
The Gut Microbiota Appears to Compensate for Seasonal Diet Variation in the Wild Black Howler Monkey...
10
HOST MICROBE INTERACTIONS The Gut Microbiota Appears to Compensate for Seasonal Diet Variation in the Wild Black Howler Monkey (Alouatta pigra) Katherine R. Amato & Steven R. Leigh & Angela Kent & Roderick I. Mackie & Carl J. Yeoman & Rebecca M. Stumpf & Brenda A. Wilson & Karen E. Nelson & Bryan A. White & Paul A. Garber Received: 9 May 2014 /Accepted: 3 December 2014 # Springer Science+Business Media New York 2014 Abstract For most mammals, including nonhuman primates, diet composition varies temporally in response to differences in food availability. Because diet influences gut microbiota composition, it is likely that the gut mi- crobiota of wild mammals varies in response to seasonal changes in feeding patterns. Such variation may affect host digestive efficiency and, ultimately, host nutrition. In this study, we investigate the temporal variation in diet and gut microbiota composition and function in two groups (N =13 individuals) of wild Mexican black howler monkeys (Alouatta pigra) over a 10-month period in Palenque National Park, Mexico. Temporal changes in the relative abundances of individual bacterial taxa were strongly correlated with changes in host diet. For exam- ple, the relative abundance of Ruminococcaceae was highest during periods when energy intake was lowest, and the relative abundance of Butyricicoccus was highest when young leaves and unripe fruit accounted for 68 % of the diet. Additionally, the howlers exhibited increased microbial production of energy during periods of reduced energy intake from food sources. Because we observed few changes in howler activity and ranging patterns dur- ing the course of our study, we propose that shifts in the composition and activity of the gut microbiota pro- vided additional energy and nutrients to compensate for changes in diet. Energy and nutrient production by the gut microbiota appears to provide an effective buffer against seasonal fluctuations in energy and nutrient intake for these primates and is likely to have a similar function in other mammal species. Keywords Alouatta . Gut microbiome . Nutrition . Seasonality Electronic supplementary material The online version of this article (doi:10.1007/s00248-014-0554-7) contains supplementary material, which is available to authorized users. K. R. Amato Program in Ecology, Evolution, and Conservation Biology, University of Illinois, Urbana, IL 61801, USA K. R. Amato (*) : S. R. Leigh Department of Anthropology, University of Colorado, Boulder, CO 80309, USA e-mail: [email protected] A. Kent Department of Natural Resources and Environmental Sciences, University of Illinois, Urbana, IL 61801, USA R. I. Mackie : B. A. White Department of Animal Sciences, University of Illinois, Urbana, IL 61801, USA R. I. Mackie : R. M. Stumpf : B. A. Wilson : B. A. White Institute for Genomic Biology, University of Illinois, Urbana, IL 61801, USA C. J. Yeoman Department of Animal and Range Sciences, Montana State University, Bozeman, MT 59717, USA R. M. Stumpf : P. A. Garber Department of Anthropology, University of Illinois, Urbana, IL 80301, USA B. A. Wilson Department of Microbiology, University of Illinois, Urbana, IL 61801, USA K. E. Nelson The J. Craig Venter Institute, Rockville, MD 20850, USA Microb Ecol DOI 10.1007/s00248-014-0554-7
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The Gut Microbiota Appears to Compensate for Seasonal Diet Variation in the Wild Black Howler Monkey...

HOST MICROBE INTERACTIONS
The Gut Microbiota Appears to Compensate for Seasonal DietVariation in the Wild Black Howler Monkey (Alouatta pigra)
Katherine R. Amato & Steven R. Leigh & Angela Kent & Roderick I. Mackie &
Carl J. Yeoman & Rebecca M. Stumpf & Brenda A. Wilson & Karen E. Nelson &
Bryan A. White & Paul A. Garber
Received: 9 May 2014 /Accepted: 3 December 2014# Springer Science+Business Media New York 2014
Abstract For most mammals, including nonhumanprimates, diet composition varies temporally in responseto differences in food availability. Because diet influencesgut microbiota composition, it is likely that the gut mi-crobiota of wild mammals varies in response to seasonalchanges in feeding patterns. Such variation may affecthost digestive efficiency and, ultimately, host nutrition.In this study, we investigate the temporal variation in dietand gut microbiota composition and function in twogroups (N=13 individuals) of wild Mexican black howlermonkeys (Alouatta pigra) over a 10-month period inPalenque National Park, Mexico. Temporal changes inthe relative abundances of individual bacterial taxa werestrongly correlated with changes in host diet. For exam-ple, the relative abundance of Ruminococcaceae washighest during periods when energy intake was lowest,and the relative abundance of Butyricicoccus was highest
when young leaves and unripe fruit accounted for 68 % ofthe diet. Additionally, the howlers exhibited increasedmicrobial production of energy during periods of reducedenergy intake from food sources. Because we observedfew changes in howler activity and ranging patterns dur-ing the course of our study, we propose that shifts inthe composition and activity of the gut microbiota pro-vided additional energy and nutrients to compensate forchanges in diet. Energy and nutrient production by the gutmicrobiota appears to provide an effective buffer againstseasonal fluctuations in energy and nutrient intake forthese primates and is likely to have a similar function inother mammal species.
Keywords Alouatta . Gut microbiome . Nutrition .
Seasonality
Electronic supplementary material The online version of this article(doi:10.1007/s00248-014-0554-7) contains supplementary material,which is available to authorized users.
K. R. AmatoProgram in Ecology, Evolution, and Conservation Biology,University of Illinois, Urbana, IL 61801, USA
K. R. Amato (*) : S. R. LeighDepartment of Anthropology, University of Colorado, Boulder,CO 80309, USAe-mail: [email protected]
A. KentDepartment of Natural Resources and Environmental Sciences,University of Illinois, Urbana, IL 61801, USA
R. I. Mackie : B. A. WhiteDepartment of Animal Sciences, University of Illinois, Urbana,IL 61801, USA
R. I. Mackie : R. M. Stumpf : B. A. Wilson :B. A. WhiteInstitute for Genomic Biology, University of Illinois, Urbana,IL 61801, USA
C. J. YeomanDepartment of Animal and Range Sciences, Montana StateUniversity, Bozeman, MT 59717, USA
R. M. Stumpf : P. A. GarberDepartment of Anthropology, University of Illinois, Urbana,IL 80301, USA
B. A. WilsonDepartment of Microbiology, University of Illinois, Urbana,IL 61801, USA
K. E. NelsonThe J. Craig Venter Institute, Rockville, MD 20850, USA
Microb EcolDOI 10.1007/s00248-014-0554-7

Introduction
All mammals rely upon mutualistic gut microbial communi-ties to provide them with energy via the formation of short-chain fatty acids (SCFAs) from otherwise indigestible dietarymaterial such as cellulose and hemicellulose [1]. Individualsgenerally acquire microbes from their mother, conspecifics,and food sources [2, 3], but even after the gut microbiota isestablished, its composition shifts over time in response toselective pressures imposed by diet, health, and physiology[4–8]. Such changes in the gut microbiota can influence hostdigestive efficiency [9–15], and variation in host diet can alsolead to functional differences in the gut microbiota by alteringthe metabolic pathways utilized [16–18]
For most mammals, including nonhuman primates, theavailability, distribution, and nutritional content of food re-sources vary temporally and spatially in response to differ-ences in microhabitat, rainfall, plant species phenology, andanthropogenic influences [e.g., 19–21]. As a result, diets andpatterns of nutrient intake can change dramatically acrossseasons and habitats [e.g., 22–26]. Given that host diet influ-ences the relative abundances and metabolic functions ofmicrobial taxa in the gut, the composition and function ofthe gut microbiota in wild mammals are likely to vary acrossseasons and habitats in direct response to these dietarychanges.
Recent research with howler monkeys (Alouatta sp.) pro-vides preliminary evidence that the gut microbial communityis impacted by changes in diet across habitats [27]. However,no study to date has examined the impact of temporal changesin diet on the gut microbiota of wild mammals [but see 28].These data are of critical importance for understanding hostnutrition and health. If an individual’s gut microbiota can shiftover the course of days, weeks, or months in response tochanges in diet [28–30], associated shifts in host digestiveefficiency may make meeting nutritional demands less chal-lenging for animals during periods of resource limitation[31–33]. If, however, the gut microbiota fails to respond tothese short-term changes in the diet or responds in a way thatnegatively impacts digestive efficiency, the effects of limitedfood and nutrient availability on host health may result inreduced birth rates, reduced juvenile growth rates, increasedmortality, and increased frequency of nutrient deficiencies anddisease [34–37].
Here, we investigate temporal variation in the diet and gutmicrobiota of wild Mexican black howler monkeys (Alouattapigra). Howler monkeys respond to seasonal decreases in theavailability of ripe fruit by exploiting hard-to-digest foodssuch as flowers, leaves, and unripe fruits [38]. Whereas ripefruit is generally considered an easy-to-digest food that is lowin structural carbohydrates, high in energy, and low in proteincontent, unripe fruits, leaves, and flowers are consideredharder-to-digest foods that are higher in protein content but
also higher in structural carbohydrates and toxins and lower inenergy [39]. Within each of these categories, there also isconsiderable species-specific variation [40]. Therefore, weexpected that changes in the consumption of both plant partsand plant species would likely result in howlers consumingnutritionally distinct diets across different periods of the year.
To compensate for changes in energy and nutrient intakeacross time, howler monkeys are reported to employ anenergy-minimizing behavioral strategy [41]. They rest for62–80 % of daylight hours and, compared to other atelines(e.g., spider monkeys, woolly monkeys, and muriquis), utilizesmall day ranges and home ranges [38]. Howlers also exercisefeeding selectivity by preferentially consuming young leavesinstead of mature leaves to increase energy and nutrient intakeand by targeting a small number of tree species each day toreduce fiber and toxin intake [42, 43] (but see [44, 45]).Additionally, although they do not possess a specialized fore-gut or cecum as is found in other primate taxa such ascolobines and indriids, howlermonkeys have increased cecumand colon volumes [46] and are characterized by relativelylong food retention times compared to other New Worldprimates of similar body mass [47–49], suggesting that theyrely heavily on microbial fermentation in the hindgut to pro-cess the fiber and anti-nutritional factors found in some leaves[50].
In this study, we utilized behavioral data describing dietand activity as well as microbial analyses of fecal samplescollected during different seasons to test three hypothesesregarding temporal patterns in wild howler monkey dietarycomposition and gut microbial community composition andfunction. (1) Temporal changes in howler monkey diet areassociated with temporal changes in the composition of thehowler monkey gut microbiota. (2) Changes in howler mon-key diet and gut microbial community composition are asso-ciated with changes in gut microbial community function,measured as SCFA and ammonia production. (3) Changes ingut microbial community function aid howlers in meetingnutritional requirements and maintaining activity levels dur-ing all periods of the year.
Methods
We collected behavioral data and fecal samples from all blackhowlers in two wild, habituated neighboring social groups(Balam and Motiepa, N=6 adult males, 4 adult females, 6juveniles) in Palenque National Park, Mexico across three 10-week blocks from September 2010 to June 2011. These sam-pling blocks corresponded loosely with changes in rainfall[51] and previously documented shifts in black howler dietat Palenque. The first block (Wet, Fruit-Dominated Period(WFD), September–November 2010) was generally associat-ed with heavier rainfall and a higher proportion of ripe fruit in
K. R. Amato et al.

the howler diet [51]. The second block (Dry, Leaf-DominatedPeriod (DLD), January–March 2011) was associated with alower proportion of fruit in the diet and a higher proportion ofyoung leaves, and the last block (Dry, Fruit,-Dominated Peri-od (DFD), April–June 2011) was associated with less rainfalland a higher proportion of ripe fruit [51].
We collected 20-min instantaneous focal individual sam-ples 5 days per week between sunrise and 5 pm (park closingtime) each day to describe howler activity and feeding pat-terns, and we utilized a handheld global positioning system torecord the location of the black howler monkey study groupposition every 30 min during data collection. Collection andanalysis of activity and feeding data are described in moredetail in the supporting information as well as in Amato andGarber [52].
We collected fecal samples every 2 weeks from each groupmember for microbiota composition analysis as well as mea-surements of volatile fatty acid (VFA) and ammonia content.Measuring fecal VFA and ammonia content provides a proxyfor VFA and ammonia production since higher production ofVFAs and ammonia normally leads to higher excretion of bothas waste [53].
To describe microbiota composition, we extracted totalDNA from fecal samples using the MOBio UltraClean SoilKit (MoBio Laboratories Inc., Carlsbad, CA, USA). Weassessed microbial community composition using automatedribosomal intergenic spacer analysis (ARISA) [54], and wegenerated microbial taxonomic data in a subset of samples(N=8 individuals at 15 time points) using pyrosequencing ofthe V1–V3 region of the 16S ribosomal RNA gene [27].Details regarding methods and bioinformatics are describedin the supporting information.
We measured fecal VFA content in millimoles per gram offeces and as a proportion of total VFAs using gas chromatog-raphy [55] after neutralizing samples preserved in 1 M NaOHwith phosphoric acid and removing particulate matter withcentrifugation. We measured ammonia content in millimolesper gram of feces using spectrophotometry according toChaney and Marbach [56]. We standardized data for bothVFA and ammonia content according to sample mass andadjusted for dilution.
We used permutational multivariate analysis of variance(PERMANOVA) to test for the effects of season on gutmicrobiota composition (ARISA), fecal VFA content, anddiet. We visualized temporal patterns in gut microbiota com-position, VFA content, and diet using partial correspondenceanalysis. Univariate statistical procedures to assess seasonalpatterns in total grams of food consumed, intake of individualdiet components, relative abundances of bacteria taxa, totalfecal VFA content, total fecal ammonia content, and concen-trations of individual VFAs are described in the supportinginformation. We used nonparametric Mantel tests to compareoverall patterns in gut microbiota composition (ARISA), fecal
VFA content, and host diet. A series of Spearman rank corre-lations were used to test for relationships between the relativeabundances of individual bacterial taxa and all measured dietdescriptors. We adjusted p values for repeated tests using asequential Bonferroni correction with the initial p=0.05 [57,58]. We performed all analyses with R with the exception ofnonparametric Mantel tests, which were performed usingPRIMER 6 for Windows (PRIMER-E, Plymouth, UK).
Results
Temporal Diet Patterns
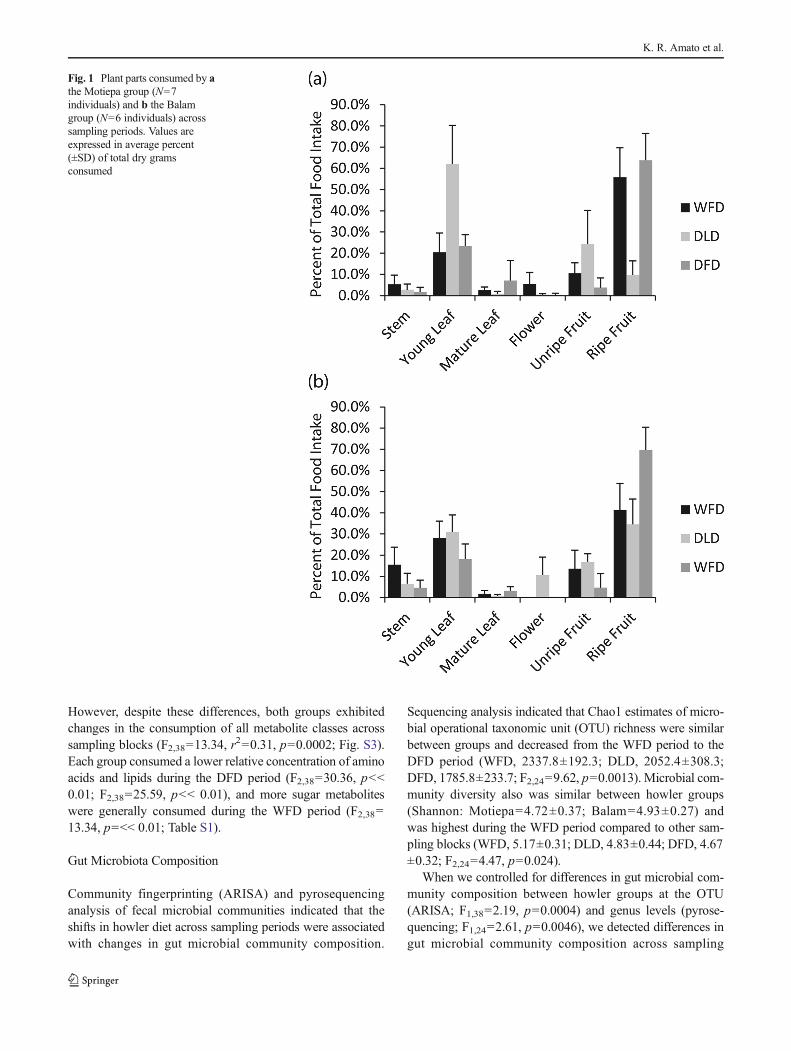
Black howler monkey diet differed markedly across samplingperiods. The howlers consumed more grams of food in theWFD period and the DLD period compared to the DFD period(F2,38=8.59, p=0.001; Table S1). Although diet compositiondiffered between groups (F2,38=4.73, p=0.004), both howlergroups consumed a higher proportion of young leaves andunripe fruits during the DLD period and more ripe fruitsduring the DFD period (F2, 38=22.06, r
2=0.16, p=0.0002;Fig. 1, S1). Dietary diversity in terms of the plant speciesconsumed was similar between groups and marginally lowerduring the DLD period compared to the WFD and DFDperiods (Table S2). Although both howler groups used manyof the same top ten plant species, the relative importance ofindividual plant species consumed differed between howlergroups (F2,38=2.69, p=0.003; Table S2). However, in bothgroups, the plant families Moraceae and Fabaceae dominatedthe diet, and the total array of plant parts and species con-sumed differed across sampling periods (F2, 38=14.99, r
2=0.31, p=0.0002, Fig. S2).
Although the Motiepa group consumed less protein thanthe Balam group (F1,38=4.32, p=0.04) during all samplingblocks, and the Balam group consumed more lipids than theMotiepa group in the DLD period (F2,38=8.21, p<0.01;Table S1), there were parallel changes in nutrient intake acrosstime in both groups. Both groups consumed fewer kilocalories(F2,38=10.27, p=0.0003), grams of protein (F2,38=17.42, p<<0.01), grams of total nonstructural carbohydrates (F2,38=17.7,p<< 0.01), and grams of neutral detergent fiber (NDF, F2,38=17.06, p<< 0.01) during the DFD period compared to theWFD period (Table S1).
When we described diet in terms of metabolites, in additionto pantothenic acid, or vitamin B5, the Motiepa group con-sumed higher average daily relative concentrations of totallipid metabolites per metabolic body weight (F2,38=7.23, p=0.011; Table S1). Also, during the WFD period, members ofthe Motiepa group consumed a higher relative concentrationof sugar metabolites than those in the Balam group, while inthe DFD period, Balam howlers consumed a higher concen-tration of sugar metabolites (F2,38=4.8, p=0.014; Table S1).
Temporal variation in the wild howler monkey gut microbiota
However, despite these differences, both groups exhibitedchanges in the consumption of all metabolite classes acrosssampling blocks (F2,38=13.34, r
2=0.31, p=0.0002; Fig. S3).Each group consumed a lower relative concentration of aminoacids and lipids during the DFD period (F2,38=30.36, p<<0.01; F2,38=25.59, p<< 0.01), and more sugar metaboliteswere generally consumed during the WFD period (F2,38=13.34, p=<< 0.01; Table S1).
Gut Microbiota Composition
Community fingerprinting (ARISA) and pyrosequencinganalysis of fecal microbial communities indicated that theshifts in howler diet across sampling periods were associatedwith changes in gut microbial community composition.
Sequencing analysis indicated that Chao1 estimates of micro-bial operational taxonomic unit (OTU) richness were similarbetween groups and decreased from the WFD period to theDFD period (WFD, 2337.8±192.3; DLD, 2052.4±308.3;DFD, 1785.8±233.7; F2,24=9.62, p=0.0013). Microbial com-munity diversity also was similar between howler groups(Shannon: Motiepa=4.72±0.37; Balam=4.93±0.27) andwas highest during the WFD period compared to other sam-pling blocks (WFD, 5.17±0.31; DLD, 4.83±0.44; DFD, 4.67±0.32; F2,24=4.47, p=0.024).
When we controlled for differences in gut microbial com-munity composition between howler groups at the OTU(ARISA; F1,38=2.19, p=0.0004) and genus levels (pyrose-quencing; F1,24=2.61, p=0.0046), we detected differences ingut microbial community composition across sampling
Fig. 1 Plant parts consumed by athe Motiepa group (N=7individuals) and b the Balamgroup (N=6 individuals) acrosssampling periods. Values areexpressed in average percent(±SD) of total dry gramsconsumed
K. R. Amato et al.
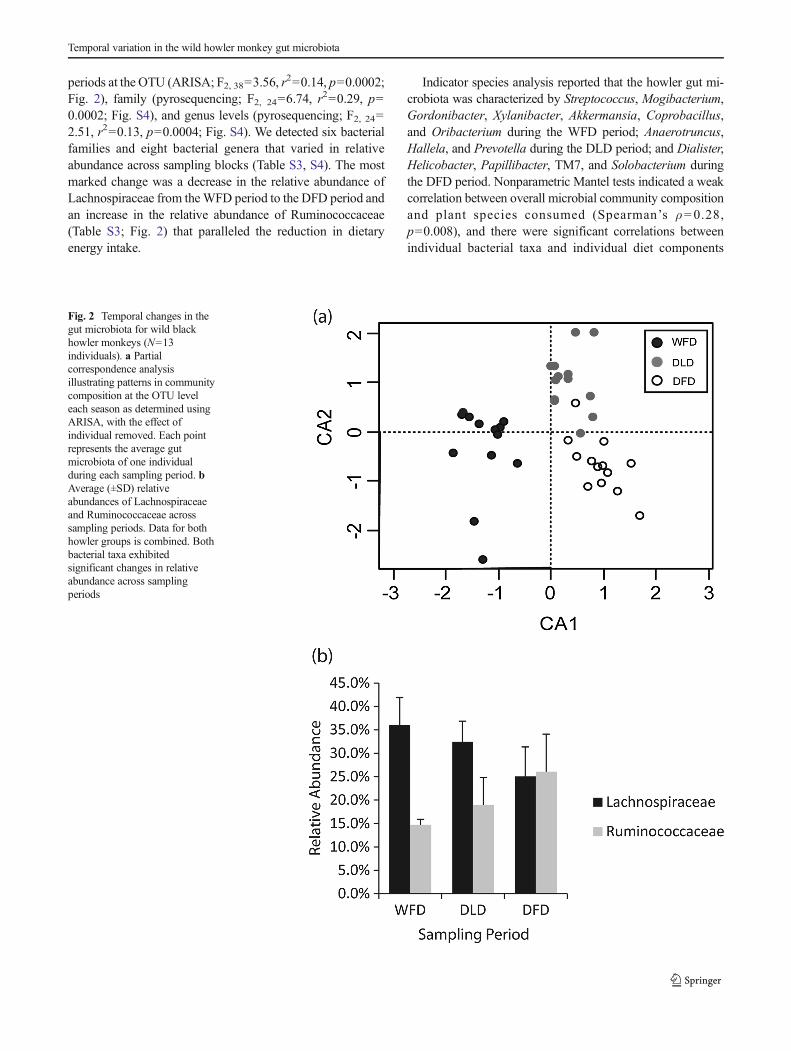
periods at the OTU (ARISA; F2, 38=3.56, r2=0.14, p=0.0002;
Fig. 2), family (pyrosequencing; F2, 24=6.74, r2=0.29, p=
0.0002; Fig. S4), and genus levels (pyrosequencing; F2, 24=2.51, r2=0.13, p=0.0004; Fig. S4). We detected six bacterialfamilies and eight bacterial genera that varied in relativeabundance across sampling blocks (Table S3, S4). The mostmarked change was a decrease in the relative abundance ofLachnospiraceae from theWFD period to the DFD period andan increase in the relative abundance of Ruminococcaceae(Table S3; Fig. 2) that paralleled the reduction in dietaryenergy intake.
Indicator species analysis reported that the howler gut mi-crobiota was characterized by Streptococcus, Mogibacterium,Gordonibacter, Xylanibacter, Akkermansia, Coprobacillus,and Oribacterium during the WFD period; Anaerotruncus,Hallela, and Prevotella during the DLD period; and Dialister,Helicobacter, Papillibacter, TM7, and Solobacterium duringthe DFD period. Nonparametric Mantel tests indicated a weakcorrelation between overall microbial community compositionand plant species consumed (Spearman’s ρ= 0.28,p=0.008), and there were significant correlations betweenindividual bacterial taxa and individual diet components
Fig. 2 Temporal changes in thegut microbiota for wild blackhowler monkeys (N=13individuals). a Partialcorrespondence analysisillustrating patterns in communitycomposition at the OTU leveleach season as determined usingARISA, with the effect ofindividual removed. Each pointrepresents the average gutmicrobiota of one individualduring each sampling period. bAverage (±SD) relativeabundances of Lachnospiraceaeand Ruminococcaceae acrosssampling periods. Data for bothhowler groups is combined. Bothbacterial taxa exhibitedsignificant changes in relativeabundance across samplingperiods
Temporal variation in the wild howler monkey gut microbiota
(Tables S5 and S6, Fig. S5). For example, Butyricicoccusrelative abundances were positively correlated to young leafand unripe fruit consumption (Table S5).
Gut Microbiota Function
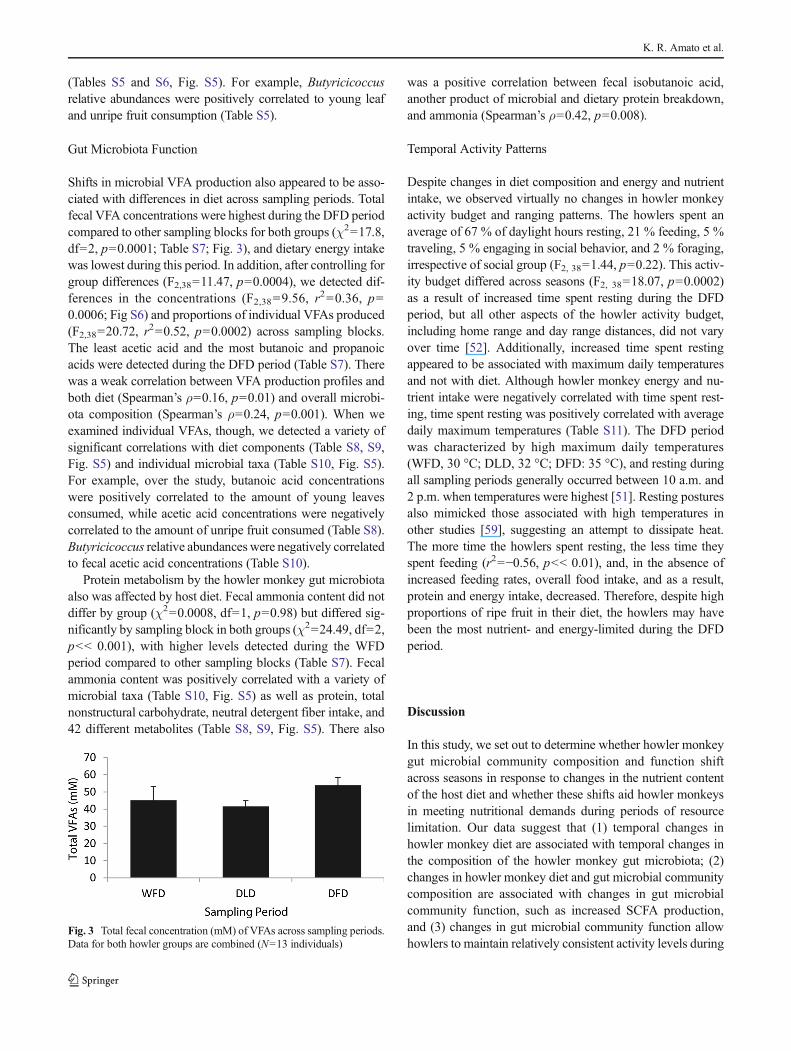
Shifts in microbial VFA production also appeared to be asso-ciated with differences in diet across sampling periods. Totalfecal VFA concentrations were highest during the DFD periodcompared to other sampling blocks for both groups (χ2=17.8,df=2, p=0.0001; Table S7; Fig. 3), and dietary energy intakewas lowest during this period. In addition, after controlling forgroup differences (F2,38=11.47, p=0.0004), we detected dif-ferences in the concentrations (F2,38=9.56, r
2=0.36, p=0.0006; Fig S6) and proportions of individual VFAs produced(F2,38=20.72, r
2=0.52, p=0.0002) across sampling blocks.The least acetic acid and the most butanoic and propanoicacids were detected during the DFD period (Table S7). Therewas a weak correlation between VFA production profiles andboth diet (Spearman’s ρ=0.16, p=0.01) and overall microbi-ota composition (Spearman’s ρ=0.24, p=0.001). When weexamined individual VFAs, though, we detected a variety ofsignificant correlations with diet components (Table S8, S9,Fig. S5) and individual microbial taxa (Table S10, Fig. S5).For example, over the study, butanoic acid concentrationswere positively correlated to the amount of young leavesconsumed, while acetic acid concentrations were negativelycorrelated to the amount of unripe fruit consumed (Table S8).Butyricicoccus relative abundances were negatively correlatedto fecal acetic acid concentrations (Table S10).
Protein metabolism by the howler monkey gut microbiotaalso was affected by host diet. Fecal ammonia content did notdiffer by group (χ2=0.0008, df=1, p=0.98) but differed sig-nificantly by sampling block in both groups (χ2=24.49, df=2,p<< 0.001), with higher levels detected during the WFDperiod compared to other sampling blocks (Table S7). Fecalammonia content was positively correlated with a variety ofmicrobial taxa (Table S10, Fig. S5) as well as protein, totalnonstructural carbohydrate, neutral detergent fiber intake, and42 different metabolites (Table S8, S9, Fig. S5). There also
was a positive correlation between fecal isobutanoic acid,another product of microbial and dietary protein breakdown,and ammonia (Spearman’s ρ=0.42, p=0.008).
Temporal Activity Patterns
Despite changes in diet composition and energy and nutrientintake, we observed virtually no changes in howler monkeyactivity budget and ranging patterns. The howlers spent anaverage of 67 % of daylight hours resting, 21 % feeding, 5 %traveling, 5 % engaging in social behavior, and 2 % foraging,irrespective of social group (F2, 38=1.44, p=0.22). This activ-ity budget differed across seasons (F2, 38=18.07, p=0.0002)as a result of increased time spent resting during the DFDperiod, but all other aspects of the howler activity budget,including home range and day range distances, did not varyover time [52]. Additionally, increased time spent restingappeared to be associated with maximum daily temperaturesand not with diet. Although howler monkey energy and nu-trient intake were negatively correlated with time spent rest-ing, time spent resting was positively correlated with averagedaily maximum temperatures (Table S11). The DFD periodwas characterized by high maximum daily temperatures(WFD, 30 °C; DLD, 32 °C; DFD: 35 °C), and resting duringall sampling periods generally occurred between 10 a.m. and2 p.m. when temperatures were highest [51]. Resting posturesalso mimicked those associated with high temperatures inother studies [59], suggesting an attempt to dissipate heat.The more time the howlers spent resting, the less time theyspent feeding (r2=−0.56, p<< 0.01), and, in the absence ofincreased feeding rates, overall food intake, and as a result,protein and energy intake, decreased. Therefore, despite highproportions of ripe fruit in their diet, the howlers may havebeen the most nutrient- and energy-limited during the DFDperiod.
Discussion
In this study, we set out to determine whether howler monkeygut microbial community composition and function shiftacross seasons in response to changes in the nutrient contentof the host diet and whether these shifts aid howler monkeysin meeting nutritional demands during periods of resourcelimitation. Our data suggest that (1) temporal changes inhowler monkey diet are associated with temporal changes inthe composition of the howler monkey gut microbiota; (2)changes in howler monkey diet and gut microbial communitycomposition are associated with changes in gut microbialcommunity function, such as increased SCFA production,and (3) changes in gut microbial community function allowhowlers to maintain relatively consistent activity levels during
Fig. 3 Total fecal concentration (mM) of VFAs across sampling periods.Data for both howler groups are combined (N=13 individuals)
K. R. Amato et al.

all periods of the year. These results highlight the importanceof the gut microbiota in wild primate nutrition and feedingecology and provide critical baseline information for under-standing the dynamic interactions between howler monkeysand their gut microbiota across time.
As hypothesized, both howler monkey diet and gut micro-bial community composition varied across sampling periods.Although additional data are necessary to fully explore themechanisms by which seasonal changes in diet impact the gutmicrobial community, in many cases, the observed patternssuggest an adaptation or adjustment of the gut microbiota tohost diet. For example, Prevotella was characteristic of theDLD period and requires both peptides and iron to stimulategrowth [60]. Because leaves consumed by howlers generallycontain more protein-binding tannins than fruits and have highiron levels (young leaf, 155.4 μg/g; fruit, 77.2 μg/g) [43, 45,61], a leaf-heavy diet during the DLD period may haveresulted in more undigested peptides and iron reaching thehowler hindgut for use by Prevotella. In addition, Prevotellacan utilize xylan efficiently [62], which is an important com-ponent of leaves. Similarly, Papillibacter relative abundanceswere positively correlated to the amount of the flavonoid,kaempferol, consumed. Because Papillibacter is related to aflavonoid-degrading anaerobe [63], increases in its abundancein response to kaempferol suggest an adaptation of the gutmicrobial community to food resources containingkaempferol. Such an adaptation would be beneficial to boththe host and the gut microbiota.
In addition to patterns in gut microbial community compo-sition, subtle changes in black howler fecal VFA profilesacross this study signal diet-related shifts in gut microbialcommunity function. For example, over the study, butanoicacid concentrations and Butyricicoccus relative abundanceswere positively correlated to the amount of young leaves andunripe fruits consumed. Because Butyricicoccus degrades fi-ber and resistant starch to produce butanoic acid [64], thesepatterns may be the result of Butyricicoccus fermenting thefiber in young leaves and unripe fruit to produce butanoicacid. Although our data did not show a relationship betweenthe amount of NDF in the howler diet and either butanoic acidconcentrations or the relative abundance of Butyricicoccus,NDF intake was positively correlated to unripe fruit intake.Also, Butyricicoccus has the ability to produce butyrate fromacetate [64], and our results indicate that Butyricicoccus rela-tive abundances were negatively correlated to fecal acetic acidconcentrations. Therefore, it appears that the combination ofhigh relative abundances of Butyricicoccus with a diet char-acterized by young leaves and unripe fruit provided the howlermonkeys with additional energy-rich butyrate from the fer-mentation of dietary fiber and the conversion of acetate tobutyrate.
Likewise, increased fecal ammonia during theWFD periodindicates a shift in gut microbiota function in response to diet.
Ammonia is produced when undigested protein (either proteinin excess of host requirements or protein bound to fiber andtannins) reaches the hindgut and is fermented by the bacterialcommunity to produce energy [65], as well as from the effluxof urea across the gut wall. Most bacteria can use ammonia asa nitrogen source when energy is not limiting, but if ammoniais produced in excess of bacterial requirements, it builds upand can be toxic to the host [66]. Therefore, increased proteinand fiber intake generally results in increased fecal ammoniaexcretion [53, 67]. It is likely that increased protein and fiberintake during the WFD period resulted in higher fecal ammo-nia concentrations. High-fiber intake is likely to have led tomore protein being indigestible in the small intestine, and bothenergy and protein intake surpassed estimated howler monkeyrequirements during this time (~0.49–0.58 MJ of energy permetabolic body weight, ~3.26 g of protein per metabolic bodyweight [52]). While excess energy (carbohydrates and lipids)can be stored in adipose tissue for later use [68, 69], excessprotein appears to have been converted to ammonia andexcreted. The positive correlation between fecal isobutanoicacid, another product of microbial and dietary protein break-down, and ammonia (Spearman’s ρ=0.42, p=0.008) supportsthis hypothesis. Relationships between ammonia and otherdiet components may be an effect of covariation in macronu-trient intake since during the WFD period, the howler mon-keys consumed the highest quantities of all macronutrientsmeasured.
Finally, an examination of black howler foraging behaviorand activity patterns suggests that the shifts we observed in gutmicrobiota function contributed to balancing howler nutrientintake under conditions of changing food availability and diet.During the study, we observed few changes in howler monkeyactivity and ranging patterns despite marked changes in dietcomposition and energy and nutrient intake. In particular, wefound no evidence that reduced nutrient and energy intakeduring the DFD period altered howler monkey activity pat-terns and ranging behavior. Because overall fecal VFA con-centrations were highest in the DFD period compared to othersampling blocks, we hypothesize that increased microbialVFA production during the DFD period aided the howlers inmeeting energy requirements despite reduced food intake. Inparticular, higher amounts of acetic acid, which can be used bythe host for fatty acid synthesis, lipogenesis, and musclemetabolism [12], and propanoic acid, which can be used toproduce blood glucose [12], may have provided howlers withthe energy necessary to maintain time spent traveling, travelsimilar distances each day, and engage in social behavior at alevel similar to the DLD and WFD periods.
It is also interesting to note that although an importantmucin degrader in the gut, Akkermansia, has been associatedwith fasting and lack of exogenous substrates for microbialmetabolism in other studies of the gut microbiota [70, 71], wedid not observe increased relative abundances of this microbe
Temporal variation in the wild howler monkey gut microbiota

in this study during the DFD period. This difference is likelydue to the fact that the amount of food consumed by thehowler monkeys was simply reduced during the DFD Period.The howler monkeys were not fasting. Therefore, exogenoussubstrates were being provided to the gut microbial commu-nity throughout each day, and mucin degradation may nothave been critical for energy production by the gut microbiota.However, further investigation of this relationship in wildprimates, especially during periods of extreme drought ornatural disasters [36, 72], is necessary to understand the im-portance of Akkermansia during extreme diet restrictionevents.
Like mantled howler monkeys (Alouatta palliata) [50], theblack howler monkeys in this study exhibited high molarproportions of acetic acid in the feces compared to otherprimates and mammals (Table S12) [73]. These results sug-gest that high proportions of acetic acid are typical of howlermonkeys and may be an important energy source. Althoughhowler monkey genetics and digestive physiology may play arole in the production of similar VFA profiles by influencingthe gut microbiota [74], it is more likely that this pattern is theresult of similar diets across the genus [75]. High proportionsof acetic acid are indicative of a high-fiber diet and a slow,efficient fermentation process [76]. While howler monkeysare not reported to consume more fiber than primates such ascolobines or gorillas, high proportions of acetic acid mayimply that soluble components of the howler monkey dietare more completely digested in the stomach and small intes-tine, and therefore, only fibrous, insoluble components arefermented in the hindgut to produce acetic acid [76]. Increasedrelative abundances of Ruminococcaceae during the DFDperiod may signal this type of efficient fermentation in re-sponse to reduced energy intake since Ruminococcaceae isenriched for cellulase genes compared to Lachnospiraceae[77].
Overall, our data suggest that the ability of the gut micro-biota to shift in response to changes in the howler diet canenhance howler nutrition by providing energy-rich SCFAsthrough the fermentation of otherwise indigestible plant car-bohydrates. However, this process is unlikely to be unique tohowler monkeys. Over 70 % of measured primate responsesto seasonality involve dietary shifts while less than 10 %involve changes in home range, day range, or activity levels[78]. Although field data and metagenomic examinations ofmicrobial metabolic processes are needed to verify thesemechanisms, energy and nutrient production by the gut mi-crobiota may provide an effective buffer against fluctuationsin energy and nutrient intake in primates and other mammalsadopting diet shifts as a strategy to endure seasonal changes infood availability. Furthermore, because dietary flexibility is astrong determinant of host fitness in new or variable environ-ments [79], understanding the ability of the gut microbiota toadapt to changes in the diet over periods of days, months, or
years and its role in maintaining host nutritional balances willprovide critical new perspectives on mammalian ecology andevolution.
Acknowledgments This study was funded by a National GeographicWaitt grant (W139-10) and a University of Illinois Dissertation TravelGrant to KRA, a University of Illinois Research Board Grant to PAG, andNational Science Foundation grant #0935347 (HOMINID) to SRL,RMS, BAW, and KEN. KRAwas also supported by a National ScienceFoundation Graduate Research Fellowship. We would like to thankAlejandro Estrada and the Universidad Nacional Autonoma de Méxicofor logistic support in the field, as well as Brianna Wilkinson and SarieVan Belle. We thank Gillian Britton for her help in the lab and ManolitoTorralba and Marcus Gillis for their help with sequencing at the J. CraigVenter Institute. We also thank CONANP, SEMARNAT, and SAGARPAin Mexico and the CDC in the USA for permits and logistic support. Wewould like to acknowledge Miguel Martínez Icó and Dave Seigler fortheir help with tree species identification and Alex Ulanov for his helpwith metabolite analyses through the Metabolomics Center at the Roy J.Carver Biotechnology Center, University of Illinois at Urbana-Cham-paign. PAG wishes to thank Chrissie, Sara, and Jenni for their love andsupport. The authors report no conflicts of interest.
References
1. Mackie RI (2002) Mutualistic fermentative digestion in the gastroin-testinal tract: diversity and evolution. Integr Comp Biol 42:319–326
2. Friswell MK, Gika H, Stratford IJ, Theodoridis G, Telfer B, WilsonID, McBain AJ (2010) Site and strain-specific variation in gut mi-crobiota profiles and metabolism in experimental mice. PLoS One 5:e8584
3. Mackie RI, Sghir A, Gaskins HR (1999) Developmental microbialecology of the neonatal gastrointestinal tract. Am J Clin Nutr 69:1035S–1045S
4. Koren O, Goodrich JK, Cullender TC, Spor A, Laitinen K, BackhedHK, Gonzalez A, Werner JJ, Angenent LT, Knight R, Backhed F,Isolauri E, Salminen S, Ley RE (2012) Host remodeling of the gutmicrobiome and metabolic changes during pregnancy. Cell 150:470–480
5. Suzuki K, Harasawa R, Yoshitake Y, Mitsuoka T (1983) Effect ofcrowding and heat stress on intestinal flora, body weight gain, andfeed efficiency of growing rats and chicks. Nippon Juigaku Zasshi45(3):331–8
6. Kolida S, Tuohy K, Gibson GR (2002) Prebiotic effects of inulin andoligofructose. Br J Nutr 87:S193–S197
7. De Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB,Massart S, Collini S, Pieraccini G, Lionetti P (2010) Impact of diet inshaping gut microbiota revealed by a comparative study in childrenfrom Europe and rural Africa. PNAS 107:14691–14696
8. Sekirov I, Russel SI, Antunes CM, Finlay BB (2010) Gut microbiotain health and disease. Physiol Rev 90:859–904
9. Brinkworth GD, Noakes M, Clifton PM, Bird AR (2009)Comparative effects of very low-carbohydrate, high-fat and high-carbohydrate, low-fat weight-loss diets on bowel habit and faecalshort-chain fatty acids on bacterial populations. Br J Nutr 101:1493–1502
10. Donohoe DR, Garge N, Zhang X, Sun W, O’Connell TM, BungerMK, Bultman SJ (2011) The microbiome and butyrate regulateenergy metabolism and autophagy in the mammalian colon. CellMetab 13:517–526
11. Duncan SH, Belenguer A, Holtrop G, Johnstone AM, Flint HJ,Lobley GE (2007) Reduced dietary intake of carbohydrates by obese
K. R. Amato et al.

subjects results in decreased concentrations of buyrate and butyrate-producing bacteria in feces. Appl Environ Microbiol 73:1073–1078
12. Flint HJ, Scott KP, Duncan SH, Louis P, Forano E (2012) Microbialdegradation of complex carbohydrates in the gut. Gut Microbes 3:289–306
13. Fraser MD, Theobald VJ, Davies DR, Moorby JM (2009) Impact ofdiet selected by cattle and sheep grazing heathland communities onnutrient supply and faecal micro-flora activity. Agric Ecosyst Environ129:367–377
14. Secor SM (2001) Regulation of digestive performance: a proposedadaptive response. Comp Biochem Physiol 128:565–577
15. Nicholson JK, Holmes E, Kinross J, Burcelin R, Gibson G, Jia W,Pettersson S (2012) Host-gut microbiota metabolic interactions. Sci336(6086):1262–1267
16. Duncan SH, Scott KP, Ramsay AG (2003) Effects of alternative dietarysubstrates on competition between human colonic bacteria in an anaer-obic fermentor system. Appl Environ Microbiol 69:1136–1142
17. Hooper LV, Midtvedt T, Gordon JI (2002) How host-microbial inter-actions shape the nutrient environment of the mammalian intestine.Annu Rev Nutr 22:283–307
18. Macfarlane S, Macfarlane GT (2003) Regulation of short-chain fattyacid production. Proc Nutr Soc 65:67–72
19. Chapman CA, Chapman LJ, Rode KD, Hauck EM, McDowell LR(2003) Variation in the nutritional value of primate foods: amongtrees, time periods, and areas. Int J Primatol 24:317–333
20. Milton K (1980) The foraging strategy of howlermonkeys. ColumbiaUniversity Press, New York
21. van Schaik CP, Terborgh J, Wright SJ (1993) The phenology oftropical forests: adaptive significance and consequences for primaryconsumers. Annu Rev Ecol Syst 24:353–377
22. Chaves OM, Stoner KE, Arroyo-Rodriguez V (2012) Differences indiet between spider monkey groups living in forest fragments andcontinuous forest in Mexico. Biotropica 44:105–113
23. Goldizen AW, Terborgh J, Cornejo F, Porras DT, Evans R (1988)Seasonal food shortage, weight loss, and the timing of births insaddle-back tamarins (Saguinus fuscicollis). J Anim Ecol 57(3):893–901
24. Nakagawa N (1997) Determinants of the dramatic seasonal changesin the intake of energy and protein by Japanese monkeys in a cooltemperate forest. Am J Primatol 41:267–288
25. Overdorff DJ, Strait SG, Telo A (1997) Seasonal variation in activityand diet in a small-bodied folivorous primate, Hapalemur griseus, insoutheastern Madagascar. Am J Primatol 43:211–223
26. Rothman JM, Dierenfeld ES, Hintz HF, Pell AN (2008) Nutritionalquality of gorilla diets: consequences of age, sex, and season.Oecologia 155:111–122
27. Amato KR, Yeoman CJ, Kent A, Carbonero F, Righini N, EstradaAE, Gaskins HR, Stumpf RM, Yildirim S, Torralba M, Gillis M,Wilson BA, Nelson KE, White BA, Leigh SR (2013) Habitat degra-dation impacts primate gastrointestinal microbiomes. ISME J 7:1344–1353
28. Williams CL, Willard S, Kouba A, Sparks D, Holmes W, Falcone J,Williams CH, Brown A (2012) Dietary shifts affect the gastrointes-tinal microflora of the giant panda (Ailuropoda melanoleuca). J AnimPhysiol Anim Nutr 97:577–585
29. Turnbaugh PJ, Ridaura VK, Faith JJ, Rey FE, Knight R, Gordon HA(2009) The effect of diet on the human gut microbiome: a metagenomicanalysis in humanized gnotobiotic mice. Sci Transl Med 1:6ra14
30. Wu GD, Chen J, Hoffmann C, Bittinger K, Chen YY, Keilbaugh SA,Bewtra M, Knights D, Walters WA, Knight R, Sinha R, Gilroy E,Gupta K, Baldassano R, Nessel L, Li H, Bushman FD, Lewis JD(2011) Linking long-term dietary patterns with gut microbialenterotypes. Sci 334:105–108
31. Wostmann BS, Larkin C, Moriarty A, Bruckner-Kardoss E (1983)Dietary intake, energymetabolism, and excretory losses of adult malegermfree Wistar rats. Lab Anim Sci 33:46–50
32. Backhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A,Semenkovich CF, Gordon JI (2004) The gut microbiota as an envi-ronmental factor that regulates fat storage. PNAS 101:15718–15723
33. Backhed F, Manchester JK, Semenkovich CF, Gordon JI (2007)Mechanisms underlying the resistance to diet-induced obesity ingerm-free mice. PNAS 104:979–984
34. Altmann SA (1998) Foraging for survival: Yearling baboons inAfrica. University of Chicago Press, Chicago
35. Gogarten JF, Brown LM, Chapman CA, Cords M, Doran-SheehyDM, Fedigan LM, Grine FE, Perry S, Pusey AE, Sterck EHM, WichSA, Wright PC (2012) Seasonal mortality patterns in non-humanprimates: implications for variation in selection pressures acrossenvironments. Evol 66:3252–3266
36. HamiltonWJ (1985) Demographic consequences of a food and watershortage to desert chacma baboons, Papio ursinus. Int J Primatol 6:451–462
37. Dunbar RIM (1980) Demographic and life history variables of apopulation of gelada baboons (Theropithecus gelada). J Anim Ecol49:485–506
38. Di Fiore A, Link A, Campbell C (2011) The Atelines: behavioral andsocioecological diversity in a New World monkey radiation. In:Campbell C, Fuentes A, MacKinnon KC, Panger M, Bearder SK(eds) Primates in perspective, 2nd edn. Oxford University Press,Oxford, pp 390–416
39. Norconk MA, Wright BW, Conklin-Brittain NL, Vinyard CJ (2009)Mechanical and nutritional properties of food as factors in platyrrhinedietary adaptations. In: Garber PA, Bicca-Marques JC, Estrada AE,Heymann EW, Strier KB (eds) South American primates, develop-ments in primatology: progress and prospects. Springer, New York,pp 279–319
40. Conklin NL, Wrangham RW (1994) The value of figs to a hind-gutfermenting frugivore: a nutritional analysis. Biochem Syst Ecol 22:137–161
41. Rosenberger AI, Strier KB (1989) Adaptive radiation of the atelineprimates. J Human Evol 18:717–750
42. Glander K (1981) Feeding patterns in mantled howling monkeys. In:Kamil A, Sargent TD (eds) Foraging behavior: ecological, ethological,and psychological approaches. Garland Press, New York, pp 231–259
43. Milton K (1979) Factors influencing leaf choice by howler monkeys:a test of some hypotheses of food selection by generalist herbivores.Am Nat 114:362–378
44. Behie AM, Pavelka MS (2012) Food selection in the black howlermonkey following habitat disturbance: implications for the impor-tance of mature leaves. J Trop Ecol 28:153–160
45. Silver SC, Ostro LET, Yeager CP, Dierenfeld ES (2000) Phytochemicaland mineral components of foods consumed by black howler monkeys(Alouatta pigra) at two sites in Belize. Zoo Biol 19:95–109
46. Chivers DJ, Hladik CM (1980) Morphology of the gastrointestinaltract in primates: comparisons with other mammals in relation to diet.J Morphol 166:337–386
47. Milton K (1981) Food choice and digestive strategies of two sym-patric primate species. Am Nat 117:496–505
48. Martins MM (2006) Comparative seed dispersal effectiveness ofsympatric Alouatta guariba and Brachyteles arachnoides inSoutheastern Brazil. Biotropica 38:57–63
49. Yumoto T, Kimura K, Nishimura A (1999) Estimation of the reten-tion times and distances of seeds dispersed by two monkey species,Alouatta seniculus and Lagothrix lagotricha, in a Colombian forest.Ecol Res 14:179–191
50. Milton K, McBee RH (1983) Rates of fermentative digestion in thehowler monkey, Alouatta palliata (Primates: Ceboidea). CompBiochem Physiol 74A:29–31
51. CONAGUA (2011) http://smn.cna.gob.mx/. http://smn.cna.gob.mx/52. Amato KR, Garber PA (2014) Nutrition and foraging strategies of the
black howler monkey (Alouatta pigra) in Palenque National Park,Mexico. Am J Primatol 76:774–787
Temporal variation in the wild howler monkey gut microbiota

53. Mackie RI, Stroot PG, Varel VH (1998) Biochemical identificationand biological origin of key odor components in livestock waste. JAnim Sci 76:1331–1342
54. Kent AD, Yannarell AC, Rusak JA, Triplett EW, McMahon KD(2007) Synchrony in aquatic microbial community dynamics.ISME J 1:38–47
55. Erwin ES,MarcoGJ, Emery EM (1961)Volatile fatty acid analysis ofblood and rumen fluid by gas chromatography. J Dairy Sci 44:1768–1771
56. Chaney AL, Marbach EP (1962) Modified reagents for the determi-nation of urea and ammonia. Clin Chem 8:130–132
57. Holm S (1979) A simple sequentially rejective multiple test proce-dure. Scand J Stat 6:65–70
58. RiceWR (1989) Analyzing tables of statistical tests. Evol 43:223–22559. Bicca-Marques JC, Calegaro-Marques C (1998) Behavioral thermo-
regulation in a sexually and developmentally dichromaticNeotropical primate, the black-and-gold howling monkey (Alouattacaraya). Am J Phys Anthr 106:533–546
60. Krieg NR, Ludwig W, Euzeby J, Whitman WB (2010) TheBacteroidetes, Spirochaetes, Tenericutes (Mollicutes),Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglmi,Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae,and Planctomycetes. Bergey’s manual of systematic bacteriology.Springer, New York, pp 25–469
61. Glander K (1979) Howling monkey feeding behavior and plantsecondary compounds: a study of strategies. In: Montgomery GG(ed) The ecology of arboreal folivores. Smithsonian Institution Press,Washington, pp 561–573
62. Dodd D, Mackie RI, Cann IK (2011) Xylan degradation, a metabolicproperty shared by rumen and human colonic Bacteroidetes. MolMicrobiol 79:292–304
63. Schleifer KH (2009) Phylum XIII. Firmicutes Gibbons and Murray1978, 5 (Firmacutes [sic] Gibbons and Murray 1978, 5). Bergey’smanual of systematic bacteriology. Springer, Dordrecht, pp 19–1317
64. Eeckhaut V, Van Immerseel FV, Teirlynch E, Pasmans F, Fievez V,Snauwaert C, Haesebrouck F, Ducatelle R, Louis P, Vandamme P(2008) Butyricicoccus pullicaecorum gen. nov., sp. nov. an anaero-bic, butyrate-producing bacterium isolated from the caecal content ofa broiler chicken. Int J Sys Evol Microbiol 58:2799–2802
65. Hungate RE (1966) The rumen and its microbes. Academic, New York66. Cotta MA, Russell JB (1997) Digestion of nitrogen in the rumen: a
model for metabolism of nitrogen compounds in gastrointestinalenvironments. In: Mackie RI, White BA (eds) Gastrointestinal mi-crobiology. Chapman and Hall, New York, pp 380–423
67. Cummings JH, Hill MJ, Bone ES, Branch WJ, Jenkins DJA (1979)The effect of meat protein and dietary fiber on colonic function andmetabolism. II. Bacterial metabolites in feces and urine. Am J ClinNutr 32:2094–2101
68. Dufour DJ, Sauther ML (2002) Comparative and evolutionary di-mensions of the energetics of human pregnancy and lactation. Am JHum Biol 14:584–602
69. Oftedal OT (2000) Use of maternal reserves as a lactation strategy inlarge mammals. Proc Nutr Soc 59:99–106
70. Sonoyama K, Fujiwara R, Takemura N, Ogasawara T, Watanabe J,Ito H, Morita T (2009) Response of gut microbiota to fasting andhibernation in Syrian hamsters. Appl Environ Microbiol 75:6451–6456
71. Costello EK, Gordon JI, Secor SM, Knight R (2010) Postprandialremodeling of the gut microbiota in Burmese pythons. ISME J 4:1375–1385
72. Behie AM, Pavelka MS (2005) The short-term effects of a hurricaneon the diet and activity of black howlers (Alouatta pigra) in MonkeyRiver, Belize. Folia Primatologia 76:1–9
73. Lambert JE, Fellner V (2012) In vitro fermentation of dietary carbo-hydrate consumed by African apes and monkeys: preliminary resultsfor interpreting microbial and digestive strategy. Int J Primatol 33:263–281
74. Buhnik-Rosenblau K, Danin-Poleg Y, Kashi Y (2011) Host geneticsand gut microbiota. In: Rosenberg E, Gophna U (eds) Beneficialmicroorganisms in multicellular life forms. Springer, Berlin, pp281–295
75. Garber PA, Righini N, Kowalewski MM (accepted) Evidence ofalternative dietary syndromes and nutritional goals in the GenusAlouatta. In: Kowalewski, MM, Garber, PA, Cortes-Ortiz, L,Urbani, B, Youlatos, D (eds.) Ecology and conservation of the mostwidely distributed Neotropical primate. Springer Press
76. Hume ID (1997) Fermentation in the hindgut of mammals. In:Mackie RI, White BA (eds) Gastrointestinal microbiology.Chapman and Hall, New York, pp 84–115
77. Biddle A, Stewart L, Blanchard J, Leschine S (2013) Untangling thegenetic basis of fibrolytic specialization by Lachnospiraceae andRuminococcaceae in diverse gut communities. Divers 5:627–640
78. Hemingway CA, Bynum N (2005) The influence of seasonality onprimate diet and ranging. In: Brockman DK, van Schaik CP (eds)Seasonality in primates: studies of living and extinct human and non-human primates. Cambridge University Press, New York, pp 57–95
79. Teaford MF, Ungar PS (2000) Diet and the evolution of the earliesthuman ancestors. PNAS 97:13506–13511
K. R. Amato et al.


















