
Differential Evolution A Simple Evolution Strategy for Fast Optimization
Upload
independentCategory
view
2download
0
The evolution of placentas and superfetation in the fishgenus Poecilia (Cyprinodontiformes: Poeciliidae:subgenera Micropoecilia and Acanthophacelus)
MARCELO N. PIRES, JEFF ARENDT and DAVID N. REZNICK*
Department of Biology, University of California, Riverside, California 900 University Avenue –Riverside, CA 92521, USA
Received 20 August 2009; accepted for publication 22 October 2009bij_1391 784..796
Complex adaptations are often found in nature, although our ability to discern how and why such traits evolvedis limited because their origin occurred in the distant past and the details of their evolution have been lost throughextinction (e.g. all placental mammals inherited their placentas from a single common ancestor that lived over100 Mya). In poeciliid fishes, placentas have evolved independently multiple times and portions of the path to theevolution of complexity can be found in living species. In the present study, we describe the life histories of sixspecies within the genus Poecilia that includes the subgenera Micropoecilia and Acanthophacelus (the guppy;Poecilia reticulata). We demonstrate that extensive placentotrophy and superfetation, the ability to simultaneouslycarry more than one developing brood, have evolved within this clade. These fish represent the third clade in whichwe have discovered the independent origin of a placenta that also includes close relatives that lack a placenta. Wediscuss possible adaptive advantages of the joint evolution of extensive placentation and superfetation in thesefishes. © 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796.
ADDITIONAL KEYWORDS: complex traits – guppy – life history – matrotrophy – placentotrophy –reproductive adaptations.
INTRODUCTION
The empirical study of the evolution of what Darwinreferred to as ‘organs of extreme perfection and com-plication’ (Darwin, 1859: 186), such as the eye, pre-sents a special challenge for two reasons. One is thatsuch organs are the product of a process that unfoldson a time scale that is much longer than our lives, soit is not directly observable. Another is that we haveno record of how the organ evolved because specieswith intermediate development of the trait areextinct. The eye of a vertebrate traces to a commonancestor that lived over 400 Mya, whereas the pla-centa of a mammal traces to a common ancestor thatlived over 100 Mya. Whatever transitions were asso-ciated with the origin of these ‘organs of extremeperfection’ have long since been lost to history, so we
must resort to indirect means to study their origin;however, logic dictates that somewhere among livingorganisms we should be able to find examples ofcomplexity that are in earlier stages of evolution suchthat evidence of the origin of the organ is retained asvariation among closely-related species. In thepresent study, we report on the discovery of suchvariation in a complex organ: the placenta in a groupof poeciliid fishes.
The fish family Poeciliidae (Rosen & Bailey, 1963)contains over 200 described species, although most ofthe impressive diversity that has been found acrossthe family is known from detailed studies of a fewspecies (Reznick & Miles, 1989). The guppy (Poeciliareticulata) is one of the most studied vertebrates andhas been a valuable model system for empiricalstudies of life-history evolution, sexual selection, andadaptation (Magurran, 2005). Among other accom-plishments, these studies have tested theoretical pre-dictions on the effects of age-specific mortality on*Corresponding author. E-mail: [email protected]
Biological Journal of the Linnean Society, 2010, 99, 784–796. With 3 figures
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796784

life-history evolution (Reznick, Bryga & Endler, 1990;Roff, 2002) and characterize the complex interactionsof pre- and post-copulatory mechanisms on sexualselection (Evans et al., 2003; Pilastro et al., 2004;Evans & Rutstein, 2008). Given the utility of thesefish for evolutionary studies and the diversityrevealed through the study of a small subset of thefamily, we have surveyed aspects of the biology of allspecies in the family. Interspecific studies create thepotential to address the evolution of a greater diver-sity of phenomena because trait variation amongspecies typically exceeds intraspecific variation.
In an early phase of our survey, we found thatextensive placentation evolved at least four timeswithin the family: once in the monospecific genusXenodexia and three times in the genus Poeciliopsis(Reznick et al., 2002; Reznick, Meredith & Collette,2007). Placentation has also been described in Het-erandria formosa (Grove & Wourms, 1991, 1994)and Phalloceros caudimaculatus (Arias & Reznick,2000). Such diversity in the development of placen-tas, often among closely-related species, provides avenue for studying the origins of biological complex-ity and its consequences. The placenta requiresadaptations at the trophic, immunological, osmo-regulatory, endocrinological, and respiratory levelsto deal with demands that are common to all pla-cental organisms. It is found across invertebrates(Woollacott & Zimmer, 1975; Huebner & Lococo,1994; Farley, 1998; Korniushin & Glaubrecht, 2003)and vertebrates (Wourms, Grove & Lombardi, 1988;Blackburn, 2006). Despite its frequent origin acrossa wide array of taxa, only a few studies have leadto the development of hypotheses for the adaptivevalue of the placenta (Trexler & DeAngelis, 2003;Crespi & Semeniuk, 2004), all of which remainlargely untested (Marsh-Matthews & Deaton, 2006;Pires et al., 2007; Banet & Reznick, 2008). The dis-covery of multiple origins of placentas and diversityin the development of placentas among closely-related species creates the potential for studyinghow and why placentas have evolved.
In the present study, we extend our family-widesurvey of reproductive biology to the close relatives ofthe guppy. Specifically, we describe the life histories offive species (Poecilia wingei, Poecilia picta, Poeciliaparae, Poecilia bifurca, Poecilia branneri) that arepart of the monophyletic cluster that includes guppiesand evaluate the presence of placentotrophy, superfe-tation, and other life-history traits. All of thesespecies but P. wingei, which was recently described(Poeser, Kempkes & Isbrücker, 2005), were classifiedin the subgenus Lebistes by Rosen & Bailey (1963).Meyer (1993) later revived an earlier classification byHubbs (1926), which was to treat P. picta, P. parae, P.bifurca, and P. branneri as the genus Micropoecilia.
He classified guppies as Lebistes, a monotypic subge-nus of Poecilia. Poeser et al. (2005) later describedPoecilia wingei (the Endler’s livebearer, Campomaguppy, or Cumaná guppy), a close relative of Poecilia(Lebistes) reticulata, and placed both species in thesubgenus Acanthophacelus rather than Lebistes.Shories, Meyer & Schartl (2009) described Poecilia(Acanthophacelus) obscura, which now represents athird species in the subgenus Acanthophacelus.Poeser et al. (2005) also placed all of Meyer’s (1993)Micropoecilia species and the newly-describedPoecilia (Lebistes) minima of Costa & Sarraf (1997)into Poecilia (Micropoecilia)
Prior to our study, extensive data were availabledescribing the life histories of guppies (Reznick &Endler, 1982) and there was one published study onthe life history of Poecilia picta (Reznick, Miles &Winslow, 1992). Poecilia picta has a very similar lifehistory to guppies; it lacks superfetation and appearsto fully provision eggs prior to fertilization. There areno published life-history data available for theremaining species. Poecilia branneri had beenreported by aquarists to produce litters of youngevery few days, which is anecdotal evidence of super-fetation, or the ability to simultaneously carry broodsin different developmental stages (Turner, 1937;Scrimshaw, 1944b). Superfetation is often associatedwith placentotrophy in the family Poeciliidae(Reznick & Miles, 1989), and so we suspected thatplacentas might also be present in these close rela-tives of guppies. Because guppies lack both placentasand superfetation, having close relatives that havethese traits would represent another independentorigin of placentas and superfetation in the familyPoeciliidae. In the present study, we describe the lifehistories of these fish. In a companion study(Meredith et al., 2010 ), we describe their phylogenet-ics, based upon the analysis of DNA sequences fromsix nuclear and two mitochondrial loci. Figure 1,derived from the companion study, shows the phylo-genetic relationships among the species examinedhere.
MATERIAL AND METHODSLIFE-HISTORY ANALYSIS: WILD-CAUGHT,
PRESERVED SPECIMENS
We dissected from four to 47 reproductive females perpopulation and from one to three populations perspecies (Table 1; the origin of each collection isdetailed in Table S1). Poecilia reticulata was repre-sented by two populations [a ‘low predation’ (JordanRiver) and a ‘high predation’ (Rincon River) locality],thus encompassing populations that express therange of life-history traits known for this species. We
PLACENTAL EVOLUTION IN POECILIA 785
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796

quantified female standard length (mm), brood size(number of embryos), and embryo dry weight (mg).The stage of development of embryos was classifiedwith a system modified from Reznick (1981) andHaynes (1995) based on fin, eye, and scale develop-ment, as well as other morphological features. When-ever permission was given from museums to desiccatefemales (which destroys the specimen), we estimatedfemale reproductive allocation (RA dry) as: (develop-ing embryos plus associated tissues)/(female total dryweight). We estimated size at maturity as theminimum millimetre size class in which two-thirds ofthe individuals contained developing embryos. We
inferred that superfetation was present when devel-oping embryos could be classified into discrete, well-separated stages of development. The degree ofsuperfetation per sample was defined as the averagenumber of developing broods within females (Table 1).The degree of superfetation per population/specieswas defined as the maximum number of broods foundwithin a female (Fig. 1); we chose this convention forcharacterizing populations/species because our labo-ratory studies reveal that the number of simulta-neous litters increases with the size (age) of theindividual and with food availability, so the maximumobserved number would come closest to representing
Cnesterodon decemmaculatus
Cnesterodon hypselurus
100
100
54
100
100
100
100
100
100
100
100
100
100
100
100
100
17
100
100
100
100
100
100
100
100
100
100
100
Superfetation Matrotrophic Index
2 55.1
7 63.8 - 99.7
2 4.4 - 10.0
1 0.76 - 0.80
1 0.61 - 0.79
1 0.84
Poecilia (Limia) dominicensis
Poecilia (Limia) melanogaster
Poecilia (Mollienesia) caucana
Poecilia (Mollienesia) latipunctata
Poecilia (Pamphorichthys) araguaiensis
Poecilia (Pamphorichthys) minor
Poecilia (Micropoecilia) bifurca
Poecilia (Micropoecilia) branneri
Poecilia (Micropoecilia) parae (French Guyana)
Poecilia (Micropoecilia) parae (Suriname)
Poecilia (Lebistes) reticulata
Poecilia (Lebistes) wingei (Unknown origin)
Poecilia (Lebistes) wingei (Venezuela)
Poecilia (Micropoecilia) picta (Venezuela)
Poecilia (Micropoecilia) picta (Trinidad)
Figure 1. Maximum posterior probability Bayesian tree (Meredith et al., 2010) recovered with the data set with 11partitions (each gene segment given own partition). Values above and below branches correspond to the mean Bayesianposterior probabilities based on two independent runs and the maximum likelihood bootstrap support percentages,respectively. Phylogenetic analyses from Hrbek et al. (2007) show Cnesterodon, our outgroup, as the genus most closelyallied to Poecilia. Superfetation values represent the maximum number of simultaneous developing broods per female foreach species. Matrotrophy index values represent the range across populations from which we obtained life-history data.Samples from which DNA was extracted did not always correspond to populations from which we obtained life-historydata.
786 M. N. PIRES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796

Tab
le1.
Des
crip
tive
stat
isti
csfo
rfe
mal
eli
fe-h
isto
rytr
aits
Spe
cies
Pop
ula
tion
N*
Min
imu
msi
zeat
mat
uri
ty(m
m)
Mea
n±
SE
fem
ale
size
(mm
)
Mea
n±
SE
fem
ale
dry
wei
ght
(mg)
Mea
n±
SE
broo
dsi
ze
Mea
n±
SE
nu
mbe
rof
deve
lopi
ng
broo
ds†
Siz
e-ad
just
edfe
cun
dity
‡,to
tal
nu
mbe
r±
SE
ofem
bryo
spe
rfe
mal
eR
epro
duct
ive
allo
cati
on(%
)
Poe
cili
are
ticu
lata
Jord
on47
/70
1717
.8±
0.3)
285
±15
2.8
±0.
21.
02.
9±
0.1
10.5
(0.6
)R
inco
n21
/26
1617
.7±
0.6
249
±27
6.4
±0.
71.
07.
4±
0.1
17.1
(1.0
)P
oeci
lia
win
gei
Lab
orat
ory
9/17
2427
.1±
1.4
819
±84
7.0
±1.
21.
01.
9±
0.2
17.6
(1,7
)P
oeci
lia
pict
aB
oat
Can
al25
/29
1920
.2±
0.4
287
±21
7.0
±0.
61.
05.
3±
0.1
17.1
(0.7
)M
aria
nn
e9/
1921
22.4
±0.
745
9±
6015
.9±
2.4
1.0
8.6
±0.
222
.8(1
.9)
Poe
cili
apa
rae
Leo
nsb
erg
10/1
226
25.7
±0.
818
.2±
1.5
1.1
±0.
16.
8±
0.2
Lel
iën
daal
5/7
2123
.5±
1.1
599
±96
13.0
±2.
71.
05.
6±
0.2
8.1
(2.8
)E
mm
astr
aat
8/15
1820
.8±
1.1
402
±86
7.6
±1.
31.
05.
3±
0.2
8.1
(1.2
)P
oeci
lia
bifu
rca
Ura
coa
4/5
1214
.0±
1.3
2.5
±0.
71.
06.
6±
0.3
Cor
opin
a14
/14
1414
.9±
0.2
106
±5
3.9
±0.
41.
4±
0.1
11.3
±0.
25.
9(1
.3)
Poe
cili
abr
ann
eri
João
Alv
es30
/30
1718
.2±
0.3
262
±12
6.0
±0.
84.
5±
0.2
23.5
±0.
16.
1(0
.4)
Cap
im20
/20
1919
.5±
0.3
3.1
±0.
34.
8±
0.2
12.1
±0.
1Ta
uá
6/7
1919
.8±
0.4
3.3
±0.
65.
0±
0.5
13.1
±0.
2
*Nu
mbe
rof
repr
odu
ctiv
efe
mal
es/t
otal
nu
mbe
rof
fem
ales
diss
ecte
d.In
the
foll
owin
gca
ses,
asa
resu
ltof
the
rem
oval
ofst
atis
tica
lou
tlie
rsor
vari
atio
nin
diss
ecti
onm
eth
odol
ogy,
the
sam
ple
size
for
ace
rtai
nva
riab
lew
asre
duce
d[P
.bif
urc
aC
orop
ina:
fem
ale
dry
wei
ght,
N=
9;re
prod
uct
ive
allo
cati
on(R
A)
dry,
N=
9;P.
bran
ner
iJo
ãoA
lves
:fe
mal
edr
yw
eigh
t,N
=15
;R
Adr
y,N
=15
;P.
para
eL
elië
nda
al:
RA
dry,
N=
4].
†At
leas
ton
ein
divi
dual
exh
ibit
ssu
perf
etat
ion
per
popu
lati
onw
hen
ever
the
aver
age
nu
mbe
rof
sim
ult
aneo
us
deve
lopi
ng
broo
dsis
grea
ter
than
1.‡E
stim
ated
fecu
ndi
tyfo
rth
eav
erag
efe
mal
esi
zeac
ross
all
popu
lati
ons,
base
don
the
back
-nat
ura
l-lo
g-tr
ansf
orm
edva
lues
from
the
mar
gin
alm
ean
sof
the
anal
ysis
ofco
vari
ance
ofn
atu
ral
log-
tran
sfor
med
fecu
ndi
tyw
ith
popu
lati
onas
fixe
dfa
ctor
.
PLACENTAL EVOLUTION IN POECILIA 787
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796

a population attribute that was independent of envi-ronmental influences. Laboratory studies in theseand other poeciliid species (Reznick & Miles, 1989;Reznick et al. 1996; Pires et al. 2007) demonstrate aclose correspondence between the degree of super-fetation identified through dissections and theinterbrood interval observed in live specimens;populations/species with a higher degree of superfe-tation have shorter intervals between the birth ofsuccessive litters. Total fecundity in species withsuperfetation equals the average brood size multi-plied by the number of developing broods.
LIFE HISTORY ANALYSIS: LABORATORY STUDIES
We augmented our analyses of wild-caught fish withlaboratory studies of wild-caught females of P. reticu-lata, P. wingei, P. picta, and three populations of P.branneri. The latter three are part of what may soonbe described as the branneri species complex, ratherthan being different populations of P. branneri(Figueiredo, 2003). Details on the origin of our labo-ratory stocks are given in the Supporting Informa-tion, Table S3.
All fish included in the present study were second-or later-generation offspring of wild-caught adults(Table S2). All had been reared under a commonlaboratory environment on the same diet, but differedin water quality. Poecilia reticulata, P. wingei, and P.picta were reared in water that had a total hardnessof approximately 200 p.p.m. and a pH of approxi-mately 8, whereas the populations of P. branneri werereared in water that had a pH of approximately 7 andtotal hardness of approximately 100 p.p.m.. Theseconditions were chosen to match the water quality oftheir native habitats. We reared newborn fish atdensities of one to six per 19-L tank on ad libitumfood until aged 25 days old. They were thereafterreared one per 8-L tank on a quantified ration of liverpaste (morning) and newly-hatched brine shrimp(afternoon) that was designed to sustain a moderaterate of growth, based on earlier research on guppies(Reznick, 1983).
We measured length and weight every other weekand examined the anal fin with aid of a dissectingmicroscope to identify sex. Fish were anaesthetizedwith MS-222 to facilitate handling. Females weremated once per week, beginning at an age of 5 weeks,by adding a mature male to the tank for a period of 2days. There is no external index of maturity forfemales, so we instead used the age at first parturi-tion as in index of maturity. For species that lackedsuperfetation, we kept all females until they producedthree broods of young plus an additional 3–5 daysafter the production of the third brood so that theycould mature a new set of ova. Species with superfe-
tation reproduce much more frequently and alwayscontain multiple litters of developing young, each at adifferent stage of development, so we instead keptthem for 60 days after the production of their firstlitter. We chose 60 days because it equals or slightlyexceeds the time interval between the production ofthe first and third broods of young by species thatlack superfetation. Our dependent variables were theage at first parturition, the number of offspring perbrood, the mean dry weight of newborn offspring, andthe average number of days between consecutivelitters.
We preserved all females in a solution of 5% form-aldehyde after the period of reproduction, then theywere dissected. Species that lack superfetation onlycontained the yolking ova that would comprise theirnext litter. Species with superfetation generally con-tained multiple litters of developing young; werecorded the stage of development, number of off-spring, and dry weight of offspring for these individu-als. The dissections yielded estimates of additionaldependent variables, including the total number ofdeveloping offspring, reproductive allocation, and thenumber of developing litters.
STATISTICAL ANALYSIS
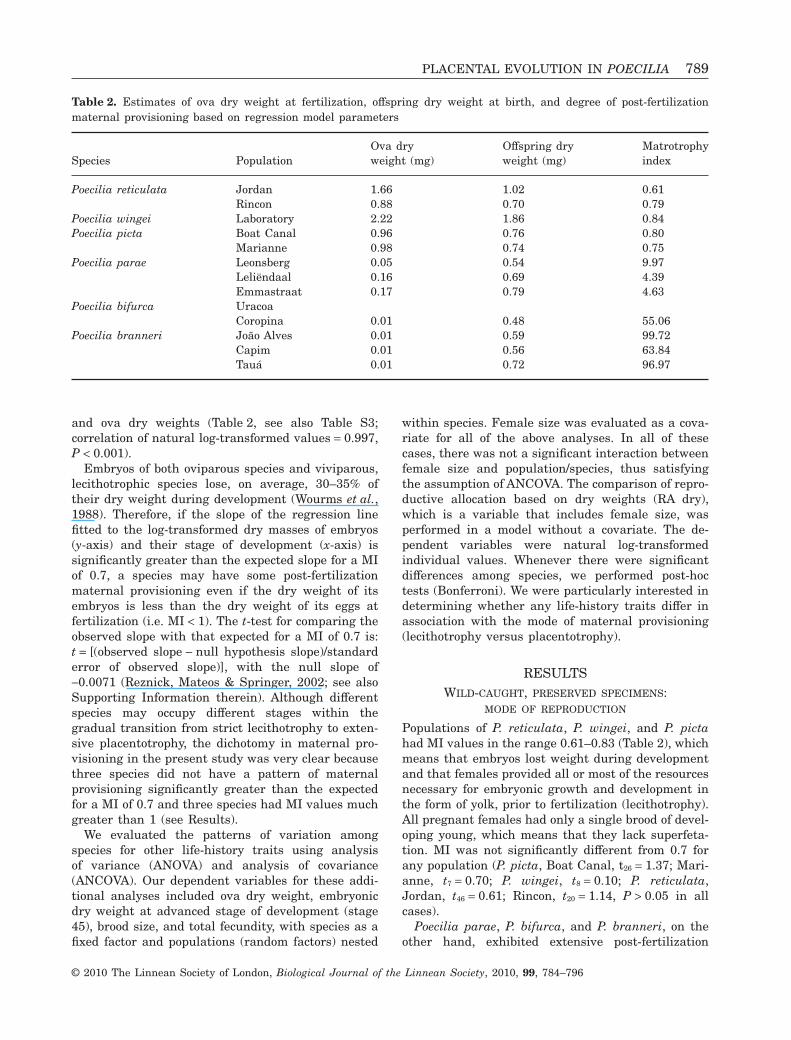
We describe the relationship between the stage ofdevelopment of embryos and their dry mass by con-verting their stage of development into a numericalscale that ranged in value from 0 (fully yolked eggs)to 45 (fully developed embryos). Other stages areassigned values in increments of five. In priorresearch on guppies (Reznick & Endler, 1982), apply-ing this scale to the relationship between embryo drymass (y-axis) and stage of development (x-axis) pro-duced a straight line with a negative slope. Applyingthe same numerical scale to all species gives us auniform frame of reference for evaluating maternalprovisioning.
The degree of placentotrophy was quantified withthe matrotrophy index (MI), which is the the drymass of an advanced embryo (stage 45) or newbornoffspring divided by the dry mass of a fully-yolked egg(stage 0). We estimated these values for all popula-tions based on the linear or quadratic regressionmodels between stage of development (independentvariable) and embryonic dry mass (dependent vari-able, natural log-transformed to meet assumptions ofregression analyses whenever needed). For popula-tions for which we had multiple measurements ofstage 0 and stage 45 embryos (or, in two instances,embryos in developmental stages 0–5 or 40), we alsoestimated MI directly. Estimated MI values based onregression models generated very similar results tothe actual ratio between average stage 45 embryos
788 M. N. PIRES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796
and ova dry weights (Table 2, see also Table S3;correlation of natural log-transformed values = 0.997,P < 0.001).
Embryos of both oviparous species and viviparous,lecithotrophic species lose, on average, 30–35% oftheir dry weight during development (Wourms et al.,1988). Therefore, if the slope of the regression linefitted to the log-transformed dry masses of embryos(y-axis) and their stage of development (x-axis) issignificantly greater than the expected slope for a MIof 0.7, a species may have some post-fertilizationmaternal provisioning even if the dry weight of itsembryos is less than the dry weight of its eggs atfertilization (i.e. MI < 1). The t-test for comparing theobserved slope with that expected for a MI of 0.7 is:t = [(observed slope - null hypothesis slope)/standarderror of observed slope)], with the null slope of-0.0071 (Reznick, Mateos & Springer, 2002; see alsoSupporting Information therein). Although differentspecies may occupy different stages within thegradual transition from strict lecithotrophy to exten-sive placentotrophy, the dichotomy in maternal pro-visioning in the present study was very clear becausethree species did not have a pattern of maternalprovisioning significantly greater than the expectedfor a MI of 0.7 and three species had MI values muchgreater than 1 (see Results).
We evaluated the patterns of variation amongspecies for other life-history traits using analysisof variance (ANOVA) and analysis of covariance(ANCOVA). Our dependent variables for these addi-tional analyses included ova dry weight, embryonicdry weight at advanced stage of development (stage45), brood size, and total fecundity, with species as afixed factor and populations (random factors) nested
within species. Female size was evaluated as a cova-riate for all of the above analyses. In all of thesecases, there was not a significant interaction betweenfemale size and population/species, thus satisfyingthe assumption of ANCOVA. The comparison of repro-ductive allocation based on dry weights (RA dry),which is a variable that includes female size, wasperformed in a model without a covariate. The de-pendent variables were natural log-transformedindividual values. Whenever there were significantdifferences among species, we performed post-hoctests (Bonferroni). We were particularly interested indetermining whether any life-history traits differ inassociation with the mode of maternal provisioning(lecithotrophy versus placentotrophy).
RESULTSWILD-CAUGHT, PRESERVED SPECIMENS:
MODE OF REPRODUCTION
Populations of P. reticulata, P. wingei, and P. pictahad MI values in the range 0.61–0.83 (Table 2), whichmeans that embryos lost weight during developmentand that females provided all or most of the resourcesnecessary for embryonic growth and development inthe form of yolk, prior to fertilization (lecithotrophy).All pregnant females had only a single brood of devel-oping young, which means that they lack superfeta-tion. MI was not significantly different from 0.7 forany population (P. picta, Boat Canal, t26 = 1.37; Mari-anne, t7 = 0.70; P. wingei, t8 = 0.10; P. reticulata,Jordan, t46 = 0.61; Rincon, t20 = 1.14, P > 0.05 in allcases).
Poecilia parae, P. bifurca, and P. branneri, on theother hand, exhibited extensive post-fertilization
Table 2. Estimates of ova dry weight at fertilization, offspring dry weight at birth, and degree of post-fertilizationmaternal provisioning based on regression model parameters
Species PopulationOva dryweight (mg)
Offspring dryweight (mg)
Matrotrophyindex
Poecilia reticulata Jordan 1.66 1.02 0.61Rincon 0.88 0.70 0.79
Poecilia wingei Laboratory 2.22 1.86 0.84Poecilia picta Boat Canal 0.96 0.76 0.80
Marianne 0.98 0.74 0.75Poecilia parae Leonsberg 0.05 0.54 9.97
Leliëndaal 0.16 0.69 4.39Emmastraat 0.17 0.79 4.63
Poecilia bifurca UracoaCoropina 0.01 0.48 55.06
Poecilia branneri João Alves 0.01 0.59 99.72Capim 0.01 0.56 63.84Tauá 0.01 0.72 96.97
PLACENTAL EVOLUTION IN POECILIA 789
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796
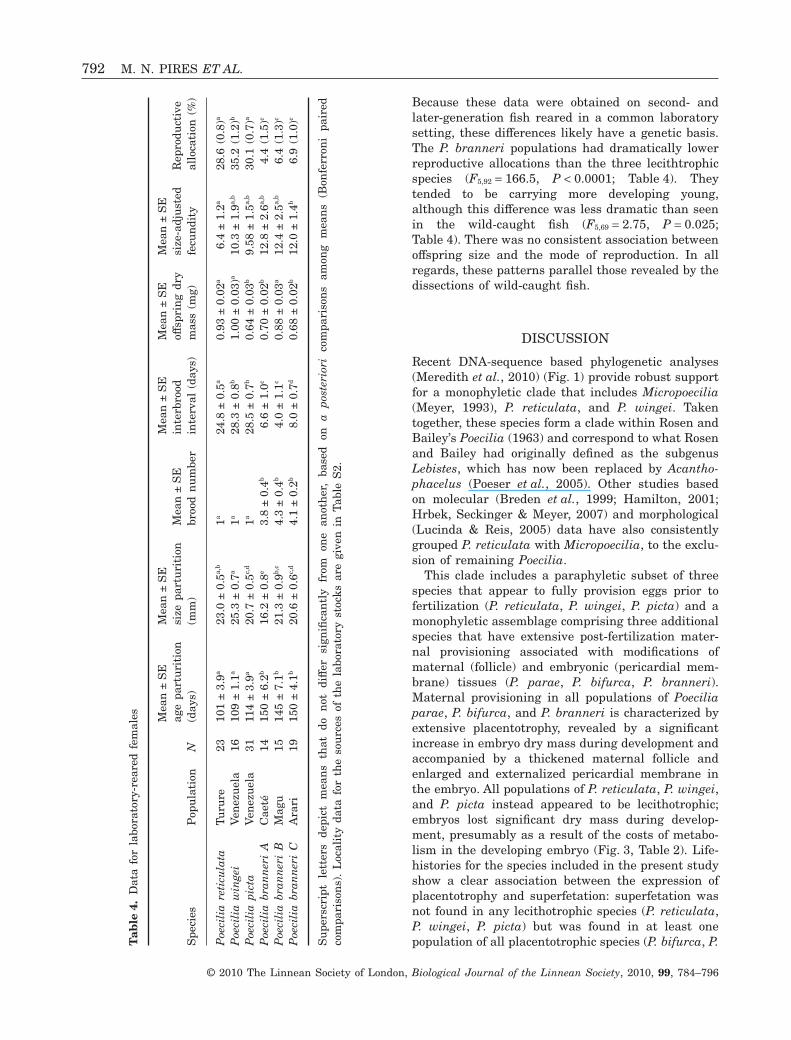
resource allocation to embryos via a follicular pla-centa (placentotrophy), with MI values in the range4.4–99.7 (Table 2, Fig. 2). They also exhibited super-fetation in various degrees (Fig. 3B, C, Table 1).Developing embryos from all placentotrophic specieswere enclosed in a thick maternal follicle and hadwell-developed pericardial membranes (Fig. 3B, D).By contrast, the developing embryos of lecithotrophicspecies were surrounded by diaphanous follicles andlacked well-developed, externalized pericardial mem-branes (Fig. 3A). These differences suggest that themembranes involved in resource transfer in thesespecies are likely similar to the membranes respon-sible for placentotrophy in other poeciliids (Turner,1940; Grove & Wourms, 1994).
Out of the 23 reproductive P. parae females dis-sected, only one exhibited superfetation (Fig. 3B, with20 ‘uneyed’ embryos and 17 ‘very late-eyed’ embryos,sensu Reznick, 1981). Superfetation was much morecommon in P. bifurca (six out of 18 reproductive
females dissected had two distinct broods) and it waspresent in all 56 reproductive females of P. branneri,in which the number of simultaneous broods was inthe range 2–7 (Table 1).
In conclusion, three of the six species (P. reticulata,P. wingei, P. picta) are lecithotrophic and lack super-fetation, whereas the other three (P. parae, P. bifurca,and P. branneri) are placentotrophic and have super-fetation. The molecular phylogeny (Fig. 1) shows thatthe latter three species are a monophyletic cladenested within the larger clade of six species, which inturn suggests that they inherited these life-historytraits from a single common ancestor.
WILD-CAUGHT, PRESERVED SPECIMENS:OTHER LIFE-HISTORY TRAITS
We quantified other features of the life history andevaluated the degree to which there were consistentdifferences in life histories between species with andwithout placentation.
First, the high MI values associated with P. parae,P. bifurca, and P. branneri were caused by a reduc-tion in the size of the ova at fertilization, ratherthan an increase in the size of embryos at birth(Fig. 3A, C, Table 2). The dry mass of eggs at fer-tilization is up to 100-fold greater in lecithotrophicspecies than in placentotrophic species (Fig. 3,Table 2). Species differed significantly in ovadry weight at fertilization (ANOVA: F3,48 = 277.8,P < 0.001; Tables 2, 3). Placentotrophic taxa pro-duced, on average, significantly smaller ova thanthe lecithotrophic taxa (P < 0.001 for all Bonferronipairwise comparisons between lecithotrophic andplacentotrophic taxa; Table 3). There was no asso-ciation between mode of maternal provisioning andthe estimated mass of full-term embryos (ANOVA,F3,16 = 2.2, P = 0.132; Tables 2, 3).
Size-adjusted average brood size differed signifi-cantly among species (ANCOVA: brood size: F5,194 =16.54, P < 0.001; female size: F1,194 = 115.43, P < 0.001;Table 3), although there was no consistent associationbetween brood size and the mode of maternal provi-sioning (Table 3).
Figure 2. Change in embryonic dry mass over develop-ment for a representative lecithotrophic species, Poeciliareticulata (Jordan population, grey diamonds) and a rep-resentative matrotrophic species, Poecilia branneri (CapimRiver population, dark circles). Stage 45 values for P.reticulata are slightly offset to facilitate identification oferror bars.
Table 3. Estimated marginal means from analysis of covariance and analysis of variance models, showing subgroups ofhomogeneous species means (a, b, c, d, and e) that did not differ significantly after Bonferroni pairwise comparisons
Poeciliareticulata
Poeciliawingei
Poeciliapicta
Poeciliaparae
Poeciliabifurca
Poeciliabranneri
Ova dry weight (mg) 1.65a 0.95a 0.17b 0.01c
Late embryo dry weight (mg) 0.80a 0.64a 0.55a 0.64a
Brood size 5.9a,b 1.0c 9.6a 9.5a,b 8.2a,b 4.1c
Reproductive allocation (%) 13.3b 14.6a,b 20.1a 7.1c 6.1c 5.9c
Fecundity 6.7c,d -2.5e 8.5b,c,d 8.2b,c,d 12.0b,c 20.4a
790 M. N. PIRES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796

Size-adjusted total number of embryos per female[(number of developing broods) ¥ (number of embryosper brood)] differed significantly among species;species with placentas and superfetation tended tohave higher fecundity than lecithotrophic species(ANCOVA: fecundity: F5,194 = 37.63, P < 0.001; femalesize: F1,194 = 72.39, P < 0.001; Table 3, Bonferroni pair-wise comparisons). The two species with the highestMI values also had the largest number of developingbroods (P. branneri and P. bifurca) and higher fecun-dities. The extreme case of P. branneri, which has thehighest size-adjusted fecundity, demonstrates that, inthe presence of superfetation, total fecundity is notcompromised by a reduction in number of young ineach brood; this species had relatively small broodsizes but had the largest degree of superfetation andlargest mean size-adjusted fecundity of all species(Table 1).
Reproductive allocation (the percent of total femaleweight that consists of developing embryos) is ameasure of the size of the ovary relative to the size ofthe female. Reproductive allocation based on dryweights (RA dry) differed significantly among species
(F5,165 = 21.87, P < 0.001). Placentotrophic speciesallocate significantly less of their total dry weightto reproduction (7.05%) compared to lecithotrophicspecies (17.01%; Tables 1, 3, Bonferroni pairwise com-parisons), despite the fact that they have the highesttotal fecundity.
LABORATORY STUDIES: LIFE HISTORIES
Our laboratory studies enabled us to augment thefield descriptions with the estimation of traits thatpertain to life cycle timing, including the age atmaturity and frequency of reproduction. The threepopulations of P. branneri were smaller and older atsexual maturity than the three lecithotrophic species.Their shorter interbrood intervals (4–8 days for P.branneri versus 25–28 days for the three leci-thotrophic species) are typical of species with super-fetation and the overlapping of developing broods. Inother respects, the life histories patterns of the P.branneri populations relative to the three leci-thotrophic species were qualitatively similar to theresults obtained for the wild-caught fish (Table 4).
Figure 3. A, Poecilia wingei ovum (far left) and embryos extracted from different females. Represented developmentalstages, from left to right: 0, 25, 30, 35, and 42.5. Ovum diameter = 1.6 mm. B, two embryos in different stages ofdevelopment (left: stage 10, ‘uneyed’; right: stage 45, ‘very late-eyed’), each representing one brood found inside a Poeciliaparae female at the time of dissection, thus characterizing superfetation. C, embryonic developmental series from Poeciliabranneri. Each embryo was removed from one of the five distinct broods found within a single female at the time ofdissection. D, Poecilia branneri embryo at advanced stage of development with maternal follicle removed, shown next tothe embryo.
PLACENTAL EVOLUTION IN POECILIA 791
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796
Because these data were obtained on second- andlater-generation fish reared in a common laboratorysetting, these differences likely have a genetic basis.The P. branneri populations had dramatically lowerreproductive allocations than the three lecithtrophicspecies (F5,92 = 166.5, P < 0.0001; Table 4). Theytended to be carrying more developing young,although this difference was less dramatic than seenin the wild-caught fish (F5,69 = 2.75, P = 0.025;Table 4). There was no consistent association betweenoffspring size and the mode of reproduction. In allregards, these patterns parallel those revealed by thedissections of wild-caught fish.
DISCUSSION
Recent DNA-sequence based phylogenetic analyses(Meredith et al., 2010) (Fig. 1) provide robust supportfor a monophyletic clade that includes Micropoecilia(Meyer, 1993), P. reticulata, and P. wingei. Takentogether, these species form a clade within Rosen andBailey’s Poecilia (1963) and correspond to what Rosenand Bailey had originally defined as the subgenusLebistes, which has now been replaced by Acantho-phacelus (Poeser et al., 2005). Other studies basedon molecular (Breden et al., 1999; Hamilton, 2001;Hrbek, Seckinger & Meyer, 2007) and morphological(Lucinda & Reis, 2005) data have also consistentlygrouped P. reticulata with Micropoecilia, to the exclu-sion of remaining Poecilia.
This clade includes a paraphyletic subset of threespecies that appear to fully provision eggs prior tofertilization (P. reticulata, P. wingei, P. picta) and amonophyletic assemblage comprising three additionalspecies that have extensive post-fertilization mater-nal provisioning associated with modifications ofmaternal (follicle) and embryonic (pericardial mem-brane) tissues (P. parae, P. bifurca, P. branneri).Maternal provisioning in all populations of Poeciliaparae, P. bifurca, and P. branneri is characterized byextensive placentotrophy, revealed by a significantincrease in embryo dry mass during development andaccompanied by a thickened maternal follicle andenlarged and externalized pericardial membrane inthe embryo. All populations of P. reticulata, P. wingei,and P. picta instead appeared to be lecithotrophic;embryos lost significant dry mass during develop-ment, presumably as a result of the costs of metabo-lism in the developing embryo (Fig. 3, Table 2). Life-histories for the species included in the present studyshow a clear association between the expression ofplacentotrophy and superfetation: superfetation wasnot found in any lecithotrophic species (P. reticulata,P. wingei, P. picta) but was found in at least onepopulation of all placentotrophic species (P. bifurca, P.T
able
4.D
ata
for
labo
rato
ry-r
eare
dfe
mal
es
Spe
cies
Pop
ula
tion
N
Mea
n±
SE
age
part
uri
tion
(day
s)
Mea
n±
SE
size
part
uri
tion
(mm
)M
ean
±S
Ebr
ood
nu
mbe
r
Mea
n±
SE
inte
rbro
odin
terv
al(d
ays)
Mea
n±
SE
offs
prin
gdr
ym
ass
(mg)
Mea
n±
SE
size
-adj
ust
edfe
cun
dity
Rep
rodu
ctiv
eal
loca
tion
(%)
Poe
cili
are
ticu
lata
Turu
re23
101
±3.
9a23
.0±
0.5a,
b1a
24.8
±0.
5a0.
93±
0.02
a6.
4±
1.2a
28.6
(0.8
)a
Poe
cili
aw
inge
iV
enez
uel
a16
109
±1.
1a25
.3±
0.7a
1a28
.3±
0.8b
1.00
±0.
03)a
10.3
±1.
9a,b
35.2
(1.2
)b
Poe
cili
api
cta
Ven
ezu
ela
3111
4±
3.9a
20.7
±0.
5c,d
1a28
.5±
0.7b
0.64
±0.
03b
9.58
±1.
5a,b
30.1
(0.7
)a
Poe
cili
abr
ann
eri
AC
aeté
1415
0±
6.2b
16.2
±0.
8e3.
8±
0.4b
6.6
±1.
0c0.
70±
0.02
b12
.8±
2.6a,
b4.
4(1
.5)c
Poe
cili
abr
ann
eri
BM
agu
1514
5±
7.1b
21.3
±0.
9b,c
4.3
±0.
4b4.
0±
1.1c
0.88
±0.
03a
12.4
±2.
5a,b
6.4
(1.3
)c
Poe
cili
abr
ann
eri
CA
rari
1915
0±
4.1b
20.6
±0.
6c,d
4.1
±0.
2b8.
0±
0.7d
0.68
±0.
02b
12.0
±1.
4b6.
9(1
.0)c
Su
pers
crip
tle
tter
sde
pict
mea
ns
that
don
otdi
ffer
sign
ifica
ntl
yfr
omon
ean
oth
er,
base
don
apo
ster
iori
com
pari
son
sam
ong
mea
ns
(Bon
ferr
oni
pair
edco
mpa
riso
ns)
.L
ocal
ity
data
for
the
sou
rces
ofth
ela
bora
tory
stoc
ksar
egi
ven
inTa
ble
S2.
792 M. N. PIRES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796

parae, P. branneri), and was clearly more pronouncedin the most placentotrophic species, P. branneri(Table 1).
The combination of these phylogenetic and life-history studies suggests that extensive maternal pro-visioning and superfetation evolved once in a commonancestor to P. parae, P. bifurca, and P. branneri. Inaddition, the phylogeny implies a progressive increasein maternal provisioning accompanied by an increasein the extent of superfetation across species.
To our knowledge, the presence of placentotrophywithin the subgenus Micropoecilia (or within any ofthe Poecilia, sensu Rosen & Bailey, 1963) has neverbeen documented. The results obtained in the presentstudy add this subgenus to the list of independentorigins of extensive placentotrophy in poeciliid fishes.Other origins are found in H. formosa (Turner, 1940;Scrimshaw, 1944a; Grove & Wourms, 1991; Schrader& Travis, 2005; MI of approximately 40–50) and threeclades in the genus Poeciliopsis (Reznick et al., 2002;13 of 21 species are placentotrophic, with MI valuesin the range 0.8–117). Placentotrophy is also found inP. caudimaculatus (Arias & Reznick, 2000; MI ofapproximately 2) and Xenodexia ctenolepis (Reznicket al., 2007; MI of approximately 3–5).
Scrimshaw (1944b) and Turner (1937) reportedanecdotal evidence that suggested the presence ofsuperfetation in P. branneri, but not in P. picta and P.parae, the other species in the Micropoecilia groupthat they examined. The results obtained in thepresent study, revealing varying degrees of supefeta-tion in P. branneri, P. bifurca, and P. parae, werederived from the dissection of wild-caught indi-viduals, so population/species differences may beconfounded with food availability and otherenvironmental effects (Thibault & Schultz, 1978;Travis et al., 1987); individuals that have access tomore food also tend to carry more developing litters ofyoung. Thus, it remains to be determined whethersuperfetation in P. parae and P. bifurca is rare, asimplied by the results obtained in the present study,or a by-product of environmental effects or femalereproductive condition. The results obtained in thelaboratory show that the differences between P. bran-neri and the three lecithotrophic species in super-fetation persist despite our controlling for foodavailability. Superfetation is known to occur in allother placentotrophic lineages of the Poeciliidae, withthe exception of P. caudimaculatus, and also in somelecithotrophic poeciliid species (Reznick & Miles,1989; Reznick et al., 2002). Across other taxa, super-fetation is found in the matrotrophic clinid (Gunn &Thresher, 1991) and zenarchopterid (Meisner &Burns, 1997; Reznick, Meredith & Collette, 2007)fishes. The common association between matrotrophyand superfetation found in poeciliid fishes (noted sub-
sequent to Scrimshaw, 1944b) thus appears to bepresent in other fish groups.
EVOLUTIONARY CONSIDERATIONS
These alternative life histories (i.e. lecithotrophywithout superfetation versus placentotrophy withsuperfetation) are associated with large differences inthe proportion of body mass that consists of develop-ing young (Tables 1, 4). Lecithotrophic species allo-cate all resources to their offspring prior tofertilization in the form of yolk. The reproductiveburden increases during development because theoffspring increase in wet mass and volume, even asdry mass declines (Ghalambor, Reznick & Walker,2004). By contrast, placentotrophic species withsuperfetation initiate a few young at a time withsmall ova and carry many small litters that increasein size as development progresses. The combinedeffect of small, staggered litters, all initiated withsmall ova, results in substantially lower values ofreproductive allocation relative to lecithotrophicspecies. Reznick, Meredith & Collette (2007) foundthe same association between matrotrophy, superfe-tation, and reduced reproductive allocation in thefreshwater halfbeaks (Zenarchopteridae).
The combination of placentotrophy with superfeta-tion also enables these species to produce as manyoffspring as (or even more offspring than) leci-thotrophic species without a sacrifice in offspring size,even when devoting less resources to reproduction atany one time (RA; Tables 1, 4). For example, thewild-caught P. branneri had the highest size-adjustedtotal fecundity and the lowest RA values of all popu-lations included in the present study (Table 1).
Thibault & Schultz (1978) hypothesized that thejoint evolution of superfetation and placentotrophycould be advantageous in species exposed to highcurrent conditions because of increased swimmingefficiency. Indeed, increased reproductive allocationcauses reduced swimming performance in P. reticu-lata (Ghalambor et al., 2004). If a reduction in repro-ductive allocation results in increased streamlining ofthe female body, then placentotrophy and superfeta-tion may serve as adaptations that increase swim-ming efficiency. The most suggestive support for thishypothesis to date is provided by the finding ofZuñiga-Vega, Reznick & Johnson (2007) indicatingthat the degree of superfetation increases in highcurrent environments in Poeciliopsis turrubarensis.
Trexler & DeAngelis (2003, in press) offer a secondexplanation for the joint evolution of superfetationand matrotrophy. Their models predict that the priorpresence of superfetation facilitates the subsequentevolution of matrotrophy and that matrotrophy ismore likely to evolve when food availability is high
PLACENTAL EVOLUTION IN POECILIA 793
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796

and constant. If resource availability fluctuates overtime, they predict that lecithotrophy without super-fetation will be the favoured life-history strategy.Again, little is known about resource availability innatural populations of these fish.
An alternative hypothesis for the evolution of pla-centation (Haig, 1993; Crespi & Semeniuk, 2004) isinstead motivated by the fundamental differencebetween placentotrophy and lecithotrophy withrespect to when mothers allocate resources to off-spring. Lecithotrophic species allocate resources toreproduction and determine offspring size prior to thefertilization of the egg. For placentotrophic species,reproductive allocation and offspring size is deter-mined mostly by resources transferred to embryosthroughout pregnancy. This shift in the timing ofallocation of resources to reproduction means that allmaternal provisioning in lecithotrophic species pre-cedes the expression of the paternal genome, whereasmost maternal provisioning in placentotrophic speciesinstead post-dates fertilization and the activation ofthe paternal genome. This overlap between maternalprovisioning and the expression of the paternalgenome creates a forum for intergenomic conflict,which is proposed to lead to an evolutionary ‘tug ofwar’ between the mother and developing embryos.Many of the complex features of mammalian placen-tas are interpretable as a product of this ongoingconflict (Haig, 1993).
Crespi & Semeniuk (2004) carry this argument astep further by proposing that the evolution of vivi-parity without post-fertilization maternal provision-ing sets the stage for the subsequent evolution of theplacenta. Viviparity is accompanied by prolonged,intimate contact between the mother and developingembryos. Trivers (1974) demonstrated that theamount of resources which is in the best interest ofthe embryo to obtain from the mother is greatercompared to that which is in the best interest of themother to allocate to offspring. Viviparity createsthe potential for offspring to gain these additionalresources. Crespi & Semeniuk (2004) propose that thereduction in maternal investment prior to fertiliza-tion and elaboration of the placenta after fertilizationare thus caused by this intergenomic conflict.
Our discovery of a new cluster of closely-relatedspecies that differ both in the presence or absence ofplacentotrophy and superfetation and in the degree towhich each of these traits has evolved in differentspecies creates opportunities for future research. Thesubgenus Micropoecilia now represents the third suchcluster, along with the northern and southern cladesof the genus Poeciliopsis (Reznick et al., 2002). Thus,they expand the potential of the family Poeciliidae toserve as a model system for studying the evolution ofcomplex traits. The fact that this clade includes
guppies, for which there are now emerging genomictools (Dreyer et al., 2007; Tripathi et al., 2009), makesthe Micropoecilia a particularly valuable group forthe future study of the evolution of the placenta, inparticular, and for addressing the more general topicof the genetics and evolution of complex traits.
ACKNOWLEDGEMENTS
We thank Y. Reynoso for dissecting some specimensused in the present study. We are grateful to F.Breden, Heather Alexander, Armando Pou, Jaap deGreef, Manfred Schartl, and the São Paulo UniversityMuseum of Zoology, Brazil (MZUSP), in particularM. de Pinna, C. R. Moreira, and C. A. Figueiredo, forproviding us with some specimens and/or tissuesamples. Live specimens of P. branneri were importedfrom Brazil under license number 0132131 BR fromIBAMA. This work was funded by the University ofCalifornia, Riverside Academic Senate and by a NSFgrant DEB0416085 to D.N.R. and M.S.S.
REFERENCES
Arias A, Reznick D. 2000. Life history of Phalloceros cau-diomaculatus: a novel variation on the theme of livebearingin the family Poeciliidae. Copeia 2000: 792–798.
Banet AI, Reznick DN. 2008. Do placental species abortoffspring? Testing an assumption of the Trexler-DeAngelismodel. Functional Ecology 22: 323–331.
Blackburn DG. 2006. Squamate reptiles as model organismsfor the evolution of viviparity. Herpetological Monographs20: 131–146.
Breden F, Ptacek MB, Rashed M, Taphorn D,Figueiredo CA. 1999. Molecular phylogeny of the live-bearing fish genus Poecilia (Cyprinodontiformes: Poecili-idae). Molecular Phylogenetics and Evolution 12: 95–104.
Costa WJEM, Sarraf A. 1997. Poecilia (Lebistes) minima, anew species of neotropical poeciliid fish from the BrazilianAmazon. Ichthyological Exploration of Freshwaters 8: 185–191.
Crespi B, Semeniuk C. 2004. Parent–offspring conflict inthe evolution of vertebrate reproductive mode. AmericanNaturalist 5: 635–653.
Darwin C. 1859. On the origin of species by means of naturalselection, or the preservation of favoured races in thestruggle for life. London: John Murray.
Dreyer C, Hoffmann M, Lanz C, Willing EM, Riester M,Worthmann N, Sprecker A, Tripathi N, Henz SR,Weigel D. 2007. ESTs and EST-linked polymorphisms forgenetic mapping and phylogenetic reconstruction in theguppy, Poecilia reticulata. BMC Genomics 8: 269.
Evans JP, Rutstein AN. 2008. Postcopulatory sexual selec-tion favours intrinsically good sperm competitors. Behav-ioral Ecology and Sociobiology 62: 1167–1173.
Evans JP, Zane L, Francescato S, Pillastro A. 2003.Directional postcopulatory sexual selection revealed by arti-ficial insemination. Nature 421: 360–363.
794 M. N. PIRES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796

Farley R. 1998. Matrotrophic adaptations and early stagesof embryogenesis in the desert scorpion Paruroctonusmesaensis (Vaejovidae). Journal of Morphology 237: 187–211.
Figueiredo CA. 2003. Análise cladística da subfamíliaPoeciliidae (Cyprinodontiformes: Poeciliidae) com ênfase nasinter-relações dos gêneros Poecilia, Limia e Pamphorichthys.PhD Thesis, Universidade de São Paulo.
Ghalambor CK, Reznick DN, Walker JA. 2004. Con-straints on adaptive evolution: The functional trade-offbetween reproduction and fast-start swimming performancein the Trinidadian guppy (Poecilia reticulata). AmericanNaturalist 164: 38–50.
Grove BD, Wourms JP. 1991. The follicular placenta of theviviparous fish, Heterandria formosa. I. Ultrastructure anddevelopment of the embryonic absorptive surface. Journal ofMorphology 209: 265–284.
Grove BD, Wourms JP. 1994. Follicular placenta of theviviparous fish, Heterandria formosa: II. Ultrastructure anddevelopment of the follicular epithelium. Journal of Mor-phology 220: 167–184.
Gunn JS, Thresher RE. 1991. Viviparity and the reproduc-tive ecology of clinid fishes (Clinidae) from temperate Aus-tralian waters. Environmental Biology of Fishes 31: 323–344.
Haig D. 1993. Genetic conflicts in human pregnancy. Quar-terly Review of Biology 68: 495–532.
Hamilton A. 2001. Phylogeny of Limia (Teleostei: Poeciliidae)based on NADH dehydrogenase subunit 2 sequences.Molecular Phylogenetics and Evolution 19: 277–289.
Haynes JL. 1995. Standardized classification of poeciliiddevelopment for life history studies. Copeia 1995: 147–154.
Hrbek T, Seckinger J, Meyer A. 2007. A phylogenetic andbiogeographic perspective on the evolution of poeciliidfishes. Molecular Phylogenetics and Evolution 43: 986–998.
Hubbs CL. 1926. Studies of the fishes of the order Cyprin-odontes. VI. Miscellaneous publications – University ofMichigan. Museum of Zoology 16: 1–87.
Huebner E, Lococo DJ. 1994. Oogenesis in a placentalviviparous onychophoran. Tissue Cell 26: 867–889.
Korniushin AV, Glaubrecht M. 2003. Novel reproductivemodes in freshwater clams: brooding and larval morphologyin Southeast Asian taxa of Corbicula (Mollusca, Bivalvia,Corbiculidae). Acta Zoologica (Stockholm) 84: 293–315.
Lucinda PHF, Reis RE. 2005. Systematics of the familyPoeciliidae Bonaparte (Cyprinodontiformes: Poeciliidae),with an emphasis on the tribe Cnesterodontini Hubbs.Neotropical Ichthyology 3: 1–60.
Magurran AE. 2005. Evolutionary ecology: the Trinidadianguppy. Oxford University Press, Oxford, UK.
Marsh-Matthews E, Deaton R. 2006. Resources and off-spring provisioning: a test of the Trexler-DeAngelis modelfor matrotrophy evolution. Ecology 87: 3014–3020.
Meisner AD, Burns JR. 1997. Viviparity in the halfbeakgenera Dermogenys and Nomorhamphus (Teleostei: Hemir-amphidae). Journal of Morphology 234: 295–317.
Meredith RW, Pires MN, Reznick DN, Springer MS. 2010.Molecular phylogenetic relationships and the evolution of the
placenta in Poecilia (Micropoecilia) (Poeciliidae: Cyprinodon-tiformes). Molecular Phylogenetics and Evolution (2009),doi:10.1016/j.ympev.2009.11.006.
Meyer MK. 1993. Reinstatement of Micropoecilia Hubbs,1926, with a redescription of M. bifurca (Eigenmann, 1909)from Northeast South America (Teleostei, Cyprinodonti-formes: Poeciliidae). Zoologische Abhandlungen 47: 121–130.
Pilastro A, Simonato M, Bisazza A, Evans JP. 2004.Cryptic female preference for colorful males is guppies.Evolution 58: 665–669.
Pires MN, McBride KE, Reznick DN. 2007. Interpopula-tion variation in life-history traits of Poeciliopsis prolifica:implications for the study of placental evolution. Journal ofExperimental Zoology Part A 307A: 113–125.
Poeser FN, Kempkes M, Isbrücker IJH. 2005. Descriptionof Poecilia (Acanthophacelus) wingei n. sp. from Paría Pen-insula, Venezuela, including notes on AcanthophacelusEigenmann, 1907 and other subgenera of Poecilia Bloch andSchneider, 1801 (Teleostei, Cyprinodontiformes, Poecili-idae). Contributions to Zoology 74: 97–115.
Reznick D. 1981. ‘Grandfather effects’: the genetics of inter-population differences in offspring size in the mosquito fish.Evolution 35: 941–953.
Reznick DN. 1983. The structure of guppy life histories: thetradeoff between growth and reproduction. Ecology 64: 862–873.
Reznick DN, Bryga H, Endler JA. 1990. Experimentallyinduced life history evolution in a natural population.Nature 346: 357–359.
Reznick DN, Callahan H, LLauredo R. 1996. Maternaleffects on offspring quality in poeciliid fishes. AmericanZoologist 36: 147–156.
Reznick D, Endler JA. 1982. The impact of predation on lifehistory evolution in Trinidadian guppies (Poecilia reticu-lata). Evolution 177: 160–177.
Reznick D, Hrbek T, Caura S, de Greef J, Roff D. 2007.Life history of Xenodexia ctenolepis: implications for lifehistory evolution in the family Poeciliidae. BiologicalJournal of the Linnean Society 92: 77–85.
Reznick D, Meredith R, Collette BB. 2007. Independentevolution of complex life history adaptations in two familiesof fishes, live-bearing halfbeaks (Zenarchopteridae, Beloni-formes) and Poeciliidae (Cyprinodontiformes). Evolution 61:2570–2583.
Reznick DN, Mateos M, Springer MS. 2002. Independentorigins and rapid evolution of the placenta in the fish genusPoeciliopsis. Science 298: 1018–1020.
Reznick DN, Miles DB. 1989. Review of life history patternsin poeciliid fishes. In: Meffe GK, Snelson JrFF, eds. Ecologyand evolution of livebearing fishes. Upper Saddle River, NJ:Prentice-Hall, Inc., 125–148.
Reznick DN, Miles DB, Winslow S. 1992. Life history ofPoecilia picta (Poeciliidae) from the island of Trinidad.Copeia 1992: 782–790.
Rodriguez CMI. 1997. Phylogenetic analyses of the tribePoeciliini (Cyprinodontiformes, Poeciliidae). Copeia 1997:663–679.
PLACENTAL EVOLUTION IN POECILIA 795
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796

Roff DA. 2002. Life history evolution. Sunderland, MA:Sinauer Associates.
Rosen DE, Bailey RM. 1963. The poeciliid fishes (Cyprin-dontiformes), their structure, zoogeography, and systemat-ics. Bulletin of the American Museum of Natural History126: 1–176.
Schrader M, Travis J. 2005. Population differences in pre-and post-fertilization offspring provisioning in the LeastKillifish Heterandria formosa. Copeia 2005: 649–656.
Scrimshaw NS. 1944a. Embryonic growth in the viviparouspoeciliid, Heterandria formosa. Biological Bulletin 87:37–51.
Scrimshaw NS. 1944b. Superfetation in poeciliid fishes.Copeia 1944: 180–183.
Shories S, Meyer MK, Schartl M. 2009. Description ofPoecilia (Acanthophacelus) obscura n. sp., (Teleostei: Poecil-lidae), a new guppy species from western Trinidad, withremarks on P. wingei and the status of the “Endler’s guppy.Zootaxa 2266: 35–50.
Thibault RE, Schultz RJ. 1978. Reproductive adaptationsamong viviparous fishes (Cyprinodontiformes: Poeciliidae).Evolution 32: 320–333.
Travis J, Farr JA, Henrich S, Cheong RT. 1987. Testingtheories of clutch overlap with the reproductive ecology ofHeterandria formosa. Ecology 68: 611–623.
Trexler JC, DeAngelis DL. 2003. Resource allocation inoffspring provisioning: an evaluation of the conditions favor-ing the evolution of matrotrophy. American Naturalist 162:574–585.
Trexler JC, DeAngelis DL. In press. Modeling the evolu-tion of complex reproductive adaptations in poeciliid fishes:matrotrophy and superfetation. In: Uribe MC, Greer HJ,eds. Viviparous fishes II. Orlando, FL: NewLife Publica-tions.
Tripathi N, Hoffmann M, Willing EM, Lanz C, Weigel D,Dreyer C. 2009. Genetic linkage map of the guppy, Poeciliareticulata, and quantitative trait loci analysis of male sizeand colour variation. Proceedings of the Royal Society ofLondon Series B, Biological Sciences 276: 2195–2208.
Trivers RL. 1974. Parent–offspring conflict. American Zoolo-gist 14: 249–264.
Turner CL. 1937. Reproductive cycles and superfetation inpoeciliid fishes. Biological Bulletin 72: 145–164.
Turner CL. 1940. Pseudoamnion, pseudochorion, and follicu-lar pseudoplacenta in poeciliid fishes. Journal of Morphol-ogy 67: 59–89.
Woollacott RM, Zimmer RL. 1975. Simplified placenta-likesystem for transport of extraembryonic nutrients duringembryogenesis of Bugula neritina (Bryozoa). Journal ofMorphology 147: 355–377.
Wourms JP, Grove BD, Lombardi J. 1988. The maternal–embryonic relationship in viviparous fishes. In: Hoar WS,Randal DJ, eds. Fish physiology. New York, NY: AcademicPress, Inc., 1–134.
Zuñiga-Vega JJ, Reznick DN, Johnson JB. 2007. Habitatpredicts reproductive superfetation and body shape in thelivebearing fish Poeciliopsis turrubarensis. Oikos 116: 995–1005.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article:
Table S1. Origin of dissection specimens for all ingroup species included in the study.Table S2. Origin of livestock that were the progenitors of the labstocks used for the laboratory life-historyassays.Table S3. Comparison between matrotrophy index (MI) estimates based on regression models and MI estimatesbased on actual average values for embryonic dry weights at early and late stages of development. Embryos atearly and/or late developmental stages were not available for all populations.
Please note: Wiley-Blackwell are not responsible for the content or functionality of any supporting materialssupplied by the authors. Any queries (other than missing material) should be directed to the correspondingauthor for the article.
796 M. N. PIRES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 784–796
Copyright © 2022 FDOKUMEN




















