aggregated lipoplexes of lipid 3 arelarge enough to get filtered first in lung (the first pass organ) andnothing passes into circulation beyond lung. The in vivo findingssummarized in Fig. 6 demonstrate for the first time dramatic in vivolinker orientation influence in liposomal gene delivery under in vivocondition.
4. Conclusions
Toward demonstrating the influence of the structural orientationof linker functionality in liposomal gene delivery under systemicsettings, we have designed a highly serum compatible and lungselective cationic transfection lipid 3. We have demonstrated thatreversed structural orientation of the amide linker functionality inthe molecular architecture of lipid 4 adversely affects its serumcompatibilityand the lung selective gene transfer property. Findingsin fluorescence resonance energy transfer and dye entrapmentexperiments, taken together, support the notion that lower rigidityand higher biomembrane fusogenicity of the liposomes of thetransfection efficient amphiphile are likely to play important rolebehind its high gene transfer efficiency. In summary, the present

M. Srujan et al. / Biomaterials 32 (2011) 5231e52405240
findings demonstrate for the first time that amide linker structuralorientation profoundly influences the serum compatibility and lungtransfection efficiencies of cationic amphiphiles.
Acknowledgments
This work was supported by the Council of Scientific andIndustrial Research, Government of India, New Delhi (NWP-0036).We thank N. M. Rao, Centre for Cellular and Molecular Biology,Hyderabad, India for providing us p-CMV-SPORT-b-Gal; MS thankCSIR and VC thank University Grant Commission, Government ofIndia, New Delhi, for providing doctoral research fellowships. Wethank Anunoy Samanta, University of Hyderabad, for his help ingetting the elemental analysis (C, H, N) of lipids 3 & 4 at Universityof Hyderabad elemental analysis facility and Sanjit Kanjilal forproviding us with CAL-B enzyme.
Appendix. Supplementary data
Supplementary data related to this article can be found online atdoi:10.1016/j.biomaterials.2011.03.059.
References
[1] Bhattacharya S, Bajaj A. Advances in gene delivery through molecular designof cationic lipids. Chem Commun (Camb) 2009;31:4632e56.
[2] Srinivas R, Samanta S, Chaudhuri A. Cationic amphiphiles: promising carriersof genetic materials in gene therapy. Chem Soc Rev 2009;38:3326e38.
[3] Karmali PP, Chaudhuri A. Cationic liposomes as non-viral carriers of genemedicines: resolved issues, open questions and future promises. Med Res Rev2007;27:696e722.
[4] Heyes JA, Duvaz DN, Cooper RG, Springer CJ. Synthesis of novel cationic lipids:effect of structural modification on the efficiency of gene transfer. J Med Chem2002;45:99e114.
[5] Fichert T, Regelin A, Massing U. Synthesis and transfection properties of novelnon-toxic monocationic lipids. variation of lipid anchor, spacer and headgroup structure. Bioorg Med Chem Lett 2000;10:787e91.
[6] Byk G, Dubertret C, Escriou V, Frederic M, Jaslin G, Rangara R, et al. Synthesis,activity, and structure-activity relationship studies of novel cationic lipids forDNA transfer. J Med Chem 1998;41:224e35.
[7] Felgner JH, Kumar R, Sridhar CN, Wheeler CJ, Tsai YJ, Border R, et al. Enhancedgene delivery and mechanism studies with a novel series of cationic lipidformulations. J Biol Chem 1994;269:2550e61.
[8] Singh RS, Mukherjee K, Banerjee R, Chaudhuri A, Hait SK, Moulik S, et al.Anchor dependency for non-glycerol based cationic lipofectins: mixed bag ofregular and anomalous transfection profiles. Chem Eur J 2002;8:900e9.
[9] Srilakshmi GV, Sen J, Chaudhuri A, Ramdas Y, Rao NM. Anchor-dependent lip-ofection with non-glycerol based cytofectins containing single 2-hydroxyethylhead groups. Biochim Biophys Acta (Biomembranes) 2002;1559:87e95.
[10] Majeti BK, Singh RS, Yadav SK, Reddy SB, Ramkrishna S, Diwan PV, et al.Enhanced intravenous transgene expression in mouse lung using cyclic-headcationic lipids. Chem Biol 2004;11:427e37.
[11] Banerjee R, Mahidhar YV, Chaudhuri A, Gopal V, Rao NM. Design, synthesis,and transfection biology of novel cationic glycolipids for use in liposomal genedelivery. J Med Chem 2001;44:4176e85.
[12] Banerjee R, Das PK, Srilakshmi GV, Chaudhuri A, Rao NM. Novel series of non-glycerol-based cationic transfection lipids for use in liposomal gene delivery.J Med Chem 1999;42:4292e9.
[13] Karmali PP, Kumar VV, Chaudhuri A. Design, syntheses and in vitro genedelivery efficacies of novel mono-di- and trilysinated cationic lipids: a struc-ture-activity investigation. J Med Chem 2004;47:2123e32.
[14] Ghosh YK, Visweswariah SS, Bhattacharya S. Nature of linkage between thecationic headgroup and cholesteryl skeleton controls gene transfectionefficiency. FEBS Lett 2000;473:341e4.
[15] Singh RS, Chaudhuri A. Single additional methylene group in the head-groupregion imparts high gene transfer efficacy to a transfection-incompetentcationic lipid. FEBS Lett 2004;556:86e90.
[16] Ghosh YK, Viswesaraiah SS, Bhattacharya S. Advantage of the ether linkagebetween the positive charge and the cholesteryl skeleton in cholesterol-basedamphiphiles as vectors for gene delivery. Bioconj Chem 2002;13:378e84.
[17] Koynova R, Li Wang, MacDonald RC. An intracellular lamellar-nonlamellarphase transition rationalizes the superior performance of some cationic lipidtransfection agents. Proc Natl Acad Sci USA 2006;103:14373e8.
[18] Rajesh M, Sen J, Srujan M, Mukherjee K, Sreedhar B, Chaudhuri A. Dramaticinfluence of the orientation of linker between hydrophilic and hydrophobiclipid moiety in liposomal gene delivery. J Am Chem Soc 2007;129:11408e20.
[19] Makwell MAK, Hass SM, Bieber LL, Tolbert NE. A modification of the Lowryprocedure to simplify protein determination in membrane and lipoproteinsamples. Anal Biochem 1978;87:206e10.
[20] Huto CM, Starostin N, West P, Mecartney ML. Acomparison of atomic forcemicroscocy (AFM) and dynamic light scattering (DLS) methods to characterizenanoparticle size distribution. J Nanopart Res 2008;10:89e96.
[21] Hübner W, Blume A. Interactions at the lipidewater interface. Chem PhysLipids 1998;96:99e123.
[22] Struck DK, Hoekstra D, Pagano RE. Use of resonance energy transfer tomonitor membrane fusion. Biochemistry 1981;20:4093e9.
[23] Regelin AE, Fankhaenel S, Gurtesch L, Prinz C, Kiedrowski G, Massing U.Biophysical and lipofection studies of DOTAP analogs. Biochim Biophys Acta2000;1464:151e64.
[24] Savva M, Chen P, Aljaberi A, Selvi B, Spelios M. In vitro lipofection with novelasymmetric series of 1,2-dialkoylamidopropane-based cytofectins containingsingle symmetric bis-(2-dimethylaminoethane) polar headgroups. BioconjugChem 2005;16:1411e22.
[25] Prata CAH, Zhao Y, Barthelemy P, Li Y, Luo D, McIntosh TJ, et al. Charge-reversal amphiphiles for gene delivery. J Am Chem Soc 2004;126:12196e7.
[26] Bell PC, Bergsma M, Dolbnya IP, Bras W, Stuart MC, Rowan AE, et al. Trans-fection mediated by gemini surfactants: engineered escape from theendosomal compartment. J Am Chem Soc 2003;125:1551e8.
[27] Zhu J, Munn RJ, Nantz MH. Self-cleaving ortho ester lipids: a new class ofpH-vulnerable amphiphiles. J Am Chem Soc 2000;122:2645e6.
[28] Kumar VV, Pichon C, Refregiers M, Guerin B, Midoux P, Chaudhuri A. Singlehistidine residue in head-group region is sufficient to impart remarkable genetransfection properties to cationic lipids: evidence for histidine-mediatedmembrane fusion at acidic pH. Gene Ther 2003;10:1206e15.
[29] Li S, Tseng WC, Stolz DB, Wu SP, Watkins SC, Huang L. Dynamic changes in thecharacteristics of cationic lipidic vectors after exposure to mouse serum:implications for intravenous lipofection. Gene Ther 1999;6:585e94.
[30] Zelphati O, Uyechi LS, Barron LG, Szoka Jr FC. Effect of serum components onthe physico-chemical properties of cationic lipid/oligonucleotide complexesand on their interactions with cells. Biochim Biophys Acta 1998;1390:119e33.
[31] Rajesh M, Marepally S, Mahidhar YV, Naidu VGM, Ramakrishna S,Chaudhuri A. Cationic glycolipids with cyclic and open galactose head groupsfor the selective targeting of genes to mouse liver. Biomaterials 2009;30:2369e84.
[32] Song YK, Liu F, Chu S, Liu D. Characterization of cationic liposome-mediatedgene transfer in vivo by intravenous administration. Hum Gene Ther 1997;8:1585e94.




















