
The impact of mental workload on inhibitory control subprocesses
9
The impact of mental workload on inhibitory control subprocesses Witold X. Chmielewski, Moritz Mückschel, Ann-Kathrin Stock, Christian Beste ⁎ Cognitive Neurophysiology, Department of Child and Adolescent Psychiatry, Faculty of Medicine of the TU Dresden, Germany abstract article info Article history: Accepted 26 February 2015 Available online 6 March 2015 Keywords: Inhibitory control Working memory NoGo-N2 NoGo-P3 Prefrontal cortex The inhibition of inappropriate responses is a function known to rely on prefrontal cortex (PFC) functioning. Similarly, working memory processes are known to rely on the PFC. Even though these processes are usually closely intertwined and the functional neuroanatomy underlying these processes is largely overlapping, the influence of working memory load on inhibitory control process has remained largely elusive. In the current study, we therefore examine how response inhibition processes are modulated by working memory load. For this, we systematically increased the working memory load of participants by integrating mental rotation processes in a Go/NoGo paradigm. To examine the system neurophysiology of these processes in detail, and to examine whether there are differential effects of working memory load on distinct response inhibition subpro- cesses, we applied event-related potentials (ERPs) in combination with source localization techniques. The data shows that after exceeding a certain threshold, inhibitory control processes are aggravated by working memory load. The neurophysiological data paralleled the behavioral data. However, it suggests that distinguish- able response inhibition subprocesses are differentially modulated by working memory load: Changes were evident in the NoGo-P3 amplitude but not in the NoGo-N2 amplitude. On a system level, this distinctive modu- lation of response inhibition subprocesses was related to differences in neural activity in the left inferior and middle frontal gyri. We show that inhibitory control processes are impaired when the working memory load sur- passes a certain threshold. This, however only applies to situations in which the necessity of inhibitory control processes cannot be easily detected on the basis of perceptual factors. © 2015 Elsevier Inc. All rights reserved. Introduction When attempting to master a task, different executive functions are required. One of them, namely inhibitory control (Bari and Robbins, 2013; Diamond, 2013), is crucial for blocking inappropriate responses. Others, like working memory (Baddeley, 1992; Baddeley and Hitch, 1974; Diamond, 2013), are a limiting factor, since only a certain capacity is available for the maintenance and processing of information that is used to perform response inhibition processes, for example. Even though these two factors are closely intertwined in daily life, it has remained widely elusive in how far increasing working memory load impacts response inhibition processes and in how far there are distinc- tive effects on dissociable response inhibition subprocesses that can be examined using neurophysiological (EEG) techniques. In the current study, we therefore examined these questions by systematically modu- lating working memory load and examining its effects on response inhi- bition processes. Traditionally, inhibitory control processes are often examined using Go/NoGo tasks (Beste et al., 2011; Casey et al., 1997; Nieuwenhuis et al., 2003; Ocklenburg et al., 2013; Rubia et al., 2001; Stock et al., 2014). In these paradigms, stimulus–response contingencies that trigger largely automatized reactions are established. These pre-potent response tendencies however have to be overcome in a subset of trials, thus chal- lenging participants to actively inhibit the intended reactions. From a system level perspective, a distributed prefrontal cortical network is assumed to mediate these response inhibition processes (Aron et al., 2003, 2004; Chamberlain et al., 2009; Chikazoe et al., 2007; Garavan et al., 1999; Hampshire et al., 2010; Munakata et al., 2011). In this network, PFC activity is assumed to reflect goal maintenance as it repre- sents and maintains abstract information required for successful inhibi- tion (Munakata et al., 2011). According to Braver and Barch (2002), response inhibition and working memory processes might both rely on top-down strengthening of task-relevant representations; i.e. the strengthening of the representation of the correct response, or respec- tively of task-relevant information. In this regard, response inhibition mechanisms rely upon functional neuroanatomical structures and pro- cesses that are also central for working memory functions (Barch et al., 1997; Cohen et al., 1997; Curtis and D'Esposito, 2003; D'Esposito et al., 2000). To examine working memory processes in the visual do- main, mental rotation paradigms have been established (Hyun and Luck, 2007; Kaufman, 2007; Shepard and Metzler, 1971, 1988). When applying these paradigms, it has been shown that on a functional neuro- anatomical level, parietal and frontal areas are essential for mental NeuroImage 112 (2015) 96–104 ⁎ Corresponding author at: Cognitive Neurophysiology, Department of Child and Adolescent Psychiatry, TU Dresden, Schubertstrasse 42, D-01309 Dresden, Germany. Fax: +49 351 458 7318. E-mail address: [email protected] (C. Beste). http://dx.doi.org/10.1016/j.neuroimage.2015.02.060 1053-8119/© 2015 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect NeuroImage journal homepage: www.elsevier.com/locate/ynimg
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of The impact of mental workload on inhibitory control subprocesses

NeuroImage 112 (2015) 96–104
Contents lists available at ScienceDirect
NeuroImage
j ourna l homepage: www.e lsev ie r .com/ locate /yn img
The impact of mental workload on inhibitory control subprocesses
Witold X. Chmielewski, Moritz Mückschel, Ann-Kathrin Stock, Christian Beste ⁎Cognitive Neurophysiology, Department of Child and Adolescent Psychiatry, Faculty of Medicine of the TU Dresden, Germany
⁎ Corresponding author at: Cognitive NeurophysiolAdolescent Psychiatry, TU Dresden, Schubertstrasse 42Fax: +49 351 458 7318.
E-mail address: christian.beste@uniklinikum-dresden.
http://dx.doi.org/10.1016/j.neuroimage.2015.02.0601053-8119/© 2015 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 26 February 2015Available online 6 March 2015
Keywords:Inhibitory controlWorking memoryNoGo-N2NoGo-P3Prefrontal cortex
The inhibition of inappropriate responses is a function known to rely on prefrontal cortex (PFC) functioning.Similarly, working memory processes are known to rely on the PFC. Even though these processes are usuallyclosely intertwined and the functional neuroanatomy underlying these processes is largely overlapping, theinfluence of working memory load on inhibitory control process has remained largely elusive. In the currentstudy, we therefore examine how response inhibition processes are modulated by working memory load. Forthis, we systematically increased the working memory load of participants by integrating mental rotationprocesses in a Go/NoGo paradigm. To examine the system neurophysiology of these processes in detail, and toexamine whether there are differential effects of working memory load on distinct response inhibition subpro-cesses, we applied event-related potentials (ERPs) in combination with source localization techniques.The data shows that after exceeding a certain threshold, inhibitory control processes are aggravated by workingmemory load. The neurophysiological data paralleled the behavioral data. However, it suggests that distinguish-able response inhibition subprocesses are differentially modulated by working memory load: Changes wereevident in the NoGo-P3 amplitude but not in the NoGo-N2 amplitude. On a system level, this distinctive modu-lation of response inhibition subprocesses was related to differences in neural activity in the left inferior andmiddle frontal gyri.We show that inhibitory control processes are impairedwhen theworkingmemory load sur-passes a certain threshold. This, however only applies to situations in which the necessity of inhibitory controlprocesses cannot be easily detected on the basis of perceptual factors.
© 2015 Elsevier Inc. All rights reserved.
Introduction
When attempting to master a task, different executive functions arerequired. One of them, namely inhibitory control (Bari and Robbins,2013; Diamond, 2013), is crucial for blocking inappropriate responses.Others, like working memory (Baddeley, 1992; Baddeley and Hitch,1974; Diamond, 2013), are a limiting factor, since only a certain capacityis available for the maintenance and processing of information that isused to perform response inhibition processes, for example. Eventhough these two factors are closely intertwined in daily life, it hasremained widely elusive in how far increasing working memory loadimpacts response inhibition processes and in how far there are distinc-tive effects on dissociable response inhibition subprocesses that can beexamined using neurophysiological (EEG) techniques. In the currentstudy, we therefore examined these questions by systematically modu-latingworkingmemory load and examining its effects on response inhi-bition processes.
Traditionally, inhibitory control processes are often examined usingGo/NoGo tasks (Beste et al., 2011; Casey et al., 1997; Nieuwenhuis et al.,
ogy, Department of Child and, D-01309 Dresden, Germany.
de (C. Beste).
2003; Ocklenburg et al., 2013; Rubia et al., 2001; Stock et al., 2014). Inthese paradigms, stimulus–response contingencies that trigger largelyautomatized reactions are established. These pre-potent responsetendencies however have to be overcome in a subset of trials, thus chal-lenging participants to actively inhibit the intended reactions. From asystem level perspective, a distributed prefrontal cortical network isassumed to mediate these response inhibition processes (Aron et al.,2003, 2004; Chamberlain et al., 2009; Chikazoe et al., 2007; Garavanet al., 1999; Hampshire et al., 2010; Munakata et al., 2011). In thisnetwork, PFC activity is assumed to reflect goalmaintenance as it repre-sents andmaintains abstract information required for successful inhibi-tion (Munakata et al., 2011). According to Braver and Barch (2002),response inhibition and working memory processes might both relyon top-down strengthening of task-relevant representations; i.e. thestrengthening of the representation of the correct response, or respec-tively of task-relevant information. In this regard, response inhibitionmechanisms rely upon functional neuroanatomical structures and pro-cesses that are also central for working memory functions (Barchet al., 1997; Cohen et al., 1997; Curtis and D'Esposito, 2003; D'Espositoet al., 2000). To examine working memory processes in the visual do-main, mental rotation paradigms have been established (Hyun andLuck, 2007; Kaufman, 2007; Shepard and Metzler, 1971, 1988). Whenapplying these paradigms, it has been shown that on a functional neuro-anatomical level, parietal and frontal areas are essential for mental

97W.X. Chmielewski et al. / NeuroImage 112 (2015) 96–104
rotation processes as a means to examine visual working memory(Alivisatos and Petrides, 1997; Cohen et al., 1996; Jordan et al., 2001;Kosslyn et al., 1998; Zacks, 2008). Especially in the inferior frontalgyrus (IFG), increased activity can be observed whenever workload isincreased (Jaeggi et al., 2007; Leung and Cai, 2007; Michael et al.,2001). Considering the functional anatomical overlap of mental rota-tion (visual working memory) and response inhibition processes, itappears likely that response inhibition processes are modulated bysystematically increasing visual working memory load, as inducedby mental rotation requirements. In line with that hypothesis, ithas been shown that increased task difficulty induces an aggravationof inhibitory control processes, even though the exact mechanismshave not been stated (Vaurio et al., 2009; Wodka et al., 2007). Previ-ous studies examining the interrelation of working memory and re-sponse inhibition processes examined this interrelation on a behaviorallevel (e.g. Grandjean and Collette, 2011), or used functionalmagnetic im-aging techniques (e.g. Mostoksky et al., 2003; Barber et al., 2013). Whileall these studies speak for an interrelations of working memory and re-sponse inhibition processes it is unclear what response inhibition sub-processes are mostly affected by the manipulation of working memoryload.
To examine in how far working memory load differentially affectsresponse inhibition subprocesses, we examined neurophysiologicaldata (event-related potentials, ERPs) and performed source localizationanalyses. Using ERPs, it is possible to distinguish between two responseinhibition subprocesses that are possibly differentially affected bymen-tal workload. These subprocesses are reflected by two event-related po-tentials (ERPs), namely the fronto-central negative–positive N2–P3complex (Beste et al., 2010b; Bokura et al., 2001; Falkenstein et al.,1999; Huster et al., 2013; Lavric et al., 2004; Smith et al., 2010). Whilethe NoGo-N2 is assumed to reflect pre-motor inhibition processes (i.e.the inhibition or revision of a motor plan/program before the actualmotor process, see Beste et al., 2010b; Falkenstein et al., 1999; JodoandKayama, 1992; Lavric et al., 2004), theNoGo-P3 is assumed to eitherreflect the act of inhibition itself (Beste et al., 2010b; Falkenstein et al.,1999; Huster et al., 2013; Schmajuk et al., 2006), a stimulus evaluationprocess (Friedman et al., 2001), or the inhibition outcome evaluation(Schmajuk et al., 2006). Aside from these two ERPs, we also examinedthe rotation-related negativity to illuminate the impact of inhibitorycontrol processes on mental rotation processes. The rotation-relatednegativity can be observed in mental rotation tasks and is thought toreflect the angular displacement of the stimulus material (Bajric et al.,1999; Band and Kok, 2000; Beste et al., 2010a; Heil and Rolke, 2002;Koshino et al., 2005).
We assumed inhibitory control processes and mental rotation pro-cesses to be interdependent processes. We therefore hypothesize thatinhibitory control processes are aggravated by systematically increasingworkload; i.e. that a stepwise increase of the rotation angle will lead toan increasing frequency of false alarms. Alongwith that, we hypothesizedthat the NoGo-N2 and NoGo-P3 show a decrease in their amplitudes as acorrelate of less efficient response inhibition under increasing workingmemory load. However, we expected that NoGo-P3 processes are moreaffected than NoGo-N2 processes, because processes reflected by theNoGo-P3 occur in a timewindow inwhich alsomental rotation processeslead to a workload-dependent modulation of neurophysiological re-sponses. Since the IFG is involved in response inhibition and workingmemory processes, we furthermore assumed the increase of workingmemory load on inhibitory trials to be reflected in IFG activity. Moreover,we assumed a reversed pattern to be observable in the rotation-relatednegativity. Since the employed task required mental rotation processesto determine whether or not inhibitory control processes are necessary,we expected the rotation-related negativity to increase with increasingrotation angle on inhibitory trials. In case the necessity of inhibitory con-trol processes is easily detectable, we also assumed the rotation-relatednegativity to be decreased due to a resulting cancelation of the mentalrotation process.
Material and methods
Participants
A total of n = 25 healthy young participants (17 females) between18 and 30 years old (mean age 25 ± 2.6) took part in the experiment.All participants had normal or corrected vision, were free of medicationand reported no psychiatric or neurological disorders. Before the testprotocol was conducted, written informed consent was obtained fromall participants. The study was approved by the institutional reviewboard of the Medical faculty of the TU Dresden and realized in accor-dance with the Declaration of Helsinki.
Task
For the task employed in this study, rotated numbers and charac-ters were used as stimuli in a Go/NoGo paradigm. To ensure the clas-sic characteristics of a Go/NoGo task, 30% NoGo trials requiring noresponse and 70% Go trials that required a specific response by theparticipants were utilized. The characters G and R and the numbers5 and 7 were employed as target stimuli due to their good visual dis-criminability and due to the fact that these stimuli are known to notevoke sex-dependent differences in the mental rotation process(Jansen-Osmann and Heil, 2007). By means of target rotation, varyingworking memory load was induced in this Go/NoGo paradigm. Equalproportions of each target stimulus were rotated by 30, 90, or 150°,thus evoking an increasing workload from the smallest to the largestrotation angle (Band and Kok, 2000; Koshino et al., 2005). Targetswere presented rotated clockwise and counter-clockwise in a normal(not mirrored) and a mirrored fashion (Beste et al., 2010a; Heil, 2002;Heil et al., 1998).
To thoroughly examine the impact of workload on response inhibi-tion, two blocks with alternating task complexity/difficulty were creat-ed. As mentioned above, numbers and letters were chosen as targets,thus establishing two target categories (numbers vs. letters) that caneasily be distinguished. In the less demanding block A, all letters re-quired a response, thus constituting the Go stimulus category. In con-trast, all numbers required no response, thus constituting the NoGostimulus category. Thus, NoGo trial processing was facilitated based onperceptual information (letter vs. number). In other words, as soon asthe target was identified as a number, mental rotation of the stimuluscould be canceled for these NoGo trials. On Go trials in block A, the pre-sentation of amirrored letter required a button presswith the left hand,while the presentation of a normal (notmirrored) letter required a righthand response. In contrast to block A, numbers and letters were utilizedas targets in the more challenging block B. Here, task complexity/difficulty was increased in comparison to block A, because participantscould no longer benefit from the category information (letter vs.number). In this block, responses had to be carried out to un-mirroredtargets, while mirrored numbers or letters required no responseand thus served as NoGo trials. It was therefore necessary to performmental rotation processes in order to decide whether or not to re-spond. These mental rotation processes increase working memoryload, thus allowing to examine the influence of mental workloadon response inhibition processes. As both blocks A and B required aresponse to normal, un-mirrored letters, transfer effects betweenthe two blocks needed to be ruled out. To do so, normal, un-mirroredletters required a left hand response in block B (as compared to aright-hand response in block A). Consequently, the presentation of nor-mal, un-mirrored numbers required a right hand response in block B. Inorder to provoke inhibition errors and to further amplify the effect ofworkload manipulation through time pressure, participants were gen-erally requested to respond as fast and accurately as possible in eachtrial during the whole task.
Each trial began with an 800 ms presentation of a fixation cross,which was followed by a 1100 ms target presentation. Irrespective of

98 W.X. Chmielewski et al. / NeuroImage 112 (2015) 96–104
correctness, target presentation was terminated when a response wasexecuted. The two blocks were presented in a counterbalanced orderand always preceded by a standardized instruction. Furthermore, anexercise with 60 trials was conducted before the experiment in orderto familiarize subjects with the task. In the subsequent experimentalblocks, 360 trials (252 Gos and 108 NoGos, equally distributed acrossthe different rotation angles and mirror conditions) were presented ina randomized order, thus summing up to a total of 720 trials in thetwo blocks. The experiment lasted for about 30 min.
EEG recording and analysis
EEG datawas recorded from60Ag/AgCl electrodes arranged in equi-distant positions. The sampling rate was 500 Hz and electrode imped-ances were kept below 5 kΩ. Afterwards a band-pass filter from 0.5 to20 Hz (with a slope of 48 dB/oct each) and a notch filter at 50 Hzwere applied and data was down-sampled to 256 Hz. Subsequently,an independent component analysis (ICA; infomax algorithm) wasconducted on the un-epoched data sets in order to remove recurringartifacts. ICA components revealing horizontal and vertical eye move-ments, blinks and pulse artifacts were then manually chosen to bediscarded. Afterwards, the EEG data was separately segmented for GoandNoGo trials: Go trialswere only taken into accountwhen the correctresponse was given in a time window of 1200 ms after target onset.Likewise, NoGo trials were only included in subsequent data analysiswhen no response was given in the period up to 1200ms after stimuluspresentation. Additionally, each condition (Go/NoGo) was further sub-divided based on the rotation, resulting in 3 segmentation categories(30°/90°/150°) each. The segmentationswere locked to the target stim-ulus, starting 200 ms before target presentation of the respective trialand ending 1200ms after its onset. Subsequently, an automated artifactrejection procedure was conducted for all segments. An activity below0.5 μV in a 200 ms period and a maximal value difference of 200 μV ina 100 ms interval were used as rejection criteria. In order to eliminatereference potential from the data, a current source density (CSD) trans-formation (Nunez and Pilgreen, 1991) re-referencing the data was thenapplied. The resulting CSD values are stated in μV/m2. An additional ad-vantage of the CSD-transformation is that it serves as a spatial filter(Nunez and Pilgreen, 1991), which makes it possible to identify elec-trodes that best reflect activity related to cognitive processes. After-wards, a baseline correction from −200 ms to 0 ms prior to targetonset was applied. For each category of segments, individual averageswere calculated for every participant. The different ERP components,namely P1 (at P7 & P8: 90–110 ms after target presentation onset), N1(at P7 & P8: 155–175 ms), Go/NoGo-N2 (at Cz: 246–380 ms), Go/NoGo-P3 (at Cz: 450–700 ms) and rotation-related negativity (at P7 &P8: 440–740 ms after target presentation) were identified by means ofthe scalp topography. To validate this choice of electrodes for subse-quent data analysis, the following procedure was applied (see also:Mückschel et al., 2014): The mean amplitudes of the ERP componentsin the corresponding search intervals (see above) at all electrode posi-tions were extracted. Subsequently, each electrode was comparedagainst an average of all other electrodes using Bonferroni-correctionfor multiple comparisons (critical threshold p= .0007). Based thereon,only electrodes showing significantly larger mean amplitudes than theremaining electrodes in at least one of the different experimentalconditions (negative or positive) were chosen. Essentially, the elec-trodes chosen that way were coherent with those found in the visualdata inspection. Moreover, the peaks of all ERP components, exceptfor the rotation-related negativity, were extracted to examine thepossible latency effects. Since mean amplitudes are more reliablethan peak amplitudes (Handy, 2005), subsequent analysis on ampli-tudes was however only conducted based on the mean amplitudes ofthe ERP components in the time intervals noted above for the differentERPs.
sLORETA analyses
Source localization was conducted using sLORETA (standardized lowresolution brain electromagnetic tomography; Pascual-Marqui, 2002).sLORETA gives a single linear solution to the inverse problem, based onextra-cranial measurements without a localization bias (Marco-Pallaréset al., 2005; Pascual-Marqui, 2002; Sekihara et al., 2005). sLORETA re-veals high convergence with fMRI data and it has been mathematicallyproven that sLORETA provides reliable results without localization bias(Sekihara et al., 2005). There is also evidence of EEG/fMRI and EEG/TMS studies underlining the validity of the sources estimated usingsLORETA (e.g. Sekihara et al., 2005; Dippel and Beste, 2015). ForsLORETA, the intracerebral volume is partitioned into 6239 voxels at5 mm spatial resolution. The standardized current density at eachvoxel is calculated in a realistic head model (Fuchs et al., 2002) usingthe MNI152 template (Mazziotta et al., 2001). In this study, the voxel-based sLORETA images were compared across conditions using thesLORETA-built-in voxel-wise randomization tests with 3000 permuta-tions, based on statistical nonparametric mapping (SnPM). Voxels withsignificant differences (p b .01, corrected for multiple comparisons)between contrasted conditions were located in the MNI-brain www.unizh.ch/keyinst/NewLORETA/sLORETA/sLORETA.htm.
Statistics
Behavioral data, i.e. Go reaction times, frequency of Go errors(i.e., missed Go trials or choice errors on Go trials) and frequency ofNoGo false alarms (i.e., button presses on NoGo trials) were analyzedwith a repeated-measures ANOVA. Behavioral data was analyzedusing the within-subject factors “block” and “rotation angle”. For theneurophysiological data, “condition” (Go/NoGo) and “electrode” (elec-trode sites at which the ERP effects were quantified) were included inthe model as additional within-subject factors. Greenhouse–Geissercorrection was applied wherever appropriate and all post-hoc testswere Bonferroni-corrected. As indicated by the Kolmogorov–Smirnovtests, all variables analyzed were normally distributed (all z b 0.9;p N .3).
Results
Behavioral data
Go reaction times (RTs), Go error frequency, and NoGo false alarmfrequency are shown in Fig. 1 for block A and block B.
Themean and standard error of themean (SEM) are reported for thedescriptive data. The ANOVA for Go reaction time data (Fig. 1A) re-vealed a main effect of block (F(1,24) = 12.17; p = .002; η2 = .336)showing that Go RTs were slower in block A (669.3 ms ± 16.5) thanin block B (630.5 ms ± 13.4). Furthermore, a main effect of rotationangle (F(1,24) = 134.11; p b .001; η2 = .848) was observed. Post-hocpaired t-tests revealed that RTs increased with the rotation angle (30°:610.0 ms ± 13.3; 90°: 636.0 ms ± 13.5; 150°: 703.1 ms ± 16.2)(all t N 7.04; p ≤ .001). No interaction of block × rotation angle couldbe detected (F(1,24) = 1.09; p = .331), as can be seen in Fig. 1A.
For Go errors (i.e., missed Go trials or choice errors on Go trials)(Fig. 1B), the ANOVA revealed a main effect of block (F(1,24) = 17.17;p b .001;η2= .417), showing that the frequency ofGo errorswas higherin block A (15.3 ± 1.5) than in block B (9.1 ± 1.1). Additionally, a maineffect of rotation angle (F(1,24) = 40.76; p b .001; η2 = .629) wasobserved. That is, the frequency of errors increased with increasing an-gular displacement (30°: 9.3 ± 1.1; 90°: 9.5 ± 1.1; 150°: 17.8 ± 1.6).Post-hoc paired t-tests however revealed that only the 150° conditiondiffered from the two other conditions (all t ≥ 6.51, p ≤ .001), whilethe 30° and 90° conditions did not differ from each other (t = 0.64;p = .529). Furthermore, an interaction of block × rotation angle(F(1,24) = 18.65; p b .001; η2 = .437) was detected as shown in
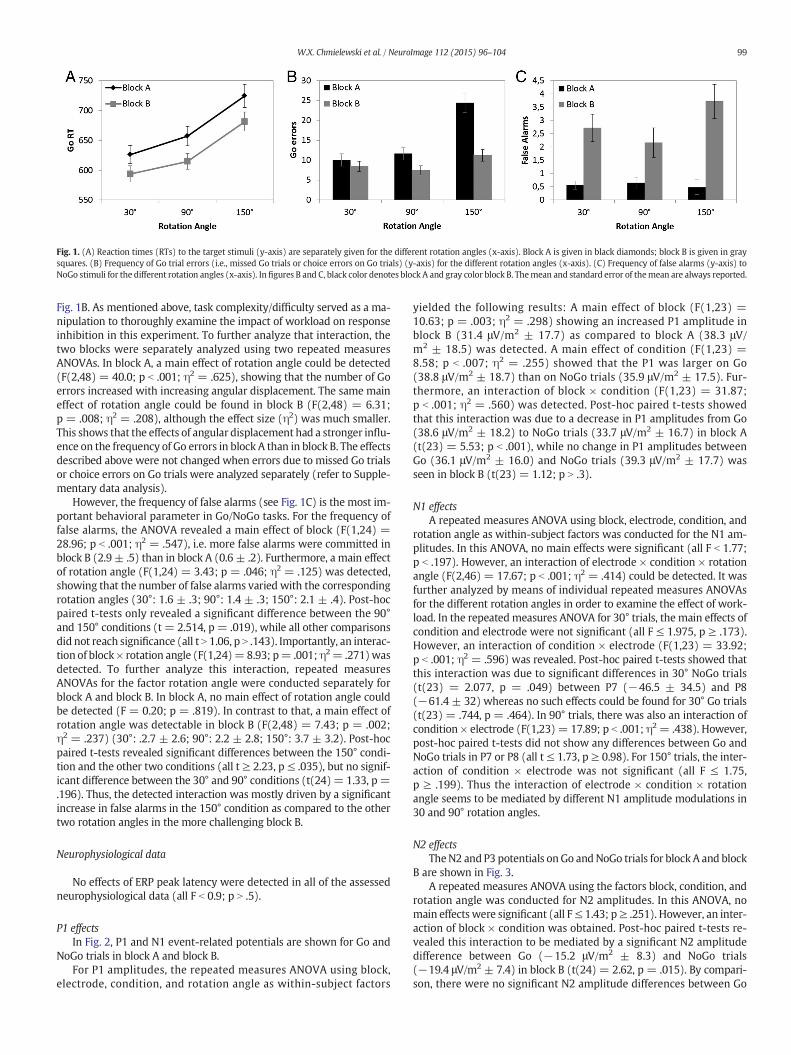
Fig. 1. (A) Reaction times (RTs) to the target stimuli (y-axis) are separately given for the different rotation angles (x-axis). Block A is given in black diamonds; block B is given in graysquares. (B) Frequency of Go trial errors (i.e., missed Go trials or choice errors on Go trials) (y-axis) for the different rotation angles (x-axis). (C) Frequency of false alarms (y-axis) toNoGo stimuli for the different rotation angles (x-axis). Infigures B and C, black color denotes block A and gray color block B. Themean and standard error of themean are always reported.
99W.X. Chmielewski et al. / NeuroImage 112 (2015) 96–104
Fig. 1B. As mentioned above, task complexity/difficulty served as a ma-nipulation to thoroughly examine the impact of workload on responseinhibition in this experiment. To further analyze that interaction, thetwo blocks were separately analyzed using two repeated measuresANOVAs. In block A, a main effect of rotation angle could be detected(F(2,48) = 40.0; p b .001; η2 = .625), showing that the number of Goerrors increased with increasing angular displacement. The same maineffect of rotation angle could be found in block B (F(2,48) = 6.31;p = .008; η2 = .208), although the effect size (η2) was much smaller.This shows that the effects of angular displacement had a stronger influ-ence on the frequency of Go errors in block A than in block B. The effectsdescribed above were not changed when errors due to missed Go trialsor choice errors on Go trials were analyzed separately (refer to Supple-mentary data analysis).
However, the frequency of false alarms (see Fig. 1C) is the most im-portant behavioral parameter in Go/NoGo tasks. For the frequency offalse alarms, the ANOVA revealed a main effect of block (F(1,24) =28.96; p b .001; η2 = .547), i.e. more false alarms were committed inblock B (2.9 ± .5) than in block A (0.6 ± .2). Furthermore, a main effectof rotation angle (F(1,24) = 3.43; p = .046; η2 = .125) was detected,showing that the number of false alarms varied with the correspondingrotation angles (30°: 1.6 ± .3; 90°: 1.4 ± .3; 150°: 2.1 ± .4). Post-hocpaired t-tests only revealed a significant difference between the 90°and 150° conditions (t = 2.514, p = .019), while all other comparisonsdid not reach significance (all t N 1.06, p N .143). Importantly, an interac-tion of block× rotation angle (F(1,24)=8.93; p= .001;η2= .271)wasdetected. To further analyze this interaction, repeated measuresANOVAs for the factor rotation angle were conducted separately forblock A and block B. In block A, no main effect of rotation angle couldbe detected (F = 0.20; p = .819). In contrast to that, a main effect ofrotation angle was detectable in block B (F(2,48) = 7.43; p = .002;η2 = .237) (30°: .2.7 ± 2.6; 90°: 2.2 ± 2.8; 150°: 3.7 ± 3.2). Post-hocpaired t-tests revealed significant differences between the 150° condi-tion and the other two conditions (all t ≥ 2.23, p ≤ .035), but no signif-icant difference between the 30° and 90° conditions (t(24)= 1.33, p =.196). Thus, the detected interaction was mostly driven by a significantincrease in false alarms in the 150° condition as compared to the othertwo rotation angles in the more challenging block B.
Neurophysiological data
No effects of ERP peak latency were detected in all of the assessedneurophysiological data (all F b 0.9; p N .5).
P1 effectsIn Fig. 2, P1 and N1 event-related potentials are shown for Go and
NoGo trials in block A and block B.For P1 amplitudes, the repeated measures ANOVA using block,
electrode, condition, and rotation angle as within-subject factors
yielded the following results: A main effect of block (F(1,23) =10.63; p = .003; η2 = .298) showing an increased P1 amplitude inblock B (31.4 μV/m2 ± 17.7) as compared to block A (38.3 μV/m2 ± 18.5) was detected. A main effect of condition (F(1,23) =8.58; p b .007; η2 = .255) showed that the P1 was larger on Go(38.8 μV/m2 ± 18.7) than on NoGo trials (35.9 μV/m2 ± 17.5). Fur-thermore, an interaction of block × condition (F(1,23) = 31.87;p b .001; η2 = .560) was detected. Post-hoc paired t-tests showedthat this interaction was due to a decrease in P1 amplitudes from Go(38.6 μV/m2 ± 18.2) to NoGo trials (33.7 μV/m2 ± 16.7) in block A(t(23) = 5.53; p b .001), while no change in P1 amplitudes betweenGo (36.1 μV/m2 ± 16.0) and NoGo trials (39.3 μV/m2 ± 17.7) wasseen in block B (t(23) = 1.12; p N .3).
N1 effectsA repeated measures ANOVA using block, electrode, condition, and
rotation angle as within-subject factors was conducted for the N1 am-plitudes. In this ANOVA, no main effects were significant (all F b 1.77;p b .197). However, an interaction of electrode × condition × rotationangle (F(2,46) = 17.67; p b .001; η2 = .414) could be detected. It wasfurther analyzed by means of individual repeated measures ANOVAsfor the different rotation angles in order to examine the effect of work-load. In the repeatedmeasures ANOVA for 30° trials, the main effects ofcondition and electrode were not significant (all F ≤ 1.975, p ≥ .173).However, an interaction of condition × electrode (F(1,23) = 33.92;p b .001; η2 = .596) was revealed. Post-hoc paired t-tests showed thatthis interaction was due to significant differences in 30° NoGo trials(t(23) = 2.077, p = .049) between P7 (−46.5 ± 34.5) and P8(−61.4 ± 32) whereas no such effects could be found for 30° Go trials(t(23) = .744, p = .464). In 90° trials, there was also an interaction ofcondition × electrode (F(1,23) = 17.89; p b .001; η2 = .438). However,post-hoc paired t-tests did not show any differences between Go andNoGo trials in P7 or P8 (all t≤ 1.73, p ≥ 0.98). For 150° trials, the inter-action of condition × electrode was not significant (all F ≤ 1.75,p ≥ .199). Thus the interaction of electrode × condition × rotationangle seems to be mediated by different N1 amplitude modulations in30 and 90° rotation angles.
N2 effectsTheN2 and P3 potentials on Go andNoGo trials for block A and block
B are shown in Fig. 3.A repeated measures ANOVA using the factors block, condition, and
rotation angle was conducted for N2 amplitudes. In this ANOVA, nomain effects were significant (all F≤ 1.43; p≥ .251). However, an inter-action of block × condition was obtained. Post-hoc paired t-tests re-vealed this interaction to be mediated by a significant N2 amplitudedifference between Go (−15.2 μV/m2 ± 8.3) and NoGo trials(−19.4 μV/m2 ± 7.4) in block B (t(24) = 2.62, p = .015). By compari-son, there were no significant N2 amplitude differences between Go
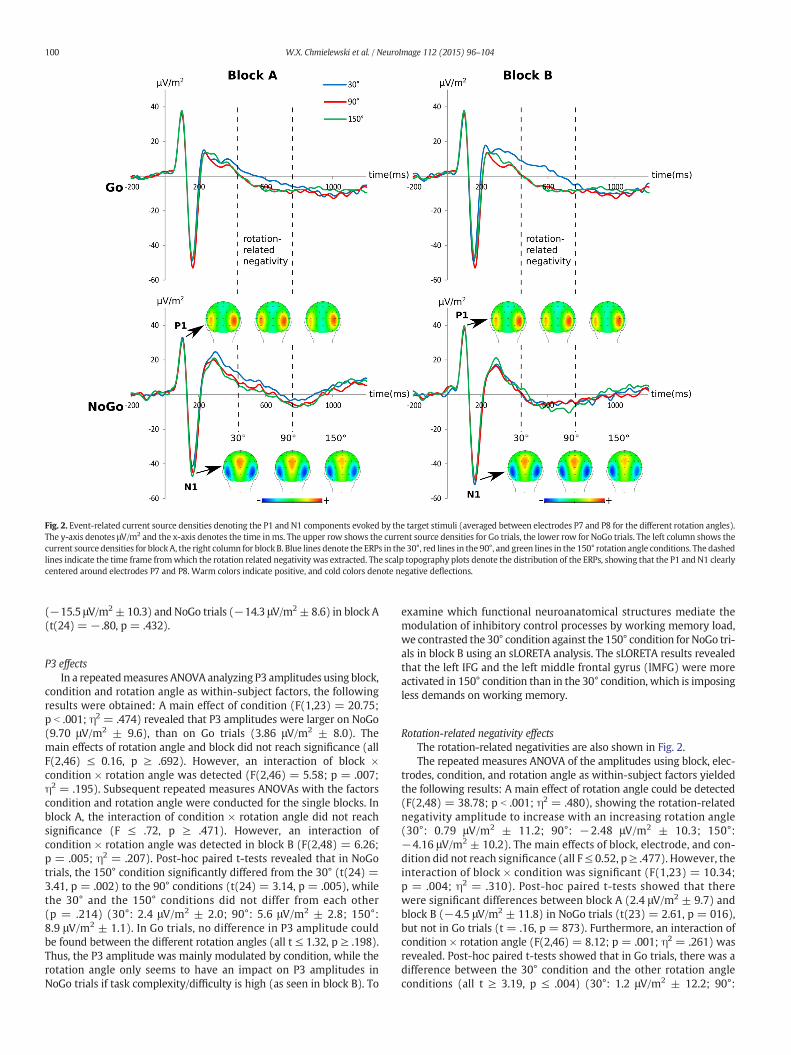
Fig. 2. Event-related current source densities denoting the P1 and N1 components evoked by the target stimuli (averaged between electrodes P7 and P8 for the different rotation angles).The y-axis denotes μV/m2 and the x-axis denotes the time inms. The upper row shows the current source densities for Go trials, the lower row for NoGo trials. The left column shows thecurrent source densities for blockA, the right column for block B. Blue lines denote the ERPs in the 30°, red lines in the 90°, and green lines in the 150° rotation angle conditions. The dashedlines indicate the time frame fromwhich the rotation related negativity was extracted. The scalp topography plots denote the distribution of the ERPs, showing that the P1 and N1 clearlycentered around electrodes P7 and P8. Warm colors indicate positive, and cold colors denote negative deflections.
100 W.X. Chmielewski et al. / NeuroImage 112 (2015) 96–104
(−15.5 μV/m2± 10.3) and NoGo trials (−14.3 μV/m2± 8.6) in block A(t(24) = − .80, p = .432).
P3 effectsIn a repeatedmeasures ANOVA analyzing P3 amplitudes using block,
condition and rotation angle as within-subject factors, the followingresults were obtained: A main effect of condition (F(1,23) = 20.75;p b .001; η2 = .474) revealed that P3 amplitudes were larger on NoGo(9.70 μV/m2 ± 9.6), than on Go trials (3.86 μV/m2 ± 8.0). Themain effects of rotation angle and block did not reach significance (allF(2,46) ≤ 0.16, p ≥ .692). However, an interaction of block ×condition × rotation angle was detected (F(2,46) = 5.58; p = .007;η2 = .195). Subsequent repeated measures ANOVAs with the factorscondition and rotation angle were conducted for the single blocks. Inblock A, the interaction of condition × rotation angle did not reachsignificance (F ≤ .72, p ≥ .471). However, an interaction ofcondition × rotation angle was detected in block B (F(2,48) = 6.26;p = .005; η2 = .207). Post-hoc paired t-tests revealed that in NoGotrials, the 150° condition significantly differed from the 30° (t(24) =3.41, p = .002) to the 90° conditions (t(24) = 3.14, p = .005), whilethe 30° and the 150° conditions did not differ from each other(p = .214) (30°: 2.4 μV/m2 ± 2.0; 90°: 5.6 μV/m2 ± 2.8; 150°:8.9 μV/m2 ± 1.1). In Go trials, no difference in P3 amplitude couldbe found between the different rotation angles (all t ≤ 1.32, p ≥ .198).Thus, the P3 amplitude was mainly modulated by condition, while therotation angle only seems to have an impact on P3 amplitudes inNoGo trials if task complexity/difficulty is high (as seen in block B). To
examine which functional neuroanatomical structures mediate themodulation of inhibitory control processes by working memory load,we contrasted the 30° condition against the 150° condition for NoGo tri-als in block B using an sLORETA analysis. The sLORETA results revealedthat the left IFG and the left middle frontal gyrus (lMFG) were moreactivated in 150° condition than in the 30° condition, which is imposingless demands on working memory.
Rotation-related negativity effectsThe rotation-related negativities are also shown in Fig. 2.The repeated measures ANOVA of the amplitudes using block, elec-
trodes, condition, and rotation angle as within-subject factors yieldedthe following results: A main effect of rotation angle could be detected(F(2,48) = 38.78; p b .001; η2 = .480), showing the rotation-relatednegativity amplitude to increase with an increasing rotation angle(30°: 0.79 μV/m2 ± 11.2; 90°: −2.48 μV/m2 ± 10.3; 150°:−4.16 μV/m2 ± 10.2). The main effects of block, electrode, and con-dition did not reach significance (all F≤ 0.52, p≥ .477). However, theinteraction of block × condition was significant (F(1,23) = 10.34;p = .004; η2 = .310). Post-hoc paired t-tests showed that therewere significant differences between block A (2.4 μV/m2 ± 9.7) andblock B (−4.5 μV/m2 ± 11.8) in NoGo trials (t(23) = 2.61, p = 016),but not in Go trials (t = .16, p = 873). Furthermore, an interaction ofcondition × rotation angle (F(2,46) = 8.12; p = .001; η2 = .261) wasrevealed. Post-hoc paired t-tests showed that in Go trials, there was adifference between the 30° condition and the other rotation angleconditions (all t ≥ 3.19, p ≤ .004) (30°: 1.2 μV/m2 ± 12.2; 90°:

Fig. 3. Event-related current source densities denoting the N2 and P3 evoked by the target stimuli at electrode Cz for the different rotation angles. The y-axis denotes μV/m2 and the x-axisdenotes the time in ms. The upper row shows the current source densities for Go trials, the lower row for NoGo trials. The left column shows the current source densities for block A, theright column for block B. Blue lines denote the ERPs in the 30°, red lines in the 90°, and green lines in the 150° rotation angle conditions. The scalp topography plots denote the distributionof the ERPs on NoGo trials, showing strong differences in an electrode cluster around electrode Cz. Warm colors indicate positive and cold colors negative deflections. The sLORETA plot(right side) denotes the sources of the difference between 30° and 150° rotated target stimuli in block B NoGo trials. The color scale denotes the critical t-values. The sources of the ERPdifferences are located in the MFG and IFG.
101W.X. Chmielewski et al. / NeuroImage 112 (2015) 96–104
−3.8 μV/m2 ± 12.1; 150°: −5.7 μV/m2 ± 11.8). The 90° and 150°conditions did not differ from each other (t b 0.9; p N .5). In NoGo trials,none of the rotation angle conditions differed from each other (allt ≤ .74, p ≥ .095) (30°: 0.4 μV/m2 ± 10.7; 90°: −1.2 μV/m2 ± 9.0;150°: −2.6 μV/m2 ± 9.0).
Discussion
In this study, we examined the impact of working memory load oninhibitory control processes. Varyingworkingmemory load was imple-mented by means of a target stimulus rotation triggering mental rota-tion processes (Beste et al., 2010a; Heil, 2002; Heil et al., 1998). Incontrast to the conceptualization by Braver and Barch (2002), inwhich only themaintenance of context information and goal representa-tions was utilized to produce a strain on workingmemory, these mentalrotation processes allowed for a systematical manipulation.
The behavioral data shows that the manipulation of inhibitory con-trol processes by means of workload augmentation was successful. In-creasing rotation angles were accompanied by increased Go trial RTsand an increased number of Go errors reflecting increased processingcosts with increasing angular displacements (Shepard and Metzler,1971). Likewise, more false alarms on NoGo trials were committed inthemore challenging block B, inwhichNoGo trials could not be as easily
identified as in block A. It may be argued, because there were fewererrors on Go trials in block B than in block A, block B is easier thanblock A and not more challenging. However, the main focus of thetask is on response inhibition and therefore on NoGo trials, makingthe rate of false alarms the most relevant parameter, since this parame-ter provides an estimate of the performance in response inhibition. Therate of false alarmswas higher in block B than in blockA. Concerning thefocus of the study on response inhibition processes, block B is moredemanding than block A. Moreover, in block A reaction times were lon-ger on Go trials than in block B, because in block A Go trial stimuli weremirrored. In block B this was not the case, since in block B only NoGo-trials weremirrored and rotated to further increase workload. Most im-portantly, the frequency of false alarms inNoGo trialswasmodulated byrotation angle, and hence by working memory load (Shepard andMetzler, 1971, 1988). It was shown that the frequency of false alarmswas highest in the 150° condition, i.e. the conditionwith the highest de-mands on mental rotation and thus working memory processes. Thisshows that increases in working memory load compromise inhibitorycontrol processes. Even more, the effect of workingmemory load on in-hibitory control processes was particularly strong when the decision ofwhether to inhibit or to execute a response was difficult (i.e., in blockB) (i.e. Vaurio et al., 2009; Wodka et al., 2007), because it could not bebased on pure perceptual stimulus features (i.e. like on the simple

102 W.X. Chmielewski et al. / NeuroImage 112 (2015) 96–104
distinction between numbers and letters in block A). Yet, a successiveincrease of false alarms with increasing rotation angle could not be ob-served, as the 30 and 90° rotation conditions did not differ from eachother. An explanation for this finding might be that people are morefamiliarized with stimuli rotated by 90°, which might facilitate the pro-cessing of such stimuli (Neely and Heath, 2010, 2011).
The neurophysiological data suggests that modulations are alreadyevident at the perceptual or attentional selection level. The P1 ampli-tude was increased in the condition with the highest rotation angle,which suggests intensified perceptual processing evoked by increasedworking memory load (Beste et al., 2010a). Furthermore, the P1 ampli-tude was larger in the more challenging block B than in the easier blockA. Also, it was larger in Go trials than in NoGo trials. The finding that theP1 was differentially modulated between Go and NoGo trials in block A,but not in block B suggests that in block B Go andNoGo trialswere prob-ably not distinguishable as quickly as in block A. This interpretation isunderlined by the increased frequency of false alarms in block B. How-ever, since no interaction of rotation angle and condition was presentfor the P1 or N1 it however seems rather unlikely that themanipulationof inhibitory control processes by means of workload superimpositionalready starts at the perceptual and attentional levels.
Rather, the neurophysiological data shows that response inhibitionsubprocesses are modulated by working memory load manipulation.In particular, the neurophysiological data suggest dissociated effects ofworking memory load on response inhibition subprocesses: In classicalGo/NoGo experiments, response inhibition processes are reflectedby the fronto-central negative–positive N2–P3 complex (Beste et al.,2010b; Bokura et al., 2001; Falkenstein et al., 1999; Lavric et al., 2004;Smith et al., 2010). The NoGo N2 is assumed to reflect pre-motor inhibi-tion of intended movements (Beste et al., 2010b; Falkenstein et al.,1999; Jodo and Kayama, 1992; Lavric et al., 2004). Interestingly, in thisexperiment no main effects of the experimental manipulations couldbe detected for the N2 amplitude. This lack of N2 modulation might beexplained by the fact that classical Go/NoGo tasks are speeded responsetasks with RTs between 250 and 400ms, where task difficulty is mainlyinduced by means of time pressure. Thus, participants are usuallyrequired to utilize pre-motor inhibition processes on intendedmotor movements in order to respond as fast and accurately as pos-sible, which is reflected in the NoGo N2 (Beste et al., 2010b, 2011;Falkenstein et al., 1999). In contrast to these paradigms, the aggrava-tion of response inhibition processes in our experiment was induced bymeans of superimposing additional working memory load (i.e. stimulusrotation). The results suggest that in the current study, the manipulationofworkingmemory load largely excludes pre-motor inhibition processes.The observed mean RTs were around 600 ms long, which surpasses theN2 latency by far. This suggests that the functional interpretation of theNoGo-N2 to reflect pre-motor inhibition processes is plausible in thecase of studies examining the inhibition of speeded motor responses insimple stimulus–response couplings, but not when response inhibitionprocesses are modulated by working memory load. This interpretationis line with other studies reporting N2 modulations when speed (incontrast to accuracy) instructions are given (Band et al., 2003; Pliszkaet al., 2000).
The results obtained for the NoGo P3 dissociate from the patternfound for the N2. The NoGo P3 is assumed to either reflect the act of in-hibition itself (Beste et al., 2010b; Falkenstein et al., 1999; Huster et al.,2013; Polich, 2007; Schmajuk et al., 2006), a stimulus evaluation pro-cess (Friedman et al., 2001), or even the evaluation of the outcome ofthe inhibition process (Schmajuk et al., 2006; Wu and Zhou, 2009). Inthis experiment, the usually increasedNoGo P3 amplitude (as comparedto the Go P3) could be observed. This supports the assumption that theNoGo P3 reflects response inhibition processes. Most importantly, andopposed to the N2, an interaction of block ×condition × rotation anglewas found. It was shown that in the more difficult block B, the NoGoP3 amplitude was decreased when workingmemory load was maximal(150°). This matches the behavioral data showing an increase in the
frequency of false alarms in this condition. This suggests that, after ex-ceeding a certain threshold, working memory load certainly impairs in-hibitory control processes. On a neuronal level, this is reflected by anattenuated NoGo P3 amplitude. Since this was only the case in themore challenging block B, this however suggests that an impairmentof inhibitory control processes only occurs when inhibitory control pro-cesses cannot be easily triggered by clearly distinguishable perceptualstimulus features (i.e. the distinction between numbers and letters inblock A). However, the finding that no effects were found for block Amay also be due to the fact that mental rotation is unnecessary for per-formance of the task in block A. Therefore, rotation angle has no effect onP3 amplitude. The finding that especially the NoGo-P3 was modulatedmay be because processes reflected by the NoGo-P3 occur in a timewin-dow in which also mental rotation processes lead to a workload-dependent modulation of neurophysiological responses. The sLORETAresults suggest that themodulations observed in block B are due to acti-vation differences in the left IFG and in the left MFG. These regions havepreviously been shown to be activated when demands on workingmemory processes are increased (Hampson et al., 2006; Jaeggi et al.,2007; Leung and Cai, 2007; McCarthy et al., 1996; Michael et al.,2001). Similarly, such lateral prefrontal regions have been shown to beimportant in response inhibition processes (Aron et al., 2003, 2004;Chamberlain et al., 2009; Chikazoe et al., 2007; Garavan et al., 1999;Hampshire et al., 2010; Munakata et al., 2011). It may be speculatedthat a co-incidence of working memory and response inhibition pro-cesses overstrains processing capacities in the left IFG and MFG andtherefore leads to modulations of response inhibition processes byworking memory load. However, in a study by Gogos et al. (2010) itwas the right and not the left MFG that was modulated during a mentalrotation task. While this may be viewed at odds with the current find-ings it needs to be noted that (i) the task applied was different betweenthe studies (i.e., there was no “inhibitory aspect” in the study by Gogoset al.), and (ii) that themethodswere different (i.e., EEG source localiza-tion in our study, fMRI in the Gogos et al. study). With respect to the lat-ter difference it needs to be stressed that themethods applied are able tospecifically examine subprocesses involved that are only a few hundredmilliseconds apart. This is not possible using fMRI and therefore the re-sults not comparable to each other.
It may be argued that in task A, mental rotation is not necessary forthe decisionwhether or not to act, which is reflected in the lack of an ef-fect of rotation on NoGo errors. In this sense, task A may be seen asbeing unrelated to the research question, as no interaction betweenmental rotation and inhibition would be expected, nor is it observed.However, mental rotation was the means to examine the role of work-ing memory load for response inhibition. In both blocks stimuli on re-sponse inhibition were rotated. In block A no rotation was necessarysince perceptual processes were sufficient to decide betweenresponding and non-responding. Yet, this needed to be shown tomake a point that mental rotation (working memory load) and notother processes were necessary in block B and are driving the effectsobtained in block B.
When assessingmental rotation processes, the rotation-related neg-ativity was observed to increase with increasing workload (i.e. rotationangle), or working memory demands, respectively (Band and Kok,2000; Beste et al., 2010a; Koshino et al., 2005). Interestingly, an interac-tion of block × condition could be observed. Here, it could be shownthat the rotation-related negativity in NoGo trials was decreased inthe less demanding block A, as compared to block B.Moreover, an inter-action of condition × rotation angle showed that the rotation-relatednegativity in the 30° condition was specifically modulated in Go trials,but not in NoGo trials. As mental rotation was irrelevant in trials ofblock A, this effect likely represents a less specific consequence of actioncancelation, rather than an influence of response control on workingmemory load. If there was such an effect, it should especially be ob-served in block B, where response inhibition and mental rotation wererequired simultaneously. The results therefore suggest that there is no

103W.X. Chmielewski et al. / NeuroImage 112 (2015) 96–104
interdependency ofworkingmemory load andmental rotation process-es. It seems that beforemost inhibitory control processes are affected byworking memory load, but not vice versa.
A limitation of the study is that the sources provided are only basedupon EEG-source localization techniques. The results on the functionalneuroanatomy are therefore not comparable in reliability to fMRI local-izations, due to the ‘inverse problem’ which cannot unambiguously besolved.
In summary, we show that inhibitory control processes are affectedby working memory processes, but unlikely vice versa. Concerning in-hibitory control processes, working memory load does not change N2-related inhibitory control processes, but P3-related inhibitory controlprocesses via the modulation of neural processes in the left IFG andleft MFG. In line with the behavioral data, we show that inhibitory con-trol processes are impairedwhen theworkingmemory load surpasses acertain threshold. This, however only applies to situations in which thenecessity of inhibitory control processes cannot be easily detected usingperceptual factors.
Conflict of interest statement
The authors declare no conflicts of interest.
Acknowledgments
This work was supported by a grant from the DeutscheForschungsgemeinschaft (DFG) BE4045/10-1 and 10-2 to C.B.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.neuroimage.2015.02.060.
References
Alivisatos, B., Petrides, M., 1997. Functional activation of the human brain during mentalrotation. Neuropsychologia 35, 111–118. http://dx.doi.org/10.1016/S0028-3932(96)00083-8.
Aron, A.R., Fletcher, P.C., Bullmore, E.T., Sahakian, B.J., Robbins, T.W., 2003. Stop-signal in-hibition disrupted by damage to right inferior frontal gyrus in humans. Nat. Neurosci.6, 115–116. http://dx.doi.org/10.1038/nn1003.
Aron, A.R., Robbins, T.W., Poldrack, R.A., 2004. Inhibition and the right inferior frontalcortex. Trends Cogn. Sci. 8, 170–177. http://dx.doi.org/10.1016/j.tics.2004.02.010.
Baddeley, A., 1992. Working memory. Science 255, 556–559. http://dx.doi.org/10.1126/science.1736359.
Baddeley, A.D., Hitch, G., 1974. Working memory. In: Bower, Gordon H. (Ed.), Psychologyof Learning and Motivation. Academic Press, pp. 47–89.
Bajric, J., Rösler, F., Heil, M., Hennighausen, E., 1999. On separating processes of eventcategorization, task preparation, and mental rotation proper in a handedness recog-nition task. Psychophysiology 36, 399–408.
Band, G.P.H., Kok, A., 2000. Age effects on response monitoring in a mental-rotation task.Biol. Psychol. 51, 201–221. http://dx.doi.org/10.1016/S0301-0511(99)00038-1.
Band, G.P.H., Ridderinkhof, K.R., van der Molen, M.W., 2003. Speed-accuracy modulationin case of conflict: the roles of activation and inhibition. Psychol. Res. 67, 266–279.http://dx.doi.org/10.1007/s00426-002-0127-0.
Barber, A.D., Caffo, B.S., Pekar, J.J., Mostofsky, S.H., 2013. Effects of working memorydemand on neural mechanisms of motor response selection and control. J. Cogn.Neurosci. 25, 1235–1248.
Barch, D.M., Braver, T.S., Nystrom, L.E., Forman, S.D., Noll, D.C., Cohen, J.D., 1997. Dissociatingworkingmemory from task difficulty in human prefrontal cortex. Neuropsychologia 35,1373–1380.
Bari, A., Robbins, T.W., 2013. Inhibition and impulsivity: behavioral and neural basis ofresponse control. Prog. Neurobiol. 108, 44–79. http://dx.doi.org/10.1016/j.pneurobio.2013.06.005.
Beste, C., Heil, M., Konrad, C., 2010a. Individual differences in ERPs duringmental rotationof characters: lateralization, and performance level. Brain Cogn. 72, 238–243.http://dx.doi.org/10.1016/j.bandc.2009.09.005.
Beste, C., Willemssen, R., Saft, C., Falkenstein, M., 2010b. Response inhibition subprocessesand dopaminergic pathways: basal ganglia disease effects. Neuropsychologia 48,366–373. http://dx.doi.org/10.1016/j.neuropsychologia.2009.09.023.
Beste, C., Ness, V., Falkenstein, M., Saft, C., 2011. On the role of fronto-striatal neural synchro-nization processes for response inhibition—evidence from ERP phase-synchronizationanalyses in pre-manifestHuntington's disease genemutation carriers. Neuropsychologia49, 3484–3493. http://dx.doi.org/10.1016/j.neuropsychologia.2011.08.024.
Bokura, H., Yamaguchi, S., Kobayashi, S., 2001. Electrophysiological correlates for responseinhibition in a Go/NoGo task. Clin. Neurophysiol. 112, 2224–2232. http://dx.doi.org/10.1016/S1388-2457(01)00691-5.
Braver, T.S., Barch, D.M., 2002. A theory of cognitive control, aging cognition, andneuromodulation. Neurosci. Biobehav. Rev. 26, 809–817. http://dx.doi.org/10.1016/S0149-7634(02)00067-2.
Casey, B.J., Trainor, R.J., Orendi, J.L., Schubert, A.B., Nystrom, L.E., Giedd, J.N., Castellanos, F.X.,Haxby, J.V., Noll, D.C., Cohen, J.D., Forman, S.D., Dahl, R.E., Rapoport, J.L., 1997. A devel-opmental functional MRI study of prefrontal activation during performance of a Go-No-Go task. J. Cogn. Neurosci. 9, 835–847. http://dx.doi.org/10.1162/jocn.1997.9.6.835.
Chamberlain, S.R., Hampshire, A., Müller, U., Rubia, K., del Campo, N., Craig, K., Regenthal,R., Suckling, J., Roiser, J.P., Grant, J.E., Bullmore, E.T., Robbins, T.W., Sahakian, B.J., 2009.Atomoxetine modulates right inferior frontal activation during inhibitory control: apharmacological functional magnetic resonance imaging study. Biol. Psychiatry 65,550–555. http://dx.doi.org/10.1016/j.biopsych.2008.10.014.
Chikazoe, J., Konishi, S., Asari, T., Jimura, K., Miyashita, Y., 2007. Activation of right inferiorfrontal gyrus during response inhibition across response modalities. J. Cogn. Neurosci.19, 69–80. http://dx.doi.org/10.1162/jocn.2007.19.1.69.
Cohen, M.S., Kosslyn, S.M., Breiter, H.C., DiGirolamo, G.J., Thompson, W.L., Anderson, A.K.,Bookheimer, S.Y., Rosen, B.R., Belliveau, J.W., 1996. Changes in cortical activity duringmental rotation. A mapping study using functional MRI. Brain 119, 89–100. http://dx.doi.org/10.1093/brain/119.1.89.
Cohen, J.D., Perlstein, W.M., Braver, T.S., Nystrom, L.E., Noll, D.C., Jonides, J., Smith, E.E.,1997. Temporal dynamics of brain activation during a working memory task. Nature386, 604–608. http://dx.doi.org/10.1038/386604a0.
Curtis, C.E., D'Esposito, M., 2003. Persistent activity in the prefrontal cortex during workingmemory. Trends Cogn. Sci. 7, 415–423. http://dx.doi.org/10.1016/S1364-6613(03)00197-9.
D'Esposito, M., Postle, B.R., Rypma, B., 2000. Prefrontal cortical contributions to workingmemory: evidence from event-related fMRI studies. Exp. Brain Res. 133, 3–11.http://dx.doi.org/10.1007/s002210000395.
Diamond, A., 2013. Executive functions. Annu. Rev. Psychol. 64, 135–168. http://dx.doi.org/10.1146/annurev-psych-113011-143750.
Dippel, G., Beste, C., 2015. A causal role of the right inferior frontal cortex in the strategies ofmulti-component behaviour. Nat. Commun. http://dx.doi.org/10.1038/ncomms7587.
Falkenstein, M., Hoormann, J., Hohnsbein, J., 1999. ERP components in Go/Nogo tasks andtheir relation to inhibition. Acta Psychol. (Amst.) 101, 267–291. http://dx.doi.org/10.1016/S0001-6918(99)00008-6.
Friedman, D., Cycowicz, Y.M., Gaeta, H., 2001. The novelty P3: an event-related brainpotential (ERP) sign of the brain's evaluation of novelty. Neurosci. Biobehav. Rev.25, 355–373. http://dx.doi.org/10.1016/S0149-7634(01)00019-7.
Fuchs, M., Kastner, J., Wagner, M., Hawes, S., Ebersole, J.S., 2002. A standardized boundaryelement method volume conductor model. Clin. Neurophysiol. Off. J. Int. Fed. Clin.Neurophysiol. 113, 702–712.
Garavan, H., Ross, T.J., Stein, E.A., 1999. Right hemispheric dominance of inhibitory con-trol: an event-related functional MRI study. Proc. Natl. Acad. Sci. 96, 8301–8306.http://dx.doi.org/10.1073/pnas.96.14.8301.
Gogos, A., Gavrilescu, M., Davison, S., Searle, K., Rossel, S.L., Bell, R., Davis, S.R., Egan, G.F.,2010. Greater superior than inferior parietal lobule activation with increasing rotationangle during mental rotation: an fMRI study. Neuropsychologia 48, 529–535.
Grandjean, J., Collette, F., 2011. Influence of response prepotency strength, general work-ing memory resources, and specific working memory load on the ability to inhibitpredominant responses: a comparison of young and elderly participants. BrainCogn. 77, 237–247.
Hampshire, A., Chamberlain, S.R., Monti, M.M., Duncan, J., Owen, A.M., 2010. The role ofthe right inferior frontal gyrus: inhibition and attentional control. NeuroImage 50,1313–1319. http://dx.doi.org/10.1016/j.neuroimage.2009.12.109.
Hampson,M., Driesen, N.R., Skudlarski, P., Gore, J.C., Constable, R.T., 2006. Brain connectivityrelated to working memory performance. J. Neurosci. 26, 13338–13343. http://dx.doi.org/10.1523/JNEUROSCI.3408-06.2006.
Handy, T.C., 2005. Event-related Potentials: A Methods Handbook. MIT Press.Heil, M., 2002. The functional significance of ERP effects during mental rotation. Psycho-
physiology 39, 535–545.Heil, M., Rolke, B., 2002. Toward a chronopsychophysiology of mental rotation. Psycho-
physiology 39, 414–422. http://dx.doi.org/10.1017/S0048577202001105.Heil, M., Rauch, M., Hennighausen, E., 1998. Response preparation begins before mental
rotation is finished: evidence from event-related brain potentials. Acta Psychol.(Amst.) 99, 217–232. http://dx.doi.org/10.1016/S0001-6918(98)00012-2.
Huster, R.J., Enriquez-Geppert, S., Lavallee, C.F., Falkenstein, M., Herrmann, C.S., 2013.Electroencephalography of response inhibition tasks: functional networks and cogni-tive contributions. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 87, 217–233.http://dx.doi.org/10.1016/j.ijpsycho.2012.08.001.
Hyun, J.-S., Luck, S.J., 2007. Visual working memory as the substrate for mental rotation.Psychon. Bull. Rev. 14, 154–158. http://dx.doi.org/10.3758/BF03194043.
Jaeggi, S.M., Buschkuehl, M., Etienne, A., Ozdoba, C., Perrig, W.J., Nirkko, A.C., 2007. Onhow high performers keep cool brains in situations of cognitive overload. Cogn.Affect. Behav. Neurosci. 7, 75–89. http://dx.doi.org/10.3758/CABN.7.2.75.
Jansen-Osmann, P., Heil, M., 2007. Suitable stimuli to obtain (no) gender differences inthe speed of cognitive processes involved in mental rotation. Brain Cogn. 64,217–227. http://dx.doi.org/10.1016/j.bandc.2007.03.002.
Jodo, E., Kayama, Y., 1992. Relation of a negative ERP component to response inhibition ina Go/No-go task. Electroencephalogr. Clin. Neurophysiol. 82, 477–482. http://dx.doi.org/10.1016/0013-4694(92)90054-L.
Jordan, K., Heinze, H.-J., Lutz, K., Kanowski, M., Jäncke, L., 2001. Cortical activations duringthe mental rotation of different visual objects. NeuroImage 13, 143–152. http://dx.doi.org/10.1006/nimg.2000.0677.

104 W.X. Chmielewski et al. / NeuroImage 112 (2015) 96–104
Kaufman, S.B., 2007. Sex differences in mental rotation and spatial visualization ability:can they be accounted for by differences in working memory capacity? Intelligence35, 211–223. http://dx.doi.org/10.1016/j.intell.2006.07.009.
Koshino, H., Carpenter, P.A., Keller, T.A., Just, M.A., 2005. Interactions between the dorsaland the ventral pathways in mental rotation: an fMRI study. Cogn. Affect. Behav.Neurosci. 5, 54–66. http://dx.doi.org/10.3758/CABN.5.1.54.
Kosslyn, S.M., Digirolamo, G.J., Thompson, W.L., Alpert, N.M., 1998. Mental rotation of ob-jects versus hands: neural mechanisms revealed by positron emission tomography.Psychophysiology 35, 151–161 (doi:null).
Lavric, A., Pizzagalli, D.A., Forstmeier, S., 2004. When “go” and “nogo” are equally fre-quent: ERP components and cortical tomography. Eur. J. Neurosci. 20, 2483–2488.http://dx.doi.org/10.1111/j.1460-9568.2004.03683.x.
Leung, H.-C., Cai, W., 2007. Common and differential ventrolateral prefrontal activity dur-ing inhibition of hand and eye movements. J. Neurosci. 27, 9893–9900. http://dx.doi.org/10.1523/JNEUROSCI. 2837-07.2007.
Marco-Pallarés, J., Grau, C., Ruffini, G., 2005. Combined ICA-LORETA analysis of mismatchnegativity. NeuroImage 25, 471–477. http://dx.doi.org/10.1016/j.neuroimage.2004.11.028.
Mazziotta, J., Toga, A., Evans, A., Fox, P., Lancaster, J., Zilles, K., Woods, R., Paus, T., Simpson,G., Pike, B., Holmes, C., Collins, L., Thompson, P., MacDonald, D., Iacoboni, M.,Schormann, T., Amunts, K., Palomero-Gallagher, N., Geyer, S., Parsons, L., Narr, K.,Kabani, N., Le Goualher, G., Boomsma, D., Cannon, T., Kawashima, R., Mazoyer, B.,2001. A probabilistic atlas and reference system for the human brain: InternationalConsortium for Brain Mapping (ICBM). Philos. Trans. R. Soc. Lond. Ser. B 356,1293–1322. http://dx.doi.org/10.1098/rstb.2001.0915.
McCarthy, G., Puce, A., Constable, T., Krystal, J.H., Gore, J.C., Goldman-Rakic, P., 1996. Acti-vation of human prefrontal cortex during spatial and nonspatial working memorytasks measured by functional MRI. Cereb. Cortex 6, 600–611. http://dx.doi.org/10.1093/cercor/6.4.600.
Michael, E.B., Keller, T.A., Carpenter, P.A., Just, M.A., 2001. fMRI investigation of sentencecomprehension by eye and by ear: modality fingerprints on cognitive processes.Hum. Brain Mapp. 13, 239–252. http://dx.doi.org/10.1002/hbm.1036.
Mostoksky, S.H., Schafer, J.G., Abrams, M.T., Goldberg, M.C., Flower, A.A., Boyce, A.,Courtney, S.M., Calhoun, V.D., Kraut, M.A., Denckla, M.B., Pekar, J.J., 2003. fMRI evi-dence that the neural basis of response inhibition is task-dependent. Brain Res.Cogn. Brain Res. 17, 419–430.
Mückschel, M., Stock, A.-K., Beste, C., 2014. Psychophysiological mechanisms of interindi-vidual differences in goal activation modes during action cascading. Cereb. Cortex 24,2120–2129. http://dx.doi.org/10.1093/cercor/bht066.
Munakata, Y., Herd, S.A., Chatham, C.H., Depue, B.E., Banich, M.T., O'Reilly, R.C., 2011. Aunified framework for inhibitory control. Trends Cogn. Sci. 15, 453–459. http://dx.doi.org/10.1016/j.tics.2011.07.011.
Neely, K.A., Heath, M., 2010. Visuomotor mental rotation: reaction time is determined bythe complexity of the sensorimotor transformations mediating the response. BrainRes. 1366, 129–140. http://dx.doi.org/10.1016/j.brainres.2010.09.096.
Neely, K.A., Heath, M., 2011. The visuomotor mental rotation task: visuomotor transfor-mation times are reduced for small and perceptually familiar angles. J. Mot. Behav.43, 393–402. http://dx.doi.org/10.1080/00222895.2011.618850.
Nieuwenhuis, S., Yeung, N., van den Wildenberg, W., Ridderinkhof, K.R., 2003. Electro-physiological correlates of anterior cingulate function in a go/no-go task: effects of re-sponse conflict and trial type frequency. Cogn. Affect. Behav. Neurosci. 3, 17–26.http://dx.doi.org/10.3758/CABN.3.1.17.
Nunez, P.L., Pilgreen, K.L., 1991. The spline-Laplacian in clinical neurophysiology: amethod toimprove EEG spatial resolution. J. Clin. Neurophysiol. Off. Publ. Am. Electroencephalogr.Soc. 8, 397–413.
Ocklenburg, S., Ness, V., Güntürkün, O., Suchan, B., Beste, C., 2013. Response inhibition ismodulated by functional cerebral asymmetries for facial expression perception.Front. Psychol. 4, 879. http://dx.doi.org/10.3389/fpsyg.2013.00879.
Pascual-Marqui, R.D., 2002. Standardized low-resolution brain electromagnetic tomogra-phy (sLORETA): technical details. Methods Find. Exp. Clin. Pharmacol. 24 (Suppl. D),5–12.
Pliszka, S.R., Liotti, M., Woldorff, M.G., 2000. Inhibitory control in children with attention-deficit/hyperactivity disorder: event-related potentials identify the processing com-ponent and timing of an impaired right-frontal response-inhibition mechanism.Biol. Psychiatry 48, 238–246. http://dx.doi.org/10.1016/S0006-3223(00)00890-8.
Polich, J., 2007. Updating P300: an integrative theory of P3a and P3b. Clin. Neurophysiol.118, 2128–2148.
Rubia, K., Russell, T., Overmeyer, S., Brammer, M.J., Bullmore, E.T., Sharma, T., Simmons, A.,Williams, S.C.R., Giampietro, V., Andrew, C.M., Taylor, E., 2001. Mappingmotor inhibi-tion: conjunctive brain activations across different versions of Go/No-Go and stoptasks. NeuroImage 13, 250–261. http://dx.doi.org/10.1006/nimg.2000.0685.
Schmajuk, M., Liotti, M., Busse, L., Woldorff, M.G., 2006. Electrophysiological activityunderlying inhibitory control processes in normal adults. Neuropsychologia 44,384–395. http://dx.doi.org/10.1016/j.neuropsychologia.2005.06.005.
Sekihara, K., Sahani, M., Nagarajan, S.S., 2005. Localization bias and spatial resolution ofadaptive and non-adaptive spatial filters for MEG source reconstruction. NeuroImage25, 1056–1067. http://dx.doi.org/10.1016/j.neuroimage.2004.11.051.
Shepard, R.N., Metzler, J., 1971. Mental rotation of three-dimensional objects. Science 171,701–703. http://dx.doi.org/10.1126/science.171.3972.701.
Shepard, S., Metzler, D., 1988. Mental rotation: effects of dimensionality of objects andtype of task. J. Exp. Psychol. Hum. Percept. Perform. 14, 3–11.
Smith, J.L., Smith, E.A., Provost, A.L., Heathcote, A., 2010. Sequence effects support the con-flict theory of N2 and P3 in the Go/NoGo task. Int. J. Psychophysiol. 75, 217–226.http://dx.doi.org/10.1016/j.ijpsycho.2009.11.002.
Stock, A.-K., Schulz, T., Lenhardt, M., Blaszkewicz, M., Beste, C., 2014. High-dose alcohol in-toxication differentially modulates cognitive subprocesses involved in response inhi-bition. Addict. Biol. http://dx.doi.org/10.1111/adb.12170.
Vaurio, R.G., Simmonds, D.J., Mostofsky, S.H., 2009. Increased intra-individual reactiontime variability in attention-deficit/hyperactivity disorder across response inhibitiontasks with different cognitive demands. Neuropsychologia 47, 2389–2396. http://dx.doi.org/10.1016/j.neuropsychologia.2009.01.022.
Wodka, E.L., Mahone, E.M., Blankner, J.G., Larson, J.C.G., Fotedar, S., Denckla, M.B.,Mostofsky, S.H., 2007. Evidence that response inhibition is a primary deficit inADHD. J. Clin. Exp. Neuropsychol. 29, 345–356. http://dx.doi.org/10.1080/13803390600678046.
Wu, Y., Zhou, X., 2009. The P300 and reward valence, magnitude, and expectancy in out-come evaluation. Brain Res. 1286, 114–122. http://dx.doi.org/10.1016/j.brainres.2009.06.032.
Zacks, J.M., 2008. Neuroimaging studies of mental rotation: a meta-analysis and review.J. Cogn. Neurosci. 20, 1–19. http://dx.doi.org/10.1162/jocn.2008.20013.




















