
The influence of molecular oxygen exposure on the biology of Prevotella intermedia , with emphasis...
10
ORIGINAL ARTICLE The influence of molecular oxygen exposure on the biology of Prevotella intermedia, with emphasis on its antibiotic susceptibility S.G. dos Santos 1 , C.G. Diniz 2 , V.L. da Silva 1 , N.C. Souza 1 , F.L. de Lima 1 , M.R.Q. Bomfim 1 , M.A.R. de Carvalho 1 and L.M. Farias 1 1 Departamento de Microbiologia, Instituto de Cie ˆ ncias Biolo ´ gicas, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil 2 Departamento de Parasitologia, Microbiologia e Imunologia, Instituto de Cie ˆ ncias Biolo ´ gicas, Universidade Federal de Juiz de Fora, Juiz de Fora, Minas Gerais, Brazil Introduction Prevotella intermedia is a rod-shaped, gram-negative, anaerobic black-pigmented bacterium that belongs to the Bacteroidaceae family (Shah and Gharbia 1992) and may be found in several kinds of clinical specimens from patients with infection in almost all the anatomic sites (Finegold 1995; Santos et al. 2003). Anaerobic therapy is based on modification of the environment to slow down the proliferation of microbes with the use of appropriate antimicrobial agents. Gener- ally, an antimicrobial drug is selected according to the knowledge of suspected pathogens, their anticipated antimicrobial susceptibility pattern and favourable phar- macokinetic properties in an environment with low pH and oxygen tension (Falagas and Siakavellas 2000). According to the literature, however, studies have docu- mented an alarming but gradual increase in resistance rates of anaerobic bacteria to several antibiotics in hospitals in the United States, Europe and elsewhere (Falagas and Siakavellas 2000; Aldridge et al. 2001; Santos et al. 2004). Keywords aerotolerance, antimicrobial susceptibility, Prevotella intermedia. Correspondence Luiz de Macedo Farias, Departamento de Microbiologia, Instituto de Cie ˆ ncias Biolo ´ gicas, Universidade Federal de Minas Gerais. Avenida Anto ˆ nio Carlos, 6627, Pampulha; CEP 31.270-901, Belo Horizonte, MG, Brazil. E-mail: [email protected] 2006 ⁄ 1487: received 25 October 2006, revised 18 December 2006 and accepted 4 January 2007 doi:10.1111/j.1365-2672.2007.03313.x Abstract Aim: This study focuses on investigating the molecular and physiological char- acteristics of Prevotella intermedia after molecular oxygen exposure (MOE) and the effect on drug susceptibility patterns. Methods and Results: Samples of P. intermedia were used as parent strains: ATCC25611 and four clinical isolates. Strains adapted to oxidative stress by MOS were obtained by the enrichment technique. Drug susceptibility was evaluated by minimal inhibitory concentrations (MIC) using agar dilution. Arbitrarily primed-polymerase chain reaction (AP-PCR) was used to evaluate the genetic diversity of all strains and physiological analyses were made by sodiumdodecylsulfate-polyacrylamide gel electrophoresis and two-dimensional electrophoresis of crude, cell-free extracts. The genetic profile showed that line- ages with altered MIC values were selected after MOE. Overall, we found signi- ficant decrease in drug susceptibility for the aero-strains against all tested antimicrobials (amoxicillin, amoxicillin + clavulanic acid, clindamycin, chlo- ramphenicol, ertapenen and metronidazole). We also observed markedly differ- ent protein expression patterns between the parent and selected aero-strains. Conclusions: MOE induces changes in the genetic profile and protein expres- sion patterns of P. intermedia that may also be linked to its drug resistance mechanisms. Significance and Impact of the Study: The effects of MOE on anaerobic bacterial physiology and behaviour may influence antimicrobial susceptibility patterns with potential consequences to antimicrobial chemotherapy. Journal of Applied Microbiology ISSN 1364-5072 882 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891 ª 2007 The Authors
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The influence of molecular oxygen exposure on the biology of Prevotella intermedia , with emphasis...

ORIGINAL ARTICLE
The influence of molecular oxygen exposure on the biologyof Prevotella intermedia, with emphasis on its antibioticsusceptibilityS.G. dos Santos1, C.G. Diniz2, V.L. da Silva1, N.C. Souza1, F.L. de Lima1, M.R.Q. Bomfim1,M.A.R. de Carvalho1 and L.M. Farias1
1 Departamento de Microbiologia, Instituto de Ciencias Biologicas, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
2 Departamento de Parasitologia, Microbiologia e Imunologia, Instituto de Ciencias Biologicas, Universidade Federal de Juiz de Fora, Juiz de Fora,
Minas Gerais, Brazil
Introduction
Prevotella intermedia is a rod-shaped, gram-negative,
anaerobic black-pigmented bacterium that belongs to the
Bacteroidaceae family (Shah and Gharbia 1992) and may
be found in several kinds of clinical specimens from
patients with infection in almost all the anatomic sites
(Finegold 1995; Santos et al. 2003).
Anaerobic therapy is based on modification of the
environment to slow down the proliferation of microbes
with the use of appropriate antimicrobial agents. Gener-
ally, an antimicrobial drug is selected according to the
knowledge of suspected pathogens, their anticipated
antimicrobial susceptibility pattern and favourable phar-
macokinetic properties in an environment with low pH
and oxygen tension (Falagas and Siakavellas 2000).
According to the literature, however, studies have docu-
mented an alarming but gradual increase in resistance
rates of anaerobic bacteria to several antibiotics in
hospitals in the United States, Europe and elsewhere
(Falagas and Siakavellas 2000; Aldridge et al. 2001;
Santos et al. 2004).
Keywords
aerotolerance, antimicrobial susceptibility,
Prevotella intermedia.
Correspondence
Luiz de Macedo Farias, Departamento de
Microbiologia, Instituto de Ciencias Biologicas,
Universidade Federal de Minas Gerais.
Avenida Antonio Carlos, 6627, Pampulha;
CEP 31.270-901, Belo Horizonte, MG, Brazil.
E-mail: [email protected]
2006 ⁄ 1487: received 25 October 2006,
revised 18 December 2006 and accepted 4
January 2007
doi:10.1111/j.1365-2672.2007.03313.x
Abstract
Aim: This study focuses on investigating the molecular and physiological char-
acteristics of Prevotella intermedia after molecular oxygen exposure (MOE) and
the effect on drug susceptibility patterns.
Methods and Results: Samples of P. intermedia were used as parent strains:
ATCC25611 and four clinical isolates. Strains adapted to oxidative stress by
MOS were obtained by the enrichment technique. Drug susceptibility was
evaluated by minimal inhibitory concentrations (MIC) using agar dilution.
Arbitrarily primed-polymerase chain reaction (AP-PCR) was used to evaluate
the genetic diversity of all strains and physiological analyses were made by
sodiumdodecylsulfate-polyacrylamide gel electrophoresis and two-dimensional
electrophoresis of crude, cell-free extracts. The genetic profile showed that line-
ages with altered MIC values were selected after MOE. Overall, we found signi-
ficant decrease in drug susceptibility for the aero-strains against all tested
antimicrobials (amoxicillin, amoxicillin + clavulanic acid, clindamycin, chlo-
ramphenicol, ertapenen and metronidazole). We also observed markedly differ-
ent protein expression patterns between the parent and selected aero-strains.
Conclusions: MOE induces changes in the genetic profile and protein expres-
sion patterns of P. intermedia that may also be linked to its drug resistance
mechanisms.
Significance and Impact of the Study: The effects of MOE on anaerobic
bacterial physiology and behaviour may influence antimicrobial susceptibility
patterns with potential consequences to antimicrobial chemotherapy.
Journal of Applied Microbiology ISSN 1364-5072
882 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891
ª 2007 The Authors

Antibiotics that are frequently used against anaerobic
bacteria include metronidazole, imipenem and merope-
nem which are active against almost all anaerobic bacteria
from clinical isolates, although resistant strains to these
agents have been sporadically reported (Aldridge et al.
2001; Santos et al. 2004; Behra-Miellet et al. 2006). In
contrast to these less common antibiotic resistance pat-
terns, there are two classes that are more commonly seen.
First, resistance to the combination of b-lactam and b-
lactamase inhibitor amoxicillin ⁄ clavulanic acid has been
noted in some multicentre in vitro antimicrobial surveys,
showing somewhat higher resistance rates (Santos et al.
2004; Behra-Miellet et al. 2006). Second, resistance to
clindamycin and cephalosporins such as cefoxitin has
been widely spread and is potentially significant (Teng
et al. 2002). On the other hand, recently developed fluor-
oquinolones such as tosufloxacin, demonstrate improved
antianaerobic activity (Takahata and Nishino 1997).
Because of its cost effectiveness, especially in developing
countries, chloramphenicol continues to be widely used
against anaerobes despite its known haemotoxicity
(Turton et al. 1999).
Oxidative stress has been defined as a disturbance in
the pro-oxidant–antioxidant balance in favour of pro-oxi-
dants (Zheng and Storz 2000). Reactive oxygen species
(ROS) such as O2–, HO and H2O2 are extremely harmful
as they easily penetrate membranes and diffuse through
cells (Zheng and Storz 2000). Thus, the alteration of nor-
mal levels of ROS may cause injury by oxidative stress
raising lesions in different tissues. Several substances can
affect ROS production by cells during oxidative processes
such as exposure to oxygen, radiation, organic peroxides,
metals and chemical substances which may generate intra-
cellular free radicals; a mechanism utilized by some anti-
microbial drugs (Zheng and Storz 2000; Albesa et al.
2004).
The alteration of oxidative metabolism and the increase
of oxygen may affect the mechanism of action of various
antibiotics. Studies indicate that diverse antibiotics can
generate an increase of O)2 in different bacterial species
(Albesa et al. 2004; Koutsolioutsou et al. 2005). Cipro-
floxacin, ceftazidime, piperacillin and chloramphenicol
stimulate oxidative stress in bacteria that may be useful in
explaining aspects of antibacterial action which are, as of
yet, unaccounted (Albesa et al. 2004; Koutsolioutsou et al.
2005).
The production of enzymes involved in oxygen meta-
bolism by an organism – such as superoxide dismutase,
catalase and peroxidase – is mirrored by variable degrees
of tolerance and virulence in some strains and, therefore,
in their pathogenic potential (Silva et al. 2005). The abil-
ity to respond to oxidative stress when exiting in its nat-
ural anaerobic environment to more oxygenated tissues as
well as to resist oxidative events from the host immune
system until anaerobic conditions are restored in the
infection site is an important acquired adaptation for
anaerobic bacteria (Lynch and Kuramitsu 2000).
Little data on the regulation of oxidative stress control
in Prevotella spp. are available. Increasing knowledge
about ROS neutralizing enzymes offers enticing prospects
for improving our understanding in several aspects of the
molecular biology, physiology and ecology of this import-
ant microbial group and how its members participate in
infectious diseases.
Our objective in the present study was to evaluate the
P. intermedia response to oxidative stress by exposing this
anaerobic bacterium to molecular oxygen and then iden-
tify the consequences to microbial biology with regards to
its antimicrobial drug susceptibility patterns.
Materials and methods
Bacterial strains
Five P. intermedia strains were used as parent strains in
this study: the reference P. intermedia ATCC25611 (Wt-
ref) and four P. intermedia strains isolated from patients
with intra-abdominal infections (Wt-34, Wt-71, Wt-96
and Wt-134).
All bacterial strains were routinely cultivated anaerobi-
cally, at 37�C (anaerobic chamber; Forma Scientific Com-
pany, Marietta, OH, USA, containing an atmosphere of
N2 85%, H2 10%, CO2 5%) in Brucella broth (BB; Difco,
Sparks, MD, USA), supplemented with 0Æ05 mg dl)1 of
hemin, 0Æ01 mg dl)1 of menadione, 5% yeast extract
(BB-S) and stored at –86�C (Holdeman et al. 1977).
Throughout the experiments, the strains were checked
by gram staining and aerotolerance tests (Holdeman et al.
1977).
Adaptation of Prevotella intermedia ATCC 25611 and
Prevotella intermedia clinical isolates to atmospheric
molecular oxygen
Strains adapted to oxidative stress were derived from the
parent P. intermedia ATCC25611 (Wt-ref) and the clinical
isolates (Wt-34, Wt-71, Wt-96 and Wt-134) by atmo-
spheric oxygen exposure, as already described (Silva et al.
2005). Briefly, a 300-ml volume of BB was inoculated
with nearly 10% of an overnight culture of each parent
strain and the resulting bacterial suspension was grown to
mid-exponential phase. The bacterial cultures were split
into two 150-ml subcultures. One of the subcultures was
kept in the anaerobic chamber as a control and the other
was placed in an aerobic shaker (Cientec; Biosan, Sao
Paulo, SP, Brazil) adjusted to 200 rev min)1 and 37�C for
S.G. dos Santos et al. Influence of molecular oxygen on Prevotella intermedia biology
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891 883

aeration and temperature. Every 6 h, up to a maximum
of 72 h, samples from both subcultures were withdrawn
and aliquots were evaluated for OD at 600 nm and were
plated after 10-fold serial dilutions, in triplicate, to count
colony-forming units per millilitre (CFU ml)1) onto
Brucella agar supplemented with 0Æ05 mg dl)1 hemin,
0Æ01 mg dl)1 menadione and 5% yeast extract (BA-S). All
plates used to recover bacteria for viable cell counts were
incubated anaerobically, at 37�C, for 48 h. The plates cor-
responding to the last point of atmospheric oxygen expo-
sure at which survival colonies were observed were used
to re-isolate P. intermedia strains to be used for further
adaptive steps. One colony of the surviving derived ones
from each bacterial strain were subcultured in BB for
24 h in an anaerobic chamber and then submitted to four
cycles of 12 h atmospheric oxygen exposure and sub-
culturing as before. The strain resulting from the fourth
re-isolation was considered as the aero-tolerant strain
(Aero-ref, Aero-34, Aero-71, Aero-96 and Aero-134).
Survival curve of Prevotella intermedia strains selected
by molecular oxygen exposure
Two volumes of 600 ml BRU-S each were inoculated
with 10% of an overnight culture (initial OD600 nm =
0Æ136) of WT-strains or aero-strains, respectively, and the
cultures were incubated in the anaerobic chamber at
37�C, until the mid-exponential phase. Then, each culture
was split into two 300-ml subcultures. One 300-ml
aliquot of each culture was kept inside the anaerobic
chamber as control and the other 300-ml aliquot was
removed and kept under aerobic conditions on a shaker
at 200 rev min)1 and 37�C. Every 6 h, up to 144 h, sam-
ples from all subcultures were withdrawn and aliquots
were evaluated for OD at 600 nm and plated onto BRU-S
after 10-fold serial dilutions, in triplicate, for viable cell
counts (CFC ml)1).
Biochemical identification and identity confirmation of
selected strains by oxygen exposure
The identification and the identity confirmation of the
bacterial strains selected after molecular oxygen exposure
(MOS), including the P. intermedia reference strain, were
based on the biochemical, physiological or enzymatic
characteristics using the Rapid-ID 32A system (Bio-
Merieux, Marcy- I’Etoile, France).
Crude cell-free protein extracts from the parent
Prevotella and their derivative strains selected after MOS
Crude cell-free protein extracts were performed according
to Shibata et al. (2005) and (Westeimeier and Naven)
2002. Pellets from mid-log phase cultures of parent (Wt)
and aero-strains were washed in sterile sucrose buffer
(10 mmol l)1 of Tris, 250 mmol l)1 of sucrose, pH 7)
and adjusted to a concentration of approximately 108
cells ml)1 (OD = 0Æ4, at 600 nm). Five-millilitre aliquots
of each cell suspension were precipitated by adding 50%
trichloroacetic acid (Sigma, St. Louis, MO, USA) to a
final concentration of 10% to prevent degradation of
proteins by proteases or endopeptidases and centrifuged
at 8500 g for 10 min at 4�C. The harvested cells
were solubilized in 4 ml of lysis solution [7 mol l)1
of urea, 2 mol l)1 of thiourea, 4% 3-[(3-cholamido-
propyl)-dimethylammonio]-1-propanesulfonate (CHAPS),
40 mmol l)1 of Tris and 1 mmol l)1 of EDTA), and were
lysed by ultrasonication (Branson sonifier 450; Branson
Ultrasonics Corp., Danbury, CT, USA), followed by cen-
trifugation for 15 min at 10 000 g (Laemmli 1970), and
then protein content was measured (PlusOne 2-D Quant
kit; Amersham Biosciences, Little Chalfont, UK).
Identification of differentially regulated proteins by
sodiumdodecylsulfate-polyacrylamide gel electrophoresis
The crude cell-free protein extracts were fractionated in
sodiumdodecylsulfate-polyacrylamide gel electrophoresis
(12% SDS-PAGE) (Laemmli 1970). Electrophoresis was
performed in Tris-glycine-SDS buffer (10 mmol l)1 of
Tris, 250 mmol l)1 of sucrose, pH 7), under constant cur-
rent of 30 mA over 4 h. Proteins were visualized by
Coomassie blue staining (Westeimeier and Naven 2002).
Identification of differentially regulated proteins by
two-dimensional electrophoresis
The crude protein extracts were submitted to two-dimen-
sional gel electrophoresis, as described before (Rocha
et al. 1996; Westeimeier and Naven 2002; Shibata et al.
2005). Briefly, the first dimension electrophoresis, based
on isoelectric focusing, was carried out using a poly-
acrylamide strip, with a pH gradient from 4Æ0 to 7Æ0(IPG; Immobiline DryStrip Kit; Amersham Pharmacia
Biotech), which was loaded with 200 lg of crude cell
extract. Electrophoresis was performed for 20 h, at 20�C,
1 mA and 5 W, using a gradient programming electro-
phoresis power supply EPS 3500 XL (Amersham Pharma-
cia Biotech), as follows: 100 V, 0Æ01 h; 100 V, 4 h; 300 V,
0Æ01 h; 300 V, 4Æ5 h; 2000 V, 5 h and 2000 V, 6Æ5 h
(Veith et al. 2001). With isoelectric focusing complete,
second dimension electrophoresis was carried by 12%
SDS-PAGE (Laemmli 1970). Finally, the gels were stained
with colloidal Coomassie brilliant blue G 250 for analysis
(Westeimeier and Naven 2002). The assays were per-
formed in triplicate.
Influence of molecular oxygen on Prevotella intermedia biology S.G. dos Santos et al.
884 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891
ª 2007 The Authors

Arbitrarily primed-polymerase chain reaction typing
Arbitrarily primed-polymerase chain reaction (AP-PCR)
typing for the Prevotella strains was based on previous
studies (Kononen et al. 1998; Matto et al. 1999). Briefly,
the isolates were grown on BRU-S plates under anaerobic
atmosphere at 37�C for 24 h. For each parent or oxygen
exposure selected strain, 10 single colonies were collected
from the plates from which DNA was extracted using
Wizard Genomic DNA Purification Kit (Promega Cor-
poration, Madison, WI, USA). Primers Opa-13 (5¢-CAG-
CACCCAC-3¢) and Opa-03 (5¢-AGTCAGCCAC-3¢) were
employed for typing because of better discriminatory
power shown in previous studies (Kononen et al. 1998;
Veith et al. 2001) (Integrated DNA Technologies, Coral-
ville, IA, USA). AP-PCR was performed using the follow-
ing mix (50-ll final volume): 2X PCR Master Mix
(Promega Corporation, Madison, WI, USA), 0Æ8 lmol l)1
of primer and 5 ll of DNA suspension (1 ⁄ 100 dilution).
Amplifications were carried out in a DNA thermal cycler
(PCR Express Thermo Hybaid, Ashford, Middlesex, UK).
An internal negative control was included in each
AP-PCR run. Samples were amplified by initial denatura-
tion at 94�C for 5 min followed by 35 cycles with dena-
turation at 94�C for 1 min, annealing at 32�C for 2 min
and extension at 72�C for 2 min, and the final extension
at 72�C for 5 min (Kononen et al. 1998; Matto et al.
1999).
Analysis of the DNA fragments amplified by AP-PCR
Amplified products were separated by PAGE stained with
silver nitrate (Oliveira et al. 1995) and digitally photo-
graphed. Then, the gels were carefully analysed in order
to determine the main fragments of the AP-PCR profiles
in comparison with the molecular size of the fragments of
a marker, 1-kb DNA Ladder (Promega Corporation). The
similarity among the AP-PCR profiles obtained with the
DNA of the parent or the oxygen exposure-selected strain
P. intermedia was analysed with the LabImage -1D gel
analysis software, version 2Æ7Æ2. Copyright 1999–2004,
Kapelan GmbH, Halle (Saale), Germany. Available at:
http://www.labimage.net (Nei 1972).
Dendrogram
AP-PCR profiles were scored based on the presence (i) or
absence (ii) of each polymorphic DNA fragment gener-
ated by each primer (OPA-3 ⁄ OPA-13) with DNA from
the parent strains and those submitted to oxidative stress.
Cluster analysis based on similarity using Nei’s (1972)
original distance was performed by the Unweighted Pair
Group Method (UPGMA) with arithmetic averages clus-
tering algorithm (Sneath and Sokal 1973) and the rand-
omization procedure as implemented in TFPGA (Tools
for Population Genetic Analyses) software package (Miller
1998).
Minimum inhibitory concentration determination
The minimum inhibitory concentration (MIC) was per-
formed using agar dilution method (Holdeman et al.
1977; NCCLS 2004). Antibiotic stock solutions were
added to melted BA (Difco) to obtain final concentra-
tions ranging from 0Æ05 to 1025Æ0 lg ml)1. The anti-
microbial drugs were selected based on microbial
characteristics and clinical relevance: metronidazole
(Oxoid Ltd, Basingstoke, Hampshire, England), ertape-
nem (Merck & Co., Inc., Whitehouse Station, NJ, USA),
clindamycin (Sigma Chemical Co., St. Louis, MO, USA),
amoxillin (Sigma), choramphenicol (Sigma) and amoxi-
cillin–clavulanate (Oxoid). All samples were grown near
108 UFC ml)1 corresponding to 0Æ5 on the MacFarland
scale, and then inoculated into BB-S (Difco) with antimi-
crobial agents, using a ‘Steers’ replicator (Steers et al.
1959). The reference strains Bacteroides fragilis (ATCC
25285) and Eubacterium lentum (ATCC 43055) were also
included as controls. The plates were incubated in an
anaerobic chamber at 37�C for 48 h. The MIC was meas-
ured as the lowest concentration able to inhibit microbial
growth as interpreted according to Clinical Laboratories
Standards Institute (formerly, National Committee for
Clinical Laboratory Standards) recommendations (NCCLS
2004).
Results
Survival curve of Prevotella intermedia ATCC 25611
(Wt-strain) and aero-strains under atmospheric oxygen
exposure
Five P. intermedia strains adapted to oxidative stress from
atmospheric oxygen exposure were obtained (Aero-ref,
Aero-34, Aero-71, Aero-96 and Aero-134) from parental
strain ATCC 25611 (Wt-ref) and the intra-abdominal
infection clinical isolates (Wt-34, Wt-71, Wt-96 and
Wt-134), after five cycles of aeration, according to the
aforementioned methods. The efficiency of the adaptive
process was demonstrated by the progressive increase in
aerotolerance of P. intermedia strains observed after each
successive cycle. To confirm the adapted phenotype, sur-
vival curves under oxidative conditions were compared
between parent and aero-strains. Viable cells were no lon-
ger observed after 48-h exposure to atmospheric oxygen
for all parent strains, whereas aero-strains were viable for
up to 96 h under the same conditions.
S.G. dos Santos et al. Influence of molecular oxygen on Prevotella intermedia biology
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891 885
Biochemical identification
Biochemical–physiological characterization of P. interme-
dia strains subjected to oxidative stress by MOS did not
show significant alterations. The resulting phenotypes
were similar to that of the reference strain P. intermedia
ATCC 25611.
Comparison between protein expression patterns of parent
Prevotella intermedia and aero-strains selected by MOS
The protein expression patterns were compared for all
tested strains by SDS-PAGE analysis. In addition, the pro-
tein expression patterns of P. intermedia ATCC 25611
(Wt-ref) and its derivative strain (Aero-ref) were also
compared using two-dimensional eletrophoresis and
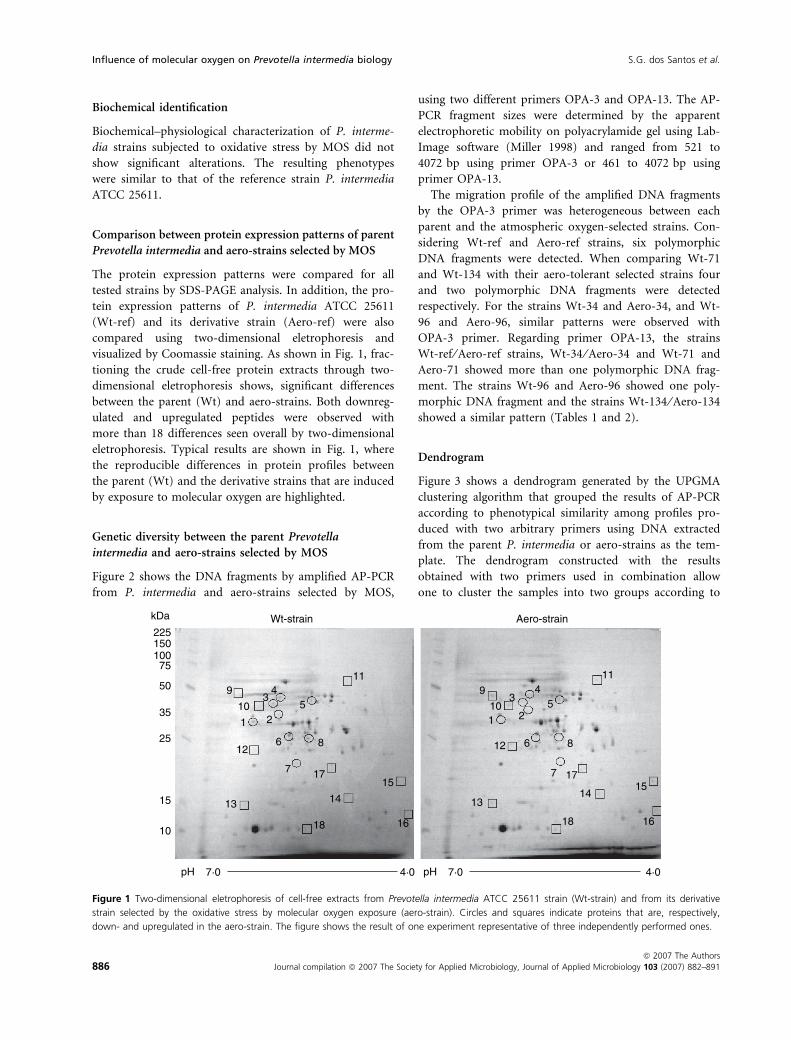
visualized by Coomassie staining. As shown in Fig. 1, frac-
tioning the crude cell-free protein extracts through two-
dimensional eletrophoresis shows, significant differences
between the parent (Wt) and aero-strains. Both downreg-
ulated and upregulated peptides were observed with
more than 18 differences seen overall by two-dimensional
eletrophoresis. Typical results are shown in Fig. 1, where
the reproducible differences in protein profiles between
the parent (Wt) and the derivative strains that are induced
by exposure to molecular oxygen are highlighted.
Genetic diversity between the parent Prevotella
intermedia and aero-strains selected by MOS
Figure 2 shows the DNA fragments by amplified AP-PCR
from P. intermedia and aero-strains selected by MOS,
using two different primers OPA-3 and OPA-13. The AP-
PCR fragment sizes were determined by the apparent
electrophoretic mobility on polyacrylamide gel using Lab-
Image software (Miller 1998) and ranged from 521 to
4072 bp using primer OPA-3 or 461 to 4072 bp using
primer OPA-13.
The migration profile of the amplified DNA fragments
by the OPA-3 primer was heterogeneous between each
parent and the atmospheric oxygen-selected strains. Con-
sidering Wt-ref and Aero-ref strains, six polymorphic
DNA fragments were detected. When comparing Wt-71
and Wt-134 with their aero-tolerant selected strains four
and two polymorphic DNA fragments were detected
respectively. For the strains Wt-34 and Aero-34, and Wt-
96 and Aero-96, similar patterns were observed with
OPA-3 primer. Regarding primer OPA-13, the strains
Wt-ref ⁄ Aero-ref strains, Wt-34 ⁄ Aero-34 and Wt-71 and
Aero-71 showed more than one polymorphic DNA frag-
ment. The strains Wt-96 and Aero-96 showed one poly-
morphic DNA fragment and the strains Wt-134 ⁄ Aero-134
showed a similar pattern (Tables 1 and 2).
Dendrogram
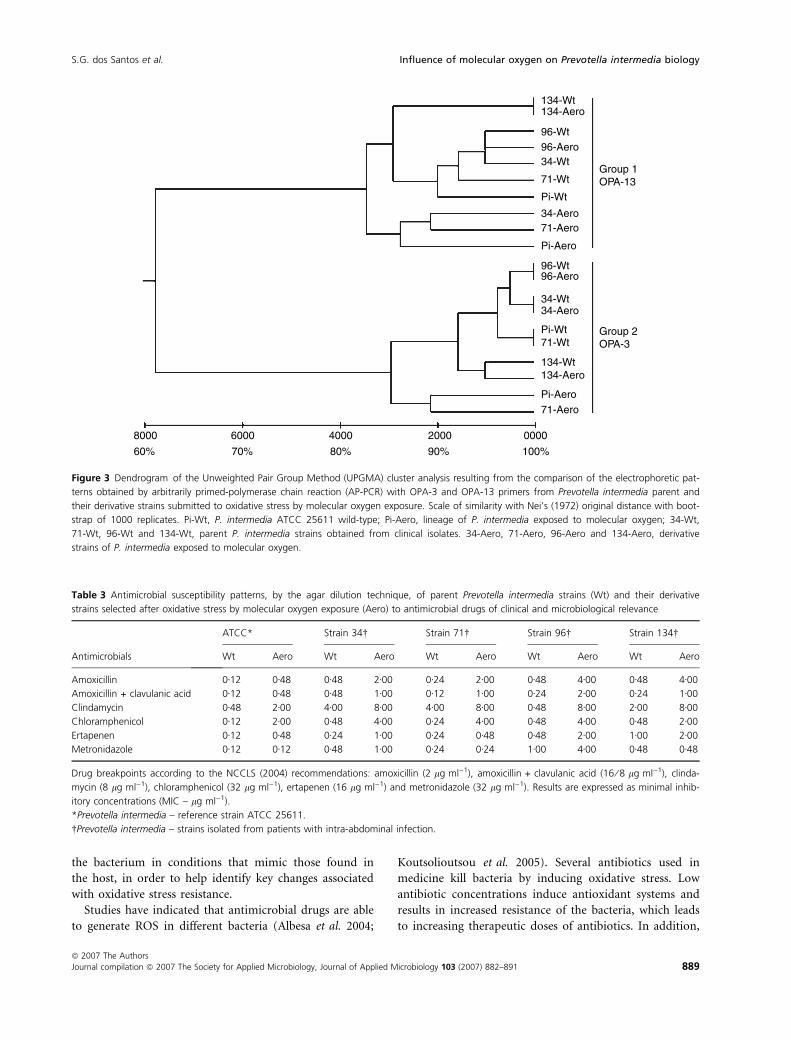
Figure 3 shows a dendrogram generated by the UPGMA
clustering algorithm that grouped the results of AP-PCR
according to phenotypical similarity among profiles pro-
duced with two arbitrary primers using DNA extracted
from the parent P. intermedia or aero-strains as the tem-
plate. The dendrogram constructed with the results
obtained with two primers used in combination allow
one to cluster the samples into two groups according to
kDa Wt-strain Aero-strain22515010075
50 9
1035
25
15
10
pH 7·0 4·0 pH 7·0 4·0
1 2
34
8
5
11 11
612
7 17
14
18
13
15 1514
13
18 16
177
6 8
549
101
3
2
12
16
Figure 1 Two-dimensional eletrophoresis of cell-free extracts from Prevotella intermedia ATCC 25611 strain (Wt-strain) and from its derivative
strain selected by the oxidative stress by molecular oxygen exposure (aero-strain). Circles and squares indicate proteins that are, respectively,
down- and upregulated in the aero-strain. The figure shows the result of one experiment representative of three independently performed ones.
Influence of molecular oxygen on Prevotella intermedia biology S.G. dos Santos et al.
886 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891
ª 2007 The Authors

the similarity between them. Group 1 contains the sam-
ples amplified with OPA-13 primer and in group 2 those
samples amplified by AP-PCR using OPA-3 primer.
MIC determinations
We observed that all P. intermedia strains (Wt) tested in
this study had an altered drug susceptibility profile after
MOS (aero-strains, Table 3). Overall, a significant
decrease in drug susceptibility of the aero-strains for all
tested antimicrobials (amoxicillin, amoxicillin + clavul-
anic acid, clindamycin, chloramphenicol, ertapenen and
metronidazole) was observed. Taken together as a mean
value for the aero-strains, if compared with that for the
parent strains, the recorded MIC value was 8Æ89 times
higher for chloramphenicol, 6Æ93 times higher for amoxi-
bp
M 1
PrimerOPA-3
PrimerOPA-3
PrimerOPA-3
PrimerOPA-3
PrimerOPA-3
PrimerOPA-13
PrimerOPA-13
PrimerOPA-13
PrimerOPA-13
PrimerOPA-13
2 1 2 3 4 3 4 5 6 5 6 7 8 7 8 9 10 9 10 M
407230542036
1018
517506
396344298
220202
Figure 2 Electrophoretic profile generated by arbitrarily primed-polymerase chain reaction (AP-PCR) by employing primers OPA-3 and OPA-13 of
Prevotella intermedia parent or oxidative stress-adapted strains. M, Molecular size of marker; lanes 1, 3, 5, 7 and 9, parent strains (ATCC 25611,
34, 71, 96 and 134, respectively); lanes 2, 4, 6, 8 and 10, derivative strains selected by exposure to molecular oxygen (ATCC 25611, 34, 71, 96
and 134, respectively).
Table 1 Relative size (in base pairs) of the specific arbitrarily primed-polymerase chain reaction (AP-PCR) products from Prevotella intermedia
parent or oxidative stress-adapted strains obtained by amplification with OPA-3 and OPA-13 primers, respectively
Tested strains
AP-PCR products*
OPA-3 primer OPA-13 primer
Wt-ref � 521, 586, 747, 1183, 1340, 1656, 1850, 1954, 4072 921, 1057, 1374, 1656, 2464, 4072
Aero-ref� 521, 528, 586, 1183, 1656, 1954, 2240 461, 586, 921, 1057, 1125, 1656, 2036, 2464
Wt-34� 521, 586, 670, 747, 1183, 1340, 1656, 1954, 4072 461, 921, 1057, 1340, 1480, 1656, 2464, 4072
Aero-34� 521, 586, 670, 747, 1183, 1340, 1656, 1954, 4072 461, 921, 1057, 1139, 1480, 1656, 2036, 2464
Wt-71� 521, 586, 747, 1183, 1340, 1656, 1850, 1954, 4072 921, 1340, 1656, 2464, 4072
Aero-71� 521, 586, 1183, 1656, 4072 1139, 1656, 2036, 2464
Wt-96� 521, 586, 670, 747, 1183, 1340, 1656, 1850, 1954, 4072 461, 586, 921, 1057, 1340, 1656, 2464, 4072
Aero-96� 521,586, 670, 747, 1183, 1340, 1656, 1850, 1954, 4072 461, 921, 1057, 1139, 1340, 1656, 2464, 4072
Wt-134� 521, 586, 670, 747, 1183, 1656, 4072 461, 921, 1057, 1139, 1183, 1340, 2036, 2464, 4072
Aero-134� 521, 586, 670, 747, 1183, 1340, 1656, 2036, 4072 461, 921, 1057, 1139, 1183, 1340, 2036, 2464, 4072
Bold numbers represent DNA bands with high intensity, as observed after electrophoresis.
*Used primers by AP-PCR.
�Reference strains of P. intermedia.
�Prevotella intermedia strains submitted to molecular oxygen exposure.
Abbreviations: Pi-Wt, P. intermedia ATCC 25611 wild-type; Pi-Aero, lineage of P. intermedia exposed to molecular oxygen; 34-Wt, 71-Wt, 96-Wt
and 134-Wt, P. intermedia strains obtained from clinical isolates; 34-Aero, 71-Aero, 96-Aero and 134-Aero, lineage of P. intermedia exposed to
molecular oxygen.
S.G. dos Santos et al. Influence of molecular oxygen on Prevotella intermedia biology
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891 887
cillin, 3Æ10 times higher for clindamycin, 4Æ57 times
higher for the combination treatment of amoxicil-
lin + clavulanic acid, 2Æ87 times higher for ertapenen and
2Æ52 times higher for metronidazole. Although there were
decreased sensitivity patterns to the antimicrobial drugs
tested, full resistance was not observed for the tested
aero-strains.
Discussion
Our study reinforces this idea as the obligate anaerobe P.
intermedia was able to develop an efficient response to
MOS that resulted in a survival time at least twice as
long.
Data generated by SDS-PAGE and two-dimensional
electrophoresis show a complex shift in the aero-strains
protein expression patterns when compared with their
parent Wt-strains band profiles as revealed by both
down- and upregulated proteins with different molecular
mass and isoelectric points (Fig. 1). Differences of these
magnitudes may have not resulted from the accumulation
of single point mutations and thus may be part of a
broader programme of genetic control that alters the
physiology to protect and prevent oxidative stress damage
to the cell. However, our results suggest that for these
bacteria the differences observed did not significantly alter
the major metabolic pathways in these cells, once no sig-
nificant alterations were observed in aero-strains based on
the biochemical and physiological or enzymatic character-
istics using the Rapid-ID 32A system.
These global changes, while not entirely understood, do
have other significant effects on the organisms, as related
to their virulence patterns, reflecting the host–bacterial
relationship, as already reported (Silva et al. 2005). Previ-
ous studies document (Rocha et al. 1996; Rocha and
Smith 1997; Fletcher et al. 2001) that environmental fac-
tors, including atmospheric conditions, may induce phy-
siological modulations as observed in this study. Also, the
influence of antimicrobial agents, as another environmen-
tal factor, on the regulation of protein expression by B.
fragilis has already been demonstrated (Diniz et al. 2004).
During entry into the host organism, anaerobic bacteria
may be exposed to high oxygen levels and may be persist-
ently stressed by oxidative bursts that originate from
the host immune response (Katsuragi et al. 2003). It is
assumed that an adaptive response to this oxidative stress
is a key factor for bacterium survival (Rocha et al. 1996).
It seems that this ability represents an important charac-
teristic for an invasive process from the natural anaerobic
environment to more oxygenated tissues of the body
(Lynch and Kuramitsu 2000; Zheng and Storz 2000; Silva
et al. 2005).
The AP-PCR method may be applied to detect poly-
morphisms in a wide variety of organisms using different
primers (George et al. 1997). This technique has been fre-
quently used for clonal analysis, being particularly useful
for determining whether isolates of the same taxa are epi-
demiologically related (Camarena et al. 1995; Song 2005).
In this study, the combination genotyping using two
arbitrary primers, showed lower discriminatory power of
AP-PCR typing with the primers OPA-3 than with OPA-
13 (Fig. 2). Similar results have also been reported with
these same primers and the AP-PCR method when
applied to detect polymorphisms in Neisseria gonorrhoeae
(Camarena et al. 1995).
As observed by the arbitrary amplifications of DNA in
all bacterial strains after MOS by AP-PCR, several altera-
tions were detected in the banding patterns generated
(Tables 1 and 2) and this fact might not be considered
merely a phenotypic adaptive response (Fig. 3). Moreover,
alterations were also observed by the cellular proteins’
electrophoregrams (Fig. 1).
Although there have been great advances in microbial
genomics and proteomics over the past decade, little is
known regarding the mechanisms of bacterial oxidative
stress adaptive response and the influences of this adapta-
tion on the micro-organism–host relationship. In this
regard, it may be useful to evaluate the aerotolerance of
Table 2 Numbers of polymorphic fragments generated by arbitrarily
primed-polymerase chain reaction (AP-PCR) using OPA-3 and OPA-13
primers
Used primers*
OPA-3 OPA-13
Total of
fragments
Polymorphic
fragments
Total of
fragments
Polymorphic
fragments
Wt-ref� 9 4 6 2
Aero-ref c 7 2 8 4
Wt-34� 9 0 8 2
Aero-34� 9 0 8 2
Wt-71� 9 4 5 3
Aero-71� 5 0 4 2
Wt-96� 10 0 8 1
Aero-96� 10 0 8 1
Wt-134� 7 0 9 0
Aero-134� 9 2 9 0
Total 84 12 73 17
*Used primers by AP-PCR.
�Reference strains of Prevotella intermedia.
�Prevotella intermedia strains submitted to molecular oxygen expo-
sure.
Abbreviations: Pi-Wt, P. intermedia ATCC 25611 wild-type; Pi-Aero,
lineage of P. intermedia exposed to molecular oxygen; 34-Wt, 71-Wt,
96-Wt and 134-Wt, P. intermedia strains obtained from clinical
isolates; 34-Aero, 71-Aero, 96-Aero and 134-Aero, lineage of P. inter-
media exposed to molecular oxygen.
Influence of molecular oxygen on Prevotella intermedia biology S.G. dos Santos et al.
888 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891
ª 2007 The Authors
the bacterium in conditions that mimic those found in
the host, in order to help identify key changes associated
with oxidative stress resistance.
Studies have indicated that antimicrobial drugs are able
to generate ROS in different bacteria (Albesa et al. 2004;
Koutsolioutsou et al. 2005). Several antibiotics used in
medicine kill bacteria by inducing oxidative stress. Low
antibiotic concentrations induce antioxidant systems and
results in increased resistance of the bacteria, which leads
to increasing therapeutic doses of antibiotics. In addition,
Table 3 Antimicrobial susceptibility patterns, by the agar dilution technique, of parent Prevotella intermedia strains (Wt) and their derivative
strains selected after oxidative stress by molecular oxygen exposure (Aero) to antimicrobial drugs of clinical and microbiological relevance
Antimicrobials
ATCC* Strain 34� Strain 71� Strain 96� Strain 134�
Wt Aero Wt Aero Wt Aero Wt Aero Wt Aero
Amoxicillin 0Æ12 0Æ48 0Æ48 2Æ00 0Æ24 2Æ00 0Æ48 4Æ00 0Æ48 4Æ00
Amoxicillin + clavulanic acid 0Æ12 0Æ48 0Æ48 1Æ00 0Æ12 1Æ00 0Æ24 2Æ00 0Æ24 1Æ00
Clindamycin 0Æ48 2Æ00 4Æ00 8Æ00 4Æ00 8Æ00 0Æ48 8Æ00 2Æ00 8Æ00
Chloramphenicol 0Æ12 2Æ00 0Æ48 4Æ00 0Æ24 4Æ00 0Æ48 4Æ00 0Æ48 2Æ00
Ertapenen 0Æ12 0Æ48 0Æ24 1Æ00 0Æ24 0Æ48 0Æ48 2Æ00 1Æ00 2Æ00
Metronidazole 0Æ12 0Æ12 0Æ48 1Æ00 0Æ24 0Æ24 1Æ00 4Æ00 0Æ48 0Æ48
Drug breakpoints according to the NCCLS (2004) recommendations: amoxicillin (2 lg ml)1), amoxicillin + clavulanic acid (16 ⁄ 8 lg ml)1), clinda-
mycin (8 lg ml)1), chloramphenicol (32 lg ml)1), ertapenen (16 lg ml)1) and metronidazole (32 lg ml)1). Results are expressed as minimal inhib-
itory concentrations (MIC – lg ml)1).
*Prevotella intermedia – reference strain ATCC 25611.
�Prevotella intermedia – strains isolated from patients with intra-abdominal infection.
8000 6000 4000 2000 0000
71-Aero
71-Aero34-Aero
Pi-Wt
34-WtGroup 1OPA-13
Group 2OPA-3
96-Aero96-Wt
134-Aero134-Wt
71-Wt
Pi-Aero
Pi-Wt
134-Aero
34-Aero
96-Aero
Pi-Aero
96-Wt
34-Wt
134-Wt
71-Wt
60% 70% 80% 90% 100%
Figure 3 Dendrogram of the Unweighted Pair Group Method (UPGMA) cluster analysis resulting from the comparison of the electrophoretic pat-
terns obtained by arbitrarily primed-polymerase chain reaction (AP-PCR) with OPA-3 and OPA-13 primers from Prevotella intermedia parent and
their derivative strains submitted to oxidative stress by molecular oxygen exposure. Scale of similarity with Nei’s (1972) original distance with boot-
strap of 1000 replicates. Pi-Wt, P. intermedia ATCC 25611 wild-type; Pi-Aero, lineage of P. intermedia exposed to molecular oxygen; 34-Wt,
71-Wt, 96-Wt and 134-Wt, parent P. intermedia strains obtained from clinical isolates. 34-Aero, 71-Aero, 96-Aero and 134-Aero, derivative
strains of P. intermedia exposed to molecular oxygen.
S.G. dos Santos et al. Influence of molecular oxygen on Prevotella intermedia biology
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891 889

the increase in antioxidant potential could generate
new bacterial strains resistant to antibiotics, particularly
those that affect bacteria via the generation of ROS
(Koutsolioutsou et al. 2001).
The strains selected in this study after MOS showed
altered drug susceptibility profiles with significant decrea-
ses, up to four times, in drug sensitivity to clindamycin
followed by chloramphenicol and amoxicillin, compared
with the parent strains. This altered susceptibility might
be related to the microbial protective effect against oxida-
tive stress as they were selected by previous oxygen expo-
sure. However, as MIC values did not achieve the break
points of the assayed drugs, antimicrobial resistance was
not observed for any strain. In general, the phenomenon
was markedly observed for clindamycin followed by chlo-
ramphenicol and amoxicillin.
As suggested before and according to our results, it is
possible that MOS may induce an upregulation in the
protective enzymes of P. intermedia that may be also
linked with drug resistance mechanisms (Koutsolioutsou
et al. 2001, 2005; Albesa et al. 2004).
However, there is still much to be learned regarding
the precise mechanisms of the adaptive response to MOS
and how they might evolve inside the host. This is a pre-
liminary observation and further prospective studies are
needed to better understand how oxidative stress may
influence the drug resistance phenomenon and conse-
quently antimicrobial chemotherapy, concerning anaer-
obic infections.
Acknowledgements
The authors are grateful to Luzia Rosa Rezende and Jose
Sergio de Barros (AT ⁄ CNPq) for their technical support
and to the institutions Conselho Nacional de Desenvolvi-
mento Cientıfico e Tecnologico (CNPq), Fundacao de
Amparo a Pesquisa do Estado de Minas Gerais (FAP-
EMIG) and Coordenacao de Aperfeicoamento de Pessoal
de Nıvel Superior (CAPES) for their financial support
and facilities.
References
Albesa, I., Becerra, C.M., Battan, P.C. and Paez, L.P. (2004)
Oxidative stress involved in the antibacterial action of dif-
ferent antibiotics. Biochem Biophys Res Commun 317, 605–
609.
Aldridge, K.E., Asheraft, D., Cambre, K., Pierson, C.L., Jenkins,
S.G. and Rosenblatt, J.E. (2001) Multicenter survey of the
changing in vitro antimicrobial susceptibility of isolates of
Bacteroides fragilis group, Prevotella, Fusobacterium, Por-
phyromonas, and Peptostreptococcus species. Antimicrob
Agents Chemother 45, 1283–1284.
Behra-Miellet, J., Dubreuil, L. and Calvet, L. (2006) Evaluation
of the in vitro activity of ertapenem and nine other com-
parator agents against 337 anaerobic bacteria. Int J Anti-
microb Agents 28, 25–35.
Camarena, J.J., Nogueira, J.M., Dasi, M.A., Moreno, F., Garcia,
R., Ledesma, E., Lorca, J. and Hernandez, J. (1995) DNA
amplification fingerprinting for subtyping Neisseria gonor-
rhoeae strains. Sex Transm Dis 22, 128–136.
Diniz, C.G., Farias, L.M., Carvalho, M.A.R., Rocha, E.R. and
Smith, C.J. (2004) Differential gene expression in a Bacter-
oides fragilis metronidazole-resistant mutant. J Antimicrob
Chemother 54, 100–108.
Falagas, E.M. and Siakavellas, S. (2000) Bacteroides, Prevotella,
and Porphyromonas species: a review of antibiotic resistance
and therapeutic options. Int J Antimicrob Agents 15, 1–9.
Finegold, S.M. (1995) Overview of clinically important anaer-
obes. Clin Infect Dis 20, 205–207.
Fletcher, J.M., Nair, S.P., Ward, J.M., Henderson, B. and
Wilson, M. (2001) Analysis of the effect of changing envi-
ronmental conditions on the expression patterns of expor-
ted surface-associated proteins of the oral pathogen
Actinobacillus actinomycetemcomitans. Microb Pathog 30,
359–368.
George, K.S., Reynolds, M.A. and Falkler, W.A. Jr (1997) Arbi-
trarily primed polymerase chain reaction fingerprinting
and clonal analysis of oral Fusobacterium nucleatum iso-
lates. Oral Microbiol Immunol 12, 219–226.
Holdeman, L.V., Catto, E.P. and Moore, W.E. (1977) Anaer-
obic Laboratory Manual, 4th edn. Virginia, USA: Institute
and State University Anaerobe Laboratory.
Katsuragi, H., Ohtake, M., Kurasawa, I. and Saito, K. (2003)
Intracellular production and extracellular release of oxygen
radicals by PMNs and oxidative stress on PMNs during
phagocytosis of periodontopathic bacteria. Odontology 91,
13–18.
Kononen, E.J., Matto, M.L., Vaisanen-Tunkelrott, M.L., Frand-
sen, E.V.G., Helander, I., Asikainen, S., Finegold, S.M. and
Jousimies-Somer, H. (1998) Biochemical and genetic char-
acterization of a Prevotella intermedia ⁄ nigrescens-like
organism. Int J Syst Bacteriol 48, 39–46.
Koutsolioutsou, A., Pena-Llopis, S. and Demple, B. (2005)
Constitutive soxR mutations contribute to multiple-antibi-
otic resistance in clinical Escherichia coli isolates. Antimic-
rob Agents Chemother 49, 2746–2752.
Koutsolioutsou, A., Martins, E.A., White, D.G., Levy, S.B. and
Demple, B. (2001) A soxRS-constitutive mutation contri-
buting to antibiotic resistance in a clinical isolate of Sal-
monella enterica (Serovar Typhimurium). Antimicrob
Agents Chemother 45, 38–43.
Laemmli, U.K. (1970) Cleavage of structural proteins during
the assembly of the head of bacteriophage T4. Nature 227,
680–685.
Lynch, M. and Kuramitsu, H. (2000) Expression and role of
superoxide dismutases (SOD) in pathogenic bacteria.
Microbes Infect 2, 1245–1255.
Influence of molecular oxygen on Prevotella intermedia biology S.G. dos Santos et al.
890 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891
ª 2007 The Authors

Matto, J., Asikainen, S., Vaisanen, M.-L., Von Troil-Linden, B.,
Kononen, E., Saarela, M., Salminen, K., Finegold, S.M., et al.
(1999) b-lactamase production in Prevotella intermedia,
Prevotella nigrescens, and Prevotella pallens genotypes and
in vitro susceptibilities to selected antimicrobial agents.
Antimicrob Agents Chemother 43, 2383–2388.
Miller, M.P. (1998) TFPGA: Tools for Population Genetic Ana-
lyses for Windows. Arizona State University, USA. <http://
www.public.asu.edu/~mmille8/>. Accessed on 15 July
2001.
National Committee for Clinical Laboratory Standards
(NCCLS) (2004) Methods for antimicrobial susceptibility
testing of anaerobic bacteria. Approved Standard – 6th edn.
M11-A6. Wayne, Pennsylvania.
Nei, M. (1972) Genetic distance between populations. Am Nat
106, 283–292.
Oliveira, M.A., Caballero, O.L., Neto, E.D., Koury, M.C.,
Romanha, A.J., Carvalho, J., Hartskeerl, R.A. and Simpson,
A.J. (1995) Use of nondenaturing silver-stained polyacryla-
mide gel analysis of polymerase chain reaction amplifica-
tion products for the differential diagnosis of Leptospira
interrogans infection. Diagn Microbiol Infect Dis 22, 343–
348.
Rocha, E.R. and Smith, C.J. (1997) Regulation of Bacteroides
fragilis katB mRNA by oxidative stress and carbon limita-
tion. J Bacteriol 179, 7033–7039.
Rocha, E.R., Selby, T., Coleman, J.P. and Smith, C.J. (1996)
Oxidative stress response in anaerobe, Bacteroides fragilis: a
role for catalase in protection against hydrogen peroxide.
J Bacteriol 178, 6895–6903.
Santos, S.G., Serufo, J.C., Silva, R.A.P, Marra, B.A., Reis,
C.M.F., Hamdan, J.S., Nicoli, J.R., Carvalho, M.A.R., et al.
(2003) Microbiologic profile of intra-abdominal infections
at Belo Horizonte, Brazil. Am J Infect Control 136, 135–
142.
Santos, S.G., Carvalho, M.A.R., Serufo, J.C., Pinto-Silva, R.A.,
Albuquerque, W., Rosa, R.V., Pires, A.F.B., Hahn, R.C.,
et al. (2004) Antimicrobial susceptibility of microorgan-
isms recovered from intraabdominal infections at Belo
Horizonte, Brazil. Am J Infect Control 32, 414–416.
Shah, H.N. and Gharbia, S.E. (1992) Biochemical and chemical
studies on strains designated Prevotella intermedia and pro-
posal of a new pigmented species Prevotella nigrescens sp.
nov. Int J Syst Bacteriol 42, 542–546.
Shibata, Y., Nakano, Y., Kawada, M., Kojima, M., Fukamachi,
S., Okano, S., Matsushira, K., Abiko, Y., et al. (2005) A
searcheable database for proteomes of oral microorgan-
isms. Oral Microbial Immunol 20, 344–348.
Silva, V.L., Diniz, C.G., Cara, D.C., Santos, S.G., Nicoli, J.R.,
Carvalho, M.A.R. and Farias, L.M. (2005) Enhanced
pathogenicity of Fusobacterium nucleatum adapted to oxi-
dative stress. Microbe Pathogen 39, 131–138.
Sneath, P.H.A. and Sokal, R.R. (1973) Numerical taxonomy:
The Principles and Practice of Numerical Classification.
San Francisco: Freeman & Co.
Song, Y. (2005) PCR-based diagnostics for anaerobic infec-
tions. Anaerobe 11, 79–91.
Steers, E., Foltz, E.L. and Graves, B.S. (1959) An inocula repli-
cating apparatus for routine testing of bacterial susceptibil-
ity to antibiotics. Antibiot Chemother 9, 307–311.
Takahata, M. and Nishino, T. (1997) Antibacterial activities of
tosufloxacin against anaerobic bacteria and the electron
micrograph of its bactericidal effects. Chemotherapy 43,
153–158.
Teng, L.J., Hsueh, P.R., Tsai, J.C., Liaw, S.J. and Luh, K.T.
(2002) High incidence of cefoxitin and clindamycin resist-
ance in Taiwan. Antimicrob Agents Chemother 46, 2908–
2913.
Turton, J.A., Yallop, D., Andrews, C.M., Fagg, R., York, M.
and William, T.C. (1999) Haemotoxicity of chlorampheni-
col succinate in the CD-1 mouse and Wistar Hanover rat.
Hum Exp Toxicol 18, 566–567.
Veith, P.D., Talbo, G.H., Slakeski, N. and Reynolds, E.C.R.
(2001) Identification of a novel heterodimeric outer mem-
brane protein of Porphyromonas gingivalis by two-dimen-
sional gel electrophoresis and peptide mass fingerprinting.
Eur J Biochem 268, 4748–4757.
Westeimeier, R. and Naven, T. (2002) Proteomics in Practice.
Weinheim, Germany: Wiley-VCH Verlag-GmbH.
Zheng, M. and Storz, G. (2000) Mechanisms for sensing and
responding to oxygen deprivation. In Bacterial Stress
Responses ed.Storz, G. and Hengge-Aronis, R. pp. 47–59.
Washington, DC: American Society for Microbiology.
S.G. dos Santos et al. Influence of molecular oxygen on Prevotella intermedia biology
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 103 (2007) 882–891 891




















